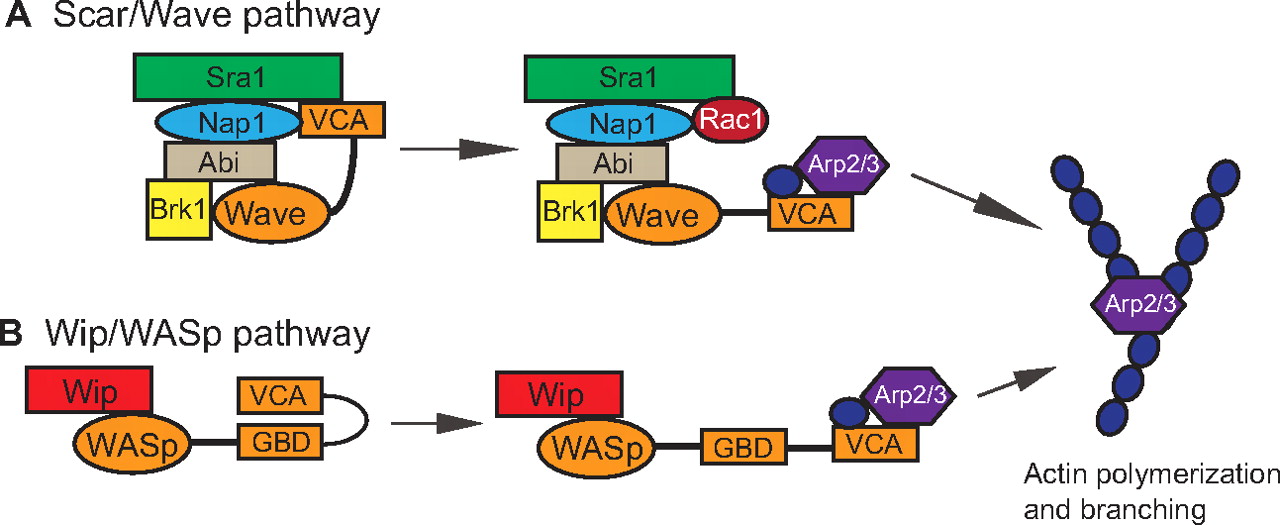
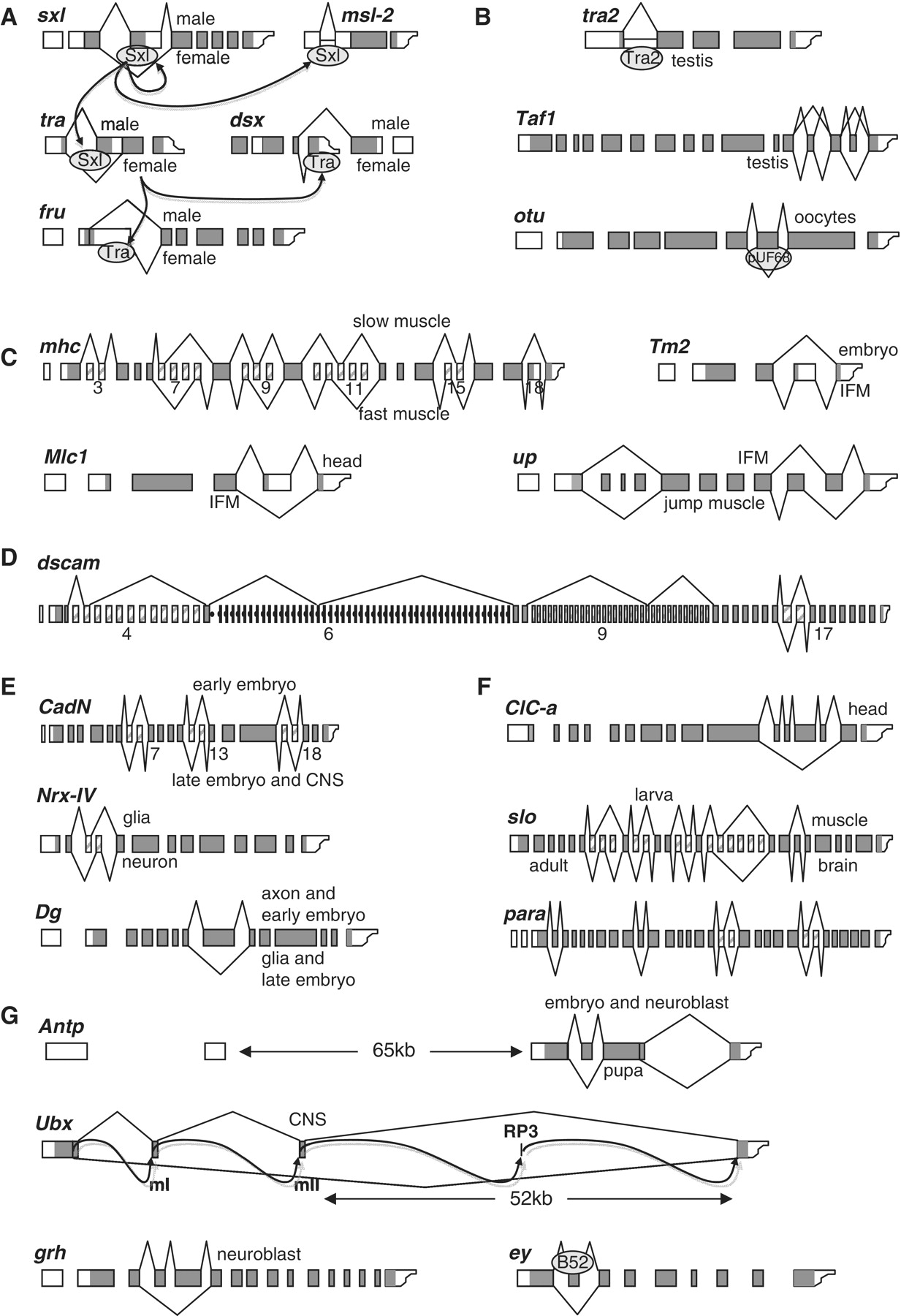
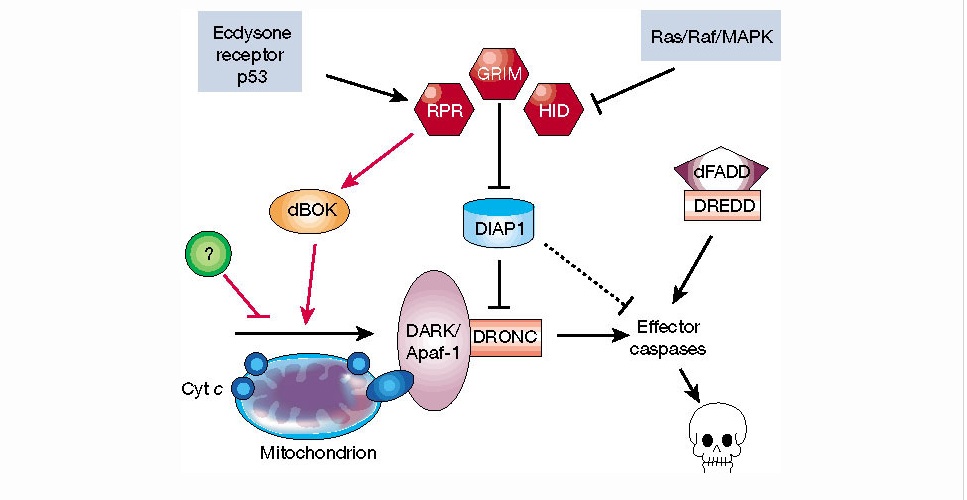
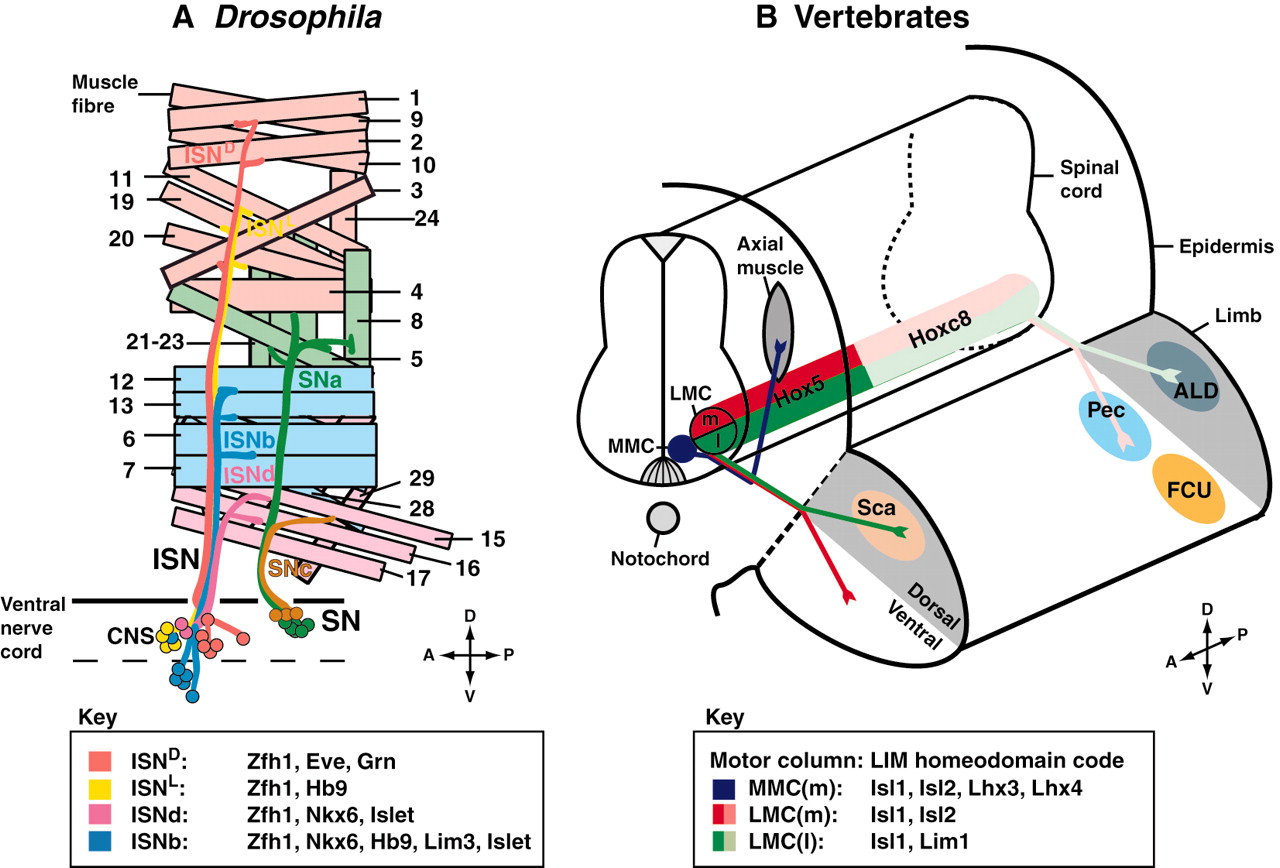
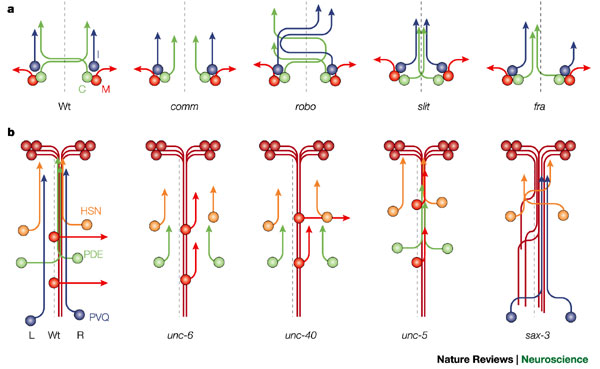
