The Interactive Fly
Genes involved in tissue and organ development
Global inhibition in head-direction neural circuits: a systematic comparison between connectome-based spiking neural circuit models
Drosophila neuroblasts are an excellent model for investigating how neuronal diversity is generated. Most brain neuroblasts generate a series of ganglion mother cells (GMCs) that each make two neurons (type I lineage), but sixteen brain neuroblasts generate a series of intermediate neural progenitors (INPs) that each produce 4-6 GMCs and 8-12 neurons (type II lineage). Thus, type II lineages are similar to primate cortical lineages, and may serve as models for understanding cortical expansion. Yet the origin of type II neuroblasts remains mysterious: do they form in the embryo or larva? If they form in the embryo, do their progeny populate the adult central complex, as do the larval type II neuroblast progeny? This study presents molecular and clonal data showing that all type II neuroblasts form in the embryo, produce INPs, and express known temporal transcription factors. Embryonic type II neuroblasts and INPs undergo quiescence, and produce embryonic-born progeny that contribute to the adult central complex. These results provide a foundation for investigating the development of the central complex, and tools for characterizing early-born neurons in central complex function (Walsh, 2017).
It has been difficult to link embryonic neuroblasts to their larval counterparts in the brain and thoracic segments owing to the period of quiescence at the embryo-larval transition, and owing to dramatic morphological changes of the CNS that occur at late embryogenesis. Recent work has revealed the embryonic origin of some larval neuroblasts: the four mushroom body neuroblasts in the central brain and about 20 neuroblasts in thoracic segments. This study used molecular markers and clonal analysis to identify all eight known type II neuroblasts in each brain lobe and show they all form during embryogenesis, perhaps the last-born central brain neuroblasts. It was not possible to identify each neuroblast individually, however, owing to their tight clustering, movements of the brain lobes, and the lack of markers for specific type II neuroblasts (Walsh, 2017).
The single previously reported embryonic type II neuroblast formed from PntP1+ neuroectodermal cells with apical constrictions called a placode. This study has not investigated this neuroectodermal origin of type II neuroblasts in much detail, but this study also observe multiple type II neuroblasts developing from PntP1+ neuroectoderm. In the future, it would be interesting to determine whether all type II neuroblasts arise from PntP1+ neuroectoderm or from neuroectodermal placodes. Interestingly, one distinguishing molecular attribute of type II neuroblasts is PntP1, which is not detected in type I neuroblasts. Thus, a candidate for distinguishing type I/type II neuroblast identity is EGF signaling, which can be detected in the three head placodes and is required for PntP1 expression. Clearly, there are more PntP1+ neuroectodermal cells than there are type II neuroblasts, and expression of an EGF negative regulator such as Argos might be necessary to divert some of these neuroectodermal cells away from type II neuroblast specification. The earliest steps of type II neuroblast formation represent an interesting spatial patterning question for future studies (Walsh, 2017).
Now that the embryonic type II neuroblasts have been identified, it is worth considering whether there are differences between embryonic and larval type II neuroblasts or their INP progeny. To date, molecular markers do not reveal any differences between embryonic and larval type II neuroblasts, with the exception that embryonic neuroblasts transiently express the temporal transcription factor Pdm. Interestingly, type I embryonic neuroblasts require Cas to close the Pdm expression window, whereas this study finds that cas mutants do not exhibit extension of the Pdm expression window in the earliest-born type II neuroblast or de novo expression of Pdm in the later-forming neuroblasts. Are there differences between embryonic and larval INPs? Larval INPs mature over a period of 6 h and then divide four to six times with a cell cycle of about 1 h. In contrast, embryonic INPs might have a more rapid maturation because Elav+ neurons are seen within 9D11+ INP lineages by stage 14, just 3 h after the first type II neuroblast forms. This study found that INPs undergo quiescence at the embryo-larval transition, as shown by the pools of INPs at stage 16 that do not stain for the mitotic marker pH3. The fate of these quiescent INPs - whether they resume proliferation, differentiate or die - remains to be determined (Walsh, 2017).
