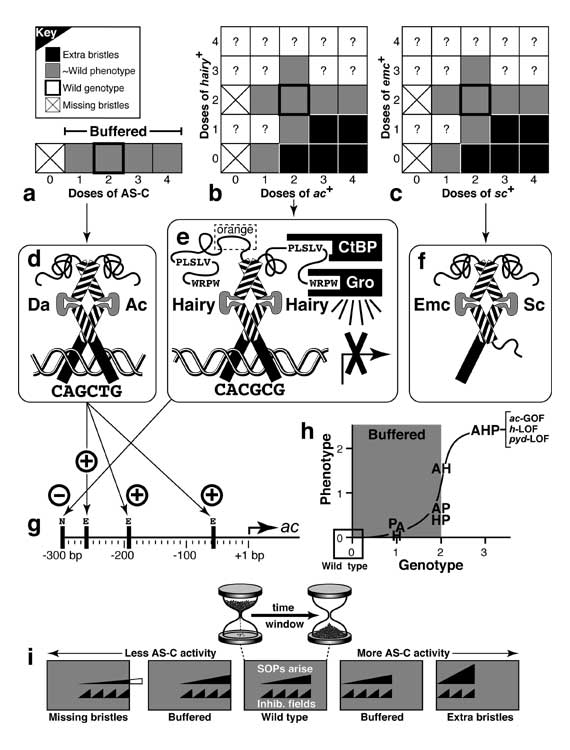
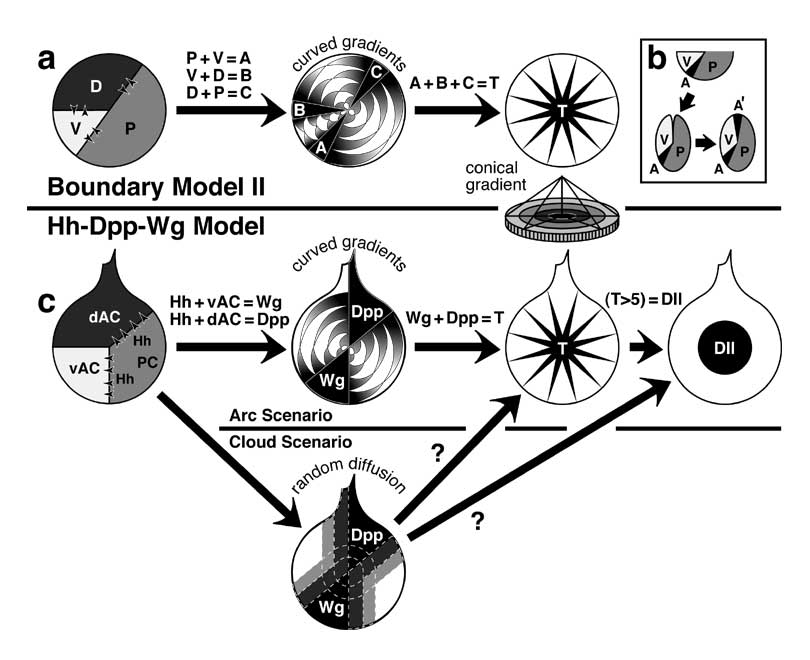
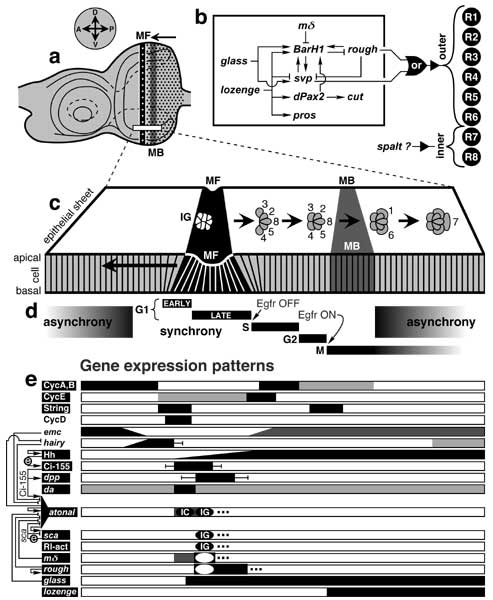
Select image to enlarge
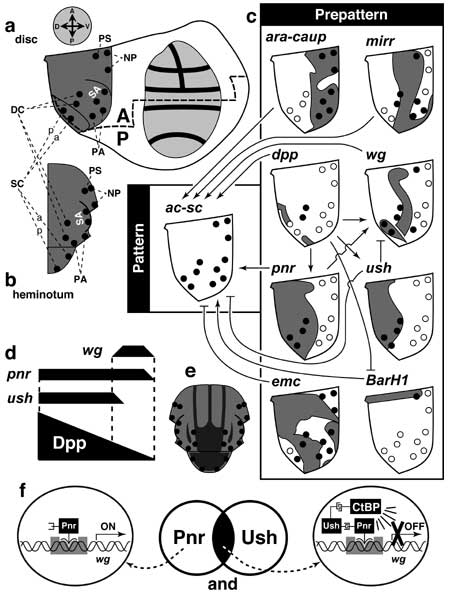
Figure 6.14
Landscape of prepattern factors that evoke macrochaetes on the notum (cf. Fig. 3.4).