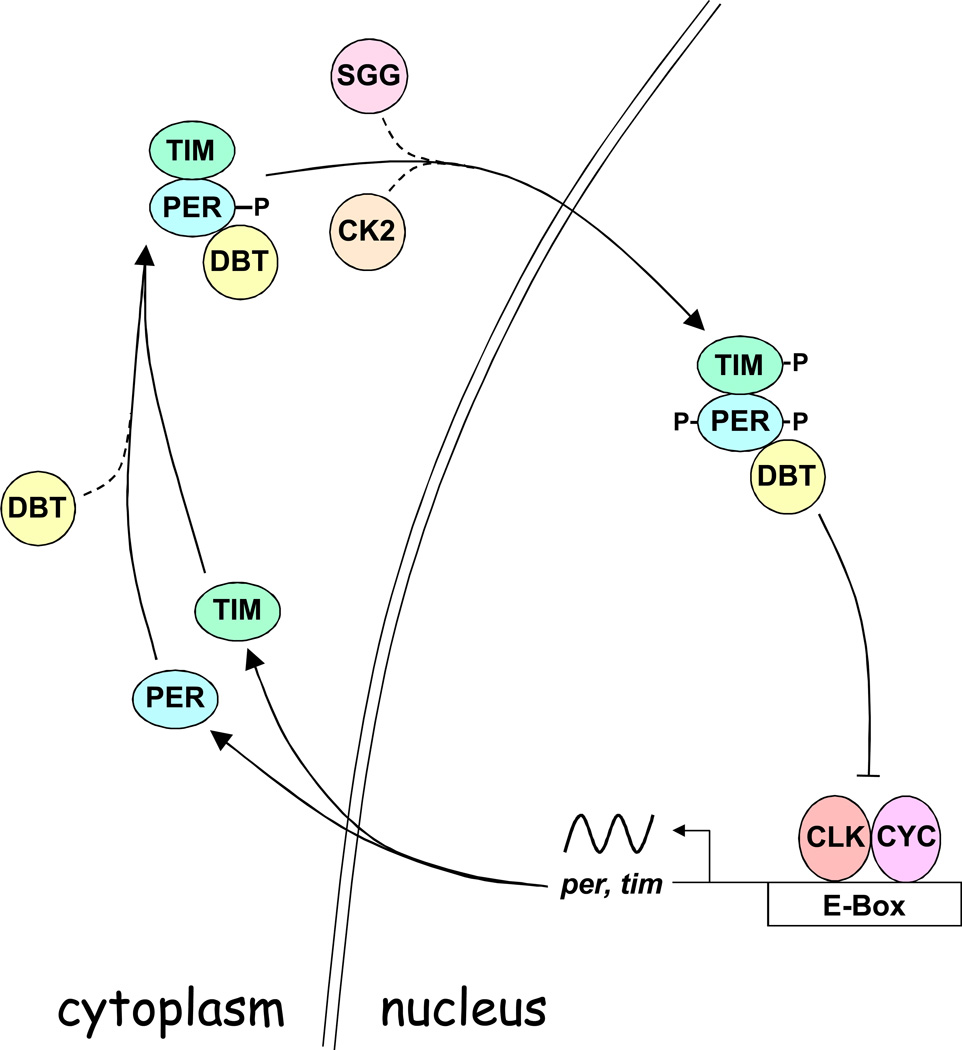
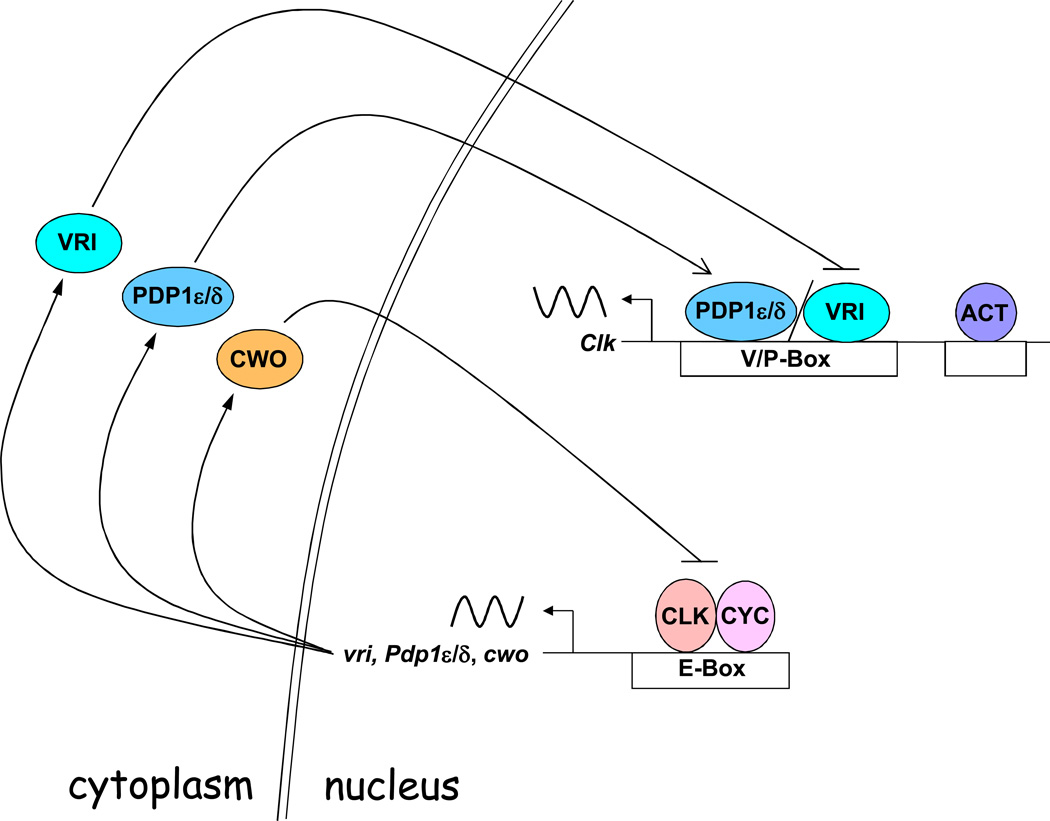
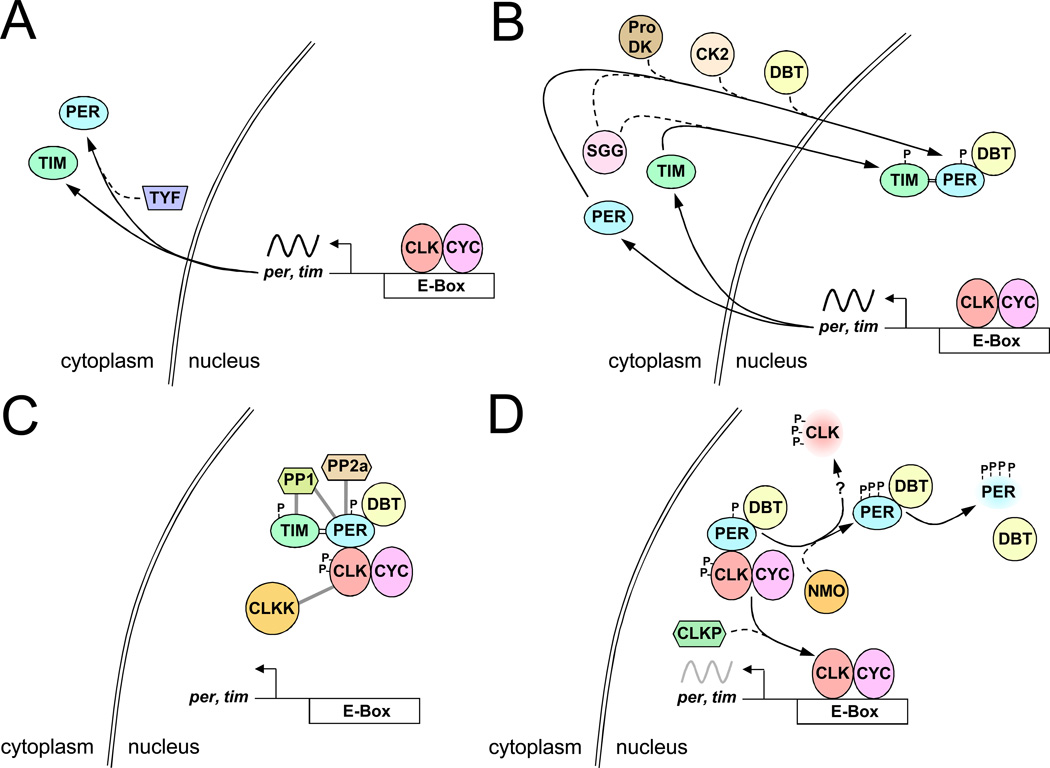
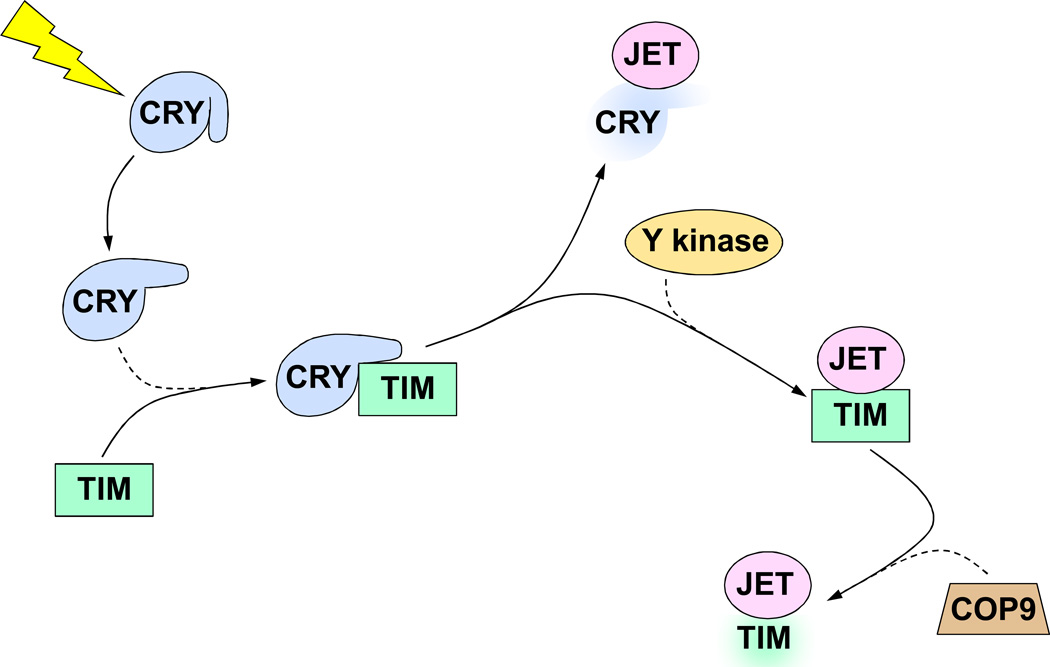
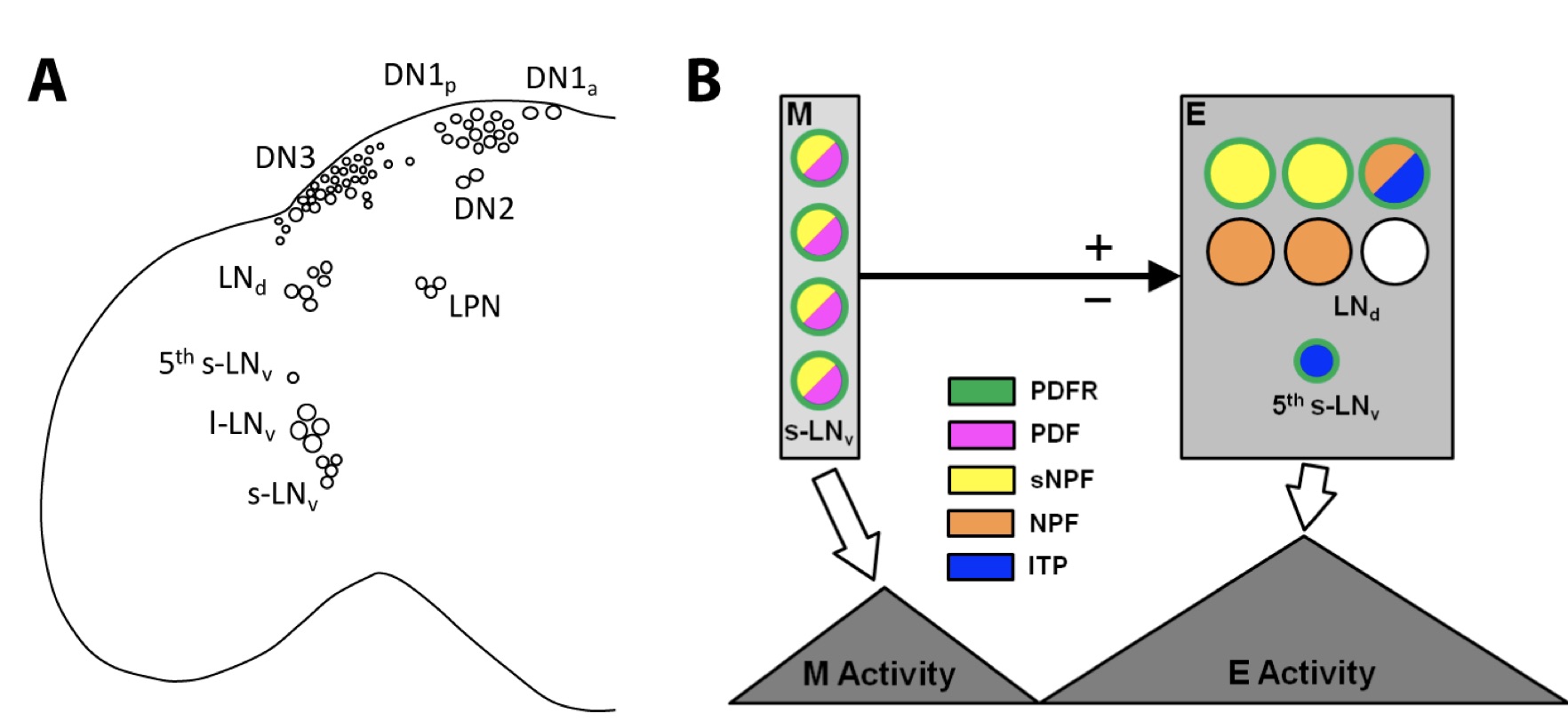
