The Interactive Fly
Genes involved in tissue and organ development
INDEX
Schematic drawing of the adult Drosophila visual system: a horizontal section of the visual system including the retina, lamina, lobula and lobula plate and a lateral view of the ommatidial unit (from Waves of differentiation in the fly visual system)
Early optic lobe development and lamina neurogenesis; With the arrival of photoreceptor axons in the optic lobe during the mid third instar larval stage, the outer proliferation center neuroepithelium undergoes a morphological change with the appearance of a deep groove—called the lamina furrow at its lateral edge. Lateral cells gradually give rise to lamina precursor cells located posteriorly of the lamina furrow. Each lamina precursor cells undergoes a terminal symmetric division to generate undifferentiated lamina neurons (from A Challenge of Numbers and Diversity: Neurogenesis in the Drosophila Optic Lobe)
Early optic lobe development and lamina neurogenesis: Medulla neurogenesis depends on the sequential conversion of neuroepithelial (NE) cells to NBs (from A Challenge of Numbers and Diversity: Neurogenesis in the Drosophila Optic Lobe)
Neuronal basis of color vision in Drosophila: Anatomical representation of color processing neurons in the fly optic lobe (from Color vision in insects: insights from Drosophila)
Visual information processing in animals with large image forming eyes is carried out in highly structured retinotopically ordered neuropils. Visual neuropils in Drosophila form the optic lobe, which consists of four serially arranged major subdivisions; the lamina, medulla, lobula and lobula plate; the latter three of these are further subdivided into multiple layers. The visual neuropils are formed by more than 100 different cell types, distributed and interconnected in an invariant highly regular pattern. This pattern relies on a protracted sequence of developmental steps, whereby different cell types are born at specific time points and nerve connections are formed in a tightly controlled sequence that has to be coordinated among the different visual neuropils. The developing fly visual system has become a highly regarded and widely studied paradigm to investigate the genetic mechanisms that control the formation of neural circuits. However, these studies are often made difficult by the complex and shifting patterns in which different types of neurons and their connections are distributed throughout development. This study has reconstructed the three-dimensional architecture of the Drosophila optic lobe from the early larva to the adult. Based on specific markers, it was possible to distinguish the populations of progenitors of the four optic neuropils and map the neurons and their connections. This paper presents sets of annotated confocal z-projections and animated 3D digital models of these structures for representative stages. The data reveal the temporally coordinated growth of the optic neuropils, and clarify how the position and orientation of the neuropils and interconnecting tracts (inner and outer optic chiasm) changes over time. Finally, the emergence of the discrete layers of the medulla and lobula complex were analyzed using the same markers (DN-cadherin, Brp) employed to systematically explore the structure and development of the central brain neuropil. This work will facilitate experimental studies of the molecular mechanisms regulating neuronal fate and connectivity in the fly visual system, which bears many fundamental similarities with the retina of vertebrates (Ngo, 2017).
Deciphering how neuronal diversity is established and maintained requires a detailed knowledge of neuronal gene expression throughout development. In contrast to mammalian brains, the large neuronal diversity of the Drosophila optic lobe and its connectome are almost completely characterized. However, a molecular characterization of this neuronal diversity, particularly during development, has been lacking. Thhis study presents insights into brain development through a nearly complete description of the transcriptomic diversity of the optic lobes of Drosophila. The transcriptome of 275,000 single cells was acquired at adult and at five pupal stages, and a machine-learning framework was built to assign them to almost 200 cell types at all time points during development. Two large neuronal populations were discovered that wrap neuropils during development but die just before adulthood, as well as neuronal subtypes that partition dorsal and ventral visual circuits by differential Wnt signalling throughout development. Moreover, it was shown that the transcriptomes of neurons that are of the same type but are produced days apart become synchronized shortly after their production. During synaptogenesis this study also resolved neuronal subtypes that, although differing greatly in morphology and connectivity, converge to indistinguishable transcriptomic profiles in adults. These datasets almost completely account for the known neuronal diversity of the Drosophila optic lobes, and serve as a paradigm to understand brain development across species (Ozel, 2020).
The optic lobes constitute two thirds of the fly brain, containing approximately 60,000 neurons per lobe and around 200 morphologically distinct neuronal types. The visual information detected by the photoreceptors is integrated in the four neuropils of the optic lobe-lamina, medulla, lobula and lobula plate-which are each divided into around 750 columns, corresponding to the approximately 750 ommatidia in the compound eye. This highly parallel processing depends on neurons that are either present at a 1:1 ratio with the number of columns (unicolumnar neurons), or present at a lower ratio but contact multiple columns to cover the entire visual map (multicolumnar neurons). This modular structure in which the same cell types are present multiple times renders the system particularly amenable to single-cell sequencing studies that rely heavily on clustering of numerous cells of the same type (Ozel, 2020).
Most optic lobe neurons are produced continuously through late larval and early pupal development from two crescent-shaped neuroepithelia known as the outer and inner proliferation centres. Immediately after birth, neurons project their processes to build the columns and the protolayers of all neuropils, which is completed by the time pupal development reaches 30% (P30). Synapse formation commences around P45 and continues until eclosion. These processes are genetically hardwired and seem to be strictly determined by the initial specification of neuronal identity, because both the morphology of specific cell types and the identity of their synaptic partners are largely invariant. Although a good understanding is had of the molecular mechanisms that specify neuronal identity, it remains unclear how morphological and functional diversity is established and maintained throughout development and how brain wiring is achieved. Characterizing both the common and cell-type-specific genetic programs that are implemented by neurons at different stages of their development is therefore essential (Ozel, 2020).
The single-cell transcriptomic atlases of the adult Drosophila optic lobe that have been previously generated by, and the bulk sequencing of isolated cell types, did not fully account for its cellular diversity. To produce an exhaustive catalogue of neurons in the adult optic lobe, 109,743 single-cell transcriptomes were obtained using the Chromium system (10x Genomics). The Seurat v.3 integration pipeline was used to remove batch effects between libraries. The single-cell transcriptomes were clustered using parameters that optimized the biological relevance of the resulting clusters. Then tested the statistical significance of the clusters was tested and improperly separated clusters that did not display biologically relevant differences in gene expression were merged, resulting in 193 final clusters (Ozel, 2020).
172 clusters were identified as optic lobe neurons and 19 clusters as glia-which were clearly separated from each other within the hierarchical tree of the clusters-and one cluster of low-quality transcriptomes. Neuronal clusters were annotated by calculating the Pearson correlations between the average gene expression of each cluster and 52 published bulk transcriptomes obtained from purified optic lobe neuronal types, and two additional neuronal types (Pm2 and T4) sequenced for this study. A clear match between a cell type and a cluster should result in an obvious correlation gap between the best and the second-best matches. The corresponding clusters were thereby identified for 53 of these 54 neurons, with only one exception (LPi3-4). In addition, the clusters corresponding to LC12, LC14, LC17, Pm1, T2a, TmY4, TmY8 and TmY14 neurons were identified using the binarized expressionof combinations of protein markers identified by antibody or reporter-line stainings. In summary, it was possible to identify 61 of the 172 neuronal clusters. The relative size of the identified clusters was consistent with the known abundance of these cell types. The very high resolution of this atlas is highlighted by the fact that distinct cluster matches were found even for rare neuronal types, such as Dm4 and Dm1, which are represented by only around 40 cells in each optic lobe. The dataset is therefore likely to contain independent clusters for almost all neuron types present at more than 30 cells per optic lobe. This criterion is satisfied by all unicolumnar and most multicolumnar neuron types; that is, by more than 95% of the cells in the dataset (Ozel, 2020).
The single-cell atlas was expanded to developing neurons. Optic lobe cells were sequenced at five pupal stages (around 30,000 cells at each stage; Methods), covering all stages of neuronal differentiation. Because developing neurons most often lack marker genes or available bulk transcriptomes, a supervised approach was adopted to annotate these datasets. A two-layer neural network was trained on a labelled adult dataset to classify P70 cells into corresponding adult clusters. This enabled sequencing of a lower number of cells at each pupal stage while retaining the high resolution of the adult dataset. The method was able to resolve less-abundant cell types that were grouped together by unsupervised clustering at P70. It was more robust in distinguishing closely related cell types than was Seurat v.3 label transfer. However, supervised classifications are inherently unfit for the identification of new cell types. To overcome this drawback and assess the accuracy of neural network classifications,they were compared to an unsupervised clustering of the P70 dataset and performed manual adjustments where appropriate (Ozel, 2020).
The earlier pupal datasets were classified sequentially backwards throughout development (from P70 to P50, P50 to P40, and so on). Because of the smaller sample sizes of the pupal datasets, a multi-task extension was used of the neural network to prevent overfitting the classifiers. At each stage, the neural network classifications were also manually assessed and adjusted before proceeding to the next step. Notably, several adult clusters corresponded to two or more unsupervised clusters at earlier time points. Using this iterative 'classify-adjust-retrain' approach, correspondences between pupal cells and each of the adult clusters were successfully assigned. The final trained models are provided in the Supplementary Data and can be used to annotate any single-cell RNA sequencing (scRNA-seq) dataset that contains optic lobe cells. As a benchmark, the classifier was tested on an atlas of the entire adult brain. The optic lobe cells were classified with high confidence and at a much higher resolution than in the original study, while the central brain neurons were clearly set aside as low-confidence predictions (Ozel, 2020).
Together, these data represent the first single-cell atlas of a complex central nervous structure at near complete saturation and throughout development. Coupled with the detailed knowledge of optic-lobe connectivity through electron microscopy reconstruction, and along with the Drosophila genetic toolkit, this provides a unique resource to investigate the general mechanisms that underlie the function and development of neuronal circuits. To facilitate the exploration of the datasets, summary tables are provided of average gene expression, binarized gene expression, and differentially expressed genes for all clusters at all stages, as well as the annotated objects with raw data, all of which can be found in the Gene Expression Omnibus (GEO) under accession number GSE142787. A list of transcription factor markers is provided that are maintained at all stages, the combinations of which uniquely identify each neuronal cluster (Ozel, 2020).
Two large neuronal clusters in the pupal datasets were identified that received very low confidence scores from the neural network classifier, which indicates that they might not exist in the adult dataset. These two clusters strongly expressed the pro-apoptotic gene sickle at P70, suggesting that the corresponding cells die during late pupal stages. Throughout development, they specifically expressed genes encoding the secreted protein Follistatin (Fs), the pro-secretory transcription factor Dimmed (dimm) and the vesicular transporter Portabella (prt). R10D10(dimm)-Gal4 and Fs-Gal4 were expressed in pupae in two large sets of cells that were also labelled by anti-Prt antibody. Their projections wrapped the dorsal and ventral edges of all optic lobe neuropils, except the lamina: each neuron possessed multiple branches that contacted-but did not enter-the neuropils at different points. These neurons could not be found in adults, and cleaved Dcp1 staining-a readout of Caspase 3 activity-confirmed that they were gradually cleared by apoptosis during late pupal stages. Activation of a FLEXAMP memory cassette at L3 stage using R10D10-Gal4 confirmed that the failure to observe these neurons in adult brains was not due to downregulation of reporter expression. Furthermore, expression of the anti-apoptotic protein p35 caused the perdurance of these neurons in adult brains. These cells were therefore named cells transient extrinsic (TE) neurons (Ozel, 2020).
Analysis of the genes that were differentially expressed between the two TE clusters revealed that only one of the clusters expressed Wnt4 whereas the other expressed Wnt10. Indeed, Wnt4-Gal4 was only expressed in the ventral cluster of TE neurons, which wetr named TEv; the Wnt10+ cluster was named TEd. Although TE neurons contacting the lamina were never observed, Fs-Gal4 was also expressed at P30 in photoreceptors from one row of ommatidia at the edge of the retina that connect to the edges of the lamina and are also apoptotically removed during development. This suggests that Follistatin, an inhibitor of Activin signalling that has been implicated in regulating neuron growth, is secreted at the edges of all neuropils by neurons that die before adulthood (Ozel, 2020).
A third TE cluster, which was denoted TEe (early), could be found only at P15 and P30. This cluster was connected to both TE clusters on uniform manifold approximation and projection (UMAP) visualization and was bsh+hth-. In L3 optic lobes, s few Bsh+Hth- cells labelled by R10D10-Gal4 were observed at the tips of the outer proliferation centre, suggesting that TE cells are produced there (the only other Bsh+ neurons are Hth+ Mi1). Bsh was also expressed in a subset of TE neurons at P30. A FLEXAMP memory cassette driven by bsh-Gal4 labelled all TE neurons, confirming that the TEe cluster contains younger bsh+ TE neurons that lose bsh expression as they mature. TE neurons expressing bsh were present in decreasing numbers until P50, but were not present at P70. It is therefore proposed that TE neurons are produced continuously from a relatively small number of progenitors at the tips of the outer proliferation centre (Ozel, 2020).
The transient nature of TE neurons, their superficial innervations and their enrichment in secretory markers are reminiscent of mammalian Cajal-Retzius cells that are essential for neuronal migration and other developmental processes. Further investigation will determine whether TE cells serve comparable functions in the development of the fly brain (Ozel, 2020).
At larval stages, the inner proliferation centre and the medial outer proliferation centre are progressively converted into neural stem cells (neuroblasts) that divide asymmetrically multiple times, each time self-renewing and producing a ganglion mother cell, which in turn divides once to generate two neurons or glia. Similar to photoreceptors that are produced sequentially from the posterior to the anterior of the eye imaginal disc, optic lobe neurons are also produced sequentially; the first-born neurons connect to the most posterior photoreceptors whereas later-born neurons connect to more anterior photoreceptors. This is also true in the lateral outer proliferation centre, in which lamina precursor cells are progressively converted into lamina monopolar neurons. As a result, neurons of the same type are produced days apart during development, and the optic lobes contain neurons at various stages of their differentiation path. Accordingly, P15 neuronal clusters displayed 'tails' of differentiating cells that expressed several genes known to be transiently expressed by newborn neurons-including Hey, nerfin-1 and zelda. Moreover, whereas all P30-P70 cells could be assigned directly to adult clusters-except for TE neurons and a few glia-like cells that were not investigated further-six clusters were found with low confidence scores from the neural network at P15 . These clusters were identified by known markers as neuroblasts, ganglion mother cells (three clusters), lamina precursor cells and undifferentiated apoptotic neurons. When visualized by UMAP, most P15 neuronal clusters converged towards the ganglion mother cell and neuroblast clusters, whereas lamina monopolar cells L1-L4 converged specifically towards the lamina precursor cell cluster. This suggests that the cluster tails are intermediate states between progenitors and differentiated neurons. Because these convergent tails containing very young neurons might often be mixtures of different cell types, they were separated from the mature neurons wherever possible and were assigned an 'immature' designation (Ozel, 2020).
Notably, these tails were not present at any of the later stages, when the clusters appeared homogenous despite containing neurons that were born more than 40 h apart. To further investigate this phenomenon, a UMAP plot was generated containing only T1 and Tm3 neurons, combined from all stages with no batch correction. Owing to continuous production of the neurons of the same type in the optic lobe, a Tm3 or T1 cell born at P0 in the P30 dataset would be the same age as a Tm3 or T1 cell born at P10 in the P40 dataset. Therefore, if neurons were to maintain age-dependent differences, cells of the same type would form a continuous trajectory across time points; however, they do not. Indeed, such a trajectory was observed only between P15 and P30, whereas all other time points remained robustly separated. To rule out batch effects between different stages, the homogeneity of the Tm3 cluster was directly assessed. Using Monocle, trajectories for Tm3 neurons were generated at both P15 and P30, and marker genes were calculated that varied with pseudotime. Analysis of these markers clearly indicated that cells were indeed ordered by age at P15. For instance, nerfin-1 was expressed only in the younger P15 cells and in almost no P30 cells, whereas expression of nicotinic Acetylcholine Receptor α7 peaked later, and the serotonin receptor gene 5-HT2A could be observed in only the most mature cells at P15. Conversely, all apparent sources of variation within the Tm3 cluster at P30 could be attributed to transcriptome quality, as measured by the percentage of mitochondrial reads observed. Therefore, the youngest (10-15 h old) and the oldest (more than 3 days old) Tm3 cells were no longer distinguishable at P30 (Ozel, 2020).
These data suggest that age information in neurons is lost within 15 h of birth, as they converge to a common transcriptomic state with older neurons of the same type. This generalizes observations that have previously been made in photoreceptors, and implies that all brain-wiring steps beyond initial neuropil targeting-that is, axon pathfinding-are executed synchronously in the optic lobe (Ozel, 2020).
Both T4 and T5 neurons have four subtypes (a, b, c and d) that each process motion in one of four cardinal directions. Their polarized dendritic arbors mirror their direction selectivity. Unsupervised clustering reliably distinguished these eight subtypes only at P50. However, supervised annotation and subclustering of T4-T5 cells showed that a and b subtypes could be separated from c and d subtypes at all stages. Many of the P50 subcluster markers turned off or lost their specificity in adult brains, which explains why T4-T5 subtypes were transcriptionally indistinguishable in adult brains. Gene Ontology analysis of these markers revealed exclusive enrichment for cell-surface-receptor terms involved in cell adhesion and axon or dendrite development (Ozel, 2020).
In addition, Dm3 and Tm9 cells were split into two subgroups only at P50 and earlier stages. Subgroups of Tm9 have not been previously described, but Dm3 cells have two known sub-populations with orthogonal dendritic orientations. Immunostainings against Bifid (Bi, also known as Omb)-which was differentially expressed between the Dm3 subclusters-showed that the dendrites of the Bi+ Dm3b cells were always oriented posterior-dorsally, whereas those of Bi- Dm3a cells were oriented posterior-ventrally. Because Dm3 subtypes differentially expressed several cell-surface molecules during synapse formation, it was asked whether they also differed in their connectivity. Analysis of the medulla connectome revealed that the two Dm3 subtypes connect differentially to several postsynaptic partners. Most notably, Dm3-Dm3 synapses were found almost exclusively between the different subtypes. Notably, Bi is also necessary and sufficient to specify the identity of T4-T5c/d subtypes that have dendrites with orthogonal directionality to those of T4-T5a/b neurons. Thus, bi may specify subtypes with orthogonal dendritic orientations in neurons that have completely different origins and properties (Ozel, 2020).
Attempting to project Dm3 or Tm9 subdivisions onto adult clusters by training binary classifiers on the P50 cells proved very unreliable, with out-of-bag errors greater than 20%. Consistently, the number of differentially expressed genes between subgroups peaked around P50 and decreased sharply thereafter. Even though T4 and T5, Dm3 and Tm9 may represent extreme cases, this increased transcriptional diversity during synapse formation is indeed a general phenomenon: Pearson correlations between the average gene expression profiles of clusters that are most similar to each other were significantly lower between P40 and P70, across all neuronal clusters. These results generalize the previous findings that olfactory projection neurons VA1d and DC3 are transcriptionally distinct during development but merge into a single cluster in adults. They also call into question the common practice of cell-type identification based solely on adult transcriptomes, and advocate for developmentally based approaches (Ozel, 2020).
To investigate why neuronal types are more easily distinguished during development, Gene Ontology analysis of the neuronal cluster markers was performed at each stage. At all stages, this consistently revealed overwhelming enrichment for receptor binding and activity terms related to axon or dendrite development and synapse formation, followed by transcription factor and ion-channel terms. Because different neurons are distinguished largely by cell-surface molecules at all ages, the origin of the increased diversity observed at mid-pupal stages was investigated. Gene Ontology analysis was performed on 'stage markers'-genes that were upregulated in neurons at a particular time point as compared to all other stages. Cell-surface-molecule terms involved in synaptogenesis and membrane potential were particularly upregulated around P50-P70, which explains the increased diversity at these stages. By contrast, early pupal markers were dominated by protein-synthesis terms and adult markers by energy-metabolism terms. This suggests that, as a general principle, the upregulation of cell-surface molecules necessary to enable synaptic specificity around P50 causes a peak of transcriptional diversity. This diversity is not maintained later, especially between subtypes that perform highly related functions and differ only on the basis of their connectivity. Moreover, P40 was enriched in molecular terms related to nuclear hormone receptor, driven by ecdysone-responsive transcription factors-suggesting that global activation of a hormonal switch triggers the upregulation of cell-type-specific cell-surface molecules at the onset of synaptogenesis (Ozel, 2020).
Differential gene expression analysis between the two Tm9 subgroups at P50 indicated that one exclusively expressed Wnt4 whereas the other expressed Wnt10, similar to the observations in TEv and TEd neurons. Because Wnt4-Gal4 expression overlapped only with ventral Tm9s, these subtypes were named Tm9v (Wnt4+) and Tm9d (Wnt10+). Sparse labelling of individual Tm9 neurons did not reveal any obvious morphological differences between these subtypes. However, differential expression of several cell-adhesion molecules at P50 raise the possibility that they differ in their connectivity. Even though these subtypes were statistically mostly indistinguishable in P70 and in adult datasets, Wnt4 and Wnt10 expression were found in separate parts of the same cluster in the t-distributed stochastic neighbour embedding (t-SNE) plot. It was also noticed that Wnt4-Gal4 expression was restricted to the ventral photoreceptors R7 and R8 in pupae, but not in adult flies. Therefore, both the receptive visual field and its downstream circuitry are partitioned by differential Wnt signalling during development (Ozel, 2020).
Flying insects are exposed to very different stimuli in their ventral (ground) compared with their dorsal (sky) visual fields, which may need to be processed differently. Dorsoventral asymmetries have previously been described in the retinas of flies, butterflies, dragonflies and honeybees and could therefore be a fundamental adaptation to flight. The current findings expand these differences to the downstream circuitry. In addition to the cell-autonomous differences during development that could enable these cells to connect with different synaptic partners, secreted Wnt ligands could differentially affect the development and function of other neurons in the ventral and dorsal parts of the brain (Ozel, 2020).
Finally, this study tested whether the differential expression of Wnt4 and Wnt10 observed in TE, Tm9 and R7 and R8 neurons applied to other seemingly homogeneous neuronal types. Two other neurons were found with dorsoventral asymmetries: Tm4 neurons and the unidentified cluster 62 could be separated into Wnt4+ and Wnt10+ populations, with only ventral Tm4 neurons overlapping with Wnt4-Gal4 expression. For both Tm4 and cluster 62, Wnt4+ and Wnt10+ cells remained separated on the t-SNE plot but were highly similar: artificially separating them yielded an average out-of-bag error across all stages of around 25%. Despite this similarity, Wnt4+ and Wnt10+ neurons differentially expressed several genes throughout development. Notably, both Tm4d and Tm9d specifically expressed 5-HT1A, encoding a serotonin receptor, in adult flies, raising the possibility that neuromodulatory signals are processed differently in ventral and dorsal visual circuits (Ozel, 2020).
This study has presented the first scRNA-seq dataset that reaches near complete saturation of any complex nervous system throughout its development. Coupled with the available optic lobe connectomes, this will provide a resource for functional studies of adult neurons as well as for the identification of new mechanisms involved in circuit formation. This analyses revealed two notable populations of pupae-specific neurons that share many characteristics with mammalian Cajal-Retzius cells and could be involved in neuropil development. They also lead to several observations that have implications on how neural circuits are built. The convergence of neuronal transcriptomes of the same type is described and previous observations were generalized of increased transcriptomic diversity in neurons during development to the entire optic lobe circuit. This increased diversity is due to a transient upregulation of cell-type specific cell-surface molecules that are involved in synapse formation, which explains how neurons with indistinguishable transcriptomes in adult brains could nevertheless serve different functions as a result of their developmental history. Finally, in an extension to the asymmetries described in the retinas of several flying invertebrates, this study demonstrated that ventral and dorsal visual circuits are subjected to differential Wnt signalling, providing potential mechanisms for the differential processing of ground and sky inputs (Ozel, 2020).
The brain consists of thousands of neuronal types that are generated by stem cells producing different neuronal types as they age. In Drosophila, this temporal patterning is driven by the successive expression of temporal transcription factors (tTFs). This study used single-cell mRNA sequencing to identify the complete series of tTFs that specify most Drosophila optic lobe neurons. It was verified that tTFs regulate the progression of the series by activating the next tTF(s) and repressing the previous one(s), and also identify more complex mechanisms of regulation. Moreover, the temporal window of origin and birth order of each neuronal type in the medulla was established Finally, this study describes the first steps of neuronal differentiation and shows that these steps are conserved in humans. That terminal differentiation genes, such as neurotransmitter-related genes, are present as transcripts, but not as proteins, in immature larval neurons (Konstantinides, 2022).
The brain is the most complex organ of the animal body. The human brain consists of over 80 billion neurons that belong to probably thousands of neuronal types. As neural stem cells age, temporal patterning allows them to generate different neuronal types in the correct order and stoichiometry. Temporal patterning in neuronal systems was first described in the Drosophila ventral nerve cord (VNC), in which a cascade of tTFs is expressed in embryonic neural stem cells (neuroblasts) as they divide and age. This concept was later expanded to the Drosophila optic lobe, with a different tTF series. It was later suggested that tTFs also contribute to the generation of neuronal diversity in different mammalian neuronal tissues, such as the retina and the cortex. However, series of tTFs are incomplete, as they were discovered by relying on existing antibodies. To generate a comprehensive description of the tTFs patterning a neural structure, a single-cell mRNA-sequencing (scRNA-seq) analysis was performed of the larval fly optic lobe (Konstantinides, 2022).
The Drosophila optic lobe is an ideal system to address how neuronal diversity is generated and how neurons proceed to differentiate. It is an experimentally manageable, albeit complex structure, for which there exists a very comprehensive catalogue of neuronal cell types. Meticulous research from the past decades has identified multiple cell types in the optic lobes based solely on morphological characters. Recent research made use of elaborate molecular genetic tools, as well as scRNA-seq, to expand the number of neuronal cell types to around 200, based on both morphology and molecular identity. Importantly, the neuroblasts that generate the medulla, which is the largest optic lobe neuropil containing around 100 neuronal types, are formed by a wave of neurogenesis over a period of days and progress through the same tTF temporal series. This means that, at any given developmental stage from mid third larval stage (L3) to early pupal stages (P15), the neurogenic region contains neuroblasts at all developmental stages (Konstantinides, 2022).
To study neuroblast and neuronal trajectories, a scRNA-seq analysis was performed of the optic lobes. 49,893 single-cell transcriptomes were obtained from 40 L3 optic lobes. The outer proliferation centre (OPC) neuroepithelium generates two optic lobe neuropils: the medulla from the medial side and the lamina from the lateral side. Medulla neuroepithelium, neuroblasts, intermediate precursors (known as ganglion mother cells (GMCs)) and neurons were arranged in a uniform manifold approximation and projection (UMAP) plot following a progression that resembled their differentiation in vivo. Similarly, lamina neuroepithelium, precursor cells and neurons were also arranged following a similar differentiation trajectory but in the opposite orientation of that of the medulla. The neuroblasts and the neurons that are generated from the inner proliferation centre followed a different trajectory in the UMAP plot (Konstantinides, 2022).
The larval single-cell dataset was merged with the annotated early P15 stage single-cell dataset. The P15 neurons mapped at the tip of each of the neuronal trajectories, which enabled identification of the corresponding neuronal types. Neurons were identified from all the neuropils of the optic lobe (lamina, medulla, lobula and lobula plate), as well as a small number of neuroblasts and neurons from the central brain that were probably retained when microdissecting the optic lobe (Konstantinides, 2022).
Next, expression was looked at of the known spatial TFs in the OPC neuroepithelium and tTFs in the neuroblasts: the spatial TFs Vsx1, Optix and Rx25 were expressed in largely non-overlapping subsets of neuroepithelial cells, and the tTFs Homothorax (Hth), Eyeless (Ey), Sloppy-paired (Slp), D and Tll were expressed in neuroblast subsets that were temporally organized in the UMAP plot (Konstantinides, 2022).
Thus, the UMAP plot recapitulated both proliferation and differentiation axes in the developing tissue: the UMAP horizontal axis represents differentiation status, whereas the vertical axis represents neuroblasts progressing through their tTF series (Konstantinides, 2022).
The larval scRNA-seq dataset provided the opportunity to look for all potential tTFs in an unbiased manner. The medulla neuroblast cluster was isolated from the scRNA-seq data and Monocle was used to reconstruct its developmental trajectory. Hth, Ey, Slp1/2, D and Tll were expressed in the previously described temporal order along the trajectory. The expression dynamics of all Drosophila TFs was examined and 14 candidate tTFs were identified, the expression of which was restricted to a specific pseudotime window, including the 6 previously known tTFs. Using antibodies or in situ hybridization for the eight newly discovered candidate tTFs and those already known in medulla neuroblasts, it was shown that their expression is indeed limited to restricted temporal windows, therefore defining new temporal windows as the neuroblasts progress through divisions (Konstantinides, 2022).
The previously known tTFs (except for Hth) contribute to the progression of the series by activating the next tTF in the cascade and repressing the previous one. To test which of the newly identified tTFs were involved in the progression of the temporal series, tTF mutant neuroblast MARCM (mosaic analysis with a repressible cell marker) clones or tTF RNA interference (RNAi) knockdowns were generated using the MZVUM-Gal4 line that is expressed in the Vsx1 domain of the OPC. Hth is expressed in the neuroepithelium and young neuroblasts, and is not required for Ey activation. Two factors were identified that regulate the expression of Ey in different ways: Erm is required to activate Ey and to inhibit Hth, whereas Opa is required for the correct timing of Ey activation. Opa also activates the expression of Oaz, which does not regulate the expression of any of the tTFs. Opa expression is repressed by Erm. Once Ey expression is initiated at the correct time by the combined action of Erm and Opa, Ey represses the expression of its activators. Thus, Erm is essential for the progression of the cascade, whereas Opa contributes to the correct timing of the expression of the next tTFs (Konstantinides, 2022).
Previous work has shown that Ey activates Slp, which in turn inhibits Ey. However, the developmental trajectory of neuroblasts uncovered a more complex situation. First, Ey activates Hbn. Hbn then represses Ey and activates Slp. Hbn also activates Scro and a second wave of Opa expression. Hbn then inhibits the expression of Erm and Scro inhibits the expression of Ey. Finally, Slp inhibits Hbn, Opa and Oaz (Konstantinides, 2022).
D expression requires both Slp and Scro. Previous work showed that in slp-mutant clones D is not expressed. Similarly, when scro was knocked down by RNAi, D was not activated. Scro is therefore important for the progression of the series, as it inhibits Ey and activates the expression of D. It remains expressed until the end of the neuroblast life. Once D is activated, it inhibits Slp and activates BarH1, which in turn activates Tll. Finally, similar to the inhibitory interaction between Tll and D previously described, Tll is sufficient but not necessary to inhibit BarH1 (Konstantinides, 2022).
This study has therefore identified most, if not all, tTFs in a developing neuronal system and show that these tTFs participate in the progression of the temporal series. Many of these interactions were confirmed by analysing the effect of tTF mis-expression on the temporal cascade (Konstantinides, 2022).
Besides their participation in the progression of the temporal series, tTFs regulate neuronal identity. Some tTFs are maintained in the neuronal subsets that are generated during their temporal window, whereas others are expressed only in newly born neurons. tTFs activate the expression of downstream neuronal transcription factors that regulate effector genes in the absence of the tTF. To test how tTFs regulate neuronal identity, whether knocking down the expression of the tTFs in neuroblasts affects the expression of neuronal transcription factors was tested. The loss of hth, ey and slp in neuroblasts leads to the loss of Bsh-, Vvl- and Toy-positive neurons, respectively. Hbn was shown to be required for the specification of Toy-, Traffic-jam (Tj)- and Orthodenticle (Otd)-positive neurons and Opa is required for the generation of TfAP-2-positive neurons. Thus, Hbn and Opa, as well as Hth, Ey and Slp, regulate neuronal diversity not only by allowing the temporal series to progress, but also by regulating the expression of neuronal transcription factors (Konstantinides, 2022).
The identified tTFs define at least 11 temporal windows in which different neurons (and glia) are generated. As they are generated, newly born neurons displace earlier born neurons away from the parent neuroblast, creating a columnar arrangement of neuronal cell bodies in the medulla cortex that represent birth order: early born neurons are located close to the emerging medulla neuropil, whereas late born neurons are closer to the surface of the brain. Neurons born in each temporal window express downstream effectors of tTFs (such as Bsh, Runt (Run) and Vvl) that were termed concentric genes due to their pattern of expression). The expression of tTFs in GMCs, and concentric genes that were previously described as well as those described in this work, in scRNA-seq neuronal clusters, together with cluster relative proximity in the UMAP plot, were used to assign the 105 neuronal clusters that comprise the medulla dataset to their predicted temporal window of origin. Proximal medulla neurons are generated in the Hth and Hth/Opa temporal windows, whereas distal medulla neurons are generated starting from the Ey temporal window. By contrast, transmedullary neurons are generated throughout most of the neuroblast life (Opa, Ey/Hbn and Slp temporal windows). Importantly, co-expression of some concentric genes is restricted to subregions of the medulla cortex, which enabled assigning the spatial origin to several medulla neuron clusters (Konstantinides, 2022).
To assess the status of all neuronal types, the expression of Apterous (Ap), which is expressed in the NotchON progeny of each GMC, was examined. Among the 105 neuronal types, 64 were NotchOFF and 41 were NotchON. As a given GMC division generates one NotchON and one NotchOFF neuron, Ap+ and Ap− neurons are intermingled in the medulla cortex. Thus, the position in the medulla cortex of concentric TFs expressed in NotchON and NotchOFF neurons enables inferrence of sister neurons, for example, Run neurons are probably sisters of TfAP-2 neurons, whereas early-born Vvl neurons are probably sisters of Knot (Kn) neurons (Konstantinides, 2022).
Finally, neurotransmitter identity was assigned to all of the medulla clusters at L3 and P15 stages. Ap expression is highly correlated with cholinergic identity, as nearly all Ap+—that is, NotchON—clusters in the dataset express ChAT and therefore have cholinergic identity, whereas most of the NotchOFF clusters are either GABAergic (most of them express Lim3)18 or glutamatergic (most of them express Tj or Fd59A). Interestingly, all the NotchOFF neurons from the same temporal window express the same neurotransmitter, independently of their spatial origin. This suggests that the temporal origin of medulla neurons and their Notch status instructs shared TF expression and neurotransmitter identity, and therefore function. In summary, this study has defined the temporal (and spatial) origin, birth order and Notch identity of all medulla cell types and highlighted the role of tTFs in regulating the generation of neural diversity (Konstantinides, 2022).
To study the first steps of neuronal differentiation after specification, the clusters from pupal stages (P15, P30, P40, P50 and P70) corresponding to the Mi1 cells were merged with the L3 scRNA-seq cluster and the GMCs most closely linked to them in the UMAP plot. Their differentiation trajectory was reconstructed, groups of genes (modules) were identified that co-vary along the entire trajectory from L3 to P70 and the Gene Ontology (GO) terms enriched in each gene module were sought. The timing of differentiation appears to follow a specific path. At L3, cell cycle genes and DNA replication genes are first expressed, as expected, from the division of GMCs. This is closely followed by genes involved in translation. Then, genes related to dendrite development and axon guidance are upregulated from late L3 until P30, stages during which the neurons direct their neurites to the appropriate neuropils. Genes that are important for neuronal function, such as neurotransmitter-related genes, synaptic transmission proteins, as well as ion channels start to be expressed as early as L3, reaching a plateau that is maintained until P15. Their expression then increases again until adulthood, when their products support neuronal function. This timing of differentiation was observed not only for Mi1 but could be generalized to all optic lobe neurons. These results indicate that not only is neuronal identity specified during the first hours of neuronal development, but their neuronal function (as indicated by the upregulation of chemical synaptic transmission terms) is also implemented very early, although the function is not required until much later. As this was unexpected, whether neurotransmitter mRNA expression observed as early as L3 was also translated was examined. Neurotransmitter-related genes, ChAT, VGlut and Gad1 mRNA are all expressed in the scRNA-seq data in non-overlapping neuronal sets and are maintained in the adult. However, protein expression at L3 was not observed. This suggests that their transcription represents a commitment to a specific neurotransmitter identity early in their development, but that other factors prevent premature translation of these mRNAs until they are needed at later stages of development (Konstantinides, 2022).
Next, whether the Drosophila optic lobe neuronal differentiation trajectory was similar to human neuronal differentiation was examined. We generated single-nucleus RNA-seq data from the human fetal cortical plate at gestational week 19. Monocle was used to reconstruct their developmental trajectory from apical progenitors to intermediate progenitors and postmitotic neurons and identified gene modules that were co-regulated along the trajectory. GO analysis uncovered a notable similarity to Drosophila. Then the expression of the GO terms that were expressed at different stages of the differentiation trajectory in Drosophila was plotred on the human cortical differentiation trajectory. Very similar dynamics were observed; the main difference was the absence of enrichment for ribosome assembly and translation-related GO terms at early stages. This could potentially be explained by the slower development of human neurons compared with those of Drosophila, leading to a slower increase in size and the fact that the divisions of the radial glia are more symmetric31 compared with those of optic lobe neuroblasts. Despite this difference, these results show that neurons follow a similar differentiation trajectory in Drosophila and humans (Konstantinides, 2022).
Although temporal patterning is a universal neuronal specification mechanism, it is unclear how it has evolved. This study investigated whether the medulla tTFs were conserved in mouse cortical radial glia using a published scRNA-seq dataset. None of the medulla neuroblast tTFs were expressed in strict temporal windows in ageing radial glia, with the exception of PAX6 (orthologue of Ey), which was enriched in older progenitors. Reciprocally, the Drosophila orthologues of the mouse temporally expressed TFs were not expressed temporally in the developing optic lobe (Konstantinides, 2022).
The mouse orthologues of the Drosophila VNC tTFs Ikzf1, Pou2f1/Pou2f2 and Casz1 are expressed temporally in mouse retinal progenitors. The expression was looked at of the optic lobe tTFs in the mouse retina in a published scRNA-seq dataset. PAX6 was constitutively expressed, MEIS2 (orthologue of Hth), ZIC5 (orthologue of Opa) and SOX12 (orthologue of D) were expressed at embryonic stage 12, while NR2E1, the orthologue of Tll (which is expressed when neuroblasts become gliogenic), was expressed late, when retinal progenitors become gliogenic and start generating Muller glia. The lack of a strict conservation of tTFs between flies and mice indicates that the acquisition of the specific temporal series occurred independently in each phylum (Konstantinides, 2022).
The comprehensive series of transcription factors described in this work and their regulatory interactions temporally pattern a developing neural structure. Most tTFs are expressed in overlapping windows, creating combinatorial codes that differentiate neural stem cells of different ages and therefore provide them with the ability to generate diverse neurons after every division. They were conservatively assigned into 11 distinct temporal windows (ten of which generate neurons) that—when integrated with spatial patterning (six spatial domains) and the Notch binary cell fate decision—can explain the generation of approximately 120 cell types, which is close to the entire neuronal type diversity of the Drosophila medulla. Moreover, this study determined the downstream TFs that were expressed in neurons produced temporally, which enabled establishment of the birth order of all medulla neurons. Moreover, a detailed transcriptomic description is provided of the first steps in the differentiation trajectory of a neuron. Terminal differentiation genes are expressed within the first 20 h of neuronal life, approximately 2-4 days before their protein products can fulfil their function. Why these genes are expressed so early remains unclear, but it is hypothesized that this reflects the commitment of neurons to a specific function. We also show that all neurons follow the same route for differentiation and that this is similar to the differentiation process in developing human cortical neurons. Thus, understanding the mechanisms of neuronal differentiation in flies can generate insight for the equivalent process in humans (Konstantinides, 2022).
Dynamically regulated cell adhesion plays an important role during animal morphogenesis. The formation of the visual system in Drosophila embryos has been used as a model system to investigate the function of the Drosophila classic cadherin, DE-cadherin, which is encoded by the shotgun (shg) gene. The visual system is derived from the optic placode, which normally invaginates from the surface ectoderm of the embryo and gives rise to two separate structures, the larval eye (Bolwig's organ) and the optic lobe. The optic placode dissociates and undergoes apoptotic cell death in the absence of Shotgun, whereas overexpression of Shotgun results in the failure of optic placode cells to invaginate and of Bolwig's organ precursors to separate from the placode. These findings indicate that dynamically regulated levels of Shotgun are essential for normal optic placode development. Overexpression of Shotgun can disrupt Wingless signaling through titration of Armadillo out of the cytoplasm to the membrane. However, the observed defects are likely the consequence of altered Shotgun mediated adhesion rather than a result of compromising Wingless signaling, since overexpression of a Shotgun-alpha-catenin fusion protein, which lacks Armadillo binding sites, causes defects similar to Shotgun overexpression. The genetic interaction between Shotgun and the Drosophila EGF receptor homolog, Egfr, was studied. If Egfr function is eliminated, optic placode defects resemble those following Shotgun overexpression, which suggests that loss of Egfr results in an increased adhesion of optic placode cells. An interaction between Egfr and Shotgun is further supported by the finding that expression of a constitutively active Egfr enhances the phenotype of a weak shg mutation, whereas a mutation in rhomboid (rho) (an activator of the Egfr ligand Spitz) partially suppresses the shg mutant phenotype. Finally, Egfr can be co-immunoprecipitated with anti-Shotgun and anti-Armadillo antibodies from embryonic protein extracts. It is proposed that Egfr signaling plays a role in morphogenesis by modulating cell adhesion (Dumstrei, 2002).
The head ectoderm of early Drosophila embryos is subdivided into several domains that realize different
morphogenetic programs. The embryonic eye field is the posterior-medial region of the procephalic neurectoderm that gives rise to the visual system, which includes the larval eye (Bolwig's organ) and adult
eye, as well as the optic lobe. Around gastrulation, cells of the eye field undergo a convergent extension directed laterally. Shortly afterwards these cells form two morphologically visible placodes, one on either side of the embryo. These optic placodes sink inside and become the optic lobe primordia, epithelial double layers attached to the posterior surface of the brain. The optic placode of a stage 12-13 embryo is V-shaped, with the anterior leg of the V representing the anterior lip, which later forms the inner anlage of the optic lobe, and the posterior leg forming the posterior lip, later forming the outer anlage. As the invagination deepens and the two lips 'sink' inside the embryo, ectodermal cells that earlier surrounded the perimeter of the optic placode approach each other and eventually form a
closed epidermal cover. Abundant cell death accompanies the closing of the head epidermis over the optic lobe anlage, and the subsequent separation of
this anlage from the epidermis. A small number of cells that originally formed part of the posterior lip of the optic placode
remain integrated in the head epidermis and form the larval eye or Bolwig's organ. As these cells move away from the optic lobe anlage their basal ends
become drawn out and form axons that constitute Bolwig's nerve (Dumstrei, 2002).
Shotgun is expressed throughout the ectoderm including the eye field and its epithelial derivatives. One would expect that normal optic
lobe development requires modulation of Shotgun activity to allow, for example, the segregation of the invaginating optic placodes from the
surrounding ectoderm. Since cell culture studies have indicated that the mammalian EGF receptor can disrupt cadherin-based adhesion, it was of interest to see whether Drosophila Egfr is expressed in the visual system to allow for such a possibility in Drosophila as well. Egfr is
expressed in a complex and dynamic pattern that closely parallels the pattern of double-phosphorylated ERK (dpERK) expression, indicating activation
of the MAP kinase signaling pathway. During stage 7 both rho (an activator of Egfr signaling) and dpERK are expressed in two stripes in the head ectoderm. The expression of dpERK in these two stripes is the result of Egfr activity. The anterior stripe corresponds to part of the head midline, while the posterior stripe reaches into the eye field. Distribution of dpERK in the two stripes becomes patchy during stage 10. At the
same time, the posterior stripe widens dorsally to overlap with part of the optic lobe placode. Finally, at the late extended germ band stage and
during germ band retraction, dpERK becomes restricted to the optic lobe placodes and cells of the dorsal head midline. This expression pattern demonstrates that Egfr activation accompanies the determination, morphogenesis and differentiation of the embryonic visual system (Dumstrei, 2002).
On the subcellular level, Egfr is expressed diffusely on the membrane of epithelial cells and neuroblasts. Egfr overlaps with Armadillo,
the Drosophila ß-catenin homolog, which is an integral component of the cadherin-catenin complex. Like Shotgun, Armadillo is
concentrated strongly in the apically located zonula adherens but is also found at lower levels in the entire lateral membranes (Dumstrei, 2002).
A second type of junction, called a septate junction, develops in Drosophila epithelial cells at a slightly later stage than the zonula adherens. Septate junctions have been implicated in maintaining epithelial stability. The Coracle protein forms part of
the septate junctional complex, and an antibody against Coracle serves as a sensitive marker for this junction. Applying this
marker to embryos of different stages it was found that all ectodermally derived epithelia express Coracle, except for the optic lobe and the invaginations
that form the stomatogastric nervous system. Accordingly, no septate junctions have been reported in previous electron microscopic
investigations of these tissues. The reliance on adherens junctions alone may make the optic lobe (and stomatogastric nervous system) susceptible to changes in the stability of these junctions; such changes occur resulting from manipulations of Shotgun and Egfr (Dumstrei, 2002).
A finely adjusted level of Shotgun is required for optic placode morphogenesis, and ß-catenin, as well as EGFR signaling, is involved in this process. Reduction in Shotgun results in dissociation of the placode around the time when it normally invaginates, suggesting that the forces exerted on the epithelial sheet while folding may disrupt cell contacts. A similar phenotype was described for other epithelial invaginations, including the Malpighian tubules and stomatogastric nervous system. Abolishing Armadillo/ß-catenin function results in a similar, if somewhat weaker phenotype. If Shotgun is overexpressed, invagination is also impaired. Cells stay together in a placode-like formation (as would be expected from 'hyperadhesive' epithelial cells), but do not noticeably constrict apically. It should be noted that the interpretation of this failure of optic placode cells to constrict is complicated by the accompanying increase in cell death in surrounding head epidermal cells. This phenomenon, in addition to a direct effect of an increased amount of Shotgun in the optic placode cells, could be part of the pathology responsible for the non-invagination phenotype. By contrast, the non-disjunction of optic lobe and larval eye is likely to be a rather direct consequence of an increased amount of Shotgun expression. Interestingly, other adhesion systems, notably the Drosophila N-CAM homolog FasII, are also involved in optic lobe-larval eye separation. Thus, the down regulation of FasII by the 'anti-adhesion' molecule Beaten path is also required for normal larval eye morphogenesis (Dumstrei, 2002).
Overexpression of Shotgun or the DE-cad-alpha-catenin fusion construct causes a dramatic change in optic lobe morphogenesis, without causing much disruption in other epithelia. It is speculated that this enhanced sensitivity of optic lobe cells towards an increased level of Shotgun may be in part due to the fact that adherens junctions form the only means of contact between optic lobe cells. In other epithelia, such as epidermis, trachea and hindgut, septate junctions form by far the more prominent junctional complex. Septate junctions have been implicated in epithelial stability. One could surmise that embryonic epithelia, as they enter the phase of differentiation during mid-embryogenesis, construct septate junctions that add to the adherens junctions developed at an earlier stage. This additional junctional complex makes late epithelia more resistant to changes in cadherins, a notion supported by the finding that blocking cadherins (by applying calcium chelators, or tyrosine kinase inhibitors) in early embryos up to stage 10 leads to a break down of epithelia, whereas it has only a small effect in later stages. The optic lobe, which does not differentiate but gives rise to a population of neuroblasts later dring the larval period, does not form septate junctions, which could account for its strong reliance on normally functioning adherens junctions (Dumstrei, 2002).
Expression of a fusion construct, DE-cad-alpha-catenin, in which the cytoplasmic domain of Shotgun is replaced by a truncated alpha-catenin, thereby preventing a reduction in the cytoplasmic pool of Arm, results in a similar phenotype as overexpressing full length Shotgun. This finding lends support to the notion that dissociation of the cadherin-catenin complex (CCC) may not occur at the interface between Shotgun and Arm or Arm and alpha-catenin. If one were to assume that dissociation occurred between any components of the CCC, one would expect a stronger phenotype, given that by overexpressing the fusion construct one not only increases the amount of Shotgun molecules interconnecting cells, but also the stability with which they are coupled to the cytoskeleton. Biochemical studies in vertebrates and Drosophila also show that phosphorylation of the CCC does not result in increased dissociation of Arm or alpha-catenin from the CCC, suggesting that the dissociation occurs distal to alpha-catenin (Dumstrei, 2002).
The strength of the CCC and other structural molecules driving morphogenesis has to be controlled in a complex spatiotemporal pattern. Numerous widely conserved signaling pathways have been implicated in this process. In vertebrate embryos, mutations of the Wnt, Shh and BMP signaling pathways result in impressive examples which tissues and organs show defects in morphogenesis. Furthermore, it became clear that frequently signaling proteins affect fundamental cellular behaviors, such as proliferation, motility, adhesiveness and survival. This prompted the hypothesis that in many developmental scenarios, the 'proximal' effect of receiving a signal could be a change in morphogenetic behavior. The discovery that one of the Wnt signal transducers, ß-catenin, leads a 'double life' as a structural component of the cadherin-catenin complex, fueled the idea that Wnt signal could directly exert an effect on the adhesiveness on the cell, an idea that is supported by cell culture experiments. However, genetic studies have demonstrated that in Drosophila, the roles of ß-catenin as a signaling transducer and a CCC component seem to be quite separate. Although it is clear that the cytoplasmic and membrane bound ß-catenin pools are in a steady state, binding of more ß-catenin to the membrane, by overexpression of Shotgun, reduces the cytoplasmic pool resulting in a wg minus phenotype. However, Wnt/Wg signaling seems to have no effect on the amount of membrane bound ß-catenin. Thus, in Drosophila, it appears that Shotgun mediated adhesion, at least under experimental conditions, interferes with Wnt/Wg signaling by competing for ß-catenin but Wnt/Wg signaling may not have a direct effect on adhesion mediated by the CCC (Dumstrei, 2002).
The findings suggest that another signaling pathway, the Egfr pathway, is involved in modulating cadherin-mediated adhesion and thereby controls morphogenesis. Egfr, similar to its function in the developing compound eye, is activated in the precursors of the larval eye and adjacent optic lobe at a stage preceding optic lobe invagination and larval eye separation. The ligand for Egfr is Spitz, which is activated by Rhomboid (Dumstrei, 2002).
In a small subset of larval eye precursors (the 'Bolwig's organ founders') loss of Egfr signaling results primarily in cell death, lending further support to the view that Egfr signaling functions generally in the ectoderm and its derivatives to maintain cell viability. Recent studies in Drosophila indicate that MAPK directly controls the expression and protein stability of the cell death regulator, Hid (W; Wrinkled). If cell death is prohibited by a deficiency of the reaper-complex, cells of the optic placode and most other embryonic cells that undergo apoptosis in Egfr loss-of-function mutants survive. Both optic lobe and Bolwig's organ express several of their normal differentiation markers, but show a characteristic 'hyperadhesive phenotype', consisting in the failure of optic Iobe invagination and Bolwig's organ separation. Based on the similarity of this phenotype to the one resulting from Shotgun overexpression, and the genetic interaction between Egfr and Shotgun mutants in the ventral ectoderm, it is proposed that Egfr activation is required in normal development to phosphorylate the CCC and thereby allows optic lobe invagination and Bolwig's organ separation to occur. This would be in line with experimental results obtained in vertebrate cell culture studies, which have demonstrated that drug- or Egfr-induced phosphorylation of the CCC leads to dissociation between CCC and cytoskeleton. Recent findings have shown that another phosphorylation event, mediated by the rho/rac GTPases, also affects adhesion by dissociating alpha-catenin from the CCC (Dumstrei, 2002).
Co-IP data indicates that Egfr is linked to the CCC in Drosophila as well. This implies that the effect of Egfr on Shotgun mediated adhesion could be a direct one that occurs at the cell membrane and does not involve MAPK signal transduction to the nucleus. It has been shown in a number of vertebrate cell culture systems that tyrosine phosphorylation of ß-catenin results in a disassembly of the CCC complex and a consecutive loss in cadherin-mediated adhesion. Phenotypically, this results in increased invasiveness of tumor cell lines, neuronal and growth cone motility. Several tyrosine kinases and phosphatases have been identified that can increase or decrease the degree of phosphorylation of the CCC. For example, v-src transfected into cultured cells phosphorylates ß-catenin and causes cells to dissociate, round up, and become more motile. Egfr also phosphorylates the CCC and forms an integral part of this complex. This opens up the possibility that growth factors, with their widespread expression and biological activity in the developing embryo, may exert part of their effect on cell behavior by modulating, in a rather direct way, cell adhesion at the membrane. Such a mechanism would account for the 'rapid mode' of control of adhesion molecules. Systems such as the optic placode of the Drosophila embryo, where matters of different cell fates are decided at the same time when morphogenetic movements change the arrangement and shape of the cells involved, constitute favorable paradigms to address how signaling systems control both processes (Dumstrei, 2002).
Developmental regulators and cell cycle regulators have to interface in order to ensure appropriate cell proliferation during organogenesis. An analysis of
the roles of the pan-neural genes deadpan and asense
defines critical roles for these genes in regulation of
mitotic activities in the larval optic lobes. Loss of deadpan results in reduced cell proliferation, while ectopic
deadpan expression causes over-proliferation. In contrast,
loss of asense results in increased proliferation,
while ectopic asense expression causes reduced proliferation.
Consistent with these observations endogenous
Deadpan is expressed in mitotic areas of the optic lobes,
and endogenous Asense is expressed in cells that will
become quiescent. Altered Deadpan or Asense expression
results in altered expression of the cyclin dependent
kinase inhibitor gene dacapo. Thus, regulation of
mitotic activity during optic lobe development may, at
least in part, involve deadpan and asense mediated regulation
of the cyclin dependent kinase inhibitor gene
dacapo (Wallace, 2000).
Optic lobes begin development during embryogenesis
between stages 11 and 12 when a group of 30-40
epidermal cells delaminates and moves from the surface
of each brain hemisphere. Once delaminated, these cells
remain inactive until the embryo hatches as a first instar
larva. This inactive state of the cells is partially mediated
by the glycoprotein Anachronism, secreted by glia surrounding
the developing optic lobe (Ebens, 1993).
In the first instar larva the cells begin to divide, a process
requiring the function of the trol gene.
These first divisions appear to be synchronous and continue through the beginning
of pupal development. A total of approximately 3,000
cells are produced in the mitotically active areas of the
optic lobe. During second instar some of the cells of the developing lamina and medulla
begin to differentiate into neurons and glia. This differentiation
is accompanied by the innervation of the
first and second optic lobes by photoreceptor axons. Their arrival and the release of
Hedgehog protein in the developing optic lobes begins
the differentiation of the lamina cells into neurons and
glia. The outer proliferation center (OPC) represents one of the major areas of mitotic
activity in the optic lobe. The OPC becomes a distinct structure at
late second instar and the cells in the OPC and the inner
proliferating center (IPC) continue to divide until all of
the photoreceptor axons have innervated the optic lobe
and initiated differentiation of the lamina precursors. The lamina furrow spreads outward in a semicircle and passes through the OPC where the cells for the developing
lamina originate. As the lamina furrow advances outward, the cells in the passing furrow arrest in G1 phase (Wallace, 2000 and references therein).
The IPC, which is the second major area of mitotic
activity in the optic lobe, forms in a crescent shape at a
more interior position of the brain with respect to the
OPC. The IPC represents a pool of cells that produces
the cells for the medulla and the lobula. The IPC cells,
however, do not divide and differentiate in as synchronous
an order as the OPC (Wallace, 2000).
To determine the functional properties of Dpn during
larval development the pUAST/Gal4 system was used to test the effects of ectopic
dpn expression. The 71B Gal4 driver line was used in this analysis, since it drives the
expression of pUAST constructs in most cells of the
second and third instar larval optic lobes. In addition,
this line also drives strong expression in the wing discs.
Bromodeoxyuridine (BrdU) incorporation and histone
H1 RNA expression were used as S phase specific markers
to detect changes in mitotic activity (Wallace, 2000).
In the larval central nervous system, ectopic Dpn
expression results in a striking increase in the size of the
brain lobes as compared to wild-type brains. In
brains with ectopic Dpn expression, an increase in the
number of mitotically active cells is apparent across the
entire surface of the enlarged brain. In addition, a breakdown
of the mitotic domain pattern that is present in
wild-type third instar optic lobes is also evident. The over-proliferation phenotype that
is associated with ectopic Dpn expression is fully penetrant.
It can range from >10 times to two- to four-fold the
size of a normal wild-type brain lobe, and appears sensitive
to the accumulation modifiers in the genome (Wallace, 2000).
HES proteins can have opposite functions from proteins
of the AS-C in neural development. ase, a member of the AS-C, has
been reported to be expressed in the developing third
instar optic lobes and loss of ase function results in
disturbances of the adult optic lobe. It was asked
whether the AS-C protein Ase can modulate mitotic
activity. To this end, the effects of ectopic
ase expression on mitotic activity in the developing
larval optic lobe were investigated.
As with Dpn, ectopic expression of Ase results in
strong expression in most cells of the second and third
larval optic lobes. This expression results in breaks in the
normally continuous pattern of S phase positive cells in
the OPC suggesting that increase and/or ectopic expression
of the Ase protein decreases mitotic activity in the optic lobe (Wallace, 2000).
It was asked whether mitotic activity is altered in third
instar larval optic lobes of dpn1 homozygous loss of
function mutants. dpn1 is an apparent null allele of the
dpn gene. In the OPC of dpn1 homozygous
third instar larvae, sporadic breaks are evident in
the normally continuous area of S phase positive cells. These breaks can vary in size and location in the OPC, but can be found in the OPC of nearly all
homozygous dpn1 mutant larvae. In addition, S phase activity in the developing lamina appears compressed and disorganized (Wallace, 2000).
To better determine whether reduction in the
amount of OPC neuroblasts in dpn1 mutants results in
a significant loss of OPC neuroblast progenies, assays were performed for developing lamina cells that represent
direct progeny of the OPC cells. If there is a reduction
in the amount of cells in the OPC, a subsequent
reduction in the amount of developing lamina cells
could be expected. Anti-Dachshund antibodies were used to mark the cells of the
developing lamina. In dpn1 homozygous mutant third
instar larvae, a reduced number of Dachshund (Dac)
positive cells is evident as compared to the wild type. Photoreceptor axons that innervate the lamina are responsible for initiating the differentiation
of the cells into neurons and glia. It was necessary to determine whether the
reduced number of cells is due to an aberrant projection
of photoreceptor axons. Anti-HRP antibodies
were used to mark the photoreceptor axons in dpn1
homozygous larvae. Overall size and morphology of
the eye disc, as well as photoreceptor axon extension
and innervation of the lamina in dpn1
larvae appears normal. One difference, however,
is that the area of innervation is smaller than wild
type. The reduced number of developing lamina cells in dpn1 loss of function larvae indeed may, therefore, be due to a reduced amount of OPC neuroblasts rather than aberrant axon projection (Wallace, 2000).
To analyze the possible involvement of ase gene function
on mitotic activity in the larval optic lobes, the S phase activity in third instar larval
brains was determined. In ase1/scb57 mutants, there is an expansion of S
phase activity to include the normally mitotic quiescent
cells between the OPC and the lamina precursor cells
(LPCs), as well as scattered S phase activity in the lamina. ase1 is a deletion of the ase coding region and scB57 is a deletion of the entire AS-C as well as the proximal complementation
group EC4 making the larva
homozygous mutant for ase and heterozygous for the
other members of the AS-C. In contrast, +/scB57 larval
optic lobes show normal S phase activity. Thus, ase loss of function mutants show an
increase in the S phase activity between the OPCs and
the LPCs, and a random pattern of increased S phase
activity in the lamina (Wallace, 2000).
If Dpn is involved in the positive regulation of mitotic
activity, as indicated by the dpn loss of function and
ectopic expression phenotypes, then Dpn would be
expected to be expressed in mitotic active areas. Endogenous dpn protein expression was examined in the wild-type larval CNS; Dpn was found to be expressed in
areas of active cell division in the optic lobes.
Dpn is expressed in the OPC of the late third instar larva
and stops at the edge of the OPC. After cells exit the
OPC, S phase activity ceases and the cells subsequently
arrest in G1 as they pass through the lamina furrow. Dpn is also expressed in the cells
of the IPC. Thus, expression of Dpn in the larval optic lobes is in agreement with a possible role as one positive regulator of the cell cycle (Wallace, 2000).
If Ase is involved in the termination of mitotic activity in
the larval optic lobes, then Ase expression would be
expected in or near areas where the cell cycle is arresting.
Ase protein expression was examined in the larval
optic lobes; Ase was found to be present in a band at the
posterior edge of the OPC that partially overlaps
with Dpn expression. Ase is expressed just before
the cells exit the OPC and cease S phase activity. These
cells then arrest in G1 phase as they pass through the
lamina furrow. Ase is also expressed
in cells of the IPC and at a low level in the lamina
furrow. The expression pattern of Ase, which
comes to a maximum at the posterior edge of the OPC,
is in agreement with a possible role for Ase in aiding cell
cycle arrest as cells leave the OPC (Wallace, 2000).
Cdk inhibitors have been shown to represent key regulators
of mitotic activity. In Drosophila a cdk inhibitor gene, dap, has been
identified that is transiently expressed during embryogenesis
in cells prior to entering their last mitosis and at
the onset of terminal differentiation. Ectopic expression of dap results in
G1 arrest, while loss of dap function has been shown to
cause one extra cell division in embryonic epidermal
cells.
Dpn appears to promote the continuation of mitotic
activity, while Ase has a role in ending cell proliferation
in the developing optic lobes. Therefore, it was asked
whether altered expression of Dpn and Ase can modulate
the expression of the dap. In wild-type third instar
larva, optic lobe expression of dap occurs in specific
domains. dap is expressed in cells of the lamina
furrow and scattered cells of the lamina. There is also
strong expression of dap in a subset of cells in the IPC
throughout third instar. In contrast, dap
expression is virtually absent from the cells of the OPC (Wallace, 2000).
The effects were determined of the loss of dpn function on
the expression of dap. In homozygous dpn1 mutant
third instar larva, expression expands into the area of the
OPC. Also, cells of the lamina begin to express dap more
strongly. In contrast, in larvae with ectopic Dpn
expression, dap expression is strongly reduced or absent
in the optic lobes of third instar larva.
Thus, dpn activity has a negative regulatory effect on the
dap RNA level (Wallace, 2000).
In homozygous ase mutant third instar larvae, there is
a strong reduction of dap RNA throughout the entire
developing optic lobe while dap expression in the developing
eye disk appears normal. The ase loss
of function phenotype demonstrates that ase activity is
necessary for the expression of dap throughout the
developing optic lobe. When Ase is ectopically expressed
in third instar optic lobes, ectopic activation of
dap expression becomes evident. Therefore,
ase activity has a positive regulatory effect on the dap
RNA level (Wallace, 2000).
It was asked whether the phenotypical effects on cell
proliferation produced by alterations of Dpn and Ase
expression may be caused, at least in part, by changes in
the levels of dap transcript. During embryogenesis, alteration in the levels of dap expression through either
ectopic expression or by loss of function, result in dramatic
changes in mitotic activity. Therefore, the mitotic
activity in optic lobes of homozygous dap6 mutant third
instar larvae were analyzed. While predominately recessive lethal, a few dap6 homozygous escapees can be viable to adulthood.
Therefore, the larval optic lobes of homozygous dap6
mutant third instar larva can be analyzed. In such
homozygous dap6 mutant larvae over-proliferation of
the cells of the optic lobes is evident. There is a
significant increase in the number of mitotically active
cells and break down of mitotic domains, as compared
to the wild type (Wallace, 2000).
The over-proliferation phenotype of dap6 null mutants
can be compared to the over-proliferation phenotype in
larvae with ectopic dpn expression,
and the associated suppression of dap expression. Although the over-proliferation in both cases is
similar, there are clearly more cells produced in the dpn
over-expressing brain lobes. This strongly indicates that
other cell cycle regulators are also likely to be affected
by the ectopic expression of dpn in the optic lobes (Wallace, 2000).
A model is proposed for mitotic control in the developing
third instar optic lobe in which cell proliferation is
modulated by a positive regulator of mitotic activity such
as Dpn and a negative regulator of mitotic activity such
as Ase. In this model, one role of Dpn and Ase would be
to interface with cell cycle regulation through the direct
or indirect modulation of dap expression. Mitotic control
during optic lobe development may involve the
following events. Cells that give rise to the optic lobe delaminate
from the neuroectoderm during embryogenesis
and remain quiescent until first instar with the help of
proteins such as Anachronism. The
mitotic activity is then initiated through a process that
requires the trol gene product and the developmental
regulator and transcription factor Eve to begin the proliferation of neuroblasts to
form the OPC. The mitotically active state of OPC cells
would be maintained in part by Dpn. In the absence of
Dpn, the cells in the OPC have a greater chance of
exiting mitosis by allowing Dap to be expressed. As cells
arrive at the edge of the OPC, Ase is expressed at high
levels, allowing the neuroblasts to become quiescent
only after they pass out of the region where Dpn is
expressed. Suppression of dap by Dpn in the OPC would
allow the neuroblasts to be mitotically active while the
increased expression of Ase at the posterior edge of the
OPC allows the neuroblasts to exit mitosis and begin
differentiation. In addition, the resulting quiescent state
needs to be maintained in the lamina; otherwise the cells
may reenter mitosis (Wallace, 2000 and references therein).
Notch signaling mediates multiple developmental decisions in Drosophila. This study examined the role of Notch signaling in Drosophila larval optic lobe development. Loss of function in Notch or its ligand Delta leads to loss of the lamina and a smaller medulla. The neuroepithelial cells in the optic lobe in Notch or Delta mutant brains do not expand but instead differentiate prematurely into medulla neuroblasts, which lead to premature neurogenesis in the medulla. Clonal analyses of loss-of-function alleles for the pathway components, including N, Dl, Su(H), and E(spl)-C, indicate that the Delta/Notch/Su(H) pathway is required for both maintaining the neuroepithelial stem cells and inhibiting medulla neuroblast formation while E(spl)-C is only required for some aspects of the inhibition of medulla neuroblast formation. Conversely, Notch pathway overactivation promotes neuroepithelial cell expansion while suppressing medulla neuroblast formation and neurogenesis; numb loss of function mimics Notch overactivation, suggesting that Numb may inhibit Notch signaling activity in the optic lobe neuroepithelial cells. Thus, these results show that Notch signaling plays a dual role in optic lobe development, by maintaining the neuroepithelial stem cells and promoting their expansion while inhibiting their differentiation into medulla neuroblasts. These roles of Notch signaling are strikingly similar to those of the JAK/STAT pathway in optic lobe development, raising the possibility that these pathways may collaborate to control neuroepithelial stem cell maintenance and expansion, and their differentiation into the progenitor cells (Wang, 2011).
This study find that Notch signaling plays an essential role in the maintenance and expansion of neuroepithelial cells in the optic lobe; it also inhibits medulla neuroblast formation. Clonal analyses of several pathway components indicate that this dual function bifurcates downstream of Su(H) with E(spl)-C only partly involved in the inhibition of medulla neuroblast formation but not the maintenance and expansion of neuroepithelial stem cells (Wang, 2011).
In the optic lobe, Notch signaling plays a role analogous to lateral inhibition during embryonic CNS development. However, the selection of neuroblasts in the OPC neuroepithelium is an all-or-none process rather than selecting individual neuroblasts from the neuroepithelium. Medulla neuroblasts are generated in a wave progressing in a medial to lateral direction in the OPC neuroepithelium with all cells at a particular position along the medial-lateral axis differentiating into neuroblasts. Interestingly, this wave of medulla neuroblast formation coincides with the down-regulation of both Delta and Notch expression in the medial cells in the OPC, which might reduce Notch signaling activity, thereby allowing medulla neuroblasts to form. What factors drive the recession of both Delta and Notch expression in the OPC neuroepithelium along the medial-lateral axis is not known. When Notch signaling is inactivated, neuroepithelial cells in the OPC change cell morphology and differentiate into medulla neuroblasts prematurely. The results indicate that Notch signaling actively controls neuroepithelial integrity, possibly by regulating the adherens junction (AJ), since in Notch pathway mutant mosaic clones in the OPC, the apical determinants PatJ, Crumbs and aPKC are cell autonomously reduced or lost and the mutant cells change to rounded or irregular morphology. Further experiments will be needed to determine how Notch signaling activity affects the maintenance of neuroepithelial integrity, particularly the stability of the adherens junction (Wang, 2011).
Is neuroblast formation also actively inhibited by Notch signaling or simply a default state of neurogenic epithelial cells? In the latter model, Notch signaling may only maintain neuroepithelial integrity and promote their expansion while medulla neuroblasts form when the neuroepithelial integrity is disrupted. The argument against this model is that changes in neuroepithelial integrity are not always accompanied with cell fate changes. In N, Dl or Su(H) mosaic clones located in the OPC neuroepithelium, it was found that in about 25% of the clones, the mutant cells changed morphology or lost apical marker expression but did not become neuroblasts (Dpn-negative), whereas in E(spl)-C mosaic clones, Dpn+ cells were prematurely induced, which indicate that the cells begin to differentiate into neuroblasts, but these cells still retained columnar epithelial cell morphology and apical marker expression. This suggests that the suppression of neuroblast formation by Notch signaling activity is separable from the maintenance of neuroepithelial integrity and that medulla neuroblast formation is actively suppressed by Notch signaling. A possible scenario is that activation of the Notch pathway turns on the E(spl)-C genes, which in turn suppress proneural gene expression in the optic lobe neuroepithelia. Indeed, at least one member in the E(spl)-C genes, E(spl)m8, appears to be activated in the neuroepithelial cells by the Notch pathway, as the E(spl)m8-lacZ reporter is expressed in a pattern similar to Delta and Notch expression in the OPC and IPC. E(spl)m8 protein and possibly additional members of the E(spl)-C may suppress the expression of proneural genes in the optic lobe. The proneural genes of the achaete-scute complex (as-c) comprise four members, achaete, scute, L'sc, and asense. achaete is not expressed in the optic lobe, but scute is expressed in both the neuroepithelial cells and neuroblasts in the OPC implying that scute expression in the neuroepithelial cells is not suppressed by Notch signaling activity. By contrast, asense is only expressed in the neuroblast and GMCs and L'sc is transiently detected in an advancing stripe of neuroepithelial cells of 1-2 cells wide that are just ahead of newly formed medulla neuroblasts. Thus, E(spl)-C proteins may suppress L'sc and/or ase expression, the release of this suppression may allow the neuroepithelial cells to begin to differentiate into medulla neuroblasts. It should be noted, however, that the removal of the E(spl)-C activity does not seem to be sufficient to allow full differentiation of neuroepithelial cells into medulla neuroblasts, suggesting that additional factors downstream of Notch signaling may be involved in the suppression of medulla neuroblast formation (Wang, 2011).
The phenotypes of Notch pathway mutants are reminiscent of those of JAK/STAT mutants. For example, inactivation of either pathway led to early depletion of the OPC neuroepithelium; either pathway inhibits neuroblast formation, and ectopic activation of either pathway promotes the growth of the OPC neuroepithelium. The remarkable phenotypic similarities in Notch and JAK signaling mutant brains suggest that these pathways may act in a linear relationship such that activation of one pathway is relayed to the second, perhaps by inducing the expression of a ligand. Alternatively, these pathways may act in parallel and converge onto some key downstream effectors or target genes. Further experiments will be needed to test whether Notch interacts with JAK/STAT and if it does, to find out where the interaction occurs during the development of the optic lobe (Wang, 2011).
The roles of Notch signaling in mammalian brain development have been studied intensely. Many Notch pathway components have been examined in knockout mice, which showed defects in brain development. Mice deficient for Notch1 or Cbf all display precocious neurogenesis during early stages of nervous system development. This has led to the view that the role of Notch signaling in the mouse brain is to maintain the progenitor state and inhibit neurogenesis. However, it is not clear from these studies whether the premature neurogenesis in Notch signaling mutant mice was caused by premature differentiation of neuroepithelial stem cells into neurons or by premature differentiation of neuroepithelial stem cells into progenitor cells, which then generated neurons. In fact, it has been proposed that Notch activation can promote the differentiation of neuroepithelial stem cells into radial glial cells, the progenitor cells that generate the majority of neurons in the cerebral cortex. This is based on the observation that ectopic Notch activation using activated forms of Notch1 and Notch3 (NICD) caused an increase in radial glial cells as compared to control. The radial glial cells resemble medulla neuroblasts in the Drosophila optic lobe in that they are both derived from neuroepithelial stem cells and undergo asymmetric division to self-renew and generate neurons, although morphologically radial glial cells are still polarized while medulla neuroblasts have lost epithelial characters and are rounded in shape. Based on the current results, it is suggested that Notch signaling maintains the pool of neuroepithelial stem cells and promotes their expansion in both Drosophila and mammals and that the precocious neurogenesis in Notch signaling mutant brains arise due to premature differentiation of the neuroepithelial stem cells into the progenitor cells (Wang, 2011).
However, ectopic Notch activation may indeed promote progenitor cell proliferation in the brain. Ectopic neuroblasts were observed in the medulla cortex when NACT was ectopically expressed by the neuroblast/GMC driver insc-Gal4, by ubiquitous expression using hs-Gal4, or when numb15 mosaic clones were induced at later larval stages when neuroblasts normally begin to form. Since the results have shown that the Notch pathway is not essential for medulla neuroblast formation or self-renewal, the ectopic neuroblasts are a novel phenotype solely induced by ectopic Notch signaling activity. This is consistent with Notch activation promoting ectopic neuroblast formation in the central brain and VNC without being required for neuroblast self-renewal in these regions of the CNS; and Notch has been shown to be an oncogene in mammals. Since the sizes of the ectopic neuroblasts were in the range of GMC or neurons, they may resemble the transit-amplifying (TA) neuroblasts that are found in the dorsal-medial region of the central brain. The origin of these ectopic neuroblasts in the medulla cortex is not clear, but it is unlikely that they are derived from differentiated medulla neurons as ectopic expression of NACT using elav-Gal4, which is active in medulla neurons, did not result in ectopic neuroblasts and by the fact that ectopic neuroblasts can be induced in numb15 mosaic clones, which could only arise from mitotically active cells that include neuroepithelial cells, medulla neuroblasts, and ganglion mother cells (GMCs), but not neurons. The ectopic neuroblasts could be generated by a transformation of GMCs into a neuroblast identity as suggested for ectopic neuroblasts in brat mutant central brains. Ectopic Notch signaling activity may even directly promote the expansion of neuroblasts after they have differentiated from the neuroepithelial cells in the OPC. In either case, ectopic Notch signaling activity may block the normal path of neuronal differentiation and lock the cells in a proliferative state. This is indeed what was observed in numb15 mosaic clones in which numerous ectopic neuroblasts were induced in the medulla cortex without generating medulla neurons. Perhaps ectopic Notch signaling activity may also promote the proliferation of neural progenitors in vertebrates, such as the radial glial cells in the mouse brain (Wang, 2011).
During neurogenesis in the medulla of the Drosophila optic lobe, neuroepithelial cells are programmed to differentiate into neuroblasts at the medial edge of the developing optic lobe. The wave of differentiation progresses synchronously in a row of cells from medial to the lateral regions of the optic lobe, sweeping across the entire neuroepithelial sheet; it is preceded by the transient expression of the proneural gene lethal of scute [l(1)sc] and is thus called the proneural wave. This study found that the epidermal growth factor receptor (EGFR) signaling pathway promotes proneural wave progression. EGFR signaling is activated in neuroepithelial cells and induces l(1)sc expression. EGFR activation is regulated by transient expression of Rhomboid (Rho), which is required for the maturation of the EGF ligand Spitz. Rho expression is also regulated by the EGFR signal. The transient and spatially restricted expression of Rho generates sequential activation of EGFR signaling and assures the directional progression of the differentiation wave. This study also provides new insights into the role of Notch signaling. Expression of the Notch ligand Delta is induced by EGFR, and Notch signaling prolongs the proneural state. Notch signaling activity is downregulated by its own feedback mechanism that permits cells at proneural states to subsequently develop into neuroblasts. Thus, coordinated sequential action of the EGFR and Notch signaling pathways causes the proneural wave to progress and induce neuroblast formation in a precisely ordered manner (Yasugi, 2010).
Loss of EGFR function in progenitor cells caused failure of L(1)sc expression and differentiation into neuroblasts (see A model of progression of the proneural wave). In addition, elevated EGFR signaling resulted in faster proneural wave progression and induced earlier neuroblast differentiation. The
activation of the EGFR signal is regulated by a transient expression of Rho, which cleaves membrane-associated Spi to generate secreted active Spi. This study also demonstrated that Rho expression itself depends on EGFR function, and thus the sequential induction
of the EGFR signal progresses the proneural wave. Clones of cells mutant for pnt were not recovered unless Minute was employed, suggesting that the EGFR pathway is required for the proliferation of neuroepithelial cells. However, the progression of the proneural wave is not regulated by the proliferation rate per se (Yasugi, 2010).
The function of the Notch signaling pathway in neurogenesis is known as the lateral inhibition. A revision of this notion has recently been proposed for mouse neurogenesis, in which levels of the Notch signal oscillate in neural progenitor cells during early stages
of embryogenesis, and thus no cell maintains a constant level of the signal. The oscillation depends mainly on a short lifetime and negative-feedback regulation of the Notch effecter protein Hes1,
a homolog of Drosophila E(spl). This prevents precocious neuronal fate determination. The biggest difficulty in analysis of Notch signaling is the
random distribution of different stages of cells in the developing ventricular zone, which is thus called a salt-and-pepper pattern. In medulla neurogenesis, however, cell differentiation is well organized spatiotemporally and the developmental process
of medulla neurons can be viewed as a medial-lateral array of progressively aged cells across the optic lobe. Such features allowed the functions of Notch to be precisely analyzed. Cells are classified into four types according to their developmental stages: neuroepithelial cells expressing PatJ, neuronal progenitor I expressing a low level of Dpn, neuronal
progenitor II expressing L(1)sc and neuroblasts expressing high levels of Dpn. The Notch signal is activated in neuronal progenitor I and II. The EGFR signal turns on in the neuronal progenitor II stage and progresses the stage by activating L(1)sc expression. Cells become neuroblasts when the Notch and EGFR signals are shut off. Cells stay as neuronal progenitor I when Notch signal alone is activated, whereas cells stay as neuronal progenitor II
when the Notch signal is activated in conjunction with the EGFR signal. Although the Notch signal is once activated, it must be turned off to let cells differentiate into neuroblasts. In neuronal
progenitor II, E(spl)-C expression is induced by Notch signaling, and the increased E(spl)-C next downregulates Dl expression and subsequent activation
of the Notch signal (Yasugi, 2010).
What does Notch do in medulla neurogenesis? It is infered that the Notch signal sustains cell fates, whereas the EGFR signal progresses the transitions of cell fate. This was well documented when a constitutively active form of each signal component was induced. EGFR, or its downstream Ras, induces expression of L(1)sc but does not fix its state, even though the constitutively active form is employed. At the same time, a constitutively active Notch sustains cell fates in a cell-autonomous manner. Constitutively active N receptors, by contrast, autonomously determine cell fates depending on the context: cells become neuronal progenitor I in the absence of EGFR and neuronal progenitor II in the presence of EGFR. The precocious neurogenesis caused by the impairment of Notch signaling suggests that Notch keeps cells in the progenitor state for a certain length of time in order to allow neuroepithelial cells to grow into a sufficient population. In the prospective spinal cord of chick
embryo (Hammerle, 2007), the development from neural stem cells to neurons progresses rostrocaudally, during which the transition from proliferating
progenitors to neurogenic progenitors is regulated by Notch signaling (Yasugi, 2010).
Although Notch plays a pivotal role in determining cell fate between neural and non-neural cells, the function may be context
dependent and can be classified into three categories. (1) Classical lateral inhibition is seen in CNS formation in embryogenesis
and SOP formation in Drosophila. Cells that once expressed a higher level of the Notch ligand maintain their cell states and become neuroblasts. (2) Oscillatory
activations are found in early development of the mouse brain (Shimojo, 2008). Progenitor cells are not destined to either cell types. (3) An association with the proneural wave found in Drosophila medulla neurogenesis as is described in this study. The Notch signal is transiently activated only once and then shuts off in a synchronized manner. The notable difference in the outcome is the ratio of neural to non-neural cells; a small number of cells from the entire population become neuroblasts or neural stem cells in the former cases (1 and 2), whereas most of the cells become
neuroblasts in the latter case (3). The differences between (1) and (2) can be ascribed at least in part to the duration of development. Hes1 expression has been shown
to oscillate within a period of 2 hours in the mouse, whereas in Drosophila embryogenesis, selection of neuroblasts from neuroectodermal cells takes place within a few hours. Thus, even if Drosophila E(spl) has a half-life time equivalent to Hes1, the selection process during embryogenesis probably finishes within a cycle of the oscillation. The process of medulla neuroblast formation continues for more than 1 day, but Notch signaling is activated
for a much shorter period in any given cell. This raises the possibility that E(spl)/Hes1 may have a similarly short half-life but outcome would depend on the developmental context (Yasugi, 2010).
The functions of EGFR and Notch described in this study resemble their roles in SOP formation of adult chordotonal organ development; the EGFR signal provides an inductive cue, whereas the Notch signal prevents premature SOP formation. In addition, restricted expression of rho and activation of the EGFR signal assure reiterative SOP commitment. Several neuroblasts are also sequentially differentiated from epidermal cells in adult chordotonal organs (Yasugi, 2010).
Unpaired, a ligand of the JAK/STAT pathway is expressed in lateral neuroepithelial cells and shapes an activity gradient that is higher in lateral and lower in the medial neuroepithelium. The JAK/STAT signal acts as a negative regulator of the progression of the proneural wave (Yasugi, 2008). This report has shown that activation of both EGFR and Notch signaling pathways depends on the activity of the JAK/STAT signal. The JAK/STAT signal probably acts upstream of EGFR and Notch signals in a non-autonomous fashion. These three signals
coordinate and precisely regulate the formation of neuroblasts (Yasugi, 2010).
Neuroepithelial cell proliferation must be carefully balanced with the transition to neuroblast (neural stem cell) to control neurogenesis. This study shows that loss of the Drosophila microRNA mir-8 (the homolog of vertebrate miR-200 family) results in both excess proliferation and ectopic neuroblast transition. Unexpectedly, mir-8 is expressed in a subpopulation of optic-lobe-associated cortex glia that extend processes that ensheath the neuroepithelium, suggesting that glia cells communicate with the neuroepithelium. Evidence is provided that miR-8-positive glia express Spitz, a transforming growth factor α (TGF-α)-like ligand that triggers epidermal growth factor receptor (EGFR) activation to promote neuroepithelial proliferation and neuroblast formation. Further, these experiments suggest that miR-8 ensures both a correct glial architecture and the spatiotemporal control of Spitz protein synthesis via direct binding to Spitz 3' UTR. Together, these results establish glial-derived cues as key regulatory elements in the control of neuroepithelial cell proliferation and the neuroblast transition (Morante, 2013).
This analysis reveals that the production by an optic-lobe-associated cortex glia of the EGFR ligand Spitz is critical for coordination of neuroepithelial proliferation and the spatiotemporal emergence of neuroblasts. External signaling from the surrounding microenvironment is a common mechanism for the regulation of stem cell number and behavior in mature tissues, but the need of a niche microenvironment during early neurogenesis was unknown (Morante, 2013).
Flies deficient for the microRNA mir-8 exhibit brain degeneration and behavioral defects. Strikingly, the microRNA is expressed in cortex glial cells lying underneath the blood-brain-barrier (subperineurial) glial layer. These glia are of large size and produce long protrusions that ensheath the developing optic lobe neuroepithelium and can be distinguished by their
selective expression of the EGFR ligand spitz as well as its modulators aos and rho (Morante, 2013).
Genetic manipulation of mir-8, spi, or aos, in this glial cell population unveiled cell nonautonomous influences of these glia on the development of the underlying neuroepithelium. Similar glial signaling to neuroblasts in the larval Drosophila brain has been demonstrated for a class of neuroblasts that remain quiescent until nutrient-responsive satellite and cortical glia reactivate their proliferation. This study identifies another population of glia cells that sustain growth of neuroepithelial cells and the neuroepithelial-neuroblast transition via a mir-8-Spitz axis (Morante, 2013).
Glial-mediated regulation of the neuroepithelium is reminiscent to the roles of mammalian astrocytes that are known to potently stimulate neurogenesis in cell culture and a component of endogenous neural stem cell niche in adult mammalian neurogenesis. Additionally, EGFR is also implicated in glial cell proliferation in Drosophila and human glioma often exhibits elevated EGFR signaling (Morante, 2013).
The notion that mir-8-Spitz-positive cortex glia constitute an anatomically and functionally distinct population of surface-associated glia cells is strongly supported by the finding that RNAi knockdown of spi in subperineurial glia using moody- Gal4 has no effect on neuroepithelial development. Spitz protein is converted to its active form by the Rhomboid protease, which is also expressed by miR-8 glia. The extracellular factor Aos limits Spitz spreading and signaling level that may influence the effects of Spitz-EGFR in the responding neuroepithelium (e.g., sustaining proliferation and preventing premature or ectopic neuroblast formation). The posttranscriptional silencing of spi mRNA by the miR-8 binding to a sequence in its 3' UTR provides another layer of regulation to fine-tune the timing, localization and/or amount of Spitz protein translation. Moreover, the distinctive architecture of miR-8-Spitz-positive cortex glia appears to be regulated by endoreplication regulators dup/ Cdt1 and the microRNA miR-8. Importantly, expression of a spitz transgene lacking its 3' UTR (and hence unable of regulating by miR-8) fully rescued the undergrowth defect caused by mir-8 overexpression in the glia cells. Therefore, it is suggested that cortex glia employ a coordinated strategy that is mediated by miR-8 to ensure that: (1) the glia establish a correct architecture to provide a continuous layer of cortex glia cells that extend long processes to the neuroepithelium; and (2) correct local (or temporal) control of Spitz protein synthesis. Given the expression of rho and aos is directly induced by EGFR signaling in other context, a feed-back signaling via EGFR may also occur in miR-8-Spitz positive glial cells, thereby contributing to the fine-tuning of Spitz protein activation and secretion (Morante, 2013).
A niche typically refers to a confined anatomical location where adult stem cells reside and provides the signals required to sustain stem cell function and number. Niches are usually composed of supporting cells that make physical contact with the stem cells and act locally. The optic-lobe-associated miR-8-Spitz-positive cortex glia appear to represent a niche that contributes signals for the growth and morphogenesis of the neuroepithelium and that constitutes a functionally distinct population of that of the bloodbrain- barrier glial cells (Morante, 2013).
Extrinsic signaling in the coordination of neuroepithelial proliferation in the developing mammalian forebrain of Foxc1 mutant mice has also been suggested. In these mice, the meninges are reduced or absent, resulting in an expansion of the neuroepithelium due to the predominance of symmetric divisions. The meninges are a source of the retinoic acid required for the transition of neuroepithelial cells into radial glia and neurons (Siegenthaler, 2009). Furthermore, meningeal cells secrete and organize the pial basement membrane, a thin sheet of extracellular matrix that covers the brain and that is enriched in a variety of growth factors. Rupture of the basement membrane in the developing brain causes type II lissencephaly, generating ectopic precursor clusters and cortical heterotopias due to impaired attachment of the radial glia to the basement membrane, resulting in a general laminar disorganization. These defects are reminiscent to those of disrupted surface glia cells described in this study (Morante, 2013).
In summary these findings suggest that neuroepithelial proliferation and the onset of neuroblasts in the developing optic lobe neuroepithelium are largely influenced by extrinsic cues via a miR-8-dependent mechanism in the overlying glia. The reprogramming of neuroepithelial cells into neural stem cells (neuroblasts) is associated with dramatic morphological and molecular changes, including the loss of epithelial determinants DE-Cadherin, Crumbs and PatJ, and enhanced expression of the Snail-family zinc-finger transcriptional repressor Worniu.
These changes are strikingly reminiscent of the events that drive the epithelial-to-mesenchymal transition (EMT), which confers a stem-like character in mammalian epithelial cells and in cancer cells and which are regulated by the miR-200 family. Indeed, downregulation of human mir-200 genes in epithelial normal and cancer cells promotes EMT and the acquisition of 'stemness' effects that are presumed to be cell-autonomous. These findings demonstrate that miR-8 can exert its effects non-cell- autonomously, opening the possibility that microRNA of the miR-200 family may play similar roles in stem cell fate niches and/or microenvironmental regulation of metastasis (Morante, 2013).
The 40,000 neurons of the medulla, the largest visual processing center of the Drosophila brain, derive from a sheet of neuroepithelial cells. During larval development, a wave of differentiation sweeps across the neuroepithelium, converting neuroepithelial cells into neuroblasts that sequentially express transcription factors specifying different neuronal cell fates. The switch from neuroepithelial cells to neuroblasts is controlled by a complex gene regulatory network and is marked by the expression of the proneural gene l'sc. This study discovered that microRNA miR-7 is expressed at the transition between neuroepithelial cells and neuroblasts. miR-7 promotes neuroepithelial cell-to-neuroblast transition by targeting downstream Notch effectors to limit Notch signaling. miR-7 acts as a buffer to ensure that a precise and stereotypical pattern of transition is maintained, even under conditions of environmental stress, echoing the role that miR-7 plays in the eye imaginal disc. This common mechanism reflects the importance of robust visual system development (Caygill, 2017).
Drosophila vision requires the accurate specification of over 80 different types of optic lobe neurons and the establishment of precise visual circuits between the neurons of the optic lobe and the photoreceptors of the eye. The medulla is the largest visual ganglion of the brain. Medulla neurons play roles in motion detection, through input from the R1-R6 photoreceptors via the lamina, and in the perception of color, via direct input from the R7 and R8 photoreceptors. The 40,000 medulla neurons originate from a pseudostratified neuroepithelium. During early development, symmetric division expands the stem cell pool. As development progresses, the medial edge of the neuroepithelium is progressively converted into asymmetrically dividing neuroblasts. Medulla neuroblasts sequentially express a series of transcription factors that specify the differentiation of the medulla neurons (Caygill, 2017).
