The Interactive Fly
Zygotically transcribed genes
Embryonic origins of a motor system: Motor dendrites form a myotopic map in Drosophila
The organisational principles of locomotor networks are less well understood than those of many sensory systems, where in-growing axon terminals form a central map of peripheral characteristics. Using the neuromuscular system of the Drosophila embryo as a model and retrograde tracing and genetic methods, principles underlying the organisation of the motor system have been uncovered. Dendritic arbors of motor neurons, rather than their cell bodies, are partitioned into domains to form a myotopic map, which represents centrally the distribution of body wall muscles peripherally. While muscles are segmental, the myotopic map is parasegmental in organisation. It forms by an active process of dendritic growth independent of the presence of target muscles, proper differentiation of glial cells, or (in its initial partitioning) competitive interactions between adjacent dendritic domains. The arrangement of motor neuron dendrites into a myotopic map represents a first layer of organisation in the motor system. This is likely to be mirrored, at least in part, by endings of higher-order neurons from central pattern-generating circuits, which converge onto the motor neuron dendrites. These findings will greatly simplify the task of understanding how a locomotor system is assembled. These results suggest that the cues that organise the myotopic map may be laid down early in development as the embryo subdivides into parasegmental units (Landgraf, 2003).
The analysis began by correlating the positions of motor neuron dendrites with the distribution of their muscle targets in the periphery. Motor neurons were retrogradely labelled in a pairwise fashion and the positions of their dendritic arbors were mapped. Because of an interest in the mechanisms that underlie the assembly of the motor system, focus was placed on stages when each motor neuron first establishes a characteristic domain of arborisation within the neuropile (early stage 17, 15h after egg-laying [AEL]) (Landgraf, 2003).
Motor axons project into the muscle field via two main nerves, the intersegmental (ISN) and the segmental nerve (SN). The transverse nerve (TN) runs along the segment border and has few motor axons. Choice of nerve root is one of several features that divide the motor neurons into two principal sets, the ISN and SN. (1) The cell bodies of SN motor neurons are located in the same segment as the muscles that they innervate, whereas ISN motor neuron somata are located in the segment next anterior (with the exception of the RP2 and two neuromodulatory efferent ventral unpaired median [VUM] neurons. (2) ISN motor neurons innervate internal muscles, which span a segment from anterior to posterior, whereas SN (and the TN) motor neurons innervate external muscles. External muscles are distinct from the internal set in several respects: (1) they are generally transverse; (2) unlike internal muscles, they require wingless (wg) signalling for their specification; (3) external (but not internal) muscles and their innervating motor neurons express the cell adhesion molecule (CAM) Connectin, with the single exception of muscle ventral transverse 1 (VT1) (Landgraf, 2003 and references therein).
In addition, ISN and SN motor neurons elaborate their dendrites in distinct regions of the neuropile. Dendrites of ISN motor neurons occupy a domain extending posteriorly from the posterior part of one neuromere into the anterior part of the next. SN motor neuron dendrites occupy a domain that lies between the domains of ISN motor neuron arbors (Landgraf, 2003).
Thus, the organisation of the body wall muscles into internal and external sets is reflected centrally in patterns of motor neuron arborisations. The innervating motor neurons project their axons through different nerves and elaborate their dendritic fields in distinct regions of the neuropile. Although dendritic arbors become progressively more elaborate and extensive over developmental time, their separate domains remain clearly recognisable and appear to be maintained at least until the motor system is fully functional (18 h AEL) (Landgraf, 2003).
Having established that there is a central representation of the muscle field, the organisation of the motor neuron dendrites was analyzed in greater detail. (1) The set of external muscles and their innervating (SN) motor neurons were examined. Muscles of similar anteroposterior positions, such as the ventral acute muscle (VA3) and the segment border muscle (SBM), are innervated by motor neurons whose dendritic arbors lie in a common region of the neuropile. Conversely, motor neurons supplying the anterior (lateral transverse 1-2 [LT1-LT2]) versus the posterior (SBM) muscles have dendritic arbors that are correspondingly separated in the anteroposterior axis of the CNS (Landgraf, 2003).
To put the idea of a regular map to the test, focus was placed on an unusual external motor neuron-muscle pair. Muscle VT1 is innervated by a TN rather than an SN motor neuron. However, VT1 lies at the same place in the anteroposterior axis as the SBM, although VT1 is ventral and the SBM more dorsal. The VT1 motor neuron dendritic field is found to overlaps with that of the SBM motor neuron. For the external set, it is concluded that differences in target muscle location in the anteroposterior axis are mapped centrally as regular differences in dendritic position, but dorsoventral distinctions are not (Landgraf, 2003).
It was next asked whether there is a similarly regular representation of the internal muscles in the developing CNS. While most external muscles are transverse and have unique anteroposterior locations, the internal muscles span the width of a segment so that positional distinctions between them are solely in the dorsoventral axis. It was found that the set of internal muscles is represented centrally by three dendritic domains. Motor neurons innervating ventral internal muscles elaborate their dendritic arbors in the anterior half of the ISN dendritic domain. Motor neurons with dorsolateral internal muscle targets (lateral longitudinal [LL] 1, dorsal acute [DA] 3, dorsal oblique 3-5 [DO3-DO5]) put their arbors into the posterior part of the ISN dendritic domain. Finally, dorsal muscles are represented by a motor neuron dendritic domain that lies between those representing ventral (anterior) and dorsolateral (posterior) internal muscle groups. Thus, the internal muscles are represented in the neuropile by three domains of dendritic arborisation that reflect their different dorsoventral locations in the periphery. Once again, it is concluded that there is a regular mapping of muscle position in the neuropile: in this case, it is positions in the dorsoventral axis peripherally that are represented centrally as differences in the anteroposterior locations of dendrites (Landgraf, 2003).
To test the idea that dendritic arbor positions relate to the distribution of muscles, an atypical motor neuron-muscle pair was examined. The RP2 motor neuron is reported to innervate dorsal muscle DA2, yet its dendrites span the domains that represent both dorsal and dorsolateral internal muscles. However, on careful analysis it was found that DA2 is, in fact, specifically innervated by a U neuron whose dendrites lie in the dorsal internal domain, whereas the RP2 axon forms endings generally on all dorsolateral and dorsal muscles by 19 h AEL. These seem to correspond to the type 1s boutons found in late larvae. Thus, the RP2 neuron puts its dendrites into a region of the neuropile that does indeed represent its targets, namely the dorsolateral and dorsal internal muscles (Landgraf, 2003).
Like the muscle field itself, the map of motor neuron dendrites is metamerically repeated. However, the boundaries of these two units are out of register with one another, since the dendrites of the motor neurons innervating internal muscles lie in the next anterior neuromere. The anterior border of the dendritic map, as defined by the extent of these anterior dendrites, coincides with the anterior margin of engrailed (en) expression. Thus, while the muscles are segmental in their organisation, the domains occupied by the dendrites of their innervating motor neurons are parasegmental (Landgraf, 2003).
To test whether genes that implement the parasegmental pattern in the epidermis are also required for the formation of the parasegmental organisation of the neuromuscular system, the formation of SN and ISN dendritic fields was studied in embryos singly mutant for the following segment polarity genes: en/invected (Df(enE)), wg (wgCX4), naked (nkd2), patched (ptc9), hedgehog (hh21), and gooseberry (Df2R(gsb)). Every one of the six different mutants that were analysed has partially aberrant patterns of neuroblasts (NBs). Nevertheless, SN and ISN motor neurons still form and can be identified by their characteristic axonal projections into the periphery. In addition, it was found that the fundamental separation between SN and ISN dendritic domains is present despite often severe perturbations in CNS structure. For example, in gsb mutant embryos, both nerve roots are frequently fused so that the SN and ISN share a common CNS exit point. Nevertheless, SN and ISN axons as well as their dendritic fields do not intermingle but remain separate. These results suggest that the subdivision of the neuropile into the principal ISN and SN dendritic domains is a robust feature of the system, which appears to be specified early in development, since the embryo subdivides into parasegmental units (Landgraf, 2003).
It was next asked what mechanisms underlie the formation of the myotopic map. Because ISN and SN motor neurons lie at different positions in the CNS and their axons grow out into the muscle field through different nerves, it is reasonable to suppose that at least the major subdivision of dendritic arborisations into internal and external domains could be a byproduct of the locations at which the motor neurons are generated and the paths taken by their growing axons. This passive mapping' explanation can be excluded by considering a single motor neuron-muscle pair, namely dorsal transverse 1 (DT1) and its innervating motor neuron. DT1 is an external muscle (by position, orientation, wg dependence, and Connectin expression), yet its motor neuron is clustered with the internal muscle innervating set and its axon (uniquely for the external muscles) grows out through the ISN. Despite its packing within the internal motor neuron' set, the DT1 motor neuron makes a long posterior projection through the internal muscle domain of the myotopic map to reach the external domain, where it arborises appropriately, reflecting the orientation and external nature of its target muscle. In contrast, motor neurons derived from the same NB as DT1 innervate neighboring internal muscles DO3-DO5 and put their dendrites in a more anterior region characteristic of the dorsolateral muscles. These findings strongly suggest that the mapping of the muscle field within the CNS is an active process of growth and arborisation that partitions dendrites into subdomains of the neuropile that are appropriate to their function, rather than a passive subdivision of available space by position of origin or axon trajectory (Landgraf, 2003).
Since dendritic arbors form after motor axons have reached their targets, the muscles could be instrumental in dictating the organisation of the central map. To test this idea, the UAS/GAL4 system was used to misexpress an activated form of Notch (Kidd et al. 1998) in the developing mesoderm, suppressing the formation of muscle founder cells while leaving other tissues intact. In such muscleless embryos, the main nerve trunks, SN and ISN, still form and project into the periphery. Retrograde labellings of these nerves show that SN and ISN motor neurons form relatively normal dendritic arbors that consistently conform to the characteristic separation of SN and ISN dendrites. Thus, the neuropile is partitioned into distinct fields of dendritic arborisation independently of the muscles. It is concluded that the mapping process is likely to be an autonomous property of the motor neurons and their neighboring cells (Landgraf, 2003).
It was next asked whether motor neuron dendritic fields could be patterned by the substrates on which they grow. In the Drosophila ventral nerve cord (VNC), motor neuron dendrites form in the dorsal-most region of the neuropile, sandwiched between longitudinal glia above and the underlying scaffold of axons. Glial cells can act as substrates for supporting and guiding axonal growth. To test whether they might also be required for the growth and spatial patterning of dendritic fields, dendritic arbors were analysed in glial cells missing (gcm) mutant embryos, which are defective in glial cell differentiation. Although the structure of the nervous system is disrupted in gcm mutant embryos and the dendritic arbors are abnormal, they continue to form in their characteristic locations and the fundamental distinction between the ISN and SN motor neuron dendritic fields is maintained. Remarkably, even the long posterior dendritic projection of the DT1 motor neuron forms and reaches its target region, the SN external muscle dendritic domain. These results suggest that the patterning of the neuropile into distinct motor neuron dendritic domains is a process that appears to be intrinsic to the motor neurons and their neighboring neurons, but does not require proper glial cell differentiation (Landgraf, 2003).
One likely explanation for the division of dendrites into separate domains is that there is a process of mutual exclusion between the arborisations of neighboring cells. Such a process of dendritic tiling has so far only been documented between particular classes of sensory neurons, but could also occur in the motor system. The idea of tiling was tested by considering two groups of motor neurons whose axons have a common trajectory, but whose dendritic fields form in adjacent territories. The DO3-DO5 and DT1 motor neurons project their dendrites posteriorly, and at their most-anterior point, these dendrites meet the axons and dendrites of the anterior corner cell (aCC) and U/CQ neurons. To show whether the aCC and U/CQ axons and/or dendrites inhibit the growth of DO3-DO5 and DT1 dendrites anteriorly, these neurons (as well as RP2 and the posterior corner cell [pCC] interneuron) were selectively ablated. Using anti-Even-skipped (Eve) staining as a marker for aCC, RP2, and U/CQs (there are an additional two medially located eve-expressing interneurons, pCC and friend of pCC [fpCC], it was found that these neurons can be selectively ablated before they form dendrites (at approximately 11 h AEL): on average, by 10.5 h AEL all but 0.6 and by 12 h AEL all but 0.06 of the seven medially located eve-expressing neurons have been ablated per half-neuromere. In no instance was a concomitant anterior expansion of the DO3-DO5 and DT1 motor neuron dendrites into the regions vacated by the aCC and U/CQ dendrites observed. It is concluded that, at least in this instance, the initial dendritic territory of one set of motor neurons (DO3-DO5 and DT1) is not defined by a process of tiling, in which they are excluded by neighboring (aCC and U/CQ) dendritic arbors. However, it is possible that the elaboration of motor neuron dendritic arbors during later developmental stages may involve interactions between neighboring dendritic territories, activity-dependent processes, or both (Landgraf, 2003).
Thus, in summary, these results suggest that the mechanisms that subdivide the neuropile into distinct dendritc domains are very robust and refractory to perturbations. They further suggest that the cues that organise the map may be laid down early in development as the embryo subdivides into parasegmental units (Landgraf, 2003).
The patterning of the motor neuron dendritic arbors in the Drosophila embryo represents a first layer of organisation in the motor system. This is likely in part to be mirrored by the endings of higher-order neurons of central pattern generating circuits, which converge onto the myotopic map. While motor neuron cell body positions may, as has been proposed for vertebrate systems, relate to the ontogeny of target muscles, the operation of mature muscles is reflected by the allegiance of corresponding motor neuron dendrites to a particular territory in the neuropile. Thus, changes in muscle operation could be accommodated by a change of allegiance of the appropriate motor neuron dendrites from one domain to another (e.g., the DT1 motor neuron-muscle pair) without the need for rewiring the underlying higher-order circuitry. Such a model resolves the apparent discrepancy between the distributions of motor neuron cell bodies centrally and target muscles in the periphery. It also implies a considerable degree of flexibility, particularly at the level of motor output, yet suggests that elements of the underlying motor circuitry may have been highly conserved (Landgraf, 2003).
Axons and dendrites differ in both microtubule organization and in the organelles and proteins they contain. This study shows that the microtubule motor dynein has a crucial role in polarized transport and in controlling the orientation of axonal microtubules in Drosophila melanogaster dendritic arborization (da) neurons. Changes in organelle distribution within the dendritic arbors of dynein mutant neurons correlate with a proximal shift in dendritic branch position. Dynein is also necessary for the dendrite-specific localization of Golgi outposts and the ion channel Pickpocket. Axonal microtubules are normally oriented uniformly plus-end-distal; however, without dynein, axons contain both plus- and minus-end distal microtubules. These data suggest that dynein is required for the distinguishing properties of the axon and dendrites: without dynein, dendritic organelles and proteins enter the axon and the axonal microtubules are no longer uniform in polarity (Zheng, 2008).
Proper cellular morphology and function depends on the polarized localization of organelles and proteins to specific subcellular compartments. This study shows that dynein plays a crucial role in dendrite arbour patterning and in organizing distinct functional compartments (the axon and dendrites) of a neuron. The position of branches within a dendritic arbour has a key role in determining the inputs a neuron receives from pre-synaptic axons or, in the case of sensory neurons, the local environment. This study shows that dynein is necessary for proper positioning of dendritic branches relative to the soma. As a motor, dynein likely influences branch formation by mediating the distribution of cargos that affect branch growth and dynamics. Notwithstanding an overall decrease in dendrite extension and branching in dynein mutants, time-lapse analysis of a few dendrites revealed that they extend normally but have fewer and less stable terminal branches, suggesting that decreased terminal branching is not simply caused by a decrease in dendrite growth. One likely explanation is that 'branching machinery' (including Golgi outposts, endosomes and potentially other proteins and/or organelles) that are normally transported distally for dendrite extension and maintenance become trapped in the proximal arbour in the dynein mutant neurons, resulting in decreased distal branching and the formation of ectopic branches close to the cell body (Zheng, 2008).
Without dynein, Golgi outposts and Pickpocket (Ppk) are present ectopically in axons, revealing a previously unappreciated role for dynein in mediating the dendrite-specific localization of organelles and proteins. One possible explanation for axonal mislocalization is that Golgi outposts and Ppk interact with a MT plus end-directed motor (e.g., kinesin) that transports them into axons in the absence of dynein. Dynein might normally transport such cargo directly to dendrites; alternatively, it is also possible that cargo first enters axons but that dynein counteracts kinesin and carries this cargo out of axons. Since Golgi outposts moving from the soma into the axon are never seen in wild type neurons, the data favour the former possibility. In contrast to the mislocalization of dendritic protein and organelles, Kin-βgal and proteins destined for the axonal terminal retain their polarized distribution, perhaps to be expected given that kinesin mediates the majority of anterograde axonal transport (Zheng, 2008).
In mammalian and fly neurons, axonal MTs are arrayed plus end-distal whereas dendritic MT orientation is mixed. A long-standing question concerns the mechanism(s) that establish and maintain different MT orientations in axons and dendrites. Loss of dynein function causes the axonal localization of Nod-βgal and retrograde movement of EB1-GFP, indicating that minus end-distal MTs are present in these mutant axons. How might dynein regulate the orientation of axonal MTs? The sliding filament model of axonal MT transport proposes that a subset of dynein in the axon is stationary (via an interaction with stable MTs and/or actin) and that dyneins motor domain interacts with short MT polymers, propelling plus end-distal MTs down the axon as the motor moves to the MT minus end. In vivo data support the idea that in addition to transporting MTs, dynein functions as a 'gatekeeper' to move minus end-distal MTs towards the soma, excluding them from the axon. Neurons lacking functional dynein would still transport MTs, likely via kinesin, but now minus end-distal MTs would infiltrate the axon. Proximal axons likely have unique properties, providing a possible explanation for how minus end distal MTs would be excluded from axons but not dendrites (Zheng, 2008).
Recent studies indicate that the trans Golgi network (TGN), which comprises part of the Golgi outposts, can also function as a MT organizing center (MTOC) and influence MT organization. Although it is conceivable that Golgi outposts mislocalized to dynein mutant axons could alter MT polarity, expressing lava lamp dominant-negative, which prevents Golgi from associating with dynein without affecting dynein function, causes Golgi outposts to mislocalize to axons without altering MT orientation (Ye, 2007). Moreover, the change in axonal MT orientation is not likely to be simply a consequence of altered axon morphology because the loss of dynein function also alters the MT orientation of class I neuron axons, which appear relatively normal. With the current level of understanding, the model in which dynein acts as a 'gatekeeper' is most consistent with the results and the findings of others (Zheng, 2008).
A fundamental strategy for organising connections in the nervous system is the formation of neural maps. Map formation has been most intensively studied in sensory systems where the central arrangement of axon terminals reflects the distribution of sensory neuron cell bodies in the periphery or the sensory modality. This straightforward link between anatomy and function has facilitated tremendous progress in identifying cellular and molecular mechanisms that underpin map development. Much less is known about the way in which networks that underlie locomotion are organised. In the Drosophila embryo, dendrites of motorneurons form a neural map, being arranged topographically in the antero-posterior axis to represent the distribution of their target muscles in the periphery. However, the way in which a dendritic myotopic map forms has not been resolved and whether postsynaptic dendrites are involved in establishing sets of connections has been relatively little explored. This study shows that motorneurons also form a myotopic map in a second neuropile axis, with respect to the ventral midline, and they achieve this by targeting their dendrites to distinct medio-lateral territories. This map is 'hard-wired'; that is, it forms in the absence of excitatory synaptic inputs or when presynaptic terminals have been displaced. The midline signalling systems Slit/Robo and Netrin/Frazzled are the main molecular mechanisms that underlie dendritic targeting with respect to the midline. Robo and Frazzled are required cell-autonomously in motorneurons and the balance of their opposite actions determines the dendritic target territory. A quantitative analysis shows that dendritic morphology emerges as guidance cue receptors determine the distribution of the available dendrites, whose total length and branching frequency are specified by other cell intrinsic programmes. These results suggest that the formation of dendritic myotopic maps in response to midline guidance cues may be a conserved strategy for organising connections in motor systems. It is further proposed that sets of connections may be specified, at least to a degree, by global patterning systems that deliver pre- and postsynaptic partner terminals to common 'meeting regions' (Mauss, 2009).
How different dendritic morphologies and territories are generated in a motor system was investigated using the neuromuscular system of the Drosophila embryo as a model. Its principal components are segmentally repeated arrays of body wall muscles (30 per abdominal half segment), each innervated by a specific motorneuron. The motorneuron dendrites are the substrate on which connections with presynaptic cholinergic interneurons form. 180 cells (on average 11.25 for each identified motorneuron and a minimum of five) were labelled, and the dendritic morphologies and territories of the motorneurons that innervate the internal muscles were charted using retrograde labelling with the lipophilic tracer dyes 'DiI'and 'DiD.' This was done in the context of independent landmarks, a set of Fasciclin 2-positive axon bundles, at 18.5 h after egg laying (AEL), when the motor system first becomes robustly functional and the geometry of motorneuron dendritic trees has become sufficiently invariant to permit quantitative comparisons (Mauss, 2009).
Three classes of motorneurons were found based on dendritic arbor morphology and territory with respect to the ventral midline: (1) motorneurons with dendrites in the lateral neuropile (between the lateral and intermediate Fasciclin 2 tracts); (2) in the lateral and intermediate neuropile (between the intermediate and medial Fasciclin 2 tracts), and (3) in the lateral, intermediate plus medial neuropile (posterior commissure) (Mauss, 2009).
Moreover, the medio-lateral positions of motorneuron dendrites correlate with the dorsal to ventral locations of their target muscles in the periphery. Motorneurons with dorsal targets (DA1, DA3, DO1-5) have their dendrites in the lateral neuropile, while those innervating ventral and lateral muscles (LL1, VL2-4, VO1-2) also have dendrites in the intermediate neuropile. Coverage of the medial neuropile is particular to motorneurons innervating the most ventral group of muscles (VO3-6). These dendritic domains are arranged in the medio-lateral axis of the neuropile in such a way that they form a neural, myotopic representation of the distribution of body wall muscles in the periphery. Only a single motorneuron deviates from this clear-cut correlation between dendritic medio-lateral position and target muscle location: MN-DA2 has dendrites not only in the lateral neuropile, like other motorneurons with dorsal targets, but also in the intermediate neuropile (Mauss, 2009).
Previously studies have shown that motorneurons in the Drosophila embryo distribute their dendrites in distinct anterior to posterior domains in the neuropile, forming a central representation of target muscle positions in the periphery. The mechanisms required for the generation of this dendritic myotopic map remain elusive. In this study, dendritic myotopic organisation was characterized in a second dimension, with respect to the ventral midline, and the main molecular mechanism that underlies the formation of this dendritic neural map were identified, namely the combinatorial action of the midline signalling systems Slit/Robo and Netrin/Frazzled (Mauss, 2009).
Neural maps are manifestations of an organisational strategy commonly used by nervous systems to order synaptic connections. The view of these maps has been largely axonocentric and focused on sensory systems, though recent studies have challenged the notion of dendrites as a 'passive' party in arranging the distribution of connections. This study has demonstrated that motorneuron dendrites generate a neural, myotopic map in a motor system and that this manifest regularity can form independently of its presynaptic partner terminals (Mauss, 2009).
An essential feature of neural maps is the spatial segregation of synaptic connections. In the Drosophila embryonic nerve cord, there is some overlap between dendritic domains in the antero-posterior neuropile axis. Overlap of dendritic territories is also evident in the medio-lateral dimension, since all motorneurons have arborisations in the lateral neuropile, though distinctions arise by virtue of dendrites in additional intermediate and medial neuropile regions. The combination of myotopic mapping in both dimensions may serve to maximise the segregation between dendrites of different motorneuron groups. For example, the dendritic domain of motorneurons with dorsal targets differs from the territory innervated by ventrally projecting motorneurons in the antero-posterior location and the medio-lateral extent. Myotopic mapping in two dimensions could also provide a degree of flexibility that could facilitate wiring up in a combinatorial fashion. For instance, muscle LL1 lies at the interface between the dorsal and ventral muscle field; its motorneuron, MN-LL1, has one part of its dendritic arbor in the lateral domain that is characteristic for dorsally projecting motorneurons, while the other part of the dendritic tree innervates the intermediate neuropile precisely where ventrally projecting motorneurons put their dendrites (Mauss, 2009).
Myotopic dendritic maps might constitute a general organisational principle in motor systems. In insects, a comparable system of organisation has now been demonstrated also for the adult motor system of Drosophila (Brierley, 2009; Baek, 2009) and a degree of topographic organisation had previously been suggested for the dendrites of motorneurons that innervate the body wall muscles in the moth Manduca sexta. In vertebrates too, there is evidence that different motor pools elaborate their dendrites in distinct regions of the spinal cord in chick, turtle, and mouse. Moreover, elegant work in the mouse has shown that differences in dendritic territories correlate with and may determine the specificity of proprioceptive afferent inputs (Mauss, 2009 and references therein).
The neural map characterised in this study is composed of three morphological classes of motorneurons with dendrites innervating either (1) the lateral or (2) the lateral and intermediate or (3) the lateral, intermediate, and medial/midline neuropile (Mauss, 2009).
The motorneuron dendrites are targeted to these medio-lateral territories by the combinatorial, cell-autonomous actions of the midline guidance cue receptors Robo and Frazzled. The formation of dendritic territories by directed, targeted growth appears to be an important mechanism that may be more widespread than previously anticipated, though the underlying mechanisms may vary. Global patterning cues have been implicated in the vertebrate cortex (Sema3A). In the zebrafish retina, live imaging has shown that retinal ganglion cells put their dendrites into specific strata of the inner plexiform layer, but the roles of guidance cues and interactions with partner (amacrine) cells have not yet been studied (Mauss, 2009).
Slit/Robo and Netrin/Frazzled mediated gating of dendritic midline crossing has been previously documented in Drosophila embryos and zebrafish. This study demonstrated that dendrites are targeted to distinct medio-lateral territories by the combinatorial, opposing actions of Robo and Frazzled and that this is the main mechanism underlying the formation of the myotopic map. Strikingly, the same signalling pathways also regulate dendritic targeting of adult motorneurons in Drosophila, suggesting this to be a conserved mechanism (Brierley, 2009). Robo gates midline crossing of dendrites and in addition, at progressively higher signalling levels, restricts dendritic targeting to intermediate and lateral territories. Frazzled, on the other hand, is required for targeting dendrites towards the midline into intermediate and medial territories. The data argue that Frazzled is expressed by representatives of all three motorneuron types. Recently, Yang (2009) has shown that expression of frazzled leads to a concomitant transcriptional up-regulation of comm, thus linking Frazzled-mediated attraction to the midline with a decrease in Robo-mediated repulsion. While this has been demonstrated for midline crossing of axons in the Drosophila embryo, this study found that, at least until 18.5 h AEL, expression of UAS-frazzled alone was not sufficient to induce midline crossing of dendrites in MN-LL1 and MN-DA3. It is conceivable that differences in expression levels and/or timing between CQ-GAL4 used in this study and egl-GAL4 used by Yang might account for the differences in axonal and dendritic responses to UAS-frazzled expression. Moreover, the widespread expression of Frazzled in motorneurons and other cells in the CNS may point to additional functions, potentially synaptogenesis, as has been shown in C. elegans (Mauss, 2009).
Strikingly, neither synaptic excitatory activity nor the presynaptic (cholinergic) partner terminals seem to be necessary for the formation of the map. The map is already evident by 15 h AEL, before motorneurons receive synaptic inputs. It also forms in the absence of acetylcholine, the main (and at that stage probably exclusive) neurotransmitter to which motorneurons respond. Moreover, motorneuron dendrites innervate their characteristic dendritic domains when the cholinergic terminals have been displaced to outside the motor neuropile. However, interactions with presynaptic partners seem to contribute to its refinement. First, it was found that dendritic mistargeting phenotypes show a greater degree of penetrance earlier (15 h AEL) than later (18.5 h AEL) in development. Secondly, when interactions with presynaptic partner terminals are reduced or absent, dendritic arbor size increases and the distinction between dendritic territories is less evident than in controls. Fine-tuning of terminal arbors and sets of connections through contact and activity-dependent mechanisms is a well-established feature of neural maps in sensory systems and the current observations suggest that this may also apply to motor systems (Mauss, 2009).
The formation of the myotopic map is the product of dendritic targeting. It is therefore intimately linked with the question of how cell type-specific dendritic morphologies are specified. For instance, changing the balance between the Robo and Frazzled guidance receptors in motorneurons is sufficient to 'convert' dendritic morphologies from one type to another. The importance of target territories for determining dendritic arbor morphology has recently been explored in a study of lobula plate tangential cells in the blowfly, where the distinguishing parameter between the dendritic trees of four functionally defined neurons were not growth or branching characteristics but the regions where neurons put their dendrites (Mauss, 2009).
Because Slit/Robo and Netrin/Frazzled signalling have been reported to affect dendritic and axonal branching as well as axonal growth, respectively, it was asked what the effect was on motorneuron dendrites of altered Robo and Frazzled levels. It was found that in the wild-type different motorneurons generate characteristically different amounts of dendritic length and numbers of branch points (MN-DA1/aCC and MN-VO2/RP1, RP2, MN-DA3 and MN-LL1). In the Drosophila embryo and larva, Slit/Robo interactions have been suggested to promote the formation of dendrites and/or branching events, similar to what has been shown for cultured vertebrate neurons. The current data on embryonic motorneurons are not compatible with this interpretation. First, when altering the levels of Robo (or Frazzled) in individual motorneurons and mistargeting their dendrites, no statistically significant changes were detected in total dendritic length or number of branch points. Instead, for MN-DA3 and MN-LL1, it was observed that dendritic arbors respond to changes in the expression levels of midline cue receptors by altering the amount of dendritic length distributed to the medial, intermediate, and lateral neuropile. Secondly, in nerve cords entirely mutant for the Slit receptor Robo an increase is seen in dendrite branching at the midline. These observations suggest that for Drosophila motorneurons Slit/Robo interactions negatively regulate the establishment and branching of dendrites and thus specify dendritic target territories by defining 'exclusion' zones in the neuropile. The quantitative data from this study suggest that dendritic morphology is the product of two intrinsic, genetically separable programmes: one that specifies the total dendritic length to be generated and the frequency of branching; the other implements the distribution of these dendrites in the target territory, presumably by locally modulating rates of extension, stabilisation, and retraction of branches in response to extrinsic signals (Mauss, 2009).
The question of how neural circuits are generated remains at the heart of developmental neurobiology. At one extreme, one could envisage that every synapse was genetically specified, the product of an exquisitely choreographed sequence of cell-cell interactions. At the other extreme, neural networks might assemble through random cell-cell interactions and feedback processes enabling functional validation. The latter view supposes that neurons inherently generate polarised processes, have a high propensity to form synapses, and arrive at a favourable activity state through homeostatic mechanisms. Current evidence suggests that, at least for most systems, circuits form by a combination of genetic specification and the capacity to self-organise (Mauss, 2009).
This study has demonstrated that the postsynaptic structures of motorneurons, the dendrites, form a neural map. It was also shown that dendrites are closely apposed to cholinergic presynaptic specialisations in their target territories, suggesting that the segregation of dendrites may be a mechanism that facilitates the formation of specific sets of connections. Strikingly, this map of postsynaptic dendrites appears to be 'hard-wired' in that it can form independently of its presynaptic partners and it is generated in response to a third party, the midline guidance cues Slit and Netrin. A comparable example is the Drosophila antennal lobe, where projection neurons form a neural map independently of their presynaptic olfactory receptor neurons, though in this sensory system the nature and source of the cue(s) remain to be determined. This study complements previous work that demonstrated the positioning of presynaptic axon terminals by midline cues, also independently of their synaptic partners. Together, these results suggest that global patterning cues set up the functional architecture of the nervous system by independently directing pre- and postsynaptic partner terminals towards common 'meeting' areas (Mauss, 2009).
Clearly, such global guidance systems deliver a relatively coarse level of specificity and there is ample evidence for the existence of codes of cell-adhesion molecules and local receptor-ligand interactions capable of conferring a high degree of synaptic specificity. Therefore, one has to ask what the contribution is of global partitioning systems in establishing patterns of connections that lead to a functional neural network. A recent study in the Xenopus tadpole spinal cord has addressed this issue. Conducting patch clamp recordings from pairs of neurons, it has been found that the actual pattern of connections in the motor circuit reveals a remarkable lack of specificity. Furthermore, the segregation of axons and dendrites into a few broad domains appears to be sufficient to generate the connections that do form and to enable the emergence of a functional network. The implication is that neurons might be intrinsically promiscuous and that targeting nerve terminals to distinct territories by global patterning cues, as has been shown in this study, is important to restrict this synaptogenic potential and thereby confer a degree of specificity that is necessary for the emergence of network function (Mauss, 2009).
Dendrite arborization patterns are critical determinants of neuronal function. To explore the basis of transcriptional regulation in dendrite pattern formation, RNA interference (RNAi) was used to screen 730 transcriptional regulators and 78 genes involved in patterning the stereotyped dendritic arbors of class I da neurons were identified in Drosophila. Most of these transcriptional regulators affect dendrite morphology without altering the number of class I dendrite arborization (da) neurons and fall primarily into three groups. Group A genes control both primary dendrite extension and lateral branching, hence the overall dendritic field. Nineteen genes within group A act to increase arborization, whereas 20 other genes restrict dendritic coverage. Group B genes appear to balance dendritic outgrowth and branching. Nineteen group B genes function to promote branching rather than outgrowth, and two others have the opposite effects. Finally, 10 group C genes are critical for the routing of the dendritic arbors of individual class I da neurons. Thus, multiple genetic programs operate to calibrate dendritic coverage, to coordinate the elaboration of primary versus secondary branches, and to lay out these dendritic branches in the proper orientation (Parrish, 2006; Full text of article).
To assay for the stereotyped dendrite arborization pattern of class I da neurons (hereafter referred to as class I neurons) in RNAi-based analysis of dendrite development, a Gal4 enhancer trap line (Gal4221) was used that is highly expressed in class I neurons and weakly expressed in class IV neurons during embryogenesis. Because of the simple and stereotyped dendritic arborization patterns of the dorsally located ddaD and ddaE, the studies of dendrite development focused on these two dorsally located class I neurons (Parrish, 2006).
To establish that RNAi is an efficient method to systematically study dendrite development in the Drosophila embryonic PNS, it was demonstrated that injecting embryos with double-stranded RNA (dsRNA) for green fluorescent protein (gfp) is sufficient to attenuate Gal-4221-driven expression of an mCD8::GFP fusion protein as measured by confocal microscopy. Next whether RNAi could efficiently phenocopy loss-of-function mutants known to affect dendrite development was tested. Similar to the mutant phenotype of short stop (shot), which encodes an actin/microtubule cross-linking protein, shot(RNAi) caused routing defects, dorsal overextension, and a reduction in lateral branching of dorsally extended primary dendrites. Likewise, RNAi of sequoia or flamingo resulted in overextension of ddaD and ddaE, RNAi of hamlet resulted in supernumerary class I neurons, and RNAi of tumbleweed resulted in supernumerary class I neurons and a range of arborization defects, consistent with the reported mutant phenotypes. Thus, RNAi is effective in generating reduction of function phenotypes in embryonic class I dendrites (Parrish, 2006).
Group A TFs that promote arborization through concerted regulation of outgrowth and branching
A group of transcriptional regulators, group A, controls the size of the dendritic field of class I neurons. RNAi of 19 TFs resulted in reduction of the field size covered by ddaD and ddaE. A reduction of coverage could be the result of a net reduction in dendrite outgrowth, branching, or both. Group A TFs have effects on both primary dendrite growth and secondary dendrite growth. For example, RNAi of the PAS-domain TF trachealess (trh) caused a minor reduction in both primary branch outgrowth and the number of lateral branches and a more marked reduction in the overall length of lateral branches. Consequently, the most distal regions of the dendritic field, especially the regions covered by lateral branches, are not innervated. By contrast, RNAi of genes such as the zinc-finger TF pygopus or the BTB/POZ-domain TF cg1841 caused more severe reduction of primary branch outgrowth as well as lateral branching and lateral branch length, resulting in a more drastic reduction of receptive field. In an extreme case, RNAi of the high mobility group gene hmgD resulted in an almost complete block of primary dendrite extension and lateral branching. In general, the genes with the most severe effects on primary branch outgrowth also have the most severe effects on branching, suggesting that these genes may function to regulate dendritic arborization overall (Parrish, 2006).
Although the genes in this class all caused qualitatively similar defects in arborization, some notable phenotypic differences are suggestive of distinct functions for some of these genes in regulating dendrite arborization. RNAi of the nuclear hormone receptors ultraspiracle (usp) and ecdysone receptor (EcR) significantly reduced primary dendrite outgrowth, but caused only modest reduction of lateral branching and lateral branch outgrowth, suggesting that branching is not absolutely dependent on proper outgrowth. Since the Usp/EcR heterodimer is responsible for ecdysone-responsive activation of transcription, as well as ligand-independent transcriptional repression, it is likely that these genes function together to promote dendrite outgrowth (Parrish, 2006).
RNAi of many group A genes resulted in embryonic lethality at a significantly higher rate than control injections. Thus, many of these genes are likely essential for embryonic development, either due to their involvement in regulating neuronal morphogenesis or due to other aspects of their functions (Parrish, 2006).
Group A transcriptional regulators that restrict dendrite arborization
In addition to genes with functions in promoting dendrite arborization, 20 group A genes were identified that regulate dendrite arborization by limiting dendrite growth and/or branching. Consistent with recent reports that loss of function of the BTB/POZ domain TF abrupt (ab) causes an increase in dendritic branching and altered distribution of branches, it was found that ab(RNAi) altered the arborization of class I dendrites. ab(RNAi) caused an increase in the number and length of lateral branches, expanding the coverage field most noticeably along the anteroposterior (AP) axis. In addition to these defects, ab(RNAi) also caused frequent cell death, consistent with the phenotype observed for a hypomorphic allele of ab (Parrish, 2006).
Increased dendritic branching also resulted from RNAi of several genes known to affect nervous system development, including Adh transcription factor 1 (Adf1), the zinc finger TF nervy (nvy), the basic helix–loop–helix (bHLH) TF deadpan (dpn), as well as genes not previously known to affect neuronal function, such as the putative transcription elongation factor Elongin c. Both Adf1 and dpn mutants have defects in larval locomotion and, in light of recent findings suggesting that da neurons may regulate aspects of larval locomotion, it is possible that dendrite defects underlie these behavioral defects. Consistent with its role in class I dendrite development, dpn is expressed in all PNS neurons. Likewise, nervy has been implicated in regulation of axon branching in motorneurons and is apparently expressed in most neurons. Thus, nervy likely regulates multiple aspects of neuronal differentiation. Finally, Elongin C may regulate transcriptional elongation but also likely functions as a component of a multimeric protein complex that includes the von Hippel-Lindau (VHL) tumor suppressor and targets specific proteins for poly-ubiquitination and degradation. Moreover, BTB/POZ domain proteins (such as cg1841 and ab) function as substrate adaptors for cullin E3 ligases. Interestingly, RNAi of a Drosophila homolog (tango) of a known VHL substrate (HIF-1) also affected dendrite arborization. It thus appears that protein degradation pathways regulate dendrite arborization (Parrish, 2006).
RNAi of the Polycomb group (PcG) genes Su(z)12, E(z), esc, or Caf1 similarly caused an increase in branch number and an expansion of the receptive field of class I neurons. Consistent with the similar RNAi phenotypes for these genes, Su(z)12, E(z), esc, and Caf1 are components of the multiprotein esc/E(z) polycomb repressor complex. One critical role for PcG-mediated gene silencing is the regulation of hox gene expression. Therefore, Polycomb-mediated regulation of hox gene expression likely contributes to arborization of class I neurons (Parrish, 2006).
RNAi of several genes affected dendrite arborization primarily by causing an increase in dorsal and lateral dendrite extension without significantly affecting branch number. For example, RNAi of the putative transcriptional repressor cg5684 caused dorsal overextension of the primary dendrite in ddaE and an overall increase in dendritic length in both ddaD and ddaE. In general, RNAi of genes that increased arborization rarely caused dendrites to cross the dorsal midline or segment borders, or increased branching more than twofold as compared with untreated neurons. It thus appears that dendritic outgrowth is further limited by neuronal growth capacity and/or other external constraints (Parrish, 2006).
Group B TFs with opposing actions on dendrite outgrowth and branching shape dendrite arbors
In contrast to the genes that coordinately affect dorsal dendrite outgrowth and lateral branching/outgrowth, a group of 21 genes (group B) were identified that have opposing effects on dendrite outgrowth and branching, suggesting that dendrite outgrowth and branching might partially antagonize one another. RNAi of 19 of these genes resulted in dorsal overextension of primary dendrites and a reduction in lateral branching/lateral branch extension. In the most severe cases, such as RNAi of the transcriptional repressor snail, dorsal overextension of almost completely unbranched dendrites was found. Like snail(RNAi), RNAi of the nuclear hormone receptor knirps, the transcriptional repressor l(3)mbt, as well as 15 other genes, all caused dorsal overextension of primary dendrites. As in the case of genes that normally limit arborization, RNAi of these genes rarely caused dendrites to cross the dorsal midline (Parrish, 2006).
In addition to the effects on primary dendrite extension, RNAi of each of these 18 genes limits the number and length of lateral dendrite branches. RNAi of some genes such as snail or knirps almost completely blocked dendrite branching, whereas RNAi of other genes such l(3)mbt had more modest effects on dendrite branching. In addition, a significant reduction of branching was noticed at the distal tip of the dorsally projected primary dendrite. In control treated stage 17 embryos, branchpoints are distributed along the primary dendrite, with the most distal branchpoint usually located within a few microns of the distal tip of the dendrite. In contrast, branching is rarely observed within 10 microns of the distal dendritic tip following RNAi of these group B genes. In some cases, such as snail(RNAi), knirps(RNAi), or l(3)mbt(RNAi), the most distal branchpoint is located 25 microns or further from the distal tip of the primary dendrite. Therefore, these TFs inhibit primary branch extension but promote lateral branching and lateral branch extension (Parrish, 2006).
In addition to identifying a large class of TFs that inhibit primary branch extension and promote lateral branching, it was also found that TFs promote dendrite extension and limit dendrite branching. RNAi of two genes, glial cells missing 2 (gcm2) and the histone acetyltransferase pcaf, caused an increase in lateral branching and a marked reduction in dorsal extension of ddaE. Thus, transcriptional pathways exist that have opposing effects on primary branch outgrowth and secondary branching, suggesting that these processes may normally antagonize one another (Parrish, 2006).
Group C TFs regulate dendrite routing
Proper dendritic routing is important for primary dendrites of ddaD and ddaE to grow in parallel toward the dorsal midline without crossing each other and for secondary branches of ddaD and ddaE to avoid the space between ddaD and ddaE. Therefore, there must be mechanisms that promote this stereotyped arborization pattern, including signals that promote anterior arborization of ddaD and posterior arborization of ddaE, as well as signals that antagonize posterior arborization of ddaD and anterior arborization of ddaE. Indeed, RNAi of 10 TFs disrupted the dendritic routing patterns of ddaD and ddaE, resulting in aberrantly oriented primary dendrites. RNAi of cg1244, bap55 (brahma associated protein of 55kD), cg9104, cg4328, and cg7417 resulted in inappropriate anterior arborization of ddaE as well as inappropriate posterior arborization of ddaD. Anterior or even ventral displacement of ddaD concomitant with anterior arborization of ddaE was also observed as well as displacement of ddaE arbors concomitant with misrouting of ddaD. Finally, reducing sens function by RNAi or genetic mutation caused extensive mixing of dendritic arbors from ddaD and ddaE, in addition to dorsal overextension of primary dendrites and an overall reduction in the number of class I neurons (Parrish, 2006).
It is also worth noting that RNAi treatment that caused reduced dendritic outgrowth often caused minor routing defects. For example, pyg(RNAi) or cg1841(RNAi) caused inappropriate routing of ddaE. The routing defects seen with these candidates may reflect a disruption of attractive/repulsive signaling that normally regulates dendrite arborization. The source of such signals is currently unknown, although of great interest (Parrish, 2006).
TFs likely exert distinct mitotic and post-mitotic functions to regulate neuron morphogenesis
TFs play critical roles in neurogenesis, and some genes that regulate neurogenesis also affect post-mitotic neuronal differentiation. Because clones of duplicated class I neurons have wild-type dendrite arborization patterns, class I dendritic arbors appear to be insensitive to cell number defects. Indeed, dendrite arborization of class I neurons in embryos carrying the temperature-sensitive neurogenic mutation Notchts (Nts) is unaffected by as much as a fivefold increase in class I neuron number, and is likely insensitive to multiplication of other da neurons as well since Nts experiments caused increased numbers of other da neurons. Furthermore, in cases where only one of the class I neurons is multiplied, the dendrites of neighboring class I neuron are unaffected. In contrast laser ablation of ddaD or ddaE or the occasional cell loss caused by RNAi of various genes did not generally cause defects in arborization of neighboring class I neurons. Therefore, analysis of class I neurons should allow study of post-mitotic functions of genes that affect neuron number (Parrish, 2006).
RNAi of several genes affected the number of class I neurons as well as morphogenesis of class I dendrites; RNAi of seven genes caused supernumerary cells and RNAi of four genes caused high penetrance cell loss in addition to dendrite defects. For example, RNAi of the zinc finger TF nerfin-1 caused an increase in neurons labeled by Gal4221 with as many as eight neurons visible in some segments. Unlike wild-type class I neurons, neurons from nerfin-1(RNAi)-treated embryos extended mostly unbranched dendrites. In many cases, the routing pattern of the dendrites appeared abnormal, but the cell number defects make it difficult to resolve the projection pattern of individual dendrites or conclusively determine whether each neuron projects the same number of primary dendrites. RNAi of six other genes, including jumeau, a winged-helix TF known to regulate neuroblast cell fate and the number of PNS neurons, similarly caused an increase in neuronal number as well as defects in dendrite morphogenesis (Parrish, 2006).
In addition to the seven genes that function to restrict class I neuron number and control dendrite morphology, three other genes are required to maintain the number of class I neurons. Reduction of their function caused a reduction of class I neurons and defects in dendrite morphogenesis in the remaining neurons. For example, RNAi of the zinc finger TF senseless (sens) reduced the number of class I neurons, consistent with previous findings that sens is required for development of most cells in the PNS. In addition, sens(RNAi) or a sens loss-of-function mutation caused an increase in dendrite outgrowth and mixing of dendrites in segments with both ddaD and ddaE present. Similarly, RNAi of the proneural bHLH TF atonal (ato) reduced the number of class I neurons, consistent with previous findings that chordotonal organs and some md neurons are absent in embryos lacking ato. Consistent with reports that ato functions in neurite arborization in the larval brain, it was also found that ato(RNAi) caused altered arborization patterns of class I dendrites. Thus, it is likely that multiple TFs that regulate neuron number also regulate aspects of post-mitotic neuronal differentiation (Parrish, 2006).
Finally, gene was found that appears to be essential for a subset of class I da neurons. RNAi of cubitus interruptus (ci), a component of the hedgehog signaling pathway, caused a loss of dorsal class I da neurons without affecting the ventral class I neuron vpda. Because ci(RNAi) causes embryonic lethality at high concentrations of dsRNA, the phenotypes in surviving embryos likely represent hypomorphic phenotypes. Loss of ddaD and ddaE is not compensated for by a concomitant increase in other md neurons since ci(RNAi) leads to an overall reduction of dorsal md neurons expressing the pan-da marker Gal4109(2)80. Therefore, ci likely promotes differentiation or survival of a subset of da neurons, including ddaD and ddaE, but not of the ventral class I neuron vpda, demonstrating that morphologically similar neurons can be molecularly distinct (Parrish, 2006).
Some TFs are continuously required to regulate class I dendrite morphology
As another indication of the hypomorphic nature of many of the alleles and maternal rescue of gene function in mutant embryos, focus was place on dendrite defects that were first apparent during larval stages. For example, a mutant allele of Drosophila Mi-2, which encodes a Hunchback-interacting ATP-dependant chromatin remodeling factor, shows only minor defects in late embryonic stages, but shows an obvious reduction in arborization by 72 h after egg laying. Since Mi-2(RNAi) demonstrates that Mi-2 is required for embryonic dendrite arborization, these findings suggest that Mi-2 is continuously required for class I neurons to maintain proper dendrite arborization patterns. Similarly, the dendritic overbranching associated with a P-element insertion allele of Adf1 was first apparent after embryonic stages, although Adf1(RNAi) caused overbranching in embryos. Class I dendritic arbors of Adf1 mutants are indistinguishable from wild-type neurons until 96 h AEL. By 144 h AEL, ddaE arbors of Adf1 mutants showed a greater than twofold increase in branch number when compared with time-matched wild-type controls. Interestingly, ddaD showed only very minor branching defects in Adf1 mutants, suggesting that ddaD and ddaE might have distinct requirements for Adf1. Similarly, mutant alleles of either E(bx) or Elongin C showed dendrite branching defects only at late larval stages. These findings indicate that Adf1, E(bx), and Elongin C are continuously required to inhibit branching in class I neurons, demonstrating that although class I neurons have very little new branching after embryogenesis, they still retain the capacity to branch (Parrish, 2006).
RNAi of some group A genes that cause reduced dendrite outgrowth and branching is epistatic to mutation of ab or sens
Since group A and B TFs regulate aspects of dendritic growth and branching, potential epistatic relationships among TFs was explored in these phenotypic classes. To do this, RNAi was used to knockdown expression of select TFs in Drosophila embryos carrying a loss-of-function mutation in either the group B/C gene senseless (sens) or the group A gene abrupt (ab). sens mutant class I dendrites overextend dorsally and have reduced lateral branching in addition to routing defects. In sens mutants, RNAi of the group A genes Su(z)12 and ab, which cause increased lateral branching following RNAi in wild-type embryos, led to an increase in lateral branching compared with injected controls. Therefore, Su(z)12 and ab function are still required to limit arborization in sens mutants, and the increased dendritic branching as a result of Su(z)12(RNAi) or ab(RNAi) is epistatic to the increased dorsal extension and reduced lateral branching of sens mutants. In contrast, RNAi of the group A genes cg1244 and cg1841, which caused reduced arborization following RNAi in wild-type embryos, led to a reduction in primary dendrite outgrowth and lateral dendrite branching compared with injected controls. Therefore, at least in the instances described above, loss of group A genes is epistatic to loss of group B genes (Parrish, 2006).
RNAi of group A genes either promoted or antagonized dendrite arborization; therefore, the effect was examined of simultaneously disrupting one group A gene that promoted and one group A gene that antagonized dendrite outgrowth and lateral branching. RNAi or a loss-of-function mutant of the group A gene ab caused increased dendritic branching and extension of class I dendrites. In addition, mutation of ab caused a significant reduction in the number of class I neurons labeled by Gal4221 that was most pronounced in the dorsal cluster of PNS neurons, consistent with the results from RNAi experiments. To facilitate epistasis analysis in ab mutants, dendrite arborization effects in vpda, the ventrally located class I neuron, were assayed. RNAi of the group A gene hmgD, which caused reduced primary dendrite outgrowth and reduced lateral branching when injected into wild-type embryos, caused a striking reduction in the number of dendritic branches and size of the receptive field of vpda in ab mutants. RNAi of the group A gene bap55 had similar effects in ab mutants, demonstrating that, at least in some cases, loss of group A genes that results in reduced arborization is epistatic to loss of group A genes that results in increased arborization. Therefore it is possible that the different classes of group A genes antagonistically regulate a common set of target genes required for dendrite arborization (Parrish, 2006).
Dendrites exhibit a wide range of morphological diversity, and their arborization patterns are critical determinants of proper neural connectivity. How different neurons acquire their distinct dendritic branching patterns during development is not well understood. This study reports that Spineless (Ss), the Drosophila homolog of the mammalian aryl hydrocarbon (dioxin) receptor (Ahr), regulates dendrite diversity in the dendritic arborization (da) sensory neurons. In loss-of-function ss mutants, class I and II da neurons, which are normally characterized by their simple dendrite morphologies, elaborate more complex arbors, whereas the normally complex class III and IV da neurons develop simpler dendritic arbors. Consequently, different classes of da neurons elaborate dendrites with similar morphologies. In its control of dendritic diversity among da neurons, ss likely acts independently of its known cofactor tango and through a regulatory program distinct from those involving cut and abrupt. These findings suggest that one evolutionarily conserved role for Ahr in neuronal development concerns the diversification of dendrite morphology (Kim, 2006).
The ss protein is present at nearly the same level in all da neurons and acts cell-autonomously to dictate their dendritic complexity, while different da neurons exhibit different sensitivity to the level of Ss, and even the bipolar td neuron can respond to elevated ss activity by increasing dendritic complexity (Kim, 2006).
Previous studies in C. elegans have demonstrated essential roles for invertebrate homologs of Ahr in neuronal cell fate determination. For example, ahr-1 regulates the differentiation program of a subclass of neurons that contact the pseudocoelomic fluid, and both ahr-1 and aha-1 specify GABAergic neuron cell fate in C. elegans. The dramatic changes in the dendrite morphologies of the da neurons, however, are not due to an all-or-nothing change in cell fate because the da neurons in ss mutants displayed normal class-specific expression patterns of the molecular markers Ab and Cut and normal axon projection patterns characteristic of individual da neurons. However, this also does not assume that a partial cell fate change has not occurred. One reflection of the ss function as a transcription factor is the altered expression levels of GFP in the class I Gal4221 reporter, with increased levels of expression in all class IV neurons and essentially no expression in the dorsal class I neuron ddaD and the ventral class I neuron vpda in ss mutants. It will be of interest to further characterize the genetic basis for this Gal4 reporter, to determine whether this regulation constitutes a partial cell fate alteration or transcriptional regulation of genes downstream from ss in the execution of adjustment of dendritic complexity (Kim, 2006).
There is an emerging theme that ss functions to diversify
neuronal differentiation by expanding the photopigment repertoire of R7 photoreceptors in the Drosophila eye and by diversifying da neuron dendritic morphologies. Recent studies have demonstrated that the entire retinal mosaic pattern required for color vision in Drosophila is regulated by ss. In the Drosophila retina, two types of ommatidia form the wild-type retinal mosaic: 'pale' and 'yellow.' In ssD115.7 mutants, the yellow ommatidial subtype is lost and normally yellow R7 cells are misspecified into the pale subtype. As a result, nearly all R7 cells adopt the pale subtype, leading to loss of the retinal mosaic pattern. Thus, the pale R7 subtype represents the R7 'default state' (Kim, 2006 and references therein).
The overall lack of dendritic diversity in the da neurons
in ss mutants is suggestive of the hypothesis that ss, an ancient, evolutionarily conserved gene, may act to convert a primordial dendrite pattern (perhaps a default state) to different complexities for different neurons in the peripheral nervous system. The loss-of-function ss phenotype in the da neuron dendrites might reflect such a primordial pattern as the dendrites in the mutant are devoid of specific morphological features that define distinct neuronal subclasses. In support of this notion, dendrites of the different classes of da neurons share similar morphological characteristics and elaborate similar numbers of total branches in ss mutants. The ability of ss to regulate the complexity and diversity of this dendrite pattern, by limiting dendritic branching to shape the simpler arbors of the class I and class II neurons and by promoting class-specific terminal branching to shape the more complex arbors of the class III and class IV neurons, is quite unique. Of the many mutants that affect multiple classes of da neurons, the great majority affect da neurons with simple or complex dendritic arbors the same way; that is, causing them to all become simpler or more complex. The ss phenotype of making simple dendritic arbors more complex and complex arbors simpler is very unusual among the many mutants affecting dendrite complexity. It thus seems likely that the distinct dendritic patterns rely not only on a cohort of gene activities specifying the mechanics of dendrite outgrowth and branching, but also a genetic program that diverts the generic primordial mode of dendritic formation to a diverse range of dendritic patterns (Kim, 2006).
How might spineless exert its functions? Unlike the homeodomain protein Cut, which promotes dendritic complexity in a specific direction, ss functions in an opposing manner in different cell types to regulate dendritic diversity. How might ss function differently in different neuronal cell types? One possibility is that ss is activated by different ligands in different neurons. ss is incapable of binding dioxin and other related compounds, suggesting that other, as yet unidentified ligands are required for its activation. Previous reports have suggested that ss and other invertebrate homologs of Ahr are activated by an endogenous ligand or that no ligand is required at all. Recent studies have shown that Ahr can accumulate in the nucleus upon activation by the second messenger cyclic AMP (cAMP), although it is not yet known whether cAMP signaling can activate ss in Drosophila. Thus, it is conceivable that ss is activated by different upstream factors in different cell types. It will be of interest to test in future studies whether in different neuronal cell types ss is activated by different ligands or upstream second messengers and whether ss acts in concert with regulatory programs for cell fate specification to dictate dendritic complexity (Kim, 2006).
In the canonical Ahr signaling pathway, Ahr requires
the appropriate cofactor for its proper function. Members of the bHLH-PAS protein family are able to heterodimerize with other bHLH-PAS proteins. Previous studies have shown that, upon ligand binding, Ahr is translocated to the nucleus, where it heterodimerizes with Arnt to form a transcriptionally active complex. However, tango, the Drosophila homolog of Arnt, is likely not required for the regulation of dendritic morphogenesis, indicating that ss is probably not acting through its canonical signaling pathway to specify dendritic complexity. In Sf9 cells, ss can act independently of tgo to enhance expression of a reporter in the absence of a ligand. Furthermore, Ahr is unable to interact with Arnt upon activation by cAMP. Although Ahr, Arnt, and the Arnt homolog Arnt2 are widely distributed throughout the rat brain, Ahr does not preferentially colocalize with either Arnt or Arnt2. Ahr is also expressed in specific regions of the rat brain where neither Arnt nor Arnt2 is expressed. These studies support the notion that ss can act independently of tgo in certain developmental contexts. Tgo can heterodimerize with other bHLH-PAS proteins in addition to ss. It is conceivable that ss may act with different heterodimerization partners to mediate its different functions in different cell types (Kim, 2006).
Ubiquitin-proteasome system (UPS) is a multistep protein degradation machinery implicated in many diseases. In the nervous system, UPS regulates remodeling and degradation of neuronal processes and is linked to Wallerian axonal degeneration, though the ubiquitin ligases that confer substrate specificity remain unknown. Having shown previously that class IV dendritic arborization (C4da) sensory neurons in Drosophila undergo UPS-mediated dendritic pruning during metamorphosis, an E2/E3 ubiquitinating enzyme mutant screen was conducted, revealing that mutation in ubcD1, an E2 ubiquitin-conjugating enzyme encoding Effete, resulted in retention of C4da neuron dendrites during metamorphosis. Further, UPS activation likely leads to UbcD1-mediated degradation of DIAP1, a caspase-antagonizing E3 ligase. This allows for local activation of the Dronc caspase, thereby preserving C4da neurons while severing their dendrites. Thus, in addition to uncovering E2/E3 ubiquitinating enzymes for dendrite pruning, this study provides a mechanistic link between UPS and the apoptotic machinery in regulating neuronal process remodeling (Kuo, 2006).
The ubiquitin-proteasome system (UPS), evolutionarily conserved for the regulation of protein turnover, targets proteins for degradation via a complex, temporally regulated process that results in proteasome-mediated destruction of polyubiquitinated proteins. There are two distinct steps involved: first, the covalent conjugation of ubiquitin polypeptide to the protein substrates, and second, the destruction of tagged proteins in the proteasome complex. The transfer of ubiquitin to a target molecule slated for degradation involves at least three enzymatic modifications: ubiquitin is first activated by the ubiquitin-activating enzyme E1; ubiquitin is then transferred to a carrier protein, a ubiquitin-conjugating enzyme E2, and finally, ubiquitin is transferred to a protein substrate bound by a ubiquitin ligase E3. There are minor variations to this enzymatic cascade, but overall, these highly specific protein-protein interactions ensure ubiquitin targeting specificity and regulate many aspects of housekeeping protein turnover and cellular maintenance. However, with the multiple regulatory layers, different parts of this complex machinery can break down. Mutations in the UPS pathway causing accumulation of nondegraded proteins have been implicated in a variety of human diseases (Kuo, 2006).
In the nervous system, aberrations in the UPS pathway have been implicated in disorders such as Alzheimer's disease, Parkinson's disease, amyotrophic lateral sclerosis, and other neurodegenerative diseases. One of the common pathological features of neurodegenerative diseases, besides neuronal loss, is local axon degeneration. For example, in the case of Wallerian degeneration in vertebrates, distal parts of a severed axon remain viable and conduct action potentials in vivo for some time before a rapid dismantling of cytoskeletal proteins and axon degeneration, and the initiation of this rapid axon degeneration involves the UPS pathway. It is thought that UPS activation can lead to microtubule depolymerization and subsequent neurofilament degradation, possibly acting in conjunction with the Ca2+-dependent protease calpain. Moreover, inhibiting UPS activity in neurons prior to severing their axons can dramatically retard degradation of the severed axons. These results suggest that a cell-intrinsic UPS pathway regulates axon stability and that pharmaceutical inactivation of the UPS may prevent axonal degeneration in disease states (Kuo, 2006 and references therein).
In Drosophila, the remodeling of neuronal processes during normal development closely resembles the pathological phenotypes in Wallerian degeneration. In the mushroom body γ neurons, extensive pruning of larval axons occurs during metamorphosis in a process regulated by glia engulfment and neuron-intrinsic UPS activity. Similarly, in the fly peripheral nervous system, the class IV dendritic arborization (C4da) neurons undergo complete pruning of their extensive larval dendrites during metamorphosis, in a process that is also regulated by UPS activity (Kuo, 2005). In both of these examples, severing of neuronal processes is preceded by microtubule depolymerization and followed by cytoplasmic blebbing and degeneration, all phenotypes resembling Wallerian degeneration. Therefore, these fly neurons represent excellent systems in which to understand the roles of the UPS in regulating neuronal axon/dendrite integrity, given the rather limited knowledge of how the UPS participates in the degradation of neuronal processes. It is not known which specific E2 ubiquitin-conjugating enzyme(s) and E3 ubiquitin ligase(s) are involved in UPS-mediated remodeling/degradation of neuronal processes, or their specific downstream target(s) (Kuo, 2006).
It has been shown that mutations in the fly ubiquitin activation enzyme (uba1) and the proteasome complex (mov34) can prevent efficient pruning of C4da neuron larval dendrites during metamorphosis (Kuo, 2005). To further investigate the role of UPS in C4da neuron dendrite remodeling, a candidate gene screen was conducted to identify the E2 ubiquitin-conjugating enzyme and the E3 ubiquitin ligase required for this process. Analysis of genetic mutants showed that UPS activation in C4da neurons likely results in UbcD1 (an E2 ubiquitin-conjugating enzyme) mediated degradation of Drosophila inhibitor of apoptosis protein 1 (DIAP1), an E3 ligase that antagonizes caspase activity. Degradation of DIAP1 leads to activation of caspase Dronc, which results in local caspase activation and cleavage of proximal dendrites in C4da neurons during metamorphosis. In addition to the identification of a set of E2/E3 ubiquitinating enzymes for C4da neuron dendrite remodeling—with the surprising finding that the UPS mediates degradation of the potent protease inhibitor DIAP1—this study also establishes a mechanistic link between the UPS and caspase pathways in regulating C4da neuron dendrite pruning (Kuo, 2006).
To identify the E2 ubiquitin-conjugating enzyme and E3 ubiquitin ligase mediating dendrite pruning of C4da neurons during metamorphosis, a candidate gene approach was taken to systematically test the roles of known E2/E3 ubiquitinating enzymes in Drosophila. A set of putative E2/E3 ubiquitinating enzyme mutations was assembled, and live imaging was used to visualize C4da neurons carrying the E2/E3 mutation via the pickpocket(ppk)-EGFP marker, which specifically labels C4da neurons during Drosophila development. Those mutants with an early lethal phase were characterized by generating mosaic analysis with a repressible cell marker (MARCM) mutant neuronal clones. Since wild-type (wt) C4da neurons during metamorphosis do not retain any larval dendrites following head eversion, as imaged 18–20 hr after puparium formation (APF), mutations that caused larval dendrite retention in C4da neurons at this stage were sought. The candidate genes tested mostly showed no defects in dendrite pruning or neuronal cell death. However, one candidate, ubcD1, showed a modest level of larval dendrite retention at 18 hr APF (Kuo, 2006).
Live imaging of wt C4da neuron MARCM clones at the start of pupariation (white pupae, WP) showed primary and secondary dendritic branching patterns typical of C4da neurons. Consistent with previous reports (Kuo, 2005; Williams, 2005), wt C4da neurons sever their larval dendrites during early metamorphosis and by 18 hr APF are devoid of any dendrites. The ubcD1 mutant C4da MARCM clones showed similar dendritic morphology to the wt clones at the onset of metamorphosis. However, at 18 hr APF, the mutant clones consistently retained intact, nonsevered larval dendrites. Thus, the UbcD1 E2 ubiquitin-conjugating enzyme is required for proper UPS-mediated dendrite pruning in C4da neurons during metamorphosis (Kuo, 2006).
UbcD1, encoded by the gene effete, regulates UPS-mediated degradation of the antiapoptotic protein DIAP1 (Treier, 1992; Wang, 1999; Ryoo, 2002). In protecting cells from apoptosis, the DIAP1 E3 ubiquitin ligase antagonizes Dronc caspase activity by regulating ubiquination and degradation of the Dronc protein. Following apoptotic stimuli, UbcD1 mediates self-ubiquination and degradation of DIAP1, allowing for subsequent Dronc caspase activation. The biochemical and genetic interactions between these molecules are well established. The baculovirus p35, which is commonly used to inhibit caspase activity in Drosophila, and does not block C4da neuron dendrite pruning (Kuo, 2005). This may seem to make the involvement of caspases in this process unlikely; however, p35 has only limited activity against the caspase Dronc. To study the effects of dronc mutation on C4da neuron dendrite pruning, two null alleles of Dronc, dronc51 and dronc11, were used. MARCM analysis of dronc mutant clones revealed that the dendrites of mutant C4da neurons appeared normal at larval stages. However, unlike wt clones, without Dronc these neurons failed to properly prune their larval dendrites during metamorphosis, and most showed relatively intact primary and secondary larval dendritic arbors at 18 hr APF. These results show that severing of primary larval dendrites from C4da neurons during early metamorphosis requires the Dronc caspase (Kuo, 2006).
During apoptosis, Dronc activation requires the degradation of the antiapoptotic/anticaspase protein DIAP1, which is downstream of UbcD1. The requirement of UbcD1 for C4da neuron larval dendrite pruning during metamorphosis, together with the finding that Dronc caspase activity is also essential, raised the question of whether UPS-mediated DIAP1 degradation is a key step that allows for the severing of larval dendrites. Because loss of DIAP1 function causes C4da neuron cell death prior to the onset of metamorphosis, this question was approached using a gain-of-function allele of diap1, diap16-3s, which has a single amino acid mutation that makes DIAP1 an inefficient substrate for UPS-mediated degradation. ppk-EGFP was crossed into the gain-of-function mutant background and live imaging was used to follow C4da neuron dendrite pruning during metamorphosis. The diap16-3s mutation did not significantly affect the ability of C4da neurons to elaborate larval dendrites. However, unlike wt C4da neurons that completely pruned their larval dendrites by 18 hr APF, C4da neurons in the diap16-3s gain-of-function mutants failed to efficiently sever larval dendrites at 18 hr APF. These results suggest that the degradation of DIAP1 during early metamorphosis is required for proper C4da neuron larval dendrite pruning. Quantitatively, mutations in the UPS pathway that modulate Dronc activity (diap16-3s and ubcD1) resulted in less severe dendrite pruning defects than dronc mutants, both in terms of total number of large dendrites attached to soma and in the length of the longest attached dendrite at 18 hr APF (Kuo, 2006).
The UbcD1-DIAP1-Dronc pathway in apoptosis is well established. Thus, it may be necessary for C4da neurons to restrict the action of this pathway to specific cellular locations in order to prune unwanted dendrites without triggering apoptosis. To address this possibility, the subcellular distribution of DIAP1 and Dronc proteins was examined in ppk-EGFP C4da neurons. During the transition from third instar larvae to white pupae at the onset of metamorphosis, as well as 2 hr APF, there was a consistent induction of nuclear DIAP1 in GFP-labeled C4da neurons. During the same period a concurrent decrease was detected in Dronc staining in the soma of C4da neurons, unlike those from the neighboring cells at 2 hr APF. These results are consistent with previous observations that C4da neurons survive through this stage of metamorphosis. However, the level of antibody staining made it difficult to monitor the distribution of DIAP1 and Dronc within the dendritic structures of the C4da neurons. Because overexpression of Dronc caused C4da neuron to undergo apoptosis prior to metamorphosis, it was not possible to use GFP-tagged Dronc to examine its distribution in these neurons during pupariation. It was therefore necessary to search for alternative means to visualize activated Dronc or its downstream caspases (Kuo, 2006).
An antibody generated against activated mammalian caspase 3 has been shown to be effective in recognizing activated caspases in Drosophila. Whereas this antibody reportedly recognizes the Drosophila effector caspase Drice, it may also cross react with other activated Drosophila caspases such as Dronc during tissue staining, because of similarities in the sequences of these caspases in the region corresponding to the peptide used to generate this antibody. Therefore this antibody was used to determine whether activated caspase is localized to the dendrites of C4da neurons during the initial severing event. At 4 hr APF, just prior to dendrite severing, antibody staining for activated caspase was consistently observed within the proximal larval dendrites of C4da neurons, especially within dendritic swellings. In the diap16-3s gain-of-function mutant that inhibits Dronc activity, as well as in ubcD1 and dronc mutant MARCM clones, C4da neurons did not show dendritic swellings or activated caspase staining in dendrites during early metamorphosis. Consistent with previous observation that C4da neurons do not remodel their axons during concurrent dendrite pruning, no activated caspase staining was seen within the axons of C4da neurons during dendrite severing. Since overexpression of p35 in these neurons did not block dendrite pruning (Kuo, 2005), it is believed this antibody staining likely recognizes activated Dronc directly or recognizes a p35-resistant caspase that is activated by Dronc. These results show that, concurrent with the nuclear upregulation of DIAP1 in C4da neurons that prevents apoptosis, there is a local activation of caspases in the dendrites, likely as a result of UPS-mediated degradation of DIAP1. The spatially restricted activation of caspases then allows the severing of proximal larval dendrites from the soma (Kuo, 2006).
This study has shown that the UPS regulates pruning of larval dendrites from C4da neurons in a cell-intrinsic manner. To better understand the molecular pathways regulating UPS-mediated pruning, a candidate E2/E3 ubiquitinating enzyme screen was conducted. In this screen an E2 ubiquitin-conjugating enzyme mutation in was uncovered ubcD1, causing dendrite pruning defects. Taken together with the extensive biochemical characterization of interactions between UbcD1, DIAP1, and Dronc, this study suggests that in C4da neurons, UPS activation leads to UbcD1-mediated degradation of E3 ubiquitin ligase DIAP1, thereby allowing Dronc caspase activation and the subsequent cleavage of larval dendrites. This work not only identifies a set of E2/E3 ubiquitinating enzymes regulating neuronal process remodeling, it also links the UPS to a hitherto unappreciated mechanism for local caspase activation in dendrites during Drosophila metamorphosis (Kuo, 2006).
The mechanistic link between the UPS and caspase activity in regulating C4da neuron dendrite pruning is unexpected. Although the UPS is known to regulate remodeling and degradation of neuronal processes, it is generally believed that this process is accomplished by degradation of cellular proteins (such as microtubules and neurofilaments) that are required to keep dendrites and axons intact. However, it was found that the UPS in C4da neurons is in fact causing the degradation of an E3 ligase, DIAP1, thereby allowing for subsequent dendrite pruning. In this case, UPS-mediated degradation of a protein does not in and of itself lead to a structural compromise in dendrites, but rather it leads to the activation of another protease that executes dendrite pruning. This two-step activation cascade, which involves both the UPS and the apoptotic machinery, may provide an additional level of control and flexibility that would not be possible if UPS alone regulated the pruning program. After all, these C4da neurons, which are specified during fly embryogenesis, maintain a highly elaborate dendritic field to receive sensory inputs throughout larval development, which lasts for several days. The maintenance of these dendrites over time requires a network of finely tuned cell-intrinsic and -extrinsic pathways. Just as important, the dendritic pruning program enables dramatic neuronal remodeling in response to profound environmental changes during metamorphosis. It is conceivable that C4da neurons evolved this dual control mechanism to prevent any accidental triggering of dendrite pruning prior to metamorphosis. Initiation of C4da neuron dendrite pruning requires cell-intrinsic ecdysone signaling, and ecdysone receptors have been shown to regulate Dronc expression. It will be of interest to determine how this UPS/caspase dendritic pruning pathway is related to the ecdysone signaling cascade (Kuo, 2006).
During metamorphosis, C4da neurons upregulate DIAP1 expression in the nucleus, which is consistent with this class of neurons surviving early stages of the metamorphosis (only one of the three C4da neurons per hemisegment, the ventral neuron, is lost at a later stage of pupariation). Remarkably, there are activated caspases within the dendrites prior to severing, and a gain-of-function diap1 mutation can block dendrite pruning, strongly implicating a local dendritic program that can activate caspases without causing apoptosis of the neuron. Although mutations in both the Dronc caspase and the UPS pathway that modulate Dronc activity (UbcD1 and DIAP1) result in retention of larval dendrites, their dendrite pruning defects differed somewhat quantitatively. Compared to dronc mutants, diap1 gain-of-function and especially ubcD1 mutants showed less retention of larval dendrites during metamorphosis. This is not surprising for diap1 gain-of-function, as it is an effective Dronc inhibitor but unlikely to be 100% efficient. UbcD1, as an E2 ubiquitin-conjugating enzyme, has wider substrate specificity than E3 ligases. Previous study showed that UbcD1 is involved in mushroom body neuroblast proliferation, so it may be involved in other UPS-mediated pathways during dendrite pruning. It is also conceivable that in the absence of UbcD1 another E2 may trigger a low level of DIAP1 degradation, allowing residual Dronc activation which results in a milder dendrite pruning phenotype in ubcD1 mutants. It is currently unclear whether UbcD1 is also required during DIAP1-mediated degradation of Dronc. However, pruning defects in the ubcD1 mutants suggest that it may not be absolutely required, since undegraded DIAP1 continues to inhibit Dronc, presumably via interaction with another E2 protein (Kuo, 2006).
How is the specificity of dendrite pruning achieved? Several possible mechanisms are proposed: first, C4da neurons do not change their axonal projections during dendrite pruning, so there could be dendrite-specific trafficking of components of the UPS, such as UbcD1, and/or the caspase Dronc. Of the known proteins that are preferentially trafficked to dendrites, these molecules have not been implicated but warrant further investigation. Second, it is also possible that activated Dronc, or another p35-resistant protease activated by Dronc, could cleave a dendrite-specific substrate. Examples are now emerging from other cellular systems, such as in sperm formation and border cell migration, in which caspases can participate in cleavage of proteins not resulting in apoptosis. Third, the dendritic pruning program takes place during drastic environmental changes that include concurrent degradation and regrowth of the overlying epidermis, activation of extracellular matrix metalloproteases, and blood phagocytes. These environmental cues likely complement the neuronal intrinsic pruning programs, but their exact relationships are not known. Experiments addressing these and other possible mechanisms should provide a greater insight into how the large-scale remodeling of C4da neuron dendrites is achieved (Kuo, 2006).
In vertebrates, the UPS pathway has been implicated in Wallerian degeneration of severed axons. In the fly, mushroom body γ neurons undergo extensive remodeling of their processes during metamorphosis. The initial stages of axon pruning in these mushroom body neurons closely resemble Wallerian degeneration, and the UPS again plays a critical role. To date, the specific ubiquitin-conjugating enzymes and ligases that mediate target protein degradation have not been identified in these systems. It will be interesting to see whether the UbcD1-DIAP1-Dronc pathway implicated in C4da neuron dendrite pruning also participates in remodeling/degradation of neuronal processes in other systems. It seems likely that more than one pathway would be employed in remodeling different neurons; a previous study excluded UbcD1 as a possible ubiquitin-conjugating enzyme regulating mushroom body γ neuron remodeling, and normal remodeling of mushroom body neuron processes in is seen dronc mutant MARCM clones during metamorphosis (Kuo, 2006).
A multilayered regulatory machinery for remodeling neurons, as uncovered in this study for C4da neurons, offers versatility and flexibility. It is conceivable that another ubiquitin ligase/caspase pair may function in an analogous UPS pathway during mushroom body neuron remodeling, potentially affording differential regulation of neuronal remodeling. Although pharmacological inhibition of mammalian caspases showed no effect on Wallerian degeneration, it would be important to assess the in vivo effectiveness of the inhibitors against a comprehensive panel of caspases. Moreover, a dual control mechanism, similar to what is proposed for C4da neuron remodeling, may coordinately regulate UPS and another protease that executes axon degradation. Conceivably, instead of having the target of the UPS directly involved in maintaining dendrite/axon stability, the executor of neuronal process degradation may involve a different protease: in the case of C4da neurons it is the caspase Dronc, and in Wallerian degeneration the relevant protease might be the Ca2+-responsive calpain. Future experiments along these lines of thinking may accelerate the identification of specific ubiquitinating enzymes involved in other areas of developmental neuronal remodeling and in diseases where the UPS pathway has been implicated. As target-specific E3 ligases are excellent candidates for pharmaceutical intervention, this approach may also help to find effective treatments for developmental and neurodegenerative diseases that involve degeneration of neuronal processes (Kuo, 2006).
Microtubules are essential to neuronal structure and function. Axonal and dendritic microtubules are enriched in post-translational modifications that impact microtubule dynamics, transport, and microtubule-associated proteins. Acetylation of alpha-tubulin lysine 40 (K40) is a prominent, conserved modification of neuronal microtubules. However, the cellular role of microtubule acetylation remains controversial. To resolve how microtubule acetylation might affect neuronal morphogenesis, endogenous alpha-tubulin was mutated in vivo using a new fly strain that facilitates the rapid knock-in of designer alpha-tubulin alleles. Leveraging this new strain, it was found that microtubule acetylation, as well as polyglutamylation and (de)tyrosination, is not essential for survival. However, it was found that dendrite branch refinement in sensory neurons relies on alpha-tubulin K40. Mutagenesis of K40 reveals moderate yet significant changes in dendritic lysosome transport, microtubule polymerization, and Futsch distribution in dendrites but not axons. These studies point to an unappreciated role for alpha-tubulin K40 and acetylation in dendrite morphogenesis. While the results are consistent with the idea that microtubule acetylation patterns microtubule function within neurons, they also suggest there may be an acetylation-independent requirement for alpha-tubulin K40 (Jenkins, 2017).
Precise patterning of dendritic fields is essential for neuronal circuit formation and function, but how neurons establish and maintain their dendritic fields during development is poorly understood. In Drosophila class IV dendritic arborization neurons, dendritic tiling, which allows for the complete but non-overlapping coverage of the dendritic fields, is established through a 'like-repels-like' behaviour of dendrites mediated by Tricornered (Trc), one of two NDR (nuclear Dbf2-related) family kinases in Drosophila. The other NDR family kinase, the tumour suppressor Warts/Lats (Wts), regulates the maintenance of dendrites; in wts mutants, dendrites initially tile the body wall normally, but progressively lose branches at later larval stages, whereas the axon shows no obvious defects. Biochemical and genetic evidence is provided for the tumour suppressor kinase Hippo (Hpo) as an upstream regulator of Wts and Trc for dendrite maintenance and tiling, respectively, thereby revealing important functions of tumour suppressor genes of the Hpo signalling pathway in dendrite morphogenesis (Emoto, 2006).
Dendritic arborization patterns are critical to a neuron's ability to receive and process impinging signals. Whereas neurons normally maintain the gross morphology of their dendrites, cortical neurons of Down's syndrome patients gradually lose dendritic branches after initially forming normal dendritic fields. Thus, neurons appear to have separate mechanisms for establishment and maintenance of their dendritic fields (Emoto, 2006).
Dendritic tiling is an evolutionarily conserved mechanism for neurons of the same type to ensure complete but non-redundant coverage of dendritic fields. In the mammalian visual system, for instance, dendrites of each retinal ganglion cell type cover the entire retina with little overlap, like tiles on a floor. In Drosophila, the dendritic arborization sensory neurons can be divided into four classes (I-IV) based on their dendrite morphology, and the dendritic field of class IV dendritic arborization neurons is shaped, in part, through a like-repels-like tiling behaviour of dendrite terminals. The NDR family kinase Trc and its activator Furry (Fry) has been identified as essential regulators of dendritic tiling and branching of class IV dendritic arborization neurons. These proteins are evolutionarily conserved and probably serve similar functions in neurons of different organisms (Emoto, 2006).
In addition to Trc, Drosophila has one other NDR family kinase, Wts, which is a tumour suppressor protein that functions in the coordination of cell proliferation and cell death in flies. To uncover the cell-autonomous functions of Wts in neurons, MARCM (mosaic analysis with a repressive cell marker) was ised to generate mCD8-GFP-labelled wts clones in a heterozygous background. Wild-type class IV neurons elaborate highly branched dendrites that cover essentially the entire body wall. Compared to wild-type ddaC (dorsal dendrite arborization neuron C) neurons, wts clones showed a severe and highly penetrant simplification of dendritic trees, with significantly reduced number (wild type, 575.1; wts, 255.6) and length (wild type, 1,457.0; wts, 590.4) of dendritic branches, and hence a greatly reduced dendritic field (Emoto, 2006).
In contrast to the severe dendritic defects caused by loss of Wts function, wts mutant ddaC axons entered the ventral nerve cord at the appropriate position and showed arborization patterns very similar to wild-type controls, with their axons terminating on the innermost fascicle and sending ipsilateral branches anteriorly and posteriorly and sometimes also a collateral branch towards the midline. Thus, Wts seems to have a crucial role in dendrite-specific morphogenesis in post-mitotic neurons (Emoto, 2006).
In proliferating cells, Wts is part of a signalling complex for tumour suppression that includes the adaptor protein Salvador (Sav) and the serine/threonine kinase Hpo. sav mutant ddaC MARCM clones were examined and dendritic defects were observed similar to wts MARCM clones. In severely affected clones (3 of 15 clones), most of the high-order branches were missing, whereas moderately affected clones (12 of 15 clones) exhibited a partial loss of their fine branches and major branches (Emoto, 2006).
To confirm that Wts and Sav function in the same pathway, genetic interaction between wts and sav in regulating dendrite morphogenesis was tested. Whereas heterozygous wts or sav mutants had no obvious dendritic phenotype, trans-heterozygous combinations of wts and sav alleles resulted in simplified dendrites similar to moderately affected sav clones. Furthermore, sav wts double mutant clones showed a severe dendrite defect comparable to wts mutant clones. Thus, Wts and Sav most probably function together in class IV neurons to regulate dendrite morphogenesis (Emoto, 2006).
The dendritic phenotypes of wts mutants and sav mutants might result from defects in branch formation and/or elongation, or loss of normally formed dendrites. Therefore ddaC dendrites were examined at different time points of larval development using the pickpocket-EGFP reporter, which is specifically expressed in class IV dendritic arborization neurons. Wild-type ddaC neurons elaborated primary and secondary dendritic branches by 24-28 h after egg laying, but large regions of the body wall were not yet covered by dendrites. By 48-52 h after egg laying, the major branches reached the dorsal midline, and the open spaces between major branches were filled with fine branches, resulting in complete dendritic coverage of the body wall. This tiling of dendrites persisted throughout the rest of larval development. In wts and sav mutants, ddaC dendrites were indistinguishable from those of wild-type controls at 24-28 h after egg laying. By 48-52 h after egg laying, wts and sav dendrites tiled the body wall as in wild type. During the next 24 h, however, dendrites of wts and sav mutants no longer tiled the body wall. Therefore, wts and sav seem to be required for maintenance of the already established tiling of dendrites (Emoto, 2006).
The loss of dendrites was further documented in live mutant larvae imaged for 30 h starting in early second instar larvae (48-50 h after egg laying). In wild-type larvae, ddaC dendrites grew steadily; the number of terminal branches increased by 23.0 over this time period. By contrast, dendrites of wts and sav mutants gradually lost their fine branches (decrease of 27.5 and 31.5, respectively) as well as some of the major branches by 78-80 h after egg laying (Emoto, 2006).
Class-IV-neuron-specific expression of wts and sav largely rescued the dendritic phenotype of wts and sav mutants, respectively, confirming that Wts and Sav act cell autonomously in class IV neurons. Furthermore, no detectable defect in patterning of the epidermis (anti-Armadillo antibody) or muscle (Tropomyosin::GFP reporter) was observed in wts or sav mutant third instar larvae. Taken together, these results indicate that the Wts/Sav signalling pathway functions in class IV neurons to maintain dendritic arborizations (Emoto, 2006).
Wts kinase activity is regulated, at least in part, by the Ste20-like serine/threonine kinase Hpo. Indeed, ddaC clones mutant for hpo exhibited simplified dendritic trees in third instar larvae, similar to wts or sav mutant clones, but showed more extensive dendritic arborizations in earlier larval stages (second to early third instar), consistent with the involvement of Hpo in the maintenance of dendrites. Notably, in hpo mutant clones at earlier developmental stages, dendritic branches were often found to overlap. Both the dendritic tiling and maintenance phenotypes were rescued by hpo expression in MARCM clones, consistent with the cell-autonomous function of Hpo in class IV neurons. Because this tiling defect in hpo mutant clones was similar to the tiling defects of trc mutant clones, whether hpo could genetically interact with trc to regulate dendritic tiling was tested. Compared with wild-type controls, trans-heterozygous combinations of trc and hpo exhibited obvious iso-neuronal as well as hetero-neuronal tiling defects, whereas wts and hpo trans-heterozygotes displayed simplified dendrites similar to wts mutants. These dendritic defects were consistently observed in multiple allelic combinations between hpo and trc or wts. In contrast, trans-heterozygous combinations of trc and wts showed no significant dendritic phenotypes. Furthermore, overexpression of wild-type Trc, but not Wts, in hpo MARCM clones partially rescued the dendritic tiling defects in class IV neurons. Thus, Hpo acts through Trc and Wts to regulate dendritic tiling and maintenance, respectively (Emoto, 2006).
Not only did Hpo interact genetically with Trc and Wts, its physical association with these NDR kinases could be detected in vivo. When Flag-tagged Trc was expressed using a nervous-system-specific Gal4 driver, anti-Flag antibodies immunoprecipitated Trc together with Hpo. Similarly, Myc-tagged Wts co-immunoprecipitated with Hpo expressed in embryonic nervous systems. Hpo co-immunoprecipitation appeared to be specific, because Misshapen, another Ste20-like kinase protein present in neurons, was not co-immunoprecipitated by anti-Flag or anti-Myc antibodies in similar experiments. These results suggest that Hpo associates with Trc and Wts in the Drosophila nervous system (Emoto, 2006).
To examine further the physical interaction between Trc and Hpo, analogous experiments were carried out in Drosophila S2 cells co-transfected with a haemagglutinin (HA)-tagged Trc construct and a Flag-tagged Hpo construct containing the full open reading frame, an amino-terminal fragment containing the kinase domain, or a carboxy-terminal fragment containing the regulatory domain. Full-length Hpo and the C-terminal portion of Hpo, but not the N-terminal fragment, were co-immunoprecipitated with Trc, suggesting that the C-terminal domain of Hpo is sufficient for Trc-Hpo complex formation (Emoto, 2006).
Hpo physically interacts with Wts and promotes Wts phosphorylation at multiple serine/threonine sites, including two sites, S920 and T1083 of Drosophila Wts, that appear to be necessary for Wts kinase activation. Indeed, Wts protein with mutations in the S920 and T1083 residues was unable to rescue the wts mutant dendritic phenotypes. Given that the corresponding phosphorylation sites in Trc are critical for Trc activation as well as control of dendritic tiling and branching, it was of interest to know whether Hpo may promote Trc phosphorylation at the critical serine and/or threonine residue. Wild-type Hpo, but not catalytically inactive Hpo or the Misshapen kinase, led to substantial incorporation of 32P-labelled phosphate into recombinant Trc or Trc with a mutation at the S292 site (S292A), but not the T449A mutant form of Trc. Analogous results were obtained with Wts. These results support a model in which Hpo associates with and phosphorylates Trc and Wts at a critical threonine residue to regulate dendritic tiling and maintenance, respectively (Emoto, 2006).
Both genetic and biochemical evidence reveals that Hpo regulates complementary aspects of dendrite development through two distinct downstream signalling pathways: the Trc kinase pathway for tiling and the Wts kinase pathway for maintenance. These studies of class IV dendritic arborization neurons, together with the recent report that Wts signalling is required for cell fate specification of photoreceptor cells in Drosophila retina, demonstrate that the Wts signalling pathway is important for post-mitotic neurons. In proliferating cells, Wts phosphorylates Yorkie (Yki), a transcriptional co-activator, to regulate cell cycle and apoptosis in growing cells. However, Yki is dispensable for Hpo/Wts-mediated dendrite maintenance. Hpo probably functions as an upstream kinase for Trc, as well as Wts, in neurons by phosphorylating a functionally essential threonine, which may also be regulated by MST3, a Ste20-like kinase closely related to Hpo. Given the evolutionary conservation of known components in the Trc and Wts signalling pathways, it will be important to identify their relevant downstream targets and explore mechanisms that coordinate the establishment and maintenance of dendritic fields, and to determine the role of Trc and Wts signalling in the mammalian nervous system (Emoto, 2006).
To cover the receptive field completely and non-redundantly, neurons of certain functional groups arrange tiling of their dendrites. In Drosophila class IV dendrite arborization (da) neurons, the NDR family kinase Tricornered (Trc) is required for homotypic repulsion of dendrites that facilitates dendritic tiling. This study reports that Sin1, Rictor, and target of rapamycin (TOR), components of the TOR complex 2 (TORC2), are required for dendritic tiling of class IV da neurons. Similar to trc mutants, dendrites of sin1 and rictor mutants show inappropriate overlap of the dendritic fields. TORC2 components physically and genetically interact with Trc, consistent with a shared role in regulating dendritic tiling. Moreover, TORC2 is essential for Trc phosphorylation on a residue that is critical for Trc activity in vivo and in vitro. Remarkably, neuronal expression of a dominant active form of Trc rescues the tiling defects in sin1 and rictor mutants. These findings suggest that TORC2 likely acts together with the Trc signalling pathway to regulate the dendritic tiling of class IV da neurons, and thus uncover the first neuronal function of TORC2 in vivo (Koike-Kumagai, 2009).
In addition to establishing dendritic coverage of the receptive field, neurons need to adjust their dendritic arbors to match changes of the receptive field. This study shows that dendrite arborization (da) sensory neurons establish dendritic coverage of the body wall early in Drosophila larval development and then grow in precise proportion to their substrate, the underlying body wall epithelium, as the larva more than triples in length. This phenomenon, referred to as scaling growth of dendrites, requires the function of the microRNA (miRNA) bantam (ban) in the epithelial cells rather than the da neurons themselves. ban in epithelial cells dampens Akt kinase activity in adjacent neurons to influence dendrite growth. This signaling between epithelial cells and neurons receiving sensory input from the body wall synchronizes their growth to ensure proper dendritic coverage of the receptive field (Parrish, 2009).
Dendrites of class IV da neurons completely and nonredundantly cover the larval body wall early in larval development, a phenomenon referred to as dendritic tiling. Once field coverage is established, dendrites continue to branch and lengthen to maintain tiling as larvae grow, providing a sensitive system for analysis of how neurons first establish and later maintain coverage of the receptive field. This study addressed the question of how late-stage dendrite growth is precisely coordinated with larval growth to maintain proper dendrite coverage of the body wall (Parrish, 2009).
To examine this process, the pickpocket-EGFP (ppk-EGFP) marker was used to monitor class IV dendrite growth before and after establishment of tiling. To quantitatively assess dendrite coverage, a metric was used that is referred to as the coverage index, the ratio of the territory covered by dendrites of a given da neuron, such as the class IV neuron ddaC, to the area of a hemisegment that harbors the da neurons. Dendrite outgrowth of class IV neurons begins at ~16 hr After Egg Laying (AEL), with class IV dendrites growing rapidly during late embryonic/early larval stages to tile the body wall between 40 and 48 hr AEL and subsequently maintaining this coverage until dendrites are pruned during metamorphosis. Between 48 hr AEL and 120 hr AEL (just prior to metamorphosis), larvae grow nearly 3-fold in length and the dorsal area of class IV receptive fields expands by more than 6-fold. Therefore, class IV dendrites grow extensively and this dendrite growth must be precisely coordinated with larval growth in order to maintain proper coverage of the receptive field (Parrish, 2009).
Class IV dendrites are located between muscle and epithelial cells. Cell divisions that give rise to larval cells are complete by mid-embryogenesis, and larval growth is achieved by increasing cell size rather than additional proliferation. Thus, all the cells that will comprise the larval body wall musculature and epithelia are in place when dendrite outgrowth begins. To simultaneously visualize growth of class IV dendrites and epithelial cells, a protein trap line was used that directs GFP expression in epithelial cells and outlines their borders (Armadillo::GFP, adherens junctions, or Neuroglian::GFP, septate junctions) in combination with ppk-GAL4 driving expression of mCD8-RFP in class IV neurons. Using these markers, growth of class IV dendrites and epithelial cells was monitored throughout embryonic/larval stages (Parrish, 2009).
Epithelial cells grow at a nearly constant rate over the time course. Likewise, the class IV neuron soma grows at a relatively constant rate. In contrast, the dendrite growth is biphasic. Initially, class IV dendrite growth outpaces growth of epithelial cells and the larva as a whole between 16 hr and 48 hr AEL, the timeframe in which class IV dendrites establish tiling. Dendrite growth slows as class IV dendrite arbors achieve complete body wall coverage, and from 48 hr to 120 hr AEL class IV dendrites grow in proportion to larval growth at a rate comparable to that of epithelial cells. This late dendrite growth will be referred to as scaling growth of dendrites (a phenomenon unrelated to synaptic scaling) to reflect the physical scaling of dendrite arbors as they grow precisely in proportion to surrounding cells and the larva as a whole in order to maintain proper coverage of the receptive field (Parrish, 2009).
To determine whether scaling growth is a general property of da neurons, dendrite growth was monitored in class I and class III da neurons, two additional morphologically distinct classes of da neurons, using the coverage index metric introduced above. Like class IV neurons, dendrites of class I and III neurons rapidly establish coverage of a characteristic region of the body wall and subsequently maintain their coverage by expanding their dendrite arbors in precise proportion to larval growth. Class III neurons cover their territory in the same timeframe as class IV neurons, first establishing receptive field coverage at about 48 hr AEL. In contrast, class I neurons covered their characteristic territory by 24 hr AEL. Thus, temporally distinct signals may regulate scaling of dendrite growth in class I and class III/IV neurons. Nevertheless, scaling growth of dendrites seems to be a general feature of da neuron development (Parrish, 2009).
Based on the fidelity of dendrite coverage in class IV neurons, a focus was placed on these neurons for studies of dendrite scaling. The finding that class IV dendrites have a rapid growth phase during establishment of tiling and a scaling phase with slower dendrite growth to maintain tiling suggests that some signal(s) attenuate dendrite growth following establishment of tiling, synchronizing growth of class IV dendritic arbors with growth of surrounding tissue. Attempts were therefore made to characterize the signaling that underlies dendrite scaling (Parrish, 2009).
