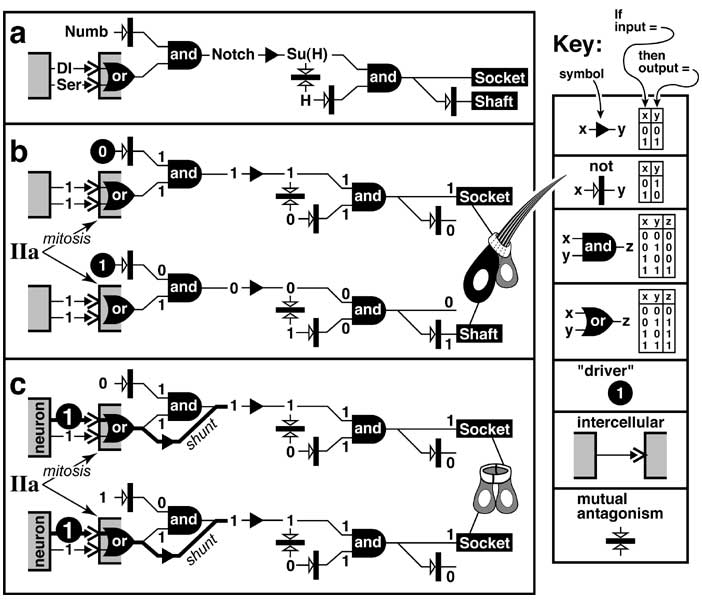
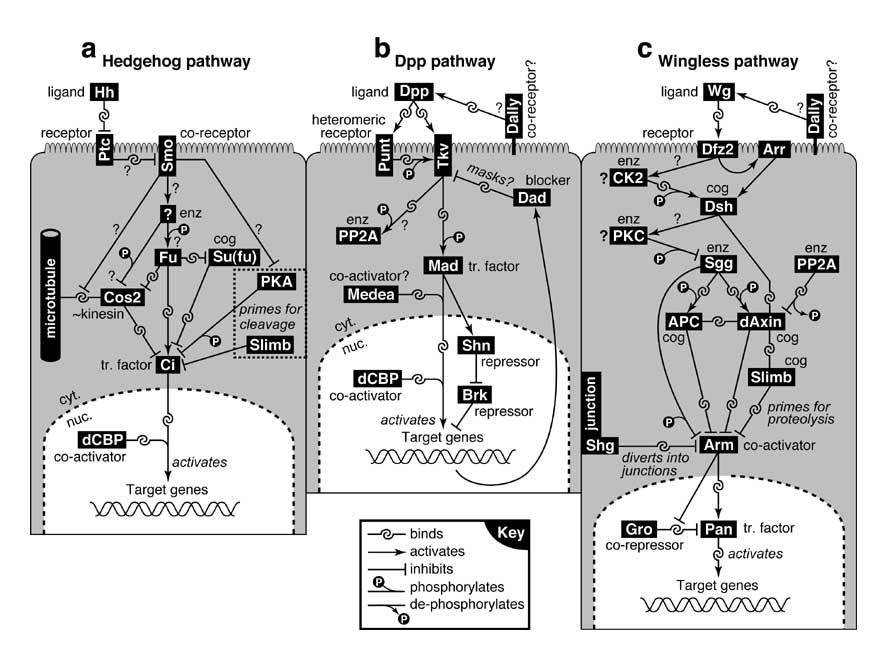
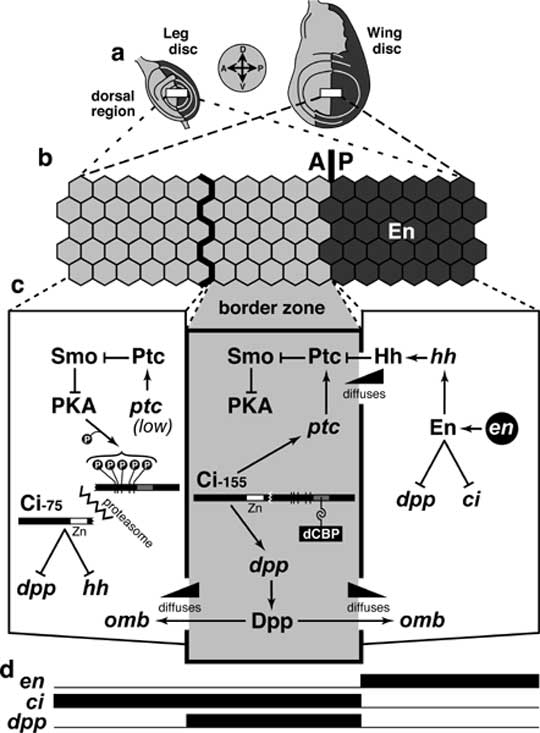
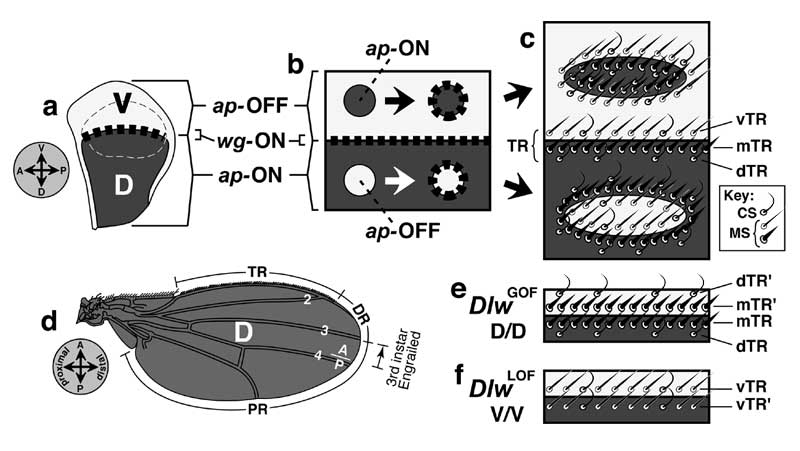
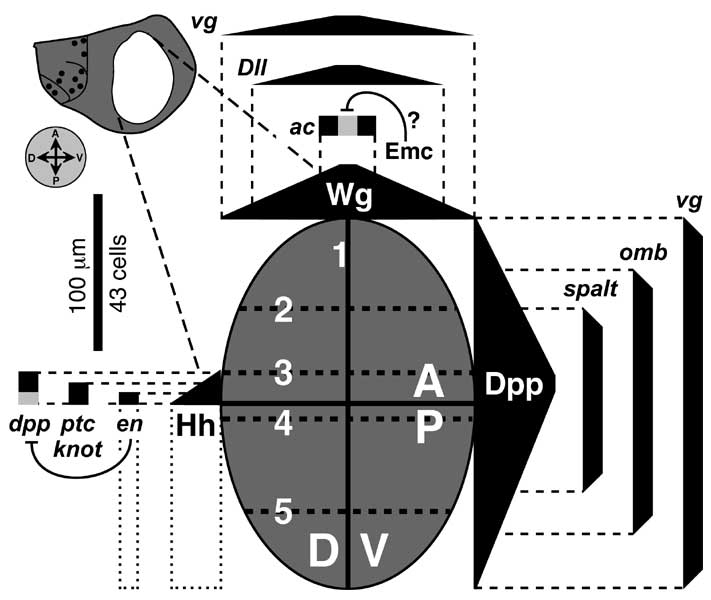
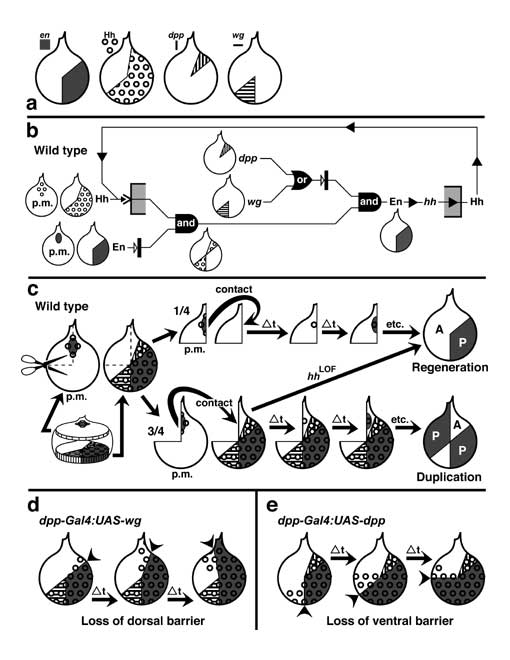
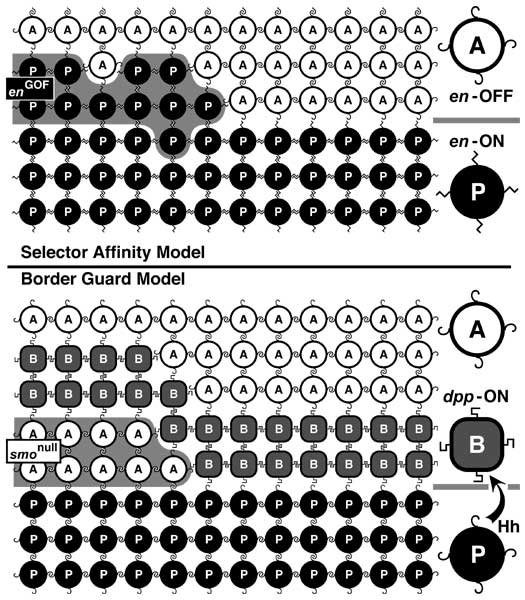