The transition from neuroepithelial cells into neuroblasts occurs in a highly ordered, sequential manner in response to expression of the proneural gene, lethal of scute (l'sc). Expression of L'sc marks a two- to three-cell-wide boundary between the neuroepithelial cells and the neuroblasts, the so-called transition zone, which moves medially across the neuroepithelium, forming a proneural wave. Within the transition zone, L'sc transiently suppresses Notch activity, triggering the switch from the symmetric, proliferative division of neuroepithelial cells to the asymmetric, differentiative division of neuroblasts. Progress of the wave is regulated by the orchestrated action of the Notch, epidermal growth factor receptor (EGFR), Fat-Hippo, and JAK/STAT signaling pathways (Caygill, 2017).
This study presents evidence here that the transition from neuroepithelial cells to neuroblasts in the developing optic lobe is buffered by the microRNA miR-7. miR-7 is expressed at the transition zone in response to epidermal growth factor (EGF) signaling and is sufficient to promote transition. miR-7 acts via repression of downstream Notch effectors to limit Notch signaling and promote timely transition. In the absence of miR-7, proneural wave progression is disrupted. This disruption becomes more severe under conditions of temperature stress, suggesting that the role of miR-7 is to act as a buffer to ensure the timely and precise transition from neuroepithelial cells to neuroblasts in the developing optic lobe (Caygill, 2017).
This study has shown that miR-7 targets members of the E(spl) family of bHLH transcription factors to define the boundaries of the transition zone, buffering the transition from neuroepithelial cell to neuroblast in the developing optic lobe. Downregulation of Notch signaling is essential for the neuroepithelial-to-neuroblast transition. L'sc provides one mechanism for Notch downregulation. miR-7, expressed in a pattern similar to that of l'sc at the transition zone, provides another, emphasizing the importance of Notch regulation at the transition zone (Caygill, 2017).
The transition zone of the proneural wave mediates the specification of the neuroblasts of the medulla, the largest ganglion of the adult Drosophila visual system. The medulla receives information directly from the R7 and R8 photoreceptors of the ommatidia. Similar to the specification of neuroblasts by the proneural wave in the optic lobe, photoreceptor differentiation is triggered by the movement of the morphogenetic furrow across the epithelium of the eye imaginal disc. As the furrow passes, expression of the proneural gene atonal (ato) is induced in a stripe that is later refined to the R8 cells by Notch-mediated lateral inhibition (Caygill, 2017).
Similar to the observations in the optic lobe, miR-7 has been shown to play a role in photoreceptor differentiation. Misexpression of miR-7 results in an increase in Ato expression and R8 cell specification, while a loss of miR-7 results in a decrease in Ato expression under conditions of temperature stress. These results show that miR-7 acts to buffer the development of both the medulla and the eye, two tissues that will directly communicate in the adult brain (Caygill, 2017).
miR-7 has been shown to target anterior open (aop; also known as yan) in the eye imaginal disc. Within the neuroepithelium, aop activity helps to repress the neuroepithelial-to-neuroblast transition. This raises the possibility that miR-7 targeting of aop could also contribute to its function in buffering the neuroepithelial-to-neuroblast transition and that aop could represent a common target during the progression of the proneural wave and the morphogenetic furrow (Caygill, 2017).
In the adult brain, each ommatidium maps to a columnar unit within the lamina and medulla, providing a retinotopic map of the visual field. Signaling from innervating photoreceptors induces the differentiation of lamina neurons. This direct communication provides a strict control of the mapping of photoreceptor and lamina neuron numbers. In contrast, while final numbers of ommatidia and medulla neurons show some co-ordination based on nutrient availability, there is no evidence for direct communication between the eye disc and the developing medulla. The presence of miR-7 in both the eye imaginal disc and the optic lobe represents an independent but conserved buffer that operates to coordinate appropriate developmental progression in each system, in spite of external environmental fluctuations. The presence of this common buffer provides robustness within each system that may contribute to ensuring the eventual connectivity required for retinotopic mapping of the visual system (Caygill, 2017).
The proper balance between symmetric and asymmetric stem cell division is crucial both to maintain a population of stem cells and to prevent tumorous overgrowth. Neural stem cells in the Drosophila optic lobe originate within a polarised neuroepithelium, where they divide symmetrically. Neuroepithelial cells are transformed into asymmetrically dividing neuroblasts in a precisely regulated fashion. This cell fate transition is highly reminiscent of the switch from neuroepithelial cells to radial glial cells in the developing mammalian cerebral cortex. To identify the molecules that mediate the transition, neuroepithelial cells were microdissected, and their transcriptional profile was compared with similarly obtained optic lobe neuroblasts. Genes encoding members of the Notch pathway were found expressed in neuroepithelial cells. Notch mutant clones are extruded from the neuroepithelium and undergo premature neurogenesis. A wave of proneural gene expression is thought to regulate the timing of the transition from neuroepithelium to neuroblast. The proneural wave transiently suppresses Notch activity in neuroepithelial cells, and inhibition of Notch triggers the switch from symmetric, proliferative division, to asymmetric, differentiative division (Egger, 2010).
In the developing mammalian cortex, neural stem cells initially divide symmetrically to produce two neural stem cells, thereby increasing the neural precursor pool. The radial glial cells subsequently divide asymmetrically to produce a neural stem cell and either a basal progenitor cell or an immature neuron. Most basal progenitor cells divide once more to generate two postmitotic neuron. The Notch signalling pathway is thought to play a role in maintaining the undifferentiated state of neuroepithelial cells, radial glia and basal progenitors, but the downstream signalling cascades activated in these cells might be differentially regulated. Neurogenesis is initiated by proneural genes, such as Mash1 and Neurogenin2 (Ngn2) (Egger, 2010).
This study shows that this sequence of neurogenic events is remarkably similar to that seen in the development of the optic lobe in Drosophila. Notch is activated in the neuroepithelial cells, which remain undifferentiated. The proneural gene l'sc is expressed within the transition zone, and levels of Delta are increased, while Notch activity is decreased. Thus neuroepithelial cells ultimately give rise to a variety of differentiated neurons, but only after they have passed through the transition zone (Egger, 2010).
Low levels of Delta expression were found thoughout the optic lobe neuroepithelium, with increased expression in the transition zone. Several Brd genes were found within the neuroepithelium. Negative regulation of Delta activity by the Brd proteins would be expected to further reduce the level of Delta signalling. This situation might be analogous to the oscillations in Delta and Ngn2 levels observed in vertebrates, and it will be interesting to assess whether the expression of Delta, HLHm5 or proneural genes also oscillate in flies. Strikingly, when Delta activity is inhibited throughout the epithelium, the premature transformation of the entire neuroepithelium into neuroblasts is observed. This suggests that neuroepithelial cells might both send and receive the Notch signal (Egger, 2010).
Higher levels of Delta were observed in the L'sc positive transition zone. High levels of Delta or Serrate can inhibit Notch signalling through cis-inhibition, suggesting one possible mechanism for the downregulation of Notch signalling at the transition zone. Interestingly, very recent results suggest that cis-inhibition can create sharp boundaries and this could be the role of the high levels of Delta that were observe in the transition zone (Egger, 2010).
Epithelial integrity might be important to maintain proliferative cell division. This study shows that Notch mutant clones are extruded from the neuroepithelium. Furthermore, expression in the optic lobe neuroepithelial cells was found of a number of genes involved in cell adhesion. Notch could regulate cell adhesion molecules at the transcriptional level, or might itself form a complex with adhesion molecules. In either case, Notch loss of function would disrupt cell adhesion and lead to the extrusion of epithelial cells. Subtypes of cadherins, such as DE-Cad, Cad99C, Fat, which were found preferentially expressed in the neuroepithelium, might be activated by Notch to maintain the neuroepithelium, and repressed by L'sc to promote neurogenesis. Notch mutant clones also upregulate expression of the neuroblast transcription factor Dpn (but not of L'sc), and divide asymmetrically only once they have delaminated from the epithelium. In contrast to Notch mutant clones, L'sc misexpression clones upregulate Dpn and switch to asymmetric division whilst still embedded within the neuroepithelium. L'sc acts, at least in part, through repression of Notch signalling, but might also induce neuroblast-specific genes directly (Egger, 2010).
JAK/STAT signalling negatively regulates the progression of the proneural wave and neurogenesis in the optic lobe. Interestingly, the ability of Notch to maintain radial glial cell fate appears to be largely dependent on functional JAK/STAT signalling. It remains to be seen whether the Notch pathway interacts with JAK/STAT in the Drosophila optic lobe (Egger, 2010).
This study has shown that the development of the Drosophila optic lobe parallels that of the vertebrate cerebral cortex, suggesting that the pathways regulating the transition from symmetric to asymmetric division might be conserved from flies to mammals. Identifying the effector genes that are regulated by Notch and L'sc, and the links between JAK/STAT and Notch signalling, will yield further insights into the molecular mechanisms that maintain an expanding neural stem cell pool and regulate the timely transition to differentiation (Egger, 2010).
A dynamic balance between stem cell maintenance and differentiation paces generation of post-mitotic progeny during normal development and maintenance of homeostasis. Recent studies show that Notch plays a key role in regulating the identity of neuroepithelial stem cells, which generate terminally differentiated neurons that populate the adult optic lobe via the intermediate progenitor cell type called neuroblast. Thus, understanding how Notch controls neuroepithelial cell maintenance and neuroblast formation will provide critical insight into the intricate regulation of stem cell function during tissue morphogenesis. This study shows that a low level of Notch signaling functions to maintain the neuroepithelial cell identity by suppressing the expression of pointedP1 gene through the transcriptional repressor Anterior open. Increased Notch signaling, which coincides with transient cell cycle arrest but precedes the expression of PointedP1 in cells near the medial edge of neuroepithelia, defines transitioning neuroepithelial cells that are in the process of acquiring the neuroblast identity. Transient up-regulation of Notch signaling in transitioning neuroepithelial cells decreases their sensitivity to PointedP1 and prevents them from becoming converted into neuroblasts prematurely. Down-regulation of Notch signaling combined with a high level of PointedP1 trigger a synchronous conversion from transitioning neuroepithelial cells to immature neuroblasts at the medial edge of neuroepithelia. Thus, changes in Notch signaling orchestrate a dynamic balance between maintenance and conversion of neuroepithelial cells during optic lobe neurogenesis (Weng, 2012).
A deregulated conversion of neuroepithelial cells into neuroblasts perturbs formation of the neuronal network and will almost certainly lead to visual impairment of the adult fly. Thus, a dynamic balance between neuroepithelial cell maintenance and differentiation plays a pivotal role during morphogenesis of the optic lobe. This study provides evidence that changes in Notch signaling regulate the dynamic balance between maintenance of neuroepithelial cells and formation of neuroblasts. A low level of Notch signaling maintains the neuroepithelial cell identity by triggering Aop-dependent repression of the pntP1 gene. Transient up-regulation of Notch signaling in transitioning neuroepithelial cells raises their threshold of response to PntP1 preventing them from precociously converting into immature neuroblasts. Finally, abrupt down-regulation of Notch signaling together with a high level of PntP1 trigger the conversion from transitioning neuroepithelial cells into immature neuroblasts at the medial edge of neuroepithelia. Thus, interplay between changes in Notch signaling and transient up-regulation of pntP1 orchestrates synchronous and progressive formation of neuroblasts in a medial-to-lateral orientation across the entire neuroepithelial swath (Weng, 2012).
Lack of Notch reporter transgene expression throughout neuroepithelia located laterally from transitioning neuroepithelial cells has been perplexing in light of recent studies reporting that Notch signaling is necessary for maintenance of their identity. One possibility might be that these Notch reporter transgenes including E(spl)mγ-GFP might not contain all necessary regulatory response elements to respond to Notch signaling in most neuroepithelial cells. Alternatively, the level of Notch signaling might simply be too low to activate the expression of the Notch reporter transgene. The second hypothesis is favored for the following reasons. Since over-expression of Notchintra is sufficient to trigger robust cell autonomous expression of E(spl)mγ-GFP in neuroepithelia located laterally from transitioning neuroepithelial cells, this transgene does contain all necessary regulatory elements to respond to a high level of Notch signaling. Furthermore, the Notch ligand Delta is expressed in a low level throughout neuroepithelia located laterally from transitioning neuroepithelial cells and Delta likely functions to trans-activate Notch signaling in these cells. In the context of trans-activation of Notch signaling by Delta, the level of the ligand correlates with the level of signaling output. Taken together, it is concluded that maintenance of the neuroepithelial cell identity requires a low level of Notch signaling (Weng, 2012).
It is proposed that Notch maintains the identity of neuroepithelial cells by activating Aop-dependent repression of the pntP1 gene. The Suppressor of Hairless protein, which is necessary for activating transcription of Notch targets genes, directly binds to the promoter of the aop gene. Furthermore, removing the Notch or aop function triggered premature conversion of neuroepithelia into neuroblasts whereas over-expressing Notch or aop prevented conversion of neuroepithelial cells into neuroblasts. Most importantly, over-expression of aop suppressed premature differentiation of Notch mutant neuroepithelial cells. Finally, heterozygosity of the pntP1 gene completely suppressed premature conversion of neuroepithelial cells into neuroblasts in a hypomorphic aop mutant genetic background. These data lead to the conclusion that Notch signaling maintains the identity of neuroepithelial cells by activating an Aop-dependent repression of pntP1. In the future, analyses of Notch and pntP1 double mutants will be necessary to confirm this regulatory mechanism (Weng, 2012).
Down-regulation of Notch signaling is necessary for formation of neuroblasts, so transient up-regulation of Notch signaling in transitioning neuroepithelial cells appears rather counterproductive. One possibility might be that up-regulation of Notch signaling paces the conversion from transitioning neuroepithelial cells into neuroblasts by increasing their threshold of response to PntP1. Consistent with this hypothesis, constitutively activated Notch signaling prevented transitioning neuroepithelial cells from becoming converted into neuroblasts despite expressing PntP1. This hypothesis was further supported by co-expression of pntP1 overcoming the blockade by constitutively activated Notch signaling and restoring conversion of transitioning neuroepithelial cells into neuroblasts. Thus, it is proposed that up-regulation of Notch signaling in transitioning neuroepithelial cells raises their threshold of response to PntP1 and functions to prevent them from becoming converted into immature neuroblasts precociously. Such an elaborated mechanism only permits transitioning neuroepithelial cells expressing the highest level of PntP1 to convert into immature neuroblasts. This mechanism is consistent with a recent study reporting that the EGF ligand is processed and secreted by cells near the medial edge of the optic lobe neuroepithelia. As a result of simple diffusion, transitioning neuroepithelial cells at the medial edge of neuroepithelia will be exposed to the highest level of the EGF ligand and will express the highest level of PntP1. As such, EGF signaling likely creates a vector field establishing the directionality of conversion from neuroepithelial cells into neuroblasts whereas Notch signaling refines the functional output of EGF signaling by raising the threshold response to PntP1 (Weng, 2012).
Many important questions arise from this highly plausible mechanism by which the interplay between Notch and EGF signaling paces synchronous conversion of neuroepithelial cells into neuroblasts one row at a time. This model will predict that immature neuroblasts immediately adjacent to transitioning neuroepithelial cells should secrete the processed EGF ligand. However, the antibody specific for the Rhomboid (Rho) protease required for proteolytic activation of the EGF protein is currently unavailable and a genomic fragment encompassing the rho-1 locus tagged with YFP did not show detectable expression in the larval optic lobe. Alternatively, a recent study shows that pntP1 is a direct target of Notch in vivo. Thus, up-regulation of Notch signaling might directly activate transcription of the pntP1 gene in transitioning neuroepithelial cells. Since Notch signaling becomes abruptly down-regulated at the medial edge of neuroepithelia, it is highly possible that the threshold of response to PntP1 also becomes lowered in the same cells. Thus, the pre-existing level of PntP1 protein will likely be sufficient to trigger the conversion from transitioning neuroepithelial cells into immature neuroblasts. More experiments including identification of the cell type from which the processed EGF ligand is released and a direct test to confirm the role of EGF signaling during conversion of neuroepithelia into neuroblasts will be key to distinguish these two possible mechanisms (Weng, 2012).
The fat-hippo signaling mechanism controls tissue growth by regulating proliferation and cell death and promotes timely differentiation of optic lobe neuroepithelial cells). While inactivation of fat-hippo signaling delays conversion of neuroepithelia into neuroblasts, removal of the downstream effecter yorkie only accelerates the conversion near the medial edge of the optic lobe neuroepithelia. Thus, fat-hippo signaling likely functions as a gatekeeper to prevent over-growth of optic lobe neuroepithelia by triggering transient cell cycle arrest. Intriguingly, transient cell cycle arrest precedes increased Notch signaling in transitioning neuroepithelial cells. Detailed studies in the future will be necessary to determine whether activation of the fat-hippo signaling might contribute to increased Notch signaling in transitioning neuroepithelial cells (Weng, 2012).
It is firmly established that neuron-glia interactions are fundamental across species for the correct establishment of a functional brain. This study found that the glia of the Drosophila larval brain display an essential non-autonomous role during the development of the optic lobe. The optic lobe develops from neuroepithelial cells that proliferate by dividing symmetrically until they switch to asymmetric/differentiative divisions generating neuroblasts. The proneural gene lethal of scute (l'sc) is transiently activated by the Epidermal Growth Factor Receptor (EGFR)/Ras signal transduction pathway at the leading edge of a proneural wave that sweeps from medial to lateral neuroepithelium promoting this switch. This process is tightly regulated by the tissue-autonomous function within the neuroepithelium of multiple signaling pathways, including EGFR/Ras and Notch. This study shows that the Notch ligand Serrate (Ser) is expressed in the glia and it forms a complex in vivo with Notch and Canoe, which colocalize at the adherens junctions of neuroepithelial cells. This complex is crucial for glial-neuroepithelial cell interactions during optic lobe development. Ser is tissue-autonomously required in the glia where it activates Notch to regulate its proliferation, and non-autonomously in the neuroepithelium where Ser induces Notch signaling to avoid the premature activation of the EGFR/Ras pathway and hence of L'sc. Interestingly, different Notch activity reporters showed very different expression patterns in the glia and in the neuroepithelium, suggesting the existence of tissue-specific factors that promote the expression of particular Notch target genes or/and a reporter response dependent on different thresholds of Notch signaling (Perez-Gomez, 2013).
Cno and its vertebrate homologues AF-6/Afadin localize at epithelial AJs where they regulate the linkage of AJs to the actin cytoskeleton by binding both actin and nectin family proteins. This study found that Cno colocalizes with Notch at the AJs of NE cells in the optic lobe proliferation centers. Notch also
colocalizes with its ligand Ser, which was detected at the glia, highly accumulated at the interface between NE cells and the surrounding glia. Co-immunoprecipitation experiments indicate the formation of a Ser-Notch-Cno complex in vivo, and the mutant analysis shows the functional relevance of such a complex at the glia neuroepithelium interface. The data presented in this study support the hypothesis that Cno may be stabilizing Notch at the AJs of NE cells, favoring the binding of Ser present in the adjacent glial cells. Indeed, in cno lof both Notch and Ser distribution is affected; this alteration is accompanied by an abnormally advanced proneural wave, a reminiscent phenotype to that shown by Notch lof optic lobes and also a similar phenotype found in this work in Ser lof. The activation of Notch pathway is essential to maintain the integrity of the neuroepithelium and to allow the correct progression of the proneural wave. The results show that glial Ser is responsible of such activation, promoting the expression of the m7-nuclacZ reporter in NE cells. In fact, the reduction of glial Ser either by knocking down epithelial cno or by expressing DNSer in the glia led to a decrease in the expression of the m7-nuclacZ reporter in NE cells and to an ectopic activation of the Ras/PntP1 pathway and of L'sc. It is proposed that this may be ultimately the cause of the proneural wave advance observed in those genotypes. Thus, the activation of Notch in the neuroepithelium by glial Ser, in nomal conditions, would be essential to avoid a premature activation of the EGFR/Ras/PntP1 pathway and hence of L'sc. Indeed, Notch has been shown to downregulate different EGFR/Ras signaling pathway components such as Rhomboid (Rho), required for the processing of the EGFR ligand Spitz, in other developmental contexts in which both pathways are actively cross-talking. Therefore, Notch activity in NE cells could be contributing to inhibit Rho, restricting its presence to the transition zone where Rho is very locally expressed (Perez-Gomez, 2013).
It was observed that in a WT condition Ser is present in all surface glia (perineurial and subperineurial), as shown by the expression of CD8::GFP (SerGal4>>UAS-CD8::GFP), and Notch, as tested by different reporters, is active in this tissue and highly reduced in Ser lof in the glia. This makes sense with the existence of a Ser-Notch mediated intercellular communication between the glial cells that comprise both the perineurial and subperineurial glia. Intriguingly, the knockin down and overexpression of cno in NE cells also had a clear effect on Notch activity in the glia, a reduction and an increase, respectively. This is more challenging to explain. As the cno lof in the NE led to a high reduction of both neuroepithelial Notch and glial Ser, the easiest explanation is that an 'excess' of unbound glial Ser is degraded and this impinges on the general thresholds of glial Ser, therefore causing a general reduction in the Notch activity in this tissue. This is an interesting field to explore in detail and is left open for future investigation (Perez-Gomez, 2013).
The activity of Notch in the neuroepithelium and in medulla NBs seems controversial. For example, Notch has been shown to be active in the neuroepithelium at low/null levels or in a 'salt and
pepper' patter. A weak/null activity of Notch has also been reported in NBs as well as a high activation. One possibility to conciliate all these results and apparently contradictory data is that different Notch target genes used as Notch activity reporters are, in fact, differentially activated in particular regions or tissues. The results support this proposal. Four different Notch reporters were used in this study. Whereas m7-nuclacZ was expressed throughout the neuroepithelium, Gbe+Su(H)lacZ was restricted to the transition zone, although both were expressed in medulla NBs along with mβ-CD2. In addition, mβ-CD2 was strongly activated in the glia, whereas the Gbe+Su(H)lacZ and the mδ-lacZ reporters were expressed at much lower levels at this location. Differential activation of Notch targets genes has been previously reported and tissue-specific factors could contribute to this differential expression. This is an intriguing scenario to analyze in the future. The in depth analysis of other Notch reporter genes in the developing optic lobe can contribute to further clarify this issue (Perez-Gomez, 2013).
At third larval instar during optic lobe development, Dl is highly restricted to 2-3 cells at the transition zone in the neuroepithelium, where Dl activates Notch. This work has found that the other ligand of Notch, Ser, is expressed in the surrounding glia at this larval stage and it is strongly accumulated at the interface with NE cells. Ser activates Notch in the neuroepithelium and this, in turn, would contribute to restrict the activation of the Ras-PntP1 pathway and L'sc to the transition zone. Intriguingly, it was observed that Ser preferentially activates the Notch target gene m7-nuclacZ in the neuroepithelium whereas Dl activates other Notch target genes, including Gbe+Su(H)lacZ, in the transition zone. For example, the overexpression of Dl in NE cells caused an ectopic activation throughout the neuroepithelium of Gbe+Su(H)lacZ, along with dpn that also behaves as a Notch target in other systems, and a repression of m7-nuclacZ . In addition, the lof of Ser in the glia caused a striking decrease in the expression of m7-nuclacZ in the neuroepithelium. One possibility to explain these observations is that the pool of Notch associated to the AJs and activated by glial Ser is subject of particular posttranslational modifications or/and is associated with other AJs proteins (including Cno) that somehow make Notch more receptive to Ser and able to activate specific target genes (i.e., m7). In this regard, it is interesting to note that Dl ectopically expressed in the glia (i.e., repoGal4>>>UAS-Dl) was not detected at the interface with NE cells, where glial Ser is highly present in contact with Notch, but Dl was restricted to the outermost surface glia (perineurial glia). This result strongly indicates that Dl cannot bind or has very low affinity for this pool of Notch at the AJs, hence being actively degraded in the subperineurial glia. This low affinity of Dl by Notch at this location further suggests that this pool of Notch at the AJs must be endowed with particular characteristics that ultimately could alter the activity properties of Su(H), explaining in turn the distinct expression pattern of Notch targets genes. Another possibility, which is not necessarily exclusive, to explain the differential activation of the Notch reporters is that they respond to different Notch thresholds. For example, m7-nuclacZ would require very low levels of Notch activation whereas Gbe+Su(H)lacZ would require high amounts of Notch signaling in NE cells. All these questions remain open for further investigation (Perez-Gomez, 2013).
A large number of neural and glial cell cell types differentiate from neuronal precursor cells during nervous system development. Two types of Drosophila optic lobe neurons, lamina and medulla neurons, are derived from the neuroepithelial (NE) cells of the outer optic anlagen. During larval development, epidermal growth factor receptor (EGFR)/Ras signaling sweeps the NE field from the medial edge and drives medulla neuroblast (NB) formation. This signal drives the transient expression of a proneural gene, lethal of scute, and its signal array is referred to as the 'proneural wave', since it is the marker of the EGFR/Ras signaling front. This study shows that the atypical cadherin Fat and the downstream Hippo pathways regulate the transduction of EGFR/Ras signaling along the NE field and, thus, ensure the progress of NB differentiation. Fat/Hippo pathway mutation also disrupts the pattern formation of the medulla structure, which is associated with the regulation of neurogenesis. A candidate for the Fat ligand, Dachsous is expressed in the posterior optic lobe, and its mutation was observed to cause a similar phenotype as fat mutation, although in a regionally restricted manner. It was also shown that Dachsous functions as the ligand in this pathway and genetically interacts with Fat in the optic lobe. These findings provide new insights into the function of the Fat/Hippo pathway, which regulates the ordered progression of neurogenesis in the complex nervous system (Kawamori, 2011).
The Fat/Hippo pathway has been known as a tumor suppressor pathway. This study and in the report of Reddy (2010), it was shown that the loss of Fat/Hippo signaling causes a delay of NB differentiation in the optic lobe. In contrast, dachs;ft double mutation, which is expected to stabilize the Fat/Hippo pathway, causes an advance of NB differentiation. This led to the question of how the Fat/Hippo pathway controls NB differentiation (Kawamori, 2011).
It has been reported that EGFR/Ras signaling is necessary and sufficient for NB induction, and its transduction is the driving force of the progress of the proneural wave. EGFR/Ras signaling sweeps the NE field through the gradual activation of Ras and its downstream EGF secretion by Rho. It was reasoned that this signal transduction is the target of the Fat/Hippo pathway in the control of NB differentiation. Indeed, ectopic expression of the EGFR/Ras signaling components RasV12 and rho was sufficient to induce NBs in the ft mutant background (Kawamori, 2011).
Which step of this cycling process does Fat/Hippo pathway mutation affect? Based on ectopic expression experiments, the Fat/Hippo pathway lies upstream of Ras and Rho, and it is expected to control the process from EGF transmission to Ras activation. The phenotypic difference produced by RasV12 and rho overexpression should be noted. When rho was expressed in the ft mutant background, several NE clones with abnormal morphology remained. This phenotype was not observed when RasV12 was expressed. In this model, RasV12 drives NB differentiation in RasV12-expressing cells in a cell-autonomous manner. In contrast, Rho activates Ras signaling in neighboring cells through the secretion of an EGF ligand. Based on these phenotypic differences, the Fat/Hippo pathway is expected to control the cell-to-cell EGF transmission, including its secretion, distribution or reception at the cell surface. This hypothesis is supported by the fact that several signaling components of the EGFR/Ras pathway, including Rho and EGFR, are localized to the apical side of epithelial tissues, and it is thought that this signal is transmitted along the apical side in epithelial tissues. It has also been reported that Fat/Hippo pathway mutations enhance the expression level of several apically localized molecules, such as aPKC, PatJ, Crumbs and E-cadherin. Thus, Fat/Hippo signaling targets could include unknown apical components that are involved in EGF transmission and this could account for the incomplete NB induction by rho overexpression. rho-expressing cells secret the EGF ligand, which diffuses in the NE surface, but Fat/Hippo pathway mutation would prevent its cell-to-cell transmission and subsequent EGFR/Ras pathway activation in the receiving cells. In this hypothesis, EGF transmission would be disturbed in the NE mutant for the Fat/Hippo pathway, causing the delay of proneural wave progress (Kawamori, 2011).
As an alternative hypothesis, the Fat/Hippo pathway could regulate signal transduction from the EGFR to Ras activation. If this is the case, the Fat/Hippo pathway regulates the intracellular signal transduction of the EGFR pathway. Many of the known targets of the Fat/Hippo pathway are components of growth regulatory, cell survival and cell adhesion molecules. There could be unknown targets that modulate other signaling pathways, including the EGFR pathway, and the NB differentiation defect would thus be caused by a failure in the activation of differentiation signals in the absence of Fat/Hippo signaling (Kawamori, 2011).
This study shows that the Fat/Hippo pathway mutation also affected the morphological character of the NE. Fat/Hippo pathway mutant clones were induced, and they often included NE tissue with a folded morphology and disrupted the medulla structure. The results showed that the Fat/Hippo pathway functions in the regulation of NB differentiation and in NE morphology are distinct, but the two functions could affect each other. The morphological defect of the NE could affect EGFR/Ras signal transduction. The possibility is discussed that the EGF ligand could be distributed along the apical membrane of the NE. The invagination of the apical membrane of the folded NE into the inner region could prevent EGF ligand signaling. There were clones with a normal NE morphology in which NB differentiation was delayed and, thus, morphological defects are not determinate, but they could promote the delay of NB induction (Kawamori, 2011).
How is the activity of the Fat/Hippo pathway regulated throughout the development of the optic lobe? Ft is a member of the cadherin family, and an extracellular molecule is expected to regulate its activity. Ds is a candidate for the Ft ligand that regulates planar cell polarity and Fat/Hippo signaling activity in other epithelial tissues. The expression of ds with a posterior-specific pattern in the developing optic lobe (Reddy, 2010) was confirmed. In the rescue experiments for the ds mutation, the expression of either ds lacking its intracellular domain (dsΔICD) or ft lacking its extracellular domain (ftΔECD) was sufficient to compensate for ds function, suggesting that Ds functions as a ligand and that Ft lies downstream of Ds in this context (Kawamori, 2011).
The phenotypes of ds and ft mutants were compared to assess whether the mutation of ds by itself accounts for the phenotype of the ft mutants. In contrast to the ft mutants that exhibited altered NB differentiation in the entire outer optic anlagen, the ds mutant phenotype was regionally specific; NB differentiation was severely delayed in the posterior region, and the development of the anterior region was not significantly affected. These differences suggest that there might be some regulatory mechanisms that control Ft activity independently of Ds in the anterior region of the optic lobe (Kawamori, 2011).
The Fat/Hippo pathway is known as a tumor suppressor pathway, and many studies related to this pathway have focused on tissue growth or cell survival. This study has reported a new function of the Fat/Hippo pathway in the regulation of neural differentiation. The Fat/Hippo pathway regulates the progress of neural differentiation signaling, and the EGFR/Ras pathway is a candidate target of this pathway. The data suggest that the Fat/Hippo pathway includes unknown targets involved in EGFR/Ras signal transduction. Further studies are required to identify the targets of the Fat/Hippo pathway and determine the interplay between Fat/Hippo and EGFR/Ras pathways, specifically in NB differentiation (Kawamori, 2011).
In Drosophila, defects in asymmetric cell division often result in the formation of stem cell derived tumors. This study shows that very similar terminal brain tumor phenotypes arise through a fundamentally different mechanism. Brain tumors in l(3)mbt mutants originate from overproliferation of neuroepithelial cells in the optic lobes caused by de-repression of target genes in the Salvador-Warts-Hippo (SWH) pathway. ChIP-seq was used to identify L(3)mbt binding sites, and it was shown that L(3)mbt binds to chromatin insulator elements. Mutating l(3)mbt or inhibiting expression of the insulator protein gene mod(mdg4) results in upregulation of SWH-pathway reporters. As l(3)mbt tumors are rescued by mutations in bantam or yorkie or by overexpression of expanded the deregulation of SWH pathway target genes is an essential step in brain tumor formation. Therefore, very different primary defects result in the formation of brain tumors, which behave quite similarly in their advanced stages (Richter, 2011).
Characterization of Drosophila genes identified in brain tumor suppressor screens has demonstrated that defects in neuroblast asymmetric cell division result in the formation of stem cell derived tumors that metastasize and become aneuploid upon transplantation. These screens also identified lethal (3) malignant brain tumor (l(3)mbt), a conserved transcriptional regulator that is also required for germ-cell formation in Drosophila. L(3)mbt binds to the cell cycle regulators E2F and Rb but the relevance of these interactions is unclear. This study shows that in Drosophila, L(3)mbt regulates target genes of the Salvador-Warts-Hippo (SWH) pathway that are important in proliferation and organ size control. The SWH-pathway is regulated by the membrane proteins Expanded (Ex) and Fat, which activate a protein complex containing the kinases Hippo and Warts to phosphorylate the transcriptional co-activator Yorkie. Yorkie acts together with the transcription factors Scalloped and Homothorax to activate proliferative genes like Cyclin E and the microRNA bantam (ban) and Drosophila inhibitor of apoptosis 1 (diap1: thread). Upon phosphorylation, Yorkie is retained in the cytoplasm and its target genes are not activated. In Drosophila the main role of the SWH-pathway is to limit proliferation in imaginal discs and its absence leads to tumorous overgrowth. In vertebrates, many homologs of key pathway members are tumor suppressors indicating that this function is conserved (Richter, 2011).
L(3)mbt contains three MBT domains which bind mono- or dimethylated histone tails. Biochemical experiments in vertebrates have suggested a role in chromatin compaction but whether this role is conserved is not known. Results published while this paper was under review have shown that germline genes are upregulated in l(3)mbt mutant brains and are necessary for tumor formation. The current data indicate that L(3)mbt is bound to insulator sequences, which affect promoter-enhancer interactions and influence transcription. In Drosophila, the proteins CTCF, CP190, BEAF-32, Su(Hw), Mod(mdg4) and GAF are found at insulator sequences but how these factors act is largely unknown (Richter, 2011).
The data presented in this study show that tumor formation in l(3)mbt mutants is initiated by the uncontrolled overproliferation of neuroepithelial cells in the optic lobes due to the upregulation of proliferation control genes normally repressed by the SWH-pathway. L(3)mbt is located at DNA sequences bound by chromatin insulators and we propose that the function of L(3)mbt as a chromatin insulator is essential for repressing SWH target genes and preventing brain tumor formation (Richter, 2011).
brat, lgl and dlg were previously identified as Drosophila brain tumor suppressors. In all cases, defects in asymmetric cell division cause a huge expansion of the neuroblast pool. In l(3)mbt mutants, however, the neuroblast pool is expanded because an upregulation of SWH target genes results in a massive expansion of neuroepithelial tissue. Why those neuroblasts proliferate indefinitely upon transplantation is currently not understood for any of those mutants (Richter, 2011).
While the SWH-pathway is essential for tumorigenesis in l(3)mbt mutants, its overactivation can not recapitulate the neuroblast tumor phenotype seen in l(3)mbt mutants (this study and Reddy, 2010). Similar to the multifactorial origin of mammalian tumors, therefore, the combined deregulation of several signaling pathways could be required. The Notch pathway could be involved as it regulates the formation of OL neuroblasts from neuroepithelia and Notch pathway gene insulator sequences are bound by L(3)mbt. Increased activity of the Jak/STAT pathway, a major regulator of OL development, was also observed. Finally, the deregulation of germline genes in l(3)mbt mutants that has been described while this manuscript was under review (Janic, 2010) could provide another exciting explanation (Richter, 2011).
The results indicate that L(3)mbt acts on insulator elements, which isolate promoters from the activity of nearby enhancers acting on other genes. the analysis showed that L(3)mbt binding sites overlap with CP190, CTCF and BEAF-32, placing the protein into what has been called the class I of chromatin insulators (Negre, 2010; Richter, 2011 and references therein).
The identification of a DNA consensus motif for a histone binding protein like L(3)mbt is highly unexpected as insulators are typically nucleosome free. Currently, the activity of these important transcriptional regulators could be explained in several ways. Either, they form physical barriers blocking the interaction between enhancers and promoters. Alternatively, they mimic promoters and compete with endogenous promoters for enhancer interaction. Finally, they could interact with each other or nuclear structures to form loop domains that regulate transcriptional activity. The data suggests another model in which insulators interact with histones on nearby nucleosomes and influence the structure of higher order chromatin. Importantly, in the regions flanking CTCF binding sites nucleosomes are enriched for histones that are mono- and di-methylated on H3K4 or mono-methylated on H3K9 or H4K20, the variants to which MBT domains can bind in vitro. As the human L(3)mbt homolog L3MBTL1 was shown to compact nucleosome arrays in vitro , a model becomes feasible in which simultaneous binding to insulators and the surrounding nucleosomes reduces flexibility and thereby restricts the ability of nearby enhancers to interact with promoters on the other side of the insulator. However, the data could equally well be worked into the other prevalent models for insulator activity. Since L(3)mbt is currently the only chromatin insulator besides CTCF that is conserved in vertebrates, analysis of its homologs will certainly allow to distinguish between those possibilities (Richter, 2011).
OL development resembles vertebrate neurogenesis. Both processes consist of an initial epithelial expansion phase followed by neurogenesis through a series of asymmetric divisions. Together with previous findings, these data demonstrate that l(3)mbt and the SWH-pathway are crucial regulators of the initial neuroepithelial proliferation phase. Interestingly, the SWH-pathway has been implicated in regulating neural progenitors in the chicken embryo and it will be exciting to test the role of mammalian L(3)mbt in this process. It is remarkable that YAP is upregulated and L3MBTL3 is deleted in a subset of human medulloblastomas. Medulloblastoma is the leading cause of childhood cancer death and investigating the role of the SWH-pathway might contribute to the progress in fighting this disastrous disease (Richter, 2011).
In the Drosophila optic lobes, the medulla processes visual information coming from inner photoreceptors R7 and R8 and from lamina neurons. It contains approximately 40,000 neurons belonging to more than 70 different types. This study describes how precise temporal patterning of neural progenitors generates these different neural types. Five transcription factors - Homothorax, Eyeless, Sloppy paired, Dichaete and Tailless - are sequentially expressed in a temporal cascade in each of the medulla neuroblasts as they age. Loss of Eyeless, Sloppy paired or Dichaete blocks further progression of the temporal sequence. Evidence is provided that this temporal sequence in neuroblasts, together with Notch-dependent binary fate choice, controls the diversification of the neuronal progeny. Although a temporal sequence of transcription factors had been identified in Drosophila embryonic neuroblasts, this work illustrates the generality of this strategy, with different sequences of transcription factors being used in different contexts (Li, 2013).
In the developing medulla, the wave of conversion of neuroepithelium into neuroblasts makes it possible to visualize neuroblasts at different temporal stages in one snapshot, with newly generated neuroblasts on the lateral edge and the oldest neuroblasts on the medial edge of the expanding crescent shaped neuroblast region. An antibody screen was conducted for transcription factors expressed in the developing medulla and five transcription factors, Hth, Ey, Slp1, D and Tll, were identified that are expressed in five consecutive stripes in neuroblasts of increasing ages, with Hth expressed in newly differentiated neuroblasts, and Tll in the oldest neuroblasts. This suggests that these transcription factors are sequentially expressed in medulla neuroblasts as they age. Neighbouring transcription factor stripes show partial overlap in neuroblasts with the exception of the D and Tll stripes, which abut each other. Previous studies have reported that Hth and Ey< were expressed in medulla neuroblasts, but they had not been implicated in controlling neuroblast temporal identities. Hth and Tll also show expression in the neuroepithelium (Li, 2013).
To address whether each neuroblast sequentially expresses the five transcription factors, their expression was examined in the neuroblast progeny. Hth, Ey and Slp1 are expressed in three different layers of neurons that correlate with birth order, that is, Hth in the first-born neurons of each lineage in the deepest layers; Ey or Slp1 in correspondingly more superficial layers, closer to the neuroblasts. This suggests that they are born sequentially in each lineage. D is expressed in two distinct populations of neurons. The more superficial population inherit D from D+ neuroblasts. D+ neurons in deeper layers (corresponding to the Hth and Ey layers) turn on D expression independently and will be discussed later. Single neuroblast clones were generated, and the expression of the transcription factors was examined in the neuroblast and its progeny. Single neuroblast clones in which the neuroblast is at the Ey+ stage include Ey+ GMCs/neurons as well as Hth+ neurons. This indicates that Ey+ neuroblasts have transited through the Hth+ stage and generated Hth+ neurons. Clones in which the neuroblast is at the D+ stage contain Slp1+ GMCs and Ey+ neurons, suggesting that D+ neuroblasts have already transited through the Slp+ and Ey+ stages. This supports the model that each medulla neuroblast sequentially expresses Hth, Ey, Slp1 and D as it ages, and sequentially produces neurons that inherit and maintain expression of the transcription factor (Li, 2013).
slp1 and slp2 are two homologous genes arranged in tandem and function redundantly in embryonic and eye development. Slp2 is expressed in the same set of medulla neuroblasts as Slp1. Slp1 and Slp2 are referred to collectively as Slp (Li, 2013).
Tll is expressed in the oldest medulla neuroblasts. The oldest Tll+ neuroblasts show nuclear localization of Prospero (Pros), suggesting that they undergo Pros-dependent cell-cycle exit at the end of their life, as in larval nerve cord and central brain neuroblasts. Tll+ neuroblasts and their progeny express glial cells missing (gcm), and the progeny gradually turn off Tll and turn on Repo, a glial-specific marker. These cells migrate towards deeper neuronal layers and take their final position as glial cells around the medulla neuropil. Thus, Tll+ neuroblasts correspond to previously identified glioblasts between the optic lobe and central brain that express gcm and generate medulla neuropil glia. Clones in which the neuroblast is at the Tll+ stage contain Hth+ neurons and Ey+ neurons, among others, confirming that Tll+ neuroblasts represent the final temporal stage of medulla neuroblasts rather than a separate population of glioblasts. Therefore, these data clearly show that medulla neuroblasts sequentially express five transcription factors as they age. The four earlier temporal stages generate neurons that inherit and maintain the temporal transcription factor present at their birth, although a subset of neurons born during the Ey, Slp or D neuroblast stages lose expression of the neuroblast transcription factor. At the final temporal stage, neuroblasts switch to glioblasts and then exit the cell cycle (Li, 2013).
Whether cross-regulation among transcription factors of the neuroblast temporal sequence contributes to the transition from one transcription factor to the next was examined. Loss of hth or its cofactor, extradenticle (exd), does not affect the expression of Ey and subsequent progression of the neuroblast temporal sequence (Li, 2013).
ey-null mutant clones were generated using a bacterial artificial chromosome (BAC) rescue construct recombined on a chromosome containing a Flip recombinase target (FRT) site in an eyJ5.71 null background. eyJ5.71 homozygous mutant larvae were also tested. In both cases, Slp expression is lost in neuroblasts, along with neuronal progeny produced by Slp+ neuroblasts, marked by the transcription factor Twin of eyeless (Toy, see below). However, neuroblast division is not affected, and Hth remains expressed in only the youngest neuroblasts and first-born neurons. Targeted ey RNA interference (RNAi) using a Vsx-Gal4 driver that is expressed in the central region of the neuroepithelium and neuroblasts gives the same phenotype. This suggests that Ey is required to turn on the next transcription factor, Slp, but is not required to repress Hth (Li, 2013).
In clones of a deficiency mutation, slpS37A, that deletes both slp1 and slp2, neuroblasts normally transit from Hth+ to Ey+, but older neuroblasts maintain the expression of Ey and do not progress to express D or Tll, suggesting that Slp is required to repress ey and activate D (Li, 2013).
Similarly, in D mutant clones, neuroblasts are also blocked at the Slp+ stage, and do not turn on Tll, indicating that D is required to repress slp and activate tll. Finally, in tll mutant clones, D expression is not expanded into oldest neuroblasts, suggesting that tll is not required for neuroblasts to turn off D. Thus, in the medulla neuroblast temporal sequence, ey, slp and D are each required for turning on the next transcription factor. slp and D are also required for turning off the preceding transcription factor (Li, 2013).
Gain-of-function phenotypes of each gene were studied. However, misexpression of Hth, Ey, Slp1 or Slp2, or D in all neuroblasts or in large neuroblast clones is not sufficient to activate the next transcription factor or repress the previous transcription factor in neuroblasts. Only misexpressing tll in all neuroblasts is sufficient to repress D expression (Li, 2013). In summary, cross-regulation among transcription factors is required for at least some of the transitions. No cross-regulation was observed between hth and ey. Because ey is already expressed at low levels in the neuroepithelium and in Hth+ neuroblasts, an as yet unidentified factor might gradually upregulate ey and repress hth to achieve the first transition. As tll is sufficient but not required to repress D expression, additional factors must act redundantly with Tll to repress D (Li, 2013).
The temporal sequence of neuroblasts described above could specify at least four neuron types plus glia (in fact more than ten neuron types plus glia considering that neuroblasts divide several times at each stage with overlaps between neighbouring temporal transcription factors). As this is not sufficient to generate the 70 medulla neuron types, it was asked whether another process increases diversity in the progeny neurons born from a neuroblast at a specific temporal stage. Apterous (Ap) is known to mark about half of the 70 medulla neuron types. In the larval medulla, Ap is expressed in a salt-and-pepper manner in subsets of neurons born from all temporal stages. In the progeny from Hth+ neuroblasts, all neurons seem to maintain Hth, with a subset also expressing Ap. However, only half of the neurons born from neuroblasts at other transcription factor stages maintain expression of the neuroblast transcription factor. For instance, in the progeny of Ey+ neuroblasts, Ey+ neurons are intermingled with about an equal number of Ey− neurons that instead express Ap. Neuroblast clones contain intermingled Ey+ and Ap+ neurons. This is also true for the progeny of Slp+ neuroblasts: Slp1+ neurons are intermingled with Slp1− Ap+ neurons. In the progeny of D+ neuroblasts, D and Ap are co-expressed in the same neurons, and they are intermingled with neurons that express neither D nor Ap. Neurons in deeper neuronal layers (corresponding to the Ey+ and Hth+ neuron layers) also express D independently, and these neurons are Ap−. The expression of Ap is stable from larval to adult stages (Li, 2013).
The intermingling of Ap+ and Ap− neurons raised the possibility that asymmetric division of GMCs gives rise to one Ap+ and one Ap− neuron. Two-cell clones were generated to visualize the two daughters of a GMC. In every case, one neuron is Ap+ and the other is Ap-, suggesting that asymmetric division of GMCs diversifies medulla neuron fates by controlling Ap expression (Li, 2013).
Asymmetric division of GMCs in Drosophila involves Notch (N)-dependent binary fate choice. In the developing medulla, the N pathway is involved in the transition from neuroepithelium to neuroblast, and loss of Su(H), the transcriptional effector of N signalling, leads to faster progression of neurogenesis and neuroblast formation. However, Su(H) mutant neuroblasts still follow the same transcription factor sequence and generate GMCs and neuronal progeny, allowing analysis of the effect of loss of N function on GMC progeny diversification. Notably, neurons completely lose Ap expression in Su(H) mutant clones. All mutant neurons born during the Hth+ stage still express Hth, but not Ap, suggesting that the NON daughters of Hth+ GMCs are the neurons expressing both Ap and Hth. In contrast to wild-type clones, all Su(H) mutant neurons born during the Ey+ neuroblast stage express Ey and none express Ap. Similarly, all mutant neurons born during the Slp+ neuroblast stage express Slp1 but lose Ap. These data suggest that, for Ey+ or Slp+ GMCs, the NOFF daughter maintains the neuroblast transcription factor expression, whereas the NON daughter loses this expression but expresses Ap. In the wild-type progeny born during the D+ neuroblast stage, Ap+ neurons co-express D. Both D and Ap are lost in Su(H) mutant clones in the D+ neuroblast progeny, confirming that D is transmitted to the Ap+ NON daughter of D+ GMCs. By contrast, the D+ Ap− neurons in the deeper layers (corresponding to the NOFF progeny born during the Ey+ and Hth+ neuroblast stages, see above) are expanded in Su(H) mutant clones at the expense of Ap+ neurons. Therefore, the deeper layer of D expression is turned on independently in the NOFF daughters of Hth+ and Ey+ GMCs (Li, 2013).
Finally, in wild type, a considerable amount of apoptotic cells were observed dispersed among neurons, suggesting that one daughter of certain GMCs undergoes apoptosis in some of the lineages. Together these data suggest that Notch-dependent asymmetric division of GMCs further diversifies neuronal identities generated by the temporal sequence of transcription factors (Li, 2013).
How does the neuroblast transcription factor temporal sequence, together with the Notch-dependent binary fate choice, control neuronal identities in the medulla? Transcription factor markers specifically expressed in subsets of medulla neurons, but not in neuroblasts, were examined including Brain-specific homeobox (Bsh) and Drifter (Dfr), as well as other transcription factors identified in the antibody screen, for example, Lim3 and Toy. Bsh is required and sufficient for the Mi1 cell fate, and Dfr is required for the morphogenesis of nine types of medulla neurons, including Mi10, Tm3, TmY3, Tm27 and Tm27Y (Hasegawa, 2011). Investigation were carried out to identify at which neuroblast temporal stage these neurons were born by examining co-expression with the inherited neuroblast transcription factors. Then whether the neuroblast transcription factors regulate expression of these markers and neuron fates was investigated. The results for each neuroblast stage are described below (Li, 2013).
Bsh is expressed in a subset of Hth+ neurons, suggesting that Bsh is in the NON daughter of Hth+ GMCs. Indeed, Bsh expression is lost in both Su(H) and hth mutant clones. Thus, both Notch activity and Hth are required for specifying the Mi1 fate, consistent with the previous report that Hth is required for the Mi1 fate. Ectopic expression of Hth in older neuroblasts is also sufficient to generate ectopic Bsh+ neurons, although the phenotype becomes less pronounced in later parts of the lineage. These data suggest that Hth is necessary and sufficient to specify early born neurons, but the competence to do so in response to sustained expression of Hth decreases over time. This is similar to embryonic CNS neuroblasts, where ectopic Hb is only able to specify early born neurons during a specific time window (Li, 2013).
Lim3 is expressed in all Ap− progeny of both Hth+ and Ey+ neuroblasts. Toy and Dfr are expressed in subsets of neurons born from Ey+ neuroblasts, as indicated by their expression in the Ey+ neuron progeny layer. The most superficial row of Ey+ Ap− neurons express Toy (and Lim3), suggesting that they are the NOFF progeny of the last-born Ey+ GMCs. Dfr is co-expressed with Ap in two or three rows of neurons that are intermingled with Ey+ neurons, suggesting that they are the NON progeny from Ey+ GMCs. In addition to these Ap+ Dfr+ neurons, Dfr is also expressed in some later-born neurons that are Ap− but express another transcription factor: Dachshund (Dac), in specific sub-regions of the medulla crescent (Li, 2013).
Whether Ey in neuroblasts regulates Dfr expression in neurons was tested. As expected, Dfr-expressing neurons are lost in ey-null mutant clones, suggesting that they require Ey activity in neuroblasts, even though Ey is not maintained in Ap+ Dfr+ neurons. Furthermore, in slp mutant clones in which neuroblasts remain blocked in the Ey+ state, the Ap+ Dfr+ neuron population is expanded into later-born neurons, suggesting that the transition from Ey+ to Slp+ in neuroblasts is required for shutting off the production of Ap+ Dfr+ neurons. In addition, Ap+ Dfr+ neurons are lost in Su(H) mutant clones. Thus, Ey expression in neuroblasts and the Notch pathway together control the generation of Ap+ Dfr+ neurons (Li, 2013).
In addition to its expression with Ey in the NOFF progeny of the last-born Ey+ GMCs, Toy is also expressed in Ap+ (NON) neurons in more superficial layers generated by Slp+ and D+ neuroblasts. Consistently, in Su(H) mutant clones, an expansion of Toy+ Ey+ neurons is seen in the Ey progeny layer, followed by loss of Toy in the Slp and D progeny layer (Li, 2013).
Tests were performed to see whether Slp is required for the neuroblasts to switch from generating Toy+ Ap− neurons, progeny of Ey+ neuroblasts, to generating Toy+ Ap+ neurons. Indeed, in slp mutant clones, the Toy+ Ap+ neurons largely disappear, whereas Toy+ Ap− neurons expand (Li, 2013).
WAp and Toy expression was examined in specific adult neurons. OrtC1-gal4 primarily labels Tm20 and Tm5 plus a few TmY10 neurons, and these neurons express both Ap and Toy. To examine whether Slp is required for the specification of these neuron types, wild-type or slp mutant clones were generated using the mosaic analysis with a repressible cell marker (MARCM) technique by heat-shocking for 1 h at early larval stage, and the number of OrtC1-gal4-marked neurons in the adult medulla was examined. In wild-type clones, OrtC1-gal4 marks ~100 neurons per medulla. By contrast, very few neurons are marked by OrtC1-gal4 in slp mutant clones. Slp is unlikely to directly regulate the Ort promoter because Slp expression is not maintained in Ap+ Toy+ neurons. Furthermore, the expression level of OrtC1-gal4 in lamina L3 neurons is not affected by slp mutation. These data suggest that loss of Slp expression in neuroblasts strongly affects the generation of Tm20 and Tm5 neurons (Li, 2013).
In summary, these data show that the sequential expression of transcription factors in medulla neuroblasts controls the birth-order-dependent expression of different neuronal transcription factor markers, and thus the sequential generation of different neuron types (Li, 2013).
Although a temporal transcription factor sequence that patterns Drosophila nerve cord neuroblasts was reported more than a decade ago, it was not clear whether the same or a similar transcription factor sequence patterns neural progenitors in other contexts. The current identification of a novel temporal transcription factor sequence patterning the Drosophila medulla suggests that temporal patterning of neural progenitors is a common theme for generating neuronal diversity, and that different transcription factor sequences might be recruited in different contexts (Li, 2013).
There are both similarities and differences between the two neuroblast temporal sequences. In the Hb-Kr-Pdm-Cas-Grh sequence, ectopically expressing one gene is sufficient to activate the next gene, and repress the previous gene, but these cross-regulations are not necessary for the transitions, with the exception of Castor. In the Hth-Ey-Slp-D-Tll sequence, removal of Ey, Slp or D does disrupt cross-regulations necessary for temporal transitions (except the Hth-Ey transition). However, in most cases these cross-regulations are not sufficient to ensure temporal transitions, suggesting that additional timing mechanisms or factors are required (Li, 2013).
For simplicity, the medulla neuroblasts are represented as transiting through five transcription factor stages, whereas in fact the number of stages is clearly larger than five. First, neuroblasts divide more than once while expressing a given temporal transcription factor, and each GMC can have different sub-temporal identities. Furthermore, there is considerable overlap between subsequent temporal neuroblast transcription factors: neuroblasts expressing two transcription factors are likely to generate different neuron types from neuroblasts expressing either one alone (Li, 2013).
Although the complete lineage of medulla neuroblasts is still being investigated, this study shows how a novel temporal sequence of transcription factors is required to generate sequentially the diverse neurons that compose the medulla. The requirement for transcription factor sequences in the medulla and in embryonic neuroblasts suggests that this is a general mechanism for the generation of neuronal diversity. Interestingly, the mammalian orthologue of Slp1, FOXG1, acts in cortical progenitors to suppress early born cortical cell fates. Thus, transcription-factor-dependent temporal patterning of neural progenitors might be a common theme in both vertebrate and invertebrate systems (Li, 2013).
During development, neural progenitors are temporally patterned to sequentially generate a variety of neural types. In Drosophila neural progenitors called neuroblasts, temporal patterning is regulated by cascades of Temporal Transcription Factors (TTFs). However, known TTFs were mostly identified through candidate approaches and may not be complete. In addition, many fundamental questions remain concerning the TTF cascade initiation, progression, and termination. This work used single-cell RNA sequencing of Drosophila medulla neuroblasts of all ages to identify a list of previously unknown TTFs, and experimentally characterize their roles in temporal patterning and neuronal specification. This study reveals a comprehensive temporal gene network that patterns medulla neuroblasts from start to end. Furthermore, the speed of the cascade progression is regulated by Lola transcription factors expressed in all medulla neuroblasts. This comprehensive study of the medulla neuroblast temporal cascade illustrates mechanisms that may be conserved in the temporal patterning of neural progenitors (Zhu, 2022).
scRNA-Seq analysis revealed the temporal progression of transcriptional profiles as medulla NBs age at single-cell resolution. Candidates of critical temporal patterning regulators included eight previously unknown TTFs, as well as TFs such as Nerfin-1 and Lola, that are also involved in the temporal patterning process. Further experimental validation of previously unknown TTFs and other crucial regulators confirmed the accuracy of the high-resolution data, supporting that scRNA-seq is a powerful tool to study the highly dynamic temporal patterning process. This analysis and further experimental investigation revealed a comprehensive temporal cascade in Drosophila medulla NBs: Hth+SoxN+dmrt99B->Opa->Ey+Erm->Ey+Opa->Slp+Scro->D->BarH1&2->Tll, Gcm (see A schematic model summarizing the medulla TTF cascade and its regulation), and also illustrated several principles that are likely conserved during the temporal patterning of neural progenitors (Zhu, 2022).
First, this study identified early temporal factors that initiate the medulla neuroblast TTF cascade. Before this study, Hth was proposed to be the only TTF at play during the earliest temporal stage. Hth is expressed in the neuroepithelium and the youngest NBs. It is necessary for the generation of Bsh neurons, but is required neither for the NE to NB transition nor for the further temporal cascade progression. Loss of Ey also does not affect the termination of Hth. These data suggested missing links between Hth and the later TTF cascade. Several previously unknown TTFs were identified that linked the whole cascade together. Two of those TTFs that start their expression in the NE, SoxN, and Dmrt99B, are also required for the first temporal fate (Bsh neurons), and Dmrt99B is required for the timely activation of Opa in the youngest NBs. Opa is then required to activate Ey and repress Hth. Interestingly, the three TTFs inherited from NE maintain their expression for different durations in NBs, as Hth is repressed by Opa and Erm, SoxN is repressed by Ey, whereas Dmrt99B expression extends until the Slp stage. Whether this differential downregulation is significant for temporal patterning is currently unknown. However, it is worth noting that the expression of mammalian orthologs of Dmrt99B, Dmrt3, and Dmrta1, also starts in symmetrically dividing early cortical progenitors (NE), and decreases gradually in asymmetrical dividing cortical progenitors due to the direct suppression by FoxG1, the mammalian ortholog of Slp1/2. Given the essential role of Dmrt99B in initiating temporal patterning in medulla neuroblast, it will be interesting to investigate whether its mammalian orthologs play conserved roles in the temporal patterning of cortical progenitors (Zhu, 2022).
Second, this study showed that a broad temporal stage can be divided into sub-temporal stages by combinations of TTFs, which determine the progeny fates. This is well-illustrated in the Ey stage. The first stripe of Opa is necessary to initiate the expression of Erm and Ey, which are then required to repress Opa in a negative feedback loop, generating a gap in Opa expression. Furthermore, the data suggest that Ey may first enhance the activation of Erm at the gap, but then possibly a higher level of Ey is required to repress Erm, either directly or indirectly. After Erm is turned off, Opa is turned back on. At the same time, Slp has been gradually activated by Ey and Scro, and when it reaches a certain level, it will repress Opa and Ey to end the Ey stage. Thus, cross-regulations among TTFs divide the Ey stage into (at least) two (sub-)temporal stages determined by the co-expression of Ey and Erm, or Ey and Opa. Different neural types are generated in these two sub-temporal stages, and the first set of neurons require both Ey and Erm, whereas the second set of neurons require both Ey and Opa. Interestingly to note, the mammalian ortholog of Erm, Fezf2, is also expressed in cortical progenitors and plays important roles in cortical neuron specification (Zhu, 2022).
Third, this study demonstrated that a TTF that is required for the switch to gliogenesis at the final stage is also required for the cell-cycle exit and termination of the medulla TTF cascade. Previously it was thought that Tll stage NBs switch to gliogenesis and then exit the cell cycle, but whether Tll indeed plays a role in these processes has not been studied. Here, the scRNA-Seq data suggested another final temporal stage marked by the expression of Gcm and Dap. Further, it was shown that BarH1 and BarH2 are required to activate both Tll and Gcm, but Tll is activated first, and when Gcm is activated, Gcm represses Tll. Gcm but not Tll is required for the NBs to switch to gliogenesis and exit the cell cycle. Gcm is well-known for its role in gliogenesis, but this study showed that it is also required and sufficient to activate Dap expression in NBs, possibly through which to promote cell-cycle exit and end the temporal progression. In vertebrate retina, scRNA-seq analysis of retinal progenitor cells identified NFI factors as required for both late-born cell fates including Muller glia and for exiting the cell cycle. As neural progenitors often switch to produce glia at the end of the lineage, it is possibly a general mechanism that factors required for the switch to gliogenesis are also required for the mitotic exit to end the temporal progression (Zhu, 2022).
Another factor that is likely involved in the final stage is Nerfin-1. The expression of Nerfin-1 is observable mostly in maturing neurons, and is required to prevent neurons from de-differentiation. However, this TF responsible for maintaining the differentiation status of neurons, is turned on in the final-stage NBs, where it may function to promote gliogenesis and help terminate the temporal cascade on time. The fast exit of the cell cycle at the final stage is likely accomplished because self-renewal repressors that usually function in GMCs and neurons, such as Prospero and Nerfin-1, gather and cooperate in the oldest NBs. Whether Nerfin-1 can be characterized as a TTF is a remaining question. Since Nerfin-1 expression in both the oldest NBs and the newly born glia is very transient, and cell cycle exit is coupled with glia generation in the oldest NBs, it is not easy to distinguish when exactly Nerfin-1 functions to contribute to the termination of the final temporal stage. The mechanism behind Nerfin-1's requirement at the final stage may be different from the mechanism used in neurons preventing their de-differentiation. One evidence is that while a previous study showed that double knockdown of Nerfin-1 and Su(H) could reduce most ectopic NBs generated by single knockdown of Nerfin-1, suggesting that Nerfin-1 represses Notch signaling in neurons to prevent their de-differentiation, there are always several ectopic NBs remaining located at the medial edge inside the double knockdown clone. The location of those ectopic NBs indicates that they are likely the oldest NBs unable to exit the cell cycle. Therefore, Nerfin-1 may function through a different mechanism in the final-stage NBs, which is not dependent on the downregulation of Notch signaling. Finally, it was shown that Nerfin-1 is not required for Gcm expression, and it remains to be determined whether Gcm regulates Nerfin-1's expression in this process (Zhu, 2022).
Fourth, complex cross-regulations were observed among TTFs that form temporal gene networks. The model for the cross-regulations between medulla TTFs was that each TTF activates the next TTF and inhibits the previous TTF from the Ey stage to the end of the cascade, exhibiting a simple combination of feedforward activation and feedback repression. However, based on the experimental evidenc,e as well as inferred from the scRNA-seq data, the cross-regulations among TTFs are more complex. One TTF is not necessarily repressed by the very next TTF, or activated by the exactly previous TTF. Hth is repressed by Opa and Erm. SoxN is repressed by Ey, while Dmrt99B is likely to be repressed by Slp or later TTFs. Tll is activated just before Gcm, however, Tll is not required for Gcm's activation. The complexity of their cross-regulation is a way to increase the number of combinations of TTFs in aging NBs, thereby increasing the number of possible neuron fates determined along with the temporal progression. However, the overall trend that early TTFs activate late TTFs, and late TTFs repress early TTFs remains valid (Zhu, 2022).
Finally, the speed of the TTF cascade progression is regulated by Lola factors expressed in all NBs. Lola proteins belong to a BTB/POZ family of proteins which have been shown to be involved in chromatin remodeling and organization. Certain isoforms of Lola are expressed in all NBs, e.g., Lola-F is activated one cell cycle earlier than Opa. WLola proteins function as a speed modulator of the temporal cascade progression. It represses the expression of Hth, facilitates the activation of Opa and the following TTFs to different extents, thereby guaranteeing a quick transition from the NE TTF network to the NB TTF network. Interestingly, the vertebrate ortholog of lola, Zbtb20, was also found to modulate the sequential generation of different neural types in cortical progenitors. Loss of Zbtb20 causes the temporal transitions to be delayed further and further, very similar to the loss of lola phenotype in this system. Thus, it is possible that lola/Zbtb20 play conserved roles in the temporal patterning of neural progenitors (Zhu, 2022).
In summary, the entire life of a medulla neuroblast from the beginning to the end was revealed in this study. This comprehensive study of the medulla neuroblast temporal cascade illustrated mechanisms that may be conserved in the temporal patterning of neural progenitors. The single-cell RNA-sequencing data provide a plethora of information that allows further exploration of the mechanisms of temporal patterning (Zhu, 2022).
Temporal patterning is an important mechanism for generating a great diversity of neuron subtypes from a seemingly homogenous progenitor pool in both vertebrates and invertebrates. Drosophila neuroblasts are temporally patterned by sequentially expressed Temporal Transcription Factors (TTFs). These TTFs are proposed to form a transcriptional cascade based on mutant phenotypes, although direct transcriptional regulation between TTFs has not been verified in most cases. Furthermore, it is not known how the temporal transitions are coupled with the generation of the appropriate number of neurons at each stage. This study used neuroblasts of the Drosophila optic lobe medulla to address these questions and show that the expression of TTFs Sloppy-paired 1/2 (Slp1/2) is directly regulated at the transcriptional level by two other TTFs and the cell-cycle dependent Notch signaling through two cis-regulatory elements. It was also shown that supplying constitutively active Notch can rescue the delayed transition into the Slp stage in cell cycle arrested neuroblasts. These findings reveal a novel Notch-pathway dependent mechanism through which the cell cycle progression regulates the timing of a temporal transition within a TTF transcriptional cascade (Ray, 2022).
Drosophila neuroblasts are temporally patterned by sequentially expressed TTFs. Although the expression pattern and mutant phenotypes suggest that TTFs form a transcriptional cascade, direct transcriptional regulation between TTFs has not been demonstrated in most cases. This work has characterized two enhancers of the slp genes that enable the expression of Slp1 and Slp2 in medulla neuroblasts. The u8772 220 bp enhancer is activated at an earlier stage relative to the d5778 850 bp enhancer. In these two enhancers, sites were identified for the previous TTF -Ey and Scro-a TTF expressed at around the same time as Slp1. Deleting either enhancer alone did not eliminate the expression of endogenous Slp1 and Slp2, suggesting that they act partially redundantly with one another. Deletion of both enhancers completely eliminates Slp1 and Slp2 expression in medulla neuroblasts but does not affect their expression in lamina neurons or glia, confirming the specificity and necessity of these two enhancers. Using GFP reporter assays, this study has shown that mutation of Ey binding sites in these enhancers abolishes reporter expression similar to genetic experiments where a loss of GFP reporter was reported within ey RNAi clones. The results are also consistent with previous studies that showed a complete loss of endogenous Slp1/2 expression in UAS-ey-RNAi expressing neuroblasts. This study also confirmed the in vivo binding of Ey to the identified enhancers of Slp by Dam-ID sequencing. The expression of the TTF Scro is initiated simultaneously as Slp1/2, and it has been shown that loss of Scro significantly reduces Slp expression level. Mutation of most probable Scro binding sites on the u8772 220 bp or d5778 850 bp enhancers led to a dramatic reduction of GFP reporter expression. Thus, the combined effect of mutating Scro binding sites on both enhancers recapitulates the observed impact of Scro knock-down on endogenous Slp1/2 expression, which is reduced expression of Slp1/2 in neuroblasts expressing UAS scro-RNAi and a consequent loss of neural fates specified by Slp1/2 in their progeny. It is interesting to note that the observation of multiple enhancers regulating Slp1/2 expression is consistent with regulation of Slp1/2 in other developmental contexts. Previous studies have noted the presence of multiple enhancers of Slp1/2 expression in the vicinity of the slp1 and slp2 coding loci. Many of these regulatory DNA segments function as stripe enhancers enabling Slp1/2 to function as pair-rule genes during embryonic segmentation. Although these enhancers share some overlapping functions and domains of activation, a full complement of stripe enhancers is required for maintaining parasegment boundaries and wingless expression (Ray, 2022).
It was previously demonstrated that although Ey is necessary for activating Slp1/2 expression it is not sufficient. There is always a time delay after the start of Ey expression to the start of Slp expression to ensure the sufficient duration of the Ey window. How is the timing controlled? From analyses of the slp1/2 enhancer sequences, several binding sites were found for the CSL transcription factor Su(H), most prominently known as a central component of the ternary Notch transcription complex and the primary DNA binding component. To confirm the involvement of the Notch pathway in regulating Slp1/2, we observed the effects of knocking down key Notch pathway components on endogenous Slp1/2 expression. In all cases a delay was observed in the expression of Ey and a further delay in the transition to the Slp1/2 stage in neuroblasts expressing the RNAi knockdowns. Mutating Su(H) binding sites in the u8772 220bp enhancer led to a loss or reduction of GFP reporter expression in neuroblasts. However, mutating Su(H) binding sites in the d5778 850 bp enhancer did not decrease the reporter expression. These results suggest that Notch signaling directly regulates Slp expression through the u8772 220 bp enhancer, but not the d5778 850 bp enhancer, and this is consistent with the delayed expression driven by the d5778 850 bp enhancer. However, Ey still plays a more critical role in activating Slp1/2 expression than the Notch pathway, since Slp1/2 are still expressed albeit later in the absence of Su(H) and other Notch components, and Notch signaling requires Ey to speed up the Ey to Slp transition. As with Ey, this study confirmed Su(H) binding to the u872 220 bp enhancer using DamID-seq. Thus, this work provided strong evidence that N signaling, a general signaling pathway involved in neuroblast development, regulates the timing of activation of a TTF gene directly. In addition, the results also raised the interesting hypothesis that Notch signaling might be involved in facilitating all temporal transitions, because the turning on of Opa and Ey is also delayed, and a further and further delay was observed in turning on of later TTFs. Whether Notch signaling regulates other TTF expression directly or indirectly still awaits further investigation. (Ray, 2022).
What might explain the delay in Slp1/2 expression in the absence of Notch signaling? Recent developments in single-molecule Fluorescence In Situ Hybridization (smFISH) technology and live imaging techniques using the MS2-MCP system have enabled studying the transcription process in molecular detail. Imaging transcription driven by Notch responsive enhancers in native contexts has shown this process to be inherently 'bursty', i.e., episodes of transcription (enhancer 'On' state) are punctuated with gaps in activity (enhancer in 'Off' state) (Falo-Sanjuan et al., 2019; Lee et al., 2019). The dosage of NICD modulates the duration of the 'On' phase in one context studied by live imaging (Falo-Sanjuan et al., 2019; Lee et al., 2019). Additionally, binding of tissue-specific regional factors to these Notch responsive enhancers may prime these enhancers and help synchronize transcription and sustain a steady transcriptional output upon Notch binding to enhancers; this helps integrate important positional cues and the perception of context (Falo-Sanjuan et al., 2019). Applying these insights to our system, we suggest that Ey may act by priming the Notch-responsive enhancer of slp providing crucial contextual information, and this is required for Notch to further activate Slp1/2 transcription, and speed up the transition. (Ray, 2022).
Notch target genes and Dpn are transcriptional repressors that act partially redundantly to maintain neuroblast identity. In type II NBs, Dpn depends on Notch signaling, and loss of Dpn causes premature differentiation. However, in type I NBs, Dpn is not lost when Notch signaling is lost, and Notch signaling seems dispensable for the self-renewing abilities of NBs. In the medulla neuroblasts, this study also observed that in Su(H) mutant clones, the clone size and neuroblast proliferation are not significantly affected. On the other hand, N signaling was observed to be dependent on cell-cycle progression, and the Notch target gene is lost when cell cycle progression is blocked. (Ray, 2022).
In the medulla, blocking cell cycle progression in neuroepithelial cells prematurely transforms them into neuroblasts, and these neuroblasts seem to be arrested or severely delayed in the TTF cascade. When the cell cycle was arrested or slowed down later in neuroblasts to preserve Ey expression, Slp expression was is still delayed. Therefore, cell cycle progression also has a role in the Ey to Slp transition. Further, this study showed that supplying Notch signaling is sufficient to rescue the delay in the Ey to Slp transition caused by cell cycle defect. Thus at the Ey to Slp transition, the cell cycle effect is mediated through the direct regulation of Slp transcription by Notch signaling. Taken together, these results suggest that in Ey stage neuroblasts, Ey is required to initiate Slp expression but not sufficient to activate it to a strong level right away, and after each asymmetric division, activation of Notch signaling in the neuroblast enhances Slp expression, until Slp expression reaches a certain level to repress Ey expression and make the transition. This can be part of a mechanism to coordinate the TTF temporal transition with the cell cycle progression to generate the appropriate number of neural progenies at a given temporal stage (Ray, 2022).
How neuronal and glial fates are specified from neural precursor cells is an important question for developmental neurobiologists. This study addresses this question in the Drosophila optic lobe, composed of the lamina, medulla, and lobula complex. It was shown that two gliogenic regions posterior to the prospective lamina also produce lamina wide-field (Lawf) neurons, which share common progenitors with lamina glia. These progenitors express neither canonical neuroblast nor lamina precursor cell markers. They bifurcate into two sub-lineages in response to Notch signaling, generating lamina glia or Lawf neurons, respectively. The newly born glia and Lawfs then migrate tangentially over substantial distances to reach their target tissue. Thus, Lawf neurogenesis, which includes a common origin with glia, as well as neuronal migration, resembles several aspects of vertebrate neurogenesis (Chen, 2016).
Neural stem cells must balance symmetric and asymmetric cell divisions to generate a functioning brain of the correct size. In both the developing Drosophila visual system and mammalian cerebral cortex, symmetrically dividing neuroepithelial cells transform gradually into asymmetrically dividing progenitors that generate neurons and glia. As a result, it has been widely accepted that stem cells in these tissues switch from a symmetric, expansive phase of cell divisions to a later neurogenic phase of cell divisions. In the Drosophila optic lobe, this switch is thought to occur during larval development. However, this study found that neuroepithelial cells start to produce neuroblasts during embryonic development, demonstrating a much earlier role for neuroblasts in the developing visual system. These neuroblasts undergo neurogenic divisions, enter quiescence and are retained post-embryonically, together with neuroepithelial cells. Later in development, neuroepithelial cells undergo further cell divisions before transforming into larval neuroblasts. These results demonstrate that the optic lobe neuroepithelium gives rise to neurons and glia over 60 h earlier than was thought previously (Hakes, 2018).
The importance of producing the correct numbers of neurons during development is illustrated by both evolutionary enhancement of cognitive capacities in larger brains, and developmental disorders of brain size. In humans, increased neuronal numbers during development is speculated to partly derive from a unique subtype of neural stem cells (NSCs) that undergo a phase of expansion through symmetric self-amplifying divisions before generating neurons. Symmetric amplification also appears to underlie adult neural stem maintenance in the mouse. However, the mechanisms regulating this behavior are unclear. This study reports the discovery of self-amplifying NSCs in Drosophila and shows that they arise by a spatiotemporal conversion of classical self-renewing NSCs. This conversion is regulated by a temporal transition in the expression of proneural transcription factors prior to cell division. A causal link was found between stem cell self-amplification and increased neuronal numbers. It was further shown that the temporal transcriptional switch controls both stem cell division and subsequent neuronal differentiation (Mora, 2018).
The development of functional organs relies on the coordinated production of cells of different identities with temporal, spatial, and numerical precision. In the brain, where information processing depends on the output of interconnected neuronal circuits, not only the ratios of different neuronal subtypes, but also absolute numbers are important for optimal function. The number of neurons in the adult brain is a direct consequence of a spatiotemporally coordinated sequence of divisions of neural stem cells (NSCs) during development. However, it remains unclear how NSCs alter their division patterns over time and whether these alterations are causal to the generation of the correct number of neurons. Less clear still is whether and how the temporal transitions in NSC division influence the differentiation of their progeny (Mora, 2018).
In both mammals and insects NSCs regulate neurogenesis through a series of self-renewing divisions. NSC division patterns can be broadly classified in five categories. In three of these, NSCs divide asymmetrically renewing themselves and giving rise to daughters that differ in their proliferation potentials: daughters that do not divide, daughters that divide once, and daughters that divide multiple times. In the other two, NSCs divide symmetrically. One type of symmetric division common to vertebrates and invertebrates signals the end of stemness through the generation of two daughter cells committed to differentiation. A second, much rarer type, expands the progenitor pool through the generation of two cells, which retain the expression of NSC markers and the ability to generate neurons. In mouse, self-renewal by symmetric division has recently been reported to be predominant during adult neurogenesis, in contrast to what is observed in embryonic stages where most NSCs divide asymmetrically. In the primate brain, embryonic self-amplifying divisions have been detected in the NSCs known as outer radial glia (oRG). Multiple lines of evidence support the hypothesis that oRGs' high abundance and proliferative capacity are critical for the vast increase of brain size in primates. However, the direct evidence for the impact of symmetric amplification of NSCs on neuronal numbers, the mechanisms that mediate the switch from self-renewal to self-amplification and then to neurogenesis, and the impact of such a switch on terminal differentiation remain unexplored (Mora, 2018).
The fruit fly Drosophila melanogaster has long been a powerful model system for the discovery of the genetic, cellular, and molecular underpinnings of the behavior of NSCs, as well as the generation and differentiation of their neuronal progeny. Drosophila NSCs are called neuroblasts (Nbs), and two major modes of neurogenesis have been described. The type I Nbs self-renew while giving rise to committed daughters called ganglion mother cells (GMCs) that in turn divide terminally to produce two neurons or glia. The type II Nbs also self-renew but produce intermediate progenitors that in turn undergo a limited number of self-renewing divisions giving rise to GMCs, which give rise to neurons. Thus, to date, all Nbs in the fly brain are thought to produce neurons by asymmetric self-renewal and no symmetrically dividing, self-amplifying, NSCs have been found (Mora, 2018).
The majority of the fly brain is dedicated to visual processing. The higher-order visual centers called the optic lobes (OLs) receive the visual input from the retina and are arranged in four neuropils called lamina, medulla, and lobula plate (LP); all four organized in retinotopic maps. OL neurons derive from two major proliferation zones, called the outer proliferation center and the inner proliferation center (IPC), containing actively dividing Nbs. The organization of the OLs is constrained by the characteristics of the compound eye, which is composed of 1~750 repetitive units of 8 photoreceptors covering the visual field and projecting to the OL in a retinotopic order. This integration of the retinal map requires a tight control of the diversity and stoichiometry of the neuronal populations. While temporal and spatial cues required to generate different types of neurons have been identified, the control of the production of large numbers of neurons is much less understood. One striking example is the motion detection neurons of the LP called T4/T5. For each of the 750 retinal units, the LP contains 8 different T4/T5 direction sensitive neurons (T4a, b, c, d, and T5a, b, c, d, respectively). Thus, the direction-selective T4/T5 lineage generates approximately 12,000 neurons, representing more than 10% of all neurons in the fly brain. How such a massive proportion of neurons is generated is entirely unknown (Mora, 2018).
Another highly conserved feature of neurogenesis is that it is regulated by a small and highly conserved set of transcription factors known as the proneural proteins. First described in Drosophila, basic loop-helix proneural factors regulate neurogenesis in insects as well as in mammals. There are three families of proneural proteins named after their founding members; the Atonal (ATH), Achaete-scute (AS), and Neurogenin families. Proneural proteins most conserved function is to provide progenitors with the neuronal fate. In addition, they have been found to promote asymmetric division, exit from cell cycle and initiation of differentiation. Whether proneural proteins can promote symmetric proliferation or if they can combine their proliferation and differentiation functions in the same neuronal linage is still unclear (Mora, 2018).
This study has identified the first symmetrically self-amplifying NSCs in Drosophila giving rise to the population of T4/T5 neurons. These Nbs are generated by a temporal conversion of asymmetrically dividing Nbs, which is accompanied by a temporal transition in proneural protein expression from the AS protein Asense (Ase) to the ATH protein Atonal (Ato). Furthermore, it was discovered that the switch from Ase to Ato is necessary and sufficient for the switch in stem cell division pattern and the generation of the correct number of neurons. Lastly, it was demonstrated that Atonal creates a quantitative change in target gene expression that is propagated throughout the lineage to ensure the commitment of T4/T5 neurons to terminal differentiation (Mora, 2018).
This is the first example of transient amplification by symmetric division of NSCs in a non-mammalian animal, namely the Drosophila fruit fly. These cells are termed type III Nbs to distinguish them from previously described Nb types 0, I, and II. Embryonic NSC symmetric expansion is common in mammals, especially in gyrencephalic mammals, where oRG are highly abundant. This includes ferrets, non-human primates, and humans, but not rodents. oRG is thought to be in part responsible for the brain size expansion that is observed in these species. Having simpler models that recapitulate at least some aspects of oRG biology could be particularly relevant to the study of fundamental questions surrounding the control of brain size. Understanding the symmetry of self-renewal is also relevant for the study of adult neurogenesis where symmetric division have recently been shown to be predominant. In this context, limited rounds of symmetric self-renewal and consuming symmetric differentiation division can explain how neurogenesis is sustained for extended periods of time. This work finds that Drosophila symmetrically amplifying Nbs expand the progenitor pool while at the same time scheduling the future terminal differentiation of their progeny. This study describes genetic and regulatory control mechanisms of these features and the consequences of interfering with such mechanisms for brain development (Mora, 2018).
The type III Drosophila Nbs described in this study are located in the visual system anlagen in a region known as the IPC, where they generate two different neuronal populations: the C2, C3, T2, and T3 neurons (C/T neurons) and the T4/T5 neurons, in that specific temporal order. IPC Nbs transit through two distinct types of proliferation: an earlier phase of type I asymmetric divisions to generate C/T neurons and a later phase of symmetric transit amplification. These two phases coincide with a change in neuronal fate and number. While late born T4/T5 constitute one of the largest lineages in the Drosophila brain, the early born C/T neurons are two times less abundant. Although it is difficult to know the exact number of symmetric divisions each upper-Nb undergoes, it is interesting to note that one symmetric amplification before the terminal production of GMCs would account for the doubling of the number of upper-Nbs compared with lower-Nbs that were observed, resulting in exactly four T4/T5 neurons per upper-Nb. A concurrent study (Pinto-Teixeira, 2018) proposes that this particular stoichiometry may be accounted for through a single terminal Nb division. While the current observations do not contradict the stoichiometry, the suggestion that there is no amplifying step prior to the terminal division is difficult to reconcile with the multiple lines of evidence presented in this study. Together with a study by Apitz (2018), these authors further show that the layer specificity of T4/T5 neurons relies on Dpp and that the T4 versus T5 fate is Notch dependent. The Apitz study further shows how the Dpp signal is maintained from NE progenitors to neurons through a temporal relay mechanism. Together, these studies open the door for understanding precisely how this very large and complex lineage combines numerical expansion, cell fate, and layer-specific targeting over a series of successive temporal developmental transitions (Zhang, 2018).
This study focuses on the temporal transition of proliferation properties and shows that they are regulated by the serial expression of two proneural proteins, Ase and Ato. Interestingly Ase and Ato had not been involved in C/T versus T4/T5 fate decision, suggesting that lineage size can be controlled independently of cell fate. Previous studies in the IPC have shown that the switch in neuronal fate depends on another temporal series of two factors called Tailless and Dichaete. It would be interesting to investigate the crosstalk between these two temporal series as a model to further understand how neuronal numbers and neuronal fate are integrated during development. The current findings provide one of the first examples of Nbs changing their proliferation properties to achieve lineage size proportions, where NSC amplification is causally linked to an increase in the number of neurons generated (Mora, 2018).
It is important to note that, in the IPC, Ato can robustly impose symmetric division when ectopically expressed. However, only a fraction of Nbs divisions are affected in its absence. This demonstrates that Ato is sufficient, but not always necessary, for symmetric division, and suggests the existence of an overlapping and independent mechanism controlling the process. Ato in this context likely acts to ensure robust transitions first to symmetric amplification and later to differentiation. It is proposed that the strong reduction of T4/T5 neuron numbers in ato mutant brains is due to an incomplete transition from asymmetric to symmetric division. However, the effect of other functions of Ato yet to be characterized, for example in ensuring neuronal survival, cannot be excluded (Mora, 2018).
The fact that Ato expression in Nbs controls the differentiation of T4/T5 neurons is demonstrated by the ectopic expression of Nbs markers and the global downregulation of differentiation genes in neurons of ato mutant animals. This resembles the de-differentiation phenotype previously found in Drosophila mutants of longitudinal lacking (lola). However, unlike lola, Ato itself is never expressed in neurons, not even transiently. It is proposed that a stable cellular memory of differentiation is initiated transcriptionally in stem cells and inherited through successive cell divisions to ensure terminal differentiation of neuronal progeny. What the mechanisms of such a memory are, how they are activated in stem cells, and how they relate to stem cell division mode are exciting questions for future investigation (Mora, 2018).
A recurring observation throughout this analyses is that quantitative, rather than all or nothing, changes in gene expression downstream of Ato control the temporal progression of developmental events. For example, premature Ato expression causes a relatively modest reduction in Ase expression, and yet suffices to induce symmetric division prematurely. Similarly, quantitative regulation of Brat levels is required for a dose-dependent maintenance of terminal differentiation in postmitotic neurons. Brat is a member of a family of evolutionarily conserved tumor suppressor proteins that regulate differentiation and growth. In type I and II Nbs, Brat is asymmetrically inherited to promote differentiation. In IPC Nbs, Brat is symmetrically inherited during the transient amplification but it does not prevent Nb gene expression. It is therefore proposed that it is the progressive accumulation through temporal quantitative regulation, rather than its expression per se, that schedules the onset and maintenance of differentiation (Mora, 2018).
How cell division and differentiation are coordinated to determine organ size is a fundamentally important but poorly understood process. In Drosophila, the intrinsic activity life time of given proneural transcription factor is both a developmental and evolutionary strategy for the control of cell number in the peripheral nervous system. During the development of mammalian telencephalon, the expression of Ascl1, the mammalian homolog of Drosophila Achaete-scute proteins such as Ase, oscillates in NSCs. These oscillations promote proliferation, while sustained expression of Ascl1 promotes neuronal differentiation. Finally, there is evidence that spatiotemporal transitions in cross-regulatory transcription factors control root meristem growth in plants. This study shows that a similar logic regulates brain size. These observations suggest that the differential, temporally restricted and quantitative regulation of transcription factors and their target genes may serve a universal role as molecular clocks underlying the coordinated temporal order of developmental events (Mora, 2018).
The Drosophila medulla, part of the visual processing center of the brain, contains more than 70 neural types generated by medulla neuroblasts which sequentially express several temporal transcription factors (TTFs), including Homothorax (Hth), eyeless (Ey), Sloppy paired 1 and 2 (Slp), Dichaete (D) and Tailless (Tll). However, it is not clear how such a small number of TTFs could give rise to diverse combinations of neuronal transcription factors that specify a large number of medulla neuron types. This study reports how temporal patterning specifies one neural type, the T1 neuron. The T1 neuron is the only medulla neuron type that expresses the combination of three transcription factors Ocelliless (Oc or Otd), Sox102F and Ets65A. Using CRISPR-Cas9 system, this study shows that each transcription factor is required for the correct morphogenesis of T1 neurons. Interestingly, Oc, Sox102F and Ets65A initiate expression in neurons beginning at different temporal stages and last in a few subsequent temporal stages. Oc expressing neurons are generated in the Ey, Slp and D stages; Sox102F expressing neurons are produced in the Slp and D stages; while Ets65A is expressed in subsets of medulla neurons born in the D and later stages. The TTF Ey, Slp or D is required to initiate the expression of Oc, Sox102F or Ets65A in neurons, respectively. Thus, the neurons expressing all three transcription factors are born in the D stage and become T1 neurons. In neurons where the three transcription factors do not overlap, each of the three transcription factors can act in combination with other neuronal transcription factors to specify different neural fates. This study shows that this way of expression regulation of neuronal transcription factors by temporal patterning can generate more possible combinations of transcription factors in neural progeny to diversify neural fates (Naidu, 2020).
T1 neurons are a class of mysterious neurons that connect the lamina and the medulla part of the optic lobe. They are uni-columnar neurons with one in each of the 800 columns of the medulla. The cell body of the T1 neuron is found in the medulla cortex, and its axon branches in a characteristic 'T' shape on the distal surface of the medulla. One branch projects through the outer optic chiasm back to the lamina and then forms a basket like structure of processes surrounding each lamina cartridge. The other branch arborizes in the M2 layer of the medulla with a dense bush like structure. T1 neuron is post-synaptic to amc (lamina amacrine cells), and the amc/T1 pathway was shown to enhance the lamina neuron L1 motion detection pathway at intermediate contrast. Depolarizing T1 neurons affected the flight steering responses to visual stimuli (Naidu, 2020).
Through screening antibodies and GFP fusion lines, this study found that Ocelliless (oc), Sox102F and Ets65A are expressed in T1 neurons, and the combination of these three TFs can distinguish T1 neurons from all other medulla neurons. Using CRISPR-Cas9 system, bi-allelic somatic mutations of each of the three TF genes were generated in T1 neurons; knock-down of each one affected different aspects of the T1 neuron morphology. Next, how the expression of each TF is controlled by temporal patterning to generate the combination code was examined. Oc expression in neurons starts in the Ey temporal stage, and continues in the Slp and D temporal stages, and Ey is required for the initiation of Oc expression in neurons; while Sox102F expression in neurons starts in the Slp temporal stage, and continues in the D temporal stage, and Slp is required for initiating the expression of Sox102F in neurons; finally, Ets65A is expressed in subsets of medulla neurons born in the D and later temporal stages, and D is required for the expression of Ets65A. Thus, the three TFs that control T1 neuron morphology initiate their expression in neurons beginning at different temporal stages controlled by different TTFs, but each of them spans a few temporal stages, and the neurons expressing all three TFs are born in the D stage and become T1 neurons. In neurons where the three transcription factors do not overlap, each of the three TFs could also act with other neuronal TFs to specify different neural fates. In this way, more combinations of TFs can be generated through temporal patterning (Naidu, 2020).