To test the capacity of dendrite scaling, the effects were examined of mutations that alter the dimensions of larvae at different developmental states on class IV dendrite growth. Alleles were chosen that survive until at least the second larval instar, allowing monitoring of dendrite coverage by class IV neurons at a time when they should have already established tiling. Overall, 35 mutant alleles were screened that cause a range of defects in larval size, shape, and growth rate. Notably, class IV dendrites properly covered the receptive field in nearly all of these mutants, accommodating a broad range of receptive field areas (ranging from 10% of wild-type [WT] in chico mutants to 120% of WT in giant [gt] mutants) and shapes. Dendrites also scaled properly in mutants defective in developmental rate, for example maintaining proper receptive field coverage in b6-22 mutants that develop slowly and persist as second instar larvae or in broad (br) mutants that persist as third instar larvae for days or even weeks. Taken together, these results demonstrate the robustness of dendrite scaling growth in class IV neurons (Parrish, 2009).
Among the few mutants that had any effect on scaling growth of dendrites, the ban mutant had the most severe dendrite overgrowth phenotype observed, with the first sign of larval growth defects at 72 hr AEL. It was reasoned that ban might be required for dendrite scaling but not earlier aspects of dendrite development, and the remainder of the study focused on the role of ban in dendrite scaling. Notably, ban encodes a miRNA and might represent a regulatory node for scaling of dendrite growth since miRNAs likely regulate expression of 100 or more target genes (Parrish, 2009).
Dendrites of individual class IV neurons occupy a larger proportion of the body wall in ban mutant third instar larvae. At 96 hr AEL, ddaC class IV neurons in ban mutants have a mean coverage index of 1.22, meaning that the receptive field of the average ddaC dendrite in ban mutant larvae is 122% of the size of the dorsal hemisegment that harbors the neuron. Thus, dendrites in ban mutants promiscuously cross boundaries that are observed by dendrites of WT neurons. For example, fewer than two dendrite branches cross the midline for a given WT class IV neuron, whereas more than 18 dendrite branches cross the midline in ban mutants. The exuberant growth of dendrites in ban mutants is manifest throughout the arbor, not just at the boundaries. However, although a coverage index of >1 is seen for ban mutant, no significant tiling defect is seen because branches that cross normal boundaries still avoid dendrites of neighboring class IV neurons. In addition to these defects in dendrite coverage, class IV neurons in ban mutants show significant increases in the number of dendrites, the density of dendrites, and overall dendrite length (data not shown). However, increased terminal dendrite branching is not sufficient to increase receptive field coverage. Several other mutants have been described that increase terminal dendrite branching in class IV neurons, and none of these mutants cause an overall increase in the size of the dendritic field. For example, furry (fry) mutations cause a 100% increase in the number/density of terminal dendrites without an accompanying increase in coverage index at 96 hr AEL. Likewise, overexpression of the small GTPase Rac drastically increases terminal dendrite branching but reduces receptive field coverage (Parrish, 2009).
The dendrite growth defects in ban mutants could reflect increased dendrite growth from early stages of development or defects specific to the scaling phase of dendrite growth. To distinguish between these possibilities, dendrite growth was monitored over a developmental time course, focusing on the coverage index and midline crossing events as metrics for growth of the dendrite arbor as a whole. Importantly, class IV dendrites in ban mutants are indistinguishable from WT during the early, rapid growth phase (through 48 hr AEL) as measured by coverage index, midline crossing events, and total dendrite branch number. However, beginning at 72 hr AEL, progressively more severe defects are noted in the coverage index and a greater number of midline crossing events in ban mutants. This late-onset exuberant dendrite growth demonstrates that ban is not causing a general growth defect since ban is dispensable for establishment of dendrite coverage. Whereas a generalized defect in dendrite growth, as seen in dendritic arbor reduction (dar; mutations that lead to defective dendritic arbors but normal axonal projections), would affect both the early (isometric) and late (scaling) phases of growth, mutations that specifically affect the scaling growth of dendrites would be dispensable for the early, rapid growth of dendritic fields. This is precisely what is seen for ban mutants. Therefore, ban is specifically required for scaling of dendrite arbors, potentially by affecting growth-inhibitory signals that normally restrict dendrite growth (Parrish, 2009).
To confirm that loss of ban causes these phenotypes, the following experiments were conducted. First, whereas heterozygosity for a ban null allele or deficiencies that span the ban locus show no obvious defects in dendrite scaling, placing ban mutations in trans to a deficiency that spans the locus, but not a nearby deficiency that does not span the ban locus, recapitulates the dendrite defects described above. Second, the ban mutant dendrite defects can be fully rescued by a ban genomic rescue transgene but not a genomic transgene in which the ban locus has been deleted. Therefore, disrupting ban function is sufficient to cause defects in scaling growth of dendrites (Parrish, 2009).
Next whether ban is required for scaling growth of dendrites was tested in other classes of da neurons. Both class I and class III neurons establish proper dendrite coverage in ban mutants. However, class III dendrites are defective in scaling of dendrite growth in ban mutants, showing a significant increase in dendrite coverage after 48 hr AEL. In contrast, larval class I dendrites show no obvious defects in dendrite coverage in ban mutants, demonstrating that ban is not required for scaling in class I neurons. The onset of scaling growth of dendrites differs by 24 hr in class I and class III/IV neurons, thus different scaling signals may operate at the two time points with ban required for the scaling growth signal for class III/IV neurons that tile (Parrish, 2009).
Next, time-lapse microscopy of single neurons was conducted to characterize the cellular basis of the ban mutant phenotype. Single class IV neurons were imaged from time-matched WT or ban mutant larvae at 24 hr intervals beginning at 72 hr AEL, just after the ban phenotype is first apparent. Dynamics were monitored of every terminal dendrite that could be unambiguously followed through the time course and dendrite growth, initiation of new dendrites, dendrite retraction, and branch loss were measured. For each of these categories, ban mutants differed from WT controls, exhibiting significantly more dendrite growth and branch initiation and significantly less dendrite retraction and branch loss. Therefore, stabilization of existing dendrites, increased dendrite growth, and increased addition of new dendrites all contribute to the defect in dendrite scaling growth of the ban mutant (Parrish, 2009).
Time-lapse studies suggest that signals normally restricting dendrite growth are largely absent in ban mutants. Attempts were made to verify this hypothesis using laser ablation assays. Previous studies showed that, following embryonic ablation of a class IV neuron, dendrites of neighboring neurons grow exuberantly to invade the unoccupied territory of the ablated neuron, with the ability of dendrites to invade unoccupied territory progressively restricted in older larvae. It was therefore important to determine whether the timing of this restricted growth potential correlates with the onset of scaling of dendrite growth and whether ban is required for restriction of the dendrite growth potential (Parrish, 2009).
Consistent with prior reports, ablating a class IV neuron at 24 hr AEL led to extensive invasion by dendrites of neighboring neurons, with 55% of the unoccupied territory covered by neighboring neurons 48 hr postablation. This ability of dendrites to grow into unoccupied territory was severely attenuated 1 day later, with dendrites of neighboring neurons invading only 23% of the unoccupied territory after ablation of a class IV neuron at 48 hr AEL . The extent of invasion was even further reduced when neurons were ablated at 72 hr AEL. Therefore, the ability of dendrites to grow beyond their normal boundaries to invade unoccupied territory is severely restricted during larval development at a time coincident with the onset of scaling of dendrite growth (Parrish, 2009).
If the restriction of dendrite growth potential in larvae is caused by scaling signals that limit dendrites to growth in proportion to body wall growth, the majority of invading activity by neighboring dendrites should be present before scaling growth ensues at 48 hr AEL. To test this prediction, class IV neurons were ablated at 24 hr AEL and invasion activity was monitored at 24 hr intervals over the next 72 hr. By 48 hr AEL, dendrites of neighboring neurons had invaded unoccupied territory, and the extent of invasion was not noticeably increased at later time points. Instead, the entire dendrite arbor of class IV neurons, including the portion that invaded unoccupied territory, scaled with larval growth after 48 hr AEL. Thus, the receptive field that is established by 48 hr AEL is maintained by scaling of dendrite growth, even in cases in which dendrites establish aberrant body wall coverage. The signals responsible for dendrite scaling growth are likely distinct from the homotypic repulsion required to establish tiling as ablation of all neighboring same-type neurons does not potentiate the ability of a class IV neuron to invade unoccupied territory. Additionally, dendrites of class I da neurons, which do not rely on homotypic repulsion to establish their coverage, also exhibit scaling growth (Parrish, 2009).
As described above, dendrite coverage is properly established in ban mutants. Importantly, unlike WT controls, following ablation at 48 hr AEL, dendrites in ban mutants extensively fill unoccupied space, with dendrites in ban mutants invading unoccupied territory just as efficiently as dendrites in WT controls ablated at 24 hr AEL. Therefore, the receptive field boundaries of class IV neurons have not been fixed in ban mutants at 48 hr AEL. Dendrites in ban mutants invade unoccupied territory more efficiently than WT controls at later time points as well. Thus, either the growth-inhibitory scaling signal is lost or dendrites are refractory to the signal in ban mutants (Parrish, 2009).
To test whether machineries for dendritic tiling contribute to the progressive reduction of a dendrite's ability to invade vacant territories, mutations of fry, which encodes a gene required for establishment of dendritic tiling and of extra sex combs (esc) and salvador (sav), which function in a common pathway to regulate stability of terminal dendrites and, consequently, maintenance of dendrite coverage, were examined for effects on dendrite invasion following neuron ablation. Unlike mutations in ban, mutations in fry, esc, or sav had no effect on the ability of dendrites to invade unoccupied territory. Moreover, consistent with the scaling signal functioning in a distinct pathway, double-mutant combinations of ban with fry or esc showed additive phenotypes. Thus, ban exerts its effects on scaling of dendrite growth independently of known pathways for establishment and maintenance of dendrite coverage (Parrish, 2009).
To further characterize the signaling required for scaling growth of dendrites, it was of interest to determine where ban functions to regulate scaling. First, whether ban is expressed in neurons, surrounding cells, or both, was examined by using a miRNA activity sensor as a reporter for ban expression in third instar larvae. A control sensor directs ubiquitous expression of GFP, including robust GFP expression in muscle, epithelial cells, and sensory neurons. The ban sensor contains two ban binding sites in the 3'UTR of the transgene, hence GFP expression is attenuated in cells that express ban. Unlike the control sensor, very little, if any, GFP expression was detected in third instar muscle cells, epithelial cells, or sensory neurons using several independent transgenic fly lines with distinct insertions of the ban sensor. Significant attenuation of the ban sensor was first observ ed in larval muscle, epithelium, and PNS neurons between 48 and 72 hr AEL, precisely at the time when dendrite defects were first observed in ban mutants, suggesting that ban activity is more pronounced during this period than at earlier time points. Notably, the attenuation of the ban sensor was dependent on ban activity, as shown by the persistent, ubiquitous expression of the sensor in ban mutant larvae. Thus, ban is likely expressed in the muscle, epithelium, and PNS neurons and may be required in any of these cell types for scaling of dendrite growth (Parrish, 2009).
To determine whether ban is required cell-autonomously for dendrite scaling, MARCM was used to generate single neuron clones homozygous for a ban mutation in a heterozygous background. ban activity was effectively dampened in MARCM clones, as indicated by derepression of the ban sensor in the clones. However, loss of ban function had no significant effect on dendrite coverage of class IV neurons. Time-lapse analysis of ban mutant class IV MARCM clones revealed no defects in dendrite coverage at any time during larval development. Furthermore, ban is dispensable in other da neurons for dendrite scaling growth. Thus, ban function in sensory neurons is dispensable for scaling growth of dendrites (Parrish, 2009).
Although scaling of dendrite growth proceeds normally, there is some reduction of overall dendrite length and the number of dendrite branches in ban mutant class IV clones. Therefore, ban likely acts cell-autonomously to promote dendrite growth and nonautonomously to limit dendrite Taking advantage and ensure proper scaling (Parrish, 2009).
A genetic rescue assay was used to test the ability of transgenic expression of ban in different tissues to rescue the dendrite growth defects of ban mutants. Consistent with MARCM results, neuronal expression of ban, using either panneuronal or PNS-specific Gal4 drivers, was not sufficient to rescue the scaling growth defect of ban mutants. Thus, ban likely functions nonautonomously in nonneuronal cells to regulate scaling of da neuron dendrite growth. Moreover, expression of ban in muscle alone could not ameliorate the dendrite defects of ban mutants. Remarkably, every time ban expression was rescued in epithelial cells, significant suppression of the exuberant dendrite growth of ban mutants was found. The three epithelial Gal4 driver lines caused reductions of dendrite growth that correlated with Gal4 expression levels in epithelial cells: arm-Gal4 caused the greatest reduction in dendrite growth and had the strongest epithelial expression, whereas twi-Gal4 displayed the lowest activity and drove epithelial Gal4 expression at the lowest level. Taking advantage of the temperature-sensitive nature of Gal4 activity, rescue activity of each epithelial Gal4 line was monitored over a graded temperature series (18°C to 29°C) and it was found that, for each driver, rescue activity was directly proportional to expression level. Therefore, epithelial ban expression is sufficient to suppress the exuberant dendrite growth of ban mutants, and the extent of dendrite growth inhibition varies with the level of ban expression in epithelial cells (Parrish, 2009).
Epithelial expression of twi-Gal4 was first apparent in larval stages, suggesting that postembryonic expression of ban in epithelia is sufficient for proper scaling of dendrite growth. Given that dendrite defects in ban mutants first appear after 48 hr AEL, it was asked whether late expression of ban would suffice for dendrite scaling. To examine the temporal requirement for ban function, a heat-shock-inducible Gal4 driver was used to express ban during larval development. Indeed, inducing ban expression at 48 hr AEL was sufficient to rescue the dendrite defects of ban mutants. These findings reinforce the notion that ban is dispensable for early aspects of dendrite development (Parrish, 2009).
Resupplying ban in tissues known to regulate larval growth, such as the fat body, prothoracic (PTTH) gland, or insulin-producing cells (IPCs), had no measurable effect on dendrite growth in ban mutants. Moreover, ablation of each of these tissues mediated by a reaper transgene caused larval growth defects without obvious dendrite growth defects. Thus, ban function in the fat body, PTTH gland, or IPCs is not sufficient to modulate scaling of dendrite growth. Altogether, these results suggest that epithelial cells are likely the major functional sites for ban in regulation of PNS dendrite scaling (Parrish, 2009).
Because ban expression in epithelial cells affects scaling growth of dendrites in a dose-dependent fashion via a mechanism that likely involves growth-inhibitory signals, it was of interest to see whether ectopic epithelial expression of ban in a WT background could further inhibit dendrite growth and thus disrupt scaling of dendrite arbors. Indeed, overexpression of ban in epithelial cells resulted in a severe reduction in dendrite growth and induced striking defects in the pattern of dendrite growth over epithelial cells, with terminal dendrites appearing to wrap around epithelial cells. Consistent with ban dosage in epithelial cells regulating the strength of dendrite growth-inhibitory signals, epithelial overexpression of ban induced more robust inhibition of dendrite growth at higher temperatures (which lead to higher levels of transgene expression) (Parrish, 2009).
Since ban expression in epithelial cells is sufficient to ensure proper scaling, it was of interest to address whether ban function in epithelial cells is necessary for scaling of dendrite growth. To this end, MARCM was used to generate ban mutant epithelial cell clones. Although it was not possible to address the contribution of epithelial ban to scaling of the entire dendrite arbor using this approach (it was only possible to generate one to four cell epithelial clones), the pattern was monitored of dendrite growth over ban mutant or WT control epithelial clones. Class IV dendrites grow extensively over epithelial cells, with multiple dendrite branches often coursing over a single epithelial cell. The epithelial nucleus was used as a landmark and dendrite growth was monitored over the epithelial cell surface shadowed by the nucleus. Although the gross morphology of epithelial cells was not obviously affected in ban mutant clones, the propensity of class IV dendrites to grow into the region shadowed by the epithelial nucleus was significantly increased for ban mutant epithelial clones when compared to WT controls or ban heterozygous epithelial cells. Therefore, ban is required in epithelial cells to ensure proper dendrite growth and placement over epithelial cells (Parrish, 2009).
To gain insight into the molecular mechanism underlying dendrite scaling, a platform was developed for microarray-based expression profiling of dissociated, FACS-isolated PNS neurons or epithelial cells. Akt and numerous other candidate genes were identified that were deregulated in PNS neurons and/or epithelial cells of ban mutant larvae. Because Akt is a well-established regulator of growth, including dendrite growth in mammalian hippocampal neurons, whether ban regulates Akt as part of the scaling program was investigated (Parrish, 2009).
Microarray experiments found that Akt expression was increased in neurons but reduced in epithelial cells of ban mutants relative to WT controls. By monitoring Akt levels in lysates of larval fillets composed mostly of muscle and epithelial cells, it was found that in the absence of ban function Akt protein levels were substantially reduced. Furthermore, Akt activity was substantially reduced as shown by reductions in active, phosphorylated Akt and phosphorylated S6K, a downstream reporter of Akt activity. Therefore, Akt expression and activity are substantially reduced in ban mutant larval lysates, likely reflecting reduced Akt function in muscle/epithelia (Parrish, 2009).
Next larval fillets were immunostained to determine whether ban influences Akt protein levels in the PNS. In WT controls, Akt is detectible only at low levels in the soma or dendrites of the PNS. By contrast, in ban mutants Akt is highly expressed in the PNS and is detectible in axons, the soma, and dendrites. Similarly, phosphorylated Akt is barely detectible in the larval PNS of WT controls but is present at high levels in the PNS of ban mutants. Therefore, ban regulates Akt expression and activity in the larval PNS (Parrish, 2009).
To test whether this effect on Akt levels reflects a neuronal requirement for ban, Akt expression levels were monitored in ban mutants in which ban expression is resupplied under the control of twist-Gal4, an experimental condition that rescues both the dendrite scaling defect and larval size defect of ban mutants. It was found that ban nonautonomously regulates Akt levels in da neurons since nonneuronal expression of ban (twist-Gal4) is sufficient to dampen the ectopic neuronal Akt expression normally seen in ban mutants. Therefore, ban likely functions in epithelia to regulate signals that influence Akt expression and activity in neurons (Parrish, 2009).
Finally, it was of interest to determine whether Akt function in class IV neurons is important for scaling of dendrite growth. Based on expression data, it was predicted that increasing Akt expression/activity in class IV neurons should cause a scaling defect similar to what is seen in ban mutants. Indeed, ectopic expression of Akt, or a constitutively active form of PI3 kinase (PI3k) that leads to activation of Akt, caused a significant increase in dendrite coverage, similar to ban mutants. Conversely, antagonizing Akt activity in class IV neurons by overexpressing Pten, a PIP3 phosphatase that functions as an inhibitor of Akt activity, by knocking down Akt expression via RNAi in class IV neurons, or by generating Akt null mutant class IV neuron MARCM clones caused a significant reduction in dendrite coverage. Therefore, Akt plays a critical role in regulating dendrite coverage (Parrish, 2009).
If increased neuronal Akt activity underlies the dendrite defects in ban mutants, then antagonizing neuronal Akt activity should suppress the dendrite overgrowth in ban mutants. This hypothesis was tested with the following three experiments. First, RNAi was used to knock down Akt expression in class IV neurons of ban mutant larvae. On its own, Akt(RNAi) causes a reduction in dendrite growth and overall coverage of the receptive field, and this phenotype is epistatic to the dendrite overgrowth seen in ban mutants. Similarly, Pten was overexpressed in class IV neurons of ban mutant larvae and it was found that the Pten-mediated reduction in dendrite coverage is epistatic to the dendrite overgrowth seen in ban mutants. Finally, class IV neurons was ablated in ban mutants in the absence or presence of neuron-specific Akt RNAi and it was found that reducing neuronal Akt expression blocks the exuberant dendrite invasion activity of ban mutants. Altogether, these results strongly suggest that ban functions in epithelial cells to regulate neuronal expression/activation of Akt, and deregulation of Akt leads to the dendrite growth defects of ban mutants (Parrish, 2009).
This study has shown that nonautonomous signals coordinate growth of dendrites with the growth of their substrate and the body as a whole. Dendrites of many types of neurons cover characteristic receptive fields, and growth of the dendrite arbor in synchrony with the receptive field in a process, referred to as scaling growth of dendrites, allows a neuron to maintain proper dendrite coverage of the receptive field. Thus, scaling growth of dendrites is likely a general mechanism to ensure fidelity of dendrite coverage (Parrish, 2009).
Dendrites of class IV neurons cover their receptive field before larval growth is complete and must maintain this coverage as the larva grows. Two properties distinguish the scaling phase of dendrite growth from the early dendrite growth when the neuron establishes receptive field coverage. During the scaling phase of growth, the dendrite arbor grows precisely in proportion to receptive field expansion (which is often achieved by animal growth). Moreover, dendrite growth is constrained by boundaries delineated when the dendrite arbor first covers the receptive field. Thus, although dendrites continue to grow, growth occurs only to maintain proportional coverage of the receptive field (Parrish, 2009).
Dendrites of Drosophila da neurons exhibit a biphasic growth profile: dendrites establish coverage of their receptive field via an early, rapid growth phase and maintain this coverage via a late scaling growth phase in which dendrites grow in proportion with epithelial cells and the animal as a whole. The miRNA ban acts in the second phase to enable scaling of dendrite growth in da neurons, ensuring that dendrites maintain proper body wall coverage. Loss of ban disrupts epithelial-derived signaling that normally modulates dendrite growth, and, as a result, dendrites remain in the 'rapid growth' phase, extending beyond their normal territories. This phenotype is reminiscent of the heterochronic phenotypes of C. elegans lin-4 and let-7 mutants in that an early developmental phase is inappropriately reiterated during a later phase (Parrish, 2009).
How broadly do miRNAs regulate developmental progressions in the nervous system? Although the developmental roles of vertebrate miRNAs have been somewhat elusive because of the vast number of miRNA-encoding genes, several studies suggest that miRNAs may serve highly specialized roles in regulating developmental transitions in neuron morphogenesis. For example, completely abrogating miRNA function causes robust defects in neuron morphogenesis but not specification in zebrafish, consistent with miRNAS regulating late aspects of neuronal differentiation. Likewise, miRNAs miR9a* and miR-124 regulate the switch in subunit composition of chromatin remodeling complexes as neural progenitors differentiate into neurons in mice. Additionally, a number of miRNAs function primarily at a very late step of neuron development to regulate activity-dependent dendrite growth and synaptic plasticity. For example, neuronal activity antagonizes miR-134, which normally inhibits growth of dendritic spines, and promotes expression of mir-132, which promotes dendritic plasticity (Parrish, 2009).
Although ban is known to regulate growth in proliferating tissues in Drosophila, ban-mediated regulation of dendrite growth likely represents a distinct mode of growth control by ban for the following reasons. First, previous studies focused on autonomous regulation of tissue growth by ban. In contrast, ban acts nonautonomously to regulate scaling of dendrite growth. Second, prior studies of ban function focused on imaginal discs where growth is achieved by increasing cell number rather than cell size. By contrast, dendrite scaling involves ban-mediated regulation of growth in differentiated, postmitotic cells. Likewise, postmitotic expression of ban in the larval eye disc can also regulate cell size. Third, ban functions downstream of the tumor suppressor kinase Hippo to control proliferation, with Hippo activating the transcription factor Yorkie, which in turn activates ban expression. Whereas hippo is required cell-autonomously for establishment and maintenance of dendrite tiling, yorkie is dispensable for dendrite growth. As to the cell-nonautonomous function of ban in dendrite scaling, Hippo is not required. Although these findings suggest that ban regulates growth of proliferating and differentiated tissues by different means, it is possible that in both scenarios ban is antagonizing expression of growth-inhibitory factors, possibly even the same factors, and removing growth inhibition has different consequences on proliferative and differentiated tissues (Parrish, 2009).
It is proposed that ban positively regulates an epithelial-derived signal that modulates neuronal Akt expression and activity to influence dendrite growth. Several observations suggest that the signal acts over a short range, possibly even via direct adhesive interactions between dendrites and epithelia or the underlying matrix. First, ban overexpression in epithelial cells but not in muscle influences growth of dendrites. Second, removing ban function from epithelial cell clones influences the distribution of dendrites over the clone but not over adjacent WT epithelial cells. Third, in addition to inhibiting the overall rate of dendrite growth, overexpressing ban in epithelial cells induces exaggerated “wrapping” of epithelial cells by terminal dendrites. Since these signals appear to preferentially regulate dendrite growth during the scaling phase, ban may modulate dendrite/epithelial adhesion during the scaling phase of dendrite development (Parrish, 2009).
Morphologically distinct classes of da neurons establish type-specific dendritic coverage of the body wall and maintain this coverage by means of dendrite scaling as larvae grow. However, arbors of different da neurons develop at different rates, with class I dendrites establishing their coverage 1 day earlier than class III and class IV dendrites. Mutations in ban disrupt scaling of dendrite growth in class III and class IV but not class I neurons, suggesting that different signals regulate dendrite scaling in distinct types of neurons. Thus, temporally distinct signals or temporally restricted sensitivity to the signals may ensure that different neurons maintain appropriate coverage of their receptive field (Parrish, 2009).
The dendritic arbors of the larval Drosophila peripheral class IV dendritic arborization neurons degenerate during metamorphosis in an ecdysone-dependent manner. This process-also known as dendrite pruning-depends on the ubiquitin-proteasome system (UPS), but the specific processes regulated by the UPS during pruning have been largely elusive. This study shows that mutation or inhibition of Valosin-Containing Protein (VCP; termed TER94 by FlyBase), a ubiquitin-dependent ATPase whose human homolog is linked to neurodegenerative disease, leads to specific defects in mRNA metabolism and that this role of VCP is linked to dendrite pruning. Specifically, it was found that VCP inhibition causes an altered splicing pattern of the large pruning gene Molecule interacting with CasL and mislocalization of the Drosophila homolog of the human RNA-binding protein TAR-DNA-binding protein of 43 kilo-Dalton (TDP-43). These data suggest that VCP inactivation might lead to specific gain-of-function of TDP-43 and other RNA-binding proteins. A similar combination of defects is also seen in a mutant in the ubiquitin-conjugating enzyme ubcD1 (Effete) and a mutant in the 19S regulatory particle of the proteasome, but not in a 20S proteasome mutant. Thus, these results highlight a proteolysis-independent function of the UPS during class IV dendritic arborization neuron dendrite pruning and link the UPS to the control of mRNA metabolism (Rumpf, 2014).
To achieve specific connections during development, neurons need to refine their axonal and dendritic arbors. This often involves the elimination of neuronal processes by regulated retraction or degeneration, processes known collectively as pruning. In the Drosophila, large-scale neuronal remodeling and pruning occur during metamorphosis. For example, the peripheral class IV dendritic arborization (da) neurons specifically prune their extensive larval dendritic arbors, whereas another class of da neurons, the class III da neurons, undergo ecdysone- and caspase-dependent cell death. Class IV da neuron dendrite pruning requires the steroid hormone ecdysone and its target gene SOX14, encoding an HMG box transcription factor. Class IV da neuron dendrites are first severed proximally from the soma by the action of enzymes like Katanin-p60L and Mical that sever microtubules and actin cables, respectively. Later, caspases are required for the fragmentation and phagocytic engulfment of the severed dendrite remnants . Another signaling cascade known to be required for pruning is the ubiquitin-proteasome system (UPS). Covalent modification with the small protein ubiquitin occurs by a thioester cascade involving the ubiquitin-activating enzyme Uba1 (E1), and subsequent transfer to ubiquitin-conjugating enzymes (E2s) and the specificity-determining E3 enzymes. Ubiquitylation of a protein usually leads to the degradation of the modified protein by the proteasome, a large cylindrical protease that consists of two large subunits, the 19S regulatory particle and the proteolytic 20S core particle. Several basal components of the ubiquitylation cascade—Uba1 and the E2 enzyme ubcD1—as well as several components of the 19S subunit of the proteasome have been shown to be required for pruning, as well as the ATPase associated with diverse cellular activities (AAA) ATPase Valosin-Containing Protein (VCP) (CDC48 in yeast, p97 in vertebrates, also known as TER94 in Drosophila), which acts as a chaperone for ubiquitylated proteins. Interestingly, autosomal dominant mutations in the human VCP gene cause hereditary forms of ubiquitin-positive frontotemporal dementia (FTLD-U) and amyotrophic lateral sclerosis (ALS). A hallmark of these diseases is the occurrence of both cytosolic and nuclear ubiquitin-positive neuronal aggregates that often contain the RNA-binding protein TAR-DNA-binding protein of 43 kilo-Dalton (TDP-43). It has been proposed that ubcD1 and VCP promote the activation of caspases during dendrite pruning via degradation of the caspase inhibitor DIAP1. However, mutation of ubcD1 or VCP inhibit the severing of class IV da neuron dendrites from the cell body, whereas in caspase mutants, dendrites are still severed from the cell body, but clearance of the severed fragments is affected. This indicates that the UPS must have additional, as yet unidentified, functions during pruning (Rumpf, 2014).
This study further investigated the role of UPS mutants in dendrite pruning. vcp mutation was shown to lead to a specific defect in ecdysone-dependent gene expression, as VCP is required for the functional expression and splicing of the large ecdysone target gene molecule interacting with CasL (MICAL). Concomitantly, mislocalization of Drosophila TDP-43 and up-regulation of other RNA-binding proteins were observed, and genetic evidence suggests that these alterations contribute to the observed pruning defects in VCP mutants. Defects in MICAL expression and TDP-43 localization are also induced by mutations in ubcD1 and in the 19S regulatory particle of the proteasome, but not by a mutation in the 20S core particle, despite the fact that proteasomal proteolysis is required for dendrite pruning, indicating the requirement for multiple UPS pathways during class IV da neuron dendrite pruning (Rumpf, 2014).
Class IV da neurons have long and branched dendrites at the third instar larval stage. In wild-type animals, these dendrites are completely pruned at 16-18 h after puparium formation (h APF). VCP mutant class IV da neurons were generated by the Mosaic Analysis with a Repressible Cell Marker (MARCM) technique for clonal analysis. Mutant vcp26-8 class IV da neurons displayed strong pruning defects and retained long dendrites at 16 h APF. Expression of an ATPase-deficient dominant-negative VCP protein (VCP QQ) under the class IV da neuron-specific driver ppk-GAL4 recapitulated the pruning phenotype and also led to the retention of long and branched dendrites at 16 h APF. VCP inhibition also causes defects in class III da neuron apoptosis. This combination of defects in both pruning and apoptosis is reminiscent of the phenotypes caused by defects in ecdysone-dependent gene expression. Indeed, overexpression of the transcription factor Sox14, which induces pruning genes, led to a nearly complete suppression of the pruning phenotype caused by VCP QQ. This genetic interaction suggested that VCP might be required for the expression of one or several ecdysone target genes during pruning (Rumpf, 2014).
How could VCP be linked to Sox14? The suppression of the vcp mutant phenotype by Sox14 overexpression could be achieved in one of several ways. Sox14 could be epistatic to VCP -- that is, VCP could be required for functional Sox14 expression -- and this effect would be mitigated by Sox14 overexpression. However, VCP could also be required for the expression of one or several Sox14 target genes, and enhanced Sox14 expression could overcome this requirement either via enhanced induction of one or several particular targets or via enhanced induction of other pruning genes, in which case Sox14 would be a bypass suppressor of VCP QQ. To distinguish between these possibilities, the effects were assessed of VCP inhibition on the expression of known genes in the ecdysone cascade required for pruning in class IV da neurons. Class IV da neuron pruning is governed by the Ecdysone Receptor B1 (EcR-B1) isoform, which in turn directly activates the transcription of Sox14 and Headcase (Hdc), a pruning factor of unknown function. Sox14, on the other hand, activates the transcription of the MICAL gene encoding an actin-severing enzyme. In immunostaining experiments, VCP QQ did not affect the expression of EcR-B1, Sox14, or Hdc at the onset of the pupal phase. However, the expression of Mical was selectively abrogated in class IV da neurons expressing VCP QQ, or in vcp26-8 class IV da neuron MARCM clones . These data indicated that VCP might affect dendrite pruning by regulating the expression of the Sox14 target gene Mical, indicating that Sox14 might act as a bypass suppressor of VCP QQ (Rumpf, 2014).
How could VCP inhibition suppress Mical expression? To answer this question, whether Mical mRNA could still be detected in class IV da neurons expressing VCP QQ was assessed. To this end, enzymatic tissue digestion and FACS sorting were used to isolate class IV da neurons from early pupae (1-5 h APF). Total RNA was then extracted from the isolated neurons, and the presence of Mical mRNA expression was assessed by RT-PCR, using control samples or samples from animals expressing VCP QQ under ppk-GAL4. The Mical gene is large (~40 kb) and spans multiple exons that are transcribed to yield a ~15 kb mRNA. To detect Mical cDNA, primer pairs spanning several exons were used for two different regions of Mical mRNA, exons 14-16 and exons 8-12. [MICAL is on the (-) strand, but the exon numbering denoted by Flybase follows the direction of the (+) strand. Therefore, exons 14-16 are upstream of exons 8-12, and the latter are closer to the 3' end of MICAL mRNA.] MICAL mRNA was detectable upon VCP inhibition in these extracts with a primer pair spanning exons 14-16. The second primer pair spanning exons 8-12 also detected MICAL mRNA in both samples, but the RT-PCR product from the VCP QQ-expressing class IV da neurons had a larger molecular weight. Sequencing of the PCR products indicated that MICAL mRNA from VCP QQ-expressing class IV da neurons contained exon 11, which was not present in Mical mRNA from the control sample. Exon 11 is absent from all predicted MICAL splice isoforms except for a weakly supported isoform designated 'Mical-RM'. It introduces a stop codon into MICAL mRNA that would lead to the truncation of the C-terminal 1,611 amino acids from Mical protein. This portion of Mical protein contains several predicted protein interaction domains such as a proline-rich region, a coiled-coil region with similarity to Ezrin/Radixin/Moesin (ERM) domains, and a C-terminal PDZ-binding motif, and is required for the interaction between Mical and PlexinA. In addition, the truncated region contains the epitope for the antibody used in the immunofluorescence experiments, thus explaining the observed lack of Mical expression upon VCP inhibition. Given that a mutant of Mical with a smaller C-terminal truncation (compared with the one induced by VCP inhibition) was not sufficient to rescue the class IV da neuron dendrite pruning defect of mical mutants, disruption of VCP function likely results in expression of a truncated Mical protein without pruning activity. Taken together, these data suggest that the observed defect in MICAL mRNA splicing contributes significantly to the pruning defects of VCP mutants (Rumpf, 2014).
How is VCP linked to alternative splicing of MICAL mRNA? A plausible mechanism for the control of an alternative splicing event would be the modulation of specific (pre)mRNA-binding proteins. VCP has recently been linked to several RNA-binding proteins: human autosomal dominant VCP mutations cause frontotemporal dementia or ALS with inclusion bodies that contain aggregated human TDP-43; a genetic screen in Drosophila identified the RNA-binding proteins Drosophila TDP-43, HRP48, and x16 as weak genetic interactors of the dominant effects of VCP disease mutants; and HuR (a human homolog of the neuronal Drosophila RNA-binding protein elav) was recently shown to bind human VCP. Of these, TDP-43 and also elav have been linked to alternative splicing in various model systems, including Drosophila. Therefore this study used available specific antibodies to assess the levels and distribution of Drosophila TDP-43 (hereafter referred to as TDP-43) and elav. TDP-43 has previously been shown to localize to the nucleus in Drosophila motoneurons and mushroom body Kenyon cells. Surprisingly, TDP-43 was largely localized to the cytoplasm in class IV da neurons, where it was enriched in a punctate pattern around the nucleus, with only a small fraction also detectable in the nucleus, a localization pattern that could be reproduced with transgenic N-terminally HA-tagged TDP-43. Elav is a known nuclear marker for Drosophila neurons; in class IV da neurons, it was somewhat enriched in nuclear punctae. The effects of VCP inhibition on these two RNA-binding proteins was assessed. Elav localization did not change notably upon VCP QQ expression. Strikingly, TDP-43 became depleted from the cytoplasm of class IV da neurons and relocalized to the nucleus upon VCP QQ expression. Closer inspection revealed that TDP-43 in VCP-inhibited neurons was now enriched in nuclear dots that often also exhibited increased elav staining. The relocalization of TDP-43 from the cytoplasm to the nucleus was also observed in vcp26-8 mutant class IV da neuron MARCM clones. Importantly, quantification and normalization of TDP-43 levels showed that VCP inhibition did not alter the absolute levels of TDP-43, suggesting that the observed effects were not a consequence of a defect in TDP-43 degradation. In fact, the only manipulation that resulted in a mild but significant increase in TDP-43 levels -- but without a change in localization -- was the expression of an RNAi directed against the autophagy factor ATG7, perhaps reflecting the degradation of cytoplasmic RNA granules through the autophagy pathway (Rumpf, 2014).
It was next asked whether manipulation of TDP-43 would affect class IV da neuron dendrite pruning. A previously characterized TDP-43 mutant, TDP-43 Q367X (28-128">28), did not display pruning defects, but overexpression of TDP-43 led to strong dendrite pruning defects at 16 h APF. In support of the hypothesis that TDP-43 acts in the same or a similar pathway as VCP during dendrite pruning, it was also found that a more weakly expressed TDP-43 transgene (UAS-TDP-43weak) and VCP A229E, a weakly dominant-active VCP allele corresponding to a human VCP disease mutation, exhibited a synergistic inhibition of pruning when coexpressed. Interestingly, manipulation of elav gave very similar results as with TDP-43: elav down-regulation by RNAi did not affect pruning, but elav overexpression led to highly penetrant pruning defects (Rumpf, 2014).
To exclude the possibility that the pruning defects induced by TDP-43 or elav overexpression were due to long-term expression and aggregation of RNA-binding proteins, TDP-43 and elav overexpression was also induced acutely (24 h before the onset of pupariation). Pruning was still inhibited in these cases. Also, overexpression of several other RNA-binding proteins did not cause pruning defects, with two exceptions: a UAS-carrying P-element in the promotor of the adjacent x16 and HRP48 genes caused a strong pruning defect when expression was induced in class IV da neurons, and levels of a GFP protein trap insertion into the x16 gene were also markedly increased in class IV da neurons expressing VCP QQ, possibly indicating a role for VCP in x16 degradation. In further support of an involvement of VCP with RNA-binding proteins during neuronal pruning processes, it was also found that VCP is required for mushroom body γ neuron axon pruning and induces the accumulation of Boule, an RNA-binding protein that had previously been shown to inhibit γ neuron axon pruning when overexpressed. Thus, the data suggest that VCP regulates a specific subset of RNA-binding proteins and that this regulatory role of VCP is associated with its role in pruning (Rumpf, 2014).
As VCP is an integral component of the UPS, it was next asked whether the role of VCP in MICAL regulation and TDP-43 localization was also dependent on ubiquitylation and/or the proteasome. To address this question, Mical levels and TDP-43 distribution was assessed in UPS mutants with known pruning defects. An ubiquitylation enzyme known to be required for pruning is the E2 enzyme ubcD1. When TDP-43 localization was assessed in larval ubcD1D73 mutant class IV da neurons, TDP-43 was again localized to the nucleus in these cells. Furthermore, a pronounced reduction of Mical expression in ubcD1D73 mutant class IV da neurons was noted during the early pupal stage, indicating that ubiquitylation through ubcD1 is involved in the regulation of TDP-43 localization and Mical expression (Rumpf, 2014).
TDP-43 localization and Mical expression were assessed in proteasome mutants. A previously characterized mutant in the Mov34 gene encoding the 19S subunit Rpn8 was used. TDP-43 was again relocalized to the nucleus in Mov34 mutant class IV da neurons, and Mical expression was absent from Mov34 mutant class IV da neurons at 2 h APF. To rigorously address whether proteasomal proteolysis was also required for TDP-43 localization and Mical expression, the effect was assessed of Pros261, a previously characterized mutation in the 20S core particle subunit Prosβ6. In contrast to Mov34 mutant class IV da neurons, Pros261 mutant class IV da neurons displayed cytoplasmic TDP-43 localization, and robust Mical expression was detected in these neurons at 2 h APF. Thus, although ubiquitylation and the 19S proteasome are both required for Mical expression and normal TDP-43 localization, proteolysis through the 20S core particle of the proteasome is not. Importantly, Pros261 MARCM class IV da neurons showed strong dendrite pruning defects at 16 h APF, as did expression of RNAi constructs directed against subunits of the 20S core particle (Rumpf, 2014).
These data indicate that there must be several ubiquitin- and proteasome-dependent pathways that are required for dendrite pruning: one pathway requires ubcD1, VCP, and the 19S regulatory particle of the proteasome, but not the 20S core particle. This pathway regulates MICAL expression. A second UPS pruning pathway does depend on proteolysis through the 20S core. In an E3 ubiquitin ligase candidate screen, cul-1/lin19 was identified as a pruning mutant. Cul-1 encodes cullin-1, a core component of a class of multisubunit ubiquitin ligases known as SCF (for Skp1/Cullin/F-box) ligases. Class IV da neurons mutant for cul-1 or class IV da neurons expressing an RNAi construct directed against cul-1 had not pruned their dendrites at 16 h APF. However, unlike with VCP, ubcD1, and Mov34, cul-1 mutation did not affect Mical expression at 2 h APF, indicating that cullin-1 is not a component of the VCP-dependent UPS pathway involved in splicing and might thus be a component of a proteolytic UPS pathway. In support of this notion, a recent report independently identified cul-1 as a pruning mutant and associated it with protein degradation (Rumpf, 2014).
It has been proposed that the E2 enzyme ubcD1 and VCP would act to activate caspases during pruning. However, the dendrite pruning defects caused by those UPS mutants are much stronger than the phenotypes caused by caspase inactivation, which mostly causes a delay in the phagocytic uptake of severed dendrites by the epidermis. Although it cannot be excluded that ubcD1 and VCP contribute to caspase activation during pruning, the new mechanism proposed in this study -- control of RNA-binding proteins and MICAL expression -- likely makes a much stronger contribution to the drastic pruning phenotypes of UPS mutants (Rumpf, 2014).
How precisely do VCP, ubcD1, and the 19S proteasome contribute to MICAL expression? The data indicate that VCP inhibition causes missplicing of MICAL mRNA that likely leads to the expression of an inactive Mical protein variant. At the same time, VCP inhibition leads to the mislocalization of TDP-43, and possibly the dysregulation of a number of other RNA-binding proteins. The fact that these phenotypes correlate in the vcp, ubcD1, and Mov34 mutants gives a strong indication that they are related. TDP-43 had previously been identified as a suppressor of the toxicity induced by a weak VCP disease allele in the Drosophila eye. In class IV da neurons, reducing the amounts of TDP-43 (with a deficiency) or elav (by RNAi) did not ameliorate the pruning defect induced by VCP inhibition. Therefore, the possibility cannot be excluded that the two proteins act in parallel rather than in an epistatic fashion. As VCP has been shown to remodel protein complexes that contain ubiquitylated proteins and is structurally similar to the 19S cap, it is interesting to speculate that VCP and the 19S cap might alter the subunit composition of ubiquitylated TDP-43-containing complexes of RNA-binding proteins, and that this activity—rather than a direct action on TDP-43 (or maybe also elav) alone—might lead to both MICAL missplicing and TDP-43 mislocalization (Rumpf, 2014).
Interestingly, autosomal dominant mutations in human VCP cause frontotemporal dementia and ALS, a hallmark of which is the formation of aggregates that contain TDP-43. Most of these aggregates are cytoplasmic (and contain TDP-43 that has relocalized from the nucleus to the cytoplasm), but VCP mutations also induce TDP-43 aggregation in the nucleus, a situation that might be similar to the situation caused by VCP inhibition in class IV da neurons. Although human VCP disease mutations have been proposed to act as dominant-active versions of VCP with enhanced ATPase activity, both the disease allele and the dominant-negative ATPase-dead VCP QQ mutant cause class IV da neuron pruning defects and TDP-43 relocalization to the nucleus of class IV da neurons and therefore act in the same direction. It is thought that VCP can only bind substrates when bound to ATP, and will release bound substrates upon ATP hydrolysis. Thus, it is conceivable that the phenotypic outcome of inhibiting the ATPase (no substrate release) should be similar to that of ATPase overactivation (reduced substrate binding or premature substrate release): in both cases, a substrate protein complex would not be properly remodeled (Rumpf, 2014).
Taken together, these results indicate the existence of a nonproteolytic function of VCP and the UPS in RNA metabolism and highlight its importance during neuronal development (Rumpf, 2014).
During development, dendrites migrate to their correct locations in response to environmental cues. The mechanisms of dendritic guidance are poorly understood. Recent work has shown that the Drosophila olfactory map is initially formed by the spatial segregation of the projection neuron (PN) dendrites in the developing antennal lobe (AL). This study reports that between 16 and 30 h after puparium formation, the PN dendrites undergo dramatic rotational reordering to achieve their final glomerular positions. During this period, a novel set of AL-extrinsic neurons express high levels of the Wnt5 protein and are tightly associated with the dorsolateral edge of the AL. Wnt5 forms a dorsolateral-high to ventromedial-low pattern in the antennal lobe neuropil. Loss of Wnt5 prevents the ventral targeting of the dendrites, whereas Wnt5 overexpression disrupts dendritic patterning. Drl/Ryk, a known Wnt5 receptor, is expressed in a dorsolateral-to-ventromedial (DL > VM) gradient by the PN dendrites. Loss of Drl in the PNs results in the aberrant ventromedial targeting of the dendrites, a defect that is suppressed by reduction in Wnt5 gene dosage. Conversely, overexpression of Drl in the PNs results in the dorsolateral targeting of their dendrites, an effect that requires Drl's cytoplasmic domain. It is proposed that Wnt5 acts as a repulsive guidance cue for the PN dendrites, whereas Drl signaling in the dendrites inhibits Wnt5 signaling. In this way, the precise expression patterns of Wnt5 and Drl orient the PN dendrites allowing them to target their final glomerular positions (Wu, 2014).
How does wiring specificity of neural maps emerge during development? Formation of the adult Drosophila olfactory glomerular map begins with patterning of projection neuron (PN) dendrites at the early pupal stage. To better understand the origin of wiring specificity of this map, this study created genetic tools to systematically characterize dendrite patterning across development at PN type-specific resolution. PNs were found to use lineage and birth order combinatorially to build the initial dendritic map. Specifically, birth order directs dendrite targeting in rotating and binary manners for PNs of the anterodorsal and lateral lineages, respectively. Two-photon- and adaptive optical lattice light-sheet microscope-based time-lapse imaging reveals that PN dendrites initiate active targeting with direction-dependent branch stabilization on the timescale of seconds. Moreover, PNs that are used in both the larval and adult olfactory circuits prune their larval-specific dendrites and re-extend new dendrites simultaneously to facilitate timely olfactory map organization. This work highlights the power and necessity of type-specific neuronal access and time-lapse imaging in identifying wiring mechanisms that underlie complex patterns of functional neural maps (Wong, 2023).
Neural maps are emergent, highly ordered structures that are essential for organizing and presenting synaptic information. Within the embryonic nervous system of Drosophila motoneuron dendrites are organized topographically as a myotopic map that reflects their pattern of innervation in the muscle field. This fundamental organizational principle exists in adult Drosophila, where the dendrites of leg motoneurons also generate a myotopic map. A single postembryonic neuroblast sequentially generates different leg motoneuron subtypes, starting with those innervating proximal targets and medial neuropil regions and producing progeny that innervate distal muscle targets and lateral neuropil later in the lineage. Thus the cellular distinctions in peripheral targets and central dendritic domains, which make up the myotopic map, are linked to the birth-order of these motoneurons. Developmental analysis of dendrite growth reveals that this myotopic map is generated by targeting. The medio-lateral positioning of motoneuron dendrites in the leg neuropil is controlled by the midline signalling systems Slit-Robo and Netrin-Fra. These results reveal that dendritic targeting plays a major role in the formation of myotopic maps and suggests that the coordinate spatial control of both pre- and postsynaptic elements by global neuropilar signals may be an important mechanism for establishing the specificity of synaptic connections (Brierley, 2009).
Neural maps are emergent, highly ordered structures that are essential for organizing and presenting synaptic information. The architecture of dendrites and the role they play in establishing connectivity within maps has been somewhat overlooked. Classic cell-labelling studies in the moth Manduca sexta revealed that the dendrites of motoneurons are topographically organized to reflect their site of innervation in the bodywall. More recent work by Landgraf and colleagues has demonstrated that motoneurons in Drosophila embryos generate a detailed dendritic (myotopic) map of body wall muscles within the CNS. Alongside these data, studies on the architecture of the spinal cord also suggest that similar design principles may play a role in organizing information in vertebrate motor systems. How such dendritic maps are built is still largely unknown. This study describes the role dendritic targeting plays in constructing a myotopic map and the molecular mechanisms that control it (Brierley, 2009).
The majority of leg motoneurons in a fly are born postembryonically and most of those are derived from a single neuroblast lineage, termed lineage 15. Perhaps the most striking feature of this lineage is its birth-order-based pattern of innervation along the proximo-distal axis of the leg. Using mosaic analysis, the sequential production was observed of four neuronal subtypes during larval life, each elaborating stereotyped axonal and dendritic projections in the adult. The axon of the first-born neuron innervates a muscle in the body wall and subsequent neurons innervate more distal targets in the leg. This organization has also been reported by Baek (2009) (Brierley, 2009).
This birth-order-based peripheral pattern of lineage 15 is mirrored in the CNS, where dendrites generate a stereotyped anatomical organization. Dendrites of early-born cells span medial to lateral territories, whereas late-born cells elaborate dendrites in the lateral neuropil and cells born between these times occupy intermediate territories. The sequential production of neuronal subtypes by neural precursor cells is a common mechanism for generating a diversity of circuit components. A similar birth-order-based specification of axonal and dendritic projection patterns has previously been described for projection neurons in the fly's olfactory system (Brierley, 2009).
The data reveal the existence of a myotopic map in the adult fly and supports the proposition that dendritic maps are a common organizing principle of all motor systems. Mauss (2009) also reveal a map in the embryonic CNS of Drosophila, where the dendrites of motoneurons are organized along the medio-lateral axis of the neuropil representing dorsoventral patterns of innervation in the body wall muscles (Brierley, 2009).
How are dendritic maps built? The myotopic map seen in the leg neuropil could be generated by two distinctly different mechanisms. Neurons could elaborate their dendrites profusely across a wide field and then remove branches from inappropriate regions or, alternatively, they could target the growth of dendrites into a distinct region of neuropil throughout development. Both mechanisms can generate cell-type-specific projection patterns as seen in the vertebrate retina. To reveal which mechanism is deployed in the leg motor system of Drosophila, single-cell clones of motoneuron subtypes generated by heatshocks at 48 and 96 h AH were imaged, since their final dendritic arborizations cover clearly distinct territories within the map. The dendrites of both elaborate branches only in territories where the mature arborizations eventually reside, which strongly supports the notion that this myotopic map is generated by targeting and not large-scale branch elimination. Importantly, this developmental timeline also revealed that the motoneurons elaborate their dendrites synchronously, regardless of the birth date of the cell. This observation suggests that a 'space-filling/occupancy based' model, where later-born neurons are excluded from medial territories by competitive interactions is unlikely. Similarly, heterochronic mechanisms where different members of the lineage experience different signalling landscapes due to differences in the timing of outgrowth are not likely either. With synchronous outgrowth dendrites experience the same set of extracellular signals, suggesting that the intrinsic properties of cells, defined by their birth order, may be more important for the generation of subtype-specific projections. Such intrinsic properties could include cell-cell recognition systems such as adhesion molecules, e.g., Dscams or classical guidance receptors, that could interpret extracellular signals. In the Drosophila embryo motoneurons also use dendritic targeting to generate a myotopic map (Brierley, 2009).
It is emerging that dendrites are guided by the same molecules that control axon pathfinding. The medio-lateral organization of leg motoneuron dendrites within the leg neuropil prompted an investigation as to whether the midline signalling molecules Slit and Netrin and their respective receptors Roundabout and Frazzled could be involved in targeting growth to specific territories (Brierley, 2009).
Using mosaic analysis it was found that both the 48 and 96 h AH motoneuron subtypes require Robo to generate their appropriate shape and position within the medio-lateral axis. When Robo was removed from the 48 h AH subtype the mean centre of arbor mass was shifted toward the midline. The dendrites of 96 h AH neurons showed a shift in distribution in the absence of Robo but still failed to reach the midline, suggesting that only part of this cell's targeting is due to repulsive cues mediated by the Robo receptor. It was predicted that if Robo levels played an instructive role in dendrite targeting it would be possible to shift dendrites laterally by cell autonomously increasing Robo. This was found to be the case in both subtypes. Taken together these data suggest that differences in the level of Robo signalling may provide a mechanism by which Slit could be differentially interpreted to allow subtype-specific targeting along the medio-lateral axis (Brierley, 2009).
The Robo receptor is part of a larger family of receptors that includes Robo2 and Robo3. This family of receptors have been found to be important for targeting axons to the appropriate longitudinal pathway in the embryonic CNS. Comm plays a key role in allowing contralaterally projecting neurons to cross the midline, and its ectopic expression (CommGOF) is known to robustly knock down Robo and Robo2 and 3. Comm was cell autonomously expressed in both lineage 15 subtypes and shifts to the midline were found in both 48 and 96 h AH neurons. For the 48 h AH neurons, Robo LOF data and CommGOF data are comparable, suggesting that Robo alone plays a major role in the positioning dendrites of these cells. In contrast, in the 96 h AH subtype RoboLOF and CommGOF effects were found to be significantly different, suggesting that the 96 h AH subtype may not only use the Robo receptor but additional Robos as well. Knockdown of Slit also supports this idea, as the branches of late-born neurons were occasionally found reaching the midline, something that was never see in RoboLOF clones. Thus, one way of establishing differences in the medio-lateral position could be through a dendritic “Robo code” where early-born cells express Robo and late-born cell express multiple Robo receptors (Brierley, 2009).
With Netrin being expressed in the midline cells during the pupal-adult transition it was asked whether attractive Netrin-Fra signalling could also contribute to positioning dendrites in the leg neuropil. When Fra was removed from the 48 h AH subtype it was found that the arborization was shifted laterally, whereas removing it from the 96 h AH subtype had little effect, and neither did the removal of Netrin A and B from the midline, suggesting that Netrin-Fra signalling may not play a role in dendritic targeting in the later-born cell. It may be that Fra is expressed in early-born cells within the lineage and then down-regulated, although it cannot be excluded that Netrin-Fra signalling was masked by the repulsion from Slit-Robo signalling. These data are consistent with Fra being a major player in targeting the dendrites of the 48 h AH cell. The fact that both Fra and Robo are required for normal morphogenesis of 48 h AH neurons raises the possibility that members of lineage 15 could use a 'push-pull' mechanism for positioning their dendrites, where the blend of receptors within a cell dictates the territory within the map that they will innervate (Brierley, 2009).
How could such subtype-specific blends of receptors be established? A number of studies have revealed that spatial codes of transcription factors are important for specifying the identity of motoneuron populations. Within lineage 15 it is possible that temporal, rather than spatial, transcription factor codes are important for regulating the blend of guidance receptors. A number of molecules have been identified that control the sequential generation of cell types within neuroblast lineages. Chief amongst these are a series of transcription factors that include Hunchback, Krüppel, Pdm, Castor and Seven-up. These temporal transcription factors are transiently expressed within neuroblasts and endow daughter neurons with distinct “temporal identities”. Castor and Seven-up are known to schedule transitions in postembryonic lineages, regulating the neuronal expression of BTB-POZ transcription factors Chinmo and Broad. It is possible that the temporal transcription factors Broad and Chinmo could control the subtype-specific expression of different Robo receptors or the Netrin receptor Frazzled in leg motoneurons. There is a precedent for this in the Drosophila embryo, where motoneuron axon guidance decisions to distal (dorsal) versus proximal (ventral) targets are orchestrated by Even-Skipped, a homeobox transcription factor, which in turn controls the expression of distinct Netrin receptor combinations (Brierley, 2009).
Studies focusing on the growth of olfactory projection neuron dendrites in Drosophila reveal that they elaborate a glomerular protomap prior to the arrival of olfactory receptor neurons suggesting that target/partner-derived factors may not be necessary for establishing coarse patterning of synaptic specificity. The global nature of the signals describe in this study and their origin in a third-party tissue is a fundamentally different situation to that where target-derived factors instruct partner cells, such as presynaptic amacrine cells signalling to retinal ganglion cell dendrites in the zebrafish retina. Furthermore, although this study shows that Slit and Netrin control the positioning of dendrites across the medio-lateral axis of the CNS, it may be that other similar guidance signals are important for patterning dendrites in other axes. There is a striking conservation of the molecular mechanisms that build myotopic maps in the embryo and pupae. Understanding the similarities and differences between these myotopic maps, from an anatomical, developmental, and functional perspective, may give insight into the evolution of motor systems and neural networks in general (Brierley, 2009).
This study found that individual leg motoneurons that lacked Robo signalling appeared to have more complex dendritic arborizations. The working hypothesis, that dendrites invaded medial territories because of a failure of Slit-Robo guidance function, did not take into account the possibility that cells may generate more dendrites due to a change in a cell-intrinsic growth program. Thus the changes seen in dendrite distribution relative to the midline could formally be a result of 'spill-over' from that increase in cell size/mass. To determine whether this was the case larger cells were generated by activating the insulin pathway in single motoneurons. It was found the dendrites of these 'large cells' remained within their normal neuropil territory, supporting the idea that the removal of Robo-Slit signalling results in a disruption in guidance, not growth. These data underline the fundamental importance of midline signals in controlling the spatial coordinates that these motoneuron dendrites occupy, i.e., that a neuron twice the size/mass of a wild-type cell is still marshalled into the same volume of neuropil (Brierley, 2009).
When the image stacks were reconstructed to look at the distribution of the dendrites in the dorso-ventral axis, it was found that the apparent increase in size was in fact a redistribution of the dendrites from ventral territories into more dorsal medial domains. This was unexpected and suggests that changes in midline signalling can also impact the organization of dendrites in the dorso-ventral axis. So CommGOF 96 h AH neurons may not only encounter novel synaptic inputs by projecting into medial territories, but they may also lose inputs from the ventral domains of neuropil they have vacated. These observations suggest that motoneurons within lineage 15 have a fixed quota of dendrites and where it is distributed in space depends on cell-intrinsic blends of guidance receptors. Taken together these data support the idea that growth and guidance mechanisms are genetically separable programs. In identified embryonic motoneurons where Slit-Robo and Netrin-Fra signalling has been disrupted, quantitative analysis reveals dendrites also show no measurable difference in their total number of branch tips or length (Mauss, 2009). Moreover, recent computational studies in larger flies reveal that dendritic arborizations generated by the same branching programs can generate very different shapes depending on how their 'dendritic span' restricted within the neuropil. Previous work in both vertebrates and Drosophila has shown that a loss of Slit-Robo signalling results in a reduction in dendrite growth and complexity, but this study found no evidence to support this (Brierley, 2009).
Neural maps and synaptic laminae are universal features of nervous system design and are essential for organizing and presenting synaptic information. How the appropriate pre- and postsynaptic elements within such structures are brought together remains a major unanswered question in neurobiology. Studies in recent years have shown that neural network development involves both hardwired molecular guidance mechanisms and activity-dependent processes; the relative contribution that each makes is still unclear. Work on the spinal cord network of Xenopus embryos revealed that seven identifiable neuron subtypes can establish connections with one another and that the key predictor of connectivity was their anatomical overlap. This could be interpreted to mean that connectivity is promiscuous and that the major requirement for the generation of synaptic specificity is the proximity of axons and dendrites. This is particularly interesting in light of the current dendrite targeting data and the observation that both sensory neurons and interneurons in Drosophila use the same midline cues to position their pre-synaptic terminals in the CNS. Moreover, a recent study has shown that Semaphorins control the positioning of axons within the dorso-ventral axis. Taken together these observations suggest that during development the coordinated targeting of both pre- and postsynaptic elements into the same space using global, third-party guidance signals could provide a simple way of establishing the specificity of synaptic connections within neural networks. This idea is akin to 'meeting places' such as the traditional rendezvous underneath the four-sided clock at Waterloo railway station where two interested parties organize to meet. Understanding how morphogenetic programs contribute to the generation of synaptic specificity is likely to be key to solving the problem of neural network formation (Brierley, 2009).
In many differentiated cells, microtubules are organized into polarized noncentrosomal arrays, yet few mechanisms that control these arrays have been identified. For example, mechanisms that maintain microtubule polarity in the face of constant remodeling by dynamic instability are not known. Drosophila neurons contain uniform-polarity minus-end-out microtubules in dendrites, which are often highly branched. Because undirected microtubule growth through dendrite branch points jeopardizes uniform microtubule polarity, this system was used to understand how cells can maintain dynamic arrays of polarized microtubules. It was found that growing microtubules navigate dendrite branch points by turning the same way, toward the cell body, 98% of the time and that growing microtubules track along stable microtubules toward their plus ends. Using RNAi and genetic approaches, it was shown that kinesin-2, and the +TIPS EB1 and APC, are required for uniform dendrite microtubule polarity. Moreover, the protein-protein interactions and localization of Apc2-GFP and Apc-RFP to branch points suggests that these proteins work together at dendrite branches. The functional importance of this polarity mechanism is demonstrated by the failure of neurons with reduced kinesin-2 to regenerate an axon from a dendrite. It is concluded that microtubule growth is directed at dendrite branch points and that kinesin-2, APC, and EB1 are likely to play a role in this process. It is proposed that kinesin-2 is recruited to growing microtubules by +TIPS and that the motor protein steers growing microtubules at branch points. This represents a newly discovered mechanism for maintaining polarized arrays of microtubules (Mattie, 2010).
Using Drosophila dendrites as a model system, this study demonstrates that growing microtubule plus ends almost always turn toward the cell body at branch points and that they track stable microtubules through branches. Kinesin-2, EB1, and APC are all required for maintaining microtubule polarity and are linked in an interaction network (Mattie, 2010).
On the basis of these results, a model for directed growth of microtubules in dendrites is proposed. Apc2 most likely contains a branch localization signal because Apc2-GFP localizes well to dendrite branches even when overexpressed. Localization of Apc2 to dendrite branch points can recruit Apc to the branch. Apc can interact with the Kap3 subunit of kinesin-2 and so could increase concentration of the motor near the branch. EB1-GFP does not concentrate at branches, therefore it is proposed that a growing microtubule plus end coated with EB1 is transiently linked, through the interaction between Apc and the EB1 tail, to kinesin-2 as it passes through the branch. SxIP motifs in Apc and Klp68D could also contribute to this interaction. Because both Kap3 and the SxIP motif in Klp68D are in the kinesin-2 tail, the motor domain would be free to walk along a nearby stable microtubule toward the plus end and cell body (Mattie, 2010).
Even a very brief application of force pulling the growing microtubule toward the cell body should be sufficient to steer growth toward the cell body. Once the tip of the microtubule turns, growth should be constrained by the dendrite walls. The association of the growing plus end with a stable microtubule would probably only need to be maintained over a distance of a micron. This model is consistent with the observations that kinesin-2 has shorter run lengths than kinesin-1 and that individual EB1 interactions with the microtubule plus end persist for less than a second (Mattie, 2010).
Observations of plus-end behavior in vivo also favor a model in which only transient interactions of the motor and microtubule plus end are involved; frequently plus ends are seen turning sharply. Stable microtubules do not accommodate such sharp turns, so the sharp turns of plus ends are not likely to occur while the plus end is tracking along a stable microtubule. Instead, they probably represent a switch from a freely growing plus end to one that is following a track (Mattie, 2010).
Directed growth of microtubules at dendrite branch points allows dendrites to maintain uniform minus-end-out polarity despite continued microtubule remodeling. This mechanism is also probably necessary to establish uniform microtubule polarity in branched dendrites, but it probably cannot account for minus-end-out polarity on its own. It is hypothesized that directed microtubule growth is used in concert with an unidentified mechanism to control microtubule polarity in dendrites. Transport of oriented microtubule pieces has been proposed to contribute to axon microtubule polarity, and a similar mechanism could play a role in dendrites. For example, a kinesin anchored to the cell cortex by its C terminus could shuttle minus-end-out microtubule seeds into dendrites. Alternatively, polarized microtubule nucleation sites could be localized within dendrites. Identifying directed growth as one mechanism that contributes to dendrite microtubule polarity should facilitate identification of the missing pieces of this puzzle (Mattie, 2010).
Because kinesin-2, APC, EB1, and polarized microtubules are found in many cell types, directed growth of microtubules along stable microtubule tracks could be very broadly used for maintaining microtubule polarity. This is a newly identified function for both the +TIP proteins and kinesin-2. APC has previously been localized to junctions between a microtubule tip and the side of another microtubule at the cortex of epithelial cells, and sliding of microtubule tips along the side of microtubules was also seen in this system, but there was no association with a particular direction of movement or overall microtubule polarity (Mattie, 2010).
Kinesin-2 has previously been shown to be enriched in the tips of growing axons in cultured mammalian neurons, and it is possible that they could mediate tracking of growing microtubules along existing microtubule tracks in the growth cone. In fact, this type of microtubule tracking behavior has been observed in axonal growth cones under low-actin conditions. Thus, directed growth of microtubules could also be important during axon outgrowth. Indeed, the same principle of directed growth by a motor protein connected to a growing microtubule plus end could be involved in aligning microtubules in many circumstances (Mattie, 2010).
The formation of neuronal dendrite branches is fundamental for the wiring and function of the nervous system. Indeed, dendrite branching enhances the coverage of the neuron's receptive field and modulates the initial processing of incoming stimuli. Complex dendrite patterns are achieved in vivo through a dynamic process of de novo branch formation, branch extension and retraction. The first step towards branch formation is the generation of a dynamic filopodium-like branchlet. The mechanisms underlying the initiation of dendrite branchlets are therefore crucial to the shaping of dendrites. Through in vivo time-lapse imaging of the subcellular localization of actin during the process of branching of Drosophila larva sensory neurons, combined with genetic analysis and electron tomography, this study has identified the Actin-related protein (Arp) 2/3 complex as the major actin nucleator involved in the initiation of dendrite branchlet formation, under the control of the activator WAVE and of the small GTPase Rac1. Transient recruitment of an Arp2/3 component marks the site of branchlet initiation in vivo. These data position the activation of Arp2/3 as an early hub for the initiation of branchlet formation (Sturner, 2019).
Microtubule minus ends are thought to be stable in cells. Surprisingly, in Drosophila and zebrafish neurons, persistent minus end growth is observed, with runs lasting over 10 min. In Drosophila, extended minus end growth depended on Patronin, and Patronin reduction disrupted dendritic minus-end-out polarity. In fly dendrites, microtubule nucleation sites localize at dendrite branch points. Therefore, it was hypothesized that minus end growth might be particularly important beyond branch points. Distal dendrites have mixed polarity, and reduction of Patronin lowered the number of minus-end-out microtubules. More strikingly, extra Patronin made terminal dendrites almost completely minus-end-out, indicating low Patronin normally limits minus-end-out microtubules. To determine whether minus end growth populated new dendrites with microtubules, dendrite development and regeneration were analyzed. Minus ends extended into growing dendrites in the presence of Patronin. In sum, these data suggest that Patronin facilitates sustained microtubule minus end growth, which is critical for populating dendrites with minus-end-out microtubules (Feng, 2019).
Microtubule plus ends exhibit rapid growth and shrinkage phases, a behavior termed dynamic instability. This behavior occurs in vitro with pure αβtubulin dimers and in vivo, where it can be regulated by plus end binding proteins and microtubule age. Minus ends also exhibit dynamic instability in vitro, although their growth rate is two to three times slower than plus ends. In vivo, however, minus ends are generally thought not to contribute to microtubule dynamics or exploration of cellular space (Feng, 2019).
In cells, the γtubulin ring complex acts as a template and nucleator to allow new microtubules to form. As long as it remains attached at the nucleating or minus end, it acts as a cap to block addition of subunits and prevent depolymerization. Free minus ends can then be generated by severing proteins. Two well-established fates for free minus ends are depolymerization and stabilization. In plants, cortical microtubules that are freed from nucleation sites depolymerize at the minus end while growing at the plus end to generate treadmilling behavior. In animal cells, depolymerization at the minus end is an important aspect of mitotic spindle dynamics, and minus end recognition by ASPM (abnormal spindle-like microcephaly associated) is important to help control depolymerization. In interphase, most free minus ends seem to be rapidly stabilized by calmodulin-regulated spectrin-associated protein (CAMSAP)/Patronin proteins (Feng, 2019).
Identification of CAMSAP/Patronin proteins was a major breakthrough in understanding the cellular stability of minus ends. They were the first proteins identified that could directly recognize free minus ends and are likely responsible for their resistance to depolymerization in interphase. The repertoire of minus end-binding proteins was recently expanded by the addition of ASPM, which can also recognize minus ends and controls their depolymerization in the spindle. It is not known whether this family of proteins also functions in post-mitotic cells like neurons (Feng, 2019).
Invertebrates have one CAMSAP/Patronin family member, termed Patronin, while vertebrates generally have three, CAMSAP1-3, with some extras where genome duplications have occurred. For example, in zebrafish, there are two each of CAMSAP1 and 2 and one CAMSAP3. Drosophila melanogaster Patronin stabilizes minus ends against depolymerization by kinesin-13 in cultured cells and in vitro. Mammalian CAMSAPs also protect minus ends from depolymerization by kinesin-13s in vitro and in cells. Overall stabilization of minus ends by CAMSAPs and Patronin is agreed upon as a critical function without which the microtubule network in cultured Drosophila cells and mammalian cells including neurons is dramatically destabilized (Feng, 2019).
The relationship between CAMSAPs/Patronin and minus end growth is more complex. In vitro CAMSAP2 and 3, as well as two domains of Patronin, suppress addition of tubulin subunits to the minus end in a concentration-dependent manner. In contrast, CAMSAP1 tracks minus ends as they grow without altering the rate of subunit addition. In cells, CAMSAP1 also tracks growing minus ends, but reduction of CAMSAP1 does not result in any change in microtubule behavior. CAMSAP2 has been described as suppressing minus end growth and also as promoting addition of short stretches of microtubule to the minus end. Although these two models for CAMSAP2 sound incompatible, they are actually not so different. Minus ends grow slowly in the presence of CAMSAP2, and this allows short stretches of CAMSAP2 to become stably associated with the microtubule. The stretches are on average 1 µ in control cells and 2 µ when katanin is depleted, and so, although they are derived from tubulin subunit addition, this does not result in much net growth at the minus end. In primary neuron cultures, stretches >10 µ of CAMSAP2 along microtubules have been observed, but growth has only been tracked for stretches of about a micron, so it is not clear how the longer stretches arise. Thus, it is still ambiguous whether extended growth at the minus end occurs in cells and, if so, how it contributes to global microtubule organization (Feng, 2019).
CAMSAP/Patronin proteins are particularly important in neurons where most, if not all, microtubules are noncentrosomal. In cultured hippocampal neurons, reduction of CAMSAP2, the major family member in this cell type, destabilizes microtubules and reduces dendrite complexity. Caenorhabditis elegans Patronin is required for maintenance of normal neuronal morphology, neuronal microtubule stability, and axon regeneration. Beyond stabilizing microtubules, it is not clear whether Patronin regulates specific aspects of microtubule organization in neurons. In epithelial cells, CAMSAP3 is responsible not only for stability of microtubules but also their polarized arrangement with minus ends concentrated at the apical surface. Neuronal microtubules are even more dramatically polarized than epithelial ones, with uniform plus-end-out polarity in axons and mixed or minus-end-out polarity in dendrites. It is therefore hypothesized that CAMSAP/Patronin proteins might function to control not only microtubule stability in neurons, but also their polarity (Feng, 2019).
Drosophila dendrites are strikingly polarized with >90% minus-end-out microtubules. It is conceptually straightforward to imagine plus-end-out processes in which fast-growing microtubule plus ends allow microtubules to populate an extending structure, while more complex models are thought to be required for population of processes with minus-end-out microtubules. In dendrites, local nucleation can generate new minus ends, and outgrowth of minus ends has not been considered as an alternative way to get minus-end-out microtubules into dendrites. However, in Drosophila neurons, nucleation sites are concentrated at dendrite branch points, so how the terminal dendrite beyond the branch point is populated with minus-end-out microtubules remains a conundrum (Feng, 2019).
Using live imaging of microtubule dynamics with tagged end-binding (EB) proteins in Drosophila and zebrafish neurons, this study identified a population of slow-growing microtubule ends that move in the opposite direction to fast-growing plus ends. In Drosophila dendrites, these slow-moving structures are labeled with Patronin, confirming that they are growing microtubule minus ends. This study demonstrates that sustained growth of minus ends requires Patronin, and in turn is important for getting minus-end-out microtubules into distal regions of mature dendrites, as well as into developing and regenerating dendrites (Feng, 2019).
In contrast to the traditional view that microtubule plus end growth is exclusively responsible for extending microtubules, this study found that in neurons, minus ends undergo extensive growth, with some individual runs lasting ≥10 min. Surprisingly, Patronin facilitates the persistent slow growth of minus ends, allowing them to elongate microns in single growth runs. Moreover, this minus end growth has critical functions in organizing the neuronal microtubule cytoskeleton, including populating growing dendrites and distal regions of mature dendrites with minus-end-out microtubules (Feng, 2019).
Neuronal DLK/JNK stress signaling is a common response to reduced Patronin in C. elegans and Drosophila. As this signaling is responsible for some of the phenotypes observed in both systems, it will be important to determine whether any of the phenotypes that have been described in mammalian neurons with reduced CAMSAP2 are downstream of DLK or JNK. Changes in microtubule dynamics are a key output of neuronal stress from expression of proteins that cause neurodegeneration in flies and mammals, so some of the phenotypes attributed to direct function of CAMSAP2 could be secondary to stress. The current data suggest that in Drosophila, Patronin directly regulates minus end extension and microtubule polarity, but that changes in microtubule dynamics may be secondary to stress signaling (Feng, 2019).
Persistent minus end growth facilitated by Patronin could be an evolutionary innovation in animals like Drosophila that have mostly minus-end-out dendrites. However, this study was able to visualize growing minus ends that behaved similarly to the ones in Drosophila dendrites in plus-end-out zebrafish sensory endings. If this phenomenon is widespread across evolutionarily distant taxa, then why has it not previously been reported? EB proteins have been used to visualize microtubule plus end growth in neurons since 2003, but slow-moving dots have not been noted before in neurons. One reason for this is likely improvements in sensitivity and signal-to-noise detection in microscopes used for live imaging. However, upon close inspection, there seem to be occasional minus ends in published EB data; for example, slow dim EB structures are present in kymographs from Drosophila and mammalian neurons. In the future, it will be important to investigate behavior of microtubule minus ends in neurons from other species in detail to determine whether minus ends grow for long periods and over substantial distances (Feng, 2019).
It is particularly interesting to compare minus end growth across species because of the expansion of the Patronin/CAMSAP family in vertebrates. In zebrafish, clear minus end growth is observed, but it is not yet known which of the five CAMSAPs (two each of CAMSAP1 and 2 and one CAMSAP3) in fish might be present at these growing minus ends. In contrast to CAMSAP2 and 3, which suppress minus end growth, CAMSAP1 tracks minus ends as they grow, so it is a good candidate. However, knockdown of CAMSAP1 has not been associated with changes in microtubule behavior in immortalized cell cultures and was not detected in neurons in the hippocampus. In contrast, CAMSAP2 knockdown has dramatic effects on microtubule stability in immortalized cells and neurons, so it is an open question what family member might be present on the growing minus ends tracked in zebrafish sensory neurons and how different types of CAMSAPs might cooperate or compete with one another (Feng, 2019).
The finding that Drosophila Patronin facilitates, rather than reduces, minus end growth in neurons is surprising in light of its previous characterization. Rather than tracking growing minus ends like CAMSAP1, it is thought to dampen minus end growth like CAMSAP2 and 3. One possible explanation for this discrepancy between Patronin behavior in vitro and in neurons is that in specific cell types or contexts, its behavior at minus ends could be regulated. In vitro, the coiled coil domain of Patronin tracks dynamic minus ends, and when the neighboring CKK domain is added, the behavior switches to growth suppression. In vivo, perhaps the effect of the CKK domain is modulated to uncover the minus end growth tracking ability of the coiled coil domain (Feng, 2019).
The neuronal phenotypes from reduction of Patronin are consistent with a role in minus end growth, and it is proposed that extended periods of minus end growth allow minus-end-out microtubules to grow into dendrites. Consistent with this idea, Patronin reduction resulted in fewer minus-end-out microtubules in distal regions of mature dendrites as well as in developing and regenerating dendrites. However, other mechanisms likely work in parallel with minus end growth to populate dendrites with microtubules (Feng, 2019).
Sliding of minus-end-out microtubules along plus-end-out ones by kinesin-6 motors was the mechanism first proposed to populate dendrites with minus-end-out microtubules in mixed polarity mammalian neurons, and the function of kinesin-6 has been confirmed more recently with newer methods. In Drosophila, these motors could play a role in dendritic microtubule organization early in development when microtubules in dendrites are mixed, but it is harder to imagine how they might function in mature minus-end-out dendrites. Analysis of Pavarotti, one of the Drosophila kinesin-6 motors, confirms that it is unlikely to play a major role in polarity of mature dendrites, and actually reduces sliding of microtubules by kinesin-1 into developing axons (Feng, 2019).
Local nucleation of microtubules in dendrites can also help populate dendrites with microtubules and, in contrast to sliding, has been shown to be important in Drosophila and C. elegans. In mammalian neurons, noncentrosomal microtubule nucleation also predominates and is known to be critical for axon growth and microtubule organization, though the localization and role of nucleation sites in dendrites has not been detailed so far. During C. elegans development, microtubule nucleation operates in parallel to minus end regulation by Patronin, and both types of minus end regulation may cooccur broadly in different cell types and species. In different contexts, nucleation or Patronin may vary in relative importance. For example, during initial dendrite outgrowth, no strong morphological defects were observed with Patronin knockdown, but during regeneration, dendrite growth was strongly perturbed. While differences in RNAi knockdown could account for the phenotypic differences, it is also possible that nucleation can compensate more completely for Patronin reduction in development compared with regeneration (Feng, 2019).
Overall, this study has demonstrated that, rather than existing as static structures, microtubule minus ends can grow in Drosophila and zebrafish neurons. Minus end growth is slow, but persistent, and up to 10 µ can be added in a single episode of growth. While both EB proteins and Patronin can bind to minus ends, they do so independently, and Patronin is critical for extended stretches of minus end growth. Without Patronin-mediated minus end growth, minus-end-out microtubules were reduced in dendrite tips and developing dendrites. Moreover, extra Patronin was sufficient to convert dendrite tips from mixed polarity to minus-end-out. Dendrite regeneration was particularly sensitive to Patronin reduction. It is concluded that minus end microtubule growth is likely to be broadly important in helping minus-end-out microtubules reach cellular regions that are distal to nucleation sites (Feng, 2019).
Dendrite arbor pattern determines the functional characteristics of a neuron. It is founded on primary branch structure, defined through cell intrinsic and transcription-factor-encoded mechanisms. Developing arbors have extensive acentrosomal microtubule dynamics, and this study reports an unexpected role for the atypical actin motor Myo6 in creating primary branch structure by specifying the position, polarity, and targeting of these events. In vivo time-lapse imaging was carried out of Drosophila adult sensory neuron differentiation, integrating machine-learning-based quantification of arbor patterning with molecular-level tracking of cytoskeletal remodeling. This revealed that Myo6 and the transcription factor Knot regulate transient surges of microtubule polymerization at dendrite tips; they drive retrograde extension of an actin filament array that specifies anterograde microtubule polymerization and guides these microtubules to subdivide the tip into multiple branches. Primary branches delineate functional compartments; this tunable branching mechanism is key to define and diversify dendrite arbor compartmentalization (Yoong, 2020).
Dendritic morphology is inextricably linked to neuronal function. Systematic large-scale screens combined with genetic mapping have uncovered several mechanisms underlying dendrite morphogenesis. However, a comprehensive overview of participating molecular mechanisms is still lacking. This study conducted an efficient clonal screen using a collection of mapped P-element insertions that were previously shown to cause lethality and eye defects in Drosophila melanogaster. Of 280 mutants, 52 exhibited dendritic defects. Further database analyses, complementation tests, and RNA interference validations verified 40 P-element insertion genes as being responsible for the dendritic defects. Twenty-eight mutants presented severe arbor reduction, and the remainder displayed other abnormalities. The intrinsic regulators encoded by the identified genes participate in multiple conserved mechanisms and pathways, including the protein folding machinery and the chaperonin-containing TCP-1 (CCT) complex that facilitates tubulin folding. Mutant neurons in which expression of CCT4 or CCT5 was depleted exhibited severely retarded dendrite growth. CCT is shown to localize in dendrites and is required for dendritic microtubule organization and tubulin stability, suggesting that CCT-mediated tubulin folding occurs locally within dendrites. This study also reveals novel mechanisms underlying dendrite morphogenesis. For example, it was shown that components of Drosophila Nogo signaling (POSH and Tango14), are required for dendrite development and that Mummy and Wech also regulate dendrite morphogenesis, potentially via Dpp- and integrin-independent pathways. Our methodology represents an efficient strategy for identifying intrinsic dendrite regulators, and provides insights into the plethora of molecular mechanisms underlying dendrite morphogenesis (Wang, 2020).
Microtubules (MTs) and F-actin (F-act) have long been recognized as key regulators of dendritic morphology. Nevertheless, precisely ascertaining their distinct influences on dendritic trees have been hampered until now by the lack of direct, arbor-wide cytoskeletal quantification. This study paired live confocal imaging of fluorescently labeled dendritic arborization (da) neurons in Drosophila larvae with complete multi-signal neural tracing to separately measure MTs and F-act. Dendritic arbor length is highly interrelated with local MT quantity, whereas local F-act enrichment is associated with dendritic branching. Computational simulation of arbor structure solely constrained by experimentally observed subcellular distributions of these cytoskeletal components generated synthetic morphological and molecular patterns statistically equivalent to those of real da neurons, corroborating the efficacy of local MT and F-act in describing dendritic architecture. The analysis and modeling outcomes hold true for the simplest (class I), most complex (class IV), and genetically altered (Formin3 overexpression) da neuron types (Nanda, 2020).
Kinetochores connect centromeric chromatin to spindle microtubules during mitosis. Neurons are post-mitotic, so it was surprising to identify transcripts of structural kinetochore (KT) proteins and regulatory chromosome passenger complex (CPC) and spindle assembly checkpoint (SAC) proteins in Drosophila neurons after dendrite injury. To test whether these proteins function during dendrite regeneration, post-mitotic RNAi was performed and dendrites or axons were removed using laser microsurgery. Reduction of KT, CPC and SAC proteins decreased dendrite regeneration without affecting axon regeneration. To understand whether neuronal functions of these proteins rely on microtubules, microtubule behavior was analyzed in uninjured neurons. The number of growing plus, but not minus, ends increased in dendrites with reduced KT, CPC and SAC proteins, while axonal microtubules were unaffected. Increased dendritic microtubule dynamics was independent of DLK-mediated stress, but was rescued by concurrent reduction of γTubulin, the core microtubule nucleation protein. Reduction of γTubulin also rescued dendrite regeneration in backgrounds containing kinetochore RNAi transgenes. It is concluded that kinetochore proteins function post-mitotically in neurons to suppress dendritic microtubule dynamics by inhibiting nucleation (Hertzler, 2020).
The kinetochore is a quintessential mitosisand meiosis-specific structure that attaches chromosomes to the mitotic spindle for segregation of genetic material to daughter cells. In many species, including Drosophila, it is built on centromeric DNA that is recognized by the histone Cid/Cenp-A that recruits inner kinetochore protein Cenp-C to serve as a binding site for the KMN network (Knl1, Mis12, and Ndc80 complexes). Of these, the Mis12 complex is most central, while the remainder make up more distal parts of the structure that is involved in connecting to microtubules. Many animals have other inner kinetochore proteins, including the constitutive centromere-associated network (CCAN), but these are absent in Drosophila (Drinnenberg, 2016). In cycling mammalian cells, inner centromere proteins including Cenp-A, Cenp-C, and CCAN are found on the centromere in interphase as well as mitosis. The Mis12 complex has some nuclear staining in interphase, but it is not as clearly punctate as CENP-A. In Drosophila, Cenp-A/Cid, Cenp-C, and Mis12 localize to centromeres in interphase and mitosis. In mammals and Drosophila, the other kinetochore components including the Knl1 and Ndc80 complexes are recruited to the kinetochore only in mitosis. Once the kinetochore is fully assembled, Nuf2 and Ndc80 in the outer kinetochore capture microtubule plus ends growing into the central spindle to allow chromosome segregation (Hertzler, 2020).
In addition to the kinetochore proteins that bridge chromosomes to microtubules, several regulatory complexes are present at the kinetochore before final bioriented microtubule attachment. The chromosome passenger complex (CPC) targets the kinase Aurora B to the centromere, where it helps correct errors in chromosome attachment to microtubules by phosphorylation of substrates including Ndc80. The targeting subunits of the CPC are borealin (or in flies borealin-related, borr), survivin (or in flies Deterin, Det), and Incenp, which are tightly associated with one another through a three-helix bundle. Aurora B is also critical for recruiting spindle assembly checkpoint (SAC) proteins to unattached kinetochores. The major effector of the SAC is cdc20/fzy, which is a subunit of the anaphase-promoting complex or cyclosome (APC/C). When bound by SAC subunits at the kinetochore, cdc20/fzy is inactive. A third complex, Rod1-Zw10-Zwilch (RZZ) complex, interacts with both SAC and KMN proteins. Together these regulatory complexes ensure that anaphase does not begin until all sister chromatids are correctly attached to opposite spindle poles. Attachment causes a shift in kinase and phosphatase balance, such that Aurora B starts to lose, and 'stripping' of the SAC by dynein connected to the RZZ complex is initiated. As a result, cdc20/Fzy is freed to activate APC/C, the ubiquitin ligase that triggers degradation of Cyclin B and Securin to initiate sister chromatid separation. KMN network proteins remain at the kinetochore to mediate attachment of separating sister chromatids to spindle microtubules (Hertzler, 2020).
Most of the proteins that make up the kinetochore and its associated regulatory complexes have not been linked to functions in interphase or postmitotic cells. However, two recent studies demonstrated neuronal defects after postmitotic reduction of kinetochore proteins, suggesting that they do function in noncycling cells (Cheerambathur, 2019; Zhao, 2019). In Caenorhabditis elegans, KMN network components localize to ciliated dendrites of amphid neurons and play a role in their extension (Cheerambathur, 2019). Although the microtubule-binding domains of Ndc80 are required to support normal amphid development, specific defects in microtubules were not detected (Cheerambathur, 2019). In Drosophila, mis12 mutants were identified in a genetic screen to identify modulators of synaptic bouton structure (Zhao, 2019). Reduction of other KMN network proteins had similar effects on neuromuscular junction structure, and intriguingly, so did targeting the centromeric histone cid/CENP-A (Zhao, 2019). Knockdown of Mis12 in primary rodent hippocampal neurons increased the number of protrusions from dendrites (Zhao, 2019), indicating that kinetochore proteins likely function in neurons broadly across evolution. Specific alterations in the neuronal cytoskeleton were not reported in this study either. It therefore remains to be determined how kinetochore proteins impact neuronal structure (Hertzler, 2020).
One of the most intense structural challenges neurons face is rebuilding axons or dendrites after injury. Key regulators of axon regeneration have been identified, including sensors like the dual leucine zipper kinase (DLK), epigenetic regulators like HDAC5, transcription factors including fos, jun, and cebp-1, and a myriad of downstream regeneration-associated genes (RAGS). However, it was only recently shown that neurons survive removal of all dendrites and can regrow a new arbor. Only a handful of genes including Akt and Ror have been linked to dendrite regeneration. To identify dendritic RAGs, RNA was isolated from Drosophila neurons after in vivo dendrite removal. Surprisingly, many kinetochore gene transcripts were coordinately up-regulated 6 h after dendrite injury. These included not only transcripts encoding structural kinetochore proteins in the inner kinetochore and KMN network, but also transcripts encoding regulatory proteins in the CPC, SAC, and RZZ complexes. Postmitotic RNA interference (RNAi) was used to confirm a role of some of these proteins in dendrite regeneration. In the same genetic backgrounds, no effect was seen on axon regeneration. In uninjured neurons, kinetochore protein reduction specifically increased microtubule plus-end number (also referred to as microtubule dynamics) in dendrites without altering other metrics of microtubule behavior such as polarity, speed, or minus-end density. Unexpectedly, axonal microtubule dynamics was unaffected. Therefore, kinetochore proteins act in a compartment-specific manner to suppress microtubule dynamics in dendrites. The dendritic microtubule phenotype and dendrite regeneration defect were rescued by concurrent reduction of γ-tubulin, suggesting that kinetochore proteins normally function to temper microtubule nucleation in dendrites. Together the data suggest that the KMN network and CPC, SAC, and RZZ complexes function together to promote dendrite regeneration and modulate dendritic microtubule dynamics through control of nucleation in postmitotic neurons (Hertzler, 2020).
On the basis of the surprising appearance of transcripts of structural and regulatory kinetochore proteins in RNASeq data from injured neurons, their role in dendrite regeneration was investigated. Regeneration after dendrite, but not axon, injury was impaired when they were reduced using postmitotic RNAi. Because kinetochore proteins function to attach, and monitor attachment, to microtubules in mitotic spindles, microtubule behavior was surveyed in neurons in which they were reduced. RNAi knockdowns of many kinetochore proteins caused an up-regulation of dendritic microtubule dynamics, an effect not seen in axons. Other parameters of microtubule behavior, including number of growing minus ends, were not affected. The increase in plus-end, but not minus-end, number suggested that nucleation rather than severing was responsible. The involvement of nucleation was confirmed by rescue of the plus end and regeneration phenotypes in neurons with reduced γ-tubulin. The increase in microtubule dynamics when kinetochore proteins were reduced suggests that they normally function to dampen nucleation of dendritic microtubules in postmitotic neurons (Hertzler, 2020).
Two recent studies indicate that kinetochore proteins are likely to function broadly in different neuron types. In one study, the authors identified Mis12 mutants in a Drosophila forward genetic screen. Other components of the KMN network complexes (Ndc80, Knl1, and Nnf1a), as well as centromeric protein Cenp-A (Cid), had similar neuromuscular junction phenotypes and reduced neuropil in the CNS (Zhao, 2019). The authors went on to show that mammalian neurons with reduced Mis12 also had structural defects, this time in dendrites (Zhao, 2019). The group reported some localization of KMN network proteins to spots in peripheral nerves and neuropil in the CNS (Zhao, 2019), but the pattern did not help to suggest how the proteins might function in neurons (Hertzler, 2020).
The other study demonstrated that Cenp-C, Ndc80, and Nuf2 localize postmitotically to the ciliated dendrites of amphid neuron bundles in C. elegans. When these proteins were knocked down with an elegant GFP-degrader system, amphid dendrite extension was impaired (Cheerambathur, 2019). Effects on egg laying and fertility were also seen. Notably, a deletion of only the microtubule-binding domains of Ndc80 was able to phenocopy the knockdowns, indicating that an interaction with microtubules likely mediates the phenotypes. Degradation of GIP2, an essential γTuRC subunit, showed the same neuron extension deficits, supporting the hypothesis that microtubules in these neurons are affected by KMN network protein knockdown, although the authors were unable to pinpoint any specific changes in the microtubule cytoskeleton (Hertzler, 2020).
The two previous studies on neuronal roles for kinetochore proteins focused on the structural components of the kinetochore, including inner centromere components Cenp-A and C and KMN network proteins (Cheerambathur, 2019; Zhao, 2019). The three regulatory complexes-CPC, RZZ, and SAC- are added to the list of kinetochore proteins with neuronal function. Moreover, it was found that players in all structural and regulatory complexes have similar roles in controlling microtubule dynamics. This suggests that, as in mitosis, these complexes function together in a single pathway in neurons. It is therefore likely that they are not only involved in attaching microtubules to something, but are also acting as sensors of microtubule behavior. If they have a similar role in neurons as in mitosis, they may recognize plus-end arrival at a specific cellular location. Again, in analogy with mitosis, plus-end arrival could trigger release of regulatory complexes and free them to send signals to other cellular locations about the status of the microtubule cytoskeleton. These signals could involve ubiquitination mediated by association of Cdc20/Fzy with an E3 ligase complex, or phosphorylation by Aurora B. One target of either output signal could be suppression of γTuRC activity (Hertzler, 2020).
One intriguing aspect of these findings is that changes were only observed in the microtubule cytoskeleton in dendrites, and dynamics in the axon was unaffected. While phenotypes in C. elegans and mammalian neurons occurred in dendrites, changes in motor axon terminals were seen in Drosophila. One possible way to reconcile the axonal and dendritic phenotypes would be if kinetochore proteins are important throughout dendrites but function specifically at synaptic regions of axons. There are several hints that microtubule nucleation might be particularly important near presynaptic sites. In Drosophila motor neurons, concentrations of γ-tubulin were seen in large synaptic boutons at the neuromuscular junction. In cultured mammalian neurons, γ-tubulin also concentrates at presynaptic sites, and increases in neuronal activity lead to increases in EB comet formation at these sites, suggesting that regulation of nucleation is important in axons. It is therefore possible that kinetochore proteins regulate presynaptic nucleation in addition to dendritic nucleation (Hertzler, 2020).
In mitotic cells, kinetochore proteins localize to tight spots around centromeric DNA, and their localization has provided invaluable clues to their function. So far, localization patterns in neurons have not helped pin down specific sites of action. In C. elegans, expression levels in amphid neurons were high enough to detect tagged proteins expressed at endogenous levels, and they were seen throughout the linear ciliated dendrite region (Cheerambathur, 2019). In contrast, expression seems low in Drosophila neurons, and beyond being present in neuropil, a pattern has not been discernible (Zhao, 2019). Attempts of this study to visualize meaningful localization in sensory neurons also did not provide any insights. Use of amplification systems like SunTag may be required to acquire meaningful information about where structural and regulatory kinetochore proteins function in neurons (Hertzler, 2020).
One possible model for neuronal kinetochore protein function is that, in analogy with mitosis, the structural and regulatory complexes colocalize at a specific cellular site, perhaps in the cell body, when microtubule plus ends are absent or in low abundance. Upon plus-end arrival, the RZZ complex in conjunction with dynein could transport regulatory proteins toward microtubule minus ends. Dendrites contain minus-end-out microtubules, so this would mean that regulatory proteins could be transported outward into dendrites to suppress microtubule dynamics distally after plus-end arrival. This type of mechanism could allow for global homeostatic control of microtubule dynamics in dendrites (Hertzler, 2020).
The exocyst complex is an important regulator of intracellular trafficking and tethers secretory vesicles to the plasma membrane. Understanding of its role in neuron outgrowth remains incomplete, and previous studies have come to different conclusions about its importance for axon and dendrite growth, particularly in vivo. To investigate exocyst function in vivo Drosophila sensory neurons were used as a model system. To bypass early developmental requirements in other cell types, neuron-specific RNAi was used to target seven exocyst subunits. Initial neuronal development proceeded normally in these backgrounds, however, this was considered to be due to residual exocyst function. To probe neuronal growth capacity at later times after RNAi initiation, laser microsurgery was used to remove axons or dendrites and prompt regrowth. Exocyst subunit RNAi reduced axon regeneration, although new axons could be specified. In control neurons, a vesicle trafficking marker often concentrated in the new axon, but this pattern was disrupted in Sec6 RNAi neurons. Dendrite regeneration was also severely reduced by exocyst RNAi, even though the trafficking marker did not accumulate in a strongly polarized manner during normal dendrite regeneration. The requirement for the exocyst was not limited to injury contexts as exocyst subunit RNAi eliminated dendrite regrowth after developmental pruning. It is concluded that the exocyst is required for injury-induced and developmental neurite outgrowth, but that residual protein function can easily mask this requirement (Swope, 2022).