Neuroblasts in the embryonic ventral nerve cord use the temporal transcription factor cascade Hb>Krüuppel>Pdm>Cas>Grh to generate neural diversity. This study shows that the type II neuroblasts are among the last neuroblasts to form in the embryonic brain and that they sequentially express only the late temporal transcription factors Pdm (in the earliest-forming neuroblast) followed by Cas and Grh (in all eight type II neuroblasts). It is unknown why most type II neuroblasts skip the early Hb>Krüppel>Pdm temporal transcription factors; perhaps it is due to their late time of formation, although several earlier-forming thoracic neuroblasts also skip Hb (NB3-3), Hb>Krüppel (NB5-5), or Hb>Krüppel->Pdm (NB6-1). This is another interesting spatial patterning question for the future. Furthermore, misexpression of the early factors (Hb and Krüppel) would be unlikely to affect the progeny produced by type II NBs during embryogenesis, as the competence window for Hb (i.e., the stage at which neuroblasts are responsive to Hb expression) closes with the loss of Dan/Danr expression in all neuroblasts at stage 12. Thus, most embryonic type II neuroblasts form after closing of the Hb competence window and would probably be unresponsive (Walsh, 2017).
Type I neuroblasts show persistent expression of the temporal transcription factors within neurons born during each window of expression (i.e. a Hb+ neuroblast divides to produce a Hb+ GMC which makes Hb+ neurons). In contrast, this study found that type II lineages do not show persistent Cas or Grh expression in INPs born during each expression window, but do contain some Cas+ neurons. Both Cas and Grh transcription factors can be seen in INPs immediately adjacent to the parental neuroblast, consistent with transient perdurance from the parental neuroblast, but they are typically lacking in INPs more distant. The function of Pdm, Cas and Grh in embryonic type II neuroblasts awaits identification of specific markers for neural progeny born during each expression window (Walsh, 2017).
During larval neurogenesis, virtually all INPs sequentially express the temporal transcription factors Dichaete->Grh->Ey. In contrast, embryonic INPs express only Dichaete. These data, together with the short time frame of embryogenesis, suggest that INP quiescence occurs during the Dichaete window, preventing expression of the later Grh->Ey cascade. Interestingly, INPs in the posterior cluster (presumptive DL1 and DL2 type II neuroblast progeny) completely lack Dichaete; this is similar to the DL1 and DL2 larval lineages, which also do not express Dichaete. It is possible that DL1/DL2 neuroblasts make INPs that generate identical progeny (and thus do not require an INP temporal cascade), or perhaps these two neuroblasts use a novel temporal cascade in both embryonic and larval stages (Walsh, 2017).
Larval type II neuroblasts produce many intrinsic neurons of the adult central complex. This study shows that embryonic INPs also produce neurons that contribute to the adult central complex. The data show ~54 neurons (64 minus 10 due to 'leaky' expression) born from embryonic-born INPs survive to adulthood and innervate the central complex. It is likely that this is an underestimate, however, because (1) 9D11-gal4 expression is lacking from a few INPs in the embryonic brain and (2) the time to achieve sufficient FLP protein levels to achieve immortalization could miss the earliest born neurons. The variation in immortalization of the widefield ellipsoid body neuron might represent a neuron born early in the type II lineages, thus unlabeled in a subset of embryos. Additionally, some embryonic-born neurons might perform important functions in the larval/pupal stages but die prior to eclosion (Walsh, 2017).
Further studies will be required to understand the function of neurons born from embryonic type II lineages. It remains to be experimentally determined whether some or all embryonic progeny of type II neuroblasts (1) remain functionally immature in both the larval and adult brain, but serve as pioneer neurons to guide larval-born neurons to establish the central complex, (2) remain functionally immature in the larval brain, but differentiate and function in the adult central complex, or (3) differentiate and perform a function in both the larval and adult CNS. It will be informative to ablate embryonic-born neurons selectively and determine the effect on the assembly of the larval or adult central complex, and their role in generating larval and adult behavior (Walsh, 2017).