This study identified a combination of three transcription factors that control T1 neuron morphology, and examined how the expression of these three transcription factors are controlled by temporal patterning of medulla neuroblasts. Oc is turned on in neurons starting in the late Ey stage, and Oc expressing neurons continue to be generated in the Slp and D stages, although the fates will be different, possibly dependent on the co-expression with other neuronal TFs. Sox102F expressing neurons start to be generated in the Slp stage and continue in the D stage. Ets65A expressing neurons are generated in the D and later temporal stages. Thus, the three TFs that control T1 neuron morphology start their expression in neurons born at different temporal stages, and require the corresponding TTF for initiation of their expression, and each neuronal TF is expressed in neurons spanning a few temporal stages. One advantage for such temporal control of neuronal TFs is that more combinations of TFs can be generated to specify different fates. For example, Toy is expressed in the N-on neuronal progeny born from the Slp and D stages, and also in some N-off progeny born from the late Ey stage neuroblasts in some regions of the medulla. Results from this study and others suggest that the subset of Sox102F neurons that do not express Oc, express Toy and Ap instead, and they are specified as Tm5 neurons. In addition, the neurons that express both Toy and Oc in the N-off progeny of some late Ey stage neuroblasts could determine another unknown neural type. Although it remains to be determined whether these TF combinations are indeed required for the corresponding neural fates, these examples do suggest that different combinations of neuronal TFs can be created that might determine different fates (Naidu, 2020).
Mutation of each of the three TFs expressed in T1 caused a certain morphological defect, similar to the morphology TFs that act in combinations to determine motor neuron morphology. For oc
and Ets65A mutant neurons, it appeared that they still maintained the T1 fate, but the morphology was abnormal. Some Sox102F mutant neurons resembled medulla intrinsic neurons, but without functional assay, it was not clear whether they were fully transformed to a normal Mi neuron fate, or they still maintained some T1 neuron charateristics but underwent dramatic morphological changes. One question is whether the combination of TFs regulate neuron morphology by simple addition (each TF determines one feature, and the simple addition of these features determines one neural type), or in a synergistic way (three TFs together can determine features not determined by either TF alone). In the case of T1, when Sox102F was removed from T1 neurons, the driver used (T1-LexA) was still expressed in the mutant neurons, but the neurons became more like medulla intrinsic neurons, and some neurons lost the projection back to the lamina. However, Sox102F is not expressed in other neurons that project back to the lamina like lamina wide field neurons (lawf 1/2) which express Hth and Eya. Instead, Sox102F is also expressed in a Transmedulla neural type (Tm5) which do not resemble T1 neurons. Thus, these results favor the synergistic action model of neuronal TFs to control neuron morphology (Naidu, 2020).
The results are consistent with the principle that integration between temporal/spatial patterning of neuroblasts and the Notch-dependent binary neuron fate choice further diversifies neural fates. This study found that T1 neurons are derived from the Notch-off hemilineage of D stage neuroblasts. In addition, although T1 neurons are uni-columnar neurons that are generated throughout the main medulla region, there is a spatial component that regulates Oc expression and neural fate specification. Neurons that co-express Oc and Forkhead are only localized in the Dpp domains. Through analyzing the sequencing data published for all medulla neurons, the neurons expressing both Oc and Fkh should become the Dm12 neuron, a multi-columnar neuron with arborizations spanning several columns. Thus, these results support the conclusion that uni-columnar neurons are generated throughout the medulla main region, while multi-columnar neurons are generated in special spatial domains determined by spatial patterning (Naidu, 2020).
In summary, this study of T1 neuron specification illustrated an example how temporal patterning of neuroblasts sequentially turns on the expression of three TFs in neuronal progeny, and generates different combinational codes to determine neural fates. In the future it will be interesting to examine how TTFs in neuroblasts regulate the expression of neuronal TFs in neurons that often span a few temporal stages. Only a subset of neurons maintain the expression of TTFs, while other neurons do not. Thus the TTFs should determine the expression of neuronal TFs already in neuroblasts. It is possible that the TTF promotes epigenetic modifications in the neuronal TF gene locus, so that the TF will be turned on in its progeny as well as in neurons born in subsequent temporal stages. It is also possible that the expression of the same neuronal TF in two subsequent temporal stages are controlled by two separate enhancers that respond to different TTFs. Addressing these questions will further advance understanding of the link between neuroblast temporal patterning and neural fate specification (Naidu, 2020).
The brain is consisted of diverse neurons arising from a limited number of neural stem cells. Drosophila neural stem cells called neuroblasts (NBs) produces specific neural lineages of various lineage sizes depending on their location in the brain. In the Drosophila visual processing centre - the optic lobes (OLs), medulla NBs derived from the neuroepithelium (NE) give rise to neurons and glia cells of the medulla cortex. The timing and the mechanisms responsible for the cessation of medulla NBs are so far not known. This study showed that the termination of medulla NBs during early pupal development is determined by the exhaustion of the NE stem cell pool. Hence, altering NE-NB transition during larval neurogenesis disrupts the timely termination of medulla NBs. Medulla NBs terminate neurogenesis via a combination of apoptosis, terminal symmetric division via Prospero, and a switch to gliogenesis via Glial Cell Missing (Gcm); however, these processes occur independently of each other. Temporal progression of the medulla NBs is mostly not required for their termination. As the Drosophila OL shares a similar mode of division with mammalian neurogenesis, understanding when and how these progenitors cease proliferation during development can have important implications for mammalian brain size determination and regulation of its overall function (Nguyen, 2024).
Robust brain development requires the tight coordination between stem cell maintenance and differentiation. In the Drosophila OL, there are two main stem cell pools, the NE cells that divide symmetrically, and the NBs. Medulla NBs are derived from the OPC NE via the proneural wave, which switch to asymmetric division where they self-renew and give rise to GMCs that will produce post-mitotic neurons and glial cells of the medulla cortex. This study shows that during early pupal development, medulla NBs terminate via a combination of mechanisms including apoptosis-mediated cell death, Pros-mediated differentiation and a switch from neurogenesis to gliogenesis. While factors such as ecdysone signalling and OxPhos of the NBs are involved in NB termination in other lineages outside of the OL, this study found that these factors do not play a significant role in the termination of medulla NBs. Instead, the main regulator of the timely NB cessation in the pupal medulla is the timing of the NE-NB conversion during larval stages. Interestingly, while med12 RNAi clones exhibited persistent NBs in the OL at 24 hr APF, this is likely due to the role of mediator protein Med12 in regulating NE-NB transition during larval development (Med12 knockdown in the medulla NBs did not cause persistent NBs). Additionally, while a specific cascade of temporal identity transcription factors in the medulla NBs has been shown to generate the cellular diversity of medulla neurons and glia, temporal progression in the medulla NBs appears to exert negligible effects on the timing of NB cessation in the pupal stages. Diseases such as microcephaly are mediated by the premature depletion of the NE pool, or through a precocious commitment to neurogenic divisions. Consistent with this, the data present evidence that during development, the regulation of the NE stem cell pool, and its timely conversion into NSCs is of critical importance for the overall size and function of the brain (Nguyen, 2024).
The data support a model whereby once the NE pool is exhausted, no new NBs can be generated. NBs transit through their temporal series to terminally differentiate into neurons and glia, which marks the end of medulla neurogenesis. Thus, unlike type I NB lineages in which NB termination is scheduled by ecdysone signalling-mediated temporal progression at the level of the NBs; in the OL, NB termination is pre-determined in the NE. While this work has only tested the role of Notch signalling in the NE for determining medulla NB termination, JAK/STAT signalling was previously demonstrated to control the proneural wave and the associated NE-NB transition, in part through Notch signalling. Therefore, it is speculated that JAK/STAT signalling will also likely influence the timing of medulla NB termination. In the OL, besides medulla NBs, NE cells also sequentially express an array of temporal factors, such as Chinmo and Broad downstream of ecdysone signalling. Moreover, inhibiting the Chinmo-to-Broad expression switch in the NE was shown to delay the NE-NB transition, resulting in tumours with persistent NBs in the adult OLs. Overall, the differences in the regulation of NB termination in the OL medulla versus other lineages may be due to some fundamental differences in the origins of these NBs. In the OL, NBs are derived from the NE, that undergoes rapid growth during larval development. In contrast, NBs from other lineages are produced directly from the neuroectoderm in the embryo without undergoing the transition from the NE. Hence, their termination mechanism acts predominantly in the NBs during pupal development (Nguyen, 2024).
Consistent with previous reports, whis study found that apoptosis occurred in a subset of medulla NBs from 12 hr to 16 hr APF. Furthermore, although apoptosis inhibition caused persistent NBs at 24 hr APF, by 48 hr APF, all NBs were still eliminated in the OL. Notably, half of the NBs undergoing apoptosis at 12 hr APF expressed the mid-temporal factor Ey, but not Slp or Tll. These data suggest an intriguing link between the expression of the mid-temporal factor and the competence of the NBs to undergo apoptosis. It is possible that the early-born medulla NBs that in turn express late temporal factors (Tll or Gcm) during pupal development, will terminate via either Pros-mediated symmetric division or gliogenic switch. Whereas the late-born NBs, that still express mid-temporal transcription factors such as Ey, will terminate via apoptosis. This mechanism could be a fail-safe paradigm to ensure the elimination of all the medulla NBs before adulthood. Whether such decision of the NBs is determined by the intrinsic factors like the temporal series, or extrinsic factors such as signalling cues from other cell types in the niche remain to be probed. Although the timing of apoptosis in the medulla NBs coincides with a small pulse of ecdysone around 10 hr APF, the inhibition of ecdysone signalling in the medulla NBs did not phenocopy the inhibition of apoptosis. Therefore, unlike neurons in the OL which require ecdysone signalling for apoptosis during pupal development, medulla NBs undergo apoptosis independently of ecdysone signalling. Altogether, while some NBs terminate proliferation through apoptosis, it is likely that apoptosis of the NBs makes a minor contribution to the termination of neurogenesis in the medulla (Nguyen, 2024).
Many medulla NBs displayed nuclear Pros expression during early pupal stages. Consistent with the essential role of Pros in promoting differentiation, pros knockdown caused the persistence of the NBs in the OL at 24 hr APF. Nevertheless, later during the pupal stages, pros knockdown was insufficient to maintain medulla NB persistence, suggesting that Pros expression in the medulla NBs is not strictly required for their termination. In addition to Pros-mediated differentiation, another mechanism responsible for the cessation of neurogenesis is the gliogenic switch. Cells triple positive for the pan-NB marker Dpn, the differentiation marker Pros, and the glial cell determinant Gcm in the early pupal medulla, which likely give rise to glial cells at the end of the temporal series. Interestingly, the inhibition of Pros did not affect gliogenesis in the larval medulla; conversely, the inhibition of Gcm did not influence Pros expression in the pupal OL. As such, these data suggest that despite Pros and Gcm co-localisation in the terminating NBs, gliogenesis and differentiation appear to act in parallel. Taken together, apoptosis, symmetric cell division and a gliogenic switch appear to be employed as fail-safe mechanisms that help eliminate the NB progenitor pool in the pupal OL. Hence, it would be interesting to explore in the future the cross-regulatory relationship between these termination programs. (Nguyen, 2024).
Drosophila larval central nervous system comprises the central brain, ventral nerve cord and optic
lobe. In these regions, neuroblasts divide asymmetrically to self-renew and generate differentiated
neurons or glia. To date, mechanisms of preventing neuron dedifferentiation are still unclear,
especially in the optic lobe. This study shows that the zinc finger transcription factor Nerfin-1 is expressed in early stage of medulla neurons and essential for maintaining their
differentiation. Loss of Nerfin-1 activates Notch signaling,
which promotes neuron-to-NB reversion. Repressing Notch signaling largely rescues dedifferentiation
in nerfin-1 mutant clones. Thus, it is concluded that Nerfin-1 represses Notch activity in
medulla neurons and prevents them from dedifferentiation (Xu, 2017).
Stem cells generate progeny that undergo terminal differentiation. In Drosophila CNS, the balance of
self-renewal and differentiation of neural stem and progenitor cells is a central issue during
development. On the other hand, the maintenance of differentiated status of post-mitotic neurons is
also crucial for tissue function and homeostasis. It is obvious that mechanisms must exist to
prevent the cells from dedifferentiation. Although proteins that function to keep differentiation
have been well studied in other cell types, few have been implicated in post-mitotic neuronal maintenance. In the central brain, loss of
Midlife crisis (Mdlc), a CCCH zinc-finger protein, results in a decrease in Pros, thus derepressing
NB genes in neurons. However, it is insufficient to make neurons revert to
proliferating NBs. Furthermore, as Pros is not expressed in medulla neurons, it is unclear whether
Mdlc has the same function in the optic lobe. On the other hand, absence of Lola leads to
neuron-to-NB reversion and tumorigenesis , but it is crucial for neuronal
maintenance only in the optic lobe. Recently, a paper reported that Nerfin-1 loss induces neuron
dedifferentiation in both central brain and VNC (Froldi, 2015). This paper
demonstrates a conserved function for Nerfin-1 in medulla neurons in the optic lobe. These findings
indicate that Nerfin-1 is expressed mainly in early-stage medulla neurons and functions to maintain
their differentiated state (Xu, 2017).
Interestingly, it was noticed that ectopic NB induced by Nerfin-1 depletion in the optic lobe appeared
much earlier than that in the central brain. Considering that Lola loss causes dedifferentiation
just in the optic lobe, it is speculated that the differentiated state of
medulla neurons is less stable, possibly owing to absence of Pros. Furthermore, different from the
mechanism in the central brain, the function of Nerfin-1 in the optic lobe requires the silencing of
Notch signaling. Neither Myc knockdown nor Tor-DN misexpression inhibits dedifferentiation caused by
Nerfin-1 loss in the medulla neurons. Thus, these findings identify a distinct regulatory
mechanism in medulla neurons and validate different regulatory modes between the optic lobe and the
rest of the CNS (Xu, 2017).
On the other hand, cell cycle genes play important roles in cell differentiation. Among them, Cyclin
E (CycE) is reported to be regulated directly by Lola-N and is involved in
the neuron dedifferentiation caused by loss of Mdlc. Thus, this study also examined
whether CycE is regulated directly by Nerfin-1 and controls cell differentiation independently of
Notch and neuroblast genes. Interestingly, CycE expression levels were upregulated dramatically in
nerfin-1159 clones, but such upregulation was mostly blocked by Notch repression.
These results suggest that CycE is not a direct target of Nerfin-1 for maintaining medulla neuron
differentiation. CycE acts downstream of Notch signaling or it is subsequently upregulated after
cell type change (Xu, 2017).
As Notch signaling is hyper-activated in nerfin-1 mutant clones, it was of interest to discover how it is
regulated. One possibility is that Notch signaling becomes constitutively activated without the
inhibition by Nerfin-1. To investigate this, Delta was knocked down upon Nerfin-1 loss and it was found that
dedifferentiation was suppressed. These results indicate that Notch signaling is not
constitutively activated and that it needs a ligand. Furthermore, Notch signal is both produced and
received by medulla neurons. At the same time, the results show that Nerfin-1 loss induces dramatic
upregulation of the expression level of Notch receptor. Thus, it is hypothesized that
Nerfin-1 suppresses the expression of the Notch receptor in normal medulla neurons and inhibits
Notch pathway activity. When Nerfin-1 is absent, expression levels of the Notch receptor increase
strikingly. The receptors then bind to Delta from the adjacent cells and activate Notch signaling in
its own. However, it is still unclear whether Notch receptor is a direct target of Nerfin-1.
Therefore, subsequent studies on Nerfin-1 may help to clarify the underlying mechanisms and
provide better understanding about neuronal maintenance (Xu, 2017).
Brain areas each generate specific neuron subtypes during development. However, underlying regional variations in neurogenesis strategies and regulatory mechanisms remain poorly understood. In Drosophila, neurons in four optic lobe ganglia originate from two neuroepithelia, the outer (OPC) and inner (IPC) proliferation centers. Using genetic manipulations, this study found that one IPC neuroepithelial domain progressively transformed into migratory progenitors that matured into neural stem cells (neuroblasts) in a second domain. Progenitors emerged by an epithelial-mesenchymal transition-like mechanism that required the Snail-family member Escargot and, in subdomains, Decapentaplegic signaling. The proneural factors Lethal of scute and Asense differentially controlled progenitor supply and maturation into neuroblasts. These switched expression from Asense to a third proneural protein, Atonal. Dichaete and Tailless mediated this transition, which was essential for generating two neuron populations at defined positions. It is proposed that this neurogenesis mode is central for setting up a new proliferative zone to facilitate spatio-temporal matching of neurogenesis and connectivity across ganglia. (Apitz, 2014).
Recent studies have distinguished three neurogenesis modes in the Drosophila CNS. First, type I neuroblasts arise from neuroepithelia and generate GMCs, which produce neuronal and glial progeny. Second, Dpn+ type II neuroblasts in the dorsomedial central brain go through a transit-amplifying Dpn+, Ase+ population, called intermediate neural precursors, which generate GMCs and postmitotic offspring. Third, lateral OPC neuroepithelial cells bypass the neuroblast stage and generate lamina precursor cells (LPCs) that divide once to produce lamina neurons. The current results provide evidence for a fourth strategy: p-IPC neuroepithelial cells give rise to progenitors that migrate to a second neurogenic domain, where they mature into type I neuroblasts. These progenitors are distinct, as they originate from the neuroepithelium, do not express markers for neuroblasts, intermediate neural precursors, GMCs or postmitotic neurons, and acquire NSC properties after completing their migration (Apitz, 2014).
Migratory progenitors arise from the p-IPC by a mechanism that shares cellular and molecular characteristics with EMT. On the basis of data on gastrulation and neural crest formation, EMT is commonly associated with cells adopting a mesenchymal state, enabling them to leave their epithelial tissue and migrate through the extracellular matrix to new locations. A recent study also reported an EMT-like process in the mammalian neocortex, whereby newborn neurons and intermediate progenitors delaminate from the ventricular neuroepithelium and radially migrate to the pial surface. This study observed that neuroepithelial cells at the p-IPC margins and migratory progenitors upregulated the Snail homolog Esg, whereas E-cad levels were decreased. Moreover, esg knockdown caused the formation of ectopic E-cad-expressing clusters adjacent to the p-IPC. Although this is a previously uncharacterized role of Drosophila esg, these findings are consistent with the requirement of two Snail transcription factors, Scratch1 and 2, and downregulation of E-cad in cortical EMT migration (Apitz, 2014).
Although TGFβ signaling is well known to induce EMT, it was unclear whether it could have such a role in the brain. Two lines of evidence are consistent with a requirement of the Drosophila family member Dpp. First, it is expressed and downstream signaling is activated in dorsal and ventral p-IPC subdomains and emerging cell streams. Second, tkv mutant cells form small neuroepithelial clusters in p-IPC vicinity. Similar to the neural crest, where distinct molecular cascades control delamination in the head and trunk, region-specific regulators may also be required in p-IPC subdomains. Because neuroblasts derived from Dpp-dependent cell streams map to defined areas in the d-IPC, this pathway could potentially couple EMT and neuron subtype specification (Apitz, 2014).
Cell migration is an essential feature of vertebrate brain development. Commonly, postmitotic immature neurons migrate from their proliferation zones to distant regions, where they further differentiate and integrate into local circuits. Examples include the radial migration of projection neurons and tangential migration of interneurons in the embryonic cortex, as well as migration of interneuron precursors in the rostral migratory stream to the olfactory bulb in adults. In contrast, IPC progenitors develop into NSCs (neuroblasts) after they migrated. A recent study found that NSCs relocating from the embryonic ventral hippocampus to the dentate gyrus act as source for adult NSCs in the subgranular zone. In addition, cerebellar granule cell precursors migrate from the rhombic lip to the external granule layer, where they proliferate during early postnatal development. The migration of neural cell types that become proliferative in a new niche could therefore constitute a more general strategy. IPC progenitors form streams of elongated, closely associated cells. Despite their different developmental state, their organization is notably similar to the neuronal chain network in the lateral walls of the subventricular zone and the rostral migratory stream in mammals, or of migratory trunk neural crest cells in chick. Further studies will need to identify the determinants directing migratory progenitors into the d-IPC (Apitz, 2014).
Several constraints could shape a neurogenesis mode that requires migratory progenitors in the larval optic lobe. The OPC is located superficially and the IPC is positioned centrally. If medulla and lobula neurons arose by neuroepithelial duplications, these new populations would need to be integrated into an ancestral visual circuit consisting of lamina and lobula plate neurons. Cellular migration may therefore be a derived feature and serve as an essential spatial adjustment of the IPC to the newly added medulla. In principle, the migratory population could consist of immature neurons. However, migratory progenitors help to establish a new superficial proliferative niche, and to align OPC and d-IPC neuroblast positions. This in turn enables the OPC and IPC to use spatially matching birth order-driven neurogenesis patterns for establishing functionally coherent connections across ganglia (Apitz, 2014).
IPC progenitors were primed to mature into neuroblasts, but were prevented to do so in cell streams. Consistently, progenitors showed weak cytoplasmic Mira expression and prematurely differentiated into neuroblasts following loss of Pcl. Although Dichaete has been shown to repress ase to maintain embryonic neuroectodermal cells in an undifferentiated state, this study did not identify such a role in the IPC. Future studies are therefore required to distinguish whether this block in neuroblast maturation is released in the d-IPC by cell-intrinsic mechanisms or locally acting signals (Apitz, 2014).
The p-IPC and d-IPC consecutively expressed three proneural factors. esg-positive p-IPC neuroepithelial cells transiently expressed L'sc as they converted into progenitors. Following arrival in the d-IPC, progenitors matured into neuroblasts, which switched bHLH protein expression from Ase to Ato. This correlated with a change in cell division orientations from toward the lamina to the optic lobe surface and the generation of two lineages, distal cells and lobula plate neurons. The progression of neuroblasts through two stages is supported by the observations that progenitors solely entered the lower d-IPC, all neuroblasts were labeled with Ase in this area, and idpp reporter gene expression in a progenitor subset persisted in both lower and upper d-IPC neuroblasts and their progeny (Apitz, 2014).
Late l'sc knockdown reduced the number of d-IPC neuroblasts and both neuron classes, whereas p-IPC formation and EMT of progenitors appeared to be unaffected. This supports the idea that l'sc promotes neuroblast formation by controlling the rate of conversion and the progenitor supply. In contrast, ase loss severely decreased the amount of lower d-IPC neuroblasts and distal cells. This revealed a central role in the maturation of progenitors into neuroblasts, endowing them with the potential to proliferate and generate a specific lineage. Although these functions are the opposite of those observed in the OPC, they align with the role of a murine Ase homolog, Achaete-scute homolog 1 (Ascl1), in the embryonic telencephalon. Ase- neuroblasts with type I proliferation patterns have not previously been described. Further underscoring the context-dependent activities of proneural bHLH factors, ato does not have the equivalent role of ase in conferring neurogenic properties to upper d-IPC neuroblasts, but acts upstream of differentiation programs controlling the projections of lobula plate neurons (Apitz, 2014).
Although Ase and Ato each regulated distinct aspects of d-IPC development, they were not required for either the transition or the extent of their expression domains. These functions were fulfilled by Dichaete and tll, whose cross-regulatory interactions were essential for the transition from Ase+ to Ato+, Dac+ expression. To link birth order and fate, temporal identity transcription factors are sequentially expressed by neuroblasts and inherited by GMCs and their progeny born during a given developmental window. Acting as the final two members of the OPC-specific series of temporal identity factors, Dichaete is required for Tll expression, whereas tll is sufficient, but not required, to inhibit Dichaete Although OPC and d-IPC neuroblasts shared the sequential expression of Dichaete and Tll, key differences include the fact that d-IPC progeny did not maintain Dichaete, that Tll was transiently expressed in newborn progeny of the upper d-IPC and was not maintained in older lineages, that Dichaete in the lower d-IPC was not required in its own expression domain for neurogenesis, and that Dichaete was required to activate tll, and tll to repress Dichaete and ase, as well as to independently upregulate Ato and Dac. Although the mechanisms that trigger the timing of the switch require further analysis, these observations support the notion that, in the d-IPC, Dichaete and tll do not function as temporal identity factors, but as switching factors between two sequential neuroblast stages. The vertebrate homologs of Dichaete and tll, Sox2 and Tlx, are essential for adult NSC maintenance and Sox2 positively regulates Tlx expression, suiggesting that core regulatory interactions between Dichaete and tll family members may be conserved (Apitz, 2014).
These studies uncovered molecular signatures for generating a migratory neural population by EMT and subsequent NSC development that are in part shared between the fly optic lobe and vertebrate cortical neurogenesis. The unexpected parallels suggest that ancestral gene regulatory cassettes imparting specific cellular properties may have been re-employed during vertebrate brain development. Analysis of p-IPC and d-IPC neurogenesis in the Drosophila optic lobe therefore opens new possibilities for systematically identifying genes regulating EMT, cell migration and sequential NSC specification (Apitz, 2014).
The Drosophila optic lobe comprises a wide variety of neurons forming laminar and columnar structures similar to the mammalian brain. The Drosophila optic lobe may provide an excellent model to investigate various processes of brain development. However, it is poorly understood how neuronal specification is regulated in the optic lobe to form a complicated structure. This study shows that the Brain-specific homeobox (Bsh) protein, which is expressed in the lamina and medulla ganglia, is involved in specifying neuronal identity. Bsh is expressed in L4 and L5 lamina neurons and in Mi1 medulla neurons. Analyses of loss-of-function and gain-of-function clones suggest that Bsh is required and largely sufficient for Mi1 specification in the medulla and L4 specification in the lamina. Additionally, Bsh is at least required for L5 specification. In the absence of Bsh, L5 is transformed into glial cells (Hasegawa, 2013).
The establishment of precise neuronal circuits is essential for correct brain function. Complex neuronal circuits contain various types of neurons that are connected intricately with one another. Processes that result in the formation of the correct circuits include the specification of neuronal types, the extension of axons to the appropriate places, and the formation of synapses with their correct partners. The specification of neuronal types is an important process in the making of complete neuronal circuits (Hasegawa, 2013).
The Drosophila visual system may serve as a powerful model for neuronal circuit formation because it has only a limited number of neurons but forms sufficiently complex neuronal circuits that can be analyzed comprehensively. In addition, neurogenetic tools that are available in Drosophila allow artificial manipulation of neuronal activity in temporal ly and spatially restricted manner. However, the full picture of development of the Drosophila optic lobe awaits further investigation (Hasegawa, 2013).
The Drosophila retina is composed of 750-800 ommatidia that contain eight types of photoreceptor neurons, denoted as R1-R8. The visual information received in the retina is transmitted to the optic lobe, which is composed of four ganglia; the lamina, medulla, lobula and lobula plate. The complex neuronal circuits in the visual center process various types of visual information, such as motion, color and shape. This paper focuses on the development of the lamina and medulla (Hasegawa, 2013).
The development of the lamina has been studied in some detail. During the third instar larva, photoreceptor neurons extend retinal axons (R axons) to the optic lobe and deliver the inductive signal Hedgehog (Hh) to the lamina precursor cells (LPCs). LPCs divide and become lamina neurons and activate the expression of Dachshund (Dac) and EGF receptor. Spitz, an EGF ligand, is delivered by R axons, received by EGFR and promotes further differentiation of lamina neurons, including the expression of Elav and the formation of lamina columns. Finally, the five types of lamina neurons, L1-L5, become closely associated with R1-R8 axons, forming a lamina cartridge. Although the differentiation of lamina neurons is understood to some extent, how the distinction among L1-L5 neurons is regulated remains unclear (Hasegawa, 2013).
The second visual ganglion, the medulla, contains 40,000 neurons forming tangentially oriented stratifications, which are defined as 10 layers. Medulla neurons are classified by their pattern of arborizations. Some neurons arborize only in the medulla (medulla intrinsic neurons, Mi-neurons), and some send projections back to the lamina (lamina wide-field neurons, Lawf-neurons). Other neurons arborize in both the medulla and the lobula (transmedullary neurons, Tm-neurons) and in the lobula complex (transmedullary Y neurons, TmY neurons). The medulla is the largest part of the optic lobe and is thought to process both color and motion. Although the medulla is considered to play an important role in visual processing, the developmental mechanisms of the medulla remain elusive. During the third instar larva, neuroblasts (NBs) located in the outer proliferation center divide to make ganglion mother cells (GMCs), which divide to produce differentiated neurons. Expression of specific transcription factors in a subset of medulla neurons was examined (Hasegawa, 2011; Morante, 2008). It has been reported previously that neurons produced from NBs express different types of transcription factors according to their birth order to form a concentric expression pattern (Hasegawa, 2011). However, how the differentiation of the medulla neurons is controlled is still unclear (Hasegawa, 2013).
The Bsx family transcription factors are widely conserved homeodomain proteins that are involved in various neuronal processes. For example, mouse Bsx regulates hyperphagia, locomotory behavior, growth, and nursing. Xenopus Bsx links daily cell cycle rhythms to pineal photoreceptors. The Drosophila Bsx protein, Brain-specific homeobox (Bsh), is expressed in the embryonic brain , and in the lamina and medulla neurons of larvae and adults. However, the molecular function of Drosophila bsh has not been studied (Hasegawa, 2013).
A previous paper showed that Bsh is expressed in the medulla and that Bsh-positive neurons differentiate into a single type of medulla intrinsic neuron, Mi1. Moreover, Hth, which is expressed in Mi1 together with Bsh, is essential for Mi1 neuron identity (Hasegawa, 2011). This study shows that Bsh is also required for Mi1 neuron specification in the medulla. bsh mutant neurons were transformed to Tm-type neurons, and overexpression of Bsh induced Mi1-like neurons. Moreover, Bsh expression was required for L4 neuronal specification in the L4 neurons of the lamina, and overexpression of Bsh in the lamina induced L4-like neurons. Therefore, Bsh may have roles in neuronal type specification in both the lamina and the medulla. Relatively weak Bsh expression found in L5 lamina neurons may be required for neuronal differentiation of L5. In the absence of bsh, L5 cells were transformed into glial cells (Hasegawa, 2013).
The previous study showed that Hth and Bsh are expressed in a concentric manner in the larval medulla. Bsh is expressed in the outer half of the Hth domain, and these Bsh/Hth double-positive neurons differentiate into a single neuronal type, Mi1 (Hasegawa, 2011). bsh is predicted to encode two isoforms of homeodomain proteins, long Bsh-PB and short Bsh-PA lacking the N-terminal domain of Bsh-PB The homeodomain is located in the C-terminal region that is shared by both isoforms. No conserved motifs are found in the N-terminal region, which is deleted in Bsh-PA (Hasegawa, 2013).
This study has shown that Bsh is expressed in Mi1 neurons, which differentiates into Tm-type neurons in bsh mutant clones. A similar neuronal transformation from local interneuron to projection neuron is observed in hth mutant clones (Hasegawa, 2011). The number of Hth positive cells was decreased in bsh homozygous animals. However, Hth expression was not affected in bsh mutant clones at larval and adult stages, suggesting that the neuronal type change observed in bsh mutants may not have been caused by reduced expression of Hth. The previous paper showed that residual Bsh is still observed in transformed hth mutant neurons. Moreover, expressing UAS-bshPB could not rescue the defect of the hth mutation, suggesting that Bsh alone cannot induce Mi1 neuron identity and both Hth and Bsh are required for Mi1 neuron identity. By contrast, Bsh expression under the control of drf-Gal4 induced Mi1-like neurons. These results seem inconsistent, however, Mi1-like neurons induced by ectopic Bsh expression were still abnormal compared to endogenous Mi1. Expressing Bsh together with Hth induced Mi1-like neurons that were more similar to endogenous Mi1, implying that both Hth and Bsh are required to generate a complete Mi1 neuron (Hasegawa, 2013).
Because Bsh is a homeodomain protein, the neuronal transformation observed in bsh mutants may be a homeotic transformation. Similar neuronal transformation is observed in mouse Hox gene mutants. However, the transformations observed in bsh mutant clones are striking compared to those observed in mouse Hox mutants. It is not yet known what happens downstream of Bsh and what kind of genes are expressed in transformed Tm-type neurons. It may be that Bsh represses the expression of unknown transcription factors that specify Tm-type neurons. Identification of such transcription factors will lead to insights into the mechanism of neuronal transformation found in bsh mutant (Hasegawa, 2013).
Bsh is expressed in L4 and L5 neurons. The results suggest that bsh mutation transforms L4 neurons into L3-like neurons, and that ectopic Bsh expression induces L4-like neurons, suggesting that Bsh is necessary and sufficient for L4 neuron specification. Although induction and development of lamina neurons are understood to some extent, almost nothing is known about neuronal type specification in the lamina. It is possible to speculate that Ap acts downstream of Bsh. However, Ap alone could not induce L4-like neurons (data not shown), suggesting that Ap is not sufficient for L4 neuron formation. Ap may cooperate with Bsh and/or other factors to specify L4 neuron identity (Hasegawa, 2013).
In bsh mutant lamina neurons, transformation into L3-like neurons was observed most often. It is not known whether this transformation is specifically oriented to L3. Other mutations that transform, for example, L1 neuron into other neurons may reveal whether L3 is the ground state of lamina neurons. At third instar, L1-L5 all express Dac, but during metamorphosis Dac expression disappears in L2 and is reduced in L5. It will be interesting to determine the mechanism that regulates the changes in Dac expression. The combination of Dac, Bsh, and Ap expression can specify L2, L4 and L5 but cannot distinguish between L1 and L3. Therefore, there must be other transcription factors that distinguish between L1 and L3 (Hasegawa, 2013).
Using L5-Gal4, it was unexpectedly found that L5 neurons are transformed to glial cells in bsh mutant clones. The transformed glial cells are found along the endogenous glial cell layers that are situated adjacent to the L5 cells in adult and pupal lamina. They are located in the lamina cortex and ensheath lamina neuron cell bodies, and are closely positioned to L5, suggesting that they are most likely proximal satellite glial cells. Transformation of L5 to glia may suggest that their developmental mechanisms are coupled to each other. It will be interesting to see if there is developmental relationship between L5 lamina neurons and proximal satellite glial cells in the developing optic lobe (Hasegawa, 2013).
Bsx regulates hyperphagia and locomotory behavior by regulating the expression of Npy and Agrp in the arcuate nucleus of the mouse hypothalamus. Bsx expression is repressed by repressor element silencing transcription factor (REST) in non-neuronal cells. Xenopus Bsx links daily cell cycle rhythms with pineal photoreceptors, but its downstream targets are not known. Xbsx expression peaks during the night and represses proliferation. Xbsx knockdown prevents the cell cycle exit of photoreceptor precursors that eventually undergo apoptosis. Xbsx overexpression increases the cell cycle exit of photoreceptor precursors and promotes their differentiation. Therefore, Bsx family proteins may have a general role in cell fate determination. If Bsh target genes are studied in Drosophila, insights into the molecular function of Bsh family proteins may be obtained in the future (Hasegawa, 2013).
These results show that Bsh is expressed in the Mi1 medulla neuron and essential for Mi1 neuron identity. Overexpression of Bsh induces Mi1-like neurons that show some of the features of the Mi1 neuron. In the lamina, Bsh is expressed in the L4 and L5 neurons and is required for L4 and L5 neuron specification. Overexpression of Bsh can induce L4-like neurons. Bsh may have roles in specifying neuronal identity in both the lamina and the medulla (Hasegawa, 2013).
The brain consists of various types of neurons that are generated from neural stem cells; however, the mechanisms underlying neuronal diversity remain uncertain. A recent study demonstrated that the medulla, the largest component of the Drosophila optic lobe, is a suitable model system for brain development because it shares structural features with the mammalian brain and consists of a moderate number and various types of neurons. The concentric zones in the medulla primordium that are characterized by the expression of four transcription factors, including Homothorax (Hth), Brain-specific homeobox (Bsh), Runt (Run) and Drifter (Drf/Vvl), correspond to types of medulla neurons. This study examined the mechanisms that temporally determine the neuronal types in the medulla primordium. For this purpose, transcription factors were sought that are transiently expressed in a subset of medulla neuroblasts (NBs, neuronal stem cell-like neural precursor cells) and identified five candidates [Hth, Klumpfuss (Klu), Eyeless (Ey), Sloppy paired (Slp) and Dichaete (D)]. The results of genetic experiments at least explain the temporal transition of the transcription factor expression in NBs in the order of Ey, Slp and D. The results also suggest that expression of Hth, Klu and Ey in NBs trigger the production of Hth/Bsh-, Run- and Drf-positive neurons, respectively. These results suggest that medulla neuron types are specified in a birth order-dependent manner by the action of temporal transcription factors that are sequential ly expressed in NBs (Suzuki, 2013).
In the embryonic central nervous system, the heterochronic
transcription factors suchas Hb, Kr, Pdm, Cas and Grh are
expressed in NBs to regulate the temporal specification of
neuronal identity. They regulate each
other to achieve sequential changes in their expression in NBs without cell-extrinsic
factors. However, expression of the embryonic heterochronic genes was not detected in the medulla
NBs.Instead this study found that Hth, Klu, Ey, Slp and D are
transiently and sequentially expressed in medulla NBs. The
expression of Hth and Klu was observed in lateral NBs, while that
of Ey/Slp and D was observed in intermediate and medial NBs,
respectively. These observations suggest that the expression of heterochronic transcription factors changes sequentially as
each NB ages, as observed in the development of the embryonic
central nervous system (Suzuki, 2013).
This study demonstrates that at least three of the
temporal factors Ey, Slp and D regulate each other to form a
genetic cascade that ensures the transition from Ey expression to D
expression in the medulla NBs. Ey expression in NBs
activates Slp, while Slp inactivates Ey expression. Similarly, Slp
expression in NBs activates D expression, while D inactivates Slp
expression. In fact, the expression of Slp is not strong in newer NBs in which Ey is strongly expressed, but is up regulated in older NBs
n which Ey is weakly expressed in the wildtype medulla. A
similar relationship is found between Slp and D, supporting the
idea that Ey, Slp and D regulate each other's expression to control
the transition from Ey-expression to D-expression. In the
embryonic central nervous system, similar interaction is mainly
observed between adjacent genes of the cascade hb-Kr-pdm-cas-grh, and this concept may also be applied to the medulla primordium. The expression pattern and function of Ey, Slp
and D suggest that they are adjacent to each other in the cascade of
transcription factor expression in medulla NBs (Suzuki, 2013).
However, no such relationship was found between Hth, Klu and
the other temporal factors.The sequential expression of Hth and
Klu could be regulated by an unidentified mechanism that is
totally different from the genetic cascade that controls the transition through Ey-Slp-D. Or, there might be unidentified temporal factors that are expressed in lateral NBs which act upstream of Hth
and Klu to regulate their expression. It is necessary to identify additional transcription factors that are transiently expressed in medulla NBs (Suzuki, 2013).
The expression of concentric transcription factors in the
medulla neurons correlates with the temporal sequence of neuron
production from the medulla NBs (Hasegawa, 2011). In the
larval medulla primordium, the neurons are located in the order of
Hth/Bsh-, Run- and Drf-positive cells from inside to outside, and
these domains are adjacent to each other (Hasegawa, 2011).
Given that NBs generate neurons toward the center of the developing medulla, Hth/Bsh-positive neurons are produced at first, and
then Run-positive and Drf-positive neurons. Thus Hth/Bsh, Run and Drf were used as markers to examine roles of Hth, Klu, Ey, Slp and D expressed in NBs in specifying types of medulla neurons.
The continuous expression of Hth and Ey from NBs to neurons
and the results of clonal analyses that visualize the progeny of NBs
expressing each one of the temporal transcription factors suggest
that the temporal windows of NBs expressing Hth, Klu and Ey approximately correspond to the production of Hth/Bsh-, Run- and
Drf- positive neurons, respectively. Indeed, the results of
the genetic study suggest that Hth and Ey
re necessary and sufficient to induce the production of Hth/Bsh-
and Drf-positive neurons,respectively (Hasegawa, 2011,
2013). Ectopic Klu expression at least induces the produc tion of Run-positive neurons (Suzuki, 2013).
Slp and D expression in NBs may correspond to the temporal
windows that produce medulla neurons in the outer domains of
the concentric zones, which are most likely produced after the
production of Drf-positive neurons. The results at least
suggest that Slp is necessary and sufficient and D is sufficient to
repress the production of Drf-positive neurons.
Identification of additional markers that are expressed in the outer
concentric zones compared to the Drf-positive domain would be
needed to elucidate the roles of Slp and D in specification of
medulla neuron types (Suzuki, 2013).
D mutant clones did not produce any significant phenotype
except for derepression of Slp expression in NBs. Drf
expression in neurons was not affected either. Since D is a
Sox family transcription factor, SoxN, another Sox family transcription factor, is a potential candidate molecule that acts together
with D in the medulla NBs. However, its
expression was found in neuroepithelia cells and lateral NBs that overlap with
Hth-positive cells but not with D-positive cells.
All the potential heterochronic transcription factors examined
in this study are expressed in three to five cell rows of NBs.
Nevertheless, one NB has been observed to produce one Bsh-
positive and one Run-positive neuron (Hasegawa, 2011).
Therefore, the expression pattern of the heterochronic transcription factors is not sufficient to explain the stable production of one
Bsh-positive and one Run-positive neuron from a single NB.The
combinatorial action of multiple temporal factors expressed in NBs
may play important roles in the specification of Bsh- and Run-
positive neurons (Suzuki, 2013).
Another possible mechanism that guarantees the production of
a limited number of the same neuronal type from multiple rows of
NBs expressing a temporal transcription factor could be a mutual
repression between concentric transcription factors expressed in
medulla neurons. For example, Hth/Bsh, Run and Drf may repress
each other to restrict the number of neurons that express either of
these transcription factors. However, expression of Run and Drf
was not essentially affected in hth mutant clones and in clones
expressing Hth (Hasegawa, 2011). Similarly, expression of
Hth and Drf was not essentially affected in clones expressing run
RNAi under the control of AyGal4, in which Run expression is
eliminated. Hth and Run expression was not affected in drf mutant clones
(Hasegawa, 2011). These results suggest that Hth/Bsh, Run and Drf do not essentially regulate each other during the formation of concentric zones in the medulla (Suzuki, 2013).
During embryonic development, the heterochronic genes
that are expressed in NBs (hb-Kr-pdm-cas-grh) are maintained and
act in GMCs to specify neuronal type. Similarly, Hth and Ey are continuously
expressed from NBs to neurons, suggesting that their expression
may also be inherited through GMCs (Hasegawa, 2011).
However, this type of regulatory mechanism may be somewhat
modified in the case of Klu, Slp and D (Suzuki, 2013).
Klu is expressed in NBs and GMCs, but not in neurons.
Slp and D are predominantly detected in NBs and neurons
visualized by Dpn and Elav, respectively. Occasionally, however, expression of D was found in putative GMCs, which are situated between NBs and neurons. Additionally, both D-positive and D-negative cells were found among Miranda-positive GMCs. Slp expression was not found
in Miranda-positive GMCs. Finally, D is expressed in
medulla neurons forming a concentric zone in addition to its
expression in medial NBs. However, D expression was abolished in
slp mutant NBs but remained in the mutant neurons,
suggesting that D expression in medulla neurons is not inherited
from the NBs. These results suggest that Slp and D expression are
not maintained from NBs to neurons and that not all the temporal
transcription factors expressed in NBs are inherited through GMCs.
However, it is possible to speculate that Klu, Slp and D regulate expression of unidentified transcription factors in NBs that are
inherited from NBs to neurons through GMCs (Suzuki, 2013).
In the Drosophila optic lobes, 800 retinotopically organized columns in the medulla act as functional units for processing visual information. The medulla contains over 80 types of neuron, which belong to two classes: uni-columnar neurons have a stoichiometry of one per column, while multi-columnar neurons contact multiple columns. This study shows that combinatorial inputs from temporal and spatial axes generate this neuronal diversity: all neuroblasts switch fates over time to produce different neurons; the neuroepithelium that generates neuroblasts is also subdivided into six compartments by the expression of specific factors (see The OPC neuroepithelium is patterned along its dorsal-ventral axis). Uni-columnar neurons are produced in all spatial compartments independently of spatial input; they innervate the neuropil where they are generated. Multi-columnar neurons are generated in smaller numbers in restricted compartments and require spatial input; the majority of their cell bodies subsequently move to cover the entire medulla. The selective integration of spatial inputs by a fixed temporal neuroblast cascade thus acts as a powerful mechanism for generating neural diversity, regulating stoichiometry and the formation of retinotopy (Erclik, 2017).
The optic lobes, composed of the lamina, medulla and the lobula complex, are the visual processing centres of the Drosophila brain. The lamina and medulla receive input from photoreceptors in the compound eye, process information and relay it to the lobula complex and central brain. The medulla, composed of ~40,000 cells, is the largest compartment in the optic lobe and is responsible for processing both motion and colour information. It receives direct synaptic input from the two colour-detecting photoreceptors, R7 and R8. It also receives input from five types of lamina neuron that are contacted directly or indirectly by the outer photoreceptors involved in motion detection (Erclik, 2017).
Associated with each of the ~800 sets of R7/R8 and lamina neuron projections are 800 medulla columns defined as fixed cassettes of cells that process information from one point in space. Columns represent the functional units in the medulla and propagate the retinotopic map established in the compound eye. Each column is contributed to by more than 80 neuronal types, which can be categorized into two broad classes. Uni-columnar neurons have arborizations principally limited to one medulla column and there are thus 800 cells of each uni-columnar type. Multi-columnar neurons possess wider arborizations, spreading over multiple columns. They compare information covering larger receptor fields. Although they are fewer in number, their arborizations cover the entire visual field (Erclik, 2017).
The medulla develops from a crescent-shaped neuroepithelium, the outer proliferation centre (OPC). During the third larval instar, the OPC neuroepithelium is converted into lamina on its lateral side and into medulla neuroblasts on its medial side. A wave of neurogenesis moves through the neuroepithelial cells, transforming them into neuroblasts; the youngest neuroblasts are closest to the neuroepithelium while the oldest are adjacent to the central brain. Neuroblasts divide asymmetrically multiple times to regenerate themselves and produce a ganglion mother cell that divides once more to generate medulla neurons. Recent studies have shown that six transcription factors are expressed sequentially in neuroblasts as they age: neuroblasts first express Homothorax (Hth), then Klumpfuss (Klu), Eyeless (Ey), Sloppy-paired 1 (Slp1), Dichaete (D) and Tailless (Tll). This temporal series is reminiscent of the Hb --> Kr --> Pdm --> Cas --> Grh series observed in Drosophila ventral nerve cord neuroblasts that generates neuronal diversity in the embryo. Indeed, distinct neurons are generated by medulla neuroblasts in each temporal window. Further neuronal diversification occurs through Notch-based asymmetric division of ganglion mother cells. In total, over 20 neuronal types can theoretically be generated using combinations of temporal factors and Notch patterning mechanisms. However, little is known about how the OPC specifies the additional ~60 neuronal cell types that constitute the medulla (Erclik, 2017).
To understand the logic underlying medulla development, late larval brains were stained with 215 antibodies generated against transcription factors and 35 genes were identifiied that are expressed in subsets of medulla progenitors and neurons. The OPC neuroepithelial crescent can be subdivided along the dorsal-ventral axis by the mutually exclusive expression of three homeodomain-containing transcription factors: Vsx1 is expressed in the central OPC (cOPC), Rx in the dorsal and ventral posterior arms of the crescent (pOPC), and Optix in the two intervening 'main arms' (mOPC). These three proteins are regionally expressed as early as the embryonic optic anlage and together mark the entire OPC neuroepithelium with sharp, non-overlapping boundaries. Indeed, these three regions grow as classic compartments: lineage trace experiments show that cells permanently marked in the early larva in one OPC region do not intermingle at later stages with cells from adjacent compartments. Of note, Vsx1 is expressed in cOPC progenitor cells and is maintained in a subset of their neuronal progeny whereas Optix and Rx are not expressed in post-mitotic medulla neurons. The OPC can be further subdivided into dorsal (D) and ventral (V) halves: a lineage trace with hedgehog-Gal4 (hh-Gal4) marks only the ventral half of the OPC, bisecting the cOPC compartment. As hedgehog is not expressed in the larval OPC, this dorsal-ventral boundary is set up in the embryo. Thus, six compartments (ventral cOPC, mOPC and pOPC and their dorsal counterparts) exist in the OPC. The pOPC compartment can be further subdivided by the expression of the wingless and dpp signalling genes. Cells in the wingless domain behave in a very distinct manner from the rest of the OPC, and have been described elsewhere (Erclik, 2017).
The Hth --> Klu --> Ey --> Slp1 --> D --> Tll temporal progression is not affected by the compartmentalization of the OPC epithelium; the same neuroblast progression throughout the entire OPC. Thus, in the developing medulla, neuroblasts expressing the same temporal factors are generated by developmentally distinct epithelial compartments (Erclik, 2017).
To test whether the intersection of the dorsal-ventral and temporal neuroblast axes leads to the production of distinct neural cell types, focus was placed on the progeny of Hth neuroblasts, which maintain Hth expression. In the late third instar, Hth neurons are found in a crescent that mirrors the OPC (see Distinct neuronal cell types are generated along the dorsal-ventral axis of the OPC). The NotchON (NON) progeny of Hth+ ganglion mother cells express Bsh and Ap, and they are distributed throughout the entire medulla crescent. In contrast, the NotchOFF (NOFF) progeny, which are Bsh−Hth+ neurons, express different combinations of transcription factors, and can be subdivided into three domains along the dorsal-ventral axis: (1) in the cOPC, NOFFHth+ neurons express Vsx1, Seven-Up (Svp) and Lim3; (2) in the pOPC, these neurons also express Svp and Lim3, but not Vsx1; (3) in the ventral pOPC exclusively, these neurons additionally express Teashirt (Tsh). NOFFHth+ cells are not observed in the mOPC. Rather, Cleaved-Caspase-3+ cells are intermingled with Bsh+ neurons. When cell death is prevented, Bsh+Hth+ cells become intermingled with neurons that express the NOFF marker Lim3, confirming that the NOFFHth+ progeny undergo apoptosis in the mOPC (Erclik, 2017).
It was therefore possible to distinguish three regional populations of Hth neurons (plus one that is eliminated by apoptosis) and a fourth population that is generated throughout the OPC. The neuronal identity of each of these populations was identified, as follows. (1) Bsh is a specific marker of Mi1 uni-columnar interneurons that are generated in all regions of the OPC. (2) To determine the identity of Hth+NOFF cOPC-derived neurons, Hth+ single cell flip-out clones were generated (using hth-Gal4) in the adult medulla. The only Hth+ neurons that are also Vsx1+Svp+ are Pm3 multi-columnar local neurons. (3) For Hth+NOFF pOPC-derived neurons, 27b-Gal4 was used; it drives expression in larval pOPC Hth+NOFF neurons and is maintained to adulthood. Flip-out clones with 27b-Gal4 mark Pm1 and Pm2 neurons, as well as Hth- Tm1 uni-columnar neurons that come from a different temporal window. Both Pm1 and Pm2 neurons (but not Tm1) express Hth and Svp. Pm1 neurons also express Tsh, which only labels larval ventral pOPC neurons (Erclik, 2017).
Thus, in addition to uni-columnar Mi1 neurons generated throughout the OPC, Hth neuroblasts generate three region-specific neuronal types: multi-columnar Pm3 neurons in the cOPC; multi-columnar Pm1 neurons in the ventral pOPC; and multi-columnar Pm2 neurons in the dorsal pOPC (Erclik, 2017).
To determine the contribution of the temporal and spatial factors to the generation of the different neuronal fates, the factors were mutated them and whether neuronal identity was lost was examined. To test the temporal axis, hth was mutated. As previously reported, Bsh expression is lost in hth mutant clones. Loss of hth in clones also leads to the loss of the Pm3 marker Svp without affecting expression of Vsx1, indicating that Vsx1 is not sufficient to activate Svp and can only do so in the context of an Hth+ neuroblast. Hth is also required for the specification of Pm1 and Pm2 in the pOPC as Svp and Tsh expression is lost in hth mutant larval clones. Ectopic expression of Hth in older neuroblasts is not able to expand Pm1, 2 or 3 fates (on the basis of the expression of Svp) into later born neurons, although it is able to expand Bsh expression. Thus, temporal input is necessary for the specification of all Hth+ neuronal fates but only sufficient for the generation of Mi1 neurons (see Temporal and spatial inputs are required for neuronal specification in the medulla. ) (Erclik, 2017).
Next, whether regional inputs are necessary and/or sufficient to specify neuronal fates in the progeny of Hth+ neuroblasts was determined. In Vsx1 RNA interference (RNAi) clones, Svp expression is lost in the cOPC but Bsh is unaffected. Additionally, Hth+Lim3+ cells are absent, suggesting that NOFF cells undergo apoptosis in these clones. Conversely, ectopic expression of Vsx1 leads to the expression of Svp in mOPC Hth+ neurons but does not affect Bsh expression. Therefore, Vsx1 is both necessary and sufficient for the specification of Pm3 fates in the larva. However, unlike the temporal factor Hth, Vsx1 does not affect the generation of Mi1 neurons (Erclik, 2017).
In Rx whole mutant larvae and in mutant clones, Svp+Lim3+Hth+ larval neurons (that is, Pm1 and Pm2 neurons) in the pOPC are lost. Additionally, the Pm1 marker Tsh is lost in ventral pOPC Hth+ cells. Consistent with the Vsx1 mutant data, larval Bsh expression is not affected by the loss of Rx. In adults, the Pm1/Pm2 markers (Svp, Tsh and 27b-Gal4) are lost in the medulla (Erclik, 2017).
Ectopic expression of Rx leads to the activation of Svp in mOPC Hth+ neurons, but does not affect the expression of Bsh. It also leads to the activation of Tsh, but only in the ventral half of mOPC Hth+ neurons, suggesting that a ventral factor acts together with Rx to specify ventral fates. Taken together, the above data show that Rx is both necessary and sufficient for the specification of Pm1/2 neurons but (like Vsx1) does not affect the generation of Mi1 neurons (Erclik, 2017).
Finally, the role of the mOPC marker Optix in neuronal specification was examined. In Optix mutant clones, Svp is ectopically expressed in the mOPC, but Bsh expression is not affected. Of note, these ectopic Svp+ neurons fail to express the region-specific Pm markers Vsx1 or Tsh (in ventral clones), which suggests that they assume a generic Pm fate. Conversely, ectopic expression of Optix leads to the loss of Svp expressing neurons in both the cOPC and pOPC but does not affect Bsh. These NOFF neurons die by apoptosis as no Lim3+ neurons are found intermingled with Bsh+Hth+NON neurons. When apoptosis is prevented in mOPC-derived neurons, Svp is not derepressed in the persisting Hth+NOFF neurons, which suggests that Optix both represses Svp expression and promotes cell death in Hth+NOFF neurons (Erclik, 2017).
The above data demonstrate that input from both the temporal and regional axes is required to specify neuronal fates. The temporal factor Hth is required for both Mi1 and Pm1/2/3 specification. The spatial genes are not required for the specification of NON Mi1 neurons, consistent with the observation that Mi1 is generated in all OPC compartments. The spatial genes, however, are both necessary and sufficient for the activation (Vsx1 and Rx) or repression (Optix) of the NOFF Pm1/2/3 neurons. Thus, Hth+ neuroblasts generate two types of progeny: NOFF neurons that are sensitive to spatial input (Pm1/2/3) and NON neurons that are refractory to spatial input (Mi1). Vsx1 expression in the cOPC is only maintained in Hth+NOFF neurons, suggesting that spatial information may be 'erased' in Mi1, thus allowing the same neural type to be produced throughout the OPC (Erclik, 2017).
Do spatial genes regulate each other in the neuroepithelium? In Vsx1 mutant clones, Optix (but not Rx) is derepressed in the cOPC epithelium. Conversely, ectopic Vsx1 is sufficient to repress Optix in the mOPC and Rx in the pOPC. Similarly, Optix, but not Vsx1, is derepressed in Rx mutant clones in the pOPC epithelium and ectopic Rx is sufficient to repress Optix in the mOPC (but not Vsx1 in the cOPC). In Optix mutant clones, neither Vsx1 nor Rx are derepressed in the mOPC epithelium, but ectopic Optix is sufficient to repress both Vsx1 in the cOPC and Rx in the pOPC. The observation that Optix is not necessary to suppress Vsx1 or Rx in the mOPC neuroepithelium is surprising because Svp is activated in a subset of Hth+ neurons in the mOPC in Optix mutant clones. Nevertheless, when cell death in the mOPC is abolished, the ectopic undead NOFF neurons express Lim3 but not Svp, which confirms that Optix represses Svp expression in mOPC neurons. Taken together, these results support a model in which Optix is sufficient to repress Vsx1 and Rx, to promote the death of Hth+NOFF neurons and to repress Pm1/2/3 fates (see Spatial genes cross-regulate each other in the OPC neuroepithelium). Vsx1 and Rx act to promote Pm3 (Vsx1) or Pm1/2 (Rx) fates but can only do so in the absence of Optix (Erclik, 2017).
These results suggest that multi-columnar neurons are generated at specific locations in the medulla crescent. However, since these neurons are required to process visual information from the entire retina in the adult medulla, how does the doral-ventral position of neuronal birth in the larval crescent correlate with their final position in the adult? Lineage-tracing experiments were performed with Vsx1-Gal4 to permanently mark neurons generated in the cOPC and with Optix-Gal4 for mOPC neurons, and the position of the cell bodies of these neurons was analyzed. In larvae, neurons from the cOPC or from the mOPC remain located in the same dorsal-ventral position where they were born. However, in adults, both populations have moved to populate the entire medulla cortex along the dorsal-ventral axis (see Neuronal movement during medulla development is restricted to multi-columnar cell types). The kinetics of cell movement during development was analyzed by following cOPC neurons. Neurons born in the cOPC remain tightly clustered until 20 h after puparium formation (P20), after which point the cell bodies spread throughout the medulla cortex. By P30 the neurons are distributed over the entire dorsal-ventral axis of the medulla cortex. In the adult, most neurons derived from the cOPC neuroepithelium are located throughout the cortex although there is an enrichment of neurons in the central region of the cortex (Erclik, 2017).
To determine whether these observed movements involve the entire neuron or just the cell body, the initial targeting of cOPC or mOPC-derived neurons in larvae was examined before the onset of cell movement. In larvae, both populations send processes that target the entire dorsal-ventral axis of the medulla neuropi. Therefore, medulla neurons first send projections to reach their target columns throughout the entire medulla. Later, remodelling of the medulla results in extensive movement of cell bodies along the dorsal-ventral axis, leading to their even distribution in the cortex (Erclik, 2017).
What is the underlying logic behind why some neurons move while others do not? Markers were studied for the Mi1 (Bsh), Pm2 (Hth+Svp+), Pm1 (Hth+Svp+Tsh+), and Pm3 (Vsx1+Svp+Hth+) populations of neurons through pupal stages and up to the adult. Mi1 neurons are generated evenly throughout the larval OPC and remain regularly distributed across the dorsal-ventral axis at all stages. The lineage-tracing experiment was repeated with Vsx1-Gal4 to follow Mi1 neurons produced by the cOPC. These Mi1 neurons remain exclusively in the centre of the adult medulla cortex, demonstrating that they do not move. In contrast, Pm3 neurons remain tightly clustered in the central region until P20, at which point they move to occupy the entire cortex (Erclik, 2017).
However, not all multi-columnar neurons have cell bodies that move to occupy the entire medulla cortex. Unlike Mi1 and Pm3, adult Pm1 and Pm2 cell bodies are not located in the adult medulla cortex but instead in the medulla rim, at the edges of the cortex. Pm1 and Pm2 markers remain clustered at the ventral (Pm1) or dorsal (Pm2) posterior edges of the medulla cortex throughout all pupal stages. In the adult, both populations occupy the medulla rim from where they send long horizontal projections that reach the entire dorsal-ventral axis of the medulla neuropil. The pOPC may be a specialized region where many of the medulla rim cell types are generated. Even though most of cOPC-derived neurons move during development, a cOPC-derived multi-columnar neuron (TmY14) was identified that sends processes targeting the entire dorsal-ventral length of the medulla neuropil but whose cell bodies remain in the central medulla cortex in the adult (Erclik, 2017).
Thus, the four populations of Hth neurons follow different kinetics: Mi1 neurons are born throughout the OPC and do not move; Pm3 neurons are born centrally and then move to distribute throughout the entire cortex; and Pm1/Pm2 neurons are born at the ventral or dorsal posterior edges of the OPC and occupy the medulla rim in adults (Erclik, 2017).
It is noted that uni-columnar Mi1 neurons, whose cell bodies do not move, reside in the distal cortex whereas multi-columnar Pm3 neurons, which move, reside in the proximal cortex. The hypothesis was thus tested that neurons whose cell bodies are located distally in the medulla cortex represent uni-columnar neurons generated homogeneously throughout the OPC that do not move. In contrast, proximal neurons, which are fewer in number and are generated in specific subregions of the medulla OPC, would be multi-columnar and move to their final position (Erclik, 2017).
It was first confirmed that neurons that move have their cell bodies predominantly in the proximal medulla cortex. The cell body position of neurons born ventrally that have moved dorsally was analyzed using the hh-Gal4 lineage trace: in the adult, the cell bodies found dorsally are mostly in the proximal medulla cortex, whereas the cell bodies in the ventral region are evenly distributed throughout the distal-proximal axis of the ventral cortex. They probably represent both distal uni-columnar neurons that did not move as well as proximal multi-columnar neurons that remained in the ventral region (Erclik, 2017).
Next the pattern of movement of Tm2 uni-columnar neurons from the ventral and dorsal halves of the OPC was analyzed using the hh-based lineage-trace. The cell bodies of Tm2 neurons are located throughout the dorsal-ventral axis in the adult medulla cortex but are co-labelled with the hh lineage marker only in the ventral half. Thus, like Mi1, Tm2 uni-columnar neurons do not move. Furthermore, uni-columnar Tm1 neurons, labelled by 27b-Gal4, are born throughout the dorsal-ventral axis of the OPC crescent with distal cell bodies in the adult, suggesting that they also remain where they were born (Erclik, 2017).
Conversely, it was asked whether neurons that are specified in only one region, such as the Vsx+ neurons of the cOPC, are multi-columnar in morphology. By sparsely labelling cOPC-derived neurons using the Vsx1-Gal4 driver, 13 distinct cell types were characterized that retain Vsx1 expression in the adult medulla. Strikingly, all are multi-columnar in morphology, further supporting the model that it is the multi-columnar neurons that move during pupal development (Erclik, 2017).
Finally, MARCM clones were generated in the OPC neuroepithelium and visualized using cell-type-specific Gal4 drivers in the adult medulla. Two classes of adult clone distribution were observed: clones in which neurons are tightly clustered, and clones in which neurons are dispersed. Consistent with the model, the clustered clones are those labelled with uni-columnar neuronal drivers, whereas the dispersed clones are those labelled with a multi-columnar driver (Erclik, 2017).
Taken together, these data demonstrate that neurons that do not move are uni-columnar (with cell bodies in the distal cortex), whereas most multi-columnar neurons (with cell bodies in the proximal cortex) move (Erclik, 2017).
This study shows that combinatorial inputs from the temporal and spatial axes act together to promote neural diversity in the medulla. Previous work has shown that a temporal series of transcription factors expressed in medulla neuroblasts allows for a diversification of the cell types generated by the neuroblasts as they age. This study now shows that input from the dorsal-ventral axis leads to further diversification of the neurons made by neuroblasts; at a given temporal stage, neuroblasts produce the same uni-columnar neuronal type globally as well as smaller numbers of multi-columnar cell types regionally. This situation is reminiscent of the mode of neurogenesis in the Drosophila ventral nerve cord, in which each neuroblast also expresses a (different) temporal series of transcription factors that specifies multiple neuronal types in the lineage. Spatial cues from segment polarity, dorsal-ventral and Hox genes then intersect to impart unique identities to each of the lineages. However, neuroblasts from the different segments give rise to distinct lineages to accommodate the specific function of each segment. In contrast, in the medulla, the entire OPC contributes to framing the repeating units that form the retinotopic map. It is therefore likely that each neuroblast produces a common set of neurons that connect to each pair of incoming R7 and R8 cells, or L1-L5 lamina neurons. This serves to produce 800 medulla columns with a 1:1 stoichiometry of medulla neurons to photoreceptors. The medulla neurons that are produced by neuroblasts throughout the dorsal-ventral axis of the OPC are thus uni-columnar The production of the same neuronal type along the entire OPC could be achieved by selectively 'erasing' spatial information in uni-columnar neurons, as observed in Mi1 neurons (Erclik, 2017).
Regional differences in the OPC confer further spatial identities to neuroblasts with the same temporal identity, and lead to specific differences in the lineages produced in the compartments along the dorsal-ventral axis of the medulla. These differences produce smaller numbers of multi-columnar neurons whose stoichiometry is much lower than 1:1. The majority of these neurons move during development to be uniformly distributed in the adult medulla cortex. This combination of regional and global neuronal specification in the medulla presents a powerful mechanism to produce the proper diversity and stoichiometry of neuronal types and generate the retinotopic map (Erclik, 2017).
Photoreceptor (PR) axons project from the retina to the optic lobe in brain and form a precise retinotopic map in the Drosophila visual system. Yet the role of retinal basal glia in the retinotopic map formation is not previously known. This study examined the formation of the retinotopic map by marking single PR pairs and following their axonal projections. In addition to confirming previous studies that the spatial information is preserved from the retina to the optic stalk and then to the optic lamina, this study found that the young PR R3/4 axons transiently overshoot and then retract to their final destination, the lamina plexus. The process of wrapping glia (WG) membrane extension was examined in the eye disc, and the WG membrane extensions also follow the retinotopic map. The WG is important for the proper spatial distribution of PR axons in the optic stalk and lamina, suggesting an active role of wrapping glia in the retinotopic map formation (Chang, 2018).
DmMANF, Drosophila melanogaster mesencephalic astrocyte-derived neurotrophic factor, is an evolutionarily conserved orthologue of mammalian MANF. This neurotrophic factor exerts many functions in the Drosophila brain, particularly those dependent on glial cells. An earlier study found that downregulation of DmMANF in glia induces degeneration of glial cells in the first optic neuropil (lamina) where DmMANF abundance is especially high. The present study observed that changes in the level of DmMANF in two types of glia, astrocyte-like glia (AlGl) and ensheathing glia (EnGl), affect activity and sleep of flies. Interestingly, a proper level of DmMANF in AlGl seems to be important in guiding processes of pigment dispersing factor (PDF)-expressing clock neurons. This is supported by THE finding that DmMANF overexpression in AlGl leads to structural changes in the architecture of the PDF-positive arborisation in the brain. Finally, it was detected that DmMANF also affects rhythms in glia themselves, since circadian oscillations in expression of the catalytic α subunit of the sodium pump in the lamina epithelial glia were abolished after DmMANF silencing. DmMANF-expressed in AlGL and EnGl seems to affect activity of neurons leading to changes in behaviour (Walkowicz, 2021).
During brain development, various types of neuronal populations are
produced from different progenitor pools to produce neuronal diversity
that is sufficient to establish functional neuronal circuits. However, the
molecular mechanisms that specify the identity of each progenitor pool
remain obscure. This study shows that Wnt signaling is essential for the specification of the identity of
posterior progenitor pools in the Drosophila visual center. In the medulla, the largest component of the visual center,
different types of neurons are produced from two progenitor pools: the
outer proliferation center (OPC) and glial precursor cells (GPCs; also
known as tips of the OPC). It was found that OPC-type neurons are produced
from the GPCs at the expense of GPC-type neurons when Wnt signaling is
suppressed in the GPCs. In contrast, GPC-type neurons are ectopically
induced when Wnt signaling is ectopically activated in the OPC. These
results suggest that Wnt signaling is necessary and sufficient for the
specification of the progenitor pool identity. It was also found that The adult optic lobe of Drosophila develops from the primordium during metamorphosis from mid-3rd larval stage to adult. Many cells die during development of the optic lobe with a peak of the number of dying cells at 24 h after puparium formation (h APF). Dying cells were observed in spatio-temporal specific clusters. This study analyzed the function of a component of the insect steroid hormone receptor, EcR, in this cell death. Expression patterns of two EcR isoforms, EcR-A and EcR-B1, were examined in the optic lobe. Expression of each isoform altered during development in isoform-specific manner. EcR-B1 was not expressed in optic lobe neurons from 0 to 6h APF, but was expressed between 9 and 48 h APF and then disappeared by 60 h APF. In each cortex, its expression was stronger in older glia-ensheathed neurons than in younger ones. EcR-B1 was also expressed in some types of glia. EcR-A was expressed in optic lobe neurons and many types of glia from 0 to 60 h APF in a different pattern from EcR-B1. Then, EcR function were genetically analyzed in the optic lobe cell death. At 0 h APF, the optic lobe cell death was independent of any EcR isoforms. In contrast, EcR-B1 was required for most optic lobe cell death after 24 h APF. It was suggested that cell death cell-autonomously required EcR-B1 expressed after puparium formation. betaFTZ-F1 was also involved in cell death in many dying-cell clusters, but not in some of them at 24 h APF. Altogether, the optic lobe cell death occurred in ecdysone-independent manner at prepupal stage and ecdysone-dependent manner after 24 h APF. The acquisition of ecdysone-dependence was not directly correlated with the initiation or increase of EcR-B1 expression (Hara, 2013).
This study analyzed the requirement of ecdysone in the optic lobe cell death. The role of ecdysone in cell death during metamorphosis has been examined in the salivary gland, larval midgut, and two types of neurons in the VNC, vCrz neurons and RP2 neurons. In the salivary gland, ecdysone triggers cell death in vitro, and the cell death required some components of ecdysone cascade, BR-C, E93, E74 and βFTZ-F1. In the midgut, cell death was induced by injection of ecdysone, and required BR-C and E93. For the cell death in vCrz neurons and RP2 neurons, ecdysone requirement was shown using EcR mutants. Among these tissues, a requirement for EcR isoforms was addressed only in the vCrz neurons and RP2 neurons. In the vCrz neurons, the cell death occurred in EcR-A or EcR-B1 mutants, but not in EcR-B1 and EcR-B2 mutant, indicating that EcR-B2 is required for this cell death. In RP2 neurons, cell death did not require EcR-A, but EcR-B1, EcR-B2 or both. Here, it was shown that the optic lobe cell death included ecdysone-independent and dependent tissues. The ecdysone-dependent cell death required EcR-B1 (Hara, 2013).
The number of dying cells in the optic lobe of EcR-B1 mutant animals at 24 and 36 h APF was much smaller than that in wild-type animals, but the number in EcR-A mutant animals was not. This finding showed that cell death in the optic lobe at these stages required EcR-B1, but not EcR-A (Hara, 2013).
Dying cells were examined in the following structures; LAD, lamina anterior dying cells; LPD, lamina posterior dying cells; LUD, lamina underlying dying cells; MALD, medulla anterolateral dying cells; MAMD, medulla anteromedial dying cells; MCD, medulla cortex dying cells; MCLD, medulla cortex lateral dying cells; MCMD, medulla cortex medial dying cells; MLBD, medulla-lamina boundary dying cells; MPLD, medulla posterolateral dying cells; PMCD, posterior medulla cortex dying cells; PMD, posteromedial dying cells; T/C, dying cells in the T/C region; LopD, dying cells in the lobula plate cortex; MMC, abnormal dying cells in the medial side of the medulla cortex; MLopD, abnormal dying cells in the medial side of the lobula plate cortex. The dependence on EcR-B1 was common among all dying cells in all clusters, except the MCMD. At 24 h APF, dying cells in the LAD, LPD, LUD, MCLD and T/C region were absent in optic lobes of most EcR-B1 mutants. In these mutants at 36 h APF, dying cells in the LUD, MLBD, MPLD and the lobula plate cortex were not evident. Similarly, at 48 h APF, MPLDs were absent from the EcR-B1 mutants. These results indicated that cell death in most clusters required EcR-B1 at stages after 24 h APF. It could not be determined whether death of MCMD was dependent on EcR-B1 because it was not clear whether the abnormal dying cells included MCMD in the medial side of the medulla cortex in EcR-B1 mutants (Hara, 2013).
In EcR-B1 mutant, a significant number of dying cells was constantly observed after 24 h APF. However, this fact does not mean that the loss of EcR-B1 delayed the timing of cell death. From 24 to 72 h APF, dying cells were mostly located in the medial side of the medulla cortex in EcR-B1 mutants and they did not include those in the clusters which would have been normally observed from 24 to 48 h APF, i.e., the LAD, LPD, LUD, MCLD, MPLD, MLBD, and dying cells in the T/C region and the lobula plate cortex. This fact strongly suggests that the optic lobe cell death was not delayed but suppressed by EcR-B1 mutation (Hara, 2013).
In some EcR-B1 mutants, enormous dying cells were observed at 72 h APF at positions where cell death would have normally occurred: the LAD, MCLD, MPLD, MLBD and T/C region. This suggests that delayed cell death can be induced at normal position without EcR-B1 at 72 h APF in these samples. It is known that experimental suppression of cell death can lead a delayed cell death by another complementally cell death mechanism. Therefore, it is possible that a complementary mechanism was induced in these samples (Hara, 2013).
It has been shown that a cell death initiator caspase, Dronc, had a EcRE in its promoter and EcR-B1 could induce Dronc expression. Indeed, the optic lobe cell death was suppressed in Dronc mutant in a preliminary experiment. Altogether, it is most likely that EcR-B1 directly controls cell death and consistently induces the death at right time in the optic lobe (Hara, 2013).
In this study, requirement of EcR-B2 function was not suggested. When the functions of all EcR isoforms were inhibited after puparium formation, the number of dying cells was larger than that in EcR-B1 mutant at 24 h APF (638.6 versus 363.8). If EcR-B2 was required for the optic lobe cell death, the number would have been less than that in the mutant. However, a function of EcR-B2 in the ecdysone-dependent cell death can still not be entirely excluded since there was a possibility that RNAi was insufficient to entirely inhibit EcR function in this experimental condition (Hara, 2013).
The number of dying cells in the optic lobes of EcR-A and EcR-B1 mutants was the same as that in wild-type animals at 0 h APF. When distribution of dying cells was examined, all clusters that were present in wild-type animals (specifically the LAD, MALD, MCD, MAMD, PMCD and dying cells in the T/C region) were also observed in the mutants. These results strongly indicated the cell death at 0 h APF was independent of both EcR-A and EcR-B1. EcR-B2 was also not required for the cell death because concurrent knockdown of all EcR isoforms via expression of hs-EcRi-11 resulted in no reduction in the number of dying cells at 0 h APF (Hara, 2013).
The above argument is relevant only for the zygotic not with maternal EcR. The contribution of maternal EcR should be tested. The result of the heat shock-inducible EcR RNAi denied the contribution of maternal EcR mRNA of all EcR isoforms. As with maternal proteins, there was no detectable EcR-B1 in any cluster or region at 0 h APF. In contrast, EcR-A was weakly expressed in all cluster regions, and these is no information about EcR-B2. Therefore, possible roles of maternal EcR-A and EcR-B2 protein cannot be excluded. However, there are no published reports of a requirement for maternal EcR proteins during metamorphosis. Taken together, these findings indicated that cell death in the optic lobe at 0 h APF is independent of any EcR. Furthermore, it seems likely this cell death is also independent of ecdysone because the number of dying cells gradually increased from 0 to 6 h APF rather than decreased, but the ecdysone titer rapidly drops and is very low during this period (Hara, 2013).
The period around 12 h APF may be a transient period when the ecdysone-dependence of cell death changes. In many EcR-B1 mutant optic lobes, the number of dying cells was the same as that in the wild type. In contrast, the number was reduced in a few mutant optic lobes and dying cells were absent in many of the clusters. These findings indicated that most of the cell deaths in many optic lobes was independent of EcR-B1, but some had become EcR-B1-dependent (Hara, 2013).
There is no previous report on ecdysone-independent cell death during metamorphosis. The ecdysone-independent cell death was limited to the early phase of metamorphosis in the optic lobe. However, this timing does not necessarily indicate that all cell death is independent of ecdysone because the larval midgut and vCrz neurons die ecdysone-dependently during this period. Therefore, ecdysone independence is a unique feature of the cell death that occurs in the optic lobe. There has been no report that the cell death that occurs during embryogenesis and larval development depends on ecdysone. Hence, it is proposed that the same mechanisms that mediate cell death during embryogenesis or larval development work for cell death during the early phase of optic lobe development (Hara, 2013).
Based on findings from many previous studies, every cell death that occurred during metamorphosis was part of the degeneration of a larval tissue and was dependent on ecdysone. These finding are understandable because ecdysone orchestrates the entire developmental process of metamorphosis. However, cell death within the optic lobe was independent of ecdysone during an early phase and then this cell death became ecdysone dependent later. This unique feature of the optic lobe cell death may be due to the fact that cell death in the optic lobe takes place during metamorphosis and is simultaneously involved in the organogenesis. So two cell death mechanisms, i.e., an organogenesis-accompanied (ecdysone-independent) mechanism and a metamorphosis-accompanied (ecdysone-dependent) mechanism may have evolved to cooperate during the optic lobe development (Hara, 2013).
The expression pattern of EcR-A and EcR-B1 was examined in this study. Expression of each isoform altered during development in isoform specific manner. However, there was no direct relationship between EcR-B1 expression and the emergence of the cell death. At 0 h APF, when cells die independent of ecdysone, EcR-B1 was not expressed in any region with clusters of dying cells. In contrast, EcR-B1 was expressed in all regions with clusters of dying cells at 12 h APF, although cell death, at this stage, was, for the most part, ecdysone independent in all clusters, except PMD. Thus, there was a temporal gap between EcR-B1 expression and ecdysone-dependent cell death. This indicates that the expression of EcR-B1 was not a direct cause that shifted cell death from an ecdysone-independent to an ecdysone-dependent one. EcR-B1 expression would be one of the requisites to make cells competent to undergo ecdysone-dependent cell death at a later time point and another mechanism following EcR-B1 expression would be required for the shift (Hara, 2013).
Although cell death in the optic lobe after 24 h APF required EcR-B1, the level of EcR-B1 expression varied among cluster regions during this period. For example, at 24 h APF, EcR-B1 was expressed weakly in the anterior region of the lamina cortex where LAD was located. On the lateral side of the medulla cortex where MCLD were present, EcR-B1 was expressed moderately. EcR-B1 was strongly expressed in the T/C region where many dying cells were present. The expression levels also varied among cluster regions at 36 and 48 h APF. All these findings indicate that the death decision, even for the ecdysone-dependent cell death, was not simply related to high EcR-B1 levels. This decision would be made within specific context of each cluster (Hara, 2013).
EcR-B1 expression was correlated with glial ensheathment in the lamina cortex, medulla cortex and T/C region. In these regions, newly-born neurons derived from the OOA or IOA compose pre-ensheathed domains. As development proceeds, they become to be surrounded by glial membrane and compose ensheathed domains of mature neurons. In particular, the ensheathed domain in lamina cortex corresponds to a region with columnar structures. In the lamina cortex, EcR-B1 was weakly expressed in the pre-ensheathed domain, while strongly expressed in the ensheathed domain. In the medulla cortex and T/C region, it was not expressed in the pre-ensheathed domains, but expressed in the ensheathed domains. These facts suggest a possibility that the glial ensheathment promotes or initiates EcR-B1 expression in the process of neuronal differentiation in these regions. This possibility is supported by the fact that EcR-B1 expression became stronger after the ensheathment as development proceeded (Hara, 2013).
With regard to cell death, the LAD, MCLD and MALD were always located near the border of the pre-ensheathed and ensheathed domains. Therefore, cell death may be linked to the entry of glial membrane in these clusters. Since this positional relationship was observed from 0 to 24 h APF, the glial ensheathement and ecdysone signaling via EcR-B1 may cooperate to induce cell death in the clusters after 12 h APF, when cell death become dependent on EcR-B1 (Hara, 2013).
Temporal patterning of neural progenitors is one of the core mechanisms generating neuronal diversity in the central nervous system. This study shows that, in the tips of the outer proliferation center (tOPC) of the developing Drosophila optic lobes, a unique temporal series of transcription factors not only governs the sequential production of distinct neuronal subtypes but also controls the mode of progenitor division, as well as the selective apoptosis of NotchOFF or NotchON neurons during binary cell fate decisions. Within a single lineage, intermediate precursors initially do not divide and generate only one neuron; subsequently, precursors divide, but their NotchON progeny systematically die through Reaper activity, whereas later, their NotchOFF progeny die through Hid activity. These mechanisms dictate how the tOPC produces neurons for three different optic ganglia. It is concluded that temporal patterning generates neuronal diversity by specifying both the identity and survival/death of each unique neuronal subtype (Bertet, 2014).
Although apoptosis is a common feature of neurogenesis in both vertebrates and Drosophila, the mechanisms controlling this process are still poorly understood. For instance, several studies in Drosophila have shown that, depending on the context, Notch can either induce neurons to die or allow them to survive during binary cell fate decisions. This is the case in the antennal lobes where Notch induces apoptosis in the antero-dorsal projecting neurons lineage (adpn), whereas it promotes survival in the ventral projecting neurons lineage (vPN). In both of these cases, the entire lineage makes the same decision whether the NotchON or NotchOFF cells survive or die. This suggests that, in this system, Notch integrates spatial signals to specify neuronal survival or apoptosis (Bertet, 2014).
This study shows that, during tOPC neurogenesis, neuronal survival is determined by the interplay between Notch and temporal patterning of progenitors. Indeed, within the same lineage, Notch signaling leads to two different fates: it first induces neurons to die, whereas later, it allows them to survive. This switch is due to the sequential expression of three highly conserved transcription factors-Dll/Dlx, Ey/Pax-6, and Slp/Fkh-in neural progenitors. These three factors have distinct functions, with Dll promoting survival of NotchOFF neurons, Ey inducing apoptosis of NotchOFF neurons, and Slp promoting survival of NotchON neurons. These data suggest that Ey induces death of NotchOFF neurons by activating the proapoptotic factor hid. Thus, Dll probably antagonizes Ey activity by preventing Ey from activating hid. The data also suggest that Notch signaling induces neuronal death by activating the proapoptotic gene rpr. Thus, Slp might promote survival of NotchON neurons by directly repressing rpr expression or by preventing Notch from activating it. In both cases, the interplay between Notch and Slp modifies the default fate of NotchON neurons, allowing them to survive. Further investigations will test these hypotheses and determine how Dll, Ey, Slp, and Notch differentially activate/repress hid and rpr (Bertet, 2014).
Although the tOPC and the main OPC have related temporal sequences, their neurogenesis is very different. This difference is in part due to the fact that newly specified tOPC neuroblasts express Dll, which controls neuronal survival, instead of Hth. Why do tOPC neuroblasts express Dll? The tOPC, which is defined by Wg expression in the neuroepithelium, is flanked by a region expressing Dpp. Previous studies have shown that high levels of Wg and Dpp activate Dll expression in the distal cells of the Drosophila leg disc. Wg and Dpp could therefore also activate Dll in the neuroepithelium and at the beginning of the temporal series in tOPC progenitors. Another difference between the main OPC and tOPC neurogenesis is that Ey and Slp have completely different functions in these regions. Indeed, unlike in the main OPC, Ey and Slp control the survival of tOPC neurons. This suggests that autonomous and/or nonautonomous signals interact with these temporal factors and modify their function in the tOPC (Bertet, 2014).
Finally, tOPC neuroblasts produce neurons for three different neuropils of the adult visual system, the medulla, the lobula, and the lobula plate. This ability could be due to the particular location of this region in the larval optic lobes. Indeed, the tOPC is very close to the two larval structures giving rise to the lobula and lobula plate neuropils-Dll-expressing neuroblasts are located next to the lobula plug, whereas D-expressing neuroblasts are close to the IPC. Interestingly, Dll and D neuroblasts specifically produce lobula plate neurons. This raises the possibility that these neuroblasts and/or the neurons produced by these neuroblasts receive signals from the lobula plug and the IPC, which instruct them to specifically produce lobula plate neurons. These nonautonomous signals could also modify the function of Ey and Slp in the tOPC (Bertet, 2014).
In summary, this study demonstrates that temporal patterning of progenitors, a well-conserved mechanism from Drosophila to vertebrates, generates neural cell diversity by controlling multiple aspects of neurogenesis, including neuronal identity, Notch-mediated cell survival decisions, and the mode of intermediate precursor division. In the tOPC temporal series, some factors control two of these aspects (Ey), whereas others have a specialized function (Dll, Slp, and D). This suggests that temporal patterning does not consist of a unique series of transcription factors controlling all aspects of neurogenesis but instead consists of multiple superimposed series, each with distinct functions (Bertet, 2014).
The arrival of retinal axons in the Drosophila brain triggers the assembly of glial and neuronal precursors into a neurocrystalline array of lamina synaptic cartridges. Retinal axons arriving
from the eye imaginal disc trigger the assembly of neuronal and glial precursors into precartridge ensembles in the crescent-shaped lamina
target field. In the eye disc, photoreceptor cells assemble into ommatidial clusters behind the morphogenetic
furrow (mf) as it moves to the anterior. The ommatidial clusters project their axon fascicles into the
crescent-shaped lamina. Neuronal precursor cells of the lamina (LPCs) are incorporated into the
axon target field at its anterior margin, which is demarcated by a morphological depression known as the lamina furrow. Glia precursor cells (GPCs) are generated in two domains that lie at the dorsal and ventral anterior margins of the prospective lamina. These glial precursors migrate into the lamina
along an axis perpendicular to that of LPC entry. Postmitotic LPCs within the lamina axon target field express the nuclear protein Dac, as revealed by anti-Dac antibody staining. Like the eye, lamina differentiation occurs in a temporal progression on the anterioposterior axis. Axon fascicles from new ommatidial R-cell clusters arrive at the anterior margin of the lamina (adjacent to the lamina furrow) and associate with neuronal and glia precursors in a vertical lamina column assembly. At the anterior of the lamina, at the trough of the lamina furrow, LPCs await a retinal axon-mediated signal in G1-phase and enter their terminal S-phase at the posterior margin of the furrow. Postmitotic (Dac-positive) LPCs assemble into columns at the posterior
margin of the furrow. In older columns at the posterior of the lamina, a subset of postmitotic LPCs express definitive neuronal markers as they become specified as the lamina neurons L1-L5. Lamina neurons L1-L4 form a stack in a superficial layer, while L5 neurons reside in a medial layer near
the R1-R6 axon termini. These neurons arise at cell-type specific positions along the column's
vertical axis. Lamina glial cells take up cell-type positions in the precartridge assemblies. Epithelial (E-glia) and marginal (Ma-glia) glia are located above and below the R1-R6 termini, respectively.
Satellite glia are interspersed among the neurons of the L1-L4 layer. The Ma-glia and E-glia layers, both located ventral to the neuronal precursor column, sandwich the R1-R6 axon termini. The
medulla neuropil serves as the target for R7/8 axons and is separated from the lamina by the medulla glia, situated just below the Ma-glia (Huang, 1998 and references).
Hedgehog is transmitted along retinal axons to serve as the inductive signal in the brain for differentiation of lamina neurons. The target of HH is wingless, which in turn targets decapentaplegic and Distal-less. The lamina is a ganglion layer of the visual center of the brain that processes information received from R1-R6 photoreceptor neurons located in the ommatidia of compound eyes. The lamina develops in a precise order, directly coupled to the arrival of retinal axons from the eye. Ectopic hh expression in the brains of eyeless flies induces lamina differentiation, but is not sufficient to induce Elav, a late marker. This suggests that HH alone is not sufficient for the later events of lamina development that include the specification of lamina neurons (Huang, 1998).
Hedgehog, a secreted protein, is an inductive signal
delivered by retinal axons for the initial steps of lamina
differentiation. In the development of many tissues,
Hedgehog acts in a signal relay cascade via the induction
of secondary secreted factors. Lamina
neuronal precursors respond directly to Hedgehog signal
reception by entering S-phase, a step that is controlled by
the Hedgehog-dependent transcriptional regulator Cubitus
interruptus. The terminal differentiation of neuronal
precursors and the migration and differentiation of glia
appear to be controlled by other retinal axon-mediated
signals. Thus retinal axons impose a program of
developmental events on their postsynaptic field utilizing
distinct signals for different precursor populations (Huang, 1998).
A number of markers distinguish glial and neuronal precursor cells from the corresponding mature cell types. The expression of optomotor-blind (omb) labels both glial precursors in the dorsal and ventral
anlagen and mature glia that have migrated into the lamina
target field. The glia cell marker Repo and the enhancer-trap lacZ insertion 3-109 are expressed by glia once they have entered the lamina target field. Cubitus interruptus (Ci), a transcriptional
mediator of Hh signaling is expressed by LPCs anterior of the lamina furrow
and by the postmitotic neuronal precursors within the lamina. The nuclear protein Dachshund is expressed only by neuronal precursors that have begun terminal differentiation and lie posterior to the lamina
furrow. Thus, Omb and Ci label the glial and neuronal precursors, respectively, while
the mature cells, following their interaction with retinal axons,
additionally express Repo and Dac. In the lamina target field of eyeless mutants (mutants that project no neurons toward the optic disc), such as eyes absent (eya) or sine oculis (so),
Dac expression is not detected and Repo expression is greatly diminshed (Huang, 1998).
The migration and early differentiation of lamina glia are independent of Hh.