Dendrite microtubules are polarized with minus-end-out orientation in Drosophila neurons. Nucleation sites concentrate at dendrite branch points, but how they localize is not known. Using Drosophila, this study found that canonical Wnt signaling proteins regulate localization of the core nucleation protein gammaTubulin (gammaTub). Reduction of frizzleds (fz), arrow (low-density lipoprotein receptor-related protein [LRP] 5/6), dishevelled (dsh), casein kinase Igamma, G proteins, and Axin reduced gammaTub-green fluorescent protein (GFP) at branch points, and two functional readouts of dendritic nucleation confirmed a role for Wnt signaling proteins. Both dsh and Axin localized to branch points, with dsh upstream of Axin. Moreover, tethering Axin to mitochondria was sufficient to recruit ectopic gammaTub-GFP and increase microtubule dynamics in dendrites. At dendrite branch points, Axin and Dsh colocalized with early endosomal marker Rab5, and new microtubule growth initiated at puncta marked with Fz, Dsh, Axin, and Rab5. It is proposed that in dendrites, canonical Wnt signaling proteins are housed on early endosomes and recruit nucleation sites to branch points (Weiner, 2020).
Neurons extend long branched processes from a central cell body. This shape is incompatible with a centrosomal microtubule organizing center (MTOC). Mature neurons are therefore among the ranks of differentiated cells that have noncentrosomal microtubule arrays. It is particularly important to understand how neuronal microtubules are organized because the distance from the primary site of synthesis in the cell body to functional sites in axons and dendrites can be large and, therefore, place heavy demands on microtubule-based transport. In humans, slight disruptions in microtubule regulators or motors can manifest as neurodegenerative disease, underscoring neuronal reliance on perfectly orchestrated microtubule-based transport (Weiner, 2020).
If neuronal microtubules are not anchored to the centrosome, how are they organized? In all neurons so far examined, axonal microtubules have their dynamic plus ends oriented away from the cell body (plus-end-out). In dendrites of vertebrate neurons, microtubules are mixed polarity. In invertebrate neurons (Drosophila and Caenorhabditis elegans), axons have the same plus-end-out microtubule organization as vertebrates, but mature dendrites have almost all minus-end-out microtubules. In immature Drosophila dendrites, microtubules are mixed polarity and only gradually resolve to the minus-end-out mature arrangement. Thus, although the final arrangement of microtubules in vertebrate and invertebrate dendrites is somewhat different, they are the same during dendrite development (Weiner, 2020).
Although the arrangement of neuronal microtubules is clearly noncentrosomal, the source of axonal and dendritic microtubules has been controversial. Two major models for generating axonal and dendritic microtubules have been proposed. The first is that neuronal microtubules are nucleated at the centrosome, or perhaps elsewhere in the cell body, and then released for transport/sliding into axons and dendrites. This model has substantial support, including recent analyses with newer techniques. For example, live imaging of microtubules with plus-tip (+TIP) tracking proteins and photoconvertible αTubulin has provided evidence for directional transport of microtubules into and out of developing axons in mammalian and Drosophila neurons (Weiner, 2020).
The second model is that nucleation sites are found outside the cell body and that microtubules are generated locally in axons and dendrites. Evidence for this model came from the observation that centrosomal γTubulin (γTub), the core microtubule nucleation protein, decreases gradually over time and that centrosome ablation does not disrupt axon formation. Similarly, the centriole is not surrounded by γTub in Drosophila neurons in vivo, and it is dispensable for neuronal microtubule organization. One way to reconcile these two models is to assume that both are important and that, very early in neuronal development, microtubule sliding can dominate, whereas later in development and in mature neurons, microtubules are primarily locally nucleated (Weiner, 2020).
In some cell types, the Golgi complex recruits nucleation sites, and small Golgi outposts can be found in both mammalian and Drosophila dendrites. Thus, it was proposed that the Golgi might act as a noncentrosomal MTOC in dendrites. However, subsequent analysis of γTub and Golgi outposts, including a strategy to deplete Golgi from dendrites, called this proposal into question (Weiner, 2020).
Within Drosophila dendrites, γTub is concentrated at branch points. A previous study identified proteins that localize a different microtubule regulator, adenomatous polyposis coli (Apc) 2, to branch points. It was reasoned that some or all of this machinery might be used to position γTub to the same region. Whether any of the Apc2 localization proteins act upstream of γTub-green fluorescent protein (GFP) in dendrites was tested. Surprisingly, a subset of Wnt signaling proteins was required to localize γTub-GFP to dendrite branch points, regulate dendritic microtubule polarity, and nucleate microtubules in dendrites in response to axon injury. The required proteins include the seven transmembrane domain Frizzled (Fz) proteins (Wnt receptors), Arrow (Arr, a Wnt coreceptor), heterotrimeric G proteins, Dishevelled (Dsh), Casein kinase I (CK1)γ, and Axin. Axin seems to be the key output protein of this pathway because it was sufficient to recruit γTub to ectopic sites in dendrites. Within branch points, Fz, Axin, and Dsh were found on puncta that colocalized with Rab5. In addition, new end-binding protein 1 (EB1) comets at polymerizing microtubule plus ends initiated from puncta marked with Fz, Arr, Dsh, Axin, and Rab5. It is proposed that Wnt signaling proteins localize to early endosomes at dendrite branch points and function there to control local microtubule nucleation. Although it has previously been shown that Wnt signaling proteins can function from endosomes, identification of microtubule nucleation as an output of endosomal Wnt proteins is quite unexpected (Weiner, 2020).
It was particularly intriguing to find integral membrane signaling proteins required for noncentrosomal microtubule nucleation. Although Wnt signaling has been linked to microtubule plus-end regulation in axon growth cones and regulation of microtubule stability and spindle orientation, the only connection to the minus end is localization of some cytoplasmic Wnt signaling proteins like Axin to the centrosome in dividing cells. This study demonstrated that a Wnt signaling pathway acts upstream of microtubule nucleation in a postmitotic cell. Not only were many canonical Wnt signaling proteins required for γTub-GFP to accumulate at branch points, but Axin and Dsh themselves concentrated at branch points. In addition, the scaffolding protein Axin was able to recruit γTub-GFP and the nucleation activator Cnn to mitochondria when tethered to them. Moreover, reduction of Wnt signaling proteins phenocopied loss of γTub in two functional nucleation assays, indicating that most or all dendritic nucleation occurs downstream of this pathway. Although this pathway seems to be the major regulator of dendritic nucleation, neurons are quite resilient to its loss under baseline conditions, and the simple ddaE neurons have normal arbor shape. This is likely because parallel pathways can be used to generate new minus ends. For example, microtubule severing can be used to generate new plus and minus ends and amplify microtubule number. In many cell types, minus ends generated when a microtubule is severed are recognized by minus-end binding proteins in the calmodulin-regulated spectrin-associated protein (CAMSAP)/Patronin family. In C. elegans, γTub-mediated microtubule nucleation has been shown to act in parallel and quite redundantly with Patronin to regulate microtubule organization. Recent work has shown that Patronin-mediated minus-end growth is an important regulator of dendritic microtubules in Drosophila, so it is possible that microtubule severing in conjunction with Patronin recruitment to minus ends can compensate for nucleation under most normal circumstances. Consistent with this hypothesis, phenotypes from reduction of nucleation or Patronin become more evident after severe stress, including axon [29] or dendrite [77, 98] injury (Weiner, 2020).
Although it was consistently found that partial loss of function (RNAi or heterozygous mutants) for fz, fz2, arr, dsh, Gao, Gas, and Axin reduced γTub localization and/or function, no evidence was found that β-catenin/Arm, the key transcription factor that is the output of canonical Wnt signaling, was involved. In addition, an Arm protein trap showed clear expression in epidermal cells but was not seen in dendritic arborization neurons (da neurons). Because Axin itself was sufficient to recruit γTub, there was no strong rationale for a transcriptional regulator to mediate signaling between fz/arr and microtubule nucleation. It is proposed that canonical Wnt signaling proteins are co-opted in dendrites to directly recruit nucleation complexes to endosomes. Because this is a variant of canonical Wnt signaling that unexpectedly seems not to involve β-catenin, this pathway is termed apocryphal Wnt signaling in reference to the Apocrypha, ancient writings found in only some versions of the Bible (Weiner, 2020).
The involvement of arr as well as dsh and Axin suggests that a signalosome might be involved in dendritic Wnt signaling. Signalosomes form when wnt ligands bind to Fz and LRP5/6 at the plasma membrane, triggering recruitment and multimerization of Dsh and Axin. The normal output of signalosome formation is release of β-catenin from the destruction complex and its subsequent stabilization and transit to the nucleus to activate transcription. Signalosomes assemble at the plasma membrane. Endocytosis generally seems to promote Wnt signaling, although in many contexts the signalosome itself is disassembled upon endocytosis. It is not clear whether signalosomes persist after endocytosis, though in some Drosophila cells, Dsh and Arr are localized to endosomes. In dendrites, puncta of Fz, Dsh, and Axin colocalized with Rab5, suggesting that a stable signaling complex is present on endosomes in mature neurons. The initiation of comets from these puncta indicates that endosomes are likely the key site where Wnt signaling proteins promote nucleation. Colocalization of tagged Golgi proteins with Rab5 suggests that the previous association between Golgi markers and nucleation could have been due to leakage into endosomes. In addition, the identification of plasma membrane proteins acting upstream of γTub in dendrites suggests a more general role for the Golgi in the cell body by controlling secretion of Arr and Fz (Weiner, 2020).
Wnt signaling receptors have been classically studied at the plasma membrane, where they bind extracellular ligands that can be autocrine or paracrine in nature. A requirement for arr and fz upstream of γTub in dendrites suggests that a Wnt ligand is likely involved. Failure of neuronal wntless knockdown to reduce γTub-GFP at branch points favors the hypothesis that the ligand may be secreted from a neighboring cell. In the embryo, wingless (wg)/Wnt-1 is made in a patch of epithelial cells adjacent to developing dendritic arborization neurons and helps pattern dendrite orientation in ddaE . It would be very interesting if surrounding cells influenced the microtubule cytoskeleton in mature neurons through Fz and Arr at the plasma membrane. This signaling pathway is particularly intriguing in the context of regeneration or during neurodegenerative disease. During axon regeneration, the initial injury response involves a nucleation-dependent increase in microtubule dynamics, which serves a neuroprotective role. Modulating Wnt signaling could therefore influence neuroprotection in dendrites. In addition, this study found that this pathway is required during dendrite regeneration to position nucleation sites in regrowing dendrites. Interestingly, G protein coupled receptors (GPCRs) represent 33% of all Food and Drug Administration-approved drug targets, and as part of this family, Fz presents a possible target (Weiner, 2020).
Local microtubule nucleation also occurs in axons. As Rab5 endosomes are present throughout axons, it will be interesting to determine whether Wnt signaling proteins can be recruited to axonal early endosomes and whether they recruit nucleation proteins in this part of the cell. It is also possible that a link between Wnt signaling, endosomes, and nucleation could exist more broadly in other cell types. Indeed, the localization of Axin to centrosomes suggests that even in mitotic cells, parts of this relationship are conserved. Intriguingly, endosomal membranes are concentrated around the centrosome, and Rab5 reduction disrupts mitosis, so it is possible that Wnt signaling proteins, endosomes, and nucleation function together at centrosomes (Weiner, 2020).
VAMP-associated protein (VAP; see Drosophila Vap33) is an endoplasmic reticulum (ER) membrane protein that functions as a tethering protein at the membrane contact sites between the ER and various intracellular organelles. Mutations such as P56S in human VAPB cause neurodegenerative diseases such as amyotrophic lateral sclerosis (ALS). However, VAP functions in neurons are poorly understood. This study utilized Drosophila olfactory projection neurons with a mosaic analysis with a repressible cell marker (MARCM) to analyze the neuronal function of VAP33, a Drosophila ortholog of human VAPB. In vap33 null mutant clones, the dendrites of projection neurons exhibited defects in the maintenance of their morphology. The subcellular localization of the Golgi apparatus and mitochondria were also abnormal. These results indicate that Vap33 is required for neuronal morphology and organelle distribution. Additionally, to examine the impact of ALS-associated mutations in neurons, human VAPB-P56S was overexpressed in vap33 null mutant clones (mosaic rescue experiments) and found that, in aged flies, human VAPB-P56S expression caused mislocalization of Bruchpilot, a presynaptic protein. These results implied that synaptic protein localization and ER quality control may be affected by disease mutations. This study provides insights into the physiological and pathological functions of VAP in neurons (Kamemura, 2021).
Dendrite
shape impacts functional connectivity and is mediated by organization
and dynamics of cytoskeletal fibers. Identifying the molecular factors
that regulate dendritic cytoskeletal architecture is therefore important
in understanding the mechanistic links between cytoskeletal
organization and neuronal function. This study identified Formin 3 (Form3) as an essential regulator of cytoskeletal architecture in nociceptive sensory neurons
in Drosophila larvae. Time course analyses reveal that Form3 is
cell-autonomously required to promote dendritic arbor complexity. form3 is required for the maintenance of a population of stable dendritic microtubules (MTs),
and mutants exhibit defects in the localization of dendritic
mitochondria, satellite Golgi, and the TRPA channel Painless. Form3
directly interacts with MTs via FH1-FH2 domains. Mutations in human
inverted formin 2 (INF2; ortholog of form3) have been causally
linked to Charcot-Marie-Tooth (CMT) disease. CMT sensory neuropathies
lead to impaired peripheral sensitivity. Defects in form3
function in nociceptive neurons result in severe impairment of noxious
heat-evoked behaviors. Expression of the INF2 FH1-FH2 domains partially
recovers form3 defects in MTs and nocifensive behavior,
suggesting conserved functions, thereby providing putative mechanistic
insights into potential etiologies of CMT sensory neuropathies (Das, 2021).
It has long been thought that microtubule disassembly, one of the earliest cellular events, contributes to neuronal pruning and neurodegeneration in development and disease. However, how microtubule disassembly drives neuronal pruning remains poorly understood. A systematic investigation was undertaken of various microtubule-destabilizing factors and exchange factor for Arf6 (Efa6) and Stathmin (Stai) were identified as new regulators of dendrite pruning in ddaC sensory neurons during Drosophila metamorphosis. Efa6 was shown to be both necessary and sufficient to regulate dendrite pruning. Interestingly, Efa6 and Stai facilitate microtubule turnover and disassembly prior to dendrite pruning without compromising the minus-end-out microtubule orientation in dendrites. Moreover, the pharmacological and genetic manipulations strongly support a key role of microtubule disassembly in promoting dendrite pruning. Thus, this systematic study highlights the importance of two selective microtubule destabilizers in dendrite pruning and substantiates a causal link between microtubule disassembly and neuronal pruning (Bu, 2021).
To reshape neuronal connectivity in adult stages, Drosophila sensory neurons prune their dendrites during metamorphosis using a genetic degeneration program that is induced by the steroid hormone ecdysone. Metamorphosis is a nonfeeding stage that imposes metabolic constraints on development. AMP-activated protein kinase (AMPK), a regulator of energy homeostasis, is cell-autonomously required for dendrite pruning. AMPK is activated by ecdysone and promotes oxidative phosphorylation and pyruvate usage, likely to enable neurons to use noncarbohydrate metabolites such as amino acids for energy production. Loss of AMPK or mitochondrial deficiency causes specific defects in pruning factor translation and the ubiquitin-proteasome system. These findings distinguish pruning from pathological neurite degeneration, which is often induced by defects in energy production, and highlight how metabolism is adapted to fit energy-costly developmental transitions (Marzano, 2021).
Axons and dendrites are distinguished by microtubule polarity. In Drosophila, dendrites are dominated by minus-end-out microtubules while axons contain plus-end-out microtubules. Local nucleation in dendrites generates microtubules in both orientations. To understand why dendritic nucleation does not disrupt polarity, this study used live imaging to analyze the fate of microtubules generated at branch points. It was found that they had different rates of success exiting the branch based on orientation: correctly oriented minus-end-out microtubules succeeded in leaving about twice as often as incorrectly oriented microtubules. Increased success relied on other microtubules in a parallel orientation. From a candidate screen, Trim9 and kinesin-5 (Klp61F) were identified as machinery that promoted growth of new microtubules. In S2 cells, EB1 recruited Trim9 to microtubules. Klp61F promoted microtubule growth in vitro and in vivo, and could recruit Trim9 in S2 cells. In summary, the data argue that Trim9 and kinesin-5 act together at microtubule plus ends to help polymerizing microtubules parallel to pre-existing ones resist catastrophe (Feng, 2021).
The almost uniform minus-end-out microtubule polarity in Drosophila dendrites has proven a useful system for the identification of mechanisms that control microtubule organization and polarity. One surprise is that multiple mechanisms operate in parallel even in this very confined space with simple microtubule layout. Two basic types of polarity control mechanisms have been identified: those that can establish microtubule polarity independently of existing microtubules, and those that act as positive feedback loops to reinforce the predominant polarity (Feng, 2021).
Two mechanisms have been identified that can autonomously contribute to dendritic minus-end-out microtubule polarity: local nucleation and minus-end growth. Very early in the development of dendritic arborization neurons in the Drosophila embryo, new dendrites are populated by plus-end-out microtubules that grow in from the cell body (Feng, 2019). The next step is for slower-growing minus ends to enter dendrites from the cell body adding minus-end-out microtubules. Dendrites then remain with mixed polarity for the rest of embryogenesis, and eventually this resolves to minus-end-out in larvae. Minus-end-out microtubules can also be generated locally in dendrites by nucleation. Like microtubule growth from the cell body, nucleation can contribute plus-end-out and minus-end-out microtubules. In mature dendritic arborization neurons, nucleation at branch points is biased towards generating minus-end-out microtubules. Several mechanisms to bias nucleation have been shown to operate in neurons, but they have not been shown to act in mature dendritic arborization neurons. In developing C. elegans sensory dendrites, nucleation sites cluster close to the tip, resulting in a short region of plus-end-out microtubules beyond the cluster and the proximal dendrite dominated by minus-end-out microtubules (Liang, 2020). However, in ddaE dendrites, nucleation sites are found not just at the dendrite tip but throughout the arbor at branch points. In ddaE dendrites, nucleation has been proposed to be biased by recruitment to only one side of Golgi outposts. However, nucleation sites have recently been shown to be recruited to endosomes rather than Golgi in ddaE dendrites (Weiner, 2020), making the earlier findings difficult to interpret. In summary, both growth from microtubule ends and nucleation of new microtubules create minus-end-out, as well as plus-end-out, microtubules in dendrites (Feng, 2021).
In addition to the de novo mechanisms that add minus-end-out microtubules to dendrites described above, feedback mechanisms reinforce the dominant polarity. One of these controls the direction of microtubule growth at branch points. As microtubules grow from distal regions of the dendrite into branch points, they encounter a choice to grow towards the cell body or away from the cell body. Kinesin-2, together with Eb1, Apc and Apc2, allows the growing plus end to track existing microtubules, reinforcing polarity. If this mechanism is eliminated, polarity in dendrites remains mixed. The final percentage of minus-end-out microtubules seems to depend on the angle of branches such that in ddaE dendrites, in which branch angles are close to 90° and microtubules can easily turn either direction at branch points, ~50% of microtubules are minus-end-out. In dendrites with more acute branch angles that help direct growing plus ends towards the cell body, 70% of microtubules remain minus-end-out, even when steering is eliminated. The mechanism described in this study, that selectively promotes growth of new microtubules out of branch points along parallel microtubules, seems to act as an additional feedback loop that helps align newly nucleated microtubules with pre-existing ones. Impairing this mechanism seems to have little impact on overall polarity in dendrites, unlike eliminating steering. Quality control of newly nucleated microtubules may have a small effect on overall polarity because nucleation itself is somewhat biased and nucleation may contribute at relatively low levels to the overall microtubule population at steady state in mature neurons. Quality control of new microtubules may exist as a backup mechanism to help maintain microtubule organization under stressful conditions like axon injury, which can upregulate nucleation (Feng, 2021).
The data suggest that Trim9 and Klp61F are both needed for new microtubules to grow in parallel bundles with pre-existing microtubules. The fact that reduction of either results in a phenotype suggests that they act together, rather than in parallel, to control plus-end behavior. A vertebrate Trim9 family member, Trim46, organizes parallel microtubules at the axon initial segment and has autonomous parallel bundling activity in vitro. A role for the single Drosophila Trim9 family member in parallel orientation of microtubules suggests that this may be an ancestral function of this family, rather than a new function that evolved with the expansion of the family in vertebrates. If this is the case, then the other vertebrate family members in the C-1 subfamily containing a microtubule-binding COS box may all engage parallel microtubules, perhaps in different cell types or subcellular regions. In vitro, Trim46 has a strong preference for interacting with parallel bundles of microtubules over individual microtubules, and so tends to accumulate some distance behind dynamic plus ends. When a depolymerizing plus end encounters a bundled region decorated with Trim46, catastrophe is strongly inhibited. Although the ability of Trim46 to rescue catastrophes would be expected to promote growth like Drosophila Trim9, the site of action seems to be somewhat different. Trim46 acts at bundled regions of microtubules behind the dynamic plus end, and Drosophila Trim9 prevents catastrophes from happening, likely at the plus end, with Eb1 and Klp61F (Feng, 2021).
The in vivo role of Klp61F in promoting continued growth of plus ends out of the branch point is consistent with previous in vitro studies showing that Eg5 dimers can accumulate at the plus end and promote polymerization. Klp61F tetramers form rods ~95 nm in length that crosslink microtubules with spacing of at least 60 nm. The spacing of microtubules bundled with Trim46 is ~37 nm. Based on these general size considerations, as well as in vitro activities of Klp61F and Trim46, the following model is proposed. Klp61F could be constantly traveling along stable bundles of microtubules. If a new microtubule approaches within 60 nm, then it could be captured. As Klp61F interacts with the growing plus end it could promote polymerization. At the same time, Trim9 could be recruited to the growing plus end by Eb1, perhaps through [S/T]-x-[I/L]-P motifs, which interact with Eb1. Both Drosophila Trim9 proteins contain two potential motifs at amino acids: the first is at 42-SALP-46 in both, and the second is 479-TILP-483 in the RB isoform and 468-TILP-472 in the RA isoform. After Klp61F grabs the growing microtubule with Eb1 and Trim9 at its tip, Trim9 could reinforce the parallel interaction. Although this model provides an initial framework for the function of Klp61F and Trim9 in dendrites, it is quite speculative and raises many additional questions. For example, if Klp61F is in constant flux towards the plus end of microtubules, how is it transported into dendrites? And do these proteins really act sequentially? If so, perhaps family members collaborate in a similar way in other cellular scenarios; for example, at the axon initial segment (Feng, 2021).
mRNP granules at adult central synapses are postulated to regulate local mRNA translation and synapse plasticity. However, they are very poorly characterized in vivo. This study presents early observations and characterization of candidate synaptic mRNP particles in Drosophila olfactory synapses; one of these particles contains a widely conserved, DEAD-box helicase, Me31B. In Drosophila, Me31B is required for translational repression of maternal and miRNA-target mRNAs. A role in neuronal translational control is primarily suggested by Me31B's localization, in cultured primary neurons, to neuritic mRNP granules that contain: (1) various translational regulators; (2) CaMKII mRNA; and (3) several P-body markers including the mRNA hydrolases, Dcp1, and Pcm/Xrn-1. In adult neurons, Me31B localizes to P-body like cytoplasmic foci/particles in neuronal soma. In addition it is present to synaptic foci that may lack RNA degradative enzymes and localize predominantly to dendritic elements of olfactory sensory and projection neurons (PNs). MARCM clones of PNs mutant for Me31B show loss of both Me31B and Dcp1-positive dendritic puncta, suggesting potential interactions between these granule types. In PNs, expression of validated hairpin-RNAi constructs against Me31B causes visible knockdown of endogenous protein, as assessed by the brightness and number of Me31B puncta. Knockdown of Me31B also causes a substantial elevation in observed levels of a translational reporter of CaMKII, a postsynaptic protein whose mRNA has been shown to be localized to PN dendrites and to be translationally regulated, at least in part through the miRNA pathway. Thus, neuronal Me31B is present in dendritic particles in vivo and is required for repression of a translationally regulated synaptic mRNA (Hillebrand, 2010).
The Me31B/Dhh1p/DDX6/CGH-1 class of DEAD box helicases is associated with many different kinds of mRNP aggregates, including maternal RNA storage granules, P-bodies, stress granules, as well as various granule subtypes observed during C. elegans germline development. In addition it is required for the assembly of P-bodies in yeast, Drosophila and mammalian cells and as well for the formation of stress granules in mammals. For these reasons, the punctate distribution of Me31B in postsynaptic dendrites is likely to indicate its presence in a specific type of synaptic mRNP particle. However, unlike Me31B-positive particles described in neurites of cultured Drosophila neurons, synaptic Me31B foci do not appear to contain the RNA hydrolases Dcp1 and Pcm/Xrn-1. Thus, they may be a distinct class of particle, which localize preferentially to postsynaptic dendrites. These represent early images of candidate mRNA storage particles at synapses in vivo. A paucity of antibodies and the challenging nature of such high-resolution immunocytochemistry in whole brain tissue has so far made it difficult to more completely characterize other components of synaptic Me31B particles as well as to establish whether Dcp1, Pcm, and Stau coexist on the same or different particle in the adult brain. Indeed even the conclusion that Me31B particles constitute a separate class must be qualified by the possibility that the visualization of two apparently distinct particle types arises from an artifact of incomplete antibody penetration into the neuropil (Hillebrand, 2010).
It is possible that synaptic Me31B particles could be analogous to recently described granules in the C. elegans germline, which contain translationally controlled mRNAs and CGH-1/Me31B but exclude decapping enzymes and the P-body protein PATR1/PAT1. Immunoprecipitation and further colocalization studies suggest that these granules can also contain PAB-1, ATX-2, or TIA-1, markers of stress granules, which in other systems, contain translation initiation factors together with mRNAs stalled in translational initiation. Thus, it is conceivable that Me31B/CGH-1-containing storage particles contain mRNAs stored in a stress-granule like state, in which the resident mRNAs are available for rapid activation (Hillebrand, 2010).
The potential separation of storage and degradative particles leads to an attractive model in which individual mRNAs may transition from being available for translational activation in a storage granule, to being targeted for degradation in a P-body like particle. This is supported as well by observations in dendrites of cultured mammalian neurons where a distinct class of RNPs contain the degradative enzyme Xrn1, which is excluded from RNPs supposedly involved in storage (Hillebrand, 2010).
At synapses, a transition between storage and degradation particles may occur by three, non-exclusive, candidate mechanisms: (1) by the remodeling of a storage mRNP to a degradative one through protein exchange; (2) by the initial exit of mRNA from the storage RNP to a translating pool, followed by its subsequent targeting to a degradative particle; or (3) the fusion of the two particles. Recent studies in Drosophila provide a possible mechanism by which a change of proteins in RNP complexes could alter its function. Two related proteins of the Lsm-family, Enhancer of Decapping 3 (EDC3), which is implicated to play a role in mRNA decay, and Trailer Hitch (Tral), which supposedly is involved in mRNA repression, interact at the same domain with the Me31B protein. This suggests that the function of Me31B complexes might be determined by the interaction with specific binding partners. Some support for the second model is provided by the observation that the synaptically localized Arc mRNA is targeted for degradation after its translation is induced by synaptic activity and also by the observation that RCK-positive particles in dendrites of cultured hippocampal neurons are transiently disassembled following BDNF stimulation. Further studies are required to understand how, when, and even whether these transitions of mRNA state occur in synapses and other biological contexts (Hillebrand, 2010).
Together with many analogous studies in yeast and mammalian cells, previous observations in Drosophila that Me31B is a repressor of maternal mRNA translation, a component of a repression pathway mediated by the bantam microRNA, and a repressor of growth of terminal dendrites, has led to a strong model that Me31B is a translational repressor protein. In contrast, recent studies in C. elegans and P. falciparum have shown that Me31B orthologs, CGH-1 and DOZI, associate with specific mRNAs and protects them from degradation (Hillebrand, 2010).
The observations in neurons indicate a function for Me31B in repressing translation of a miRNA regulated, dendritically localized reporter mRNA in vivo. This is consistent with two related lines of data. First, it is consistent with the known function for Me31B in repression of miRNA-target genes in Drosophila wing imaginal cells as well as for its human homolog RCK in mammalian cultured cells. Second, the correlation observed between loss of synaptic Dcp1 puncta and upregulation of the CaMKII reporter, is consistent with observations in hippocampal cultured cells, where observed disassembly of 'dendritic P-bodies' induced by synaptic stimulation has been proposed to underlie the temporally coincident translation of localized mRNAs (Hillebrand, 2010).
Thus, this study suggestd a simple model in which neuronal Me31B, as well as its homologs in other metazoa, mediates the formation of synaptic mRNP particles that contain locally repressed mRNAs. And that synaptic stimulation-induced disassembly of these particles is one aspect of the mechanism of local translational control (Hillebrand, 2010).
One key goal of future studies will be to understand the composition and dynamics of dendritic mRNPs in vivo. This will be aided by genetic techniques to replace endogenous translational control molecules with genetically encoded, fluorescently tagged variants that retain functional and localization patterns of the endogenous proteins. When coupled with procedures to induce local protein synthesis in dendrites, such reagents will allow analysis of functionally relevant particle dynamics in vivo. In addition, by eliminating the need for antibodies whose use may be associated with artifacts of inclusion and exclusion, such reagents may provide more direct insight into the real nature of synaptic mRNPs in vivo (Hillebrand, 2010).
A second goal is to understand the mechanism by which Me31B regulates the expression of CaMKII reporter levels in vivo. Although Me31B has been shown to be required for the miRNA pathway it is also required for other forms of translational repression, for example in S. cerevisiae that does not have miRNAs. Similarly, although the reporter used in this study is miRNA regulated, the same UTR also has binding sites for translational regulators that may operate independently of miRNAs. Thus, important and linked goals of future studies are to understand mechanisms by which the CaMKII UTR is regulated in dendrites and how Me31B engages with these mechanisms of neuronal translational control (Hillebrand, 2010).
The Drosophila olfactory system exhibits very precise and stereotyped wiring that is specified predominantly by genetic programming. Dendrites of olfactory projection neurons (PNs) pattern the developing antennal lobe before olfactory receptor neuron axon arrival, indicating an intrinsic wiring mechanism for PN dendrites. These wiring decisions are likely determined through a transcriptional program. This study found that loss of Brahma associated protein 55 kD (Bap55) results in a highly specific PN mistargeting phenotype. In Bap55 mutants, PNs that normally target to the DL1 glomerulus mistarget to the DA4l glomerulus with 100% penetrance. Loss of Bap55 also causes derepression of a GAL4 whose expression is normally restricted to a small subset of PNs. Bap55 is a member of both the Brahma (BRM) and the Tat interactive protein 60 kD (Tip60) ATP-dependent chromatin remodeling complexes. The Bap55 mutant phenotype is partially recapitulated by Domino and Enhancer of Polycomb mutants, members of the TIP60 complex. However, distinct phenotypes are seen in Brahma and Snf5-related 1 mutants, members of the BRM complex. The Bap55 mutant phenotype can be rescued by postmitotic expression of Bap55, or its human homologs BAF53a and BAF53b. These results suggest that Bap55 functions through the TIP60 chromatin remodeling complex to regulate dendrite wiring specificity in PNs. The specificity of the mutant phenotypes suggests a position for the TIP60 complex at the top of a regulatory hierarchy that orchestrates dendrite targeting decisions (Tea, 2011).
The stereotyped organization of the Drosophila olfactory system makes it an attractive model to study wiring specificity. The first olfactory processing center is the antennal lobe, a bilaterally symmetric structure at the anterior of the Drosophila brain. It is composed of approximately 50 glomeruli in a three-dimensional organization. Each olfactory projection neuron (PN) targets its dendrites to one of those glomeruli to make synaptic connections with a specific class of olfactory receptor neurons. Each PN sends its axon stereotypically to higher brain centers (Tea, 2011).
During development, the dendrites of PNs pattern the antennal lobe prior to axons of olfactory receptor neurons. The specificity of PN dendrite targeting is largely genetically pre-determined by the cell-autonomous action of transcription factors, several of which have been previously described. Furthermore, chromatin remodeling factors have been shown to play an important role in PN wiring (Tea, 2010), although very little is currently known about their specific functions. This study reports a genetic screen for additional factors that regulate PN dendrite wiring specificity; Brahma associated protein 55 kD (Bap55) was identified as a regulator of PN dendrite wiring specificity as part of the TIP60 chromatin remodeling complex (Tea, 2011).
Bap55 is an actin-related protein, the majority of which physically associates with the Brahma (BRM) chromatin remodeling complex in Drosophila embryo extracts. There are two distinct BRM complexes: BAP (Brahma associated proteins; homologous to yeast SWI/SNF) and PBAP (Polybromo-associated BAP; homologous to yeast RSC), both of which contain Brahma, Bap55, and Snf5-Related 1 (Snr1). The human homologs of the BAP and PBAP complexes are called the BAF (Brg1 associated factors) and PBAF (Polybromo-associated BAF) complexes, respectively. The BRM/BAF complexes are members of the SWI/SNF family of ATP-dependent chromatin-remodeling complexes, and have been shown to both activate and repress gene transcription, in some cases, of the same gene (Tea, 2011).
In experiments purifying proteins in complex with tagged Drosophila Pontin in S2 cells, Bap55 was also co-purified as a part of the TIP60 complex, as determined by mass spectrometry. The TIP60 histone acetyltransferase complex has been shown to be involved in many processes, including both transcriptional activation and repression. The complex contains many components, including Bap55, Domino (Dom), and Enhancer of Polycomb (E(Pc)). Dom, homologous to human p400, is the catalytic DNA-dependent ATPase; its ATPase domain is highly similar to Drosophila Brahma and human BRG1 ATPase domains. E(Pc) is homologous to human EPC1 and EPC2 and is an unusual member of the Polycomb group; it does not exhibit homeotic transformations on its own, but rather enhances mutations in other Polycomb group genes (Tea, 2011).
This study provides evidence that Bap55 functions as a part of the TIP60 complex rather than the BRM complex in postmitotic PNs to control their dendrite wiring specificity (Tea, 2011).
To further understanding of dendrite wiring specificity in Drosophila olfactory PNs, a MARCM-based forward genetic screen was performed using piggyBac insertional mutants. MARCM allows visualization and genetic manipulation of single cell or neuroblast clones in an otherwise heterozygous background, permitting the study of essential genes in mosaic animals. In this screen, GH146-GAL4 was used to label a single PN born soon after larval hatching, which in wild-type (WT) animals always projects its dendrites to the dorsolateral glomerulus DL1 in the posterior of the antennal lobe. The DL1 PN also exhibits a stereotyped axon projection, forming an L-shaped pattern in the lateral horn, with additional branches in the mushroom body calyx. A mutant, called LL05955, was identified in which DL1 PNs mistargeted to the dorsolateral glomerulus DA4l in the anterior of the antennal lobe. This phenotype is strikingly specific, with 100% penetrance. Arborization of mutant axons, however, was not obviously altered. The piggyBac insertion site was identified using inverse PCR and Splinkerette PCR. LL05955 is inserted into the coding sequence of Bap55, encoding a homolog of human BAF53a and BAF53b. Precise excision of the piggyBac insertion reverted the dendrite mistargeting phenotype, confirming that disruption of the Bap55 gene causes the dendrite mistargeting (Tea, 2011).
In addition to causing DL1 mistargeting, Bap55 mutants also display neuroblast clone phenotypes. In WT, GH146-GAL4 can label three distinct types of PN neuroblast clones generated in newly hatched larvae. Two of these clones, the anterodorsal neuroblast clone and the lateral neuroblast clone, possess cell bodies that lie dorsal or lateral to the antennal lobe, respectively. PNs from these two lineages project their dendrites to stereotyped and nonoverlapping subsets of glomeruli in the antennal lobe. The third type of clone, the ventral neuroblast clone, has cell bodies that lie ventral to the antennal lobe and dendrites that target throughout the antennal lobe due to the inclusion of at least one PN that elaborates its dendrites to all glomeruli (Tea, 2011).
In Bap55-/- PNs, anterodorsal neuroblast clones display a mild reduction in cell number, and their dendrites are abnormally clustered in the anterior dorsal region of the antennal lobe, including the DA4l glomerulus. Lateral neuroblast clones display a severe reduction in cell number, and the remaining dendrites are unable to target to many glomeruli throughout the antennal lobe. Ventral neuroblast clones display a mild reduction in cell number and a reduced dendrite mass throughout the antennal lobe. During development, the lateral neuroblast first gives rise to local interneurons before switching to produce PNs; in mutants affecting cell proliferation, this property of the lateral neuroblast displays as a severe reduction in GH146-labeled PNs. The severely reduced cell number in Bap55 mutants suggests that Bap55 is essential for neuroblast proliferation or neuronal survival. In the anterodorsal and ventral neuroblasts, PN numbers are not drastically changed; thus, the phenotypes indicate that Bap55 is important for dendrite targeting in multiple classes of PNs (Tea, 2011).
In WT, Mz19-GAL4 labels a subset of the GH146-GAL4 labeling pattern. It labels a small number of PNs derived from two neuroblasts, which can be clearly identified in WT clones generated in newly hatched larvae. Anterodorsal neuroblast clones target their dendrites to the VA1d glomerulus, as well as the DC3 glomerulus residing immediately posterior to VA1d (not easily visible in confocal stacks). Lateral neuroblast clones target all dendrites to the DA1 glomerulus. Unlike GH146-GAL4, WT Mz19-GAL4 never labels ventral neuroblast clones because it is not normally expressed in those cells (Tea, 2011).
In Bap55 mutant PN clones, however, Mz19-GAL4 labels additional PNs in anterodorsal, lateral, and ventral clones compared to their WT counterparts. This phenotype suggests that some Mz19-negative PNs now express Mz19-GAL4. In anterodorsal clones, Mz19-GAL4 labels additional cells, although not as many as are labeled by GH146-GAL4. The PNs also mistarget their dendrites to the anterior antennal lobe, similar to mutant GH146-GAL4 anterodorsal neuroblast clones. WT lateral neuroblast clones normally contain GH146-positive PNs and GH146-negative local interneurons. In Bap55-/- lateral neuroblast clones, Mz19-GAL4 predominantly labels local interneurons that send their processes to many glomeruli throughout the antennal lobe and do not send axon projections to higher brain centers. Lateral clones also show ectopic PN labeling with a lower frequency. The Bap55 mutant appears to cause derepression of Mz19-GAL4, resulting in labeled local interneurons. Ventral neuroblast clones are never labeled in WT Mz19-GAL4, yet are labeled in Bap55 mutants. This further indicates a derepression of the Mz19-GAL4 labeling pattern (Tea, 2011).
Unlike GH146-GAL4, WT Mz19-GAL4 never labels single cell clones when clone induction is performed in newly hatched larvae. This is because Mz19-GAL4 is not expressed in the DL1 PN, the only GH146-positive cell generated during this heat shock time of clone generation. However, in Bap55 mutant PN clones, Mz19-GAL4 ectopically labels single cell anterodorsal PN clones targeting to the DA4l glomerulus, which show an L-shaped pattern in the lateral horn with branches in the mushroom body calyx, similar to GH146-GAL4 labeling. The simplest interpretation is that this compound phenotype reflects first a derepression of Mz19-GAL4 in the DL1 PN, and second a DL1 to DA4l mistargeting phenotype in Bap55 mutants (Tea, 2011).
To test whether Bap55 functions in the neuroblast or postmitotically in PNs, GH146-GAL4, which expresses only in postmitotic PNs, was used to express UAS-Bap55 in a Bap55-/- single cell clone. The dendrite mistargeting phenotype was shown to be rescued to the WT DL1 glomerulus and it is concluded that Bap55 functions postmitotically to regulate PN dendrite targeting. The axon phenotype remains the stereotypical L-shaped pattern (Tea, 2011).
The Drosophila Bap55 protein is 70% similar and 54% identical to human BAF53a and 71% similar and 55% identical to human BAF53b. BAF53a and b are 91% similar and 84% identical to each other. Using GH146-GAL4 to express human BAF53a or b in a Bap55-/- single cell clone, it was found that the human homologs can effectively rescue the Bap55-/- dendrite mistargeting phenotype. Interestingly, both also cause the de novo DM6 dendrite and ventral axon mistargeting phenotypes in 6 out of 19 cases for BAF53a and 2 out of 32 cases for BAF53b. Thus, human BAF53a and b can largely replace the function of Drosophila Bap55 in PNs (Tea, 2011).
To address whether Bap55 functions as a part of the BRM complex in PN dendrite targeting, two additional BRM complex mutants were tested for their PN dendrite phenotypes. First, Brahma (brm), the catalytic ATPase subunit of the BRM complex, which is required for the activation of many homeotic genes in Drosophila, was tested. Null mutations have been shown to decrease cell viability and cause peripheral nervous system defects. RNA interference knockdown of brm in embryonic class I da neurons revealed dendrite misrouting phenotypes, although not identical to the Bap55 phenotype. The human homologs of brm, BRM and BRG1 (Brahma-related gene-1), both have DNA-dependent ATPase activity. Inactivation of BRM in mice does not yield obvious neural phenotypes, but inactivation of BRG1 in neural progenitors results in reduced proliferation. BRG1 is likely to be required for various aspects of neural development, including proper neural tube development (Tea, 2011).
In PNs, brm mutants displayed anterodorsal single cell clone mistargeting to non-stereotyped glomeruli throughout the antennal lobe, with each clone differing from the next. This is in contrast to the highly stereotyped DA4l mistargeting of Bap55 mutants. brm-/- neuroblast clones also displayed phenotypes where dendrites make small, meandering projections throughout the antennal lobe, which does not resemble the Bap55-/- phenotype. They additionally exhibit a perturbed cell morphology phenotype, which is markedly more severe than the Bap55 mutant phenotype (Tea, 2011).
Next, Snr1, a highly conserved subunit of the BRM complex, was tested. It is required to restrict BRM complex activity during the development of wing vein and intervein cells and functions as a growth regulator. Its human homolog, SNF5, is strongly correlated with many cancers, yet little is known about its specific role in neurons (Tea, 2011).
In PNs, Snr1 mutants displayed similar phenotypes to brm mutants. The single cell clones displayed mistargeting to non-stereotyped glomeruli throughout the antennal lobe, with each clone demonstrating a unique phenotype. The neuroblast clones exhibited small meandering dendrites throughout the antennal lobe, which also showed extremely perturbed cell morphology. These results, especially the non-sterotyped single cell clone phenotypes, indicate that the BRM complex functions differently from Bap55 in controlling PN dendrite targeting (Tea, 2011).
brm and Snr1 mutants were further analyzed with Mz19-GAL4 to determine whether their phenotypes resembled the Bap55 mutant phenotype of derepression. It was not possible to generate any labeled clones, indicating one of three possibilities: increased cell death, severe cell proliferation defects, or repression of the Mz19-GAL4 labeling pattern. In any of the three cases, the phenotype does not resemble the Bap55-/- mutant phenotype of abnormal activation of Mz19-GAL4 in single cell or neuroblast clones,
indicating that the BRM complex functions differently from Bap55 in PNs (Tea, 2011).
In the same screen in which the Bap55 mutation was identified, LL05537, a mutation in dom that resulted in a qualitatively similar phenotype to Bap55 mutants was identified. dom-/- DL1 PNs split their dendrites between the posterior glomerulus DL1 and the anterior glomerulus DA4l. Their axons exhibit a WT L-shaped pattern in the lateral horn (Tea, 2011).
The LL05537 allele contains a piggyBac insertion in an intron just upstream of the translation start of dom. Because the piggyBac insertion contains splice acceptor sites and stop codons in all three coding frames, this allele likely disrupts all isoforms of dom. Similarly to Bap55, the piggyBac insertion site was identified using inverse PCR and Splinkerette PCR. Precise excision of the piggyBac insertion reverted the dendrite targeting phenotype, confirming that disruption of the dom gene causes the dendrite mistargeting. In addition, a BAC transgene that contains the entire dom transcriptional unit rescued the dom-/- mutant phenotypes (Tea, 2011).
Dom is the catalytic DNA-dependent ATPase of the TIP60 complex and has been shown to contribute to a repressive chromatin structure and silencing of homeotic genes. Dom is a member of the SWI/SNF family and its ATPase domain is highly similar to the Drosophila Brahma and human BRG1 ATPase domains. The human homolog of Dom is p400, which is important for regulating nucleosome stability during repair of double-stranded DNA breaks and an important regulator of embryonic stem cell identity (Tea, 2011).
To determine whether Bap55 and Dom genetically interact, UAS-Bap55 was expressed in a dom-/- PN. This manipulation did not significantly alter the dendrite phenotype. The axon branching pattern also was not altered (Tea, 2011).
Another component of the TIP60 complex, E(Pc), was also examined. In Drosophila, E(Pc) is a suppressor of position-effect variegation and heterozygous mutations in E(Pc) result in an increase in homologous recombination over nonhomologous end joining at double-stranded DNA breaks. Following ionizing radiation, heterozygous animals also exhibit higher genome stability and lower incidence of apoptosis. Yet little is known about its role in neurons (Tea, 2011).
In this study, it was found that E(Pc)-/- DL1 PN dendrites also mistarget to the anterior glomerulus DA4l and exhibit the stereotyped L-shaped axon pattern in the lateral horn. A BAC transgene that contains the entire E(Pc) transcription unit rescued the E(Pc) mutant phenotypes. To determine whether Bap55 and E(Pc) genetically interact, UAS-Bap55 was expressed in an E(Pc)-/- DL1 PN. This manipulation caused the dendrites to split between the DA4l and DM6 glomeruli, and resulted in axons targeting ventrally to the lateral horn (Tea, 2011).
Neuroblast clones mutant for dom also exhibit dendrite mistargeting phenotypes to inappropriate glomeruli throughout the antennal lobe. Anterodorsal and lateral neuroblast clones show a very mild reduction in cell number and their dendrites do not target to the full set of proper glomeruli. Ventral neuroblast clones, when compared to WT, exhibit incomplete targeting throughout the antennal lobe (Tea, 2011).
Further analysis of dom mutants by labeling with Mz19-GAL4 revealed the same derepression as in Bap55 mutants. dom mutant Mz19-GAL4 PN clones also label anterodorsal, lateral, and ventral neuroblast clones with phenotypes similar to GH146-GAL4 labeled neuroblast clones. In anterodorsal and lateral neuroblast clones, Mz19-GAL4 labels a large number of PNs that target to many glomeruli throughout the antennal lobe, although the cell number is smaller than GH146-GAL4 labeling. Ventral neuroblast clones are never labeled in WT Mz19-GAL4, yet are labeled in dom mutants. Mz19-GAL4 also labels single cell clones that split their dendrites between the DA4l and DL1 glomeruli and form the stereotypical L-shaped axon pattern in the lateral horn. As in Bap55 mutants, this compound phenotype likely results from ectopic labeling of a DL1 PN, which further mistargets to DA4l (Tea, 2011).
The E(Pc) phenotypes in GH146 and Mz19-GAL4 labeled neuroblast clones, as well as Mz19-GAL4 labeled single cell clones, displayed similar phenotypes to dom as described above. The phenotypic similarities in single cell clone dendrite mistargeting and derepression of a PN-GAL4 in mutations that disrupt Bap55, dom and E(Pc) strongly suggest that these three proteins act together in the TIP60 complex to regulate PN development (Tea, 2011).
This study has demonstrated a similar role for three members of the TIP60 complex in olfactory PN wiring. The TIP60 complex plays a very specific role in controlling dendrite wiring specificity, with a precise mistargeting of the dendrite mass in Bap55, dom, and E(Pc) mutants. This specific DL1 to DA4l mistargeting phenotype has only been seen in these three mutants, out of approximately 4,000 other insertional and EMS mutants screened, supporting the conclusion that the TIP60 complex has a specific function in controlling PN dendrite targeting. TIP60 complex mutants show discrete glomerular mistargeting, rather than randomly distributed dendrite spillover to different glomeruli. In contrast, perturbation of individual cell surface receptors often leads to variable mistargeted dendrites that do not necessarily obey glomerular borders, possibly reflecting the combinatorial use of many cell surface effector molecules. Even transcription factor mutants yield variable phenotypes. Interestingly, BRM complex mutants yield non-stereotyped phenotypes in PNs. No stereotyped glomerular targeting was seen for brm or Snr1 mutant dendrites; each PN spreads its dendrites across different glomeruli. These data suggest that different chromatin remodeling complexes play distinct roles in regulating neuronal differentiation. The uni- or bi-glomerular targeting to specific glomeruli implies that the TIP60 complex sits at the top of a regulatory hierarchy to orchestrate an entire transcriptional program of regulation (Tea, 2011).
This study suggests a function for Bap55 in Drosophila olfactory PN development as a part of the TIP60 complex rather than the BRM complex. Another possibility could be that Bap55 also serves as the interface between the BRM and TIP60 complexes. While loss of core BRM complex components results in a more general defect, loss of Bap55 could specifically disrupt interactions with the TIP60 complex but maintain other BRM complex functions, causing a more specific targeting phenotype mimicking loss of TIP60 complex components (Tea, 2011).
Interestingly, both human BAF53a and b can significantly rescue the Bap55-/- phenotype. Though in mammals BAF53a is expressed in neural progenitors and BAF53b is expressed in postmitotic neurons, they can perform the same postmitotic function in Drosophila PNs. Further, both can function with the TIP60 complex in PNs to regulate wiring specificity. These data suggest that the functions for BAF53a and b (in neural precursors and postmitotic neurons, respectively) diverge after the evolutionary split between vertebrates and insects (Tea, 2011).
The discrete glomerular states of the mistargeting phenotypes may suggest a role for the TIP60 complex upstream of a regulatory hierarchy determining PN targeting decisions. It is possible that disrupting various components changes the composition of the complex. Additionally, overexpression of Bap55 in various mutant backgrounds might alter the sensitive stoichiometry of the TIP60 complex, resulting in targeting to different but still distinct glomeruli (Tea, 2011).
Several mutants have been identified that cause DL1 PNs to mistarget to areas near the DM6 glomerulus (Tea, 2010). Interestingly, WT DM6 PNs have the most similar lateral horn axon arborization pattern to DL1 PNs. It is hypothesized that the transcriptional code for DM6 is similar to that of DL1, which is at least partially regulated by the TIP60 complex. The genes described in this manuscript are the only mutants that have yielded specific DA4l mistargeting to date. It is possible that the targeting 'code' for DA4l, DL1, and DM6 may be most similar, such that perturbation of the TIP60 complex might result in reprogramming of dendrite targeting. PNs have previously been shown to be pre-specified by birth order. Yet DA4l is born in early embryogenesis, DL1 is born in early larva, and DM6 is born in late larva. This implies that the TIP60 transcriptional code does not correlate with PN birth order. The mechanisms by which the TIP60 complex specifies PN dendrite targeting remain to be determined (Tea, 2011).
This study has characterize PN phenotypes of mutants in the BRM and TIP60 complexes, with a focus on Bap55, which is shared by the two complexes. The TIP60 complex was found to play a very specific role in regulating PN dendrite targeting; mutants mistarget from the DL1 to the DA4l glomerulus. This specific mistargeting phenotype suggests that TIP60 controls a transcriptional program important for making dendrite targeting decisions (Tea, 2011).
As the primary sites of synaptic or sensory input in the nervous system, dendrites play an essential role in processing neuronal and sensory information. Moreover, the specification of class specific dendrite arborization is critically important in establishing neural connectivity and the formation of functional networks. Cytoskeletal modulation provides a key mechanism for establishing, as well as reorganizing, dendritic morphology among distinct neuronal subtypes. While previous studies have established differential roles for the small GTPases Rac and Rho in mediating dendrite morphogenesis, little is known regarding the direct regulators of these genes in mediating distinct dendritic architectures. This study demonstrates that the RhoGEF Trio is required for the specification of class specific dendritic morphology in dendritic arborization (da) sensory neurons of the Drosophila peripheral nervous system (PNS). Trio is expressed in all da neuron subclasses and loss-of-function analyses indicate that Trio functions cell-autonomously in promoting dendritic branching, field coverage, and refining dendritic outgrowth in various da neuron subtypes. Moreover, overexpression studies demonstrate that Trio acts to promote higher order dendritic branching, including the formation of dendritic filopodia, through Trio GEF1-dependent interactions with Rac1, whereas Trio GEF-2-dependent interactions with Rho1 serve to restrict dendritic extension and higher order branching in da neurons. Finally, it was shown that de novo dendritic branching, induced by the homeodomain transcription factor Cut, requires Trio activity suggesting these molecules may act in a pathway to mediate dendrite morphogenesis. Collectively, these analyses implicate Trio as an important regulator of class specific da neuron dendrite morphogenesis via interactions with Rac1 and Rho1 and indicate that Trio is required as downstream effector in Cut-mediated regulation of dendrite branching and filopodia formation (Iyer, 2012).
This analysis demonstrates that Trio functions in promoting and refining class specific dendritic arborization patterns via GEF1- and GEF2-dependent interactions with Rac1 and Rho1, respectively. It was also demonstrated that Trio is required in mediating Cut induced effects on dendritic branching and filopodia formation suggesting that these molecules may operate in a common pathway to direct dendritic morphogenesis. Giniger and colleagues (NINDS/NIH) have likewise been investigating Trio function in da neurons via a non-overlapping, complementary experimental approach, and that they arrived at conclusions regarding Trio function largely consistent with those reported in this study (Iyer, 2012).
Previous studies have demonstrated that Trio functions via its GEF1 domain in mediating the regulation of axon morphogenesis by modulating Rac1 activity, however much less is known regarding the potential in vivo functional role(s) of the Trio GEF2 domain. Intriguingly, a previous study demonstrated that trio mutant neuroblast clones display a neurite overextension phenotype from the dendritic calyx region of the mushroom body which strongly resembled the dendrite-specific overextension phenotype observed in RhoA mutant mushroom body clones suggesting that RhoA/Rho1 activation may be required for restricting dendritic extension. In Drosophila da neurons, trio loss-of-function analyses reveal a reduction in dendritic branching in three distinct da neuron subclasses (class I, III, and IV), indicating a functional role for Trio in promoting dendritic branching. However, class specific differences are observed with Trio gain-of-function studies in which Trio overexpression in class I neurons increases dendritic branching, whereas in class III neurons there is no change in overall dendritic branching, but rather a redistribution of branches, and in class IV there is a reduction in overall dendritic branching. The basis for these differences appear to lie in the observation that refinement of dendritic branching in da neurons is subject to the opposing roles of Rac1 and Rho1 activation via Trio-GEF1 and Trio-GEF2, respectively, where Trio-GEF1 activity promotes higher order dendritic branching, whereas Trio-GEF2 activity restricts higher order branching and also limits overall dendritic length/extension (Iyer, 2012).
One of the key distinctions between class I versus class III and IV neurons relates to inherent differences in normal dendritic branching complexity and the relative roles of dynamic actin cytoskeletal based processes in these neurons which are known to mediate higher order branching including the dendritic filopodia of class III neurons and fine terminal branching in class IV neurons, whereas the class I neurons do not normally exhibit this degree of higher order branching and are predominantly populated by stable, microtubule-based primary and secondary branches. As such, Trio overexpression in these distinct subclasses may yield different effects on overall dendritic branching morphology based upon the normal distribution of actin cytoskeleton within these subclasses leading to unique effects on class specific dendritic architecture. Both loss-of-function and gain-of-function results support this hypothesis as the predominant effects are restricted to actin-rich higher order branching, whereas the primary branches populated by microtubles are relatively unaffected. This is further supported by the demonstration that trio knockdown suppresses Cut induced formation of actin-rich dendritic filopodia. Moreover, phenotypic analyses revealed that co-expression of Cut and Trio-GEF1 synergistically enhance dendritic branching in class I neurons likely due to increased activation of Rac1, whereas co-expression of Cut and Trio-GEF2 lead primarily to increased dendritic extension likely due to increased activation of Rho1. Thus, Trio mediated regulation of Rac1 and/or Rho1 signaling has the potential for sculpting dendritic branching and outgrowth/extension depending upon the combinatorial and opposing effects of Rac1 and Rho1 (Iyer, 2012).
In contrast to Cut, which has been shown to be differentially expressed in da neuron subclasses and exert distinct effects on class specific dendritic arborization, this study has demonstrated that Trio is expressed in all da neuron subclasses and can exert distinct effects on class specific dendritic branching. For example, in all subclasses examined, loss-of-function analyses indicate Trio is required to promote dendritic branching and yet individual subclasses exhibit strikingly distinct dendritic morphologies. These results suggest that Trio is generally required in each of these subclasses to regulate branching, however alone is insufficient to drive these class specific morphologies solely via activation of Rac1 and/or Rho1 signaling. One logical hypothesis is that differential expression of RhoGAP family members in distinct da neuron subclasses may work in concert with Trio to refine class specific morphologies. The potential for combinatorial activity between Trio and various RhoGAPs is significant given that 20 RhoGAPs have been defined in the Drosophila genome. For example, given that class I da neurons exhibit a simple branching morphology which becomes more complex when Trio or Trio-GEF1 domains are overexpressed, perhaps there is higher expression of Rac-inactivating GAPs in class I neurons that function in limiting dendritic branching, whereas in the more complex class III or IV da neurons, there may be lower expression of RacGAPs. Since overexpression of Trio-GEF2 reduces dendritic branching complexity in all three da neuron subclasses analyzed, it might be predicted that Rho1 activation limits dendritic branching and that therefore the expression of RhoGAPs may be modulated to facilitate branching in class III and IV neurons relative to class I neurons. In concert, differential expression of RacGAPs and RhoGAPs together with the uniform expression of Trio in all da neuron subclasses could potentially account for differential levels of activation/inactivation of Rac1 and/or Rho1 in individual subclasses and thereby influence overall class specific dendritic architecture (Iyer, 2012).
In support of this hypothesis, class-specific microarray analyses conducted in class I, III, and IV da neurons indeed reveal differential gene expression levels for most of the 20 known RhoGAP family members at a class-specific level. These expression analyses reveal one trend whereby select RhoGAP encoding genes are upregulated in the more complex class III and IV da neurons relative to the simple class I da neurons, whereas select RacGAP encoding genes are downregulated in complex neurons relative to simple neurons. Moreover, it is known that individual RhoGAPs display differential specificities for Rac, Rho and Cdc42 in vivo, such that a given RhoGAP may function in activating one or more of these small G proteins thereby increasing the potential for fine-tuning activation levels of a particular G protein at a class specific level. Furthermore recent studies provide direct evidence of the importance of RhoGAP family members in regulating da neuron dendritic morphogenesis. Analyses of the tumbleweed (tum) gene, which encodes the GTPase activating protein RacGAP50C, demonstrate that tum mutants display excessive da neuron dendritic branching. The dendritic phenotype observed in tum mutant da neurons is strikingly similar to that observed with Trio-GEF1 overexpression which also leads to excessive dendritic branching. Together these data suggest that Trio-GEF1 functions in activating Rac1 to promote dendritic branching whereas Tum/RacGAP50C function in inactivating Rac1 via its GTPase activity and thereby limit dendritic branching. In contrast, mutant analyses of the RhoGAP encoding gene, crossveinless-c, whose target in da neurons is the Rho1 small G protein, reveal defects in directional growth of da neuron dendrites. These results indicate that Crossveinless-C is required to inactivate Rho1 in order to promote directional dendritic growth and further suggest that a failure to inactivate Rho1 leads to restricted dendritic growth consistent with the phenotypes observed with Trio-GEF2 overexpression in all da neuron subclasses examined. These results, together with those presented herein, suggest that potential combinatorial activity of Trio and RhoGAP family proteins may converge in shaping the class specific dendritic architecture. Ultimately, future functional studies will be required to validate this hypothesis (Iyer, 2012).
While previous studies have revealed Trio acts in concert with Abl and Ena in coordinately regulating axon guidance, the same regulatory relationship does not appear to operate in da neuron dendrites as Abl has been shown to function in limiting dendritic branching and the formation of dendritic filopoda, whereas both Ena functions in promoting dendritic branching. This study demonstrates that Trio functions in promoting dendritic branching, consistent with Ena activity, but in da neuron dendrites works in an opposite direction to Abl. These findings suggest that, at least in da neuron dendrites, Trio may operate in either an Abl-independent pathway or that Trio and Abl may exhibit a context dependent regulatory interaction that is distinctly different in dendrites versus axons (Iyer, 2012).
Drosophila Transmembrane channel-like (Tmc) is a protein that functions in larval proprioception. The closely related TMC1 protein is required for mammalian hearing and is a pore-forming subunit of the hair cell mechanotransduction channel. In hair cells, TMC1 is gated by small deflections of microvilli that produce tension on extracellular tip-links that connect adjacent villi. How Tmc might be gated in larval proprioceptors, which are neurons having a morphology that is completely distinct from hair cells, is unknown. This study has used high-speed confocal microscopy both to measure displacements of proprioceptive sensory dendrites during larval movement and to optically measure neural activity of the moving proprioceptors. Unexpectedly, the pattern of dendrite deformation for distinct neurons was unique and differed depending on the direction of locomotion: ddaE neuron dendrites were strongly curved by forward locomotion, while the dendrites of ddaD were more strongly deformed by backward locomotion. Furthermore, GCaMP6f calcium signals recorded in the proprioceptive neurons during locomotion indicated tuning to the direction of movement. ddaE showed strong activation during forward locomotion, while ddaD showed responses that were strongest during backward locomotion. Peripheral proprioceptive neurons in animals mutant for Tmc showed a near-complete loss of movement related calcium signals. As the strength of the responses of wild-type animals was correlated with dendrite curvature, it is proposed that Tmc channels may be activated by membrane curvature in dendrites that are exposed to strain. These findings begin to explain how distinct cellular systems rely on a common molecular pathway for mechanosensory responses (He, 2019).
For stimuli in motion, sensory systems must encode the direction of movement. This is perhaps best studied in the visual system, where neurons in the vertebrate and invertebrate retina are activated by moving edges in a visual scene. In the retina, specific neurons are tuned to be activated by stimuli moving in a preferred direction but are inhibited by stimuli with non-preferred motion. More poorly understood is how mechanosensory systems might encode the direction of movement. Nevertheless, direction selectivity has been observed in several mechanosensory systems. In the best-understood example, the hair cells of the inner ear show a preferred mechanosensory response when the actin-rich bundles of stereocilia are displaced toward the microvilli on the taller side of the bundle. Another example is found in the neurons that innervate the mechanosensory bristles of adult Drosophila. These neurons are activated by forces that displace the bristle toward the body, but not by displacements away from the body. Similarly, texture sensing in the adult fly proboscis involves a directional deflection of taste bristles that depends on Drosophila Tmc (Zhang, 2016). Low-threshold mechanoreceptors with lanceolate endings that innervate hair follicles in the mouse respond preferentially to deflection of hairs in the caudal to rostral direction (He, 2019).
This study has also discovered another example of preferred directional mechanosensory responses in identified non-ciliated sensory neurons of the Drosophila larva. The ddaE neuron shows preferential responses to forward locomotion, while the ddaD neuron responds preferentially to backward locomotion. Interestingly, the molecular basis of these mechanosensory responses depends on the Drosophila Tmc gene, which encodes
a putative ion channel gene that is homologous to a pore forming subunit of the mechanotransduction channel of mammalian hearing (TMC1) (He, 2019).
In the hair cell, direction selectivity is an emergent property of the actin-rich bundle of stereovilli. The villi possess extracellular tip-links that transmit tugging forces to the mechanosensory channels localized near the tips of the actin bundles. The tip-link tension that is needed for mechanosensory channel gating is generated when the bundle is deflected toward the tallest side, but not when deflected toward the shortest side. A dimeric TMC1 protein complex comprises an ion channel that may be activated by the tugging forces of the tip-link. It is remarkable that Drosophila proprioceptive neurons, which bear no apparent structural resemblance to the inner ear hair cell, rely on a homologous gene (Tmc) for mechanosensory responses that are direction sensitive. These observations raise interesting questions for future study. How can Tmc family channel members function for mechanosensation in such structurally distinct cells as class I neurons and hair cells? Do class I neurons possess extracellular or intracellular links that are involved in activating the Tmc channels? If not, it may be that membrane curvature or tension alone is an important feature for the activation Tmc channels. The latter idea is consistent with proposed models for activation of mechanosensory transduction channels via the forces imposed on them by the plasma membrane (He, 2019).
An additional question that comes from these studies underlies finding the mechanism that generates the preferred direction responses of the class I neurons. Several potential possibilities are envisioned that are not mutually exclusive. The first possibility is that the direction preference is entirely explained by the magnitude of dendrite curvature that occurs in the different neurons during forward and backward movement. Estimates of dendrite curvature were found to be higher in ddaE relative to ddaD during forward locomotion and higher in ddaD than in ddaE during backward locomotion. Thus, in the current experiments, the degree of curvature was correlated with the strength of the calcium signals that was observed in the different neurons during movement. Although the total curvature, and the peak GCaMP signals, were higher for the cells in the preferred direction, these findings may not provide a complete explanation for the direction-selective responses. For instance, evidence for possible differences in adaptation mechanisms is found in the sustained recordings on the tracking microscope, which revealed a higher baseline calcium level in neurons that were responding to prolonged bouts of movement in the preferred direction (He, 2019).
A second possibility would invoke a circuit mechanism that involves inhibition. The current results have shown that the dendrite deformations observed in ddaE and ddaD occur at distinct phases of the segmental contraction cycle. During forward locomotion, ddaE dendrites deform earlier than those of ddaD, and the dendrites of ddaD deform earlier during reverse locomotion. Thus, the more strongly activated neuron is the first to experience deformation, and it is possible that inhibition of the less strongly activated cell occurs during the delay. This model has similarities to the mechanisms that allow starburst amacrine cells to shape responses of direction-selective ganglion cells of the vertebrate retina (He, 2019).
A third possibility is that dendrite deformations that progress in a distal to proximal direction are more strongly activating than
those that progress in proximo-distal direction. Ionic currents that progress from distal to proximal might summate at a spike initiation zone reflected by calcium signals at the cell soma. In contrast, proximal-to-distal dendrite deformations would show reduced summation since the currents would progress in a direction that is moving away from the cell body. This model predicts passive dendrites in class I neurons that lack strongly voltage-gated currents.
Fourth, as with other mechanosensory systems the cellular transduction machinery of the class I neurons may be constructed with an inherent asymmetry that causes it to be more sensitive to the forces that are generated in the preferred direction of movement. This model is appealing due to the involvement of the Tmc family of ion channels in the mechanically driven responses of both the class I neurons and hair cells of the inner ear. Thus, the cellular ultrastructure of the Tmc-dependent transduction machinery of class I neurons will be a fascinating subject for future study (He, 2019).
Finally, the results indicate that the responses of the class I neurons are consistent with the previously proposed mission-accomplished model, but this study adds into this model the feature of direction selectivity. The highest responses of the neurons coincide with the phase of the segmental contraction cycle in which the muscles of the segment are most fully contracted (i.e., mission accomplished). The timing of this peak class I response may facilitate the progression of the wave of neural activity in the larval ganglion to initiate contraction of the next segment, and the signals may also help to terminate the contraction of the preceding segment and within the contracting segment of the traveling wave. It is noteworthy that neurons of the larval ganglion have been identified that show specific activity during bouts of forward locomotion and backward locomotion, respectively. In addition, the larva has a suite of neurons beyond ddaE and ddaD that are thought to participate in proprioception. These neurons include the chordotonal neurons, the bipolar dendritic neurons, and possibly the dmd1 neuron. The activities of many of these neurons (such as the bipolar dendritic neurons, dmd1, and the class I cells ddaD, ddaE, and vpda) have been recently investigated using SCAPE microscopy of moving larvae (see the accompanying paper by Vaadia (2019), and the results indicate that each cell shows a relatively unique response that is timed to various phases of the forward locomotion contraction cycle (as was also seen with ddaE and ddaD). As the larval connectome mapping proceeds, it will be interesting to determine how sensory input from each of these neurons impacts CNS circuits that are specifically engaged during forward and backward locomotion, respectively (He, 2019).
Neurotransmitter receptors and ion channels shape the biophysical properties of neurons, from the sign of the response mediated by neurotransmitter receptors to the dynamics shaped by voltage-gated ion channels. Therefore, knowing the localizations and types of receptors and channels present in neurons is fundamental to understanding of neural computation. This study developed two approaches to visualize the subcellular localization of specific proteins in Drosophila: the flippase-dependent expression of GFP-tagged receptor subunits in single neurons and 'FlpTag', a versatile new tool for the conditional labelling of endogenous proteins. Using these methods, the subcellular distribution of the receptors GluClα, Rdl, and Dα7 and the ion channels Para and Ih in motion-sensing T4/T5 neurons of the Drosophila visual system was investigated. A strictly segregated subcellular distribution of these proteins a sequential spatial arrangement of glutamate, acetylcholine, and GABA receptors was discovered along the dendrite that matched the previously reported EM-reconstructed synapse distributions (Fendl, 2020).
How neural circuits implement certain computations in order to process sensory information is a central question in systems neuroscience. In the visual system of Drosophila, much progress has been made in this direction: numerous studies examined the response properties of different cell-types in the fly brain and electron microscopy studies revealed the neuronal wiring between them. However, one element crucial to understanding is still missing; these are the neurotransmitter receptors used by cells at the postsynaptic site. This knowledge is essential since neurotransmitters and corresponding receptors define the sign and the time-course of a connection, that is whether a synapse is inhibitory or excitatory and whether the signal transduction is fast or slow. The same neurotransmitter can act on different receptors with widely differing effects for the postsynaptic neuron. Glutamate for instance is mainly excitatory, however, in invertebrates it can also have inhibitory effects when it acts on a glutamate-gated chloride channel, known as GluClα. Recently, it has also been shown that acetylcholine, usually excitatory, might also be inhibitory in Drosophila, if it binds to the muscarinic mAChR-A receptor. Hence, knowledge inferring the type of transmitter receptor at a synapse is essential for understanding of the way neural circuits process information (Fendl, 2020).
Moreover, voltage-gated ion channels shape synaptic transmission and the integration of synaptic inputs by defining the membrane properties of every neural cell type. The voltage-gated calcium channel cacophony, for instance, mediates influx of calcium ions that drives synaptic vesicle fusion at presynaptic sites. Voltage-gated sodium channels like Paralytic (Para) are important for the cell's excitability and the generation of sodium-dependent action potentials. The voltage-gated channel Ih influences the integration and kinetics of excitatory postsynaptic potentials. However, only little is known about how these channels are distributed in neurons and how this shapes the neural response properties (Fendl, 2020).
One of the most extensively studied neural circuits in Drosophila is the motion vision pathway in the optic lobe and the underlying computation for direction-selectivity. The optic lobe comprises four neuropils: lamina, medulla, lobula, and lobula plate. As in the vertebrate retina, the fly optic lobe processes information in parallel ON and OFF pathways. Along the visual processing chain, T4/T5 neurons are the first neurons that respond to visual motion in a direction selective way. T4 dendrites reside in layer 10 of the medulla and compute the direction of moving bright edges (ON-pathway). T5 dendrites arborize in layer 1 of the lobula and compute the direction of moving dark edges (OFF-pathway). The four subtypes of T4/T5 neurons (a, b, c, d), project axon terminals to one of the four layers in the lobula plate, each responding only to movement in one of the four cardinal directions, their preferred direction (Fendl, 2020).
How do T4/T5 neurons become direction-selective? Both T4 and T5 dendrites span around eight columns collecting signals from several presynaptic input neurons, each of which samples information from visual space in a retinotopic manner. The functional response properties of the presynaptic partners of T4/T5 have been described in great detail along with their neurotransmitter phenotypes. T4 dendrites receive glutamatergic, GABAergic and cholinergic input, whereas T5 dendrites receive GABAergic and cholinergic input only. These input synapses are arranged in a specific spatial order along T4/T5 dendrites (Fendl, 2020).
Which receptors receive this repertoire of different neurotransmitters at the level of T4/T5 dendrites? Recently, several RNA-sequencing studies described the gene expression pattern of nearly all cell-types in the optic lobe of the fruit fly including T4/T5 neurons. T4/T5 neurons were found to express numerous receptor subunits of different transmitter classes and voltage-gated ion channels at various expression strengths. However, RNA-sequencing studies do not unambiguously answer the above question for two reasons: mRNA and protein levels are regulated in complex ways via post-transcriptional, translational, and protein degradation mechanisms making it difficult to assign protein levels to RNA levels. Secondly, standard RNA-sequencing techniques cannot provide spatial information about receptor localizations, hence, they are not sufficient to conclude which transmitter receptors receive which input signal. Both shortcomings could in principle be overcome by antibody staining since immunohistochemical techniques detect neurotransmitter receptors at the protein level and preserve spatial information. However, high-quality antibodies are not available for every protein of interest and may have variable affinity due to epitope recognition. Furthermore, labeling ion channels via antibodies and ascribing expression of a given channel to a cell-type in dense neuronal tissue remains challenging. The disadvantages of the above techniques highlight the need for new strategies for labeling neurotransmitter receptors in cell types of interest (Fendl, 2020).
This study employed existing and generated new genetic methods to label and visualize ion channels in Drosophila. For endogenous, cell-type-specific labeling of proteins, a generalizable method called FlpTag was developed that expresses a GFP-tag conditionally. Using these tools, the subcellular distribution was determined of the glutamate receptor subunit GluClα, the acetylcholine receptor subunit Dα7, and the GABA receptor subunit Rdl in motion-sensing T4/T5 neurons. These receptor subunits were differentially localized between dendrites and axon terminals. Along the dendrites of individual T4/T5 cells, the receptor subunits GluClα, Rdl, and Dα7 reveal a distinct distribution profile that can be assigned to specific input neurons forming synapses in this area. Furthermore, it was demonstrated the generalizability of the FlpTag approach by generating lines for the metabotropic GABA receptor subunit Gaba-b-r1 and the voltage-gated ion channels para and Ih. The strategies described in this study can be applied to other cells as well as other proteins to reveal the full inventory and spatial distribution of the various ion channels within individual neurons (Fendl, 2020).
Neurotransmitter receptors are essential neuronal elements that define the sign and temporal dynamics of synaptic connections. For understanding of complex neural circuits, it is indispensable to examine which transmitter receptor types are used by the participating neurons and to which compartment they localize. This study developed FlpTag, a generalizable method for endogenous, cell-type-specific labeling of proteins. Alongside several GFP-tagged UAS-lines, the newly developed FlpTag lines were developed to explore the distribution of receptor subunits GluClα, Rdl, Dα7, Gaba-b-r1 and voltage-gated ion channels Para and Ih in motion-sensing T4/T5 neurons of the visual system of Drosophila. These ion channels were found to be localized to either the dendrite, the axonal fiber or the axon terminal. Even at the level of individual dendrites, GluClα, Rdl and Dα7 were differentially distributed precisely matching the locations where T4 and T5 neurons sample signals from their glutamatergic, cholinergic, or GABAergic input neurons, respectively (Fendl, 2020).
Working with Drosophila as model organism bears some unrivaled advantages when it comes to genetic tools. The MiMIC and FlyFos libraries, for instance, are large-scale approaches of enormous value for the fly community as they provide GFP-tagged protein lines for thousands of Drosophila genes including several neurotransmitter receptors and voltage-gated ion channels. Recently, Kondo expanded these existing libraries with T2A-Gal4 insertions in 75 neurotransmitter receptor genes that can also be exchanged by the fluorescent protein tag Venus (Kondo, 2020). While all these approaches tag genes at their endogenous locus, none of them are conditional, for example they cannot be applied in a cell-type-specific manner. Hence, ascribing the expression of the pan-neuronally tagged proteins to cell-types of interest are challenging in dense neuronal tissue (Fendl, 2020).
To overcome these difficulties, two conditional strategies were used for the investigation of membrane protein localizations in the cell types of interest, T4 and T5 neurons. First, GFP-tagged UAS-lines were developed for GluClα and Rdl, and an existing UAS-Dα7::GFP line was tested. As stated above, aberrant localization of overexpressed proteins can occur, however, this is not always the case. Overexpression of UAS-GluClα::GFP shows a similar receptor localization pattern as both MiMIC and FlpTag endogenous lines, thus, validating the use of UAS-GluClα::GFP for studying receptor distribution. Additionally, previous studies reported that the UAS-Dα7::GFP line showed proper localization of the acetylcholine receptor to endogenous synapses when compared to antibody stainings or endogenous Bruchpilot (Brp) puncta. This study confirmed confirmed this finding and further showed that Dα7::GFP presumably localizes only to cholinergic synapses. Overexpressing Dα7::GFP in a medulla neuron that is devoid of endogenous Dα7 demonstrated that Dα7::GFP localized to apparent cholinergic synapses. Hence, the UAS-Dα7::GFP line can be used to study the distribution of cholinergic synapses, but not the exact composition of cholinergic receptor subunits. A recent study showed that quantitatively the levels of the postsynaptic density protein PSD95 change when overexpressed, but qualitatively the localization is not altered. Altogether, this suggests that tagged overexpression lines can be used for studying protein localizations, but they have to be controlled carefully and drawn conclusions might be different for every line (Fendl, 2020).
Ideally, a tool for protein tagging should be both endogenous and conditional. This can be achieved by introducing an FRT-flanked STOP cassette upstream of the gene of interest which was engineered with an epitope tag or fluorescent protein. Only upon cell-type specific expression of Flp, the tagged protein will be expressed in a cell-type specific manner. This genetic strategy was utilized by two independent studies to label the presynaptic protein Brp, the histamine channel Ort and the vesicular acetylcholine transporter VAChT. Recently, a new approach based on the split-GFP system was utilized for endogenous, conditional labeling of proteins in two independent studies. However, all these aforementioned approaches are not readily generalizable and easily applicable to any gene of interest (Fendl, 2020).
The FlpTag strategy presented in this study overcomes these caveats by allowing for endogenous, conditional tagging of proteins and by offering a generalizable toolbox for targeting many genes of interest. Similar to the conditional knock-out tools FlpStop and FlipFlop, FlpTag utilizes a FLEx switch to conditionally control expression of a reporter gene, in this case GFP. Likewise, FlpTag also easily integrates using the readily available intronic MiMIC insertions. This study attempted to generate FlpTag lines for six genes, GluClα, Rdl, Dα7, Gaba-b-r1, para and Ih. Four out of these six lines yielded conditional GFP-tagged protein lines (GluClα, Gaba-b-r1, para, Ih). The FlpTag cassette was injected in MI02620 for Rdl and MI12545 for Dα7, but no GFP expression was detected across the brain. The MiMiC insertion sites used for Rdl and Dα7 seem to be in a suboptimal location for tagging the protein (Fendl, 2020).
As of now, there are MiMIC insertions in coding introns for more than 2800 genes available, which covers approximately 24% of neuronal genes. Additionally, the attP insertion sites generated in the study by Kondo provide possible landing sites for the FlpTag cassette for 75 neurotransmitter receptor genes (Kondo, 2020). Transmembrane proteins such as neurotransmitter receptors form complex 3D structures making fluorescent tagging especially difficult. Neither the MiMIC insertion sites, nor the target sites of the Kondo study at the C-terminus of several transmitter receptor genes, ensure a working GFP-tagged protein line. For genes of interest lacking a suitable MiMIC insertion site a homology directed repair (HDR) cassette was generated that utilizes CRISPR/Cas9-mediated gene editing to integrate the FlpTag cassette in any desired gene locus. The plasmid consists of the FlpTag cassette flanked by multiple cloning sites for the insertion of homology arms (HA). Through HDR the FlpTag cassette can be knocked-in into any desired locus. Taken together, the FlpTag cassette is a generalizable tool that can be integrated in any available attP-site in genes of interest or inserted by CRISPR-HDR into genes lacking attP landing sites. This allows for the investigation of the endogenous spatial distributions of proteins, as well as the correct temporal dynamics of protein expression (Fendl, 2020).
Further, the FlyFos project demonstrated that most fly lines with an extra copy of GFP-tagged protein-coding genes worked normally and GFP-tagged proteins could be imaged in living fly embryos and pupae. In principle, live-imaging of the GFP-tagged lines that were created could be performed during different developmental stages of the fruit fly. In general, the tools generated in this study can be used as specific postsynaptic markers, visualizing glutamatergic, GABAergic, and cholinergic synapses with standard confocal light microscopy. This extends the existing toolbox of Drosophila postsynaptic markers for studying the localization and development of various types of synapses (Fendl, 2020).
T4/T5 neurons combine spatiotemporal input from their presynaptic partners, leading to selective responses to one of the four cardinal directions. Numerous studies investigated the mechanisms underlying direction-selective responses in T4/T5 neurons, yet the computation is still not fully understood. At an algorithmic level, a three-arm detector model is sufficient to describe how direction-selective responses in T4/T5 neurons arise. This model relies on the comparison of signals originating from three neighboring points in space via a delay-and-compare mechanism. The central arm provides fast excitation to the neuron. While one flanking arm amplifies the central signal for stimuli moving along the preferred direction, the other inhibits the central signal for stimuli moving along the null direction of the neuron. Exploring the neurotransmitter receptors and their distribution on T4/T5 dendrites allows defining the sign as well as the temporal dynamics of some of the input synapses to T4/T5 (Fendl, 2020).
According to the algorithmic model, an excitatory, amplifying input signal on the distal side of T4/T5 dendrites was expected. This study found that T4 cells receive an inhibitory, glutamatergic input from Mi9 via GluClα, which, at first sight, seems to contradict expectation. However, since Mi9 has an OFF-center receptive field, this glutamatergic synapse will invert the polarity from Mi9-OFF to T4-ON. Theoretically, in darkness, Mi9 inhibits T4 via glutamate and GluClα, and this inhibition is released upon an ON-edge moving into its receptive field. The concomitant closure of chloride channels and subsequent increased input resistance in T4 cells results in an amplification of a subsequent excitatory input signal from Mi1 and Tm3. As shown by a recent modeling study, this biophysical mechanism can indeed account for preferred direction enhancement in T4 cells (Borst, 2018). Some studies failed to detect preferred direction enhancement in T4/T5 neurons and they proposed that the enhanced signal in PD seen in GCaMP recordings could be a result from a non-linear calcium-to-voltage transformation. If this was really the case, the role of Mi9 and GluClα must be reconsidered and future functional experiments will shed light onto this topic (Fendl, 2020).
Nevertheless, Strother (2017) showed that the RNAi- knock-down of GluClα in T4/T5 neurons leads to enhanced turning responses on the ball set-up for faster speeds of repeating ON and OFF edges (Strother, 2017). Although this observation cannot answer the question about preferred direction enhancement in T4 cells, it indicates that both T4 and T5 receive inhibitory input and that removal of such create enhanced turning responses at the behavioral level. In line with these observations, the glutamate receptor GluClα was also found in T4/T5 axon terminals. A possible functional role of these inhibitory receptors in the axon terminals could be a cross-inhibition of T4/T5 cells with opposite preferred directions via lobula plate intrinsic neurons (LPis). Glutamatergic LPi neurons are known to receive a cholinergic, excitatory signal from T4/T5 neurons within one layer and to inhibit lobula plate tangential cells, the downstream postsynaptic partners of T4/T5 neurons, via GluClα in the adjacent oppositely tuned layer. This mechanism induces a motion opponent response in lobula plate tangential cells and increases their flow-field selectivity. In addition, LPi neurons could also inhibit T4/T5 neurons presynaptically at their axon terminals via GluClα in order to further sharpen the flow-field selectivity of lobula plate tangential cells. Taken together, exploring the subcellular distribution of GluClα in T4/T5 neurons highlights its differential functional roles in different parts of these cell types (Fendl, 2020).
Secondly, the Dα7 signal in the center of T4/T5 dendrites discovered in this study, corresponds to ionotropic, cholinergic input from Mi1 and Tm3 for T4, and Tm1, Tm2 and Tm4 for T5. These signals correspond to the central, fast, excitatory arm of the motion detector model. As T4 and T5 express a variety of different ACh receptor subunits, the exact subunit composition and underlying biophysics of every cholinergic synapse on T4/T5 dendrites still awaits further investigations (Fendl, 2020).
Third, inhibition via GABA plays an essential role in creating direction-selective responses in both T4 and T5 neurons by providing null direction suppression. Computer simulations showed that direction selectivity decreases in T4/T5 motion detector models without this inhibitory input on the null side of the dendrite. This study shows that T4 and T5 neurons possess the inhibitory GABA receptor subunit Rdl mainly on the proximal base on the null side of their dendrites, providing the synaptic basis for null direction suppression. The metabotropic GABA receptor subunit Gaba-b-r1 was not detected in T4/T5 neurons using the newly generated FlpTag Gaba-b-r1 line. Finally, all of the receptor subunits GluClα, Rdl and Dα7 investigated in this study are ionotropic, fast receptors, which presumably do not add a temporal delay at the synaptic level. In the detector model described above, the two outer arms provide a slow and sustained signal, and such properties are already intrinsic properties of these input neurons. However, it cannot be excluded that slow, metabotropic receptor subunits for acetylcholine or GABA (e.g. Gaba-br2) which are also present in T4/T5 and could induce additional delays at the synaptic level (Fendl, 2020).
Furthermore, the subcellular distribution was investigated of the voltage-gated ion channels Para and Ih in T4/T5 neurons. Para, a voltage-gated sodium channel, was found to be distributed along the axonal fibers of both T4 and T5 neurons. As Para is important for the generation of sodium-dependent action potentials, it will be interesting for future functional studies to investigate, if T4/T5 really fire action potentials and how this shapes their direction-selective response. Further, Ih, a voltage-gated ion channel permeable for several types of ions, was detected in T4/T5 dendrites using the FlpTag strategy. Ih channels are activated at negative potentials below -50 mV and as they are permeable to sodium and potassium ions, they can cause a depolarization of the cell after hyperpolarization. Loss-of-function studies will unravel the functional role of the Ih channel for direction-selective responses in T4/T5 neurons (Fendl, 2020).
Since the ability to combine synaptic inputs from different neurotransmitters at different spatial sites is common to all neurons, the approaches described in this study represent an important future perspective for other circuits. The tools can be used to study the ion channels GluClα, Rdl, Dα7, Gaba-b-r1, para and Ih in any given Drosophila cell-type and circuit. Furthermore, the FlpTag tool box can be used to target many genes of interest and thereby foster molecular questions across fields (Fendl, 2020).
The techniques described in this study can be transferred to other model organisms as well, to study the distribution of different transmitter receptors. For instance, in the mouse retina - similar to motion-sensing T4/T5 neurons in the fruit fly - so-called On-Off direction-selective ganglion cells receive asymmetric inhibitory GABAergic inputs from presynaptic starburst amacrine cells during null-direction motion. A previous study investigated the spatial distribution of GABA receptors of these direction-selective ganglion cells using super-resolution imaging and antibody staining. Additionally, starburst amacrine cells also release ACh onto ganglion cells which contributes to the direction-selective responses of ganglion cells. Thus, mapping the distribution of ACh receptors on direction-selective ganglion cells will be the next important step to further investigate cholinergic transmission in this network (Fendl, 2020).
Overall, this study has demonstrated the importance of exploring the distributions of neurotransmitter receptors and ion channels for systems neuroscience. The distinct distributions in T4/T5 neurons discovered in this study and the resulting functional consequences expand knowledge of the molecular basis of motion vision. Although powerful, recent RNAseq studies lacked information about spatial distributions of transmitter receptors which can change the whole logic of wiring patterns and underlying synaptic signs. Future studies can use this knowledge to target these receptors and directly probe their role in functional experiments or incorporate the gained insights into model simulations. However, this study is only highlighting some examples of important neural circuit components: expanding the approaches described in this study to other transmitter receptors and ion channels, as well as gap junction proteins will reveal the full inventory and the spatial distributions of these decisive determinants of neural function within an individual neuron (Fendl, 2020).
Cell recognition molecules are key regulators of neural circuit assembly. The Dscam family of recognition molecules in Drosophila has been shown to regulate interactions between neurons through homophilic repulsion. This is exemplified by Dscam1 and Dscam2, which together repel dendrites of lamina neurons, L1 and L2, in the visual system. By contrast, this study shows that Dscam2 directs dendritic targeting of another lamina neuron, L4, through homophilic adhesion. Through live imaging and genetic mosaics to dissect interactions between specific cells, Dscam2 was shown to be required in L4 and its target cells for correct dendritic targeting. In a genetic screen, Dscam4 was identified as another regulator of L4 targeting which acts with Dscam2 in the same pathway to regulate this process. This ensures tiling of the lamina neuropil through heterotypic interactions. Thus, different combinations of Dscam proteins act through distinct mechanisms in closely related neurons to pattern neural circuits (Tadros, 2016).
This paper shows that Dscam2 and Dscam4 act together to regulate dendritic targeting. The genetic data presented in this study, coupled with previous biochemical studies, are consistent with Dscam2 using homophilic interactions to adhere L4 dendrites to multiple lamina neurons. In all other contexts examined previously, Dscam2 promotes repulsion. To further understand the adhesive role of Dscam2 in circuit assembly, a genetic screen was carried out to identify other genes with a similar phenotype. This led to the identification of Dscam4 mutants with an L4 targeting phenotype indistinguishable from that of Dscam2. Dscam2 and Dscam4 promote adhesion between L4 dendritic filopodia and their target fascicles, as revealed through live imaging. In this way, these proteins regulate the tiling of L4 dendritic processes across the lamina neuropil. Thus, Dscam family members act in different combinations through either adhesion or repulsion to pattern neural circuits (Tadros, 2016).
Previously, it was reported that Dscam2 regulates tiling of the axon terminals of L1 neurons in the medulla. It was proposed that this arrangement of processes is a consequence of homotypic interactions (i.e., interactions between cells of the same type) between L1 axon terminals in adjacent columns. This model was based on three observations. First, L1 axon terminals initially expand to contact those of their immediate neighbors and then retract. Second, the axon terminals of Dscam2 mutant L1 neurons remain expanded, invading the territory of their neighbors, in genetically mosaic animals. And third, wild-type L1s also exhibit the same defect when confronted with Dscam2 mutant lamina neurons in adjacent columns (Tadros, 2016).
This paper has uncovered a role for Dscam2 in regulating L4 dendritic tiling that occurs in a mechanistically different fashion. Developmental and reverse MARCM experiments suggested that Dscam2 mediates L4 tiling in a manner analogous to its role in L1 axons, that is via Dscam2-mediated homotypic interactions between L4 dendrites innervating the same cartridge. Additional genetic mosaic studies, however, ruled this out. By modifying MARCM (i.e., DL-MARCM), it was possible to assess the targeting of wild-type L4 dendrites encountering dendrites from neighboring L4 mutant neurons. In all cases, these wild-type dendrites targeted normally. Thus, while L4 tiling relies upon Dscam2-mediated homophilic recognition, it does not result solely from homotypic interactions between neighboring L4 dendrites (Tadros, 2016).
Developmental analyses raised the alternative possibility that Dscam2 is used by L4 dendrites to promote interactions with the target fascicle. Indeed, targeting defects were observed in L4 dendrites innervating cartridges with mutant L1 or L2 neurons using DL-MARCM. Thus, in this context, tiling arises from heterotypic interactions. As the penetrance of this cell non-autonomous phenotype was weaker than removing Dscam2 from L4, it is proposed that L1 and L2 act redundantly, perhaps with other lamina neurons (e.g. L3 and L5 which were not tested), to promote Dscam2-dependent recognition between L4 dendrites and the target fascicle. This is consistent with the expression of Dscam2B and Dscam2A in L1 and L2 neurons, respectively, and the redundant role for these isoforms in L4 uncovered through the mosaic analysis of isoform-specific knockin alleles. Thus, L4 tiling arises through heterotypic interactions via Dscam2-mediated homophilic recognition. This is consistent with studies in both vertebrates and invertebrates in which cell ablation experiments indicate that, in some contexts, tiling may arise through heterotypic interactions (Tadros, 2016).
As previously reported, removal of Dscam2 from either L1 or L2 results in ectopic branching of these neurons. This observation raised the possibility that wild-type L4s follow these branches in a Dscam2-independent fashion and that this underlies the Dscam2 non-autonomous effect on wild-type L4s. Although this cannot be out, the interpretation is preferred that these data reflect a Dscam2-dependent effect independent of the ectopic L1 or L2 phenotype for two reasons. First, of the two instances in which a wild-type L4 produced an extra branch upon encountering a Dscam2 mutant L1, one L1 did not form an ectopic branch. And second, in most cases where either L1 (2/3) or L2 (18/23) displayed an extra branch phenotype, the associated L4 did not (Tadros, 2016).
The functional significance of this tiling and its disruption in Dscam2 mutants remain open questions. A recent behavioral study has suggested that L4 functions in responding to progressive, i.e., front-to-back, as opposed to regressive motion. This response would be consistent with L4's asymmetric dendritic arrangement, having one dendrite projecting anteriorly and two posteriorly. While this is an attractive model, another study using slightly different methodology suggests no role for L4 in directional selectivity. Critically assessing the importance of the spatial arrangement of L4 dendrites on behavior will require selective removal of Dscam2 from all L4 neurons in an otherwise wild-type background (Tadros, 2016).
Dscam paralogs promote cell adhesion in vitro. However, in all cases examined previously, genetic and phenotypic analyses of Dscam1 and Dscam2 mutations are consistent with a repulsive function for these proteins upon homophilic binding in vivo. In addition, gain-of-function studies in Drosophila support this view. Targeted expression of single Dscam1 isoforms promotes dendritic repulsion between class I and class III dendritic arborization (da) sensory neurons. This repulsive interaction is converted into adhesion by removing the cytoplasmic domain or selectively removing specific motifs, underscoring the role of cytoplasmic signaling in converting Dscam1-mediated homophilic binding to repulsion. And finally, forced expression of either Dscam2A or Dscam2B isoforms in closely apposed neurons in the visual system promotes repulsion between them (Tadros, 2016).
By contrast, several observations argue that Dscam2 and Dscam4 promote adhesion between L4 dendrites and their target fascicles. In fixed samples, wild-type L4 dendrites target to lamina axon fascicles, while Dscam2, Dscam4, or Dscam2/4 double mutant dendrites extend laterally along the lattice of R cell growth cones surrounding the target fascicles. Indeed, using multicolor flip out (MCFO), extensive intermingling of L4 dendrites and L1 processes was observed in the target fascicle. In live imaging, L4 dendritic processes become progressively anchored to their target fascicles. By contrast, in mutants, the interactions between L4 dendrites and the target fascicles were transient, while interactions with R cell growth cones were sustained. This tight association in mutants could reflect a loss of repulsive interactions between L4 dendrites and R cells. Genetic mosaic analysis, however, indicate that this is highly unlikely. Together, these data support a model in which Dscam2 and Dscam4 act together to promote adhesion between L4 dendritic growth cones and their lamina targets. In the absence of this Dscam2/Dscam4-mediated adhesion, an adhesive interaction between L4 dendrites and the surrounding R cell growth cones is suggested to be unmasked. Indeed, transcript profiling of R cells and L4 neurons during development suggest that they both express several recognition molecules that are candidates for mediating this interaction (Tadros, 2016).
