Select image to enlarge
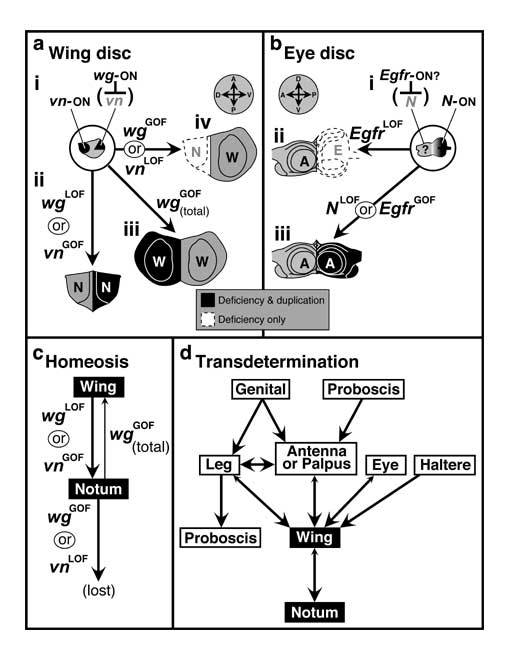
Figure 6.9
Bipolar duality of the wing disc (i.e., wing vs. notum, a) and eye disc (i.e., eye vs. antenna, b), homeotic tendencies in the wing disc (c), and how these tendencies may explain the wing-to-notum trend in transdetermination (d). N.B.: The key for a (ii-iv) and b (ii-iii) straddles the a/b dividing line, and prospective adult axes are given in the compasses above (right wing disc, left eye disc). The letter N in plain type here represents the heminotum, while an italicized N signifies the gene Notch.
a. Genetic control of wing-vs.-notum identity in the wing disc. i. Wing disc (gray) showing regions (solid black) where vein (vn, dorsal spot) and wingless (wg, ventral sector) are transcribed in 2nd instar. Both vn and wg encode diffusible signaling molecules. Although Vn and Wg could probably diffuse into one another's territory and exert mutual inhibition, the chief link that appears critical is Wg —|vn. ii-iv. Abnormal discs resulting from LOF or GOF changes in vn or wg expression. ii. N/N double-heminotum phenotype resulting from wgLOF mutations. The wing portion of the disc has transformed to notal identity. Evidently, vn is expressed by default when Wg is absent. N/N duplications can also be evoked by skinheadLOF [3371], vgLOF [2252] and NLOF [886] (cf. Table 8.1). Although vnGOF has not yet been tested, it is assumed to also behave this way because: (1) Vn is the only essential ligand for Egfr in wing disc development [3025, 3928, 4604], (2) N/N defects can be created by misexpressing Egfr (via omb-Gal4) [4543], and (3) wing-to-notum transformations are seen in EgfrGOF clones [4543]. iii. 'W/W' double-wing phenotype resulting from misexpressing wg throughout the disc (via T80-Gal4:UAS-wg) [3025]. The notal portion of the disc has transformed to wing identity. Apparently, Wg suppresses vn within the dorsal domain (Wg—|vn) and hence triggers the 'wing subroutine' there as well as in wg's native (ventral) domain. A partial W/W trait is rarely seen with vnLOF alleles (omitted), which instead typically cause notal loss (as in iv) [3801, 3929]. Its rarity is likely due to a tendency of wg to remain OFF in the absence of Vn. iv. 'W/-' (notumless) phenotype resulting from vnLOF mutations [4543] or wg misexpression throughout the pouch (via sd-Gal4:UAS-wg) [2252]. The difference in wgGOF effects (iii vs. iv) may be due to a threshold of Wg concentration below which vn can be turned OFF without inducing wing development. Forcing wg to be expressed along the A/P border (via dpp-Gal4:UAS-wg) can yield either outcome (iii or iv) [2252, 3025, 3104].
b. Genetic control of eye-vs.-antenna identity in the eye disc (cf. Fig. 7.1). i. Eye disc (gray) showing the region (dark gradient) where Notch is transcribed in 2nd instar [2353]. Egfr's transcriptional area is unknown; it may complement Notch's. ii. 'A/-' (eyeless) phenotype caused by LOF conditions of the EGFR pathway [2353] due to overexpressing the EGFR inhibitors anterior open, tramtrack, or Bar-C (also caused by wgGOF). See Ch. 7 for 'early eye' genes whose LOF alleles cause a similar phenotype. GOF conditions in the Notch pathway (NGOF) can cause smaller, abnormal eyes (but see [2362]). iii. 'A/A' double-antenna phenotype resulting from LOF effects (DN = dominant negative) in the Notch pathway (NDN, DlDN, SerDN, or mastermindDN, but not Su(H) or m8DN) or from GOF effects in the EGFR pathway (EgfrGOF, spitzGOF, Ras1GOF, dRafGOF, or pointedGOF, but not dMAPKGOF or dMEKGOF) [2353]. The eye portion of the disc has transformed to antennal identity. A comparaably complete transformation of the opposite kind (E/E) has never been found (but cf. Ch. 8)
c. Summary of data from a.ii-iv in terms of histotype identities. Cells can undergo a wing-to-notum switch in two ways (thicker arrow denotes greater likelihood) but a notum-to-wing switch in only one way.
d. Switches in histotype fate seen during long-term culture of disc fragments. Double-tipped arrows mean that switching can occur in either direction (size of arrowhead denotes relative frequency). Top-to-bottom order reflects the favored flow of events, so the notal fate is a 'sink' for the system. The tendency of wing tissue to transdetermine to notum (d) may be due to the ease with which such a transformation can occur homeotically (c). 'Proboscis' appears twice (as per original data [1421]), which poses a paradox for any scheme that tries to rank histotypes hierarchically. Although no arrow is shown from eye to antenna, such switches probably do occur during culture since (1) antennal fragments duplicate themselves while eye fragments regenerate antennae [1404, 1406, 1423, 3810] and (2) LOF mutations in various genes transform eye to antennal tissue (but not vice versa; cf. Table 8.1) [2362]. Third-instar vnLOF leg discs can transdetermine to wing (not shown) -- even though mature vnLOF larvae lack wing discs -- because they bypass the 2nd-instar need for vn [4768].
Schematic in a.i is based on [3928, 3929, 4543] for vn-ON and [2252, 2253, 3089, 3104, 4683] for wg-ON. Cartoons in a.ii-iv and b.iii are sketches of discs pictured in [2252, 3104, 4543]: (a.ii), [3025, 3104] (a.iii), [2252, 3929] (a.iv), and [2353] (b.iii). Panel b.i is based on [3382], and d is redrawn from [1421] with one arrow omitted (from unidentified tissues to a genital state). See also App. 7.
|
|
{kind=link}