Enhanced transcription of the putative Hh receptor, patched (ptc) is a universal characteristic of Hh signal reception. All classes of glia in the lamina region upregulate
ptc expression in an hh-dependent fashion. These cells are thus Hh-responsive. All three classes of lamina glia, as well as medulla glia,
that express a ptc-lacZ reporter construct are in close proximity
to Hh-bearing retinal axons. Glia cell ptc reporter gene expression is not observed in hh- animals. This raises the question of whether Hh signal
reception is responsible for the migration and/or subsequent
maturation of glia cells. To determine whether the migration of glial precursors into the lamina target field is Hh-dependent, the distribution of Omb-positive cells was examined in hh- animals. In the wild type, a trail of Omb-positive cells delineates a path of glia migration from the dorsal and vental anlagen.
Is glia precursor migration Hh-dependent? This was investigated
by examining the distribution of Omb-positive cells
in hh1 mutant animals. hh1 is a regulatory mutation that specifically
affects hh expression in the visual system. In hh1 animals,
approximately 12 columns of ommatidia initiate differentiation
in the eye imaginal disc before the anterior progression of the
morphogenetic furrow ceases. hh1
retinal axons lack Hh immunoreactivity by the time they reach
the lamina target field and thus the Hh-dependent steps of LPC
maturation fail to occur in hh1 animals. Omb staining reveals a relatively
normal number of glia precursors in the lamina target field of
hh1 animals, despite the absence of Dac induction. The Omb-positive cells are distributed uniformly along the dorsoventral axis among the retinal axon fascicles, but appear
more closely spaced than in the wild type. A likely explanation
for this spacing defect is the absence of the neuronal precursors
that would constitute the majority of lamina cells at this point in development. To determine whether the glial precursors that enter the lamina target field in hh- animals express a retinal innervation-dependent
marker, their expression of Repo was examined. In hh1 animals, the Omb-positive
cells within the lamina also express Repo. Moreover, the Repo-positive cells occupy proper layers above and below the R1-R6 axon termini expected for satellite, marginal and
epithelial glia, though the lack of markers specific for these three glia types precludes an unambiguous determination of glial cell type. The presence of marginal and epithelial glia is
consistent with the observation that R1-R6 growth cones terminate in their proper positions between these layers in hh- animals. The ectopic expression of Hh in the brains of `eyeless'
animals is sufficient to induce the initial steps of LPC
maturation in the absence of retinal axons. However, neither Hh
nor the Hh-mediated events of LPC maturation are sufficient
for glia cell migration and maturation (Huang, 1998).
The activities of a number of Hh signal transduction pathway components are now
well characterized. Mutations at these loci have been shown to either mimic or block Hh signal
reception in a cell-autonomous fashion. Examining the cellular requirements for these genes in mosaic animals should help illuminate the cellular circuitry that mediates the
Hh-dependent events of lamina development. The seven-pass transmembrane protein encoded by
smoothened (smo) acts as a positive effector of Hh signal reception, downstream of
the Hh receptor Patched. If Hh exerts its effects directly on LPCs, it would be expected that loss of smo function should block the entry of G1-phase LPCs into S-phase and/or
prevent the expression of Hh-dependent markers of lamina differentiation such as Dac. Inducing smo mutant clones reveals that with respect to lamina differentiation, smo acts cell autonomously.
smo clones that extended to the posterior of the lamina are
rare. It is possible that LPCs that cannot respond to Hh are not
readily incorporated into the lamina and displaced by smo+
LPCs. LPCs that are unable to respond to Hh might be eliminated by cell death (Huang, 1998).
A hallmark of Hh signal reception in many Drosophila
tissues is an increase in immunoreactivity to the C-terminal
portion of the protein Ci, a transcriptional mediator of Hh
signaling. This enhanced Ci immunoreactivity is due to inhibition of Ci proteolytic processing, a cellular
response to Hh signal reception. LPCs posterior to the lamina furrow display the enhanced Ci
immunoreactivity that would be predicted for Hh signal
reception by LPCs. In animals in which hh-
retinal axons innervate the lamina target field, cells posterior
to the lamina furrow display a level of Ci immunoreactivity
equivalent to the basal level detected anterior to the furrow, indicating that the increased Ci observed in the wild type is Hh-dependent. In smo mosaic animals, smo cells either
anterior or posterior to the lamina furrow display a basal level
of Ci immunoreactivity, while smo + cells
immediately adjacent to the portion of smo clones within the
lamina display the high Hh-dependent level.
The initial response of LPCs to the arrival of Hh-bearing
retinal axons would appear to be entry into S-phase at the
lamina furrow. To determine whether cell cycle progression is
directly dependent on Hh signal reception, the
incorporation of bromodeoxyuridine (BrdU) into S-phase cells was examined
in smo mosaic animals. In the wild type, LPCs that have
entered their terminal S-phase form a discrete and continuous
band at the posterior margin of the lamina furrow. In animals lacking photoreceptor innervation (due to defective hh expression in the eye disc) or animals
in which photoreceptor axons lacking functional Hh enter the
lamina target field, only a low background of scattered S-phase
cells are detected. It is unclear whether the products of these scattered divisions are
incorporated into the lamina (i.e., that these cells are indeed
LPCs). In smo 3 mosaic animals, mutant clones that include the
posterior margin of the lamina furrow lack S-phase LPCs. In contrast, the scattered S-phase cells anterior to the lamina furrow, and the distribution of S-phase cells in other
proliferation centers, such as the OPC, are unaffected by the
loss of smo function. At the lamina furrow, smo+ cells
bordering smo clones are often found in S-phase. Thus, in
sum, smo+ behaves as a cell-autonomous requirement for
LPCs to initiate the Hh-dependent steps of lamina differentiation (Huang, 1998).
The Hh receptor Ptc, a multiple-pass membrane protein, and
the cAMP-dependent protein kinase (PKA) normally maintain
the Hh signal transduction pathway in a repressed state.
Loss-of-function mutations in either of these genes mimic Hh
signal reception and result in the cell autonomous activation of
Hh target genes in many tissues. LPCs harboring mutations for
either pka or ptc undergo differentiation cell-autonomously
and independently of retinal innervation. Mutant cells anterior to
the furrow do not differentiate precociously. This observation
is consistent with the consequences of ectopic Hh expression
in an the lamina in mutants lacking retinal innervation of the lamina.
Hh expression in regions anterior to the lamina furrow does not
induce precocious lamina differentiation, as though competence to respond to Hh is acquired by G1-phase LPCs at the anterior margin of the lamina furrow. Within the lamina
target field, wild-type cells neighboring the pka or ptc
mutant cells are never observed to express Dac. Thus
activation of the Hh pathway by loss-of-function in either gene
results in a strictly autonomous induction of LPC maturation.
These results permit the conclusion that the terminal cell
division and differentiation of LPCs both require the direct reception of the Hh signal (Huang, 1998).
In a number of instances, pattern formation mediated by Hh is
accompanied by cell division. The well-defined pattern of Hh-induced
cell division in the lamina provides an opportunity to
determine the point at which the Hh signal reception engages
the cell cycle machinery. LPC cell cycle progression and cell fate
determination are jointly controlled by the transcriptional regulator Cubitus interruptus.
Biochemical and epistasis experiments have placed the zinc finger molecule Ci
downstream of all other hh signaling pathway components. Ci
has been shown to bind directly to the regulatory sequences of
Hh-responsive genes. Should all Hh-mediated events of LPC maturation be found to
depend on Ci function, it could be concluded that, at least with
regard to cell proliferation and the expression of differentiation
markers, there is no branchpoint within the signaling pathway. To examine the requirement for Ci, two
recombinant constructs were used that result in either dominant Ci gain-of-function or loss-of-function phenotypes. Overexpression of the wild-type Ci gene results in a gain-of-function phenotype
that mimics activation of the Hh signaling pathway. Expression of an amino terminal fragment of Ci (hereafter
referred to as DN-Ci) results in a dominant loss-of-function
phenotype, as the normal in vivo function of this portion of the
molecule appears to be transcriptional repression of Hh target
genes. With either construct, genetically engineered ectopic expression in the lamina
region results in the expected phenotype with respect to the
lamina differentiation marker Dac. Dac expression in cells
posterior of the lamina furrow is strongly reduced or
undetectable in cells expressing DN-Ci. Conversely,
the ectopic expression of wild-type Ci results in the induction
of Dac-positive cells in the lamina target field of
animals lacking innervation from the developing eye. The effects observed with either construct
are strictly cell autonomous. Thus the results with ectopic Ci and DN-Ci expression are
consistent with the expectation that Ci modulates Hh signaling activity directly in LPCs (Huang, 1998).
To determine whether Hh signaling acts via Ci to regulate
the G1- to S-phase transition of LPCs at the lamina furrow, the incorporation of BrdU into S-phase cells was examined in animals harboring clones expressing either of the
two constructs described above. Cells expressing DN-Ci at the posterior margin of the lamina furrow fail to enter S-phase. Where clones of DN-Ci-expressing cells traversed
the lamina furrow, S-phase LPCs are absent, while S-phase LPCs are observed
immediately outside of the clone. Moreover, the effect on cell division is
limited to the LPCs at the lamina furrow. No defects are observed in other proliferation zones
such as the OPC or IPC, the other major proliferation centers of the optic lobe, when they
contain DN-Ci-expressing cells. Conversely, the induction of lamina differentiation by ectopic Ci
expression in flies lacking retinal input into the lamina is accompanied by the entry of LPCs into S-phase at
the lamina furrow. At the point when lamina differentiation is induced in the absence of retinal axons by ectopic Hh expression, ectopic Ci expression triggers a posterior-to-anterior pattern of differentiation such that S-phase LPCs are found at the anterior margin. In sum, these observations indicate that the induction of cell division by Hh occurs via the transcriptional regulation of Hh target genes (Huang, 1998).
To investigate a possible involvement of synaptic machinery in Drosophila visual system development, the effects of a
loss of function of neuronal synaptobrevin (n-syb), a protein required for synaptic vesicle release, were studied. Expression of tetanus toxin light chain
(which cleaves neuronal synaptobrevin) and genetic mosaics were used to analyze neuropil pattern formation and levels of
selected neural adhesion molecules in the optic lobe. Targeted tetanus toxin light chain (TeTxLC) expression in the developing optic lobe results
in disturbances of the columnar organization of visual neuropils and of photoreceptor terminal morphology. IrreC-rst
immunoreactivity in neuropils is increased after widespread expression of toxin. In photoreceptors, targeted toxin expression results in increased Fasciclin II and
chaoptin but not IrreC-rst immunoreactivity. Axonal pathfinding and programmed cell death are not affected. In genetic mosaics, patches of photoreceptors that lack
neuronal synaptobrevin exhibit the same phenotypes observed after photoreceptor-specific toxin expression. These results demonstrate the requirement of neuronal
synaptobrevin for regulation of cell adhesion molecules and development of the fine structure of the optic lobe. A possible causal link to fine-tuning processes that
may include synaptic plasticity in the development of the Drosophila CNS is discussed (Hiesinger, 1999).
The finding of an onset of n-syb expression in the first half of pupation poses the question of whether synapses actually start to function so early during optic lobe
development. Neuronal activity plays a major role during vertebrate visual system development. A critical period of 1 d after
eclosion has been demonstrated for experience-dependent developmental plasticity in the Drosophila. It has not yet been shown
whether synaptic plasticity in the Drosophila CNS extends to pupation or whether neurotransmitters are released before any form of neuronal activity. Assuming the
involvement of such processes, the following time scale would be expected: first, expression and localization of proteins of the vesicle release machinery; second,
release of neurotransmitter independent or dependent on spontaneous activity; and third, release of neurotransmitter dependent on evoked activity. Given the early
immunoreactivity of specific synaptic vesicle cycle proteins such as n-syb and synaptotagmin before P + 25% (25% through the pupal period), the synaptic vesicle cycle appears to be available for
more than half of pupal development before first evoked photoreceptor responses occur at P + 82%. Morphological analysis has revealed a brief interval of intense
synapse formation in the lamina of Musca starting ~P + 62% and peaking at P + 74%. Although this time window does
not necessarily correspond to the first occurrence of synapses in the optic lobe of Drosophila, and the heterogeneity of optic lobe neurons should also be taken into consideration, it
may indicate that n-syb is expressed long before synapses are morphologically recognizable (Hiesinger, 1999 and references).
Apparently, not all processes between target selection and the establishment of functional connectivity are yet known. The demonstration of the dependence of
neuropil patterning on NO release shows a process of terminal development in a similar time window as the neuropil patterning
defects observed in the current study. With regard to the current study, one possibility would be the involvement of n-syb in the release of neurotransmitters or other factors before or
during synapse development. histamine is synthesized in photoreceptors extending from cultured imaginal disks. Histamine or
other substances released by growth cones after arrival in their target layers might exert functions necessary for the establishment of a regular terminal pattern (Hiesinger, 1999).
Lack of functional n-syb has no obvious influence on target selection and the development of largely overlapping terminals of R7 and R8 cells. In contrast, further
development of terminal fine structure between P + 25 and P + 50% is significantly disturbed, indicating its involvement in a fine-tuning process. This early onset of
n-syb function shows that either n-syb is involved in nonsynaptic processes taking place soon after target recognition, or synapses form earlier in the Drosophila
optic lobe than is generally believed. Because this time window occurs significantly before the observed upregulation of Fasciclin II in active toxin-expressing
photoreceptors, the morphological changes do not depend on this CAM (Hiesinger, 1999).
Cell adhesion molecules play multiple roles during optic lobe development. Most fruitfully investigated are functions during axon guidance and target recognition and synaptic plasticity. The finding of increased Fasciclin II immunoreactivity under conditions of blocked neurotransmitter release corresponds to previous studies that have shown the opposite
effect with an opposite approach: apCAM is downregulated after application of serotonin, and synaptic Fasciclin II is reduced in
mutants with abnormally high neuronal activity. Although it has been demonstrated that apCAM is downregulated via endocytosis, the mechanism of activity-dependent Fasciclin II downregulation at the Drosophila neuromuscular junction remains unknown. Possible
downregulation mechanisms to be considered include endocytosis, extracellular cleavage, and reduced transcription or translation in combination with a continuous
turnover of the protein. The upregulation of two different types of CAMs (Fasciclin II and chaoptin) in the same cell type under conditions of blocked neurotransmitter release poses the
question of the specificity of the mechanism. In the absence of functional n-syb, increased numbers of docked synaptic vesicles accumulate presynaptically. Assuming that this would result in a significant sequestration of membrane material and that the synaptic vesicle cycle is continuously
replenished from cell surfaces carrying adhesion molecules, deactivation of n-syb could result in decreased intake of CAMs and thus increased CAM
immunoreactivity. However, current understanding of the recycling mechanism in the synaptic vesicle cycle and different localization of CAM
isoforms does not support this hypothesis. Alternatively, specifically CAMs on active terminals and fibers could be
downregulated to serve as markers for the competence of the synapses for sprouting (Hiesinger, 1999).
In wild-type third instar larvae, Fasciclin II is found on R7 and R8 retinal axons. During parts of pupation, Fasciclin II is
detectable at low levels on photoreceptor cell bodies. It is possible that Fasciclin II is never completely downregulated from R7
and R8 terminals but is mostly below threshold for visualization with confocal microscopy. Upregulation of Fasciclin II levels in photoreceptors lacking functional
n-syb after P + 75% may thus be attributable to an accumulation of the protein, when its downregulation would normally occur via an n-syb-dependent mechanism
as part of a continuous protein turnover. The finding that IrreC-rst immunoreactivity remains unaltered in photoreceptors without functional n-syb but is increased in proximal neuropils after widespread
TeTxLC expression can be interpreted in two different ways: either IrreC-rst protein is not present on photoreceptor terminals at the addressed time of pupation, or
the n-syb-dependent CAM downregulation mechanism has a different molecular specificity in photoreceptors than in other optic lobe cells. During axonal pathfinding
IrreC-rst is expressed on photoreceptors. In pupal stages IrreC-rst is localized to
rhabdomeres but not to axons and cell bodies of photoreceptors during the second half of pupation. Because rhabdomeres are unique to this cell type and seem to
be a preferred localization for IrreC-rst in photoreceptors, a cell-specific distribution that excludes terminals appears more likely than a specific CAM regulation
mechanism for photoreceptors. Taken together, the results presented here clearly show a requirement of n-syb for optic lobe development. Either n-syb has a previously unknown activity-independent function, or
synaptic transmission is involved in optic lobe development, or both (Hiesinger, 1999 and references).
Information processing relies on precise patterns of synapses between neurons. The cellular recognition mechanisms regulating this specificity are poorly understood. In the medulla of the Drosophila visual system, different neurons form synaptic connections in different layers. This study identifies candidate cell recognition molecules underlying this specificity. Using RNA sequencing (RNA-seq), it was shown that neurons with different synaptic specificities express unique combinations of mRNAs encoding hundreds of cell surface and secreted proteins. Using RNA-seq and protein tagging, it was demonstrated that 21 paralogs of the Dpr family, a subclass of immunoglobulin (Ig)-domain containing proteins, are expressed in unique combinations in homologous neurons with different layer-specific synaptic connections. Dpr interacting proteins (DIPs), comprising nine paralogs of another subclass of Ig-containing proteins, are expressed in a complementary layer-specific fashion in a subset of synaptic partners. The study proposes that pairs of Dpr/DIP paralogs contribute to layer-specific patterns of synaptic connectivity (Tan, 2015).
This study defined a network of interacting Drosophila cell surface proteins in which a 21-member IgSF subfamily, the Dprs, binds to a nine-member subfamily, the DIPs. The structural basis of the Dpr-DIP interaction code appears to be dictated by shape complementarity within the Dpr-DIP binding interface. Each of the six dpr and DIP genes examined is expressed by a unique subset of larval and pupal neurons. In the neuromuscular
system, interactions between Dpr11 and DIP-γ affect presynaptic terminal development, trophic factor responses, and neurotransmission. In the visual system, dpr11 is selectively expressed by R7 photoreceptors that use Rh4 opsin (yR7s). Their primary synaptic targets, Dm8 amacrine neurons, express DIP-γ. In dpr11 or DIP-γ mutants, yR7 terminals extend beyond their normal termination zones in layer M6 of the medulla. DIP-γ is also required for Dm8 survival or differentiation. These findings suggest that Dpr-DIP interactions are important determinants of synaptic connectivity (Carrillo, 2015). This study has defined a network of interacting Drosophila IgSF CSPs in which 21 Dpr proteins bind to 9 DIPs. The structure of the Dpr-DIP complex resembles that of neural and immune cell adhesion complexes. Each of the six dpr and DIP genes examined in this study is expressed by a different subset of neurons in the larval VNC and pupal OL. In the larval neuromuscular system, Dpr11 and its binding partner DIP-γ regulate presynaptic terminal development and neurotransmission. In the pupal OL, they are required for normal formation of synapses between a Dpr11-expressing sensory neuron and a DIP-γ expressing interneuron (Carrillo, 2015).
The crystal structure shows that Dprs and DIPs belong to a group of IgSF CSPs that interact via their N-terminal Ig domains. These include immune cell receptors such as CD2, CD58, JAML, CAR, B7-1, and CTLA-4, and Nectin/Nectin-like (Necl) proteins. The nine Nectin/Necls interact with each other, forming a small network. Although DIPs resemble Nectins/Necls, their closest vertebrate counterpart is the five-member IgLON subfamily, which is also expressed in neurons. Dprs have no clear mammalian orthologs. DIPs and Dprs are distinguished from IgLONs and Nectins in that their interactions are across subfamilies, not within a subfamily. The closest structural homolog of the Dpr-DIP complex is the SYG-1-SYG-2 complex, known to be involved in synapse specification (Carrillo, 2015).
The Dpr-DIP complex has an interface involving no charge pairs, suggesting that binding specificity is encoded through shape complementarity. The Dpr-DIP interaction code may be created by substitution of larger or smaller residues within the binding interface in order to create more or less complementary surfaces between individual interacting Dpr-DIP pairs. This differs substantially from the electrostatic complementarity model, in which receptor-ligand specificity is created primarily through hydrogen bonding interactions and salt bridges. Interestingly, for Dscam homophilic interactions, where each of the many thousands of possible variants binds primarily to itself, both electrostatic and shape complementarity play crucial roles. Each Dscam variant has to find a single binding solution, which is a task that can be solved in many ways. By contrast, the complex cross-reactivity observed for Dpr-DIP interactions may impose restrictions on encoding of specificity that mandate the selection of shape complementarity as the primary mechanism (Carrillo, 2015).
The larval neuromuscular system is a genetic model system for glutamatergic synapses in mammals. In mutants lacking either Dpr11 or DIP-γ, NMJs contain many small clustered boutons called satellites. The satellite bouton phenotypes are rescued by either pre- or postsynaptic expression of the proteins. mEPSP amplitude and frequency are increased to similar extents in dpr11 and DIP-γ mutants. These data, together with the fact that the two loci genetically interact, indicate that the two proteins have linked functions, and suggest that the phenotypes are due to loss of Dpr11-DIP-γ adhesion complexes (Carrillo, 2015).
BMPs are trophic factors for mammalian neurons, and retrograde BMP signaling controls NMJ arbor growth in Drosophila. Satellites are observed in mutants in which BMP signaling is upregulated. Consistent with this, presynaptic pMad staining, which reports on the magnitude of the BMP signal, is increased in dpr11 mutants, and dpr11 and DIP-γ interact with genes encoding BMP signaling components (Carrillo, 2015).
Each dpr and DIP examined is expressed in a unique subset of neurons that project to specific layers in the OL neuropils. Identifying these neurons can define relationships between dpr/DIP expression and synaptic connectivity, because detailed synaptic maps for units of the first two areas of the OL, the La and Me, have been created using electron microscopic reconstruction (Carrillo, 2015).
Axons of UV-sensitive R7 photoreceptors synapse in layer M6 of the Me onto Dm8, Tm5a, Tm5b, and other targets. dpr11 is selectively expressed by yR7s, which express Rh4 opsin and are in ~70% of ommatidia. Dpr11 is the first cell surface protein to be associated with a subclass of R7s. DIP-γ is expressed by Dm8s, which arborize in M6 and receive more R7 synapses than any other neurons (Carrillo, 2015).
To examine whether formation of synapses between yR7s and Dm8s involves interactions between Dpr11 and DIP-γ, a marker for existing active zones, Brp-shortmCherry, was expressed in yR7s. In control animals, yR7 terminals are bulb-shaped and regularly arranged in M6. In dpr11 and DIP-γ mutants, the main bodies of yR7 terminals have altered shapes, and active zone and membrane markers are found in extensions projecting into deeper Me layers. These data suggest that synapses between yR7 and its M6 targets do not form normally in the absence of Dpr11 or DIP-γ. Because most M6-projecting DIP-γ-positive cells seen in the FLP-out analysis are Dm8s, and because Dpr11's other partner, DIP-β, does not label M6, it is infered that the loss of Dpr11 or DIP-γ is likely to primarily affect yR7-Dm8 synapses in M6 (Carrillo, 2015).
In DIP-γ mutants, there are large gaps in M6 labeling by DIP-γ or Dm8 reporters. The number of OrtC2b+, DIP-γ+ cells is reduced by >3-fold, suggesting that most DIP-γ-expressing Dm8s die. Alternatively, they might turn off expression of the OrtC2b-GAL4 driver, although this seems less likely. This effect on cell fate suggests that DIP-γ is required for reception of a neurotrophic signal. Since dpr11 mutants have no DIP-γMiMIC M6 gaps, implying that they have normal numbers of OrtC2b+, DIP-γ+ cells, this signal might be communicated through Dprs 15, 16, and/or 17, the other Dprs that bind to DIP-γ. Other OL neurons are also dependent on trophic factors for survival. R cell growth cones secrete the Jelly Belly (Jeb) ligand, which binds to its receptor Alk on L3 neurons, and L3s die in the absence of Jeb or Alk. The functions of DIP-γ in mediating normal development of yR7-Dm8 connectivity, as assayed by displacement of the active zone marker in yR7s, may be distinct from its roles in Dm8 survival, because about half of the overshoots in DIP-γ mutants appear to grow through a Dm8 arbor labeled by the DIP-γ reporter (Carrillo, 2015).
dpr11 is expressed by subsets of direction-selective T4 and T5 neurons that arborize in the Lop layers activated by front-to-back and back-to-front motion, and DIP-γ is expressed by three LPTCs, which receive synaptic input from T4s and T5s. These data suggest that Dpr11 and DIP-γ expression patterns might have evolved to facilitate assembly of synaptic circuits for specific sensory responses: near-UV vision for yR7-Dm8 connections and movement along the anterior-posterior axis for T4/T5 subset-LPTC connections. In a conceptually similar manner, a specific type of vertebrate amacrine neuron, VG3-AC, forms synapses on W3B retinal ganglion cells, which are specialized for detecting object motion. Both VG3-ACs and W3B-RGCs selectively express the IgSF protein Sidekick2 (Sdk2), and Sdk2-mediated homophilic adhesion is required for their connectivity (Carrillo, 2015).
An accompanying paper on gene expression in La neurons (Tan, 2015) presents ten instances in which a La neuron expressing a Dpr is synaptically connected to a Me neuron expressing a DIP to which that Dpr binds in vitro. In nine of these, as well as in the two cases described in this study (yR7 -> Dm8 and T4/T5 -> LPTC), the Dpr is in the presynaptic neuron and the DIP in the postsynaptic neuron. Each dpr and DIP gene examined in the two papers is expressed in a different subset of OL neurons, each of which projects to a distinct set of neuropil layers, and neurons can express multiple Dprs or DIPs or a combination of the two (Tan, 2015). This means that there are hundreds of different synaptic matches in the OL that could potentially be programmed by the Dpr-ome network. Dprs and DIPs are also expressed by subsets of neurons in other areas of the larval and pupal brain. These expression patterns, together with the phenotypic data presented here for one Dpr-DIP binding pair, suggest that Dpr-DIP interactions are likely to be important determinants of synaptic connectivity during brain development (Carrillo, 2015).
Neurons form precise patterns of connections. The cellular recognition mechanisms regulating the selection of synaptic partners are poorly understood. As final mediators of cell-cell interactions, cell surface and secreted molecules (CSMs) are expected to play important roles in this process. To gain insight into how neurons discriminate synaptic partners, the transcriptomes were profiled of 7 closely related neurons forming distinct synaptic connections in discrete layers in the medulla neuropil of the fly visual system. The sequencing data revealed that each one of these neurons expresses a unique combination of hundreds of CSMs at the onset of synapse formation. 21 Ig domain paralogs of the defective proboscis extension response (see Drosophila Piecing Together the Extracellular Puzzle) family were shown to be expressed in a unique cell-type-specific fashion, consistent with the distinct connectivity pattern of each neuron profiled. Expression analysis of their cognate binding partners, the 9 members of the Dpr interacting protein (DIP) family, revealed complementary layer-specific expression in the medulla, suggestive of interactions between neurons expressing Dpr and those expressing DIP in the same layer. Through coexpression analysis and correlation to connectome data, neurons expressing DIP were identified as a subset of the synaptic partners of the neurons expressing Dpr. It is proposed that Dpr-DIP interactions regulate patterns of connectivity between the neurons expressing them (Marta, 2016).
Binding between DIP and Dpr neuronal
recognition proteins has been proposed to regulate synaptic connections
between lamina and medulla neurons in the Drosophila visual system. Each
lamina neuron was previously shown to express many Dprs. This study
demonstrates, by contrast, that their synaptic partners typically
express one or two DIPs, with binding specificities matched to the
lamina neuron-expressed Dprs. A deeper understanding of the molecular
logic of DIP/Dpr interaction requires quantitative studies on the
properties of these proteins. A quantitative affinity-based DIP/Dpr
interactome was generated for all DIP/Dpr protein family members. This
revealed a broad range of affinities and identified homophilic binding
for some DIPs and some Dprs. These data, along with full-length
ectodomain DIP/Dpr and DIP/DIP crystal structures, led to the
identification of molecular determinants of DIP/Dpr specificity. This
structural knowledge, along with a comprehensive set of quantitative
binding affinities, provides new tools for functional studies in vivo (Cosmanescu, 2018).
Brains from flies to humans comprise vast numbers of different types of neurons interconnected by networks of precisely patterned synaptic connections. Currently, the molecular mechanisms underlying the specification of neural circuit assembly are poorly understood. The predominant model, based on Roger Sperry's 'chemoaffinity hypothesis,' postulates that neurons make specific connections with their targets based on interactions between specific cell surface molecules. Cell-cell recognition proteins are often members of families diversified in evolution by gene duplication to yield numerous members, each bearing a canonical binding interface characteristic of the family. For such protein families, binding between members is often promiscuous, and it is the distinctive strength of binding, or binding affinity, that underlies the differential biological functions of each protein. Understanding the logic underlying the patterning of neural networks will require determination of the binding affinities of cell recognition proteins, their expression patterns, their signaling properties, and gain- and loss-of-function genetic analyses (Cosmanescu, 2018).
In Drosophila, two families of cell-recognition proteins of the immunoglobulin superfamily (IgSF), the 21-member Dpr (defective proboscis extension response) and the 11-member DIP (Dpr interacting proteins) families, have many of the properties expected of proteins controlling synaptic specificity. Members of each family are expressed in subsets of neurons throughout the developing nervous system (Carrillo, 2015, Tan, 2015). Within the Drosophila visual system, the five lamina monopolar neurons, L1-L5, as well as the R7 and R8 photoreceptor cells each express unique combinations of Dpr proteins. Cognate DIPs were found to be expressed in some of their synaptic partners in the medulla, suggesting a potential role in synaptic targeting (Carrillo, 2015, Tan, 2015). It remains unclear from these earlier studies, however, whether each medulla neuron type expresses many DIPs as observed for Dpr expression in lamina neurons or only a more limited repertoire of them (Cosmanescu, 2018).
In the accompanying paper, single Dm12 neurons with DIP-α null mutations exhibit robust defects in target-layer specificity in a wild-type background, and misexpression of cognate Dpr ligands dramatically re-specifies these connections (Xu, 2018). In addition to targeting phenotypes, DIP/Dpr interactions also play a role in cell survival. Loss of DIP-γ (Carrillo, 2015) as well as its binding partner Dpr11 in R7 neurons (Xu, 2018) leads to a reduction in the number of Dm8 neurons. In both cases, cell loss results from apoptosis during development (Xu, 2018), consistent with the idea that DIP/Dpr interactions may influence the regulation of apoptosis. The number of DIP and Dpr paralogs, their patterns of expression within the brain, and the complexity of the DIP/Dpr interactome allude to a widespread and complex role in patterning neural circuitry (Cosmanescu, 2018).
High-throughput in vitro binding experiments using an ELISA-based assay revealed a heterophilic interaction network between members of the two families, where all but two members of the DIP family were found to interact with individual or subsets of Dprs. Although such assays are effective at identifying heterophilic binding, technical constraints of the method often select against the detection of homophilic interactions. Furthermore, these assays utilized multimerized chimeras to increase binding affinities so as to enable robust detection; as a consequence, however, this method inherently obscures the native molecular binding affinities, yielding binary results that provide a yes or no answer as to whether an interaction takes place (Cosmanescu, 2018).
Do binding affinities of adhesion proteins significantly impact interactions between cells? Differential affinities can have clear effects on signaling between adherent cells: for example, T cells bearing receptors with different affinities for peptide-major histocompatibility complex (MHC) complexes on antigen-presenting cells adopt different developmental fates. With respect to selectivity of cellular interactions, type I classical cadherin family proteins provide a typical example of the role of affinity: each type I cadherin family member binds to all other type I family members, yet the differences in affinity of each pairwise interaction dictate their distinct adhesive and cell-patterning functions. Thus, for protein families with promiscuous binding, in which selectivity is dictated mainly through the differential pairwise binding affinities of different family members, quantitative measures are required to understand their function (Cosmanescu, 2018).
In the nervous system, binding affinities of cell-cell recognition proteins have been shown to control the targeting of neurites to their appropriate partners. For example, members of the two-protein family of Ig-like sidekick (Sdk) proteins are expressed in specific layers within the inner plexiform region of the mouse retina during synapse formation. In vitro, Sdk1 and Sdk2 bind heterophilically through a canonical interface, but their homophilic affinities are stronger. Despite their heterophilic binding, the higher affinity of the respective homophilic interactions appears to determine their synaptic targeting activities. By contrast, within this same region of the retina, the type II cadherin family members cadherin-8 and cadherin-9, which show distinctive heterophilic and homophilic affinities to other type II cadherin family members, appear to rely on heterophilic rather than homophilic binding for proper layer-specific targeting. Thus, differential molecular binding affinities of both cadherins and Ig superfamily proteins contribute to synaptic patterning (Cosmanescu, 2018).
DIP-Dpr binding specificity is controlled by interactions between their immunoglobulin-like extracellular domains. The extracellular regions of Dpr family members consist of two tandem Ig-like domains, and the extracellular region of DIP family members consists of three tandem Ig-like domains. The crystal structure of a two-domain fragment of DIP-α in complex with the membrane-distal Ig1 domain of Dpr6 revealed the Ig1-Ig1 interaction to be characterized by a buried core of hydrophobic residues and an extensive network of hydrogen bonds (Carrillo, 2015). The interaction topology of this complex shares a strong resemblance to other complexes of Ig-like cell adhesion molecules, including those of vertebrate nectins and C. elegans SYGs, both of which have roles in nervous system development. Interestingly, members of the nectin and SYG-related protein families exhibit both homophilic and heterophilic binding (Cosmanescu, 2018).
As a step toward understanding how DIP and Dpr protein families contribute to neural circuit assembly, this study sought to extend understanding of both the binding affinities of DIPs/Dprs and the neuron-specific localization of DIPs in the Drosophila visual system. The multi-color flip out (MCFO) technique was used to provide a more extensive map of DIP expression in the medulla. To assess the biophysical properties of interactions between protein family members, surface plasmon resonance (SPR) was used to determine binding affinities for all DIP-Dpr interactions, identified DIPs and Dprs that form homodimers, and identified specificity-determining residues in DIP-Dpr interfaces that had not previously been noticed. This new knowledge was used to design site-directed mutants with defined intermolecular binding affinities for in vivo functional experiments reported in the accompanying paper (Xu, 2018). Biophysical studies raise the intriguing possibility that DIP/Dpr interactions function over a wide range of affinities to regulate neural circuit assembly throughout the Drosophila nervous system (Cosmanescu, 2018).
Developing axons and dendrites encounter the processes of perhaps hundreds of different neuronal cell types and must select appropriate synaptic partners from a myriad of neuronal processes. RNA sequencing technologies have revealed that developing neurons express hundreds of cell surface proteins, many of which bind in vitro to proteins known to be expressed on neighboring cells. Identifying which interactions are important and understanding how their expression patterns and binding interactions contribute to the specificity, complexity, and function of neural circuits remains a central challenge in developmental neuroscience (Cosmanescu, 2018).
Families of cell surface proteins with related ectodomains and differences in binding specificity provide one way of generating diverse patterns of connectivity. As opposed to Dscams and protocadherins, which are expressed stochastically to provide neurons with single cell identities that form the basis of self-avoidance, this study envisions that selective recognition between synaptic partners relies on deterministic mechanisms of gene regulation to ensure the appropriate cell-type-specific pairing of ligands and receptors. Indeed, it is the deterministic expression of matching DIP/Dpr pairs in some synaptic partners in the visual system that led to the idea that DIP/Dpr interactions might influence synaptic specification (Carrillo, 2015; Tan, 2015). A previous study demonstrated through mRNA sequencing and genetic tagging methods that Dprs were expressed in a dynamic and complex way in developing lamina neurons (Tan, 2015). Each lamina neuron expresses a discrete combination of numerous Dprs. It was also shown that some synaptic partners of lamina neurons, specific medulla neurons, express cognate-interacting DIP proteins. This study extended these observations through a systematic analysis of eight of the 11 DIPs using the MCFO technique. Of the 60 neuronal cell types analyzed, 26 expressed a single DIP, 12 expressed two DIPs, and one expressed four DIPs (i.e., 39/60 or 65% of the neurons express at least one of the eight DIPs). Assuming the remaining three DIPs, for which gene-trap GAL4s are not yet available (i.e., DIP-ι, -κ, and -λ), are expressed in a similar fashion, it is estimated some ~90% of the 60 different medulla neuron types considered in this study express one or, less frequently, two DIPs (Cosmanescu, 2018).
By comparing the synaptic connectivity maps between lamina and medulla neurons, the expression patterns of DIPs and Dprs, and the DIP/Dpr interactome, many DIP/Dpr pairs expressed in synaptic partners were identified. Lamina neurons form synapses on many different medulla neuron types; for instance, lamina L3 neurons express many Dprs and form synapses with over 10 different medulla neurons, many of which express DIPs that bind to Dprs expressed in L3. It appears then that lamina neuron outputs diverge to synapse with multiple partners. By contrast, medulla neurons express a more limited set of DIPs. For instance, Dm4 neurons only express DIP-α and form synapses with on the order of 20 L3 neurons, which express, among other Dprs, Dpr6 and Dpr10, high-affinity ligands for DIP-α. L3 is by far the predominant input to Dm4. The inputs into Dm4, therefore, are convergent. Indeed, information from multiple lamina neurons of the same type frequently converge onto a single DIP-expressing Dm neuron (Cosmanescu, 2018).
A clear pattern emerges whereby multiple Dprs on lamina neurons may promote connections to multiple targets, whereas a single DIP expressed on Dm neurons, for instance, accommodates convergence of many different neurons of the same type onto a single partner. Overall, this arrangement mirrors the interactome, where a single type of DIP tends to interact with high affinity to multiple Dprs, but in general a single Dpr exhibits high-affinity binding to one type of DIP. A similar trend is seen with both Tm and TmY neurons; they typically form connections with more different types of neurons than Dm, but fewer than lamina neurons. Interestingly, about half of the Tm and TmY neurons analyzed (10/21) express more than one DIP, whereas only one of 18 Dm neurons expressed more than one DIP (Cosmanescu, 2018).
Quantitative biophysical and structural studies enabled the identification of residues in DIPs and Dprs that control their binding specificity. Grouping DIPs and Dprs according to their cross-family binding interactions facilitated the identification of resides at positions in the sequence that were correlated with the binding preferences of different specificity groups. Most of these specificity residues are charged or polar in contrast to the conclusion of Carrillo (2015) that shape complementarity was the dominant determinant of inter-subgroup specificity. As discussed above, part of the discrepancy is due to their focus on the hydrophobic core of the interface although most distinct specificity determinants are located in the periphery (Cosmanescu, 2018).
The specificities of DIP-Dpr interactions are partially overlapping and grouped by phylogeny, with interaction affinities spanning approximately two orders of magnitude. Three main DIP affinity groups and DIP-δ, which forms a one-member group, emerge with cognate Dpr interactions mainly falling within a single DIP group, with sparser and weaker interactions between groups. These groupings became clear only when binding affinities were incorporated and false positive and negative interactions removed (e.g., removal of Dpr16/Dpr17 with DIP-ε and addition of DIP-κ and DIP-λ interactions). Quantitative binding affinities were also crucial for assigning the 'primary' DIP-binding specificities (the DIP[s] with highest interaction affinity) for groups of Dprs, which was used in the identification of specificity determinants. Dprs with similar binding preferences are closely related with a few exceptions, and DIPs within each of the three main groups are also close in phylogeny, with the exception of DIP-γ and DIP-κ. Indeed, single mutants in dpr6 and dpr10, which are phylogenetic nearest neighbors with similar DIP-binding profiles, show weaker phenotypes than null mutations, inactivating their common binding partner, DIP-α (Cosmanescu, 2018).
Like other families of cell surface proteins with related ectodomains, DIPs and Dprs bind through canonical interfaces common to all family members. Because interactions between members of such diversified protein families rely on a common binding mode, many family members might be expected to bind one another, albeit with different affinities. Thus, DIP and Dpr proteins engage in promiscuous interactions, as has also been observed for other protein families implicated in targeting, e.g., type I and type II cadherins, sidekicks, nectins, synCAMs, and Drosophila irre cell recognition module (IRM) proteins. The binding properties of these protein families differ significantly from the strict homophilic recognition observed for stochastically expressed multi-domain repulsion proteins (i.e., Dscam and clustered protocadherins). These achieve recognition only when all interacting domains are matched with their cognate partners, leading to an all-or-none binding specificity. Multi-domain interfaces may be required to achieve precise fine-tuning to avoid the promiscuity that is characteristic of two-domain interfaces. In contrast, wide-ranging affinities in protein families, such as Dprs and DIPs, may be exploited by developing neurons to sculpt neural circuitry in different ways (Cosmanescu, 2018).
The demonstration that some DIPs and Dprs form homodimers adds another layer to the potential regulatory complexity of interactions between these proteins. DIP homodimerization affinities are in the range of 22-35 μM, with Dpr homodimerization affinities ranging from 39 to 71 μM. The homodimerization affinity of a DIP can be significantly weaker than with its heterophilic binding to Dpr partners (DIP-α), equivalent to the strongest heterophilic interactions of its group (DIP-ζ), or stronger than its heterophilic interactions (DIP-η). For Dprs, in each case the homodimer affinities that were determined were substantially weaker than their heterophilic DIP interactions. Crystal structure and mutational analyses reveal that DIP/DIP and DIP/Dpr interfaces are largely overlapping. The Dpr/Dpr dimer structure has not yet been determined. Although AUC was used to identify homodimers, in principle, heterophilic DIP-DIP and Dpr-Dpr interactions could also form, though this study did not seek to identify such potential interactions. Indeed, Özkan, 2013, detected Dpr3-Dpr7 and Dpr5-Dpr6 heterophilic interactions in their high-throughput interaction study (Cosmanescu, 2018).
In principle, some DIPs and Dprs could function in cell-cell recognition driven by homophilic rather than heterophilic interactions. In support of this possibility, genetic rescue studies indicate that, in some contexts, homophilic interactions can substitute for heterophilic binding. For example, DIP-α overexpression in DIP-α-interacting neurons reduces Dm4 cell loss by apoptosis in Dpr6/10 null mutants (Xu, 2018). In some contexts, competition between homophilic and heterophilic binding partners could play a regulatory role in controlling interactions between neurons, as has been suggested for Sdks and nectins. Interestingly, germline knockin mutants of a homophilic binding-deficient form of DIP-α designed in this study led to a 50% increase in synapse number for Dm4 neurons. These findings are consistent with the notion that complex regulatory roles may modulate DIP/Dpr interactions during circuit assembly, and these, in turn, may regulate cell number and neuronal morphogenesis, as well as the distribution, number, and specificity of synaptic connections (Cosmanescu, 2018).
Altogether, these findings provide a firm biophysical basis for the exploration, through genetic analysis, of the role of DIP/Dpr interactions in neural circuit assembly. Moving forward, it is now possible to design DIP and Dpr mutants that abrogate, increase, or decrease homophilic and heterophilic interactions so as to allow a detailed exploration of the role of binding affinities in neural circuit assembly (Cosmanescu, 2018).
Neurons in the developing brain express many different cell adhesion molecules (CAMs) on their surfaces. CAM-binding affinities can vary by more than 200-fold, but the significance of these variations is unknown. Interactions between the immunoglobulin superfamily CAM DIP-α and its binding partners, Dpr10 and Dpr6, control synaptic targeting and survival of Drosophila optic lobe neurons. This study designed mutations that systematically change interaction affinity and analyze function in vivo. Reducing affinity causes loss-of-function phenotypes whose severity scales with the magnitude of the change. Synaptic targeting is more sensitive to affinity reduction than is cell survival. Increasing affinity rescues neurons that would normally be culled by apoptosis. By manipulating CAM expression together with affinity, this study shows that the key parameter controlling circuit assembly is surface avidity, which is the strength of adherence between cell surfaces. It is concluded that CAM binding affinities and expression levels are finely tuned for function during development (Xu, 2022).
In the developing Drosophila visual system, glia migrate into stereotyped positions within the photoreceptor axon target fields and provide positional information for photoreceptor axon guidance. Conversely, glial migration depends on photoreceptor axons, as glia precursors stall in their progenitor zones when retinal innervation is eliminated. These results support the view that this requirement for retinal innervation reflects a role of photoreceptor axons in the establishment of an axonal scaffold that guides glial cell migration. Optic lobe cortical axons extend from dorsal and ventral positions toward incoming photoreceptor axons and establish at least four separate pathways that direct glia to proper destinations in the optic lobe neuropiles. Photoreceptor axons induce the outgrowth of these scaffold axons. Most glia do not migrate when the scaffold axons are missing. Moreover, glia follow the aberrant pathways of scaffold axons that project aberrantly, as occurs in the mutant dachsous. The local absence of glia is accompanied by extensive apoptosis of optic lobe cortical neurons. These observations reveal a mechanism for coordinating photoreceptor axon arrival in the brain with the distribution of glia to multiple target destinations, where they are required for axon guidance and neuronal survival (Dearborn, 2004).
Attempts were made to determine the location of progenitors that give rise to the distinct types of migratory glia and the neurons that form their migratory pathways. The Wingless expressing cells of the dorsal and ventral domains are located in areas of complex gene expression controlled by Wingless (Wg) signaling activity. Adjacent
to the Wg domains are non-overlapping cell populations that express the
TGF-ß family member Decapentaplegic (Dpp). Both the Wg- and
Dpp-positive cell populations express the transcription factor Optomotor Blind (Omb). Dachsous (Ds), a Cadherin family member, is expressed in a graded fashion with respect to the Wg
domains. These three genes, though expressed in different patterns, are under the control of Wg activity (Dearborn, 2004 and references therein).
It is concluded that Drosophila optic lobe glia use axon
fascicles as migratory guides and that the extension of these axon fascicles is induced by the ingrowth of photoreceptor axons from the developing retina. The migratory
scaffold axons emerge from optic lobe regions that are in close proximity to sites where glial cells originate; both arise in the dorsal and ventral domains where cells express the morphogen Wingless.
When the scaffold axons were eliminated by the autonomous expression of an activated Ras transgene, glia failed to migrate and stalled at the borders of their progenitor sites. Extensive cortical cell apoptosis ensued. When the scaffold axons projected aberrantly (in animals mutant for the cadherin Dachsous), glia followed the
aberrant routes to incorrect destinations. The longstanding observation that glial migration does not occur in eyeless mutant Drosophila might thus be explained by an indirect mechanism in which innervation controls the establishment of an axon scaffold necessary to direct glial migration (Dearborn, 2004).
The Drosophila visual system consists of the compound eyes and the
optic ganglia in the brain. Among the eight photoreceptor (R) neurons, axons
from the R1-R6 neurons stop between two layers of glial cells in the lamina,
the most superficial ganglion in the optic lobe. Although it has been
suggested that the lamina glia serve as intermediate targets of R axons,
little is known about the mechanisms by which these cells develop. DPP signaling has been shown to play a key role in this process. dpp is expressed
at the margin of the lamina target region, where glial precursors reside. The
generation of clones mutant for Medea, the DPP signal transducer, or
inhibition of DPP signaling in this region results in defects in R neuron
projection patterns and in the lamina morphology; these defects are caused by defects in the differentiation of the lamina glial cells. glial cells
missing is expressed shortly after glia precursors start to
differentiate and migrate. Its expression depends on DPP; gcm is
reduced or absent in dpp mutants or Medea clones, and
ectopic activation of DPP signaling induces ectopic expression of gcm
and Repo. In addition, R axon projections and lamina glia development are
impaired by the expression of a dominant-negative form of gcm,
suggesting that gcm indeed controls the differentiation of lamina
glial cells. These results suggest that DPP signaling mediates the maturation
of the lamina glia required for the correct R axon projection pattern by
controlling the expression of gcm (Yoshida, 2005).
dpp is expressed in the dorsal and ventral margins of the
posterior region of the optic lobe, adjacent to the cells expressing
wg, which induces dpp expression. Glial cells in the
lamina target region arise from these regions and migrate into the lamina
target region as they contact R axons. Axons from R1-R6
neurons stop between two rows of glial cell layers, the epithelial and
marginal layers, and form the lamina plexus. The third row of glial cells, the medulla glia, is located just beneath the marginal glia. The homeodomain
protein Repo is expressed in these glial cells (Yoshida, 2005).
The expression pattern of dpp-lacZ, an enhancer-trap
allele of dpp, was compared with the expression pattern of Repo. At a stage prior to glia differentiation and migration, expression of the dpp reporter is detected in the dorsal and ventral margins of the lamina target region. dpp continues to be expressed at the margins of the lamina target region throughout the third larval instar (Yoshida, 2005).
wg at the posterior-most domain induces the expression of
dpp and omb. Some wg-expressing cells extend projections
towards the lamina target region. These cells extend scaffold axons along
which the lamina glia migrate. Thus, it was possible that the wg signal is involved in the migration and/or differentiation of lamina glia. However, partial elimination of Wg activity with a wgts allele does not cause a specific defect in glia migration.
Therefore, wg may play a role in organizing domains in the visual
cortex by activating/repressing various genes, rather than contributing to the
generation of specific cell types (Yoshida, 2005).
Medea is required for lamina glia development.
Medea encodes a co-SMAD and mediates a range of DPP/BMP/TGFß
signaling events. In addition to dpp, four related genes --
glass bottom boat (gbb), screw, activin and
activin2 -- have been identified in Drosophila. GBB signals
through TKV/Saxophone (SAX) and Wishful Thinking (WIT) type I and type II
receptors, respectively. Activin uses Baboon as a type
I receptor, and Punt and WIT as type II receptors. Brains mutant for gbb and wit were examined, but no defects in lamina glia development were observed. It is concluded that it is highly likely that
dpp is the ligand responsible for lamina glia development. However,
the possibility that one or more of the DPP-related ligands
acts redundantly in this process cannot be excluded (Yoshida, 2005).
In the embryo, gcm initiates the specification of glial cells from
neural cells of various lineages. gcm expression is strictly
controlled to ensure the correct separation of glial versus neuronal cell
fate. Analysis of the cis-regulatory elements of gcm
suggests that gcm expression depends on multiple regulatory elements
to allow the control of lineage-specific transcription and autoregulation. The analysis carried out in this study suggests that a different situation exists in the optic lobe; gcm is expressed in the glia and the lamina neuronal cells, and is required for the differentiation of these cell types. In addition, differentiation is controlled differently in the lamina and in the glia. In the lamina, gcm expression seems to be controlled by hh, and in the glia, by dpp. These results suggest that gcm is controlled and functioning in a different manner in the optic lobe. Uncovering the mechanisms of the control and function of gcm would probably prove an intriguing focus for future research (Yoshida, 2005).
DPP and its vertebrate homolog BMP play crucial roles in many aspects of
development by controlling patterning, cell growth and differentiation. This
analysis reveals a role for DPP signaling in lamina glia differentiation in
the Drosophila visual system. DPP has also been reported to function
in several aspects of visual center development; for instance, DPP signaling
has been shown to be involved in the proliferation and migration of the
subretinal glia in eye disc development, which plays an important role in the
R axon navigation. In addition, defects have been reported in the medulla neuropile in dpp mutant animals, suggesting a role for dpp in neuronal fate specification. Furthermore, tkv is expressed in lamina precursor cells just ahead of the lamina furrow, where these cells meet R axons and start to differentiate. Although this possibility is one of the
things that prompted an examination of the role of DPP signaling in lamina
development, no defects were uncovered when Mad or
Medea clones were generated in the OPC or the lamina. Moreover,
dpp appears to be expressed in the inner proliferation center (IPC),
which will form the lobula, in addition to its expression in the dorsal and
ventral marginal domains. Thus, dpp may be required for some aspects
of lobula development. Unfortunately, this cannot be easily addressed at this
moment because of a lack of appropriate markers. Further study of the
requirements for dpp in the lamina, the medulla, the lobula and other
cell types could lead to a more comprehensive understanding of how DPP
signaling controls differentiation and other events during development of the
visual system (Yoshida, 2005).
Coordinated development of neurons and glia is essential for the establishment of neuronal circuits during embryonic development. In the developing Drosophila visual system, photoreceptor (R cell) axons and wrapping glial (WG) membrane extend from the eye disc through the optic stalk into the optic lobe. Extensive studies have identified a number of genes that control the establishment of R-cell axonal projection pattern in the optic lobe. The molecular mechanisms directing the exit of R-cell axons and WG membrane from the eye disc, however, remain unknown. This study shows that integrins are required in R cells for the extension of R-cell axons and WG membrane from the eye disc into the optic stalk. Knockdown of integrins in R cells but not WG caused the stalling of both R-cell axons and WG membrane in the eye disc. Interfering with the function of Rhea (i.e. the Drosophila ortholog of vertebrate talin and a key player of integrin-mediated adhesion), caused an identical stalling phenotype. These results support a key role for integrins on R-cell axons in directing R-cell axons and WG membrane to exit the eye disc (Ren, 2022).
glia display diverse morphologies, both across and within glial classes. This study explored the relationship between glial morphology and transcriptional signature using the Drosophila CNS, where glia are categorised into 5 main classes (outer and inner surface glia, cortex glia, ensheathing glia, and astrocytes). Single-cell RNA sequencing data of Drosophila glia were analysedin 2 well-characterised tissues from distinct developmental stages containing distinct circuit types: the embryonic ventral nerve cord (VNC) (motor) and the adult optic lobes (sensory). This analysis identified a new morphologically and transcriptionally distinct surface glial population in the VNC. However, many glial morphological categories could not be distinguished transcriptionally, and indeed, embryonic and adult astrocytes were transcriptionally analogous despite differences in developmental stage and circuit type. While extensive within-class transcriptomic diversity was detected for optic lobe glia, this could be explained entirely by glial residence in the most superficial neuropil (lamina) and an associated enrichment for immune-related gene expression. In summary, this study generated a single-cell transcriptomic atlas of glia in Drosophila, and this extensive in vivo validation revealed that glia exhibit more diversity at the morphological level than was detectable at the transcriptional level (Lago-Baldaia, 2023).
Glial cells are essential for proper nervous system development and function. To understand glial development and function, glial cells were comprehensively annotated in a single-cell mRNA-sequencing (scRNAseq) atlas of the developing Drosophila visual system. This allowed study of their developmental trajectories, from larval to adult stages, and to understand how specific types of glia diversify during development. For example, neuropil glia that are initially transcriptionally similar in larvae, split into ensheathing and astrocyte-like glia during pupal stages. Other glial types, such as chiasm glia change gradually during development without splitting into two cell types. The analysis of scRNA-seq led to the discovery that the transcriptome of glial cell bodies can be distinguished from that of their broken processes. The processes contain distinct enriched mRNAs that were validated in vivo. Therefore, this study have identified most glial types in the developing optic lobe and devised a computational approach to identify mRNA species that are localized to cell bodies or cellular processes (Ferreira, 2023).
The formation of neuronal connections requires the precise guidance of developing axons toward their targets. In the Drosophila visual system, photoreceptor neurons (R cells) project from the eye into the brain. These cells are grouped into some 750 clusters comprised of eight photoreceptors or R cells each. R cells fall into three classes: R1 to R6, R7, and R8. Posterior R8 cells are the first to project axons into the brain. Using a microarray-based molecular screen as a starting point, this study identified the early and transient expression of Robo3 in R8 growth cones. Loss of Robo3 demonstrated a specific axon guidance choice point at an early stage of optic lobe innervation. In the absence of Robo3, posterior R8 growth cones inappropriately extend across Slit-expressing glial cells joining axon fascicles of the C+T lobula neurons, instead of remaining alongside the glial process as they extend into the lamina. This early repulsive function of Robo3 plays a crucial role in segregating axons and thereby contributes to the orderly assembly of columnar units comprising the fly visual system (Pappu, 2011).
The microarray data, coupled with antibody staining and the identification of a robo3-Gal4 enhancer trap, identified Robo3 as an R8-specific guidance receptor. Robo3 expression is transient in the R8 growth cone and prolonging Robo3 expression in R8 axons results in defects in R8 targeting. The microarray analysis suggests that restricted Robo3 expression during early stages of R8 differentiation occurs downstream of the transcription factors Sens and Run. However, the expression of both Sens and Run persists beyond the expression of Robo3. For example, during mid- to late-pupal development, Sens regulates both targeting of R8 axons to their final target layer in the medulla and the expression of R8-specific opsins. Therefore, other mechanisms must exist to control the expression of Robo3 in R8 (Pappu, 2011).
The importance of the tightly regulated expression of Robo receptors is emerging as a central theme in axon guidance. Indeed, previous studies have revealed a set of discrete posttranslational mechanisms controlling Robo functions both in vertebrates and invertebrates. For example, commissural axons in the fly embryo express Robo1 transcripts before crossing the midline, but Robo1 protein in these axons is sequestered into endosomes by the action of Commissureless protein, thereby preventing precocious repulsion from the midline and, thus, allowing these axons to cross. Subsequent up-regulation of Robo1 prevents them from recrossing. In vertebrates, alternative splicing of a divergent Robo receptor Rig1/Robo3, perhaps coupled with translational regulation, governs the switch from midline attraction to repulsion (Pappu, 2011).
The regulated expression of Robo3 in the R8 photoreceptors is similar to its expression in the chordotonal neurons in the embryonic peripheral nervous system. Sens is activated downstream of Ato in both chordotonal and in R8 neurons, suggesting that a conserved transcriptional program regulates Robo3 expression in these neurons. In a broader sense, these findings raise the possibility that conserved regulatory cassettes exist, which link specific transcriptional hierarchies controlling neuronal differentiation with specific constellations of downstream guidance receptors controlling wiring specificity (Pappu, 2011).
Posterior-most R8 neurons face at least three different guidance choices as they extend from the eye disk into the developing optic lobe. These early choices have a profound effect on later aspects of visual system assembly. First, R8 axons must navigate to the posterior of the eye disk and enter the optic stalk. This process is facilitated, in part, by retinal basal glial cells at the posterior edge of the eye disk. If glial cells are displaced anteriorly, R-cell axon fascicles project away from the optic stalk rather than toward it. Although it seems likely that this directional choice relies upon R8, it is not known whether posterior growth requires R8-specific functions or whether all retinal neurons are endowed with this function (Pappu, 2011).
Second, R8 axons from each ommatidium must possess molecular mechanisms to retain their individuality. As the R8 axons extend down the optic stalk they form a tight fascicle. Fasciculation is transient, however, because R8s defasciculate as they exit the optic stalk. R8 defasciculation relies on two cell surface receptors, Flamingo (Fmi) and Golden Goal (Gogo) that are expressed in the R8 growth cones as they exit the optic stalk. Fmi and Gogo mediate repulsive interactions between R8 axons, and thus play a key role ensuring that columns remain as separate modules. These repulsive interactions between R8 axons of adjacent columns also explain why axons from later-born (anterior) R8 neurons are not affected in robo33 mutant optic lobes; only the posterior R8 axons traverse through the optic lobe with access to the glial cell boundaries that separate them from the C+T lobular neuron axons (Pappu, 2011).
Third, in this article it was demonstrated that posterior R8 axons rely upon Robo3 to prevent inappropriate fasciculation with C+T lobula neurons. This process requires early, transient, and specific expression of Robo3 in R8 growth cones and is likely to require the reciprocal expression of Slit in the glial cells that these posterior R8 axons encounter when they enter the optic lobe. Thus, the posterior R8 axons are unique because they navigate a choice point that is not encountered by later arriving, more anterior R8 axons. The robo33 mutant phenotype described in this study is reminiscent of the loss of another Ig receptor encoded by the irregular chiasm/roughest (irre-C) locus. Whether IrreC acts in the same molecular pathway as Robo3 and, indeed, whether it acts in photoreceptor growth cones or lamina neurons is not known (Pappu, 2011).
In summary, Fmi, Gogo, and Robo3 play crucial roles in R8s in regulating fascicle organization, which provides the structural basis for columnar organization of the visual system. Although Fmi and Gogo mediate interactions between axons of the same class of cells (R8s), Robo3 prevents axons from one class of neurons (R8s) from inappropriately associating with a different class of axons (C+T lobular neurons) projecting into the same neuropil along a different pathway. Given the cellular complexity of columns (e.g., medulla columns comprise more than 50 axons from many different neuronal subclasses) and the stereotyped organization of axons and synaptic connections within them, it is speculated that many additional cell-surface proteins must act in a coordinated fashion in space and time to promote the orderly assembly of columnar units (Pappu, 2011).
Does Robo3 function in the R8 rely on Slit? This is indeed the most parsimonious model for Robo3 function in R8. Slit expression is detected around glial cells separating the posterior R8 growth cones from C+T lobula neurons. Although R8 projection defects seen in slit mutants are similar to those seen in robo3 mutants, they are more severe. In contrast to robo3, slit mutant optic lobes are extremely disorganized, arguing that Slit has a broader role in neuropil organization. Indeed, mutants deficient in all three Robo proteins exhibit cell migration defects, which largely phenocopy the loss of Slit. It has been proposed that Slit provides a repellent function in the optic lobe preventing cell migration between cell populations in the lamina and lobula. However, the residual Robo3 function in the robo31 hypomorphic allele available for study at that time masked the robo3 phenotypes uncovered in this study. Thus, although slit mutants uncover a broader role for Slit-Robo signaling in many aspects of optic lobe development and patterning, the unique robo3 mutants described in this study uncover a specific role for the Robo3 receptor in R8 axon guidance (Pappu, 2011).
The repulsive role of Robo3 in R8 neurons in response to locally secreted Slit is proposed in this study to be analogous to the role of Robo1 in the guidance of ipsilateral longitudinal pioneer axons in the ventral nerve cord. Robo1 is expressed on the growth cones of ipsilateral pioneer axons and prevents these axons from crossing the midline in response to Slit secreted from midline glia. In contrast, Robo3 is not expressed in the growth cone of commissural and longitudinal pioneer axons and is dispensable for the midline crossing during the development of the embryonic ventral nerve cord. Thus, the function of Robo3 in posterior R8s is analogous to the function of Robo1 in embryonic ipsilateral pioneers. However, analyses of knock-in mutants indicate that Robo1 and Robo3 must be sensitive to cell-type-specific regulatory functions. Robo1 has a unique role in midline repulsion of ipsilateral pioneers as it cannot be functionally replaced by Robo2 or Robo3. In contrast, as reported in this study, either Robo1 or Robo2 can functionally replace Robo3 in the R8s. Thus, repulsive signaling downstream of ipsilateral pioneers in the embryo is dependent on unique structural features of the Robo1 protein, but repulsion of posterior R8 axons does not depend on unique structural features of Robo3. Instead it is the unique and context-specific expression of Robo3 that allows it to determine R8 axon guidance and in a broader context function in the orderly assembly of a subset of columnar elements in the visual circuit (Pappu, 2011).
During development of the Drosophila visual center, photoreceptor cells extend their axons (R axons) to the lamina ganglion layer, and trigger proliferation and differentiation of synaptic partners (lamina neurons) by delivering the inductive signal Hedgehog (Hh). This inductive mechanism helps to establish an orderly arrangement of connections between the R axons and lamina neurons, termed a retinotopic map because it results in positioning the lamina neurons in close vicinity to the corresponding R axons. The bHLH-PAS transcription factor Single-minded (Sim) is induced by Hh in the lamina neurons and is required for the association of lamina neurons with R axons. In sim mutant brains, lamina neurons undergo the first step of differentiation but fail to associate with R axons. As a result, lamina neurons are set aside from R axons. The data reveal a novel mechanism for regulation of the interaction between axons and neuronal cell bodies that establishes precise neuronal networks (Umetsu, 2006).
Most axons in the brain establish topographic maps in which the arrangement
of synaptic connections maintains the relationships between neighboring cell
bodies. A notable model of topographic map formation is the visual
system, where the relay of visual information from the retina to the visual
center must be arranged in a spatially ordered manner through the topographic
connections of retinal axons with their midbrain target, which is the optic
tectum (OT) in lower vertebrates and the superior colliculus (SC) in mammals.
This topographic map is termed a retinotopic map. Many studies have shown that
Ephrin protein family members, acting through their Eph receptors, play
pivotal roles in the establishment of the retinotopic map. In
the mouse and the chick, for example, the retinal ganglion cells (RGCs) extend
their axons to the OT/SC, and the low-to-high anteroposterior gradient of
ephrin A in the target limits the posterior extension of growth cones at
various positions, dependent on the EphA level of each RGC (Umetsu, 2006).
The Drosophila visual system has also provided insight into
topographic mapping. In Drosophila, the projections of photoreceptor
neurons (R cells) themselves induce development of the corresponding
postsynaptic neurons. The Drosophila visual system consists of the
compound eyes and the three optic ganglia: the lamina, the medulla and the
lobula complex. Each of the approximately 750 ommatidial units comprising the
compound eye contain six outer photoreceptors (R1-R6) and two inner
photoreceptors (R7, R8). R1-R6 cells send their axons to the first optic
ganglion, the lamina, whereas R7 and R8 cells send axons through the lamina to
the second ganglion, the medulla. R1-R6 cells in each ommatidium make
stereotypic connections with particular lamina neurons. Synaptic units in the lamina are referred to as lamina
cartridges. During the initial step of the assembly of a lamina cartridge, an
arriving photoreceptor axon (R axon) fascicle forms a pre-cartridge ensemble,
the 'lamina column', with a set of five lamina neurons. Formation of the
ensemble results in a one-to-one correspondence of ommatidia to column units,
and is fundamental to the subsequent establishment of intricate synaptic
connections. Development of the lamina is tightly regulated by
the projection of R axons. Failure in eye formation results in concurrent loss
of the lamina, as in a normal brain, lamina neurogenesis is directly coupled
to the arrival of R axons. Both R
cell differentiation and ommatidial assembly progress in a
posterior-to-anterior direction across the eye disc. Differentiated R cells
begin to send their axons to the brain in the same sequential order,
reflecting their position in the retina along the anteroposterior and the
dorsoventral axes. Wnt signaling plays a role in regulating projections along
the dorsoventral axis (Umetsu, 2006).
As the axons from each new row of ommatidial R cell clusters arrive in the
lamina, a corresponding group of lamina precursor cells (LPCs) undergo a final
division and initiate differentiation into lamina neurons. In the first step
of their neurogenesis, direct contact with R axons triggers the transition of
G1-phase LPCs into S phase. Both the G1-S transition and the initial specification
into a lamina neuron are induced by Hedgehog (Hh), which is delivered by
arriving R axons, and the next step in lamina differentiation is induced by
the Spitz signaling molecule, which is also delivered by R axons. Hh
expressed in R cells functions as a signal for photoreceptor development as well:
secreted Hh induces anterior precursor cells to enter the pathway of R cell
specification (Umetsu, 2006).
Thus, the retinotopic map along the anteroposterior axis of the lamina
seems to be established autonomously and in a posterior-to-anterior order, as
newly specified R cells send their axons to the lamina layer and make lamina
columns. Each ommatidial unit sends a set of R axons as a single bundle to the
lamina along the pre-existing fascicle that has been just projected. Then, the
axon bundles are enveloped by the processes of newly induced lamina neurons. This step is key to forming the one-to-one associations
between R axon bundles and their corresponding lamina neurons. This study shows that the activity of Single-minded (Sim) is required for developing lamina
neurons to establish an association with the corresponding R axons and, hence,
to form the lamina column. sim encodes a basic-helix-loop-helix-PAS
(bHLH-PAS) transcription factor and is induced by Hh provided by the R axons.
In sim mutant brains, the developing lamina neurons fail to associate
with R axon bundles, resulting in a failure to establish connections between R
axons and lamina neurons. It is inferred that sim programs developing
lamina neurons to express a molecule(s) that is required for the association
with R axons (Umetsu, 2006).
Retinotopic mapping in Drosophila provides unique insights into
neuronal network formation not only because of its tight coupling to the
control of development, but also because of the interactions between axons and
neuronal cell bodies. The interactions observed
stand in sharp contrast to what has been found for other models of axon
guidance, where the growth cones of axons respond to a variety of attractive
or repulsive guidance cues to navigate to their synaptic target cells. The
cues include the netrins, Slits, semaphorins and ephrins, and the restricted expression pattern of these cues and the
reactivity of growth cones play pivotal roles in the establishment of the
proper synaptic connections. In this context, postsynaptic cells are seen as
mere providers of guidance/adhesion molecules, passively awaiting the arrival
of a growth cone. In other words, it is conceivable that presynaptic growth
cones seek their targets dynamically, whereas postsynaptic cells remain
static. Unlike the roles of presynaptic axons, the cellular behaviors of
postsynaptic cells in the establishment of synaptic targeting are poorly
understood. This study proposes another possible model for the establishment of
topographic neuronal connections in which postsynaptic cells dynamically
interact with presynaptic axons (Umetsu, 2006).
Thus, Sim, a target of Hh, is required for at least the first
step of lamina column formation; namely, the incorporation of developing
lamina neurons into the area where R axons project and lamina columns mature,
an area referred to as the assembling domain. This model for Sim is based on
four observations. First, sim2/simry75
brains have a reduced number of lamina neurons in the assembling domain,
leaving an abnormally large number of premature lamina neurons behind in the
pre-assembling domain. Second, in clonal analysis, sim2
clones fail to be recovered in the assembling domain (similar to
smo1 clones). Third, lamina neuron-specific inhibition of
Sim function causes R axon bundles to be tightly packed and lamina neurons to
be excluded from R axon bundles. And fourth, overexpression of sim in
lamina neurons causes precocious incorporation of lamina neurons into the
assembling domain (Umetsu, 2006).
In case of overexpression, neither expansion of the assembling domain nor increase
in the number of lamina neurons relative to the number of R axon bundles was
observed, even though lamina neurons prematurely incorporated into the
assembling domain. This is probably because a reduced number of lamina neurons
were generated. In fact, loss of E2F expression was observed at the lamina
furrow in NP6099-GAL4 UAS-sim brains. The onset of incorporating lamina neurons into the
assembling domain might be linked to an inhibition of cell proliferation.
However, this is thought to be unlikely for two reasons: (1) lamina neurons did not
show any extra E2F signal in the sim mutant brain in spite of an
increase in unincorporated lamina neurons; and (2) lamina neurons ectopically expressing a cell cycle-braking
factor, the Drosophila p21/p27 homolog dacapo (dap)
cause the precocious incorporation of lamina neurons. Thus, a
direct link between cell cycle regulation and the incorporation of lamina
neurons is less plausible (Umetsu, 2006).
An alternative model, the 'time lag' model, is proposed. There appears to be
a lag between the onset of sim expression and the onset of
incorporation of lamina neurons. Differentiating lamina neurons are held
temporarily in the pre-assembling domain and then the proper amount of lamina
neurons are coordinately integrated into columns as more R axons are projected. Thus, it is speculated
that a certain degree of accumulation of the Sim/dARNT heterodimer in nuclei
is needed to exert cellular function. Consistent with this idea, graded
accumulation of Sim is observed, with lower Sim levels in anterior (younger)
lamina neuron nuclei and higher levels in posterior (older) lamina neuron
nuclei.
Overexpression of Sim in lamina neurons would thus cause higher levels of
accumulation of the protein in young lamina neurons and facilitate their
incorporation into the assembling domain. Interestingly, overexpression of the
wild-type dARNT did not have any detectable effects, suggesting that Sim
accumulation is a limiting factor for cell incorporation (Umetsu, 2006).
The mechanism of neuronal maturation and that of assembly of lamina neurons
are independent, although both are under the control of Hh signaling. Disruption of
sim did not affect the differentiation and proliferation of lamina
neurons. Correspondingly, neither the incorporation of lamina
neurons into the lamina column nor the expression of sim were
affected by dac mutation. The cellular function required for assembling the column or the
function of Sim at the cellular level is still not known. Electron microscopic observations by have revealed an intriguing behavior of lamina neurons at
the early pupal stage; large processes extending from lamina neurons engulf R1
and R6 axons of newly incoming R axon bundles. This may be the key step in lamina column formation and
interaction between the R axons and lamina neurons. Sim may regulate genes
required for process formation, interaction with R axons and/or events that
follow shortly after, since lamina neurons seem to fail to make interactions with
R axons from the beginning in the sim mutant background. Sim is
expressed in the midline cells of the CNS throughout neurogenesis in the
Drosophila embryo and is required for the proper differentiation of
the midline cells into mature neurons and glial cells. Midline
precursor cells undergo synchronized cell division and then transform into the
bottle-shaped cells, in which the nuclei migrate internally and leave a
cytoplasmic projection joined to the surface of the embryo. The sim
mutant midline cells fail to delaminate from the epidermal cell layer. Cells
do not make the normal bottle-like shape and, instead, they appear rounded. In
addition, overexpression of sim can induce other cell types to
exhibit midline morphology. sim may thus regulate the transcription of a set
of genes required for morphological changes, which in turn are required for
interaction between cells, both in the lamina and during embryonic CNS
development (Umetsu, 2006).
Although sim expression is regulated by Hh
signaling, this does not answer the question of whether sim function
is sufficient to confer on cells the ability to be incorporated into the
assembling domain. Whether smo mutant clones can be
recovered in the assembling domain was examined by forcing sim expression in
smo clones using the MARCM technique. However, smo mutant
clones expressing sim were not recovered in the assembling domain. This suggests that additional factors are involved in lamina
neuron assembly. Hh may also contribute to specification of the difference in
affinity between lamina neurons and R axons and/or between anterior and
posterior lamina neurons. In Drosophila wing discs, the Hh signal
differentiates the affinity of anterior compartment cells from that of the
posterior compartment cells, thereby maintaining the compartment border (Umetsu, 2006).
An active role is proposed for postsynaptic cells in making a topographic
map of the Drosophila visual system. Targeted expression of the
dominant-negative form of the Sim partner in the lamina neurons clearly showed
a role for postsynaptic cells in assembling lamina columns. This presumably
affects an early step of assembly. It is not known if Sim
function is also required for later steps in more mature lamina neurons. The
forced expression of the dominant-negative Sim partner in the posterior lamina
neurons had no effect, although it may simply be that the level of expression
of the dominant-negative form of dARNT was not sufficient to have an
observable effect on Sim function. In the lamina column, the
R axon bundle associates with a precisely arranged row of five lamina neurons.
No mechanisms for the development and formation of this stereotypic structure
have been revealed. Another signal might be provided from the R axons with
lamina neurons, and/or intrinsic structures of the R axons might play a role
in this architecture. An intriguing property of
postsynaptic muscle cells for axonal targeting has been observed: the muscle cells bear numerous
postsynaptic filopodia ('myopodia') during motoneuron targeting.
They showed that postsynaptic cells actively contribute to synaptic
matchmaking by direct, long-distance communication. Together with what has
been learned about myopodia in neuromuscular synapse formation, the curent findings
reveal an active role for postsynaptic cells for the establishment of precise
neural networking (Umetsu, 2006).
Sim belongs to the family of bHLH-PAS transcription factors, whose members
function in many developmental and physiological processes, including
neurogenesis, tissue development, toxin metabolism, circadian rhythms, response to hypoxia, and hormone receptor function. bHLH-PAS
proteins usually function as dimeric DNA-binding protein complexes. The most
common functional unit is a heterodimer. These heterodimers consist of one
partner that is broadly expressed, and another whose expression is regulated
spatially, temporally or by the presence of inducers. Sim and the bHLH-PAS
protein dARNT heterodimerize to bind to their responsive element, the CME (CNS
midline enhancer element), to activate target gene transcription. In
this complex, dARNT is the general dimerization partner and Sim is the
tissue-specific partner (Umetsu, 2006).
The Drosophila Sim and mammalian Sim1 and Sim2 proteins are highly
conserved in their amino-terminal halves, which contain a bHLH and a PAS
domain. Murine Sim1
and Sim2 are also expressed in both proliferative and postmitotic zones of the
central nervous system at different stages of neural development. These zones
of expression include the longitudinal basal plate of the diencephalon (Sim1
and Sim2), the mesencephalon (Sim1), the zona limitans intrathalamica (Sim1
and Sim2) and the portion of the spinal cord that flanks the floor plate
(Sim1). Sim2 maps
to the region responsible for Down Syndrome (DS) on Chromosome 21.
Interestingly, Sim2 is also expressed in non-neuronal tissues, including
branchial arches and the developing limb, which are primordia of tissues and
organs where DS abnormalities are frequently observed (Umetsu, 2006).
Given the important roles of sim in Drosophila
development and the expression of Sim2 in cell types that are affected in DS
individuals, it was proposed that Sim2 may play a causative role in DS.
However, because of a lack of direct evidence and the existence of other
candidate genes, this remains speculative. Cells expressing sim
during Drosophila development and Sim2-positive cells affected in DS
seem to be able to migrate. The conserved role of Sim may enable cells to migrate
and/or interact with surrounding cells in the various tissues, including the
central nervous system. It will thus be intriguing to search for extra
cellular targets of Sim regulation with the hope of elucidating mechanisms
that underlie the behavior of Sim-expressing cells (Umetsu, 2006).
microRNAs (miRNAs) are small, conserved, non-coding RNAs that contribute to the control of many different cellular processes, including cell fate specification and growth control. Drosophila bantam, a conserved miRNA, is involved in several functions, such as stimulating proliferation and inhibiting apoptosis in the wing disc. This study reports the detailed expression pattern of bantam in the developing optic lobe, and demonstrates a new, essential role in promoting proliferation of mitotic cells in the optic lobe, including stem cells and differentiated glial cells. Changes in bantam levels autonomously affect glial cell number and distribution, and non-autonomously affect photoreceptor neuron axon projection patterns. Furthermore, bantam promotes the proliferation of mitotically active glial cells and affects their distribution, largely through down regulation of the T-box transcription factor, optomotor-blind (omb, Flybase, bifid). Expression of omb can rescue the bantam phenotype, and restore the normal glial cell number and proper glial cell positioning in most Drosophila brains. These results suggest that bantam is critical for maintaining the stem cell pools in the outer proliferation center and glial precursor cell regions of the optic lobe, and that its expression in glial cells is crucial for their proliferation and distribution (Li, 2012).
These results provide evidence that bantam is important for stem cell maintenance in the optic lobe. First, bantam shows high expression in the OPC and GPC areas in the optic lobe, where stem cells are located. Second, bantam is critical for cell proliferation in the OPC and GPC areas. banΔ1/banΔ1 null mutants have smaller brains with a dramatic decrease in the proliferation in the OPC and GPC. On the other hand, bantam over expression causes brain size to increase, along with increased proliferation in the OPC and GPC. During development, it is very important to maintain a constant stem cell population while differentiated cells are produced. In Drosophila, the central nervous system is derived from neural stem cells called neuroblasts. The optic lobe neuroepithelia are important as they maintain the pool of optic lobe neuroblasts with symmetric division. Misregulation of the self-renewing capacity of the neuroblasts is related to brain tumors; however, the mechanism underlying the precise regulation of proliferation and differentiation of the neuroepithelia and neuroblasts is not well known. miRNAs are known to be crucial for stem cell maintenance in other tissues. When the miRNA processing machinery is affected by loss of Dicer-1 (Dcr-1), which is essential for generating mature miRNAs from their corresponding precursors, stem cells cannot be maintained and are lost rapidly in the Drosophila ovary. These dcr-1 mutant stem cells are delayed in G1 to S transition. bantam was reported to be important for germline stem cell (GSC) maintenance in adult Drosophila, but the detailed underlying mechanism remains to be determined. It will be interesting to learn how bantam affects the cell cycle machinery of stem cells in the OPC and GPC regions. bantam has been known to promote cell proliferation in other tissues as well. The ability of bantam to promote cell proliferation in various tissues suggests that bantam might target molecules that directly, but negatively, affect cell-cycle machinery. Recently, a report showed that bantam targets Mei-P26, which has ubiquitin ligase activity, causing the oncogene c-Myc to degrade in the wing imaginal disc. c-Myc can respond to different growth factors to promote cell proliferation through positive regulation of the transcription factor E2F, which is a common G1-S master regulator, and is involved in regulating the expression of a number of genes required for G1-S progress. Future experiments studying whether bantam employs this same mechanism in regulating the cell cycle of stem cells in the optic lobe will be informative (Li, 2012).
It was also found that bantam is required for glial cell growth in the optic lobe. Glial cell numbers in the optic lobe were greatly increased, in a cell-autonomous manner, by an over expression of bantam. Conversely, a loss of bantam led to a dramatic decrease in glial cells in the optic lobe. During normal development, development of glial cells in the optic lobe is controlled by both extrinsic and intrinsic mechanisms. Glial cell numbers increase rapidly during the third instar larval stage due to the mitosis of differentiated glia, and, more significantly, the proliferation of precursor cells. bantam was found to increase proliferation of both glia precursor cells. This work also provides evidence that bantam's function on glial cell numbers is dependent on its negative regulation of omb in a small subgroup of differentiated glial cells, as evidenced by the ability of omb to rescue bantam's effect on glial cell numbers and distribution. Omb is a T-box transcription factor, highly conserved in all metazoans. The T-box family appears to play critical roles in development, including specification of the mesoderm and morphogenesis in the heart and limbs. In the Drosophila optic lobe, omb is expressed in a subgroup of glial cells that are required for their proper positioning and morphology. However, the downstream targets of omb responsible for these functions are not clear. Future experiments to determine if the same mechanism is employed in the brain need to be performed (Li, 2012).
It is thought that bantam does not affect glial cell differentiation because the loss of bantam in null mutants still maintains Repo-positive differentiated glial cells. Transcriptional regulators, such as Glial cells missing (Gcm) and its closely related homolog Gcm2, have been well-studied for their roles in glial cell differentiation in the embryonic and postembryonic nervous system of Drosophila. Gcm/Gcm2 are considered to be at the top of the hierarchy for initiating the differentiation of all glial cells. Their downstream targets for maintaining terminal glial cell differentiation include repo, pointed and tramtrack. With antibody staining for Repo, no obvious defects were seen in larvae caused by bantam, further supporting the idea that bantam increases glial cell numbers independent of Gcm-Repo (Li, 2012).
Besides promoting glial cell numbers, bantam also affects the mobility of glial cells, as an increase was observed in glial cells located under the lamina furrow, the migrating path for glial cells. When bantam was over-expressed, the three-layer organization of glial cells was disturbed. R-cell axon-derived signals were reported to be required for glial cell proliferation and migration in the lamina. However, the results demonstrated that glial cell defects by bantam are cell-autonomous, as neuronal over expression of bantam did not show any affect on glial cells. So far, nonstop, which encodes an ubiquitin-specific protease, was the only gene reported to be required in laminal glial cells for migration. Future experiments to determine bantam's target genes responsible for glial cell migration will be of interest (Li, 2012).
Topographic maps, which maintain the spatial order of neurons in the order of their axonal connections, are found in many parts of the nervous system. This study focused on the communication between retinal axons and their postsynaptic partners, lamina neurons, in the first ganglion of the Drosophila visual system, as a model for the formation of topographic maps. Post-mitotic lamina precursor cells differentiate upon receiving Hedgehog signals delivered through newly arriving retinal axons and, before maturing to extend neurites, extend short processes toward retinal axons to create the lamina column. The lamina column provides the cellular basis for establishing stereotypic synapses between retinal axons and lamina neurons. This study identified two cell-adhesion molecules: Hibris, which is expressed in post-mitotic lamina precursor cells; and Roughest, which is expressed on retinal axons. Both proteins belong to the nephrin/NEPH1 family. Evidence is provided that recognition between post-mitotic lamina precursor cells and retinal axons is mediated by interactions between Hibris and Roughest. These findings revealed mechanisms by which axons of presynaptic neurons deliver signals to induce the development of postsynaptic partners at the target area. Postsynaptic partners then recognize the presynaptic axons to make ensembles, thus establishing a topographic map along the anterior/posterior axis (Sugie, 2010).
This study shows that cell recognition between pre- and postsynaptic neurons via the Hbs-Rst interaction is required for the establishment of precise retinotopic mapping. During the development of the Drosophila visual center, presynaptic photoreceptors extend their axons to the lamina layer. Postsynaptic lamina precursor cells (pLPCs) start to differentiate in response to Hh delivered through newly arriving R axons. They then express Hbs, which interacts with Rst on R axons (see Model for the specific interaction between R axons and pLPCs mediated by an interaction between Hbs and Rst). This Hbs-Rst interaction is required for lamina column assembly, which underlies the topographic connections of the synapses along the anteroposterior axis (Sugie, 2010).
The process of lamina column assembly is unique in that presynaptic neurons regulate the development of postsynaptic partners in the target area, and the somata of postsynaptic neurons recognize the presynaptic axons at the developing stage well before neurite formation. This mechanism appears to be an efficient and accurate way to make a topographic map along the anterior/posterior axis. In addition, unlike the well-known axon guidance process, in which growth cones search for their targets, postsynaptic cells actively contribute to the pre- and postsynaptic interactions via direct communication. The changes in the Hbs localization that are associated with rst mutation were not only observed in pLPCs adjacent to R axons, but also in pLPCs far from R axons. This finding could be ascribed to the fact that pLPCs that are distant from R axons can contact R axons through their protrusions. Hbs might be preferentially localized at the protrusions of pLPCs that interact with R axons. The behavior of pLPCs is analogous to that of developing muscle cells, which extend filopodia to the axonal targeting of innervating motoneurons (Sugie, 2010).
Tests were performed to see whether the cell-adhesion mechanism mediated by Hbs and Rst was sufficient to rescue the sim phenotypes. Induction of exogenous hbs in pLPCs did not rescue sim loss-of-function mutants. Consistent with this finding, overexpression of sim using the NP6099-Gal4 driver caused the premature incorporation of pLPCs into the assembling domain, but overexpression of hbs did not. These results suggest that other molecules under the control of sim must be required for lamina column assembly (Sugie, 2010).
hbs expressed in photoreceptor cells does not play an essential role in lamina column assembly. The reason that Hbs originating in R axons does not interfere with the Hbs-Rst association remains unknown. The intracellular interaction of the two proteins might be blocked in R axons as a result of alternative subcellular localization and/or steric hindrance, or additional intermediates might be required for Hbs function in pLPCs, but not in R axons (Sugie, 2010).
Nephrin and NEPH1 homolog proteins tend to be located on opposing cell membranes so that they are brought into close apposition. This arrangement underlies the amazingly similar patterns of immunoreactivity in the eye disc, wing disc and somatic muscle as well as in the pupal optic lobe. These proteins are located in opposing cell membranes in the lamina. Consistent with previous studies, Hbs and Sns proteins were expressed in pLPCs, whereas Rst and Kirre were expressed in R axons; however, Hbs was also expressed in R axons. Recent studies have demonstrated that proteins of the nephrin and NEPH subfamilies are also expressed in neighboring cell types in vertebrate nervous systems. These observations reveal the conservation of nephrin/NEPH1 expression patterns across tissues and species (Sugie, 2010).
Previous work has identified SYG-1, a homolog of Rst, Kirre and NEPH1, as well as SYG-2, a homolog of Hbs, Sns and nephrin, which are necessary for synaptic specificity in Caenorhabditis elegans. The first Ig domain of SYG-1 and the first five Ig domains of SYG-2 are necessary and sufficient for binding and synapse formation in vivo. Similarly, it was found that the extracellular domain of Hbs and the first Ig domain of Rst are important for the association of pLPCs with R axons. These observations show remarkable functional conservation of the restricted domains of Drosophila and C. elegans nephrin/NEPH1 homologs (Sugie, 2010).
Further study of the preferential cell adhesion between nephrin/NEPH1 homolog proteins may reveal a common mechanism underlying the interaction between pre- and postsynaptic neurons in both Drosophila and vertebrate brains (Sugie, 2010).
A shared feature of many neural circuits is their organization into synaptic layers. However, the mechanisms that direct neurites to distinct layers remain poorly understood. This study identified a central role for Netrins and their receptor Frazzled in mediating layer-specific axon targeting in the Drosophila visual system. Frazzled is expressed and cell autonomously required in R8 photoreceptors for directing their axons to the medulla-neuropil layer M3. Netrin-B is specifically localized in this layer owing to axonal release by lamina neurons L3 and capture by target neuron-associated Frazzled. Ligand expression in L3 is sufficient to rescue R8 axon-targeting defects of Netrin mutants. R8 axons target normally despite replacement of diffusible Netrin-B by membrane-tethered ligands. Finally, Netrin localization is instructive because expression in ectopic layers can retarget R8 axons. It is proposed that provision of localized chemoattractants by intermediate target neurons represents a highly precise strategy to direct axons to a positionally defined layer (Timofeev, 2012).
Recent studies identified at least four molecular mechanisms that control layer-specific targeting in the nervous system by cell-cell interactions independently of neural activity. First, combinatorial expression of homophilic cell surface molecules promotes the recognition and stabilization of contacts between matching branches of pre- and postsynaptic neuron subsets. For instance, four members of the immunoglobulin superfamily of cell adhesion molecules, Sidekick 1 and 2 and Dscam and DscamL, are expressed and required in subsets of bipolar, amacrine, and retinal ganglion cells for targeting to different inner plexiform sublayers (IPLs) in the chick retina. In Drosophila, the leucine-rich repeat protein Caps may play an analogous role, as it is specifically expressed in R8 cells and target layers M1-M4 and, thus, could promote homophilic interactions to stabilize connections within correct columns and layers. Second, concise temporal transcriptional control is used to regulate the levels of ubiquitous cell surface molecules and, thus, adhesiveness of afferent and target neurons to balance branch growth and targeting. This mechanism is supported by findings in the fly visual system where the transcription factor Sequoia controls R8 and R7 axon targeting by the temporal regulation of N-Cadherin (CadN) expression levels. Third, repellent guidance cues are utilized to exclude projections from some layers, as has been shown for membrane-bound Semaphorin family members and Plexin receptors in the IPL of the mouse retina. Fourth, recent studies also implicated the graded expression of extracellular matrix-bound guidance cues such as Slit in the organization of layered connections in the zebrafish tectum. The current findings for the essential role of Netrins and Fra in visual circuit assembly provide evidence for a different strategy: a localized chemoattractant guidance cue is used to single out one layer, thus providing precise positional information required for layer-specific axon targeting of cell types expressing the receptor. Unlike in the ventral nerve cord, where the Netrin/Fra guidance system controls growth across the midline, in the visual system, it mediates target recognition by promoting axon growth into but not past the Netrin-positive layer (Timofeev, 2012).