Although a model is favored in which Dscam2 and Dscam4 mediate selective adhesive interactions with the lamina fascicle, the data do not rule out a model in which Dscam2 and Dscam4 regulate L4 targeting in a permissive fashion. For instance, Dscam2/4 may restrict the exploratory phase to fascicles in the immediate vicinity through adhesive interactions with both lamina axons and R cell growth cones. Indeed, in live imaging, mutant L4 dendrites extend to additional fascicles on either side of their normal targets at a very early stage of filopodial exploration. In this model, another cell recognition molecule, acting in an instructive way, would promote association of L4 dendrites with the lamina axon fascicle. Characterization of other genes acting in this process and higher resolution live imaging may allow critical distinguishion between an instructive and a permissive role for Dscam2 and Dscam4 in L4 dendritic targeting (Tadros, 2016).
It is becoming increasingly clear that Drosophila Dscams regulate dendritic patterning in different ways. Dscam1 is alternatively spliced to give rise to greater than 19,000 ectodomain isoforms, with greater than 18,000 exhibiting homophilic binding specificity. This property, coupled with probabilistic expression, endows each neuron with a unique identity and allows for discrimination between self and non-self (i.e., self-avoidance). Dscam2, on the other hand, is alternatively spliced to give rise to only two isoforms. These act in combination with Dscam1 isoforms to regulate the appropriate association of L1 and L2 dendrites at multiple contact synapses. Here, L1 and L2 express different Dscam2 isoforms in a mutually exclusive fashion. It is thought that the unique identity acquired by the dendrites of these cells, through alternative splicing of both Dscam1 and Dscam2, allows them to appropriately discriminate between self and non-self at multiple contact synapses. In these contexts, Dscam proteins promote repulsion. As this study has shown, Dscam2 in combination with Dscam4 promotes adhesive interactions during dendritic targeting. This may reflect a unique property of a Dscam2/Dscam4 protein complex, differences in output of different spliced variants of the cytoplasmic domains, or differences in levels of expression. Precedents for each of these potential mechanisms have been reported in other intercellular signaling proteins. Additional experiments will be needed to distinguish between these different possibilities (Tadros, 2016).
In summary, this study has shown that two different combinations of Dscam proteins regulate the patterning of different dendritic elements in the developing lamina in diverse ways. As all four Dscam proteins are broadly expressed within the developing lamina, distinct paralogs, and isoforms of them, either alone or in combination may pattern dendrites in different ways. Furthermore, as Dscam proteins also interact with other proteins in both cis and trans, they have the potential to pattern circuits in many different ways (Tadros, 2016).
Dendrites possess distinct structural and functional properties that enable neurons to receive information from the environment as well as other neurons. Despite their key role in neuronal function, current understanding of the ability of neurons to regenerate dendrites is lacking. This study characterizes the structural and functional capacity for dendrite regeneration in vivo in adult animals and examines the effect of neuronal maturation on dendrite regeneration. Focus was placed on the class IV dendritic arborization (c4da) neuron of the Drosophila sensory system, which has a dendritic arbor that undergoes dramatic remodeling during the first 3 d of adult life and then maintains a relatively stable morphology thereafter. Using a laser severing paradigm, regeneration was monitored after acute and spatially restricted injury. The capacity for regeneration was found to be present in adult neurons but diminished as the animal aged. Regenerated dendrites recovered receptive function. Furthermore, it was found that the regenerated dendrites show preferential alignment with the extracellular matrix (ECM). Finally, inhibition of ECM degradation by inhibition of matrix metalloproteinase 2 (Mmp2) to preserve the extracellular environment characteristics of young adults led to increased dendrite regeneration. These results demonstrate that dendrites retain regenerative potential throughout adulthood and that regenerative capacity decreases with aging (DeVault, 2018).
Dendrite pathology is frequently observed in various neurodegenerative diseases (NDs). Although previous studies identified several pathogenic mediators of dendrite defects that act through loss of function in NDs, the underlying pathogenic mechanisms remain largely unexplored. A search for additional pathogenic contributors to dendrite defects in NDs identified Relish/NF-kappaB as a novel gain-of-toxicity-based mediator of dendrite defects in animal models for polyglutamine (polyQ) diseases and amyotrophic lateral sclerosis (ALS). In a Drosophila model for polyQ diseases, polyQ-induced dendrite defects require Dredd/Caspase-8-mediated endoproteolytic cleavage of Relish to generate the N-terminal fragment, Rel68, and subsequent Charon-mediated nuclear localization of Rel68. Rel68 alone induced neuronal toxicity causing dendrite and behavioral defects, and two novel transcriptional targets, Tup and Pros, were identified that mediate Rel68-induced neuronal toxicity. Finally, it was shown that Rel68-induced toxicity also contributes to dendrite and behavioral defects in a Drosophila model for ALS. Collectively, these data propose disinhibition of latent toxicity of Relish/NF-κB as a novel pathogenic mechanism underlying dendrite pathology in NDs (Han, 2020).
Microtubule nucleation is essential for proper establishment and maintenance of axons and dendrites. Centrosomes, the primary site of nucleation in most cells, lose their function as microtubule organizing centers during neuronal development. How neurons generate acentrosomal microtubules remains unclear. Drosophila dendritic arborization (da) neurons lack centrosomes and therefore provide a model system to study acentrosomal microtubule nucleation. This study investigated the origin of microtubules within the elaborate dendritic arbor of class IV da neurons. Using a combination of in vivo and in vitro techniques, it was found that Golgi outposts can directly nucleate microtubules throughout the arbor. This acentrosomal nucleation requires gamma-tubulin and CP309, the Drosophila homolog of AKAP450, and contributes to the complex microtubule organization within the arbor and dendrite branch growth and stability. Together, these results identify a direct mechanism for acentrosomal microtubule nucleation within neurons and reveal a function for Golgi outposts in this process (Ori-McKenney, 2012).
Microtubules are organized into dynamic arrays that serve as
tracks for directed vesicular transport and are essential for the
proper establishment and maintenance of neuronal architecture. The organization and nucleation of microtubules must be highly regulated in order to generate and maintain such
complex arrays. Nucleating complexes,
in particular, are necessary because spontaneous
nucleation of new tubulin polymers is kinetically limiting both
in vivo and in vitro. Gamma(Γ)-tubulin is
a core component of microtubule organization centers and has
a well-established role in nucleating spindle and cytoplasmic
microtubules. Previous studies have proposed
that in mammalian neurons, microtubules are nucleated by
γ-tubulin at the centrosome, released by microtubule severing
proteins, and then transported into developing neurites by motor
protein. Indeed, injection of antibodies against
γ-tubulin or severing proteins inhibited axon outgrowth in
neurons cultured for one day in vitro (DIV1) (Ori-McKenney, 2012).
However, proper neuron development and maintenance may
not rely entirely on centrosomal sites of microtubule nucleation.
Although the centrosome is the primary site of microtubule
nucleation at DIV2, it loses its function as a microtubule-organizing
center during neuronal development.
In mature cultured mammalian neurons (DIV 11-12), γ-tubulin
is depleted from the centrosome, and the majority of microtubules
emanate from acentrosomal sites. In
Drosophila dsas-4 mutants that lack centrioles, organization of eye-disc neurons and axon outgrowth are normal in third-instar
larvae. Within the Drosophila peripheral
nervous system (PNS), although dendritic arborization neurons
contain centrioles, they do not form functional centrosomes,
and laser ablation of the centrioles does not perturb microtubule
growth or orientation (Nguyen, 2011). These results indicate
that acentrosomal generation of microtubules contributes to
axon development and neuronal polarity. How and where acentrosomal
microtubule nucleation may contribute to the formation
and maintenance of the more complex dendrites, and what
factors are involved in this nucleation is unknown. Dendritic
arborization (da) neurons provide an excellent system for investigating
these questions. They are a subtype of multipolar
neurons in the PNS of Drosophila melanogaster which produce
complex dendritic arrays and do not contain centrosomes. Based on their
patterns of dendrite projections, the da neurons have been
grouped into four classes (I-IV) with branch complexity and arbor
size increasing with class number. Class IV da neurons are ideal
for studying acentrosomal microtubule nucleation because they
have the most elaborate and dynamic dendritic arbor, raising
intriguing questions about the modes of nucleation for its growth
and maintenance (Ori-McKenney, 2012).
One potential site of acentrosomal microtubule nucleation
within these neurons is the Golgi complex. A number of studies
have shown that the Golgi complex can nucleate microtubules
in fibroblasts. Although, in these cell
types, the Golgi is tightly coupled to the centrosome, it does
not require the centrosome for nucleation. It does, however,
require γ-tubulin, the centrosomal protein AKAP450, and the
microtubule binding proteins CLASPs. When the Golgi is fragmented upon
treatment with nocodazole, the dispersed Golgi ministacks can
still promote microtubule nucleation, indicating that these individual
ministacks contain the necessary machinery for nucleation (Ori-McKenney, 2012 and references therein).
In both mammalian and Drosophila neurons, the Golgi
complex exists as Golgi stacks located within the soma and
Golgi outposts located within the dendrites. In cultured
mammalian hippocampal neurons, these Golgi outposts are
predominantly localized in a subset of the primary branches; however, in Drosophila class IV da neurons, the Golgi outposts appear throughout the dendritic arbor,
including within the terminal branches (Ye, 2007). The Golgi
outposts may provide membrane for a growing dendrite branch,
as the dynamics of smaller Golgi outposts are highly correlated
with dendrite branching and extension. However, the majority of larger Golgi outposts
remains stationary at dendrite branchpoints and could have
additional roles beyond membrane supply. It is unknown whether Drosophila Golgi outposts
contain nucleation machinery similar to mammalian Golgi
stacks. Such machinery could conceivably support microtubule
nucleation within the complex and dynamic dendritic arbor.
This study identifies a direct mechanism for acentrosomal
microtubule nucleation within the dendritic arbor and reveal
a role for Golgi outposts in this process. Golgi outposts contain
both γ-tubulin and CP309, the Drosophila homolog of AKAP450,
both of which are necessary for Golgi outpost-mediated microtubule
nucleation. This type of acentrosomal nucleation contributes
not only to the generation of microtubules at remote
terminal branches, but also to the complex organization of
microtubules within all branches of the dendritic arbor. Golgi
outposts are therefore important centers of acentrosomal microtubule
nucleation, which is necessary to establish and maintain
the complexity of the class IV da neuronal arbor (Ori-McKenney, 2012).
This study has addressed how microtubules are organized and nucleated
within the complex arbor of class IV da neurons and how
essential these processes are for dendrite growth and stability.
Microtubule organization within different subsets of branches
in da neurons must require many levels of regulation. This
study has identified the first direct mechanism for
acentrosomal microtubule nucleation within these complex
neurons and has uncovered a role for Golgi outposts in this process.
The data are consistent with the observation that pericentriolar
material is redistributed to the dendrites in mammalian neurons
(Ferreira, 1993) and that γ-tubulin is depleted from the
centrosome in mature mammalian neurons (Stiess, 2010).
This suggests that the Golgi outposts may be one structure
involved in the transport of centriole proteins such as γ-tubulin
and CP309. This study found that microtubule nucleation from these Golgi
outposts correlates with the extension and stability of terminal
branches, which is consistent with the observation that
EB3 comet entry into dendritic spines accompanies spine enlargement
in mammalian neurons (Jaworski, 2009). It is
striking that microtubule organization in shorter branches, but
not primary branches, mimics the organization in mammalian
dendrites, with a mixed microtubule polarity in the secondary
branches and a uniform plus end distal polarity in the terminal
branches. Kinesin-2 and certain +TIPS are
necessary for uniform minus end distal microtubule polarity in
the primary dendrites of da neurons. Golgi
outpost mediated microtubule nucleation could also contribute
to establishing or maintaining this polarity both in the terminal
branches and in the primary branches. It will be of interest to
identify other factors that may be involved in organizing microtubules
in different subsets of branches in the future (Ori-McKenney, 2012).
In vivo and in vitro data support a role for Golgi outposts in
nucleating microtubules at specific sites within terminal and
primary branches. However, it is noted that not all EB1 comets
originate from Golgi outposts, indicating other possible mechanisms
of generating microtubules. One potentially important source of microtubules is the
severing of existing microtubules by such enzymes as katanin
and spastin, both of which are necessary for proper neuronal
development. It is likely that both microtubule
nucleation and microtubule severing contribute to the
formation of new microtubules within the dendritic arbor;
however, the current studies suggest that Golgi-mediated nucleation
is especially important for the growth and maintenance of the
terminal arbor. In γ-tubulin and CP309 mutant neurons, the
primary branches contain a similar number of EB1 comets, but
only a small fraction of the terminal branches still contain EB1
comets. This result indicates that severing activity or other sources
of nucleation may suffice for microtubule generation within
the primary branches, but γ-tubulin mediated nucleation is
crucial in the terminal branches. As a result, the terminal branch
arbor is dramatically reduced by mutations compromising the
γ-tubulin nucleation activity at Golgi outposts (Ori-McKenney, 2012).
It is important to note that Golgi outposts are present in the
dendrites, but not in the axons of da neurons; thus, this mode
of nucleation is dendrite specific and likely contributes to the
difference in microtubule arrays in axons and dendrites. While
the axon is one long primary branch with uniform microtubule
polarity, the dendrite arbor is an intricate array of branches
where microtubule polarity depends on branch length.
Therefore, this more elaborate branched structure may have
evolved a variety of nucleation mechanisms, including Golgi
outpost nucleation and microtubule severing. Intriguingly, in
da neurons lacking cytoplasmic dynein function, the Golgi
outposts are mislocalized to the axon, which appears branched
and contains microtubules of mixed polarity (Zheng, 2008). It is speculated that in these mutants, Golgi-mediated microtubule nucleation within the axon is contributing to the mixed microtubule orientation and formation of ectopic dendrite-like branches (Ori-McKenney, 2012).
Only a subpopulation of Golgi outposts could support microtubule
nucleation both in vivo and in vitro. The results show
that Golgi outpost mediated microtubule nucleation is restricted
to stationary outposts and dependent upon γ-tubulin and CP309, but why
some outposts contain these proteins while others do not is unknown. γ-tubulin
and CP309 could be recruited to the Golgi
outposts in the cell body and transported on the structure into the dendrites, or they
could be recruited locally from soluble pools throughout the dendritic arbor.
Golgi outposts are small enough to be trafficked into terminal branches that are 150-300 nm in diameter, and therefore may provide
an excellent vehicle for transporting nucleation machinery to
these remote areas of the arbor. It will be interesting to determine
how these nucleation factors are recruited to the Golgi outposts (Ori-McKenney, 2012).
It has been previously shown that GM130 can recruit AKAP450
to the Golgi complex, but whether the first coiled-coil domain
of the Drosophila AKAP450 homolog, CP309, can also bind
GM130 is unknown. Interestingly, this study has observed that
predominantly stationary Golgi outposts correlated with EB1
comet formation, indicating that this specific subpopulation
may contain γ-tubulin and CP309. What other factors may be
necessary to properly position the Golgi outposts at sites such
as branchpoints, and how this is achieved will be a fascinating
direction for future studies (Ori-McKenney, 2012).
Whether the acentrosomal microtubule nucleation uncovered
in this study also occurs in the dendrites of mammalian neurons
is a question of great interest. Golgi outpost distribution in
cultured hippocampal neurons is significantly different than
that in da neurons, and
hippocampal neurons do not form as elaborate arbors as da
neurons. However, other types of mammalian neurons form
much more complex dendritic arbors and may conceivably require acentrosomal nucleation for the growth and perpetuation of the dendrite branches (Ori-McKenney, 2012).
This study provides the first evidence that Golgi outposts
can nucleate microtubules at acentrosomal sites in neurons,
shedding new light on the longstanding question about the
origin of the microtubule polymer in elongated neuronal processes.
This source of nucleation contributes to the complex
organization of microtubules within all branches of the
neuron, but is specifically necessary for terminal branch
development. It is thus conclude that acentrosomal microtubule
nucleation is essential for dendritic branch growth and
overall arbor maintenance of class IV da neurons, and that Golgi outposts
are important nucleation centers within the dendritic arbor (Ori-McKenney, 2012).
Neuronal dendrite branching is fundamental for building nervous systems. Branch formation is genetically encoded by transcriptional programs to create dendrite arbor morphological diversity for complex neuronal functions. In Drosophila sensory neurons, the transcription factor Abrupt represses branching via an unknown effector pathway. Targeted screening for branching-control effectors identified Centrosomin, the primary centrosome-associated protein for mitotic spindle maturation. Centrosomin repressed dendrite branch formation and was used by Abrupt to simplify arbor branching. Live imaging revealed that Centrosomin localized to the Golgi cis face and that it recruited microtubule nucleation to Golgi outposts for net retrograde microtubule polymerization away from nascent dendrite branches. Removal of Centrosomin enabled the engagement of wee Augmin activity to promote anterograde microtubule growth into the nascent branches, leading to increased branching. The findings reveal that polarized targeting of Centrosomin to Golgi outposts during elaboration of the dendrite arbor creates a local system for guiding microtubule polymerization (Yalgin, 2015).
Neurons primarily receive inputs through their dendrite arbors. The shape and complexity of the dendrite arbor, which is elaborated during differentiation, enables the neuron to properly cover its receptive field and establishes the positions of inputs into the arbor. Disruptions to dendritic branching can precipitate intellectual disability and psychiatric disorders (Yalgin, 2015).
Arbor morphology is regulated for each neuron class to support its structural and functional requirements3; it is genetically encoded, being linked to class specification by transcriptional programs. For example, in Drosophila, the single unbranched dendrite of external sensory neurons is specified over an alternative multipolar dendritic arborization (da) neuron fate by the Prdm transcription factor Hamlet. Similarly, the proneural transcription factor Ngn2 regulates multiple aspects of pyramidal neuron development in the mammalian cortex, including the specification of a characteristic apical dendrite, whereas Cux1, Cux2 and SatB2 link dendrite development to cortical layer-specific developmental program (Yalgin, 2015).
Dendrite development is controlled in a neuron class-specific manner to create differences in arbor morphology and complexity. Class-specific dendrite targeting is regulated via the activity of transmembrane adhesion proteins. For example, in C. elegans, class-specific expression patterns of the transcription factors MEC-3, AHR-1 and ZAG-1 regulate the morphology of mechanosensory neurons, and MEC-3 promotes differential expression of the Claudin-like membrane protein HPO-30 to enable lateral branch stabilization. Drosophila da neurons exist in four classes, of which class I neurons express Abrupt (Ab), which defines their simple arbor shape, and class IV express Knot and Cut, which together promote the complex morphology of this class. The EGF-repeat factor Ten-m is co-regulated by both Knot and Ab to control the direction of branch outgrowth in both class I and IV neurons (Yalgin, 2015).
Contrasting activities of Knot, Cut and Ab in da neurons emphasize that altering dendrite branching is fundamental for regulating arbor complexity. Knot and Cut promote branch formation; conversely, Ab represses branch formation. Little is understood about how modulatory control over branching is achieved (Yalgin, 2015).
Microtubules polymerize via the addition of Tubulin dimers, primarily at the plus end. In axons, microtubules polymerize in an anterograde direction, providing a protrusive force for outgrowth. Microtubule polymerization also drives axon branch formation, as precursors only transform into branches after microtubule invasion. Mature dendrites have a predominantly minus-ends-out microtubule array, nevertheless recent studies have identified that anterograde microtubule polymerization events can initiate or extend branches, or modulate the size of dendritic spines. In addition, the re-initiation of dendrite growth and branch formation following injury uses upregulation of microtubule polymerization and its polarization in the anterograde direction (Yalgin, 2015).
This study examined whether class-specific transcription factors regulate branch promotion and repression by controlling microtubule organization during arbor development. In da sensory neurons, microtubule nucleation and polarity can be assayed in vivo using transgenic markers. Using genetic manipulation of class I and class IV da neurons, this study found that Ab controls class-specific differences in the localization of microtubule minus-end-directed markers in the da neuron arbor. By assaying Ab-mediated changes in the expression of a set of candidate microtubule regulators and using chromatin immunoprecipitation (ChIP), this study identified Cnn (Centrosomin) as an effector of Ab action. Cnn-centered control mechanisms, analogous to those that cluster microtubule nucleation events to create the mitotic spindle, are used in growing dendrites to regulate branching and to create class-specific arbor complexity (Yalgin, 2015).
Centrosomin repressed dendrite branch formation and was used by Abrupt to simplify arbor branching. Live imaging revealed that Centrosomin localized to the Golgi cis face and that it recruited microtubule nucleation to Golgi outposts for net retrograde microtubule polymerization away from nascent dendrite branches. Removal of Centrosomin enabled the engagement of wee Augmin activity to promote anterograde microtubule growth into the nascent branches, leading to increased branching. The findings reveal that polarized targeting of Centrosomin to Golgi outposts during elaboration of the dendrite arbor creates a local system for guiding microtubule polymerization (Yalgin, 2015).
Neuronal dendrite branching is fundamental for building nervous systems. Branch formation is genetically encoded by transcriptional programs to create dendrite arbor morphological diversity for complex neuronal functions. In Drosophila sensory neurons, the transcription factor Abrupt represses branching via an unknown effector pathway. Targeted screening for branching-control effectors identified Centrosomin, the primary centrosome-associated protein for mitotic spindle maturation. Centrosomin repressed dendrite branch formation and was used by Abrupt to simplify arbor branching. Live imaging revealed that Centrosomin localized to the Golgi cis face and that it recruited microtubule nucleation to Golgi outposts for net retrograde microtubule polymerization away from nascent dendrite branches. Removal of Centrosomin enabled the engagement of wee Augmin activity to promote anterograde microtubule growth into the nascent branches, leading to increased branching. The findings reveal that polarized targeting of Centrosomin to Golgi outposts during elaboration of the dendrite arbor creates a local system for guiding microtubule polymerization (Yalgin, 2015).
Intracellular mRNA localization is a conserved mechanism for spatially regulating protein production in polarized cells, such as neurons. The mRNA encoding the translational repressor Nanos (Nos) forms ribonucleoprotein (RNP) particles that are dendritically localized in Drosophila larval class IV dendritic arborization (da) neurons. In nos mutants, class IV da neurons exhibit reduced dendritic branching complexity. This study investigated the mechanism of dendritic nos mRNA localization by analyzing requirements for nos RNP particle motility in class IV da neuron dendrites. Dynein motor machinery components were shown to mediate transport of nos mRNA in proximal dendrites. Two factors, the RNA-binding protein Rumpelstiltskin and the germ plasm protein Oskar function in da neurons for formation and transport of nos RNP particles. nos was shown to regulate neuronal function, most likely independently of its dendritic localization and function in morphogenesis. These results reveal adaptability of localization factors for regulation of a target transcript in different cellular contexts (Xu, 2013).
This study has combined a method that allows live imaging of mRNA in intact Drosophila larvae with genetic analysis to investigate the mechanism underlying transport of nos mRNA in class IV da neurons. Live imaging over the short time periods allowed has provided a snapshot into the steady-state behavior of nos*RFP particles in the proximal dendrites of mature da neurons. The results indicate that anterograde transport of nos RNP particles into and within da neuron dendrites is mediated by dynein and is consistent with the minus-end out model for microtubule polarity in the proximal dendrites of da neurons (Zheng, 2008). This model predicts that bidirectional trafficking would be mediated by opposite polarity motors and the predominance of retrograde movement of nos*RFP particles when dynein function is partially compromised is consistent with this. Moreover, Rab-5 endosomes, whose accumulation in class IV da neuron dendrites is dynein-dependent, also exhibit bidirectional movement (Satoh, 2008), suggesting that different cargos may use similar dendritic transport strategies. Unfortunately, the severe defects caused by loss of kinesin have thus far hampered confirmation of a role for kinesin in these events (Xu, 2013).
The observed bidirectional movement of nos RNP particles resembles the constant bidirectional transport observed for dendritic mRNAs near synapses in hippocampal neurons. In contrast to da neurons, proximal dendrites of mammalian neurons have mixed microtubule polarity so that bidirectional trafficking could be mediated by a single motor that switches microtubules or by switching between the activities of plus-end and minus-end motors. The association of kinesin with neuronal RNP granule components and inhibition of CaMKIIα RNA transport by dominant-negative inhibition of kinesin has implicated kinesin as the primary motor for dendritic mRNA transport. However, a recent study showed that dynein mediates unidirectional transport of vesicle cargoes into dendrites of cultured hippocampal neurons as well as bidirectional transport within the dendrites. Whether dynein plays a role in RNP particle transport in mammalian dendrites as it does in Drosophila neurons remains to be determined (Xu, 2013).
Despite its prevalence, the role of bidirectional motility is not yet clear. A recently proposed 'sushi belt' model suggests that neuronal RNP particles traffic back and forth along the dendrite until they are recruited by an active synapse and disassembled for translation (Doyle, 2011). Although da neuron dendrites do not receive synaptic input, this continual motility may provide a reservoir of nos mRNA that can be rapidly mobilized for translation locally in response to external signals that regulate dendrite branching (Xu, 2013).
These studies have shown that nos mRNA can be adapted for different localization mechanisms depending on cellular context: diffusion/entrapment in late oocytes that lack a requisite polarized microtubule cytoskeleton and microtubule-based transport during germ cell formation in the embryo and in class IV da neurons. Surprisingly, Rump and Osk are specifically required for nos localization in both oocytes and da neurons, suggesting that they function in the assembly or recognition of a fundamental nos RNP that can be adapted to both means of localization. However, because it is not possible to distinguish individual particles within the cell body, the possibility cannot be ruled out that Rump and/or Osk mediate coupling of nos RNP particles to dynein motors rather than particle formation. Within the germ plasm, nos associates with Vasa (Vas), a DEAD-box helicase, and is transported together with Vas into germ cells. Although dendritic branching complexity is reduced in vas mutants, no effect on dendritic localization of nos RNP particles was detected, suggesting that only a subset of germ plasm components are shared by neuronal localization machinery. A role for osk in learning and memory was proposed based on the isolation of an enhancer trap insertion upstream of osk in a screen for mutants with defective long-term memory, but osk function in memory formation has not been directly tested. Notably, however, a recent study showed that the osk ortholog in the cricket Gryllus bimaculatus functions in development of the embryonic nervous system rather than in germ cell formation. Thus, the ancestral function of osk appears to be in neural development, whereas its role in germ plasm formation is a later adaptation in higher insects. The results showing that Osk protein function is not limited to Dipteran germ plasm organization but also plays an important role in neuronal development and function supports this idea (Xu, 2013).
The data indicate that the Nos/Pum complex is not only required for da neuron morphogenesis, but also for nociceptive function. However, nociception does not appear to require local function of Nos/Pum in the dendrites and reduced dendritic branching does not necessarily correlate with a deficit in nociception. These results suggest that morphogenesis and function are regulated separately and that Nos/Pum plays a second role in regulating the somatic translation of proteins required for the nociceptive response. Systematic identification of Nos/Pum targets will be essential to further investigate these different roles (Xu, 2013).
Golgi complexes (Golgi) play important roles in the development and function of neurons. Not only are Golgi present in the neuronal soma (somal Golgi), they also exist in the dendrites as Golgi outposts. Previous studies have shown that Golgi outposts serve as local microtubule-organizing centers and secretory stations in dendrites. It is unknown whether the structure and function of Golgi outposts differ from those of somal Golgi. This study show in Drosophila that, unlike somal Golgi, the biochemically distinct cis, medial, and trans compartments of Golgi are often disconnected in dendrites in vivo. The Golgi structural protein Golgi matrix protein 130 kD ortholog (GM130) is responsible for connecting distinct Golgi compartments in soma and dendritic branch points, and the specific distribution of GM130 determines the compartmental organization of dendritic Golgi in dendritic shafts. It was further shown that compartmental organization regulates the role of Golgi in acentrosomal microtubule growth in dendrites and in dendritic branching. This study provides insights into the structure and function of dendritic Golgi outposts as well as the regulation of compartmental organization of Golgi in general (Zhou, 2014).
Dendritic Golgi outposts have been previously shown to
participate in the patterning of dendritic branches of
Drosophila da neurons. To test whether Golgi compartmental
organization might contribute to dendritic branching, Golgi compartmental organization was examined in several
mutants with dendritic branching defects. Loss-of-function
mutations of the transcription factor
dar1, which reduces
dendritic branching in class III da (C3 da) neurons (Ye, 2011) led to
a decrease in the percentage of multicompartmental Golgi in
dendrites without affecting the Golgi in the soma and branch
points. In contrast, overexpression of Knot, a transcription factor known to
increase dendritic growth, did not change the
compartmental organization in dendrites. These results suggest that certain regulators of dendritic branching and growth may act by regulating the compartmental organization of dendritic Golgi outposts (Zhou, 2014).
A recent study suggests that dendritic Golgi outposts
regulate dendritic branching by functioning as acentrosomal
nucleation sites for microtubules. In light of the current findings
that the Golgi outposts comprise two populations, one with
single compartments and the other with multiple compartments, it was asked whether the structural organization of Golgi outposts regulates microtubule growth. Microtubule growth was examined in da neurons in vivo by time-lapse imaging of
EB1-GFP. EB1 binds to growing microtubule plus
ends and moves in a way that resembles comets (hence
termed 'EB1-GFP comets') as microtubules grow. The association of microtubule growth initiation
with multicompartment outposts was compared to that with single-compartment outpost by live imaging the presence of ManII-EBFP
and GalT-TagRFP together with EB1-GFP in wild-type da
neurons. The number of microtubule initiation events associated with the Golgi outposts containing both medial- and
trans-Golgi compartments was significantly greater than the
events associated with single-compartment outposts. Consistent with the result that Golgi outposts at
branch points contain multiple compartments, branch points
also initiated more microtubule growth than the single-
compartment outposts in dendritic shafts. In the
dendritic shafts, 57.8% of microtubule growth initiation sites
were associated with Golgi outposts containing both
medial and trans compartments, compared to 8.9% for
medial- only and 13.3% for trans- only outposts. 20% of
dendritic microtubule growth initiation sites were either not
associated with dendritic Golgi outposts or associated with
Golgi outposts that were below detection sensitivity.
These results raised the possibility that connecting multiple
Golgi compartments promotes microtubule growth in vivo.
Because introducing dGM130 into dendrites connects the
cis, medial, and trans compartments of dendritic Golgi, the number of initiation events
of microtubule growth in the dendrites were compared between control and
dGM130-overexpressing neurons. The number of microtubule
growth initiation events was significantly increased in the dendritic shafts of neurons overexpressing dGM130 compared to control. The increase was largely due to
events associated with dGM130-containing multicompartment Golgi outposts.
In dGM130 null mutant neurons, microtubule growth initiation events were reduced in distal dendrites. Because Golgi compartments in dGM130-deficient neurons were dispersed in branch points, microtubule growth initiation at these branch points was examined (Zhou, 2014).
Consistently, microtubule growth initiation at dendritic branch
points was dramatically suppressed by
dGM130 mutations. In contrast, loss of
dGMAP, another Golgi structural protein, did not affect microtubule growth initiation. Taken together, these results suggest
that the dGM130-mediated compartmental organization of
dendritic Golgi outposts regulates microtubule growth in
dendrites (Zhou, 2014).
The role of dGM130 was also assessed
in dendritic branching was also assessed.
The total number of dendritic branch points as well as the
number of higher-order branches (fourth order and up) was
significantly reduced in dGM130
mutant class III da neurons
but increased in dGM130-overexpressing neurons of the
same type. These results suggest that
dGM130, and possibly compartmental organization of Golgi
outposts, is a factor that determines the number of higher-
order dendritic branches (Zhou, 2014).
The mechanism underlying the microtubule growth regulated by dGM130 is currently unclear. dGM130 might regulate
microtubule growth through three different mechanisms. First,
different Golgi compartments may each serve a unique role
in microtubule nucleation. Thus, multicompartment Golgi
serve as a functional scaffold for the microtubule nucleation
machinery. Second, GM130, rather than multicompartment
Golgi, may be responsible for initiating microtubule growth.
Third, it is also possible that dGM130 and compartmental
organization of Golgi regulates microtubule growth indirectly
through other Golgi functions such as membrane trafficking.
Previous studies on mammalian hippocampal neurons have
shown that ribbon-like Golgi stacks that are disconnected
from somal Golgi and are positive for GM130 are located
only in the soma and proximal dendrites. This has led to
the speculation that Golgi outposts might exist only in
proximal dendrites. However, this speculation contradicts
the proposal that membrane proteins are synthesized locally
at synapses in distal dendrites and ultrastructural and
immunofluorescence studies showing the presence of membranous organelles positive for Golgi markers. The findings described in this study reconcile this contradiction by
showing that Golgi in the soma and those in the dendrites
assume different compartmental organizations (Zhou, 2014).
Microtubule-organizing centers (MTOCs) often play a central role in organizing the cellular microtubule networks that underlie cell function. In neurons, microtubules in axons and dendrites have distinct polarities. Dendrite-specific Golgi outposts, in particular multi-compartment outposts, have emerged as regulators of acentrosomal microtubule growth, raising the question of whether outposts contribute to establishing or maintaining the overall polarity of the dendritic microtubule cytoskeleton. Using a combination of genetic approaches and live imaging in a Drosophila model, this study found that dendritic microtubule polarity is unaffected by eliminating known regulators of Golgi-dependent microtubule organization including the cis-Golgi matrix protein GM130, the fly AKAP450 ortholog pericentrin-like protein (plp), and centrosomin (cnn). This indicates that Golgi outposts are not essential for the formation or maintenance of a dendrite-specific cytoskeleton. However, the over-expression of GM130, which promotes the formation of ectopic multi-compartment units, is sufficient to alter dendritic microtubule polarity. Axonal microtubule polarity is similarly disrupted by the presence of ectopic multi-compartment Golgi outposts. Notably, multi-compartment outposts alter microtubule polarity independently of microtubule nucleation mediated by the gamma-tubulin ring complex (gamma-TuRC). Thus, although Golgi outposts are not essential to dendritic microtubule polarity, altering their organization correlates with changes to microtubule polarity. Based on these data, it is proposed that the organization of Golgi outposts is carefully regulated to ensure proper dendritic microtubule polarity (Yang, 2020).
GM130 is a matrix protein that is conserved in metazoans and involved in the architecture of the Golgi apparatus. In neurons, Golgi apparatus and dendritic Golgi outposts (GOs) have different compartmental organizations, and GM130 localization is present in both, indicating that GM130 has a unique Golgi-targeting mechanism. This study investigated the Golgi-targeting mechanism of the GM130 homologue, dGM130, using in vivo imaging of Drosophila dendritic arborization (da) neurons. The results showed that two independent Golgi-targeting domains (GTDs) with different Golgi localization characteristics in dGM130, together determined the precise localization of dGM130 in both the soma and dendrites. GTD1, covering the first coiled-coil region, preferentially targeted to somal Golgi rather than GOs; whereas GTD2, containing the second coiled-coil region and C-terminus, dynamically targeted to Golgi in both soma and dendrites. These findings suggest that there are two distinct mechanisms by which dGM130 targets to the Golgi apparatus and GOs, underlying the structural differences between them, and further provides new insights into the formation of neuronal polarity (Cheng, 2023).
The directional flow of information in neurons depends on compartmentalization: dendrites receive inputs whereas axons transmit them. Axons and dendrites likewise contain structurally and functionally distinct subcompartments. Axon/dendrite compartmentalization can be attributed to neuronal polarization, but the developmental origin of subcompartments in axons and dendrites is less well understood. To identify the developmental bases for compartment-specific patterning in dendrites, a screen was carried out for mutations that affect discrete dendritic domains in Drosophila sensory neurons. From this screen, mutations were identified that affected distinct aspects of terminal dendrite development with little or no effect on major dendrite patterning. Mutation of one gene, raw, affected multiple aspects of terminal dendrite patterning, suggesting that Raw might coordinate multiple signaling pathways to shape terminal dendrite growth. Consistent with this notion, Raw localizes to branch-points and promotes dendrite stabilization together with the Tricornered (Trc) kinase via effects on cell adhesion. Raw independently influences terminal dendrite elongation through a mechanism that involves modulation of the cytoskeleton, and this pathway is likely to involve the RNA-binding protein Argonaute 1 (AGO1), as raw and AGO1 genetically interact to promote terminal dendrite growth but not adhesion. Thus, Raw defines a potential point of convergence in distinct pathways shaping terminal dendrite patterning (Lee, 2015).
Although the concept of positional information was first applied to embryonic development, intracellular positional information governs morphogenesis of individual cells as well. For example, positioning the nucleus at the cell center and growth zones at the cell periphery depends on positional information from the microtubule cytoskeleton in Schizosaccharomyces pombe. Several lines of evidence support the existence of distinct subcompartments in axons and dendrites, but the forms of intracellular positional information and the coordinate systems that guide the development of these subcompartments have not been extensively characterized. Results from this screen and other studies suggest that at least two types of positional information govern C4da dendrite patterning. First, terminal branch distribution along the proximal-distal axis depends on microtubule-based processes; perturbing microtubule-based transport leads to a distal-proximal shift in the distribution of terminal dendrites in C4da arbors. Interestingly, modulating the activity of the F-actin nucleator Spire also affects terminal dendrite positioning along the proximal-distal axis, suggesting that multiple pathways contribute to the fidelity of branch placement. Second, terminal dendrites rely on dedicated programs that may act locally to regulate terminal dendrite patterning. The observation that different pathways regulate different aspects of terminal dendrite development suggests that multiple signaling systems exist for the local control of dendrite growth (Lee, 2015).
This study identified raw as a key regulator of terminal dendrite patterning. raw encodes a membrane protein that accumulates at branch-points and coordinately regulates terminal dendrite adhesion/stability via a pathway that involves Trc and terminal dendrite elongation via a pathway that is likely to involve cytoskeletal remodeling and AGO1. Raw therefore provides a potential point of integration for external signals that regulate these downstream growth programs. These pathways could be responsive to the same signal -- for example, Raw association with an extracellular ligand or a co-receptor -- or could be spatially/sequentially segregated. Identification of additional raw-interacting genes should help clarify the architecture of these signaling pathways (Lee, 2015).
Raw regulates cell-cell signaling, and in gonad morphogenesis Raw modulates Cadherin-based interactions between somatic gonadal precursor cells and germ cells, in part by localizing Armadillo to the cell surface. Likewise, the data support a role for Raw in promoting Trc activation by localizing Trc to the plasma membrane. Thus, one plausible model for Raw function in dendrite development is that it interacts with an extracellular signal, which might be a component of the ECM or a cell surface protein on epithelial cells, and signals together with a co-receptor to stimulate downstream pathways for adhesion and cytoskeletal remodeling. Several analogous signaling systems involving interactions with the epidermis that influence terminal dendrite or sensory axon patterning have been described, but how many of these signaling systems are at work in a given neuron, and how Raw interfaces with other signaling pathways, remain to be determined (Lee, 2015).
Although Raw has no obvious vertebrate counterpart, stretches of the ECD bear similarity to mucins and leucine-rich repeat proteins, one of which might serve an analogous function. Moreover, components of both downstream signaling pathways that this study identified are conserved in vertebrates and play known roles in dendrite patterning, including roles in the local control of dendrite growth: the Trc orthologs NDR1/2 (STK38/STK38L) regulate aspects of dendrite branch and spine morphogenesis, and Argonaute proteins mediate miRNA-mediated control of dendrite patterning, in part through local effects on translation. Additionally, dendrites contain structures related to P-granules, and Argonaute proteins may influence local translation in P-granules as well. Thus, versions of the Raw-regulated signaling pathways might control terminal dendrite patterning in vertebrates (Lee, 2015).
During developmental remodeling, neurites destined for pruning often degenerate on-site. Physical injury also induces degeneration of neurites distal to the injury site. Prompt clearance of degenerating neurites is important for maintaining tissue homeostasis and preventing inflammatory responses. This study shows that in both dendrite pruning and dendrite injury of Drosophila sensory neurons, epidermal cells rather than hemocytes are the primary phagocytes in clearing degenerating dendrites. Epidermal cells act via Draper-mediated recognition to facilitate dendrite degeneration and to engulf and degrade degenerating dendrites. Using multiple dendritic membrane markers to trace phagocytosis, it was shown that two members of the CD36 family, croquemort (crq) and debris buster (dsb), act at distinct stages of phagosome maturation for dendrite clearance. These findings reveals the physiological importance of coordination between neurons and their surrounding epidermis, for both dendrite fragmentation and clearance (Han, 2014).
Removal of nonfunctional or damaged tissues is an important
biological process during tissue remodeling or repair. This study
shows that, for Drosophila class IV da neurons in the periphery,
degenerating dendrites in both dendrite pruning and injury
models are removed by neighboring epithelial cells rather than
professional phagocytes. By developing multiple dendritic
markers that label phagosomes differentially,
the clearance of degenerating dendrites was established as an in vivo model to
study phagocytosis. With these tools, key players
in engulfment and phagosome maturation were analyzed, and roles
of the CD36 family members Crq and Dsb were elucidated. This study further reveals
that, as phagocytes, epidermal cells actively participate in
not only the removal but also the fragmentation of degenerating
dendrites (Han, 2014).
Professional phagocytes such as macrophages in vertebrates
and plasmatocytes in Drosophila dispose the majority of
apoptotic cells in development, as well as invading microorganisms
during infection. However,
nonprofessional phagocytes may take charge when
macrophages or other professional phagocytes are absent or
cannot easily access cell corpses, as in apoptosis of rat lens
cells, follicular atresia, and degeneration of Drosophila egg chambers induced by
protein deprivation. This scenario
does not apply to Drosophila da neuronal dendrites, which are
exposed to circulating plasmatocytes in the hemolymph and
sessile plasmatocytes clustered around da neuron somas. Indeed, previous observation
of dendrite debris engulfment by plasmatocytes during
dendrite pruning has led to the conclusion that plasmatocytes
clear pruned dendrites. The current finding
that clearance of degenerating dendrites is mainly carried out
by epidermal epithelial cells demonstrates that nonprofessional
phagocytes are not just a substitute for professional phagocytes
in their absence. Rather, plasmatocytes and epidermal cells
probably carry out different functions reflecting specialization
of cellular functions. The removal of pruned dendrites by
Drosophila epidermal cells perhaps can be seen as a parallel
to the clearance of photoreceptor outer segments by retinal
pigment epithelial cells; in both cases
epithelial cells maintain homeostasis of the nervous system
as part of their physiological functions. The observation that
epidermal cells are also responsible for clearing injured dendrites
indicates that the same cellular mechanism is also used
to cope with perturbations in the peripheral nervous system (Han, 2014).
Epithelial cells may profoundly influence the development of
dendritic arbors of da neurons. During larval development,
growing epithelial cells signal to the dendritic arbors so they
can grow proportionally to epithelial cells in order to maintain
the same coverage of receptive fields of the sensory neurons, a
phenomenon known as dendritic scaling.
Epithelial cells also contribute to the patterning of dendritic
arbors of da neurons by tethering dendrites to the 2D space of
the extracellular matrix so that dendrites have to avoid sister
dendrites from the same neuron (self-avoidance) or dendrites
from neighboring like-neurons (tiling). The finding that epithelial cells mediate the clearance of degenerating dendrites substantially adds to the
growing list of dendrite properties regulated by epithelial cells (Han, 2014).
The vertebrate CD36 family members CD36 and scavenger receptor
class B type I (SR-BI) mediate phagocytosis of apoptotic
cells and microbial pathogens in vitro. The Drosophila CD36
family member Crq is required for efficient phagocytosis of cell
corpses in embryos and mediates binding of apoptotic cells by in vitro cultured cells,
leading to its proposed role as a receptor for apoptotic cells. This study shows that in epithelial cells crq is required for
phagosome maturation but not for the engulfment of degenerating
dendrites. As loss of crq does not completely abolish the
engulfment of apoptotic cells in the embryo, it is possible that the cell-corpse clearance defect in crq mutant embryos may be a consequence of blocked phagosome
maturation. An alternative possibility is that Crq may be required
for engulfment and/or phagosome maturation of apoptotic cells
by embryonic macrophages but only required for phagosome
maturation of pruned or injured dendrites by epithelial cells.
This could be due to the fact that macrophages have to actively
search for and bind apoptotic cells, while epithelial cells engulf
neighboring debris. Further experiments will be needed to determine
whether Crq also plays a role in phagosome maturation
during phagocytosis by macrophages (Han, 2014).
Loss of Crq function resulted in the fusion of dendrite-derived
phagosomes accompanied with a failure of degradation of
phagosome contents, most likely due to inefficient delivery of
degradation machineries to late phagosomes. As phagosomes
normally acquire hydrolases and other phagolysosomal components
by fusing with endosomes and lysosomes, it is hypothesized
that Crq suppresses homotypic phagosome fusion to promote
fusion between phagosomes and late endosomes/lysosomes.
Homotypic phagosome fusion rarely happens during normal
phagocytosis but is induced by infection of bacterial pathogens
such as Helicobacter pylori and Chlamydia trachomatis; the ability of different strains
of H. pylori to induce phagosome fusion correlates with the
virulence and intracellular survival of these bacteria. Therefore, regulation of
the balance between homotypic phagosome fusion and heterotypic
fusion between phagosomes and late endosomes/lysosomes
is probably critical for the degradation of internalized materials (Han, 2014).
The Drosophila genome encodes fourteen CD36 family members.
Besides the involvement of Crq in phagocytosis, another
member Pes mediates mycobacteria infection. This study found that the CD36 family member
Dsb regulates late stages of phagosome maturation. Interestingly,
LIMP-2, the mammalian CD36 family member with the
highest homology to Dsb, is an intrinsic lysosomal protein
required for the degradation of Listeria in phagosomes. Dsb and LIMP-2 thus appear to have
evolutionarily conserved functions in phagosome maturation (Han, 2014).
Phagocytes not only clear cell corpses but may also engulf still-living
cells and promote cellular degeneration in many contexts. This study shows that efficient degeneration of dendrites requires the coordination with phagocytic epithelial
cells. One mechanism for such coordination is the Drpr-mediated
recognition of degenerating dendrites by epidermal phagocytes
that form actin-rich membrane structures wrapping around
the dendrites to facilitate their fragmentation. In the postnatal
mouse brain, microglia actively induce apoptosis of Purkinje
cells by producing superoxide ions. It
remains to be determined whether nonprofessional phagocytes
such as epidermal cells also promote neurite degeneration by
emitting diffusible agents (Han, 2014).
Double-bromo and extraterminal (BET) domain proteins regulate dendrite morphology and mechanosensory function
A complex array of genetic factors regulates neuronal dendrite morphology. Epigenetic regulation of gene expression represents a plausible mechanism to control pathways responsible for specific dendritic arbor shapes. By studying the Drosophila dendritic arborization (da) neurons, this study discovered a role of the double-bromodomain and extraterminal (BET) family proteins in regulating dendrite arbor complexity. A loss-of-function mutation in the single Drosophila BET protein encoded by female sterile 1 homeotic [fs(1)h] causes loss of fine, terminal dendritic branches. Moreover, fs(1)h is necessary for the induction of branching caused by a previously identified transcription factor, Cut (Ct), which regulates subtype-specific dendrite morphology. Finally, disrupting fs(1)h function impairs the mechanosensory response of class III da sensory neurons without compromising the expression of the ion channel NompC, which mediates the mechanosensitive response. Thus, these results identify a novel role for BET family proteins in regulating dendrite morphology and a possible separation of developmental pathways specifying neural cell morphology and ion channel expression. Since the BET proteins are known to bind acetylated histone tails, these results also suggest a role of epigenetic histone modifications and the 'histone code,' in regulating dendrite morphology (Bagley, 2014).
Dendrites are the primary site of information input to neural circuits, and the shape of dendritic arbors influences the electrophysiological responses of neurons. Due to the existence of highly diverse morphologies among different neuronal subtypes, a question of the relationship between form and function arises: By understanding how the shape of a neuron is specified, it is possible to understand how morphology relates to neural function and how altered morphology relates to dysfunction (Bagley, 2014).
Neurons can be defined by their physiology, morphology, and gene expression. Neuronal diversity is thought to arise from the combinatorial expression of genetic determinants. The dendritic arborization (da) sensory neurons of the Drosophila peripheral nervous system (PNS) constitute a powerful system to study genetic determinants of dendritic arbor morphology. In particular, the use of Drosophila genetic techniques to study the specification of stereotyped, subtype-specific dendritic arbor shapes resulted in the identification of multiple transcription factors, encoded by abrupt (ab), knot/collier (kn/col), spineless (ss), and cut (ct), which regulate dendritic arbor morphology. However, large-scale genomic analyses comparing the transcriptomes of various neural subtypes indicate a daunting amount of varied gene expression and implicate regulation by multiple transcription factors. Thus, a particular neuronal morphology is likely the result of coordination between multiple genomic programs (Bagley, 2014).
The post-translational modification of histone tails involves three types of molecules: The 'writers' add methyl, acetyl, or phospho groups and consist of histone methyl transferase (HMT), histone acetyltransferase (HAT), and kinase enzymes. The 'erasers' remove these modifications and include demethylases (DMTs), histone deacetyltransferases (HDACs), and phosphatases. Finally, the 'readers' are scaffolding proteins that recognize and bind acetyl, methyl, or phosphate modifications to position the 'writer' and 'eraser' enzymes along with transcriptional machinery to the correct genomic position and thereby modify gene expression. The discovery that the Polycomb repressor complex, which binds methylated histone tails, regulates da sensory neuron dendrite morphology indicates a role of histone methylation in dendrite development and a notion supported by the recent finding that the chromodomain Y-like (CDYL) protein negatively regulates dendritic complexity (Qi, 2014). Regarding a role of histone acetylation in dendrite morphogenesis, both HDAC and HAT activities have been implicated in regulating dendrite morphology. Specifically, the Drosophila HDAC1/2 homolog Rpd3 regulates class I da sensory neuron morphology and olfactory PN dendritic targeting. In addition, HDAC2 suppresses dendritic spine density of hippocampal CA1 and dentate granule neurons. The HAT enzyme Pcaf also regulates class I da sensory neuron dendrite morphology. A different HAT enzyme, CREB-binding protein (CBP), regulates the developmental pruning of class IV da sensory neuron dendrites, and mutations in the human homolog CREBBP cause the mental retardation syndrome Rubenstein-Taybi. While these studies indicate a definite role of 'writers' and 'erasers' of histone modifications in regulating dendrite morphogenesis, the role of 'reader' scaffolding proteins associated with histone acetylation has not been thoroughly investigated (Bagley, 2014).
Double-bromo and extraterminal (BET) domain-containing proteins bind acetylated histone tails (Umehara 2010a; Umehara 2010b) and modulate gene expression. In mice, mutations in one BET family member, BRD2, cause neural tube closure defects, behavioral abnormalities, and altered interneuron numbers. In addition, in certain human genomic population studies, mutations in BRD2 have been associated with juvenile myoclonic epilepsy and photosensitivity, which is frequently observed in idiopathic generalized epilepsies. In the current study, evidence is provided for a role of the Drosophila homolog of BRD2, encoded by female sterile 1 homeotic [fs(1)h], in regulating dendrite morphology and sensory function (Bagley, 2014).
This study examined the role of fs(1)h in dendritic development. The effect of a loss-of-function allele [fs(1)h1112] was examined on the morphology of class III da sensory neurons in the Drosophila PNS. Overall, fs(1)h1112 causes a reduction in dendritic arbor complexity, most notably in the finer, higher-order branches. It was possible to partially rescue this reduced morphological complexity by reintroducing Drosophila Fsh-S or the human homolog (huBRD2) proteins. Furthermore, one aspect of the genetic mechanism of action for fs(1)h was found to be regulating the expression of ct (and possibly other genes in the pathway) in multiple da neuron subtypes as well as subtype-specific transcription factors, such as Abrupt for class I and Knot/Collier for class IV da neurons, which in turn affect subtype-specific dendrite development. The data show that fs(1)h regulates genetic pathways controlling dendritic arbor development but does not specify which ion channels are expressed. Finally, the results suggest that the subtype-specific spike morphology is important for an optimal response to relevant sensory stimuli in the mechanosensitive class III da neurons (Bagley, 2014).
The development of a dendritic arbor involves multiple steps beginning with differentiation, where a neuronal precursor acquires a neural fate. Next, neurites begin to extend, and a neuron becomes polarized as neurites are designated as axon or dendrite. The immature axons and dendrites continue to grow as the neuron and the nervous system develop. Initially, the dendritic arbors are simple, with only a few primary dendrites, but as development progresses, the number of branches and overall arbor size increase. The terminal dendrite branches are dynamic throughout development, exhibiting growth, retraction, or stability. In addition, as the animal body size increases, the dendritic field area increases, and therefore a dendritic arbor must scale accordingly. Thus, dendritic development involves a complex plethora of processes, and dendritic morphology could be altered by affecting any of these processes. For instance, if the balance of dendrite dynamics is shifted such that retraction is greater than growth, then dendritic branching will become reduced over time. This appears to be the case in fs(1)h1112 mutants, since an increase was observed in retracting branches with no change in growth as well as a decrease in the proportion of stable branches in dendritic arbors of fs(1)h1112 mutant da neuron clones compared with wild-type clones. Alternatively, if scaling of the dendritic arbor is affected, the size of the dendritic arbor will become disproportionately small as the body size of the animal increases throughout development. This does not seem to occur in fs(1)h1112 mutants because the primary dendrites of fs(1)h1112 arbors exhibited growth throughout development, although at a delayed rate. Instead, the number of spikes in class III da neuron arbors was reduced early in development and remained reduced throughout development, probably due to the increased amount of dendritic branch retraction and reduced stability. Since the primary dendritic branches were not affected to a large degree by loss of fs(1)h function, it is concluded that the major role of fs(1)h in dendritic development is to regulate dendritic complexity at the level of higher-order dendritic spikes. Moreover, the data suggest that fs(1)h affects dendritic arbor complexity by modulating the dynamics of terminal dendritic branches (Bagley, 2014).
In the da neurons, many molecules are known to regulate dendrite morphology. In particular, Ct, Ss, Ab, and Kn have been shown to regulate subtype-specific morphology of the four classes of da sensory neurons, and these proteins act in parallel genetic pathways. Moreover, the expression of Ct and Ss regulates class III da neuron spike morphology. This study observed a loss of Ct expression in fs(1)h1112 mutant class III da neuron clones, which suggests that fs(1)h regulates the induction or maintenance of Ct expression throughout class III da neuron development. However, reintroducing Ct expression to class III da neuron fs(1)h1112 clones did not rescue the nearly absent spike morphology. Therefore, the class III da neuron dendritic phenotype caused by loss of fs(1)h cannot be solely attributed to the loss of Ct protein. Since it is thought that Ct is a component of a genetic pathway responsible for subtype-specific dendritic arbor development, it is possible that fs(1)h regulates Ct expression as well as expression of genes necessary for the Ct pathway to affect dendritic morphology. Therefore, the relationship between ct and fs(1)h does not appear to be a linear pathway, and fs(1)h might regulate both upstream and downstream components of ct. These data indicating that fs(1)h is necessary for the Ct-induced overbranching and spike formation in class I da neuron dendrites support the idea that fs(1)h regulates the expression of downstream components of the Ct pathway, which are necessary for Ct-induced overbranching and spike formation. This hypothesis also explains why reintroducing Ct expression to fs(1)h1112 clones fails to rescue the dendrite phenotype. It is also known that Ct and Rac1 act synergistically to produce spike morphology. This study examined Rac1 overexpression in a fs(1)h1112 mutant background and found that Rac1 expression significantly rescued the loss of spikes in class III da neurons. However, Rac1-induced overbranching in class I da neurons was not affected by fs(1)h1112. Therefore, fs(1)h does not appear to regulate genes downstream from Rac1 but does regulate genes downstream from ct. Since these pathways are known to converge in order to regulate dendritic spike formation, the current data suggest that fs(1)h may be a crucial link between these two pathways. One possible scenario is that ct and Rac1 regulate parallel pathways, but ct may regulate the level of Rac1 expression such that increased Rac1 expression facilitates the formation of spikes. In this model, the results support the hypothesis that fs(1)h is necessary for the ct potentiation of Rac1 expression, which explains why increased expression of Rac1 with UAS-Rac1 causes a rescue of the class III da neuron dendritic phenotype in fs(1)h1112 mutants. Recent evidence indicates a role for reduced Rac1 expression in social defeat and depressive behavior in mice, possibly through regulating dendritic spine morphology (Golden, 2013). In these behavioral paradigms, reduced Rac1 expression occurred with altered epigenetic marks such that transcriptionally permissive histone H3 acetylation was reduced, while repressive histone H3 methylation was increased. Moreover, administering a class 1 HDAC inhibitor mitigated the reduced Rac1 expression. Thus, these data suggest that Rac1 expression can be regulated by histone acetylation. It is possible that epigenetic reader proteins, such as BET family proteins like fs(1)h, bind acetylated histone marks in the Rac1 promoter to recruit transcriptional machinery and in turn enhance Rac1 expression (Bagley, 2014).
In addition, overexpression of UAS-Fsh-S in class I da neurons did not cause an overbranching phenotype similar to UAS-Ct. In fact, there was no alteration of class I morphology, suggesting that Fsh-S is not sufficient to induce necessary components of the ct pathway to alter dendrite morphology. However, overexpression of Fsh-S in class III and class IV da neurons did cause a decrease in dendritic spike numbers. These data indicate that dendrite morphology may be sensitive to the amount of Fsh-S expression, which was confirmed by modulating the amount of overexpression by reducing GAL4/UAS activity with lower temperature. This may explain why it is possible to achieve only a partial rescue of the fs(1)h1112 dendritic phenotype with UAS-Fsh-S expression and why overexpression causes a dendritic phenotype similar to the phenotype caused by loss of Fsh-S. In support of this expression level hypothesis, it was observed that Fsh-S overexpression can reduce Ct-induced branching in class I da neurons. Since BRD2 is known to be part of a protein complex (Denis 2006), it is possible overexpression causes a gain-of-function or dominant-negative effect by altering the availability of complex components (Bagley, 2014).
Another possible explanation for the partial rescue of Fsh-S expression concerns the developmental timing of expression. Since these experiments were completed using MARCM, GAL80 is expressed until mitotic recombination occurs to generate the mutant clones. It is likely that GAL80 protein may persist for some time after the clones are formed, and the presence of GAL80 would block GAL4/UAS activity. Therefore, UAS-induced Fsh-S expression may occur at a delayed stage in embryonic development, which could produce a partial rescue. In actuality, a combination of both expression level and developmental timing probably explains the partial rescue of the fs(1)h1112 phenotype (Bagley, 2014).
While this study focused on the role of fs(1)h in regulating class III da neuron dendrite morphology, phenotypes were observed in other classes of the da neurons as well as expression of Fsh-S in all da neuron classes. In fs(1)h1112 mutants, a loss of Ct expression was observed in all da neurons that normally express Ct (classes II, III, and IV), suggesting that fs(1)h regulates Ct expression broadly among different neural subtypes. A loss of the class I-specific transcription factor Ab and the class IV-specific transcription factor Kn/Col wer also observed. Thus, it appears that fs(1)h can regulate the expression of subtype-specific gene expression among various neuron subtypes. The loss of Ct or the loss of Kn/Col could explain the reduction in class IV da neuron dendritic arbor complexity, and this further illustrates the pleiotropic nature of the fs(1)h1112 phenotype. The loss of Ab from class I da neurons should produce an increase in dendritic complexity, but interestingly, this did not occur in fs(1)h1112 mutants. Thus, these results consistently suggest that fs(1)h is necessary for dendritic arbor complexity, probably by regulating the expression of many different genes. In this manner, fs(1)h could act as a necessary gate for the gene expression responsible for establishing dendritic complexity (Bagley, 2014).
How can fs(1)h regulate gene expression? Histone modifications are a diverse set of post-translational modifications that produce a code whereby epigenetic reader proteins bind these modified histone tails with specificity for particular modifications, such as methylation, acetylation, or phosphorylation. Previous structural studies have shown that the bromodomains of BET family proteins form a hydrophobic pocket enveloping acetylated histone tails (Umehara 2010a; Umehara, 2010b). Moreover, histone acetylation is largely, but not exclusively, regarded as a mark for transcriptional activation. Therefore, fs(1)h may be required for transcriptional activation of gene expression, which has been shown in vitro with respect to Ubx (Chang, 2007). The current data suggest that fs(1)h is required for ct expression and is in agreement with the hypothesis that fs(1)h is a transcriptional activator. It is possible that expression of other genes in the ct pathway also depends on histone acetylation modifications for transcriptional activation, and this activation may require Fsh-S. This would explain the observed nonlinear genetic relationship between ct and fs(1)h. In addition, the results indicate a necessary, but not sufficient, role of fs(1)h in regulating gene expression. This may indicate that BET family proteins require histone acetylation marks to be established but that these scaffold reader proteins do not actively alter histone tail modifications (Bagley, 2014).
Histone modifications, termed the histone code, vary among different cell types and constitute a genome-wide mechanism for coordinating gene expression programs. This is intriguing because fs(1)h contains bromodomains that require histone acetylation to be first established at specific genomic regions in order to influence transcription at these regions. The current results suggest BET family proteins as candidates for reading this histone code to allow the development of dendritic complexity. It is important to note that although many proteins are observed with altered expression in fs(1)h1112 mutant da neurons, some proteins were unaltered, such as the mechanosensitive ion channel NompC. Furthermore, even though the Ct-induced overbranching in class I da neurons was blocked by fs(1)h1112, the Ct-induced NompC expression was normal. These data indicate some specificity to the action of fs(1)h in regulating dendritic morphology but not ion channel specification. It is possible that epigenetic 'reader' proteins, such as the BET proteins, coordinate the activity of many genetic pathways but with relevance to a specific outcome, such as regulating dendritic arbor morphology. In this model, the epigenetic 'readers' provide coordination and specificity of genome-wide histone marks to regulate particular aspects of neural cell biology. Moreover, it is conceivable that the specific genes regulated by fs(1)h could vary among different cell types depending on the cell type-specific histone code. This is supported by the different effects of UAS-Fsh-S overexpression in class I versus class III and IV da neurons as well as the loss of expression of cell type-specific transcription factors (Ab and Kn/Col) in fs(1)h1112 mutants. Currently, there is no atlas of the histone code for individual neural subpopulations. However, as the technology for conducting these types of analyses improves for distinct cell populations, it is conceivable that future studies can provide an answer to how cell type-specific histone modifications affect neural subtype-specific dendritic arbor morphologies (Bagley, 2014).
Finally, the results suggest that the specific morphological shape of the class III da neuron dendrites is important for their ability to appropriately respond to sensory stimuli. The results indicate that pathways regulating dendrite morphology, such as the ct pathway, are reduced in fs(1)h mutants, but other pathways involved in axon morphogenesis or cell type-specific physiology, such as NompC channel expression, remain active. Moreover, the number of spike protrusions correlates with the number of APs produced in response to a mechanosensitive stimulus. This was also observed in another study (Tsubouchi 2012) involving manipulation of the number of spiked protrusions through modulating Rac1 activity. In that study, the gentle touch response increases as spike numbers increase, causing elevated calcium activity detectable with GCaMP fluorescence imaging. Conversely, decreasing the spike numbers results in a decrease of the gentle touch response and calcium activity. One potential caveat to this study is that Rac1 can modulate many aspects of dendritic cell biology through modulating actin cytoskeletal dynamics, and therefore it is unclear whether manipulating Rac1 activity alters the electrophysiological properties or localization of ion channels such as NompC. The finding of a correlation between dendritic spike number and gentle touch/electrophysiological responses in fs(1)h mutant neurons with normal appearance of NompC expression implicates dendritic morphology in regulating touch sensitivity (Bagley, 2014).
Interestingly, NompC is expressed in fs(1)h1112 mutants, and its distribution throughout the dendritic arbor resembles that of wild-type neurons. While nompC mutants lack a mechanosensory response, neurons lacking fs(1)h still respond to mechanical stimuli, but the magnitude of the response (number of APs) is reduced for a given stimulus intensity. At the behavioral level, this manifests as a reduced response to gentle touch. Therefore, the data suggest that the unique dendritic spike morphology of class III dendrites contributes to their mechanical sensitivity (Bagley, 2014).
While various proteins involved in epigenetic regulation of gene expression have been implicated in dendrite morphogenesis, this study provides evidence that 'readers' of acetylated histone marks regulate dendrite morphology by demonstrating the involvement of BET family proteins in this process. Given the complexity of achieving a comprehensive view of molecularly defined neural subtypes, it is necessary to identify genome-wide mechanisms for molecular diversity that regulate dendritic morphology in order to further understand how morphological diversity is specified. Epigenetic regulators are an intriguing possibility in this endeavor, and future studies comparing gene expression profiles in mutants for regulators of histone modifications among neurons with varied morphologies may be one step forward in answering this fundamental question (Bagley, 2014).
A subset of sound-detecting Johnston's Organ neurons (JONs) in Drosophila melanogaster that express the transcription factors Engrailed (En) and Invected (Inv) form mixed electrical and chemical synaptic inputs onto the giant fiber (GF) dendrites. These synaptic connections are detected by trans-synaptic Neurobiotin (NB) transfer and by colocalization of Bruchpilot-short puncta. Misexpressing En postmitotically in a second subset of sound-responsive JONs causes them to form ectopic electrical and chemical synapses with the GF, in turn causing that postsynaptic neuron to redistribute its dendritic branches into the vicinity of these afferents. A simple electrophysiological recording paradigm was introduced for quantifying the presynaptic and postsynaptic electrical activity at this synapse, by measuring the extracellular sound-evoked potentials (SEPs) from the antennal nerve while monitoring the likelihood of the GF firing an action potential in response to simultaneous subthreshold sound and voltage stimuli. Ectopic presynaptic expression of En strengthens the synaptic connection, consistent with there being more synaptic contacts formed. Finally, RNAi-mediated knockdown of En and Inv in postmitotic neurons reduces SEP amplitude but also reduces synaptic strength at the JON-GF synapse. Overall, these results suggest that En and Inv in JONs regulate both neuronal excitability and synaptic connectivity (Pezier, 2014).
The dendrite of the sensory neuron is surrounded by support cells and is composed of two specialized compartments: the inner segment and the sensory cilium. How the sensory dendrite is formed and maintained is not well understood. Hook-related proteins (HkRP) like Girdin, DAPLE, and Gipie are actin-binding proteins, implicated in actin organization and in cell motility. This study shows that the Drosophila melanogaster single member of the Hook-related protein family, Girdin, is essential for sensory dendrite formation and function. Mutations in girdin were identified during a screen for fly mutants with no mechanosensory function. Physiological, morphological, and ultra-structural studies of girdin mutant flies indicate that the mechanosensory neurons innervating external sensory organs (bristles) initially form a ciliated dendrite that degenerates shortly after, followed by the clustering of their cell bodies. Importantly, it was observed that Girdin is expressed transiently during dendrite morphogenesis in three previously unidentified actin-based structures surrounding the inner segment tip and the sensory cilium. These actin structures are largely missing in girdin. Defects in cilia are observed in other sensory organs such as those mediating olfaction and taste, suggesting that Girdin has a general role in forming sensory dendrites in Drosophila. These suggest that Girdin functions temporarily within the sensory organ and that this function is essential for the formation of the sensory dendrites via actin structures (Ha, 2015).
This study reports the identification and characterization of girdin mutants and their role in dendrite formation. This study shows a novel function of Girdin during sensory neuron development in Drosophila, suggesting that Girdin is essential for morphogenesis of sensory dendrites. This conclusion stems from several lines of evidence. First, girdin101 exhibits complete unc-type uncoordination, a phenotype that is present only in flies with mutations in the sensory neuron or its attachment to the bristle. Second, girdin101 flies have no mechanoreceptor currents (MRC) but a normal transepithelial potentials (TEP), the socket and the bristle appear normal, and the dendrite sheath is present, suggesting that, although the support cells of the sensory organ are present, the neuron itself is defective. Third, in girdin101, GFP-tubulin labeling exhibits dramatic defects in dendrite and cilium morphogenesis but a normal morphology of the cell body and axon. Finally, Girdin-GFP is present in relatively high intensity in the sensory organ at the general location of the dendritic tip during its morphogenesis (Ha, 2015).
This finding suggests a role for Girdin in dendrite formation and neuron cell body positioning. During the early stages of mechanosensory neuron development, the morphology of the cell body, dendrite, and cilium all appear normal. Only later in development, the ciliated dendrite begins shortening. This degeneration continues until the dendrites are fully absent and the cell bodies become clustered. Since the cell bodies in girdin101 tend to form a cluster after their dendrites degenerate, it is possible that the dendrite is essential for positioning of the sensory neuron cell body. Alternatively, since Girdin is important in cell migration in many cell types (Enomoto, 2005; Ghosh, 2008), it is possible that the mislocalization of the cell body is due to a function of Girdin in cell migration. One possible explanation for the clustering of the cell bodies is that cell bodies migrate along their axons until they bundle. In C. elegans, dendrites of ciliated sensory cells are extended in a retrograde fashion where the cell body migrates away from the dendritic tip, which remains anchored in place. A similar mechanism that regulates cell-body positioning during mechanosensory neuronal development may also occur in Drosophila (Ha, 2015).
Girdin shares some similarities with NompA: both are essential for fly viability, coordinated behavior, and MRC, while both are not essential for bristle or socket formation and TEP. Both have a similar dendrite formation defect in which the tip of the sensory dendrite terminates at a significant distance from the bristle base and socket. Finally, both NompA and Girdin localize inside the sensory organ at the vicinity of the dendritic tip. Taking together, these phenotypes and localization suggest that Girdin is associated with the function of NompA to attach the dendritic tip to the base of the bristle for the mechanical transduction of mechanosensory signals. However, since Girdin is expressed only transiently in the sensory organ, it is likely to have only a developmental role in forming the mechanosensory organ rather than a direct role in mechanosensation as is proposed for NompA. Also, since girdin101 sensory neurons showed a more severe defect in dendrite morphology than nompA, Girdin might have additional functions beyond temporarily connecting the dendrite to the bristle shaft. One such additional function of Girdin may be to stabilize the dendrite as it differentiates (Ha, 2015).
Girdin is an intracellular, actin-binding protein that associates with the plasma membrane or cytoplasm of epithelial cells. In the sensory organ cells, Girdin-GFP is present in three actin structures surrounding the tip of the inner segment and the sensory cilium. Phalloidin staining relative to neurons expressing GFP-tubulin suggests that the cap-like and loop-like structures of actin are found outside the sensory neuron and in the support cells. Since actin colocalizes with Girdin-GFP, this suggests that Girdin-GFP is a component of the support cells at these two locations. On the other hand, the tube-like structure of actin and Girdin-GFP appears to localize near the inner segment and may be part of the neuron or sheath cell (Ha, 2015).
The Girdin-GFP-labeled tube-like structure is found at the tip of the inner segment, a specialized location that has cell-cell contacts with the sheath cell. The Girdin-GFP-labeled cap-like structure is found at the tip of the cilium where the dendritic sheath forms during neuron differentiation. The Girdin-GFP-labeled loop-like structure is found away from the neuronal tip as determined by neurons expressing GFP-tubulin, and the loop-like structure diameter is too large to fit within the dimensions of either the sensory neuron or the sheath cell. Therefore, it is likely that the loop-like structure is inside either the shaft or the socket support cells, which are bigger and are found at that location. Which of these three Girdin-GFP/actin structures is essential for normal dendrite formation is currently not known (Ha, 2015).
Based on the current findings and known functions of Girdin, a model for Girdin function in sensory organs during sensory neuron development is proposed (see A model for Girdin function during sensory neuron development). This study shows that Girdin functions near the tip of the dendrite at three locations. In these locations, Girdin regulates the formation of actin structures that may temporarily stabilize the dendritic tip and is essential for its connection to the dendritic sheath. In girdin mutants, dendrite degeneration is likely due to a failure in complete development of the dendrite after its initiation. Once the tip of the dendrite is destabilized and disappears, the neuron cell body recedes toward the axon until it meets other sensory cell bodies and forms clusters where the individual axon meets to form a bundle. This model can serve to direct further studies of Girdin (Ha, 2015).
Because of the similarities between fly sensory neurons and human photoreceptor cells, Girdin may have a similar function in the formation of rod and cone dendrites. Rod and cone photoreceptors are capable of surviving throughout a life span, yet they are sensitive to various insults that result in their degeneration, leading to gradual loss of visual acuity and blindness. Since RPE cells surround the tip of the photoreceptors and are essential for forming and maintaining the morphology of the photoreceptors, studying the role of Girdin in RPE cell lines may provide more insights into understanding how RPE cells regulate the morphology of photoreceptor cells. By better understanding the mechanisms of how ciliated a sensory neuron functions and forms its structure in Drosophila, it is hoped to assist in confronting retinal diseases and provide insight to treat them (Ha, 2015).
Girdin shares some similarities with NompA: both are essential for fly viability, coordinated behavior, and MRC, while both are not essential for bristle or socket formation and TEP. Both have a similar dendrite formation defect in which the tip of the sensory dendrite terminates at a significant distance from the bristle base and socket. Finally, both NompA and Girdin localize inside the sensory organ at the vicinity of the dendritic tip. Taking together, these phenotypes and localization suggest that Girdin is associated with the function of NompA to attach the dendritic tip to the base of the bristle for the mechanical transduction of mechanosensory signals. However, since Girdin is expressed only transiently in the sensory organ, it is likely to have only a developmental role in forming the mechanosensory organ rather than a direct role in mechanosensation as is proposed for NompA. Also, since girdin101 sensory neurons showed a more severe defect in dendrite morphology than nompA, Girdin might have additional functions beyond temporarily connecting the dendrite to the bristle shaft. One such additional function of Girdin may be to stabilize the dendrite as it differentiates (Ha, 2015).
Neurons exhibit extreme diversity in size, but whether large neurons have specialized mechanisms to support their growth is largely unknown. The SLC36 amino acid transporter Pathetic (Path) has been identified as a factor required for extreme dendrite growth in neurons. Path is broadly expressed, but only neurons with large dendrite arbors or small neurons that are forced to grow large require path for their growth. To gain insight into the basis of growth control by path, this study generated additional alleles of path and further examined the apparent specificity of growth defects in path mutants. Prior finding that loss of path function imposes an upper limit on neuron growth was conformed, and additionally it was found that path likely limits overall neurite length rather than dendrite length alone. Using a GFP knock-in allele of path, additional tissues were identified where path likely functions in nutrient sensing and possibly growth control. Finally, it was demonstrated that path regulates translational capacity in a cell type that does not normally require path for growth, suggesting that path may confer robustness on growth programs by buffering translational output. Altogether, these studies suggest that Path is a nutrient sensor with widespread function in Drosophila (Lin, 2016).
How growth programs are tailored to meet the demands of large neurons is largely unknown. This study demonstrates that the SLC36 transporter Path is required for growth of axons and dendrites in large neurons. Although the precise mechanism of action remains to be determined, the amino-terminal intracellular domain is essential for Path function in neuron growth control (Lin, 2015) and it is hypothesized that Path transduces nutrient signals via protein-protein interactions between its N-terminal intracellular domain and unidentified signaling proteins to modulate translational output . In light of prior studies suggesting a link between SLC36 transporters and TORC1 signaling and the observation that co-overexpression of Path and Rheb potentiates dendrite growth (Lin, 2015) it is proposed that Path may promote neuron growth, in part, through regulation of TORC1 activity. However, given the observations that Path primarily localizes to the plasma membrane and that mutations in TORC1 components cause more modest neuron growth deficits than mutation of path, it seems likely that Path engages downstream pathways other than TORC1 to promote neuron growth. Although the identity of these pathways is currently unknown, eukaryotic initiation factor 2 (eIF2) activity is regulated by a suite of kinases (eIF2K) that tune translation levels in response to environmental stresses, including amino acid deficiency (Lin, 2016).
Although Path is broadly expressed, path is dispensable for growth in most cell types under well-fed laboratory conditions. One prediction of this observation is that Path selectively affects translation in large neurons. The results from this study refute this notion; cells that do not require path for growth still require path for maximum translational output. How can the disparity in growth phenotypes between neurons and other cells explaned? First, Path may play a more significant role in translational control of neurons than other cell types. Drosophila has several uncharacterized genes that encode SLC36 transporters, and these gene products may function redundantly to control growth together with Path in some cell types. Consistent with this notion, overexpression of CG1139, which encodes another SLC36 transporter, drives ommatidial overgrowth in the eye (Goberdhan, 2005) Second, the residual translational capacity in path mutants is likely sufficient to support normal growth in many cell types, at least under certain conditions. Similarly, loss of Drosophila 4E-BP (Thor) function reduces overall translation without any obvious effect on animal growth in well-fed conditions. Intriguingly, environmental stresses such as hypoxia or starvation uncover a requirement for Thor in growth control. Similarly, it is predicted that stresses that compromise translational efficiency should sensitize cells to a requirement for path function, therefore an exciting future direction will be to determine whether path exerts state-dependent functions on growth control (Lin, 2016).
Neurons typically assume multipolar, bipolar, or unipolar morphologies. Little is known about the mechanisms underlying the development of these basic morphological types. This study shows that the Kruppel-like transcription factor Dar1 determines the multipolar morphology of postmitotic neurons in Drosophila. Dar1 is specifically expressed in multipolar neurons and loss of dar1 gradually converts multipolar neurons into the bipolar or unipolar morphology without changing neuronal identity. Conversely, misexpression of Dar1 or its mammalian homolog in unipolar and bipolar neurons causes them to assume multipolar morphologies. Dar1 regulates the expression of several dynein genes and nuclear distribution protein C (nudC), which is an essential component of a specialized dynein complex that positions the nucleus in a cell. These genes were shown to be required for Dar1-induced multipolar neuron morphology. Dar1 likely functions as a terminal selector gene for the basic layout of neuron morphology by regulating both dendrite extension and the dendrite-nucleus coupling (Wang, 2015).
Ramon y Cajal placed neurons into three major morphological types based on the number of dendrites connected to the soma (i.e., primary dendrites): unipolar, bipolar, and multipolar and this classification system is universally applicable to different species throughout evolution. Multipolar neurons, like mammalian pyramidal neurons, develop more than one primary dendrite. In contrast, bipolar neurons are defined as having a single primary dendrite that may (e.g., cerebellar Purkinje cells) or may not (e.g., photoreceptors) branch out into an elaborate dendritic arbor. Finally, unipolar neurons such as DRG neurons in vertebrates and the majority of CNS neurons in invertebrates extend a single primary neurite, which usually bifurcates into dendritic and axonal branches (Wang, 2015).
Multipolar morphology separates the dendritic arbor into distinct fields around the soma, which has an impact, not only on the passive current spread and processing of electrical signals in the neuron), but also on the types of synaptic or sensory inputs that the neuron receives . In addition, the three basic morphologies of neurons are relevant to the distinct organizational principles used in both the nervous systems of different animal species and in different parts of a single nervous system. Although all three morphological types are found in different species throughout evolution, the majority of neurons in invertebrates are unipolar, whereas the majority of those in vertebrates are multipolar (Wang, 2015).
In the insect CNS, unipolar neurons extend a single process from the soma to a synapse-enriched neuropil and then bifurcate into dendrites that arborize locally and an axon that typically projects to other neuropil areas or target tissues. Unipolar organization of neuronal processes allows the formation of synaptic connections away from the location of the neuronal cell body, so it is likely an alternative strategy for neuronal migration, which is rare in the insect CNS but common in the vertebrate CNS. Despite the importance of these fundamental organizations of neuronal processes, very little progress has been made toward understanding the molecular and cellular programs that lead postmitotic neurons to develop multipolar, bipolar, or unipolar morphologies since their description a century ago (Wang, 2015).
This study shows that the transcription factor Dar1 determines the multipolar morphology of postmitotic neurons in Drosophila. Dar1 is selectively expressed in postmitotic multipolar neurons and is required for these neurons to assume the multipolar morphology. Ectopic expression in unipolar or bipolar neurons leads to multipolar morphology. Dar1 regulates the expression of several dynein genes and nudC, which is an essential component of a specialized dynein complex that positions the nucleus in a cell. It is further shown that this evolutionarily conserved complex is required for multipolar morphology of neurons. These results suggest that dar1 likely functions as a terminal selector gene for the basic layout of neuron morphology (Wang, 2015).
The universal morphological organization of neuronal dendrites and axons-in the form of the unipolar, bipolar, and multipolar morphologies-is important for information processing in neurons and for the wiring of neural circuits. It has generally been assumed that the formation of the basic morphological types of neurons is determined by the number of dendrites growing out from the cell body (the 'outgrowth model'). This study shows that this model alone is insufficient to explain the formation of multipolar morphology. Nuclear positioning is introduced as a factor in determining the multipolar neuron morphology, and it is proposed that Dar1 determines multipolar morphology by regulating both dendrite extension and primary dendrite-nucleus coupling (Wang, 2015).
A novel, instructive role is reported for Dar1 in determining the multipolar morphology of postmitotic neurons without changing cell fate. First, despite the dendritic defects, axon morphology (Ye, 2011) and targeting are unchanged in dar1 mutant neurons. Second, ectopic expression of Dar1 in postmitotic neurons leads to supernumerary primary dendrites. Third, the remaining dendrites in dar1-/- neurons still follow the branching pattern assumed by wild-type neurons (Ye, 2011). Fourth, dar1 mutations do not affect the expression of neuron type-specific markers. Based on extensive studies in C. elegans, Hobert proposed the concept of terminal selector genes (Hobert, 2008). A terminal selector gene is required for determining specific aspects of a neuron's identity by regulating the expression of genes responsible for these characteristics such as those encoding neurotransmitter receptors, enzymes in a neurotransmitter synthesis pathway, and structural proteins. Loss of a terminal selector gene results in the loss of a specific aspect of the neuron type without affecting the overall neuronal identity. Dar1 plays such a function in the basic layout of neuronal morphology and thus is likely a 'terminal selector gene' for neuronal morphology (Wang, 2015).
Based on the findings in this study, it is proposed that generating neuronal multipolar morphology requires, not only dendritic extension, but also a coupling mechanism between the nucleus and the dendrites. Dar1 promotes both dendritic growth and dendrite-nucleus coupling. Therefore, its misexpression converts unipolar neurons into neurons with multipolar morphology. The results presented in this study raise the interesting possibility that a specialized dynein complex in multipolar neurons with components that are transcriptionally regulated by Dar1 couples the nucleus with the primary dendrites. If the primary dendrite-nucleus coupling is weakened, then the nucleus may move to a different location in relation to the dendrites and axons. It is speculated that there might also be an active or passive force that pulls the nucleus toward the axon, opposing the force that couples the primary dendrites and the nucleus. Consistent with this model, the remaining single primary dendrites of all bipolar-shaped da neurons-caused by loss of dar1, reduced functions of dynein, or nuclear positioning complex-are those that project in the direction opposite the axon. This observation again rules out the possibility that the reduction in number of primary dendrites of dar1-/- da neurons is the result of reduction in dendrite growth. If that were the case, then the remaining single primary dendrites would likely project in random directions and not solely away from the axon. Further studies are needed to determine the cellular and molecular basis of the dendrite-nucleus coupling (Wang, 2015).
Several prior studies have demonstrated that neurons switch between different morphological types during development. These observations suggest that the acquisition of basic morphological types in many neurons includes intermediate morphologies with nucleus-primary dendrite relationships that are different from those seen in the mature neurons. It will be interesting to investigate whether the activity of the nuclear positioning complex and its regulators play a role in these developmental changes in morphological type (Wang, 2015).
In summary, this study offers a novel model for understanding the establishment of the three basic morphological types of neurons. Starting from genetic analysis of the KLF transcription factor Dar1, the study not only uncovers an instructive factor that determines the multipolar morphology of neurons, but also provide a mechanistic model showing that the position of the nucleus is critical for establishing multipolar neuron morphology. This study also demonstrates that the basic morphological types are determined by intrinsic molecular mechanisms in postmitotic neurons rather than in precursor cells. The model presented in this study may also be applied to explaining the changes in basic morphological type during neuron development. This study therefore opens the door for a unifying theory of basic structural organization in neurons (Wang, 2015).
During development, certain Drosophila sensory neurons undergo dendrite pruning that selectively eliminates their dendrites but leaves the axons intact. How these neurons regulate pruning activity in the dendrites remains unknown. This study identifies a coiled-coil protein Spindle-F (Spn-F) that is required for dendrite pruning in Drosophila sensory neurons. Spn-F acts downstream of IKK-related kinase Ik2 in the same pathway for dendrite pruning. Spn-F exhibits a punctate pattern in larval neurons, whereas these Spn-F puncta become redistributed in pupal neurons, a step that is essential for dendrite pruning. The redistribution of Spn-F from puncta in pupal neurons requires the phosphorylation of Spn-F by Ik2 kinase to decrease Spn-F self-association, and depends on the function of microtubule motor dynein complex. Spn-F is a key component to link Ik2 kinase to dynein motor complex, and the formation of Ik2/Spn-F/dynein complex is critical for Spn-F redistribution and for dendrite pruning. These findings reveal a novel regulatory mechanism for dendrite pruning achieved by temporal activation of Ik2 kinase and dynein-mediated redistribution of Ik2/Spn-F complex in neurons (Lin, 2015)
The precise assembly of neural circuits is crucial for the nervous system to function properly. The developing nervous systems often start with a primitive prototype, characterized by exuberant branches and excessive connections. Thus, further remodeling is required to refine the developing nervous systems to maturity. Neuronal pruning, one such remodeling mechanism, is a highly regulated self-destruct process that eliminates excessive neuronal branches in the absence of cell death. Pruning is widely observed in the nervous systems of both vertebrates and invertebrates, that not only ensures precise wiring during development, but also allows for adjustment of neuronal connections in response to injury and disease. Various studies have shown that defects in developmental pruning affect the function of the nervous systems in C. elegans and Drosophila. Moreover, a progressive loss of neurites far ahead of cell death is commonly observed in many neurodegenerative disorders. Thus, any dysregulation of pruning activity even at the level of individual neurons would bring catastrophic consequences to the nervous systems. Although the primary triggers for developmental pruning and pruning that ensues upon neuronal injury and disease are diverse, the downstream machinery that eliminates neuronal processes shared some common features. For example, microtubule disruption is the earliest cellular event observed in all types of pruning, and the ubiquitin-proteasome system is required in all circumstances (Lin, 2015)
During Drosophila metamorphosis, substantial neuronal remodeling takes place in both the central and peripheral nervous systems. Most of the larval peripheral neurons die during metamorphosis, whereas few, including some class IV dendritic arborization (C4da) neurons, survive and undergo large-scale dendrite pruning. Dendrite pruning of the dorsal C4da neuron ddaC starts with severing of the proximal dendrites at 4-6 h APF (after puparium formation). Subsequently these disconnected dendrites become fragmented and eventually eliminated by the surrounding epidermal cells by 16-18 h APF. In contrast to the central brain mushroom body (MB) gamma neurons where both larval dendrites and axons are pruned during development, the peripheral C4da neurons specifically prune their dendrites keeping the axons intact. The molecular basis for how the pruning activity is confined to the dendrites of C4da neurons remains unknown. It was reasoned that molecular differences between dendrites and axons should be considered for such differential pruning activity in C4da neurons. It is known that microtubule polarity is different in the dendrites and axons of neurons, including in the Drosophila sensory neurons. For example, C4da neurons have polarized microtubules in their proximal dendrites predominantly with microtubule minus end pointing away from the cell body, but have an opposite polarity in their axons. This difference in microtubule polarity is essential for maintaining the proper function and compartmental identities of dendrites and axons, and might be an important determinant for spatially restricting pruning activity in the dendritic compartments of C4da neurons. Based on this assumption, some molecules are required to connect the pruning activity with the distinctive microtubule polarity of the dendrites in C4da neurons during dendrite pruning (Lin, 2015)
Previous studies have shown that dendrite pruning in C4da neurons is initiated by the steroid hormone ecdysone and its heterodimeric receptors, ecdysone receptor B1 (EcR-B1) and Ultraspiracle (Usp). Through transcriptional regulation of sox14, ecdysone signaling activates the Sox14 target gene mical, which encodes a cytoskeletal regulator, to regulate dendrite pruning. A few other molecules mediating specific cellular activities have been shown to participate in dendrite pruning of C4da neurons, such as the ubiquitin-proteasome system, caspases, matrix metalloproteases, microtubule severing proteins and mediators of dendritic calcium transients. Previous studies identified Ik2 kinase, a homologue of vertebrate IKK-ε in Drosophila, that plays an essential role in dendrite pruning of pupal neurons, and further demonstrated that Ik2 is sufficient to induce precocious dendrite severing in larval neurons. Ik2 is the only known molecule sufficient to induce premature dendrite severing in larvae, reflecting a central role of Ik2 kinase in dendrite pruning. Therefore, this study aimed to elucidate the mechanism by which Ik2 kinase signaling is transduced and regulated in Drosophila sensory neurons during dendrite pruning (Lin, 2015)
To elucidate the mechanism of Ik2 kinase signaling, candidate molecules were sought that mediate Ik2 signals during dendrite pruning. Several lines of evidence suggested that Spn-F, a coil-coiled protein, is a good candidate. Firstly, spn-F mutant flies showed defects in developing oocytes and bristles (Abdu, 2006), similar to the phenotypes observed in ik2 mutants (Shapiro, 2006). Secondly, Spn-F physically interacts with Ik2 (Dubin-bar, 2008). It implied that ik2 and spn-F may act in the same pathway during oogenesis and bristle morphogenesis, and the possibility has been raised that a similar pathway might also be involved in dendrite pruning of C4da neurons. This study demonstrates Spn-F plays a key role in linking Ik2 kinase to microtubule motor dynein complex for dendrite pruning. Spn-F acts downstream of Ik2 kinase in the same pathway for dendrite pruning. Spn-F is shown to displays a punctate pattern in larval neurons and these Spn-F puncta become dispersed in pupal cells. The redistribution of Spn-F from puncta is essential for dendrite pruning, and depends on the activity of Ik2 kinase and the function of microtubule motor dynein complex. These data also demonstrate that Spn-F not only links Ik2 to dynein motor complex, but also mediates the formation of Ik2/Spn-F/dynein complex, that is critical for Spn-F punctum disassembly and dendrite pruning (Lin, 2015)
In addition to apoptosis, neurons have a second self-destruct program in their axons for axonal pruning during development and in response to neuronal injury and disorders. This study proposes a third self-destruct program, which is mediated by Ik2 kinase activity in Drosophila sensory neurons, specific for dendrite pruning. Ik2 is essential for dendrite severing in pupal C4da neurons (Lee, 2009), and currently is the only known molecule sufficient to cause precocious dendrite severing in larval cells (Lee, 2009), indicating that Ik2 activation must be regulated temporally. For temporal regulation, ecdysone signaling plays a key role in dendrite pruning (Williams, 2005; Lee, 2009). These studies show that no Ik2 activation is detected in pupal C4da neurons with impaired ecdysone signaling and thus places Ik2 kinase downstream of ecdysone signaling. Microarray studies have identified ik2 as one of the ecdysone/EcR up-regulated genes in brain MB γ neurons during axon pruning (Hoopfer, 2008). This suggests one possible mechanism where ecdysone/EcR regulates Ik2 activation through increasing ik2 expression in C4da neurons. Although Ik2 kinase activity is crucial for oogenesis and bristle morphogenesis (Shapiro, 2006; Otani, 2011), the activation mechanisms of Ik2 kinase in both processes remain unknown. Since pruning activity is considered as a self-destruct program, how to regulate this activity spatially in subcellular compartments within individual neurons is an intriguing issue to investigate. This study identifies Spn-F and cytoplasmic dynein complex as critical regulators of Ik2-mediated dendrite pruning activity in C4da neurons (Lin, 2015)
It is known that endogenous Spn-F exhibits a punctate pattern in nurse cells (Abdu, 2006), consistent with the observation of punctate Spn-F-GFP in larval C4da neurons. The formation of Spn-F puncta in cells is through self-association, and does not depend on the integrity of microtubule network or the function of cytoplasmic dynein. Since Ik2 could form oligomers in cells, the interaction between Ik2 and Spn-F might also play a role in Spn-F puncta formation. Indeed, it was observed that SpnF-ΔCC3-GFP has normal interaction with either SpnF-ΔCC3 or full-length Spn-F, but formed fewer puncta than the wild type Spn-F-GFP did in larval neurons. Therefore, the Spn-F puncta formation could be attributed not only to Spn-F self-association, but also to Ik2/Spn-F interaction and Ik2 oligomerization (Lin, 2015)
In larval C4da neurons, Ik2 kinase is inactive and associates with Spn-F as puncta in the cytosol. After puparium formation, Ik2 kinase becomes activated promptly and phosphorylates Spn-F in C4da neurons. This Ik2-dependent phosphorylation on Spn-F decreases Spn-F self-association, and subsequently the numbers and sizes of Spn-F puncta were reduced. One may question that protein degradation might contribute to decrease the numbers and sizes of Spn-F puncta in C4da neurons during dendrite pruning. It was known that Ik2 promotes caspase inhibitor DIAP1 degradation via proteasomes during the development of sensory organ precursors; therefore, Ik2 might promote Spn-F degradation in C4da neurons during dendrite pruning. However, Ik2 overexpression does not alter the protein level of Spn-F in either S2 or germline cells (Dubin-Bar, 2008). Thus, protein degradation by proteasomes is unlikely the mechanism leading to decreased Spn-F puncta after Ik2 activation. Since P-Ik2 signals were indistinguishable between wild-type and Dhc64C RNAi neurons, it is reasonable to presume that both Ik2 activation and Spn-F phosphorylation occur normally in dynein mutant neurons. No significant differences were found between the pruning defects of C4da neurons in spn-F mutants and that in spn-F mutants with Dhc-RNAi, and between the pruning phenotypes observed in Dhc mutants and that in Dhc mutants with ik2-RNAi. These findings further support that Ik2, Spn-F and dynein complex function together in the same pathway in dendrite pruning of C4da neurons. However, the finding of Spn-F puncta in mutant pupal neurons with impaired dynein function indicated that dynein is required for Spn-F redistribution after Ik2 activation. Furthermore, Spn-F remains punctate in S2 cells with Ik2 overexpression even after microtubule depolymerization and inhibition of dynein function, suggesting that dynein might redistribute Ik2/Spn-F complexes via transporting complexes toward the minus ends of microtubules in C4da neurons during dendrite pruning. The results in this study and studies in germline cells, (the fact that more Spn-F puncta accumulated in nurse cells with colchicine treatment and with Dhc mutation) (Abdu, 2006), favor the mechanism of protein redistribution for Spn-F punctum reduction in dendrite pruning of C4da neurons (Lin, 2015)
It has been shown that during Drosophila bristle elongation, directional transport of activated Ik2 and of Spn-F to the bristle tips, where the microtubule minus ends are concentrated, requires the function of cytoplasmic dynein, and Spn-F acts as an adaptor to link Ik2 to dynein complexes. These are similar to the current findings that both Ik2 activation and dynein complex are essential for Spn-F redistribution, and Spn-F plays a central role in the formation of Ik2/Spn-F/dynein complex, which is crucial for Spn-F redistribution and for dendrite pruning in C4da neurons. However, the studies in bristle elongation indicating that spn-F acts upstream of ik2 (Otani, 2015) disagree with the current finding that ik2 acts upstream of spn-F in dendrite pruning. The discrepancy between the epistasis analyses of ik2 and spn-F in these two different processes might be due to different cell-type specific factors in these two types of cells that affect the morphological readouts in genetic studies. Moreover, this study demonstrated that Ik2-dependent phosphorylation of Spn-F decreases Spn-F self-association, promotes Spn-F redistribution, and finally leads to dendrite pruning in C4da neurons (Lin, 2015)
The activated Ik2 signals accumulate at the microtubule minus ends in cells with polarized microtubule distribution, such as oocytes, follicle cells and bristles. This is consistent with the conclusion that dynein transports activated Ik2 toward microtubule minus ends in C4da neurons. Since Drosophila sensory neurons have polarized microtubules in their proximal dendrites predominantly with microtubule minus end pointing away from the cell body, these studies revealed a possible mechanism that Spn-F and minus-end directed motor dynein complex confine Ik2-dependent pruning activity to the somatodendritic compartments of C4da neurons. During Drosophila bristle elongation, the accumulation of endogenous Spn-F observed at the bristle tip, where the microtubule minus ends are enriched, led to an examination of Spn-F-GFP signals along the dendrites of C4da neurons during dendrite pruning. However, no enriched of Spn-F-GFP signals in the proximal dendrites, where dendrite severing is expected to occur, was observed, by live imaging during pruning. Previous studies (Lee, 2009) showed that microtubules are first disassembled in the proximal dendrites of C4da neurons during dendrite severing. This local disassembly of microtubules is suppressed in ik2 mutant neurons. Since the current genetic studies indicate that both ik2 and spn-F act in the same pathway of dendrite pruning, tests were performed to see whether local microtubule disassembly happens normally in spn-F mutants. Local breakage of microtubules found in the proximal dendrites of C4da neurons was also suppressed in spn-F RNAi mutants, suggesting that Spn-F, like Ik2, plays a role in dendrite severing that involves local microtubule disassembly. However, the molecular mechanisms by which activated Ik2 and Spn-F lead to dendrite severing in the proximal dendrites of C4da neurons will be an important question for future studies (Lin, 2015)
It is known that there is no decrease in cell death in wing discs with ik2 knockdown and in ik2 mutant embryos (Kuranaga, 2006), indicating that the primary function of Ik2 is not involved in the apoptotic pathway during development. However, ectopic Ik2 activation by overexpression leads to cell death in fly compound eyes (Kuranaga, 2006) and in C4da neurons (Lee, 2009), suggesting that excessive Ik2 kinase signaling could trigger a crosstalk with signaling molecules in apoptotic pathways and result in apoptosis. It is known that Ik2 kinase regulates the nonapoptotic function of caspase through promoting DIAP1 degradation (Kuranaga, 2006). In a similar manner, the confinement of activated Ik2 kinase in the dendritic compartments might restrict the detected caspase activity in the degenerating dendrites after separating from the soma of C4da neurons during dendrite pruning. Therefore, this raises a possibility that de-regulation of pruning activity in neurons may trigger a crosstalk with molecules in apoptotic pathways and lead to undesired cell death during neuronal injury and disorders. Recently, a caspase cascade, including caspase 3 and 6, was identified in mice to play a role in developmental axon pruning and in sensory axon pruning after trophic factor withdrawal. Moreover, activated caspase 6 was detected in human patient brains of Alzheimer and Huntington diseases long before cell death, highlighting a critical role in regulating caspase activity in both diseases. Understanding the regulatory mechanisms that confine pruning activity into proper subcellular compartments of the neuron might provide molecular insights into the pathogenesis of neural disorders (Lin, 2015)
In Drosophila neurons, kinesin-2, EB1 and Apc are required to maintain minus-end-out dendrite microtubule polarity, and it has been proposed they steer microtubules at branch points. Motor-mediated steering of microtubule plus ends could be accomplished in two ways: 1) by linking a growing microtubule tip to the side of an adjacent microtubule as it navigates the branch point (bundling), or 2) by directing a growing microtubule after a collision with a stable microtubule (collision resolution). Using live imaging to distinguish between these two mechanisms, this study found that reduction of kinesin-2 did not alter the number of microtubules that grew along the edge of the branch points where stable microtubules are found. However, reduction of kinesin-2 or Apc did affect the number of microtubules that slowed down or depolymerized as they encountered the side of the branch opposite to the entry point. These results are consistent with kinesin-2 functioning with Apc to resolve collisions. However, they do not pinpoint stable microtubules as the collision partner as stable microtubules are typically very close to the membrane. To determine whether growing microtubules were steered along stable ones after a collision, the behavior was analyzed of growing microtubules at dendrite crossroads where stable microtubules run through the middle of the branch point. In control neurons, microtubules turned in the middle of the crossroads. However, when kinesin-2 was reduced some microtubules grew straight through the branch point and failed to turn. It is proposed that kinesin-2 functions to steer growing microtubules along stable ones following collisions (Weiner, 2016).