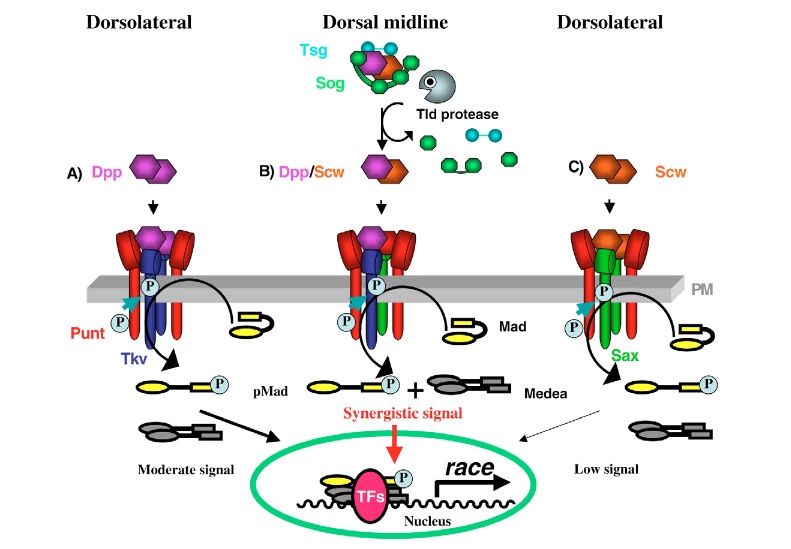
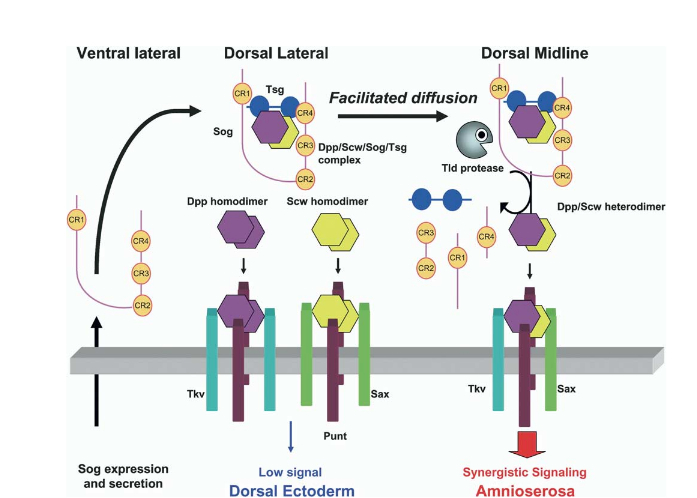
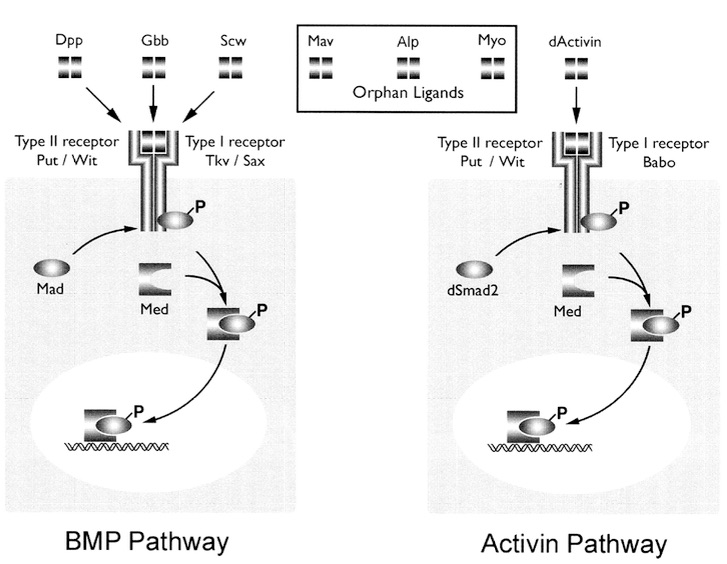
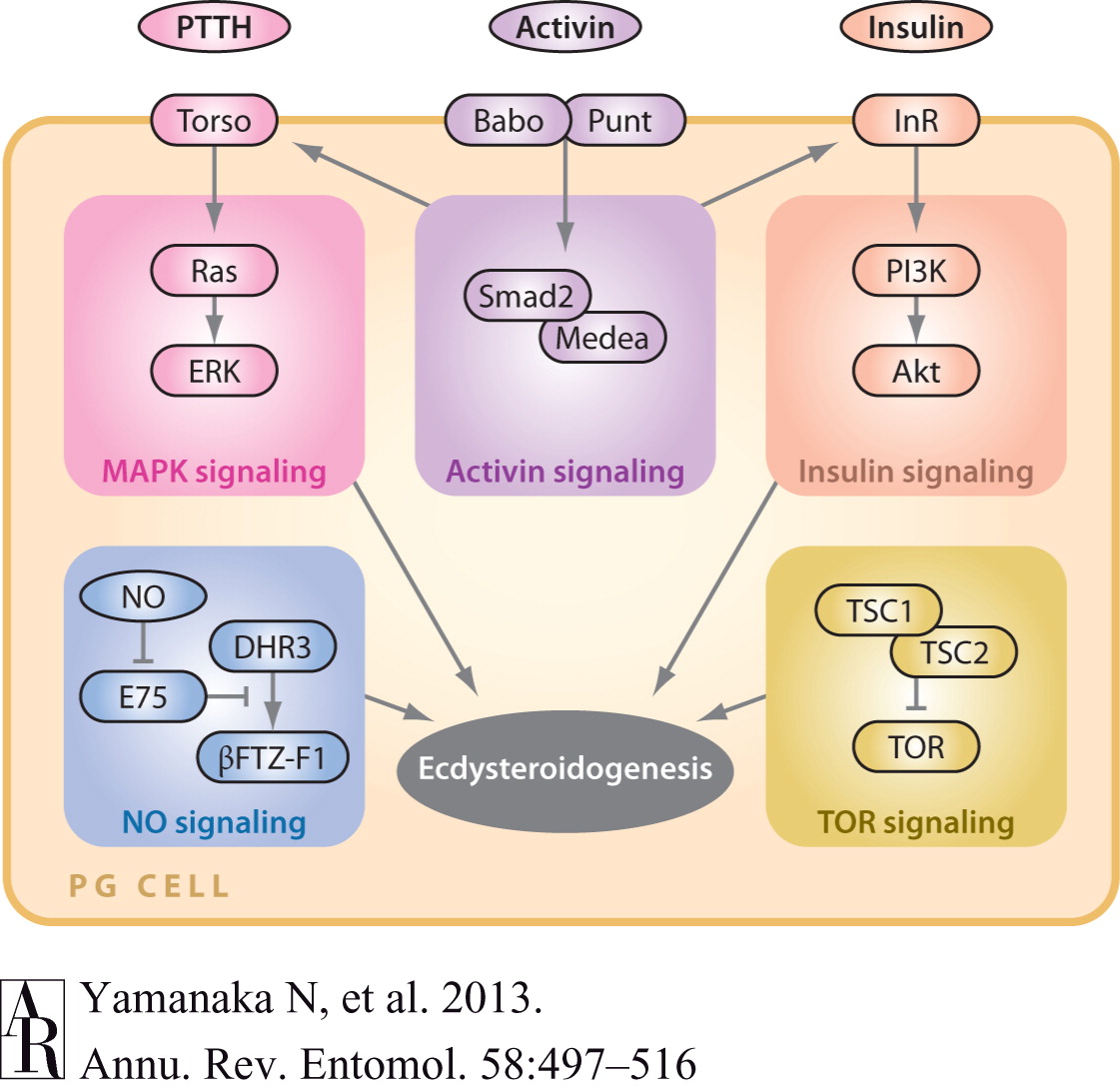
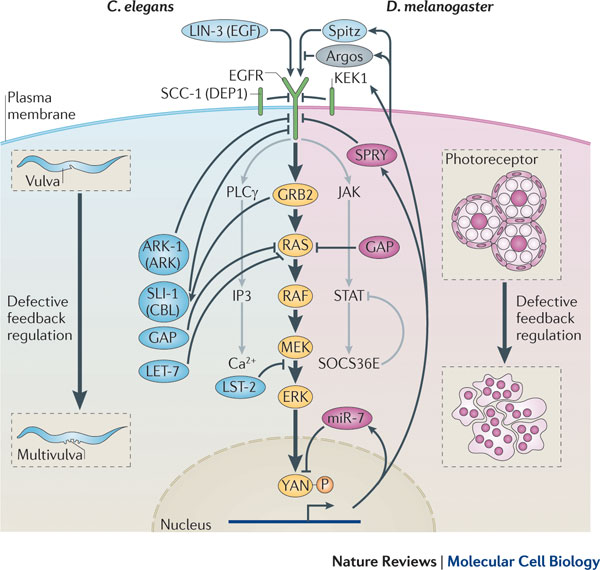