Rescue experiments support the model that Netrins are primarily provided by the axon terminals of lamina neurons L3 in the M3 layer. During early pupal stages, Fra-positive R8 axons pause in their temporary layer at the distal medulla neuropil border. From midpupal development onward, upon release from this block, Fra-positive R8 axons are guided to the Netrin-expressing M3 layer (Timofeev, 2012).
Axons can use intermediate target cells either along their trajectory to guide them toward their target areas or within the target area to bring putative synaptic partners into close vicinity. Although R8 axons and lamina neurons L3 terminate closely adjacent to each other in the same layer, they have been described to not form synaptic connections with each other but to share common postsynaptic partners such as the transmedullary neuron Tm9. Thus, the results suggest that layer-specific targeting of R8 axons relies on the organizing role of lamina neurons L3 as intermediate targets in the M3 layer rather than direct interactions with postsynaptic partners. Consistent with this notion, axons of lamina neurons L3 timely extend between the temporary layers of R8 and R7 axons from early pupal stages onward, and targeting of their axons is independently controlled by other cell surface molecules such as CadN. Further studies will need to identify potential Fra-positive synaptic partners in the medulla and test whether this guidance receptor equally controls targeting of their dendritic branches, thus bringing pre- and postsynaptic neurites into the same layer. Additional mechanisms likely mediate cell-cell recognition and synaptic specificity, as electron microscopic analysis showed that presynaptic sites in R8 axons are not restricted to the M3 layer but distributed along the axon (Timofeev, 2012).
Netrins are diffusible guidance cues acting both at long range in a gradient and at short range when immobilized. Consistent with studies in the Drosophila embryo, it was observed in this study that NetB in the visual system acts at short range, as R8 axon targeting is normal when solely membrane-tethered NetB is available at near-endogenous levels. Secreted Netrins are converted into a short-range signal because they are locally released by lamina neurons L3 and prevented to diffuse away through a Fra-mediated capturing mechanism. Filopodial extensions could enable R8 growth cones to bridge the distance to NetB-expressing lamina neuron L3 axon terminals (Timofeev, 2012).
Although in principle Netrins could be secreted by both dendritic and axonal arbors of complex neurons, the results support the notion that axon terminals are the primary release sites to achieve layer-specific expression. This may be mediated by a cargo transport machinery along polarized microtubules similar to that used by synaptic proteins or neurotransmitters. Consistently, recent findings in C. elegans identified proteins involved in motor cargo assembly and axonal transport as essential for Netrin localization and secretion. Intermediate target neurons may thus constitute an important strategy to draw afferent axons into a layer, if guidance cues are preferentially released by axon terminals and not by dendritic branches of synaptic partner neurons. Netrin-releasing lamina neurons L3 form dendritic spines in the lamina and axon terminals in the medulla. Similarly, Netrin-positive transmedullary neuron subtypes such as Tm3 and Tm20 form dendritic branches in the medulla and extend axons into the lobula. Thus, a mechanism, whereby neurons in one brain area organize the connectivity in the next, may be used at least twice in the visual system (Timofeev, 2012).
Knockdown of fra in the target area strongly reduced NetB in the M3 layer, supporting the notion that a receptor-mediated capturing mechanism controls layer-specific Netrin accumulation. Despite the use of multiple genetic approaches, no R8 axon-targeting errors were observed when manipulating Fra levels exclusively in target . This could be attributed to the technical limitation that knockdown is incomplete owing to the activity of the ey enhancer in around 50% of medulla neurons. However, as lamina neurons L3 continue to locally release Netrins, remaining ligands may likely be sufficient to guide fully responsive R8 axons to their target layer (Timofeev, 2012).
Unlike in the fly embryonic CNS, where Netrins are captured by Fra and presented to growth cones expressing a Netrin receptor other than Fra, or in C. elegans, where Unc-6 is captured at the dendrite tips of nociceptive neurons by Unc-40 to interact with Unc-5 (Smith, 2012), genetic analyses indicate that fra is required in R8 axons. Hence, Netrins captured by Fra-positive target neurons may either be presented to Fra-expressing R8 axons in a dynamic fashion, or R cell- and target neuron-derived Fra interact with Netrins in a ternary complex in trans. This is conceivable since (1) the vertebrate counterpart Netrin-1 shows a high binding affinity for DCC; (2) DCC can bind Netrins with multiple domains (DCC, fourth and fifth fibronectin type III domains; Netrins, Laminin N-terminal (LamNT) and three Laminin-type epidermal growth factor [EGF]-like domains); and (3) at least in cis, Netrins can bind and aggregate multiple DCC ectodomain molecules. Ligand capture and presentation by receptors have also been reported for F-spondin and lipoprotein receptor-related protein (LRP) at the vertebrate floor plate. Netrins have previously been shown to promote exocytosis and recruitment of their receptor to distinct subcellular locations on cell surfaces. Moreover, in the visual system, Netrins may increasingly draw neurites of Fra-positive target neurons into layer M3, which in turn could promote further ligand accumulation. Thus, additional feedback loops may contribute to the specific enrichment of both Netrins and Fra in the M3 layer (Timofeev, 2012).
R8 axon targeting involves multiple successive steps: (1) the selection of the retinotopically correct column; (2) pausing in the temporary layer; (3) timely release from the temporary layer and extension of a filopodium; (4) bypassing of incorrect neuropil layers; (5) correct identification and targeting to the M3 layer; (6) stabilization of connections in the correct layer and column and transformation of growth cones into mature terminals; and (7) formation of the correct repertoire of synaptic contacts. Strong early defects would likely impact on subsequent steps (Timofeev, 2012).
Within this sequence of events, interactions of Golden goal (Gogo) and Flamingo (Fmi) in cis within R8 axons and in trans with Fmi-positive neuronal processes in the emerging M1, M2, and lower M3 layers have been shown to contribute to the timely release of R8 growth cones from their temporary layer and, consequently, enable correct targeting to the M3 layer (steps 3 and 6). Caps may specifically promote cell-cell recognition and stabilize interactions between R8 axons and target neuron branches within their correct column and target layer (step 6). However, an alteration of adhesiveness may not be sufficient to promote the extension of filopodia toward the correct layer, and additional attractive guidance forces are required. The Netrin/Fra guidance system is well suited to play such a role by providing the necessary positive forces directing filopodia toward deeper layers and by promoting recognition of a single layer at a given position (steps 4 and 5). This notion is supported by observations that loss of Fra or Netrins causes many R8 axons to stall at the distal medulla neuropil border and to terminate at interim positions in layers M1/M2. Furthermore, ectopic expression of membrane-tethered NetB is sufficient to retarget a significant proportion of R8 axons. Unlike Caps and Gogo/Fmi, whose ectopic expression can promote targeting of some R7 axons to the M3 layer, Fra was not sufficient to redirect R7 axons from the M6 to the M3 layer. A likely explanation is that the effects of R7-specific guidance determinants cannot be overwritten, or essential cooperating receptors or downstream components of Fra present in R8 are missing in R7 cells. Furthermore, overexpression of Fra causes many R8 axons to stall at the medulla neuropil border, suggesting that tight temporal regulation of receptor levels in R8 axons is essential for the integration of an additional potential repellent input (Timofeev, 2012).
Together, these findings in the Drosophila visual system suggest that the dynamic coordinated actions of chemotropic guidance cues and cell adhesion molecules contribute to layer-specific targeting of specific cell types. A similar molecular mechanism relying on Netrins or other localized attractive guidance cues and their receptors may be more widely used for the assembly of laminated circuits (Timofeev, 2012).
Neural circuit formation relies on interactions between axons and cells within the target field. While it is well established that target-derived signals act on axons to regulate circuit assembly, the extent to which axon-derived signals control circuit formation is not known. In the Drosophila visual system, anterograde signals numerically match R1-R6 photoreceptors with their targets by controlling target proliferation and neuronal differentiation. This study demonstrates that additional axon-derived signals selectively couple target survival with layer specificity. Jelly belly (Jeb) produced by R1-R6 axons was shown to interact with its receptor, Anaplastic lymphoma kinase (Alk), on budding dendrites to control survival of L3 neurons, one of three postsynaptic targets. L3 axons then produce Netrin, which regulates the layer-specific targeting of another neuron within the same circuit. It is proposed that a cascade of axon-derived signals, regulating diverse cellular processes, provides a strategy for coordinating circuit assembly across different regions of the nervous system (Pecot, 2014).
This study demonstrates that Jeb/Alk signaling regulates the survival of L3 neurons, one of several postsynaptic targets of R1-R6 neurons. Jeb is expressed in R1-R6 growth cones and acts at short range, prior to synapse formation, through the Alk receptor tyrosine kinase localized on budding L3 dendrites within the lamina neuropil. Jeb/Alk signaling is highly selective, as the survival of other R1-R6 postsynaptic targets (i.e., L1 and L2) is not affected when signaling is disrupted. This study also showed that, at a later stage of development, L3 growth cones produce Netrin within the medulla, which is required for the targeting of R8 growth cones to the M3 layer. It is speculated that a cascade of growth-cone-derived signals acting across different brain regions provides a general strategy for the assembly of neural circuits (Pecot, 2014).
In many regions of the developing nervous system, neurons are produced in excess, and significant cell death occurs after axons innervate their targets. In vertebrates, it is well established that target-derived neurotrophins, such as nerve growth factor, regulate neuronal numbers. These factors are produced by target neurons in limiting amounts and locally promote survival in a retrograde manner through receptors localized on axon terminals, providing a mechanism for matching the number of axons to targets. In recent years, diverse classes of molecules have been shown to control neuronal survival during development. Anterograde sources of trophic factors may also regulate survival, as denervation has been shown to induce excessive target neuron cell death. Indeed, several signals, including BDNF, are transported, in some contexts, in an anterograde manner within axons. In addition, the overexpression of BDNF in afferents can rescue cell death within the target field, and the disruption of BDNF through function blocking antibodies has been reported to decrease the number of target neurons within the rat superior colliculus. As BDNF may be produced by both axons and cells within the superior colliculus, it remains unclear whether endogenous axon-derived BDNF, and thus anterograde signaling, is required to regulate neuron survival (Pecot, 2014).
Although a role for target-derived retrograde trophic factors in vertebrate neural development was established many decades ago, trophic factors have only recently been shown to regulate neuronal development in Drosophila. Three Drosophila proteins, Neurotrophin 1, Neurotrophin 2, and Spatzle, are distantly related to vertebrate neurotrophins, and it has been shown that, like their vertebrate counterparts, they function as target-derived retrograde survival signals. Unlike their vertebrate homologs, however, which act through receptor tyrosine kinases, fly neurotrophins promote cell survival through Toll-like receptors (Pecot, 2014).
Although Jeb bears no significant homology to fly or vertebrate neurotrophins, Jeb acts through a receptor tyrosine kinase, Alk, which is distantly related to vertebrate neurotrophin receptors or Trks. Alk was originally identified as part of a fusion protein associated with large cell anaplastic lymphoma. Its role in mammals remains poorly understood. Drosophila Alk was initially found to regulate visceral mesoderm development through interaction with Jeb, and subsequently, Jeb/Alk signaling has been shown to regulate diverse cellular processes. Recent studies in vertebrates and Drosophila demonstrated that disrupting Alk function causes a decrease in the number of neurons. While in the vertebrate studies Alk's mechanism of action was not established, in Drosophila, Alk was shown to antagonize pathways that restrict neurogenesis under conditions of nutrient deprivation. Whether Jeb and Alk regulate neuronal survival in contexts outside of L3 development is not known, although Alk is widely expressed in the developing visual system, and Jeb is expressed by several populations of neurons, in addition to photoreceptors (Pecot, 2014).
The cellular specificity of the Jeb/Alk requirement is particularly surprising. Indeed, at all R1-R6 synapses containing L3 postsynaptic elements, L1 and L2 neurons each contribute a single postsynaptic element juxtaposing the same presynaptic site on R cell axons. In the absence of Jeb/Alk signaling, however, only L3 neurons die. The mechanisms that underlie this selectivity are not known. Alk is broadly expressed in the lamina, suggesting specificity may be controlled at the level of downstream signaling or that other trophic signals act redundantly with Jeb to control L1 and L2 survival. Collectively, the findings reported in this study demonstrate that anterograde Jeb/Alk signaling acts selectively to control L3 survival, providing direct evidence that anterograde signaling regulates target neuron survival in vivo (Pecot, 2014).
Several lines of evidence indicate that signaling between Jeb, expressed by R1-R6 growth cones, and Alk, localized to budding L3 dendrites, controls L3 survival between 20-40 hr APF. First, Alk mutant L3 neurons, or wild-type L3 neurons innervated by jeb mutant R1-R6 axons, die between 20-40 hr APF. Second, R cell populations containing only R1-R6 neurons are sufficient for L3 survival. Third, Alk and Jeb are expressed in a complementary fashion at the appropriate time on budding L3 dendrites and R1-R6 growth cones, respectively. And finally, L3 degeneration begins within budding L3 dendrites juxtaposed to R1-R6 growth cones. The temporal requirement for Alk/Jeb signaling corresponds to a critical and fascinating phase of lamina circuit assembly (Pecot, 2014).
R1-R6 growth cones form connections with lamina neurons in three discrete steps. First, R1-R6 growth cones from the same ommatidium associate with a single cartridge of differentiating lamina neurons. Second, through a highly stereotyped reassortment process occurring between 24-38 hr APF, these six growth cones diverge from one another and project locally to six different developing cartridges. As a consequence of this rearrangement, the R1-R6 cells that 'see' the same point in space form connections with L1, L2, and L3 neurons within the same cartridge. And third, R1-R6 then commence synapse formation at 45 hr APF, and this process continues until eclosion (~96 hr). Thus, L3 death in jeb and Alk mutants occurs prior to synapse formation, during the process of R1-R6 growth cone rearrangement. The suppression of L3 death by expression of the caspase inhibitor p35 argues that during normal development Jeb/Alk signaling acts to inhibit caspase activity. Which caspases contribute to L3 death, and whether caspases antagonize other cellular processes necessary for wiring, is not known. Regardless of how Jeb/Alk signaling functions at the molecular level, it acts to ensure that visual input from R1-R6 neurons is transmitted to the L3 pathway (Pecot, 2014).
These findings and the work of others suggest a logic underlying neural circuit assembly within the Drosophila visual system. The retina, lamina, and medulla are distinct yet interconnected regions comprising columnar modules (i.e., ommatidia, cartridges, and columns, respectively) that are matched topographically between each region. Within each module, intrinsic mechanisms and intercellular interactions control cell fate determination. For instance, R8 neurons provide a discrete locally acting signal to induce R7 development in the developing retina, while in the medulla, Notch/Delta interactions between daughter cells generated from the same ganglion mother cell promote acquisition of distinct cell fates. Superimposed upon these interactions are axon-derived signals that coordinate development between matched modules from different regions. Together, these mechanisms organize the assembly of columnar units in multiple regions (i.e., super columns), each processing visual information captured from a discrete region of the visual field. Indeed, the modular assembly of these super columns spanning different regions of the visual system reflects the function of these circuits in the parallel processing of visual information (Pecot, 2014).
R cell growth cones produce signals that regulate diverse cellular processes in the developing lamina. Hedgehog drives lamina neuronal precursors through their final division; cell adhesion proteins promote the association of columns of lamina neurons with R cell axon fascicles; EGF induces lamina neuron differentiation; a yet-to-be-identified signal regulates the development of lamina glia; and Jeb selectively regulates L3 survival. Thus, axon-derived signals act at multiple levels and in a cell-type-specific manner to regulate target development (Pecot, 2014).
Axon-derived signals also coordinate circuit assembly across topographically matched modules. Within medulla columns, L3 growth cones produce Netrin in the M3 layer, which controls the targeting of R8 growth cones to M3. Importantly, Netrin production by L3 occurs after Jeb, released from R1-R6 cells in topographically corresponding lamina cartridges, promotes L3 survival. Thus, Netrin indirectly relies upon prior Jeb signaling. As the L3 and R8 axon terminals within each medulla column transmit information captured from the same point in space to the same layer (M3) and share several postsynaptic targets, the developmental mechanisms giving rise to this circuit may reflect functional relationships between these neurons. Thus, signals produced by axons coordinate assembly of circuits between different brain regions (Pecot, 2014).
It is envisioned that intercellular signaling cascades, analogous to what are described in this study, organize other circuit modules in the fly visual system [e.g., ON (L1) and OFF (L2) circuits] comprising different cell types. As many regions of the vertebrate nervous system, including the neocortex, spinal cord, and retina, are also arranged in a hierarchically repetitive fashion, this raises the intriguing possibility that similar strategies may coordinate the development of these structures (Pecot, 2014).
In many regions of the central nervous systems, such as the fly optic lobes and the vertebrate cortex, synaptic circuits are organized in layers and columns to facilitate brain wiring during development and information processing in developed animals. Postsynaptic neurons elaborate dendrites in type-specific patterns in specific layers to synapse with appropriate presynaptic terminals. The fly medulla neuropil is composed of 10 layers and about 750 columns; each column is innervated by dendrites of over 38 types of medulla neurons, which match with the axonal terminals of some 7 types of afferents in a type-specific fashion. This report details the procedures to image and analyze dendrites of medulla neurons. The workflow includes three sections: (1) the dual-view imaging section combines two confocal image stacks collected at orthogonal orientations into a high-resolution 3D image of dendrites; (2) the dendrite tracing and registration section traces dendritic arbors in 3D and registers dendritic traces to the reference column array; (3) the dendritic analysis section analyzes dendritic patterns with respect to columns and layers, including layer-specific termination and planar projection direction of dendritic arbors, and derives estimates of dendritic branching and termination frequencies. The protocols utilize custom plugins built on the open-source MIPAV (Medical Imaging Processing, Analysis, and Visualization) platform and custom toolboxes in the matrix laboratory language. Together, these protocols provide a complete workflow to analyze the dendritic routing of Drosophila medulla neurons in layers and columns, to identify cell types, and to determine defects in mutants (Ting, 2017).
The precise recognition of appropriate synaptic partner neurons is a critical step during neural circuit assembly. However, little is known about the developmental context in which recognition specificity is important to establish synaptic contacts. This study shows that in the Drosophila visual system, sequential segregation of photoreceptor afferents, reflecting their birth order, lead to differential positioning of their growth cones in the early target region. By combining loss- and gain-of-function analyses it was demonstrated that relative differences in the expression of the transcription factor Sequoia regulate R cell growth cone segregation. This initial growth cone positioning is consolidated via cell-adhesion molecule Capricious in R8 axons. Further, the initial growth cone positioning was shown to determine synaptic layer selection through proximity-based axon-target interactions. Taken together, this study demonstrates that birth order dependent pre-patterning of afferent growth cones is an essential pre-requisite for the identification of synaptic partner neurons during visual map formation in Drosophila (Kulkarni, 2016).
This study demonstrates that the early, birth order dependent, segregation of R cell growth cones determines later synaptic layer identity in the Drosophila visual system. Small inter-ommatidial differences in Sequoia levels organize R cell growth cones within a layer whereas large intra-ommatidial differences segregate growth cones between layers. Changes in the positioning of growth cones directly correlate with changes in synaptic layer selection without affecting the expression of known cell-type specific targeting molecules (see Role of early growth cone patterning in synaptic layer selection). These results highlight the importance of initial afferent growth cone positioning for visual map formation prior to synaptic partner recognition (Kulkarni, 2016).
An early shift in the R8 growth cones to R7 position, induced by a short pulse of Sequoia expression, allows them to recognize the R7 target cell Dm8 as synaptic partner later during the development. Similarly, if R7 growth cones fail to segregate from R8 growth cones, they terminate together with R8 axons in the M3 synaptic layer independent of their intrinsic differentiation and targeting program. This extension of Frazzled-negative R7 axons towards layer M3 could be explained by the default setting of R7 axons to tightly fasciculate and follow the R8 pioneer axons towards the target region (Kulkarni, 2016).
R cell growth cone segregation can be controlled by axon-target interactions or axon-axon interactions or both. Although there is no experimental demonstration for direct R7-R8 afferent interactions, the following set of data indicate that such interactions occur during development. Support for direct interaction between R7-R8 afferents comes from Maurel-Zaffran (2001), where the expression of LAR as membrane tethered ligand in R8 cells alone (using an R8-specific Gal4 driver line) could induce a response from R7 axons, indicating a direct signalling between R8-R7 axons. Further, induced Capricious expression in R8 and R7 cells, in Capricious null background (therefore resulting in a target region without any Capricious expression) is sufficient to mis-target R7 axons to layer M3, again indicating a direct R7-R8 afferent interaction (Berger-Müller, 2013). The current study shows that final target layer of sequoia mutant R7 axons depends on the targeting of R8 axons, further suggesting that mis-targeting of sequoia mutant R7 axons to ectopic synaptic layer is the consequence of segregation defect rather than a change in target layer recognition (Kulkarni, 2016).
Interactions among afferent axons have been implicated in the assembly of visual and olfactory circuits in vertebrates as well as invertebrates. It has recently been shown that Eph-Ephrin signalling mediates local sorting of RGC axons in mammalian visual system. Notch signalling was demonstrated to play a role in spacing of DCN cluster neuron axons via neighbour axon interactions. But, whether these afferent interactions influence synaptic partner recognition is not known (Kulkarni, 2016).
This study shows that in the Drosophila visual system, relative levels of Sequoia determine the segregation of afferent R7/R8 growth cones within or between layers. By creating Seqhigh-Seqlow R cell combinations using Sequoia gain-of-function R7 mosaics it was observed that difference in Sequoia levels among neighbouring cells could induce growth cone segregation. The endogenous differences in Sequoia levels most likely arise as a result of the temporal sequence of R cell specification, suggesting a self-patterning mechanism in early visual circuit assembly. How the relative differences in Sequoia levels in the nuclei of R cells translate into growth cone segregation remains elusive. This study has tested candidate signalling pathways including Semaphorin/Plexin, TGF-beta ligand Activin and its receptor Baboon and Notch but did not find evidence for a critical role in initial growth cone segregation. This suggests a so far unknown molecular mechanism in which the growth state of an axon is directly coupled to differential growth cone adhesion. As it was possible to demonstrate a cell-autonomous function of Sequoia in R8 for columnar segregation as well as in R7 for layer segregation, a mechanistic model related to the concept of cell competition is envisioned, in which strong cell-cell interactions induce cell-autonomous responses (Kulkarni, 2016).
The initial segregation of afferent growth cones into distinct positions is then consolidated by expression of Capricious in R8 axons in the same posterior-to-anterior pattern in which they arrive in the medulla. It is speculated that Capricious mediated growth cone consolidation serves two purposes: 1. It removes the temporal difference in the arrival of R8 axons and 2. It maintains R8 axons in the position where they are responsive to subsequent NetrinB signal provided by L3 neurites. This is supported by two different sets of results: First, as was shown in this study, the displacement of R8 growth cones to deeper medulla position leads to their mis-targeting to layer M6 in spite of normal Frazzled expression in these R8 cells. Second, the ectopic expression of Frazzled in R7 cells cannot re-direct them to M3 layer in response to localized NetrinB signal present in the superficial position but R8 axons can be re-directed to a different layer (M1/M2) by ectopic expression of localized NetrinB in a position deeper to the superficial R8 medulla position. Taken together, these observations suggest that M3 layer targeting via L3-mediated NetrinB signalling requires R8 axons to be positioned superficially in the medulla further underscoring the importance of R8 growth cone consolidation in this position (Kulkarni, 2016).
Previous studies have identified several molecules necessary for M6 targeting of R7 axons, including Liprin-alpha/beta/gamma, PTP69D and D-Lar. The loss of these molecules specifically affects the stabilization of R7 growth cones during the second step of targeting. Additionally, these molecules along with N-Cadherin have been shown to be critical for establishment of synaptic contacts between pre and post-synaptic neurons. This study confirmed previous observations, that R7 growth cones are in close proximity with their primary post-synaptic target neurons, Dm8, at the end of growth cone segregation. This raises the possibility that targeting of R7 axons to M6 layer, later in the development, could be the direct result of R7->Dm8 contacts mediated by N-Cadherin. Recently it was shown that N-Cadherin function is necessary for stabilizing R7 growth cones in the deeper medulla position but not for targeting and subsequent extension of R7 axons to M6 layer seems to be a result of passive dislocation. Additional support for the role of N-Cadherin in the formation and maintenance of R7-->Dm8 contacts, following their initial segregation from R8 growth cones, comes from the observation that early expression of Sequoia in CadN mutant R7 cells under weak elav-Gal4 driver can rescue the mis-targeting of R7 growth cones in the superficial medulla position along with R8 growth cones at 24 hr APF, but fails to rescue the later mis-targeting to layer M3 eventually resulting in a mis-targeting phenotype identical to CadN mutant R7 axons. Interestingly, the R8 growth cones initially mis-positioned in the deeper medulla eventually mis-target to layer M6 and form synaptic contacts with Dm8. In addition, these R8 cells, with axons mis-targeted to layer M6, do not show changes in any of their known cell-type specific molecules including early specifier of cell identity (Senseless), guidance receptors (Frazzled, Capricious) and sensory receptors (Rh6). In addition, no expression of R7 specific molecules (Prospero, R3, Rh4) can be detected. Thus, the R8 cells interact with Dm8 neurons most likely via ubiquitously expressed molecules such as N-Cadherin expressed in both, R7 as well as R8, cells. This is supported by the observation that N-Cadherin is required for stabilization of R8 axons at the layer M6 (Kulkarni, 2016).
It was observed that R8 axons form functional synapses with Dm8, a known R7 target neuron, in the layer M6. This raises the fundamental question of how synaptic layer selection influences synaptic partner recognition. The cellular complexity of potential post-synaptic target layer encountered by ingrowing R cell axons has not been fully determined, leaving room for selective recognition for synaptogenesis within a layer. In fact, it has been shown that within M6, R7 axons form synapses with Dm8 but not with Tm5c which also arborize the M6 layer. In addition, this study has identified various medulla columnar neurons within M6 that are not contacted by R7 axons. Similarly in layer M3 R8 and L3 select distinct post-synaptic partners (Kulkarni, 2016).
The types of neurons present in the medulla at the time of R8 and R7 axon innervation have not been fully identified. Based on published data, the medulla neurons are generated in temporal fashion and therefore they likely innervate the medulla at different time points. Experiments presented in this study support a developmental scenario in which the medulla context for arriving R cell axons reduces the complexity of synaptic partner selection. For example R8 and L3 have different arrival times at M3, thereby would encounter a different local environment of potential post-synaptic partners competent for synaptogenesis. It is plausible that some form of temporal co-ordination of afferent axons and their post-synaptic partner cell neurites would actually simplify the synaptic partner matching. The concept of temporal identity would argue that R7 and R8 axons arriving at the same medulla position approximately the same time, as shown in the Sequoia gain-of-function background, will pick the same synaptic partners exemplified by Dm8. Support for such proximity-based axon-target interaction for synaptogenesis comes from earlier analysis of ectopic axons in Drosophila as well as Zebrafish (Kulkarni, 2016).
From an evolutionary perspective, such proximity-induced synapse formation has several advantages over mechanisms that require regulation and expression of distinct sets of cell recognition molecules. Considering R7 as the most recently added cell to the precursor ommatidium: During development, R7 is recruited using mechanisms similar to R8 and therefore possesses default R8 specification program. However, this default R8 program is suppressed to facilitate R7 specification. Thus, a temporally separated, novel R7 cell is generated with basic neuronal differentiation similar to that of an R8 cell. Interestingly the temporal difference in the R8/R7 differentiation is then translated into Sequoia mediated layer segregation of their growth cones, with Sequoia expression being part of common differentiation program. Thus, the evolutionary recent R7 cell seems to recognize its synaptic targets via pan neuronal molecules like N-Cadherin as part of the default neuronal differentiation program, instead of the invention of an additional recognition code (Kulkarni, 2016).
Establishment of synaptic connections in the neuropils of the developing nervous system requires the coordination of specific neurite-neurite interactions (i.e., axon-axon, dendrite-dendrite and axon-dendrite interactions). The molecular mechanisms underlying coordination of neurite-neurite interactions for circuit assembly are incompletely understood. This study identified a novel Ig superfamily transmembrane protein that was named Borderless (Bdl), as a novel regulator of neurite-neurite interactions in Drosophila. Bdl induces homotypic cell-cell adhesion in vitro and mediates neurite-neurite interactions in the developing visual system. Bdl interacts physically and genetically with the Ig transmembrane protein Turtle, a key regulator of axonal tiling. These results also show that the receptor tyrosine phosphatase leukocyte common antigen-related protein (LAR) negatively regulates Bdl to control synaptic-layer selection. It is proposed that precise regulation of Bdl action coordinates neurite-neurite interactions for circuit formation in Drosophila (Cameron, 2013).
The presence of numerous axons and dendrites in the neuropils of the developing CNS makes it a daunting task for establishing specific synaptic connections. Studies over the last two decades have identified a number of cell-surface recognition molecules that mediate specific neurite-neurite interactions for circuit assembly. That many cell-surface recognition molecules are present broadly in developing neuropils throughout embryonic development, however, raises the question how the action of cell-surface recognition molecules is modulated temporally to ensure accuracy in circuit formation (Cameron, 2013).
The assembly of visual circuits in Drosophila is an attractive model for understanding the general mechanisms underlying spatiotemporal control of neurite-neurite interactions. The Drosophila adult visual system is comprised of the compound eye and the optic lobe. The compound eye consists of ∼800 ommatidia, each containing six outer photoreceptor neurons (R1-R6) for processing motion and two inner photoreceptor neurons (R7 and R8) for processing color. R1-R6 axons form synaptic connections in the superficial lamina layer, and R7 and R8 axons project through the lamina into the deeper medulla layer, where they are organized into ∼800 regularly spaced columns. Each R7 and R8 axon from the same ommatidium terminate in a topographic manner in two synaptic layers within the same column. The R8 axon terminates within the M3 layer, and the R7 axon terminates in the deeper M6 layer (Cameron, 2013).
Visual circuit assembly in Drosophila involves complex neurite-neurite interactions. Specific recognition between R-cell axons and their target layers in the optic lobe have been shown to be required for synaptic-layer selection. Visual circuit assembly also requires the interactions among R-cell axons. Selection of postsynaptic targets by R1-R6 axons in the lamina requires specific axon-axon interactions. The assembly of medulla columns requires modulation of both heterotypic and homotypic axon-axon adhesion. For instance, receptor tyrosine phosphatases LAR and protein tyrosine phosphatase 69D (PTP69D) are reported to be involved in negatively regulating the adhesion between R7 and R8 axons for facilitating R7 synaptic-layer selection. And Ig-superfamily transmembrane proteins Dscam2 and Turtle (Tutl) prevent homotypic axon-axon terminal adhesion for tiling L1 and R7 axons, respectively. The exact mechanisms by which those cell-surface recognition molecules negatively regulate axon-axon adhesion, however, remain unknown (Cameron, 2013).
The role of a novel Ig-superfamily transmembrane protein Borderless (Bdl) in Drosophila was investigated in this study. Bdl is expressed in the developing visual system, and functions as a cell-surface recognition molecule to mediate neurite-neurite interactions. The receptor tyrosine phosphatase LAR and the Ig-superfamily transmembrane protein Tutl are key regulators of Bdl-mediated axon-axon interactions in controlling synaptic-layer selection and axonal tiling, respectively. The results shed new light on spatiotemporal control of cell-surface recognition molecules for coordinating circuit assembly (Cameron, 2013).
Tiling and self-avoidance, two cellular mechanisms discovered in the early 1980s, are important for patterning neuronal circuitry. Previous studies have identified several cell-surface recognition molecules, such as Dscam, Tutl, Protocadherins, MEGF10, and MEGF11, that mediate homotypic neurite-neurite interactions in tiling and self-avoidance. These cell-surface recognition molecules may act by mediating homotypic repulsion or de-adhesion between adjacent same-type neurites. For instance, molecular and genetic analyses of fly Dscam1 support a role for Dscam1 in mediating homotypic repulsion in dendritic self-avoidance, whereas mammalian Dscams appear to mediate de-adhesion by interfering with some unknown cell-type-specific cell adhesion molecules. The exact mechanisms by which these cell-surface recognition molecules mediate homotypic repulsion or de-adhesion, however, remains elusive (Cameron, 2013).
Several lines of evidence implicate Bdl as a target of Tutl in regulating R7 axonal tiling. First, overexpression of Bdl induced an R7 tiling phenotype similar to that in tutl mutants. Second, Tutl associates with Bdl in cultured cells. And third, loss of bdl rescued the tiling phenotype in tutl mutants. It is proposed that Tutl-mediated surface recognition counteracts the affinity between adjacent R7 axonal terminals by interacting with Bdl. The association of Tutl with Bdl may downregulate the level and/or adhesive activity of Bdl, thus allowing the separation of adjacent R7 axonal terminals. Since co-overexpression of Tutl and Bdl did not affect Bdl-mediated cell-cell aggregation in culture nor the Bdl-overexpression-induced tiling phenotype in flies, it is speculated that the regulation of Bdl by Tutl requires the involvement of additional regulatory molecules. Future studies are needed to determine the exact mechanism by which Tutl downregulates the function of Bdl. It will also be of interest to determine whether other cell-surface recognition molecules implicated in tiling and self-avoidance (e.g., Dscam and Protocadherins), function similarly to modulate certain cell adhesion molecules (Cameron, 2013).
The receptor tyrosine phosphatase LAR and its mammalian homologs have been shown to play important roles in axon guidance, neuronal target selection, and presynaptic development. In the developing Drosophila visual system, LAR is required for target selection of R1-R6 axons in the lamina, and synaptic-layer selection of R7 axons in the medulla. The action of LAR in R7 synaptic-layer selection reportedly involves both stabilization of axon-target interactions and down-regulation of adhesion between R7 and R8 axons. LAR-mediated axon-target interactions may involve the binding between LAR on R7 axons and an unknown ligand in the target layer, which in turn modulates the interaction between LAR and its cytoplasmic domain-binding partner Liprin to stabilize axon-target interactions. It is also reported that LAR negatively regulates an unknown cell adhesion molecule to decrease adhesion between R7 and R8 axons for facilitating synaptic-layer selection of R7 axons (Cameron, 2013).
The current results suggest strongly that LAR downregulates adhesion between R7 and R8 axons by negatively regulating Bdl. That LAR inhibited Bdl-mediated cell-cell adhesion without affecting the level of Bdl suggests that LAR inhibits adhesive activity of Bdl. Although the role of LAR in mediating axon-target interactions requires its binding to Liprin via the cytoplasmic domain, negative regulation of Bdl by LAR appears to involve a Liprin-independent mechanism. This is supported by in vitro analysis showing that a LAR mutant lacking the cytoplasmic domain also inhibited Bdl-mediated adhesion. Consistently, a previous study showed that R8-specific expression of a truncated LAR mutant lacking the cytoplasmic domain in LAR mutants could partially rescue the R7 mistargeting phenotype. LAR may directly modulate Bdl to downregulate R7-R8 adhesion, or act indirectly by interacting with other proteins. Future studies are needed to distinguish between these possibilities (Cameron, 2013).
Although negative regulation of Bdl-mediated axon-axon interactions is necessary for R7 synaptic-layer selection and tiling, it remains unclear how the presence of Bdl contributes to the formation of the R-cell axonal projection pattern in the fly visual system. Cell adhesion molecules, such as NCAM/FasII and L1-CAM/Neuroglian, have been shown to mediate selective fasciculation in axonal pathfinding. Similarly, Bdl-mediated axon-axon interactions may facilitate the projections of R7 and/or R1-R6 axons along the pioneer R8 axon. That the R-cell projection pattern remained normal in bdl mutants may be due to the presence of redundant genes. Functional redundancy among different cell adhesion molecules seems to be common in the developing nervous system, which may account for no or subtle phenotypes in mutants defective in a number of cell adhesion molecules (Cameron, 2013).
In conclusion, this study study identifies Bdl as a novel and important regulator of neurite-neurite interactions in the developing visual system. Tuning of Bdl-mediated axon-axon interactions in axonal tiling and synaptic-layer selection presents an excellent example for modulating the action of cell adhesion molecules in ensuring accuracy in circuit assembly. It is highly likely that similar mechanisms are employed for circuit assembly in mammalian nervous systems (Cameron, 2013).
How neurons form synapses within specific layers remains poorly understood. In the Drosophila medulla, neurons target to discrete layers in a precise fashion. This study demonstrates that the targeting of L3 neurons to a specific layer occurs in two steps. Initially, L3 growth cones project to a common domain in the outer medulla, overlapping with the growth cones of other neurons destined for a different layer through the redundant functions of N-Cadherin (CadN) and Semaphorin-1a (Sema-1a). CadN mediates adhesion within the domain and Sema-1a mediates repulsion through Plexin A (PlexA) expressed in an adjacent region. Subsequently, L3 growth cones segregate from the domain into their target layer in part through Sema-1a/PlexA-dependent remodeling. Together, these results and recent studies argue that the early medulla is organized into common domains, comprising processes bound for different layers, and that discrete layers later emerge through successive interactions between processes within domains and developing layers (Pecot, 2013).
Although the growth cones of L1, L3, and L5 neurons target to different layers, they initially overlap within a common domain in the outer medulla. Based on biochemical interactions and the mistargeting phenotypes and protein expression patterns described in this paper, it is envisioned that CadN-dependent adhesive interactions restrict processes to the outer medulla and that PlexA-expressing tangential neurons prevent Sema-1a expressing growth cones from projecting into the inner medulla. L2 and L4 growth cones also appear to initially target to a common domain within the distal outer medulla, but do not require Sema-1a and CadN for this targeting step and thus utilize an alternative mechanism. Interestingly, the morphology of L2 and L4 neurons does rely on Sema-1a and CadN function, indicating that within lamina neurons, these molecules regulate different aspects of targeting. This is supported by the expression of Sema-1a and CadN in all lamina neuron subclasses during development (Pecot, 2013).
In mice separate channels encoding light increments (ON) and decrements (OFF) are spawned in the outer retina and relayed to different sublaminas of the inner plexiform layer (IPL). The current findings are reminiscent of recent studies in the mouse IPL (Matsuoka, 2011) in which Kolodkin and colleagues demonstrated that the processes of different subclasses of PlexA4-expressing amacrine cells are segregated to different OFF layers and that this requires both PlexA4 and Sema6A. Although these proteins act in a more traditional fashion as a receptor and ligand, respectively, they are expressed in a complementary fashion early in development when the developing neuropil is very thin, with PlexA expressed in the nascent OFF layer and Sema6A in the developing ON layers. This raises the intriguing possibility that, as in the medulla, different cells initially target to common domains, from which they then segregate into discrete layers. As Cadherin proteins are differentially expressed in a layered fashion in the developing IPL and defects in targeting are incomplete in both Sema6A and PlexA4 mutants (Matsuoka, 2011), it is possible that, as in the medulla, Semaphorin/Plexin repulsion acts in parallel with cadherin-based adhesion to control layer-specific patterning within the developing IPL (Pecot, 2013).
Taken together, these studies suggest that the restriction of processes to a common domain prior to their segregation into distinct layers may be a developmental strategy used in both the medulla and the vertebrate IPL. This step-wise process may represent a more general strategy for reducing the molecular diversity required to establish synaptic connections by limiting the potential synaptic partners that growth
cones and nascent dendritic arbors encounter within the developing neuropil (Pecot, 2013).
After targeting to a common domain within the outer medulla, L3 growth cones undergo stereotyped changes in shape and position that lead to segregation into the M3 layer. Initially, L3 growth cones are spear-like, spanning much of the depth of the incipient outer medulla. They then expand and elaborate a myriad of filopodia before resolving into flattened synaptic terminals within the M3 layer. This transformation is marked by two prominent steps: extension of processes from one side of the lateral region of the growth cone into the incipient M3 layer and retraction of the leading edge of the growth cone from the incipient M5 layer (part of the domain shared by L1 and L5 growth cones) (Pecot, 2013).
It has been suggested that CadN may regulate the extension within M3, as this step is partially perturbed in CadN mutant growth cones. However, as CadN mutations affect the initial position of L3 growth cones within the outer medulla, the extension defect within the M3 layer may be indirect. By contrast, in sema-1a mutant growth cones, initial targeting is indistinguishable from wild-type, so defects in retraction away from the incipient M5 layer are likely to reflect a direct role for Sema-1a in this later step in growth cone reorganization. PlexA RNAi phenocopies a sema-1a null mutation and, thus, PlexA is also required for retraction and is likely to function on medulla tangential fibers, where it is most strongly expressed. In support of this, the tip of the L3 growth cone that retracts is in close proximity to these PlexA-expressing fibers (Pecot, 2013).
The function of Sema-1a/PlexA signaling in sculpting L3 growth cones appears to be distinct mechanistically from the earlier role it plays in confining the growth cones to a common domain. During initial targeting, PlexA acts as a barrier to L3 growth cones and prevents them from projecting beyond the outer medulla. Thus, at this early step, Sema-1a/PlexA interaction provides a stop signal for the leading edge of L3 (uncovered in double mutants with CadN). In the second step, however, Sema-1a/PlexA signaling promotes retraction into the M3 layer. How these diverse outputs of Sema-1a/PlexA signaling arise is unclear. Sema-1a may be coupled to different downstream effectors at each step, modified by association with other receptor subunits, or may be modulated by other extracellular signaling pathways (Pecot, 2013).
CadN may also play a role in the retraction of L3 growth cones away from the domain shared with L1 and L5 growth cones. In early pupal stages, disrupting CadN function, while leaving growth cone morphology largely spear-like, causes L3 axons to project deeper within the medulla. Under these conditions, Sema-1a function is sufficient to prevent the growth cones from extending beyond the outer medulla. Subsequently, CadN mutant L3 growth cones fail to move away from the outer medulla's proximal edge into the developing M3 layer and thus remain within the most proximal layer, M6. This suggests that CadN, while acting in parallel with Sema-1a to restrict L3 growth cones to the outer medulla initially, may also be required at later stages for movement of the L3 leading edge into the M3 layer. As CadN has been shown previously to regulate neurite outgrowth over cultured astrocytes, it may be required for L3 growth cones to move along adjacent processes. However, the initial projection of L3 axons into the medulla is not affected by CadN mutations, indicating that other components control this process. It also remains possible that the defect in growth cone retraction results indirectly from CadN's earlier role in targeting; this earlier role may account for the defects in growth cone extension within M3 (Pecot, 2013).
Disrupting CadN function in different neurons affects targeting in unique ways. For example, L5 axons lacking CadN target to the proper layer, but extend inappropriately within the layer into neighboring columns (Nern, 2008). In addition, CadN mutant R7 growth cones display abnormal morphology and, in contrast to mutant L3 growth cones, initially target correctly, but retract to a more superficial medulla region. Collectively, these findings demonstrate that CadN regulates divergent features of growth cone targeting in different contexts. This likely reflects molecular diversity between different growth cones and illustrates the importance of understanding how molecules act in combination to generate target specificity (Pecot, 2013).
These studies add to previous findings suggesting that column assembly relies on a precisely orchestrated sequence of interactions between different neuronal cell types (Nern, 2008; Timofeev, 2012). This study shows that, as L1, L3, and L5 growth cones expressing Sema-1a enter the medulla, they meet the processes of newly arriving tangential fibers expressing PlexA, which acting in parallel with CadN, prevents extension of these growth cones into the inner medulla. This timing may permit other Sema-1a-expressing growth cones to extend into the inner medulla at earlier stages; these growth cones may then use Sema-1a/PlexA signaling for patterning connections in the inner medulla or deeper neuropils of the lobula complex. Subsequent sculpting of the L3 growth cone, mediated by Sema-1a/PlexA and perhaps CadN, leads to its reorganization into an expanded terminal within M3. As L3 growth cones become restricted to the M3 layer, Netrin, secreted from L3 growth cones, becomes concentrated within the M3 layer, and this, in turn, attracts R8 growth cones to the M3 layer, as recently described by Salecker and colleagues (Timofeev, 2012; Pecot, 2013 and references therein).
Given the extraordinary cellular complexity of the medulla neuropil, with over 100 different neurons forming connections in different medulla layers, and the few mechanistic clues to layer specific targeting that have emerged so far, a complex interplay between different sets of neurons is envisioned to be required to assemble the medulla circuit. The availability of specific markers for many of these neurons, techniques to follow the expression of even widely expressed proteins at the single cell level as is described in this study, and the ability to genetically manipulate single cells during development provide a robust system for uncovering the molecular logic regulating the layered assembly of axon terminals, dendritic arbors, and synaptic connectivity (Pecot, 2013).
In Drosophila, the four inner photoreceptor neurons
exhibit overlapping but distinct spectral sensitivities
and mediate behaviors that reflect spectral preference.
A genetic strategy, Tango-Trace, permits the identification of the
connections of the four chromatic photoreceptors.
Each of the four stochastically distributed chromatic
photoreceptor subtypes make distinct connections
in the medulla with four different TmY cells. Moreover,
each class of TmY cells forms a retinotopic
map in both the medulla and the lobula complex,
generating four overlapping topographic maps that
could carry different color information. Thus, the
four inner photoreceptors transmit spectral information
through distinct channels that may converge in
both the medulla and lobula complex. These projections
could provide an anatomic basis for color vision and may relay information about color to motion sensitive areas. Moreover, the Tango-Trace strategy may be applied more generally to identify neural circuits in the fly brain (Jagadish, 2014).
Visual stimuli are detected by photoreceptors in the retina and
transmitted to the brain to generate an internal representation
of the visual world. The brain must then translate this representation
of stimulus features into visually guided behaviors. In Drosophila, the retina resembles a crystalline lattice comprised of 750 precisely ordered units, the ommatidia.
Each ommatidium contains eight photoreceptor neurons
(R1-R8). The outer photoreceptor neurons, R1-R6, express the
Rh1 opsin and are thought to receive achromatic visual stimuli
that ultimately inform the fly about the form, position, and movement
of objects in the visual world. There are two types of ommatidia
that differ in the opsins expressed by the inner photoreceptor,
R7 and R8. In Pale (p) ommatidia, R7 expresses the near
UV-sensitive opsin Rh3 and the R8 cell expresses the blue-sensitive
opsin Rh5. In Yellow (y) ommatidia, R7 cells contain the far
UV-sensitive opsin Rh4, whereas the R8 cell expresses the
green-sensitive opsin Rh6. The existence of the four types of inner photoreceptor neurons, each with overlapping but distinct spectral sensitivities,
has implicated these neurons in the recognition of chromatic
visual information (Jagadish, 2014).
Drosophila exhibit phototactic behaviors, strongly preferring
UV to green light, a preference that is not observed in flies in
which neurotransmitter release is blocked in R7 cells. These observations suggest
that the chromatic inner photoreceptors elicit behaviors
that reflect spectral preference, but the response to light of
distinct wavelengths does not constitute color vision.
Color vision requires the ability to distinguish light of distinct
spectral composition independent of intensity. The principle of
univariance argues that a single photoreceptor cannot distinguish
different wavelengths from different intensities of light. Color vision therefore requires a neural system capable of comparing the inputs from photoreceptor neurons in
the retina that exhibit different spectral sensitivities. In mammals,
this comparison is apparent early in the visual pathway,
with a subset of retinal ganglion cells exhibiting color opponency,
a feature that reflects opposing neural responses to
input from different types of photoreceptor cells. In flies, connections between
photoreceptors do not facilitate a comparison of inputs. Color vision would therefore require a comparison of
different photoreceptor inputs in downstream visual processing
centers. However, the connections of the four inner photoreceptors
are largely unidentified and it is unknown whether p and y ommatidia project to identical or distinct downstream circuits (Jagadish, 2014).
The achromatic R1-R6 neurons project axons to cartridges
within the lamina, an optic lobe structure immediately below
the retina. The R7 and R8 axons course through the lamina and synapse on second-order
neurons within a column in the medulla. The R7 and R8 cells from
a single ommatidium project axons to the same column and the
topographic organization of the columns maintains retinotopic
order. Anatomic studies continue to reveal a vast complexity
of richly arborizing intrinsic neurons and projection neurons in
the medulla that have seriously hindered the identification of
synaptic partners of R7 and R8 neurons. EM studies suggest
connections between R7, a medullary projection neuron Tm5
and a medullary intrinsic neuron Dm8. Moreover, genetic studies have shown that Dm8 is necessary for UV spectral preference. Tm neurons in the medulla project their axons to only one of the two neuropils of the lobula complex. TmY cells in the medulla
project a branched axon to both the lobula and the lobula plate. The lobula plate contains neurons that respond
strongly to motion and loom sensitive neurons have
been identified in the lobula plate, but the function of the remaining neural structures within the lobula complex remains obscure (Jagadish, 2014).
In other insects such as bees that exhibit clear behavioral
evidence for color vision, color opponent neurons have been
identified with electrophysiologic recordings in the inner layers
of both the medulla and lobula as well as by imaging studies in
the anterior optic tubercle. Color opponent neurons provide
an anatomic substrate for a comparison of the different photoreceptors.
Efforts to identify the chromatic neural circuits that
could underlie color vision in Drosophila have failed to identify
color opponent neurons in the optic lobe. Moreover, behavioral
studies have not provided convincing evidence for color vision
in the fly (Jagadish, 2014).
A genetic strategy, Tango-Trace, has been developed that permits
tracing of functional synaptic connections of the R7
and R8 photoreceptor neurons in the optic lobe of Drosophila.
These studies reveal that each of the four stochastically distributed
chromatic photoreceptor subtypes makes a different functional
synaptic connection with four different TmY cells in the
medulla. The four distinct TmY cells all project to the innermost
layer of the lobula and extend axons more diffusely to multiple
layers within the lobula plate. The observation that the four inner
photoreceptors transmit spectral information through
distinct channels that can connect to one another in both the
medulla and the lobula complex may provide the anatomic
substrate for color vision. Moreover, the Tango-Trace strategy
used to trace connections in the dense medullary neuropil may be applied more generally to identify neural circuits in the fly brain (Jagadish, 2014).
The stochastically distributed inner photoreceptor subtypes each contact a unique
postsynaptic TmY cell. The TmY cells, although anatomically
distinct in the proximal medulla, share several anatomic features
in the lobula and lobula plate. In the lobula plate, each
TmY projects to all four layers, whereas in the lobula, arborizations
are restricted to the innermost layer. Moreover, each of the
four different TmY cells maintains a retinotopic map from the
medulla to the lobula complex. Registered images of isosurfaces
of different TmYs show that contiguous maps are likely to
overlap. Thus, four overlapping topographic maps transmit
different color information from individual inner photoreceptor
subtypes in the retina to the lobula complex. These
observations demonstrate that the four inner photoreceptor
subtypes process chromatic visual information in separate
and parallel pathways. A given photoreceptor, however, cannot
distinguish wavelength differences from intensity differences
and color vision therefore requires that signals from the different
dedicated TmYs are compared by convergent processing
downstream (Jagadish, 2014).
In the trichromatic mammalian retina, each of the different
cones synapse on a different bipolar cell, a feature most clearly
illustrated by the S-type cones that synapse on postsynaptic
S-ON bipolar cells. Thus, independent parallel
information channels continue from the cone to bipolar cell and
L versus M opponency, as well as L-M versus S opponency is
observed in the retinal ganglion cells (Nassi,
2009). This functional organization in the mammalian retina resembles
the parallel channels between the inner photoreceptors
of the fly and the four TmY cells. Opponency in Drosophila would
therefore be apparent in downstream targets including the proximal
medulla or lobula complex. In the honeybee, several types
of color opponent neurons have been detected by electrophysiologic
recordings in the medulla and lobula, two sites of axon arborization we
observe for the four TmY cells. Whatever the site of integration
of the four distinct photoreceptor channels, the identification of
independent output neurons for the four photoreceptor types
suggests an anatomic substrate for the integration necessary
for color analysis (Jagadish, 2014).
Individual inner photoreceptors express only one of the four
rhodopsin subtypes, dictated by the stochastic expression of
specific transcription factors. Each photoreceptor then projects
retinotopically to a single column in the medulla. The stochastically distributed
photoreceptor subtypes then contact a unique postsynaptic
TmY target with precision. How is this map established? In one
model, all columns might contain dendrites from all four
potential TmY targets, but an identity code of synaptic specificity
cues between photoreceptors and TmY cells assures that only
the correct synapses form. Alternatively, each photoreceptor
subtype may induce the differentiation of its cognate TmY target,
thereby matching pre- and postsynaptic partners. Finally, the
specificity of connections may be initiated by columnar guidance
cues and TmY receptors (Jagadish, 2014).
The four TmY cells postsynaptic to each of the chromatic
photoreceptor types arborize across all layers of the lobula plate
and this may afford a new site of integration of inner and outer
photoreceptor signals. Inner photoreceptors provide inputs to
motion-sensitive pathways by forming direct, electrical connections
with the axon terminals of outer photoreceptors, enhancing
the robustness of motion vision. These connections
arise at an early stage in visual processing that necessarily
precedes comparisons between inner photoreceptor
signals and therefore cannot provide color information to motion
detecting circuits. Rather, these early connections can broaden
the spectral tuning of these pathways. The lobula plate outputs
of the TmY cells identified in this study may act redundantly with
these peripheral interactions, providing another neural mechanism
to enhance motion detection. However, as the arbors of
these TmY cells span small numbers of neighboring columns in
the proximal medulla, these cells might also represent a site of
integration between different spectral inputs. Thus, the projections
of these cells to the lobula plate could relay spatially
restricted information about color to motion sensitive areas.
Although turning responses evoked by optic flow are color
insensitive, lobula plate neurons in fruit
flies and larger Diptera can be tuned to motion in different directions,
complex patterns of optic flow, as well as looming objects. Moreover, distinct motion sensitive circuits guide different behavioral responses. Thus, the pathways defined by these TmY cells might adjust the sensitivity of lobula plate neurons to
motion in a color-dependent fashion, providing a mechanism
for tuning particular motion-sensitive behaviors to specific spectral
environments (Jagadish, 2014).
Tango-Trace was developed to identify postsynaptic targets
of the chromatic photoreceptor neurons. The modular design
of Tango-Trace allows a more general application to identify
postsynaptic partners of virtually any genetically defined subpopulation
of presynaptic cells. A variant Tango mapping
approach has been developed to identify sites of dopamine-mediated
neuromodulation in the brain.
The only requirement for Tango tracing is the knowledge of
the neurotransmitter released by the presynaptic neuron allowing
the identification and genetic modification of its cognate
GPCR essential for Tango-Trace. Tango-Trace affords several
advantages as a general approach to trace neural pathways
in the fly brain. First, the identification of postsynaptic targets
relies on exogenous genes introduced into the fly to create
a novel signaling pathway independent of the activation of
potential confounding endogenous cell-signaling events. Moreover,
this approach transforms rapidly adapting signaling
events into a more stable and amplifiable cellular response.
In addition, the transcriptional readout of the Tango system
not only permits the expression of reporters that allow the
visualization of neurons, but also manipulatable effectors
including ion channels as well as Ca2+ indicators in restricted
populations of postsynaptic neurons. Tango-Trace has particular
advantages in the fly brain where tangled neuropils,
without apparent structure, predominate and the anatomic
proximity of processes cannot reliably predict functional connections.
GRASP, which depends upon
synaptic level proximity, affords an alternative tracing approach
but does not directly report a functional synapse, an important
feature inherent in Tango-Trace (Jagadish, 2014).
One concern remaining with the Tango approach surrounds
the question of sensitivity. This study detected four TmY cells and
two interneurons, Dm8 and Mia, postsynaptic to the inner photoreceptors.
The complete repertoire of neurons
in the medulla, postsynaptic to the inner photoreceptors is not known and
therefore the effectiveness of labeling for all
postsynaptic targets cannot be determined. Over 70 neuronal cell types have been
identified in the medulla and the identification of direct targets
of R7 and R8 by electron microscopic reconstruction has preliminarily
described Dm8, Tm5, Tm9 as well as L1,
L2, L3, Dm2, Mi4, Mi9, Mi15, and Tm20 as postsynaptic targets. At present, the EM reconstruction of
medulla columns is incomplete because current efforts have
focused on the circuits involved in motion detection downstream
of outer photoreceptors. Thus, resolution
of apparent discrepancies among the various EM reconstruction
efforts, and between the EM data and Tango trace, may emerge
upon a complete EM reconstruction of a medulla column. The
inability to detect targets other than the four TmYs in Tango-
Trace may reflect inefficiencies or bias in the identification of
postsynaptic targets. This notwithstanding, the identification of
the four distinct TmYs and the independent confirmation of their
connectivity by functional imaging argues strongly for their participation in specific parallel pathways postsynaptic to the chromatic photoreceptors (Jagadish, 2014).
In Drosophila, color vision and wavelength-selective behaviors are mediated by the compound eye's narrow-spectrum photoreceptors, R7 and R8, and their downstream neurons, Tm5a/b/c and Tm20, in the second optic neuropil, or medulla. These chromatic Tm neurons project axons to a deeper optic neuropil, the lobula, which in insects has been implicated in processing and relaying color information to the central brain. The synaptic targets of the chromatic Tm neurons in the lobula are not known, however. Using a modified GRASP (GFP reconstitution across synaptic partners) method to probe connections between the chromatic Tm neurons and 28 known and novel types of lobula neurons, the visual projection neurons LT11 and LC14, and the lobula intrinsic neurons Li3 and Li4, were identified anatomically as synaptic targets of the chromatic Tm neurons. Single-cell GRASP analyses revealed that Li4 receives synaptic contacts from over 90% of all four types of chromatic Tm neurons while LT11 is postsynaptic to the chromatic Tm neurons with only modest selectivity and at a lower frequency and density. To visualize synaptic contacts at the ultrastructural level, a 'two-tag' double labeling method was developed and applied to label LT11's dendrites and the mitochondria in Tm5c's presynaptic terminals. Serial electron microscopic reconstruction confirmed that LT11 receives direct contacts from Tm5c. This method would be generally applicable to map the connections of large complex neurons in Drosophila and other animals (Lin, 2015).
Color vision is an important sensory capability of humans and many animals. It relies on color opponent processing in visual circuits that gradually compare the signals of photoreceptors with different spectral sensitivities. In Drosophila, this comparison begins already in the presynaptic terminals of UV-sensitive R7 and longer wavelength-sensitive R8 inner photoreceptors that inhibit each other in the medulla. How downstream neurons process their signals is unknown. This study reports that the second order medulla interneuron Dm8 is inhibited when flies are stimulated with UV light and strongly excited in response to a broad range of longer wavelength (VIS) stimuli. Inhibition to UV light is mediated by histaminergic input from R7 and expression of the histamine receptor ort in Dm8, as previously suggested. However, two additional excitatory inputs antagonize the R7 input. First, activation of R8 leads to excitation of Dm8 by non-canonical photoreceptor signaling and cholinergic neurotransmission in the visual circuitry. Second, activation of outer photoreceptors R1-R6 with broad spectral sensitivity causes excitation in Dm8 through the cholinergic medulla interneuron Mi1, which is known for its major contribution to the detection of spatial luminance contrast and visual motion. In summary, Dm8 mediates a second step in UV/VIS color opponent processing in Drosophila by integrating input from all types of photoreceptors. These results demonstrate novel insights into the circuit integration of R1-R6 into color opponent processing and reveal that chromatic and achromatic circuitries of the fly visual system interact more extensively than previously thought (Pagni, 2021).
A universal principle of sensory perception is the progressive transformation of sensory information from broad non-specific signals to stimulus-selective signals that form the basis of perception. To perceive color, human brains must transform the wavelengths of light reflected off objects into the derived quantities of brightness, saturation and hue. Neurons responding selectively to hue have been reported in primate cortex, but it is unknown how their narrow tuning in color space is produced by upstream circuit mechanisms. To enable circuit level analysis of color perception, this study report sthe discovery of neurons in the Drosophila optic lobe with hue selective properties. Using the connectivity graph of the fly brain, a connectomics-constrained circuit model was constructed that accounts for this hue selectivity. Unexpectedly, the model predicts that recurrent connections in the circuit are critical for hue selectivity. Experiments using genetic manipulations to perturb recurrence in adult flies confirms this prediction. These findings reveal the circuit basis for hue selectivity in color vision (Christenson, 2023).
Complicated neuronal circuits can be genetically encoded, but the underlying developmental algorithms remain largely unknown. This study describes a developmental algorithm for the specification of synaptic partner cells through axonal sorting in theDrosophila visual map. This approach combines intravital imaging of growth cone dynamics in developing brains of intact pupae and data-driven computational modeling. These analyses suggest that three simple rules are sufficient to generate the seemingly complex neural superposition wiring of the fly visual map without an elaborate molecular matchmaking code. This computational model explains robust and precise wiring in a crowded brain region despite extensive growth cone overlaps and provides a framework for matching molecular mechanisms with the rules they execute. Finally, ordered geometric axon terminal arrangements that are not required for neural superposition are a side product of the developmental algorithm, thus elucidating neural circuit connectivity that remained unexplained based on adult structure and function alone (Langen, 2015).
A central question in neuroscience is how neural circuits self-organize into functional structures during development. The wiring of compound eyes to the brain of flies provides a fascinating model system for studying this question. In particular, the neural superposition eye, such as found in advanced flies, is characterized by a complicated wiring diagram: each point in visual space is captured by multiple photoreceptors from different ommatidia that converge upon the same synaptic unit (cartridge) in the brain; different photoreceptors within the same ommatidium view different points in visual space and project to neighboring cartridges. The correct pooling of axon terminals viewing the same point in space into a single cartridge increases sensitivity without loss of spatial resolution compared with simpler, ancestral eye types. The developmental process underlying neural superposition is remarkable, because each individual axon, among thousands of neighboring axons in the brain, must be sorted together with those few axons that receive input from the same point in visual space (Langen, 2015).
A classic model of neural superposition is found in the Drosophila compound eye, which contains ~800 ommatidia. Each ommatidium projects a bundle of eight photoreceptor (retinula or R-cell) axons into the brain. Six of these photoreceptors, R1-R6 (the focus of the current study) form the primary visual map in the lamina (first optic neuropil) of the fly brain. The R1-R6 axons from one bundle that receive input from six different points in visual space are denoted A-F (Langen, 2015).
After neural superposition is established, the R-cells have a precise organization of the six subtypes around the circumference of cartridges, that is, R1 neighbors R2, which neighbors R3, etc., referred to as 'rotational stereotypy'. The precision of rotational stereotypy is noteworthy, as the six axon terminals in a cartridge carry the same input information and synapse with the same postsynaptic target cells. Hence, rotational stereotypy is not a functional requirement for neural superposition and increases the demands placed on the sorting problem from 800 cartridges to 4,800 (800 × 6 R1-R6) precise terminal positions. The role, development, and evolutionary origin of this wiring precision are unknown (Langen, 2015).
The neural superposition wiring diagram has a 'canonical' pattern of six R-cell axon terminals per cartridge. An equator from anterior to posterior divides the compound eye, as well as the wiring pattern in the lamina, into dorsal and ventral halves. The wiring patterns in each half of the lamina are opposite to one another with respect to the equator axis. As a consequence, six rows of "non-canonical" cartridges exist at the equator that contain stereotypic compositions of seven or eight R1-R6 cell axon terminals. The three different types of equator cartridges also exhibit rotational stereotypy, each with a distinct pattern. As in the case of canonical cartridges, the function of the rotational stereotypic arrangement of photoreceptor terminals within the equator cartridges is unknown. It is unclear which common developmental rules or mechanisms might robustly encode the canonical cartridges, as well as the three types of equator cartridges (Langen, 2015).
The Drosophila visual system is an example of a genetically encoded neural circuit in which a developmental sorting step precedes and ensures synaptic specificity between input neurons and their targets. Many aspects of the developmental sorting step have been characterized in detail, including the formation of an initial grid by lamina cells. Previous studies have suggested the possibility of simple developmental rules underlying this sorting process. Furthermore, work in recent years has revealed molecular mechanistic insight into how differential adhesion of guidance receptors may play a key role in growth cone sorting. However, no rule set or algorithm has been formulated that is sufficient to generate precise neural superposition in canonical cartridges and equator cartridges. Two key challenges have been (1) the inability to monitor the dynamic sorting process live in developing flies and (2) lack of quantitative, data-driven models to conceptualize or test understanding of this apparently complicated process (Langen, 2015).
This study reports live imaging of R1-R6 growth cone dynamics in intact developing pupa and the derivation of a model that summarizes the conceptual understanding of the development of neural superposition. It is proposed that three simple rules are sufficient to provide a solution to the neural superposition sorting problem. Systematic tests of these rules in a computational model reveal that the same rule set leads to precise superposition and the three types of cartridges observed at the equator (Langen, 2015).
This study describes a developmental algorithm for the axonal sorting of ~4,800 presynaptic cells in the primary visual map of Drosophila. The work suggests that the neural superposition wiring diagram found in adult fly brains can be established through simple, local pattern formation principles without the need for an elaborate molecular matchmaking code. Codification of the developmental algorithm reveals quantitative constraints and provides a conceptual framework for molecular mechanisms that execute these rules (Langen, 2015).
The current findings, together with previous studies, support the following developmental algorithm.
Rule 1: The Scaffolding Rule:
Incoming rows of axon bundles from individual ommatidia are organized in a repeating pattern of evenly spaced semi-circles. This pattern and spacing of original axon arrival points provide a scaffold that remains stable during the entire process of growth cone sorting and is required for neural superposition. The future target areas are encircled by the anchored heels and thus already defined prior to growth cone movements. How the precision of the scaffold pattern develops is unknown. The scaffold is likely to instruct the extension angle through non-autonomous R-cell interactions within a bundle. Such intra-bundle interactions have been proposed to play a more prominent role than do interactions across bundles (Schwabe, 2013). To which extent the geometric arrangement of heels observed in the scaffold is influenced by their axonal arrangements within the bundles or by other cells within the target area is unclear (Langen, 2015).
Rule 2: The Extension Rule: All R1-R6 growth cones extend synchronously with speeds and angles specific to their R-cell subtype during the 5-10 hr of extension. The extension is unaffected by highly varying environments at the equator and thus is unlikely to depend on R-cell front interactions. However, precise extension dynamics may require permissive R-cell heel interactions and recognition of other cells that are equally distributed across the equator as instructive guides. It is unlikely that R-cell growth cones simply extend toward attractive cues at the target regions because the growth cones can overlap with several target regions throughout their sorting (including the target regions closest to the heels). Based on these observations, it is considered that the extension process of the bipolar R-cell growth cones differs from the classic view of growth cone movements toward attractive targets (Langen, 2015).
Rule 3: The Stop Rule: The target regions defined by the scaffold (and the L-cells therein) provide possible, but poor, targets for R-cell fronts to stop extending, because those R-cell fronts overlap with multiple targets simultaneously and throughout their extension. In addition, all R-cell fronts increasingly overlap with other R-cell fronts throughout their extension. The computational model reveals that stop rules based on R-cell front overlap function even without any target-derived cues and are more robust than a 'target only' model under the same conditions. A target model using coincidence detection of overlap with other R fronts, as well as target L-cells, performs best. R-cell front interaction is predicted to be part of the stop rule because of reduced robustness at the equator and because of the rotational stereotypy of R-cell terminal positions within cartridges. These two observations also argue against 'no interaction' stop rules. However, the results do not rule out the existence of a synchronously applied stop signal that could act as part of a combinatorial stop rule. The precise nature and molecular correlate of the stop rule remain unknown (Langen, 2015).
Previous work has revealed important insights into further constraints of these rules. Most importantly, the 180°rotation of a single bundle results in 180° rotated extension angles. This finding is consistent with the model. In addition, differential subtype dependencies have been unraveled, where R1, R2, R5 and R6 targeting depend on R3 and R4, but not the other way round. Whether this dependency arises from the scaffolding, extension, or stop rule remains to be determined. It is not yet known whether reconciliation of the current model with these observations arises from constraining existing rules or requires new ones (Langen, 2015).
After growth cone sorting is complete, a process of centripetal growth commences synchronously from all R-cell fronts and these then generate R-cell terminal columns orthogonal to the lamina plexus. This columnar extension preserves and freezes the relative positions of R-cell fronts in the lamina plexus; the resulting columns of R-cell terminals then define the adult lamina (Langen, 2015).
Complicated wiring diagrams can originate through the iterative execution of simple rules. Early brain development is associated with genetically encoded pattern formation rules, while later phases of synapse specification often depend on neuronal activity. It is unclear which level of synaptic partner specification can be achieved through simple, genetically encoded developmental rules. This study has focused on the identification of such rules and their quantitative constraints using the genetically hard-wired Drosophila visual map as a model (Langen, 2015).
Much previous elegant work has focused on searching for molecular codes underlying synaptic partner specification. Such codes may be characterized by either many molecular cues (e.g., olfactory systems) or fewer molecular cues that are dynamically localized (e.g., the fly's visual system). The current work on identifying an underlying developmental algorithm provides a framework for matching these molecular mechanisms with the rules they execute. For example, recent studies on guidance receptors of the Cadherin family have provided strong evidence for a role of differential adhesion in R-cell growth cone sorting (Schwabe, 2013; Schwabe, 2014). Specifically, R-cell growth cones interact through differential adhesion of the protocadherin Flamingo, both within the same bundle. The current data are consistent with the idea that Flamingo-dependent differential adhesion between R-cell heels prior to extension determines the extension angle (thus exercising a role in the scaffolding rule). In contrast, interactions between moving R-cell fronts are unlikely to instruct extension itself (no role in the extension rule). However, studies on the guidance receptor N-Cadherin suggest a role for the interaction of R-cell growth cones with L-cells in the target cartridge. These findings are consistent with a role of N-Cadherin-mediated interactions between R-cell fronts and L-cells as part of the stop rule and thereby indicate that L-cell interactions contribute to the stop rule. These interpretations of roles of Flamingo in the R-cell heel (as part of the scaffolding rule) and N-Cadherin in the R-cell front (as part of the stop rule) are further supported by their subcellular localization within the growth cone (Schwabe, 2013; Langen, 2015 and references therein).
Finally, the model supports R-cell front interactions as part of the stop rule. It is unclear to which degree this interaction is based on differential adhesion. How molecular signal integration is implemented to utilize the substantial increasing overlap of R-cell fronts as a stop signal remains to be discovered (Langen, 2015).
Both the equator and the rotational stereotypy of R-cell terminals have received little recent attention in the study of growth cone sorting and neural superposition, perhaps because they appear to be complications of an already complicated wiring problem. In particular, the findings of four types of rotational stereotypy within cartridges across the entire lamina have not been addressed in the literature since their discovery more than 40 years ago. The stereotypic arrangement of R1-R6 terminals in cartridges that encode precise neural superposition increases the apparent number of target slots 6-fold; yet, this arrangement is not required for neural superposition, given that all six carry the same information and synapse with the same output neurons. This study shows that evolutionary selection of the developmental algorithm that ensures precise axon sorting required for neural superposition wiring is sufficient to establish rotational stereotypy. While it is possible that rotational stereotypy may serve a function independent of neural superposition, selection for such a putative unknown function is not required to explain its occurrence. Hence, the fly's visual map provides an example for a neuronal circuit whose connectivity map can only be understood through its developmental context. Knowledge of a circuit's developmental algorithm may more generally help to explain aspects of neuronal circuits that cannot be derived from the study of the adult wiring diagrams alone (Lengen, 2015).
Precise control of neurite guidance during development is essential to ensure proper formation of neuronal networks and correct function of the central nervous system (CNS). How neuronal projections find their targets to generate appropriate synapses is not entirely understood. Although transcription factors are key molecules during neurogenesis, their entire function during the formation of networks in the CNS is not known. This study used the Drosophila melanogaster optic lobe as a model for understanding neurite guidance during development. The function of Sox102F/SoxD, the unique Drosophila orthologue of the vertebrate SoxD family of transcription factors, was assessed. SoxD is expressed in immature and mature neurons in the larval and adult lobula plate ganglia (one of the optic lobe neuropils), but is absent from glial cells, neural stem cells and progenitors of the lobula plate. SoxD RNAi knockdown in all neurons results in a reduction of the lobula plate neuropil, without affecting neuronal fate. This morphological defect is associated with an impaired optomotor response of adult flies. Moreover, knocking down SoxD only in T4/T5 neuronal types, which control motion vision, affects proper neurite guidance into the medulla and lobula. These findings suggest that SoxD regulates neurite guidance, without affecting neuronal fate (Contreras, 2018).
The Drosophila melanogaster visual system is composed of the retina and the optic lobe, which is divided into four ganglia: lamina, medulla, lobula and lobula plate. The visual inputs travel from the retinal photoreceptors through different optic lobe neurons, where this information is processed, triggering behavioural responses. Correct connectivity between optic lobe neurons is fundamental for sensing visual information. In the past few years, several studies have characterised how transcription factors regulate the development and neuronal composition of the optic lobe. However, while the development of the lamina and medulla has been extensively studied, research has only recently been focused on the development of the lobula complex (lobula and lobula plate) (Contreras, 2018).
The Sox (Sry Box) family of transcription factors is a key regulator of embryonic development. These transcription factors bear a conserved DNA binding domain known as the SRY-related High Mobility Group-box (HMG-box), which was first described in the Sry protein that is fundamental for sex determination in mammals. The Sox family of proteins is subdivided into nine groups, depending on the amino acid composition of their HMG-box. Vertebrate genomes encode approximately 20 members of the Sox family, whereas only eight members have been described in the fruit fly. The Sox transcription factors do not activate or repress gene expression themselves, but act together with partner factors that determine the modulation of target genes (Contreras, 2018).
Sox proteins are important during neural development and different groups of Sox transcription factors are responsible for similar neurodevelopmental processes across species. For instance, SoxB1 group members work in early neurogenesis in vertebrates and invertebrates. The vertebrate SoxB1 protein, Sox2, participates in early events of central and peripheral nervous system development. In a similar manner, the SoxB1 orthologues in Drosophila, SoxNeuro (SoxN) and Dichaete, are also required for proper neurogenesis and the formation of neural stem cells during development. On the other hand, SoxD proteins are generally involved later during nervous system development. Vertebrate Sox5 and Sox6 regulate neural stem cell proliferation, neuronal diversity, neuronal migration and projection formation. Similarly, the Drosophila SoxD orthologue is necessary for the development of the nervous system and loss of SoxD function affects synaptic bouton development at the neuromuscular junction and dendritic arborisation in sensory neurons (Contreras, 2018).
This study analysed the role of the Drosophila melanogaster orthologue of the SoxD family: Sox102F/SoxD during optic lobe development. SoxD is expressed in all optic lobe ganglia and is involved in the morphogenesis of the lobula plate neuropil. RNAi-mediated SoxD knockdown in developing neurons severely alters the morphology of the lobula plate ganglia. These morphological defects are not a consequence of changes in the fate of lobula plate neurons, but result from an alteration in the normal pattern of axon and dendrite formation. Associated with the defects in lobula plate morphology, the fly optomotor response is also impaired upon SoxD downregulation in lobula plate neurons. These results are consistent with the observation that Sox5 is involved in neuronal migration and axon pathfinding in mice, denoting that SoxD function is evolutionary conserved (Contreras, 2018).
According to a phylogenetic analysis, Sox102F has high homology to human Sox5, Sox6 and Sox13 transcription factors; therefore, it is proposed to rename this protein SoxD. During larval brain development, SoxD is expressed in neurons of the medulla and the lobula complex, while in the lamina, SoxD is transiently expressed before LPC differentiation into neurons. SoxD knockdown in all neurons or in lobula plate neurons severely affects the morphology of the lobula plate neuropil, impairing fly optomotor behaviour. Thus, this study shows that SoxD is required for the control of lobula plate T4 and T5 neurite guidance without affecting neuronal fate. Finally, it was demonstrated that misexpression of SoxD is sufficient to alter neurite guidance in photoreceptors and mushroom body Kenyon cells (Contreras, 2018).
The lobula plate is one of the less explored ganglia of the Drosophila visual system. Although neurogenesis has been described in some detail, later stages of development and the mechanisms directing neuronal subtype specification are starting to be described (Pinto-Teixeira, 2018). The lobula plate has an important role in motion detection in insects, while the lobula has a role in the integration of optic stimuli and the behavioural response. Two important neuronal populations in this regard are T4 and T5 neurons, which gather information from the medulla and lobula respectively and make their synaptic outputs in the lobula plate. Upon SoxD knockdown in all neurons (Elav-GAL4), specifically in T4/T5 neurons (IPC-GAL4 plus R42F06-Gal4) or only in T5 neurons (R42H07-GAL4), defects are observed in lobula plate morphology with increasing severity correlated with the number of affected neurons. This result suggests that SoxD may be also required for the developing of other lobula plate neurons and not only for T4/T5 neurons. In addition to RNAi-mediated knockdown experiments, a combination of soxD alleles was used that reduces the gene dosage. This hypomorphic condition showed a similar but milder phenotype, supporting the specificity of this phenotype to SoxD function. Furthermore, analysing SoxD knockdown phenotype in T4 or T5 neurons, alterations were observed in axon/dendrite guidance that could explain the motion perception defects of the adult animal (Contreras, 2018).
Recently, Li (2017) showed that SoxD is involved in neuronal development and degeneration. The Li study demonstrated that SoxD is required for synaptic bouton development at the neuromuscular junction, dendritic arborisation in sensory neurons, olfactory behaviour and climbing. This study describes a similar role for SoxD in the development of the lobula plate, but no evidence was found of apoptosis upon SoxD knockdown in larval stages, suggesting that the phenotype observed was not due to loss of lobula plate neurons but to neurite mistargeting during development. Moreover, it wasshowm that optomotor behaviour is also affected after SoxD knockdown, complementing Li's behavioural observations. This evidence lends support for the role of SoxD in axon and dendrite guidance in different types of neurons of the CNS (Contreras, 2018).
The mechanisms by which lobula plate neurite guidance is controlled by SoxD are unknown. Recently, it was described that Atonal promotes the differentiation of T4 and T5 neurons, while Notch signalling activity discriminates between T4 and T5 neuronal fates. This study observed that loss of SoxD did not affect T4/T5 differentiation markers, suggesting that the function of SoxD lays downstream of the T4/T5 fate decision. Thus, it is proposed that SoxD controls final stages of neuronal differentiation during development (Contreras, 2018).
The overgrowth of neurites observed upon SoxD knockdown in T4 and T5 neurons may result from defects in sensing an inhibitory guidance cue that restrict their growth into the medulla or the lobula. In accordance to this hypothesis, SoxD misexpression strongly affects neurite guidance in at least two different systems: photoreceptors and Kenyon cells. This supports the possible role of SoxD on sensing guidance cues. Additionally, the presence of the synaptic terminal marker in T5 dendrites after SoxD knockdown, suggests problems in neurite differentiation that could contribute to the guidance defects. Interestingly, the R42H07-GAL4 driver used to target T5 neurons was generated using an enhancer from the soxD locus. This enhancer does not recapitulate the entire expression of soxD, which is also expressed in T4 neurons and other lobula plate neurons. However, SoxD knockdown was associated to an increase of the GFP fluorescence driven by R42H07-GAL4, suggesting that SoxD may negatively regulate this enhancer (Contreras, 2018).
The regulation of neurite guidance may not be the only role of Drosophila SoxD during development. Over-expression of SoxD in embryonic neuroblasts and RNAi-mediated knockdown of SoxD in glial cells were reported to severely disrupt embryonic CNS development. Surprisingly, this study did not observe expression of SoxD in glial cells in the larval brain, while it remains unknown whether SoxD is expressed in embryonic glia. Furthermore, SoxD is also relevant for the function of other organs. SoxD knockdown in cardiac cells affects heart anatomy and function, while SoxD-RNAi expressed in wing discs increased the size of the longitudinal veins L2 and L3, and the marginal vein (Contreras, 2018).
Future work should address the signalling pathways upstream of SoxD activation and the SoxD targets that govern the morphogenesis of lobula plate neurons. Interestingly, Sox5 knockout mice show defects in axonal pathfinding of corticothalamic neurons, similar to Drosophila T4/T5 neurons, suggesting a conserved role of SoxD proteins in neurite guidance (Contreras, 2018).
A recent paper depicted the role of Sox5 in the regulation of the Collapsin Response Mediator Protein (CRMP), an intracellular protein involved in neurite guidance. Interestingly the authors showed that Sox5 gain of function reduces neurite guidance through CRMP in hippocampal neurons, shedding some light to the molecular mechanism involved. Future work should address the conservation of this regulation (Contreras, 2018).
Finally, human Sox5 has been implicated in a number of diseases and intellectual disability in humans. Several studies report mutations and deletions in the sox5 locus that are linked to developmental defects. Therefore, using the fly as a model for neurite guidance may be valuable in determining the biological impact of these mutations in the onset of neurological diseases (Contreras, 2018).
Stereotyped synaptic connections define the neural circuits of the brain. In vertebrates, stimulus-independent activity contributes to neural circuit formation. It is unknown whether this type of activity is a general feature of nervous system development. This study reports patterned, stimulus-independent neural activity in the Drosophila visual system during synaptogenesis. Using in vivo calcium, voltage, and glutamate imaging, it was found that all neurons participate in this spontaneous activity, which is characterized by brain-wide periodic active and silent phases. Glia are active in a complementary pattern. Each of the 15 of over 100 specific neuron types in the fly visual system examined exhibited a unique activity signature. The activity of neurons that are synaptic partners in the adult was highly correlated during development. It is proposed that this cell-type-specific activity coordinates the development of the functional circuitry of the adult brain (Akin, 2019).
Synaptic connections between neurons determine how neural circuits process information. Understanding how the specificity of these connections is established is a central challenge in neurobiology. In vertebrates, cell-autonomous genetic programs and neural activity (both evoked and spontaneous) contribute to synapse development. Spontaneous activity has been observed throughout the developing CNS. Retinal waves are the best characterized examples of spontaneous activity. In mammals, retinal waves occur between the completion of axon guidance and eye opening, and the activity propagates from the retina to higher-order visual centers. Significant progress has been made toward characterizing the organizing principles of spontaneous activity in the developing vertebrate brain, and the precise developmental role of this activity is an area of active interest.
By contrast to vertebrates, brain development in invertebrates has been thought to occur largely independent of activity. In the Drosophila visual system, photoreceptor neurons can develop the wild-type complement of synapses in a stimulus-independent manner. However, the existence and significance of spontaneous activity during invertebrate brain development remains an open question (Akin, 2019).
Some of the most detailed understanding of brain development in the fly comes from the visual system. Visual information from the compound eye is topographically relayed to the optic neuropils -- the lamina, medulla, and lobula complex, each organized into layers and columns. Over 100 different neuronal cell types form precise synaptic connections, typically with several different cell types as revealed by 3D electron microscopy (EM) reconstructions (Akin, 2019).
Visual system development in the fly takes place during the last stage of larval development and the ensuing 100 h of metamorphosis, or pupal development. In the first 50 h after puparium formation (hAPF), a dynamic self-assembly process mediated by cell-surface molecules brings most of the cell types of the visual system to where they belong in the adult brain, ready for synaptogenesis. Over the remaining 50 h of pupal development, synapse assembly proceeds in parallel with the upregulation of genes involved in neural activity and new sets of cell recognition molecules, and vast networks comprising distant cells are assembled through specific synaptic connections and compatible gene expression profiles (e.g., matching neurotransmitter systems and receptors). Little is known about the molecules and mechanisms that coordinate this period of brain development (Akin, 2019).
This study reports the discovery and initial characterization of patterned, stimulus-independent neural activity (PSINA) in the developing fly visual system. Three distinct stages of PSINA were uncovered: a periodic stage (55 to 65 hAPF), a turbulent stage (70 hAPF to the final hour of pupal development), and an adult stage that persists alongside mature stimulus responses through at least five days after eclosion. In the periodic stage, which coincides with the onset of synaptogenesis in the fly CNS, each of the 15 neuronal cell types analyzed exhibited stereotyped and distinct activity patterns. Many adult synaptic partners had correlated activity, which depended on synaptic release (Akin, 2019).
Distinct periodic calcium dynamics were also seen in astrocytes. Astrocytes in the developing adult brain elaborate processes that infiltrate the neuropil during synapse formation. Ablating astrocytes significantly reduces the total synapse count in the brain, supporting a role for these cells in regulating synaptogenesis. The finding that astrocytes exhibit cycles of calcium activity alongside PSINA raises the possibility that astrocytes and PSINA contribute to the formation, specificity, or maturation of synapses within the visual system (Akin, 2019).
Work on retinal waves supports a role for stimulus-independent activity in synaptic development. In the vertebrate visual system, retinal ganglion cell (RGC) projections target the lateral geniculate nucleus (LGN) and the superior colliculus (SC), where they create retinotopic maps of the visual field and segregate based on the eye of origin. Retinotopy and eye-specific segregation in the LGN and SC are refined during cholinergic retinal waves. Early work in the field established that pharmacological manipulation of spontaneous activity in the cat retina disrupts the organization of RGC projections in the LGN. Later studies, using progressively more refined methods, have shown that disrupting the cholinergic circuit of starburst amacrine cells and RGCs largely eliminates retinal waves and leads to defects in the refinement of retinotopy and eye-specific segregation of RGC projections (Akin, 2019).
The diversity of cell types in the vertebrate retina (as many as 30 for RGCs alone) is comparable to the fly visual system. Whether there is cell-type-specific texture to the retinal waves similar to PSINA described in this study is not known, although broad classes of RGCs and cone bipolar cells have been shown to exhibit temporally offset firing patterns. With improving genetic handles for distinct retinal cell types and ongoing efforts at describing the high-resolution connectome, it will be possible to explore cell-type-specific patterns and the contribution of retinal waves to retinal circuitry (Akin, 2019).
In Drosophila, peristaltic contractions of body wall muscles have recently been appreciated as part of broad neuronal activity during embryonic development. This activity is similar to PSINA observed during pupal development with respect to periodicity and timing relative to synapse formation and refinement. Preventing motor neurons from participating in this neuronal activity, or disrupting calcium-dependent intracellular signaling, results in ectopic synapses (Akin, 2019).
Based on previous studies in the fly visual system and the role of spontaneous activity in other systems, a general conceptual framework is proposed for the role of PSINA in regulating the assembly of the adult connectome. The results suggest that some adult synaptic pairings are already established by 55 hAPF, soon after the first pre-synapses can be detected and coincident with the onset of PSINA. The global coordination of PSINA indicates that an early connectome, one that must be built through largely contact-dependent interactions of over 100 different neuronal cell types, is present at this time. While the level of organization achieved through such activity-independent mechanisms is astonishing, the early connectome may still be an approximation of what is required in the adult (Akin, 2019).
PSINA, by orchestrating cellular communication at temporal and spatial scales inaccessible to other signaling mechanisms, may act to refine this draft connectome to complete the self-assembly of the brain. The diversity of spatiotemporal patterns in PSINA presents many opportunities to establish circuit-specific topographic integration. Sweeps of activity repeatedly coursing through the brain through different 'channels' could link distinct sets of neurons to direct coordinated morphological changes and sculpt cell-cell contacts, strengthen synapses with correct targets while weakening incorrect pairings, and control transcription programs that direct circuit refinement. PSINA may act as a 'dress rehearsal' for neural networks, preparing for 'opening night' at the completion of development. Individual cells know their own lines, with whom they interact, and their respective positions on the stage; however, repeated practice of each scene ensures that each of the cast can perform as part of a whole ensemble (Akin, 2019).
It is remarkable that as a process that engages most, if not all, of the CNS, PSINA is the collective output of the genetically hardwired developmental programs of individual neurons. Thus, the organizing principles, driving forces, and functional significance of PSINA at the level of circuits, cells, and molecules should be discoverable through genetic analysis. Undertaking this effort in the fly visual system, where structures analogous to the vertebrate retinal plexiforms, LGN, and SC are compactly organized in a single microscopic field of view and for which the EM-derived connectome is available, may yield valuable insights into whether and how PSINA affects synaptic specificity and circuit maturation. It is expected that the ever-expanding genetic toolkit of Drosophila, complemented with improvements in genomic-transcriptomic analysis and imaging technology, will offer a robust experimental track toward understanding PSINA's contribution to brain development and function (Akin, 2019).
Drosophila R7 UV photoreceptors (PRs) are divided into yellow (y) and pale (p) subtypes. yR7 PRs express the Dpr11 cell surface protein and are presynaptic to Dm8 amacrine neurons (yDm8) that express Dpr11's binding partner DIP-γ, while pR7 PRs synapse onto DIP-γ-negative pDm8. Dpr11 and DIP-g expression patterns define 'yellow' and 'pale' color vision circuits. This study examined Dm8 neurons in these circuits by electron microscopic reconstruction and expansion microscopy. DIP-γ and dpr11 mutations affect the morphologies of yDm8 distal ('home column') dendrites. yDm8 neurons are generated in excess during development and compete for presynaptic yR7 PRs, and interactions between Dpr11 and DIP-γ are required for yDm8 survival. These interactions also allow yDm8 neurons to select yR7 PRs as their appropriate home column partners. yDm8 and pDm8 neurons do not normally compete for survival signals or R7 partners, but can be forced to do so by manipulation of R7 subtype fate (Menon, 2019).
Visual motion detection in sighted animals is essential to guide behavioral actions ensuring their survival. In Drosophila, motion direction is first detected by T4/T5 neurons. Their axons innervate one of the four lobula plate layers. How T4/T5 neurons with layer-specific representation of motion-direction preferences are specified during development is unknown. This study shows that diffusible Wingless (Wg) between adjacent neuroepithelia induces its own expression to form secondary signaling centers. These activate Decapentaplegic (Dpp) signaling in adjacent lateral tertiary neuroepithelial domains dedicated to producing layer 3/4-specific T4/T5 neurons. T4/T5 neurons derived from the core domain devoid of Dpp signaling adopt the default layer 1/2 fate. Dpp signaling induces the expression of the T-box transcription factor Optomotor-blind (Omb), serving as a relay to postmitotic neurons. Omb-mediated repression of Dachshund transforms layer 1/2- into layer 3/4-specific neurons. Hence, spatio-temporal relay mechanisms, bridging the distances between neuroepithelial domains and their postmitotic progeny, implement T4/T5 neuron-subtype identity (Apitz, 2018).