In many differentiated cells, microtubules are organized into polarized noncentrosomal arrays, yet few mechanisms that control these arrays have been identified. For example, mechanisms that maintain microtubule polarity in the face of constant remodeling by dynamic instability are not known. Drosophila neurons contain uniform-polarity minus-end-out microtubules in dendrites, which are often highly branched. Because undirected microtubule growth through dendrite branch points jeopardizes uniform microtubule polarity, this study used this system to understand how cells can maintain dynamic arrays of polarized microtubules. Growing microtubules navigate dendrite branch points by turning the same way, toward the cell body, 98% of the time, and growing microtubules track along stable microtubules toward their plus ends. Using RNAi and genetic approaches, this study shows that kinesin-2, and the +TIPS EB1 and APC, are required for uniform dendrite microtubule polarity. Moreover, the protein-protein interactions and localization of Apc2-GFP and Apc-RFP to branch points suggests that these proteins work together at dendrite branches. The functional importance of this polarity mechanism is demonstrated by the failure of neurons with reduced kinesin-2 to regenerate an axon from a dendrite. It is concluded that microtubule growth is directed at dendrite branch points and that kinesin-2, APC, and EB1 are likely to play a role in this process. It is propose that Kinesin-2 is recruited to growing microtubules by +TIPS and that the motor protein steers growing microtubules at branch points. This represents a newly discovered mechanism for maintaining polarized arrays of microtubules (Mattie, 2010).
Most cells in multicellular organisms contain polarized noncentrosomal microtubule arrays. In interphase mammalian cultured cells, microtubules are nucleated at the centrosomal microtubule organizing center (MTOC), and plus ends grow towards the cell periphery. However, in many differentiated cells, minus ends are not focused at a centrosomal MTOC. In epithelial cells, a major population of microtubules has minus ends focused at the apical side and plus ends at the basal side. In muscle cells, minus ends spread out around the nuclear envelope, and neurons have perhaps the simplest and most strikingly polarized noncentrosomal microtubule arrays. The mechanisms that organize these noncentrosomal microtubule arrays are poorly understood (Mattie, 2010).
Neurons have two types of processes that extend from the cell body: axons and dendrites. Dendrites primarily receive signals from other neurons or the environment, and axons send signals to other neurons or output cells. One basic difference between axons and dendrites is the arrangement of microtubules. In axons microtubules are arranged into an overlapping array of uniform polarity plus-end-out microtubules. In dendrites of cultured mammalian neurons microtubules have mixed orientation near the cell body. In dendrites of Drosophila neurons 90%-95% of microtubules have minus ends distal to the cell body. As the dendritic array in Drosophila is very simple, and extremely different from a centrosomal array, This study used it as a model system to identify mechanisms that organize polarized noncentrosomal microtubules (Mattie, 2010).
It is not known how uniform dendrite microtubule polarity is established or maintained. Models for generating the plus-end-out axonal microtubule array focus on sliding of microtubules by motor proteins. In mammalian neurons, microtubules are thought to be nucleated in the cell body at the centrosome, then released from the centrosome and transported down the axon in the correct orientation by motors including dynein. Models to account for mixed orientation microtubules in dendrites of cultured neurons have also been proposed. In this case, the kinesin MKLP1 (Kif23) has been proposed to transport minus-end-out microtubules into dendrites along plus-end-out microtubules. The current study identified a new mechanism that is required for uniform microtubule polarity in dendrites (see Interactions between kinesin-2 and +TIPs, and localization of Apc2-GFP to dendrite branch points). As it uses conserved, generally expressed proteins, it could play a role in maintaining microtubule polarity in many other cell types (Mattie, 2010).
Using Drosophila dendrites as a model system, this study demonstrates that growing microtubule plus ends almost always turn towards the cell body at branch points, and that they track stable microtubules through branches. Kinesin-2, EB1 and APC are all required to maintain microtubule polarity and are linked in an interaction network. Based on these results, a model is proposed for directed growth of microtubules in dendrites. Apc2 most likely contains a branch localization signal because Apc2-GFP localizes well to dendrite branches even when overexpressed. Localization of Apc2 to dendrite branch points can recruit Apc to the branch. Apc can interact with the Kap3 subunit of kinesin-2, and so could increase concentration of the motor near the branch. EB1-GFP does not concentrate at branches, so it is proposed that a growing microtubule plus end coated with EB1 is transiently linked to kinesin-2 as it passes through the branch, through the interaction between Apc and the EB1 tail. SxIP motifs in Apc and Klp68D could also contribute to this interaction. As both Kap3 and the SxIP motif in Klp68D are in the kinesin-2 tail, the motor domain would be free to walk along a nearby stable microtubule towards the plus end and cell body (Mattie, 2010).
Even a very brief application of force pulling the growing microtubule towards the cell body should be sufficient to steer growth towards the cell body. Once the tip of the microtubule turns, growth would be constrained by the dendrite walls. The association of the growing plus end with stable microtubule would probably only need to be maintained over a distance of a micron. This model is consistent with the observations that kinesin-2 has shorter run lengths than kinesin-1, and that individual EB1 interactions with the microtubule plus end persist less than a second (Mattie, 2010).
Observations of plus end behavior in vivo also favor a model in which only transient interactions of the motor and microtubule plus end are involved, because plus ends turning sharply are frequently seen. Stable microtubules do not accommodate such sharp turns, so the sharp turns of plus ends are not likely to occur while the plus end is tracking along a stable microtubule. Instead they likely represent a switch from a freely growing plus end to one that is following a track (Mattie, 2010).
Directed growth of microtubules at dendrite branch points allows dendrites to maintain uniform minus-end-out polarity despite continued microtubule remodeling. This mechanism is also likely necessary to establish uniform microtubule polarity in branched dendrites, but probably cannot account for minus-end-out polarity on its own. It is hypothesized that directed microtubule growth is used in concert with an unidentified mechanism to control microtubule polarity in dendrites. Transport of oriented microtubule pieces has been proposed to contribute to axon microtubule polarity and a similar mechanism could play a role in dendrites. For example, a kinesin anchored to the cell cortex by its C-terminus could shuttle minus-end-out microtubule seeds into dendrites. Alternately, polarized microtubule nucleation sites could be localized within dendrites. Identifying directed growth as one mechanism that contributes to dendrite microtubule polarity should facilitate identification of the missing pieces of this puzzle (Mattie, 2010).
Because kinesin-2, APC, EB1 and polarized microtubules are found in many cell types, directed growth of microtubules along stable microtubule tracks could be very broadly used for maintaining microtubule polarity. This is a newly identified function for both the +TIP proteins and kinesin-2. APC has previously been localized to junctions between a microtubule tip and the side of another microtubule at the cortex of epithelial cells, and sliding of microtubule tips along the side of microtubules was also seen in this system, but there was no association with a particular direction of movement or overall microtubule polarity (Mattie, 2010).
As kinesin-2 has previously been shown to be enriched in tips of growing axons in cultured mammalian neurons, as has APC, it is possible that they could mediate tracking of growing microtubules along existing microtubule tracks in the growth cone. In fact, this type of microtubule tracking behavior has been observed in axonal growth cones under low actin conditions. Thus directed growth of microtubules could also be important during axon outgrowth. Indeed, the same principle of directed growth by a motor protein connected to a growing microtubule plus end could be used to align microtubules in many circumstances (Mattie, 2010).
Pruning that selectively removes unnecessary neurites without causing neuronal death is essential for sculpting the mature nervous system during development. In Drosophila, ddaC sensory neurons specifically prune their larval dendrites with intact axons during metamorphosis. However, it remains unknown about an important role of ER-to-Golgi transport in dendrite pruning. In a clonal screen this study identified Yif1, an uncharacterized Drosophila homologue of Yif1p that is known as a regulator of ER-to-Golgi transport in yeast. This study shows that Yif1 is required for dendrite pruning of ddaC neurons but not for apoptosis of ddaF neurons. The Yif1-binding partner Yip1 was identified that is also crucial for dendrite pruning. Yif1 forms a protein complex with Yip1 in S2 cells and ddaC neurons. Yip1 and Yif1 colocalize on ER/Golgi and are required for the integrity of Golgi apparatus and outposts. Moreover, two GTPases Rab1 and Sar1, known to regulate ER-to-Golgi transport, were shown to be essential for dendrite pruning of ddaC neurons. Finally, these data reveal that ER-to-Golgi transport promotes endocytosis and downregulation of cell adhesion molecule Neuroglian and thereby dendrite pruning (Wang, 2018).
The secretory pathway involves three elementary organelles: the endoplasmic reticulum (ER), the Golgi apparatus and the trans-Golgi network (TGN). Via the secretory pathway, protein and lipid supplies are provided to facilitate the specification and outgrowth of dendrites and axons during neuronal development. In hippocampal neurons, the disruption of the secretory pathway through suppressing the post-Golgi trafficking or normal Arf1 function leads to a dendrite-outgrowth defect. Disruption of ER-to-Golgi transport by inhibiting Sar1 activity leads to shortened axons in mammalian neurons. In Drosophila, the disruption of ER-to-Golgi transport by mutating Sar1 or Rab1 dramatically inhibits dendrite arbor elaboration with normal axonal elongation in sensory neurons (Ye, 2007). In addition to facilitating dendrites/axon outgrowth, the secretory pathway is also essential for the maintenance of dendrite arbors after neuron maturation. In stark contrast to its roles in neuronal growth and maintenance, a recent study has reported that Arf1/Sec71-mediated post-Golgi trafficking process is also crucial for dendrite pruning: a regressive event (Wang, 2017). However, a potential role for ER-to-Golgi transport in dendrite pruning remained elusive (Wang, 2018).
This study identified two novel genes, Yif1 and Yip1, that play important roles in dendrite pruning in ddaC sensory neurons during early metamorphosis. Their respective homologs are known to regulate ER-to-Golgi transport in yeast and mammals. Yif1 is shown to associates with Yip1 in S2 cells and ddaC neurons. Moreover, Yip1 and Yif1 colocalize on the ER and cis-Golgi, and both are required for the integrity of the Golgi apparatus and outposts. The small GTPases Rab1 and Sar1, two key regulators of ER-to-Golgi transport, were identified as crucial for dendrite pruning of ddaC neurons. Importantly, the data indicate that the ER-to-Golgi transport promotes dendrite pruning partly via endocytosis and downregulation of the cell-adhesion molecule Neuroglian (Nrg). Thus, these data argue that some yet to be-identified molecules might be secreted into the dendrites to trigger Nrg internalization and dendrite pruning (Wang, 2018).
Drosophila Yif1 and Yip1 belong to the same protein family as yeast Yif1p/Yip1p. Yeast Yip1p was first identified as a Ypt1p/Rab1-interacting protein and Yif1p was further discovered as a Yif1p-interacting factor. Via their C-terminal regions, Yif1p and Yip1p form a heteromeric integral membrane complex. These two proteins localize on the Golgi membranes and regulate ER-to-Golgi protein trafficking and secretion. Yip1p-Yif1p was also reported to regulate the fusion process between ER-derived vesicles and the Golgi apparatus. This complex was proposed to serve as a receptor on the Golgi membranes for vesicle docking and fusion. In mammals, Yif1B was discovered as a binding partner of the 5-HT1A serotonin receptor and localized on intermediate compartments of the Golgi. Partial knockdown of Yif1B specifically disturbs the targeting of the 5-HT1A receptor to the distal dendrites, whereas complete knockout of Yif1B leads to the disruption of Golgi integrity. Likewise, mammalian Yip1 proteins were also discovered as trafficking regulators between ER exit sites, intermediate compartment and cis-Golgi (Wang, 2018).
This study shows that Drosophila Yif1 and Yip form a protein complex in vivo and are functionally relevant during dendrite pruning. First, these two proteins associate in both S2 cells and ddaC sensory neurons, as revealed in co-IP experiments and BiFC assays. Second, they colocalize on ER and cis-Golgi, and importantly their localizations are mutually dependent. Third, both of them are required for the integrity of Golgi apparatus, similar to the COPII-mediated ER-to-Golgi trafficking regulators, such as Rab1 or Sar1. Finally, removal of either of them caused the same phenotypes in terms of dendrite pruning, identical to those observed in Rab1 or Sar1 mutant neurons. Thus, it is plausible to suggest that Drosophila Yif1 and Yip1, like their homologs, can regulate the ER-to-Golgi transport to promote dendrite pruning. Consistent with these findings, previous genome-wide RNAi screens reported Yif1 as a potential component of the secretory pathway that might be involved in neural outgrowth and morphology (Wang, 2018).
Growing evidence indicates that the secretory pathway has been shown to regulate dendrite growth and maintenance in both Drosophila and mammals. The ER-to-Golgi transport is an early step of the canonical secretory pathway. Suppression of the ER-Golgi transport inhibits dendrite outgrowth in the developing neurons. Either disrupting Golgi apparatus or attenuating Golgi outposts alters dendrite morphology and outgrowth in mature neurons. A previous study illustrated that the small GTPas Arf1, which is known to regulate post-Golgi trafficking in mammals, is important for proper dendrite pruning of ddaC sensory neurons. Moreover, this present study describes Yif1 and Yip1 as two new regulators of dendrite pruning in sensory neurons. Yif1 and Yip1 appear to localize on ER and cis-Golgi, which is compatible with their roles in the ER-to-Golgi protein transport like their yeast and mammalian homolog. Yif1 and Yip1 can facilitate the biogenesis of secretory vesicles and affect the proper Golgi structure in ddaC neurons. Consistently, Rab1 and Sar1, which are two key components of the COPII vesicles, are also essential for dendrite pruning. Thus, it is tempting to hypothesize that some yet to be-identified molecules might be specifically secreted into the dendrites to promote dendrite pruning. Interestingly, mammalian Yif1B functions as a scaffold protein to recruit the 5-HT1A receptor together with Yip1A and Rab6, probably in the same trafficking vesicles and specifically targets the receptor to the distal dendrites in rat neurons. However, the current results suggest that this machinery might not be required for dendrite pruning of ddaC neurons. First, although the N-terminal 50 amino acid region of mammalian Yif1B is required for targeting the 5-HT1A receptor, this study found that a similar N-terminal fragment of Yif1 is dispensable for dendrite pruning because the N-terminal deleted Yif1 transgenes completely rescued the pruning defects in Yif110-46 mutant neurons. In addition, the ddaC neurons underwent dendrite pruning normally in mutant ddaC neurons derived from a null Rab6 mutant or RNAi expression. Future work would focus on the identification of secreted molecules that trigger dendrite pruning (Wang, 2018).
This study shows that ER-Golgi transport facilitates Nrg endocytosis prior to dendrite pruning at prepupal stage. It is also possible that ER-Golgi transport directly regulates the secretion of Nrg towards the plasma membrane at larval stages. Because of a lack of an anti-Nrg antibody against its extracellular domain, this possibility could not be examined. To bypass its early role in protein secretion, Rab1DN or Sar1DN expression was induced to temporally inhibit the EG-to-Golgi transport at late larval stage, when Nrg exocytosis was completed under normal ER-to-Golgi transport. Multiple lines of evidence were provided indicating that the ER-Golgi transport facilitates Nrg endocytosis and downregulation before the onset of dendrite pruning. First, the blockade of ER-Golgi transport caused by loss of Yif1/Yip1 function or Rab1DN/Sar1DN induction leads to a significant increase of Nrg proteins in the somas, dendrites and axons, similar to that in Rab5 mutant neurons. Moreover, when the ER-Golgi transport was inhibited upon Rab1DN/Sar1DN induction, Nrg was no longer redistributed to endosomes, indicating defective Nrg endocytosis. Finally, the pruning defects caused by loss of Yif1 or Yip1 function were significantly suppressed by Nrg knockdown. It was hypothesized that, in response to activation of ecdysone signalling, some unknown cell-surface or secreted molecules might be secreted via the canonical secretory pathway to facilitate Nrg endocytosis and thereby promote dendrite pruning. Over 1000 cell-surface or secreted molecules exist in Drosophila. Future studies will be necessary to identify such secreted molecules as potential 'eat me' signals for initiating Nrg endocytosis and dendrite pruning (Wang, 2018).
L1-CAM Nrg is the only known transmembrane protein that is endocytosed during dendrite pruning in ddaC neurons. It is conceivable that ER-to-Golgi transport likely generally affects endocytosis of various transmembrane proteins. A previous study has elegantly showed that an increase of general endocytosis occurs prior to dendrite pruning; loss of Rab5 function results in blockade of general endocytosis and thereby a dendrite pruning defect. Loss of Rab5 function leads to general endocytosis defects, including the formation of enlarged Avl-positive endosomes in ddaC neurons. These Rab5 mutant phenotypes were suppressed by the blockade of ER-to-Golgi transport, suggesting that aberrant ER-to-Golgi transport generally affects endocytosis, in addition to Nrg endocytosis. Thus, ER-to-Golgi transport may also generally facilitate Rab5-dependent endocytosis during dendrite pruning of ddaC sensory neurons (Wang, 2018).
Pruning of unspecific neurites is an important mechanism during neuronal morphogenesis. Drosophila sensory neurons prune their dendrites during metamorphosis. Pruning dendrites are severed in their proximal regions. Prior to severing, dendritic microtubules undergo local disassembly, and dendrites thin extensively through local endocytosis. Microtubule disassembly requires a katanin homologue, but the signals initiating microtubule breakdown are not known. This study shows that the kinase PAR-1 is required for pruning and dendritic microtubule breakdown. The data show that neurons lacking PAR-1 fail to break down dendritic microtubules, and PAR-1 is required for an increase in neuronal microtubule dynamics at the onset of metamorphosis. Mammalian PAR-1 is a known Tau kinase, and genetic interactions suggest that PAR-1 promotes microtubule breakdown largely via inhibition of Tau also in Drosophila. Finally, PAR-1 is also required for dendritic thinning, suggesting that microtubule breakdown might precede ensuing plasma membrane alterations. The results shed light on the signaling cascades and epistatic relationships involved in neurite destabilization during dendrite pruning (Herzmann, 2017).
The kinase PAR-1 is part of a pathway for microtubule disassembly during dendrite pruning. The data show that PAR-1 acts to enhance microtubule dynamics specifically during the early pupal phase. In the absence of PAR-1, c4da neurons accumulate stable microtubules at a time when control neurons have already degraded most of their dendritic microtubules. Genetic data suggest that Tau is a major target for PAR-1 in this process and that PAR-1 is required during pruning to remove, or inactivate Tau. It is known that Tau itself stabilizes microtubules; therefore, Tau inhibition likely serves to destabilize microtubules. Interestingly, Tau removal might also serve to activate the katanin homologue Kat-60L1 during dendrite pruning. This is an attractive possibility because Tau, but not the Futsch homolog MAP1B, has been shown to be a potent katanin inhibitor in mammalian cells, exactly matching the observed genetic interactions with PAR-1 during dendrite pruning. Tau also becomes depleted from mammalian sensory neuron axons after trophic
support withdrawal in an in vitro pruning model system. Tau depletion was not sufficient to induce pruning in mammalian sensory neurons, matching observations in c4da neurons. However, while not sufficient, it is interesting to speculate that Tau inactivation might also be required for pruning in mammalian neurons (Herzmann, 2017).
The data suggest that PAR-1 acts specifically during the pupal phase, but PAR-1 protein levels do not seem to increase at this stage. PAR-1 can be activated through phosphorylation by upstream kinases such as LKB1. lkb1 mutants showed only mild pruning defects that likely cannot fully explain the stronger defects caused by PAR-1 downregulation. Interestingly, PAR-1 interacts genetically with ik2, another kinase required for dendrite pruning. Thus, PAR-1 activation during dendrite pruning might depend on the interplay of several kinases. Given the temporal specificity of the PAR-1 effect, it is interesting to speculate that PAR-1 might be directly activated by a ecdysone-responsive factor (Herzmann, 2017).
It was also found that loss of PAR-1 prevents several processes at the dendritic plasma membrane during the pruning process: It prevented the local loss of membrane-associated Ank2XL from proximal dendrites, abrogated Ca2+ transients, and displayed strong enhancing genetic interactions with the thinning factor Shibire. As the genetic data indicate that Tau is the primary PAR-1 target during dendrite pruning, this suggests that microtubule breakdown is required for these plasma membrane alterations. In this scenario, the data actually suggest that microtubule disruption is closely linked to plasma membrane alterations, such that it is interesting to speculate that microtubule loss might trigger local endocytosis and thinning formation during dendrite pruning. Thus, a model is proposed where PAR-1, via Tau and possibly Kat-60L1, promotes microtubule disruption. In this model, these processes are placed epistatically over plasma membrane alterations during dendrite pruning (Herzmann, 2017).
Large scale neurite pruning is an important specificity mechanism during neuronal morphogenesis. Drosophila sensory neurons prune their larval dendrites during metamorphosis. Pruning dendrites are severed in their proximal regions, but how this spatial information is encoded is not clear. Dendrite severing is preceded by local breakdown of dendritic microtubules through PAR-1-mediated inhibition of Tau. This study investigated spatial aspects of microtubule breakdown during dendrite pruning. Live imaging of fluorescently tagged tubulin shows that microtubule breakdown first occurs at proximal dendritic branchpoints, followed by breakdown at more distal branchpoints, suggesting that the process is triggered by a signal emanating from the soma. In fly dendrites, microtubules are arranged in uniformly oriented arrays where all plus ends face towards the soma. Mutants in kinesin-1 and -2, which are required for uniform microtubule orientation, cause defects in microtubule breakdown and dendrite pruning. These data suggest that the local microtubule organization at branch points determines where microtubule breakdown occurs. Local microtubule organization may therefore contribute spatial information for severing sites during dendrite pruning (Herzmann, 2018).
The physiological degeneration of synapses, axons or dendrites without loss of the parent neuron is known as pruning. Pruning is an important developmental mechanism that is used to ensure specificity of neuronal connections and to remove developmental intermediates. Whereas the mechanisms of neurite outgrowth and synapse formation have been studied in some detail, comparably little is known about the mechanisms underlying pruning (Herzmann, 2018).
In holometabolous insects, the nervous system is remodeled on a large scale during metamorphosis. In the peripheral nervous system (PNS) of Drosophila, several types of sensory neurons undergo either apoptosis or prune their larval processes in an ecdysone-dependent manner. The sensory class IV dendritic arborization (c4da) neurons completely and specifically prune their long and branched larval dendrites at the onset of the pupal phase by a degenerative mechanism, while their axons remain intact. Dendrite pruning is induced in a cell-autonomous fashion by the steroid hormone ecdysone and proceeds in a stereotypical fashion. Dendrites are first severed at proximal sites close to the cell body between 5 and 10 h after puparium formation (APF). Severed dendrites are then fragmented and phagocytosed by the epidermal cells surrounding them. First signs of dendrite pruning are visible at 2-3 h APF, when proximal dendrites take on an irregular appearance with beadings and thinnings. In these regions, microtubules are disassembled locally. Plasma membrane retrieval through increased local endocytosis also contributes to thinning of proximal dendrites. Live imaging and genetic data suggest that local microtubule breakdown precedes membrane thinning. Microtubule disassembly is therefore the earliest known local sign of dendrite pruning. Microtubule disassembly requires the kinase PAR-1, which mediates inhibition of the microtubule-associated protein Tau. Furthermore, dendrite pruning also requires the microtubule-severing enzyme Katanin p60-like 1 (Kat-60L1), possibly also downstream of PAR-1 and Tau (Herzmann, 2018).
Microtubules are polar rods with so-called 'plus' and 'minus' ends. This nomenclature reflects the fact that the rates of both growth and shrinkage are greater at the plus ends. Microtubules in larval c4da neuron dendrites are oriented uniformly with their plus ends facing the soma. This uniform orientation is particularly prevalent in the primary and secondary dendrites, while higher order dendrites can also have microtubules with mixed orientation. Uniform dendritic plus-end-in microtubule orientation is known to depend on plus-end-directed motors of the kinesin family. In c4da neurons, uniform plus-end-in orientation depends on kinesin-2, which can bind to microtubule plus ends via EB1 and thereby promotes their orientation. In C. elegans, kinesin-1 was shown to be required for plus-end-in dendritic microtubule orientation. Kinesin-1 cannot bind to plus ends, and it was suggested that it instead moved wrongly oriented microtubules out of dendrites via microtubule sliding, or that it could be anchored to the dendritic cell cortex and could thus move microtubules out of the dendrite via its motor domain (Herzmann, 2018).
Given that microtubule breakdown occurs very early during the pruning process in a spatially confined manner, it is likely that this process involves spatial cues for pruning, such as the restriction of the destructive process to proximal over distal dendrites, or the sparing of the axon. This study addressed the apparent spatial regulation of microtubule disassembly during dendrite pruning by live imaging of fluorescently marked tubulin markers as well as in genetic studies. Gaps in dendritic microtubules were found to occur first at proximal dendrite branchpoints and later at more distal ones. Kinesin-2 is required for efficient c4da neuron dendrite pruning and microtubule disassembly, and EB1 manipulation was found to modulate the phenotypes associated with other microtubule disassembly factors, indicating that the uniform plus-end-in orientation of dendritic microtubules is also required for efficient microtubule breakdown and dendrite pruning. This is further supported by the observation that mutation of kinesin-1 also affects dendritic microtubule orientation, disassembly, and dendrite pruning in c4da neurons. It is proposed that microtubule disassembly depends on a signal emanating from the soma, and that the local microtubule organization at dendrite branchpoints, including the uniform plus-end-in orientation, favors microtubule disassembly there. The data indicate that dendritic microtubule organization may represent an important spatial cue for severing site selection during dendrite pruning (Herzmann, 2018).
Microtubule disassembly is the earliest known local destabilizing process during c4da neuron dendrite pruning and therefore a likely candidate to determine proximal severing. This study has begun to address the spatial regulation of microtubule disassembly during c4da neuron dendrite pruning. Using time-lapse live imaging of fluorescently tagged tubulin, it was found that gaps in the fluorescent microtubule signal develop preferentially at branchpoints of primary dendrites, and usually extend into the smaller side branches, rather than in the primary branch. Moreover, the temporal occurrence of these gaps correlates with proximity to the soma, i.e. microtubule gaps initially develop at the first branchpoint and later at the more distal branchpoints. These observations are consistent with a microtubule-destabilizing signal emanating from the soma. Previous work has shown that microtubule disassembly is induced by PAR-1-mediated Tau inhibition (Herzmann, 2017). PAR-1 activity is often induced by phosphorylation. Previous data suggested that PAR-1 is specifically activated for microtubule disassembly at the onset of the pupal phase, possibly by ecdysone (Herzmann, 2017). It is therefore plausible that the signal emanating from the soma is activated PAR-1. In such a model, activation of PAR-1 in the soma would contribute to the preference for microtubule disruption (and later dendrite severing) in proximal dendrites (Herzmann, 2018).
This study also found that the uniform plus-end-in orientation of dendritic microtubules is required for efficient microtubule disassembly and dendrite pruning. kinesin-2 and kinesin-1, both affecting dendritic microtubule orientation, are required for dendrite pruning. Manipulation of EB1, which caused less severe dendrite pruning defects by itself, strongly modified the defects seen upon manipulation of the microtubule dynamics regulator PAR-1. Interestingly, microtubule orientation (and hence the localization of the outermost microtubule plus ends) correlates well with the sites of degeneration and the degeneration mode in several models of large-scale pruning. For example, degeneration often starts from the distal end in axons, where microtubules have uniform plus-end-out orientation. As in c4da neurons, microtubule disassembly is an early event during large-scale axon pruning and is therefore likely to carry intrinsic spatial information crucial for pruning as well. This was shown to be the case in pruning axons of Drosophila mushroom body γ neurons and for pruning motoneuron axons at the mammalian neuromuscular junction. These axons retract, i.e. they shrink from their distal ends. Intriguingly, and in an exactly opposite manner to c4da neuron dendrites, these axons degenerate from the distal tips, and degeneration stops at the next branchpoint, again implying input from neurite branchpoints in the spatial regulation of large-scale pruning (Herzmann, 2018).
What could be special about microtubules at branchpoints? Although branchpoints could be localization sites for spatial pruning regulators, it is interesting to speculate that the local microtubule organization at branchpoints might suffice. For dendrites, uniformly oriented microtubules within the microtubule array would presumably enable the formation of larger gaps if the plus ends of the array microtubules have similar positions. Alternatively, microtubules at branchpoints could be more likely to be bent, and might therefore be more susceptible to shrinkage or catastrophe. Also, the overlap between the microtubule arrays of the primary and side branches may be smaller than that within an array in a straight branch, enabling gaps to occur at these sites even with relatively little microtubule shrinkage. Of note, previous data suggest that PAR-1 acts through inhibition of Tau to promote microtubule breakdown (Herzmann, 2017), and Tau was recently shown to promote microtubule bundling. Thus, PAR-1 activation might lead to 'unbundling' of microtubules at branchpoints. Furthermore, it could also be speculated that local concentrations of activated PAR-1 might increase as the diameter of the dendrite side branch decreases, which would also favor microtubule disassembly at proximal branchpoints. In order to distinguish between these models, it will be important to better characterize the local microtubule organization at dendritic branchpoints (Herzmann, 2018).
Previously it has been proposed that microtubule disassembly acts upstream of membrane thinning. Dendrite thinning was observed to be ~55% during the time of microtubule breakdown. Importantly, dendrites can often thin out to ~20% of their original diameter before rupturing, indicating that microtubule loss correlates with the onset of dendrite thinning (Herzmann, 2018).
Taken together, these data reveal that the spatial regulation of neurite pruning depends on the local microtubule organization. The data are consistent with a model in which microtubule disassembly carries the spatial information for pruning and therefore contributes to the selection of both degeneration mode and severing sites (Herzmann, 2018).
Phagocytic clearance of degenerating dendrites or axons is critical for maintaining tissue homeostasis and preventing neuroinflammation. Externalized phosphatidylserine (PS) has been postulated to be an "eat-me" signal allowing recognition of degenerating neurites by phagocytes. This study shows that in Drosophila, PS is dynamically exposed on degenerating dendrites during developmental pruning and after physical injury, but PS exposure is suppressed when dendrite degeneration is genetically blocked. Ectopic PS exposure via phospholipid flippase knockout and scramblase overexpression induced PS exposure preferentially at distal dendrites and caused distinct modes of neurite loss that differ in larval sensory dendrites and in adult olfactory axons. Surprisingly, extracellular lactadherin that lacks the integrin-interaction domain induced phagocyte-dependent degeneration of PS-exposing dendrites, revealing an unidentified bridging function that potentiates phagocytes. These findings establish a direct causal relationship between PS exposure and neurite degeneration in vivo (Sapar, 2018).
Drosophila class IV ddaC neurons selectively prune all larval dendrites to refine the nervous system during metamorphosis. During dendrite pruning, severing of proximal dendrites is preceded by local microtubule (MT) disassembly. This study identified an unexpected role of Mini spindles (Msps), a conserved MT polymerase, in governing dendrite pruning. Msps associates with another MT-associated protein TACC, and both stabilize each other in ddaC neurons. Moreover, Msps and TACC are required to orient minus-end-out MTs in dendrites. It was further shown that the functions of msps in dendritic MT orientation and dendrite pruning are antagonized by the kinesin-13 MT depolymerase Klp10A. Excessive MT depolymerization, which is induced by pharmacological treatment and katanin overexpression, also perturbs dendritic MT orientation and dendrite pruning, phenocopying msps mutants. Thus, this study demonstrated that the MT polymerase Msps is required to form dendritic minus-end-out MTs and thereby promotes dendrite pruning in Drosophila sensory neurons (Tang, 2020).
The evolutionarily conserved Glycogen Synthase Kinase 3β (GSK3β), a negative regulator of microtubules, is crucial for neuronal polarization, growth and migration during animal development. However, it remains unknown whether GSK3β regulates neuronal pruning, which is a regressive process. This study reports that the Drosophila GSK3β homologue Shaggy (Sgg) is cell-autonomously required for dendrite pruning of ddaC sensory neurons during metamorphosis. Sgg is necessary and sufficient to promote microtubule depolymerization, turnover and disassembly in the dendrites. Although Sgg is not required for the minus-end-out microtubule orientation in dendrites, hyperactivated Sgg can disturb the dendritic microtubule orientation. Moreover, pharmacological and genetic data suggest that Sgg is required to promote dendrite pruning at least partly via microtubule disassembly. This study showed that Sgg and Par-1 kinases act synergistically to promote microtubule disassembly and dendrite pruning. Thus, Sgg and Par-1 might converge on and phosphorylate a common downstream microtubule-associated protein(s) to disassemble microtubules and thereby facilitate dendrite pruning (Dzaki, 2022).
Mechanical forces actively shape cells during development, but little is known about their roles during neuronal morphogenesis. Developmental neurite pruning, a critical circuit specification mechanism, often involves neurite abscission at predetermined sites by unknown mechanisms. Pruning of Drosophila sensory neuron dendrites during metamorphosis is triggered by the hormone ecdysone, which induces local disassembly of the dendritic cytoskeleton. Subsequently, dendrites are severed at positions close to the soma by an unknown mechanism. This study found that ecdysone signaling causes the dendrites to become mechanically fragile. Severing occurs during periods of increased pupal morphogenetic tissue movements, which exert mechanical forces on the destabilized dendrites. Tissue movements and dendrite severing peak during pupal ecdysis, a period of strong abdominal contractions, and abolishing ecdysis causes non-cell autonomous dendrite pruning defects. Thus, these data establish mechanical tearing as a novel mechanism during neurite pruning (Kramer, 2023).
Localizing messenger RNAs at specific subcellular sites is a conserved mechanism for targeting the synthesis of cytoplasmic proteins to distinct subcellular domains, thereby generating asymmetric protein distributions necessary for cellular and developmental polarity. However, the full range of transcripts that are asymmetrically distributed in specialized cell types and the significance of their localization, especially in the nervous system, are not known. This study used the EP-MS2 method, which combines EP transposon insertion with the MS2/MCP in vivo fluorescent labeling system to screen for novel localized transcripts in polarized cells, focusing on the highly branched Drosophila class IV dendritic arborization neurons. Of a total of 541 lines screened, 55 EP-MS2 insertions were identified producing transcripts that were enriched in neuronal processes, particularly in dendrites. The 47 genes identified by these insertions encode molecularly diverse proteins and are enriched for genes that function in neuronal development and physiology. RNAi-mediated knockdown confirmed roles for many of the candidate genes in dendrite morphogenesis. It is proposed that the transport of mRNAs encoded by these genes into the dendrites allows their expression to be regulated on a local scale during the dynamic developmental processes of dendrite outgrowth, branching, and/or remodeling (Misra, 2016).
Neurons develop highly stereotyped receptive fields by coordinated growth of their dendrites. Although cell surface cues play a major role in this process, few dendrite specific signals have been identified to date. An in vivo RNAi screen in Drosophila class IV dendritic arborization (C4da) neurons identified the conserved Ret receptor (Ret oncogene), known to play a role in axon guidance, as an important regulator of dendrite development. The loss of Ret results in severe dendrite defects due to loss of extracellular matrix adhesion, thus impairing growth within a 2D plane. Evidence is provided that Ret interacts with integrins to regulate dendrite adhesion via rac1. In addition, Ret is required for dendrite stability and normal F-actin distribution suggesting it has an essential role in dendrite maintenance. Novel functions are proposed for Ret as a regulator in dendrite patterning and adhesion distinct from its role in axon guidance (Soba, 2015).
Accurate functional connectivity and sensory perception require proper development of the neuronal dendritic field, which ultimately determines the (sensory) input a specific neuron can receive and detect. Thus, coordinated dendrite growth and patterning is important for establishing the often complex, but highly stereotyped organization of receptive fields. Two of the organizing principles in dendrite development are self-avoidance and tiling. While self-avoidance describes the phenomenon of recognition and repulsion of isoneuronal dendritic branches, tiling refers to the complete yet non-redundant coverage of a receptive field by neighboring neurons of the same type. Both phenomena have been described in different systems across species including the mouse, zebrafish, medicinal leech, Caenorhabditis elegans, and Drosophila melanogaster (Soba, 2015).
Dendritic patterning by self-avoidance, tiling, and other mechanisms is thought to be mediated by cell surface receptors and cell adhesion molecules (CAMs), which play a pivotal role in integrating environmental and cellular cues into appropriate growth and adhesion responses. Many such receptors, prominently Robo and Ephrin receptors, have well understood roles in axon guidance. Although some of these axonal cues including Robo/Slit play a role in dendrite development as well, dendritic surface receptors and their functions are not fully characterized to date. Recent efforts have yielded some progress in this area. Down's syndrome cell adhesion molecule (Dscam) has been shown to regulate dendrite self-avoidance in Drosophila. Studies on protocadherins have revealed that they play an important role in dendrite self-avoidance in mammals. In C. elegans, sax-7/L1-CAM and menorin (mnr-1) form a defined pattern in the surrounding hypodermal tissue to guide PVD sensory neuron dendrite growth via the neuronal receptor dma-1. However, given the complexity and stereotypy of dendritic arbors within individual neuronal subtypes, it is important to search for additional signals for directing dendrite growth (Soba, 2015).
The Drosophila peripheral nervous system (PNS) has served as an excellent model which has helped to elucidate several molecular mechanisms regulating dendrite development. The larval PNS contains segmentally repeated dendritic arborization (da) neurons which have been classified as class I-IV according to their increasing dendritic complexity. All da neuron classes feature highly stereotyped sensory dendrite projections. Moreover, all da neurons exhibit self-avoidance behavior allowing them to develop their individual receptive fields without overlap. It has been demonstrated that all da neuron classes require Dscam for dendrite self-avoidance. In addition, the atypical cadherin flamingo and immunoglobulin super family (IgSF) member turtle might play a more restricted role in C4da neuron self-avoidance. Netrin and its receptor frazzled have also been shown to act in parallel to Dscam in class III da neurons ensuring their proper dendritic field size and location by providing an attractive growth cue which is counterbalanced by self-avoidance. For tiling, no surface receptor has been identified to date. However, the conserved hippo and tricornered kinases, and more recently the torc2 complex, have been implicated in C4da neuron tiling, as the loss of function of these genes results in iso- and hetero-neuronal crossing of dendrites (Soba, 2015).
Recent work has further shown that dendrite substrate adhesion plays an essential role in patterning. Da neuron dendrites are normally confined to a 2D space through interaction with the epithelial cell layer and the extracellular matrix (ECM) on the basal side of the epidermis. 2D growth of da neuron dendrites requires integrins, as loss of the α-integrin mew (multiple edomatous wing) or ß-integrin mys (myospheroid) results in dendrites being freed from the 2D confinement due to detachment from the ECM. Thus, they can avoid dendrites by growing into the epidermis leading to 3D crossing of iso- and hetero-neuronal branches . Integrins are therefore essential to ensure repulsion-mediated self-avoidance and tiling mechanisms, which restrict growth of dendrites competing for the same territory. How integrins are recruited to dendrite adhesion sites and whether they cooperate with other cell surface receptors is unknown (Soba, 2015).
To identify novel receptors required for generating complex, stereotypical dendritic fields, an in vivo RNAi screen was performed for cell surface molecules in C4da neurons. The Drosophila homolog of Ret (rearranged during transfection) was identified as a patterning receptor of C4da dendrites. Loss of Ret function in C4da neurons severely affects dendrite coverage, dynamics, growth, and adhesion. In particular, dendrite stability and 2D growth are impaired resulting in reduced dendritic field coverage and abnormal 3D dendrite crossing, respectively. These defects can be completely rescued by Ret expression in C4da neurons. It was further shown that Ret interaction with integrins is needed to mediate C4da dendrite-ECM adhesion, but not dendrite growth. These data suggest that Ret together with integrins acts through the small GTPase rac1, which is required for dendrite adhesion and 2D growth of C4da neuron dendrites as well. This study thus describes a novel role for the Ret receptor in dendrite development and adhesion by direct receptor crosstalk with integrins and its downstream signals (Soba, 2015).
This study provides evidence that Ret is a regulator of dendrite growth and patterning of C4da neurons. Ret is a conserved receptor tyrosine kinase (RTK) expressed in the nervous system of vertebrates and D. melanogaster , and has been shown to have a number of important functions in nervous system development and maintenance: it regulates motor neuron axon guidance (Kramer, 2006), dopaminergic neuron maintenance and regeneration, and mechanoreceptor differentiation and projection to the spinal cord and medulla. Ret signaling is activated by binding to glial cell line derived neurotrophic factor (GDNF) family ligands and their high affinity co-receptors, the GDNF family receptors (GFRα). Ret also plays an important role in human development and disease as loss of function mutations of Ret lead to Hirschprung's disease displaying colonic aganglionosis due to defective enteric nervous system development. Conversely, Ret gain of function mutations are causal for autosomal dominant MEN2 (multiple endocrine neoplasia type 2) type medullary thyroid carcinoma (Soba, 2015 and references).
Prior to this study, Ret has not been implicated in dendrite development. This study shows that Ret is required specifically for 2D growth of C4da neurons by regulating integrin dependent dendrite-ECM adhesion. Normally, C4da neuron dendrites are virtually always in contact with the ECM and the basal surface of the epithelium lining the larval cuticle, and thus tightly sandwiched between the two compartments. In both integrin and Ret mutants, dendrite-ECM adhesion is impaired. Ret and integrins can co-localize in dendrites and thus likely form a functional complex that could induce and maintain adhesion of dendrites to the ECM. Since Ret loss of function primarily leads to detached terminal dendrite branches, it is tempting to speculate that Ret might be required to recruit integrins to sites of growing dendrites to promote ECM interaction. This is supported by the colocalization of Ret and integrins on the dendrite surface. Their cooperative interaction could thus ensure proper adhesion of growing branches and, conversely, the fidelity of self-avoidance and tiling (Soba, 2015).
These results also highlight the importance of integrating different guidance and adhesion cues to achieve precise neuronal patterning. This has so far only been studied in axon guidance in vivo. Interestingly, vertebrate Ret has been shown to cooperate with Ephrins to ensure high fidelity axon guidance in motor neurons by mediating attractive EphrinA reverse signaling (Kramer, 2006; Bonanomi, 2012). Similar mechanisms may conceivably be employed for growing dendrites, which also encounter a multitude of attractive, repulsive, and adhesive cues that have to be properly integrated. Besides pathways acting independently or in a parallel fashion, an emerging view is that receptors exhibit direct crosstalk to integrate incoming signals. So far, only parallel receptor pathways like Dscam and Netrin-Frazzled signaling in class III da neurons or Dscam/integrins have been identified co-regulating dendrite morphogenesis. The current data show that the Ret receptor and integrins integrate dendrite adhesion and growth by collaborative interaction of the two cell surface receptors. The molecular and genetic link between Ret and integrins suggests that in this case direct receptor crosstalk plays a major role in their function. How exactly these cell surface receptors cooperate and interact remains to be elucidated. Integrins have been shown to display extensive crosstalk with other signaling receptors, including RTKs. Although integrins are involved in adhesion of virtually all cell types, the underlying signaling and recruitment of integrins to sites of adhesion in vivo is complex and not completely understood. It has been suggested that integrin and growth factor receptor crosstalk can occur by concomitant signaling, collaborative activation, or direct activation of associated signaling pathways. For example, matrix-bound VEGF can induce complex formation between VEGFR2 and β1-integrin with concomitant targeting of β1-integrin to focal adhesions in endothelial cells. The current findings of biochemical interaction and colocalization of Ret with the α/β-integrins mys and mew in C4da neuron dendrites argue in favor of direct receptor interaction and subsequent activation of a common signaling pathway (Soba, 2015).
Integrins and RTKs like Ret do share some of the same intracellular signaling components. These comprise, among others, the MAPK (mitogen-activated protein kinase) pathway, Pi3-Kinase (Pi3K), and Rho family GTPases including Rac1. Previous studies provide evidence for Ret-integrin-Rac1 interplay in vitro showing that Ret can enhance integrin mediated adhesion and induce Rac1 dependent lamellipodia formation in cell culture models. In primary chick motor neurons, Rac1 is involved in neurite outgrowth on the integrin substrates laminin and fibronectin. Interestingly, Rac1 has previously been shown to regulate dendrite branching in C4da neurons, however a role in dendrite adhesion in vivo has not been described before. This study shows that Rac1 is required for dendrite-ECM adhesion similarly to what has been described for integrins, and Ret and integrin dependent adhesion was genetically linked with Rac1 function. In Drosophila, MAPK, Src and PI3K can be activated by constitutively active Ret overexpression in the compound eye. Moreover, novel inhibitors of Ret signaling targeting Raf, Src, and S6-Kinase (S6K) prevent lethality induced by Ret over-activation in a Drosophila multiple endocrine neoplasia (MEN2) model. Interestingly, S6K has been shown to be involved in dendrite growth but not tiling in C4da neurons. It remains to be shown if these pathways play a direct role in Ret function in dendrite adhesion and growth (Soba, 2015).
Notwithstanding important commonalities, Ret function in C4da neurons cannot be fully explained by crosstalk with integrins and rac1. Reduced dendritic field coverage, likely due to the observed increase in dendrite turnover, is only evident in Ret but not in integrin or rac1 mutant C4da neurons. Moreover, increasing integrin expression in a Ret mutant background did not rescue dendrite coverage defects, albeit it prevented dendrite crossing. These findings indicate that Ret has additional functions in dendritic branch growth and stability that require as yet unknown extracellular and intracellular mediators. This is also supported by the aberrant F-actin localization in neurons lacking Ret. In this study, Ret dependent intracellular effectors are likely important for F-actin assembly to support directed dendrite growth and stabilization, and their localization and activity might be deregulated in the absence of Ret (Soba, 2015).
Drosophila Ret is a highly conserved molecule, its cognate vertebrate ligand GDNF, however is not (Airaksinen, 2006). In addition, Drosophila Ret can neither bind GDNF nor transduce GDNF signaling, although it has been shown to contain a functional tyrosine kinase domain. In mammals, the GFRα co-receptors are essential components of GDNF/Ret signaling. A Drosophila GFR-like homolog (dGFRL) has recently been characterized and was found to function and interact with the NCAM homolog FasII. Therefore, it appears that Ret's functional interaction partners in dendrite development differ significantly from the previously described co-factors in other systems. It is interesting to speculate that a yet undiscovered Ret ligand is involved in Ret mediated dendrite growth and branch stabilization, which might have implications for mammalian Ret function as well: due to its role in the maintenance of dopaminergic neurons and motor axon growth in mouse, adhesion related signaling via integrins could well be important during these processes. Moreover, the formation of a dorsal root ganglia derived mechanosensory neurons and their afferent and efferent fiber growth and innervation depends on Ret expression. It will be interesting to investigate the functional interplay of Ret and integrins in central and peripheral target innervation and neurite maintenance in these systems, given the interdependent function of Ret and integrins in sensory dendrite growth as shown in this study (Soba, 2015).
In summary, this study describes a novel role for the Ret receptor in dendrite branch growth and stability in Drosophila C4da neurons. This role involves cell-autonomous effects of Ret on ECM adhesion, and F-actin localization in these neurons. Moreover, dendritic adhesion defects attributable to Ret have been linked to integrin and rac1 function featuring a novel and possibly conserved mode of action for Ret in dendrite development (Soba, 2015).
As the nervous system develops, there is an inherent variability in the connections formed between differentiating neurons. Despite this variability, neural circuits form that are functional and remarkably robust. One way in which neurons deal with variability in their inputs is through compensatory, homeostatic changes in their electrical properties. This study shows that neurons also make compensatory adjustments to their structure. The development of dendrites on an identified central neuron (aCC) was studied in the late Drosophila embryo at the stage when it receives its first connections and first becomes electrically active. At the same time, the distribution of presynaptic sites on the developing postsynaptic arbor was charted. Genetic manipulations of the presynaptic partners demonstrate that the postsynaptic dendritic arbor adjusts its growth to compensate for changes in the activity and density of synaptic sites. Blocking the synthesis or evoked release of presynaptic neurotransmitter results in greater dendritic extension. Conversely, an increase in the density of presynaptic release sites induces a reduction in the extent of the dendritic arbor. These growth adjustments occur locally in the arbor and are the result of the promotion or inhibition of growth of neurites in the proximity of presynaptic sites. Evidence is provided that suggest a role for the postsynaptic activity state of protein kinase A in mediating this structural adjustment, which modifies dendritic growth in response to synaptic activity. These findings suggest that the dendritic arbor, at least during early stages of connectivity, behaves as a homeostatic device that adjusts its size and geometry to the level and the distribution of input received. The growing arbor thus counterbalances naturally occurring variations in synaptic density and activity so as to ensure that an appropriate level of input is achieved (Tripodi, 2008).
Since cholinergic neurons provide the only known excitatory input to Drosophila motor neurons in the embryo, the effect of the lack of neurotransmitter (synthesis) in the cholinergic neurons was examined on the development of the aCC dendritic arbor. Acetyl choline is not synthesized in animals mutant for choline acetyl transferase (Chal13), which are therefore immobile and unable to hatch. The development of aCC dendritic arborisations was examined in animals homozygous for the null mutation Chal13. In Cha mutant embryos, it was found that the development of the aCC dendritic arbor proceeds normally until 16 h AEL. However, in the interval between 16 to 18 h AEL, Cha mutants, unlike controls, fail to reduce the rate of dendritic growth. As a result, at 18 h AEL, the extent of the aCC dendritic arbor is increased by about 26% in Cha mutants as compared to controls. It is concluded that in the window of development, when in normal embryos acetyl choline-dependent excitation of aCC first begins and the rate of dendritic growth declines, the absence of neurotransmitter in presynaptic neurons allows postsynaptic growth to continue linearly at an undiminished constant rate. These findings suggest that the aCC dendritic arbor reacts to the loss of synaptic input by increasing in size. A consequence of this increase in overall dendritic length is that it allows the dendritic arbor to explore a larger portion of the neuropile than in normal animals. In fact, it was observed that in Cha mutants, the dendritic arbor of aCC extends into regions of the neuropile that are not normally invaded in control conditions (Tripodi, 2008).
Growth adjustments by the aCC arbor appear to operate sequentially at two levels. In the first instance, the dendritic arbor determines the presence or absence of presynaptic partners. This event is independent of presynaptic activity. It is also a local event that appears to affect primarily neurites receiving presynaptic sites, which act as a local stop-growing signal. The second step depends on the activity of the synapse. In normal conditions, presynaptic sites are able to inhibit growth, both in synaptic neurites as well as neighbouring nonsynaptic sister neurites. Interestingly, the dendritic arbor does not measure the efficacy of synapses (since there is no significant change in arbor size in Ace mutants as compared to wild type), but simply determines whether a synapse is active or not. However, the possibility of there being a compensatory response to reduced efficacy cannot be excluded. This phase of activity-dependent inhibition of dendritic growth is mediated by postsynaptic activation of PKA (Tripodi, 2008).
The morphology of dendrites is likely to be an important determinant of the connectivity state of a nervous system. It is not by chance that even in complex nervous systems, one of the most distinctive feature of different classes of neurons is probably the morphology of their dendritic arbors. Indeed, the anatomical discrimination of different classes of neurons, based on their dendritic morphology, has often anticipated and predicted molecular, electrophysiological, and computational differences (Tripodi, 2008).
Therefore, an appreciation of the logic governing the assembly of a nervous system must include an understanding of how dendritic morphology is acquired. Although many of the distinctive features that differentiate the dendritic morphologies of different classes of neurons are likely to be cell-autonomously and genetically determined, the effect of partner-derived cues, as shown in this study, can have a substantial impact in modulating these features. This investigation was initiated by analysing the effect of presynaptic transmission in shaping the morphology of the postsynaptic dendritic arbor. Previous studies on this issue have not reached a clear agreement on the role of activity in regulating dendritic growth. Even though in some instances it has been reported that activity has no effect on regulating arbor growth, in the majority of studies, activity emerges clearly as an essential modulator of dendritic remodelling. The main issue has been whether the incoming presynaptic input acts as a trophic factor that promotes arbor growth or whether it delivers a stop-growing signal. Unfortunately, with a few notable exceptions, many of these studies were carried out in different animals, in different neural populations, at different developmental stages, and by using different experimental approaches (genetic manipulation or pharmacological treatments), thus making it difficult to find a common theme in the results. One major source of variation that could explain the differences in the results of previous studies is likely to be the developmental stage at which manipulations were applied. For instance, a single class of tectal neurons in Xenopus tadpoles appears to respond in opposite ways to presynaptic input at different developmental stages. In immature neurons, presynaptic input acts as a growth-promoting signal, whereas in mature neurons, it acts as a stop-growing or stabilization signal. In this study, no evidence was found for a growth-promoting effect of synaptic activity. Instead, it was shown that synaptic input inhibits dendritic growth starting from the earliest stages at which neurotransmission occurs (16-18 h AEL). Altering activity before the onset of evoked synaptic transmission causes no dendritic phenotype (14-16 h AEL). It appears that developing Drosophila embryonic motor neurons behave like mature tectal neurons in Xenopus (Tripodi, 2008).
The difference in the effect obtained at different stages in Xenopus tectal neurons nicely correlates with a change in their molecular characteristics. At later stages, when synaptic input acts as a stop-growing signal, the tectal neurons in Xenopus have acquired the ability to respond to local calcium increases via the activation of a calcium-dependent protein kinase CamKII. In Drosophila, aCC appears be sensitive to the activation state of a different protein kinase, PKA (which is also regulated, albeit indirectly, by intracellular calcium levels) throughout the interval of development that was studied (Tripodi, 2008).
A great deal of attention has been given to global changes in postsynaptic activity and how these changes might regulate global dendritic growth. However, although this is an extremely interesting question, it is difficult to imagine how global variations in the state of activation of the whole dendritic arbor or the soma might contribute to fine-tuning the morphology of dendritic arbors. Far more compelling would be a system that was able to calculate and respond to local changes in activity levels. It is well known that calcium levels can be altered locally at the synaptic site following synaptic input. It is also known that changes in dendritic levels of calcium can induce dramatic changes in dendritic morphology. Nevertheless, there have been few investigations of how dendritic morphology might be altered locally by synaptic activity (Tripodi, 2008).
Because the system used allows the study a single identified postsynaptic neuron whose presynaptic input can be altered, this study begins to address the issue of local versus global changes of dendritic morphology induced by synaptic activity. Analyzing the branching pattern of the dendritic arbor with respect to the position of its synaptic inputs highlights some interesting and unexpected features. By simply looking in wild-type animals, it is clear that neurites bearing presynaptic sites branch less than nonsynaptic neurites of the same arbor. Through experimental manipulation, this study has shown that this local inhibition of the growth and branching of synaptic neurites appears to be mediated by contact between pre- and postsynaptic partners and does not require evoked transmission. The lack of this contact-dependent inhibition of dendritic growth is already apparent when the terminals normally presynaptic to aCC are mistargeted before they can form functional synapses at 16 h AEL. Moreover, this effect cannot be attributed to an interference with transmission, since Cha mutants of the same stage (16 h AEL) do not show this dendritic overgrowth phenotype. Loss of neurotransmitter release, on the other hand, can also induce an overgrowth of the postsynaptic dendritic arbor, though only after synapses would have been active for 2 h in normal conditions (i.e., 18 h AEL). It was found that this neurotransmitter-dependent overgrowth is due to increased extension of nonsynaptic segments that are immediately adjacent to neurites receiving presynaptic sites. Thus, neurotransmitter release at the synaptic sites acts on the nonsynaptic sister neurites to inhibit their extension (Tripodi, 2008).
It is concluded that the geometry of the dendritic tree is regulated by two partner-dependent mechanisms. First, contact with presynaptic terminals locally inhibits dendritic growth and branching in an activity-independent fashion. A second inhibitory effect requires evoked presynaptic neurotransmitter release. It extends from these sites to the immediate proximity, affecting nonsynaptic sister branches. This second 'neighbourhood effect' could be mediated by local increases in dendritic calcium levels (Tripodi, 2008).
Neurotransmission-dependent variations in intracellular calcium levels are an attractive mechanism that might implement activity-dependent local rearrangement of the dendritic arbor geometry. Therefore the role of the protein kinase PKA in dendritic remodelling was investigated, since its activity is regulated directly or indirectly by calcium levels. PKA is activated by increases in the intracellular levels of cAMP. It has been shown that levels of cAMP are finely regulated by intracellular levels of calcium, which in turn respond to levels of presynaptic neurotransmitter release (Tripodi, 2008).
PKA signalling in postsynaptic cells has been shown to mediate homeostatic responses. For instance, at the neuromuscular junction, postsynaptic PKA signalling modulates quantal size (Davis, 1998), while centrally, it mediates homeostatic change in the electrical excitability of aCC neurons following alterations to presynaptic input (Baines, 2003). This study has shown that PKA activity is also a potent modulator of dendritic morphology. PKA signalling is probably downstream of presynaptic transmission mediating the inhibition of dendritic growth. Overexpression of a constitutively active form of PKA (PKAact) is able to rescue the dendritic overgrowth phenotype that ensues in the absence of presynaptic transmission in Cha mutants. However, overexpression of PKAact in aCC in control animals has no measurable effect on dendritic development, suggesting that PKA signalling operates to saturation under normal levels of synaptic input. Although neurotransmission is clearly one signal that regulates dendritic development through downstream PKA signalling, it is not necessarily the only one (Tripodi, 2008).
The term homeostasis has classically been used to refer to compensatory variations in the electrical properties of neurons that tend to counterbalance changes in their synaptic input. This paper proposes that neurons might combine the homeostatic regulation of electrical properties with compensatory structural adjustments of their dendritic geometry. It is argued that variations in the morphology of aCC dendritic arbors represent such compensatory adjustment of dendritic growth and branching (Tripodi, 2008).
These experimental observations indicate that eliminating synaptic input induces a compensatory dendritic overgrowth in the postsynaptic neuron, whereas an increase in the density of active synapses induces the opposite effect. Interestingly, increasing the overall level of neurotransmitter released at presynaptic terminals without altering the density of presynaptic sites on the arbor does not induce compensatory adjustments of the postsynaptic arborisation. This suggests that the compensatory changes in dendritic arbor morphology that were observed act to compensate for variations in the density of active synaptic release sites rather than variations in the global state of dendritic or neuronal depolarization. This is in agreement with observations that changes in dendritic morphology in response to changes in presynaptic input or synaptic density appear to be implemented locally rather than across the entire arbor. This structural homeostasis, therefore, seems to work on a local scale, allowing particular regions of the dendritic arbor to compensate for variation in their inputs while leaving other regions of the arbor substantially unchanged. This could be an effective mechanism for neurons that use distinct regions of their dendritic arbor to independently compute different inputs (Tripodi, 2008).
It will be interesting to understand whether electrical homeostasis and what is proposed to be a structural homeostasis operate in concert to shape the postsynaptic response, or whether one or the other is preferentially deployed depending on the circumstances. At the moment, it is not possible to answer this question, and more experiments are required (Tripodi, 2008).
Nonetheless, one can begin to envisage how PKA signalling in the context of electrical homeostasis might be integrated with the structural homeostasis that was shown in this study. As shown by Baines (2003), postsynaptic overexpression of a constitutively active form of PKA induces a change in the electrical properties of aCC, resulting in a decrease in excitability. Overexpression of a PKA inhibitor, on the other hand, does not modulate the excitability of aCC. This study has described a perfect mirror image of this situation, namely that the expression of a PKA inhibitor induces a dendritic overgrowth phenotype, while expression of a constitutively active form of PKA does not. Bringing these two lines of observations together suggests the following model: following a decrease in presynaptic input (and PKA signalling), the postsynaptic neuron expands its receptive field so as to increase the number of presynaptic sites that it contacts. In these circumstances, an increase in postsynaptic excitability would not be required. On the other hand, following an increase in presynaptic input, the postsynaptic neuron decreases its excitability (via increased PKA signalling) without the need to reduce its receptive field (Tripodi, 2008).
The view is favoured that electrical and structural homeostatic mechanisms might indeed be integrated by neurons. This would enable the cells to implement compensatory changes that resulted in adjustments to their electrical characteristics and dendritic geometry, so as to ensure that in an inherently variable environment an adequate pattern and level of connectivity and excitability is achieved (Tripodi, 2008).
Little is known about how the distinct architectures of dendrites and axons are established. From a genetic screen, dendritic arbor reduction (dar) mutants were isolated with reduced dendritic arbors but normal axons of Drosophila neurons. dar2, dar3, and dar6 genes were identified as the homologs of Sec23, Sar1, and Rab1 of the secretory pathway. In both Drosophila and rodent neurons, defects in Sar1 expression preferentially affected dendritic growth, revealing evolutionarily conserved difference between dendritic and axonal development in the sensitivity to limiting membrane supply from the secretory pathway. Whereas limiting ER to Golgi transport resulted in decreased membrane supply from soma to dendrites, membrane supply to axons remained sustained. It was also shown that dendritic growth is contributed by Golgi outposts (see Horton, 2003), which are found predominantly in dendrites. The distinct dependence between dendritic and axonal growth on the secretory pathway helps to establish different morphology of dendrites and axons (Ye, 2007).
This study has demonstrated that growing dendrites and axons display different sensitivity to changes in the activity of the secretory pathway. These findings add to the accumulating evidence that the secretory pathway is involved in cell polarity and provide one of the first evidence of the importance of the satellite secretory pathway in dendrite development (Ye, 2007).
The secretory pathway is important for cell polarity. For example, during the cellularization of Drosophila embryos, membrane growth is tightly controlled in a polarized fashion. New membrane produced through the secretory pathway is predominantly added to the apical side early in polarization and to the lateral side at later stages. Other examples of targeted membrane transport during cell polarization include the apical-basolateral polarization of Madin-Darby canine kidney epithelial cells, cell cycle of the budding yeast, and cell migration. Thus, membrane trafficking is tightly controlled in order to supply membrane to specific subcellular compartments of polarized cells (Ye, 2007).
The establishment of dendritic and axonal arbors is a major compartmentalization process of neurons. Before dendrites and axons are formed, selective delivery of post-Golgi vesicles to one neurite precedes the specification of that neurite as the axon. After axon specification, large amount of plasma membrane is added to these two compartments with distinct architecture. Whether the secretory pathway is involved in the differential growth of dendrites and axons is an important but poorly understood question. Horton (2005) found that blockade of post-Golgi trafficking by expressing a dominant-negative protein kinase D1 (PKD-KD) causes a preferential reduction in dendritic growth. Although this result may appear similar to the current results from manipulation of Sar1 function, there is in fact an important difference. PKD is required for cargo trafficking to the basolateral but not the apical membrane in epithelial cells. Since the targeting of dendritic proteins shares some of the mechanisms of the basolateral protein transport in epithelial cells, PKD-KD may only affect dendrite-specific cargo transport. This study assessed how membrane resource is allocated between growing dendrites and axons when global membrane production is limited. It is possible that the efficiency of dendrite-and axon-specific transport, including those mediated by PKD, changes differently in response to global restriction of membrane resource (Ye, 2007).
The plasma membrane of cells can be inserted via exocytosis and internalized through endocytosis. The expansion of plasma membrane in growing dendrites and axons is achieved through the interplay between these two antagonistic processes. Before dendrites are formed in cultured hippocampal neurons, membrane is selectively added to the growth cone of growing axons as well as minor neurites that will later become dendrites. In more mature neurons, there is insertion of new membrane to the axonal growth cone but not to the dendritic growth cone. This raises the possibility that plasma membrane is added to either multiple sites along the dendritic surface or throughout the dendritic surface. It is conceivable that dendritic Golgi outposts play a role in such membrane addition given their requirement for dendritic growth as shown in this study (Ye, 2007).
It has been reported that plasma membrane of cultured hippocampal neurons is internalized throughout the dendrites but only in the presynaptic terminals in axons. It was also found that essentially all internalized structures, from both dendritic and axonal surface, move in the retrograde direction to the soma. The rate of membrane removal from dendritic and axonal surfaces were compared and it was found that endocytosis is more prominent in dendrites than in axons. The plasma membrane of dendrites was endocytosed 8.27 times faster than that of axons at 3 days in vitro. Thus, the demand for membrane supply to dendrites is likely greater than what appears based on the length of dendrites (Ye, 2007).
The difference in the ways that membrane is supplied to growing dendrites and axons is poorly understood. Notwithstanding much progress made on dendritic traffic of the temperature sensitive mutant of the vesicular stomatitis virus glycoprotein (VSVG) fused to GFP (Horton, 2003; Horton, 2005), the preferential targeting of VSVG to dendrites precludes its application for comparing dendritic and axonal membrane dynamics. This work compared the membrane supply from soma to dendrites and axons using the FRAP analysis. Fluorescence recovery was sampled in the dendritic and axonal segment 5 microm away from the soma as an indicator of the amount of the membrane passing though this segment. The sampling approach was taken because even with moderate magnification (20X) only a fraction of the arbors can be included in the imaging field, precluding the monitoring of fluorescence recovery along the entire dendritic and axonal arbor. This FRAP analysis revealed that, when the secretory pathway function is limiting, membrane supply from soma to growing dendrites is preferentially affected (Ye, 2007).
Notably, the secretory pathway with reduced activity could still provide sufficient membrane to support axonal growth at least during the period described in this study. It is possible that there are mechanisms involving post-Golgi carriers to allocate different amount of membrane to dendrites and axons. Alternatively, endocytosed dendritic membrane can also support axonal growth via the transcytosis pathway. Lastly, trafficking that bypasses COPII-based ER to Golgi transport, although not widely observed, might also contribute to axonal growth (Ye, 2007).
The finding that Golgi outposts are present in dendrites has generated great interests in the function of these structures. Two main functions have been proposed for Golgi outposts. One hypothesis is that they are involved in local protein translation (Steward, 2003). This is supported by the presence of protein synthesis machinery in the dendrites and findings of local translation of membrane proteins in dendrites suggest that Golgi outposts, together with somatic Golgi complex, participate in forming the apical dendrites of pyramidal neurons (Ye, 2007).
In this study, the dynamics of Golgi outposts were monitored in intact neurons of live Drosophila larvae, and it was observed that the direction of outpost movement correlates with dendrite branch dynamics. This suggests that Golgi outposts are probably involved in both the extension and retraction of a dendritic branch. Indeed, laser damaging of the outposts in a branch reduced branch dynamics, making the dendrite excessively stable, which will lead to retarded growth during the expansion of dendritic arbors. It should be noted that, in the laser damaging experiments, the nature and extent of damages to Golgi outposts are unclear. It is indeed technically challenging to manipulate organelle function in a small, localized subcellular region. The laser damaging experiment is considered one of the first attempts of such manipulations. Although it can be concluded that Golgi outposts are the most sensitive locations for affecting dendritic branch dynamics among the various target locations, alternative approaches are needed to test the function of Golgi outposts. The present study complemented the laser damaging experiment with genetic manipulations to redistribute Golgi outposts. It was found that dendritic branching patterns changed with the redistribution of Golgi outposts by LvaDN and Lva-RNAi. High level of LvaDN expression led to the strongest defect in Golgi outpost distribution and dendritic branching pattern. Although it is difficult to rule out the contribution of non-specific effects of LvaDN, there is very good correlation between the distribution of Golgi outposts and that of dendritic branches in neurons with varying extent of disruption of Lva function. These findings underscore the importance of Golgi outposts in dendritic growth (Ye, 2007).
It remains unclear how Golgi outposts are produced, transported to dendrites, and largely excluded from axons. The results on Lva suggest that they are probably produced in the soma and transported to dendrites. The presence of Golgi outposts in axons of LvaDN-expressing neurons suggest that they might be actively transported out of axons under normal condition. The absence of Golgi outposts and exuberant branches in the proximal axons of neurons expressing Lva-RNAi is likely due to partial interference of Lva function as similar phenotype was seen in neurons expressing low level of LvaDN. The increase in the size of Golgi outposts and dendritic branches in neurons expressing high level of LvaDN possibly reflect the requirement of Lva-dynactin interaction for the budding of Golgi outposts, a phenomenon that was observed in this study(Ye, 2007).
Although it is difficult to rule out the presence of outposts that are below the detection sensitivity in this study, axonal Golgi outposts, if any, must be dramatically smaller and/or fewer than dendritic outposts. Therefore, the requirement of Golgi outposts for dendritic growth provides a mechanism to differentially control dendrite and axon growth (Ye, 2007).
The secretory pathway is possibly under regulation to control dendritic growth
The distinct dependence of dendrites and axons on the secretory pathway, as well as the involvement of Golgi outposts for local dendritic dynamics, raise the possibility that the secretory pathway may be regulated to influence the elaboration of dendrites. Such regulation might involve both genetic programs for specifying intrinsic differences in dendritic patterning of different neurons and activity-dependent modifications of dendritic arbors (Ye, 2007).
Several molecules are known to be specifically involved in dendritic but not axonal growth. Some of these molecules function in a neural activity-dependent fashion, such as the calcium/calmodulin-dependent protein kinase IIα (CaMKIIα), the transcription factor NeuroD, and the Ca2+-induced transcriptional activator CREST, while others are neural activity-independent such as bone morphogenetic protein 7 (BMP-7) and Dasm1. It will be important to find out whether the secretory pathway contributes to their regulation of dendritic growth (Ye, 2007).
In summary, this study has demonstrated that dendritic and axonal growth exhibit different sensitivity to changes in membrane supply from the secretory pathway. These findings raise a number of questions regarding membrane trafficking in dendrite and axon development. Answers to these questions will provide cell biological basis for understanding how the tremendous diversity of neuron morphology is achieved during development and how the changes in morphology happen in pathological conditions (Ye, 2007).
During development, dendrites arborize in a field several hundred folds of their soma size, a process regulated by intrinsic transcription program and cell adhesion molecule (CAM)-mediated interaction. However, underlying cellular machineries that govern distal higher-order dendrite extension remain largely unknown. This study shows that Numb-associated kinase (Nak), a clathrin adaptor-associated kinase, promotes higher-order dendrite growth through endocytosis. In nak mutants, both the number and length of higher-order dendrites are reduced; these characters phenocopied by disruptions of clathrin-mediated endocytosis. Nak interacts genetically with components of the endocytic pathway, colocalizes with clathrin puncta and is required for dendritic localization of clathrin puncta. More importantly, these Nak-containing clathrin structures preferentially localize to branching points and dendritic tips that are undergoing active growth. Evidence is presented that the Drosophila L1-CAM homolog Neuroglian is a relevant cargo of Nak-dependent internalization, suggesting that localized clathrin-mediated endocytosis of CAMs facilitates the extension of nearby higher-order dendrites (Yang, 2011; see video abstract).
Postmitotic neurons elaborate highly branched, tree-like
dendrites that display distinct patterns in accordance with their
input reception and integration. Therefore, regulation of dendrite
arborization during development is crucial for neuronal function
and physiology. Dendrite morphogenesis proceeds in two
main phases: lower-order dendrites first pioneer and delineate
the receptive field and then higher-order dendrites branch out
to fill in gaps between existing ones (Jan, 2010). This
process is exemplified by the morphogenesis of Drosophila
dendritic arborization (da) neurons, which have a roughly fixed
pattern of lower-order dendrites in early larval stages. Higher-order
dendrites then branch out to reach the order of more
than six, covering the entire epidermal area. These distinct phases of dendrite arborization are manifested by the difference in underlying cytoskeletal composition.
While lower-order dendrites are structurally supported by rigid
microtubules, higher-order dendrites contain actin and loosely
packed microtubules. It is thought
that the structural flexibility of higher-order dendrites allows
dynamic behaviors like extension, retraction, turning and stalling
to explore unfilled areas (Yang, 2011).
The da neurons are classified into four types (I-IV) according to
branching pattern and complexity of dendrites. The most complex class IV da neurons have a unique pattern, in which few branches are sent out from proximal
dendrites, while dendrites grow extensively in distal regions. Polarized growth of higher-order dendrites requires specialized cellular machineries. For instance, disruption
of the ER-to-Golgi transport in class IV ddaC neurons preferentially
shreds higher-order dendrites, suggesting that the
secretory pathway is needed to sustain membrane addition
during dendrite formation. Golgi outposts, hallmark
of the satellite secretory pathway in dendrites, move anterogradely
and retrogradely during extension and retraction of
terminal dendrites, respectively. Arborization in the distal field
demands active transport systems mediated by microtubule-based
motors, as mutations in dynein light intermediate chain
(dlic) or kinesin heavy chain (khc) fail to elaborate branches in
the distal region of class IV ddaC neurons. The transport of Rab5-positive endosomes
allows branching of distal dendrites, suggesting that the endocytic
pathway also has a role in dendrite morphogenesis (Yang, 2011).
The growth of higher-order dendrites seems to require
elevated level of endocytosis. Endocytosis is more active in dendrites
than in axons in cultured hippocampal neurons. Dynamic
assembly and disassembly of clathrin-positive structures, indicative
of active endocytosis, are seen at dendritic shafts and tips
of young hippocampal neurons. These clathrin-positive structures
become stabilized in mature neurons. Endocytosis is known to regulate the polarized distribution of the cell adhesion molecule NgCAM in hippocampal neurons,
which is first transported to the somatodendritic membrane
and then transcytosed to the axonal surface.
Endocytosis is also important for transporting NMDAR to
synaptic sites during their formation in dendrites of young cortical
neurons. The NMDAR packets transported along microtubules
are intermittently exposed to the membrane surface by cycles
of exocytosis and endocytosis, at sites coinciding with the clathrin
'hotspots'. Endocytosis can regulate the activities of transmembrane receptors whose signaling activity is important to dendrite growth and maintenance. For instance, the neurotrophin-Trk receptor-mediated signaling that depends on endocytosis could be importantfor dendrite morphogenesis. However,
how endocytosis regulates dendrite morphogenesis is not yet clear (Yang, 2011).
Clathrin-mediated endocytosis (CME) is the major route for
selectively internalizing extracellular molecules and transmembrane
proteins from the plasma membrane. Transmembrane cargos
destined for internalization are recruited into clathrin-coated
pits through interaction with appropriate clathrin adaptors. One
such accessory factor is adaptor protein 2 (AP2), a heterotetrameric
complex consisting of a, b, m and s subunits. AP2-dependent cargo recruitment can be
regulated by reversible protein phosphorylation by actin-related
kinase (Ark) family serine/threonine kinases. In yeast, Ark family genes are known to influence endocytosis by phosphorylating Pan1, an Eps15 homolog,
to regulate actin dynamics. Mammals contain two Ark family genes, cyclin G-associated kinase (GAK) and adaptor-associated kinase 1 (AAK1) and both have been
implicated in vesicular transport. GAK, best known for its role in the disassembly of clathrin coats from clathrin-coated vesicles, has multiple functions during clathrin cycle. AAK1 has been shown to bind the a subunit of AP2, phosphorylate the cargo-binding m2 subunit and promote receptor-mediated transferrin uptake. AAK1 also participates in transferrin receptor recycling from the early/sorting endosome in a kinase activity-dependent manner (Yang, 2011).
Numb-associated kinase (Nak), the Drosophila Ark family
member, contains the conserved Ark kinase domain and several
motifs (DPF, DLL and NPF) mediating interactions with endocytic
proteins. To study the function of Nak in development, nak deletion mutants and RNAi lines were generated and it was shown that depletion
of nak activity in da neurons disrupts higher-order dendrite
development. This function of Nak in dendritic morphogenesis
is likely mediated through CME, as Nak exhibits specific genetic
interactions with components of CME, colocalizes with clathrin
in dendritic puncta and is required for the presence of clathrin
puncta in distal higher-order dendrites. More importantly, live-imaging
analysis shows that the presence of these clathrin/Nak
puncta at basal branching sites correlates with extension of
terminal branches. In addition, evidence is presented that the
localization of Neuroglian (Nrg) in higher-order dendrites requires
Nak, implying that regional internalization of a cell adhesion
molecule is crucial for dendrite morphogenesis (Yang, 2011).
This study has shown that disruption of nak during dendrite arborization of da
neurons significantly reduces both number and length of dendritic
branches. Multiple classes of da neurons were analyzed
for the lack of Nak activity, which suggests that its general role
in higher-order dendrite morphogenesis. The function of Nak in
dendrite arborization is required cell autonomously, as dendritic
defects in nak mutants could be rescued by neuron-specific
expression of wild-type Nak (Yang, 2011).
Several lines of evidence suggest a functional link between
Nak and AP2, the endocytosis-specific clathrin adaptor, in
dendrite morphogenesis. First, coimmunoprecipitation results
show that Nak predominantly associates with AP2. Second,
Nak colocalizea well with GFP-Clc and alpha-adaptin but not with
AP1 and AP3 in S2 cells. Third, neuron-specific
depletion of AP2 mimics the dendritic defect in nak
mutants and reduction of AP2 gene dose enhances nak-induced
dendritic defect. These genetic interactions are specific,
as mutations in components of AP1 (AP47SAE-10) and AP3
(garnet1) showed no enhancement of nak-associated dendritic
phenotypes (Yang, 2011).
Mutations in Nak DPF motifs that are known to interact with
alpha-adaptin (DPF-to-AAA), reducing interaction with AP2, render
Nak incapable of rescuing the dendritic defects. As AP2 acts
to recruit clathrin to endocytic sites, this functional link between
Nak and AP2 implies that the dendritic defect in nak mutants is
caused by the disruption of Clathrin-mediated endocytosis (CME). Consistent with this notion, mutations in Chc also interact genetically with nak in dendrite
morphogenesis and Nak and clathrin are colocalized in
dendrites. Thus, it is suggested that Nak functions through CME
to promote dendrite development. Being an Ark family kinase
implicated in CME, Nak might function similarly to AAK1 that is
known to regulate the activities of clathrin adaptor proteins via
phosphorylation in cultured mammalian cells). Consistently, it was shown that Nak kinase activity is indispensable for its ability to rescue dendritic
defects. Disrupting dynamin activity in shits1-expressing neurons
exhibited stronger defects than nak mutants. In addition to endocytosis,
dynamin is known to act in the secretory pathway. Given the known role of the
secretory pathway in dendrite morphogenesis, it is possible
that only endocytic aspect is disrupted in nak mutants, but
both secretory and endocytic aspects are affected in shi mutants (Yang, 2011).
Clathrin- and Nak-positive structures in da neurons are preferentially localized to the branching points of higher-order dendrites. Unlike
Rab4, Rab5 and Rab11 that are mobile in dendrites, these clathrin/Nak puncta
are stationary. Importantly, it was possible to correlate the localization of these stationary clathrin/Nak puncta with motility of local terminal dendrites. The clathrin
puncta in higher-order dendrites probably represent sites where populations
of clathrin-coated vesicles actively participate in endocytosis. Consistent with this,
these clathrin-positive structures are enriched with PI4,5P2, which is known to
assemble endocytic factors functioning in the nucleation of clathrin-coated pits. The proximity and tight association between localized endocytic
machinery and polarized growth have been described in several systems,
including the extension of root hair tips, the budding of yeast cells and the navigation
of axonal growth cones. Thus, while the mechanism remains to be determined, the requirement of CME in cellular growth appears conserved (Yang, 2011).
How does regionalized endocytosis contribute to dendrite
branching? It is proposed that region-specific internalization and
recycling of the cell adhesion molecule Nrg is a mechanism for
generating local Nrg concentration optimized for higher-order
dendrite morphogenesis. In the advance of mammalian axonal
growth cones, adherent L1 can provide the tracking force for
growth cone extension. As the growth cone
advances, L1 is endocytosed in the central region to release
unnecessary adhesion and recycled back to the peripheral
region. Similarly, continuously recycling of Nrg along the dendritic
membrane may help its delivery to growing dendrites that
potentially function in promoting dendrite extension or stabilizing
newly formed dendrites. Excessive Nrg in higher-order dendrites
as in da neurons overexpressing Nrg may inhibit dendrite arborization
by generating superfluous adhesion. Thus, Nak-mediated
endocytosis could alleviate this inhibition by internalizing Nrg
from the cell surface, allowing dendrite elongation (Yang, 2011).
Arborization of higher-order dendrites in Drosophila da neurons
requires branching out new dendrites and elongation of existing
ones, which requires two other cellular machineries. First,
transporting the branch-promoting Rab5-positive organelles to
distal dendrites by the microtubule-based dynein transport
system is essential for branching activity. In the absence of Rab5 activity, dendritic
branching is largely eliminated and lacking the dynein transport
activity limits branching activity to proximal dendrites. Second,
the satellite secretory pathway contributes to dendrite growth
by mobilizing Golgi outposts to protruding dendrites. Similar to Rab proteins, the Golgi outposts labeled by ManII-GFP were only partially colocalized with YFP-Nak and their dendritic distribution is independent of Nak
activity. Also, in lva-RNAi larvae in which the transport
of Golgi outposts is disrupted, YFP-Nak puncta
were localized normally to distal dendrites. These
findings suggest that localization of Golgi outposts in dendrites
is not dependent on Nak activity and localization of YFP-Nak
is not dependent on transport of Golgi outposts. It is envisioned
that arborization of dendrites is achieved by transporting the
branch-promoting factors like Rab5 distally via the dynein transport
system. Following the initiation of new branches, dendrite
extension requires growth-promoting activity provided by the
anterograde Golgi outposts and localized clathrin puncta to
promote local growth. To actively distribute clathrin puncta in
distal dendrites that are far away from the soma, Nak can participate
in the condensation of efficient endocytosis into the punctate
structures in higher-order dendrites. It is possible that both
stationary Nak/clathrin puncta and secretory Golgi outposts are
spatially and temporally coupled to promote extension of dendrites,
thus coordinating several events like adhesion to the
extracellular matrix, membrane addition/extraction, cargo transport,
and cytoskeletal reorganization, eventually building up the
sensory tree in the target field (Yang, 2011).
Pruning of unnecessary axons and/or dendrites is crucial for maturation of the nervous system. However, little is known about cell adhesion molecules (CAMs) that control neuronal pruning. In Drosophila, dendritic arborization neurons, ddaCs, selectively prune their larval dendrites. This study reports that Rab5/ESCRT-mediated endocytic pathways are critical for dendrite pruning. Loss of Rab5 or ESCRT function leads to robust accumulation of the L1-type CAM Neuroglian (Nrg) on enlarged endosomes in ddaC neurons. Nrg is localized on endosomes in wild-type ddaC neurons and downregulated prior to dendrite pruning. Overexpression of Nrg alone is sufficient to inhibit dendrite pruning, whereas removal of Nrg causes precocious dendrite pruning. Epistasis experiments indicate that Rab5 and ESCRT restrain the inhibitory role of Nrg during dendrite pruning. Thus, this study demonstrates the cell-surface molecule that controls dendrite pruning and defines an important mechanism whereby sensory neurons, via endolysosomal pathway, downregulate the cell-surface molecule to trigger dendrite pruning (Zhang, 2014).
Endocytic pathways profoundly regulate turnover and homeostasis of various cell-surface adhesion proteins and guidance receptors in the developing nervous systems. Perturbation of endocytic pathways often leads to a variety of neurodegenerative diseases, such as frontotemporal dementia, amyotrophic lateral sclerosis, Alzheimer's disease, lysosomal storage diseases, and Niemann-Pick disease. In Drosophila, the endolysosomal pathway is activated in neighboring glia to engulf degenerating axon/dendrite fragments for their subsequent breakdown during pruning, suggesting a non-cell-autonomous role. This study reports that Rab5 and the ESCRT complexes, two key endocytic regulators, cell autonomously promote dendrite pruning in ddaC neurons. Consistent with these findings, the endocytic pathways also play a cell-autonomous role in axon pruning of MB γ neurons. How do Rab5/ESCRT-dependent endocytic pathways facilitate dendrite pruning in ddaC neurons at the cellular level? This study has identified a cell-surface adhesion protein, namely the L1-type CAM Nrg, as a target of Rab5/ESCRT-dependent endocytic pathways (Zhang, 2014).
Drosophila Nrg and the mammalian L1-type CAMs regulate axonal growth and guidanc, synaptic stability and function, and axon/dendrite morphogenesis. Mutations in the human L1 CAM gene have been reported to cause a broad spectrum of neuronal disorders. This study has identified the Drosophila L1-type CAM Nrg as the key cell-surface molecule that inhibits dendrite pruning in ddaC neurons. The extracellular domains of the L1-type CAMs can regulate cell-cell adhesion via homophilic and/or heterophilic interactions, whereas their intracellular domains can link the proteins with F-actin/spectrins to stabilize the cytoskeletal architecture. In C. elegans, a ligand-receptor complex of cell adhesion molecules containing the nematode Nrg homolog controls dendrite-substrate adhesion to stabilize and pattern dendritic arbors in certain sensory neurons. In Drosophila, Nrg-mediated cell adhesion plays an essential role in stabilizing synapse growth and maintenance at the larval neuromuscular junction. Likewise, Nrg may also mediate adhesion of the dendrites to their adjacent epidermis to stabilize the dendritic architecture in ddaC sensory neurons, whereas downregulation of Nrg may reduce dendritic adhesion/stability and result in disassembly of dendrites. Consistent with the potential adhesive role, structure-function analysis indicates that the ECD of Nrg is important for its function in stabilizing dendrite and/or inhibiting dendrite pruning in ddaC neurons. The fact that overexpression of the ICD-deleted Nrg protein partially rescued the nrg14 mutant phenotype suggests that, in addition to the adhesion function of the ECD, the ICD of Nrg may recruit cytoskeletal components to stabilize dendritic branches in ddaC neurons. The model is therefore favored that the adhesive role of Nrg is a potential mechanism for inhibiting pruning in ddaC sensory neurons (Zhang, 2014).
Another major class of CAMs, integrins, were shown to regulate dendrite-substrate interactions and anchor ddaC dendritic arbors to the extracellular matrix. However, unlike Nrg, integrins do not accumulate on enlarged endosomes in Rab5 or ESCRT ddaC neurons, implying that integrins are not regulated by Rab5/ESCRT-dependent endocytic pathways in ddaC neurons. Moreover, other cell-surface molecules Robo and N-Cad, albeit regulated by the endolysosomal pathway in motor neurons, photoreceptors, or sensory neurons, are dispensable for normal progression of dendrite pruning in ddaC neurons. Thus, this study highlights an important role of the L1-type CAM Nrg in inhibiting dendrite pruning of ddaC sensory neurons (Zhang, 2014).
Interestingly, loss of nrg function causes precocious dendrite pruning without affecting the axonal integrity and connectivity in ddaC neurons, underscoring a specific requirement of Nrg in stabilizing the dendrites, but not the axons. Downregulation of Nrg may reduce dendritic adhesive properties of ddaC sensory neurons and thereby make the dendritic architecture more susceptible to pruning. It is conceivable that Nrg-independent mechanisms may be utilized to protect the axonal structure from the pruning machinery in ddaC neurons. Moreover, both nrg loss of function and gain of function did not affect axon pruning in MB γ neurons (data not shown), further supporting the conclusion that Nrg plays a specific role in dendrite pruning in ddaC sensory neurons. Future studies may elucidate whether and how Nrg mediates its dendritic adhesive properties to inhibit dendrite pruning (Zhang, 2014).
In summary, this study shows that Rab5/ESCRT-dependent endocytic pathways facilitate dendrite pruning of ddaC neurons by downregulating the Drosophila L1-type CAM Nrg during early metamorphosis. This study also demonstrated the role of the cell-surface adhesion protein Nrg in inhibiting dendrite pruning in ddaC sensory neurons. Thus, this study opens the door for further studies of the functions of cell-surface molecules in the regulation of dendritic adhesion during neuronal remodeling (Zhang, 2014).
Localized signaling in neuronal dendrites requires tight spatial control of membrane composition. Upon initial synthesis, nascent secretory cargo in dendrites exits the endoplasmic reticulum (ER) from local zones of ER complexity that are spatially coupled to post-ER compartments. Although newly synthesized membrane proteins can be processed locally, the mechanisms that control the spatial range of secretory cargo transport in dendritic segments are unknown. This study, carried out in mammalian neuronal cell cultures, monitored the dynamics of nascent membrane proteins in dendritic post-ER compartments under regimes of low or increased neuronal activity. In response to activity blockade, post-ER carriers are highly mobile and are transported over long distances. Conversely, increasing synaptic activity dramatically restricts the spatial scale of post-ER trafficking along dendrites. This activity-induced confinement of secretory cargo requires site-specific phosphorylation of the kinesin motor Kif17 (see Drosophila KIF17) by Ca2+/calmodulin-dependent protein kinases (CaMK) (see for example Drosophila CaMKII). Thus, the length scales of early secretory trafficking in dendrites are tuned by activity-dependent regulation of microtubule-dependent transport (Hunus, 2014. PubMed ID: 24931613).
Dendrites lengthen by several orders of magnitude during neuronal development, but how membrane is allocated in dendrites to facilitate this growth remains unclear. This study reports that Ras opposite (Rop), the Drosophila ortholog of the key exocytosis regulator Munc18-1, is an essential factor mediating dendrite growth. Neurons with depleted Rop function exhibit reduced terminal dendrite outgrowth followed by primary dendrite degeneration, suggestive of differential requirements for exocytosis in the growth and maintenance of different dendritic compartments. Rop promotes dendrite growth together with the exocyst, an octameric protein complex involved in tethering vesicles to the plasma membrane, with Rop-exocyst complexes and exocytosis predominating in primary dendrites over terminal dendrites. By contrast, membrane-associated proteins readily diffuse from primary dendrites into terminals, but not in the reverse direction, suggesting that diffusion, rather than targeted exocytosis, supplies membranous material for terminal dendritic growth, revealing key differences in the distribution of materials to these expanding dendritic compartments (Peng, 2015).
Pruning, whereby neurons eliminate their exuberant neurites, is central
for the maturation of the nervous system. In Drosophila, sensory neurons, ddaCs, selectively prune their larval dendrites without
affecting their axons during metamorphosis. However, it is unknown whether
the secretory pathway plays a role in dendrite pruning. This study shows
that the small GTPase Arf1, an important regulator of secretory pathway, is specifically required for dendrite pruning of ddaC/D/E sensory neurons but dispensable for apoptosis of ddaF neurons. Analyses of the GTP and GDP-locked forms of Arf1 indicate that the cycling of Arf1 between GDP-bound and GTP-bound forms is essential for dendrite pruning. Sec71
was identified as a guanine nucleotide exchange factor for Arf1 that
preferentially interacts with its GDP-bound form. Like Arf1, Sec71 is also
important for dendrite pruning, but not apoptosis, of sensory neurons.
Arf1 and Sec71 are interdependent for their localizations on Golgi.
Finally, Sec71/Arf1-mediated trafficking process is a prerequisite for Rab5-dependent
endocytosis to facilitate endocytosis and degradation of the cell adhesion molecule Neuroglian (Wang, 2017).
In the developing nervous systems, neurons often extend excessive neurites and form superfluous connections at early stages. Subsequent removal of those exuberant or inappropriate neurites without causing the death of parental neurons, a process known as pruning, is crucial for the refinement of neural circuits at late developmental stages. Neuronal pruning is a conserved process widely occurring in both vertebrates and invertebrates. In vertebrates, many neurons in the neocortex, neuromuscular system and hippocampal dendate gyrus prune their unwanted neurites to control the proper wiring of the nervous systems. In invertebrates, such as Drosophila, the nervous systems undergo drastic remodelling during metamorphosis, a transition stage from a larva to an adult fly. In the central nervous system (CNS), mushroom body (MB) γ neurons prune their dorsal and medial axon branches as well as entire dendrites. In the peripheral nervous system (PNS), some dorsal dendritic arborization (da) neurons, ddaC, ddaD and ddaE, selectively eliminate their larval dendrites without affecting their axons, whereas ddaF neurons are apoptotic during early metamorphosis. The pruning event involves both local degeneration and retraction, resembling neurodegeneration associated with brain injury and neurodegenerative diseases. Thus, a complete understanding of cellular and molecular mechanisms of developmental pruning would shed some light on pathological neurodegeneration following neurological diseases and injury (Wang, 2017).