Select image to enlarge
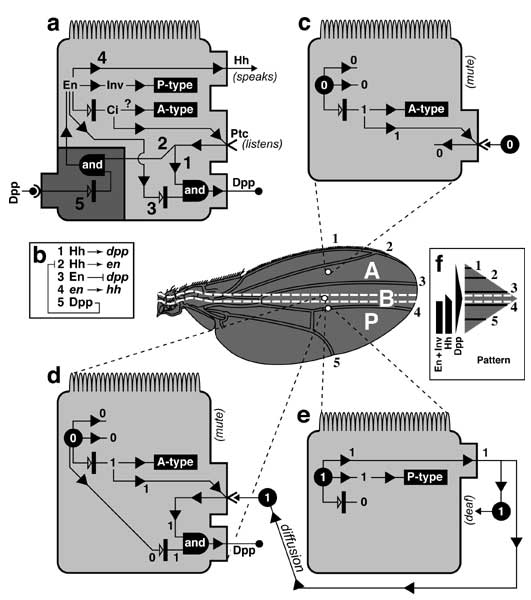
Figure 6.4
Circuitry that enforces A vs. P identities of wing cells and causes expression of dpp at the A/P interface. Abbreviations: Ci (Cubitus Interruptus), Dpp (Decapentaplegic), En (Engrailed), Hh (Hedgehog), Inv (Invected), Ptc (Patched).
a. Generic wing disc cell (cf. Fig. 2.7 for icons) showing the main circuit that operates throughout disc development (light gray), plus a 'plug-in' module that works only in late 3rd instar (dark gray) [1647]. Numbers refer to links in b.
