Select image to enlarge
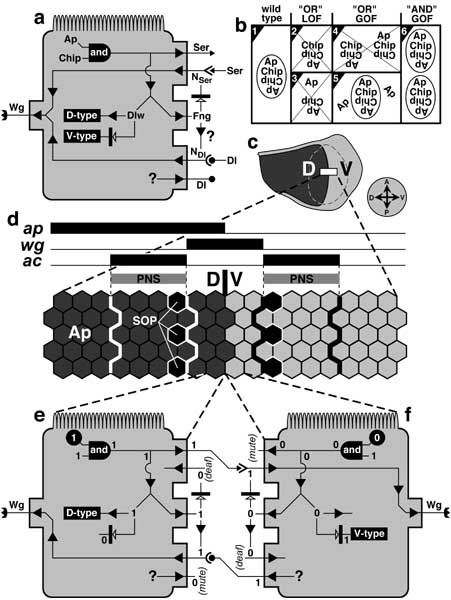
Figure 6.8
Circuitry that enforces D vs. V identities of wing cells and turns wg ON at the D/V line.
a. Generic wing disc cell (cf. Fig. 2.7 for icons) showing the main control circuit for Wg output. Abbreviations: Ap (Apterous), Dl (Delta), Dlw (Dorsal Wing), Fng (Fringe), N (Notch), Ser (Serrate), Wg (Wingless). NSer and NDl are hypothetical Fng-modulated forms of Notch that are receptive to Ser or Dl [990, 1250]. Fng's damping of the Ser-N trigger has been well documented in vivo [1250, 3246], as has its facilitation of Dl-N binding in vitro [510]. The circuit works as follows. If ap is ON (e), then the cell expresses Ser and Fng and adopts D-type identity (via Dlw) but cannot receive Ser signals because Fng blocks the Ser-N interaction. If ap is OFF (f), then the cell expresses Dl (Dl's upstream controls are uncertain [2253]) and adopts V-type identity. Dlw (under Ap control) should allow D vs. V cells to interpret symmetric Wg gradients asymmetrically. The circuit allows two types of discourse: Ser-signal 'speakers' with NSer 'listeners' or Dl-signal 'speakers' with NDl 'listeners'. Both dialogues should turn wg ON. Validation of the circuit's logic comes from (1) heating t.s. NLOF larvae, which erases wg and cut expression in border cells [2839, 3689] and (2) expressing Ser or fng in the D region of apLOF discs, which restores Notch signaling and a wg-ON stripe but not D identity (hence creating V/V symmetric wings) [2850, 3158].
b. Stoichiometry of Ap-Chip interactions and etiology of abnormal traits. Wings look wild-type when Ap and Chip are equimolar (boxes 1 and 6) or when Ap is in excess (box 5), but not otherwise (X'd boxes). Ap binds DNA (not shown) but can only activate target genes when it forms a tetramer (oval) with Chip (box 1). The tetramer is thought to be a Chip dimer with an Ap at each end. Reducing the amount of Ap (box 2) or Chip (box 3) lowers the number of functional complexes. Excess Chip (box 4) diverts Ap into inert trimers, whereas excess Ap (box 5) has no such effect. Raising the dose of both partners ('AND' GOF) maintains the balance and makes excess tetramers (box 6 shows a doubling). The counterintuitive LOF-GOF genetic data only make sense in terms of this sort of jigsaw-puzzle logic. See text for further details. Oddly, Ap and Chip show no such interdependence in leg discs [3474].
c. Mature right wing disc. Shading denotes D (dark) vs. V (light) compartments, and dashed line delimits the pouch.
d. Enlarged part of the anterior D/V border (box in c). Hexagons are cells. All cells in the D region express Ap. Zigzag lines delineate 3 zones (horizontal bars above): a median stripe where wg is ON (also cut and N: not shown) and flanking 'proneural stripes' (PNS) where achaete (ac) is ON (also scute, with intense Dl and Ser but meager N: not shown) [988]. Sense organ precursors (SOPs, black cells) arise in each PNS along the medial (high-Wg) edge [3689]. In cutLOF discs the wg-ON cells express ac [882, 988], so Cut's normal role must be to split the otherwise broad ac-ON stripe into two parts.
e, f. Schematics of cells flanking the D/V line. The V cell is drawn as if flipped around to face the D cell. States of variables (cf. a) are recorded as '1' (present and active) or '0' (absent or inactive). Black circles indicate determining factors. 'Mute' and 'deaf' denote inability to send or receive particular signals (Ser above, Dl below). The functioning of the circuit is explained under a above.
Circuitry in a is compiled from [1040, 2216, 2252, 3089, 3689] in general, [1217] for Chip, [1991, 1992, 2253] for Fringe, and [4325] for Dlw. Panel b illustrates data from [1217, 2851, 4456]. It is not known whether the tetramer's symmetry is rotational (as shown) or reflective. Panel d is adapted from [882, 2079, 3689, 3690, 3979]; and the dialogue cartoon (e, f) is based on [988, 994, 1074, 2839, 3246].
N.B.: Input/output ports (raised rectangles) on the cell surface are at arbitrary apical-basal levels (apex laced with microvilli). The reciprocity of the D/V dialogue is a realization of Boundary Model II (cf. Fig. 5.4a) [2808], whereas signaling is one-way across the A/P line (cf. Fig. 6.3).
|
|
{kind=link}
{kind=link}
{kind=link}