In Drosophila, ddaC sensory neurons have emerged as an attractive model system to elucidate the molecular and cellular mechanisms of dendrite-specific pruning during early metamorphosis. In response to the steroid-molting hormone 20-hydroxyecdysone (ecdysone)
at the late larval stage, ddaC neurons sever the proximal region of their dendrites and subsequently undergo rapid fragmentation of the severed dendrites as well as dendritic clearance via phagocytosis. It has been well documented that the Ecdysone Receptor and its co-receptor Ultraspiracle are required to activate the expression of several downstream targets to initiate dendrite pruning. Endocytic pathways have been identified that are critical for dendrite pruning. Rab5/Avalanche and ESCRT complexes, the components of the endocytic pathways, are required for downregulation of the L1-type cell adhesion molecule (L1-CAM) Neuroglian (Nrg). Nrg is drastically redistributed to endosomes and its protein levels are strongly downregulated prior to pruning, suggesting that massive Nrg endocytosis promotes dendrite pruning. It is conceivable that Nrg endocytosis might be triggered by secreted ligands/signals through the secretory pathway. However, it is completely unknown whether the secretory pathway, an opposite route of the endocytic pathway, also plays a role in dendrite pruning of ddaC neurons (Wang, 2017).
The primary sites of the secretory pathway consist of the endoplasmic reticulum (ER), the Golgi apparatus and the trans-Golgi network in eukaryotic cells. Newly synthesized membrane proteins and lipids exit from the ER, pass through the Golgi complexes and are delivered to the plasma membrane via the post-Golgi exocytosis. In developing neurons, the continuous addition of membrane proteins and lipids via the secretory pathway plays a key role in the outgrowth and elongation of dendrites and axons. Disruption of the ER-to-Golgi transport leads to the inhibition of dendritic or axonal growth in Drosophila sensory neurons and rodent hippocampal neurons. In an attempt to isolate novel players of dendrite pruning, a large-scale RNA interference (RNAi) screen was carried out, and ADP-ribosylation factor 1(Arf1), also known as ADP-ribosylation factor at 79F, was identified as an important player for dendrite pruning in ddaC sensory neurons. Arf1 is a small GTPase and belongs to the Class I Arf family. It has been reported that Arf1 can recruit COPI coat proteins on cis-Golgi and clathrin adaptor proteins, such as AP-1, AP-3, and GGAs, on trans-Golgi in a GTP- dependent manner, and thereby facilitate vesicle formation and trafficking. Studies from yeast and mammals indicate that Arf1 is activated by two conserved families of guanine nucleotide exchange factors (GEFs), including the Gea1/GBF1 family on cis-Golgi and Sec7/BIG family on trans-Golgi. Arf1 cycles between GDP- and GTP-bound forms, and both the GTP- and GDP-locked forms can interfere with its functions and disrupt secretory trafficking. In mammalian hippocampal neurons, overexpression of the GTP-locked form of Arf1 (Arf1Q71L), which abolishes Arf1 activity, inhibits dendrite growth. The mammalian Arf1GEF, BIG2, is required for vesicle trafficking and mutations in human BIG2 gene lead to autosomal recessive periventricular heterotopia with microcephaly (ARPHM), a brain disorder characterized by defective neural proliferation and migration. Thus, various studies have documented that the secretory pathway plays a critical role in neurite growth and extension in developing neurons. However, very little is known about its role in regulating neurite pruning, a developmental degenerative process (Wang, 2017).
This study reports the identification of Arf1 as an important player of dendrite pruning in ddaC sensory neurons. The cycling of Arf1 between GDP-bound and GTP-bound forms is essential for dendrite pruning. Sec71 was identified as a GEF for Arf1 in Drosophila. Sec71, like Arf1, is cell-autonomously required for dendrite pruning of ddaC/D/E sensory neurons but not for apoptosis of ddaF neurons during early metamorphosis. Arf1 and Sec71 co-localize on Golgi apparatus and regulate secretory vesicle
biogenesis in ddaC neurons. Furthermore, it was shown that Sec71/Arf1-dependent secretory pathway acts upstream of Rab5-dependent endocytosis and facilitates the internalization and downregulation of the cell adhesion molecule Nrg prior to dendrite pruning. Thus, this study demonstrates a novel and important role of Arf1/Sec71-mediated secretory pathway in promoting developmental pruning via the regulation of Nrg endocytosis (Wang, 2017).
The small G protein Arf1 regulates vesicular trafficking in eukaryotes and is activated on cis-Golgi by the Gea1/GBF1 family of Arf1GEF or on trans-Golgi by the Sec7/BIG1 family . It has been reported that Drosophila Arf1 regulates hematopoietic niche maintenance, blood cell precursor differentiation in vivo, planar cell polarity, and lamellipodium formation in S2 cells. Arf1 and other Golgi proteins were reported to exhibit upregulation of their transcripts in axon pruning of MB γ neurons during the larval-pupal transition. This study reports an important role of Arf1 in regulating dendrite pruning of ddaC sensory neurons. Arf1 puncta overlap with the trans- Golgi marker GalT and partially with the cis-Golgi marker GM130, suggesting that Arf1 is primarily localized on trans-Golgi in ddaC sensory neurons. A specific GEF for Arf1 has not been identified in Drosophila. In vivo and in vitro data provide compelling evidence that Sec71 is a GEF for Arf1 in Drosophila. First, Sec71 was co-localized with Arf1 on Golgi and their localizations were inter-dependent in ddaC neurons. Second, Sec71 preferentially bound to the GDP-bound form of Arf1 instead of GTP-bound form. Third, in the GEF assays Sec71 accelerated the release of GDP from Arf1. Fourth, both Arf1 and Sec71 are required for dendrite pruning, as loss of Sec71 or Arf1 led to comparable pruning defects in ddaC sensory neurons. Finally, the expression of Arf1 fully restored WP dendrite morphology and rescued the pruning defects in Sec71 RNAi ddaC neurons. Thus, Sec71 is specifically required for the GDP-to-GTP exchange of Arf1.
Structure-function analyses indicate that DCB domain of Sec71 is important for its targeting on Golgi. In contrast, another small GTPase Arl1 was reported to bind to the N-terminal region of Sec71 (DCB and HUS1 domains) and recruit Sec71 on trans-Golgi apparatus in Drosophila S2 cells, and mammalian Arl1 is required for the recruitment of BIG1/2 (mammalian homologues of Sec71) on trans-Golgi (Wang, 2017).
Secretory pathway plays a novel and important role in governing neurite pruning
Extensive studies have attempted to understand roles of post-Golgi trafficking in outgrowth and elaboration of dendrites in growing neurons. Post-Golgi trafficking is polarized toward apical dendrites of rodent hippocampal neurons and selectively regulates the growth of dendrites. The dynamics of the Golgi outposts, mediated by the Golgin Lava Lamp, dynein-dynactin complex and Leucine- rich repeat kinase (Lrrk), is important for dendrite growth in Drosophila class IV da neurons. A small GTPase Rab10, which mediates post-Golgi vesicle trafficking, regulates dendrite growth and branching of multi-dendritic sensory neurons in both C. elegans and Drosophila (Wang, 2017).
This study provides compelling evidence to demonstrate that post-Golgi trafficking plays a crucial role in proper dendrite pruning in sensory neurons. First, the key small GTPase Arf1 was identified that is important for post-Golgi trafficking regulates secretory vesicle biogenesis and dendrite pruning in sensory neurons during metamorphosis. Second, a Sec7-domain-containing protein Sec71 acts as a specific GEF for Arf1 and co- localizes with Arf1. Like Arf1, Sec71 is also an essential factor for regulating dendrite pruning. Third, given that both Arf1 and Sec71 also regulate dendrite growth and arborization in ddaC neurons, a critical role of Arf1 and Sec71 in dendrite pruning was further controlled
using the Gene-Switch system. Pulse induction of Arf1T31N or Sec71DN at the middle third instar larval stage when the complete larval dendrite arbors form in ddaC neurons consistently caused much more severe dendrite pruning defects. These results highlight that the secretory pathway play separable roles in two distinct processes, namely dendrite growth and dendrite pruning. Arf1 was reported to regulate post-Golgi secretion by recruiting its downstream effectors, including the clathrin adaptors AP-1 and AP-3, and GGA. Post-Golgi trafficking pathways include the transport from Golgi to plasma membrane (exocyst complex-mediated), from Golgi to early/sorting endosomes (AP-1-meditated), from Golgi to late endosomes (Golgi-localized Gamma-ear containing Arf-binding protein or GGA-mediated) as well as from Golgi to lysosomes (AP-3-mediated). It is conceivable that at least one of these post-Golgi trafficking routes is involved in dendrite pruning of sensory neurons (Wang, 2017).
It has been reported previously that Rab5/ESCRT-dependent endocytic pathways facilitate dendrite pruning by downregulating the L1-CAM Nrg in ddaC neurons during metamorphosis. In MB γ neurons, the JNK pathway promotes axon pruning by downregulating another adhesion molecule Fasciclin II. These studies suggest a general mechanism whereby cell adhesion molecules are internalized and downregulated to destabilize dendrites and/or axons during neurite pruning. However, the mechanism that triggers Nrg endocytosis is poorly understood. This study demonstrated that Arf1/Sec71-mediated secretory pathway promotes endocytosis and downregulates Nrg prior to dendrite pruning. First, while Nrg levels were strongly reduced prior to dendrite pruning, loss of Arf1 or Sec71 led to elevated levels of Nrg protein in dendrites, axons and soma, comparable to Rab5 mutant neurons. Second, Nrg was no longer redistributed on FYVE-positive endosomes in Arf1 or Sec71 mutant ddaC neurons, suggesting a blockage of Nrg endocytosis. Third, while Rab5 mutant neurons exhibited robust ubiquinated protein aggregates and enlarged endosomes, further removal of either Arf1 or Sec71 suppressed these rab5 mutant phenotypes, suggesting that the secretory pathway acts upstream of Rab5 to positively regulates endocytosis. Finally, knockdown of Nrg significantly suppressed the dendrite pruning defect in Arf1 or Sec71 mutant neurons, supporting the notion that the secretory pathway promotes Nrg endocytosis and downregulation. Thus, the secretory pathway not only secretes the cell adhesion molecules to the dendrite surface and stabilize dendrites, but also unexpectedly promotes the internalization and turnover of the adhesion molecules. It is conceivable that in response to ecdysone pulse, the secretory pathway might be required to specifically secrete an as-yet-unidentified ligand to trigger massive endocytosis of the L1-CAM Nrg and thereby leads to degeneration of larval dendrites. Further studies may continue to elucidate what ligand or secreted protein promotes Nrg endocytosis (Wang, 2017).
Dendrite development is crucial in the formation of functional neural networks. Recent studies have provided insights into the involvement of secretory transport in dendritogenesis, raising the question of how the secretory pathway is controlled to direct dendritic elaboration. This study identified a functional link between transcriptional regulatory programs and the COPII secretory machinery in driving dendrite morphogenesis in Drosophila dendritic arborization (da) sensory neurons. MARCM analyses and gain-of-function studies reveal cell-autonomous requirements for the COPII coat protein Sec31 in mediating da neuron dendritic homeostasis. The homeodomain protein Cut transcriptionally regulates Sec31 in addition to other components of COPII secretory transport, to promote dendrite elaboration, accompanied by increased satellite secretory endoplasmic reticulum (ER) and Golgi outposts primarily localized to dendritic branch points. A novel functional role was established for the transcription factor CrebA in regulating dendrite development ; Cut initiates a gene expression cascade through CrebA that coordinately affects the COPII machinery to mediate dendritic morphology (Iyer, 2013).
Dendrite aberration is a common feature of neurodegenerative diseases caused by protein toxicity, but the underlying mechanisms remain largely elusive. This study shows that nuclear polyglutamine (polyQ) toxicity resulted in defective terminal dendrite elongation accompanied by a loss of Golgi outposts (GOPs) and a decreased supply of plasma membrane (PM) in Drosophila class IV dendritic arborization (da) (C4 da) neurons. mRNA sequencing revealed that genes downregulated by polyQ proteins included many secretory pathway-related genes, including COPII genes regulating GOP synthesis. Transcription factor enrichment analysis identified CREB3L1/CrebA, which regulates COPII gene expression. CrebA overexpression in C4 da neurons restores the dysregulation of COPII genes, GOP synthesis, and PM supply. Chromatin immunoprecipitation (ChIP)-PCR revealed that CrebA expression is regulated by CREB-binding protein (CBP), which is sequestered by polyQ proteins. Furthermore, co-overexpression of CrebA and Rac1 synergistically restores the polyQ-induced dendrite pathology. Collectively, these results suggest that GOPs impaired by polyQ proteins contribute to dendrite pathology through the CBP-CrebA-COPII pathway (Chung, 2017).
Neurons have tremendously higher amounts of plasma membrane (PM) than other cell types, due to their highly elongated morphology. Recently, dendritic satellite organelles have been reported to be involved in the maintenance of integrity and dynamics of the PM in distal dendrites out of reach from somatic perinuclear organelles. The dendritic organelles have functional overlaps with the canonical perinuclear organelles. However, the restricted space appears to require the miniaturization of the satellite organelle systems, such as the endoplasmic reticulum (ER) and Golgi outposts (GOPs), compared to those localized around the nucleus, suggesting that the dendritic organelles may show different functional capacity from their perinuclear counterparts. Additionally, the distal areas of the dendrites may have quite different cellular environments, such as variations in the concentration of ions and proteins, compared to the perinuclear region. These differences may account for the vulnerability of dendrites over other neuronal domains to neuropathological insults, including protein toxicity conferred by the accumulation of mutant or misfolded proteins in neurons (Chung, 2017).
In Drosophila dendritic arborization (da) sensory neurons, GOPs were found to be located in dendrites, but not in axons, suggesting that GOPs may act as local stations to supply membranes to the PM in nearby dendrites (Ye, 2007). The synthesis of GOPs was reported to be regulated by secretory pathway-related genes, such as Sec31 and the transcription factor Cut (Iyer, 2013). Additionally, GOPs were shown to be transported toward or away from the soma by machinery consisting of a lava lamp protein (LVA), a golgin coiled-coil adaptor protein, and a motor dynein-dynactin protein complex, and their transport was found to be closely associated with the dynamics of dendrite growth. Moreover, GOPs have been implicated in the growth and maintenance of the dendrite arbor. For example, laser ablation of local GOPs decreased branch dynamics in da neurons (Ye, 2007), and the disruption of GOP trafficking blocked dendrite growth in developing hippocampal neurons, leading to the decreased length of dendrites (Chung, 2017).
Despite such functional roles of the GOPs in the growth and maintenance of the dendrite arbor under non-pathological conditions, it still remains unclear whether the GOPs can also contribute to dendrite aberration observed under pathological conditions induced by protein toxicity. Previously, nuclear proteotoxicity has been reported to cause severe dendrite pathology, but the mechanisms underlying dendrite pathology caused by protein toxicity remain largely elusive.
Therefore this study investigated a mechanistic link of the GOPs to dendrite pathology caused by toxic nuclear polyQ proteins using Drosophila da sensory neurons that have been extensively utilized by various researchers as an in vivo cellular model for the study of the mammalian neuronal dendrite system with high fidelity dendrite pathology (Chung, 2017).
Toxic nuclear polyQ proteins cause terminal dendrite defects in C4 da neurons. This study examined morphological characteristics of dendrites of class IV (C4) da neurons expressing pathogenic Machado-Joseph's Disease (MJD) protein, MJDtr-78Q (78Q), known to induce nuclear protein toxicity. For this analysis, dendrite images of C4 da neurons expressing 78Q (78Q OE) and wild-type C4 da neuron controls were compared. A mechanistic model is proposed that links toxic nuclear polyQ proteins to the impaired elongation of terminal dendrites in 78Q OE through the CBP-CrebA-COPII-GOP pathway. Toxic nuclear polyQ proteins decreased the number of GOPs in the dendrites of C4 da neurons. mRNA sequencing revealed that the secretory pathway, including the COPII pathway, was a major pathway that was downregulated by toxic polyQ proteins. The disruption of the COPII pathway decreased GOP synthesis. TF enrichment analysis identified CrebA as a key regulator of the genes involved in the COPII pathway. CrebA overexpression restored the downregulation of the COPII pathway and the loss of GOPs caused by toxic polyQ proteins. It was further found that CBP regulated CrebA expression cooperatively with Cut and that toxic polyQ proteins interfered with CBP upstream of CrebA in the regulation of GOP synthesis. Finally, the co-overexpression of CrebA and Rac1 synergistically restored polyQ-induced defects in dendrite branching and elongation (Chung, 2017).
According to a mechanistic model, however, how the overexpression of Cut, unlike CBP, can potentially contribute to the restoration of GOPs is unclear, although it cooperatively regulates CrebA expression with CBP and interacts with CBP at the protein level. This can be explained by alterations in the stoichiometry between CBP and Cut for their complex formation under polyQ-expressed conditions. The results showed that polyQ proteins appeared to sequester CBP, but not Cut. The sequestration of CBP by polyQ proteins leads to a decreased amount of CBP available for the CBP-Cut complex formation. Accordingly, CBP overexpression can increase the amount of non-sequestered CBP under polyQ-expressed conditions, thereby restoring the CBP-Cut complex formation. By contrast, Cut overexpression does not contribute to an increase in complex formation because it has no effect on the depletion of CBP by polyQ proteins. Moreover, Rac1 was included as a component in the mechanistic model. Rac1 was connected to the polyQ-CBP-CrebA pathway based on the following results. First whether polyQ expression affected the expression of Rac1 was tested and a significant decrease in Rac1 expression was found in polyQ-expressing fly heads compared to w1118. Then TFs were sought that could be affected by polyQ expression and could regulate Rac1 expression. Previously, Cut was shown to regulate Rac1 expression. Thus, to examine the effect of polyQ proteins on Cut, whether polyQ proteins altered the intracellular distribution of Cut proteins was checked in polyQ-expressing da neurons, and no noticeable alteration was found in its distribution. As previously mentioned, Cut interacted with CBP in fly heads. Furthermore, Rac1 was shown to be epigenetically suppressed by H3K27me3, which can be antagonized by CBP-dependent H3K27ac. These data together provide a mechanistic connection of the CBP-Cut-Rac1 pathway to the polyQ-CBP-CrebA pathway through CBP (Chung, 2017).
This study proposed a polyQ-CBP-CrebA pathway for polyQ-induced terminal dendrite defects. However, an alternative possibility that dendrite perturbation may precede through CBP-CrebA-independent pathways, which then can result in altered gene expression, cannot be excluded. Consistent with such a possibility, nuclear polyQ proteins were shown to sequester nuclear pore complex proteins, potentially impairing the transport of proteins and RNA, independently of CBP or CrebA (Grima, 2017). Moreover, nuclear polyQ proteotoxicity induces the mis-localization of several TFs (e.g., TBP, Sp1, and p53) other than CBP, leading to the altered expression of their target genes. The perturbed functions of nuclear pore complexes and TFs may result in dendrite defects via CBP-CrebA-independent mechanisms and consequently alterations in gene expression. However, the restoration of GOP synthesis and PM protein supply via the overexpression of CBP or CrebA and the restoration of dendritic arborization defects via the co-overexpression of CrebA and Rac1 in polyQ-expressing neurons demonstrates that the CBP-CrebA pathway should be one of the important pathways that functionally regulates the polyQ-induced dendrite defects. Nevertheless, the relative importance of the CBP-CrebA pathway to the pathways that include other TFs affected by polyQ proteins can be questioned. However, this study found that the overexpression of the three key TFs (HLH106, gt, and Dref) did not restore the number of GOPs in polyQ-expressing neurons. Moreover, the effect of Sp1, which is known to be downregulated by polyQ proteins, on GOP synthesis was examined, and Sp1 knockdown did not significantly decrease the number of GOPs compared to the controls. Furthermore, it was also found that the overexpression of Cut and Scr, known upstream TFs of CrebA, failed to restore the loss of GOPs in polyQ-expressing neurons comparable to CrebA overexpression. Therefore, the data suggest that the mechanism by which polyQ toxicity induces dendrite pathology can be attributed to the polyQ-CBP-CrebA pathway proposed in this study (Chung, 2017).
The loss of GOPs was one of the most apparent dendrite defects caused by polyQ proteins. Thus, among many cellular processes downregulated by the polyQ proteins, this study focused on three cellular processes, (1) vesicle-mediated transport (VT) and (2) membrane organization (MO) and lipid biosynthetic process (LP), that were considered to be related to GOPs. However, these processes can also be related to other secretory-pathway-related processes in addition to GOP synthesis. For example, neuropeptide maturation and trafficking were previously shown to be modulated by the secretory pathway, suggesting the potential association of neuropeptide-containing vesicle trafficking with the three processes. Thus the effect of polyQ proteins on neuropeptide-containing vesicle trafficking was examined by measuring the amount of neuropeptide-containing vesicles in w1118 and polyQ-expressing neurons using a reporter. PolyQ expression was found to decrease the amount of ANF-EMD vesicles in dendrites and soma compared to controls, suggesting that polyQ perturbs the function of the secretory pathway related to neuropeptide-containing vesicle trafficking in addition to GOP synthesis. Additionally, the three processes can be associated with the synthesis of somatic Golgi. However, the data showed a larger loss of ManII-eGFP-positive puncta in the dendrites of C4 da neurons compared to the soma, suggesting that the three processes can be considered to be related more to GOP synthesis than to somatic Golgi synthesis. Thus, although the polyQ-induced downregulation of genes associated with the abovementioned three processes may impact the secretory pathway in general, the results appear to indicate that the synthesis of GOPs responds more to the downregulation of those genes compared to the synthesis of somatic Golgi (Chung, 2017).
In mRNA-sequencing analysis, fly heads were used to identify genes affected by toxic polyQ proteins. Considering that fly heads contain no C4 da neurons, alterations in gene expression obtained from mRNA sequencing in polyQ-expressing fly heads, compared to w1118 fly heads, should be defined by non-C4 da cells. Due to a small number (~50) of C4 da neurons per larva, however, it is challenging to isolate a sufficient number (>105 cells) of C4 da neurons for mRNA sequencing. Instead of C4 da neurons, fly heads were used for mRNA sequencing to obtain a clue for alteration of gene expression by polyQ proteins. Moreover, mRNA sequencing identified 5,385 differentially expressed genes between polyQ-expressed and control samples, of which 5,325 (98.9%) were downregulated. Such a large number and high percentage of downregulated genes suggests that there might be widespread cell death in polyQ-expressing fly brains. Thus, the extent of cell loss was checked in the polyQ-expressing adult fly brains on which mRNA sequencing was performed by measuring the number of cells that displayed cleaved caspase-3 through immunohistochemistry analysis. PolyQ-expressing brains showed minimal cell loss compared to controls in the condition where mRNA sequencing was performed. Additionally, no observable decrease was found in rRNA in polyQ-expressed adult fly heads compared to w1118 heads. These data suggest that the downregulation of the large number of genes is not likely to be caused by cell death. One alternative explanation can be the previously reported dysregulation of the following histone modifications caused by polyQ proteins: impaired acetylation of H3K27 and dysregulated tri-methylation of H3K9 and H3K27. These dysregulated histone modifications may result in the downregulation of 98.9% of DEGs through a global epigenetic silencing (Chung, 2017).
The normal length of the polyQ tract in humans has been reported to be less than 36 for the MJD protein. In flies, numerous studies have used the truncated MJD protein with only 27 polyQ repeats (MJDtr-27Q) as a control. However, in this study, w1118 rather than 27Q OE was used as the control for 78Q OE in most of the experiments because 27Q OE exhibited strong dendrite pruning defects compared to 78Q OE. A previous study also showed that Httex1p-Q93 caused strong pruning defects in C4 da neurons, whereas Httex1p-Q20 caused no such defects. Taken together, these data suggest that the pruning defects are likely not due to the context of the host proteins but to polyQ toxicity. However, there appears to be a threshold length (between 20Q and 27Q) of the polyQ tract to confer such polyQ toxicity. Nevertheless, further detailed studies should be performed to understand the mechanism underlying the protein toxicity of MJDtr-27Q to induce pruning defects (Chung, 2017).
Finally, this study identified three different nuclear polyQ toxicity models that exhibited decreased GOP synthesis and PM protein supply, indicating a potentially shared mode of nuclear proteotoxicity. Of note, the accumulation of misfolded proteins in the nucleus has been observed in other neuronal diseases or during aging. Thus, the proposed mechanistic model underlying terminal dendrite defects caused by the accumulation of nuclear toxic polyQ proteins can provide significant insights into other neuronal maladies linked to nuclear proteotoxicity and aging that can be further examined in detailed functional studies (Chung, 2017).
Neural activity has profound effects on the development of dendritic structure. Mechanisms that link neural activity to nuclear gene expression include activity-regulated factors, such as CREB, Crest (Ca2+-responsive transactivator, a syntaxin-related nuclear protein that interacts with CREB-binding protein and is expressed in the developing brain) or Mef2, as well as activity-regulated immediate-early genes, such as fos and jun. This study investigates the role of the transcriptional regulator AP-1, a Fos-Jun heterodimer, in activity-dependent dendritic structure development. Genetic manipulation, imaging and quantitative dendritic architecture analysis were combined in a Drosophila single neuron model, the individually identified motoneuron MN5. First, Dalpha7 nicotinic acetylcholine receptors (nAChRs) and AP-1 are required for normal MN5 dendritic growth. Second, AP-1 functions downstream of activity during MN5 dendritic growth. Third, using a newly engineered AP-1 reporter it was demonstrated that AP-1 transcriptional activity is downstream of Dalpha7 nAChRs and Calcium/calmodulin-dependent protein kinase II (CaMKII) signaling. Fourth, AP-1 can have opposite effects on dendritic development, depending on the timing of activation. Enhancing excitability or AP-1 activity after MN5 cholinergic synapses and primary dendrites have formed causes dendritic branching, whereas premature AP-1 expression or induced activity prior to excitatory synapse formation disrupts dendritic growth. Finally, AP-1 transcriptional activity and dendritic growth are affected by MN5 firing only during development but not in the adult. These results highlight the importance of timing in the growth and plasticity of neuronal dendrites by defining a developmental period of activity-dependent AP-1 induction that is temporally locked to cholinergic synapse formation and dendritic refinement, thus significantly refining prior models derived from chronic expression studies (Vonhoff, 2013).
By combining genetic and neuroanatomical tools with imaging in a single-cell model, the adult MN5 in Drosophila, this study demonstrates that: (1) AP-1 is transcriptionally active during all stages of postembryonic motoneuron dendritic growth, (2) AP-1 and excitatory cholinergic inputs are required for normal dendrite growth in MN5, (3) AP-1 transcriptional activity is enhanced via a CaMKII-dependent mechanism by increased neural activity during pupal development but not in the adult, and (4) both activity and AP-1 can promote or inhibit dendritic branching, depending on the developmental stage.
AP-1 is required for normal MN5 dendrite growth downstream of activity and CaMKII (Vonhoff, 2013).
Although AP-1 has been thought to regulate dendrite development in an activity-dependent manner via global changes in gene expression, probably in a calcium-dependent manner as described for CREB or Crest, direct evidence for this hypothesis was sparse (Vonhoff, 2013).
This study demonstrated that excitatory cholinergic input to MN5 and AP-1 transcriptional activity were required for normal dendrite growth of MN5 during pupal life. MN5 total dendritic length and branch numbers were significantly reduced (~50%) by inhibition of AP-1 [by Jbz (a dominant-negative form of Jun) expression] and in Dα nAChR mutants. Conversely, overexpression of AP-1 or increased MN5 excitability as induced by potassium channel knockdown (by EKI) increased dendritic branching (Duch, 2008). Clearly, AP-1 acted downstream of activity as inhibition of AP-1 by Jbz completely attenuated EKI (electrical knock-in) mediated dendritic growth and branching (Vonhoff, 2013).
A new AP-1 reporter was employed to measure activity-induced AP-1 transcriptional activity by imaging, and to gain insight into the pathway that might connect MN5 activity to AP-1-dependent transcription. Although the detection threshold of this reporter might be too low to detect small changes in AP-1 activity, sensitivity was sufficient to reliably report increased AP-1 activity following overexpression of fos and jun, inhibition of AP-1 transcriptional activity by Jbz expression, and changes in AP-1 activity as induced by various manipulations of cellular signaling. Therefore, the reporter was deemed suitable for testing changes in AP-1 transcriptional activity in MN5 (Vonhoff, 2013).
Targeted expression of TrpA1 channels in MN5 allowed the induction of firing in vivo by temperature shifts during selected developmental periods. Activation of MN5 during pupal life for 36 hours (P9 to adult) or longer (P5 to adult) caused significant increases in AP-1-induced nuclear GFP fluorescence. By contrast, in adults neither similar nor longer durations of TrpA1 activation resulted in any detectable increase in AP-1 reporter-mediated nuclear GFP fluorescence in MN5. Similarly, live imaging in semi-intact adult preparations did not reveal any detectable AP-1 activity upon acute TrpA1 activation for various durations. This indicated that activity-dependent AP-1 activation was restricted to pupal life. However, whether AP-1 activation in the adult MN5 occurred upon patterned activity was not tested. Spaced stimuli that reflect endogenous activity patterns are required for insect motoneuron axonal and dendritic development and can regulate mammalian neuron dendritic morphology. However, during flight, MN5 fires tonically at frequencies between 5 and 20 Hz, a pattern that is well reflected by temperature-controlled TrpA1 channel activation. Therefore, adult flight behavior is unlikely to induce AP-1 activity, which is involved in dendrite and synapse development (Freeman, 2010). This is consistent with the assumption that dendritic structure is fairly stable in the adult (Vonhoff, 2013).
cAMP and Jun N-terminal kinase (Jnk) have been implicated as potential links between activity and AP-1 activation. Cell culture studies on Drosophila larval motoneurons and giant neurons demonstrate a role of calcium. This study showed that Dα7 nAChRs, which are highly permeable to calcium, were required for normal MN5 dendritic growth. Combining genetic manipulation of Dα7 nAChRs, AP-1 and CaMKII with imaging of AP-1 reporter activity revealed that CaMKII was required downstream of Dα7 nAChRs to cause AP-1-dependent transcription. These data show that activity-dependent calcium influx through nAChRs might activate AP-1 during pupal life via a CaMKII-dependent mechanism in vivo.
Activity and AP-1 can promote or inhibit dendritic growth during pupal life, depending on timing (Vonhoff, 2013).
In larval motoneurons, AP-1 is required for dendritic overgrowth as induced by artificially increased activity (Hartwig, 2008). In MN5, AP-1 is required downstream of nAChRs and CaMKII for normal dendritic growth. By contrast, premature expression of AP-1 in MN5 inhibited dendritic growth. These data were consistent with the hypothesis that timing is the crucial factor. First, P103.3 and D42 both caused similar overgrowth but exhibited fairly different expression patterns. Second, C380-GAL4 and Dα7 nAChR-GAL4 both inhibited MN5 dendrite growth but expressed in largely different sets of neurons. Therefore, the common factor of C380 and Dα7 nAChR on the one hand and D42 and P103.3 on the other hand was timing. Third, shifting the timing of C380-GAL4-driven AP-1 expression to later stages prevented dendritic defects. Fourth, imposed activity prior to P5 by TrpA1 activation also inhibited dendritic branching. Dendritic defects as induced by imposed premature activity were rescued by inhibition of AP-1 via Jbz expression in MN5 (Vonhoff, 2013).
MN5 early dendritic growth starts at early pupal stage 5 (P5), and expression of Dα7 nAChRs begins 2.5 hours later, at mid stage P5. Similarly, Xenopus optic tectal and turtle cortical neurons receive glutamatergic and GABAergic inputs as soon as the first dendrites are formed. In vertebrates, early synaptic inputs and neurotransmitters play essential roles in dendrite development. The current data are consistent with the hypothesis that the endogenous expression of nAChRs caused increased activity throughout the developing motor networks, which, in turn, upregulated AP-1-dependent transcription and dendritic growth via a CaMKII-dependent mechanism. During zebrafish spinal cord development, activity is required for strengthening functional central pattern generator (CPG) connectivity. As dendrites are the seats of input synapses to motoneurons, an activity-dependent component in motoneuron dendritic growth that follows early synaptogenesis might function to refine dendrite shape during the integration into the developing CPG (Vonhoff, 2013).
As animals grow, their nervous systems also increase in size. How growth in the central nervous system is regulated and its functional consequences are incompletely understood. These questions were explored using the larval Drosophila locomotor system as a model. In the periphery, at neuromuscular junctions, motoneurons are known to enlarge their presynaptic axon terminals in size and strength, thereby compensating for reductions in muscle excitability that are associated with increases in muscle size. This work examined how motoneurons change in the central nervous system during periods of animal growth. I was found that within the central nervous system motoneurons also enlarge their postsynaptic dendritic arbors, by the net addition of branches, and that these scale with overall animal size. This dendritic growth is gated on a cell-by-cell basis by a specific isoform of the steroid hormone receptor ecdysone receptor-B2, for which functions have thus far remained elusive. The dendritic growth is accompanied by synaptic strengthening and results in increased neuronal activity. Electrical properties of these neurons, however, are independent of ecdysone receptor-B2 regulation. It is proposed that these structural dendritic changes in the central nervous system, which regulate neuronal activity, constitute an additional part of the adaptive response of the locomotor system to increases in body and muscle size as the animal grows (Zwart, 2013).
The implications of these observations are twofold: First,
neurons can deploy structural changes in their dendritic trees as
a central mechanism with which to regulate and adjust levels of
neuronal activity; second, in terms of connectivity, the size of the
postsynaptic dendritic arbor seems to be decisive in determining
the number of connections that neurons form among available
presynaptic terminals (Zwart, 2013).
Increases in body or organ size
are normally accompanied by matching changes in innervation,
required to maintain appropriate neuronal control. One of the
best-studied examples is the neuromuscular junction where
increases in muscle size lead to biophysical changes in muscle
physiology, which are compensated for by a matching enlargement
of the neuromuscular junction. The neural
and cellular mechanisms that regulate these homeostatic adjustments
have been studied in detail. This study has identified a potential additional, central
mechanism associated with adjustment to growth in the neuromuscular
system: In the central nervous system, motoneurons
also enlarge their postsynaptic dendritic arbors as animals increase
in body size; this leads to increased synaptic drive and thus
prolonged periods of bursting activity (Zwart, 2013).
Morphometric quantifications of dendritic arbors during
larval stages, show that motoneuron dendritic trees increase their
overall dendritic length proportionately to body size. The growth
of these arbors occurs by addition of new dendritic segments that
'fill in' existing territory, as well as at the perimeter of the tree,
thus widening its reach. Such scaling growth of dendritic arbors
in relation to body size is a widespread phenomenon and has
previously been observed in different types of nerve cells, including
Purkinje, pyramidal, olfactory mitral cells and sympathetic
ganglionic neurons (Zwart, 2013).
An interesting discovery of this study
is that the growth that normally occurs during larval stages is
regulated cell-autonomously. Specifically, this study established that
the B2 isoform of the ecdysone steroid hormone receptor is
required cell-autonomously in motoneurons for normal dendritic
growth. Expression of a dominant-negative form of EcRB2,
UAS-EcR-B2W650A, in single cells prevents the characteristic
increase in motoneuron dendritic arbor size during the second
larval instar stage and seems to arrest neural arbors structurally at
a young larval stage, despite being embedded in an otherwise
normally developing ganglion. Electrical excitability of aCC and
RP2 motoneurons, however, are not affected by expression of
EcR-B2W650A. It was found EcR-B2 is the only isoform expressed
in the larval nerve cord during early larval stages, in agreement
with and complementary to previously published data. Although different functions have been ascribed to the other two EcR isoforms, A and B1, the role of
the B2 isoform had until now remained unknown. This study has
uncovered an important role for the EcR-B2 isoform in nervous
system development, namely to permit growth in larval stages.
The role of EcR-B2 is intrepeted as being a permissive factor for
dendritic growth for two reasons. First, the pattern of dendritic
growth during larval stages does not follow ecdysteroid titers
but may be exponential. Second, precocious or overexpression of
the wild-type form of EcR-B2 does not cause abnormal dendritic
growth. Because the nervous system is one of the most metabolically
expensive tissues, it is conceivable that gating the
decision through EcR-B2 on whether or not neurons grow may
provide a strategy to synchronize neural growth with the growth
of the animal as a whole, as it progresses from one developmental
stage to the next. Indeed, production of ecdysteroids in the
Drosophila larva is under nutritional control (Zwart, 2013).
It is likely that other signaling pathways determine the extent
to which neurons grow. For example, in rat superior cervical
ganglion cells dendritic growth correlates with peripheral target
size and NGF has been implicated. In Drosophila,
three neurotrophic factors have been identified, expressed in
subsets of body-wall muscles at embryonic stages, although none
has thus far been reported to be expressed in the dorsal musculature,
whose innervating aCC and RP2 motoneurons were analyzed
in this study. Other muscle and associated glia-derived
retrograde regulators of neuromuscular junctions include the
TGF-β homologs Glass bottom boat and Maverick and the Wnt
family member Wingless. It is conceivable
that these could regulate the growth of motoneuron dendritic
arbors in synchrony with that of presynaptic axon terminals. Indeed,
it was fiynd that, in addition to its role in regulating dendritic
growth, EcR-B2 is also required for normal neuromuscular
junction growth, suggesting that EcR-B2 itself regulates the development
of both pre- and postsynaptic compartments (Zwart, 2013).
As the Drosophila larva grows around 100-fold in surface
area with matching increases in body wall muscle size, appropriate
levels of muscle depolarization have to be maintained.
The larval body-wall muscles are virtually isopotential, and
as their input resistance goes down with increasing muscle size,
presynaptic output at the neuromuscular junction increases in
a compensatory fashion: By adjusting both terminal size and
synaptic strength the amplitude of postsynaptic responses in the
muscle is maintained. Similarly, at the growing vertebrate neuromuscular junction, motor endplates expand as muscles enlarge, enhancing neurotransmitter release.
The findings of this study show that in addition to the well-characterized
regulation of neuromuscular junction strength, in the central
nervous system, motoneurons also adjust the size of their dendritic
terminals. Previous studies have demonstrated that during the
initial assembly of the locomotor network in the embryo neurons
deploy their dendritic arbors as structural homeostatic devices,
adjusting their extent to compensate for naturally occurring
variations in the density of synaptic partner terminals. This study has shown that in subsequent larval stages, when animals grow rapidly, extension and elaboration of dendritic arbors leads to greater numbers of presynaptic inputs and thus
increased synaptic drive. Specifically, with progression from the
second to the third larval instar the duration of action potential
bursts approximately doubles. Longer action potential burst
periods might increase and potentially prolong muscle contractions;
they could also enhance facilitation. Furthermore,
the time course of excitatory junctional potentials (EJPs)
at the Drosophila neuromuscular junction changes as muscles
enlarge: Time constants describing both the rise and fall of
EJPs increase, which, taking into account the bursting input the
muscle receives, will result in a larger envelope of depolarization
as the animals grows. It is therefore conceivable that the increased
number of action potentials fired per burst that were observed in this study affects the strength of the neuromuscular synapse by enhancing both the process of facilitation and the envelope of depolarization (Zwart, 2013).
Having identified EcR-B2 as a regulator
of dendritic growth of motoneurons, this study has investigated
how dendritic growth relates to synaptic drive in this
system. During normal development, motoneurons increase their
dendritic arbor proportionally to animal body size. A biophysical
consequence of increased neuronal size is increased capacitance
and decreased input resistance, both of which reduce the cell’s
intrinsic excitability. Indeed, this study found that motoneurons in
third instar larvae (48 h ALH) have larger dendritic arbors and
are less excitable than smaller cells of younger, second instar
(24 h ALH) animals. It was also found that increases in dendritic
arbor size are accompanied by increases in the frequency of spontaneous
mEPSPs, suggesting that the dendritic growth
during larval development facilitates the addition of synapses.
It was asked how these changes in synaptic input, from the
second to the third larval instar, might lead to extended bursting
periods. Most likely, this requires the addition of synapses from new
premotor partner neurons. Interestingly, at 48 h ALH EcRB2W650A–
expressing motoneurons, despite being located within an
otherwise unmanipulated nervous system with its third instar
complexity and density of presynaptic release sites, are indistinguishable
from younger neurons in a younger ganglion, in
terms of dendritic arbor size, distribution of dendritic branches in
the neuropil, number of synaptic sites on these, and activity
patterns. This suggests that the size and geography
of these dendritic arbors is decisive in determining their
connectivity. Because increases were found in the duration of action
potential bursts over developmental time, it is likely that as they
grow aCC and RP2 motoneuron arbors establish new presynaptic
contacts from additional interneurons, some of which may be in
adjacent segments. These could include segmental homologs of
those with whom they already form connections more proximally
at earlier stages. Such a scenario could extend the duration of the
synaptic drive, as was observe: As each wave of activity passes
through the nerve cord during locomotion cycles, synapses in
adjacent segments would have different timings that when combined
on one dendritic arbor could lead to prolonged periods of
action potential bursts. Indeed, tentative evidence has shown that
larval Drosophila motoneurons change their connectivity qualitatively,
in that neurons begin to show inhibitory responses during
larval development (Zwart, 2013).
Comparable strategies have been documented in other systems.
For instance, the substantia nigra compacta dopaminergic
neurons change their dendritic architecture to alter the number
and identity of synapses they receive and thereby also their
functional properties within the network. In the case of the
aCC and RP2 motoneurons in Drosophila that are studied
here in this work, the identity of their excitatory presynaptic partners has not
yet been characterized beyond being cholinergic and so at
this point cannot be resolved conclusively (Zwart, 2013).
The electrical excitability of aCC/RP2 is not affected by expression of EcRB2W650A. The excitability of a cell is determined by the
input resistance, the sum of all leak currents at rest, and the
voltage-sensitive conductances that generate the action potential.
The input resistance of a cell is normally inversely proportional
to its size. Consistent with this notion, this study found that
during normal development the electrical excitability of these
neurons decreases during the second larval instar stage, as cells
increase in size. Moreover, no changes were recorded in the relationship
between membrane potential and action potential
firing, suggesting there is no net change in the voltage-sensitive
conductances that generate the action potential, and this aspect
is not affected by EcR-B2W650A expression.
However, the inverse relationship between cell size and excitability
no longer holds for cells expressing EcR-B2W650A: At 48 h
ALH, the intrinsic excitability of these comparatively small cells,
which are reduced both in dendritic arbor and soma size, is similar to that of their age-matched, larger,
control counterparts. These findings are compatible
with at least two scenarios. First, it is possible that the location of
the action potential initiation zone, a key determinant of excitability
of these neurons, changes from the second to the third
larval instar stage. For example, the positioning of the action
potential initiation zone relative to the proximal primary neurite,
which integrates dendritic currents, may change as nerve cords
enlarge, and this could be independent of EcR-B2 signaling in
individual motoneurons. However, in agreement with an earlier
study, this study found that the intrinsic excitability of aCC and RP2
motoneurons strongly correlates with the amplitude of synaptic
input (78). In this model, EcR-B2W650A–expressing neurons would
undergo homeostatic adjustment to remain within the normal
range of neuronal activity. Given that no changes were detected in
the voltage-sensitive conductances that generate the action potential,
but measured a reduction in membrane resistance over
developmental time, leak channels could be involved (Zwart, 2013).
The dendrites of neurons undergo dramatic reorganization in response to developmental and other cues, such as stress and hormones. Although their morphogenesis is an active area of research, there are few neuron preparations that allow the mechanistic study of how dendritic fields are established in central neurons. Dendritic refinement is a key final step of neuronal circuit formation and is closely linked to emergence of function. This is a study of a central serotonergic neuron in the Drosophila brain, the dendrites of which undergo a dramatic morphological change during metamorphosis. Using tools to manipulate gene expression in this neuron, the refinement of dendrites during pupal life was examined. This study shows that the final pattern emerges after an initial growth phase, in which the dendrites function as 'detectors', sensing inputs received by the cell. Consistent with this, reducing excitability of the cell through hyperpolarization by expression of K(ir)2.1 results in increased dendritic length. Sensory input, possibly acting through NMDA receptors, is necessary for dendritic refinement. These results indicate that activity triggers Wnt signaling, which plays a 'pro-retraction' role in sculpting the dendritic field: in the absence of sensory input, dendritic arbors do not retract, a phenotype that can be rescued by activating Wnt signaling. These findings integrate sensory activity, NMDA receptors and Wingless/Wnt5 signaling pathways to advance understanding of how dendritic refinement is established. The maturation of sensory function is shown to interact with broadly distributed signaling molecules, resulting in their localized action in the refinement of dendritic arbors (Singh, 2010).
This study focuses on a specific phase during the metamorphosis of the dendrites of a central serotonergic neuron, in which excess growth is removed by a process that has been termed refinement. Genetic analyses using loss-of-function mutants and RNAi-mediated knockdown of specific genes has led to a postulated a link between neuronal activity, synaptic input and Wnt signaling in this process. The sparse dendrites innervating the adult antennal lobe,
present on the wide-field serotonergic neurons (CSDn) during the larval stage, are removed early in pupation by pruning, followed by a period of exuberant growth. The arrival of sensory neurons at the antennal lobe correlates well with when growth of the CSDn dendrites ceases and removal of the excess branches occurs. The CSDn must be active for the refinement process to occur, as refinement fails when neuronal activity is inhibited or when the sensory neurons are absent. Phenotypes observed in the latter case can be rescued by ectopic activation of the neuron using the temperature-sensitive dTrp-A1 channel. It is suggested that activity within the CSDn, possibly together with activity in presynaptic neurons, acts to provide the correlated activity required to trigger activation of NMDARs. Knockdown of NMDARs affects the refinement process, although identifying its specific action requires further study. A possible consequence of the activity-dependent process is activation of the Wg pathway, as the phenotype observed in aristalless mutants can also be rescued by ectopic expression of Dishevelled (Dsh) in the CSDn. It seems unlikely that activity within the CSDn leads to the release of Wnt ligands, but rather that dendrites respond locally to Wnt ligands in the region of a dendrite that is receiving input. Although other interpretations of the data are possible, a hypothesis is favored whereby specific synapses are stabilized as a result of correlated neuronal activity, and that excess dendritic branches are removed by Wnt signaling (Singh, 2010).
Perturbations in neuronal activity can be compensated by changes at multiple levels, including alterations in the expression of ion channels and in synaptic strength. Tripodi (2008) provides evidence for structural homeostasis whereby alterations in afferent input during development can be compensated by changes in dendritic geometry. This suggests that dendritic arbors serve as sensors for input levels, thus allowing the self-organization of circuits that is necessary for robust behavioral outputs (Tripodi, 2008). The current studies in the CSDn support these observations: reduced activation of the cell by targeted expression of Kir2.1 results in a greatly enlarged dendritic field in the adult. This phenotype can be explained by a mechanism in which the absence of electrical activity results in a failure of the signaling mechanisms that stop growth of the arbors and that remove additional branches. Reduced excitability could also drive the homeostatic mechanisms towards making more arbors and to suppress the refinement program (Singh, 2010).
Dendritic growth and refinement are closely associated with input activity and synapse formation during development. Activity-dependent development of circuits is thought to utilize mechanisms similar to those involved in Hebbian learning and plasticity. NMDARs are ideal candidates for detecting correlated pre- and postsynaptic activity, which is crucial in the Hebbian model of learning and plasticity. Strengthening of synapses, as in this study, leads to the stabilization and extension of dendrites, whereas weakening of synapses leads to the destabilization and elimination of dendritic branches (Espinosa, 2009; Cline, 2008; Constantine-Paton, 1998). During vertebrate hippocampal development, NMDAR activation has been shown to limit synapse number and reduce dendritic complexity. The stabilization of a particular synapse or arbor possibly attenuates the formation of new branches or synapses, thus limiting further dendritic growth. In such a scenario, knocking down NMDAR levels would be expected to result in increased dendritic complexity, as indeed has been observed in this study. The mechanism by which 'appropriately connected' synapses are strengthened, whereas suboptimal contacts are eliminated, needs to be studied in thus system. In other systems, Ca2+, which is released upon NMDAR activation, impinges on various intracellular effectors that regulate dendritic morphogenesis. In addition, selective stabilization/destabilization of dendritic arbors could be affected by the local release of growth factors in response to activity (Singh, 2010).
This study shows that activity-dependent activation of the Wnt pathway facilitates retraction of dendritic arbors. Arbors that receive appropriate input are somehow protected and stabilized. These experiments suggest that Wnt-dependent refinement functions through a non-nuclear pathway and could act by impinging directly on cytoskeletal dynamics (Schlessinger, 2009; Salinas, 2008). Disruption of the microtubule cytoskeleton is a key feature of dendritic pruning in Drosophila during metamorphosis. GSK3β (Shaggy in Drosophila) an intracellular inhibitor of the Wnt pathway, has been shown to act as a sensor of inputs for neuronal activity (Chiang, 2009) and a potent regulator of microtubule dynamics in axons. In the Drosophila embryonic CNS, the Src family of tyrosine kinases (SFKs) is required for Wnt5/Drl-mediated signaling. Interestingly, SFKs seem to act as a crucial point of convergence for multiple signaling pathways that enhance NMDAR activity and hence are thought to act as molecular hubs for the control of NMDARs. It is tempting to envisage a scenario in which there is cross-talk between Wnt5/Drl signaling-mediated activation of SFKs and NMDAR signaling during refinement (Singh, 2010).
In summary, this study shows that the dendritic refinement of a central modulatory serotonergic neuron is regulated by electrical activity, NMDAR and Wnt signaling. Similar mechanisms have been implicated in dendritic growth and refinement of excitatory neurons in vertebrates. This study provides a model neuron preparation in which the dendritic growth and refinement of a modulatory neuron can be analyzed genetically. It was demonstrated that the dendrites of CSDn receive input from sensory neurons from the arista, supporting previous suggestions that mechanosensory input could alter sensitivity to odorant stimuli. In both Drosophila (Dacks, 2009) and the mammalian olfactory bulb (Petzold, 2009), serotonin gates the odor-evoked sensory response. CSDn sends projections to higher brain centers and multiglomerular projections to the contralateral antennal lobe and hence it is likely to influence the overall properties of the olfactory circuit. This study suggests that the structural and resulting functional properties of this neuron emerge from an interaction between partner neurons, together with input from intrinsic and extrinsic cues (Singh, 2010).
Brain development requires correct targeting of multiple thousand synaptic terminals onto staggeringly complex dendritic arbors. The mechanisms by which input synapse numbers are matched to dendrite size, and by which synaptic inputs from different transmitter systems are correctly partitioned onto a postsynaptic arbor, are incompletely understood. By combining quantitative neuroanatomy with targeted genetic manipulation of synaptic input to an identified Drosophila neuron, this study shows that synaptic inputs of two different transmitter classes locally direct dendrite growth in a competitive manner. During development, the relative amounts of GABAergic and cholinergic synaptic drive shift dendrites between different input domains of one postsynaptic neuron without affecting total arbor size. Therefore, synaptic input locally directs dendrite growth, but intra-neuronal dendrite redistributions limit morphological variability, a phenomenon also described for cortical neurons. Mechanistically, this requires local dendritic Ca2+ influx through Dα7 nAChRs or through low-voltage-activated channels following GABAA receptor-mediated depolarizations (Ryglewski, 2017).
Patterns of synaptic connectivity are remarkably precise and complex. Single-cell RNA sequencing has revealed a vast transcriptional diversity of neurons. Nevertheless, a clear logic underlying the transcriptional control of neuronal connectivity has yet to emerge. This study focused on Drosophila T4/T neurons, a class of closely related neuronal subtypes with different wiring patterns. Eight subtypes of T4/T neurons are defined by combinations of two patterns of dendritic inputs and four patterns of axonal outputs. Single-cell profiling during development revealed distinct transcriptional programs defining each dendrite and axon wiring pattern. These programs were defined by the expression of a few transcription factors and different combinations of cell surface proteins. Gain and loss of function studies provide evidence for independent control of different wiring features. It is proposed that modular transcriptional programs for distinct wiring features are assembled in different combinations to generate diverse patterns of neuronal connectivity (Kurmangaliyev, 2019).
T4/T5 neurons share a common developmental origin, physiological function, and general morphology, but differ in their precise wiring patterns and preferred stimulus. There are eight morphological subtypes of T4/T5 neurons in each column of the lobula plate (LoP) neuropil, comprising the most abundant cell type in the fly visual system. These subtypes can be classified into two quartets of subtypes based on dendritic inputs: the four T4 subtypes share a common set of dendritic inputs in the medulla, and the four T5 subtypes share a different set of dendritic inputs in the lobula (see Single-cell sequencing reveals eight transcriptionally distinct populations of T4/T5 neurons). T4 neurons respond to ON stimuli (i.e. bright edges moving against a dark background) and T5 to OFF stimuli (i.e., dark edges moving across a bright background). T4/T5 neurons can also be classified into four pairs of subtypes (a-d) based on the location of their axon terminals within a given column in layers a-d of the LoP. Each pair responds selectively to visual motion in one of four cardinal directions: posterior, anterior, upwards, and downwards, respectively. Although transcriptional profiling of the adult Drosophila brain revealed a common transcriptional signature for all T4/T5 neurons, genetic programs for individual subtypes have not been identified. This study hypothesized that identification of gene expression programs for individual T4/T5 subtypes during circuit assembly would provide insight into the genetic programs regulating discrete wiring features (Kurmangaliyev, 2019).
This study reports that independent transcriptional programs define the dendritic inputs and axonal outputs of T4/T5 neurons. Gain and loss of function studies indicate that these programs control their corresponding morphological features. These findings suggest that the modular assembly of separate dendritic and axonal transcriptional programs contributes to the diversity of wiring patterns in complex nervous systems (Kurmangaliyev, 2019).
A unique attribute of T4/T5 neurons is that the same dendritic and axonal wiring patterns are reiteratively used among different subtypes; each neuron can be described by a unique combination of one of four types of axonal outputs and one of two types of dendritic inputs. It was anticipated that this property of T4/T5 neurons would provide an opportunity to assess the relationship between specific genetic programs and fundamental features of neuronal architecture (Kurmangaliyev, 2019).
Unsupervised analysis revealed that separable transcriptional programs correlate with these specific wiring features. This study demonstrates through gain and loss of function experiments that these programs control specific axonal targeting features, which are separable from other features (e.g., dendrite targeting). These programs can be re-assembled in a modular fashion to generate neuronal subtypes with different combinations of wiring features. A modular transcriptional architecture may provide a general strategy for discrete modifications to neuronal connectivity in development and evolution (Kurmangaliyev, 2019).
A common T4/T5 neuronal identity is defined by a unique combination of TFs expressed in all subtypes (e.g., Lim1, Drgx, acj6). Perturbation of TFs expressed in all subtypes disrupts overall organization of T4/T5 neurons, including both dendritic and axonal morphologies. This common T4/T5 transcriptional program is further diversified by separable feature-specific transcriptional programs. These programs are defined by three binary (ON/OFF) TF expression patterns, with two TF patterns defining the four axonal outputs and one TF pattern defining the two dendritic inputs. In this way, modular TF codes defining common and feature-specific transcriptional programs give rise to eight T4/T5 subtypes (Kurmangaliyev, 2019).
Four pairs of T4/T5 subtypes with shared axonal outputs (and different dendritic inputs) each target one of four LoP layers, a-d. The ultimate layered architecture of neuropils develops through sequential lamination into increasing numbers of layers. Together with previous results, the findings suggest that the lamination of T4/T5 axonal outputs occurs via two distinct processes, each controlled by a separate TF. Binary expression of bi is required for lamination of the broad a/b from c/d LoP domains (Apitz, 2018), whereas binary expression of grn is required for sublamination of each of these two domains into separate LoP layers. Importantly, perturbation of each TF exclusively disrupts the corresponding lamination step, while not affecting other morphological features of T4/T5 neurons. Similarly, two quartets of subtypes with shared dendritic inputs (and different axonal outputs) were defined by binary expression of TfAP-2. Arborization of dendrites in M10 (T4) or Lo1 (T5) occurs during initial neurite guidance steps, preceding the developmental stages covered in this study. It is hypothesized that differentially expressed genes (DEGs) between T4 and T5 subtypes identified in this analysis contribute to the connections with two distinct sets of presynaptic partners (Kurmangaliyev, 2019).
The binary expression patterns of TFs also mirror the developmental lineages of T4/T5 neurons. a/b and c/d subtypes arise from bi- and bi+ progenitor populations. Neuroblasts from each population undergo two terminal Notch-dependent asymmetric divisions to give rise to the eight subtypes (Pinto-Teixeira, 2018). These divisions correspond to binary expression patterns of grn and TfAP-2, respectively, which act with Notch signaling to regulate wiring. Remarkably, despite divergent developmental trajectories separated by multiple divisions and distinct progenitor pools, all T4 and all T5 subtypes converge onto the same transcriptional programs associated with two types of dendritic inputs. Three regulatory dichotomies could also reflect the evolutionary origin of T4/T5 subtypes and correspond to consecutive duplications of ancestral cell types and circuits (Kurmangaliyev, 2019).
Each axonal and dendritic transcriptional program is characterized by a specific pattern of TFs, as well as a set of cell surface proteins (CSPs), many of which are implicated in regulating wiring in other developmental contexts. These include Ig superfamily proteins in which different paralogs exhibit discrete heterophilic binding specificities, including the beat/side and the dpr/DIP interacting protein families. Interestingly, dynamic expression of these proteins in neurons with shared wiring features was developmentally coordinated. It is envisioned that the synaptic specificity of T4/T5 dendrites and axons are determined by the combined activity of these recognition molecules through interactions with synaptic partners. Future experiments utilizing gain and loss of function analysis, either alone or different combinations, will provide insights into the cellular recognition mechanisms by which synaptic specificity is established (Kurmangaliyev, 2019).
The composite morphological properties of T4/T5 subtypes allowed identification, and thus decouple transcriptional programs for dendrite and axon wiring. Combining separate dendritic and axonal programs, and variations on them, may contribute to the diversification of synaptic specificity in different neuronal subtypes across complex nervous systems (Kurmangaliyev, 2019).
Cytoplasmic polyadenylation element binding proteins (CPEBs) are widely conserved proteins that regulate the length of poly(A) tails in the cytoplasm, regulate translation, and regulate mRNA transport. While CPEBs are best known for regulating maternal messages in oocytes, CPEBs also have roles in many other cell types including neurons. This study extends the knowledge of the roles of CPEBs in neurons by showing that the Drosophila CPEB-encoding gene, orb, is required for proper dendrite development in larval sensory dendritic arborization neurons. Furthermore, evidence is provided that orb is important for neuron cell fate specification (Olesnicky, 2020).
The main lysosomal protease cathepsin D (cathD) is essential for maintaining tissue homeostasis via its degradative function, and its loss leads to ceroid accumulation in the mammalian nervous system, which results in progressive neurodegeneration. Increasing evidence implies non-proteolytic roles of cathD in regulating various biological processes such as apoptosis, cell proliferation, and migration. Along these lines, this study shows that cathD is required for modulating dendritic architecture in the nervous system independent of its traditional degradative function. Upon cathD depletion, class I and class III arborization (da) neurons in Drosophila larvae exhibited aberrant dendritic morphology, including over-branching, aberrant turning, and elongation defects. Re-introduction of wild-type cathD or its proteolytically-inactive mutant dramatically abolished these morphological defects. Moreover, cathD knockdown also led to dendritic defects in the adult mushroom bodies, suggesting that cathD-mediated processes are required in both the peripheral and central nervous systems. Taken together, these results demonstrate a critical role of cathD in shaping dendritic architecture independent of its proteolytic function (Zhang, 2020).
Establishing appropriate sizes and shapes of dendritic arbors is critical for proper wiring of the central nervous system. This study reports that Insulin-like Peptide 2 (DILP2) locally activates transiently expressed insulin receptors in the central dendrites of Drosophila Dm8 amacrine neurons to positively regulate dendritic field elaboration. DILP2 is expressed in L5 lamina neurons, which have axonal terminals abutting Dm8 dendrites. Proper Dm8 dendrite morphogenesis and synapse formation required insulin signaling through TOR (target of rapamycin) and SREBP (sterol regulatory element-binding protein), acting in parallel with previously identified negative regulation by Activin signaling to provide robust control of Dm8 dendrite elaboration. A simulation of dendritic growth revealed trade-offs between dendritic field size and robustness when branching and terminating kinetic parameters were constant, but dynamic modulation of the parameters could mitigate these trade-offs. It is suggested that antagonistic DILP2 and Activin signals from different afferents appropriately size Dm8 dendritic fields (Luo, 2020).
This study has shown that insulin/TOR/SREBP signaling positively regulates dendritic development. Insulin-like peptide DILP2 from L5 axons signals through insulin receptors on Dm8 dendrites to promote dendritic expansion via the canonical TOR pathway and SREBP. The roles of the insulin/TOR signaling pathway in cell growth and differentiation have been very well established. However, this is the first study to definitively demonstrate that insulin/TOR/SREBP signaling regulates dendritic field sizes during development. In Xenopus tadpoles, insulin receptor is expressed in retina and optic tectum, where it regulates synapse density and visual circuit function. However, the source of insulin or insulin-like growth factor (IGF) and the signaling pathway have not been described in developing Xenopus tectum. In vitro studies in cultured cells suggest that insulin/IGF signaling can not only regulate dendrite spine morphology and neurite outgrowth, but also function as a synaptic component at the postsynaptic density (PSD). It would be interesting to determine whether afferent-derived insulin/IGF is also utilized for dendritic development in vertebrate nervous systems (Luo, 2020).
The PI3K/AKT/TOR signaling pathway is known to mediate a broad spectrum of functions through two major complexes, TORC1 and TORC2, which act through different and varied effectors. In cultured hippocampal neurons, the PI3K-TOR pathway regulates dendrite size and branching patterns, presumably by interacting with the microtubule plus-end-tracking protein CLIP-170 to enhance crosstalk between CLIP-170 and the actin-binding protein IQGAP1 (Swiech, 2011). Dm8 dendritic development appears to involve the TORC1 complex and one of its downstream targets, SREBP. Removing the TORC2 component, Rictor, or TORC1 targets other than SREBP had no significant effect on Dm8 dendritic field size. In contrast, SREBP mutant Dm8 neurons had a reduced dendritic field size, and overexpression of SREBP rescued the dendritic phenotype of Tor mutants. While the possibility cannot be ruled out that other TOR targets play an additional or minor role, the results suggest that TOR works primarily through SREBP to regulate Dm8 dendritic development. Recent studies showed that SREBP enhances the transcription of cholesterol and fatty acid synthesis enzymes to regulate dendritic development of Da neurons in the peripheral nervous system. Thus, mounting evidence suggests that lipid synthesis may be a key regulatory point for controlling dendritic development in both central and peripheral nervous systems (Luo, 2020).
Using a novel receptor-based activity-dependent GRASP (R-synGRASP) technique, this study found synaptic defects between R7 photoreceptors and mutant Dm8 neurons that lack Insulin/TOR pathway signaling components. While the R-GRASP method revealed that Tor or chico mutant Dm8 dendrites formed membrane contacts with R7 axonal terminals in numbers that correlated with the dendritic field sizes, the results from the R-synGRASP method suggested these contacts do not constitute active synapses. Notably, Pten mutant Dm8 neurons appear to form more active synapses with peripheral R7 photoreceptors than central R7 photoreceptors, in contrast to wild types. At this stage of inquiry, the nature of such defects remains unclear. The presynaptic structures of R7 photoreceptors appear to be unaltered, as judged by the localization of the active zone marker, Bruchpilot. The level of histamine receptors on dendrites is significantly reduced in Tor- but not chico mutant Dm8 neurons, suggesting that reduced receptor levels might only partially account for the lack of R-synGRASP signal. It has been shown that insulin signaling regulates protein trafficking in various neuronal models, and mutations in SREBP and its upstream regulator easily shocked (eas) have been associated with excitability defects. Further studies will be required to resolve the nature of the synaptic defects in Tor and chico mutant Dm8 neurons (Luo, 2020).
Antagonistic regulation of Dm8 dendritic expansion by afferent-derived factors ensures robust size control
A previous study showed that R7-derived Activin negatively regulates the expansion of Dm8 dendritic fields (Ting, 2014). This action is countered by positive regulation by L5-derived DILP2 revealed in this study. Despite the antagonistic effects on Dm8 dendritic expansion, both Activin and DILP2 are derived from afferents, delivered to the growth cones, and likely act on dendrites at close range. Both L3 and L5 lamina neurons express DILP2, but RNAi-mediated knockdown of dlip2 in L5 neurons alone is sufficient to reduce Dm8 dendritic expansion. This result suggests that L3-derived DILP2 is not necessary for normal development of Dm8 dendrites, even though the axonal terminals of L3 and L5 are only few micrometers apart. Similarly, Activin derived from R7 and R8 photoreceptors acts respectively on Dm8 and Tm20 neurons, even though the R7 and R8 axonal terminals are also only separated by a few micrometers (Ting, 2014). Furthermore, both Activin and DILP2 appear to have multiple functions in the developing optic lobe. In addition to its function on Dm8 dendritic development, R7-derived Activin functions in an autocrine fashion to control R7 axonal tiling. Glia-derived DILPs regulate lamina neuron differentiation, while L3 and L5 neurons express DILP2, which also affects photoreceptor axonal tiling. It is thus tempting to speculate that afferent-derived factors tend to function at short range on multiple targets during distinct developmental stages (Luo, 2020).
The results from genetic interaction experiments further argue that Activin and DILP2 signal in developing Dm8 dendrites through parallel pathways. Interestingly, the antagonistic regulation of Dm8 dendritic expansion by two afferent-derived factors is required for elaboration of appropriately large and robust Dm8 dendritic fields - removing both signaling events rendered highly variable sizes of Dm8 dendritic fields. While antagonistic regulation has been shown to control a wide range of biological processes, including gene and protein expression, this work provides the first example of antagonistic regulation for robust control of dendritic field sizes (Luo, 2020).
How does antagonistic regulation by insulin and Activin signaling ensure appropriate and robust sizes of Dm8 dendritic fields? Several lines of evidence suggest the involvement of spatiotemporal restriction of insulin signaling. Strong insulin receptor expression was detected in Dm8 neurons at early pupal stages (20-40 hr APF), when the cells had just begun to expand their dendritic arbors, but expression disappeared by late pupal stages (60 hr APF), before the dendrites expanded to adult sizes. Furthermore, the insulin receptors in developing Dm8 neurons were largely restricted to the central dendritic field, corresponding to one single column. Thus, insulin signaling likely occurs only over a short developmental period in response to insulin signal from one or a few L5 neurons to transiently promote Dm8 dendritic expansion during the early pupal stages (Luo, 2020).
To further understand how antagonistic regulation might control dendritic field sizes, simulations were carried out of Dm8 dendritic development using a kinetic Monte Carlo method. The variance of dendritic field sizes correlated with the difference between two key dendritic development parameters, branching and terminating rates: high branching and low terminating rates favor dendritic field expansion at the cost of high variability, while low branching and high terminating rates lead to small but robust dendritic field sizes. This study found that elaboration of large and consistent dendritic fields could be achieved by spatial and/or temporal segregation of growing and terminating phases. A previous study using in vivo imaging of dendritic dynamics showed that dendrites are elaborated at different rates according to the developmental stage. Thus, it is speculated that spatiotemporal restriction of insulin signaling, coupled with Activin-mediated negative regulation, allows Dm8 neurons to elaborate a large and consistent dendritic field. The large and robust sizes of Dm8 dendritic fields not only ensure a consistent receptive field for approximately 14 R7 photoreceptor inputs, but also can receive negative feedback signals to generate a center-surround signal structure (Luo, 2020).
During prolonged nutrient restriction, developing animals redistribute vital nutrients to favor brain growth at the expense of other organs. In Drosophila, such brain sparing relies on a glia-derived growth factor to sustain proliferation of neural stem cells. However, whether other aspects of neural development are also spared under nutrient restriction is unknown. This study shows that dynamically growing somatosensory neurons in the Drosophila peripheral nervous system exhibit organ sparing at the level of arbor growth: Under nutrient stress, sensory dendrites preferentially grow as compared to neighboring non-neural tissues, resulting in dendrite overgrowth. These neurons express lower levels of the stress sensor FoxO than neighboring epidermal cells, and hence exhibit no marked induction of autophagy and a milder suppression of Tor signaling under nutrient stress. Preferential dendrite growth allows for heightened animal responses to sensory stimuli, indicative of a potential survival advantage under environmental challenges (Poe, 2020).
Dendrite morphology is necessary for the correct integration of inputs that neurons receive. The branching mechanisms allowing neurons to acquire their type-specific morphology remain unclear. Classically, axon and dendrite patterns were shown to be guided by molecules, providing deterministic cues. However, the extent to which deterministic and stochastic mechanisms, based upon purely statistical bias, contribute to the emergence of dendrite shape is largely unknown. This issue was addressed using the Drosophila class I vpda multi-dendritic neurons. Detailed quantitative analysis of vpda dendrite morphogenesis indicates that the primary branch grows very robustly in a fixed direction, though secondary branch numbers and lengths showed fluctuations characteristic of stochastic systems. Live-tracking dendrites and computational modeling revealed how neuron shape emerges from few local statistical parameters of branch dynamics. This study reports key opposing aspects of how tree architecture feedbacks on the local probability of branch shrinkage. Child branches promote stabilization of parent branches, although self-repulsion promotes shrinkage. Finally, it was shown that self-repulsion, mediated by the adhesion molecule Dscam1, indirectly patterns the growth of secondary branches by spatially restricting their direction of stable growth perpendicular to the primary branch. Thus, the stochastic nature of secondary branch dynamics and the existence of geometric feedback emphasize the importance of self-organization in neuronal dendrite morphogenesis (Palavalli, 2020).
Dendrites require precise and timely delivery of protein substrates to distal areas to ensure the correct morphology and function of neurons. Many of these protein substrates are supplied in the form of ribonucleoprotein (RNP) complex consisting of RNA-binding proteins (RBPs) and mRNAs, which are subsequently translated in distal dendritic areas. This study investigated how Drosophila sensory neurons respond to the dysregulation of a disease-associated RBP, Ataxin-2 (ATX2), which leads to dendritic defects. ATX2 plays a crucial role in spacing dendritic branches for the optimal dendritic receptive fields in Drosophila class IV dendritic arborization (C4da) neurons, where both expression level and subcellular location of ATX2 contribute significantly to this effect. Translational upregulation through the expression of eukaryotic translation initiation factor 4E (eIF4E) further enhanced the ATX2-induced dendritic phenotypes. Additionally, this study found that the expression level of another disease-associated RBP, fragile X mental retardation protein (FMRP), decreased in both cell bodies and dendrites when neurons were faced with aberrant upregulation of ATX2. Finally, this study revealed that the PAM2 motif of ATX2, which mediates its interaction with poly(A)-binding protein (PABP), is potentially necessary for the decrease of FMRP in certain neuronal stress conditions. Collectively, these data suggest that dysregulation of RBPs triggers a compensatory regulation of other functionally-overlapping RBPs to minimize RBP dysregulation-associated aberrations that hinder neuronal homeostasis in dendrites (Cha, 2020).
While many regulators of axon regeneration have been identified, very little is known about mechanisms that allow dendrites to regenerate after injury. Using a Drosophila model of dendrite regeneration, a candidate screen was performed of receptor tyrosine kinases (RTKs), and a requirement was found for RTK-like orphan receptor (Ror). This study confirmed that Ror was required for regeneration in two different neuron types using RNA interference (RNAi) and mutants. Ror was not required for axon regeneration or normal dendrite development, suggesting a specific role in dendrite regeneration. Ror can act as a Wnt coreceptor with frizzleds (fzs) in other contexts, so this study tested the involvement of Wnt signaling proteins in dendrite regeneration. Knockdown of fz, dishevelled (dsh), Axin, and gilgamesh (gish) also reduced dendrite regeneration. Moreover, Ror was required to position Dsh and Axin in dendrites. Recently studies found that Wnt signaling proteins, including Dsh and Axin, localize microtubule nucleation machinery in dendrites. It is therefore hypothesized that Ror may act by regulating microtubule nucleation at baseline and during dendrite regeneration. Consistent with this hypothesis, localization of the core nucleation protein gammaTubulin was reduced in Ror RNAi neurons, and this effect was strongest during dendrite regeneration. In addition, dendrite regeneration was sensitive to partial reduction of gammaTubulin. It is conclude that Ror promotes dendrite regeneration as part of a Wnt signaling pathway that regulates dendritic microtubule nucleation (Ney, 2020).
Using a candidate screening approach, Ror was identified as a regulator of dendrite regrowth after injury in two different Drosophila neurons. Axon regeneration and developmental dendrite outgrowth were unaffected in Ror mutant animals. This injury-specific phenotype is consistent with expression of Ror in the nervous system without detectable defects in nervous system architecture in mutants. In C. elegans and mammals, Ror has been associated with some subtypes of Wnt signaling, and in flies, it has been shown to bind Wnt ligands and Fz2. Based on this link between Ror and Wnt signaling, Wnt signaling proteins were tested for a role in dendrite regeneration. The subset that affected regenerative growth included those involved broadly in Wnt signaling like the Frizzleds, Gish (CK1γ), and Dsh and those involved more specifically in canonical Wnt signaling like Arr, low-density lipoprotein related-receptor protein 5/6 (LRP5/6), and Axin but notably did not include Armadillo (Arm; β-catenin), the output of canonical Wnt signaling. Serendipitously, this subset matched the proteins wetr identified in a screen for factors required to position microtubule nucleation sites in dendrites. So, although the outputs of Ror-mediated Wnt signaling have typically been elusive, this study showed that Ror acts upstream of dendritic microtubule nucleation, and this likely mediates its effect on dendrite regeneration. Overall, a model is proposed in which Ror acts together with other Wnt receptors to localize the scaffolding proteins Dsh and Axin to dendrites, and these, in turn, promote microtubule nucleation throughout dendrite arbors. Because microtubules are structural elements and tracks for transport, generation of microtubules throughout dendrites is likely required for rapid regrowth after injury (Ney, 2020).
Microtubule nucleation also occurs locally in axons and in uninjured dendrites so why are axon regeneration and dendrite development normal in Ror mutants? For axon regeneration, one possibility is that Wnt signaling proteins do not act upstream of nucleation in this compartment. However, axon regeneration is also more resistant to partial loss of γTubulin than dendrite regeneration is, suggesting that axon regeneration is generally less sensitive to nucleation levels than dendrite regeneration rather than just resistant to Ror loss. Alternatively, the relatively high stability of axonal microtubules relative to dendritic ones could account for the difference in sensitivity to reduced nucleation. Axonal microtubules tend to be longer and turn over less than dendritic ones, meaning that the demands on nucleation may be lower in axons. If this is the case, a different explanation is needed for the resilience of dendrite development to Ror loss. One possibility is that Ror only functions to position nucleation sites during dendrite regeneration, but not during development. However, this does not seem to be the case because γTubulin-GFP branch point localization was lower in uninjured dendrites when Ror was knocked down. Instead, the idea is favored that Ror-mediated nucleation acts in parallel to other pathways that are sufficient for microtubule generation in uninjured neurons. For example, microtubule severing followed by Patronin-mediated microtubule minus-end growth could maintain microtubule number under normal conditions. Patronin is a microtubule minus-end-binding protein that facilitates persistent growth of minus ends into dendrites during development and regeneration. In C. elegans, Patronin has been shown to act in parallel to nucleation. If a new microtubule were released from a nucleation site by a severing protein and recognized by Patronin, the nucleation site could act catalytically, and very few would be required. Only under extreme conditions, such as those imposed by regrowth of dendrites after injury, would local nucleation become essential (Ney, 2020).
While the specific deficit of dendrite regeneration in Ror mutants could be due to increased demands on nucleation that surpass a phenotypic threshold only after dendrite injury, the fact that Ror is a signaling receptor raises the intriguing possibility that it could also respond to injury signals. The closest phylogenetic neighbors to the Ror family of RTKs are the tropomyosin receptor kinases (Trks), which have been lost in evolution in flies and worms. One major function of Trk receptors is to couple neuronal survival to target innervation. Target tissues secrete neurotrophins that bind Trks on neurons to generate signaling endosomes that are transported to the cell body to promote survival. The involvement of Ror in dendrite regeneration suggests that it could also link a neuronal survival/adaptation response to the state of surrounding cells. Ror function has been tightly linked to Wnt binding, so it is likely that a Wnt is also involved in this context. Wnt5a-Ror signaling can work in an autocrine loop, but because neuronally expressed Wntless RNAi does not affect γTubulin localization, it is thought more likely that a ligand is secreted from a surrounding cell. It is therefore possible that surrounding cells influence dendrite regeneration through Ror-controlled microtubule nucleation (Ney, 2020).
Neurogenesis is achieved through a sequence of steps that include specification and differentiation of progenitors into mature neurons. Frequently, precursors migrate to distinct positions before terminal differentiation. The Slit-Robo pathway, formed by the secreted ligand Slit and its membrane bound receptor Robo, was first discovered as a regulator of axonal growth. However, today, it is accepted that this pathway can regulate different cellular processes even outside the nervous system. This study describes the participation of the Slit-Robo pathway in the development of motion sensitive neurons of the Drosophila visual system. Slit and Robo receptors are expressed in different stages during the neurogenesis of motion sensitive neurons. Furthermore, it was found that Slit and Robo regulate multiple aspects of their development including neuronal precursor migration, cell segregation between neural stem cells and daughter cells and formation of their connectivity pattern. Specifically, loss of function of slit or robo receptors in differentiated motion sensitive neurons impairs dendritic targeting, while knocking down robo receptors in migratory progenitors or neural stem cells leads to structural defects in the adult optic lobe neuropil, caused by migration and cell segregation defects during larval development. Thus, this work reveals the co-option of the Slit-Robo signaling pathway in distinct developmental stages of a neural lineage (Guzman-Palma, 2021).
Neurons utilize plasticity of dendritic arbors as part of a larger suite of adaptive plasticity mechanisms. This explicitly manifests with motoneurons in the Drosophila embryo and larva, where dendritic arbors are exclusively postsynaptic and are used as homeostatic devices, compensating for changes in synaptic input through adapting their growth and connectivity. Reactive oxygen species (ROS) has been identified as novel plasticity signals instrumental in this form of dendritic adjustment. ROS correlate with levels of neuronal activity and negatively regulate dendritic arbor size. This study investigated NADPH oxidases as potential sources of such activity-regulated ROS and implicate Dual Oxidase (but not Nox), which generates hydrogen peroxide extracellularly. It was further shown that the aquaporins Bib and Drip, but not Prip, are required for activity-regulated ROS-mediated adjustments of dendritic arbor size in motoneurons. These results suggest a model whereby neuronal activity leads to activation of the NADPH oxidase Dual Oxidase, which generates hydrogen peroxide at the extracellular face; aquaporins might then act as conduits that are necessary for these extracellular ROS to be channeled back into the cell where they negatively regulate dendritic arbor size (Dhawan, 2021).
Memory-relevant neuronal plasticity is believed to require local translation of new proteins at synapses. Understanding this process requires the visualization of the relevant mRNAs within these neuronal compartments. This study used single-molecule fluorescence in situ hybridization to localize mRNAs at subcellular resolution in the adult Drosophila brain. mRNAs for subunits of nicotinic acetylcholine receptors and kinases could be detected within the dendrites of co-labeled mushroom body output neurons (MBONs) and their relative abundance showed cell specificity. Moreover, aversive olfactory learning produced a transient increase in the level of CaMKII mRNA within the dendritic compartments of the γ5β'2a MBONs. Localization of specific mRNAs in MBONs before and after learning represents a critical step towards deciphering the role of dendritic translation in the neuronal plasticity underlying behavioral change in Drosophila (Mitchell, 2021).
Mammalian CaMKII mRNA is transported to neuronal dendrites, where it is locally translated in response to neuronal activity. Drosophila CAMKII is critical for behavioral plasticity and is also thought to be locally translated. However, fly CAMKII mRNAs have not been directly visualized within individual neurons. Therefore this study first hybridized CaMKII smFISH probes to whole-mount brains and imaged the mushroom body (MB) calyx, a recognizable neuropil containing the densely packed dendrites of ~2000 KCs and their presynaptic inputs from ~350 cholinergic olfactory projection neurons using a standard spinning disk confocal microscope. To detect and quantify mRNA within the 3D volume of the brain, a FIJI-compatible custom-built image analysis tool was developed that segments smFISH image data and identifies spots within the 3D volume using a probability-based hypothesis test. This enabled detection of mRNAs with a false discovery rate of 0.05. CaMKII smFISH probes labeled 56 ± 5 discrete puncta within each calyx. In comparison, smFISH probes directed to the α1 nicotinic acetylcholine receptor (nAChR) subunit labeled 33 ± 2 puncta in the calyx. Puncta were diffraction limited and the signal intensity distribution was unimodal, indicating that they represent single mRNA molecules (Mitchell, 2021).
Drosophila learning is considered to be implemented as plasticity of cholinergic KC-MBON synapses. To visualize and quantify mRNA specifically within the dendritic field of the γ5β'2a and γ1pedc>α/β MBONs, a membrane-tethered UAS-myr::SNAP reporter transgene was expressed using MBON-specific GAL4 drivers. This permitted simultaneous fluorescent labeling of mRNA with smFISH probes and the MBON using the SNAP Tag. To correct for chromatic misalignment that results from imaging heterogenous tissue at depth, brains were also co-stained with the dsDNA-binding dye Vybrant DyeCycle Violet (VDV). VDV dye has a broad emission spectrum so labeled nuclei can be imaged in both the SNAP MBON and smFISH mRNA channels. This triple-labeling approach allowed quantification and correction of any spatial mismatch between MBON and smFISH channels in x, y, and z planes, which ensures that smFISH puncta are accurately assigned within the 3D volume of the MBON dendritic field (Mitchell, 2021).
Using this smFISH approach, an average of 32 ± 2 CaMKII mRNAs was detected within the dendrites of γ5β'2a MBONs. However, in contrast to the calyx, no nAChRα1 was detected in γ5β'2a MBON dendrites. This differential localization of the CaMKII and nAChRα1 mRNAs within neurons of the mushroom body is indicative of cell specificity. To probe mRNA localization in MBONs more broadly, a single YFP smFISH probe set and a collection of fly strains harboring YFP insertions in endogenous genes were used. YFP insertions in the CaMKII, PKA-R2, and Ten-m genes were selected as test cases and the localization of their YFP-tagged mRNAs was compared between γ5β'2a MBON and γ1pedc>α/β MBON dendrites (Mitchell, 2021).
The CaMKII::YFP allele is heterozygous in flies also expressing myr::SNAP in MBONs. Therefore, YFP smFISH probes detected half the number of CaMKII mRNAs in γ5β'2a MBON dendrites compared to CaMKII-specific probes. Importantly, YFP probes hybridized to YFP-negative control brains produced background signal that was statistically distinguishable in brightness from genuine smFISH puncta. Comparing data from YFP-negative and YFP-positive samples allowed definition of the false discovery rate to be 14% when using YFP-directed probes. These results indicate that the YFP probes are specific and that the YFP insertion does not impede localization of CaMKII mRNA. A similar abundance of CaMKII::YFP was detected in the dendritic field of γ5β'2a and the γ1 dendritic region of γ1pedc>α/β MBONs. In contrast, more PKA-R2 mRNAs were detected in the dendrites of γ5β'2a MBONs compared to γ1pedc>α/β MBONs. Importantly, the relative abundance of dendritically localized CaMKII and PKA-R2 mRNAs did not simply reflect the levels of these transcripts detected in the MBON somata. In addition, Ten-m mRNAs was not detected in either γ5β'2a or γ1pedc>α/β MBON dendrites, although they were visible in neighboring neuropil and at low levels in the MBON somata. These results suggest that CaMKII and PKA-R2 mRNAs are selectively localized to MBON dendrites (Mitchell, 2021).
Although nAChRα1 mRNA was not detected within γ5β'2a MBON dendrites, prior work has shown that nAChR subunits, including nAChRα1, are required in γ5β'2a MBON postsynapses to register odor-evoked responses and direct odor-driven behaviors. Since the YFP insertion collection does not include nAChR subunits, nAChRα5 and nAChRα6-specific smFISH probes were designed. These probes detected nAchRα5 and nAchRα6 mRNAs within γ5β'2a and γ1pedc>α/β MBON dendrites, with nAchRα6 being most abundant. Importantly, nAchRα1, nAchRα5, and nAchRα6 were detected at roughly equivalent levels in the γ5β'2a and γ1pedc>α/β MBON somata. Therefore, the selective localization of nAchRα5 and nAchRα6 mRNA to MBON dendrites indicates that these receptor subunits may be locally translated to modify the subunit composition of postsynaptic nAChR receptors (Mitchell, 2021).
Localized mRNAs were on average 2.8x more abundant in γ5β'2a relative to the γ1 region of γ1pedc>α/β MBON dendrites. Therefore whether this apparent differential localization correlated with dendritic volume and/or the number of postsynapses between these MBONs was tested. Using the recently published electron microscope volume of the Drosophila 'hemibrain', the dendritic volume of the γ5β'2a MBON was calculated to be 1515.36 nm3 and the γ1 region of the γ1pedc>α/β MBON was calculated to be 614.20 nm3. In addition, the γ5β'2a regions of the γ5β'2a MBON dendrite contain 30,625 postsynapses, whereas there are only 17,020 postsynapses in the γ1 region of the γ1pedc>α/β MBON. Larger dendritic field volume and synapse number is therefore correlated with an increased number of localized nAchRα5, nAchRα6, and PKA-R2 mRNAs. The correlation, however, does not hold for CaMKII mRNA abundance. Selective localization of mRNAs to MBON dendrites therefore appears to be more nuanced than simply reflecting the size of the dendritic arbor, the number of synapses, or the level of transcripts detected throughout the cell (Mitchell, 2021).
Whether CaMKII::YFP mRNA abundance in γ5β'2a and γ1pedc>α/β MBONs was altered following aversive learning was tested. mRNA in the somata and nuclei of these MBONs was quantified. Transcriptional activity is indicated by a bright nuclear transcription focus. Flies were initially subjected to four conditions: (1) an 'untrained' group that was loaded and removed from the T-maze but not exposed to odors or shock; (2) an 'odor only' group, exposed to the two odors as in training but without shock; (3) a 'shock only' group that was handled as in training and received the shock delivery but no odor exposure; and (4) a 'trained' group that was aversively conditioned by pairing one of the two odors with shock. Fly brains were extracted 10 min, 1 hr, or 2 hr after training and processed for smFISH (Mitchell, 2021).
CaMKII mRNA increased significantly in γ5β'2a MBON dendrites 10 min after training compared to all control groups. Including an additional 'unpaired' experiment, where odor and shock presentation was staggered, confirmed that the increase at 10 min after training requires coincident pairing of odor and shock. Moreover, levels returned to baseline by 1 hr and remained at that level 2 hr after training. CaMKII mRNAs in γ5β'2a MBON somata showed a different temporal dynamic, with transcripts peaking 1 hr after training, albeit only relative to untrained and odor only controls. The proportion of γ5β'2a nuclei containing a CaMKII transcription focus did not differ between treatments, suggesting that the transcript increase in the somata is not correlated with the number of actively transcribing γ5β'2a nuclei, at least at the timepoints measured. In addition, the mean brightness of γ5β'2a transcription foci did not change across treatments, although the variation was substantial. An increase of dendritically localized CaMKII mRNAs could result from enhanced trafficking or through the release of transcripts from protein bound states, which would increase smFISH probe accessibility and hence spot brightness. Since the brightness of CaMKII mRNA spots detected in the dendrites of γ5β'2a MBONs did not change with treatment, it is concluded that the increased abundance likely results from altered traffic (Mitchell, 2021).
Assessing CaMKII mRNA abundance in γ1pedc>α/β MBONs after learning did not reveal a change in mRNA abundance in the dendrites or somata between trained flies and all control groups at all timepoints measured. These results indicate specificity to the response observed in the γ5β'2a MBONs (Mitchell, 2021).
Since CaMKII protein is also labeled with YFP in CaMKII::YFP flies, protein expression was assessed by measuring YFP fluorescence intensity specifically within the MBON dendrites. This analysis did not reveal a significant difference in fluorescence intensity across treatments. However, since smFISH provides single-molecule estimates of mRNA abundance, a similar level of single-molecule sensitivity may be required to detect subcellular resolution changes in protein copy number. Moreover, new synthesis and replacement of specific isoforms of CaMKII could radically change local kinase activity, even without an observable change in overall abundance (Mitchell, 2021).
Early studies in Drosophila demonstrated that broad disruption of CAMKII function impaired courtship learning. In contrast, later studies that manipulated activity more specifically in olfactory projection neurons or particular classes of KCs reported a preferential loss of middle-term or long-term olfactory memory. This study focused on two subtypes of MBONs, that are known to exhibit changes in odor-evoked activity after a single trial of aversive olfactory conditioning. Whereas γ1pedc>α/β MBON responses to the previously shock-paired odor are depressed immediately after aversive learning, prior studies observed a learning-related increase of the conditioned odor response of γ5β'2a MBONs, likely resulting from a release of feedforward inhibition from γ1pedc>α/β MBONs. It is therefore speculated that the specific change in CaMKII mRNA abundance in the γ5β'2a MBONs after aversive learning might be a consequence of network-level potentiation of their activity, such as would result from a release from inhibition. Since CAMKII local translation-dependent plasticity is expected to underlie more extended forms of memory, it will be interesting to investigate whether the training-evoked change in CaMKII mRNA abundance in the γ5β'2a MBON dendrites contributes to later aversive memory formation and maintenance. This may be possible with MBON-specific targeting of CAMKII mRNAs that contain the long 3'UTR, which is essential for dendritic localization and activity-dependent local translation (Mitchell, 2021).
Lipid shuttling between neurons and glia contributes to the development and function, and stress responses of the nervous system. To understand how a neuron acquires its lipid supply from specific lipoproteins and their receptors, A combined genetic, transcriptome, and biochemical analyses was performed in the developing Drosophila larval brain. This study reports, the astrocyte-derived secreted lipocalin Glial Lazarillo (GLaz), a homolog of human Apolipoprotein D (APOD), and its neuronal receptor, the brain-specific short isoforms of Drosophila lipophorin receptor 1 (LpR1-short), cooperatively mediate neuron-glia lipid shuttling and support dendrite morphogenesis. The isoform specificity of LpR1 defines its distribution, binding partners, and ability to support proper dendrite growth and synaptic connectivity. By demonstrating physical and functional interactions between GLaz/APOD and LpR1, this study elucidated molecular pathways mediating lipid trafficking in the fly brain, and provide in vivo evidence indicating isoform-specific expression of lipoprotein receptors as a key mechanism for regulating cell-type specific lipid recruitment (Yin, 2021).
Lipid trafficking and homeostasis are critical for the development and maintenance of the nervous system. These processes are mediated by a large set of molecular carriers shuttling a diverse group of lipid cargos in and out of designated cell types and cellular compartments. In the central nervous system (CNS), lipid homeostasis heavily relies on neuron-glia cross talk. Studies in mammalian systems have indicated that, besides direct anatomical interactions, glia also supply neurons with metabolic substrates, antioxidants, and trophic factors through secretion. Intriguingly, apolipoproteins are among the most abundant secretory factors that are produced and released by mammalian astrocytes, a group of glial cells with complex morphology and highly branched structures that are intimately associated with synapses, suggesting a critical role for glia-derived lipoprotein and their lipid cargos in synapse formation and function. This notion is supported by studies in cultured mammalian CNS neurons, where glia-derived cholesterol and phospholipids are essential for synaptogenesis. In addition, recent findings in the Drosophila system also indicate essential functions of glia in synapse formation and neurotransmission, although the link between neuron-glia lipid transport and synaptic function has yet to be established (Yin, 2021).
Characterized by their high metabolic rate and elaborate morphology, neurons require a continuous lipid supply throughout their lifetime. How lipoproteins and their receptors mediate neuron-glia lipid shuttling to meet that demand has been a long-standing question in the neurobiology field. Numerous studies over the past decades have demonstrated the critical functions of CNS lipid trafficking in synapse development and cognitive functions. In the mammalian system, deficiencies in either apolipoproteins or their receptors lead to both structural and functional deficits in the brain. For example, Apolipoprotein E (ApoE) delivers cholesterol, amyloid‑β, and other hydrophobic molecules to neurons through its interaction with Very Low-Density Lipoprotein Receptor (VLDLR) and Apolipoprotein E Receptor 2 (ApoER2). While the Apolipoprotein E (ApoE) knockout mice display significantly reduced dendrite size and synapse number as well as impaired learning and memory, both VLDLR and ApoER2 knockout animals also show deficits in cerebellar morphology and impaired contextual fear conditioning and long-term potentiation. Genetic studies of other lipid transport proteins and receptors, including APOD, Niemann-Pick Type C (NPC), Low-Density Lipoprotein Receptor (LDLR), and Low-density lipoprotein Receptor-related Protein 1 (LRP1), further support the importance of lipid trafficking in the proper development and function of the nervous system (Yin, 2021).
Due to the diversity of lipid transport proteins and lipoprotein receptors, as well as the complexity of their tissue- and cell-specific distributions, cellular and molecular mechanisms underlying neuron-glia lipid shuttling have not been well characterized in vivo under physiological conditions. Recent findings in Drosophila highlight the protective functions of neuron-glia metabolic coupling in neurons experiencing oxidative stress or enhanced activity, demonstrating how neurons transfer lipids into glia for detoxification and storage. Similarly, observations made in the mammalian system also provided evidence illustrating fatty acid (FA) transport into astrocytes mediated by ApoE and the importance of neuronal lipid clearance. In contrast, much less is known about how neurons acquire lipid cargos from glia-derived lipoproteins under normal conditions, especially during development, when neurite outgrowth and synapse formation produce a high lipid demand. Attempts were made to fill this gap by determining the functional significance and regulatory mechanisms underlying neuronal lipid uptake using the Drosophila larval brain as a model system (Yin, 2021).
In Drosophila, the Apolipoprotein B (ApoB) family lipoprotein Apolipophorin (ApoLpp) is a major hemolymph lipid carrier and has two closely related lipophorin receptors (LpRs), LpR1 and LpR2, both of which are homologs of mammalian LDLR family proteins. Notably, Drosophila LpRs have multiple isoforms produced by alternative splicing and differential promoter usage. In the fly imaginal disc and oocyte, long isoforms of LpRs (LpR-long) interact with lipid transfer particles (LTP, Apoltp) and mediate endocytosis-independent neutral lipid uptake, while short isoforms of LpRs (LpR-short) neither bind to LTP, nor mediate lipid uptake in these peripheral tissues. In contrast, previous genetic studies revealed specific expression of LpR-short in larval ventral lateral neurons (LNvs), a group of visual projection neurons, and its functions in supporting dendrite development and synaptic transmission (Yin, 2018). This observation is validated by a recent study performed in a cultured Drosophila neuronal cell line, where the LpR-dependent lipid uptake was directly visualized using fluorescently labeled ApoLpp (Matsuo, 2019). Is there a functional significance behind the isoform-specific expression of LpRs? How do short isoforms of LpRs mediate lipid uptake in neurons? These are the questions this study aims to address (Yin, 2021).
This study focused on the LpR1 gene and uncover its isoform-specific expression in neurons and its functions in regulating brain lipid content. Through systematic genetic and biochemical analyses, Glial Lazarillo (GLaz), an astrocyte-derived secreted lipocalin and a homolog of human APOD, was identified as a binding partner for the brain-specific LpR1-short; their cooperative functions are revealed in supporting dendrite morphogenesis, synaptic transmission, and lipid homeostasis in the developing larval brain. In adult Drosophila, GLaz/APOD is found in CNS glia and has been shown to regulate stress resistance and contribute to longevity. Recent studies also demonstrated that GLaz mediates neuron to glia lipid transfer and facilitates neuronal lipid clearance. By identifying GLaz's function in neural development and its direct interaction with LpR1, this study not only uncovered a pair of molecular carriers mediating neuron-glia lipid shuttling in the Drosophila CNS but also presents evidence supporting isoform-specific expression as a key mechanism for regulating the tissue distribution and ligand specificity of neuronal lipoprotein receptors. This in turn leads to the stage- and cell-type-specific regulation of lipid uptake (Yin, 2021).