Visual signals received by the retina are generally not stationary because objects in the environment and/or the bodies of animals move. To detect motion, visual circuits perform complex spatio-temporal comparisons that convert luminance changes collected by photoreceptors into signals containing information about direction or speed. Despite the seemingly divergent anatomy of vertebrate and insect visual systems, they display remarkable parallels in the computations underlying motion vision and the neuronal elements performing them. In most sighted animals, this involves neurons that respond to motion signals in specific directions. Direction-selectivity emerges from differences in the connectivity of their dendrites. Motion-direction preferences by their axons are represented by layer-specific innervation. Thus, anatomical characteristics such as layer-specificity seem to be intricately linked with motion-directionality. However, how these are implemented during circuit development is poorly understood (Apitz, 2018).
The Drosophila visual system has emerged as a powerful model for elucidating the neural circuits and computations underlying motion detection. Photoreceptors (R-cells) in the retina extend axons into the optic lobe consisting of the lamina, medulla, lobula plate, and lobula. Neuronal projections in these ganglia are organized into retinotopically arranged columnar units. The medulla, lobula plate, and lobula are additionally subdivided into synaptic layers. They are innervated by more than a 100 neuronal subtypes that extract different visual features in parallel pathways. T4 and T5 lobula plate neurons are the first direction-selective circuit elements. Each optic lobe hemisphere contains ~5300 T4/T5 neurons. T4 dendrites arborize within medulla layer 10, and T5 dendrites in lobula layer Lo1. Their axons project to one of the four lobula plate layers, thereby defining four different neuron subtypes each. Axons segregate according to their motion-direction preferences. Thus, front-to-back, back-to-front, upward, and downward cardinal motion directions are represented in lobula plate layers. T4 neurons are part of the ON motion detection pathway reporting brightness increments, while T5 neurons are part of the OFF pathway reporting brightness decrements. Distinct neuron sets in the lamina and medulla relay ON and OFF information to T4 and T5 neurons. Direction-selectivity emerges within T4/T5 dendrites and involves the non-linear integration of input from these upstream neurons for enhancement in the preferred direction and suppression in the null-direction. Dendritic arbors of the four T4 neuron subtypes have characteristic orientations, that correlate with the direction preferences of lobula plate layers innervated by their axons. Thus, direction-selectivity involves the establishment of neuron subtypes, each with distinct spatial connectivities. This study addresses when and how T4 and T5 neuron subtypes with different layer identities are specified during development (Apitz, 2018).
Optic lobe neurons originate from two horseshoe-shaped neuroepithelia, called the outer and inner proliferation centers (OPC and IPC). These are derived from the embryonic optic lobe placode and expand by symmetric cell divisions during early larval development. At the late 2nd instar larval stage, neuroepithelial (NE) cells from the medial OPC edge begin to transform into medulla neural stem cells, called neuroblasts (Nbs). These undergo asymmetric divisions to self-renew and give rise to ganglion mother cells (GMCs), which divide to generate two neurons or glia. Apposing the OPC, two dorsal and ventral NE domains, called the glial precursor cell (GPC) areas, produce neuron subtypes associated with all ganglia. At the mid 3rd instar larval stage, the lateral OPC begins to generate lamina neurons (Apitz, 2018).
The IPC generates lobula and lobula plate neurons, including T4/T5 neurons from the early 3rd instar larval stage onward. Recent studies showed that NE cells in one domain, the proximal (p-)IPC, convert into progenitors in an epithelial-mesenchymal transition (EMT)-like process. Progenitors migrate to a second proliferative zone, the distal (d-)IPC, where they mature into Nbs. These transition through two competence windows to first produce C and T neurons, corresponding to C2 and C3 ascending neurons connecting the medulla and lamina, as well as T2/T2a and T3 neurons connecting the medulla and lobula, and then T4/T5 lobula plate neurons. Cross-regulatory interactions between Dichaete (D) and Tailless (Tll) control the switch in Nb competence defined by the sequential expression of the proneural bHLH transcription factors Asense (Ase) and Atonal (Ato). The latter is co-expressed with the retinal determination protein Dachshund (Dac). The molecular mechanisms that control layer-specific T4/T5 neuron subtype identities within this sequence of developmental events occurring at different locations have remained elusive (Apitz, 2018).
T4/T5 neuron diversity resulting in differential layer-specificity could be achieved by postmitotic combinatorial transcription factor codes upstream of distinct guidance molecules. Although not mutually exclusive, layer-specificity of T4/T5 neurons could also be determined by temporal differences in the expression of common postmitotic determinants, similar to the birth-order dependent R-cell growth cone segregation strategy described in the medulla. This study provides evidence for another mechanism, whereby layer-specific T4/T5 neuron subtype identity is determined early in the p-IPC neuroepithelium. Their specification depends on two relay mechanisms involving Wnt and Bone morphogenetic protein (Bmp) signaling and transcription factor interactions. These establish and translate the spatial patterning of NE cells into postmitotic neuronal subtype identities to bridge distances inherent to this particular neurogenesis mode (Apitz, 2018).
The spread of Wg is dispensable for patterning of many tissues. However, this study uncovered a distinct requirement for diffusible Wg in the nervous system, where it orchestrates the formation of T4/T5 neurons innervating lobula plate layers 3/4. Their generation depends on inductive mechanisms that are relayed in space and time. The spatial relay consists of a multistep-signaling cascade across several NE domains: Wg from the GPC areas induces wg expression in the s-IPC and Nb lineage adjacent to ventral and dorsal p-IPC subdomains; this secondary Wg source activates dpp expression. Dpp signaling mediates EMT of migratory progenitors from these subdomains. The p-IPC core produces Dac-positive layer 1/2 specific T4/T5 neurons. Dpp signaling in p-IPC NE subdomains triggers a temporal relay across intermediate cellular states by inducing omb. Omb in turn suppresses Dac, conferring layer 3/4 identity to postmitotic T4/T5 neurons (Apitz, 2018).
When Wg is membrane-tethered, the first step of this cascade is disrupted. This defect is not caused by decreased signaling activity of NRT-Wg protein in wg{KO;NRT-wg} flies. First, wild-type Wg signaling activity inside the GPC areas and the adjacent OPC was not affected. Second, in allele switching experiments, ectopic expression of a highly active UAS-NRT-wg transgene in the GPC areas was unable to rescue. By contrast, restoring wild-type wg function in the GPC areas was able to rescue, supporting the notion that Wg release and spread from the GPC areas are required to induce its own expression in the s-IPC and the Nb clone (Apitz, 2018).
Although Wg release is essential, the range of action is likely limited. Wg expression in the s-IPC commences in early 3rd instar larvae, when it is still in close proximity with the GPC. Half of the wg{KO;NRT-wg} flies showed residual dpp expression in one progenitor stream at the 3rd instar larval stage and a 25% reduction of T4/T5 neurons, correlating with three lobula plate layers in adults. The other half lacked dpp-lacZ expression and showed a 50% reduction of T4/T5 neurons correlating with two remaining layers. While this partial phenotypic penetrance is not fully understood, NRT-Wg likely partially substituted for Wg because of the initial close proximity of the GPC areas and the s-IPC and Nb clone. Occasional residual NRT-Wg expression in the s-IPC argues against an all-or-nothing inductive event and suggests a model, whereby cell-intrinsic signaling thresholds have to be reached. Theoretically, the dpp expression defect in the p-IPC of wg{KO;NRT-wg} flies could reflect the dependence on long-range Wg from the GPC areas. However, as this study has shown, IPC-specific wg knockdown leads to dpp loss in the p-IPC. Propagation of sequential Wnt signaling could explain long-range activities. Moreover, sequentially acting primary and secondary sources of Wg have been described in the developing Drosophila eye, suggesting that the regulatory mechanism observed in the optic lobe might be employed in several contexts. The different outcomes of early and late allele wg to NRT-wg allele switching indicate that Wg secretion is required for the induction but not long-term maintenance of wg expression in the s-IPC. The GPC areas become rapidly separated from the s-IPC and Nb clone by compact rows of newly generated neurons. As part of a relay system, diffusible Wg may therefore be required to bridge distances over a few cell diameters during the initial phase of neurogenesis. The s-IPC in wg{KO;NRT-wg} flies expressed Hth and generated two neuron clusters as in wild-type. Thus, the sole function of wg in the s-IPC is to relay the GPC-derived Wg signal to induce dpp expression in the p-IPC. Since Wg release is not required in the GPC areas to induce dpp in the adjacent OPC, this secondary wg function in the s-IPC is most likely juxtacrine (Apitz, 2018).
Compared to approximately 80 medulla neuron subtypes derived from the OPC, the specification of 13 distinct subtypes originating from the p-IPC appears simple. However, the distinct mechanisms employed are surprisingly complex. Previous work has shown that cross-regulatory interactions between D and tll regulate a Nb competence switch from generating early-born C2, C3, T2, T2a, and T3 neurons to eight distinct layer-specific T4/T5 subtypes. Ato and Dac are expressed in the second Nb competence window and depend on tll. Functional studies showed that dac mutant T4/T5 neurons adopted early-born T2/T3 neuron-like morphologies. Similarly, ato mutant T4/T5 neurons displayed neurite connectivity defects. Notably, simultaneous knockdown of dac and ato resulted in the absence of T4/T5 neurons, demonstrating that both are required together for the ability of d-IPC Nbs to produce new neuron subtypes in the second competence window (Apitz, 2018).
Dac is initially expressed in all T4/T5 neurons but only maintained in layer 1/2 innervating subtypes. This suggests that an essential step for the specification of layer 3/4 innervating neurons is the downregulation of Dac and the suppression of the T4/T5 default neuron fate, i.e., layer 1/2 identity. Although the mode of this inhibitory mechanism depends on the outcome of the Nb-specific switching mechanism in the d-IPC, it is already primed in p-IPC NE cells. Thus, layer-specificity and therefore motion-directionality are determined early in the NE precursors of T4/T5 neurons. Molecularly, it involves the Omb-mediated relay of Dpp-signaling-dependent NE cell patterning information across intermediate cell states to postmitotic T4/T5 neurons resulting in the repression of Dac. In contrast to the OPC, this study found no link between NE patterning in the p-IPC and Notch-dependent differential apoptosis of region-specific T4/T5 subtypes. Instead, Notch controls the choice between T4 and T5 identity, likely during the second competence window, indicating that the distinction between layer 1/2 and 3/4 fates precedes T4 and T5 neuron specification (Apitz, 2018).
The mechanisms controlling the maintenance of omb expression, and Omb-mediated downregulation of Dac are unclear. Hypotheses regarding the latter have to be reconciled with the fact that dac, together with ato, is required for the formation of all T4/T5 neurons and hence is expressed in all d-IPC Nbs during the second competence window. Omb and Dac are initially co-expressed in Nbs and young T4/T5 neurons, suggesting that Omb does not directly repress dac transcription. Yet, expression of the dacp7d23 enhancer trap Gal4 line showed that dac is only transcribed in layer 1/2 neurons in adults. A possible scenario is that Omb could break Dac autoregulation by triggering degradation of Dac. Since T-box genes can act as transcriptional activators and repressors and their effects are influenced by various co-factors, future studies will need to explore the molecular details underlying Omb-mediated repression of Dac. It will also be important to determine whether layer 3/4 specification is mediated solely by Dac downregulation, or whether omb has additional instructive roles (Apitz, 2018).
Consistent with the observation that C2 and C3 neurons have distinct developmental origins, this study found that Nbs derived from the Dpp-expression domain produce C2 and possibly T2a neurons during the first Nb competence window, while the core p-IPC generates C3, T2, and T3 neurons. dac mutant T4/T5 neurons adopt T2/T3-like morphologies suggesting that this is the default neuron fate in this neuron group. While Omb is maintained in C&T neurons derived from the Dpp-expression domain, Dac is not expressed, suggesting that Omb interacts with other molecular determinants in these neurons. While this study did not explore how layer 1 and 2 neurons or layer 3 and 4 neurons become distinct from each other because of the lack of specific markers, the data suggest a possible contribution of Ato/Dac and Notch signaling, as these are active within the d-IPC. Findings in a concurrent study of Pinto-Teixeira (2018) align with the current data concerning the role of Dpp and Notch signaling. Furthermore, a second study of Mora (2018) reported an additional role for Ato in controlling the transient amplification of d-IPC Nbs by symmetric cell division to ensure that the correct number of T4/T5 neurons is produced. It will be fascinating to identify the transcriptional targets of Notch, Ato/Dac, and Omb that mediate ganglion- and layer-specific targeting of T4/T5 dendrites and axons, respectively. Finally, future behavioral studies of layer 3/4-deficient flies will address to what extent direction selectivity is affected or compensatory mechanisms are in place (Apitz, 2018).
Signaling centers, also called organizers, pattern tissues in a non-autonomous fashion. The vertebrate roof plate and the cortical hem, for instance, both release Wnts and Bmps to pattern NE cells in the developing dorsal spinal cord and in the surrounding forebrain, respectively. In the Drosophila visual system, the GPC areas express wg and pattern the OPC by inducing dpp expression in adjacent dorsal and ventral OPC subdomains. Together with the current insights into the function of GPC-derived wg in IPC patterning and neurogenesis, this firmly establishes the GPC areas as local organizers of optic lobe development. At the onset of neurogenesis, wg is first expressed in the GPC areas followed by the s-IPC, explaining the well-established delay in neurogenesis between the IPC and OPC. Wg release from the GPC areas could coordinate the timely onset of neurogenesis in the OPC and IPC to safeguard the alignment of matching partner neurons across several retinotopically organized neuropils. The intercalation of new-born neurons between both neuroepithelia may have driven the need for a relay system using primary and secondary sources of Wg. Wg induces Dpp to subdivide the adjacent OPC and p-IPC NE into distinct regions as basis for generating neuronal diversity. The temporal relay mediated by Omb represents an efficient strategy to pass the memory of spatial NE patterning information by Dpp signaling on to postmitotic neurons generated at a distance. It is thus intricately tuned to the distinct neurogenesis mode of the p-IPC essential for spatially matching birth-order-dependent neurogenesis between the OPC and IPC. Interestingly, the progressive refinement of NE patterning by the induction of secondary signaling centers plays a central role in vertebrate brain development. Furthermore, similar signaling cascades have been recently identified in mammalian optic tissue cultures where sequential Wnt and Bmp signaling induces the expression of the Omb-related T-box transcription factor Tbx5 to specify dorsal retinal NE cells. Hence, such cascades could represent conserved regulatory modules that are employed repeatedly during invertebrate and vertebrate nervous system development (Apitz, 2018).
Patterns of synaptic connectivity are remarkably precise and complex. Single-cell RNA sequencing has revealed a vast transcriptional diversity of neurons. Nevertheless, a clear logic underlying the transcriptional control of neuronal connectivity has yet to emerge. This study focused on Drosophila T4/T neurons, a class of closely related neuronal subtypes with different wiring patterns. Eight subtypes of T4/T neurons are defined by combinations of two patterns of dendritic inputs and four patterns of axonal outputs. Single-cell profiling during development revealed distinct transcriptional programs defining each dendrite and axon wiring pattern. These programs were defined by the expression of a few transcription factors and different combinations of cell surface proteins. Gain and loss of function studies provide evidence for independent control of different wiring features. It is proposed that modular transcriptional programs for distinct wiring features are assembled in different combinations to generate diverse patterns of neuronal connectivity (Kurmangaliyev, 2019).
T4/T5 neurons share a common developmental origin, physiological function, and general morphology, but differ in their precise wiring patterns and preferred stimulus. There are eight morphological subtypes of T4/T5 neurons in each column of the lobula plate (LoP) neuropil, comprising the most abundant cell type in the fly visual system. These subtypes can be classified into two quartets of subtypes based on dendritic inputs: the four T4 subtypes share a common set of dendritic inputs in the medulla, and the four T5 subtypes share a different set of dendritic inputs in the lobula (see Single-cell sequencing reveals eight transcriptionally distinct populations of T4/T5 neurons). T4 neurons respond to ON stimuli (i.e. bright edges moving against a dark background) and T5 to OFF stimuli (i.e., dark edges moving across a bright background). T4/T5 neurons can also be classified into four pairs of subtypes (a-d) based on the location of their axon terminals within a given column in layers a-d of the LoP. Each pair responds selectively to visual motion in one of four cardinal directions: posterior, anterior, upwards, and downwards, respectively. Although transcriptional profiling of the adult Drosophila brain revealed a common transcriptional signature for all T4/T5 neurons, genetic programs for individual subtypes have not been identified. This study hypothesized that identification of gene expression programs for individual T4/T5 subtypes during circuit assembly would provide insight into the genetic programs regulating discrete wiring features (Kurmangaliyev, 2019).
This study reports that independent transcriptional programs define the dendritic inputs and axonal outputs of T4/T5 neurons. Gain and loss of function studies indicate that these programs control their corresponding morphological features. These findings suggest that the modular assembly of separate dendritic and axonal transcriptional programs contributes to the diversity of wiring patterns in complex nervous systems (Kurmangaliyev, 2019).
A unique attribute of T4/T5 neurons is that the same dendritic and axonal wiring patterns are reiteratively used among different subtypes; each neuron can be described by a unique combination of one of four types of axonal outputs and one of two types of dendritic inputs. It was anticipated that this property of T4/T5 neurons would provide an opportunity to assess the relationship between specific genetic programs and fundamental features of neuronal architecture (Kurmangaliyev, 2019).
Unsupervised analysis revealed that separable transcriptional programs correlate with these specific wiring features. This study demonstrates through gain and loss of function experiments that these programs control specific axonal targeting features, which are separable from other features (e.g., dendrite targeting). These programs can be re-assembled in a modular fashion to generate neuronal subtypes with different combinations of wiring features. A modular transcriptional architecture may provide a general strategy for discrete modifications to neuronal connectivity in development and evolution (Kurmangaliyev, 2019).
A common T4/T5 neuronal identity is defined by a unique combination of TFs expressed in all subtypes (e.g., Lim1, Drgx, acj6). Perturbation of TFs expressed in all subtypes disrupts overall organization of T4/T5 neurons, including both dendritic and axonal morphologies. This common T4/T5 transcriptional program is further diversified by separable feature-specific transcriptional programs. These programs are defined by three binary (ON/OFF) TF expression patterns, with two TF patterns defining the four axonal outputs and one TF pattern defining the two dendritic inputs. In this way, modular TF codes defining common and feature-specific transcriptional programs give rise to eight T4/T5 subtypes (Kurmangaliyev, 2019).
Four pairs of T4/T5 subtypes with shared axonal outputs (and different dendritic inputs) each target one of four LoP layers, a-d. The ultimate layered architecture of neuropils develops through sequential lamination into increasing numbers of layers. Together with previous results, the findings suggest that the lamination of T4/T5 axonal outputs occurs via two distinct processes, each controlled by a separate TF. Binary expression of bi is required for lamination of the broad a/b from c/d LoP domains (Apitz, 2018), whereas binary expression of grn is required for sublamination of each of these two domains into separate LoP layers. Importantly, perturbation of each TF exclusively disrupts the corresponding lamination step, while not affecting other morphological features of T4/T5 neurons. Similarly, two quartets of subtypes with shared dendritic inputs (and different axonal outputs) were defined by binary expression of TfAP-2. Arborization of dendrites in M10 (T4) or Lo1 (T5) occurs during initial neurite guidance steps, preceding the developmental stages covered in this study. It is hypothesized that differentially expressed genes (DEGs) between T4 and T5 subtypes identified in this analysis contribute to the connections with two distinct sets of presynaptic partners (Kurmangaliyev, 2019).
The binary expression patterns of TFs also mirror the developmental lineages of T4/T5 neurons. a/b and c/d subtypes arise from bi- and bi+ progenitor populations. Neuroblasts from each population undergo two terminal Notch-dependent asymmetric divisions to give rise to the eight subtypes (Pinto-Teixeira, 2018). These divisions correspond to binary expression patterns of grn and TfAP-2, respectively, which act with Notch signaling to regulate wiring. Remarkably, despite divergent developmental trajectories separated by multiple divisions and distinct progenitor pools, all T4 and all T5 subtypes converge onto the same transcriptional programs associated with two types of dendritic inputs. Three regulatory dichotomies could also reflect the evolutionary origin of T4/T5 subtypes and correspond to consecutive duplications of ancestral cell types and circuits (Kurmangaliyev, 2019).
Each axonal and dendritic transcriptional program is characterized by a specific pattern of TFs, as well as a set of cell surface proteins (CSPs), many of which are implicated in regulating wiring in other developmental contexts. These include Ig superfamily proteins in which different paralogs exhibit discrete heterophilic binding specificities, including the beat/side and the dpr/DIP interacting protein families. Interestingly, dynamic expression of these proteins in neurons with shared wiring features was developmentally coordinated. It is envisioned that the synaptic specificity of T4/T5 dendrites and axons are determined by the combined activity of these recognition molecules through interactions with synaptic partners. Future experiments utilizing gain and loss of function analysis, either alone or different combinations, will provide insights into the cellular recognition mechanisms by which synaptic specificity is established (Kurmangaliyev, 2019).
The composite morphological properties of T4/T5 subtypes allowed identification, and thus decouple transcriptional programs for dendrite and axon wiring. Combining separate dendritic and axonal programs, and variations on them, may contribute to the diversification of synaptic specificity in different neuronal subtypes across complex nervous systems (Kurmangaliyev, 2019).
Direction-selective T4/T5 neurons exist in four subtypes, each tuned to visual motion along one of the four cardinal directions. Along with their directional tuning, neurons of each T4/T5 subtype orient their dendrites and project their axons in a subtype-specific manner. Directional tuning, thus, appears strictly linked to morphology in T4/T5 neurons. How the four T4/T5 subtypes acquire their distinct morphologies during development remains largely unknown. This study investigated when and how the dendrites of the four T4/T5 subtypes acquire their specific orientations, and profiled the transcriptomes of all T4/T5 neurons during this process. This revealed a simple and stable combinatorial code of transcription factors defining the four T4/T5 subtypes during their development. Changing the combination of transcription factors of specific T4/T5 subtypes resulted in predictable and complete conversions of subtype-specific properties, i.e. dendrite orientation and matching axon projection pattern. Therefore, a combinatorial code of transcription factors coordinates the development of dendrite and axon morphologies to generate anatomical specializations differentiating subtypes of T4/T5 motion-sensing neurons (Hoermann, 2020).
The development of neuronal morphology relies on the interplay between cell-intrinsic factors, i.e. genetic programmes, and extracellular cues, e.g. growth factors. This study investigated the genetic programmes underlying the acquisition of the different morphologies defining the four T4/T5 subtypes, which are essential for detecting visual motion along the four cardinal directions. The development of T4/T5 subtype-specific morphologies relies on a postmitotic combinatorial code of transcription factors. In particular, Grain acts together with different transcription factors in T4/T5b and T4/T5c subtypes to coordinate dendrite and axon morphogenesis in order to differentiate their morphologies from those of T4/T5a and T4/T5d, respectively. The coordinated regulation of different aspects of neuron morphogenesis by the same set of transcription factors might provide a general strategy to ensure the establishment of precise neuron wiring patterns during development. Furthermore, the combinations of transcription factors controlling the development of subtype-specific properties do not appear to regulate morphological properties that are common to all T4/T5 subtypes, i.e. the restriction of dendrites and axons to single neuropil layers. The acquisition of these morphological properties is controlled postmitotically by two transcription factors of the Sox family, SoxN and Sox102F. Therefore, distinct sets of transcription factors control differentially subtype-specific properties and properties that are shared by all T4/T5 subtypes, shedding light on the developmental strategies that ensure that each neuron acquires its complete morphological signature (Hoermann, 2020).
How do the expression patterns of T4/T5 postmitotic transcription factors arise during development? SoxN and Sox102F expression in all T4/T5 subtypes arises from temporal patterning of the neuroblasts that are precursors of T4/T5 neurons. Spatial patterning of the neuroepithelium that generates T4/T5-producing neuroblasts results in the specific expression of omb in neuroblasts that are precursors of T4/T5c,d neurons. The expression of omb is further relayed to postmitotic developing T4/T5c,d neurons, in which it represses dac expression. In addition, each Omb− neuroblast (precursor of T4/T5a,b), as well as each Omb+ neuroblast (precursor of T4/T5c,d), divides to produce two ganglion mother cells, only one of which has Notch activity. Only ganglion mother cells with Notch activity generate T4/T5a and T4/T5d neurons (Pinto-Teixeira, 2018). The transcriptional programmes downstream of this Notch-dependent fate decision remain elusive. Notch activity has been shown to repress grain in the aCC motoneuron of the Drosophila embryo. In agreement with this, grain is not expressed in T4/T5a,d neurons originating from ganglion mother cells with Notch activity. Together, these observations suggest that the specific expression of grain in postmitotic T4/T5b,c neurons could result from the Notch-dependent fate decision occurring during the final division of T4/T5-producing neuroblasts. Future studies will need to investigate how the T4/T5 subtype-specific expression of grain, as well as of ab and pros, is achieved during development (Hoermann, 2020).
The current data revealed that only one transcription factor, Grain, defines T4/T5b,c neurons during development. By contrast, T4/T5a,b and T4/T5c,d are each defined by two transcription factors: Dac and Ab are co-expressed in T4/T5a,b, whereas Omb and Pros are co-expressed in T4/T5c,d. These transcription factors with overlapping expression patterns might play redundant roles. Alternatively, they might be specialised to control different aspects of development. Systematic manipulations of the expression patterns of these transcription factors will be needed to address these possibilities, as well as to further elucidate how they act in a combinatorial manner to determine the different morphologies of the four T4/T5 neuron subtypes (Hoermann, 2020).
Transcription factors control dendrite growth, in part by controlling the expression of genes relevant for sensing extrinsic cues. This study found many cell-membrane proteins with T4/T5 subtype-specific expression patterns that might result from the action of the combinatorial code of transcription factors that was uncovered in this study. In agreement with the results of a recent publication, the vast majority of cell-membrane proteins with subtype-specific expression patterns in T4 neurons exhibited the same expression patterns and dynamics in T5 neurons. These mostly included receptors, ligands, regulators of various signalling pathways, and cell-adhesion molecules, some of which have been shown to be involved in axon guidance, dendrite patterning and/or synaptic specificity in Drosophila. It is hypothesised that those cell-membrane proteins with stable subtype-specific expression patterns during, at least, the first phase of dendrite growth are the most likely candidates to regulate the development of the four dendrite orientations in a combinatorial way. However, they might also control other subtype-specific properties, e.g. axon projection patterns and connectivity with distinct postsynaptic neurons. Collectively, these data indicate that the four T4 and T5 subtypes share combinations of transcription factors and downstream effector genes that might control the development of four dendrite orientations. Yet, T4 dendrites grow in the medulla and T5 dendrites grow in the lobula. One exciting possibility is that both neuropils share extrinsic cues conveying directional information to the dendrites of T4 and T5 neurons, which might also be used as universal guideposts by other neuronal cell types that must develop oriented dendrites (Hoermann, 2020).
The dendrites of all T5 subtypes extend across the same number of neuropil columns to connect to the same set of presynaptic functionally distinct neurons signalling luminescence changes from neighbouring points in the visual space, but in a spatial order that is subtype specific. The same holds true for the dendrites of all T4 subtypes (Shinomiya, 2019). As a simplified example, T4a connects to Mi4 in column 1, Mi1 in column 2 and Mi9 in column 3, whereas T4b connects to Mi9 in column 1, Mi1 in column 2 and Mi4 in column 3. What could the minimal set of developmental instructions look like to ensure such a specific wiring? Interestingly, the dendrites of the four T4 and T5 subtypes all show a clear and distinct orientation with respect to the extrinsic coordinates of the neuropil that they occupy. The dendrites' intrinsic coordinates define three compartments: proximal, medial and distal. With respect to these intrinsic coordinates, the wiring of all T4 and T5 subtypes is identical. In the above example, both T4a and T4b connect to Mi4 on the proximal, to Mi1 on the medial and to Mi9 on the distal part of their dendrite. Thus, once the compartmentalization of synapses from different inputs along their dendrites is controlled by cell-intrinsic mechanisms, the decisive point that differentiates between the subtypes is how they distinctly orient their dendrite. By growing their dendrites along different extrinsically defined directions, they could all apply the same genetic programmes to connect to a set of input neurons. This would lead to a spatial arrangement of synaptic inputs that is different for each subtype with respect to the extrinsic coordinates of the neuropil, thus supporting the detection of motion across four different directions but identical within the intrinsic coordinates of the neurons' dendrite (Hoermann, 2020).
It is envisaged that the manipulation of the genetic programmes controlling dendrite orientation in T4/T5 neurons will allow these ideas to be addressed systematically. Studying how the four T4/T5 neuron subtypes acquire their morphologies provides a great opportunity to link development, anatomy and function in a neuronal type that performs a computation that is conserved across visual systems, which might uncover universal blueprints of neural wiring (Hoermann, 2020).
The layered compartmentalization of synaptic connections, a common feature of nervous systems, underlies proper connectivity between neurons and enables parallel processing of neural information. However, the stepwise development of layered neuronal connections is not well understood. The medulla neuropil of the Drosophila visual system, which comprises 10 discrete layers (M1 to M10), where neural computations underlying distinct visual features are processed, serves as a model system for understanding layered synaptic connectivity. The first step in establishing layer-specific connectivity in the outer medulla (M1 to M6) is the innervation by lamina (L) neurons of one of two broad, primordial domains that will subsequently expand and transform into discrete layers. Previous work found that the transcription factor dFezf (Earmuff) cell-autonomously directs L3 lamina neurons to their proper primordial broad domain before they form synapses within the developing M3 layer. This study shows that dFezf controls L3 broad domain selection through temporally precise transcriptional repression of the transcription factor slp1 (sloppy paired 1). In wild-type L3 neurons, slp1 is transiently expressed at a low level during broad domain selection. When dFezf is deleted, slp1 expression is up-regulated, and ablation of slp1 fully rescues the defect of broad domain selection in dFezf-null L3 neurons. Although the early, transient expression of slp1 is expendable for broad domain selection, it is surprisingly necessary for the subsequent L3 innervation of the M3 layer. DFezf thus functions as a transcriptional repressor to coordinate the temporal dynamics of a transcriptional cascade that orchestrates sequential steps of layer-specific synapse formation (Santiago, 2021).
For most model organisms in neuroscience, research into visual processing in the brain is difficult because of a lack of high-resolution maps that capture complex neuronal circuitry. The microinsect Megaphragma viggianii, because of its small size and non-trivial behavior, provides a unique opportunity for tractable whole-organism connectomics. This study imaged the Drosophila whole head using serial electron microscopy. Its compound eye was reconstructed, and the optical properties of the ommatidia as well as the connectome of the first visual neuropil-the lamina was analyze. Compared with the fruit fly and the honeybee, Megaphragma visual system is highly simplified: it has 29 ommatidia per eye and 6 lamina neuron types. Features are reported that are both stereotypical among most ommatidia and specialized to some. By identifying the "barebones" circuits critical for flying insects, these results will facilitate constructing computational models of visual processing in insects (Chua, 2023).
Advances in brain connectomics have demonstrated the extraordinary complexity of neural circuits. Developing neurons encounter the axons and dendrites of many different neuron types and form synapses with only a subset of them. During circuit assembly, neurons express cell-type-specific repertoires comprising many cell adhesion molecules (CAMs) that can mediate interactions between developing neurites. Many CAM families have been shown to contribute to brain wiring in different ways. It has been challenging, however, to identify receptor-ligand pairs directly matching neurons with their synaptic targets. This study integrated the synapse-level connectome of the neural circuit with the developmental expression patterns and binding specificities of CAMs on pre- and postsynaptic neurons in the Drosophila visual system. To overcome the complexity of neural circuits, focus was placed on pairs of genetically related neurons that make differential wiring choices. In the motion detection circuit, closely related subtypes of T4/T5 neurons choose between alternative synaptic targets in adjacent layers of neuropil. This choice correlates with the matching expression in synaptic partners of different receptor-ligand pairs of the Beat and Side families of CAMs. Genetic analysis demonstrated that presynaptic Side-II and postsynaptic Beat-VI restrict synaptic partners to the same layer. Removal of this receptor-ligand pair disrupts layers and leads to inappropriate targeting of presynaptic sites and postsynaptic dendrites. It is proposde that different Side/Beat receptor-ligand pairs collaborate with other recognition molecules to determine wiring specificities in the fly brain. Combining transcriptomes, connectomes, and protein interactome maps allow unbiased identification of determinants of brain wiring (Yoo, 2023).
Dysregulation of HDAC4 expression and/or nucleocytoplasmic shuttling results in impaired neuronal morphogenesis and long-term memory in Drosophila melanogaster. A recent genetic screen for genes that interact in the same molecular pathway as HDAC4 identified the cytoskeletal adapter Ankyrin2 (Ank2). This study sought to investigate the role of Ank2 in neuronal morphogenesis, learning and memory. Ank2 is expressed widely throughout the Drosophila brain where it localizes predominantly to axon tracts. Pan-neuronal knockdown of Ank2 in the mushroom body, a region critical for memory formation, resulted in defects in axon morphogenesis. Similarly, reduction of Ank2 in lobular plate tangential neurons of the optic lobe disrupted dendritic branching and arborization. Conditional knockdown of Ank2 in the mushroom body of adult Drosophila significantly impaired long-term memory (LTM) of courtship suppression, and its expression was essential in the γ neurons of the mushroom body for normal LTM. In summary, this study provides the first characterization of the expression pattern of Ank2 in the adult Drosophila brain and demonstrates that Ank2 is critical for morphogenesis of the mushroom body and for the molecular processes required in the adult brain for the formation of long-term memories (Schwartz, 2023).
Animals navigating in natural environments must handle vast changes in their sensory input. Visual systems, for example, handle changes in luminance at many timescales, from slow changes across the day to rapid changes during active behavior. To maintain luminance-invariant perception, visual systems must adapt their sensitivity to changing luminance at different timescales. This study demonstrates that luminance gain control in photoreceptors alone is insufficient to explain luminance invariance at both fast and slow timescales and reveal the algorithms that adjust gain past photoreceptors in the fly eye. Imaging and behavioral experiments were combined with computational modeling to show that downstream of photoreceptors, circuitry taking input from the single luminance-sensitive neuron type L3 implements gain control at fast and slow timescales. This computation is bidirectional in that it prevents the underestimation of contrasts in low luminance and overestimation in high luminance. An algorithmic model disentangles these multifaceted contributions and shows that the bidirectional gain control occurs at both timescales. The model implements a nonlinear interaction of luminance and contrast to achieve gain correction at fast timescales and a dark-sensitive channel to improve the detection of dim stimuli at slow timescales. Together, this work demonstrates how a single neuronal channel performs diverse computations to implement gain control at multiple timescales that are together important for navigation in natural environments (Ketkar, 2023).
Sensitivity to numbers is a crucial cognitive ability. The lack of experimental models amenable to systematic genetic and neural manipulation has precluded discovering neural circuits required for numerical cognition. This study demonstrates that Drosophila flies spontaneously prefer sets containing larger numbers of objects. This preference is determined by the ratio between the two numerical quantities tested, a characteristic signature of numerical cognition across species. Individual flies maintained their numerical choice over consecutive days. Using a numerical visual conditioning paradigm, it was found that flies are capable of associating sucrose with numerical quantities and can be trained to reverse their spontaneous preference for large quantities. Finally, this study showed that silencing lobula columnar neurons (LC11) reduces the preference for more objects, thus identifying a neuronal substrate for numerical cognition in invertebrates. This discovery paves the way for the systematic analysis of the behavioral and neural mechanisms underlying the evolutionary conserved sensitivity to numerosity (Bengochea, 2023).
Visual systems extract multiple features from a scene using parallel neural circuits. Ultimately, the separate neural signals must come together to coherently influence action. This study characterized a circuit in Drosophila that integrates multiple visual features related to imminent threats to drive evasive locomotor turns. Using genetic perturbation methods, a pair of visual projection neurons (LPLC2) and descending neurons (DNp06) were identified that underlie evasive flight turns in response to laterally moving or approaching visual objects. Using two-photon calcium imaging or whole-cell patch clamping, these cells were shown to indeed respond to both translating and approaching visual patterns. Furthermore, by measuring visual responses of LPLC2 neurons after genetically silencing presynaptic motion-sensing neurons, it was shown that their visual properties emerge by integrating multiple visual features across two early visual structures: the lobula and the lobula plate. This study highlights a clear example of how distinct visual signals converge on a single class of visual neurons and then activate premotor neurons to drive action, revealing a concise visuomotor pathway for evasive flight maneuvers in Drosophila (Kim, 2023).
The development of neuronal connectivity requires stabilization of dynamic axonal branches at sites of synapse formation. Models that explain how axonal branching is coupled to synaptogenesis postulate molecular regulators acting in a spatiotemporally restricted fashion to ensure branching toward future synaptic partners while also stabilizing the emerging synaptic contacts between such partners. This study investigated this question using neuronal circuit development in the Drosophila brain as a model system. Epidermal growth factor receptor (EGFR) activity is required in presynaptic axonal branches during two distinct temporal intervals to regulate circuit wiring in the developing Drosophila visual system. EGFR is required early to regulate primary axonal branching. EGFR activity is then independently required at a later stage to prevent degradation of the synaptic active zone protein Bruchpilot (Brp). Inactivation of EGFR results in a local increase of autophagy in presynaptic branches and the translocation of active zone proteins into autophagic vesicles. The protection of synaptic material during this later interval of wiring ensures the stabilization of terminal branches, circuit connectivity, and appropriate visual behavior. Phenotypes of EGFR inactivation can be rescued by increasing Brp levels or downregulating autophagy. In summary, this study identified a temporally restricted molecular mechanism required for coupling axonal branching and synaptic stabilization that contributes to the emergence of neuronal wiring specificity (Dutta, 2023).
The complexity of neuronal circuit wiring patterns, driven to a significant extent by the degree of axonal branching during development, is thought to be key to the emergence of behavior. In the mammalian motor cortex for example, axon branch complexity can allow a single neuron to innervate very distant ipsi- and contra-lateral cortical and sub-cortical targets. The unique branching pattern of L-fibers supports the nocturnal lifestyle of the sweat bee M. genalis, while intrinsic variation in medulla dorsal cluster neurons
(M-DCN) connectivity underlies individual variation in Drosophila visual response behavior. Understanding the spatial, temporal, and mechanistic coordination of axonal branching and synaptogenesis during development is critical for a more complete description of the emergence of both conserved patterns and individual variation of neuronal circuit diagrams (Dutta, 2023).
This study asked if, when, and how molecular factors that play key roles in regulating axon branching, synapse formation, and membrane degradation interact to produce specific presynaptic patterns, neuronal circuit connectivity and behavior using the developing Drosophila visual system as a model. This diverse list of cellular processes is tied together by their collaborative effects during the development of synaptic connectivity. Axo-dendritic branching and synapses formation have long been known to depend on each other during synaptotropic growth of branches based on synapse stabilization. For example, branch stabilization during synaptotropic growth has also been shown to depend on synaptic activity and locally restricted calcium signals in developing Drosophila dendrites. Developmental interactions of axonal and dendritic processes are a major contributor, and can sometimes predict, the adult synaptic connectivity. By contrast, cell biological processes like cytoskeletal dynamics or membrane degradation have long been described as a "permissive" basis for more "instructive" molecular mechanisms of synaptic specification. However, to the extent that such cell biological mechanisms directly contribute to branching and synapse formation, they become parts of a composite instruction that cannot be pinned on a single molecular mechanism but require the consideration of several collaborating factors to understand a neuron’s choice to branch and form a synapse. Hence, an integrative analysis of molecular recognition, signaling, and cell biological machinery is necessary to mechanistically understand how branching contributes to adult synaptic connectivity (Dutta, 2023).
Numerous molecular and cellular mechanisms have been implicated in the spatiotemporal control of how synaptic partners are brought together for synapse formation. Over the last 25 years investigating the role of EGFR in nervous systems has demonstrated its roles in neural stem cell maintenance, astrocyte and oligodendrocyte maturation, axon regeneration, and more recently in neurite outgrowth and branching. In addition, roles have been demonstrated for the cell biological regulation of filopodial dynamics and autophagic degradation in establishing specific synaptic connectivity. How such basic cellular processes are coupled to the process of patterning axonal branches and how the three processes—branching, synaptogenesis, and autophagy—are coordinated in time to establish circuit-specific wiring diagrams has remained unclear. These processes are indeed coupled but only during a very specific temporal interval through EGFR activity preventing the autophagic degradation of the synaptic AZ protein Brp. Inactivation of EGFR during this specific critical interval in late development, but not before or after, causes Brp degradation, circuit wiring changes, and altered visually driven behaviors (Dutta, 2023).
An interesting observation is the difference in dynamics upon EGFR inactivation and knockdown of Brp. While both result in increased secondary branches, loss of Brp leads to a more dynamic state of these branches even in the adult brain. In contrast, suppression of EGFR function causes these secondary branches to be largely static, even though EGFR inactivation also causes Brp loss. This is likely because suppression of EGFR function also impairs actin dynamics which are required for filopodial dynamics, while Brp loss does not affect EGFR activity and thus does not impair actin dynamics. These observations underline the critical importance of live imaging for discovering and interpreting otherwise seemingly similar phenotypes. These observations are also consistent with the genetic hierarchy observed whereby EGFR regulates Fluorescently tagged BrpD3 (or Brpshort) levels, but Brp does not appear to regulate EGFR levels. Again, live imaging combined with temporally restricted inactivation of EGFR allowed dissection of the temporal logic of this genetic hierarchy and discover the role of EGFR activity as coupling mechanism between branching dynamics, local degradation, and synapse formation (Dutta, 2023).
Observations show that the coupling between branching, autophagy, and synaptogenesis is only required during a very specific temporal interval which, perhaps not surprisingly, coincides with active synaptogenesis in the developing fly brain. Analysis of the temporal sequence of trafficking of various molecules indicates that this developmental critical interval is opened by the spatial and temporal coincidence of EGFR and BrpD3 in exploratory branches. Similarly, it is likely that the critical interval ends once synaptic contacts between DCNs and their postsynaptic cells have been established. What remains to be explored is how exactly synapse formation ends the interactive feedback between branch dynamics and synapse degradation. One possibility is that postsynaptic dendrites provide molecular signals to presynaptic axonal branches to limit AZ degradation. Another possibility is that the initiation of spontaneous activity alters local endolysosomal recycling to reduce degradation and favor maintenance, for example, through the activity-dependent regulation of local translation of resident mRNAs. A third possibility is the late arrival of presynaptic proteins that protect the AZ, reduce autophagy, or alter EGFR function. Clearly, none of these mechanisms are mutually exclusive, and it is speculated that a combination of such feedback mechanisms would be the best way to ensure a robust closing of the developmental critical interval discovered in this study. Flies, similar to vertebrates, undergo a critical period of post-natal experience-dependent synaptic pruning with unknown molecular mechanism. It would be interesting to explore whether the mechanism uncovered in this study also plays a role in synaptic regulation and neural circuit plasticity during that critical period. It is possible that similar molecular modules are used re-iteratively to regulate synaptic homeostasis and presynaptic branching in experience-independent and -dependent brain development (Dutta, 2023).
Previous work has shown that an increase in local autophagy also led to a decrease of adult synapses in R7 photoreceptor neurons, albeit by a different mechanism. In R7 photoreceptors, local autophagosome formation at the tips of synaptogenic filopodia is accompanied by engulfment of synaptic seeding factors but not Brp, followed by filopodial collapse and thus less filopodial availability to form synapses. In DCNs, axonally localized autophagosome formation is accompanied by engulfment of Brp but not synaptic seeding factors, leading to a destabilization and reduction of mature synapses. Hence, the spatiotemporal-specific roles of autophagy in different types of axon terminals during synapse formation are highly context specific, involve different substrates, and yet lead to similar outcomes. An obvious contextual difference between R7s and DCNs is that only the latter form branched axons and employ synaptotropic-like branch stabilization through mature synapses (Dutta, 2023).
Increasing evidence supports the important roles that stochastic and/or noisy molecular processes play in the early phases of establishing wiring specificity prior to the final step of spatially and temporally restricted local synapse formation and stabilization. While neuronal circuit wiring patterns across the animal kingdom are highly robust, they, almost without exception, show at least some degree of intrinsic variation within and between individuals. Noise in genetically encoded program can ensure robustness of highly reproducible wiring patterns, for example, when stochastic exploration ensures partner finding. Correspondingly, axon branch initiation in DCNs is noisy and exploratory, while the final axonal pattern is robustly stereotypic. The evidence presented in this work demonstrates the importance of intrinsic local control of the feedback between axonal branching and synapse formation to ensure the robust outcome. Such local feedback regulation ensures that genetically encoded noisy molecular and cellular processes such as filopodial growth and retraction and synaptic seeding are coordinated in time and space to produce conserved, robust, yet individually variable, non-random neuronal circuit diagrams (Dutta, 2023).
This study described a strategy to study single synapses between identified cell types by combining genetic tools, EM-based connectomics, and protein localization through expansion microscopy. To devise probes for specific synapses, tools were developed to tag and map the distribution of endogenous NR subunits of receptors for the major neurotransmitters in Drosophila in single neurons and at single synapses between neurons. These studies highlighted the complexity of different NR subunit distributions in dendrites, the developmental dynamics of NR accumulation at different synapses, and their association with other proteins at subsets of synapses. These tagged receptors provide a resource for studies focusing on subcellular localization of neuronal proteins, the assembly of synapses and synaptic plasticity. The unique features of wiring specificity of T4 and T5 dendrites suggest that in some developmental contexts targeting of NRs to discrete dendritic domains provides a spatial map of molecular determinants controlling synaptic specificity (Sanfilippo, 2023).
The approach to conditionally tag receptors through modification of genomic loci preserves their endogenous mRNA expression levels, patterns, and temporal dynamics. Epitope tags were inserted into poorly conserved and unstructured regions of variable length within a large cytoplasmic loop found in all cys-loop receptors. The distribution of the tagged NRs accurately reflects the localization of endogenous receptors. Tagged NRs selectively localized at sites adjacent to presynaptic partners (i.e., Brp and Cac) when visualized by either ExLLSM and ExLSM, and the distribution of receptors matched the neurotransmitter specificity of the neurons providing synaptic inputs. The increasing availability of genomes for comparative sequence analysis and the development of structure prediction tools such as AlphaFold facilitate the identification of sites to successfully tag other classes of NRs, as well as other synaptic proteins. The extensive synaptic connectivity maps in flies, reagents for manipulating specific cell types, and the array of endogenously tagged NR subunits reported in this study provides an opportunity to characterize the distribution of these receptors in many different circuits regulating a broad range of brain functions and behaviors (Sanfilippo, 2023).
The distribution of seven different tagged NR subunits for glutamate, GABA, and acetylcholine in the cys-loop superfamily in direction-sensitive T4 neurons was striking. The stereotyped patterns of each class of receptor matched the pattern of the neurotransmitters used by their respective presynaptic inputs. Unexpectedly, different NRs to the same neurotransmitter also localized to different domains receiving input from different presynaptic partners. This was seen for both cholinergic (excitatory) and GABAergic (inhibitory) synapses. The differential distribution of NR subunits to different spatial domains may contribute to the unique computational features of T4 dendrites. How these domains are established in neurons with diverse and often complex morphologies is not known. Perhaps the segregation of proteins to different domains may at a mechanistic level share features in common with the establishment of cell polarity domains in other cell types including apical basal polarity in epithelial cells (Sanfilippo, 2023).
Select cys-loop GABA receptors that differ in NR subunit composition are found in distinct domains in pyramidal cells of the mouse cortex. It is likely that the complexity of receptor distributions in mammals also extends to different domains within the same dendrites as is described in this study. In the mammalian brain there is a great diversity of cys-loop family of GABA receptors (i.e., GABAAR), with 19 distinct genes encoding GABAAR subunits. Thus, tagging approaches in the mouse similar to what is reported in this study in the fly may provide a way to uncover the spatial distributions of NR subunits and combinations of them at different synapses. As these receptors have different physiological properties, their spatial distributions may contribute to understanding information processing in dendrites of mammalian neurons (Sanfilippo, 2023).
Analysis of T4 neurons raised the intriguing possibility that in some developmental contexts neurotransmitter receptors themselves may serve as recognition molecules specifying the pattern of pre-synaptic inputs. In these dendrites, each presynaptic neuron type forms synapses within restricted domains along the proximodistal axis and these patterns correspond to the distribution of different NR subunits. Mi9 neurons, for instance, only form synapses within the distal domains of T4 dendrites while closely related Mi1 and Mi4 neurons form synapses in middle and proximal regions, respectively. This specificity is particularly striking as the dendritic arbors of many T4s overlap extensively such that each input axon terminal contacts the entire range of dendritic domains and yet makes synapses only within restricted spatial domains. A simple model to account for this specificity is that targeting of NR subunits to specific dendritic domains provides a spatial map of molecular signposts recognized by cell surface recognition proteins selectively expressed on the surface on different presynaptic terminal arbors. Alternatively, specific molecular determinants may be arranged along the proximodistal axis in discrete dendritic domains, and these may recruit both synaptic inputs and subsets of NR subunits to these sites. In either case, these studies suggest a close relationship between the targeting of NR subunits to different spatial domains and the specificity of synaptic inputs (Sanfilippo, 2023).
This model is consistent with studies in mammals. NRs have been shown to associate with other postsynaptic proteins that promote adhesion between pre, and postsynaptic membranes and these complexes may be selectively localized. This study, for instance, reports the identification of a transmembrane protein, Mmd, in the distal domains of T4 dendrites in close association with GluClα receptor. A mammalian homolog of Mmd, Adam22, also co-localizes to synaptic glutamate receptors, albeit of a different class, and promotes adhesion between presynaptic and postsynaptic membranes. Alternatively, there is evidence that the extracellular domains of NR subunits can directly interact with proteins on the presynaptic membrane. For instance, the extracellular domain of the iGluR subunit GluA1 can interact with presynaptic neuronal pentraxin receptors, and this interaction can support the formation of synapses in a heterologous system. Similarly, the N terminal of the α1 GABAAR subunit has been shown to interact with neurexin-2β and modulate GABAAR function. Together these observations raise the notion that, in some cases, the NR subunits themselves are intimately involved in matching of pre and postsynaptic membranes (Sanfilippo, 2023).
The complexity of neural circuit structure has become increasingly clear with the completion of dense EM connectomes (MICrONS Consortium, 2023; Scheffer et al., 2020; Shapson-Coe et al., 2021; Shinomiya et al., 2019; Takemura et al., 2013). Extensive studies have argued that different domains along the proximo-distal axis of T4 dendrites play a crucial role in direction-specific motion processing (Groschner et al., 2022; Gruntman et al., 2018; Strother et al., 2017). The identification of different NRs and combinations of them within different domains provides an opportunity to understand computations at the molecular level (Sanfilippo, 2023).
How these specific molecular domains in dendrites form and their patterns of synaptic inputs emerge during development remains enigmatic. Localization of NR subunits to different domains at early stages of dendrite development raises the possibility that this molecular diversity contributes to determining the spatial distribution of specific synapses in dendrites. The use of tagged NRs and cell type specific manipulation with EM-based connectomics, ExLSM, genetics and biochemical methods provides a way of understanding how NRs become localized to specific domains and linking these to the specificity of synaptic inputs (Sanfilippo, 2023).
Cowley, B. R., Calhoun, A. J., Rangarajan, N., Ireland, E., Turner, M. H., Pillow, J. W., Murthy, M. (2024). Mapping model units to visual neurons reveals population code for social behaviour. Nature, 629(8014):1100-1108 PubMed ID: 38778103
The rich variety of behaviours observed in animals arises through the interplay between sensory processing and motor control. To understand these sensorimotor transformations, it is useful to build models that predict not only neural responses to sensory input but also how each neuron causally contributes to behaviour. This study demonstrates a novel modelling approach to identify a one-to-one mapping between internal units in a deep neural network and real neurons by predicting the behavioural changes that arise from systematic perturbations of more than a dozen neuronal cell types. A key ingredient that is introduced is 'knockout training', which involves perturbing the network during training to match the perturbations of the real neurons during behavioural experiments. This approach was applied to model the sensorimotor transformations of Drosophila melanogaster males during a complex, visually guided social behaviour. The visual projection neurons at the interface between the optic lobe and central brain form a set of discrete channels, and prior work indicates that each channel encodes a specific visual feature to drive a particular behaviour. This model reaches a different conclusion: combinations of visual projection neurons, including those involved in non-social behaviours, drive male interactions with the female, forming a rich population code for behaviour. Overall, this framework consolidates behavioural effects elicited from various neural perturbations into a single, unified model, providing a map from stimulus to neuronal cell type to behaviour, and enabling future incorporation of wiring diagrams of the brain into the model (Cowley, 2024).
This study develop knockout training, a novel solution to identify a one-to-one mapping between internal units in a deep neural network (DNN) and real neurons in the brain of a fly. The model makes predictions about how neurons respond to sensory stimuli and drive behaviour. Although silencing each LC neuron type (LC neuron types receive input from the lobula and lobula plate in the optic lobe and send axons to optic glomeruli in the central brain) )on its own may have a small to medium effect on behaviour, the 1-to-1 network infers how the LC types work together as a population to drive the courtship behaviour of the male. The model extends beyond findings from direct recordings of LC neuron, even in behaving flies. The 1-to-1 network provides information on LC visual responses in freely behaving flies (not head-fixed, as is required for recordings) engaging in natural social interactions and can generate LC responses to any arbitrary visual stimulus. In fact, it was demonstrate that the 1-to-1 network predicts actual responses to stimuli that the model had not seen during training. The model also makes testable predictions about which combinations of LC types are both necessary and sufficient for specific courtship behaviours. A major new finding of this work is which and to what extent LC neuron types contribute to song production, an integral part of courtship guided by visual feedback. Given that the same visual stimulus sequence can drive multiple LC types, this neuron-to-behaviour relationship is not readily inferred from LC recordings alone. The 1-to-1 network is the first large-scale hypothesis of how the LC types work together to encode stimuli and contribute to behaviour; this model and code is shared with the community to inspire future experiments and models (Cowley, 2024).
The main conclusion of this study is that the complex courtship behaviour of the male relies on combinations of visual projections neurons-including those also involved in non-social behaviours. However, the extent to which other behaviours beyond those observed during courtship also rely on a population code is not yet known. Knockout training on the LC types could easily be applied to other visuomotor behaviours (for example, escape responses or flight) to make direct comparisons. Given the extent of interconnectivity between LC types and convergence of LC types onto common downstream cell types, it is posited that population coding for behaviour, particularly in natural contexts, might be the norm. By contrast, for behaviours that rely on quick and robust processing, such as escape from a predator, the arrangement of LC types into optic glomeruli may facilitate the fast readout of specific channels. One issue raised by the use of a multiplexed code is how the fly brain produces the correct behaviour at the correct time. For example, LPLC2 neurons synapse onto the giant fibre neuron to drive an escape take-off, but the 1-to-1 network predicts that this same cell type encodes female size and contributes to the forward velocity of the male during courtship; recent work has also found LPLC2 contributes to evasive flight turns. Future experiments are needed to understand how the same LC cell type can contribute to different behaviours in different contexts (Cowley, 2024).
This modelling approach comes with limitations. For example, if silencing an LC type does not lead to a noticeable change in behaviour, the 1-to-1 network cannot infer the tuning of that LC type. In addition, many silenced LC types resulted in stronger-not weaker-courtship, suggesting that these LC neurons may act partially as distractors to prevent relentless pursuit of the female. This approach also found some mismatches between real LC responses and the responses of the 1-to-1 network; although this may be owing to differences in internal state between freely moving males during natural courtship (training data for the model) versus head-fixed males passively viewing stimuli (neural recordings), training on neural data and behavioural data together may help to improve both neural and behavioural prediction. An experimental limitation of using natural behaviour arises because the statistics of the visual experience cannot be matched between LC-silenced and control males (for example, an LC9-silenced male spends much less time near the female); future experiments can use virtual reality or robotic females to present identical stimulus sequences to control and silenced males (Cowley, 2024).
Following recent studies using Deep Neural Networks (DNNs) to predict responses of visual neurons, this study used DNNs in the 1-to-1 network that are highly expressive function approximators but lack biological realism. The model-agnostic knockout training procedure can be used to train more biologically inspired models that incorporate constraints from the FlyWire connectome and emerging male brain wiring diagrams to include recurrent connections, lateral connections between LC types and delays. An intriguing future direction is to apply this framework to other bottlenecks within the Drosophila brain, such as the descending and ascending neurons that link the brain and nerve cord. and in more complex systems for which genetic control over cell types is available. This work shows that constraining models with causal perturbations of neurons during complex behaviour is an important ingredient in revealing the relationships between stimulus, neurons and behaviour (Cowley, 2024).
Dm9 neurons (for information on Dm9 neurons, go to Google AI
and query Dm9 neurons Drosophila) in the Drosophila optic lobe have been proposed as functional homologs of horizontal cells in the outer retina of vertebrates. This study combined genetic dissection of neuronal circuit function, two-photon calcium imaging in Dm9 and inner photoreceptors, and immunohistochemical analysis to reveal novel insights into the functional role of Dm9 in early visual processing. These experiments show that Dm9 receive input from all four types of inner photoreceptors R7p, R7y, R8p, and R8y. Histamine released from all types R7/R8 directly inhibits Dm9 via the histamine receptor Ort, and outweighs simultaneous histamine-independent excitation of Dm9 by UV-sensitive R7. Dm9 in turn provides inhibitory feedback to all R7/R8, which is sufficient for color-opponent processing in R7 but not R8. Color opponent processing in R8 requires additional synaptic inhibition by R7 of the same ommatidium via axo-axonal synapses and the second Drosophila histamine receptor HisCl1. Notably, optogenetic inhibition of Dm9 prohibits color opponent processing in all types of R7/R8 and decreases intracellular calcium in photoreceptor terminals. The latter likely results from reduced release of excitatory glutamate from Dm9 and shifts overall photoreceptor sensitivity toward higher light intensities. In summary, these results underscore a key role of Dm9 in color opponent processing in Drosophila and suggest a second role of Dm9 in regulating light adaptation in inner photoreceptors. These novel findings on Dm9 are indeed reminiscent of the versatile functions of horizontal cells in the vertebrate retina (Schnaitmann, 2024).
Color vision facilitates the identification of food sources, conspecifics, predators, and prey and supports a wide range of behaviors. It requires at least two photoreceptor classes with different spectral sensitivities and neural circuits that antagonistically compare their signals. Neurons that perform such comparisons are excited by certain types of photoreceptors and inhibited by others, resulting in color opponent responses that are a hallmark of color vision (Schnaitmann, 2024).
Recent research demonstrates that color opponent processing emerges already in the presynaptic terminals of photoreceptors in the visual system of different vertebrates and insects. Thus, evolutionary highly distant animals like monkeys and flies might employ similar circuit mechanisms to accomplish the first stage in color opponent processing (Schnaitmann, 2024).
In the retina of vertebrates, color opponent responses in photoreceptor terminals are mediated by spectral feedback inhibition from horizontal cells (HCs). HCs are inhibitory interneurons that laterally extend their processes into the outer plexiform layer to sample glutamatergic photoreceptor input. Light decreases glutamate release from vertebrate photoreceptors, reduces ionotropic glutamate receptor activation and thereby hyperpolarizes HCs. In turn, HCs then modulate glutamate release from their presynaptic input elements by parallel inhibitory feedback mechanisms and are thought to provide feedforward inhibition to bipolar cells dendrites. By their feedback, HCs mediate surround inhibition of the characteristic center-surround receptive fields of photoreceptor terminals and ON- and OFF-bipolar cells, that enhance spatial and chromatic contrast. In addition, HCs are important players in "global" adaptation by reversible electric coupling. They form low-resistance electric networks and the extent of coupling is controlled by ambient light and circadian rhythmxxtxxx. Finally, HCs have also been implicated in locally restricted feedback inhibition to individual photoreceptor terminals (Chapot et al., 2017a). Because of their many functions in spectral processing and adaptation, HCs are considered multi-purpose interneurons of the outer vertebrate retina (Schnaitmann, 2024).
In the fruit fly Drosophila, each of the ~780 ommatidia of the compound eye harbors six outer photoreceptors R1-R6 and a genetically determined tandem of pale (p) or yellow (y) inner photoreceptors R7/R8 (Figure 1A). Expression of rhodopsin 1 (rh1) and a UV-sensitizing photopigment make R1-R6 sensitive to a broad spectral range, from UV to green (Figure 1B; colors named according to human perception). R1-R6 transmit luminance information to the lamina, and enable downstream circuitries to detect visual motion and further achromatic visual features . However, R1-R6 additionally contribute to spectral processing and color vision (Schnaitmann, 2024).
R7/R8 project to the distal layers M1-M6 of the medulla where they convey the major input to the color vision circuitry. Pale and Yellow photoreceptors (p and y) R7/R8 are stochastically distributed over the main part of the eye. p R7/R8 express rh3/rh5 with maximum sensitivities in the short-UV/blue spectral range. y R7/R8 express rh4/rh6 with maximum sensitivity in the long-UV/green. Two recent studies found that color opponent processing in R7/R8 terminals relies on a concerted action of two mechanisms. Mutual synaptic inhibition between R7 and R8 of the same ommatidium (same medulla column), and simultaneous feedback inhibition from postsynaptic neurons in the medulla, in particular multi-columnar Dm9 neurons that are post- and presynaptic to R7 and R8. R7 and R8 terminals of an individual medulla column mutually inhibit each other via release of the neurotransmitter histamine and activation of postsynaptic HisCl1 receptors (R7/R8 axo-axonal inhibition) (Schnaitmann et al., 2018). To this point, p and y R7/R8 tandems can be considered spectral analyzers that preserve the spatial resolution of the eye. This is because the light-sensitive rhabdomeres of R7 and R8 are stacked on top of each other, both photoreceptors share the same optical axis, and they simultaneously sample the same spot in visual space. Spectral feedback inhibition that enables additional spectral comparisons that exceed pure p- and y-antagonism was suggested for all R7/R8 photoreceptor types, mediated by Dm9 cells (Heath et al., 2020). Dm9 ramify their neurites in the distal layers M1-M6 of ~7 neighboring columns of the medulla, where they closely intermingle with R7/R8 axons (Figures 1D-F). Electron microscopic reconstruction revealed that Dm9 are strongly connected to R7 and R8 being post- and presynaptic, and that Dm9 are postsynaptic to L3 lamina monopolar cells that relay luminance information from R1-R6 (Takemura et al., 2015; Ketkar et al., 2020; Kind et al., 2021). RNA-seq on Dm9 suggests that they use Ort-type histamine-receptors to detect histamine released by R7/R8 (Davis et al., 2020). Most important, calcium imaging experiments with genetic block of activity in Dm9 (cell-specific expression of Kir) or cell specific rescue of Ort receptor expression in Dm9 in histamine receptor mutant flies showed that Dm9 are required and sufficient for color opponent processing in R8p, respectively (Heath et al., 2020). Based on these findings on R8p and a computational model, it was proposed that Dm9 mediate similar spectral feedback inhibition in R8y, R7p, and R7y, and that Dm9 add an inhibitory surround to the ON-center responses of all R7/R8 (Heath et al., 2020).
This study employed a combined genetic and physiological approach to interfere with the functional properties of selected neurons in the early visual system of Drosophila, and to record calcium dynamics in same or different neuron types (Dm9 or terminals of inner photoreceptors R7/R8). Calcium recording is performed during visual stimulation of experimental animals using fly-back stimulation, targeted expression of the genetically encoded ratiometric calcium indicator Twitch-2C, and in vivo two-photon laser scanning microscopy. These experiments were complemented with the precise subcellular detection of key-proteins of synaptic function in genome-edited flies (Schnaitmann, 2024).
Activation of all four inner photoreceptor types elicits strong ort-dependent inhibition in Dm9, whereas activation of R1-R6 does not elicit detectable activity changes. This inhibition outweighs additional histamine-independent excitation of Dm9 by UV-sensitive R7 photoreceptors. Calcium imaging demonstrates that Dm9 indeed contribute to spectral feedback inhibition in all four inner photoreceptor types. However, only in R7 terminals feedback from Dm9 is sufficient to generate color opponent responses. In terminals of R8, feedback inhibition from Dm9 must coincide with axo-axonal inhibition from the intra-ommatidial partner R7 to generate color opponent responses. Optogenetic inhibition of Dm9 that mimics Dm9's responses to light, was associated with strong inhibition of all R7/R8 photoreceptor types. Cell-specific conditional labeling of multi epitope-tagged vGluT suggests that Dm9 release glutamate. Dm9 therefore likely mediates feedback inhibition by reducing excitatory glutamatergic input to R7/R8 in response to light increments. By this functional loop, Dm9 counteracts light induced depolarization in R7/R8 terminals. Finally, it is reported that persistent optogenetic inhibition of Dm9 abolishes color opponent processing in R7/R8 and shifts their sensitivity to higher light intensities. It is proposed that Drosophila Dm9 cells have important roles in both, color opponent processing and adaptation in presynaptic photoreceptor terminals, reminiscent of the versatile functions of HCs in the outer vertebrate retina (Schnaitmann, 2024).
Inner photoreceptors R7/R8 transmit the major signals to the color vision circuitry in Drosophila and recent calcium imaging studies demonstrated that R7/R8 spectrally inhibit each other by two parallel circuit mechanisms. First, reciprocal inhibition occurs between R7 and R8 photoreceptors of the same ommatidium, mediated by the neurotransmitter histamine that triggers the activation of HisCl1 receptors. Second, neurons of the medulla detect R7/R8 input and provide feedback inhibition to R7/R8. Multicolumnar Dm9 cells that sample the input from several R7 and R8 were identified as the medulla neurons that convey feedback inhibition to R8p. Based on combinatorial norpA--rescue experiments, serial EM-analysis, and RNA-seq, this function of Dm9 was conferred to all four types of inner photoreceptors and pivotal for a computational model used to explain color opponent processing in the peripheral visual system of the fly (Schnaitmann, 2024).
This study combined in vivo two-photon calcium imaging with genetic circuit manipulation to further elucidate the role of Dm9 in early visual processing. The results demonstrate that all four types of inner photoreceptors R7/R8 convey visual input to Dm9. Dm9 cells exhibit reduced calcium responses indicative of inhibition to all tested spectral stimuli that scale with stimulus intensity. However, the calcium responses of Dm9 were more complex than expected. R7 and R8 provide strong inhibitory histaminergic input to Dm9 via Ort receptor activation. This histamine-/Ort-mediated inhibition outweighs additional, previously unrecognized excitatory input from R7 that does not involve the neurotransmitter histamine. The physiological role of excitatory input from UV-sensitive R7 remains to be analyzed (Schnaitmann, 2024).
The results demonstrate that Dm9 cells indeed have a key role in color opponent processing in all four R7/R8 photoreceptor types, as previously suggested. This conclusion is substantiated by different calcium imaging experiments. First, Ort-rescue in Dm9 demonstrates that histaminergic signaling from R7/R8 is sufficient to mediate spectral inhibition in both R7 types, whereas the same processing in both R8 types requires Dm9 in addition to simultaneous activation of HisCl1 receptors in R8 by direct inhibition from the partner R7. Second, optogenetic inhibition of Dm9 abolishes color opponent processing in all four types of R7/R8. Notably, optogenetic inhibition of Dm9 strongly decreases the calcium responses of inner photoreceptors R7/R8 to light and shifts their sensitivity to higher light intensities. Because optogenetic inhibition mimics the effect of light, it is proposed that Dm9 simultaneously contribute to the adaptation of inner R7/R8 to ambient light.
Such a dual role would make Dm9 even more "horizontal cell-like" than previously thought (see Proposed circuit diagram for the multiple interactions between R7, R8, and Dm9a) (Schnaitmann, 2024).
The observed histaminergic input from all R7/R8 photoreceptor types to Dm9 is in line with the reported strong synaptic connections between these cells that were revealed by serial EM-analysis. According to these studies, Dm9 are additionally postsynaptic to lamina L3 neurons that receive strong synaptic input from the outer photoreceptors R1-R6 . However, under the current experimental conditions, light activation of R1-R6 did not elicit any detectable calcium responses in Dm9, which is consistent with results of previous studies that did not find any contribution of R1-R6 to color opponent processing in R7/R8. Whether and how R1-R6 and L3 contribute to processing in Dm9 therefore remains unknown (Schnaitmann, 2024).
Activation of R7 by light elicited excitation in Dm9 in the absence of both histamine synthesis and histamine receptor expression. R8 cells have recently been implicated in (histamine-independent) excitatory cholinergic signaling to medulla neurons, in particular of the circadian clock circuitry in the medulla. Do R7 photoreceptors also use cholinergic signaling? Reports on the expression of choline acetyltransferase (ChAT) in R7 are controversial. Low amounts of mRNA encoding the vesicular acetylcholine transporter (VAChT) in R7 and mRNA encoding ACh-receptors in Dm9. Similarly, sign conserving electrical synapses or ephaptic signaling could be at work, as reported for horizontal cell feedback to vertebrate photoreceptors. Notably, ephaptic signaling has also been described between R1-R6 and lamina monopolar cells of the fly visual system (Schnaitmann, 2024).
Previous studies as well as new results demonstrate that color opponent processing in all inner photoreceptor types is mediated by two parallel circuit mechanisms. These mechanisms are not redundant. The HisCl1-dependent reciprocal inhibition between R7/R8 generates p- and y-specific opponent processing that retains the resolution of the compound eye. In contrast, Dm9 and Ort-dependent feedback inhibition enables additional inter-ommatidial processing in R8p and possibly all inner photoreceptor types. Based on its multicolumnar ramifications, Dm9 was furthermore proposed to mediate spatial surround-inhibition in inner photoreceptors. However, initial experimental analyses of the spatial layout of the receptive field of inner photoreceptors revealed simple ON receptive fields and no evidence for classical ON-center/OFF-surround receptive fields (Schnaitmann, 2024).
Interestingly, the two mechanisms operate with different strength in R7 and R8. ort expression in visual circuits and hisCl1 expression in R7, respectively, are each sufficient to mediate color opponent processing in R7 terminals. In contrast, color opponent processing in R8 requires simultaneous ort-mediated signaling to medulla circuits together with i>hisCl1-mediated inhibition from the partner R7. In line with these results and the experiments on R8p performed by Heath et al. (2020), This study reports that Dm9 is required for color opponent processing in both R8 photoreceptor types. In contrast to the current results, another study reported that ort expression in Dm9 is sufficient for color opponency in R8p and that additional HisCl1 is not required. This study investigated sufficiency and requirement of Dm9 for color opponent processing in R8p and in the remaining R7/R8 photoreceptor types. In agreement with previous results, rescue of ort expression in Dm9 in hisCl1- mutant flies is sufficient for color opponency in R7 but not in R8 that additionally require HisCl1 (Figure 3). Such functional differences might be caused by differences in the presented light stimuli, in particular intensity, spectral composition, background illumination and the adaptational state of experimental flies (Schnaitmann, 2024).
An apparent inconsistency was observed in the current results on the requirement of Dm9 in color opponent processing in R7. Previous work on ort- mutant flies shows that flies lacking Dm9-inhibition still show color opponent processing in R7 that is mediated via the HisCl1-pathway. In contrast, GTACR1 (light-gated anion channelrhodopsin) inhibition of Dm9 prohibited color opponent processing in R7 in flies with intact HisCl1 signaling . A significant difference between these two experimental approaches lies in the different activity of Dm9. In the ort- mutant background, Dm9 exhibits aberrant excitatory responses to visual stimuli. In contrast, optogenetic manipulation induces persistent inhibition. The latter decreases the overall responsiveness of R7 by a so far unknown mechanism and likely results in a physiological condition in which additional HisCl1-mediated inhibition fails (Schnaitmann, 2024)
By what mechanism do Dm9 cells mediate feedback inhibition to R7/R8 photoreceptors? Optogenetic excitation of Dm9 revealed excitatory input to R7. The current experiments with GTACR1 recapitulate photoreceptor-mediated inhibition in Dm9 and reveal a strong decrease in intracellular calcium in all four inner photoreceptor types under these conditions (Figure 5). This suggests that Dm9 do mediate feedback inhibition, most likely through a decrease in the release of glutamate. The latter is suggested by the presence of mRNA for vGluT in Dm9, and cell-specific conditional labeling of vGluT protein in Dm9 neurites (Figure 1K). Suitable ionotropic glutamate receptors including EKAR might be expressed in all photoreceptor types of Drosophila. In the lamina, EKAR mediates feedback from lamina intrinsic amacrine cells to R1-R6 photoreceptors which helps to maintain light sensitivity under low ambient light. Similarly, excitatory signaling from Dm9 to photoreceptors is required to retain sensitivity to low intensity stimuli (Schnaitmann, 2024)
The norpA and histamine receptor rescue approach is the gold standard that is used in the field to characterize the contribution of specific photoreceptor types (or postsynaptic cells) to visual processing in Drosophila. That these genetic manipulations elicit certain developmental alterations and compensatory mechanisms cannot be excluded. However, in contrast to the vertebrate retina, light detection is seemingly not required for intact development and function of the Drosophila visual system. Even rearing flies in complete darkness yields only small behavioral changes. It is proposed that reducing light sensitivity to a single type of photoreceptor (norpA-rescue) should elicit even smaller changes or no effect at all (Schnaitmann, 2024)
Finally, future studies are required to reveal whether or how the demonstrated roles of Dm9 in spectral processing and adaptation are relevant for the execution of intact visually guided behavior. It is speculated that, in the absence of glutamatergic signaling from Dm9 to inner photoreceptor terminals, spectral and other visual behaviors should in particular be affected at low intensities of ambient light (Schnaitmann, 2024)
Do Drosophila Dm9 cells functionally resemble vertebrate horizontal cells?
Dm9 cells have been postulated as “horizontal-cell-like” neurons due to their anatomical features, positioning, connectivity in the peripheral visual system, findings from calcium imaging, and predictions generated by a computational model. The current calcium recordings demonstrate that Dm9 cells indeed provide important contributions to spectral inhibition in all four R7/R8 photoreceptor types and corroborate pivotal constraints of the above mentioned model. The global decrease in the calcium responses and shift to higher light sensitivity that were observed in R7/R8 terminals upon optogenetic inhibition of Dm9 further suggest that Dm9 have an additional role in adaptation. Dm9 and horizontal cells have similar positions in the visual circuitry, accomplish multiple tasks in early visual processing and therefore share more functional commonalities than previously expected (Schnaitmann, 2024)
How neurons form appropriately sized dendritic fields to encounter their presynaptic partners is poorly understood. The Drosophila medulla is organized in layers and columns and innervated by medulla neuron dendrites and photoreceptor axons. This study shows that three types of medulla projection (Tm) neurons extend their dendrites in stereotyped directions and to distinct layers within a single column for processing retinotopic information. In contrast, the Dm8 amacrine neurons form a wide dendritic field to receive approximately 16 R7 photoreceptor inputs. R7- and R8-derived Activin selectively restricts the dendritic fields of their respective postsynaptic partners, Dm8 and Tm20, to the size appropriate for their functions. Canonical Activin signaling promotes dendritic termination without affecting dendritic routing direction or layer. Tm20 neurons lacking Activin signaling expand their dendritic fields and aberrantly synapse with neighboring photoreceptors. It is suggested that afferent-derived Activin regulates the dendritic field size of their postsynaptic partners to ensure appropriate synaptic partnership (Ting, 2014).
Studies of the past decade have revealed the molecular mechanisms by which photoreceptor and lamina axons target to specific medulla layers and restricting their axonal terminals to their retinotopic columns. However, obtaining comparable information on their synaptic partners, the dendrites of the medulla neurons, has been stymied by the morphological complexity of the dendrites and the lack of genetic and analytical tools. This study reports new techniques for visualizing and analyzing the dendritic structures of the medulla neurons. The dual-imaging technique reported in this study requires only regular confocal microscopes to generate 3D images of dendrites, and the registration method exploits the regular array structure of the medulla to permit the standardization of dendritic patterns for analyses. In addition, recent studies have begun to unravel the transcription programs that govern the development of diverse medulla cell types and serial EM reconstruction studies have illustrated their complex synaptic circuits. These advances are paving the way to studies on dendrite patterning in three-dimensional space and the establishment of synaptic partnerships between axons and dendrites in the Drosophila medulla (Ting, 2014).
Morphometric analyses of Tm neurons revealed dendritic properties distinct from those of the da sensory neurons in the PNS. Different classes of da neurons have distinguishable morphometric parameters, such as branching geometry, and their distinguishing dendritic morphological features are correlated with specific branching orders. In contrast, the three Tm neuron types have similar dendritic branching and terminating frequencies, and share similar dendritic geometrical properties. As such, the standard morphometrics are ineffective in differentiating Tm types. Instead, the type-specific dendritic attributes of Tm neurons, revealed by registration, are directly related to the layer and column structure of the medulla. These distinctions likely reflect the different functions of the dendrites of da and Tm neurons serve: da dendrites are free-ending sensory branches that tile the two-dimensional body wall, whereas Tm dendrites receive synaptic inputs from retinotopically directed afferents organized in layers and columns (Ting, 2014).
Examination of the dendritic attributes of Tm neurons provides insights into how dendritic morphology is achieved in these neurons. The axons of Tm9 and Tm2 were localized to the posterior side of their cognate columns, and their dendrites extended anteriorly to innervate their cognate columns, while Tm20 axons were at the anterior side of the column and their dendrites projected posteriorly. These type-specific axonal localizations are probably a consequence of their ontogeny: Tm9 and Tm2 axons are fasciculated, and these neurons are presumably derived from one or closely related cell lineages. Layer-specific termination of dendrites, the most prominent type-specific attribute of medulla neurons, matches the layer-specific termination of their presynaptic partners. Previous developmental studies have showed that columns and layers form as the different types of afferents sequentially innervate the medulla during development. The observation that type-specific dendritic attributes are directly related to medulla layers and columns thus led to the hypothesis that different types of medulla neurons respond differentially to various afferent-derived cues to pattern dendrites (Ting, 2014).
This study has revealed that Activin signaling regulates the dendritic field size of the columnar neuron Tm20 and the wide-field amacrine neuron Dm8. TGF-β family morphogens have been shown to regulate a broad range of neurodevelopmental processes, including proliferation and cell-fate determination during early stage of development as well as the promotion of axonal development through a non-canonical pathway involving Lim kinase. This study has found that Activin signals through the canonical pathway to regulate dendritic patterning in Tm20 and Dm8 neurons: the transcription factor dSmad2 was functionally required for Activin-dependent dendritic patterning, and the lack of Activin signaling caused a global dendritic phenotype (Ting, 2014).
The specificity of Activin signaling in regulating dendritic patterning is underscored by the distinct dendritic patterning defects, as well as the absence of cell-fate transformation and axonal targeting defects, observed in mutant neurons. Activin-signaling-deficient Tm20 neurons still expressed the appropriate transcription factors and markers, and projected their axons to the correct target layer in the lobula; mutant Dm8 elaborated their dendrites in the appropriate medulla layer M6. Instead, lack of Activin signaling reduced dendritic terminating frequency and expanded dendritic fields while the planar projection orientations and layer-specific terminations were largely unaffected. It is speculated that Activin signaling leads to the transcription regulation of as-yet-unidentified factor(s) that facilitate the termination of dendritic arbors (Ting, 2014).
Several lines of evidence suggest that Activin for Tm20 and Dm8 is, at least in part, provided by their respective presynaptic partners, R8 and R7. R7 and R8, but not R1-R6, expressed Activin during pupal stages, and RNAi-mediated knockdown of Activin in photoreceptors resulted in expanded dendritic fields in the Dm8 and Tm20 neurons. The cell-specific requirement for Activin is underscored by the observations that the genetic ablation of R7s phenocopied babo in Dm8 but not in Tm20 neurons, and that stalling R8 axons in the superficial M1 layer (thereby preventing the delivery of R8-derived Activin to the M3 layer) and Activin knock-down in R8s, disrupt the normal dendritic patterning of Tm20. It is noted that either Activin-RNAi or R8 stalling caused milder Tm20 dendritic phenotypes than did removing Baboon but that their combination did not significantly enhance Tm20 dendritic phenotypes. Thus, there is likely a secondary and partially redundant Activin source, for example, other medulla neurons, for Tm20. Nonetheless, the specific requirement of afferents indicates that R8-derived Activin cannot substitute for that from R7s, and vice-versa, even though these terminals reside only a few micrometers apart. It is therefore suggested that R7- and R8-derived Activin functions, in a short-range or layer-restricted fashion, to regulate the dendritic patterning of Dm8 and Tm20, respectively (Ting, 2014).
Photoreceptor-derived morphogens, such as Hedgehog, EGF, and Jelly Belly, have been shown to act, in an anterograde fashion early in development, to regulate the proliferation and differentiation of their target area. The current finding, that photoreceptor-derived Activin regulates the dendrite development of their synaptic targets, further suggests anterograde signaling as an effective mechanism for coordinating afferent-target development, even late in development. The wide-field neuron Dm8, which mediates innate UV preference, pools approximately 16 R7 inputs and relays the information to a few Tm5c neurons in the center of their dendritic field, while the Tm20 neurons form synapses in one-to-one correspondence with R8s, and process retinotopic information. Conceivably, the formation of appropriate dendritic field sizes and the correct synaptic partnership is critical for the functions of Tm20 and Dm8 neurons. It is reasoned that the dendritic tiling mediated by mutual repulsion, an important regulatory feature of the da neurons, alone is not suitable for restricting Tm20s' and Dm8s' dendritic fields because Tm20's dendrites arborized in three-dimensional space, and Dm8's dendrites overlap extensively with their neighbors'. The use of afferent-derived Activin to regulate dendritic patterning provides an adaptable and self-compensating mechanism for afferents to control the dendritic field sizes of their synaptic targets (Ting, 2014).
Establishing appropriate sizes and shapes of dendritic arbors is critical for proper wiring of the central nervous system. This study reports that Insulin-like Peptide 2 (DILP2) locally activates transiently expressed insulin receptors in the central dendrites of Drosophila Dm8 amacrine neurons to positively regulate dendritic field elaboration. DILP2 is expressed in L5 lamina neurons, which have axonal terminals abutting Dm8 dendrites. Proper Dm8 dendrite morphogenesis and synapse formation required insulin signaling through TOR (target of rapamycin) and SREBP (sterol regulatory element-binding protein), acting in parallel with previously identified negative regulation by Activin signaling to provide robust control of Dm8 dendrite elaboration. A simulation of dendritic growth revealed trade-offs between dendritic field size and robustness when branching and terminating kinetic parameters were constant, but dynamic modulation of the parameters could mitigate these trade-offs. It is suggested that antagonistic DILP2 and Activin signals from different afferents appropriately size Dm8 dendritic fields (Luo, 2020).
This study has shown that insulin/TOR/SREBP signaling positively regulates dendritic development. Insulin-like peptide DILP2 from L5 axons signals through insulin receptors on Dm8 dendrites to promote dendritic expansion via the canonical TOR pathway and SREBP. The roles of the insulin/TOR signaling pathway in cell growth and differentiation have been very well established. However, this is the first study to definitively demonstrate that insulin/TOR/SREBP signaling regulates dendritic field sizes during development. In Xenopus tadpoles, insulin receptor is expressed in retina and optic tectum, where it regulates synapse density and visual circuit function. However, the source of insulin or insulin-like growth factor (IGF) and the signaling pathway have not been described in developing Xenopus tectum. In vitro studies in cultured cells suggest that insulin/IGF signaling can not only regulate dendrite spine morphology and neurite outgrowth, but also function as a synaptic component at the postsynaptic density (PSD). It would be interesting to determine whether afferent-derived insulin/IGF is also utilized for dendritic development in vertebrate nervous systems (Luo, 2020).
The PI3K/AKT/TOR signaling pathway is known to mediate a broad spectrum of functions through two major complexes, TORC1 and TORC2, which act through different and varied effectors. In cultured hippocampal neurons, the PI3K-TOR pathway regulates dendrite size and branching patterns, presumably by interacting with the microtubule plus-end-tracking protein CLIP-170 to enhance crosstalk between CLIP-170 and the actin-binding protein IQGAP1 (Swiech, 2011). Dm8 dendritic development appears to involve the TORC1 complex and one of its downstream targets, SREBP. Removing the TORC2 component, Rictor, or TORC1 targets other than SREBP had no significant effect on Dm8 dendritic field size. In contrast, SREBP mutant Dm8 neurons had a reduced dendritic field size, and overexpression of SREBP rescued the dendritic phenotype of Tor mutants. While the possibility cannot be ruled out that other TOR targets play an additional or minor role, the results suggest that TOR works primarily through SREBP to regulate Dm8 dendritic development. Recent studies showed that SREBP enhances the transcription of cholesterol and fatty acid synthesis enzymes to regulate dendritic development of Da neurons in the peripheral nervous system. Thus, mounting evidence suggests that lipid synthesis may be a key regulatory point for controlling dendritic development in both central and peripheral nervous systems (Luo, 2020).
Using a novel receptor-based activity-dependent GRASP (R-synGRASP) technique, this study found synaptic defects between R7 photoreceptors and mutant Dm8 neurons that lack Insulin/TOR pathway signaling components. While the R-GRASP method revealed that Tor or chico mutant Dm8 dendrites formed membrane contacts with R7 axonal terminals in numbers that correlated with the dendritic field sizes, the results from the R-synGRASP method suggested these contacts do not constitute active synapses. Notably, Pten mutant Dm8 neurons appear to form more active synapses with peripheral R7 photoreceptors than central R7 photoreceptors, in contrast to wild types. At this stage of inquiry, the nature of such defects remains unclear. The presynaptic structures of R7 photoreceptors appear to be unaltered, as judged by the localization of the active zone marker, Bruchpilot. The level of histamine receptors on dendrites is significantly reduced in Tor- but not chico mutant Dm8 neurons, suggesting that reduced receptor levels might only partially account for the lack of R-synGRASP signal. It has been shown that insulin signaling regulates protein trafficking in various neuronal models, and mutations in SREBP and its upstream regulator easily shocked (eas) have been associated with excitability defects. Further studies will be required to resolve the nature of the synaptic defects in Tor and chico mutant Dm8 neurons (Luo, 2020).
Antagonistic regulation of Dm8 dendritic expansion by afferent-derived factors ensures robust size control
A previous study showed that R7-derived Activin negatively regulates the expansion of Dm8 dendritic fields (Ting, 2014). This action is countered by positive regulation by L5-derived DILP2 revealed in this study. Despite the antagonistic effects on Dm8 dendritic expansion, both Activin and DILP2 are derived from afferents, delivered to the growth cones, and likely act on dendrites at close range. Both L3 and L5 lamina neurons express DILP2, but RNAi-mediated knockdown of dlip2 in L5 neurons alone is sufficient to reduce Dm8 dendritic expansion. This result suggests that L3-derived DILP2 is not necessary for normal development of Dm8 dendrites, even though the axonal terminals of L3 and L5 are only few micrometers apart. Similarly, Activin derived from R7 and R8 photoreceptors acts respectively on Dm8 and Tm20 neurons, even though the R7 and R8 axonal terminals are also only separated by a few micrometers (Ting, 2014). Furthermore, both Activin and DILP2 appear to have multiple functions in the developing optic lobe. In addition to its function on Dm8 dendritic development, R7-derived Activin functions in an autocrine fashion to control R7 axonal tiling. Glia-derived DILPs regulate lamina neuron differentiation, while L3 and L5 neurons express DILP2, which also affects photoreceptor axonal tiling. It is thus tempting to speculate that afferent-derived factors tend to function at short range on multiple targets during distinct developmental stages (Luo, 2020).
The results from genetic interaction experiments further argue that Activin and DILP2 signal in developing Dm8 dendrites through parallel pathways. Interestingly, the antagonistic regulation of Dm8 dendritic expansion by two afferent-derived factors is required for elaboration of appropriately large and robust Dm8 dendritic fields - removing both signaling events rendered highly variable sizes of Dm8 dendritic fields. While antagonistic regulation has been shown to control a wide range of biological processes, including gene and protein expression, this work provides the first example of antagonistic regulation for robust control of dendritic field sizes (Luo, 2020).
How does antagonistic regulation by insulin and Activin signaling ensure appropriate and robust sizes of Dm8 dendritic fields? Several lines of evidence suggest the involvement of spatiotemporal restriction of insulin signaling. Strong insulin receptor expression was detected in Dm8 neurons at early pupal stages (20-40 hr APF), when the cells had just begun to expand their dendritic arbors, but expression disappeared by late pupal stages (60 hr APF), before the dendrites expanded to adult sizes. Furthermore, the insulin receptors in developing Dm8 neurons were largely restricted to the central dendritic field, corresponding to one single column. Thus, insulin signaling likely occurs only over a short developmental period in response to insulin signal from one or a few L5 neurons to transiently promote Dm8 dendritic expansion during the early pupal stages (Luo, 2020).
To further understand how antagonistic regulation might control dendritic field sizes, simulations were carried out of Dm8 dendritic development using a kinetic Monte Carlo method. The variance of dendritic field sizes correlated with the difference between two key dendritic development parameters, branching and terminating rates: high branching and low terminating rates favor dendritic field expansion at the cost of high variability, while low branching and high terminating rates lead to small but robust dendritic field sizes. This study found that elaboration of large and consistent dendritic fields could be achieved by spatial and/or temporal segregation of growing and terminating phases. A previous study using in vivo imaging of dendritic dynamics showed that dendrites are elaborated at different rates according to the developmental stage. Thus, it is speculated that spatiotemporal restriction of insulin signaling, coupled with Activin-mediated negative regulation, allows Dm8 neurons to elaborate a large and consistent dendritic field. The large and robust sizes of Dm8 dendritic fields not only ensure a consistent receptive field for approximately 14 R7 photoreceptor inputs, but also can receive negative feedback signals to generate a center-surround signal structure (Luo, 2020).