The anterior visual pathway (AVP) conducts visual information from the medulla of the optic lobe via the anterior optic tubercle (AOTU) and bulb (BU) to the ellipsoid body (EB) of the central complex. This paper analyzes the formation of the AVP from early larval to adult stages. The immature fiber tracts of the AVP, formed by secondary neurons of lineages DALcl1/2 and DALv2, assemble into structurally distinct primordia of the AOTU, BU, and EB within the late larval brain. During the early pupal period (P6-P48) these primordia grow in size and differentiate into the definitive subcompartments of the AOTU, BU, and EB. The primordium of the EB has a complex composition. DALv2 neurons form the anterior EB primordium, which starts out as a bilateral structure, then crosses the midline between P6 and P12, and subsequently bends to adopt the ring shape of the mature EB. Columnar neurons of the central complex, generated by the type II lineages DM1-4, form the posterior EB primordium. Starting out as an integral part of the fan-shaped body (FB) primordium, the posterior EB primordium moves forward and merges with the anterior EB primordium. This paper documents the extension of neuropil glia around the nascent EB and BU and analyzes the relationship of primary and secondary neurons of the AVP lineages (Lovick, 2017).
The mushroom body (MB) is a well-characterized associative memory structure within the Drosophila brain. Analyzing MB connectivity using multiple approaches is critical for understanding the functional implications of this structure. Using the genetic anterograde transsynaptic tracing tool, trans-Tango, this study identified divergent projections across the brain and convergent downstream targets of the MB output neurons (MBONs). This analysis revealed at least three separate targets that receive convergent input from MBONs: other MBONs, the fan-shaped body (FSB), and the lateral accessory lobe (LAL). A multilayer circuit is described, both anatomically and functionally, in which inhibitory and excitatory MBONs converge on the same genetic subset of FSB and LAL neurons. This circuit architecture enables the brain to update and integrate information with previous experience before executing appropriate behavioral responses. This use of trans-Tango provides a genetically accessible anatomical framework for investigating the functional relevance of components within these complex and interconnected circuits (Scaplen, 2021).
The MB is a high-level integration center in the Drosophila brain with an established role in learning and memory. The iterative nature of converging and diverging MB neural circuits provides an excellent example of the anatomical framework necessary for complex information processing. For instance, on a rapid timescale, interactions between MB compartments could generate different output patterns to drive behavior, whereas on a slower timescale, interactions between MB compartments could reevaluate memories of a context (Scaplen, 2021).
This study sought to map the projections from the MB using the genetic anterograde transsynaptic technique, trans-Tango. The connectivity of MBONs is reported across multiple subjects in both males and females and the variability in connectivity is highlighted that potentially exists across animals. This study complements the ongoing efforts of EM reconstruction of a whole brain of a single female fruit fly and confirms previous anatomical predictions. Although the complete EM dataset of an adult fly brain has been an invaluable resource that significantly accelerated the mapping of the neural circuits underlying innate and learned behaviors, the massive undertaking of acquiring a full EM dataset renders it impractical to perform for multiple individuals. Thus, trans-Tango, expands the value of the EM reconstruction data by examining circuit connectivity across multiple individuals. Further, trans-Tango can be readily adapted to functional studies in which the activity of the postsynaptic neurons is altered by expressing optogenetic/thermogenetic effectors or monitored by expressing genetically encoded sensors. Tracing studies reported in this study serve as the foundation for these future experiments (Scaplen, 2021).
These studies reveal that the MB circuits are highly interconnected with multiple regions of converging projections both within and downstream of the MB. These experiments also show diverging projections in the downstream postsynaptic targets. A multilayer circuit is identified that includes GABAergic and cholinergic MBONs that converge on the same subset of FSB and LAL neurons. This circuit architecture allows for rapid updating of the online processing of sensory information before executing behavior. Further, this circuit organization is likely a conserved motif among insects (Scaplen, 2021).
Successive levels of convergence and divergence across the brain permit functional flexibility. Like the mushroom body, cerebellar circuits in mammals exhibit large divergence in connectivity, and this can support diverse types of synaptic plasticity. Previous neuroanatomical work in insects described divergent afferent and efferent MB neurons, although the extent of this divergence was unknown. The data revealed varying levels of divergence of postsynaptic connections of MBONs across the brain. Every one of the analyzed MBONs had postsynaptic partners projecting to multiple brain regions. Further, nearly the entire superior protocerebrum as well as portions of the inferior protocerebrum received input from at least one MBON, providing opportunities for comprehensive integration of signals from the MBON network (Scaplen, 2021).