Lipid-mediated communication between glia and neurons is essential for brain lipid homeostasis and serves critical functions in neural development and synaptic function. Using the developing Drosophila larval brain as a model, this study investigated how neurons acquire their lipid supply from neighboring astrocytes and the regulatory mechanisms associated with the neuron-glia lipid trafficking. Genetic and RNA-seq analyses reveal that lipid uptake in fly neurons is mediated largely by short isoforms of the LpR1 receptor, which recruits a lipid complex through direct interactions with astrocyte-derived apolipoprotein GLaz/APOD. This study identifies specific molecular carriers mediating neuron-glia lipid shuttling and reveals the isoform-specific expression of lipoprotein receptors as a mechanism that determines cell-type-specific recruitment of distinct lipid cargos (Yin, 2021).
Exon mapping of cell-specific RNA-seq libraries revealed that only short isoforms of LpR1 are expressed in LNvs and are upregulated by chronic elevation of neuronal activity. Expanding upon those studies, additional tissue-specific RNA-seq datasets were examined and qFISH analyses was proformed, which strengthened the conclusion that the short-isoforms of LpR1 are CNS-specific and are predominately expressed in neurons, while the long-isoforms of LpR1 are mainly expressed in peripheral tissues and mediate endocytosis-independent lipid uptake. In addition, genetic and biochemical analyses further reveal the functional significance of isoform specificity, including its impact on the receptor's distribution, binding partners, and ability to support specific types of lipid trafficking. These distinctions highlight transcriptional regulation as a key mechanism controlling the cell-type-specific distribution of lipoprotein receptors, their lipid cargos, and uptake efficacy. Results obtained from this study clearly indicate that the molecular and functional diversity of lipoprotein receptors far exceeds current understandings, which are mostly based on DNA sequence analyses. Single-cell transcriptome analyses with sufficient resolution to identify isoform-specific splicing events could potentially reveal the capacity and specificity of the lipid uptake machinery within each individual cell type, helping arrive at a better understanding of the regulatory mechanisms underlying the cell-type-specific lipid recruitment (Yin, 2021).
Lipid shuttling between neurons and glia contributes to energy balance, neural protection, synapse development, and function, and potentially utilizes conserved molecular machinery that is shared among different organisms. Studies in Drosophila and murine neuron-glia coculture systems have demonstrated that neuronal activity alters the metabolic programs of both neurons and glia, leading to the transfer of neuronal lipids into glia for detoxification and storage in the form of lipid droplets. Notably, in both systems, vertebrate ApoE is able to function as the lipid carrier supporting neuronal lipid transfer and lipid droplet accumulation in glia (Yin, 2021).
On the other hand, in the developing larval CNS, this study found an increased lipid demand in neurons coping with chronically elevated input activity. This increase is likely met, at least partially, by enhancing neuronal lipid uptake through the activity-induced upregulation of LpR1 expression. These observations demonstrate a strong influence imposed by activity on neurons' capacity for lipid recruitment through its effects on lipoprotein receptors. Together with earlier studies, these findings suggest that both sides of neuron-glia lipid shuttling are regulated by neuronal activity, and the regulatory mechanisms controlling the expression level of lipoprotein receptors and their interactions with specific ligand molecules likely have functional significance in activity-dependent structural and functional plasticity in the nervous system (Yin, 2021).
Recent studies in adult Drosophila compound eyes have demonstrated that GLaz is involved in lipid transfer from neuron to glia, while the current studies illustrated the interaction between GLaz and LpR1 and a possible role for GLaz in delivering lipid to neurons. Therefore, GLaz appears to be involved in both sides of the lipid trafficking and potentially serves as a key molecular target for regulatory mechanisms controlling lipid homeostasis in the fly brain (Yin, 2021).
The basic structure, molecular composition, and developmental processes of a synapse are shared among vertebrate and invertebrate systems. While synaptogenesis in mammalian neurons relies on cholesterol production by glial cells and its delivery by ApoE-containing lipoprotein complexes, whether the cholesterol and/or ApoE-like lipid carriers are required for building synapses in Drosophila neurons is not known. Importantly, the Drosophila genome does not contain a homolog of ApoE. It is also reported that flies do not have the ability to synthesize cholesterol, and only obtain it through their diet to produce essential hormones. The contrast between these two systems suggests exciting possibilities for studying the lipids involved in synapse construction by understanding the differences and similarities between lipid recruitment in Drosophila vs. mammalian neurons (Yin, 2021).
This study demonstrates that the short isoforms of LpR1 recruit lipids and support dendrite morphogenesis through their interactions with the astrocyte-secreted lipoprotein GLaz, the homolog of human APOD. Given the strong dendrite development phenotype, as well as the reduced life span and stress resistance observed in GLaz loss-of-function mutants, GLaz's hypothesized lipid cargo is likely a critical contributor towards synapse development and neuronal functions in the fly CNS. Currently, only a limited number of putative lipid ligands have been identified for GLaz's homologs; grasshopper Lazarillo binds to retinoic acid and fatty acids, and human APOD binds to arachidonic acid (AA) and progesterone (PG). Whether these lipids or other types of hydrophobic ligands bind to Drosophila GLaz has not been studied. Similar to human APOD, this study observed dimeric and tetrameric GLaz in the larval brain extract. This suggests that, instead of only being able to accommodate a small hydrophobic ligand as a single unit, the GLaz protein could participate in different types of lipoprotein complexes and exhibit distinctive behaviors in vivo through its multimeric forms (Yin, 2021).
APOD's function in the CNS has long been underestimated, despite the fact that APOD is highly elevated during aging and neural injury. When examining recent human and mouse astrocyte RNA-seq data, it was found that, although ApoE is the highest expressing apolipoprotein in mouse astrocytes, the most abundant apolipoprotein expressed in human astrocytes is APOD, strongly suggesting its functional importance in the CNS. In both Drosophila and mammalian systems, GLaz/APOD are produced by astrocytes and have both anti-oxidation and anti-inflammatory protective functions. Given the similarities between GLaz's and APOD's functional and biochemical properties, these findings on the GLaz-LpR1 interaction in Drosophila may facilitate the identification of a mammalian lipoprotein receptor that interacts with APOD and provide new insights into its protective role in aging and neurodegenerative disorders (Yin, 2021).
The adult Drosophila mushroom body (MB) is one of the most extensively studied neural circuits. However, how its circuit organization is established during development is unclear. This study provides an initial characterization of the assembly process of the extrinsic neurons (dopaminergic neurons and MB output neurons) that target the vertical MB lobes. The cellular mechanisms guiding the neurite targeting of these extrinsic neurons were probed, and it was demonstrate that Semaphorin 1a is required in several MB output neurons for their dendritic innervations to three specific MB lobe zones. This study reveals several intriguing molecular and cellular principles governing assembly of the MB circuit (Lin, 2022).
The MB is one of the most intensively studied structures in the fly brain. Its complex and organized circuit architecture has provided important clues to its operational logic. However, in contrast to the extensive investigations of its functions, how the MB circuit architecture is established during development has been little explored. This study provides an initial characterization of MB circuit assembly and identifies Sema1a as an important guidance molecule that directs dendritic innervations of multiple MBONs in three MB lobe zones. Below, several implications of this study relating to the wiring principles of the MB circuit are discussed, and a hypothetical model for how DAN axons and MBON dendrites are modularly assembled into the MB lobes is presented (Lin, 2022).
The most intriguing feature of the organization of the MB circuit is the zonal innervation of the MB lobes by DAN axons and MBON dendrites. The borders of the zones are distinct, with minimal overlap between DAN axons or MBON dendrites in the neighboring zones. Given such a highly organized neural network, elaborate interactions among the extrinsic neurons might be expected. For example, dendritic tiling, as observed between dendritic arborization (da) neurons in fly larvae, might be required for the formation of zonal borders between MBON dendrites, and match-ups between DAN axons and MBON dendrites in the same zone might be important for these neurites to establish proper zonal innervation patterns. However, the results suggest that the targeting and elaboration networks of DAN axons and MBON dendrites are largely independent, at least for those projecting to the MB vertical lobes. In the α'2 zone, where innervation by DAN axons precedes that by MBON dendrites, ablation of DANs does not affect zonal elaboration of the MBON dendrites. Moreover, upon ablation of one type of DAN or MBON in a given zone, morphologies of the neighboring neurites appear to be normal. Therefore, the extent and location of zonal network elaboration by DAN axons and MBON dendrites in the vertical lobes do not depend on interactions between these extrinsic neurons (Lin, 2022).
In each MB lobe zone, the DAN axons and MBON dendrites form synapses with each other and the KC axons. Given that DANs and MBONs do not depend on each other to form zonal networks, could KCs be responsible? The results support the importance of the KCs in the zonal organization of the DAN axons and MBON dendrites. Aberrant branching of KCs in alpha lobes absent (ala) mutant brains resulted in some MBs lacking the vertical lobes. When this occurred, most DAN axons and MBON dendrites that normally innervate these lobes do not form zonal arborization. Without the MB vertical lobes, MBON-α'2 dendrites are rerouted to other zones in the horizontal lobes and form potential synaptic connections with the local DAN axons. Importantly, this reorganization of the MBON dendrites requires the presence of the KCs (Lin, 2022).
It is still possible that the KC axons and the lobes they form simply provide an anchoring point on which DAN axons and MBON dendrites grow, and that the extent of their arborization is determined cell-autonomously as an intrinsic property. However, the axonal innervation pattern of PPL1 DANs argues against this possibility. PPL1-α'3 and PPL1-α'2α2 axons enter the MB vertical lobes at almost the same location but specifically occupy distinct zones on opposite sides of the entry point, suggesting the existence of local positional cues in the lobes to guide the innervation of DAN axons. Furthermore, overexpression of sema1a in DANs directs their dendrites to specific MB lobe zones, and importantly, the arborization of these rerouted dendrites is confined to their respective zones. Since these DAN dendrites normally do not innervate the MB lobe, there likely exist local positional cues that interact with Sema1a-expressing dendrites to guide their zonal arborization (Lin, 2022).
What could be the sources of these positional cues? KCs are good candidates because they synapse with DAN axons and MBON dendrites and are essential for zonal arborization of these neurites. However, since KCs provide the main framework of the MB lobes, their manipulation may affect the organization of other cell types in the MB lobes that could also be potential sources of the positional cues. Electron microscopy-based reconstructions of the MB circuit have provided a comprehensive catalog for the neurons that innervate the MB lobe. In addition to KCs, DANs, and MBONs, the MB lobes are innervated by one dorsal paired medial (DPM) neuron, one anterior paired lateral (APL) neuron, two SIFamide-expressing neurons, and two octopaminergic neurons. These neurons do not exhibit zonal innervation patterns in the MB lobes; the SIFamide- and octopamine-expressing neurons only sparsely innervate the MB lobes, and the neurites of APL and DPM ramify the entire MB lobes. The MB lobes are also populated by glia, which is another potential source of the positional cues. Although their sources remain undetermined, the results suggest that the MB lobes are likely prepatterned with positional cues to guide the zonal elaboration of the MBON and DAN neurites (Lin, 2022).
Sema1a is an evolutionarily conserved guidance molecule that functions as a ligand or receptor depending on the cellular context. The data suggest that Sema1a functions as a receptor in MBONs to regulate dendritic targeting in a zone-specific manner. Loss of sema1a activity preferentially affects MBON dendrites in the α'3, α'1, and β'2 zones. Even for MBONs that innervate multiple zones, such as MBON-β2β'2a and MBON-γ5β'2a, reducing sema1a activity in these neurons selectively impacts their dendrites in the β'2a zone. Since Sema1a is not differentially localized in the dendrites of these MBONs, the guidance cues that Sema1a responds to might primarily be present in the β'2a zone, with additional guidance signals working collaboratively to sort the dendrites from these MBONs into Sema1a-sensitive and -insensitive zones. Not all MBONs innervating these Sema1a-sensitive zones require Sema1a. For MBON-γ2α'1 and MBON-α'1 that both innervate the α'1 zone, loss of sema1a only affects dendritic innervation by MBON-α'1 but not MBON-γ2α'1. Therefore, how Sema1a functions is also cell-type-specific. Taken together, these results imply that multiple positional cues may be present in each MB lobe zone, with each MBON being equipped with multiple sensors that work in concert to respond to those cues. Moreover, given that Sema1a is broadly expressed in many neurons in the developing brain, including the KCs, Sema1a likely acts with other proteins or signaling molecules to determine guidance specificity (Lin, 2022).
Sema1a has been shown to mediate both neurite attraction and avoidance. Currently, it is unclear which of the two mechanisms underlies its guidance of MBON dendrites. For MBONs whose zonal dendritic innervation requires sema1a, their dendrites can still project to areas nearby their target zones when sema1a activity was removed. Hence, the cues that guide these MBON dendrites are likely to be short-ranged. However, overexpression of sema1a in PPL1-α'2α2 DANs can redirect their dendrites to innervate zones far away from their original location, suggesting that the guidance cues may also exert long-distance functionality. The data indicate that Sema1a functions as a receptor in MBONs. Identification of Sema1a ligands and determining their distributions in the MB lobes are critical steps toward understanding how Sema1a instructs the zonal innervation of MBON dendrites. Plexin A (PlexA) or secreted Semaphorin 2a and 2b (Sema2a and Sema2b) are known ligands for the Sema1a receptor. This study has tested if these canonical ligands of Sema1a are required for the dendritic innervations of β'2- and α'3-projecting MBONs. However, dendritic innervations by these MBONs were minimally affected in homozygous sema2a/2b double mutant flies or when PlexA was knocked down either pan-neuronally or in glia. Therefore, the canonical Sema1a ligands do not seem to play a role in MBON dendritic targeting. However, it remains to be determined if PlexA and Sema2a/2b function redundantly in this system or if an unidentified noncanonical ligand is involved (Lin, 2022).
Although the molecular nature of the positional cues in the MB lobes that organize the zonal patterns of DAN and MBON neurites awaits discovery, the data suggest that these cues likely work in a combinatorial manner. Supporting evidence for this notion comes from the observation that MBON-α'1 and MBON-γ2α'1 use sema1a-dependent and -independent mechanisms to innervate the α'1 zone, indicating that this zone may present at least two different guidance cues. Furthermore, sema1a is expressed in multiple MBONs that innervate distinct MB lobe zones. This pattern could potentially be explained if the positional cues attracting Sema1a-positive neurites appear sequentially in these zones (i.e., so that the zone an MBON innervates is determined by the developmental timing of the MBON). However, the finding that the ectopic innervations of MBON-α'2 in the β'2 and α'1-like zones of the ala brain occur simultaneously argues against that possibility. Therefore, the Sema1a-sensitive zones likely harbor additional zone-specific guidance cues that work in combination with Sema1a to diversify guidance specificity (Lin, 2022).
The observation that the mistargeted MBON-α'2 dendrites in ala mutant brains innervate other zones in the α'β' lobe, but not those in the αβ and γ lobes, has also prompted a hypothesis that there might be general attraction cues emanating from the α'β' lobes for all α'β' lobe-projecting MBONs, separating them from MBONs targeting αβ and γ lobes. Therefore, a hypothetical model is proposed whereby multiple hierarchically-organized positional cues are presented in the MB lobe zones, with these cues acting in concert to pattern zonal innervation by DAN axons and MBON dendrites in the MB lobes (Lin, 2022).
Membrane trafficking is essential for sculpting neuronal morphology. The GARP and EARP complexes are conserved tethers that regulate vesicle trafficking in the secretory and endolysosomal pathways, respectively. Both complexes contain the Vps51, Vps52, and Vps53 proteins, and a complex-specific protein: Vps54 in GARP and Vps50 in EARP. In Drosophila, both complexes were found to be required for dendrite morphogenesis during developmental remodeling of multidendritic class IV da (c4da) neurons. Having found that sterol accumulates at the trans-Golgi network (TGN) in Vps54KO/KO neurons, genes that regulate sterols and related lipids at the TGN were investigated. Overexpression of oxysterol binding protein (Osbp) or knockdown of the PI4K four wheel drive (fwd) exacerbates the Vps54KO/KO phenotype, whereas eliminating one allele of Osbp rescues it, suggesting that excess sterol accumulation at the TGN is, in part, responsible for inhibiting dendrite regrowth. These findings distinguish the GARP and EARP complexes in neurodevelopment and implicate vesicle trafficking and lipid transfer pathways in dendrite morphogenesis (O'Brien, 2023).
Xie, Q., Li, J., Li, H., Udeshi, N. D., Svinkina, T., Orlin, D., Kohani, S., Guajardo, R., Mani, D. R., Xu, C., Li, T., Han, S., Wei, W., Shuster, S. A., Luginbuhl, D. J., Quake, S. R., Murthy, S. E., Ting, A. Y., Carr, S. A. and Luo, L. (2022). Transcription factor Acj6 controls dendrite targeting via a combinatorial cell-surface code. Neuron. PubMed ID: 35613619
Transcription factors specify the fate and connectivity of developing neurons. This study investigated how a lineage-specific transcription factor, Acj6, controls the precise dendrite targeting of Drosophila olfactory projection neurons (PNs) by regulating the expression of cell-surface proteins. Quantitative cell-surface proteomic profiling of wild-type and acj6 mutant PNs in intact developing brains, and a proteome-informed genetic screen identified PN surface proteins that execute Acj6-regulated wiring decisions. These include canonical cell adhesion molecules and proteins previously not associated with wiring, such as Piezo, whose mechanosensitive ion channel activity is dispensable for its function in PN dendrite targeting. Comprehensive genetic analyses revealed that Acj6 employs unique sets of cell-surface proteins in different PN types for dendrite targeting. Combined expression of Acj6 wiring executors rescued acj6 mutant phenotypes with higher efficacy and breadth than expression of individual executors. Thus, Acj6 controls wiring specificity of different neuron types by specifying distinct combinatorial expression of cell-surface executors (Xie, 2022).
Developmental remodeling of neurite is crucial for the accurate wiring of neural circuits in the developing nervous system in both vertebrates and invertebrates, and may also contribute to the pathogenesis of neuropsychiatric disorders, for instance, autism, Alzheimer's disease (AD), and schizophrenia. However, the molecular underpinnings underlying developmental remodeling are still not fully understood. This study has identified DnaJ-like-2 (Droj2), orthologous to human DNAJA1 and DNAJA4 that is predicted to be involved in protein refolding, as a developmental signal promoting dendrite sculpting of the class IV dendritic arborization (C4da) sensory neuron in Drosophila. It was further shown that Arf102F, a GTP-binding protein previously implicated in protein trafficking, serves downstream of Droj2 to govern neurite pruning of C4da sensory neurons. Intriguingly, these data consistently demonstrate that both Droj2 and Arf102F promote the downregulation of the conserved L1-type cell-adhesion molecule Neuroglian anterior to dendrite pruning. Mechanistically, Droj2 genetically interacts with Arf102F and promotes Neuroglian downregulation to initiate dendrite severing. Taken together, this systematic study sheds light on an unprecedented function of Droj2 and Arf102F in neuronal development (Rui, 2023).
Neuronal target recognition is performed by numerous cell-surface transmembrane proteins. Correct folding of these proteins occurs in the endoplasmic reticulum (ER) lumen of the neuronal cells before being transported to the plasma membrane of axons or dendrites. Disturbance in this protein folding process in the ER leads to dysfunction of neuronal cell surface molecules, resulting in abnormal neuronal targeting. This study reports that the ER-resident protein Meigo in Drosophila, governs the dendrite targeting of olfactory projection neurons (PNs) along the mediolateral axis of the antennal lobe by regulating Toll-6 localization. Loss of Meigo causes Toll-6 mislocalization in the PNs and mediolateral dendrite targeting defects, which are suppressed by Toll-6 overexpression. Furthermore, this study found that the ER-chaperone protein, Gp93, also regulates the mediolateral targeting of PN dendrites by localization of the Toll-6 protein. Gp93 overexpression in the PN homozygous for the meigo mutation, partially rescued the dendrite targeting defect, while meigo knockdown decreased Gp93 expression levels in cultured cells. These results indicate that the ER-proteins Meigo and Gp93 regulate dendrite targeting by attenuating the amount and localization of cell surface receptors, including Toll-6, implying the unexpected but active involvement of ER proteins in neural wiring (Kamemura, 2022).
Mitochondria are key contributors to the etiology of diseases associated
with neuromuscular defects or neurodegeneration. How changes in cellular
metabolism specifically impact neuronal intracellular processes and cause
neuropathological events is still unclear. This study dissects the
molecular mechanism by which mitochondrial
dysfunction induced by Prel
aberrant function mediates selective dendritic
loss in Drosophila melanogaster class IV dendritic arborization
neurons. Using in vivo ATP imaging, it was found that neuronal cellular
ATP levels during development are not correlated with the progression of
dendritic loss. By searching for mitochondrial stress signaling pathways
that induce dendritic loss it was found that mitochondrial dysfunction is
associated with increased eIF2α
phosphorylation, which is sufficient to induce dendritic pathology in
class IV arborization neurons. It was also observed that eIF2α
phosphorylation mediates dendritic loss when mitochondrial dysfunction
results from other genetic perturbations. Furthermore, mitochondrial
dysfunction induces translation repression in class IV neurons in an eIF2α
phosphorylation-dependent manner, suggesting that differential translation
attenuation among neuron subtypes is a determinant of preferential
vulnerability (Tsuyama, 2017).
Neurons receive information along dendrites and send signals along axons to synaptic contacts. The factors that control axon regeneration have been examined in many systems, but dendrite regeneration has been largely unexplored. This study reports that, in intact Drosophila larvae, a discrete injury that removes all dendrites induces robust dendritic growth that recreates many features of uninjured dendrites, including the number of dendrite branches that regenerate and responsiveness to sensory stimuli. However, the growth and patterning of injury-induced dendrites is significantly different from uninjured dendrites. Regenerated arbors cover much less territory than uninjured neurons, fail to avoid crossing over other branches from the same neuron, respond less strongly to mechanical stimuli, and are pruned precociously. Finally, silencing the electrical activity of the neurons specifically blocks injury-induced, but not developmental, dendrite growth. By elucidating the essential features of dendrites grown in response to acute injury, this work builds a framework for exploring dendrite regeneration in physiological and pathological conditions (Thompson-Peer, 2016).
After injury, severed dendrites and axons expose the 'eat-me' signal phosphatidylserine (PS) on their surface while they break down. The degeneration of injured axons is controlled by a conserved Wallerian degeneration (WD) pathway, which is thought to activate neurite self-destruction through Sarm-mediated nicotinamide adenine dinucleotide (NAD(+)) depletion. While neurite PS exposure is known to be affected by genetic manipulations of NAD(+), how the WD pathway coordinates both neurite PS exposure and self-destruction and whether PS-induced phagocytosis contributes to neurite breakdown in vivo remain unknown. This study shows that in Drosophila sensory dendrites, PS exposure and self-destruction are two sequential steps of WD resulting from Sarm activation. Surprisingly, phagocytosis is the main driver of dendrite degeneration induced by both genetic NAD(+) disruptions and injury. However, unlike neuronal Nmnat loss, which triggers PS exposure only and results in phagocytosis-dependent dendrite degeneration, injury activates both PS exposure and self-destruction as two redundant means of dendrite degeneration. Furthermore, the axon-death factor Axed is only partially required for self-destruction of injured dendrites, acting in parallel with PS-induced phagocytosis. Lastly, injured dendrites exhibit a unique rhythmic calcium-flashing that correlates with WD. Therefore, both NAD(+)-related general mechanisms and dendrite-specific programs govern PS exposure and self-destruction in injury-induced dendrite degeneration in vivo (Ji, 2022).
Physical insults to the nervous system often disrupt neuronal connectivity and function by damaging dendritic or axonal processes of neurons. Injured axons break down through a series of stereotypical events collectively called Wallerian degeneration (WD). Dendrites undergo a similar program of degeneration after injury (AI). Before neurons can regenerate their processes and restore connections, the debris from damaged neurites has to be promptly cleared by phagocytes, which are cells that engulf dead cells or cell debris. Inefficient clearance can lead to neuroinflammation and further exacerbate the damage to the surrounding tissues. Although WD is mainly considered to be a neurite-intrinsic, self-destructive process, whether phagocytosis actively contributes to degeneration of injured neurites in vivo, rather than merely passively removing neuronal debris, remains unclear (Ji, 2022).
WD is governed by an evolutionarily conserved pathway, which is also called 'axon-death' pathway, because it was discovered in studies focused primarily on axon degeneration in Drosophila and rodents. In this pathway, injury induces activation of the E3 ubiquitin ligase Highwire/Phr1 in severed axons, which in turn causes degradation of nicotinamide mononucleotide adenyltransferase (Nmnat), an enzyme required for the synthesis of nicotinamide adenine dinucleotide (NAD+). The decrease of NAD+ resulting from Nmnat degradation, together with an accumulation of the NAD+ precursor nicotinamide mononucleotide (NMN), activates Sarm/SARM1, a sterile alpha/Armadillo/Toll-Interleukin receptor homology domain protein. Due to its NADase activity, Sarm/SARM1 is thought to subsequently cause local NAD+ depletion in injured axons. In Drosophila, axundead (axed) is required downstream of Sarm for axon degeneration of olfactory receptor neurons and wing sensory neurons. The loss of axed blocks axon degeneration even when Sarm is dominantly activated, suggesting that Axed activation is a key switch of WD. In addition, pebbled (peb) encodes a Drosophila transcription factor required for axon degeneration of glutamatergic but not cholinergic sensory neurons in the wing. Although how exactly Axed and/or NAD+ depletion lead to axon breakdown is still mysterious, it is generally believed that Sarm activity initiates a neurite-intrinsic self-destruction program that ultimately is responsible for WD of axons. While the WD pathway is primarily characterized in axons, evidence suggests that NAD+ reduction is also an essential step in injury-induced dendrite degeneration. However, which components of the WD pathway are conserved in dendrites remains unknown (Ji, 2022).
Neuronal debris is recognized by resident phagocytes of the nervous system through specific 'eat-me' signals exposed on the neuronal surface. A highly conserved eat-me signal is phosphatidylserine (PS), a negatively charged phospholipid normally found in the inner leaflet of the plasma membrane of healthy cells. During apoptosis, PS is externalized to the outer leaflet of the plasma membrane to mark the cell for engulfment (Sapar, 2018). Genetic analyses of certain PS-binding bridging molecules and cell membrane receptors in mice and zebrafish suggest that PS recognition contributes to the phagocytosis of neurons. Similarly, clearance of injured axons and dendrites in Drosophila requires Draper (Drpr), an engulfment receptor that binds to PS. PS exposure has also been directly observed on injured axons of mouse neurons in vitro and on injured dendrites of Drosophila sensory neurons in vivo. Although axonal PS exposure can be induced independently of axon degeneration in vitro, ectopically induced PS exposure resulted in engulfment-dependent neurite reduction of otherwise healthy neurons in both the central nervous system and the peripheral nervous system (PNS) of Drosophila, pointing to a dominant effect of PS exposure in inducing phagocytosis. Recent studies revealed a link between neuronal PS exposure and the WD pathway. Overexpression (OE) of WldS, a fusion protein containing the full-length murine Nmnat1, in Drosophila sensory neurons suppresses PS exposure of injured dendrites. In addition, Sarm1 ablation and NAD+ supplementation in neuronal culture reduce PS exposure on injured axons. These observations raise two important questions: How does the WD pathway regulate and coordinate both neurite PS exposure and self-destruction? What are the relative contributions of PS-mediated phagocytosis and neurite self-destruction in WD in vivo (Ji, 2022)?
To address these questions, Drosophila class IV dendritic arborization (C4da) neurons on the larval body wall, an established in vivo model of injury-induced dendrite degeneration, were used. In this system, degenerating dendrites of C4da neurons are phagocytosed by epidermal cells through the engulfment receptor Drpr. This study shows that PS exposure and dendrite self-destruction are two distinct steps downstream of Sarm activation and that in vivo, phagocytosis is the main driving force of dendrite degeneration induced by injury and Nmnat loss-of-function (LOF). Furthermore, unlike in axons, Axed is not required for dendrite degeneration: it contributes to injury-induced dendrite self-destruction but is not involved in dendrite PS exposure. Lastly, dendrite degenerations induced by injury and Nmnat LOF differ in phagocytosis dependency, membrane disruption, and dendrite calcium dynamics. Injured dendrites exhibit a unique rhythmic calcium-flashing, which is disrupted by WldS OE and axed loss, indicating a potential role of calcium-flashing in dendrite self-destruction (Ji, 2022).
This study investigated mechanisms of dendrite degenerations caused by injury and genetic activation of the WD pathway in the Drosophila PNS. Although neurite phagocytosis has been observed after neuronal injury both in vivo and in vitro, WD is generally considered as a result of neurite self-destruction triggered by NAD+ depletion. However, the current results using engulfment-deficient drpr LOF strongly suggest that PS-mediated phagocytosis is the main driving force for WD-related dendrite degeneration in vivo. It is solely responsible for dendrite degeneration of Nmnat KO neurons and greatly accelerates the degeneration of SarmGOF OE neurons. In injury, phagocytosis is responsible for at least half of the dendrite fragmentation by 10 h AI and may contribute more to dendrite breakdown than self-destruction at earlier stages. Supporting this idea, ectopic PS exposure on injured dendrites is sufficient to revert the blockage of dendrite fragmentation by WldS OE or Sarm KO, even though ectopically induced PS exposure is much lower than natural PS exposure on injured dendrites. Time-lapse analyses of PS exposure, the final calcium surge, and dendrite rupture also support the possibility that PS-mediated phagocytosis breaks down injured dendrites at the time of dendrite fragmentation. Therefore, at least in the context of dendrite injury, phagocytosis is the major factor driving WD, while self-destruction acts as a secondary mechanism to ensure complete fragmentation (Ji, 2022).
NAD+ reduction is thought to be responsible for neuronal PS exposure and neurite self-destruction during WD. How may NAD+ reductions coordinate the two different events? The results of this study support a hypothesis that NAD+ disruption controls PS exposure and neurite self-destruction in two separate steps of WD. In the existing model, Sarm activation is believed to cause catastrophic NAD+ depletion that is sufficient to initiate neurite self-destruction. However, this study found that downstream of Sarm activation and before the initiation of self-destruction, neurites first expose PS to engage in phagocytosis-mediated nonautonomous degeneration. Therefore, in a revised model, dendrites respond to at least three distinct, increasingly severe levels of NAD+ reduction by eliciting different molecular events. Between the NAD+ level required for Sarm activation (SA level) and the level that initiates self-destruction (the self-destruction level), Sarm activity lowers NAD+ to a level that causes neurons to expose PS on their surface (which is called the PSE level). This PS exposure is sufficient to cause phagocytosis-mediated dendrite degeneration, which can be completely prevented by blocking engulfment activity of phagocytes. However, below the self-destruction level, neurites spontaneously fragment even in the absence of phagocytosis (Ji, 2022).
This model is consistent with an apparent correlation between the expected kinetics of NAD+ reduction and the severity of neurite degeneration. Nmnat KO is expected to cause slow NAD+ reduction, due to gene perdurance and the time required for natural NAD+ turnover, and correspondingly causes engulfment-dependent dendrite degeneration only in late third instar larvae. In contrast, SarmGOF OE should lead to a more-rapid NAD+ depletion and in fact causes engulfment-dependent dendrite degeneration as early as the first instar and dendrite self-destruction by the third instar. Injury is known to cause even more rapid NAD+ reduction in axons (20) and is correlated with the fastest dendrite degeneration-initiation at around 4 h AI and completion usually by 10 h AI. Directly validating this model will likely require a sensitive NAD+ indicator that can measure NAD+ levels in dendrites in vivo (Ji, 2022).
How may NAD+ reduction cause PS exposure? A direct consequence of NAD+ loss is the decline of neurite ATP levels due to the requirement of NAD+ in glycolysis and oxidative phosphorylation. Consistent with ATP reduction playing a role in inducing PS exposure, suppressing mitochondria ATP synthesis in dorsal root ganglia (DRG) culture caused gradual axonal PS exposure. However, how ATP reduction may induce PS exposure remains elusive. Although the maintenance of membrane PS asymmetry by flippases requires ATP, flippase KO in C4da neurons causes a much-milder PS exposure than injury, suggesting that mechanisms other than flippase inhibition must be contributing to the rapid PS exposure seen AI. Identifying the PS transporters responsible for PS exposure on injured neurites will be a key step for revealing the mechanisms of NAD+ regulation of PS exposure (Ji, 2022).
Genetic analyses in Drosophila identified Axed as a key switch of WD, whose activity is absolutely required for axon degeneration caused by injury and Sarm GOF. How does Axed regulate neurite degeneration? The data suggest that Axed is not required for PS-mediated phagocytosis but contributes to the self-destruction of injured dendrites, placing its activation below the self-destruction level of NAD+ in the model. Surprisingly, Axed seems to play a minor role in dendrite degeneration, as its LOF only slowed down but did not block self-destruction, indicating the existence of other factors that promote self-destruction of injured dendrites (Ji, 2022).
By exploring the different mechanisms employed in Nmnat KO- and injury-induced dendrite degenerations, dynamic calcium activities were discovered that are only present in injured dendrites, including a calcium-flashing pattern prior to any obvious degenerative event and a final calcium surge that coincides with dendrite fragmentation. Calcium surge at the time of neurite fragmentation is a shared feature between injured axons of zebrafish and injured dendrites of Drosophila da neurons. Although calcium influx is required for WD and may activate calcium-dependent lipid scramblases, time-lapse analyses suggest that the final calcium surge is more likely a result of phagocytosis-induced membrane rupture rather than the cause of fragmentation. In comparison, the calcium-flashing soon after the injury is unique to dendrites and may play an active role in dendrite degeneration in ways similar to the compartmentalized calcium-flashing that occurs during developmental pruning of C4da neurons. Consistent with this possibility, the calcium-flashing is suppressed by WldS OE and axed KO. Interestingly, these two manipulations block the calcium-flashing in opposite ways, with WldS OE dampening the calcium level and axed KO keeping the calcium level high. This distinction may be a useful clue for understanding the regulation of PS exposure and self-destruction. For example, it is possible that elevated calcium levels prepare dendrites for PS exposure and drastic changes of calcium levels promote dendrite self-destruction (Ji, 2022).
Previous studies have shown that axons and dendrites share common features in neurite degeneration: they both undergo PS exposure and WD AI, they are both subject to PS exposure-induced degeneration, and injury-induced PS exposure and degeneration of both can be blocked by manipulations to increase NAD+ levels. These analyses on Nmnat and Sarm additionally show that dendrites are similar to axons in SARM-dependent spontaneous degeneration associated with NMNAT deficiency. Unexpectedly, the results also reveal important differences between dendrites and axons. First, while Axed is absolutely required for axon degeneration in injury and in Sarm GOF, it is only partially involved in dendrite degeneration AI. Second, calcium-flashing is found in dendrites that are injured or undergoing developmental pruning but not in injured axons. These results suggest that dendrites and axons utilize both shared and neurite type-specific programs in degeneration.
Lastly, as impairment of NAD+ metabolism is a general feature of neurodegenerative disorders including Leber congenital amaurosis, Alzheimer's disease, Parkinson's disease, and retinal degenerations, phagocytosis may play important roles in the pathogenesis of these diseases through dysregulated neuronal PS exposure (Ji, 2022).
Phagocytic clearance of degenerating neurons is triggered by "eat-me" signals exposed on the neuronal surface. The conserved neuronal eat-me signal phosphatidylserine (PS) and the engulfment receptor Draper (Drpr) mediate phagocytosis of degenerating neurons in Drosophila. However, how PS is recognized by Drpr-expressing phagocytes in vivo remains poorly understood. Using multiple models of dendrite degeneration, this study shows that the Drosophila chemokine-like protein Orion can bind to PS and is responsible for detecting PS exposure on neurons; it is supplied cell-non-autonomously to coat PS-exposing dendrites and to mediate interactions between PS and Drpr, thus enabling phagocytosis. As a result, the accumulation of Orion on neurons and on phagocytes produces opposite outcomes by potentiating and suppressing phagocytosis, respectively. Moreover, the Orion dosage is a key determinant of the sensitivity of phagocytes to PS exposed on neurons. Lastly, mutagenesis analyses show that the sequence motifs shared between Orion and human immunomodulatory proteins are important for Orion function. Thus, these results uncover a missing link in PS-mediated phagocytosis in Drosophila and imply conserved mechanisms of phagocytosis of neurons (Ji, 2023).
Phagocytic clearance of degenerating neurons is triggered by 'eat-me' signals exposed on the neuronal surface. The conserved neuronal eat-me signal phosphatidylserine (PS) and the engulfment receptor Draper (Drpr) mediate phagocytosis of degenerating neurons in Drosophila. However, how PS is recognized by Drpr-expressing phagocytes in vivo remains poorly understood. Using multiple models of dendrite degeneration, this study shows that the Drosophila chemokine-like protein orion can bind to PS and is responsible for detecting PS exposure on neurons; it is supplied cell-non-autonomously to coat PS-exposing dendrites and to mediate interactions between PS and Drpr, thus enabling phagocytosis. As a result, the accumulation of orion on neurons and on phagocytes produces opposite outcomes by potentiating and suppressing phagocytosis, respectively. Moreover, the orion dosage is a key determinant of the sensitivity of phagocytes to PS exposed on neurons. Lastly, mutagenesis analyses show that the sequence motifs shared between orion and human immunomodulatory proteins are important for orion function. Thus, these results uncover a missing link in PS-mediated phagocytosis in Drosophila and imply conserved mechanisms of phagocytosis of neurons (Ji, 2023).
Phagocytosis of apoptotic and degenerative neurons is essential for the development and homeostasis of the nervous system. Abnormal phagocytosis is also associated with neuroinflammation and neurodegenerative diseases. Neuronal debris is recognized and cleared by resident phagocytes of the nervous system through 'eat-me' signals exposed on the neuronal surface. A conserved 'eat-me' signal is phosphatidylserine (PS), a negatively charged phospholipid normally kept in the inner leaflet of the plasma membrane by P4-ATPase flippases. During neurite degeneration and apoptosis, PS is externalized to the outer surface of neuronal membranes. The exposed PS dominantly triggers phagocytosis of neurons. Besides clearing neuronal debris, PS-mediated phagocytosis also drives the degeneration of injured neurites and neurons with certain genetic perturbations. In the central nervous system (CNS), local PS exposure enables microglia-mediated synaptic elimination. Thus, the regulation and recognition of neuronal PS exposure are critical for the development and homeostasis of the nervous system (Ji, 2023).
Drosophila has been an important model organism for studying neuronal phagocytosis. In Drosophila, Draper (Drpr) is the best-known receptor responsible for phagocytosis of neurons. As a homolog of the Caenorhabditis elegans engulfment receptor CED-1 and the mammalian engulfment receptors Jedi-1 and MEGF10, Drpr is involved in many contexts of neuronal phagocytosis, including the clearance of apoptotic neurons during embryonic development, axon and dendrite pruning during neuronal remodeling, injury-induced neurite degeneration, and removal of destabilized boutons at neuromuscular junctions. Despite the well-known importance of Drpr in sculpting the nervous system, how Drpr recognizes degenerating neurons in vivo is still unclear (Ji, 2023).
Recently, the secreted protein orion was discovered as being required for the developmental pruning and clearance of Drosophila mushroom body (MB) axons. orion shares a CX3C motif with mammalian CX3CL1 (also known as fractalkine), which is required for the elimination of synapses in the mouse barrel cortex. CX3CL1 is known as a chemokine because of its ability to direct migration of leukocytes and microglia. Thus, orion represents the first known chemokine-like molecule in Drosophila and shares conserved functions with CX3CL1 in the remodeling of the nervous system. However, how orion and CX3CL1 function in the phagocytosis of neurons is still unknown (Ji, 2023).
This study examined orion's function in the phagocytosis of Drosophila class IV dendritic arborization (C4da) neurons, a well-established in vivo model for studying PS-mediated phagocytosis. In vivo evidence is presented that the transmembrane engulfment receptor Drpr relies on orion to sense PS on neurons. orion is secreted by peripheral nonneural tissues and serves as a nonautonomous permissive factor for phagocytosis of neurons. orion binds to PS-exposing neurons and is also enriched on phagocytes overexpressing Drpr. Strikingly, a membrane-tethered orion expressed by neurons dominantly induces phagocytosis even in the absence of PS exposure, while orion accumulation on the surface of phagocytes makes phagocytes blind to PS-exposing neurons. Importantly, the dosage of orion determines the sensitivity of phagocytes to neuronal PS exposure. Lastly, this study established that the motifs orion shares with human chemokines and neutrophil peptides are critical for engulfment of PS-exposing neurons. These findings reveal key mechanisms of PS recognition in Drosophila and imply potentially conserved roles of chemokines in PS-mediated phagocytosis of neurons (Ji, 2023).
By triggering phagocytosis, the recognition of PS exposed on neurons is a critical event during neurodegeneration and clearance. Although several studies have implied the involvement of Drpr in this process in Drosophila, how Drpr mediates PS recognition in vivo is unclear. This study presents several lines of in vivo evidence that strongly indicate that the Drosophila chemokine-like orion is a PS-binding bridging molecule that enables Drpr to respond to neuronal PS exposure. First, orion is required for multiple scenarios of Drpr-dependent phagocytosis of sensory dendrites and functions upstream of Drpr. Second, orion binds to PS-exposing cell surfaces. This study shows that orion binds to neurons and epidermal cells that expose PS as a result of tissue-specific KO of the PS flippase ATP8A. In addition, ATP8A OE, which retains PS in the inner membrane leaflet, eliminates orion binding on epidermal cells and late-larval dendrites, suggesting that this binding is PS-dependent. Importantly, orion outcompetes Annexin V for binding to injured dendrites, suggesting that orion may directly interact with PS in vivo. Third, overexpressed Drpr proteins can trap orion on the cell surface, suggesting that Drpr interacts with orion in vivo. Lastly, when expressed in neurons, membrane-tethered orion bypasses the requirement for PS in inducing Drpr-dependent engulfment, but when expressed in phagocytes, membrane-tethered orion blocks PS-induced engulfment. Based on these observations, it is proposed that orion functions as a bridging molecule between PS and Drpr (Ji, 2023).
Previously, SIMU, a PS-binding transmembrane protein expressed by Drosophila embryonic phagocytes, was proposed to be a bridging molecule. orion and SIMU contribute to phagocytosis through distinct mechanisms. First, SIMU is expressed by phagocytes to allow them to tether apoptotic cells, while orion is secreted from many peripheral tissues and functions as an opsonin to enable phagocytosis. Second, SIMU is a membrane protein that shares homology with Drpr but functions at a different step in apoptotic neuron clearance compared to Drpr. In contrast, as a secreted protein, orion functions at the same step of phagocytosis as Drpr. Therefore, SIMU behaves more like a tethering receptor, while orion is functionally analogous to PS-bridging molecules in other species. Although the analyses on the engulfment of somatosensory dendrites, the ubiquitous roles of PS and Drpr in phagocytosis and the broad expression patterns of orion suggest that orion may be widely involved in PS-mediated phagocytosis in Drosophila. This view is supported by findings that orion deposited in the hemolymph can mediate phagocytosis in distant tissues and that the accumulation of orion on epidermal cells turns these cells into targets of phagocytosis (Ji, 2023).
Although the role of PS exposure in inducing phagocytosis has been well documented, what determines the sensitivity of phagocytes to PS is much less understood. This study discovered that the available level of orion is a determinant of phagocyte sensitivity to PS in Drosophila. Reducing the dosage of functional orion by one half makes phagocytes blind to dendrites with moderate levels of PS exposure, but the reduced orion does not affect the ability of phagocytes to engulf dendrites that display high levels of PS exposure (i.e., injury). Conversely, an extra copy of the orion locus enhances the ability of phagocytes to engulf dendrites that have mild PS exposure (i.e., CDC50 KO). These results suggest that endogenous orion is likely expressed at a finely balanced level to enable the right amount of phagocytosis: Too much orion may cause unintended phagocytosis of stressed cells that display mild PS exposure, while too little orion may interfere with efficient clearance of sick cells or structures that are beyond rescue. Consistent with this idea, endogenous orion is expressed at a low level during larval development, but is dramatically up-regulated during metamorphosis, a time when large-scale tissue remodeling and clearance take place (Ji, 2023).
orion is required for axonal pruning and clearance of MB γ Kenyon neurons during metamorphosis. In that context, orion is expressed in the remodeling MB neurons and functions as a 'find-me' signal for glia to penetrate the axon bundles and engulf axonal debris. In contrast, in the larval peripheral nervous system (PNS), orion is supplied by many nonneural tissues and functions as a permissive signal for phagocytosis of sick or broken dendrites. This distinction is likely due to two differences between the larval PNS and the remodeling CNS. First, in the PNS, the dendrites of da neurons are exposed to the hemolymph and are readily accessible to orion secreted from other tissues, whereas in the CNS, the axons are more tightly packed and may be harder for extrinsic orion to access. Neuron-derived orion would thus be more effective than extrinsic orion for promoting phagocytosis in the CNS. Intriguingly, orion expression was detected in a small number of neurons in the larval ventral nerve cord. It will be interesting to find out whether these neurons are particularly subject to degeneration. Second, compared to the larval PNS where degenerative events are rare, the nervous system undergoing metamorphosis has a much greater demand for phagocytosis. Turning on orion expression in neurons may thus be required for efficient clearance of all pruned neurites. Consistent with this idea, orion expression was also in a subset of da neurons during metamorphosis (Ji, 2023).
Fluorescent PS probes based on AV and Lact are widely used to visualize PS exposure in cell culture and live animals and have been crucial for many discoveries in PS biology. LactC1C2 is known to have a higher affinity to PS than AV. Previously additional differences were observed between these two proteins. Compared to LactC1C2, which coats the dendrite surface well, AV tends to traffic to endocytic vesicles inside PS-exposing dendrites, consistent with the ability of the AV complex to induce endocytosis upon PS binding. Importantly, unlike AV, which does not alter the kinetics of neurite degeneration, LactC1C2 binding to PS-exposing dendrites potentiates Drpr-dependent degeneration. This latter observation previously led us to hypothesize that LactC1C2 may contain unknown sequences that interact with Drpr. This study showed that the effect of LactC1C2 in exacerbating dendrite degeneration depends on orion, suggesting instead that LactC1C2 may indirectly promote phagocyte/dendrite interactions by enhancing orion function. One possible mechanism is that LactC1C2 binding on the plasma membrane further disrupts the membrane and causes more PS exposure that can subsequently be detected by endogenous orion (Ji, 2023).
Compared to AV and LactC1C2, orion displays unique lipid-binding properties. The in vivo evidence suggests that orion may have a higher affinity for PS than AV, as orion efficiently outcompetes AV in binding to injured dendrites. In addition, overexpressed orion binds to healthy epidermal cells and dendrites (albeit the latter only in wandering 3rd instar larvae), while AV and LactC1C2 do not. Althoughr in vitro evidence suggests that orion may have an intrinsic affinity for phospholipid bilayers, in vivo orion binding to epidermal cells and WT dendrites largely depends on PS exposure. One possibility is that other endogenous factors modify orion-lipid interactions and make orion more specific to PS in vivo (Ji, 2023).
The surprising finding that overexpressed orion binds to peripheral tissues and dendrites suggests that these cells may expose PS under physiological conditions, perhaps at a level too low to detect by AV and LactC1C2. Thus, low levels of PS exposure may be much more prevalent in vivo than previously thought. Intriguingly, binding of overexpressed orion induces degeneration of wild-type dendrites but does not cause obvious phagocytosis of other nonneural tissues, suggesting that neurons may be more vulnerable to PS-induced phagocytosis than other cell types (Ji, 2023).
Recently, many human chemokines were found to bind to PS exposed on apoptotic vesicles and serve as 'find-me' signals to attract phagocytes. orion shares the CX3C motif with the mammalian chemokine CXC3L1 and has also three glycosaminoglycan putative binding sequences, a hallmark of chemokine activity. Even though direct binding to PS has not been demonstrated for CXC3L1, this chemokine is required for microglia-mediated synapse elimination after whisker lesioning, a process likely involving PS exposure. In addition, orion contains a RRY motif commonly found in human neutrophil peptides, small antimicrobial peptides important for innate immunity. Both CX3C and RRY motifs are important for orion function in mediating phagocytosis of neurons. Although orion does not show global sequence homologies to mammalian immunomodulatory proteins, its interaction partner Drpr has mammalian homologs that are involved in the phagocytosis of neurons through unknown mechanisms . Thus, the common features between orion and human proteins indicate that a functional conservation may exist between PS-sensing mechanisms in insects and humans (Ji, 2023).
Suboptimal nutrition imposes developmental constraints on infant animals, which marshal adaptive responses to eventually become mature adults. Such responses are mounted at multiple levels from systemic to cellular. Little is known about how growth of postmitotic and morphologically complex cells, such as neurons, is controlled by nutritional status. This question was addressed using Class I and Class IV dendritic arborization neurons in Drosophila larvae. Class IV neurons have been shown to sense nociceptive thermal, mechanical and light stimuli, whereas Class I neurons are proprioceptors. Larvae were reared on diets with different protein and carbohydrate content throughout larval stages, and how morphologies of Class I or Class IV neurons were affected was examined. Dendritic arbors of Class IV neurons became more complex when larvae were reared on a low-yeast diet, which contains lower amounts of amino acids and other ingredients, compared to a high-yeast diet. In contrast, such low-yeast-dependent hyperarborization was not seen in Class I neurons. The physiological and metabolic implications of the hyperarborization phenotype are discussed in relation to a recent hypothesis that Class IV neurons sense protein-deficient stress and to this characterization of how the dietary yeast contents impacted larval metabolism (Watanabe, 2016).
Neurons develop their dendrites in tight relation to their connection and computation requirements. Thus, dendrite morphologies display sophisticated type-specific patterns. From the cell biological and developmental perspective, this raises the question of at which level different neuronal types might use shared mechanisms to assemble their dendrites. And, conversely, how are specialized structures achieved in different neuronal types? To start addressing these question computational and comparative cell biological approaches were combined. It was found that two distinct growth programs are required to achieve models that faithfully reproduce the dendrite organization of c3da neurons. The models single out the STBs that are also molecularly identifiable as unique structures, displaying specific localization of actin and Singed. By combining time-lapse in vivo imaging and genetic analyses, this study sheds light on the machinery that controls the dynamic formation of those branchlets (Sturner, 2022).
The complex interplay of AMPs generates highly adaptive actin networks. In fact, in contrast to earlier unifying models, it is now clear that even the same cell can make more than one type of filopodium-like structure. This study characterized the effect of the loss of six AMPs on the morphology and dynamics of one specific type of dendritic branchlet, the STB of c3da neurons. With this information, a molecular model for branchlet dynamics in vivo is delineated in the developing animal. Similar approaches to model the molecular regulation of actin in dendrite filopodia have been taken recently for cultured neurons. The advantage of the present approach is that it relies directly on the effect of the loss of individual AMPs in vivo, preserving the morphology, dynamics, and adhesive properties of the branchlets, and non-cell-autonomous signals remain present (Sturner, 2022).
The combination of FRAP experiments and the localization of Singed/Fascin on the extending STBs indicated that actin is organized in a tight bundle of mostly uniparallel fibers in the STBs. This organization is thus very different from that of dendritic filopodia of hippocampal neurons in culture. The actin filaments in the bundle appear to be particularly stable in the c3da-neuron STBs, as the actin turnover that this study revealed by FRAP analysis was 4 times slower than that reported in dendrite spines of hippocampal neurons in vitro and 20-fold slower than in a lamellipodium of melanoma cells in vitro. It is nonetheless in line with previous data on stable c3da-neuron STBs and with bundled actin filaments of stress fibers of human osteosarcoma cells. Treadmilling was observed, similar to that of filopodia at the leading edge, with a retrograde flow rate 30 times slower than in filopodia of hippocampal cells and comparable to rates observed for developing neurons in culture lacking the mammalian homologues of Twinstar and actin-depolymerization factor (ADF)/Cofilin. Slower actin kinetics could be related to the fact that neurons differentiating in the complex 3D context of a developing animal are being imaged. Recent quantification of actin treadmilling in a growth cone of hippocampal neurons in 3D culture, however, did not produce differences with 2D-culture models(Sturner, 2022).
The alterations of MB and STB morphology and dynamics caused by the loss of individual AMP functions reported in this study can now be combined with preceding molecular knowledge about these conserved factors to produce a hypothetical model of the actin regulation underlying STB dynamics. Dendrite structure and time-lapse imaging point to an essential role of Twinstar/Cofilin for the initiation of a branchlet, in agreement with previous literature. Drosophila Twinstar/Cofilin is a member of the ADF/Cofilin protein family, with the capacity of severing actin filaments but with poor actin-filament-depolymerizing activity. It is thus proposed that Twinstar/Cofilin localized at the base of c3da STBs can induce a local fragmentation of actin filaments that can then be used as substrate by the Arp2/3 complex. In fact, in c4da neurons, Arp2/3 localizes transiently at the site where the branchlets will be formed, and its presence strongly correlates with the initiation of branchlet formation. Previous and present time-lapse data point to the role of Arp2/3 in the early phases of branchlet formation. Thus, it is suggested that localized activity of Arp2/3 generates a first localized membrane protrusion (Sturner, 2022).
Given the transitory localization of Arp2/3, this study interrogated the role of additional actin nucleators in this context. From an RNAi-supported investigation, Capu was identified as potential modifier of c3da STBs. Capu displays complex interactions with the actin-nucleator Spire during oogenesis, involving cooperative and independent functions of these two molecules. An increase in Spire levels correlates with a smaller dendritic tree and inappropriate, F-actin-rich, and shorter dendrites in c4da neurons. In this study, though, the loss of Spire function did not yield a detectable phenotype in c4da neurons. In c3da neurons, it was found that Capu and Spire support the formation of new branchlets and display a strong genetic interaction in the control of the number and length of MBs and STBs and surface area. Thus it is suggested that they cooperatively take over the nucleation of linear actin filaments possibly producing the bundle of uniparallel actin filaments. Mutants for capu showed changes in the positioning of dendritic branches, not observed in spire mutants, which could mean that Capu localization defines the sites of Capu/Spire activity. However, Spire seems to promote branch dynamics, suggesting additional independent functions of Spire possibly not related to nucleation, given that Spire itself is a weak actin nucleator. While there is no clear indication in vivo for the molecular mechanisms supporting this function, an actin-severing activity of Spire was reported in vitro. The role of Spire on STB dynamics appears to be consistent with favoring actin destabilization or actin dynamics (Sturner, 2022).
Singed/Fascin bundles actin filaments specifically in the c3da neuron STBs and gives these branches their straight conformation. The localization of Singed/Fascin in the c3da STBs correlates with their elongation. While the complete loss of singed function suppressed dynamics, the mild reduction in protein levels analyzed in this study led to more frequent STB elongations and retractions. Further, the branchlets extended at the wrong angles and displayed a tortuous path. Singed/Fascin controls the interaction of actin-filament bundles with Twinstar/Cofilin and can enhance Ena binding to barbed ends. Thus, in addition to generating mechanically rigid bundles, it can modulate actin dynamics by regulating the interaction of multiple AMPs with actin. It is speculated that the retraction and disappearance of the STB could be due to Singed/Fascin dissociating from the actin filaments, possibly in combination with Spire and Twinstar/Cofilin additionally severing actin filaments. In fact, the presence of detectable Twinstar/Cofilin along the c3da STBs was recently reported (Sturner, 2022).
Ena is important for restricting STB length, and it inhibits the new formation and extension of STBs. This appears to be a surprising function for Ena that is in contrast to its role in promoting actin-filament elongation or to its capacity of supporting the activation of the WAVE regulatory complex. Similar to what was previously reported for ena-mutant c4da neurons, a balance between elongation and branching was also observed in c3da neurons. In Drosophila macrophages, Ena was shown to associate with Singed/Fascin within lamellipodia. In line with these recent data, it is suggested that Ena might closely cooperate with Singed to form tight actin bundles that slow down STB elongation (Sturner, 2022).
Taken together, a comprehensive molecular model of dendrite-branch dynamics for the STBs of c3da neurons was put forward. In this analysis, the role of extracellular signals on the regulation of the dynamics of STBs was excluded, for simplicity. Nonetheless, such signals are likely to have a profound effect, particularly on the regulation of elongation and stabilization of STBs in relation to their target substrate. In addition, similar to what has been suggested for c1da neurons, the distribution of MBs in the target area might follow guidance cues that were not included in the analysis, such as permissive signals that specifically guide c4da neurons to tile the body wall or promote appropriate space filling (Sturner, 2022).
The investigation of morphological parameters in combination with genetic analysis has proven extremely powerful to reveal initial molecular mechanisms of dendrite differentiation. Early studies, though, have been limited in the description power of their analysis concentrating on just one or two parameters (e.g., number of termini and total dendrite length). This limitation has been recognized and addressed in more recent studies (Sturner, 2022).
A major outcome of the present and previous work is the establishment of powerful tools for a thorough and comparative quantitative morphological analysis of different mutant groups. A detailed tracing of neuronal dendrites of the entire dendritic tree or a certain area of the tree in a time series with a subsequent automatic analysis allows a precise description of mutant phenotypes. This study additionally generated tools for extracting quantitative parameters of the dynamic behavior of dendrite branches from time-lapse movies based on a novel branch registration software. This time-lapse tool yields an automated quantification after registration detecting branch types and their dynamics. Moreover, the tool operates in the same framework as the tracing and morphological analysis. These tools available within the TREES toolbox, and their use to support comparative analysis among datasets is encouraged (Sturner, 2022).
What are the fundamental principles that define dendrite elaboration and which constraints need to be respected by neurons in establishing their complex arbors? Models based on local or global rules have been applied to reproduce the overall organization of dendritic trees, including da neurons. The c3da model is based on the fundamental organizing principle that dendrites are built through minimizing cable length and signal conduction times. This general rule for optimal wiring predicts tight scaling relationships between fundamental branching statistics, such as the number of branches, the total length, and the dendrite's spanning field (Sturner, 2022).
This study found that c3da neurons respect the general developmental SFGT or MST models when stripped of all their STBs. However, the characteristic STBs of c3da dendrites did not follow this scaling behavior. Instead, a second growth program had to be applied to add the STBs to this basic structure, respecting their number, total length, and distribution. The two-step model developed in this work suggests that while main dendritic trees have common growth rules, the dendritic specializations of different neuronal cell types do not necessarily have the same constraints. This view is compatible with findings in a companion paper showing, in c1da neurons, a specialized branch-retraction step following an initial growth step. In the two-step c3da dendrite model, the resulting synthetic morphologies resemble the real dendritic trees including those of five out of the six AMP mutant dendritic trees without any changes to the model parameters. The two-step model uses, for example, the reduced total length and reduced surface area of mutants for singed and twinstar and grows synthetic trees that have the same distribution of branch lengths and amounts as expected for those mutants. The synthetic trees corresponding to the twinstar mutant have less STBs than any other AMP mutant synthetic tree, consistent with the real mutant phenotypes (Sturner, 2022).
This work indicates that a combination of thorough statistical analysis (such as using the presented morphometrics) and models, like the one developed in this study, can help capture the fundamental principles that govern dendrite differentiation. Together with genetics analysis and systematic cell biology approaches, this type of study can deliver quantitative predictions for molecular models of dendrite elaboration (Sturner, 2022).
In conclusion, this study has put forward the hypothesis that neuronal dendrites are built based on common, shared growth programs. An additional refinement step is then added to this scaffold, allowing each neuron type to specialize based on its distinctive needs in terms of number and distribution of inputs. In the exemplary case of c3da neurons, this study investigated molecular properties of these more-specialized growth programs and proposed a first comprehensive model of actin regulation that explains the morphology and dynamics of branchlets (Sturner, 2022).
Most of the AMPs studied are essential, and all perform multiple functions during the course of development. Clearly, in these experiments, the acute function of each AMP in the process of STB formation and during STB dynamics has not been isolated. Rather, the progressive reduction of functional protein in MARCM clones or during the development of homozygous animals might represent a confounding factor. Future studies will be aimed at using and developing tools for acute protein-function inactivation in vivo to add to the toolbox (Sturner, 2022).
Neurons develop their dendrites in tight relation to their connection and computation requirements. Thus, dendrite morphologies display sophisticated type-specific patterns. From the cell biological and developmental perspective, this raises the question of at which level different neuronal types might use shared mechanisms to assemble their dendrites. And, conversely, how are specialized structures achieved in different neuronal types? To start addressing these question computational and comparative cell biological approaches were combined. It was found that two distinct growth programs are required to achieve models that faithfully reproduce the dendrite organization of c3da neurons. The models single out the STBs that are also molecularly identifiable as unique structures, displaying specific localization of actin and Singed. By combining time-lapse in vivo imaging and genetic analyses, this study sheds light on the machinery that controls the dynamic formation of those branchlets (Sturner, 2022).
The complex interplay of AMPs generates highly adaptive actin networks. In fact, in contrast to earlier unifying models, it is now clear that even the same cell can make more than one type of filopodium-like structure. This study characterized the effect of the loss of six AMPs on the morphology and dynamics of one specific type of dendritic branchlet, the STB of c3da neurons. With this information, a molecular model for branchlet dynamics in vivo is delineated in the developing animal. Similar approaches to model the molecular regulation of actin in dendrite filopodia have been taken recently for cultured neurons. The advantage of the present approach is that it relies directly on the effect of the loss of individual AMPs in vivo, preserving the morphology, dynamics, and adhesive properties of the branchlets, and non-cell-autonomous signals remain present (Sturner, 2022).
The combination of FRAP experiments and the localization of Singed/Fascin on the extending STBs indicated that actin is organized in a tight bundle of mostly uniparallel fibers in the STBs. This organization is thus very different from that of dendritic filopodia of hippocampal neurons in culture. The actin filaments in the bundle appear to be particularly stable in the c3da-neuron STBs, as the actin turnover that this study revealed by FRAP analysis was 4 times slower than that reported in dendrite spines of hippocampal neurons in vitro and 20-fold slower than in a lamellipodium of melanoma cells in vitro. It is nonetheless in line with previous data on stable c3da-neuron STBs and with bundled actin filaments of stress fibers of human osteosarcoma cells. Treadmilling was observed, similar to that of filopodia at the leading edge, with a retrograde flow rate 30 times slower than in filopodia of hippocampal cells and comparable to rates observed for developing neurons in culture lacking the mammalian homologues of Twinstar and actin-depolymerization factor (ADF)/Cofilin. Slower actin kinetics could be related to the fact that neurons differentiating in the complex 3D context of a developing animal are being imaged. Recent quantification of actin treadmilling in a growth cone of hippocampal neurons in 3D culture, however, did not produce differences with 2D-culture models(Sturner, 2022).
The alterations of MB and STB morphology and dynamics caused by the loss of individual AMP functions reported in this study can now be combined with preceding molecular knowledge about these conserved factors to produce a hypothetical model of the actin regulation underlying STB dynamics. Dendrite structure and time-lapse imaging point to an essential role of Twinstar/Cofilin for the initiation of a branchlet, in agreement with previous literature. Drosophila Twinstar/Cofilin is a member of the ADF/Cofilin protein family, with the capacity of severing actin filaments but with poor actin-filament-depolymerizing activity. It is thus proposed that Twinstar/Cofilin localized at the base of c3da STBs can induce a local fragmentation of actin filaments that can then be used as substrate by the Arp2/3 complex. In fact, in c4da neurons, Arp2/3 localizes transiently at the site where the branchlets will be formed, and its presence strongly correlates with the initiation of branchlet formation. Previous and present time-lapse data point to the role of Arp2/3 in the early phases of branchlet formation. Thus, it is suggested that localized activity of Arp2/3 generates a first localized membrane protrusion (Sturner, 2022).
Given the transitory localization of Arp2/3, this study interrogated the role of additional actin nucleators in this context. From an RNAi-supported investigation, Capu was identified as potential modifier of c3da STBs. Capu displays complex interactions with the actin-nucleator Spire during oogenesis, involving cooperative and independent functions of these two molecules. An increase in Spire levels correlates with a smaller dendritic tree and inappropriate, F-actin-rich, and shorter dendrites in c4da neurons. In this study, though, the loss of Spire function did not yield a detectable phenotype in c4da neurons. In c3da neurons, it was found that Capu and Spire support the formation of new branchlets and display a strong genetic interaction in the control of the number and length of MBs and STBs and surface area. Thus it is suggested that they cooperatively take over the nucleation of linear actin filaments possibly producing the bundle of uniparallel actin filaments. Mutants for capu showed changes in the positioning of dendritic branches, not observed in spire mutants, which could mean that Capu localization defines the sites of Capu/Spire activity. However, Spire seems to promote branch dynamics, suggesting additional independent functions of Spire possibly not related to nucleation, given that Spire itself is a weak actin nucleator. While there is no clear indication in vivo for the molecular mechanisms supporting this function, an actin-severing activity of Spire was reported in vitro. The role of Spire on STB dynamics appears to be consistent with favoring actin destabilization or actin dynamics (Sturner, 2022).
Singed/Fascin bundles actin filaments specifically in the c3da neuron STBs and gives these branches their straight conformation. The localization of Singed/Fascin in the c3da STBs correlates with their elongation. While the complete loss of singed function suppressed dynamics, the mild reduction in protein levels analyzed in this study led to more frequent STB elongations and retractions. Further, the branchlets extended at the wrong angles and displayed a tortuous path. Singed/Fascin controls the interaction of actin-filament bundles with Twinstar/Cofilin and can enhance Ena binding to barbed ends. Thus, in addition to generating mechanically rigid bundles, it can modulate actin dynamics by regulating the interaction of multiple AMPs with actin. It is speculated that the retraction and disappearance of the STB could be due to Singed/Fascin dissociating from the actin filaments, possibly in combination with Spire and Twinstar/Cofilin additionally severing actin filaments. In fact, the presence of detectable Twinstar/Cofilin along the c3da STBs was recently reported (Sturner, 2022).
Ena is important for restricting STB length, and it inhibits the new formation and extension of STBs. This appears to be a surprising function for Ena that is in contrast to its role in promoting actin-filament elongation or to its capacity of supporting the activation of the WAVE regulatory complex. Similar to what was previously reported for ena-mutant c4da neurons, a balance between elongation and branching was also observed in c3da neurons. In Drosophila macrophages, Ena was shown to associate with Singed/Fascin within lamellipodia. In line with these recent data, it is suggested that Ena might closely cooperate with Singed to form tight actin bundles that slow down STB elongation (Sturner, 2022).
Taken together, a comprehensive molecular model of dendrite-branch dynamics for the STBs of c3da neurons was put forward. In this analysis, the role of extracellular signals on the regulation of the dynamics of STBs was excluded, for simplicity. Nonetheless, such signals are likely to have a profound effect, particularly on the regulation of elongation and stabilization of STBs in relation to their target substrate. In addition, similar to what has been suggested for c1da neurons, the distribution of MBs in the target area might follow guidance cues that were not included in the analysis, such as permissive signals that specifically guide c4da neurons to tile the body wall or promote appropriate space filling (Sturner, 2022).
The investigation of morphological parameters in combination with genetic analysis has proven extremely powerful to reveal initial molecular mechanisms of dendrite differentiation. Early studies, though, have been limited in the description power of their analysis concentrating on just one or two parameters (e.g., number of termini and total dendrite length). This limitation has been recognized and addressed in more recent studies (Sturner, 2022).
A major outcome of the present and previous work is the establishment of powerful tools for a thorough and comparative quantitative morphological analysis of different mutant groups. A detailed tracing of neuronal dendrites of the entire dendritic tree or a certain area of the tree in a time series with a subsequent automatic analysis allows a precise description of mutant phenotypes. This study additionally generated tools for extracting quantitative parameters of the dynamic behavior of dendrite branches from time-lapse movies based on a novel branch registration software. This time-lapse tool yields an automated quantification after registration detecting branch types and their dynamics. Moreover, the tool operates in the same framework as the tracing and morphological analysis. These tools available within the TREES toolbox, and their use to support comparative analysis among datasets is encouraged (Sturner, 2022).
What are the fundamental principles that define dendrite elaboration and which constraints need to be respected by neurons in establishing their complex arbors? Models based on local or global rules have been applied to reproduce the overall organization of dendritic trees, including da neurons. The c3da model is based on the fundamental organizing principle that dendrites are built through minimizing cable length and signal conduction times. This general rule for optimal wiring predicts tight scaling relationships between fundamental branching statistics, such as the number of branches, the total length, and the dendrite's spanning field (Sturner, 2022).
This study found that c3da neurons respect the general developmental SFGT or MST models when stripped of all their STBs. However, the characteristic STBs of c3da dendrites did not follow this scaling behavior. Instead, a second growth program had to be applied to add the STBs to this basic structure, respecting their number, total length, and distribution. The two-step model developed in this work suggests that while main dendritic trees have common growth rules, the dendritic specializations of different neuronal cell types do not necessarily have the same constraints. This view is compatible with findings in a companion paper showing, in c1da neurons, a specialized branch-retraction step following an initial growth step. In the two-step c3da dendrite model, the resulting synthetic morphologies resemble the real dendritic trees including those of five out of the six AMP mutant dendritic trees without any changes to the model parameters. The two-step model uses, for example, the reduced total length and reduced surface area of mutants for singed and twinstar and grows synthetic trees that have the same distribution of branch lengths and amounts as expected for those mutants. The synthetic trees corresponding to the twinstar mutant have less STBs than any other AMP mutant synthetic tree, consistent with the real mutant phenotypes (Sturner, 2022).
This work indicates that a combination of thorough statistical analysis (such as using the presented morphometrics) and models, like the one developed in this study, can help capture the fundamental principles that govern dendrite differentiation. Together with genetics analysis and systematic cell biology approaches, this type of study can deliver quantitative predictions for molecular models of dendrite elaboration (Sturner, 2022).
In conclusion, this study has put forward the hypothesis that neuronal dendrites are built based on common, shared growth programs. An additional refinement step is then added to this scaffold, allowing each neuron type to specialize based on its distinctive needs in terms of number and distribution of inputs. In the exemplary case of c3da neurons, this study investigated molecular properties of these more-specialized growth programs and proposed a first comprehensive model of actin regulation that explains the morphology and dynamics of branchlets (Sturner, 2022).
Most of the AMPs studied are essential, and all perform multiple functions during the course of development. Clearly, in these experiments, the acute function of each AMP in the process of STB formation and during STB dynamics has not been isolated. Rather, the progressive reduction of functional protein in MARCM clones or during the development of homozygous animals might represent a confounding factor. Future studies will be aimed at using and developing tools for acute protein-function inactivation in vivo to add to the toolbox (Sturner, 2022).
Dendritic morphology underlies the source and processing of neuronal signal inputs. Morphology can be broadly described by two types of geometric characteristics. The first is dendrogram topology, defined by the length and frequency of the arbor branches; the second is spatial embedding, mainly determined by branch angles and straightness. It has been previously demonstrated that microtubules and actin filaments are associated with arbor elongation and branching, fully constraining dendrogram topology. This study relates the local distribution of these two primary cytoskeletal components with dendritic spatial embedding. 167 sensory neurons from the Drosophila larva encompassing multiple cell classes and genotypes were reconstructed and analyzed. It was observe that branches with a higher microtubule concentration tend to deviate less from the direction of their parent branch across all neuron types. Higher microtubule branches are also overall straighter. F-actin displays a similar effect on angular deviation and branch straightness, but not as consistently across all neuron types as microtubule. These observations raise the question as to whether the associations between cytoskeletal distributions and arbor geometry are sufficient constraints to reproduce type-specific dendritic architecture. Therefore, this study create a computational model of dendritic morphology purely constrained by the cytoskeletal composition measured from real neurons. The model quantitatively captures both spatial embedding and dendrogram topology across all tested neuron groups. These results suggest a common developmental mechanism regulating diverse morphologies, where the local cytoskeletal distribution can fully specify the overall emergent geometry of dendritic arbors (Nanda, 2023).
The development of cell-type-specific dendritic arbors is integral to the proper functioning of neurons within their circuit networks. This study examined the regulatory relationship between the cytosolic chaperonin CCT, key insulin pathway genes, and an E3 ubiquitin ligase (Cullin1) in homeostatic dendritic development. CCT loss of function (LOF) results in dendritic hypotrophy in Drosophila Class IV (CIV) multidendritic larval sensory neurons, and CCT has recently been shown to fold components of the TOR (Target of Rapamycin) complex 1 (TORC1), in vitro. Through targeted genetic manipulations, this study has confirmed that LOF of CCT and the TORC1 pathway reduces dendritic complexity, while overexpression of key TORC1 pathway genes increases dendritic complexity in CIV neurons. Both CCT and TORC1 LOF significantly reduce microtubule (MT) stability. CCT has been previously implicated in regulating proteinopathic aggregation, thus CIV dendritic development was examined in disease conditions as well. Expression of mutant Huntingtin leads to dendritic hypotrophy in a repeat-length-dependent manner, which can be rescued by TORC1 disinhibition via Cullin1 LOF. Together, these data suggest that Cullin1 and CCT influence dendritic arborization through regulation of TORC1 in both health and disease (Lottes, 2023).
A major gap in understanding of sensation is how a single sensory neuron can differentially respond to a multitude of different stimuli (polymodality), such as propio- or nocisensation. The prevailing hypothesis is that different stimuli are transduced through ion channels with diverse properties and subunit composition. In a screen for ion channel genes expressed in polymodal nociceptive neurons, this study identified Ppk26, a member of the trimeric degenerin/epithelial sodium channel (DEG/ENaC) family, as being necessary for proper locomotion behavior in Drosophila larvae in a mutually dependent fashion with coexpressed Ppk1 (Pickpocket), another member of the same family. Mutants lacking Ppk1 and Ppk26 were defective in mechanical, but not thermal, nociception behavior. Mutants of Piezo, a channel involved in mechanical nociception in the same neurons, did not show a defect in locomotion, suggesting distinct molecular machinery for mediating locomotor feedback and mechanical nociception (Gorczyca, 2014).
This study has identified Ppk26, an DEG/ENaC channel subunit that is coexpressed with Ppk1 in class IV da neurons. Consistent with the model that Ppk26 and Ppk1 may be subunits of the same channel, it was found that Ppk1 and Ppk26 colocalize in class IV da neurons, they form a complex in heterologous expression systems, and they show nonadditive and nonredundant mutant phenotypes in vivo. Ppk26 protein was found in somatic, dendritic, and axonal compartments, and plasma membrane insertion was observed in terminal dendrites. Ppk1 and Ppk26 were reciprocally required for normal trafficking and/or insertion to the plasma membrane, further supporting the notion that these two channel subunits interact in vivo. This study shows that, as is the case for MEC Degenerin channels, mutations at the Degenerin position of Ppk26 lead to loss of class IV da neuron integrity. It was also found that Ppk26 function plays essential roles in normal larval locomotion, particularly in turning behavior (Gorczyca, 2014).
Overexpression of EGFP-tagged MEC channels in C. elegans has been reported to result in a punctate localization, leading to suggestions that each of these puncta represents a mechanosensory apparatus. In light of the speculation that contact of dendrites with subcuticular epidermis is part of the apparatus that senses mechanical stimuli, it is intriguing that both Ppk1 and Ppk26 were found on the surface of distal and higher-order dendrites, consistent with channel function in this compartment. Mutants in Ppk1 and Ppk26 showed defects in the frequency of turning of freely crawling larvae. Moreover, loss of function of either or both Ppk26 and Ppk1 had the same effect on larval locomotion. These findings support the notion that Ppk26 and Ppk1 may act in the same pathway—perhaps in the same channel complex—in mechanosensation that is important for proper locomotion (Gorczyca, 2014).
Class IV da neurons are known to express the DEG/ENaC channel subunit Ppk1 (Adams, 1998), an observation that was confirmed in this study. Given that DEG/ENaC channels are trimeric ion channels that typically require the assembly of different subunits for proper function, the specific coexpression of Ppk26 in class IV da neurons raises the possibility that Ppk1 and Ppk26 may correspond to different subunits of the same mechanosensory channel in vivo. This is supported by the findings of biochemical interactions between Ppk1 and Ppk26, their null mutant phenotypes that indicate nonredundancy, and by the reciprocal requirements of Ppk1 and Ppk26 for normal trafficking or insertion to the plasma membrane . The mechanism for this mutual dependence is currently unknown (Gorczyca, 2014).
Notably, inspection of the developmental expression profile of ppk1 RNA and ppk26 RNA revealed a similar time course, with the absolute levels of RNA for ppk26 twice as high as those of RNA for ppk1 throughout development. Although it is uncertain that this quantitative difference in RNA levels reflects a similar quantitative difference in the levels of Ppk1 and Ppk26 subunits, it is tempting to speculate that two Ppk26 subunits and one Ppk1 subunit may be assembled to form a surface-expressed trimeric channel (Gorczyca, 2014).
Studies in many systems have suggested that mutations in the Degenerin domain of DEG/ENaC lead to loss of cell integrity and perhaps degeneration; however, this behavior has not yet been observed in Drosophila Pickpocket family members. The Degenerin position is localized in the wrist region, close to the mouth of the pore. Thus, it has been suggested that Degenerin mutations change the properties of the channel, increasing open time probability and perhaps shifting its ion selectivity from Na+ to Ca2+. Given the tight regulation of Ca2+ within cells and its involvement in critical cellular processes, this increase in Ca2+ permeability may lead to loss of cellular homeostasis, in a process that has been dubbed excitotoxicity. Consistent with these findings, it was also observed that when a Degenerin mutation was introduced into Ppk26 that was overexpressed in class IV da neurons, it resulted in a marked reduction in dendritic arbor size. This suggests that the Ppk26-Deg mutation leads to toxicity in the sensory neurons, as has been observed in DEG/ENaC channels in C. elegans, but not in Ppk1. Whereas it is likely that the function of the pore structure of these channels is evolutionarily conserved, it is unclear why Ppk26-Deg has a more potent effect than Ppk1-Deg. One possibility is that the Ppk26-Deg residue makes a larger contribution to pore structure than Ppk1-Deg, because of its intrinsic structure or because of a potential 2:1 stoichiometry in vivo (Gorczyca, 2014).
Larval locomotion is likely regulated by sensory input provided by sensory neurons in the body wall, which may in turn modulate the motoneurons innervating the body wall. The C. elegans mechanosensitive TRPN channel TRP-4 acts in the DVA neuron to coordinate bending behavior and body posture through positive and negative modulation. It seems likely that class IV da neurons likewise provide some information for sensory modulation of locomotion through a mechanosensory mechanism, but future work will need to determine if this is indeed the case (Gorczyca, 2014).
Proprioception is a mechanosensory process involving sensory neurons that transduce the mechanical information related to body position or characteristics of the environment for the generation of appropriate behavioral output, such as the turning locomotor behavior that is essential for foraging larvae. This study has identified a member of the DEG/ENaC family of proteins, Ppk26, which acts together with Ppk1 likely as subunits of a channel important for mechanosensation. The results suggest that perhaps a major site of mechanosensory transduction is located in class IV da neuron dendritic processes. The behavioral phenotypes of larvae with Ppk1 and Ppk26 knockdown in class IV da neurons, as well as the respective null mutants or double mutants, suggest that a deficit in these channels interferes with the ability of the animal to execute proper turning behavior, raising the possibility that the two subunits could be involved in proprioceptively sensing the deformation of the cuticle. Whether class IV neurons function as proprioceptors still needs to be directly demonstrated, and future experiments will be needed to address the relationship between class IV neural activity and body position (Gorczyca, 2014).
Numerous studies have implicated the class IV da neurons in both thermal and mechanical nociception behavior. This study found that whereas Ppk1 and Ppk26 are important for mechanical nociception behavior, they are dispensable for thermal nociception behavior. While Ppk1 and Ppk26 channels indeed moonlight during two processes in the same neuron, namely, mechanical nociception and proprioception, these channels must be playing a specific role as they are not involved in thermal response by the same neuron (Gorczyca, 2014).
Whereas the class I da neurons and bd neurons are implicated in proprioception for the regulation of sequential contractions use the TRPN channel NompC as the sensor, class IV da neurons rely on the DEG/ENaC channel likely composed of Ppk26 and Ppk1 for the regulation of turning behavior as well as mechanical nociception, perhaps through sensing a mechanical signal at the cuticle. Interestingly, this study found that Piezo, a bona fide mechanotransducing ion channel involved in class IV da neuron mechanotransduction and required for mechanical nociception, does not appear to be involved in turning behavior, suggesting that different combinations of ion channels may serve different mechanosensory functions in the same neuron (Gorczyca, 2014).
The fruit fly Drosophila melanogaster has provided important insights into how sensory information is transduced by transient receptor potential (TRP) channels in the peripheral nervous system (PNS). However, TRP channels alone have not been able to completely model mechanosensitive transduction in mechanoreceptive chordotonal neurons (CNs). This study shows that, in addition to TRP channels, the sole voltage-gated sodium channel (Na(V)) in Drosophila, Para, is localized to the dendrites of CNs. Para is localized to the distal tip of the dendrites in all CNs, from embryos to adults, and is colocalized with the mechanosensitive TRP channels No mechanoreceptor potential C (NompC) and Inactive/Nanchung (Iav/Nan). Para localization also demarcates spike initiation zones (SIZs) in axons and the dendritic localization of Para is indicative of a likely dendritic SIZ in fly CNs. Para is not present in the dendrites of other peripheral sensory neurons. In both multipolar and bipolar neurons in the PNS, Para is present in a proximal region of the axon, comparable to the axonal initial segment (AIS) in vertebrates, 40-60 μm from the soma in multipolar neurons and 20-40 μm in bipolar neurons. Whole-cell reduction of para expression using RNAi in CNs of the adult Johnston's organ (JO) severely affects sound-evoked potentials (SEPs). However, the duality of Para localization in the CN dendrites and axons identifies a need to develop resources to study compartment-specific roles of proteins that will enable us to better understand Para's role in mechanosensitive transduction (Ravenscroft, 2023).
Nutrition in early life has profound effects on an organism, altering processes such as organogenesis. However, little is known about how specific nutrients affect neuronal development. Dendrites of class IV dendritic arborization neurons in Drosophila larvae become more complex when the larvae are reared on a low-yeast diet compared to a high-yeast diet. A systematic search for key nutrients revealed that the neurons increase their dendritic terminal densities in response to a combined deficiency in vitamins, metal ions, and cholesterol. The deficiency of these nutrients upregulates Wingless in a closely located tissue, body wall muscle. Muscle-derived Wingless activates Akt in the neurons through the receptor tyrosine kinase Ror, which promotes the dendrite branching. In larval muscles, the expression of wingless is regulated not only in this key nutrient-dependent manner, but also by the JAK/STAT signaling pathway. Additionally, the low-yeast diet blunts neuronal light responsiveness and light avoidance behavior, which may help larvae optimize their survival strategies under low-nutritional conditions. Together, these studies illustrate how the availability of specific nutrients affects neuronal development through inter-organ signaling (Kanaoka, 2023).
Dendrites are highly complex 3D structures that define neuronal morphology and connectivity and are the predominant sites for synaptic input. Defects in dendritic structure are highly consistent correlates of brain diseases. However, the precise consequences of dendritic structure defects for neuronal function and behavioral performance remain unknown. This study probed dendritic function by using genetic tools to selectively abolish dendrites in identified Drosophila wing motoneurons without affecting other neuronal properties. These motoneuron dendrites were unexpectedly found to be dispensable for synaptic targeting, qualitatively normal neuronal activity patterns during behavior, and basic behavioral performance. However, significant performance deficits in sophisticated motor behaviors, such as flight altitude control and switching between discrete courtship song elements, scale with the degree of dendritic defect. These observations provide the first direct evidence that complex dendrite architecture is critically required for fine-tuning and adaptability within robust, evolutionarily constrained behavioral programs that are vital for mating success and survival. It is speculated that the observed scaling of performance deficits with the degree of structural defect is consistent with gradual increases in intellectual disability during continuously advancing structural deficiencies in progressive neurological disorders (Ryglewski, 2014).
The Drosophila sex pheromone cVA elicits different behaviors in males and females. First- and second-order olfactory neurons show identical pheromone responses, suggesting that sex genes differentially wire circuits deeper in the brain. Using in vivo whole-cell electrophysiology, this study has shown that two clusters of third-order olfactory neurons have dimorphic pheromone responses. One cluster responds in females; the other responds in males. These clusters are present in both sexes and share a common input pathway, but sex-specific wiring reroutes pheromone information. Regulating dendritic position, the Fruitless transcription factor both connects the male-responsive cluster and disconnects the female-responsive cluster from pheromone input. Selective masculinization of third-order neurons transforms their morphology and pheromone responses, demonstrating that circuits can be functionally rewired by the cell-autonomous action of a switch gene. This bidirectional switch, analogous to an electrical changeover switch, provides a simple circuit logic to activate different behaviors in males and females (Kohl, 2013).
This study reveals principles of neural circuit organization and
development that are of general significance. First, it was shown
that two populations of neurons, present in both sexes, show
reciprocal, sex-specific responses to the same stimulus. Second,
it was demonstrated that these responses result from differential
wiring of a common input to different outputs. Together,
these results define an elegant principle of neural circuit organization:
a developmental circuit switch directly analogous to
an electrical changeover (or single pole, double throw, SPDT)
switch that efficiently reroutes a common input signal to one of
two possible outputs. This model appears directly applicable
to sex-specific processing of mouse pheromones, including
ESP1 and Darcin (Haga, 2010; Stowers,
2010), but not to Caenorhabditis elegans ascarosides, where
recent data suggest wiring differences may not be required. The electrical
changeover switch is the prototype for a wide-range of electrical
switches in which concerted changes involving three or more
contacts reroute signals; it is very likely
that neural circuits, including those involved in pheromone processing,
contain more complex switches or assemblies of multiple
switches that elaborate on the basic mechanism that are described in this study. Indeed, over 700
sites of dimorphic neuronal overlap have been identified that may form such switches
in other sensory pathways, multimodal interneurons, or motor
circuits across the fly brain (Cachero, 2010).
Third, sex-specific placement of target neuron
dendrites were identified as the primary cellular basis of the switch that is described in this study. This contrasts with earlier studies of this circuit
that proposed that axonal dimorphism or neurons
present only in one sex were the key
dimorphic element. Regarding axonal dimorphism, Datta
(2008) hypothesized that a male-specific extension of DA1 PN
axon terminals is the basis of differential wiring in this system,
and Ruta (2010) subsequently proposed that this extension
synapses with the dendrites of aSP-f LHNs in males. The large
shifts in dendritic position that were observed in aSP-f and aSP-g
neurons mean the male-specific extension of DA1 PNs cannot
be sufficient for rewiring. Is it necessary? In mosaic masculinization
experiments, aSP-f and aSP-h neurons adopt male
morphology and pheromone responses in a brain in which other
neurons (including DA1 PNs) are female. Therefore, the male-specific
ventral extension is either not necessary for differential
wiring or is a secondary consequence of changes in the dendrites
of post-synaptic LHNs. Of course, this extension may
increase contact between DA1 PNs and aSP-f and aSP-h
LHNs, strengthening responses of those LHNs in males. All three
mechanisms (dendritic and axonal dimorphisms, dimorphic cell
numbers) are likely relevant to different degrees in different
circuits (Kohl, 2013).
Fourth, having defined this bi-directional switch, it was demonstrated
that its male form is specified by the fruitless gene. This transcription factor has a dual function, coordinating
the disconnection of one group of target neurons and
the connection of the other. Fifth, it was shown that masculinization
of third-order neurons alone is sufficient for functional rewiring.
Although previous studies have demonstrated a cell-autonomous
effect of fruitless on neuronal morphology, this study now demonstrates a difference
in functional connectivity. This is surprising because many would
predict that connectivity changes would depend on coordinate
regulation of genes in synaptic partner neurons. Such simplicity
has evolutionary implications: it may allow variation in circuit
structure and ultimately in behavior, through evolution of cis-regulatory
elements, as previously shown for somatic characters,
such as wing spots (Kohl, 2013).
Sixth, studies of pheromone processing in general and cVA
processing in particular have emphasized a labeled line processing
model. However, the current data indicate that both narrowly (aSP-f)
and broadly tuned (aSP-h) cVA-responsive neurons coexist in
males. Likewise in females, aSP-g neurons respond to cVA
and general odors, such as vinegar, but only cVA responses
depend on the Or67d receptor. It will be very interesting to determine
the circuit origin and behavioral significance of this integration
of odor channels. For example, it seems reasonable to
speculate that coincidence of cVA and food odors could interact
in a supralinear way to promote female courtship or egg laying.
This parallels the convergence in the lateral horn of a labeled
line responsive to non-cVA fly odors (Or47b/VA1lm neurons)
and one responsive to a specific food odorant, phenylacetic
acid, that acts as a male aphrodisiac (Kohl, 2013).
This study naturally raises additional questions. The action of
fruitless within fewer than 5% of the neurons in the fly brain
can specify behavior, and this study now shows that it can reroute pheromone signals
within those neurons. But what is the behavioral relevance of
this particular bidirectional switch? Testing this will require the
development of sensitive behavioral assays of cVA processing
and a reliable genetic approach to control this switch without
affecting the many other dimorphic elements in sensory and
motor circuits. Indeed, it remains to be seen whether flipping a
single switch in sensory processing is sufficient to engage motor
behavior typical of the opposite sex without masculinizing downstream
circuitry. It is noted that it is possible to force the production of courtship song by activating
fruitless-positive neurons in headless females, but this was almost
never successful in intact females (Kohl, 2013).
Another open question concerns the functional significance of
female aSP-f and male aSP-g neurons, which do not respond to
cVA or other tested odors. Do they receive input at all? One possibility,
based on in silico analysis of the brain-wide 3D maps is that they receive gustatory input,
perhaps from contact pheromones, although further work is
necessary to test this hypothesis. Finally, which genes does
fruitless regulate in order to differentially wire the switch? Clonal transformation experiments strongly support the earlier
proposal that male and female aSP-f/g/h
clusters are generated by neuroblasts common to both sexes
but that those neurons develop in a sex-specific manner. Therefore,
cell-surface molecules required for dendritic guidance are
plausible targets. It will be intriguing to see if the same fru-dependent
factor(s) direct(s) male aSP-f and female aSP-g dendrites to
the ventral lateral horn and, more generally, whether fruitless
acts on conserved downstream targets across all the dimorphic
neurons in the fly brain (Kohl, 2013).
Activity-dependent modifications strongly influence neural development. However, molecular programs underlying their context and circuit-specific effects are not well understood. To study global transcriptional changes associated with chronic elevation of synaptic activity, cell-type-specific transcriptome profiling was performed of Drosophila ventral lateral neurons (LNvs) in the developing visual circuit, and activity-modified transcripts were identified that are enriched in neuron morphogenesis, circadian regulation, and lipid metabolism and trafficking. Using bioinformatics and genetic analyses, activity-induced isoform-specific upregulation was validated of Drosophila lipophorin receptors LpR1 and LpR2, the homologs of mammalian low-density lipoprotein receptor (LDLR) family proteins. Furthermore, these morphological and physiological studies uncovered critical functions of neuronal lipophorin receptors (LpRs) in maintaining the structural and functional integrities in neurons challenged by chronic elevations of activity. Together, these findings identify LpRs as molecular targets for activity-dependent transcriptional regulation and reveal the functional significance of cell-type-specific regulation of neuronal lipid uptake in experience-dependent plasticity and adaptive responses (Yin, 2018).
This study investigated experience-dependent regulation of dendrite plasticity using cell-type-specific transcriptome profiling followed by transgenic RNAi screens. Candidate genes were identified that are subject to activity-dependent transcriptional regulation and function in regulating dendrite development in the Drosophila CNS. Additionally, combined bioinformatics and genetic analyses revealed isoform-specific expression of lipoprotein receptors LpR1 and LpR2 in LNvs and uncovered their roles in supporting dendrite morphogenesis and synaptic functions in the CNS. Together, these findings provide in vivo evidence for neuronal lipoprotein receptors serving as targets of activity-dependent transcriptional regulation, a previously unrecognized component of the neuronal homeostatic mechanism that maintains structural and functional integrity in response to chronic elevation of input activity (Yin, 2018).
Activity-dependent transcriptional factors serve important functions in synapse development, maturation, and elimination as well as dendritic and axonal outgrowth. However, a molecular understanding of circuit and context-specific transcriptional events induced by sensory experience is just starting to emerge. This cell-type-specific RNA-seq analyses identified over 200 experience-modified transcripts, among which are previously identified activity-dependent genes, including CrebB, Hr38, dnc, and Irk1, as well as many candidate genes that have not been previously linked to activity-dependent regulation or been functionally characterized in neurons. Therefore, this study generated a large number of potential targets for future molecular studies on experience-dependent dendrite plasticity. Besides the list of activity-modified transcripts, this analyses also revealed isoform-specific regulation of LpR genes, demonstrating the possibility of studying alternative splicing events associated with chronic alterations of activity using the RNA-seq dataset (Yin, 2018).
Both insect LpRs and mammalian LDLR family proteins have multiple isoforms generated by alternative splicing events and the usage of alternative promoters. In the fly imaginal disc and oocyte, long isoforms of LpRs acquire their lipoprotein cargo by interacting with lipid transfer particles (LTP), which stabilize the receptor-lipoprotein complex on the cell surface and possibly facilitate lipolysis. In these non-neuronal tissues, long isoforms of LpRs are also solely responsible for endocytosis-independent neutral lipid uptake. The function of the short isoforms, however, is unclear. When overexpressed in imaginal discs, the short isoform of LpR2 localizes in early endosomes and mediates uptake of lipoproteins through endocytic activities similar to those observed in mammalian LDLRs. These observations suggest that isoform-specific transcriptional regulation generates LpRs with diverse properties and functionalities (Yin, 2018).
These studies in LNvs indicate that the short-isoform of LpR2 has an endosomal localization in neurons and the ability to reconstitute LpR2 function in supporting dendrite growth. In conjunction with previous studies, these results suggest that isoform-specific transcriptional regulation may lead to distinct modes of lipoprotein intake in cells expressing different isoforms. It is proposed that the short isoforms of LpR1 and LpR2 expressed in neurons are endocytic lipoprotein receptors responsible for neuronal lipid uptake. In the CNS, lipid trafficking and homeostasis involve neural-glia interactions, which likely alter local concentrations of lipoprotein complexes and generate unique demands for lipoprotein uptake compared to non-neuronal tissues. Future in vivo imaging studies on CNS lipoprotein trafficking combined with functional studies of different LpR isoforms will contribute to understanding of the molecular regulation of neuronal lipid uptake (Yin, 2018).
Lipids are essential building blocks for plasma membranes and vesicles. They also have versatile roles in regulating cellular metabolism and mediating signaling transductions. In the nervous system, neuronal lipid uptake and recycling involve complex neuron-glia interactions that are critical for lipid homeostasis in the brain. Although neurons have the intrinsic ability to synthesize lipids, glia-derived cholesterol and phospholipids are essential for the formation and maintenance of the synapse in mammalian CNS neurons. Recent studies in the Drosophila system also demonstrated that glial lipid droplet formation protects neural stem cells from systematic stress in the developing Drosophila larval brain and that the inability of neurons to transport lipids for glial lipid droplet formation leads to accelerated neurodegeneration. However, whether and how neuronal lipid uptake contributes to normal development and activity-induced plasticity events remain unknown (Yin, 2018).