a. Fate map of a mature right wing disc (simplified from Fig. 6.1). Dots are actual SOP sites [1925]. Dashed line is the A/P compartment boundary. Directions (cf. compass above): A, anterior; P, posterior; D, dorsal; V, ventral. Within the notal region (dark shading) the future scutellum (posterior end) is demarcated by a thin solid line.
b. Heminotum, with macrochaete sites labeled. Abbreviations: 'a' or 'p' (ant. or post. members per pair), DC (dorsocentrals), NP (notopleurals), PA (post-alars), PS (presutural), SA (supra-alars), SC (scutellars) [2560]. The configuration of SOPs matches the adult pattern, except that scutellar SOPs rearrange ('p' moves posterior to 'a').
c. Heminotal domains (shaded) where certain 'prepattern genes' are expressed in mature discs. Boundaries are not as sharp as depicted -- especially for emc, whose profile of expression has many subtle gradations (cf. Fig. 6.2) including a background level (not shown). Solid circles are bristle sites affected by LOF mutations (cf. Table 6.2); remaining unfilled circles are unaffected. Genetic interactions are indicated by connecting wires (—> activation; —| inhibition). Abbreviations (see references for expression patterns and circuitry): ara-caup (araucan and caupolican) [1536], emc (extramacrochaetae) [913] (see [205]Δ for expression and [1349]Δ for bristle effects), BarH1 (postnotal expression domain not shown) [3763], dpp (decapentaplegic) [3764, 4369], mirr (mirror) [2170], pnr (pannier) [614, 3764, 4369]Δ, ush (u-shaped) [3764, 4369]Δ, wg (wingless) [3764, 4369]Δ. The DC cluster, which forms far from the dpp-ON stripe, depends on both Dpp and Wg signals [3373]. The 'Dpp —|wg' link inferred from LOF-GOF studies [3763]Δ is mediated indirectly by Ush [3764, 4369]. Hence, it is omitted, as is a 'Wg —> BarH1' link that only works in part of the notum [3763]. The 'ush —|wg' link requires pnr (not shown; see below) [3764, 4369]. The genes ara and caup are each expressed like the depicted irorF209 Iro-C ('ara-caup') enhancer trap [1536]. In co-stained specimens, their expression appears more complementary to that of pnr than depicted here [614]. Indeed, dorsal pnrLOF clones turn ON Iro-C, indicating that 'pnr —| Iro-C' (not shown) [614]. Moreover, such clones sort out from the epithelium -- implying a difference in affinity [614]. The overall circuitry suggests that Pnr sets the V limit of the wg-ON stripe (excluding the scutellar spot), while Ush sets its D limit [3764]. In fact, the wg-ON stripe extends a few cells beyond the pnr-ON area in late 3rd instar [614] due to a 'Wg —>wg' link (not shown) that is disabled in wgLOF or dshLOF discs [4369]. The Gal4 insertion line em462, which exhibits a wg-like A-P stripe [615], is omitted because the patterning effects of its host gene are unknown.
d. Dpp gradient that supposedly governs the notal region. Dpp turns ON ush and pnr at different thresholds (high vs. low). Dpp also turns ON wg, but this activation is overridden where Pnr and Ush overlap. The DC cluster is regulated similarly but lies more dorsally than the wg-ON stripe [3373] -- presumably because the AS-C's DC cis-enhancer responds to slightly higher levels of Pnr and Ush than wg [1380].
e. 'Trident' pattern of notal pigmentation, which is variably detectable in wild-type stocks [2982, 4511]. The pattern may reflect a control at the yellow or ebony locus that relies on the kinds of prepattern agents in c [1435, 1442, 1443, 2968, 3055]. This pattern was the subject of Morgan's earliest (1909) artificial selection experiments with Drosophila [471, 2283]. The outer stripes of the trident are strikingly prominent in the dipteran genus Zaprionus [65]. See also the beautiful 'picture wing' flies of Hawaii [669, 4397].
f. Model for how the '{Pnr AND Ush} —|wg' link is implemented at a molecular level. Circles (center) are a Venn diagram (cf. Fig. 6.10). In the absence of Ush, Pnr is a transcriptional activator (left) [1380]. Ush is thought to mediate repression by recruiting dCtBP (right) [3764] as does Hairy (cf. Fig. 3.12). Direct binding of Pnr and Ush to the wg promoter has not yet been shown [1380]. This diagram is oversimplified because Pnr and Pnr-Ush probably do not compete for the same DNA sites (but see [4369]): excess Pnr (from C765-Gal4:UAS-pnr) apparently cannot displace Pnr-Ush heterodimers [1380].
Panel c is adapted from [3373, 3764, 3954, 4368]. Registration of the various domains follows [3763] (but see [615]). The thresholds in d summarize results of [3764, 4369], and the model in f is based on data in [1380, 1671, 3764].
N.B.: None of the boundaries in c restricts cell lineage [521, 615, 1373, 3007, 3143] (cf. the Cabaret Metaphor; Ch. 4): the sole constraint on lineage in the heminotum is the A/P boundary [1376, 1377]. The stripes where Dpp activates target genes are oriented at acute angles relative to the dpp-ON stripe -- perhaps due to modulated diffusion rates (cf. the Arc Scenario; Fig. 5.4). Like ac and sc here (c), the proneural gene atonal is regulated by a zoo of upstream factors in the eye (Fig. 7.9e), but that menagerie is more like a traveling circus since it migrates across the disc.
|
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}