Multiple feedforward and feedback circuits exist within the MB. The current data revealed at least two MBONs that receive convergent input from multiple MBONs and are also reciprocally connected. The convergent MBON input to β'2mp is especially interesting as cholinergic (MBON γ2α'1), GABAergic (MBON γ3β'1), and glutamatergic (MBON γ5β'2a) MBONs drive opposing behaviors. For instance, activation of the cholinergic or GABAergic MBON results in naive odor preference, whereas activation of the glutamatergic MBON results in robust naive avoidance. Similarly, the cholinergic MBON activity mediates aversive associations, whereas glutamatergic MBON activity mediates appetitive associations and extinction of aversive (Scaplen, 2021).
Considering that MBON β'2mp receives convergent input from these parallel and opposing pathways, it likely serves as a decision hub by integrating activity to modulate cue-induced approach and avoidance behavior. How MBON β'2mp integrates information across MBONs and drives behavioral responses remains to be determined. Naive activation of MBON β'2mp does not appear to influence behavioral choice, it instead acts as a sleep suppressor. Inhibition of MBON β'2mp during sleep enhances long-term memory. Separately, local protein synthesis within MBON β'2mp, has been implicated in the consolidation of long-term memory. This makes MBON β'2mp an ideal model for understanding how sleep and memory signals might be integrated at a molecular level. It should be mentioned that MBON γ3β'1 reportedly acts as a sleep activator and local protein synthesis within this MBON is also important for the consolidation of long-term memory. Thus, MBON γ3β'one likely also plays a role in integrating sleep and memory signals through its reciprocal connections MBON β'2mp (Scaplen, 2021).
This provides a well-characterized anatomical framework to understand how opposing memories are acquired, consolidated, expressed and updated. Since the roles of these converging MBONs in naive and learned behaviors are state dependent, it is hypothesized that MBON γ3β'1 and MBON β'2mp, both receiving convergent input from other MBONs, providing opportunities for feedforward networks to update information processing depending on the state of the animal (Scaplen, 2021).
Some of the feedback connections originally hypothesized to exist in the MB were between MBONs and DANs. The current analysis revealed neurons postsynaptic to MBONs that are TH positive. Recent studies that combined EM annotation and calcium imaging to identify specific MBON-DAN connections suggest extensive recurrent connectivity between MBONs and DANs, validating these findings. For example, previous studies using both GFP Reconstitution Across Synaptic Partners (GRASP) and EM annotation revealed that MBON α1 and DAN α1 are synaptically connected. This study similarly identified a few DAN neurons that innervate the horizontal MB lobes within the MBON α1 postsynaptic signal. A recent study showed that the 20 DANs that innervate the γ5 MB compartment are clustered into five different subtypes that innervate distinct anatomical regions within the γ5 compartment. According to this study, only one of the γ5 DANs receives direct recurrent feedback from γ5β'2a MBONs. Based on these recent anatomical characterizations, it is believed that the TH+ neurons within the postsynaptic signal of γ5β'2a are the γ5 DANs (Scaplen, 2021).
The FSB is the largest substructure of the central complex, and it serves as a sensory-motor integration center. The FSB comprises nine horizontal layers that are innervated by large-field neurons. Previous work in blow flies and, later work in Drosophila, predicted that the FSB was postsynaptic to output neurons of the MB. The current data confirm that the large-field, tangential neurons of the dorsal FSB are postsynaptic to the majority of MBONs. Although there exists some variation across brains, glutamatergic and GABAergic MBONs predominately project to FSB layers 4 and 5, whereas cholinergic MBONs mainly project to FSB layer 6. Connections between MBONs and FSB were consistent across different split-GAL4 lines that have overlapping expression patterns. Similar extensive direct connectivity between these MBONs and the dorsal FSB, especially layers 4 and 5, were found in the recently annotated EM hemibrain dataset. Together, these observations suggest that the connectivity between the MB and FSB are structurally, and perhaps in some cases functionally, conserved across insect species (Scaplen, 2021).