BMP
signaling IV
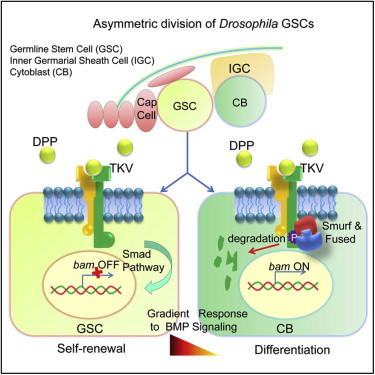
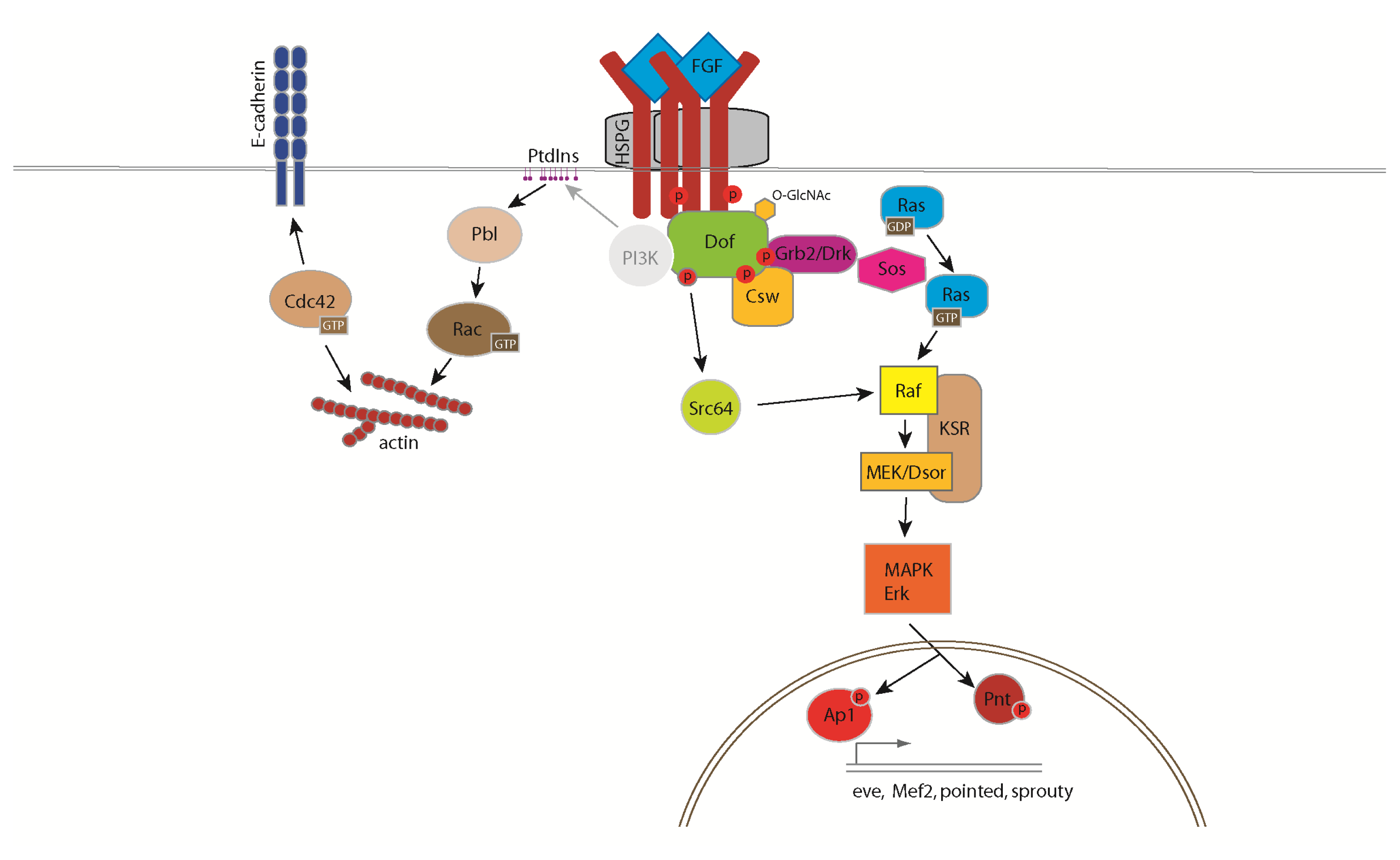
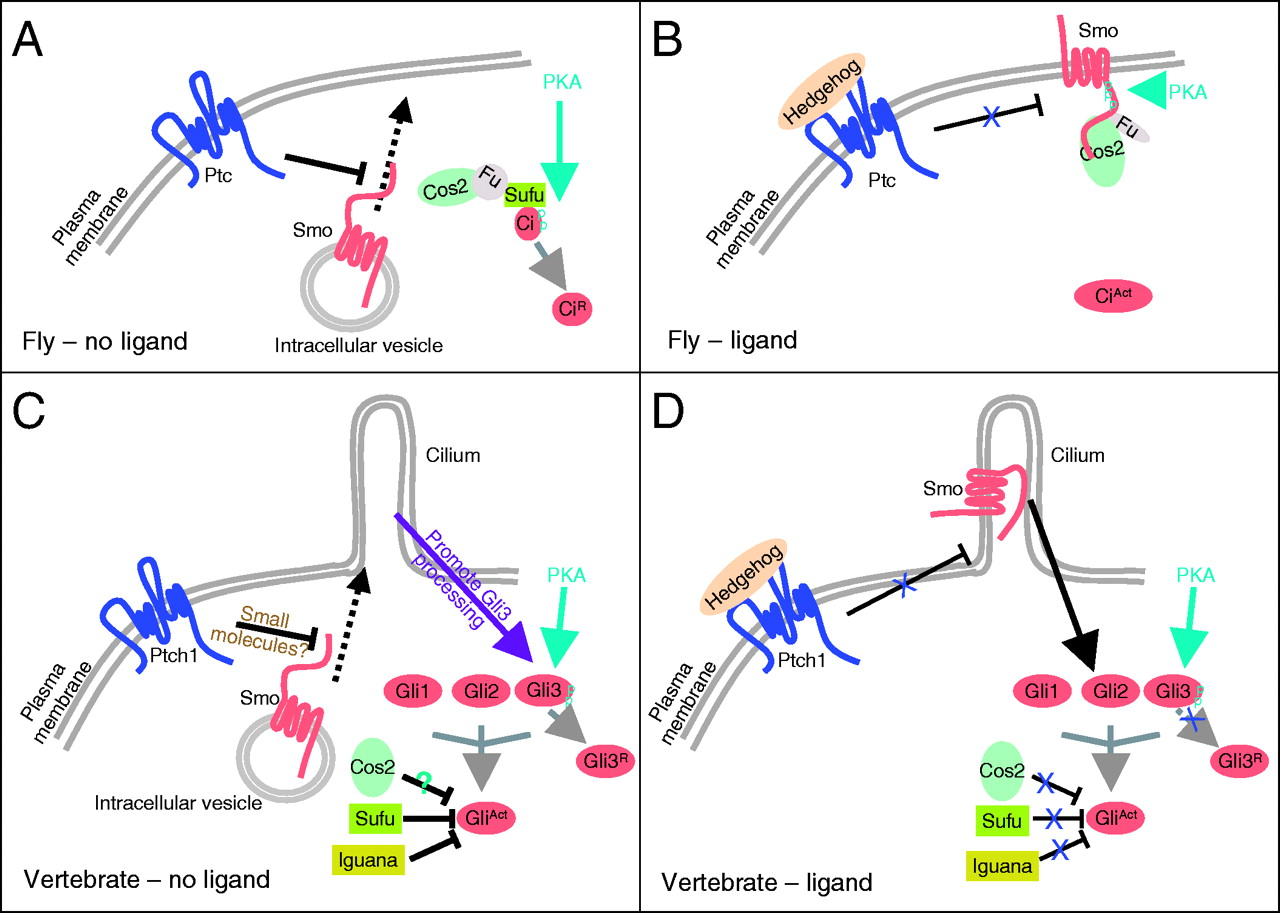
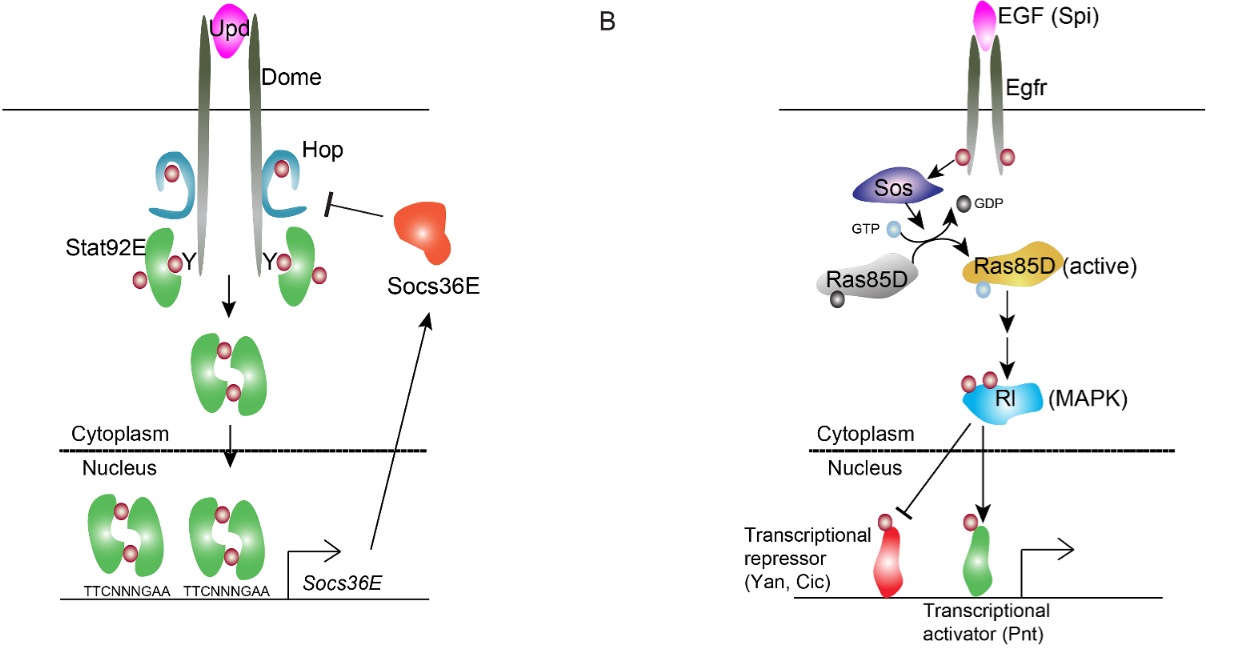