b. Major links, redrawn using epistasis symbols (cf. Figs. 5.6 and 5.7). Link 2 (receipt of Hh signal activates en) only becomes operative in late 3rd instar, when the en-ON state spreads into the A compartment to fill the zone marked 'B' in the wing diagram (cf. Fig. 6.7) [350]. Because Link 2 requires more Hh than Link 1 (not shown) [4136], the band of dpp-ON cells induced by Hh extends farther anteriorly than the band of en-ON cells, and Link 3 then turns OFF dpp where these bands overlap (cf. Fig. 6.3). Link 5 is a 'safety switch' that prevents a runaway cycle of en (Link 2) and hh (Link 4) activation that would convert all cells to a P-type state (cf. Minotaur Scenario; Fig. 5.10c-e; Ch. 5). Link 5 can be overridden by excess Hh (resulting in a 5-fold wider en-ON band) [2992], so an analog pressure clamp would be a more apt analogy than a digital switch. Literally, the clamp is probably the receipt of a maximal Dpp signal by Tkv-Punt receptors, but it might instead be an inability of dpp-ON cells to turn ON en. All we really know is that en-ON states can't traverse dpp-ON tissue. Link 4 is disabled in the B zone [2992] -- because hh is more sensitive to repression by Ci-75 than to activation by Ci-155 [2980]. In any event, the plug-in module is incidental to the grand scheme and is omitted from futher consideration.
c-e. Schematics of cells in 3 different districts of gene expression along the A-P axis before late 3rd instar. 'A' denotes the anterior A compartment beyond the range of diffusible Hh, 'B' is the border zone (of A-type cells) that responds to Hh by forming a stripe of dpp-ON cells, and 'P' is the P compartment. States of variables (cf. a) are recorded as '1' (present) or '0' (absent), though thresholds matter (discussed above). Black circles indicate decisive factors. In plain English, the logic of the main circuit is as follows. If en is OFF (c and d), then the cell takes the Ci route, deploys Ptc (Hh's receptor), and adopts A-type identity, though an agent other than Ci may mediate the latter. If en is ON (e), then the cell secretes Hh and turns ON inv which causes P-type identity. One-way signaling between Hh-secreting 'speakers' and Ptc-competent 'listeners' (d) establishes a dpp-ON stripe in the B zone (cf. Fig. 6.5). Although P cells are bathed in Hh, Link 1 can't work there since (1) they lack Ptc and so are deaf to Hh [328, 4229], (2) they lack Ci (due to En —|ci) and so can't activate dpp (Ci —>dpp link not shown) [1827, 2832, 4229], and (3) En represses dpp directly by binding its cis-enhancers [2980, 3747]. Since cell states are set by whether en is ON and Hh is received, these variables (black circles in c-e) act like a binary code, where the ordered pairs (en ON?; Hh bound?) are 00, 01, and 10.
f. Profiles of protein distributions (black bars or triangles) before late 3rd instar relative to districts of gene expression, which are demarcated on a realistic wing at left and schematic at right. Rear dashed line is A/P boundary (after [497]). Invected (under En control) lets A vs. P cells interpret symmetric Dpp gradients differently to make an asymmetric pattern (i.e., veins 1-3 vs. 4-5) [3934, 3935].
Circuitry in a is based on [643, 1647, 1659, 3935, 4229, 4848], and the gradients in panel f are adapted from [4229, 4734]. The logic of this circuit is confirmed by using dpp-Gal4 to drive UAS-hh. These conditions entrap the system in an 'infinite loop': every time an A-type cell is instructed by Hh to turn ON dpp (d), it must now also turn ON hh, leading to another round of Hh signaling that enlarges the hh-ON area still farther. The outcome of this robotic iteration is a grotesque multiplication of bristles and sensilla that are normally limited to the A/P boundary [2992].
N.B.: Input/output ports (raised rectangles) on the cell surface are at arbitrary apical-basal levels (apex denoted by microvilli). The en —>hh link is written in gene symbols because -- unlike En —|dpp -- it may be indirect [224] since En is normally a repressor (cf. Fig. 5.7 legend). The role of inv is unclear since null alleles have no apparent effect [358]. For other examples of 'safety switches' that override morphogen inputs, see [2176, 3733]
|
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}