These studies demonstrate the function of LpRs in supporting dendrite growth. Reducing or eliminating LpRs in LNvs leads to a significant decrease in dendrite volume. Counterintuitively, LpRs are upregulated in LL conditions, which also generate reductions in LNv dendrite volume. Although an increased LpR level in the LL condition does not increase dendrite size, it is critical for preventing a further reduction in dendrites and a loss of physiological functions, suggesting that LpRs counteract activity-induced morphological and functional alterations. Based on these observations and current knowledge of lipid homeostasis in the larval CNS, it is proposed that activity-dependent regulation of LpRs serves as a homeostatic compensatory mechanism to augment the capacity for neuronal lipoprotein uptake in response to chronic elevation of input activity. Lipoprotein complexes recruited by LpRs are either released by glia or captured from the circulating hemolymph. This model is supported by RNA-seq studies, which revealed that a number of lipid-metabolism-related genes are downregulated in the constant light condition, including FASN1, Dgk, CG31140, GlcAT-S, fwd, and retm. LNv transcriptome analysis indicated a strong impact of elevated input activity on lipid metabolism, which may lead to altered lipid homeostasis and increased demand for lipid uptake. In vivo studies on activity-induced modifications of lipid homeostasis and trafficking will improve understanding of the compensatory role of LpR upregulation in neuronal adaptive responses (Yin, 2018).
Taken together, these studies strongly suggest that neuronal lipoprotein receptors are important components of activity-regulated neural plasticity and adaptive responses. In addition, transcriptome studies on neuronal-specific responses induced by excessive input activity provide molecular insights for studies related to human epilepsy and seizure disorders and potentially reveal the previously unappreciated role of altered neuronal lipoprotein uptake in neurological disorders associated with the dysregulation of lipid homeostasis (Yin, 2018).
Monensin Sensitive 1 (Mon1) is a component of the Mon1:Ccz1 complex that mediates Rab5 to Rab7 conversion in eukaryotic cells by serving as a guanine nucleotide exchange factor for Rab7 during vesicular trafficking. Mon1 activity modulates the complexity of Class IV dendritic arborization (da) neurons during larval development. Loss of Mon1 function leads to an increase in arborization and complexity, while increased expression, leads to reduced arborization. The ability of Mon1 to influence dendritic development is possibly a function of its interactions with Rab family GTPases that are central players in vesicular trafficking. Earlier, these GTPases, specifically Rab1, Rab5, Rab10, and Rab11 have been shown to regulate dendritic arborization. This study has conducted genetic epistasis experiments, by modulating the activity of Rab5, Rab7, and Rab11 in da neurons, in Mon1 mutants, and has demonstrated that the ability of Mon1 to regulate arborization is possibly due to its effect on the recycling pathway. Dendritic branching is critical for proper connectivity and physiological function of the neuron. An understanding of regulatory elements, such as Mon1, as demonstrated in this study, is essential to understand neuronal function (Harish, 2019).
Obtaining neuron transcriptomes is challenging; their complex morphology and interconnected microenvironments make it difficult to isolate neurons without potentially altering gene expression. Multidendritic sensory neurons (md neurons) of Drosophila larvae are commonly used to study peripheral nervous system biology, particularly dendrite arborization. This study sought to test if EC-tagging, a biosynthetic RNA tagging and purification method that avoids the caveats of physical isolation, would enable discovery of novel regulators of md neuron dendrite arborization. Aims in this study were twofold: discover novel md neuron transcripts and test the sensitivity of EC-tagging. RNAs were biosynthetically tagged by expressing CD:UPRT (a nucleobase-converting fusion enzyme) in md neurons and feeding 5-ethynylcytosine (EC) to larvae. Only CD:UPRT-expressing cells are competent to convert EC into 5-ethynyluridine-monophosphate which is subsequently incorporated into nascent RNA transcripts. Tagged RNAs were purified and used for RNA-sequencing. Reference RNA was prepared in a similar manner using 5-ethynyluridine (EUd) to tag RNA in all cells and negative control RNA-seq was performed on "mock tagged" samples to identify non-specifically purified transcripts. Differential expression analysis identified md neuron enriched and depleted transcripts. Three candidate genes encoding RNA-binding proteins (RBPs) were tested for a role in md neuron dendrite arborization. Loss-of-function for the m6A-binding factor Ythdc1 did not cause any dendrite arborization defects while RNAi of the other two candidates, the poly(A) polymerase Hiiragi and the translation regulator Hephaestus, caused significant defects in dendrite arborization. This work provides an expanded view of transcription in md neurons and a technical framework for combining EC-tagging with RNA-seq to profile transcription in cells that may not be amenable to physical isolation (Aboukilila, 2020).
Dendrite pruning of Drosophila sensory neurons during metamorphosis is induced by the steroid hormone ecdysone through a transcriptional program. In addition, ecdysone activates the eukaryotic initiation factor 4E-binding protein (4E-BP) to inhibit cap-dependent translation initiation. To uncover how efficient translation of ecdysone targets is achieved under these conditions, the requirements for translation initiation factors during dendrite pruning were assessed. The canonical cap-binding complex eIF4F was found to be dispensable for dendrite pruning, but the eIF3 complex and the helicase eIF4A are required, indicating that differential translation initiation mechanisms are operating during dendrite pruning. eIF4A and eIF3 are stringently required for translation of the ecdysone target Mical, and this depends on the 5' UTR of Mical mRNA. Functional analyses indicate that eIF4A regulates eIF3-mRNA interactions in a helicase-dependent manner. It is proposed that an eIF3-eIF4A-dependent alternative initiation pathway bypasses 4E-BP to ensure adequate translation of ecdysone-induced genes (Rode, 2018).
Pruning, the developmentally controlled degeneration of synapses and neurites without loss of the parent neuron, is an important mechanism used to specify neuronal connections or to remove developmental intermediates. In holometabolous insects like Drosophila, the nervous system is remodeled during metamorphosis in response to the steroid hormone ecdysone. In the peripheral nervous system (PNS), the sensory class IV dendritic arborization (c4da) neurons completely prune their long and branched larval dendrites at the onset of the pupal phase, while their axons stay intact. C4da neuron dendrite pruning involves the specific destabilization of the dendritic cytoskeleton and plasma membrane and phagocytosis of severed dendrites by surrounding epidermal cells (Rode, 2018).
Ecdysone induces c4da neuron dendrite pruning through the hormone receptors EcR-B1 and ultraspiracle (Usp), which activate the transcription of pruning genes. Among these are headcase, a pruning gene of unknown function, and SOX14, an HMG box transcription factor that activates transcription of MICAL, encoding an actin-severing enzyme. Regulation of MICAL expression also involves the ubiquitin-proteasome system at a posttranscriptional level (Rode, 2018).
In addition to transcriptional activation of target genes, several lines of evidence suggest that ecdysone also regulates global translation rates through activation of the translation inhibitor eukaryotic initiation factor 4E-binding protein (4E-BP). In the Drosophila fat body, this occurs transcriptionally through FOXO, while in c4 da neurons, ecdysone inhibits the insulin and Target of Rapamycin (TOR) pathway to activate 4E-BP posttranslationally (Rode, 2018).
4E-BP inhibits translation initiation, the rate-limiting step of protein synthesis, by sequestering the cap-binding protein eIF4E. During canonical translation initiation, eIF4E binds to the 7-methylguanosine (m7Gppp) cap of eukaryotic mRNAs and then forms the so-called eIF4F complex by recruiting eIF4G, an adaptor that binds the 43S preinitiation complex (PIC), containing the 40S small ribosomal subunit, and the helicase eIF4A, which is thought to resolve hairpin structures in the 5' UTRs of mRNAs. This enables the 43S complex to scan 5' UTRs for the initiation codon, where it is joined by the large ribosomal subunit and translation can start. While eIF4A's role has been mainly linked to 5' UTR hairpins, it can also stimulate translation of mRNAs with unstructured 5' UTRs. Moreover, eIF4A is more abundant than eIF4E, suggesting that it has functions beyond the eIF4F complex (Rode, 2018).
Activated 4E-BP binds to eIF4E and prevents eIF4F assembly, thus inhibiting ribosome recruitment to mRNAs and globally dampening translation rates under stress or during development. Interestingly, 4E-BP affects translation of some mRNAs more than others. To explain this, eIF4E-independent translation initiation mechanisms have been proposed. One such mechanism could depend on internal ribosome entry sites (IRESs) that bypass the requirement for the m7Gppp cap. For example, the mRNAs of the Drosophila cell death factors reaper and hid may contain IRES sequences in their 5' UTRs that allow them to be translated under stress (Rode, 2018).
Alternative cap recognition mechanisms have also been proposed under conditions of high 4E-BP activity. In particular, the initiation factor eIF3, a 13-subunit complex, could provide a mechanism for eIF4E-independent initiation (Lee, 2015; Lee, 2016). It binds to the small ribosomal subunit as part of the 43S PIC, and it is thought to act downstream of eIF4G in mRNA recruitment. However, eIF3 dependence varies between mRNAs, and eIF3 can even suppress translation of some targets (Lee, 2015). Importantly, it was recently shown that translation of some 4E-BP-resistant mRNAs depends on an eIF3-based cap recognition activity in the eIF3d subunit that is stimulated by hairpin motifs in the 5' UTR (Lee, 2016). Other eIF3 subunits have also been shown to interact with the cap (Rode, 2018).
Given that ecdysone inhibits eIF4E-dependent translation, this study asked whether there are mechanisms that ensure the translation of ecdysone target mRNAs. To this end, the requirements for translation initiation factors during c4da neuron dendrite pruning was assessed. The canonical eIF4F components eIF4E and eIF4G were found not to be required for c4da neuron dendrite pruning, while the helicase eIF4A and the eIF3 complex are. Both eIF4A and eIF3 are required for Mical expression, and this specificity is conferred by the 5' UTR of Mical mRNA. Further biochemical analyses suggest that eIF4A regulates the interaction between eIF3 and the Mical 5' UTR. It is proposed that eIF4A/eIF3 constitute a 4E-BP bypass mechanism that ensures the adequate translation of ecdysone-induced genes in c4da neurons (Rode, 2018).
Developmental control of translation rate is required under various conditions. One well-characterized regulatory mechanism is through 4E-BP, which inhibits assembly of the cap-binding eIF4F complex. Despite the obvious need for global translation control during development, it is also clear that there must be exceptions to such regulation. Several lines of evidence suggest that global, eIF4E-dependent translation is downregulated by ecdysone during the pupal phase and that this is important for c4da neuron dendrite pruning. How downregulation of eIF4E-dependent translation contributes to dendrite pruning is not clear. TOR activity (and hence eIF4E-dependent translation) is associated with neurite regrowth after pruning in a Drosophila model for neuronal remodeling, and beta-actin mRNA was identified as a 4E-BP target in vertebrate neurons. General suppression of eIF4E-dependent translation may, therefore, serve to prevent precocious neurite growth or neurite stabilization through increased actin polymerization (Rode, 2018).
Despite the need for translation downregulation during dendrite pruning, ecdysone-induced mRNAs must still be efficiently translated. This study found that c4 da neuron dendrite pruning does not depend on the eIF4F subunits eIF4E and eIF4G, but instead on eIF3 and eIF4A. In keeping with a specific effect on dendrite-pruning genes, Mical mRNA was identified as the crucial target for eIF3 and eIF4A. The data suggest that this specificity is encoded in the 5' UTR of Mical mRNA, as a UAS-GFP reporter containing the Mical 5' UTR showed consistently stronger dependence on eIF4A and eIF3 than a regular UAS-GFP reporter. The important role of the Mical 5' UTR is also supported by the observation that Sox14 overexpression (which induces endogenous Mical mRNA) did not rescue the pruning defects induced by eIF4A RNAi, while overexpression of Mical from a UAS transgene (and thus lacking the endogenous 5' UTR) did. It is tempting to speculate that eIF3-eIF4A recognition signals may be abundant in 5' UTRs of ecdysone-induced genes (Rode, 2018).
Several lines of evidence indicate that translation initiation of pupal pruning factors in c4 da neurons is still cap dependent: for one, overexpression of a cap-binding-deficient eIF3d mutant causes dominant dendrite-pruning defects, and in vitro translation of a 5' UTRMical reporter mRNA depends on a functional cap. While physical interactions were observed between eIF3 and a 5' UTR Mical reporter mRNA in S2 cells, cap binding by eIF3 could not be directly demonstrated in vivo. eIF3 does not bind to the isolated cap structure, and a biochemical cap-binding assay for eIF3 would require crosslinking eIF3 with a purified mRNA with a radioactively labeled cap. To further investigate developmental control of translation initiation in the future, it would be interesting to set up such an assay to address whether the Mical mRNA cap is also recognized via eIF3d or another eIF3 subunit (Rode, 2018).
Sox14 expression seemed resistant to the inhibition of either eIF4E or eIF3, but this study found that these pathways can mediate Sox14 expression in a redundant fashion. Sox14 is upstream of the Cul-1 ubiquitin ligase that activates 4E-BP in c4da neurons. Its mRNA may be adapted to this position in the pruning pathway, as it could still use the regular eIF4F pathway early during the pupal phase and the eIF3 pathway later. Mical translation may only start when 4E-BP activity is already high, hence explaining its strong eIF3 dependence (Rode, 2018).
Translation of long mRNAs is sensitive to the eIF4A cofactor eIF4B (Sen, 2016), and eIF4A dependence is also in part conferred by sequences in the coding region. eIF4B manipulation did not cause dendrite pruning defects, but the Mical construct used to rescue the pruning defects induced by eIF4A knockdown lacks an internal region non-essential for pruning. It is, therefore, possible that internal regions of the long Mical mRNA also contribute to its dependence on eIF4A (Rode, 2018).
The strong similarities between the phenotypes caused by the manipulation of eIF4A and eIF3 suggested that these two factors cooperate functionally. eIF4A and eIF3 can be found in an eIF4A ATPase-dependent complex and that eIF4A clamping on the mRNA prevents eIF3 release from a 5' UTRMical reporter mRNA. Two recent in vitro studies found functional interactions between eIF4A and eIF3 in the context of canonical eIF4F-dependent translation initiation (Yourik, 2017, Sokabe, 2017): first, eIF3 stimulates eIF4A ATPase activity via its eIF3g subunit to promote PIC maturation (Yourik, 2017); and, second, eIF4A ATPase activity was required to reposition the eIF3j subunit within the PIC during maturation (Sokabe and Fraser, 2017). this study now demonstrate genetically that eIF4A has an eIF3-related function independently of eIF4F. These data showing that eIF3 and eIF4A interact in an ATPase-dependent manner and that eIF4A helicase activity is required for dendrite pruning are consistent with both the above proposals (Rode, 2018).
Taken together, these data suggest that eIF3-eIF4A are part of a bypass mechanism that ensures translation of crucial ecdysone-induced mRNAs in the absence of an eIF4E-dependent translation initiation during developmental neuronal remodeling in the Drosophila PNS (Rode, 2018).
Brain development requires correct targeting of multiple thousand synaptic terminals onto staggeringly complex dendritic arbors. The mechanisms by which input synapse numbers are matched to dendrite size, and by which synaptic inputs from different transmitter systems are correctly partitioned onto a postsynaptic arbor, are incompletely understood. By combining quantitative neuroanatomy with targeted genetic manipulation of synaptic input to an identified Drosophila neuron, this study shows that synaptic inputs of two different transmitter classes locally direct dendrite growth in a competitive manner. During development, the relative amounts of GABAergic and cholinergic synaptic drive shift dendrites between different input domains of one postsynaptic neuron without affecting total arbor size. Therefore, synaptic input locally directs dendrite growth, but intra-neuronal dendrite redistributions limit morphological variability, a phenomenon also described for cortical neurons. Mechanistically, this requires local dendritic Ca2+ influx through Dα7 nAChRs or through low-voltage-activated channels following GABAA receptor-mediated depolarizations (Ryglewski, 2017).
These results are consistent with competition of GABAergic and cholinergic inputs for a given amount of dendrites. It is proposed that total dendrite size is determined by global cues, such as the transcriptional identity of the neuron, hormone signals, and overall firing activity, all of which regulate dendritic length and branch numbers in many types of neurons. Manipulations that increase MN5 firing activity cause dendrite overgrowth via activation of CaMKII- and AP1-dependent transcription. MN5 firing causes global Ca2+ influx into the soma and the dendrites. Conversely, knockdown of the Drosophila Cav2 homolog cacophony reduces somatic and dendritic Ca2+ influx upon firing and decreases dendrite arbor size. Therefore, overall MN5 firing activity and global Ca2+ influx regulate total dendrite length, likely together with steroid hormones. This study showa that the total amount of available dendrite becomes locally redistributed within the arbor by competitive synaptic mechanisms. A mechanistic separation between global overall growth and local fine branching control might enable the formation and maintenance of dendritic branches at sites of newly formed input synapses while preventing dendrite overgrowth, thus enabling opportunistic functional connections with minimal dendrites (Ryglewski, 2017).
Competitive interactions between GABAergic and cholinergic inputs may optimize dendritic gestalt to partition synaptic inputs from different transmitter systems to segregated postsynaptic input domains. Whenever synaptic input to the GABAergic or to the cholinergic domain is increased, dendrites shift toward this domain and away from the other one. Vice versa decreased input to one domain causes dendrite shift to the other domain. Increasing input to both domains has no effect, thus indicating synaptic activity-dependent competition. Although regulation of dendrite growth by synaptic activity has previously been reported for excitatory cholinergic and glutamatergic, as well as excitatory and inhibitory GABAergic synapses, intra-neuronal dendrite shifts that reshape the entire tree have not been demonstrated (Ryglewski, 2017).
However, it must be noted that not all neurons contain distinctly different input domains, but some receive intermingled inputs of different transmitter classes. In addition, this analysis focused on the most abundant excitatory and inhibitory inputs to MN5, which segregate to different input domains (Kuehn, 2013). MN5 is a flight motorneuron that is monopolar with more than 4000 dendritic branches. MN5 likely receives also glutamatergic inputs and potentially also aminergic modulatory inputs. It remains unclear whether such input classes are also targeted to specific input domains, or whether these also compete for dendrites. Nonetheless, the current findings are likely of general interest, because segregated input domains for different transmitter classes also exist in mammalian pyramidal neurons. Although the underlying mechanisms have not been addressed in the mammalian brain, size reductions in pyramidal neuron apical dendrites are accompanied by size increases of the basal dendrites and vice versa. Moreover, the relative sizes of pyramidal neuron dendritic sub-trees are counterbalanced and stabilize total dendritic length. Similarly, for the Drosophila MN5, previously studies have reported high size variability of individual dendritic sub-trees while counterbalance between sub-tree sizes keeps total dendrite size variability low. This study provides evidence that competitive interactions between different synaptic inputs locally direct arbor growth in different input domains. This may be a general means to fine tune the gestalt of neurons with segregated input domains. The data on an identified Drosophila motoneuron may provide an entry point into understanding the mechanism by which presynaptic partners may instruct the re-distribution of postsynaptic dendrites between different input domains in a 'synaptotropic' manner (Ryglewski, 2017).
Although the mechanisms by which competition between GABAergic and cholinergic inputs locally directs dendrite growth require further studies, the current data indicate that both competitors employ the same intracellular signaling mechanism. In retinal ganglion cells, newly formed dendritic branches become selectively stabilized at sites of cholinergic input synapses from amacrine cells, and strictly local dendritic Ca2+ signals are required for this process. This study also found local dendritic Ca2+ signals at sites of cholinergic and at sites of GABAergic input, although there is no direct live imaging evidence of local branch stabilization by these signals. However, any manipulation, be it presynaptic or postsynaptic, that affects synaptic input to either the cholinergic or the GABAergic domain changes arbor size in that domain. Local dendritic Ca2+ signals at cholinergic synapses are expected, because the Dα7nAChR conducts Ca2+. By contrast, the Rdl GABAAR is a chloride channel. During vertebrate embryonic development, GABAAR activation is initially excitatory because of elevated intracellular chloride concentrations. This study found that such a developmental shift of the chloride reversal potential to more negative values at mature stages likely also exists in insect neurons and thus, may be more conserved than previously known (Ryglewski, 2017).
As a result, GABAAR activation during Drosophila pupal stages causes depolarization, which in turn, induces local dendritic Ca2+ influx through DmαG LVA Ca2+ channels. LVA channels are required for GABA-mediated dendrite redistribution, because GABAergic input fails to direct dendrites to the GABAergic domain in DmαG null mutants. This interpretation is consistent with a recent study demonstrating that depolarizing GABAergic input selectively promotes apical dendrite growth via Ca2+ influx during neonatal cortical neuron development. Therefore, directing dendrite arbor growth via GABA-mediated Ca2+ signals takes place in flies and mammals (Ryglewski, 2017).
In agreement with findings that basic flight motoneuron function is retained even with a minimal set of dendrites, intra-neuronal dendrite shifts as caused by either Rdl or nAChR overexpression did not alter firing rates or wingbeat frequencies during tethered flight. By contrast, adaptive adjustments of motoneuron firing frequencies in response to visual input during flight behavior were impaired. The defect was more severe with increased excitation and dendrite shifts to the cholinergic domain than with increased inhibition and dendrite shifts to the GABAergic domain. By contrast, motoneuron firing frequency adjustments were not affected following conditional receptor expression in mature neurons that caused no dendrite shifts. This suggested that the impairment was at least in part caused by dendrite redistribution, although this interpretation requires caution. First, possible indirect effects on network properties to result from receptor overexpression in MN5 through development cannot be completely excluded. Second, although the amount of receptor overexpression can be estimated, light microscopy analysis cannot rule out potential differences between conditional and chronic receptor expression. However, the data indicate that the balance between excitatory cholinergic input to the proximal dendritic domain and inhibitory GABAergic input to the distal domain may play a critical role for adaptive adjustments of motoneuron firing rates during behavior (Ryglewski, 2017).
Directing more dendrites to the input domain of increased GABAergic synaptic input and away from the domain of decreased cholinergic input, or vice versa, seems to contradict the concept of homeostasis. Why to shift more dendrites to sites with many synaptic partners, but away from sites with lower synapse densities? First, a mechanism that locally matches the amount of available dendritic surface to the amount of synapses is in accordance to minimizing cost. Maximum synapse numbers on minimal dendritic length has been proposed as a general principle in cortex. Second, redistribution of dendrites to domains with high synapse density may prevent superfluous increases of total arbor length. Naming dendrite redistribution while maintaining total length 'dendritic conservation' is suggested. Can dendritic conservation function in parallel to homeostatic control of excitability? Tight regulation of excitation-inhibition ratios is essential for normal circuit function. Providing more dendrites to inhibitory or to excitatory input domains, depending on GABAergic or cholinergic synaptic activity during arbor growth does not preclude compensatory adjustments of neuronal excitability on the levels of synapse strength or ion channel expression, both of which have been reported conserved mechanisms from invertebrates to mammals. In fact, even with forced receptor overexpression or massive thermogenetic stimulation of synaptic inputs through development, both of which caused dendrite shifts, multiple aspects of basic flight motor performance were normal. Therefore, at least to a certain degree, intra-neuronal dendrite shifts may operate in concert with homeostatic adjustments to keep overall excitability in check (Ryglewski, 2017).
Animal locomotion depends on proprioceptive feedback, which is generated by mechanosensory neurons. A genetic screen for impaired walking was performed in Drosophila, and a gene, stumble (stum), was isolated. The Stum protein has orthologs in animals ranging from nematodes to mammals and is predicted to contain two transmembrane domains. Expression of the mouse orthologs of stum in mutant flies rescued their phenotype, which demonstrates functional conservation. Dendrites of stum-expressing neurons in legs were stretched by both flexion and extension of corresponding joints. Joint angles that induced dendritic stretching also elicited elevation of cellular Ca(2+) levels-not seen in stum mutants. Thus, this study has identified an evolutionarily conserved gene, stum, which is required for transduction of mechanical stimuli in a specific subpopulation of Drosophila proprioceptive neurons that sense joint angles (Desai, 2014).
Animal locomotion is achieved by coordination of motor activity according to proprioceptive mechanosensory inputs. In Drosophila, mechanosensation is mediated either by ciliated or multidendritic receptor neurons. Multidendritic neurons can respond to direct application of mechanical force to their membranes. It is less clear, however, how multidendritic mechanosensory neurons can be tuned to one mechanical modality, such as joint angle, and disregard other mechanical stimuli that may originate from external impacts or changes in the shape of muscles during contraction. In order to identify genes involved in proprioceptive sensation, uncoordination was screened for in a collection of ethyl methanesulfonate-mutagenized Drosophila lines. Lines that exhibited walking impairments were selected, and the phenotype severity was quantified by measuring climbing speed. Three lines, 204, 922, and 4487, were identified that showed lack of coordination in homozygous flies and did not complement one another, which suggested that they represent alleles of the same gene. Two of the lines, 204 and 4487, showed severe uncoordination, and the phenotype in 922 was mild. Deficiency mapping pointed to a gene, CG30263, predicted to encode a large protein with 1870 or 1959 amino acids (depending on the splice variant). Because the mutants had a walking impairment phenotype, this gene was named stumble (stum). The stum ortholog in humans is C1orf95, and it was categorized as a member of the SPEC3 family (UNIPROT, INTERPRO), with unknown function. The stum gene was sequenced in the three mutant lines, and it was found that stum204, stum4487, and stum922 had stop codons at amino acid positions 171, 202, and 1081, respectively (Desai, 2014).
Transgenic flies that express stum cDNA in neurons were generated and it was found that the stum phenotype was partially rescued, which indicated that the mutations in stum were underlying the uncoordination phenotype. It is noted that the mouse ortholog of stum (National Center for Biotechnology Information, the U.S. National Institutes of Health, reference sequence: NP_001074696.1), which is only 141 amino acids long and shares 33% sequence identity with Drosophila stum, was also able to substantially rescue the uncoordination phenotype. Therefore, the function of stum appears to be conserved between distant animal species. Moreover, the rescue of the phenotype with such a short form of stum suggests that the C-terminal region of the fly protein constitutes the functional core (Desai, 2014).
Proprioceptive defects in adult flies have been attributed to malfunction of type I (ciliated) mechanoreceptor neurons. To test whether such defects also underlie the phenotype in the stum mutant, electrophysiological recordings were performed of mechanical responses from the ciliated mechanoreceptor neuron of the anterior notopleural bristle. Type I neurons of stum mutants were indistinguishable from controls. Therefore, unlike known proprioception mutants, the phenotype in stum mutants does not arise from a general defect in type I mechanoreceptor neurons. To identify which cells give rise to the phenotype, the genomic regulatory region of stum was used to drive a CD8 fused to green fluorescent protein (CD8GFP) reporter [stum-Gal4 driving the upstream activation sequence (UAS)–CD8GFP]. It was found that stum expression in the legs was localized to three labeled neurons: one at the femur-tibia joint, the second at the tibia-tarsus joint, and the third spanning the second tarsal segment. The cell bodies of these stum-expressing neurons were located near the distal end of each leg segment, and their dendrites terminated at the corresponding joints (Desai, 2014).
To study whether there are stum-expressing cells within the ventral nerve cord (VNC), it was examined in flies that express CD8GFP using stum-Gal4. The only fluorescent signal in the VNC originated from the axons of the leg neurons. The axons terminated within the neuropil that corresponds to each particular leg, branching into a bowl shape. This pattern is typical of neurons that take part in proprioception, such as the hair plate neurons. Therefore, the stum-expressing cells in the Drosophila body have characteristics of proprioceptive neurons that sense a property of specific joints (Desai, 2014).
Confocal imaging was performed of the dendritic region of stum-expressing neurons, and it was found that, close to its tip, the dendrite branches toward the lateral aspect of the joint, and this side branch terminates at a short distance from the cuticle. The tips of dendrite were not associated with a cuticular structure or a scolopale. These structural features are typical features of type II (multidendritic) neurons but are incompatible with type I mechanoreceptor neurons that terminate with a ciliary structure. Thus, stum uncoordination mutations affect type II mechanoreceptor neurons. Furthermore, the entire dendritic terminal of these neurons was located in a region that is devoid of musculature, which suggests that they do not sense the mechanical properties of muscles. Taken together, these data suggest that stum-expressing neurons sense a mechanical property of the joint (Desai, 2014).
To test whether stum-positive neurons encode joint angles, high-resolution imaging of the tibia-tarsus joint area was performed at different angles. At each angle of the joint, the total length of the sensory dendrite and its side branch was measured. It was found that the total dendrite length had a minimum typically at 130o to 170o, and it increased when the joint was shifted to either more obtuse or more acute angles. These morphological changes indicate that these neurons are mechanically affected by the position of the joint. Because the tip of the side branch is stationary, it is likely that the change in total length results from the coupling of the tip of the main dendrite to the motion of the distal joint segment. The position of the dendrite and the susceptibility of its morphology to joint angle suggest that the role of stum-positive neurons is to sense and encode the angle of the joint (Desai, 2014).
Ca2+ fluorescence was measured while forcing the tibia-tarsus joint to different angles, and the Ca2+ fluorescence in stum-expressing neurons was found to correlate with the angle of the joint. As in the morphological changes, the responses correlated with joint angle in a U-shaped manner, where both acute and obtuse angles induced increasing Ca2+ elevations. These results indicate that the stum-positive neurons encode proprioceptive information about the angle of joints. It is noteworthy that a similar U-shaped encoding of joint angles has also been described in receptor neurons of mammalian joints, which suggests that sensing the deviations from a neutral joint range is universally critical for motor function (Desai, 2014).
The walking impairment in the stum mutant fly suggests that the gene is necessary for generating the proper proprioceptive responses in stum-expressing neurons. To test whether the stum mutations affect coupling between joint angles and dendritic stretching, the stretching was quantitated in mutant flies that have stum-expressing neurons labeled with CD8GFP. In the stum mutant, the dendritic stretching in response to joint angles was comparable to that of control flies. Thus, stum is not required for the mechanical coupling between joint angles and stretching of the sensory dendrites (Desai, 2014).
Although the dendritic stretching was not significantly affected in the stum mutant, the Ca2+ responses to both acute and obtuse joint angles were abolished in the mutant. Therefore, it is concluded that stum is essential for transducing dendrite stretching into cellular responses (Desai, 2014).
The morphology of stum-expressing neurons were examined, and it was found that, although axons and cell bodies of stum mutants were indistinguishable from controls, the sensory dendrite in mutants exhibited abnormalities. Most notably, in some of the stum-expressing neurons the tip of the dendrite was overgrown and extended into the distal segment of the joint. Thus, the absence of stum leads to a morphological defect, possibly because of the lack of mechanical responsiveness. The occasional morphological differences may account for the slight difference in the stretching profile between control and mutant dendrites. The fraction of neurons demonstrating the abnormal morphology in the mutant increased from the day of eclosion to the following day. As the addition of morphological changes takes place after eclosion, it is possible that stum-dependent activity is essential for late shape determination that can take place in adult multidendritic neurons (Desai, 2014).
Transgenic flies were generated that express a GFP fused with the N terminus of Stum under UAS regulation (UAS-GFP-Stum). The Stum fusion protein was found to be specifically localized to the distal part of the sensory dendrite, although it did not accumulate substantially in any other part of the cell. The fluorescent signal started at the region of bifurcation and extended to both distal tips of the dendrite. This specific localization suggests that Stum functions in the part of the dendrite that senses stretching (Desai, 2014).
Taken together, stum expression in mechanosensory neurons, Stum localization to the sensory dendrite, and the abolition of responses to stretching in the stum mutant suggest that stum has an essential role in mediating mechanical sensing in receptor neurons. Because the Stum protein in most species is very small and because Drosophila stum is expressed in limited populations of receptor neurons, it is proposed that stum is not the mechanically activated channel. Rather, stum may serve as an accessory module that is essential for the proper localization or function of the transduction channels (Desai, 2014).
The stretch-receptor neurons that express stum present an elegant engineering solution for generating specificity to the modality of mechanical stimulus. The distal part of their dendrite bifurcates into two branches whose tips are anchored to parts of the joint that shift their relative positions. Sensing the stretching only between the two dendritic tips may tune the nerve responses to joint motions and filter out the effect of irrelevant mechanical impacts. This specificity enables the sensory neuron to relay reliable proprioceptive information to the central nervous system (Desai, 2014).
Alternative splicing mediated by RNA-binding proteins (RBPs) is emerging as a fundamental mechanism for the regulation of gene expression. Alternative splicing has been shown to be a widespread phenomenon that facilitates the diversification of gene products in a tissue specific manner. Although defects in alternative splicing are rooted in many neurological disorders, only a small fraction of splicing factors have been investigated in detail. This study finds that the splicing factor Caper is required for the development of multiple different mechanosensory neuron subtypes at multiple life stages in Drosophila melanogaster. Disruption of Caper function causes defects in dendrite morphogenesis of larval dendrite arborization neurons, neuronal positioning of embryonic proprioceptors, as well as the development and maintenance of adult mechanosensory bristles. Additionally, Caper dysfunction was found to result in aberrant locomotor behavior in adult flies. Transcriptome-wide analyses further support a role for Caper in alternative isoform regulation of genes that function in neurogenesis. This results provide the first evidence for a fundamental and broad requirement for the highly conserved splicing factor Caper in the development and maintenance of the nervous system and provide a framework for future studies on the detailed mechanism of Caper mediated RNA regulation (Olesnicky, 2017).
Adequate responses to noxious stimuli causing tissue damages are essential for organismal survival. Class IV neurons in Drosophila larvae are polymodal nociceptors responsible for thermal, mechanical, and light sensation. Importantly, activation of Class IV provoked distinct avoidance behaviors, depending on the inputs. Noxious thermal stimuli, but not blue light stimulation, was shown to cause a unique pattern of Class IV, which were composed of pauses after high frequency spike trains and a large Ca2+ rise in the dendrite (the Ca2+ transient). Both of these responses depended on two TRPA channels and the L-type voltage-gated calcium channel (L-VGCC), showing that the thermosensation provokes Ca2+ influx. The precipitous fluctuation of firing rate in Class IV neurons enhanced the robust heat avoidance. It is hypothesized that the Ca2+ influx can be a key signal encoding a specific modality (Terada, 2016).
Nociceptive neurons of Drosophila melanogaster larvae are characterized by highly branched dendritic processes whose proper morphogenesis relies on a large number of RNA-binding proteins. Post-transcriptional regulation of RNA in these dendrites has been found to play an important role in their function. This study investigated the neuronal functions of two putative RNA modification genes, RluA-1 and RluA-2, which are predicted to encode pseudouridine synthases. RluA-1 is specifically expressed in larval sensory neurons while RluA-2 expression is ubiquitous. Nociceptor-specific RNAi knockdown of RluA-1 caused hypersensitive nociception phenotypes, which were recapitulated with genetic null alleles. These were rescued with genomic duplication and nociceptor-specific expression of UAS-RluA-1-cDNA As with RluA-1, RluA-2 loss of function mutants also displayed hyperalgesia. Interestingly, nociceptor neuron dendrites showed a hyperbranched morphology in the RluA-1 mutants. The latter may be a cause or a consequence of heightened sensitivity in mutant nociception behaviors (Song, 2020).
Spatial arrangement of different neuron types within a territory is essential to neuronal development and function. How development of different neuron types is coordinated for spatial coexistence is poorly understood. In Drosophila, dendrites of four classes of dendritic arborization (C1-C4da) neurons innervate overlapping receptive fields within the larval epidermis. These dendrites are intermittently enclosed by epidermal cells, with different classes exhibiting varying degrees of enclosure. The role of enclosure in neuronal development and its underlying mechanism remain unknown. This study shows that the membrane-associated protein Coracle acts in C4da neurons and epidermal cells to locally restrict dendrite branching and outgrowth by promoting enclosure. Loss of C4da neuron enclosure results in excessive branching and growth of C4da neuron dendrites and retraction of C1da neuron dendrites due to local inhibitory interactions between neurons. It is proposed that enclosure of dendrites by epidermal cells is a developmental mechanism for coordinated innervation of shared receptive fields (Tenenbaum, 2017).
The systematic variation of diameters in branched networks has tantalized biologists since the discovery of da Vinci's rule for trees. Da Vinci's rule can be formulated as a power law with exponent two: The square of the mother branch's diameter is equal to the sum of the squares of those of the daughters. Power laws, with different exponents, have been proposed for branching in circulatory systems (Murray's law with exponent 3) and in neurons (Rall's law with exponent 3/2). The laws have been derived theoretically, based on optimality arguments, but, for the most part, have not been tested rigorously. Using superresolution methods to measure the diameters of dendrites in highly branched Drosophila class IV sensory neurons, this study has found that these types of power laws do not hold. In their place, a different diameter-scaling law was developed: The cross-sectional area is proportional to the number of dendrite tips supported by the branch plus a constant, corresponding to a minimum diameter of the terminal dendrites. The area proportionality accords with a requirement for microtubules to transport materials and nutrients for dendrite tip growth. The minimum diameter may be set by the force, on the order of a few piconewtons, required to bend membrane into the highly curved surfaces of terminal dendrites. Because the observed scaling differs from Rall's law, it is proposed that cell biological constraints, such as intracellular transport and protrusive forces generated by the cytoskeleton, are important in determining the branched morphology of these cells (Liao, 2021).
Coordination of dendrite growth with changes in the surrounding substrate occurs widely in the nervous system and is vital for establishing and maintaining neural circuits. However, the molecular basis of this important developmental process remains poorly understood. To identify potential mediators of neuron-substrate interactions important for dendrite morphogenesis, this study undertook an expression pattern-based screen in Drosophila larvae, which revealed many proteins with expression in dendritic arborization (da) sensory neurons and in neurons and their epidermal substrate. Reporters for Basigin, a cell surface molecule of the immunoglobulin (Ig) superfamily previously implicated in cell-cell and cell-substrate interactions, are expressed in da sensory neurons and epidermis. Loss of Basigin in da neurons led to defects in morphogenesis of the complex dendrites of class IV da neurons. Classes of sensory neurons with simpler branching patterns were unaffected by loss of Basigin. Structure-function analyses showed that a juxtamembrane KRR motif is critical for this function. Furthermore, knock down of Basigin in the epidermis led to defects in dendrite elaboration of class IV neurons, suggesting a non-autonomous role. Together, these findings support a role for Basigin in complex dendrite morphogenesis and interactions between dendrites and the adjacent epidermis (Shrestha, 2021).
Class I ventral posterior dendritic arborisation (c1vpda) proprioceptive sensory neurons respond to contractions in the Drosophila larval body wall during crawling. Their dendritic branches run along the direction of contraction, possibly a functional requirement to maximise membrane curvature during crawling contractions. Although the molecular machinery of dendritic patterning in c1vpda has been extensively studied, the process leading to the precise elaboration of their comb-like shapes remains elusive. To link dendrite shape with its proprioceptive role, a long-term, non-invasive, in vivo time-lapse imaging was perfomred of c1vpda embryonic and larval morphogenesis to reveal a sequence of differentiation stages. Computer models and dendritic branch dynamics tracking were used to propose that distinct sequential phases of stochastic growth and retraction achieve efficient dendritic trees both in terms of wire and function. This study shows how dendrite growth balances structure-function requirements, shedding new light on general principles of self-organisation in functionally specialised dendrites (Ferreira Castro, 2020).
Disruptions in lipid homeostasis have been observed in many neurodevelopmental disorders that are associated with dendrite morphogenesis defects. However, the molecular mechanisms of how lipid homeostasis affects dendrite morphogenesis are unclear. This study found that easily shocked (eas), which encodes a kinase with a critical role in phospholipid phosphatidylethanolamine (PE) synthesis, and two other enzymes in this synthesis pathway are required cell autonomously in sensory neurons for dendrite growth and stability. Furthermore, this study shows that the level of Sterol Regulatory Element-Binding Protein (SREBP) activity is important for dendrite development. SREBP activity increases in eas mutants, and decreasing the level of SREBP and its transcriptional targets in eas mutants largely suppresses the dendrite growth defects. Furthermore, reducing Ca(2+) influx in neurons of eas mutants ameliorates the dendrite morphogenesis defects. This study uncovers a role for EAS kinase and reveals the in vivo function of phospholipid homeostasis in dendrite morphogenesis (Meltzer, 2017).
A growing body of evidence suggests that lipid metabolism and homeostasis is important for nervous system development. This study provides evidence that EAS kinase and SREBP signaling pathway are required for dendrite growth during development. The results do not rule out the possibility that a reduction in PE levels could also independently contribute to the eas dendritic phenotype, due to a lack of PE supply for expanding the plasma membrane during dendrite growth. Interestingly, although both reduced and increased SREBP activity led to the same dendrite reduction phenotype, the underlying mechanism may be different. Reduced lipid production caused by low SREBP activity might impair the lipid supply for growing dendrites, while aberrantly increased lipid production by SREBP might lead to lipotoxicity. Since the homologues of both genes are highly conserved, it will be interesting to determine their roles in dendrite morphogenesis within the developing mammalian nervous system (Meltzer, 2017).
It was observed that terminal dendrite dynamics, but not initial establishment of the dendritic territory, is perturbed in eas mutants. It is possible that terminal dendrite growth and stability rely more on PE, while the primary and secondary branches do not. It is also possible that although PE synthesis is impaired in eas mutants, neurons still have enough PE to support initial dendrite growth. As the animal grows, neurons cannot supply enough PE for the exuberant terminal dendrite growth. It would be of interest to identify how impairment in PE specifically affects terminal dendrite dynamics in future studies (Meltzer, 2017).
Reducing the level of cacophony, a subunit of the Cav2 voltage-gated Ca2+ channel, ameliorates the dendrite morphogenesis defects in easKO mutants. In contrast, reducing SREBP signaling largely rescues the dendrite phenotypes but does not suppress the adult seizure-like phenotype. It is conceivable that the part of the neural circuit that heavily relies on SREBP signaling for dendrite morphogenesis during development does not contribute to the seizure-like behavior. Consistent with this hypothesis, expressing eas acutely in neurons in adult eas flies suppresses the seizure-like phenotype. It thus appears that alterations in neuronal Ca2+ influx contribute to not only seizure-like phenotypes but also dendrite morphogenesis defects during larval development (Meltzer, 2017).
In summary, this study uncovers an important role for phospholipid PE synthesis in neural development, and they further reveal a molecular pathway involving conserved molecules for phospholipid homeostasis as being important in the regulation of dendrite morphogenesis (Meltzer, 2017).
Highly motile dendritic filopodia are widely present in neurons at early developmental stages. These exploratory dynamic branches sample the surrounding environment and initiate contacts with potential synaptic partners. Although the connection between dendritic branch dynamics and synaptogenesis is well established, how developmental and activity-dependent processes regulate dendritic branch dynamics is not well understood. This is partly due to the technical difficulties associated with the live imaging and quantitative analyses of these fine structures using an in vivo system. A method was establised to study dendrite dynamics using Drosophila larval ventral lateral neurons (LNvs), which can be individually labeled using genetic approaches and are accessible for live imaging. Taking advantage of this system, protocols were developed to capture branch dynamics of the whole dendritic arbor of a single labeled LNv through time-lapse live imaging. Post-processing was performed to improve image quality through drift correction and deconvolution, followed by analyzing branch dynamics at the single-branch level by annotating spatial positions of all branch terminals. Lastly, R scripts and specific parameters were developed to quantify branch dynamics using the coordinate information generated by the terminal tracing. Collectively, this protocol allows achieving of a detailed quantitative description of branch dynamics of the neuronal dendritic arbor with high temporal and spatial resolution. The methods developed in this study are generally applicable to sparsely labeled neurons in both in vitro and in vivo conditions (Sheng, 2019).
Critical periods (brief intervals during which neural circuits can be modified by activity) are necessary for proper neural circuit assembly. Extended critical periods are associated with neurodevelopmental disorders; however, the mechanisms that ensure timely critical period closure remain poorly understood. This study defined a critical period in a developing Drosophila motor circuit and identified astrocytes as essential for proper critical period termination. During the critical period, changes in activity regulate dendrite length, complexity and connectivity of motor neurons. Astrocytes invaded the neuropil just before critical period closure, and astrocyte ablation prolonged the critical period. Finally, a genetic screen was used to identify astrocyte-motor neuron signalling pathways that close the critical period, including Neuroligin-Neurexin signalling. Reduced signalling destabilized dendritic microtubules, increased dendrite dynamicity and impaired locomotor behaviour, underscoring the importance of critical period closure. Previous work defined astroglia as regulators of plasticity at individual synapses. This study shows that astrocytes also regulate motor circuit critical period closure to ensure proper locomotor behaviour (Ackerman, 2021).
Critical periods are brief windows during which neural circuit activity can modify the morphological properties of neurons, producing permanent changes to circuit structure and function. Critical periods integrate multiple forms of plasticity to modify neural circuits. 'Homeostatic plasticity' encompasses changes to synapse number, structure and function across an entire neuron, as well as changes to long-range connectivity. Whereas homeostatic plasticity can occur in the adult brain, substantial activity-dependent remodelling peaks in early development. Indeed, failure to terminate critical period plasticity is linked to neurodevelopmental disorders such as autism and epilepsy. Although putative critical period disorders present with motor defects, the field has largely focused on sensory circuits. To that end, this study developed a novel critical period model in a developing motor circuit (Ackerman, 2021).
This study focused on two well-characterized Drosophila motor neurons, aCC and RP2, which are segmentally repeated in the central nervous system. These motor neurons are susceptible to activity-induced remodelling, although pioneering studies used chronic activity manipulations and did not define an end point for homeostatic plasticity. This study expressed the anion channelrhodopsin GtACR215 specifically in the aCC-RP2 motor neurons using the Gal4-upstream activation system (UAS) system and delivered acute 1-h windows of silencing, terminating at progressively later times in development. Silencing motor neurons for the last hour of embryogenesis (stage 17) increased aCC-RP2 dendritic volume at 0 h after larval hatching (ALH), whereas silencing for 1 h at later stages showed progressively less of an effect, with no remodelling occurring at 8 h ALH or beyond. By contrast, acute 1-h windows of activation using the channelrhodopsin Chrimson resulted in significant loss of motor neuron dendrites at 0 h ALH; activation at 8 h ALH and beyond had little or no effect. Activity-induced changes to dendrite length for single-cell RP2 clones [using the MultiColor FlpOut (MCFO) system] showed similar results Note that these experiments used far shorter periods of tonic activation than past studies. Although Tonic activity manipulations were primarily used, identical results were observed using 600 ms:400 ms pulses of activation or silencing, as well as thermogenetics to activate (via TrpA1) or silence [using the temperature-sensitive shibire gene (shibirets)] motor neurons. Notably, dendrite loss following acute activation could be rescued by a 22-h period of dark rearing, indicating that activity induces dendrite plasticity and not excitotoxicity. Together, these experiments define a critical period for activity-dependent motor dendrite plasticity represent the first analyses of motor circuit critical period closure within the central nervous system (Ackerman, 2021).
In vertebrates, homeostatic plasticity functions on a slow timescale, from hours to days. To determine the timescale for motor neuron dendrite expansion following GtACR2 silencing, aCC-RP2 motor neurons were silenced for 15 min, 1 h or 4 h in stage 17 embryos, terminating silencing at 0 h ALH. Larvae were then immediately dissected and dendritic morphology was assessed in single, well-spaced RP2 neurons using MCFO17. Increased dendritic arbor size and complexity following 1 h and 4 h of silencing were used. These results were confirmed using shibirets. By contrast, embryonic Chrimson activation resulted in decreased dendrite length and complexity at 0 h ALH after as little as 15 min of activation. Furthermore, using live imaging, significant dendrite retraction was observed within 12 min of Chrimson activation. The fact that silencing required more time to show an effect is not surprising, as extension requires generation of new membrane. It is concluded that activity-induced remodelling of Drosophila motor neurons occurs within minutes, much more quickly than previously documented for homeostatic plasticity in mammals (Ackerman, 2021).
This study showed that motor neurons scale dendrite length according to activity. An important question is whether these morphological changes are accompanied by changes in excitatory or inhibitory synaptic inputs. The excitatory cholinergic neuron A18b and inhibitory GABAergic neuron A23a were examined that are synaptically coupled to aCC-RP2 dendrites in a larval transmission electron microscopy (TEM) reconstruction. To quantify excitatory and inhibitory synapse number by light microscopy, a functionally inactive pre-synaptic marker, Bruchpilotshort::Cherry (Brp), was expressed in excitatory cholinergic neuron A18b or inhibitory GABAergic neuron A23a using the complementary LexA-LexAop binary expression system. A23a-inhibitory GABAergic synapses onto aCC-RP2 dendrites were examined, quantifying cell-type specific Brp puncta overlapping with aCC-RP2 dendritic membrane (putative synapses) using published standards. All critical period manipulations terminated at 4 h ALH (stage matched to the TEM data). It was found that 1 h of motor neuron silencing reduced the number of inhibitory synapses between A23a and aCC-RP2 dendrites. Silencing for a longer period (4 h) also yielded a significant increase in A18b excitatory synapses. Decreasing motor neuron activity thus leads to a compensatory reduction of inhibitory inputs and a corresponding increase in excitatory inputs to rebalance network activity. A18b excitatory cholinergic synapse numbers onto aCC-RP2 dendrites were quantified after activation or silencing. Motor neuron activation was found to significantly decreased numbers of A18b excitatory synapses onto aCC-RP2 dendrites following 1 h and 4 h manipulations. A significant increase in inhibitory synapse number following extended motor neuron activation was observed, possibly owing to insufficient dendritic membrane after activity-induced dendrite retraction. Increasing motor neuron activity thus leads to a compensatory reduction of excitatory pre-synaptic inputs. Finally, a functionally inactive reporter of excitatory post-synaptic densities (Drep2::GFP or Drep2::mStrawberry) was observed, specifically in aCC-RP2, and scaling of synapses was observed accross the entire dendritic arbor in response to altered activity—reduced excitatory post-synapses followed motor neuron activation, whereas increased excitatory post-synapses followed motor neuron silencing during the critical period. Of note, homeostatic scaling of motor neuron synapses did not occur after critical period closure. In sum, motor neurons scale excitatory and inhibitory inputs relative to their level of activity during the critical period (Ackerman, 2021).
The mechanisms that close critical periods remain poorly defined. Drosophila astrocytes infiltrate the neuropil at late embryogenesis and progressively envelop motor neuron synapses as the critical period closes. To test whether astrocytes promote critical period closure, all astrocytes were genetically ablated and optogenetics was used to assay for extension of critical period plasticity at 8 h ALH. Astrocyte elimination was confirmed by loss of the astrocyte marker Gat3. As expected, controls closed the critical period by 8 h ALH. By contrast, astrocyte ablation extended dendrite plasticity following Chrimson activation or GtACR2 silencing up to 8 h ALH. This effect was not observed at earlier stages, indicating that astrocytes do not constitutively dampen plasticity. Additionally, it was found that control motor dendrites were less dynamic after critical period closure, but that astrocyte ablation extends dendrite filopodial dynamicity. It is concluded that astrocytes are required for the transition from dynamic to stable filopodia and concurrent critical period closure (Ackerman, 2021).
To determine how astrocytes close the critical period, the astrocyte-specific alrm-gal4 was used to perform a targeted UAS RNA-mediated interference (RNAi) knockdown screen. Flies were assayed for critical period extension following 1 h of Chrimson activation from 7-8 h ALH. Four genes were identified that were required in astrocytes for timely critical period closure: gat (regulates excitatory-inhibitory balance), chpf [synthesizes chondroitin sulfate proteoglycans (CSPGs)] and the Neuroligins (Nlg) 4 and 2 (Ackerman, 2021).
Neuroligins are cell-adhesion proteins that are known to regulate astrocyte morphogenesis. In Drosophila, astrocyte-specific knockdown of nlg2 (the mouse orthologue is known as Nlgn1) had no effect on astrocyte volume or tiling, suggesting a more specific defect in astrocyte-motor neuron signalling. Knockdown of the remaining critical period regulators had variable effects on astrocyte morphology but all extended the critical period. Neuroligins bind cell adhesion proteins called Neurexins. RNAi against nrx-1 was used, which is known to bind both Nlg2 and Nlg4, specifically in aCC-RP2 motor neurons, and critical period extension was observed; this is consistent with astrocyte Nlg2 and motor neuron Nrx-1 acting in a common pathway to close the critical period. Motor neuron-specific RNAi knockdown of the CSPG receptor Lar also extended critical period plasticity. Notably, while Nrx-1 is often pre-synaptic, there is evidence for dendritic localization of these receptors. Furthermore, antibody staining for endogenous Nrx-1 and Nlg2 revealed localization of this receptor-ligand pair on motor dendrites and astrocytes, respectively. Finally, cell-type-specific overexpression of Nrx-1 and Nlg2 could induce precocious critical period closure (assayed by Chrimson activation from 3-4 h ALH). It is concluded that Nlg2-Nrx-1 ligand-receptor signalling between astrocytes and motor neurons is required for timely critical period closure (Ackerman, 2021).
How does Nlg2-Nrx-1 signalling close the critical period? The balance of excitatory to inhibitory synapses in neural circuits can instruct critical period timing. Additionally, numbers of excitatory synapses are decreased following astrocyte-specific knockout of neuroligins in mouse. This study observed no significant changes in excitatory-inhibitory balance following knockdown of nlg2 in astrocytes, suggesting that critical period closure is not dependent on Nlg2-mediated excitatory-inhibitory synapse balance (Ackerman, 2021).
Alternatively, Nrx-1 can promote microtubule stability in axons of motor neurons, suggesting a mechanism for critical period closure involving microtubule stabilization. To test this hypothesis, Chrimson::mVenus was used to activate and visualize aCC-RP2 dendrite membranes at 0 h ALH (peak critical period), and Cherry::Zeus to visualize stable microtubules during and after dendritic retraction. In live preparations, dendrites showed a reduction in Cherry::Zeus intensity immediately preceding activity-dependent retraction, suggesting that microtubule collapse in distal branches can induce dendrite retraction. In fixed preparations, this study found that proximal dendrites with the highest levels of stable microtubules were protected from activity-dependent retraction. Of note, overexpression of Nrx-1 was sufficient to increase both stable microtubules and stable dendrites at 4 h ALH. It is proposed that Nlg2 in astrocytes binds Nrx-1 in motor neurons to stabilize dendritic microtubules and close the critical period (Ackerman, 2021).
In mammals, inappropriate critical period extension has long-term effects on nervous system function. Indeed, this study observed persistent changes in motor neuron connectivity at least 24 h following acute motor neuron activation at the end of the critical period, which lead to an assay for long-term effects on behaviour. The critical period was transiently extended until 12 h ALH (4 h beyond control critical period closure), and then behaviour was assayed 1.5 days later. Control larvae showed persistent linear locomotion; by contrast, larvae with extended critical periods due to transient knockdown of motor neuron genes showed excessive turning, leading to abnormal spiralling behaviour. Similar but less severe effects were seen in larvae following knockdown of astrocyte genes. It is concluded that a modest extension of the critical period can, in some cases, lead to long-lasting alteration in locomotor behaviour (Ackerman, 2021).
Astrocytes regulate synaptogenesis, synaptic pruning and synaptic efficacy. Within critical periods, astrocyte signalling can tune neuronal plasticity, but its role in critical period closure was not known. This study identified astrocytes as promoting closure of a motor critical period, and defined a series of astrocyte-motor neuron signalling pathways required to close the critical period. Based on previous literature, it is hypothesized that astrocytes could modify critical period closure through regulation of excitatory-inhibitory balance or extracellular matrix composition. Consistent with mammalian studies, it was found that perturbing excitatory-inhibitory balance through astrocyte-specific RNAi of the sole GABA transporter gat was sufficient to extend critical period plasticity. Furthermore, it was found that decreasing signalling from inhibitory extracellular matrix CSPGs through RNAi knockdown of Chondroitin polymerizing factor (Chpf) in astrocytes extended critical period plasticity. Thus, astrocytes use similar strategies in Drosophila and mammals to regulate critical period timing. Unexpectedly, this study also identified astrocyte-derived Neuroligins and their neuronal partner Nrx-1 as instrumental for critical period closure. In sum, this study have identified a key role of astrocytes in closure of a motor critical period required for locomotor function (Ackerman, 2021).
In Drosophila neurons, uniform minus-end-out polarity in dendrites is maintained in part by kinesin-2-mediated steering of growing microtubules at branch points. Apc links the kinesin motor to growing microtubule plus ends and Apc2 recruits Apc to branch points where it functions. Because Apc2 acts to concentrate other steering proteins to branch points, it is of interest to understand how Apc2 is targeted. From an initial broad candidate RNAi screen, Miro (a mitochondrial transport protein), Ank2, Axin, spastin and Rac1 were required to position Apc2-GFP at dendrite branch points. YFP-Ank2-L8, Axin-GFP and mitochondria also localized to branch points suggesting the screen identified relevant proteins. By performing secondary screens, it was found that energy production by mitochondria was key for Apc2-GFP positioning and spastin acted upstream of mitochondria. Ank2 seems to act independently from other players, except its membrane partner, Neuroglian (Nrg). Rac1 likely acts through Arp2/3 to generate branched actin to help recruit Apc2-GFP. Axin can function in a variety of wnt signaling pathways, one of which includes heterotrimeric G proteins and Frizzleds. Knockdown of Gas, Gao, Fz and Fz2, reduced targeting of Apc2 and Axin to branch points. Overall these data suggest that mitochondrial energy production, Nrg/Ank2, branched actin generated by Arp2/3 and Fz/G proteins/Axin function as four modules that control localization of the microtubule regulator Apc2 to its site of action in dendrite branch points (Weiner, 2018).
Abdu, U., Bar, D. and Schupbach, T. (2006). spn-F encodes a novel protein that affects oocyte patterning and bristle morphology in Drosophila. Development 133: 1477-1484. PubMed ID: 16540510
Aboukilila, M. Y., Sami, J. D., Wang, J., England, W., Spitale, R. C. and Cleary, M. D. (2020). Identification of novel regulators of dendrite arborization using cell type-specific RNA metabolic labeling. PLoS One 15(12): e0240386. PubMed ID: 33264304
Ackerman, S. D., Perez-Catalan, N. A., Freeman, M. R. and Doe, C. Q. (2021). Astrocytes close a motor circuit critical period. Nature 592(7854): 414-420. PubMed ID: 33828296
Apitz, H. and Salecker, I. (2018). Spatio-temporal relays control layer identity of direction-selective neuron subtypes in Drosophila. Nat Commun 9(1): 2295. PubMed ID: 29895891
Baek, M. and Mann, R. (2009). Lineage and birth date specify motor neuron targeting and dendritic architecture in adult Drosophila. J. Neurosci. 29: 6904-6916. PubMed ID: 19474317
Baines, R. A. (2003). Postsynaptic protein kinase A reduces neuronal excitability in response to increased synaptic excitation in the Drosophila CNS. J Neurosci 23: 8664-8672. PubMed ID: 14507965
Bagley, J. A., Yan, Z., Zhang, W., Wildonger, J., Jan, L. Y., Jan, Y. N. (2014) Double-bromo and extraterminal (BET) domain proteins regulate dendrite morphology and mechanosensory function. Genes Dev 28: 1940-1956. PubMed ID: 25184680
Bhattacharjee, S., Iyer, E. P. R., Iyer, S. C., Nanda, S., Rubaharan, M., Ascoli, G. A. and Cox, D. N. (2023). The Zinc-BED Transcription Factor Bedwarfed Promotes Proportional Dendritic Growth and Branching through Transcriptional and Translational Regulation in Drosophila. Int J Mol Sci 24(7). PubMed ID: 37047316
Borst, A. (2018). A biophysical mechanism for preferred direction enhancement in fly motion vision. PLoS Comput Biol 14(6): e1006240. PubMed ID: 29897917
Boulanger, A., Thinat, C., Zuchner, S., Fradkin, L. G., Lortat-Jacob, H., Dura, J. M. (2021). Axonal chemokine-like Orion induces astrocyte infiltration and engulfment during mushroom body neuronal remodeling. Nat Commun. 12(1):1849. PubMed ID: 33758182
Brierley, D. J., Blanc, E., Reddy, O. V., VijayRaghavan, K. and Williams, D. W. (2009). Dendritic targeting in the leg neuropil of Drosophila: the role of midline signalling molecules in generating a myotopic map. PLoS Biol. 7(9): e1000199. PubMed ID: 19771147
Bu, S., Yong, W. L., Lim, B. J. W., Kondo, S. and Yu, F. (2021). A systematic analysis of microtubule-destabilizing factors during dendrite pruning in Drosophila. EMBO Rep: e52679. PubMed ID: 34338441
Cachero, S., Ostrovsky, A. D., Yu, J. Y., Dickson, B. J. and Jefferis, G. S. (2010). Sexual dimorphism in the fly brain. Curr Biol 20: 1589-1601. PubMed ID: 20832311
Cha, I. J., Lee, D., Park, S. S., Chung, C. G., Kim, S. Y., Jo, M. G., Kim, S. Y., Lee, B. H., Lee, Y. S. and Lee, S. B. (2020). Ataxin-2 Dysregulation Triggers a Compensatory Fragile X Mental Retardation Protein Decrease in Drosophila C4da Neurons. Mol Cells 43(10): 870-879. PubMed ID: 33115979
Chang, Y. L., King, B., Lin, S. C., Kennison, J. A. and Huang, D. H. (2007). A double-bromodomain protein, FSH-S, activates the homeotic gene ultrabithorax through a critical promoter-proximal region. Mol Cell Biol 27: 5486-5498. PubMed ID: 17526731
Cheerambathur, D. K., Prevo, B., Chow, T. L., Hattersley, N., Wang, S., Zhao, Z., Kim, T., Gerson-Gurwitz, A., Oegema, K., Green, R. and Desai, A. (2019). The kinetochore-microtubule coupling machinery is repurposed in sensory nervous system morphogenesis. Dev Cell 48(6): 864-872 e867. PubMed ID: 30827898 ''
Cheng, G., Chang, J., Gong, H. and Zhou, W. (2023). A distinct Golgi-targeting mechanism of dGM130 in Drosophila neurons. Front Mol Neurosci 16: 1206219. PubMed ID: 37333614
Chiang, A., et al. (2009). Neuronal activity and Wnt signaling act through Gsk3-β to regulate axonal integrity in mature Drosophila olfactory sensory neurons. Development 136: 1273-1282. PubMed ID: 19304886
Chung, C. G., Kwon, M. J., Jeon, K. H., Hyeon, D. Y., Han, M. H., Park, J. H., Cha, I. J., Cho, J. H., Kim, K., Rho, S., Kim, G. R., Jeong, H., Lee, J. W., Kim, T., Kim, K., Kim, K. P., Ehlers, M. D., Hwang, D. and Lee, S. B. (2017). Golgi outpost synthesis impaired by toxic polyglutamine proteins contributes to dendritic pathology in neurons. Cell Rep 20(2): 356-369. PubMed ID: 28700938
Cline, H. and Haas, K. (2008). The regulation of dendritic arbor development and plasticity by glutamatergic synaptic input: a review of the synaptotrophic hypothesis. J. Physiol. 586: 1509-1517. PubMed ID: 18202093
Constantine-Paton, M. and Cline, H. T. (1998). LTP and activity-dependent synaptogenesis: the more alike they are, the more different they become. Curr. Opin. Neurobiol. 8: 139-148. PubMed ID: 9568401
Dacks A. M., et al. (2009). Serotonin modulates olfactory processing in the antennal lobe of Drosophila. J. Neurogenet. 23. 366-377. PubMed ID: 19863268
Das, R., Bhattacharjee, S., Letcher, J. M., Harris, J. M., Nanda, S., Foldi, I., Lottes, E. N., Bobo, H. M., Grantier, B. D., Mihaly, J., Ascoli, G. A. and Cox, D. N. (2021). Formin 3 directs dendritic architecture via microtubule regulation and is required for somatosensory nociceptive behavior. Development 148(16). PubMed ID: 34322714
Datta, S. R., Vasconcelos, M. L., Ruta, V., Luo, S., Wong, A., Demir, E., Flores, J., Balonze, K., Dickson, B. J. and Axel, R. (2008). The Drosophila pheromone cVA activates a sexually dimorphic neural circuit. Nature 452: 473-477. PubMed ID: 18305480
Davis, G. W., DiAntonio, A., Petersen, S. A. and Goodman, C. S. (1998). Postsynaptic PKA controls quantal size and reveals a retrograde signal that regulates presynaptic transmitter release in Drosophila. Neuron 20: 305-315. PubMed ID: 9491991
Denis, G. V., McComb, M. E., Faller, D. V., Sinha, A., Romesser, P. B. and Costello, C. E. (2006). Identification of transcription complexes that contain the double bromodomain protein Brd2 and chromatin remodeling machines. J Proteome Res 5: 502-511. PubMed ID: 16512664
Desai, B. S., Chadha, A. and Cook, B. (2014). The stum gene is essential for mechanical sensing in proprioceptive neurons. Science 343(6176): 1256-9. PubMed ID: 24626929
DeVault, L., Li, T., Izabel, S., Thompson-Peer, K. L., Jan, L. Y. and Jan, Y. N. (2018). Dendrite regeneration of adult Drosophila sensory neurons diminishes with aging and is inhibited by epidermal-derived matrix metalloproteinase 2. Genes Dev 32(5-6): 402-414. PubMed ID: 29563183
Dhawan, S., Myers, P., Bailey, D. M. D., Ostrovsky, A. D., Evers, J. F. and Landgraf, M. (2021). Reactive Oxygen Species Mediate Activity-Regulated Dendritic Plasticity Through NADPH Oxidase and Aquaporin Regulation. Front Cell Neurosci 15: 641802. PubMed ID: 34290589
Doyle, M. and Kiebler, M. A. (2012). A numbers game underpins cytoplasmic mRNA transport. Nat Cell Biol 14: 333-335. PubMed ID: 22469827
Dubin-Bar, D., Bitan, A., Bakhrat, A., Kaiden-Hasson, R., Etzion, S., Shaanan, B. and Abdu, U. (2008). The Drosophila IKK-related kinase (Ik2) and Spindle-F proteins are part of a complex that regulates cytoskeleton organization during oogenesis. BMC Cell Biol 9: 51. PubMed ID: 18796167
Duch, C., Vonhoff, F. and Ryglewski, S. (2008). Dendrite elongation and dendritic branching are affected separately by different forms of intrinsic motoneuron excitability. J Neurophysiol 100: 2525-2536. PubMed ID: 18715893
Dzaki, N., Bu, S., Lau, S. S. Y., Yong, W. L. and Yu, F. (2022). Drosophila GSK3β promotes microtubule disassembly and dendrite pruning in sensory neurons. Development 149(22). PubMed ID: 36264221
Emoto, K., Parrish, J. Z., Jan, L. Y. and Jan, Y. N. (2006). The tumour suppressor Hippo acts with the NDR kinases in dendritic tiling and maintenance. Nature 443(7108): 210-3. PubMed ID: 16906135
Espinosa, J. S., Wheeler, D. G., Tsien, R. W. and Luo, L. (2009). Uncoupling dendrite growth and patterning: single-cell knockout analysis of NMDA receptor 2B. Neuron 62: 205-217. PubMed ID: 19409266
Fendl, S., Vieira, R. M. and Borst, A. (2020). Parallel Visual Conditional protein tagging methods reveal highly specific subcellular distribution of ion channels in motion-sensing neurons. Elife 9. PubMed ID: 33079061
Feng, C., Thyagarajan, P., Shorey, M., Seebold, D. Y., Weiner, A. T., Albertson, R. M., Rao, K. S., Sagasti, A., Goetschius, D. J. and Rolls, M. M. (2019). Patronin-mediated minus end growth is required for dendritic microtubule polarity. J Cell Biol 218(7):2309-2328. PubMed ID: 31076454
Feng, C., Cleary, J. M., Kothe, G. O., Stone, M. C., Weiner, A. T., Hertzler, J. I., Hancock, W. O. and Rolls, M. M. (2021). Trim9 and Klp61F promote polymerization of new dendritic microtubules along parallel microtubules. J Cell Sci. PubMed ID: 33988240
Ferreira, A., Palazzo, R. E. and Rebhun, L. I. (1993). Preferential dendritic localization of pericentriolar material in hippocampal pyramidal neurons in culture. Cell Motil Cytoskeleton 25: 336-344. PubMed ID: 8402954
Ferreira Castro, A., Baltruschat, L., Sturner, T., Bahrami, A., Jedlicka, P., Tavosanis, G. and Cuntz, H. (2020). Achieving functional neuronal dendrite structure through sequential stochastic growth and retraction. Elife 9. PubMed ID: 33241995
Freeman, A., Bowers, M., Mortimer, A. V., Timmerman, C., Roux, S., Ramaswami, M. and Sanyal, S. (2010). A new genetic model of activity-induced Ras signaling dependent pre-synaptic plasticity in Drosophila. Brain Res 1326: 15-29. PubMed ID: 20193670
Grima, J. C., Daigle, J. G., Arbez, N., Cunningham, K. C., Zhang, K., Ochaba, J., Geater, C., Morozko, E., Stocksdale, J., Glatzer, J. C., Pham, J. T., Ahmed, I., Peng, Q., Wadhwa, H., Pletnikova, O., Troncoso, J. C., Duan, W., Snyder, S. H., Ranum, L. P. W., Thompson, L. M., Lloyd, T. E., Ross, C. A. and Rothstein, J. D. (2017). Mutant Huntingtin Disrupts the Nuclear Pore Complex. Neuron 94(1): 93-107 e106. PubMed ID: 28384479
Golden, S. A., Christoffel, D. J., Heshmati, M., Hodes, G. E., Magida, J., Davis, K., Cahill, M. E., Dias, C., Ribeiro, E., Ables, J. L., Kennedy, P. J., Robison, A. J., Gonzalez-Maeso, J., Neve, R. L., Turecki, G., Ghose, S., Tamminga, C. A. and Russo, S. J. (2013). Epigenetic regulation of RAC1 induces synaptic remodeling in stress disorders and depression. Nat Med 19: 337-344. PubMed ID: 23416703
Gorczyca, D. A., Younger, S., Meltzer, S., Kim, S. E., Cheng, L., Song, W., Lee, H. Y., Jan, L. Y. and Jan, Y. N. (2014). Identification of Ppk26, a DEG/ENaC channel functioning with Ppk1 in a mutually dependent manner to guide locomotion behavior in Drosophila. Cell Rep 9: 1446-1458. PubMed ID: 25456135
Guzman-Palma, P., Contreras, E. G., Mora, N., Smith, M., Gonzalez-Ramirez, M. C., Campusano, J. M., Sierralta, J., Hassan, B. A. and Oliva, C. (2021). Slit/Robo Signaling Regulates Multiple Stages of the Development of the Drosophila Motion Detection System. Front Cell Dev Biol 9: 612645. PubMed ID: 33968921
Ha, A.1, Polyanovsky, A. and Avidor-Reiss, T. (2015). Drosophila Hook-Related Protein (Girdin) is essential for sensory dendrite formation. Genetics [Epub ahead of print]. PubMed ID: 26058848
Haga, S., Hattori, T., Sato, T., Sato, K., Matsuda, S., Kobayakawa, R., Sakano, H., Yoshihara, Y., Kikusui, T. and Touhara, K. (2010). The male mouse pheromone ESP1 enhances female sexual receptive behaviour through a specific vomeronasal receptor. Nature 466: 118-122. PubMed ID: 20596023
Han, C., Song, Y., Xiao, H., Wang, D., Franc, N. C., Jan, L. Y. and Jan, Y. N. (2014). Epidermal cells are the primary phagocytes in the fragmentation and clearance of degenerating dendrites in Drosophila. Neuron 81(3):544-60. PubMed ID: 24412417
Han, M. H., Kwon, M. J., Ko, B. S., Hyeon, D. Y., Lee, D., Kim, H. J., Hwang, D. and Lee, S. B. (2020). NF-kappaB disinhibition contributes to dendrite defects in fly models of neurodegenerative diseases. J Cell Biol 219(12). PubMed ID: 33090185
Hanus, C., Kochen, L., Tom Dieck, S., Racine, V., Sibarita, J. B., Schuman, E. M. and Ehlers, M. D. (2014). Synaptic control of secretory trafficking in dendrites. Cell Rep 7: 1771-1778. PubMed ID: 24931613
Hartwig, C. L., Worrell, J., Levine, R. B., Ramaswami, M. and Sanyal, S. (2008). Normal dendrite growth in Drosophila motor neurons requires the AP-1 transcription factor. Dev Neurobiol 68: 1225-1242. PubMed ID: 18548486
Hertzler, J. I., Simonovitch, S. I., Albertson, R. M., Weiner, A. T., Nye, D. M. R. and Rolls, M. M. (2020). Kinetochore proteins suppress neuronal microtubule dynamics and promote dendrite regeneration. Mol Biol Cell: mbcE20040237T. PubMed ID: 32673176
Herzmann, S., Krumkamp, R., Rode, S., Kintrup, C. and Rumpf, S. (2017). PAR-1 promotes microtubule breakdown during dendrite pruning in Drosophila. EMBO J 36(13): 1981-1991. PubMed ID: 28554895
Herzmann, S., Gotzelmann, I., Reekers, L. F. and Rumpf, S. (2018). Spatial regulation of microtubule disruption during dendrite pruning in Drosophila. Development [Epub ahead of print]. PubMed ID: 29712642
Kanaoka, Y., Onodera, K., Watanabe, K., Hayashi, Y., Usui, T., Uemura, T. and Hattori, Y. (2023). Inter-organ Wingless/Ror/Akt signaling regulates nutrient-dependent hyperarborization of somatosensory neurons. Elife 12. PubMed ID: 36647607
Ji, H., Sapar, M. L., Sarkar, A., Wang, B. and Han, C. (2022). Phagocytosis and self-destruction break down dendrites of Drosophila sensory neurons at distinct steps of Wallerian degeneration. Proc Natl Acad Sci U S A 119(4). PubMed ID: 35058357
Ji, H., Wang, B., Shen, Y., Labib, D., Lei, J., Chen, X., Sapar, M., Boulanger, A., Dura, J. M. and Han, C. (2023). The Drosophila chemokine-like Orion bridges phosphatidylserine and Draper in phagocytosis of neurons. Proc Natl Acad Sci U S A 120(24): e2303392120. PubMed ID: 37276397
Kamemura, K., Chen, C. A., Okumura, M., Miura, M. and Chihara, T. (2021). Amyotrophic lateral sclerosis-associated VAP33 is required for maintaining neuronal dendrite morphology and organelle distribution in Drosophila. Genes Cells. PubMed ID: 33548103
Kamemura, K., Moriya, H., Ukita, Y., Okumura, M., Miura, M. and Chihara, T. (2022). Endoplasmic reticulum proteins Meigo and Gp93 govern dendrite targeting by regulating Toll-6 localization. Dev Biol 484: 30-39. PubMed ID: 35134382
Kondo, S., Takahashi, T., Yamagata, N., Imanishi, Y., Katow, H., Hiramatsu, S., Lynn, K., Abe, A., Kumaraswamy, A. and Tanimoto, H. (2020). Neurochemical organization of the Drosophila brain visualized by endogenously tagged neurotransmitter receptors. Cell Rep 30(1): 284-297 e285. PubMed ID: 31914394
Kramer, R., Wolterhoff, N., Galic, M. and Rumpf, S. (2023). Developmental pruning of sensory neurites by mechanical tearing in Drosophila. J Cell Biol 222(3). PubMed ID: 36648440
Liang, X., Kokes, M., Fetter, R. D., Sallee, M. D., Moore, A. W., Feldman, J. L. and Shen, K. (2020). Growth cone-localized microtubule organizing center establishes microtubule orientation in dendrites. Elife 9. PubMed ID: 32657271
Liao, M., Liang, X. and Howard, J. (2021). The narrowing of dendrite branches across nodes follows a well-defined scaling law. Proc Natl Acad Sci U S A 118(27). PubMed ID: 34215693
Lin, C. H., Senapati, B., Chen, W. J., Bansal, S. and Lin, S. (2022). Semaphorin 1a-mediated dendritic wiring of the Drosophila mushroom body extrinsic neurons. Proc Natl Acad Sci U S A 119(12): e2111283119. PubMed ID: 35286204
Lottes, E. N., Ciger, F. H., Bhattacharjee, S., Timmins-Wilde, E. A., Tete, B., Tran, T., Matta, J., Patel, A. A. and Cox, D. N. (2023). CCT and Cullin1 regulate the TORC1 pathway to promote dendritic arborization in health and disease. bioRxiv. PubMed ID: 37577581
Marzano, M., Herzmann, S., Elsbroek, L., Sanal, N., Tarbashevich, K., Raz, E., Krahn, M. P. and Rumpf, S. (2021). AMPK adapts metabolism to developmental energy requirement during dendrite pruning in Drosophila. Cell Rep 37(7): 110024. PubMed ID: 34788610
Nanda, S., Bhattacharjee, S., Cox, D. N. and Ascoli, G. A. (2023). Local Microtubule and F-Actin Distributions Fully Constrain the Spatial Geometry of Drosophila Sensory Dendritic Arbors. Int J Mol Sci 24(7). PubMed ID: 37047715
Nye, D. M. R., Albertson, R. M., Weiner, A. T., Hertzler, J. I., Shorey, M., Goberdhan, D. C. I., Wilson, C., Janes, K. A. and Rolls, M. M. (2020). The receptor tyrosine kinase Ror is required for dendrite regeneration in Drosophila neurons. PLoS Biol 18(3): e3000657. PubMed ID: 32163406
O'Brien, C. E., Younger, S. H., Jan, L. Y. and Jan, Y. N. (2023). The GARP complex prevents sterol accumulation at the trans-Golgi network during dendrite remodeling. J Cell Biol 222(1). PubMed ID: 36239632
Ravenscroft, T. A., Jacobs, A., Gu, M., Eberl, D. F. and Bellen, H. J. (2023). The Voltage-Gated Sodium Channel in Drosophila, Para, Localizes to Dendrites As Well As Axons in Mechanosensitive Chordotonal Neurons. eNeuro 10(6). PubMed ID: 37328295
Rui, M., Kong, W., Wang, W., Zheng, T., Wang, S., Xie, W. (2023). Droj2 Facilitates Somatosensory Neurite Sculpting via GTP-Binding Protein Arf102F in Drosophila. Int J Mol Sci, 24(17) PubMed ID: 37686022
Sapar, M. L., Ji, H., Wang, B., Poe, A. R., Dubey, K., Ren, X., Ni, J. Q. and Han, C. (2018). Phosphatidylserine Externalization Results from and Causes Neurite Degeneration in Drosophila. Cell Rep 24(9): 2273-2286. PubMed ID: 30157423
Shrestha, B. R., Burgos, A. and Grueber, W. B. (2021). The Immunoglobulin Superfamily Member Basigin Is Required for Complex Dendrite Formation in Drosophila. Front Cell Neurosci 15: 739741. PubMed ID: 34803611
Song, W., Ressl, S. and Tracey, W. D. (2020). Loss of Pseudouridine Synthases in the RluA Family Causes Hypersensitive Nociception in Drosophila. G3 (Bethesda). PubMed ID: 33028630
Sturner, T., Ferreira Castro, A., Philipps, M., Cuntz, H. and Tavosanis, G. (2022). The branching code: A model of actin-driven dendrite arborization. Cell Rep 39(4): 110746. PubMed ID: 35476974'''
Strother, J. A., Wu, S. T., Wong, A. M., Nern, A., Rogers, E. M., Le, J. Q., Rubin, G. M. and Reiser, M. B. (2017). The emergence of directional selectivity in the visual motion pathway of Drosophila. Neuron 94(1): 168-182 e110. PubMed ID: 28384470
Swope, R. D., Hertzler, J. I., Stone, M. C., Kothe, G. O. and Rolls, M. M. (2022). The exocyst complex is required for developmental and regenerative neurite growth in vivo. Dev Biol 492: 1-13. PubMed ID: 36162553
Tadros, W., Xu, S., Akin, O., Yi, C. H., Shin, G. J., Millard, S. S. and Zipursky, S. L. (2016). Dscam proteins direct dendritic targeting through adhesion. Neuron 89: 480-493. PubMed ID: 26844831
Tenenbaum, C. M., Misra, M., Alizzi, R. A. and Gavis, E. R. (2017). Enclosure of dendrites by epidermal cells restricts branching and permits coordinated development of spatially overlapping sensory neurons. Cell Rep 20(13): 3043-3056. PubMed ID: 28954223
Terada, S., Matsubara, D., Onodera, K., Matsuzaki, M., Uemura, T. and Usui, T. (2016).. Neuronal processing of noxious thermal stimuli mediated by dendritic Ca influx in somatosensory neurons. Elife 5: e12959. PubMed ID: 26880554
Thompson-Peer, K. L., DeVault, L., Li, T., Jan, L. Y. and Jan, Y. N. (2016). In vivo dendrite regeneration after injury is different from dendrite development. Genes Dev 30: 1776-1789. PubMed ID: 27542831
Ting, C. Y., McQueen, P. G., Pandya, N., Lin, T. Y., Yang, M., Reddy, O. V., O'Connor, M. B., McAuliffe, M. and Lee, C. H. (2014). Photoreceptor-derived activin promotes dendritic termination and restricts the receptive fields of first-order interneurons in Drosophila. Neuron 81(4): 830-846. PubMed ID: 24462039
Treier, M. Seufert, W. and Jentsch, S. (1992). Drosophila UbcD1 encodes a highly conserved ubiquitin-conjugating enzyme involved in selective protein degradation. EMBO J. 11: 367-372. PubMed ID: 1310935
Tripodi, M., Evers, J. F., Mauss, A., Bate, M. and Landgraf, M. (2008). Structural homeostasis: Compensatory adjustments of dendritic arbor geometry in response to variations of synaptic input. PLoS Biol. 6(10): e260. PubMed ID: 18959482
Tsubouchi, A., Caldwell, J. C. and Tracey, W. D. (2012). Dendritic filopodia, Ripped Pocket, NOMPC, and NMDARs contribute to the sense of touch in Drosophila larvae. Curr Biol 22: 2124-2134. PubMed ID: 23103192
Tsuyama, T., Tsubouchi, A., Usui, T., Imamura, H. and Uemura, T. (2017). Mitochondrial dysfunction induces dendritic loss via eIF2α phosphorylation. J Cell Biol 216: 815-834. PubMed ID: 28209644
Umehara, T., Nakamura, Y., Jang, M. K., Nakano, K., Tanaka, A., Ozato, K., Padmanabhan, B. and Yokoyama, S. (2010a). Structural basis for acetylated histone H4 recognition by the human BRD2 bromodomain. J Biol Chem 285: 7610-7618. PubMed ID: 20048151
Umehara, T., Nakamura, Y., Wakamori, M., Ozato, K., Yokoyama, S. and Padmanabhan, B. (2010b). Structural implications for K5/K12-di-acetylated histone H4 recognition by the second bromodomain of BRD2. FEBS Lett 584: 3901-3908. PubMed ID: 20709061
Vonhoff, F., Kuehn, C., Blumenstock, S., Sanyal, S. and Duch, C. (2013). Temporal coherency between receptor expression, neural activity and AP-1-dependent transcription regulates Drosophila motoneuron dendrite development. Development 140: 606-616. PubMed ID: 23293292
Wang, Q., Wang, Y. and Yu, F. (2018). Yif1 associates with Yip1 on Golgi and regulates dendrite pruning in sensory neurons during Drosophila metamorphosis. Development. PubMed ID: 29769219
Wang, S. L., Hawkins, C. J., Yoo, S. J., Muller, H. A. and Hay, B. A. (1999). The Drosophila caspase inhibitor DIAP1 is essential for cell survival and is negatively regulated by HID. Cell 98: 453-463. PubMed ID: 10481910
Wang, X., Zhang, M. W., Kim, J. H., Macara, A. M., Sterne, G., Yang, T. and Ye, B. (2015). The Kruppel-like factor Dar1 determines multipolar neuron morphology. J Neurosci 35: 14251-14259. PubMed ID: 26490864
Wang, Y., Zhang, H., Shi, M., Liou, Y.C., Lu, L. and Yu, F. (2017). Sec71 functions as a GEF for the small GTPase Arf1 to govern dendrite pruning of Drosophila sensory neurons. Development 144(10):1851-1862. PubMed ID: 28420712
Wang, Y. H., Ding, Z. Y., Cheng, Y. J., Chien, C. T. and Huang, M. L. (2020). An Efficient Screen for Cell-Intrinsic Factors Identifies the Chaperonin CCT and Multiple Conserved Mechanisms as Mediating Dendrite Morphogenesis. Front Cell Neurosci 14: 577315. PubMed ID: 33100975
Watanabe, K., Furumizo, Y., Usui, T., Hattori, Y. and Uemura, T. (2016). Nutrient-dependent increased dendritic arborization of somatosensory neurons. Genes Cells [Epub ahead of print]. PubMed ID: 27868313
Weiner, A. T., Lanz, M. C., Goetschius, D. J., Hancock, W. O. and Rolls, M. M. (2016). Kinesin-2 and Apc function at dendrite branch points to resolve microtubule collisions. Cytoskeleton (Hoboken) 73: 35-44. PubMed ID: 26785384
Weiner, A. T., Seebold, D. Y., Michael, N. L., Guignet, M., Feng, C., Follick, B., Yusko, B. A., Wasilko, N. P., Torres-Gutierrez, P. and Rolls, M. M. (2018). Identification of proteins required for precise positioning of Apc2 in dendrites. G3 (Bethesda) 8(5):1841-1853. PubMed ID: 29602811
Weiner, A. T., Seebold, D. Y., Torres-Gutierrez, P., Folker, C., Swope, R. D., Kothe, G. O., Stoltz, J. G., Zalenski, M. K., Kozlowski, C., Barbera, D. J., Patel, M. A., Thyagarajan, P., Shorey, M., Nye, D. M. R., Keegan, M., Behari, K., Song, S., Axelrod, J. D. and Rolls, M. M. (2020). Endosomal Wnt signaling proteins control microtubule nucleation in dendrites. PLoS Biol 18(3): e3000647. PubMed ID: 32163403
Williams, D. W. and Truman, J. W. (2005). Cellular mechanisms of dendrite pruning in Drosophila: insights from in vivo time-lapse of remodeling dendritic arborizing sensory neurons. Development 132: 3631-3642. PubMed ID: 16033801
Wong, K. K. L., Li, T., Fu, T. M., Liu, G., Lyu, C., Kohani, S., Xie, Q., Luginbuhl, D. J., Upadhyayula, S., Betzig, E. and Luo, L. (2023). Origin of wiring specificity in an olfactory map revealed by neuron type-specific, time-lapse imaging of dendrite targeting. Elife 12. PubMed ID: 36975203
Wu, Y., Helt, J. C., Wexler, E., Petrova, I. M., Noordermeer, J. N., Fradkin, L. G. and Hing, H. (2014). Wnt5 and drl/ryk gradients pattern the Drosophila olfactory dendritic map. J Neurosci 34: 14961-14972. PubMed ID: 25378162
Xu, X., Brechbiel, J. L. and Gavis, E. R. (2013). Dynein-Dependent Transport of nanos RNA in Drosophila Sensory Neurons Requires Rumpelstiltskin and the Germ Plasm Organizer Oskar. J Neurosci 33: 14791-14800. PubMed ID: 24027279
Yalgin, C., Ebrahimi, S., Delandre, C., Yoong, L. F., Akimoto, S., Tran, H., Amikura, R., Spokony, R., Torben-Nielsen, B., White, K. P. and Moore, A. W. (2015). Centrosomin represses dendrite branching by orienting microtubule nucleation. Nat Neurosci 18(10):1437-45. PubMed ID: 26322925
Yang, L., Garbe, D. S. and Bashaw, G. J. (2009). A frazzled/DCC-dependent transcriptional switch regulates midline axon guidance. Science 324: 944-947. PubMed ID: 19325078
Yang, S. Z. and Wildonger, J. (2020). Golgi Outposts Locally Regulate Microtubule Orientation in Neurons but Are Not Required for the Overall Polarity of the Dendritic Cytoskeleton. Genetics. PubMed ID: 32265236
Yang, W. K., et al. (2011). Nak regulates localization of clathrin sites in higher-order dendrites to promote local dendrite growth. Neuron 72(2): 285-99. PubMed ID: 22017988
Ye, B., Zhang, Y., Song, W., Younger, S. H., Jan, L. Y. and Jan, Y. N. (2007). Growing dendrites and axons differ in their reliance on the secretory pathway. Cell 130: 717-729. PubMed ID: 17719548
Ye, B., Kim, J. H., Yang, L., McLachlan, I., Younger, S., Jan, L. Y. and Jan, Y. N. (2011). Differential regulation of dendritic and axonal development by the novel Kruppel-like factor Dar1. J Neurosci 31: 3309-3319. PubMed ID: 21368042
Yin, J., Gibbs, M., Long, C., Rosenthal, J., Kim, H. S., Kim, A., Sheng, C., Ding, P., Javed, U. and Yuan, Q. (2018). Transcriptional regulation of lipophorin receptors supports neuronal adaptation to chronic elevations of activity. Cell Rep 25(5): 1181-1192.e1184. PubMed ID: 30380410
Yin, J., Spillman, E., Cheng, E. S., Short, J., Chen, Y., Lei, J., Gibbs, M., Rosenthal, J. S., Sheng, C., Chen, Y. X., Veerasammy, K., Choetso, T., Abzalimov, R., Wang, B., Han, C., He, Y. and Yuan, Q. (2021). Brain-specific lipoprotein receptors interact with astrocyte derived apolipoprotein and mediate neuron-glia lipid shuttling. Nat Commun 12(1): 2408. PubMed ID: 33893307
Yoong, L. F., Lim, H. K., Tran, H., Lackner, S., Zheng, Z., Hong, P. and Moore, A. W. (2020). Atypical myosin tunes dendrite arbor subdivision. Neuron. PubMed ID: 32155441
Yu, S. Y., Yoo, S. J., Yang, L., Zapata, C., Srinivasan, A., Hay, B.A. and Baker, N. E. (2002). A pathway of signals regulating effector and initiator caspases in the developing Drosophila eye. Development 129: 3269-3278. PubMed ID: 12070100
Zhang, H., Wang, Y., Wong, J. J., Lim, K. L., Liou, Y. C., Wang, H., Yu, F. (2014). Endocytic pathways downregulate the L1-type cell adhesion molecule Neuroglian to promote dendrite pruning in Drosophila. Dev Cell 30: 463-478. PubMed ID: 25158855
Zhang, Y. V., Aikin, T. J., Li, Z. and Montell, C. (2016). The Basis of Food Texture Sensation in Drosophila. Neuron 91(4): 863-877. PubMed ID: 27478019
Zhang, T., Cheng, D., Wu, C., Wang, X., Ke, Q., Lou, H., Zhu, L., Wang, X. D., Duan, S. and Liu, Y. J. (2020). Lysosomal hydrolase Cathepsin D non-proteolytically modulates dendritic morphology in Drosophila. Neurosci Bull. PubMed ID: 32170568
Zhao, G., Oztan, A., Ye, Y. and Schwarz, T. L. (2019). Kinetochore proteins have a post-mitotic function in neurodevelopment. Dev Cell. PubMed ID: 30827899
Zheng, Y., Wildonger, J., Ye, B., Zhang, Y., Kita, A., Younger, S. H., Zimmerman, S., Jan, L. Y. and Jan, Y. N. (2008). Dynein is required for polarized dendritic transport and uniform microtubule orientation in axons. Nat Cell Biol 10: 1172-1180. PubMed ID: 18758451
Zhou, W., Chang, J., Wang, X., Savelieff, M. G., Zhao, Y., Ke, S. and Ye, B. (2014). GM130 is required for compartmental organization of dendritic Golgi outposts. Curr Biol 24(11): 1227-33. PubMed ID: 24835455
Zwart, M. F., Randlett, O., Evers, J. F. and Landgraf, M. (2013). Dendritic growth gated by a steroid hormone receptor underlies increases in activity in the developing Drosophila locomotor system. Proc Natl Acad Sci U S A. PubMed ID: 24043825
date revised: 25 August 2023
Genes involved in organ development
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
{kind=link}
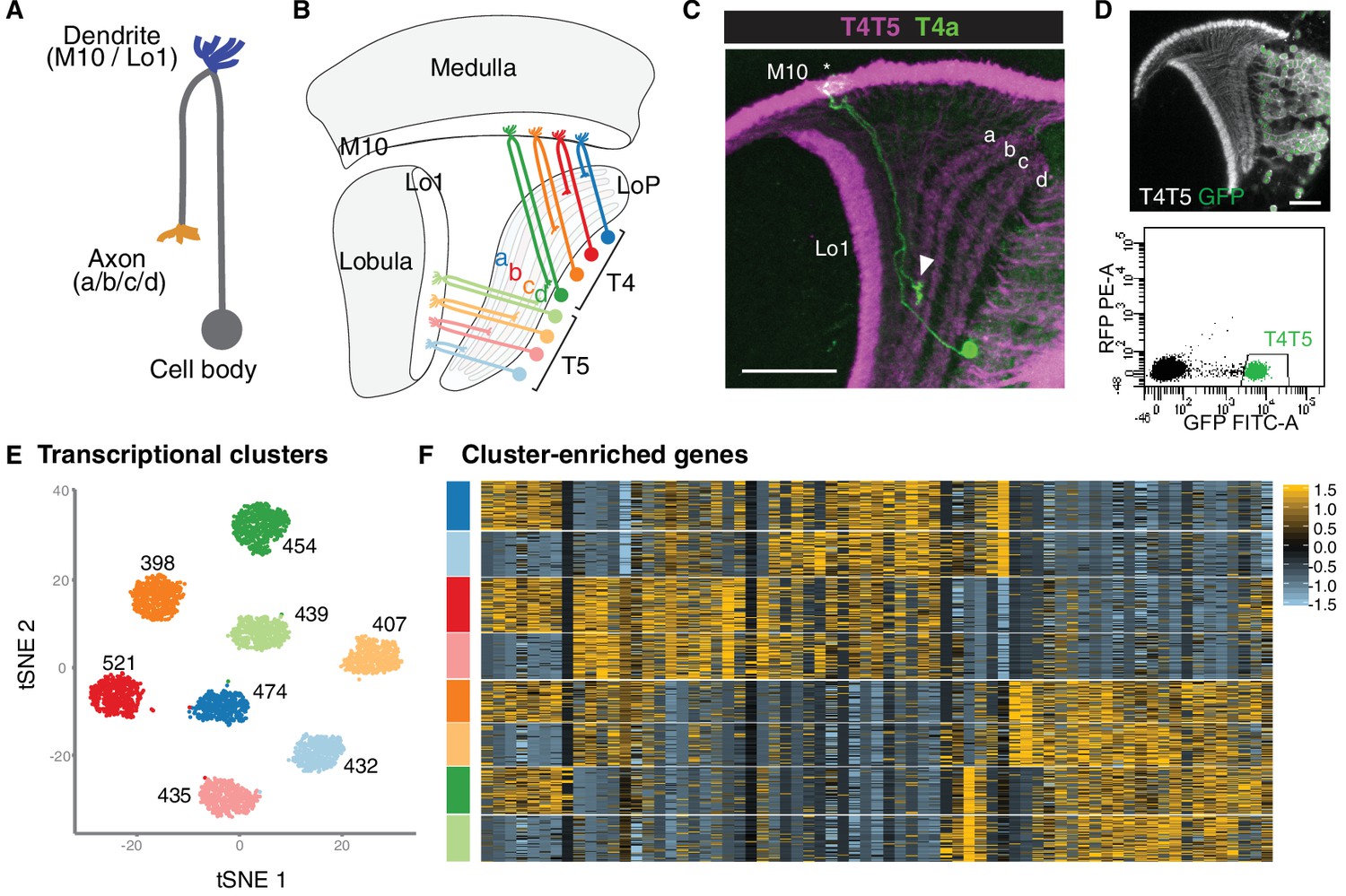{kind=link}