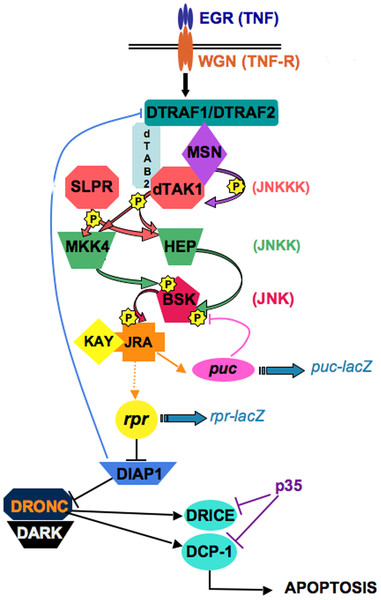
BMP signaling in the Drosophila
ovary. In the Drosophila ovary, BMP signaling from the niche represses GSC differentiation by blocking the
transcription of the differentiation factor Bam. Fused (Fu) functions in concert with the E3 ligase Smurf to regulate
ubiquitination and proteolysis of the BMP receptor Thickveins in
cystoblasts (CBs). This regulation generates a steep gradient of
BMP activity between GSCs and CBs, allowing for bam
expression in CBs and concomitant differentiation (see Xia, 2010).
Xia, L., Jia, S., Huang, S., Wang, H.,
Zhu, Y., Mu, Y., Kan, L., Zheng, W., Wu, D., Li, X., Sun, Q.,
Meng, A. and Chen, D. (2010). The Fused/Smurf complex
controls the fate of Drosophila germline stem cells by
generating a gradient BMP response. Cell 143: 978-990. PubMed ID:
21145463