Detecting motion is a feature of all advanced visual systems, nowhere more so than in flying animals, like insects. In flies, an influential autocorrelation model for motion detection, the elementary motion detector circuit (EMD), compares visual signals from neighboring photoreceptors to derive information on motion direction and velocity. This information is fed by two types of interneuron, L1 and L2, in the first optic neuropile, or lamina, to downstream local motion detectors in columns of the second neuropile, the medulla. Despite receiving carefully matched photoreceptor inputs, L1 and L2 drive distinct, separable pathways responding preferentially to moving 'on' and 'off' edges, respectively. Serial electron microscopy (EM) identifies two types of transmedulla (Tm) target neurons, Tm1 and Tm2, that receive apparently matched synaptic inputs from L2. Tm2 neurons also receive inputs from two retinotopically posterior neighboring columns via L4, a third type of lamina neuron. Light microscopy reveals that the connections in these L2/L4/Tm2 circuits are highly determinate. Single-cell transcript profiling suggests that nicotinic acetylcholine receptors mediate transmission within the L2/L4/Tm2 circuits, whereas L1 is apparently glutamatergic. It is proposed that Tm2 integrates sign-conserving inputs from neighboring columns to mediate the detection of front-to-back motion generated during forward motion (Takemura, 2011).
Given that both L2 and L4 express Choline acetyltransferase (Cha) and are thus genotypically qualified to synthesize acetylcholine and provide cholinergic input to Tm2, the expression of acetylcholine receptors in Tm2 was profiled. This proved more complex than for L2 and L4. In addition to Dα7 and Dβ1 nAcR shared with L2 and L4, Tm2 also expressed Dα1/2 and Dβ2 nAcR. The exclusive expression of nicotinic rather than muscarinic receptors (nAcR not mAcR) in Tm2 suggests that both L2 and L4 provide fast excitatory inputs to Tm2. It was also found that Tm2 expressed Cha but not VGlut, indicating that, like L2 and L4, Tm2 is also genotypically cholinergic. In summary, these data predict that both synaptic connections in the L2/L4/Tm2 network are mediated by excitatory acetylcholine systems, and therefore sign-conserving (Takemura, 2011).
While either the L1 or L2 channel alone can mediate rudimentary motion detection, each also responds differentially in walking flies, and in head-yaw assays the L2 pathway is preferentially tuned to front-to-back motion. Although the connections between L4 and L2 along the anteroposterior direction might account for this front-to-back preference, these connections are reciprocal so that information also flows from posterior to anterior, while L2's activity fails to reveal asymmetrical responses. Between L2's two targets, only Tm2 receives two additional L4 inputs from neighboring posterior columns; Tm1 does not. These L2/L4/Tm2 connections are highly determinate, underscoring a critical role in connecting neighboring L2 channels along the AP direction, in what is arguably the most important motion direction for flies since it occurs during forward flight. Interestingly, other flies have a Tm neuron closely resembling Drosophila's Tm2 morphologically, for example Tm1 in the calliphorid Phaenicia. This is proposed to receive L2 inputs, suggesting that an L2/L4/Tm2 network might be conserved in higher Diptera (Takemura, 2011).
Tm2 could conceivably serve as half of the EMD's multiplier stage, comparing the temporally delayed input from collateral L4s with the cognate signal from L2. However, electrophysiological investigations on calliphorid 'Tm1' neurons, which resemble morphologically Drosophila's Tm2, have yet to provide strong evidence for this role. An alternative interpretation is that the L2/L4/Tm2 network serves instead as a prefilter in the preprocessing stage while Tm2's output feeds into the multiplier stage. The topology and sign-conserving nature of L4/Tm2 connections suggest the spatial summation of neighboring visual signals, which could increase light sensitivity at the expense of spatial acuity. It has been suggested that under low luminance conditions, neighboring visual signals are pooled prior to their interaction at the multiplier stage, while at higher luminance levels nearest-neighbor interactions dominate motion detection. Alternatively, the L4/Tm2 connections could convert visual signals sampled from the hexagonal ommatidial array into an orthogonal coordinate upon which motion signals can be derived. Differentiating between these possibilities must await future investigations that combine genetic and electrophysiological approaches (Takemura, 2011).
Visual objects can be discriminated by static spatial features such as luminance or dynamic features such as relative movement. Flies track a solid dark vertical bar moving on a bright background, a behavioral reaction so strong that for a rigidly tethered fly, the steering trajectory is phase advanced relative to the moving bar, apparently in anticipation of its future position. By contrast, flickering bars that generate no coherent motion, or whose surface texture moves in the direction opposite to the bar generate steering responses that lag behind the stimulus. It remains unclear how the spatial properties of a bar influence behavioral response dynamics. A dark bar defined by its luminance contrast to the uniform background drives a co-directional steering response that is phase-advanced relative to the response to a textured bar defined only by its motion relative to a stationary textured background. The textured bar drives an initial contra-directional turn and phase-locked tracking. The qualitatively distinct response dynamics could indicate parallel visual processing of a luminance versus motion-defined object. Calcium imaging shows that T4/T5 motion detecting neurons are more responsive to a solid dark bar than a motion defined bar. Genetically blocking T4/T5 neurons eliminates the phase-advanced co-directional response to the luminance-defined bar, leaving the orientation response largely intact. It is concluded that T4/T5 neurons mediate a co-directional optomotor response to a luminance defined bar, thereby driving phase-advanced wing kinematics, whereas separate unknown visual pathways elicit the contra-directional orientation response (Keles, 2018).
Direction-selective neurons respond to visual motion in a preferred direction. They are direction-opponent if they are also inhibited by motion in the opposite direction. In flies and vertebrates, direction opponency has been observed in second-order direction-selective neurons, which achieve this opponency by subtracting signals from first-order direction-selective cells with opposite directional tunings. This study reports direction opponency in Drosophila that emerges in first-order direction-selective neurons, the elementary motion detectors T4 and T5. This opponency persists when synaptic output from these cells is blocked, suggesting that it arises from feedforward, not feedback, computations. These observations exclude a broad class of linear-nonlinear models that have been proposed to describe direction-selective computations. However, they are consistent with models that include dynamic nonlinearities. Simulations of opponent models suggest that direction opponency in first-order motion detectors improves motion discriminability by suppressing noise generated by the local structure of natural scenes (Badwan, 2019).
Nonlinear, multiplication-like operations carried out by individual nerve cells greatly enhance the computational power of a neural system, but understanding of their biophysical implementation is scant. This study pursued this problem in the Drosophila melanogaster ON motion vision circuit, in which the membrane potentials of direction-selective T4 neurons and of their columnar input elements were recorded in response to visual and pharmacological stimuli in vivo. Electrophysiological measurements and conductance-based simulations provide evidence for a passive supralinear interaction between two distinct types of synapse on T4 dendrites. This multiplication-like nonlinearity arises from the coincidence of cholinergic excitation and release from glutamatergic inhibition. The latter depends on the expression of the glutamate-gated chloride channel GluClα in T4 neurons, which sharpens the directional tuning of the cells and shapes the optomotor behaviour of the animals. Interacting pairs of shunting inhibitory and excitatory synapses have long been postulated as an analogue approximation of a multiplication, which is integral to theories of motion detection, sound localization and sensorimotor control (Groschner, 2022).
Nervous systems rely on nonlinearities to process information. A multiplication-like operation-possibly the simplest form of nonlinearity-is implicated in the transformation of eye-centric into head-centric coordinates, the localization of sound, the combination of multisensory signals and the detection of visual motion. The biophysical underpinnings of such an operation in a single neuron are by and large unclear. One exception is the looming detector of locusts, in which-just like on a slide rule-the sum of two logarithmically scaled signals is exponentially transformed into spike rates. Other multiplicative synaptic interactions involve NMDA receptors. Both mechanisms are contingent on threshold-like nonlinearities in the current-voltage relationships of ion channels: the gating of tetrodotoxin-sensitive sodium channels in the former and the magnesium block of NMDA receptors in the latter case. This study describes a multiplication-like nonlinearity that is independent of thresholds (Groschner, 2022).
Using the visual circuit of the fruit fly as an example, this study took advantage of the neurons' compact sizes, their known connectivity and an ability to manipulate them genetically to study the biophysical basis of the multiplication step in a Hassenstein-Reichardt detector. The membrane potentials of ON motion-sensitive T4 neurons and of their columnar input elements were measured in response to a defined set of visual stimuli. Measurements of both pre- and postsynaptic voltages obviated the need for assumptions regarding the temporal dynamics of input signals when modelling the detector's output. The voltage responses of T4 neurons were reproduced rather faithfully by a passive conductance-based model. Discrepancies between simulation and reality could be due to selective synaptic delays or the 15% of dendritic inputs from wide-field TmY15 and CT1 neurons, which were not taken into account. In the model, as in the data, the supralinearity arises from the coincidence of excitation and release from shunting inhibition. Such 'multiplicative disinhibition' constitutes the inverse operation of divisive inhibition. It is free from the voltage dependence that often limits threshold-based systems and less sensitive to changing signal amplitudes. More broadly, theory invokes multiplication as a strategy to gate information flow. The passive biophysical mechanism that is proposed could lend itself to other systems, such as the logical conjunction of chemosensory signals or the gating of cortical afferent. Motion vision in flies may provide one of many cases of multiplicative disinhibition (Groschner, 2022).
A neuron is conventionally regarded as a single processing unit. It receives input from one or several presynaptic cells, transforms these signals, and transmits one output signal to its postsynaptic partners. Exceptions exist: amacrine cells in the mammalian retina or interneurons in the locust mesothoracic ganglion are thought to represent many electrically isolated microcircuits within one neuron. An extreme case of such an amacrine cell has recently been described in the Drosophila visual system. This cell, called CT1, reaches into two neuropils of the optic lobe, where it visits each of 700 repetitive columns, thereby covering the whole visual field. Due to its unusual morphology, CT1 has been suspected to perform local computations, but this has never been proven. Using 2-photon calcium imaging and visual stimulation, this study found highly compartmentalized retinotopic response properties in neighboring terminals of CT1, with each terminal acting as an independent functional unit. Model simulations demonstrate that this extreme case of compartmentalization is at the biophysical limit of neural computation (Meier, 2019).
The visual system of the fruit fly comprises the eye with about 700 facets and the optic lobe. The latter consists of four layers of retinotopically organized neuropils called lamina, medulla, lobula, and lobula plate. Within each column of both medulla and lobula, CT1 receives input from a number of local interneurons and makes output synapses onto elementary motion-detecting T4 and T5 cells. T4 cells are sensitive to bright moving edges and have their dendrites in the most proximal layer of the medulla. T5 cells respond preferentially to dark moving edges and extend their dendrites into the posterior layer of the lobula. Both come in four subtypes, one for each of the four cardinal directions of visual motion. For each subtype, the synapses of CT1 are located precisely on the null direction side of T4 and T5 dendrites. Therefore, CT1 has been speculated to be of functional relevance for creating direction selectivity within T4 and T5, in particular because it receives excitatory, cholinergic input and releases the typically inhibitory neurotransmitter GABA, which may introduce a sign inversion. To support motion detection, however, the columnar processes of CT1 must act as spatially independent, local processing units. Curiously, these structures are connected to major branches via ultra-thin, 0.1-µ-wide processes that form loops and, thus, are about 10 times longer than needed to bridge the distance between neighboring columns (Meier, 2019).
To measure the receptive fields of individual processes in CT1, calcium levels were used - as a proxy for neural activity - recorded under a 2-photon microscope during visual stimulation with a white-noise bar pattern. By correlating the fluorescence signal of a given neural process with the stimulus intensity at each location in space and time, a spatiotemporal receptive field was obtained. As shown in a representative experiment, each compartment of CT1 has its own confined receptive field with a small half-width, comparable to the size of the interommatidial angle of the fly eye. Furthermore, receptive field centers are displaced with respect to neighboring compartments. Processes within the medulla are ON-sensitive and the ones in the lobula are OFF-sensitive, just like their postsynaptic partner cells, T4 and T5. To assess the spatiotemporal response properties more closely, such measurements were taken from many compartments of CT1 cellsm and their receptive fields were averaged after alignment to their center. The spatial profile of the processes in the medulla reveal an ON-center with a full-width at half maximum of 9.1° and temporal band-pass characteristics. The spatial profile in the lobula reveals an OFF-center with a half-width of 7.2°. Receptive field sizes determined at different locations in visual space were rather similar. To test the selectivity of the compartments to contrast polarity as well as their potential direction selectivity, bright and dark edges moving in 12 directions were used. CT1 compartments respond selectively to either moving ON (medulla) or OFF (lobula) edges, irrespective of the direction of motion (Meier, 2019).
This extreme compartmentalization requires strong electrical isolation of neural processes within neighboring columns. Given the minute absolute distances in the fruit fly brain, it was wondered whether this is at all possible. A realistic, electrically passive compartmental model of CT1 was built, based on electron microscopy reconstructions. It consists of 11 terminals in the medulla that are connected via thin, looping branches to several thicker parental branches. Using biophysically plausible membrane parameters and direct current (DC) injections into one columnar compartment at a time, the distribution of membrane voltage across the whole model cell was investigated. Irrespective of the stimulated column, the voltage drops steeply to a level of around 20% in all remaining compartments, indicating strong electrical isolation of each CT1 terminal from its neighbors. As a control, the same current injection into the tip of the thick main branch was calculated. As expected, the voltage spreads more uniformly under these conditions and drops to only about 50% of the peak voltage was examined. Next, how the compartmentalization of the model cell depends on the choice of parameters for the axial (Ra) and transmembrane (Rm) resistance. To this end, current injections into one compartment were simulated, and the average relative membrane potential was measured in all other terminals while varying the values for Ra and Rm. The model only retains a high degree of compartmentalization if Ra is rather high or if Rm is low. How this translates into receptive field properties of an isolated CT1 terminal, as determined in imaging experiments, was examined. To test this, synaptic input into one compartment was simulated, assuming three Gaussian input signals of 7° half-width, separated by 5°. Testing the receptive field of the central of three columnar CT1 compartments, a spatial receptive field half-width was found of 9.6°. Hence, the receptive field is largely determined by the spatial sensitivity of the input and closely replicates the experimental findings. In order to detect visual movement, local motion detectors must receive input signals that are separated in space and in time. To probe whether CT1 terminals fulfill this requirement, the model cell was presented with a sine grating. When the stimulus was stationary, it was observed that the membrane potential in neighboring terminals changed according to the phase of the grating, further corroborating the results from the receptive field analysis. Once the grating was moved in one direction, voltage responses of individual terminals were modulated robustly at the temporal frequency of the stimulus. Critically for motion detection, phase delays between signals were preserved. This illustrates that CT1 terminals retain information about the spatial as well as the temporal characteristics of the input signal and further demonstrates their usefulness in the motion detection pathway (Meier, 2019).
Using functional 2-photon calcium imaging and anatomically realistic compartmental modeling, strong evidence is provided for an extreme case of electrically isolated sub-compartments in a large-field amacrine cell, which functions as many hundred independent processing units. This has important implications for the potential role of CT1 in motion processing in the visual system of Drosophila. Recently, several studies have demonstrated that the computation of direction selectivity in the fly brain is based on both preferred direction enhancement and null direction suppression. As a result, presynaptic elements to the elementary motion detectors T4 and T5 must contribute both excitation (on the preferred side) and inhibition (on the null side). Although evidence for inhibitory, columnar inputs to T4 has been shown before, the only cell type in the OFF pathway potentially providing null direction suppression is the GABAergic wide-field neuron CT1. In this study, it was demonstrated that CT1 can indeed perform local computations and is therefore a promising candidate for the direction selectivity circuit, whose exact role has to be further investigated in the future (Meier, 2019).
In these experiments, changes in calcium concentration were measured, whereas in the modeling section, membrane voltage distributions were calculated. Potential discrepancies between these two readouts of neural activity need to be kept in mind. It is possible that the transformation from underlying membrane polarization to voltage-gated calcium dynamics either enhances or reduces compartmentalization between terminals through compressive or expansive channel activation functions. Any model of this transformation would require knowledge of channel properties and distribution that is currently not available. However, a recent study has shown that, if anything, calcium signaling tends to be more compartmentalized than voltage in the fly visual system. Finally, because synaptic output depends on calcium concentration at release sites, the strong compartmentalization that was observe in the calcium measurements should be of immediate functional relevance (Meier, 2019).
In summary, these results show that, in order to ensure an electrical decoupling of the different processes of an amacrine cell in the small brain of the fly, connecting branches have to be long and thin. Additionally, either specific axial resistance has to be unusually high or membrane resistance must be at the low end of the spectrum. This demonstrates that such an extreme case of compartmentalization is at the biophysical limit of neural computation (Meier, 2019).
Neurotransmitter receptors and ion channels shape the biophysical properties of neurons, from the sign of the response mediated by neurotransmitter receptors to the dynamics shaped by voltage-gated ion channels. Therefore, knowing the localizations and types of receptors and channels present in neurons is fundamental to understanding of neural computation. This study developed two approaches to visualize the subcellular localization of specific proteins in Drosophila: the flippase-dependent expression of GFP-tagged receptor subunits in single neurons and 'FlpTag', a versatile new tool for the conditional labelling of endogenous proteins. Using these methods, the subcellular distribution of the receptors GluClα, Rdl, and Dα7 and the ion channels Para and Ih in motion-sensing T4/T5 neurons of the Drosophila visual system was investigated. A strictly segregated subcellular distribution of these proteins a sequential spatial arrangement of glutamate, acetylcholine, and GABA receptors was discovered along the dendrite that matched the previously reported EM-reconstructed synapse distributions (Fendl, 2020).
How neural circuits implement certain computations in order to process sensory information is a central question in systems neuroscience. In the visual system of Drosophila, much progress has been made in this direction: numerous studies examined the response properties of different cell-types in the fly brain and electron microscopy studies revealed the neuronal wiring between them. However, one element crucial to understanding is still missing; these are the neurotransmitter receptors used by cells at the postsynaptic site. This knowledge is essential since neurotransmitters and corresponding receptors define the sign and the time-course of a connection, that is whether a synapse is inhibitory or excitatory and whether the signal transduction is fast or slow. The same neurotransmitter can act on different receptors with widely differing effects for the postsynaptic neuron. Glutamate for instance is mainly excitatory, however, in invertebrates it can also have inhibitory effects when it acts on a glutamate-gated chloride channel, known as GluClα. Recently, it has also been shown that acetylcholine, usually excitatory, might also be inhibitory in Drosophila, if it binds to the muscarinic mAChR-A receptor. Hence, knowledge inferring the type of transmitter receptor at a synapse is essential for understanding of the way neural circuits process information (Fendl, 2020).
Moreover, voltage-gated ion channels shape synaptic transmission and the integration of synaptic inputs by defining the membrane properties of every neural cell type. The voltage-gated calcium channel cacophony, for instance, mediates influx of calcium ions that drives synaptic vesicle fusion at presynaptic sites. Voltage-gated sodium channels like Paralytic (Para) are important for the cell's excitability and the generation of sodium-dependent action potentials. The voltage-gated channel Ih influences the integration and kinetics of excitatory postsynaptic potentials. However, only little is known about how these channels are distributed in neurons and how this shapes the neural response properties (Fendl, 2020).
One of the most extensively studied neural circuits in Drosophila is the motion vision pathway in the optic lobe and the underlying computation for direction-selectivity. The optic lobe comprises four neuropils: lamina, medulla, lobula, and lobula plate. As in the vertebrate retina, the fly optic lobe processes information in parallel ON and OFF pathways. Along the visual processing chain, T4/T5 neurons are the first neurons that respond to visual motion in a direction selective way. T4 dendrites reside in layer 10 of the medulla and compute the direction of moving bright edges (ON-pathway). T5 dendrites arborize in layer 1 of the lobula and compute the direction of moving dark edges (OFF-pathway). The four subtypes of T4/T5 neurons (a, b, c, d), project axon terminals to one of the four layers in the lobula plate, each responding only to movement in one of the four cardinal directions, their preferred direction (Fendl, 2020).
How do T4/T5 neurons become direction-selective? Both T4 and T5 dendrites span around eight columns collecting signals from several presynaptic input neurons, each of which samples information from visual space in a retinotopic manner. The functional response properties of the presynaptic partners of T4/T5 have been described in great detail along with their neurotransmitter phenotypes. T4 dendrites receive glutamatergic, GABAergic and cholinergic input, whereas T5 dendrites receive GABAergic and cholinergic input only. These input synapses are arranged in a specific spatial order along T4/T5 dendrites (Fendl, 2020).
Which receptors receive this repertoire of different neurotransmitters at the level of T4/T5 dendrites? Recently, several RNA-sequencing studies described the gene expression pattern of nearly all cell-types in the optic lobe of the fruit fly including T4/T5 neurons. T4/T5 neurons were found to express numerous receptor subunits of different transmitter classes and voltage-gated ion channels at various expression strengths. However, RNA-sequencing studies do not unambiguously answer the above question for two reasons: mRNA and protein levels are regulated in complex ways via post-transcriptional, translational, and protein degradation mechanisms making it difficult to assign protein levels to RNA levels. Secondly, standard RNA-sequencing techniques cannot provide spatial information about receptor localizations, hence, they are not sufficient to conclude which transmitter receptors receive which input signal. Both shortcomings could in principle be overcome by antibody staining since immunohistochemical techniques detect neurotransmitter receptors at the protein level and preserve spatial information. However, high-quality antibodies are not available for every protein of interest and may have variable affinity due to epitope recognition. Furthermore, labeling ion channels via antibodies and ascribing expression of a given channel to a cell-type in dense neuronal tissue remains challenging. The disadvantages of the above techniques highlight the need for new strategies for labeling neurotransmitter receptors in cell types of interest (Fendl, 2020).
This study employed existing and generated new genetic methods to label and visualize ion channels in Drosophila. For endogenous, cell-type-specific labeling of proteins, a generalizable method called FlpTag was developed that expresses a GFP-tag conditionally. Using these tools, the subcellular distribution was determined of the glutamate receptor subunit GluClα, the acetylcholine receptor subunit Dα7, and the GABA receptor subunit Rdl in motion-sensing T4/T5 neurons. These receptor subunits were differentially localized between dendrites and axon terminals. Along the dendrites of individual T4/T5 cells, the receptor subunits GluClα, Rdl, and Dα7 reveal a distinct distribution profile that can be assigned to specific input neurons forming synapses in this area. Furthermore, it was demonstrated the generalizability of the FlpTag approach by generating lines for the metabotropic GABA receptor subunit Gaba-b-r1 and the voltage-gated ion channels para and Ih. The strategies described in this study can be applied to other cells as well as other proteins to reveal the full inventory and spatial distribution of the various ion channels within individual neurons (Fendl, 2020).
Neurotransmitter receptors are essential neuronal elements that define the sign and temporal dynamics of synaptic connections. For understanding of complex neural circuits, it is indispensable to examine which transmitter receptor types are used by the participating neurons and to which compartment they localize. This study developed FlpTag, a generalizable method for endogenous, cell-type-specific labeling of proteins. Alongside several GFP-tagged UAS-lines, the newly developed FlpTag lines were developed to explore the distribution of receptor subunits GluClα, Rdl, Dα7, Gaba-b-r1 and voltage-gated ion channels Para and Ih in motion-sensing T4/T5 neurons of the visual system of Drosophila. These ion channels were found to be localized to either the dendrite, the axonal fiber or the axon terminal. Even at the level of individual dendrites, GluClα, Rdl and Dα7 were differentially distributed precisely matching the locations where T4 and T5 neurons sample signals from their glutamatergic, cholinergic, or GABAergic input neurons, respectively (Fendl, 2020).
Working with Drosophila as model organism bears some unrivaled advantages when it comes to genetic tools. The MiMIC and FlyFos libraries, for instance, are large-scale approaches of enormous value for the fly community as they provide GFP-tagged protein lines for thousands of Drosophila genes including several neurotransmitter receptors and voltage-gated ion channels. Recently, Kondo expanded these existing libraries with T2A-Gal4 insertions in 75 neurotransmitter receptor genes that can also be exchanged by the fluorescent protein tag Venus (Kondo, 2020). While all these approaches tag genes at their endogenous locus, none of them are conditional, for example they cannot be applied in a cell-type-specific manner. Hence, ascribing the expression of the pan-neuronally tagged proteins to cell-types of interest are challenging in dense neuronal tissue (Fendl, 2020).
To overcome these difficulties, two conditional strategies were used for the investigation of membrane protein localizations in the cell types of interest, T4 and T5 neurons. First, GFP-tagged UAS-lines were developed for GluClα and Rdl, and an existing UAS-Dα7::GFP line was tested. As stated above, aberrant localization of overexpressed proteins can occur, however, this is not always the case. Overexpression of UAS-GluClα::GFP shows a similar receptor localization pattern as both MiMIC and FlpTag endogenous lines, thus, validating the use of UAS-GluClα::GFP for studying receptor distribution. Additionally, previous studies reported that the UAS-Dα7::GFP line showed proper localization of the acetylcholine receptor to endogenous synapses when compared to antibody stainings or endogenous Bruchpilot (Brp) puncta. This study confirmed confirmed this finding and further showed that Dα7::GFP presumably localizes only to cholinergic synapses. Overexpressing Dα7::GFP in a medulla neuron that is devoid of endogenous Dα7 demonstrated that Dα7::GFP localized to apparent cholinergic synapses. Hence, the UAS-Dα7::GFP line can be used to study the distribution of cholinergic synapses, but not the exact composition of cholinergic receptor subunits. A recent study showed that quantitatively the levels of the postsynaptic density protein PSD95 change when overexpressed, but qualitatively the localization is not altered. Altogether, this suggests that tagged overexpression lines can be used for studying protein localizations, but they have to be controlled carefully and drawn conclusions might be different for every line (Fendl, 2020).
Ideally, a tool for protein tagging should be both endogenous and conditional. This can be achieved by introducing an FRT-flanked STOP cassette upstream of the gene of interest which was engineered with an epitope tag or fluorescent protein. Only upon cell-type specific expression of Flp, the tagged protein will be expressed in a cell-type specific manner. This genetic strategy was utilized by two independent studies to label the presynaptic protein Brp, the histamine channel Ort and the vesicular acetylcholine transporter VAChT. Recently, a new approach based on the split-GFP system was utilized for endogenous, conditional labeling of proteins in two independent studies. However, all these aforementioned approaches are not readily generalizable and easily applicable to any gene of interest (Fendl, 2020).
The FlpTag strategy presented in this study overcomes these caveats by allowing for endogenous, conditional tagging of proteins and by offering a generalizable toolbox for targeting many genes of interest. Similar to the conditional knock-out tools FlpStop and FlipFlop, FlpTag utilizes a FLEx switch to conditionally control expression of a reporter gene, in this case GFP. Likewise, FlpTag also easily integrates using the readily available intronic MiMIC insertions. This study attempted to generate FlpTag lines for six genes, GluClα, Rdl, Dα7, Gaba-b-r1, para and Ih. Four out of these six lines yielded conditional GFP-tagged protein lines (GluClα, Gaba-b-r1, para, Ih). The FlpTag cassette was injected in MI02620 for Rdl and MI12545 for Dα7, but no GFP expression was detected across the brain. The MiMiC insertion sites used for Rdl and Dα7 seem to be in a suboptimal location for tagging the protein (Fendl, 2020).
As of now, there are MiMIC insertions in coding introns for more than 2800 genes available, which covers approximately 24% of neuronal genes. Additionally, the attP insertion sites generated in the study by Kondo provide possible landing sites for the FlpTag cassette for 75 neurotransmitter receptor genes (Kondo, 2020). Transmembrane proteins such as neurotransmitter receptors form complex 3D structures making fluorescent tagging especially difficult. Neither the MiMIC insertion sites, nor the target sites of the Kondo study at the C-terminus of several transmitter receptor genes, ensure a working GFP-tagged protein line. For genes of interest lacking a suitable MiMIC insertion site a homology directed repair (HDR) cassette was generated that utilizes CRISPR/Cas9-mediated gene editing to integrate the FlpTag cassette in any desired gene locus. The plasmid consists of the FlpTag cassette flanked by multiple cloning sites for the insertion of homology arms (HA). Through HDR the FlpTag cassette can be knocked-in into any desired locus. Taken together, the FlpTag cassette is a generalizable tool that can be integrated in any available attP-site in genes of interest or inserted by CRISPR-HDR into genes lacking attP landing sites. This allows for the investigation of the endogenous spatial distributions of proteins, as well as the correct temporal dynamics of protein expression (Fendl, 2020).
Further, the FlyFos project demonstrated that most fly lines with an extra copy of GFP-tagged protein-coding genes worked normally and GFP-tagged proteins could be imaged in living fly embryos and pupae. In principle, live-imaging of the GFP-tagged lines that were created could be performed during different developmental stages of the fruit fly. In general, the tools generated in this study can be used as specific postsynaptic markers, visualizing glutamatergic, GABAergic, and cholinergic synapses with standard confocal light microscopy. This extends the existing toolbox of Drosophila postsynaptic markers for studying the localization and development of various types of synapses (Fendl, 2020).
T4/T5 neurons combine spatiotemporal input from their presynaptic partners, leading to selective responses to one of the four cardinal directions. Numerous studies investigated the mechanisms underlying direction-selective responses in T4/T5 neurons, yet the computation is still not fully understood. At an algorithmic level, a three-arm detector model is sufficient to describe how direction-selective responses in T4/T5 neurons arise. This model relies on the comparison of signals originating from three neighboring points in space via a delay-and-compare mechanism. The central arm provides fast excitation to the neuron. While one flanking arm amplifies the central signal for stimuli moving along the preferred direction, the other inhibits the central signal for stimuli moving along the null direction of the neuron. Exploring the neurotransmitter receptors and their distribution on T4/T5 dendrites allows defining the sign as well as the temporal dynamics of some of the input synapses to T4/T5 (Fendl, 2020).
According to the algorithmic model, an excitatory, amplifying input signal on the distal side of T4/T5 dendrites was expected. This study found that T4 cells receive an inhibitory, glutamatergic input from Mi9 via GluClα, which, at first sight, seems to contradict expectation. However, since Mi9 has an OFF-center receptive field, this glutamatergic synapse will invert the polarity from Mi9-OFF to T4-ON. Theoretically, in darkness, Mi9 inhibits T4 via glutamate and GluClα, and this inhibition is released upon an ON-edge moving into its receptive field. The concomitant closure of chloride channels and subsequent increased input resistance in T4 cells results in an amplification of a subsequent excitatory input signal from Mi1 and Tm3. As shown by a recent modeling study, this biophysical mechanism can indeed account for preferred direction enhancement in T4 cells (Borst, 2018). Some studies failed to detect preferred direction enhancement in T4/T5 neurons and they proposed that the enhanced signal in PD seen in GCaMP recordings could be a result from a non-linear calcium-to-voltage transformation. If this was really the case, the role of Mi9 and GluClα must be reconsidered and future functional experiments will shed light onto this topic (Fendl, 2020).
Nevertheless, Strother (2017) showed that the RNAi- knock-down of GluClα in T4/T5 neurons leads to enhanced turning responses on the ball set-up for faster speeds of repeating ON and OFF edges (Strother, 2017). Although this observation cannot answer the question about preferred direction enhancement in T4 cells, it indicates that both T4 and T5 receive inhibitory input and that removal of such create enhanced turning responses at the behavioral level. In line with these observations, the glutamate receptor GluClα was also found in T4/T5 axon terminals. A possible functional role of these inhibitory receptors in the axon terminals could be a cross-inhibition of T4/T5 cells with opposite preferred directions via lobula plate intrinsic neurons (LPis). Glutamatergic LPi neurons are known to receive a cholinergic, excitatory signal from T4/T5 neurons within one layer and to inhibit lobula plate tangential cells, the downstream postsynaptic partners of T4/T5 neurons, via GluClα in the adjacent oppositely tuned layer. This mechanism induces a motion opponent response in lobula plate tangential cells and increases their flow-field selectivity. In addition, LPi neurons could also inhibit T4/T5 neurons presynaptically at their axon terminals via GluClα in order to further sharpen the flow-field selectivity of lobula plate tangential cells. Taken together, exploring the subcellular distribution of GluClα in T4/T5 neurons highlights its differential functional roles in different parts of these cell types (Fendl, 2020).
Secondly, the Dα7 signal in the center of T4/T5 dendrites discovered in this study, corresponds to ionotropic, cholinergic input from Mi1 and Tm3 for T4, and Tm1, Tm2 and Tm4 for T5. These signals correspond to the central, fast, excitatory arm of the motion detector model. As T4 and T5 express a variety of different ACh receptor subunits, the exact subunit composition and underlying biophysics of every cholinergic synapse on T4/T5 dendrites still awaits further investigations (Fendl, 2020).
Third, inhibition via GABA plays an essential role in creating direction-selective responses in both T4 and T5 neurons by providing null direction suppression. Computer simulations showed that direction selectivity decreases in T4/T5 motion detector models without this inhibitory input on the null side of the dendrite. This study shows that T4 and T5 neurons possess the inhibitory GABA receptor subunit Rdl mainly on the proximal base on the null side of their dendrites, providing the synaptic basis for null direction suppression. The metabotropic GABA receptor subunit Gaba-b-r1 was not detected in T4/T5 neurons using the newly generated FlpTag Gaba-b-r1 line. Finally, all of the receptor subunits GluClα, Rdl and Dα7 investigated in this study are ionotropic, fast receptors, which presumably do not add a temporal delay at the synaptic level. In the detector model described above, the two outer arms provide a slow and sustained signal, and such properties are already intrinsic properties of these input neurons. However, it cannot be excluded that slow, metabotropic receptor subunits for acetylcholine or GABA (e.g. Gaba-br2) which are also present in T4/T5 and could induce additional delays at the synaptic level (Fendl, 2020).
Furthermore, the subcellular distribution was investigated of the voltage-gated ion channels Para and Ih in T4/T5 neurons. Para, a voltage-gated sodium channel, was found to be distributed along the axonal fibers of both T4 and T5 neurons. As Para is important for the generation of sodium-dependent action potentials, it will be interesting for future functional studies to investigate, if T4/T5 really fire action potentials and how this shapes their direction-selective response. Further, Ih, a voltage-gated ion channel permeable for several types of ions, was detected in T4/T5 dendrites using the FlpTag strategy. Ih channels are activated at negative potentials below -50 mV and as they are permeable to sodium and potassium ions, they can cause a depolarization of the cell after hyperpolarization. Loss-of-function studies will unravel the functional role of the Ih channel for direction-selective responses in T4/T5 neurons (Fendl, 2020).
Since the ability to combine synaptic inputs from different neurotransmitters at different spatial sites is common to all neurons, the approaches described in this study represent an important future perspective for other circuits. The tools can be used to study the ion channels GluClα, Rdl, Dα7, Gaba-b-r1, para and Ih in any given Drosophila cell-type and circuit. Furthermore, the FlpTag tool box can be used to target many genes of interest and thereby foster molecular questions across fields (Fendl, 2020).
The techniques described in this study can be transferred to other model organisms as well, to study the distribution of different transmitter receptors. For instance, in the mouse retina - similar to motion-sensing T4/T5 neurons in the fruit fly - so-called On-Off direction-selective ganglion cells receive asymmetric inhibitory GABAergic inputs from presynaptic starburst amacrine cells during null-direction motion. A previous study investigated the spatial distribution of GABA receptors of these direction-selective ganglion cells using super-resolution imaging and antibody staining. Additionally, starburst amacrine cells also release ACh onto ganglion cells which contributes to the direction-selective responses of ganglion cells. Thus, mapping the distribution of ACh receptors on direction-selective ganglion cells will be the next important step to further investigate cholinergic transmission in this network (Fendl, 2020).
Overall, this study has demonstrated the importance of exploring the distributions of neurotransmitter receptors and ion channels for systems neuroscience. The distinct distributions in T4/T5 neurons discovered in this study and the resulting functional consequences expand knowledge of the molecular basis of motion vision. Although powerful, recent RNAseq studies lacked information about spatial distributions of transmitter receptors which can change the whole logic of wiring patterns and underlying synaptic signs. Future studies can use this knowledge to target these receptors and directly probe their role in functional experiments or incorporate the gained insights into model simulations. However, this study is only highlighting some examples of important neural circuit components: expanding the approaches described in this study to other transmitter receptors and ion channels, as well as gap junction proteins will reveal the full inventory and the spatial distributions of these decisive determinants of neural function within an individual neuron (Fendl, 2020).
Diverse sensory systems, from audition to thermosensation, feature a separation of inputs into ON (increments) and OFF (decrements) signals. In the Drosophila visual system, separate ON and OFF pathways compute the direction of motion, yet anatomical and functional studies have identified some crosstalk between these channels. This well-studied circuit was used to ask whether the motion computation depends on ON-OFF pathway crosstalk. Using whole-cell electrophysiology, visual responses of T4 (ON) and T5 (OFF) cells were recorded and their composite ON-OFF receptive fields were mapped: it was found that they share a similar spatiotemporal structure. A biophysical model was fit to these receptive fields that accurately predicts directionally selective T4 and T5 responses to both ON and OFF moving stimuli. This model also provides a detailed mechanistic explanation for the directional preference inversion in response to the prominent reverse-phi illusion. Finally, the steering responses of tethered flying flies was used to validate the model's predicted effects of varying stimulus parameters on the behavioral turning inversion (Gruntman, 2021).
Sensory systems flexibly adapt their processing properties across a wide range of environmental and behavioral conditions. Such variable processing complicates attempts to extract a mechanistic understanding of sensory computations. This is evident in the highly constrained, canonical Drosophila motion detection circuit, where the core computation underlying direction selectivity is still debated despite extensive studies. This study measured the filtering properties of neural inputs to the OFF motion-detecting T5 cell in Drosophila. Stimulus-dependent changes in the shape of these signals, which become more biphasic under specific conditions. Summing these inputs within the framework of a connectomic-constrained model of the circuit demonstrates that these shapes are sufficient to explain T5 responses to various motion stimuli. Thus, these stimulus- and state-dependent measurements reconcile motion computation with the anatomy of the circuit. These findings provide a clear example of how a basic circuit supports flexible sensory computation (Kohn, 2021).
Self-motion generates visual patterns on the eye that are important for navigation. These optic flow patterns are encoded by the population of local direction-selective cells in the mouse retina, whereas in flies, local direction-selective T4/T5 cells are thought to be uniformly tuned. How complex global motion patterns can be computed downstream is unclear. This study shows that the population of T4/T5 cells in Drosophila encodes global motion patterns. Whereas the mouse retina encodes four types of optic flow, the fly visual system encodes six. This matches the larger number of degrees of freedom and the increased complexity of translational and rotational motion patterns during flight. The four uniformly tuned T4/T5 subtypes described previously represent a local subset of the population. Thus, a population code for global motion patterns appears to be a general coding principle of visual systems that matches local motion responses to modes of the animal's movement (Henning, 2022).
Flying insects exhibit remarkable navigational abilities controlled by their compact nervous systems. Optic flow, the pattern of changes in the visual scene induced by locomotion, is a crucial sensory cue for robust self-motion estimation, especially during rapid flight. The best-known optic-flow sensitive neurons are the large tangential cells of the dipteran lobula plate. This study reports the comprehensive reconstruction and identification of the Lobula Plate Tangential Neurons in an Electron Microscopy (EM) volume of a whole Drosophila brain. This catalog of 58 LPT neurons (per brain hemisphere) contains many neurons that are described here for the first time and provides a basis for systematic investigation of the circuitry linking self-motion to locomotion control. Leveraging computational anatomy methods, it is estimated the visual motion receptive fields of these neurons and compared their tuning to the visual consequence of body rotations and translational movements. These neurons were matched, in most cases on a one-for-one basis, to stochastically labeled cells in genetic driver lines, to the mirror-symmetric neurons in the same EM brain volume, and to neurons in an additional EM data set. Using cell matches across data sets, the integration of optic flow patterns by neurons downstream of the LPTs was analyzed and most central brain neurons were found to establish sharper selectivity for global optic flow patterns than their input neurons. Furthermore, it was found that self-motion information extracted from optic flow is processed in distinct regions of the central brain, pointing to diverse foci for the generation of visual behaviors (Zhao, 2023).
Animals have evolved sophisticated visual circuits to solve a vital inference problem: detecting whether or not a visual signal corresponds to an object on a collision course. Such events are detected by specific circuits sensitive to visual looming, or objects increasing in size. Various computational models have been developed for these circuits, but how the collision-detection inference problem itself shapes the computational structures of these circuits remains unknown. Inspired by the distinctive structures of LPLC2 neurons (ultra-selective looming detecting neuron, lobula plate/lobula columnar, type II (LPLC2) in Drosophila) in the visual system of Drosophila, this study built anatomically-constrained shallow neural network models and trained them to identify visual signals that correspond to impending collisions. Surprisingly, the optimization arrives at two distinct, opposing solutions, only one of which matches the actual dendritic weighting of LPLC2 neurons. Both solutions can solve the inference problem with high accuracy when the population size is large enough. The LPLC2-like solutions reproduces experimentally observed LPLC2 neuron responses for many stimuli, and reproduces canonical tuning of loom sensitive neurons, even though the models are never trained on neural data. Thus, LPLC2 neuron properties and tuning are predicted by optimizing an anatomically-constrained neural network to detect impending collisions. More generally, these results illustrate how optimizing inference tasks that are important for an animal's perceptual goals can reveal and explain computational properties of specific sensory neurons (Zhou, 2022).
Many sighted animals solve this inference problem with high precision, thanks to robust loom-selective neural circuits evolved over hundreds of millions of years. The neuronal mechanisms for response to looming stimuli have been studied in a wide range of vertebrates, from cats and mice to zebrafish, as well as in humans. In invertebrates, detailed anatomical, neurophysiological, behavioral, and modeling studies have investigated loom detection, especially in locusts and flies. An influential mathematical model of loom detection was derived by studying the responses of the giant descending neurons of locusts. This model established a relationship between the timing of the neurons' peak responses and an angular size threshold for the looming object. Similar models have been applied to analyze neuronal responses to looming signals in flies, where genetic tools make it possible to precisely dissect neural circuits, revealing various neuron types that are sensitive to looming signals (Zhou, 2022).
However, these computational studies did not directly investigate the relationship between the structure of the loom-sensitive neural circuits and the inference problem they appear to solve. On the one hand, the properties of many sensory circuits appear specifically tuned to the tasks that they are executing. In particular, by taking into account relevant behaviors mediated by specific sensory neurons, experiments can provide insight into their tuning properties. On the other hand, computational studies that have trained artificial neural networks to solve specific visual and cognitive tasks, such as object recognition or motion estimation, have revealed response patterns similar to the corresponding biological circuits or even individual neurons. Thus, in this study it is asked whether the properties associated with neural loom detection can be reproduced simply by optimizing shallow neural networks for collision detection (Zhou, 2022).
The starting point for the computational model of loom detection is the known neuroanatomy of the visual system of the fly. In particular, the loom-sensitive neuron LPLC2 (lobula plate/lobula columnar, type 2) has been studied in detail (Wu, 2016). These neurons tile visual space, sending their axons to descending neurons called the giant fibers (GFs), which trigger the fly's jumping and take-off behaviors. Each LPLC2 neuron has four dendritic branches that receive inputs at the four layers of the lobula plate (LP). The retinotopic LP layers host the axon terminals of motion detection neurons, and each layer uniquely receives motion information in one of the four cardinal directions. Moreover, the physical extensions of the LPLC2 dendrites align with the preferred motion directions in the corresponding LP layers. These dendrites form an outward radial structure, which matches the moving edges of a looming object that expands from the receptive field center. Common stimuli such as the wide-field motion generated by movement of the insect only match part of the radial structure, and strong inhibition for inward-directed motion suppresses responses to such stimuli. Thus, the structure of the LPLC2 dendrites favors responses to visual stimuli with edges moving radially outwards, corresponding to objects approaching the receptive field center (Zhou, 2022).
The focus of this paper is to investigate how loom detection in LPLC2 can be seen as the solution to a computational inference problem. Can the structure of the LPLC2 neurons be explained in terms of optimization-carried out during the course of evolution-for the task of predicting which trajectories will result in collisions? How does coordination among the population of more than 200 LPLC2 neurons tiling a fly's visual system affect this optimization? To answer these questions, simple anatomically-constrained neural network models were built that receive motion signals in the four cardinal directions. The model using artificial stimuli to detect visual objects on a collision course with the observer. Surprisingly, optimization finds two distinct types of solutions, with one resembling the LPLC2 neurons and the other having a very different configuration. How each of these solutions detects looming events and where they show distinct individual and population behaviors were analyzed. When tested on visual stimuli not in the training data, the optimized solutions with filters that resemble LPLC2 neurons exhibit response curves that are similar to those of LPLC2 neurons measured experimentally. Importantly, although it only receives motion signals, the optimized model shows characteristics of an angular size encoder, which is consistent with many biological loom detectors, including LPLC2. These results show that optimizing a neural network to detect looming events can give rise to the properties and tuning of LPLC2 neurons (Zhou, 2022).
The radially structured dendrites of the LPLC2 neuron in the LP can account for its response to motion radiating outward from the receptive field center. The results show that the logic of this computation can be understood in terms of inferential loom detection by the population of units. In particular, for an individual detector unit, an inward structure can make a better loom detector than an outward structure, since it is sensitive to colliding objects originating from a wider array of incoming angles. As the number of units across visual space increases, the performance of the outward-sensitive receptive field structure comes to match the performance of the inward solutions. As the number of units increases, the inhibitory component of the outward solutions also becomes broader as the population size becomes larger, which is crucial for reproducing key experimental observations, such as peripheral inhibition. The optimized solutions depend on the number of detectors, and this is likely related to the increasing overlap in receptive fields as the population grows. This result is consistent with prior work showing that populations of neurons often exhibit different and improved coding strategies compared to individual neurons. Thus, understanding anatomical, physiological, and algorithmic properties of individual neurons can require considering the population response. The solutions that were found to the loom inference problem suggest that individual LPLC2 responses should be interpreted in light of the population of LPLC2 responses (Zhou, 2022).
Our results shed light on discussions of η-like (encoding angular size) and ρ-like (encoding angular velocity) looming sensitive neurons in the literature. In particular, these optimized models clarify an interesting but puzzling fact: LPLC2 neurons transform their inputs of direction-selective motion signals to computations of angular size (Ache, 2019). Consistent with this tuning, the model also shows a linear relationship between the peak time relative to collision and the R/η ratio, which should be followed by loom sensitive neurons that encode angular size. In both cases, these properties appear to be the simple result of training the constrained model to reliably detect looming stimuli (Zhou, 2022).
The units of the outward solution exhibit sparsity in their responses to looming stimuli, in contrast to the denser representations in the inward solution. During a looming event, in an outward solution, most of the units are quiet and only a few adjacent units have very large activities, reminiscent of sparse codes that seem to be favored, for instance, in cortical encoding of visual scenes. Since the readout of the model is a summation of the activities of the units, sparsity does not directly affect the performance of the model, but is an attribute of the favored solution. For a model with a different loss function or with noise, the degree of sparsity might be crucial. For instance, the sparse code of the outward model might make it easier to localize a hit stimulus, or might make the population response more robust to noise (Zhou, 2022).
Experiments have shown that inhibitory circuits play an important role for the selectivity of LPLC2 neurons. For example, motion signals at the periphery of the receptive field of an LPLC2 neuron inhibit its activity. This peripheral inhibition causes various interesting response patterns of the LPLC2 neurons to different types of stimuli. However, the structure of this inhibitory field is not fully understood, and the model provides a tool to investigate how the inhibitory inputs to LPLC2 neurons affect circuit performance on loom detection tasks. The strong inhibition on the periphery of the receptive field arises naturally in the outward solutions after optimization. The extent of the inhibitory components increases as more units are added to models. The broad inhibition appears in the model to suppress responses to the non-hit stimuli, and as in the data, the inhibition is broader than one might expect if the neuron were simply being inhibited by inward motion. These larger inhibitory fields are also consistent with the larger spatial pooling likely to be supplied by inhibitory LPi inputs (Zhou, 2022).
The synthetic stimuli used to train models in this study were unnatural in two ways. The first way was in the proportion of hits and non-hits. Training was carried out with 25% of the training data representing hits. The true fraction of hits among all stimuli encountered by a fly is undoubtedly much less, and this affects how the loss function weights different types of errors. It is also clear that a false-positive hit (in which a fly might jump to escape an object not on collision course) is much less penalized during evolution than a false-negative (in which a fly doesn't jump and an object collides, presumably to the detriment of the fly). It remains unclear how to choose these weights in the training data or in the loss function, but they affect the receptive field weights optimized by the model (Zhou, 2022).
The second issue with the stimuli is that they were caricatures of stimulus types, but did not incorporate the richness of natural stimuli. This richness could include natural textures and spatial statistics, which seem to impact motion detection algorithms. This richness could also include more natural trajectories for approaching objects. Another way to enrich the stimuli would be to add noise, either in inputs to the model or in the model's units themselves. This was explored briefly by adding self-rotation-generated background motion; under those conditions, both solutions were present but optimized outward solutions performed better than the inward solutions. This indicates that the statistics of the stimuli may play an important role in selecting solutions for loom detection. However, it remains less clear what the true performance limits of loom detection are, since most experiments use substantially impoverished looming stimuli. Moreover, it is challenging to characterize the properties of natural looming events. An interesting future direction will be to investigate the effects of more complex and naturalistic stimuli on the model's filters and performance, as well as on LPLC2 neuron responses themselves (Zhou, 2022).
For simplicity, the models did not impose the hexagonal geometry of the compound eye ommatidia. Instead, it was assumed that the visual field is separated into a Cartesian lattice with 5° spacing, each representing a local motion detector with two spatially separated inputs. This simplification alters slightly the geometry of the motion signals compared to the real motion detector receptive fields. This could potentially affect the learned spatial weightings and reproduction of the LPLC2 responses to various stimuli, since the specific shapes of the filters matter. Thus, the hexagonal ommatidial structure and the full extent of inputs to T4 and T5 might be crucial if one wants to make comparisons with the dynamics and detailed responses of LPLC2 neurons. However, this geometric distinction seems unlikely to affect the main results of how to infer the presence of hit stimuli (Zhou, 2022).
The model requires a field of estimates of the local motion. Here, the simplest model was used - the Hassenstein-Reichardt correlator model Equation 3 - but the model could be extended by replacing it with a more sophisticated model for motion estimation. Some biophysically realistic ones might take into account synaptic conductances and could respond to static features of visual scenes. Alternatively, in natural environments, contrasts fluctuate in time and space. Thus, if one includes more naturalistic spatial and temporal patterns, one might consider a motion detection model that could adapt to changing contrasts in time and space (Zhou, 2022).
Although the outward filter of the unit emerges naturally from the gradient descent training protocol, that does not mean that the structure is learned by LPLC2 neurons in the fly. There may be some experience dependent plasticity in the fly eye , but these visual computations are likely to be primarily genetically determined. Thus, one may think of the computation of the LPLC2 neuron as being shaped through millions of years of evolutionary optimization. Optimization algorithms at play in evolution may be able to avoid getting stuck in local optima, and thus work well with the sort of shallow neural network found in the fly eye (Zhou, 2022).
This study focused on the motion signal inputs to LPLC2 neurons, and other inputs to LPLC2 neurons were neglected, such as those coming from the lobula that likely report non-motion visual features. It would be interesting to investigate how this additional non-motion information affects the performance and optimized solutions of the inference units. For instance, another lobula columnar neurons, LC4, is loom sensitive and receives inputs in the lobula. The LPLC2 and LC4 neurons are the primary excitatory inputs to the GF, which mediates escape behaviors. The inference framework set out here would allow one to incorporate parallel non-motion intensity channels, either by adding them into the inputs to the LPLC2-like units, or by adding in a parallel population of LC4-like units. This would require a reformulation of a probabilistic model. Notably, one of the most studied loom detecting neurons, the lobula giant movement detector (LGMD) in locusts, does not appear to receive direction-selective inputs, as LPLC2 does. Thus, the inference framework set out here could be flexibly modified to investigate loom detection under a wide variety of constraints and inputs, which allow it to be applied to other neurons, beyond LPLC2 (Zhou, 2022).
The perception of motion is an important function of vision. Neural wiring diagrams for extracting directional information have been obtained by connectome reconstruction. Direction selectivity in Drosophila is thought to originate in T4/T5 neurons through integrating inputs with different temporal filtering properties. Through genetic screening based on synaptic distribution, a new type of TmY neuron, termed TmY-ds, was isolated that forms reciprocal synaptic connections with T4/T5 neurons. Its neurites responded to grating motion along the four cardinal directions and showed a variety of direction selectivity. Intriguingly, its direction selectivity originated from temporal filtering neurons rather than T4/T5. Genetic silencing and activation experiments showed that TmY-ds neurons are functionally upstream of T4/T5. These results suggest that direction selectivity is generated in a tripartite circuit formed among these three neurons-temporal filtering, TmY-ds, and T4/T5 neurons, in which TmY-ds plays a role in the enhancement of direction selectivity in T4/T5 neurons (Zhao, 2022).
The HisCl1 histamine receptor acts in photoreceptors to synchronize Drosophila behavioral rhythms with light-dark cycles"
In Drosophila, the clock that controls rest-activity rhythms synchronizes with light-dark cycles through either the blue-light sensitive cryptochrome (Cry) located in most clock neurons, or rhodopsin-expressing histaminergic photoreceptors. This study shows that, in the absence of Cry, each of the two histamine receptors Ort and HisCl1 contribute to entrain the clock whereas no entrainment occurs in the absence of the two receptors. In contrast to Ort, HisCl1 does not restore entrainment when expressed in the optic lobe interneurons. Indeed, HisCl1 is expressed in wild-type photoreceptors and entrainment is strongly impaired in flies with photoreceptors mutant for HisCl1. Rescuing HisCl1 expression in the Rh6-expressing photoreceptors restores entrainment but it does not in other photoreceptors, which send histaminergic inputs to Rh6-expressing photoreceptors. These results thus show that Rh6-expressing neurons contribute to circadian entrainment as both photoreceptors and interneurons, recalling the dual function of melanopsin-expressing ganglion cells in the mammalian retina (Alejevski, 2019).
The Drosophila sleep–wake rhythms are controlled by a brain circadian clock that includes about 150 clock neurons. Light synchronizes the clock neuronal network through cell-autonomous and non-cell-autonomous light input pathways. Cry is a blue-light sensitive photoreceptor protein that is expressed in most clock neurons. In the absence of Cry, flies do not phase-shift their behavioral rhythms in response to a short light pulse but still synchronize to light–dark (LD) cycles. Only flies devoid of both Cry and rhodopsin-expressing photoreceptors fail to entrain to LD cycles. Six different rhodopsins (Rhs) have been characterized in the Drosophila photoreceptive structures, which include the compound eye, the Hofbauer-Buchner (H-B) eyelet, and ocelli. The compound eye strongly contributes to circadian photoreception, whereas a modest contribution appears to be brought by the H-B eyelet and the ocelli. A circadian function has been recently associated with the yet poorly characterized rhodopsin 7, although its exact contribution and localization in the brain and/or the eye remains controversial. In addition to entrainment, the visual system controls other features of the clock neuron network by conveying light information to either promote or inhibit the behavioral output of specific clock neuron subsets (Alejevski, 2019).
The compound eye includes about 800-unit eyes (ommatidia), each of which contains eight photoreceptors. The six Rh1-expressing outer photoreceptors (R1–6) are involved in motion detection and project to the lamina neuropile of the optic lobe. The two inner photoreceptors (R7–8) are important for color detection and project to the medulla. They express four different rhodopsins and thus define two types of ommatidia: 'pale' (p) ommatidia (30%) include a Rh3-expressing R7 and a Rh5-expressing R8, whereas 'yellow' (y) ommatidia (70%) include a Rh4-expressing R7 and a Rh6-expressing R8. Each extra-retinal H-B eyelet contains four Rh6-expressing photoreceptors that project to the accessory medulla, in the vicinity of key pacemaker neurons, the ventral lateral neurons (LNvs) that produce the pigment-dispersing factor (PDF) neuropeptide9,20–24. Each of the three ocelli contains about 80 photoreceptors that express Rh225. The Drosophila rhodopsins cover a wide range of wavelengths from 300 nm to 600 nm18,19, with only Rh1 and Rh6 being sensitive to red light (Alejevski, 2019).
Rhodopsin-dependent circadian entrainment involves two downstream signaling pathways, the canonical one that relies on the phospholipase C encoded by the no receptor potential A gene (norpA)2 or an unknown pathway that does not contribute in very low light levels. All but Rh2- and Rh5- expressing photoreceptors support synchronization in very low light, and at least Rh1, Rh5, and Rh6 can signal through the NorpA-independent pathway. Photoreceptors of the compound eye are histaminergic but the H-B eyelet expresses both histamine and acetylcholine. Although the two neurotransmitters might contribute to circadian entrainment, flies devoid of Cry and histidine decarboxylase do not synchronize their rest–activity rhythms with LD cycles. This suggests that besides Cry, there is no histamine-independent pathway to entrain the clock (Alejevski, 2019).
Two genes encoding histamine-gated chloride channels, ora transientless (ort) and Histamine-gated chloride channel subunit 1 (HisCl1), have been identified in Drosophila. The ort-null mutants are visually blind and their electroretinograms have no ON and OFF transients. In contrast, HisCl1 mutants show increased OFF transients, whereas slower responses were observed in the postsynaptic laminar monopolar cells. Based on transcriptional reporters, ort expression in the optic lobes was observed in neurons of both the lamina and medulla/lobula neuropils. Based on reporter gene expression, HisCl1 was localized in glial cells of the lamina. However, recent work reported expression in photoreceptors, in particular in the R7 and R8 inner photoreceptor subtypes. Indeed, Ort and HisCl1 support color opponency between the two subtypes of 'inner' photoreceptors, the ultraviolet (UV)-sensitive R7 and non-UV-sensitive R8, with HisCl1 and Ort mediating direct and indirect inhibition, respectively. The histaminergic pathways that are involved in circadian entrainment are unknown and are the subject of the present study. The results show that both Ort and HisCl1 define two different pathways for circadian entrainment. Whereas Ort contributes through its expression in the interneurons of the optic lobe, HisCl1 mostly contributes through its expression in the Rh6-expressing retinal photoreceptors. The work thus reveals that Rh6-expressing neurons contribute to light-mediated entrainment as both photoreceptors and interneurons (Alejevski, 2019).
This work reveals that the Cryptochrome-independent entrainment of rest–activity rhythms relies on distinct pathways that are contributed by the two histamine receptors Ort and HisCl1. Whereas Ort mediates circadian entrainment through the optic lobe interneurons that are involved in visual functions, HisCl1 defines a new photoreceptive pathway through Rh6-expressing photoreceptors. Although both receptors mediate synchronization with a shifted LD cycle, it seems likely that the two pathways will show differences in specific light conditions. It was not possible to rescue Ort function with HisCl1 expression in the ort-expressing cells, whereas the Ort could replace HisCl1 in Rh6 photoreceptors. It is possible that HisCl1 has a lower affinity for histamine with Rh6 cells receiving more neurotransmitter than optic lobe interneurons. Alternatively, interneurons could sufficiently differ from photoreceptors for their physiology or specific receptor-interacting protein content, preventing HisCl1 from working efficiently. HisCl1 downregulation in Rh6 cells slows down synchronization and flies with HisCl1134 mutant eyes synchronize very poorly with advanced LD cycles, and fail to synchronize with delays. It cannot be excluded that non-photoreceptor cells contribute to HisCl1-dependent entrainment but other pathways appear to have a modest contribution if any (Alejevski, 2019).
HisCl1 is expressed in the H-B eyelet, which could thus contribute to this synchronization pathway. However, the cell-killing experiments indicate that H-B eyelet is not required for HisCl1-mediated synchronization through Rh6 cells. In the recently described color opponency mechanism, retinal R7 cells inhibit R8 and vice versa through HisCl1 expression in the photoreceptors (Schnaitmann, 2018). It is supposed that HisCl1-dependent clock synchronization is also mediated by the hyperpolarization of Rh6-expressing cells. How this hyperpolarization interacts with the light-induced depolarization in Rh6 photoreceptors to result in a synchronization message to the clock neurons remains to be understood. Since only Rh6-expressing R8 and not the other inner photoreceptors contribute to this circadian photoreception pathway, Rh6 cells might have specific connections with downstream interneurons. Such specificity has been described for color vision where each of the four inner photoreceptor subtypes connects to a different type of TmY interneuron in the Medulla. This study shows that HisCl1 expression in Rh6 cells supports synchronization with red light, in the absence of Rh1, indicating that an intra-Rh6-photoreceptor circuit is sufficient. This indicates that Rh6-expressing R8 photoreceptors play a dual photoreceptor/interneuron role in this pathway (Model for the retinal input pathways to the brain clock). Whether the same individual cells have the two roles is unknown, although the HisCl1-dependent color opponency mechanism suggests that it could be the case. It is also unclear whether all Rh6-expressing R8 photoreceptors or only a fraction of them contribute to circadian synchronization. The results imply that, in addition to histaminergic neurotransmission, Rh6-expressing photoreceptors can talk to downstream interneurons through histamine-independent neurotransmission. A recent transcriptomics study indeed revealed the expression of cholinergic markers in R7 and R8 cells, supporting cholinergic transmission in the inner photoreceptors, in addition to histaminergic transmission (Alejevski, 2019).
The data indicate that histaminergic inputs from both outer and inner photoreceptors converge to Rh6 cells to contribute to circadian entrainment. It is possible that some of these inputs rely on Rh7, which seems to be expressed in Rh6-expressing photoreceptors, according to transcriptional reporter data. Putative connections between photoreceptors have been described in Drosophila and other insects. How R1–6 photoreceptors might be connected to Rh6-expressing R8 cells remains difficult to understand, but a few putative contacts between presynaptic outer cells and postsynaptic inner cells have been observed in Musca. The intra-retinal functional connectivity that this study reports in Drosophila is reminiscent to the circuit logic of circadian entrainment in the mammalian retina, where intrinsically photoreceptive retinal ganglion cells express the melanopsin photopigment in addition to receiving inputs from rods and cones. Interestingly, melanopsin appears to share light-sensing properties with the rhabdomeric photoreceptors of invertebrates. It has been shown that the mammalian circadian clock can synchronize with day–night cycles by tracking light color changes in addition to light intensity changes. It will be interesting to investigate the possible contribution of the dual function of Rh6-expressing photoreceptors to integrating different color cues into the retinal information that is sent to the clock (Alejevski, 2019).
The circuitry underlying the detection of visual motion in Drosophila melanogaster is one of the best studied networks in neuroscience. Lately, electron microscopy reconstructions, algorithmic models, and functional studies have proposed a common motif for the cellular circuitry of an elementary motion detector based on both supralinear enhancement for preferred direction and sublinear suppression for null-direction motion. In T5 cells, however, all columnar input neurons (Tm1, Tm2, Tm4, and Tm9) are excitatory. So, how is null-direction suppression realized there? Using two-photon calcium imaging in combination with thermogenetics, optogenetics, apoptotics, and pharmacology, this study discovered that it is via CT1, the GABAergic large-field amacrine cell, where the different processes have previously been shown to act in an electrically isolated way. Within each column, CT1 receives excitatory input from Tm9 and Tm1 and provides the sign-inverted, now inhibitory input signal onto T5. Ablating CT1 or knocking down GABA-receptor subunit Rdl significantly broadened the directional tuning of T5 cells. It thus appears that the signal of Tm1 and Tm9 is used both as an excitatory input for preferred direction enhancement and, through a sign inversion within the Tm1/Tm9-CT1 microcircuit, as an inhibitory input for null-direction suppression (Braun, 2023).
Vision is critical for the regulation of mating behaviors in many species. This study discovered that the Drosophila ortholog of human GABA (A) -receptor-associated protein (GABARAP; Atg8a) is required to fine-tune male courtship by modulating the activity of visual feedback neurons, lamina tangential cells (Lat). GABARAP is a ubiquitin-like protein that regulates cell-surface levels of GABA (A) receptors. Knocking down GABARAP or GABA (A) receptors in Lat neurons or hyperactivating them induces male courtship toward other males. Inhibiting Lat neurons, on the other hand, delays copulation by impairing the ability of males to follow females. Remarkably, the human ortholog of Drosophila GABARAP restores function in Lat neurons. Using in vivo two-photon imaging and optogenetics, Lat neurons were shown to be functionally connected to neural circuits that mediate visually-guided courtship pursuits in males. This work reveals a novel physiological role for GABARAP in fine-tuning the activity of a visual circuit that tracks a mating partner during courtship (Mabuchi, 2023).
An important step in neural information processing is the transformation of membrane voltage into calcium signals leading to transmitter release. However, the effect of voltage to calcium transformation on neural responses to different sensory stimuli is not well understood. This study used in vivo two-photon imaging of genetically encoded voltage and calcium indicators, ArcLight and GCaMP6f, respectively, to measure responses in direction-selective T4 neurons of female Drosophila Comparison between ArcLight and GCaMP6f signals reveals calcium signals to have a significantly higher direction selectivity compared with voltage signals. Using these recordings, a model was built which transforms T4 voltage responses into calcium responses. Using a cascade of thresholding, temporal filtering and a stationary nonlinearity, the model reproduces experimentally measured calcium responses across different visual stimuli. These findings provide a mechanistic underpinning of the voltage to calcium transformation and show how this processing step, in addition to synaptic mechanisms on the dendrites of T4 cells, enhances direction selectivity in the output signal of T4 neurons. Measuring the directional tuning of postsynaptic vertical system (VS)-cells with inputs from other cells blocked, this study found that, indeed, it matches the one of the calcium signal in presynaptic T4 cells (Mishra, 2023).
To survive, animals must convert sensory information into appropriate behaviours. Vision is a common sense for locating ethologically relevant stimuli and guiding motor responses. How circuitry converts object location in retinal coordinates to movement direction in body coordinates remains largely unknown. This study shows through behaviour, physiology, anatomy and connectomics in Drosophila that visuomotor transformation occurs by conversion of topographic maps formed by the dendrites of feature-detecting visual projection neurons (VPNs) into synaptic weight gradients of VPN outputs onto central brain neurons. This gradient motif transforms the anteroposterior location of a visual looming stimulus into the fly's directional escape. Specifically, it was discovered that two neurons postsynaptic to a looming-responsive VPN type promote opposite takeoff directions. Opposite synaptic weight gradients onto these neurons from looming VPNs in different visual field regions convert localized looming threats into correctly oriented escapes. For a second looming-responsive VPN type, graded responses along the dorsoventral axis were demonstrated. This synaptic gradient motif generalizes across all 20 primary VPN cell types and most often arises without VPN axon topography. Synaptic gradients may thus be a general mechanism for conveying spatial features of sensory information into directed motor outputs (Dombrovski, 2023).
Tracking visual objects while maintaining stable gaze is complicated by the different computational requirements for figure-ground discrimination, and the distinct behaviors that these computations coordinate. Drosophila melanogaster uses smooth optomotor head and body movements to stabilize gaze, and impulsive saccades to pursue elongated vertical bars. Directionally selective motion detectors T4 and T5 cells provide inputs to large-field neurons in the lobula plate, which control optomotor gaze stabilization behavior. This study hypothesized that an anatomically parallel pathway represented by T3 cells, which provide inputs to the lobula, drives bar tracking body saccades. Physiological and behavioral experiments were combined to show that T3 neurons respond omnidirectionally to the same visual stimuli that elicit bar tracking saccades, silencing T3 reduced the frequency of tracking saccades, and optogenetic manipulation of T3 acted on the saccade rate in a push-pull manner. Manipulating T3 did not affect smooth optomotor responses to large-field motion. These results show that parallel neural pathways coordinate smooth gaze stabilization and saccadic bar tracking behavior during flight (Frighetto, 2023).
Visual motion estimation is a canonical neural computation. In Drosophila, recent advances have identified anatomic and functional circuitry underlying direction-selective computations. Models with varying levels of abstraction have been proposed to explain specific experimental results but have rarely been compared across experiments. This studys used the wealth of available anatomical and physiological data to construct a minimal, biophysically inspired synaptic model for Drosophila's first-order direction-selective T4 cells. This model relates mathematically to classical models of motion detection, including the Hassenstein-Reichardt correlator model. Numerical simulation was used to test how well this synaptic model could reproduce measurements of T4 cells across many datasets and stimulus modalities. These comparisons include responses to sinusoid gratings, to apparent motion stimuli, to stochastic stimuli, and to natural scenes. Without fine-tuning this model, it sufficed to reproduce many, but not all, response properties of T4 cells. Since this model is flexible and based on straightforward biophysical properties, it provides an extensible framework for developing a mechanistic understanding of T4 neural response properties. Moreover, it can be used to assess the sufficiency of simple biophysical mechanisms to describe features of the direction-selective computation and identify where understanding must be improved (Zavatone-Veth, 2020).
In visual systems, neurons adapt both to the mean light level and to the range of light levels, or the contrast. Contrast adaptation has been studied extensively, but it remains unclear how it is distributed among neurons in connected circuits, and how early adaptation affects subsequent computations. This study investigated temporal contrast adaptation in neurons across Drosophila's visual motion circuitry. Several ON-pathway neurons showed strong adaptation to changes in contrast over time. One of these neurons, Mi1, showed almost complete adaptation on fast timescales, and experiments ruled out several potential mechanisms for its adaptive properties. When contrast adaptation reduced the gain in ON-pathway cells, it was accompanied by decreased motion responses in downstream direction-selective cells. Simulations show that contrast adaptation can substantially improve motion estimates in natural scenes. The benefits are larger for ON-pathway adaptation, which helps explain the heterogeneous distribution of contrast adaptation in these circuits (Matulis, 2020).
Neurons can change their gain in response to their inputs. In the visual system, as scenes move across the retina, neurons may encounter many different luminances over time. Neurons can adapt to the mean of these inputs and also to the range of these inputs, or the temporal contrast. Contrasts can vary widely within a scene or across scenes. Many retinal neurons change their gain in ways that amplify low-contrast signals relative to high-contrast ones. In these cases, the adaptation has been proposed to maximize information transfer to downstream circuits. However, it remains unclear how contrast adaptation is distributed among parallel processing streams in visual circuits and how early changes in gain affect specific downstream computations. This study investigated contrast adaptation in Drosophila's visual motion circuits, where powerful genetic tools and known connectivity allow investigation both the breadth of contrast adaptation and its consequences for circuit computations (Matulis, 2020).
The benefits of contrast adaptation have often been framed in terms of efficient coding. Neurons have a limited range of responses but often receive a wide range of input signals. By adapting to the distribution of inputs, neurons can make efficient use of their limited output range to encode a fuller range of inputs; this can be formalized in terms of transmitted information. In this view, contrast adaptation is beneficial because it permits cells to represent inputs from both high- and low-contrast scenes using the full range of their responses. This study examined the role of contrast adaptation in circuitry associated with direction-selective (DS) signals and use simulations to understand contrast adaptive properties in the context of specific downstream motion computations (Matulis, 2020).
Contrast adaptation has been studied extensively in vertebrate retina. There, photoreceptors and horizontal cells do not show contrast adaptation, but downstream cells, including bipolar, amacrine, and retinal ganglion cells, exhibit varying degrees of contrast adaptation. Adaptation can occur quickly, on the scale of 100 s of milliseconds, or slowly, on the scale of several seconds. This adaptation has been found to occur cell intrinsically, mediated by membrane channels, as well as by upstream synaptic depletion (Matulis, 2020).
In the fly eye, light is detected by photoreceptors, and, from there, signals are split into parallel pathways, which relay information about the visual scene with varying degrees of rectification and delay. These ON and OFF pathways converge onto the cells T4 and T5, which respond direction selectively to light edges and dark edges, respectively, and are required for optomotor orientation and speed regulation. In flies, contrast adaptation has been observed in wide-field DS cells, which receive inputs from the local motion detectors, T4 and T5. When these wide-field cells are presented with high-contrast sinusoidal stimuli, they reduce the gain in their responses to stimuli in their preferred direction. The adapting stimulus can be presented in any direction, suggesting that this mechanism depends on upstream, non-DS cells. In a different wide-field DS cell, studies have characterized another form of adaptation to stimulus variance. There, experiments examined responses to stimulus velocity, rather than to contrast, and found that cells strongly adapted to the variance of the velocity distribution. Thus, in flies, adaptation to contrast and, more generally, to stimulus variance exists in wide-field DS cells. But contrast adaptation has not been observed in neurons upstream of them (Matulis, 2020).
This study measured contrast adaptation in the neurons upstream of motion detection. Focus was placed on one strongly adapting neuron, Mi1, and it was shown to adapt roughly as fast as it responds. Silencing was used to rule out several potential mechanisms for the contrast adaptation, including inheritance from specific other cells and synaptic depletion in a primary presynaptic partner. A simple phenomenological model can account for this neuron's fast changes in gain. Downstream of Mi1, contrast adaptation was examine in the neurons T4 and T5. Simulations show how adaptation to the stimulus contrast could substantially improve motion detection computations, an effect that is stronger in ON motion pathways (Matulis, 2020).
Detecting the direction of motion contained in the visual scene is crucial for many behaviors. However, because single photoreceptors only signal local luminance changes, motion detection requires a comparison of signals from neighboring photoreceptors across time in downstream neuronal circuits. For signals to coincide on readout neurons that thus become motion and direction selective, different input lines need to be delayed with respect to each other. Classical models of motion detection rely on non-linear interactions between two inputs after different temporal filtering. However, recent studies have suggested the requirement for at least three, not only two, input signals. This study comprehensively characterized the spatiotemporal response properties of all columnar input elements to the elementary motion detectors in the fruit fly, T4 and T5 cells, via two-photon calcium imaging. Between these input neurons, large differences were found in temporal dynamics. Based on this, computer simulations show that only a small subset of possible arrangements of these input elements maps onto a recently proposed algorithmic three-input model in a way that generates a highly direction-selective motion detector, suggesting plausible network architectures. Moreover, modulating the motion detection system by octopamine-receptor activation, the temporal tuning of T4 and T5 cells was found to be shifted toward higher frequencies, and this shift can be fully explained by the concomitant speeding of the input elements (Arenz, 2017).
The perception of visual motion is critical for animal navigation, and flies are a prominent model system for exploring this neural computation. In Drosophila, the T4 cells of the medulla are directionally selective and necessary for ON motion behavioral responses. To examine the emergence of directional selectivity, genetic driver lines were developed for the neuron types with the most synapses onto T4 cells. Using calcium imaging, it was found that these neuron types are not directionally selective and that selectivity arises in the T4 dendrites. By silencing each input neuron type, which neurons are necessary for T4 directional selectivity and ON motion behavioral responses were identified. Tthe sign of the connections between these neurons and T4 cells were determined using neuronal photoactivation. These results indicate a computational architecture for motion detection that is a hybrid of classic theoretical models (Strother, 2017).
The behavioral state of an animal can dynamically modulate visual processing. In flies, the behavioral state is known to alter the temporal tuning of neurons that carry visual motion information into the central brain. However, where this modulation occurs and how it tunes the properties of this neural circuit are not well understood. This study shows that the behavioral state alters the baseline activity levels and the temporal tuning of the first directionally selective neuron in the ON motion pathway (T4) as well as its primary input neurons (Mi1, Tm3, Mi4, Mi9). These effects are especially prominent in the inhibitory neuron Mi4, and this study shows that central octopaminergic neurons provide input to Mi4 and increase its excitability. It was further shown that octopamine neurons are required for sustained behavioral responses to fast-moving, but not slow-moving, visual stimuli in walking flies. These results indicate that behavioral-state modulation acts directly on the inputs to the directionally selective neurons and supports efficient neural coding of motion stimuli (Strother, 2018).
Sensory systems sequentially extract increasingly complex features. ON and OFF pathways, for example, encode increases or decreases of a stimulus from a common input. This ON/OFF pathway split is thought to occur at individual synaptic connections through a sign-inverting synapse in one of the pathways. This study showed that ON selectivity is a multisynaptic process in the Drosophila visual system. A pharmacogenetics approach demonstrates that both glutamatergic inhibition through GluClα and GABAergic inhibition through Rdl mediate ON responses. Although neurons postsynaptic to the glutamatergic ON pathway input L1 lose all responses in GluClalpha mutants, they are resistant to a cell-type-specific loss of GluClα. This shows that ON selectivity is distributed across multiple synapses, and raises the possibility that cell-type-specific manipulations might reveal similar strategies in other sensory systems. Thus, sensory coding is more distributed than predicted by simple circuit motifs, allowing for robust neural processing (Molina-Obando, 2019).
Animals rely on their sensory systems to process behaviorally relevant information. One common feature of sensory systems is the sequential processing of information to extract complex features from simple inputs. For example, in the visual system, photoreceptors detect light and then downstream neurons progressively extract distinct features, such as contrast, direction of motion, form, or specific objects. Sensory pathways diverge into pathways that become selective for increasingly specific features (Molina-Obando, 2019).
One prominent example is the split into ON and OFF pathways, where individual neurons become selective to either increases (ON) or decreases (OFF) in a signal. Such an ON/OFF dichotomy enables more efficient coding of stimuli in the visual system and occurs across many different species and sensory modalities, such as vision, olfaction, audition, thermosensation, and electrolocation. Examples of how the split into ON and OFF pathways is implemented in sensory information processing have already been described. In the vertebrate retina, ON and OFF pathways split downstream of glutamatergic photoreceptors where ionotropic glutamate receptors on OFF bipolar cells maintain the sign of the response in the OFF pathway, and the metabotropic glutamate receptor mGluR6, located on ON bipolar cells, inverts the sign in the ON pathway. In the olfactory system of C. elegans, an odor response can be split into parallel pathways in which glutamate-gated chloride channels mediate the ON response. While these transformations are thought to occur at specific synapses, connectomics data reveals that neural circuits are intricate and that many of the possible neuronal connections are realized. This argues that important signal transformations might actually be distributed across wider circuit motifs (Molina-Obando, 2019).
In the Drosophila visual system, ON and OFF pathways functionally split in the first order lamina interneurons, but the physiological specialization occurs one synaptic layer further downstream. In brief, information travels from the retina, which houses the photoreceptors, through three optic ganglia: the lamina, the medulla, and the lobula complex, comprising lobula and lobula plate (see Figure 1 at the following site: ON pathway medulla neurons that receive graded, glutamatergic input). Contrast is encoded by the transient response of photoreceptors, and downstream lamina neurons amplify the contrast-sensitive signal component. Then, distinct ON and OFF pathways are required to detect contrast increments and decrements, respectively. In the lamina, L1 is the major input to the ON pathway, whereas L2 and L3 feed into the OFF pathway. The assignment of L1, L2, and L3 to ON and OFF pathways originates from neuronal silencing studies. However, all lamina neurons receiving direct input from photoreceptors depolarize to the offset of light and hyperpolarize to the onset of light, thus passing on information about both ON and OFF. Voltage or calcium signals in most downstream medulla neurons then selectively report only one type of contrast polarity. The major ON pathway medulla neurons Mi1 and Tm3, for example, selectively respond with depolarization or an increase in calcium signal to ON. In the OFF pathway, most neurons instead selectively respond to OFF stimuli, retaining the response polarity of their lamina inputs. Therefore, ON selectivity requires a sign inversion between the L1 input and its postsynaptic partners Mi1 and Tm3. Previous work suggested that the L1 input to the ON pathway is glutamatergic, whereas L2 and L3, the two major inputs to the OFF pathway, are cholinergic. This suggests that glutamate might also be used as an inhibitory neurotransmitter to implement ON/OFF dichotomy in the fly visual system. However, the molecular and cellular mechanisms implementing this signal transformation are not known in Drosophila visual circuitry (Molina-Obando, 2019).
This study has identified the mechanisms underlying splitting of the ON and OFF pathways in the Drosophila visual system. As expected from the major input to the ON pathway being glutamatergic, broad GluClα function is required for all ON responses in medulla neurons or downstream direction-selective cells. However, individual cell types downstream of the glutamatergic L1 input are resilient to a cell-type-specific loss of GluClα, demonstrating that ON selectivity is computed in a distributed manner. This study further showed that both the glutamate-gated chloride channel GluClα and the GABA-gated chloride channel Rdl are widely expressed in the visual system and together mediate ON responses. Thus, ON selectivity is a multisynaptic computation that is established across distributed circuits (Molina-Obando, 2019).
This work shows that visual responses in the first ON-selective neuron of the Drosophila visual system uses a combination of GluClα and Rdl receptors. This reveals a new biophysical mechanism through which ON and OFF pathway dichotomy can be established. While pharmacology can be used to deduce the function of specific molecular mechanisms, these approaches are often not specific to one protein. GluCls and GABARs belong to the same receptor family of ligand-gated chloride channels and have closely related structure and phylogeny. All known noncompetitive antagonists like Picrotoxin, γ-HCH, dieldrin, EBOB and fibronil target both receptor types although the actions are weaker in GluCls compared to GABARs. Along these lines, PTX was thought to affect GABAA receptor at low concentrations, and additionally affect GluCls at high concentrations in vitro and in vivo. This study use of PTX-insensitive alleles for glutamate and GABA-gated chloride channels making possible the deduction that, in vivo, GluClα is already blocked by PTX at lower concentrations than previously thought, and that both GluClα and Rdl play critical roles for ON responses in the Drosophila visual system. These pharmacogenetic experiments using toxin-insensitive alleles prove to be a powerful tool to unambiguously assign specific effects to individual channels (Molina-Obando, 2019).
One benefit of the use of two inhibitory transmitter systems might be the distribution of sensory coding across parallel synapses. GluClα and Rdl also appear to have very different channel dynamics. Interestingly, PTX-insensitive GluClα and Rdl alleles predominantly rescue different aspects of the visual responses. Whereas GluClαS278T predominantly rescued the peak response in all medulla layers, RdlMDRR mainly rescued the plateau response. This is consistent with the results and with previous oocyte recordings revealing that GluClα is fast desensitizing. It is also consistent with in vivo recordings of inhibitory glutamate currents in the honeybee. In contrast, GABA receptors stay open throughout the period in which the transmitter is present. Thus, the use of different inhibitory receptors might allow different aspects of a temporally structured stimulus to be encoded. This is consistent with the finding that two different types of inhibition are also in place in the vertebrate retina. There, GABAergic and glycinergic inhibition diversify the response properties of bipolar cells through a direct influence on temporal and spatial features (Molina-Obando, 2019).
While both receptors appear to be broadly expressed in many cell types of the visual system, they could be co-expressed with different transporters and channels, and interact with different molecular partners, further diversifying their role. Another common strategy to generate functional diversity is the bringing together of different receptor subunits with certain homology. Both mammalian GlyR and GABAA receptors can function as hetero-oligomers made up of different subunits and thus generating functional diversity. There are at least three different GluCl subtypes in C. elegans that can be combined. In Drosophila, only one gene coding for a glutamate-gated chloride channel has been identified. Although alternative splicing and post-transcriptional modifications could alter channel function, all known isoforms are identical in their functional domains. However, heteropentameric channels composed of mixed Rdl and GluClα subunits have been suggested biochemically. Such a potential presence of hybrid channels might also explain the higher in vivo sensitivity of GluClα to PTX in some cell types. Finally, two distinct inhibitory transmitter systems might be suitable for individual changes during evolution, allowing for adaptation to specific contextual constraints (Molina-Obando, 2019).
The current experiments revealed that GluClα is not exclusively required in a cell-autonomous manner for ON responses, since loss of GluClα function in Mi1 or Tm3 individually does not lead to a loss of ON responses. It is unlikely that this is due to an incomplete loss of function, since independent genetic tools (FlpStop and RNAi) that both disrupted GluClα expression substantially at the mRNA level gave the same result. Furthermore, the same FlpStop allele effectively abolished all ON responses when GluClα function was disrupted within its entire expression pattern. Additionally, a PTX-resistant Rdl channel can mediate ON responses in a PTX background, although L1 is not GABAergic. Together, these results suggest that ON selectivity is not a monosynaptic computation, but that parallel functional pathways can even compensate for the loss of the major synaptic connection that links L1 directly to Mi1 or Tm3. Thus, the emergence of ON selectivity is more distributed than suggested by minimal core circuit motifs. One synaptic layer further downstream, optogenetic activation of Mi1 and Tm3 most strongly contributes to T4/T5 responses. However, the current data further show that T4/T5 neurons still respond to ON stimuli when both Mi1 and Tm3 responses are completely blocked by PTX, arguing that other neurons also significantly contribute to T4/T5 responses under visual stimulation and suggesting that coding is again more distributed at this stage (Molina-Obando, 2019).
Based on connectomics, one can speculate about candidates for the implementation of these parallel circuit motifs between L1 and Mi1 and Tm3. The lamina neuron L5 and the GABAergic feedback neurons C2 and C3 receive L1 inputs and could be part of an interconnected local microcircuit. Intercolumnar neurons, not present in the current connectome datasets, like Pm or Dm neurons, might also be involved and are likely glutamatergic. In fact, there are close to 100 cell types in the visual system and ~60 medulla neurons, but their role is so far unknown. Sensory pathway splits in the periphery are one of the most fundamental steps in sensory processing. Turning this into a process that parallel pathways can achieve might make this important feature extraction step robust to perturbations (Molina-Obando, 2019).
T4 flash responses in a GluClα-deficient background show an increase in calcium signal during the OFF epoch and a decrease during the ON epoch. For a long time, the mechanisms that generate direction-selective responses in T4/T5 neurons were thought to rely on feedforward excitatory mechanisms. Recently, it was suggested that these direction-selective cells in the fly visual system also implement mechanisms that rely on null-direction suppression. Whereas electrophysiological recordings showed inhibition in T4 when the trailing edge of the receptive field was specifically stimulated, whole-cell recording experiments of T4/T5 neurons are daunting and this is the first time that calcium imaging data directly reveals inhibition in response to single ON flashes. Since glutamatergic inhibition via GluClα was disrupted in this experimental context, the data suggests that this is due to GABAergic inhibition. Several neuronal candidates could make inhibitory synapses onto T4 dendrites. Based on connectomics and neurotransmitter identity, neurons like Mi4, C3, CT1 or TmY15 give direct input and are GABAergic. Alternatively, this decrease in calcium signal in T4 might come from a lack of excitatory inputs in a GluClα mutant background. Interestingly, Mi1 and Tm3 themselves show inhibition in response to light when GluClα is blocked. However, this effect is more pronounced at their dendrites than in their output layer and shows different kinetics. The current work might thus help uncover a GABAergic inhibitory input to T4 that is more strongly apparent in the absence of Mi1 and Tm3 excitation, and could ultimately reveal the circuit implementation for the inhibitory component of T4/T5 receptive fields. Furthermore, the data also reveals an increase in calcium during OFF stimulation. The major inputs to T4 are themselves rectified. However, rectification in T4 might not be purely inherited by its inputs but also further strengthened at the T4 dendrites. The current findings thus suggest that glutamatergic inhibition contributes to establishing or maintaining contrast selectivity in T4 (Molina-Obando, 2019).
Both GluClα and Rdl are ionotropic ligand-gated receptors. While ionotropic receptors also implement the ON and OFF pathway split in C. elegans chemosensation, examples in vertebrate vision, olfaction and gustation require metabotropic receptors. Ionotropic receptors appear to be more common in insects than in vertebrates. Furthermore, glutamate-gated chloride channels have independently arisen three times within invertebrate clades and are present in arthropods, molluscs and flatworms, arguing for a strong evolutionary benefit. Ionotropic receptors mediate rapid transduction events at scales smaller than a millisecond, whereas metabotropic ones are in the millisecond to second range and last longer, from seconds to several minutes, due to an enzymatic secondary cascade previous to channel opening (Betz, 1990; Shiells, 1994). The evolutionary choice of the specific glutamatergic inhibitory system needs to match the sensory processing speed required for accurate behavioral responses in these species. For example, at the photoreceptor level, invertebrate phototransduction is faster than vertebrate phototransduction thanks to sophisticated molecular strategies. Also, the latency of olfactory sensory neurons responses in mammals is longer than that observed in insects. One advantage that metabotropic receptors have over ionotropic receptors is further amplification of the signal. The distributed circuit architecture proposed in this study might therefore strengthen signaling in a system that uses ionotropic signaling (Molina-Obando, 2019).
This study has shown that ON selectivity is not a monosynaptic process as described in other systems. Although acute pharmacological block or a systemic loss of function of GluClα abolished all ON responses in different neurons, cell-type-specific mutants retained intact ON responses, revealing that sensory coding is distributed in the fly visual system. This not only highlights the power of fly genetics but sheds new light onto the mechanisms of ON selectivity in other systems, since conclusions about ON and OFF pathway splits being mediated by specific monosynaptic processes in systems such as the vertebrate retina or the C. elegans chemosensory system relied on systemic loss-of-function approaches. Several of these systems allow for cell-type-specific manipulations using genetic approaches. It will be interesting to revisit these systems and ask if coding is similarly distributed across multiple synapses in different sensory systems and organisms (Molina-Obando, 2019).
Inhibitory interactions and columnar inputs to an object motion detector in Drosophila
The direction-selective T4/T5 cells innervate optic-flow processing projection neurons in the lobula plate of the fly that mediate the visual control of locomotion. In the lobula, visual projection neurons coordinate complex behavioral responses to visual features, however, the input circuitry and computations that bestow their feature-detecting properties are less clear. A highly specialized small object motion detector, LC11, was studied, and its responses were shown to be suppressed by local background motion. LC11 expresses GABA-A receptors that serve to sculpt responses to small objects but are not responsible for the rejection of background motion. Instead, LC11 is innervated by columnar T2 and T3 neurons that are themselves highly sensitive to small static or moving objects, insensitive to wide-field motion and, unlike T4/T5, respond to both ON and OFF luminance steps (Keles, 2020).
The cellular mechanisms of motion vision have become rapidly advanced owing to genetic, optogenetic, and in vivo imaging tools developed in Drosophila melanogaster. As in the mammalian retina, the fly optic lobe segregates ON and OFF polarity luminance changes into parallel cellular pathways. The ON- and OFF-selective pathways supply directionally selective columnar T4 and T5 neurons, respectively. The terminals of these small-field retinotopic motion detectors innervate the third optic ganglion, the lobula plate, where their synaptic output is integrated within the large planar dendrites of projection neurons that map specific wide-field patterns of optic flow onto descending pre-motor neurons to coordinate visual behavior (Keles, 2020).
In parallel to the motion vision pathway of the lobula plate, projection neurons identified in the lobula have been shown to encode moving features such as edges or objects to influence complex visual behaviors. Roughly 20 classes of lobula columnar neurons (LCs) project to the protocerebrum where axon terminals of each class form tight glomerular neuropils. Individual LC11 neurons as well as the glomerular ensemble are highly responsive to small contrasting objects moving in any direction across the ipsilateral field of view. Unlike the output cell types of the lobula plate, little is known about how the receptive field properties of LC11 arise. This study investigated the interactions between background motion and object responses in LC11, has identified a role for GABA-mediated inhibition in shaping object detection by LC11, and identifies presynaptic inputs to LC11. Columnar neurons T2 and T3 projecting from the medulla and terminating in the second and third layers of the lobula overlap with LC11 dendrites. T2 and T3 synapse with dendrites of LC11, and T3 supplies excitatory input to LC11. Finally, it was demonstrated that T2 and T3 neurons are highly selective for small objects, are suppressed by wide-field background motion, and unlike T4/T5, show full-wave rectified ON-OFF excitatory responses to rapid transitions in luminance (Keles, 2020).
In vertebrates, neurons in the retina partially encode object information, but fail to discriminate flicker from coherent motion. Yet, higher-order neurons in the mouse superior colliculus respond strongly only to moving stimuli. Similarly, this study found that T2/T3 neurons are selective for small objects, but respond to ON and OFF flicker as well, whereas downstream LC11 is responsive to object motion, not stationary flicker. It is proposed that LC11 computes continuous object motion from local ON-OFF transients conveyed by T2/T3. Future work on examining the spatiotemporal patterning of columnar inputs to LC11, as well as the cognate neurotransmitters and receptors should reveal how these computations are achieved (Keles, 2020).
In prior work (Keles, 2017), it was demonstrated that bath applied PTX, which selectively blocks chloride currents carried by GABA-A channels or glutamate channels, resulted in LC11 displaying uncharacteristic responses to elongated bars and gratings. This result was predicted under the presumption that inhibition actively filtered wide-field input from LC11. Curiously, in the same preparation, the small object responses for LC11 were essentially eradicated. How can global loss of inhibition by bath applied PTX explain both enhanced wide-field responses and diminished small object responses in LC11? Several lines of evidence suggest that postsynaptic inhibitory neuromodulation acts on LC11 in a center-surround fashion. LC11 expresses both acetylcholine receptors and a GABA-gated chloride channel subunit Rdl. Blocking GABA-A mediated synaptic currents by genetic disruption of Rdl specifically in LC11 neurons results in a decrease in response amplitude to the smallest object tested, and yet, importantly, had no effect on the normal attenuated responses to bars or normal absence of wide-field grating responses. The results support a working model in which Rdl knockout unmasks an ON-pathway input while decreasing the normal OFF object response of LC11. These properties could be explained by an ON-pathway GABAergic input to LC11 through Rdl that normally occludes ON excitation and disinhibits OFF responses. The corollary is that suppression of responses to large objects or wide-field motion occurs upstream of these object detectors. Indeed, LC11 appears to inherit its sensitivity to small object motion from excitatory T2/T3 inputs, perhaps themselves having surround inhibition mechanisms similar to T4/T5. There appears to be two mechanisms of action that are disrupted by PTX application on LC11 receptive field properties (Keles, 2017): crossover inhibition in T2/T3, which would explain their size-tuning, and local inhibition on LC11 that normally enhances small object responses. Thus, it is proposed that upon PTX delivery abnormal bar and wide-field motion responses are conveyed from T2/T3 to LC11 and small object responses are no longer boosted (Keles, 2020).
The importance of dynamic, stimulus-specific inhibition for spatial vision has been elucidated by other studies. In mice, cortical V1 center-surround receptive fields reveal stronger inhibitory currents than excitatory currents in both the surround and center, while inhibitory currents are spread more laterally than excitatory currents. In a visual collision detection circuit in the locust, feedforward inhibitory neurons actively encode dynamical variables such as object angular size. The inhibitory GABA-A receptor subunit Rdl is expressed by nearly all neurons of the fly visual system so far tested, highlighting the ubiquity and importance of inhibition for spatial vision (Keles, 2020).
T2 and T3 neurons share several key features with LC11. First, both show significantly larger responses to small solid objects than to single object edges or elongated bars, with virtually no response to moving wide-field gratings. In the large calliphorid fly Phaenicia sericata, T2 neurons have been examined with intracellular sharp electrodes, which showed that these columnar neurons depolarize to the OFF-phase of flicker, and hyperpolarize to the ON-phase. This contrasts to the GCaMP6f recordings in Drosophila, in which T2 is excited by both ON and OFF luminance transitions. Additionally, in Phaenicia T2 responded robustly to 80 x 62° moving gratings, whereas in Drosophila no response was observed in either T2 or T3 to gratings that filled the 108 x 63° display. The T2a cell type, with similar anatomy but different presynaptic inputs to T2, may show responses more closely matching those from larger flies (Keles, 2020).
An important feature of Drosophila T2/T3 neurons is that unlike T4 and T5 columnar motion detectors, which act as half-wave rectifiers that segregate ON and OFF edge stimuli, respectively, both T2 and T3 neurons show full-wave rectification in that they are excited by both ON and OFF phases of flicker. Notably, T5 shows similar amplitude responses to the OFF edges generated by either a solid two-edged dark object or a single moving OFF edge, whereas T3 responses are markedly stronger for the solid object presenting an OFF-ON sequence than to a single progressing OFF edge. T3 appears to receive input from a combination of neurons that reside in the ON and OFF pathways, including Mi1 and Tm3, providing a possible explanation for this result. Full-wave rectification of ON and OFF stimuli is consistent with single point correlation computations proposed to comprise elementary small target motion detectors (ESTMDs), which underlie the high performance object detection seen in lobula wide-field STMD neurons of hoverflies and dragonflies. Future work must explore the mechanisms that shape responses in T2 and T3, and how the spatiotemporal patterns of input from T2 and T3 confer discrimination of object motion from flicker in LC11 (Keles, 2020).
The identification of neurotransmitter type used by a neuron is important for the functional dissection of neuronal circuits. In the model organism Drosophila melanogaster, several methods for discerning the neurotransmitter systems are available. This study expanded the toolbox for the identification of cholinergic neurons by generating a new line FRT-STOP-FRT-VAChT::HA that is a conditional tagged knock-in of the VAChT gene in its endogenous locus. Importantly, in comparison to already available tools for the detection of cholinergic neurons, the FRT-STOP-FRT-VAChT::HA allele also allows for identification of the subcellular localization of the cholinergic presynaptic release sites in a cell-specific manner. The newly generated FRT-STOP-FRT-VAChT::HA line was used to characterize the Mi1 and Tm3 neurons in the fly visual system and found that VAChT is present in the axons of the both cell types, suggesting that Mi1 and Tm3 neurons provide cholinergic input to the elementary motion detectors, the T4 neurons (Pankova, 2017).
Detecting the orientation and movement of edges in a scene is critical to visually guided behaviors of many animals. What are the circuit algorithms that allow the brain to extract such behaviorally vital visual cues? Using in vivo two-photon calcium imaging in Drosophila, this study describes direction selective signals in the dendrites of T4 and T5 neurons, detectors of local motion. This circuit performs selective amplification of local light inputs, an observation that constrains motion detection models and confirms a core prediction of the Hassenstein-Reichardt correlator (HRC). These neurons are also orientation selective, responding strongly to static features that are orthogonal to their preferred axis of motion, a tuning property not predicted by the HRC. This coincident extraction of orientation and direction sharpens directional tuning through surround inhibition and reveals a striking parallel between visual processing in flies and vertebrate cortex, suggesting a universal strategy for motion processing (Fisher, 2015).
Each time a locomoting fly turns, the visual image sweeps over the retina and generates a motion stimulus. Classic behavioral experiments have suggested that flies use active neural-circuit mechanisms to suppress the perception of self-generated visual motion during intended turns. Direct electrophysiological evidence, however, has been lacking. This study found that visual neurons in Drosophila receive motor-related inputs during rapid flight turns. These inputs arrive with a sign and latency appropriate for suppressing each targeted cell's visual response to the turn. Precise measurements of behavioral and neuronal response latencies support the idea that motor-related inputs to optic flow-processing cells represent internal predictions of the expected visual drive induced by voluntary turns. Motor-related inputs to small object-selective visual neurons could reflect either proprioceptive feedback from the turn or internally generated signals. These results in Drosophila echo the suppression of visual perception during rapid eye movements in primates, demonstrating common functional principles of sensorimotor processing across phyla (Kim, 2015).
Humans scanning a visual scene show periods of stable gaze punctuated by rapid eye movements called saccades. During saccades, the visual image translates briskly over the retina and the nervous system employs mechanisms to suppress the perception of such self-induced motion stimuli so as to help perceive the outside environment as stationary. Primate saccades represent just one example in which sensorimotor processing must distinguish between self-generated sensory stimulation, also known as reafference, from externally generated stimulation, or exafference. In a few model systems, there is now even a developed understanding of how neuronal circuits distinguish exafference from reafference. For example, male crickets have an identified interneuron that activates during chirps and inhibits auditory neurons to prevent them from responding to the chirp, whereas weakly electric fish use circuitry in a cerebellum-like structure to subtract the predicted sensory input, resulting from the animal's own electric organ discharge from the incoming electrical sensory stream.
One common scenario in which the suppression of reafference is essential, but remains poorly understood, is in the inhibition of stability reflexes during voluntary locomotor turns. Consider a flying fly. Analogously with human eye movements, the fly punctuates periods of stable flight with rapid turns called body saccades. Between saccades, the fly employs an optokinetic reflex, also known as the optomotor response, to help maintain stable flight. In this reflex, wide-field visual motion, say, to the right, is interpreted as being caused by an erroneous turn of the fly to the left (perhaps caused by a gust of wind or noise in the flight motor) and a corrective rightward turn is elicited. Although this reflex is important for stability, if it were always active, it would act against any intended change to the locomotor trajectory. This begs the question of how then do flies ever turn (Kim, 2015).
Guided by simple behavioral experiments, von Holst and Mittelstaedt famously postulated that with each motor command to initiate a voluntary locomotor turn, also known as an efference, flies send a copy of the command, an efference copy, to their visual system. This efference-copy signal was postulated to have the correct sign and magnitude for silencing the reafferent visual input caused by voluntary turns, thereby preventing the optomotor response from kicking in. Subsequent behavioral experiments have continued to argue that locomoting insects send efference copies to their visual system or make use of internal models in their visuomotor processing; however, electrophysiological evidence has been scarce (Kim, 2015).
This study found that Drosophila visual neurons received motor-related inputs during voluntary body saccades. These inputs were similar in magnitude, but opposite in sign, to the expected reafferent visual input caused by saccades. Responses of wide field-sensitive neurons in the visual lobe and a new class of small object-selective cells in the central brain were strongly suppressed during intended flight turns. Other visual cells, whose receptive-field properties were such that they should not respond to wide-field motion during saccades, were only mildly affected. These findings demonstrate cell type-tailored signals that are appropriate for silencing reafferent visual responses during voluntary locomotor turns in flies, as predicted by von Holst and Mittelstaedt 65 years ago (Kim, 2015).
Drosophila visual neurons receive both visually driven and motor-related inputs with voluntary saccades. Motor-related inputs were tailored in sign and latency to effectively suppress neuronal responses to the reafferent visual motion resulting from saccades. Motor-related modulations are observed in cells that arborize in two late stages of visual processing, the lobula plate and the optic glomeruli, and these modulations are sufficiently strong to cancel, rather than just weakly modulate, visual signaling (Kim, 2015).
Von Holst and Mittelstaedt were careful to distinguish silencing of expected visual motion, the computation they believed to take place in fly brains, from complete blindness during voluntary turns. At the cellular level, some visual neurons, such as VS1-2 cells, did not receive prominent motor-related inputs (SRPs or saccade related potentials) during voluntary saccades, and other cells, such as horizontal system north (HSN) cells, were modified in a direction-selective manner that should have allowed them to still respond to unexpected (exafferent) visual motion. Thus, although flies suppress visual input during rapid flight turns, it is unlikely that they are completely blind supporting von Holst and Mittelstaedt's model. In many scenarios, it might make sense for motion-sensitive neurons to actually sense reafference; however, because HSN cells likely contribute to optomotor stability, eliminating reafference in their output signal during voluntary turns is sensible. It has been argued, based on behavioral experiments, that flying flies completely ignore certain visual motion during saccades or selectively ignore visual motion depending on whether this motion is in the expected or unexpected direction. The current data provide a plausible cellular explanation for these behavioral results (Kim, 2015).
Although the fly visual system, overall, shows cell type-specific and direction-selective silencing, it is not yet clear whether von Holst and Mittelstaedt's computation is instantiated in its full form in the Vm of single cells. To do so, each visual neuron would receive a saccade-associated input that is not just of the correct sign, but also of the correct time-varying magnitude to exactly cancel the expected visual drive associated with each voluntary turn. Activating such a negative-image input would require that fly brains instantiate a forward model to predict the visual drive that each neuron will experience from a given saccade. In realizing such a model, flies should scale their silencing signals by an internal estimate of the velocity time course of the upcoming saccade (using a so-called inverse model), as well as by how strongly the current visual environment (for example, a forest or a fog) is expected to drive each visual neuron during each saccade. If flies make use of internal models of this sort, it will be important to determine how they are implemented in the nervous system and how widespread their influence is on sensory processing and behavior (Kim, 2015).
At face value, the fact that SRPs grew in magnitude when flies generated saccades in the context of a preferred steady-state stimulus compared with a uniformly lit screen and the fact that this growth of SRP magnitude was quantitatively matched to the level of ongoing visual drive supports the forward model idea. Note, however, that if saccade-related inputs activate a consistent membrane conductance, independent of visual context, for saccades of a certain direction and size, then saccade-related potentials will naturally grow in size as the cell's Vm moves further away from the reversal potential associated with that conductance. Given that ongoing visual stimulation causes cells to depolarize or hyperpolarize from the resting Vm, SRPs may grow in magnitude as a result of this reason alone. Future work will be needed to differentiate this simple biophysical explanation for why SRPs grow in magnitude during ongoing visual drive, which may represent a rudimentary implementation of a forward model, from a more sophisticated process in which the strength of motor-related conductances in visual neurons are actively scaled on the basis of the structure of the visual environment. Notably, HS cells appear to show similar visual responses to moving natural scenes over a wide range of contrast levels and arrangements of local features40. This fact may allow an efference copy system to get away with injecting silencing signals that have a consistent time-varying profile for saccades of a given direction and magnitude, independent of the structure of the visual environment. Flies may also continuously calibrate the strength of their efference copy signal based on the difference between predicted and experienced sensory feedback (Kim, 2015).
Regardless of whether motor-related inputs are scaled by the structure of the visual environment, an important associated question is whether, in a fixed visual environment, the motor-related inputs are scaled by the magnitude and duration of each saccade. Because torque is not measured in current platform, even if L-R WBA acts as a decent proxy, and because tethered-flight saccades are known to have altered dynamics relative to free-flight saccades, it is difficult to provide a definitive answer to this question with current methods (Kim, 2015).
Classically, only neurons that contribute to optomotor stability, such as HSN cells, should be silenced during voluntary turns. This study found that small object-selective optic-glomeruli interneurons (OGINs) were also silenced. Why might this be? Object-selective OGINs may contribute to behaviors such as small object avoidance during flight or tracking of conspecifics during Drosophila courtship. Distinguishing reafference from exafference would seem to be critical for such object-orienting behaviors to prevent behavioral responses to object motion on the retina caused by the fly's own movements. When a locomoting fly turns in a cluttered visual environment, the image of the entire cluttered panorama, not just that of the small object, translates globally on the retina, and such a global stimulus, simulated by the grating in receptive field mapping experiments, will not excite these OGINs. Thus, the native stimulus selectivity of these cells already helps to distinguish exafferent from reafferent object motion. However, if a single object were situated on a sparse background, such as a spider against a homogenous blue sky, OGINs may very well respond to the reafferent motion of such an object during a locomotor turn and the motor-related silencing mechanism that is described would abrogate this deleterious sensory response (Kim, 2015).
Mechanistically, SRPs in OGINs are consistent with an inhibitory input that arrives on each saccade, either directly to these cells or to upstream neurons, to reduce feedforward excitatory drive. Because OGINs are spiking cells (although their spikes are often very small when measured at the soma), the role of SRPs in these neurons may be to simply eliminate spike output during saccades rather than to activate a precisely time-varying negative-image input. Indeed, SRPs in OGINs lasted ~400-500 ms, which is longer than the time course of the expected reafference during a typical saccade. If OGIN SRPs are the result of mechanosensory feedback associated with saccades, this feedback signal would be expected to be prolonged in tethered flight, as tethered-flight saccades lasted longer (~300-500 ms) than free-flight saccades (~50-150 ms). By contrast, HSN cells are non-spiking neurons that signal both with hyperpolarizations and depolarizations of Vm. As such, HSN cells received both depolarizing and hyperpolarizing SRPs, whose duration (~150-200 ms) was more closely matched to the expected reafference from a typical saccade. SRPs in HSN cells may serve a function closer to that of a negative image of the expected visual drive (Kim, 2015).
The average SRP in HSN cells begins ~30 ms before the wings initiate a saccade, arguing for an internal (rather than sensory feedback) origin for these signals. However, if the sole function of motor-related inputs to HSN cells were to silence visual reafference, one might expect the efference-copy signal to kick in only after the body starts turning, once reafferent visual input is arriving at the cell. One intriguing possibility is that the early component of the motor-related input to HSN cells might help to actually drive the voluntary turn by injecting a small pre-charging signal into the optomotor reflex system, hijacking its natural coupling to the neck and flight motor systems. The sign of this early component is consistent with this possibility, although a rigorous test will require a specific manipulation of the SRPs to the optomotor system, which likely includes many more cells than just HS cells. Flies stabilize flight not just with vision, but also with mechanosensory inputs from their modified hindwings, called halteres, and a similar pre-charging idea was postulated to occur in the haltere stability system during turns, a hypothesis that should now be revisited (Kim, 2015).
Dynamic modulations of visual signaling have been studied primarily during saccadic eye movements in primates. The current results open the door to studying cellular mechanisms for similar processes in Drosophila. For vision research, flies have already offered key insights into state-dependent sensory processing and the circuit basis for direction selectivity. Drosophila may now help in understanding how brains build forward models and how they use these models to modify sensory processing. The tiny fly brain may not perform these tasks in exactly the same manner as the primate brain. However, the advanced genetic and physiological tools in Drosophila should allow for a detailed cellular- and circuit-level description of how the fly brain models and predicts the outside world. This description could yield insight on how similar predictive processes are implemented in all brains (Kim, 2015).
Animals typically combine inertial and visual information to stabilize their gaze against confounding self-generated visual motion, and to maintain a level gaze when the body is perturbed by external forces. In vertebrates, an inner ear vestibular system provides information about body rotations and accelerations, but gaze stabilization is less understood in insects, which lack a vestibular organ. In flies, the halteres, reduced hindwings imbued with hundreds of mechanosensory cells, sense inertial forces and provide input to neck motoneurons that control gaze. These neck motoneurons also receive input from the visual system. Head movement responses to visual motion and physical rotations of the body have been measured independently, but how inertial information might influence gaze responses to visual motion has not been fully explored. In this study, the head movement responses to visual motion were measured in intact and haltere-ablated tethered flies to explore the haltere's role in modulating visually-guided head movements in the absence of rotation. It is noted that visually-guided head movements occur only during flight. Although halteres are not necessary for head movements, the amplitude of the response is smaller in haltereless flies at higher speeds of visual motion. This modulation occurred in the absence of rotational body movements, demonstrating that the inertial forces associated with straight tethered flight are important for gaze-control behavior. The cross-modal influence of halteres on the fly's responses to fast visual motion indicates that the haltere's role in gaze stabilization extends beyond its canonical function as a sensor of angular rotations of the thorax (Mureli, 2017).
When confronted with a large-field stimulus rotating around the vertical body axis, flies display a following behavior called 'optomotor response.' As neural control elements, the large tangential horizontal system (HS) cells of the lobula plate have been prime candidates for long. This study applied optogenetic stimulation of HS cells to evaluate their behavioral role in Drosophila. To minimize interference of the optical activation of channelrhodopsin-2 with the visual perception of the flies, a bistable variant was used called ChR2-C128S. By applying pulses of blue and yellow light, it was first demonstrated electrophysiologically that lobula plate tangential cells can be activated and deactivated repeatedly with no evident change in depolarization strength over trials. It was next shown that selective optogenetic activation of HS cells elicits robust yaw head movements and yaw turning responses in fixed and tethered flying flies, respectively (Haikala, 2013).
To safely navigate through space particularly fast-moving animals face the challenging need of constantly integrating changing information about the environment and self-motion. Flies are capable of impressively robust, precise, and fast flight maneuvers that largely depend on visual and mechanosensory information (Haikala, 2013).
Depending on the species, a bilateral set of 20-60 large neurons, termed lobula plate tangential cells (LPTCs), located in the posterior part of the optic lobes, are considered critical for vision-based estimation of self-motion. LPTCs are individually identifiable and have large receptive fields, sometimes covering more than one hemisphere of visual space. Their most distinguishing characteristic is their directional tuning to visual wide-field motion, termed optic flow (Haikala, 2013).
Among the LPTCs, three horizontal system (HS) cells sensitive to horizontal wide-field motion, such as occurring during rotation about the vertical body axis, have long been prime candidates to control compensatory yaw head movements and body turns of the fly. This suggestive notion rests on the following observations: (1) Mutant fruit flies in which LPTCs are missing or defective show a strong reduction in their optomotor response. (2) Cutting HS cell axons in flies or laser ablation of HS/vertical system precursor cells in larvae significantly affects the optomotor response of adult flies. (3) Extracellular electrical stimulation of the lobula plate region where HS cells are located elicits yaw turning responses. While all of these findings support the idea that HS cells control optomotor responses, it has not been conclusively shown that activation of HS cells is sufficient to evoke yaw optomotor behavior (Haikala, 2013).
This study analyzed the role of HS cells for head movements and flight-turning responses by stimulating them optogenetically in Drosophila. When a fly is visually stimulated by a pattern rotating around the vertical axis, its HS cells depolarize on the side where motion progresses from front to back, and the fly displays syndirectional head movements and flight turns. If HS cells control both these optomotor responses, unilateral optogenetic depolarization of HS cells alone should mimic these behaviors. To avoid behavioral artifacts due to activation of photoreceptors during optical stimulation of HS cells, this study generated flies expressing the bistable channelrhodopsin-2 variant ChR2-C128S, a depolarizing cation channel that can be switched between an open and closed configuration with brief light pulses of different wavelengths. Expressing ChR2-C128S in HS cells via a selective driver line enabled control of the cells in a similar way as visual motion does. When HS cells were activated in one brain hemisphere, flies turned their heads toward the stimulated side. Likewise, tethered flying flies displayed unilateral wing beat changes indicative of flight-turning responses in the same direction. The results strongly suggest that HS cell activity alone is sufficient to elicit yaw head movements and flight-turning responses in Drosophila (Haikala, 2013).
In recent years, optogenetic methods have been successfully applied in a number of studies, involving different species such as Caenorhabditis elegans, zebrafish, and mice. Nonetheless, its use in Drosophila has been rather limited, in particular for analyzing visual circuits, perhaps in part because light delivery to activate light-sensitive ion channels inevitably leads to direct stimulation of the photoreceptors and thus interferes with the visual processing to be studied. To avoid this problem, optogenetic stimulation has been performed in blind flies. This strategy, however, raises potential concerns as to whether the circuits are compromised by visual deprivation. Moreover, it precludes the simultaneous visual probing of the system. In this respect, the development of switchable, bistable channelrhodopsin-2 variants offers a powerful solution. As this study has shown, one of these variants (ChR2-C128S) allows for prolonged and repeated excitation of large motion-sensitive neurons in the Drosophila visual system by delivery of pulses of light with the appropriate wavelength (Haikala, 2013).
While previous work in Drosophila and Calliphora has already put forward the idea that HS cells control yaw optomotor behavior, well in agreement with their visual response properties, only one study actually tested their requirement directly by lesioning HS cell axons unilaterally and recording the yaw torque response to visual motion. The resulting behavioral phenotype nicely supports the assumed role of HS cells. Nonetheless, it was acknowledged that axons of other lobula plate output neurons could have also been severed. The current complementary experiments indicate that unilateral optogenetic stimulation of the three HS cells is sufficient to evoke both yaw head movements as well as flight-turning responses. While light pulses were visually perceived by flies and led to transient physiological and behavioral artifacts, consistent long-lasting responses were only observed in animals expressing functional ChR2-C128S, in agreement with the bistable nature of this channel. The direction of the behavioral responses toward the stimulated side can be readily interpreted such that the flies attempt to counteract perceived unintended yaw body rotations. The weaker response levels in visually intact flies compared to actual visually evoked behavior and robust optogenetic responses in blind flies could be explained by unspecific prolonged perturbances of visual circuits by blue light pulses that potentially attenuate optogenetic effects in visually intact flies. Alternatively, stationary visual signals from the inactive arena that provide stable reference points might antagonize the effect of HS cell activation. It should also be noted that visual stimuli were presented to both eyes, leading to hyperpolarization of HS cells in one hemisphere and depolarization of HS cells in the other hemisphere. In contrast, optogenetic stimulation of HS cells involved the cells within only one hemisphere. The information provided by unilateral HS cell activation is partly ambiguous and might signal yaw rotation, but also sideway or forward translation. In fact, HS cells have been shown to encode translating optic flow provided by both eyes during intersaccadic flight intervals, which may serve to extract depth information. Furthermore, HS cells might be functionally specialized, and processing the outputs in various combinations could be used to recover different aspects of self-motion-induced optic flow. For instance, HSN and HSE with dorsal and equatorial receptive fields, respectively, receive contralateral input tuned to back-to-front motion, rendering them sensitive to sideward translation in addition to yaw rotation, particularly so when activity is bilaterally subtracted. In contrast, HSS lacks contralateral input, and summating HSS activity from both hemispheres in higher processing centers might be used to recover parameters associated with forward translation. Although it is not possible to resolve such functional specializations, the fact that unilateral stimulation of all three HS cells produces clear turning responses toward the stimulated side supports the notion that the HSE and HSN cells, and potentially the HSS cell, are important for controlling compensatory yaw rotations of head and body. Nonetheless, it is well known that HS cells are part of a larger network of tangential cells in the lobula plate. There is no doubt that under natural conditions, more dynamic and finer-grained activity patterns in HS and perhaps other neurons than was possible to induce optogenetically are required to reduce ambiguities related to horizontal optic flow and to guide flies safely through their environment. It is also interesting to note that neither cessation of moving patterns nor ChR2-C128S channel closure by longer-wavelength light, both leading to repolarization of HS cells, immediately reestablishes baseline head angle and wing beat amplitudes. This might indicate that these behavioral parameters require active stimulation along the opposite direction and the concomitant depolarization of the contralateral HS cells to return to baseline (Haikala, 2013).
The results contribute to an emerging picture of how visually guided behavioral patterns are generated in flies. Certain visual cues contained in the raw image sequence such as direction of motion are extracted by intricate parallel circuits of the optic lobes and integrated and conveyed toward appropriate motor circuits via a cohort of wide-field projection neurons, each of which is endowed with a unique visual tuning profile. As an analogous example to the HS system, FOMA-1 cells of the lobula complex are tuned to looming stimuli and consequently elicit escape responses when activated (de Vries, 2012). In blow flies, part of the pathway downstream of the HS cells has been identified for the neck motor system via a motor neuron of the ventral cervical nerve, constituting a straight-forward link between unilateral HS cell activation and neck muscle contraction. In contrast, much less is known about the circuits underlying turning behavior during flight. Unilateral change of wing beat amplitude is controlled by small steering muscles, some of which have been shown to respond to horizontal wide-field motion in a direction-selective way. Such muscles are therefore probable downstream targets of the HS system, but the neural elements that convey the visual information to them remain to be functionally characterized (Haikala, 2013).
The fact that HS cell responses are remarkably robust while optomotor behavior can be quite variable as explicitly shown, for instance, for Calliphora illustrates that visual reflexes are not unimodal all-or-nothing events, but rather are subject to multimodal modification as well as to central gating. This might reflect the ethological need to conditionally modify or disable optomotor compensation by conflicting visual cues, other sensory modalities, or during certain internally generated flight maneuvers like saccades. Identification of further constituents of the optomotor pathways would provide opportunities to elucidate the neural basis for such multimodal stimulus integration, modulation, and gating of the optomotor response. To this end, flies expressing bistable channelrhodopsin-2 in combination with specific driver lines can help to establish the participation of candidate neurons. Using channelrhodopsin-2 variants with longer time constants than ChR2-C128S, the consequence of activating certain neurons could perhaps even be tested in freely behaving, unrestrained animals (Haikala, 2013).
The extraction of directional motion information from changing retinal images is one of the earliest and most important processing steps in any visual system. In the fly optic lobe, two parallel processing streams have been anatomically described, leading from two first-order interneurons, L1 and L2, via T4 and T5 cells onto large, wide-field motion-sensitive interneurons of the lobula plate. Therefore, T4 and T5 cells are thought to have a pivotal role in motion processing; however, owing to their small size, it is difficult to obtain electrical recordings of T4 and T5 cells, leaving their visual response properties largely unknown. This problem was circumvented by means of optical recording from these cells in Drosophila, using the genetically encoded calcium indicator GCaMP5. This study finds that specific subpopulations of T4 and T5 cells are directionally tuned to one of the four cardinal directions; that is, front-to-back, back-to-front, upwards and downwards. Depending on their preferred direction, T4 and T5 cells terminate in specific sublayers of the lobula plate. T4 and T5 functionally segregate with respect to contrast polarity: whereas T4 cells selectively respond to moving brightness increments (ON edges), T5 cells only respond to moving brightness decrements (OFF edges). When the output from T4 or T5 cells is blocked, the responses of postsynaptic lobula plate neurons to moving ON (T4 block) or OFF edges (T5 block) are selectively compromised. The same effects are seen in turning responses of tethered walking flies. Thus, starting with L1 and L2, the visual input is split into separate ON and OFF pathways, and motion along all four cardinal directions is computed separately within each pathway. The output of these eight different motion detectors is then sorted such that ON (T4) and OFF (T5) motion detectors with the same directional tuning converge in the same layer of the lobula plate, jointly providing the input to downstream circuits and motion-driven behaviours (Maisak, 2013).
Most of the neurons in the fly brain are dedicated to image processing. The respective part of the head ganglion, called the optic lobe, consists of several layers of neuropile called lamina, medulla, lobula and lobula plate, all built from repetitive columns arranged in a retinotopic way. Each column houses a set of identified neurons that, on the basis of Golgi staining, have been described anatomically, first by Santiago Ramon y Cajal (see Cajal, 1915), in great detail. Owing to their small size, however, most of these columnar neurons have never been recorded from electrophysiologically. Therefore, their specific functional role in visual processing is still largely unknown. This fact is contrasted by rather detailed functional models about visual processing inferred from behavioural studies and recordings from the large, electrophysiologically accessible output neurons of the fly lobula plate (tangential cells). As the most prominent example of such models, the Reichardt detector derives directional motion information from primary sensory signals by multiplying the output from adjacent photoreceptors after asymmetric temporal filtering. This model makes a number of rather counter-intuitive predictions all of which have been confirmed experimentally. Yet, the neurons corresponding to most of the circuit elements of the Reichardt detector have not been identified so far. This study focused on a set of neurons called T4 and T5 cells which, on the basis of circumstantial evidence, have long been speculated to be involved in motion detection. However, it is unclear to what extent T4 and T5 cells are directionally selective or whether direction selectivity is computed or enhanced within the dendrites of the tangential cells. Another important question concerns the functional separation between T4 and T5 cells; that is, whether they carry equivalent signals, maybe one being excitatory and the other inhibitory on the tangential cells, or whether they segregate into directional- and non-directional pathways or into separate ON- and OFF-motion channels (Maisak, 2013).
To answer these questions, Gal4-driver lines specific for T4 and T5 cells were combined with GCaMP5 (Akerboom, 2012) and the visual response properties were optically recorded using two-photon fluorescence microscopy. In a first series of experiments, a driver line labelling both T4 and T5 cells was used. A confocal image revealed clear labelling in the medulla (T4 cell dendrites), in the lobula (T5 cell dendrites), as well as in four distinct layers of the lobula plate, representing the terminal arborizations of the four subpopulations of both T4 and T5 cells. These four layers of the lobula plate can also be seen in the two-photon microscope when the calcium indicator GCaMP5 is expressed. After stimulation of the fly with grating motion along four cardinal directions (front-to-back, back-to-front, upwards and downwards), activity is confined to mostly one of the four layers, depending on the direction in which the grating is moving. The outcome of all four stimulus conditions can be combined into a single image by assigning a particular colour to each pixel depending on the stimulus direction to which it responded most strongly. From these experiments it is clear that the four subpopulations of T4 and T5 cells produce selective calcium signals depending on the stimulus direction, in agreement with previous deoxyglucose labelling. Sudden changes of the overall luminance evokes no responses in any of the layers. However, gratings flickering in counter-phase lead to layer-specific responses, depending on the orientation of the grating (Maisak, 2013).
The retinotopic arrangement of this input to the lobula plate is demonstrated by experiments where a dark edge was moved within a small area of the visual field only. Depending on the position of this area, activity of T4 and T5 cells is confined to different positions within the lobula plate. Consequently, when moving a bright vertical edge horizontally from back to front, activity of T4 and T5 cells is elicited sequentially in layer 2 of the lobula plate. These two experiments also demonstrate that T4 and T5 cells indeed signal motion locally. Next, the question of where direction selectivity of T4 and T5 cells arises was investigated; that is, whether it is already present in the dendrite, or whether it is generated by synaptic interactions within the lobula plate. This question is hard to answer, as the dendrites of both T4 and T5 cells form a dense mesh within the proximal layer of the medulla (T4) and the lobula (T5), respectively. However, signals within the inner chiasm where individual processes of T4 and T5 cells can be resolved in some preparations show a clear selectivity for motion in one over the other directions. Such signals are as directionally selective as the ones measured within the lobula plate, demonstrating that the signals delivered from the dendrites of T4 and T5 cells are already directionally selective (Maisak, 2013).
To assess the particular contribution of T4 and T5 cells to the signals observed in the above experiments, driver lines specific for T4 and T5 cells, respectively, were used. Applying the same stimulus protocol and data evaluation as described above, identical results were obtained as before for both the T4- as well as the T5-specific driver line. It is concluded that T4 and T5 cells each provide directionally selective signals to the lobula plate. Thus, both T4 and T5 cells can be grouped, according to their preferred direction, into four subclasses covering all four cardinal directions, reminiscent of ON–OFF ganglion cells of the rabbit retina (Maisak, 2013).
Next whether T4 cells respond differently to T5 cells was addressed. To answer this question, moving edges instead of gratings were used with either positive (ON edge, brightness increment) or negative (OFF edge, brightness decrement) contrast polarity as visual stimuli. It was found that T4 cells strongly responded to moving ON edges, but showed little or no response to moving OFF edges. This is true for T4 cells terminating in each of the four layers. The opposite was found for T5 cells. T5 cells selectively responded to moving OFF edges and mostly failed to respond to moving ON edges. Again, this was found for T5 cells in each of the four layers. Next, whether there are any other differences in the response properties between T4 and T5 cells was addressed by testing the velocity tuning of both cell populations by means of stimulating flies with grating motion along the horizontal axis from the front to the back at various velocities covering two orders of magnitude. T4 cells revealed a maximum response at a stimulus velocity of 30° s-1, corresponding to a temporal frequency of 1 Hz. T5 cell responses showed a similar dependency on stimulus velocity, again with a peak at a temporal frequency of 1 Hz. Thus, there is no obvious difference in the velocity tuning between T4 and T5 cells. As another possibility, T4 cells might functionally differ from T5 cells with respect to their directional tuning width. To test this, flies were stimulated with gratings moving into 12 different directions, and the relative change of fluorescence was evaluated in all four layers of the lobula plate. Using the T4-specific driver line, an approximate half width of 60-90° of the tuning curve was found, with the peak responses in each layer shifted by 90°. No decrease of calcium was detectable for grating motion opposite to the preferred direction of the respective layer. When the experiments were repeated using the T5-specific driver line, a similar dependence of the relative change of fluorescence was found on the stimulus direction. It is concluded that T4 cells have the same velocity and orientation tuning as T5 cells. The only functional difference that was detected remains their selectivity for contrast polarity (Maisak, 2013).
The finding about the different preference of T4 and T5 cells for the polarity of a moving contrast makes the strong prediction that selective blockade of T4 or T5 cells should selectively compromise the responses of downstream lobula plate tangential cells to either ON or OFF edges. To test this prediction, the output of either T4 or T5 cells was blocked via expression of the light chain of tetanus toxin, and the responses of tangential cells via somatic whole-cell patch was recorded to moving ON and OFF edges. In response to moving ON edges, strong and reliable directional responses were observed in all control flies. However, T4-block flies showed a strongly reduced response to ON edges, whereas the responses of T5-block flies were at the level of control flies. When moving OFF edges were used, control flies again responded with a large amplitude. However, the responses of T4-block flies were at the level of control flies, whereas the responses of T5-block flies were strongly reduced. These findings are reminiscent of the phenotypes obtained from blocking lamina cells L1 and L2 (Joesch, 2010) and demonstrate that T4 and T5 cells are indeed the motion-coding intermediaries for these contrast polarities on their way to the tangential cells of the lobula plate. Whether the residual responses to ON edges in T4-block flies and to OFF edges in T5-block flies are due to an incomplete signal separation between the two pathways or due to an incomplete genetic block in both fly lines is currently unclear (Maisak, 2013).
To address the question of whether T4 and T5 cells are the only motion detectors of the fly visual system, or whether they represent one cell class, in parallel to other motion-sensitive elements, tethered flies walking on an air-suspended sphere were used, and and they were stimulated by ON and OFF edges moving in opposite directions. As in the previous experiments, T4 and T5 cells were blocked specifically by selective expression of the light chain of tetanus toxin. During balanced motion, control flies did not show significant turning responses to either side. T4-block flies, however, strongly followed the direction of the moving OFF edges, whereas T5-block flies followed the direction of the moving ON edges. In summary, the selective preference of T4-block flies for OFF edges and of T5-block flies for ON edges not only corroborates the findings about the selective preference of T4 and T5 cells for different contrast polarities, but also demonstrates that the signals of T4 and T5 cells are indeed the major, if not exclusive, inputs to downstream circuits and motion-driven behaviours (Maisak, 2013).
Almost a hundred years after T4 and T5 cells have been anatomically described, this study reports their functional properties in a systematic way. Using calcium as a proxy for membrane voltage, this study found that both T4 and T5 cells respond to visual motion in a directionally selective manner and provide these signals to each of the four layers of the lobula plate, depending on their preferred direction. Both cell types show identical velocity and orientation tuning which matches the one of the tangential cells. The strong direction selectivity of both T4 and T5 cells is unexpected, as previous studies had concluded that the high degree of direction selectivity of tangential cells is due to a push–pull configuration of weakly directional input with opposite preferred direction. Furthermore, as the preferred direction of T4 and T5 cells matches the preferred direction of the tangential cells branching within corresponding layers, it is currently unclear which neurons are responsible for the null-direction response of the tangential cells. As for the functional separation between T4 and T5 cells, this study found that T4 cells selectively respond to brightness increments, whereas T5 cells exclusively respond to moving brightness decrements. Interestingly, parallel ON and OFF motion pathways had been previously postulated on the basis of selective silencing of lamina neurons L1 and L2 (Joesch, 2010). Studies using apparent motion stimuli to probe the underlying computational structure arrived at controversial conclusions: whereas some studies concluded that there was a separate handling of ON and OFF events by motion detectors, others did not favour such a strict separation. The present study directly demonstrates the existence of separate ON and OFF motion detectors, as represented by T4 and T5 cells, respectively. Furthermore, the results anatomically confine the essential processing steps of elementary motion detection -- that is, asymmetric temporal filtering and nonlinear interaction -- to the neuropile between the axon terminals of lamina neurons L1 and L2 (Joesch, 2013) and the dendrites of directionally selective T4 and T5 cells. The dendrites of T4 and T5 cells might well be the place where signals from neighbouring columns interact in a nonlinear way, similar to the dendrites of starburst amacrine cells of the vertebrate retina (Maisak, 2013).
Neuronal computation underlying detection of visual motion has been studied for more than a half-century. In Drosophila, direction-selective T4/T5 neurons show supralinear signal amplification in response to stimuli moving in their preferred direction, in agreement with the prediction made by the Hassenstein-Reichardt detector. Nevertheless, the molecular mechanism explaining how the Hassenstein-Reichardt model is implemented in T4/T5 cells has not been identified yet. The present study utilized cell type-specific transcriptome profiling with RNA-seq to obtain a complete gene expression profile of T4/T5 neurons. The expression was analyzed of genes that affect neuronal computational properties and can underlie the molecular implementation of the core features of the Hassenstein-Reichardt model to the dendrites of T4/T5 neurons. Furthermore, the acquired RNA-seq data was used to examine the neurotransmitter system used by T4/T5 neurons. Surprisingly, co-expression of the cholinergic markers and the vesicular GABA transporter was observed in T4/T5 neurons. Previously undetected expression of vesicular GABA transporter was documented in T4/T5 cells using VGAT-LexA knock-in line. The provided gene expression dataset can serve as a useful source for studying the properties of direction-selective T4/T5 neurons on the molecular level (Pankova, 1016).
How neurons become sensitive to the direction of visual motion represents a classic example of neural computation. Two alternative mechanisms have been discussed in the literature so far: preferred direction enhancement, by which responses are amplified when stimuli move along the preferred direction of the cell, and null direction suppression, where one signal inhibits the response to the subsequent one when stimuli move along the opposite, i.e. null direction. Along the processing chain in the Drosophila optic lobe, directional responses first appear in T4 and T5 cells. Visually stimulating sequences of individual columns in the optic lobe with a telescope while recording from single T4 neurons, this study found both mechanisms at work implemented in different sub-regions of the receptive field. This finding explains the high degree of directional selectivity found already in the fly's primary motion-sensing neurons and marks an important step in understanding of elementary motion detection (Haag, 2016).
In the visual motion pathways contained within the fly's optic lobe, two cell types-T4 and T5-are the first known relay neurons to signal small-field direction-selective motion responses (see Graphical abstract). These cells then feed into large tangential cells that signal wide-field motion. Recent studies have identified two types of columnar neurons in the second neuropil, or medulla, that relay input to T4 from L1, the ON-channel neuron in the first neuropil, or lamina, thus providing a candidate substrate for the elementary motion detector (EMD). Interneurons relaying the OFF channel from L1's partner, L2, to T5 are so far not known, however. This study report that multiple types of transmedulla (Tm) neurons provide unexpectedly complex inputs to T5 at their terminals in the third neuropil, or lobula. From the L2 pathway, single-column input comes from Tm1 and Tm2 and multiple-column input from Tm4 cells. Additional input to T5 comes from Tm9, the medulla target of a third lamina interneuron, L3, providing a candidate substrate for L3's combinatorial action with L2. Most numerous, Tm2 and Tm9's input synapses are spatially segregated on T5's dendritic arbor, providing candidate anatomical substrates for the two arms of a T5 EMD circuit; Tm1 and Tm2 provide a second. Transcript profiling indicates that T5 expresses both nicotinic and muscarinic cholinoceptors, qualifying T5 to receive cholinergic inputs from Tm9 and Tm2, which both express choline acetyltransferase (ChAT). It is hypothesized that T5 computes small-field motion signals by integrating multiple cholinergic Tm inputs using nicotinic and muscarinic cholinoceptors (Shinomiya, 2014).
The L2 pathway has been identified as the substrate for detecting moving dark edges. L2's partner cell, L4, and its common medulla target, Tm2, are both essential components of this dark-edge pathway prior to the computation of directionality, which occurs first in the dendrites of T5. Tm2 and L4 cells both respond with a nondirectional increase in activity to moving dark (OFF) edges, and silencing of both either singly or in combination also abolishes the response to moving dark edges in downstream LPTCs. The anatomical receptive fields described in this report for the input terminals to T5 cell dendrites are compatible with Tm2's being wired as one of the two arms of an EMD circui and with Tm1, Tm9, or both as the other. The existence of two EMD circuits would require that the inputs from Tm1/Tm2 and Tm9/Tm2 have aligned vector angles, since each T5 receives both circuits but must respond to dark-edge motion in only one of the four cardinal directions. This alignment is clear for three of the T5 cells plotted in detail, but less so for T5-08. Clearly, additional detailed plots of T5 anatomical receptive fields are needed (Shinomiya, 2014).
Even though this scheme is still highly speculative, nonlinear interaction between two input arms of the EMD circuits is the computational basis of local motion-detection models, while specific computational models favor different types of interactions: either multiplication or facilitation for the Hassenstein-Reichardt model or inhibition for the Barlow-Levick model (see Motion sensing in vision) (Shinomiya, 2014).
A parallel may be seen in the vertebrate retina. There, turtle B10 bipolar cells use ionotropic and metabotropic glutamate receptors to signal, respectively, L and M cone inputs, a dual deployment that has been suggested to form the basis for B10 red-ON, blue/green-OFF color opponency. The expression of both nicotinic and muscarinic cholinoceptors in T5 may provide a similar means to integrate multiple cholinergic Tm inputs and so compute small-field motion signals. Vertebrate muscarinic receptors are coupled to G proteins and various downstream signaling pathways to regulate a broad spectrum of cellular functions, including neuronal excitability. In Drosophila, agonist activation of mAchR-A acts via the IP3 pathway to increase calcium release from internal stores, which elsewhere is reported in turn to activate high-conductance calcium-dependent potassium (BK) channels (the Slowpoke channel), leading to membrane hyperpolarization. It was therefore interesting that, using single-cell RT-PCR, slowpoke transcripts were detected in both T4 and T5, consistent with a previous immunohistochemical study that Slowpoke is expressed in the optic lobe, including the lobula. It is proposed that these two postsynaptic events, mAchR-A-mediated increased intracellular calcium and mAchR activation of Slowpoke channels, occur at postsynaptic sites activated by different inputs distributed over the T5 dendritic arbor. Given the relative temporal inflexibility of an excitatory nicotinic cholinoceptor synaptic response and the fact that their anatomy qualifies Tm2/Tm9 and Tm2/Tm1 input pairs as two independent pairs of EMD input arms to T5 that share Tm2, it seems most reasonable that T5 uses the nicotinic receptor inputs for fast excitation from Tm2 (as the instantaneous signal) and compares this with a slow inhibitory input from either Tm9 or Tm1 (as the delayed signal). This arrangement could allow T5 to inherit motion information with two temporal characteristics provided that Tm1 and Tm9 have their own time delay. The coupling of the inhibitory inputs to secondary messenger system also provides a potential mechanism to adapt the temporal delay filter. In addition, the activation of Drosophila mAchR-A in cultured cells also induces a secondary calcium influx, potentially originating from an extracellular calcium pool, while the activation of vertebrate muscarinic receptors has been shown to inhibit potassium channels, including those of the Kv7 (KCNQ/M) type, and to lower the excitability threshold. It is therefore possible that an interaction between the nicotinic and muscarinic inputs provides some form of multiplication, as suggested in the Hassenstein-Reichardt model. Further definition of the roles of the Tm1, Tm2, and Tm9 input cells as input arms to different EMD circuits and determination of the synaptic mechanisms for detecting moving dark edges must obviously await additional genetic silencing experiments and electrophysiological recordings from all these cells, which this study now clearly identifies as synaptic inputs to T5 in Drosophila (Shinomiya, 2014).
In the fly optic lobe, approximately 800 highly stereotypical columnar microcircuits are arranged retinotopically to process visual information. Differences in cellular composition and synaptic connectivity within functionally specialized columns remain largely unknown. This study describes the cellular and synaptic architecture in medulla columns located downstream of photoreceptors in the dorsal rim area (DRA), where linearly polarized skylight is detected for guiding orientation responses. Only in DRA medulla columns both R7 and R8 photoreceptors target to the bona fide R7 target layer where they form connections with previously uncharacterized, modality-specific Dm neurons: two morphologically distinct DRA-specific cell types (termed Dm-DRA1 and Dm-DRA2) stratify in separate sublayers and exclusively contact polarization-sensitive DRA inputs, while avoiding overlaps with color-sensitive Dm8 cells. Using the activity-dependent GRASP and trans-Tango techniques, it was confirmed that DRA R7 cells are synaptically connected to Dm-DRA1, whereas DRA R8 form synapses with Dm-DRA2. Finally, using live imaging of ingrowing pupal photoreceptor axons, it was shown that DRA R7 and R8 termini reach layer M6 sequentially, thus separating the establishment of different synaptic connectivity in time. It is proposed that a duplication of R7-->Dm circuitry in DRA ommatidia serves as an ideal adaptation for detecting linearly polarized skylight using orthogonal e-vector analyzers (Sancer, 2019a).
Natural scenes contain complex visual cues with specific features, including color, motion, flicker and position. It is critical to understand how different visual features are processed at the early stages of visual perception to elicit appropriate cellular responses, and even behavioral output. The visual orientation response induced by flickering stripes in a novel behavioral paradigm in Drosophila melanogaster. This study found that free walking flies exhibited bandpass orientation response to flickering stripes of different frequencies. The most sensitive frequency spectrum was confined to low frequencies of 2-4 Hz. Through genetic silencing, this study showed that lamina L1 and L2 neurons, which receive visual inputs from R1-R6 neurons, were the main components in mediating flicker-induced orientation behavior. Moreover, specific blocking of different types of lamina feedback neurons Lawf1, Lawf2, C2, C3 and T1 modulated orientation responses to flickering stripes of particular frequencies, suggesting that bandpass orientation response was generated through cooperative modulation of lamina feedback neurons. Furthermore, this study found that lamina feedback neurons Lawf1 were glutamatergic. Thermal activation of Lawf1 neurons could suppress neural activities in L1 and L2 neurons, which could be blocked by the glutamate-gated chloride channel inhibitor picrotoxin (PTX). In summary, lamina monopolar neurons L1 and L2 are the primary components in mediating flicker-induced orientation response. Meanwhile, lamina feedback neurons cooperatively modulate the orientation response in a frequency-dependent way, which might be achieved through modulating neural activities of L1 and L2 neurons (Yuan, 2020).
The optic lobes of the fruit fly Drosophila melanogaster form a highly wired neural network composed of roughly 130.000 neurons of more than 80 different types. This study use the optic lobe of the fruit fly as a paradigm to understand how neuroblasts generate multiple neuron types. A large-scale lineage bioinformatics analysis was performed using the graph theory. A large collection of cell clones were generated that genetically label the progeny of neuroblasts, and a database was built to draw graphs showing the lineage relationships between cell types. By establishing biological criteria that measures the strength of the neuronal relationships and applying community detection tools, eight clusters of neurons were identified. Each cluster contains different cell types that are posed to be the product of eight distinct classes of neuroblasts. Three of these clusters match the available lineage data, supporting the predictive value of the analysis. Finally, it was shown that the neuronal progeny of a neuroblast do not have preferential innervation patterns, but instead become part of different layers and neuropils. This study established a new methodology that helps understanding the logic of Drosophila brain development and can be applied to the more complex vertebrate brains (Del Valle Rodriguez, 2020).
Sensory systems need to reliably extract information from highly variable natural signals. Flies, for instance, use optic flow to guide their course and are remarkably adept at estimating image velocity regardless of image statistics. Current circuit models, however, cannot account for this robustness. This study demonstrates that the Drosophila visual system reduces input variability by rapidly adjusting its sensitivity to local contrast conditions. Functional properties were exhaustively mapped of neurons in the motion detection circuit; local responses were found to be compressed by surround contrast. The compressive signal is fast, integrates spatially, and derives from neural feedback. Training convolutional neural networks on estimating the velocity of natural stimuli shows that this dynamic signal compression can close the performance gap between model and organism. Overall, this work represents a comprehensive mechanistic account of how neural systems attain the robustness to carry out survival-critical tasks in challenging real-world environments (Drews, 2020).
aThe anatomy of many neural circuits is being characterized with increasing resolution, but their molecular properties remain mostly unknown. This study characterized gene expression patterns in distinct neural cell types of the Drosophila visual system using genetic lines to access individual cell types, the TAPIN-seq method to measure their transcriptomes, and a probabilistic method to interpret these measurements. These tools were used to build a resource of high-resolution transcriptomes for 100 driver lines covering 67 cell types. Combining these transcriptomes with recently reported connectomes helps characterize how information is transmitted and processed across a range of scales, from individual synapses to circuit pathways. Examples are described that include identifying neurotransmitters, including cases of apparent co-release, generating functional hypotheses based on receptor expression, as well as identifying strong commonalities between different cell types (Davis, 2020).
Visual perception scales with changes in the visual stimulus, or contrast, irrespective of background illumination. However, visual perception is challenged when adaptation is not fast enough to deal with sudden declines in overall illumination, for example, when gaze follows a moving object from bright sunlight into a shaded area. This study shows that the visual system of the fly employs a solution by propagating a corrective luminance-sensitive signal. In vivo 2-photon imaging and behavioral analyses were used to demonstrate that distinct OFF-pathway inputs encode contrast and luminance. Predictions of contrast-sensitive neuronal responses show that contrast information alone cannot explain behavioral responses in sudden dim light. The luminance-sensitive pathway via the L3 neuron is required for visual processing in such rapidly changing light conditions, ensuring contrast constancy when pure contrast sensitivity underestimates a stimulus. Thus, retaining a peripheral feature, luminance, in visual processing is required for robust behavioral responses (Ketkar, 2020).
Sensory systems have evolved to detect changes rather than absolute inputs from their environment. For example, while it is possible to forget that glasses are perched on top of our head, insect landing on our skin can be felt. Similarly, visual systems are well suited to detect changes in light intensity. In many contexts, changes in perception are proportional to relative changes in a stimulus (Weber contrast). The ability to process such changes in sensory input under changing conditions is crucial to the behavior and survival of many organisms (Ketkar, 2020).
In vision, the most basic characteristic of a visual stimulus is the distribution of luminance, which is the luminous intensity per unit area. Rather than responding to luminance, many cells in the visual system instead respond to contrast. Contrast can be defined in the temporal or spatial domain. This study examined temporal contrast, which corresponds to the relative change in luminance (Weber contrast) over time at a single point in space. This study describes a mechanism that ensures contrast constancy, a visual phenomenon that allows discrimination of a visual feature from its background solely based on its contrast, regardless of the global light level (Ketkar, 2020).
Many visual systems are well suited to function at dusk, dawn, or in daylight as well as in rapidly changing environments. In both vertebrates and invertebrates, photoreceptor adaptation is a key mechanism that ensures contrast computation irrespective of background illumination. Photoreceptor adaptation happens across different timescales that range from tens of milliseconds to tens of seconds. However, animals frequently encounter close-to-instantaneous changes in luminance that far exceed photoreceptor adaptation. For example, during self-motion, saccadic eye movements, or when gaze follows a moving object into the shade, background luminance changes within milliseconds. A failure to adjust light sensitivity equally fast would lead to inaccurate calculation of contrast and a misinterpretation of salient cues. Therefore, the timescale of adaptation might limit the capability of the visual system to ensure contrast constancy in fast-changing light conditions, and thus may be alone insufficient to explain robust visual behaviors under changing conditions (Ketkar, 2020).
Core visual circuits have been mapped in the fruit fly Drosophila, which responds to visual cues at luminances spanning orders of magnitude. Briefly, information passes from the retina through the lamina and medulla to the lobula complex. Visual information is computed in 800 parallel channels, together forming a retinotopic array. Downstream of photoreceptors, visual circuitry splits into pathways that are specialized to detect contrast increments (ON) or decrements (OFF). The lamina neuron L1 forms the principal input to the ON pathway, whereas L2 and L3 are major input neurons to the OFF pathway. Together, L1-L3 are referred to as large monopolar cells (LMCs). Based on behavioral experiments, L2 and L3 were considered to mediate fast and slow motion-processing pathways, respectively. However, while L1 and L2 show transient responses to light and biphasic filtering properties, L3 responses are sustained and display a monophasic linear temporal filter. This argues that L2 and L3 are sensitive to entirely different features of the visual scene (Ketkar, 2020).
In flies, voltage recordings have shown that the photoreceptor cells respond to prolonged, bright illumination with an initial transient phase, which encodes contrast, followed by a luminance-sensitive plateau, which lasts for the duration of the stimulus. Downstream LMCs are thought to amplify photoreceptor contrast signals while discarding information about constant illumination. Such contrast responses in LMCs have been extensively characterized in flies that were adapted to different mean luminances. However, it is not known how the visual system deals with contrast signals in a non-adapted state. Ideally, an animal should be able to show invariant responses to contrast under rapidly changing light conditions. To achieve contrast constancy under such conditions, information about illumination might be advantageous. Interestingly, in the vertebrate retina, luminance sensitivity is retained past photoreceptors at the bipolar cell to amacrine cell synapse. Furthermore, intrinsically photosensitive retinal ganglion cells can contribute to image-forming visual functions. However, in the Drosophila visual system, a luminance-sensitive component has not been described past photoreceptors. Furthermore, it is not known whether luminance indeed contributes to image processing past the initial detection stage (Ketkar, 2020).
This study has uncover a luminance-sensitive pathway in the Drosophila visual system. Contrast-sensitive neuronal responses alone are insufficient to account for behavioral responses to changing visual stimuli, arguing for the presence of a corrective signal that scales contrast-sensitive responses when light levels suddenly decline. Through a series of in vivo calcium-imaging experiments, this study show that the two input neurons to the OFF pathway, L2 and L3, encode contrast and luminance, respectively. Luminance information from L3 is necessary for appropriate behavioral responses to visual cues when background luminance suddenly becomes dim. This is true across a range of adaptation states. The data demonstrate that luminance information is retained in a specialized L3 pathway and used as a corrective signal that ensures contrast constancy. This work thus highlights a novel mechanism that allows accurate image processing under dynamically changing light conditions (Ketkar, 2020).
This study has demonstrated that contrast sensitivity alone is not sufficient to explain the behavioral response to visual stimuli. Luminance, the primary input to the visual system, is additionally required for behavioral responses. This study has shown that the two OFF pathway neurons, L2 and L3, are sensitive to fundamentally different visual features, contrast, and luminance. The presence of a luminance-sensitive pathway boosts behavioral performance where contrast pathways prove inadequate, demonstrating a role for luminance in image processing. The specific requirement of luminance signals in behavior, along with the underlying L3 physiology, are consistent across a range of adaptation states. Together, the data suggest that visual processing can handle changing light conditions because the basic luminance feature is passed on by the early fly visual system and is utilized to modulate further computations in a behaviorally relevant way (Ketkar, 2020).
This study has shown that a sustained component in the lamina neuron L3 is sensitive to luminance. First-order visual interneurons have been characterized extensively, and canonical LMCs are generally thought to be 'amplified and more transient versions of photoreceptor responses' and, therefore, contrast sensitive. This contrast sensitivity is visible in L2 calcium responses. However, it was noted that L3 is physiologically distinct and has a more pronounced sustained component and a slower temporal filter compared with those of L1 and L2. Electrophysiological recordings showed that the L3 membrane potential did not return to baseline upon prolonged visual stimulation. The data show that this is more pronounced for calcium signals and that L3 responds to a step change in luminance with a step change in GCaMP signal. Luminance information is retained in a specialized L3-mediated pathway in parallel to contrast-sensitive pathways to facilitate visual processing under conditions that are challenging for purely contrast-sensitive pathways (Ketkar, 2020).
The parallel presence of sustained (tonic) and transient (phasic) responses has been observed in different neural systems. Examples include proprioceptive neurons, where tonic and phasic neurons encode joint position and movement, or mechanosensory neurons from Johnston's organ, where tonic and phasic responses encode antennal deflection and vibration, respectively (Ketkar, 2020).
The findings suggest that a behavioral response to a stimulus of varying temporal contrast in the Drosophila OFF pathway is computed in two phases -- first, contrast-sensitive LMCs, such as L2, report the luminance difference relative to the adapted luminance, and this signal is then subjected to corrective modulation with the luminance information from the L3 pathway. In species ranging from flies to humans, photoreceptor adaptation mechanisms operate on different timescales, typically a faster one that takes a fraction of a second and a slower one that lasts several seconds or minutes. These processes allow visual systems to operate under a wide range of luminances to detect contrast as a salient feature in the environment. However, fast changes in mean luminance caused by a shadow or by an animal's own movement would cause high contrasts to elicit only very small photoreceptor responses if the adaptation state was not yet fully caught up. Consequently, signals encoded by contrast-sensitive neurons would underrepresent the physical contrast. The presence of a luminance-sensitive pathway appears to overcome these limitations to ensure accurate motion detection in sudden dim light. This could happen through excitatory interactions between the L3 and the L2 pathways. At low luminance, when L3 is most active, such an interaction would amplify signals from contrast-sensitive neurons and thereby allow for a behaviorally adequate response. Such an interaction between luminance- and contrast-sensitive channels might be a general mechanism implemented in visual systems to explain robust behavioral responses when adaptation timescales are insufficient (Ketkar, 2020).
Luminance and contrast signals must ultimately be combined to control motion-guided behaviors. This could for example happen in the transmedullary (Tm) or medulla intrinsic (Mi) neurons that are postsynaptic to LMCs. One major postsynaptic partner of L3, Tm9, possesses similarly sustained responses to those seen in L3. Interestingly, Tm9 has wide receptive field properties, potentially receiving input from several neighboring columns. Thus, Tm9 could serve to integrate over space to identify a change in illumination and help to distinguish global from local changes in signal. Tm9 responses further depend on the biphasic ON pathway input L1, arguing that transient and sustained responses are combined at this stage. All other major medulla OFF pathway interneurons receive predominant input from L2 and have transient response kinetics. Medulla neurons of the ON pathway can also be separated into sustained and transient types, and connectomics suggests that L3 could provide luminance information to both ON and OFF pathways. Thus, relative lamina neuron contributions might determine downstream response properties. Further integration could happen in the dendrites of the downstream direction-selective T4 and T5 cells. Luminance information might simply act on contrast signals to scale them before they are spatiotemporally compared. The addition of a DC component also enhanced predictions of motion responses to slow apparent motion stimuli and reverse-phi illusion. Luminance-sensitive L3 responses may thus provide the neural substrates for conveying this DC component to downstream visual circuitry. Finally, L3 feeds into the color pathway by for example providing input to Dm9 cells and could thus help establish color constancy (Ketkar, 2020).
The finding that the presence of luminance information downstream of photoreceptors allows for appropriate behavioral response could suggest a selective advantage for animals living in certain environments. It will be interesting to see if different species with specific ethological constraints have evolved similar luminance-sensitive pathways and how they facilitate visual behaviors. The requirement for such a pathway appears to be greater in animals encountering broad luminance variance over short time periods. Although local mean and variance in natural scenes vary independently, a positive correlation between them sometimes exists because of shadows. Because objects in bright sunlight tend to cast darker shadows, diurnal animals are more likely to encounter wide variance and to possess a luminance-sensitive channel. Nocturnal animals, on the other hand, may require luminance information to tackle challenges posed by low light levels. Therefore, luminance information preserved beyond photoreceptors might facilitate behavior in different ways depending on the species' ethology (Ketkar, 2020).
Physiologically different LMC subtypes have been described in different species. For example, LMCs in bumblebees and butterflies exhibit sustained components, and motion-sensitive neurons perform better than photoreceptor responses predict in the nocturnal hawkmoth. Interestingly, presynaptic L2 and L3 calcium signals are highly reminiscent of sustained and transient calcium signals in bipolar cells of the vertebrate retina. Information about luminance is retained past vertebrate photoreceptors at the rod bipolar cell to the AII amacrine cells synapse. The rod bipolar cell pathway is an important player specifically in dim light conditions. Together, these findings suggest that luminance information in the vertebrate retina is also used to aid image processing. In line with this idea, human eyes come across substantial variation in luminance every few milliseconds when inspecting natural scenes with saccades. It has been suggested that gain control to luminance fluctuations also occurs within the retinal circuitry and past photoreceptor adaptation. Therefore, the finding that the visual system uses luminance information to aid contrast computation may reveal a general image processing strategy utilized across phyla (Ketkar, 2020).
Many visual animals have innate preferences for particular wavelengths of light, which can be modified by learning. Drosophila's preference for UV over visible light requires UV-sensing R7 photoreceptors and specific wide-field amacrine neurons called Dm8. This study identified three types of medulla projection neurons downstream of R7 and Dm8 and has shown that selectively inactivating one of them (Tm5c) abolishes UV preference. Using a modified GRASP method to probe synaptic connections at the single-cell level, each Dm8 neuron was shown to form multiple synaptic contacts with Tm5c in the center of Dm8's dendritic field but sparse connections in the periphery. By single-cell transcript profiling and RNAi-mediated knockdown, it was determined that Tm5c uses the kainate receptor Clumsy to receive excitatory glutamate input from Dm8. It is concluded that R7s-->Dm8-->Tm5c form a hard-wired glutamatergic circuit that mediates UV preference by pooling approximately 16 R7 signals for transfer to the lobula, a higher visual center (Karuppudurai, 2014).
Understanding how visual systems translate light impulses into adaptively tuned percepts to guide behavior is a central goal in neurobiology. The Drosophila visual system, with its amenability to genetic manipulation, has enabled increasingly deep investigation of the molecular and cellular basis of visual-driven behaviors, including the spectral preference for UV light examined in this study. UV spectral preference has previously been shown to require first-order interneurons in the visual medulla (i.e., the wide-field amacrine Dm8 neurons) that receive inputs from multiple UV-sensing photoreceptors. This study shows that a subclass of Tm5 neurons, called Tm5c, receive excitatory glutamatergic input from Dm8 neurons through the kainate-type receptor Clumsy. Glutamatergic signaling, both to and by Tm5c, was shown to be necessary for normal UV preference. Together these results define not only critical elements of the molecular and cellular machinery underlying UV preference, but also patterns of connectivity and information flow at the first several processing stations of this important visual circuit (Karuppudurai, 2014).
By sparse reconstruction of serial-section transmission EM (ssTEM), Previous work identified Dm8 as the major postsynaptic partner for R7. This study shows that Tm5c interneurons are required for transducing Dm8's signal to the lobula, a higher visual center, in the UV preference pathway. The thin and complex dendrites of Tm5c make them challenging to resolve by ssTEM, due to the limited axial resolution (∼50 nm) of this method. To visualize Dm8->Tm5c and R8->Tm5c synapses, this study therefore resorted to the GRASP technique. To differentiate synaptic contacts from mere membrane contacts and to visualize the spatial distribution of Dm8->Tm5c synapses, the GRASP method was adapted to permit single-cell identification of presumptive presynaptic neurons in which active zones were fluorescently tagged. By applying this single-cell GRASP method, it was demonstrated that each Dm8 neuron has multiple synaptic contacts to one (or, at most a few) Tm5c neuron in the center of its dendritic field but sparse synaptic contacts in the periphery. The nature of synaptic signaling at the R8->Tm5c and Dm8->Tm5c contacts was established by single-cell transcript profiling and functional studies. Tm5c expresses the histamine-gated chloride channel Ort, and restoring Ort expression in Tm5c in an ort mutant background drove strong green preference. Dm8 expresses VGlut, and Tm5c correspondingly expresses glutamate-gated ionotropic receptors. RNAi-mediated knockdown of VGlut in Dm8, or the Clumsy iGluR in Tm5c, abolished UV preference. It is believed that the approach taken in this study, which combines single-cell GRASP, transcript profiling, RNAi-mediated knockdown, and behavioral assays, could be profitably applied to the dissection and characterization of other complex neural circuits (Karuppudurai, 2014).
Using single-cell transcript profiling, four kainate-type glutamate receptor subunits (Clumsy, CG11155, DKaiRIC, and DKaiRID) expressed in Tm5c were identified. These four iGluRs and CG9935 share sequence homology and domain structures with vertebrate kainate-type iGluRs (GluK1/2/3 and GluK4/5). RNAi-mediated knockdown further revealed that Clumsy is functionally required in Tm5c for UV preference. This demonstrates that kainate-type iGluRs function in the Drosophila CNS. In vertebrates, functional kainate receptors assemble tetramerically as dimers of dimers; GluK1-3 are capable of forming functional homotetramers while GluK4/5 are obligatory heteromers. Fly kainate receptor subunits share with vertebrates the key hydrophobic resides at the dimerization interfaces, suggesting that they assemble in a similar way to the vertebrate receptors. RNAi knockdown of CG11155, DKaiRIC, or DKaiRID did not enhance UV preference defects caused by RNAi knockdown of Clumsy. However, simultaneous RNAi knockdown of all three iGluR subunits significantly reduced UV preference, suggesting that they are functionally redundant. It is thus suggested that Clumsy forms functional heterotetramers with one of the other kainate receptors in Tm5c to mediate UV preference. Further in vitro assembly and electrophysiological studies will be needed to determine the exact subunit composition of the functional iGluRs. Ionotropic glutamate receptors in flies have been exclusively studied at the neuromuscular junction, in part as a surrogate model for CNS glutamate synapses. Functional identification of kainate-type iGluRs in the fly visual system, in combination with the robust UV preference behavior they mediate, opens the door to studying the assembly, function, and regulation of this important class of glutamate receptors in the Drosophila CNS (Karuppudurai, 2014).
As the wide-field Dm8 neurons have no presynaptic sites or axonal projections outside of the external medulla, they depend on medulla projection neurons to transduce signals to higher visual centers. While all three subtypes of Tm5 neurons appear to be postsynaptic to Dm8 and therefore are capable of transducing the Dm8 signal to the lobula, this study shows that only Tm5c is functionally required for UV preference. Inactivating Tm5a/b or Dm8 in addition to Tm5c did not enhance UV preference defects, suggesting that Dm8's function in UV preference is solely communicated through Tm5c. Tm5c differs from Tm5a/b in axonal morphology and neurotransmitter usage. It is thus suggested that Tm5c has unique synaptic functions and/or targets in that visual compartment that account for its role in UV preference (Karuppudurai, 2014).
The spatial organization of the R7s->Dm8->Tm5c circuit suggests a neural pooling mechanism for UV preference. Each Dm8 amacrine neuron has a large dendritic field that receives ∼16 R7 inputs, while a single Tm5c is present in most, if not all, medulla columns and receives direct retinotopic inputs from R8s. Single-cell GRASP experiments reveal that each Dm8 forms multiple synaptic contacts with one, or at most a few, Tm5c neurons in the center of Dm8's dendritic field but forms few synapses with Tm5c outside of the center. By pooling 16 R7 inputs to a single Tm5c, Dm8 could increase UV sensitivity by up to 16-fold at some cost in spatial resolution. It is interesting to note that the amplification magnitude of the R7s->Dm8->Tm5c circuit depends primarily on the size of the Dm8's dendritic field, which is negatively regulated by R7-derived Activin during development: excess Activin reduces Dm8's dendritic field size while lack of Activin enhances it (Ting, 2014). It is thus tempting to speculate that size of the Dm8 arbor, and thus the trade off between UV sensitivity and spatial resolution, has been adjusted in the course of insect evolution to meet each insect's ecological needs (Karuppudurai, 2014).
In addition to the excitatory glutamate input from Dm8, Tm5c neurons also receive inhibitory histaminergic inputs directly from R8 photoreceptors. Thus, the R7s->Dm8->Tm5c pooling circuit is superimposed on the retinotopic circuit R8->Tm5c. Tm5c expresses Ort, and restoring Ort expression in Tm5c in various ort mutant backgrounds rescued green phototaxis. Thus, both direct (R8->Tm5c) and indirect (R7s->Dm8->Tm5c) pathways transduce sign-inverting signals to Tm5c and both pathways are capable of driving phototaxis. However, RNAi-mediated knockdown of ort in Tm5c, which prevents the reception of R8 inputs, did not affect normal UV preference. This observation is consistent with previous conclusions that the indirect pathway through R7s->Dm8 is both required and sufficient for optimal UV preference, at least under the test condition, and that multiple redundant pathways mediate green phototaxis. While the direct pathway is not involved in UV preference, it might play a role in true color vision. Notably, chloride ions are a known modulator for kainate receptor and the direct pathway signals through the histamine-gated chloride channel Ort. Given that multiple pathways function redundantly in true color vision, differentiating these possibilities must await single-unit electrophysiological recording and/or new genetic techniques to segregate their functions (Karuppudurai, 2014).
Color and polarization provide complementary information about the world and are detected by specialized photoreceptors. However, the downstream neural circuits that process these distinct modalities are incompletely understood in any animal. Using electron microscopy, this study had systematically reconstructed the synaptic targets of the photoreceptors specialized to detect color and skylight polarization in Drosophila, and light microscopy was used to confirm many of the findings. Known and novel downstream targets were identified that are selective for different wavelengths or polarized light, and their projections were followed to other areas in the optic lobes and the central brain. The results revealed many synapses along the photoreceptor axons between brain regions, new pathways in the optic lobes, and spatially segregated projections to central brain regions. Strikingly, photoreceptors in the polarization-sensitive dorsal rim area target fewer cell types, and lack strong connections to the lobula, a neuropil involved in color processing. This reconstruction identifies shared wiring and modality-specific specializations for color and polarization vision, and provides a comprehensive view of the first steps of the pathways processing color and polarized light inputs (Kind, 2021).
Systematic reconstruction of all synaptic inputs and outputs of identified, functionally specialized Drosophila photoreceptors (pale and yellow R7-8 and two dorsal rim area photoreceptors, R7-DRA and R8-DRA) provides a comprehensive inventory of the first steps of the color and polarization pathways, from which all the computations of the dependent behaviors stem. These data revealed core connectomic motifs shared across column types, multiple new photoreceptor targets, and uncovered additional cell types as being connected to specific photoreceptor subtypes conveying specific color and polarization information to the central brain (Kind, 2021).
This study confirmed previously reported synaptic partners of the inner photoreceptors in the non-DRA medulla and identified new photoreceptor targets. As prior reconstructions were incomplete, it was unclear whether the unidentified connections were mainly onto new target neurons or represented more connections onto known cell types; reconstructions revealed both types of omissions. One functionally important set of missed connections are the synapses between inner photoreceptors from the same central and DRA ommatidia, which was found to be stronger than previously reported, due to significant numbers of synapses outside the medulla. These synapses likely contribute to color-opponent responses seen in central R7 and R8 terminals and the polarization-opponent signals measured from DRA photoreceptors. The reconstructions also support a larger-scale opponent process mediated by multicolumnar Dm9 cells, which also formed some synapses outside the medulla neuropil (Kind, 2021).
Other cell types also received inner photoreceptor input outside the medulla, notably the lamina monopolar cells L1 and L3. These lamina connections indicate that chromatic comparisons arising from R7 and R8 may feed into the motion vision pathway, and identifies a new site for interplay between the 'color' and 'motion' pathways. Together, these observations suggest that synapses in an unexpected location, outside the main synaptic layers of the medulla, could play a significant role in early visual processing (Kind, 2021).
In the non-DRA medulla, this study found, for the first time in an EM study, strong synaptic connections between R7s and MeTu cells that project to the AOTU, confirming previous claims based on light microscopy. This finding may reconcile disparate observations, such as a role in wavelength-specific phototaxis for cells matching MeTu morphology, as well as measurements of color-sensitive signals in the AOTU of bees. Previous anatomical studies partitioned MeTu cells into distinct subclasses that terminate in discrete subdomains of the AOTU. This study identified modality-specific MeTu-DRA cells that only integrate from the polarization-sensitive R7-DRA photoreceptors, while avoiding synaptic contacts with color-sensitive pale or yellow R7s. MeTu and MeTu-DRA cells target adjacent subdomains within the small unit of the AOTU, in agreement with proposals that parallel channels convey different forms of visual information from the eye to the central complex via the AOTU (Kind, 2021).
This study also identified connections from inner photoreceptors to several cell types either not previously described or not known to be photoreceptor targets, thus setting up a clear expectation that these cells should contribute to color or polarization vision. ML1 is a new, major target of R8, a cell type that connects the medulla to the lobula via a previously unknown, non-columnar pathway and the central brain. Previous studies have identified an important role for Tm5a/b/c and Tm20 cell types for chromatic processing in the lobula, and this study has confirmed that these cell types are targets of R7-8. Whether these Tm neurons and ML1 cells have common targets in the lobula, feed into shared central pathways or contribute to separate channels remain open questions; the lobula arbors of the Tm and ML1 cells are mostly in different layers arguing against direct synaptic interactions between the cells (Kind, 2021).
The reconstruction of DRA photoreceptor targets confirmed the modality-specific connectivity of Dm-DRA cell types within layer M6: Dm-DRA1 to R7-DRA, and Dm-DRA2 to R8-DRA. The R7-DRA and R8-DRA cells respond to orthogonal orientations of polarized light at each location of the DRA, so Dm-DRA1 and Dm-DRA2 likely process orthogonal e-vector orientations spatially averaged by pooling over ~10 ommatidia. The data also revealed additional DRA pathways into the PLP in the central brain via VPN-DRA cells, as well as to the contralateral DRA, via MeMe-DRA cells. Such interhemispheric connections have been demonstrated in larger insects, but not in Drosophila, and their synaptic input was not known. Interocular transfer contributes to navigation by desert ants that can see the celestial polarization but not visual landmarks, but the interactions between the DRA regions remain poorly understood. The identification of MeMe-DRA neurons may enable the mechanisms of such phenomena to be explored (Kind, 2021).
This study has additionally identified as pale-specific the aMe12 and ML-VPN1 cell types, and the yellow bias of Tm5c and the pale bias of Mi9. Detailed analysis of Dm8 inputs confirmed that these neurons receive most of their photoreceptor input in a central home column, consistent with pale and yellow subtypes of Dm8 cells having distinct chromatic properties. The selective photoreceptor input to these cell types, combined with input to columnar cells from a single photoreceptor subtype, indicates that wavelength-specific information is maintained in the medulla and lobula. The projections of aMe12 and ML-VPN1 to the central brain indicate the possibility that wavelength-specific photoreceptor responses are directly conveyed into the central brain by these cells, although they likely integrate input from other chromatically sensitive cell types (Kind, 2021).
By focusing on a small number of columns, this study has delivered a near-complete picture of local connectivity, but the possibility cannot be ruled out that certain cell types may have been overlooked. For instance, an arbitrary threshold was chosen of two to three synapses below which no attempts were made to reconstruct synaptic targets to the extent required to uniquely identify them. As a result this study may, in principle, have missed large cells that receive small, but significant inputs across many columns. Furthermore, regional specializations in the optic lobes, such that specific cell types might be found outside of the seed columns, cannot be ruled out. For example, some MeTu cells are only found in the dorsal third of the medulla, where incoming R7 cells are known to co-express Rh3 and Rh4 rhodopsins (Kind, 2021).
Taken together, the data presented in this study provides access to the full complement of R7 and R8 photoreceptor targets from functionally specialized optical units. By reconstructing these local circuits within a full-brain EM volume, it was possible to establish the complete morphology of large, multicolumnar cell types, that are strongly connected to photoreceptors, but had eluded previous connectomic reconstruction efforts. The sparse tracing approach that was implemented and described in this study enabled efficient identification of the complete set of upstream and downstream partners of the inner photoreceptors, in a manner that is complementary to the dense connectomes generated from smaller-scale medulla volumes. As an example of this synergistic usage of complementary data sets, this study has returned to the 7-column data and used whole-cell morphologies to match the bodies of previously unidentified photoreceptor targets. In that process strong candidates were established for MeTu, ML1, and perhaps for aMe12 and several aspects of their connectivity in the Zheng (2018) data set were confirmed. In so doing, it is now possible to have access to the additional connectivity data provided by the 7-column dense reconstruction. While full exploration and follow-up analyses of these combined data is beyond the scope of this work, the combined analysis of the Zheng (2018), and the medulla-7-column connectome revealed several intriguing connectivity patterns, including new candidate paths for the integration of output from different photoreceptor types. For example, the MeTu cells that are postsynaptic to R7 also receive indirect R8 input via the R8 target Mi15, and ML1 combines direct R8 input with indirect input from outer photoreceptors via lamina neurons. As further connectome data sets are completed, this comparative interplay between data sets with unique advantages and limitations will be an important step in both cross-validating and extending the applicability of all related data sets (Kind, 2021).
Reconstruction of the DRA photoreceptor targets has provided the first EM-based connectomic data set for modality-specific cell types likely to process skylight information in any insect and will be important for developing refined models of skylight navigation. Core motifs shared between DRA and central columns are prime candidates for circuit elements that perform computations, such as establishing opponency, that are key for both polarization and color processing, whereas cell types with preferential connections to either pale or yellow columns are promising candidates for the study of specific aspects of color processing in the insect brain. This comprehensive catalog of the neurons carrying signals from R7 and R8 photoreceptors deeper into the brain establishes a broad foundation for further studies into the mechanistic basis of color vision and its contributions to perception and behavior (Kind, 2021).
Connectomics has become a standard neuroscience methodology in a few model animals, with the visual system being a popular target of study. Combining connectomics with circuit and behavioral physiology, recent studies on the color vision of the fruit fly Drosophila melanogaster have focused on the mechanisms underlying early wavelength processing in the optic ganglia. However, the color vision capabilities of D. melanogaster are limited, compared with many flower-visiting insects. For example, a butterfly Papilio xuthus has six spectral classes of photoreceptors. Each ommatidium contains nine photoreceptors in one of three fixed combinations, making the eye an array of three spectrally distinct ommatidia types. Behaviorally, P. xuthus can detect 1 nm differences in light wavelength across the spectrum from ultraviolet to red, outperforming humans. What is the neuronal basis of such precise color vision? How does such a system evolve? Addressing these questions requires comparative studies at the circuit level. This study performed a connectome analysis in the first optic ganglion, the lamina, of P. xuthus. The lamina comprises cartridges, each typically containing nine photoreceptor axons from a single ommatidium and four second-order neurons. Abundant inter-photoreceptor connections were found that are absent in the lamina of D. melanogaster. Connections were identified between neighboring cartridges, particularly those receiving inputs from spectrally distinct ommatidia. The linear summation of synaptic connections well explains the spectral sensitivity of photoreceptors and second-order neurons in the lamina (Matsushita, 2022).
Sensory systems rely on neuromodulators, such as serotonin, to provide flexibility for information processing as stimuli vary, such as light intensity throughout the day. Serotonergic neurons broadly innervate the optic ganglia of Drosophila. This study mapped of patterns of serotonin receptors in the visual system, focusing on a subset of cells with processes in the first optic ganglion, the lamina. Serotonin receptor expression was found in several types of columnar cells in the lamina including 5-HT2B in lamina monopolar cell L2, required for spatiotemporal luminance contrast, and both 5-HT1A and 5-HT1B in T1 cells, whose function is unknown. Subcellular mapping with GFP-tagged 5-HT2B and 5-HT1A constructs indicated that these receptors localize to layer M2 of the medulla, proximal to serotonergic boutons, suggesting that the medulla neuropil is the primary site of serotonergic regulation for these neurons. Exogenous serotonin increased basal intracellular calcium in L2 terminals in layer M2 and modestly decreased the duration of visually induced calcium transients in L2 neurons following repeated dark flashes, but otherwise did not alter the calcium transients. Flies without functional 5-HT2B failed to show an increase in basal calcium in response to serotonin. 5-HT2B mutants also failed to show a change in amplitude in their response to repeated light flashes but other calcium transient parameters were relatively unaffected. While serotonin receptor expression in L1 neurons was not detected, they, like L2, underwent serotonin-induced changes in basal calcium, presumably via interactions with other cells. These data demonstrate that serotonin modulates the physiology of interneurons involved in early visual processing in Drosophila (Sampson, 2020).
Appropriate termination of the photoresponse in image-forming photoreceptors and downstream neurons is critical for an animal to achieve high temporal resolution. Although the cellular and molecular mechanisms of termination in image-forming photoreceptors have been extensively studied in Drosophila, the underlying mechanism of termination in their downstream large monopolar cells remains less explored. This study shows that synaptic ACh signaling, from both amacrine cells (ACs) and L4 neurons, facilitates the rapid repolarization of L1 and L2 neurons. Intracellular recordings in female flies show that blocking synaptic ACh output from either ACs or L4 neurons leads to slow repolarization of L1 and L2 neurons. Genetic and electrophysiological studies in both male and female flies determine that L2 neurons express ACh receptors and directly receive ACh signaling. Moreover, the results demonstrate that synaptic ACh signaling from both ACs and L4 neurons simultaneously facilitates ERG termination. Finally, visual behavior studies in both male and female flies show that synaptic ACh signaling, from either ACs or L4 neurons to L2 neurons, is essential for the optomotor response of the flies in high-frequency light stimulation. This study identifies parallel synaptic ACh signaling for repolarization of L1 and L2 neurons and demonstrates that synaptic ACh signaling facilitates L1 and L2 neuron repolarization to maintain the optomotor response of the fly on high-frequency light stimulation (Wu, 2021).
A fascinating question in neuroscience is how ensembles of neurons, originating from different locations, extend to the proper place and by the right time to create precise circuits. This study investigated this question in the Drosophila visual system, where photoreceptors re-sort in the lamina to form the crystalline-like neural superposition circuit. The repeated nature of this circuit allowed establishment of a data-driven, standardized coordinate system for quantitative comparison of sparsely perturbed growth cones within and across specimens. Using this common frame of reference, the extension was investigated of the R3 and R4 photoreceptors, which is the only pair of symmetrically arranged photoreceptors with asymmetric target choices. Specifically, it was found that extension speeds of the R3 and R4 growth cones are inherent to their cell identities. The ability to parameterize local regularity in tissue organization facilitated the characterization of ensemble cellular behaviors and dissection of mechanisms governing neural circuit formation (Ji, 2021).
{kind=link}
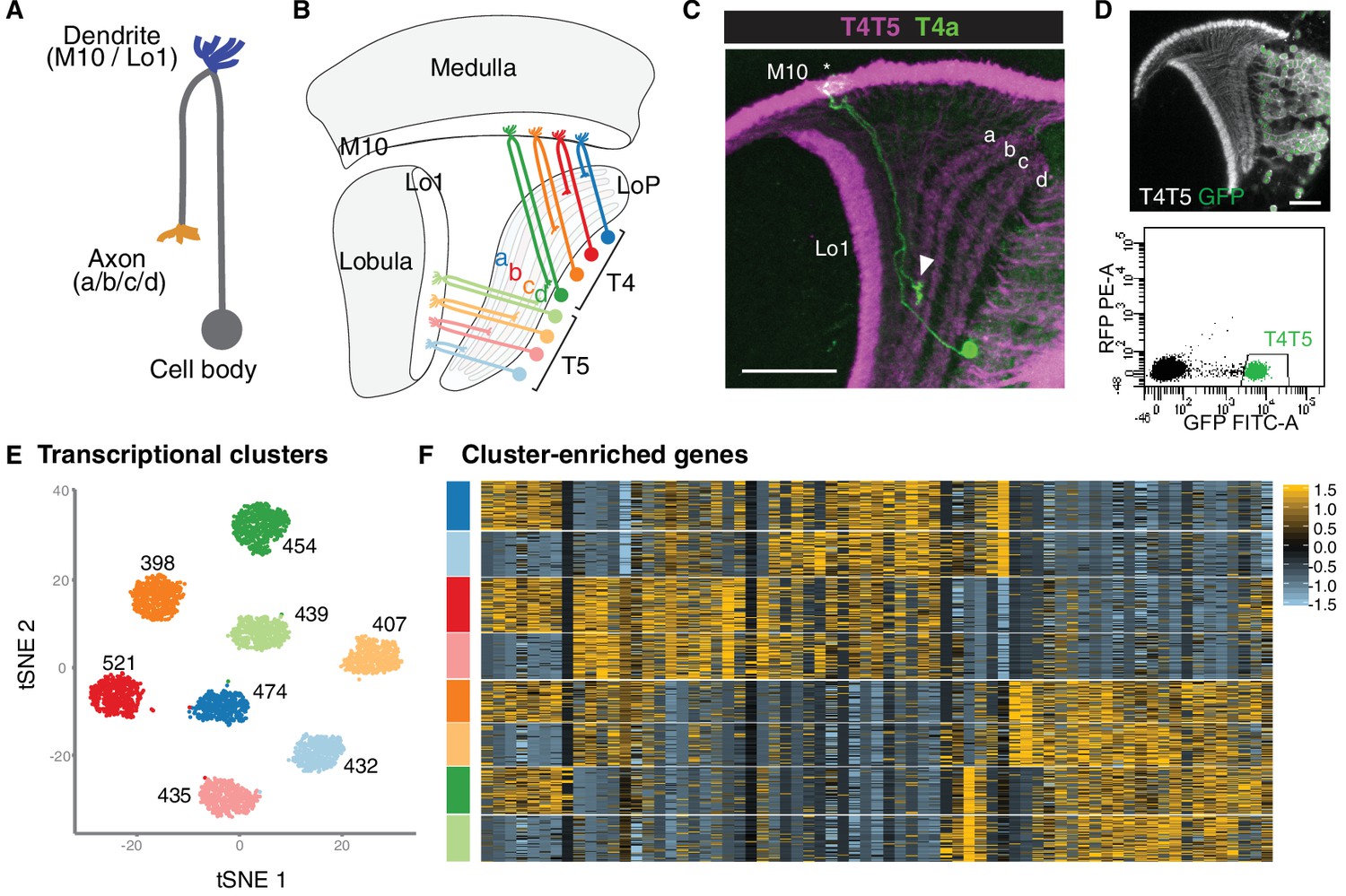{kind=link}