How are FSB layers 4/5 and 6 functionally distinct? The dorsal FSB has a well-established role in modulating sleep and arousal, locomotor control, courtship, and visual memory. FSB layer 5 has been specifically implicated in processing information regarding elevation in a foraging- and rutabaga-dependent manner. More recent studies have implicated the dorsal FSB in processing nociceptive information. FSB layer 6 plays a specific role in avoidance of a conditioned odor, whereas layers 4 and 5 respond to aversive stimuli and are responsible for innate, but not conditioned, avoidance. Moreover, recent connectome data suggest that differences exist in the postsynaptic connections of layers 4/5 and 6 as well. Overall, there is high degree of interconnectivity within the FSB. The predominate output of FSB layer 6 neurons are other FSB neurons. In fact, many FSB layer 6 neurons project exclusively to other FSB neurons. In contrast, FSB layer 4 neurons send direct projections to other brain structures -- CRE, SMP, and LAL -- in addition to projecting to other FSB neurons. The connections with the LAL position the FSB layer 4 to directly influence downstream motor output signals prior to executing behavior. Recent EM analysis also suggests that some FSB layer 6 neurons synapse back onto PAM DAN neurons. This connectivity is in line with the associative role in conditioned nociception avoidance described for FSB layer 6 (Scaplen, 2021).
Interestingly, this study found that the pattern of FSB postsynaptic targets of the MBONα1 is dissimilar to other glutamatergic MBONs. FSB layers 4/5 and 6 are not present in the MBON α1 postsynaptic signal. Instead, MBON α1 project to neurons that innervate the ventral and most dorsal aspect of the FSB. The ventral FSB is implicated in innate avoidance of electric shock, and recent data suggest that its activity is tuned to airflow cues for orientation during flight. Artificial activation of MBON α1 does not result in significant avoidance behavior. However, it has been implicated in the acquisition, consolidation, and expression of 24 hr long-term sucrose memory. It is possible that MBON α1 provides appetitive valence signals to the ventral FSB to guide goal-directed flight. Functionally validating the role of MBON α1 and its relationship with its putative downstream neurons is key to appreciating how learning signals can drive behavioral decisions (Scaplen, 2021).
More research is necessary to further understand the functional role of different FSB layers and how information is integrated across these layers. Based on the anatomical data, it is clear that although the MB and FSB can function in parallel during memory formation, they act as parts of a dynamic system to integrate information and adjust behavioral responses (Scaplen, 2021).
The LAL is an important premotor waystation for information traveling from the central complex to descending neurons innervating thoracic motor centers across insects. Accordingly, the LAL has been implicated in orientation to pheromones in the moth, flight in the locust and dragonfly, locomotion in Drosophila stimulus-directed steering in Drosophila, the cockroach, cricket, and moth and in response to mechanosensory stimuli in the locust. In the moth, recordings from neurons innervating the LAL have a characteristic 'flip-flop' firing property, which is thought to mediate walking command. More recent work has suggested a functional organization whereby the neurons in the upper division of the LAL receive convergent input from the protocerebrum and neurons in the lower division generate locomotor command (Scaplen, 2021).
The current data show that the MB network converges with the protocerebrum input, thereby providing an opportunity for MBONs to indirectly influence descending motor outputs. It was also demonstrated that two MBONs (γ3β'1 and γ2α'1) synapse on the same subset of LAL and FSB cells, revealing a convergent circuit that connects both structures. Further, in support of anatomical observations, optogenetic activation of MBON γ2α'1 resulted in activation of both LAL and FSB layer four neurons. Given that MBON γ3β'1 is GABAergic, the equivalent experiment was not performed for this neuron. Thus, understanding the functional consequences of these inhibitory connections will require further investigation. Interestingly, despite the fact that MBON γ3β'1 and γ2α'1 express different neurotransmitters and innervate different MB compartments, their manipulation has similar behavioral phenotypes: both promote sleep, and artificial activation of either results in naive preference. Further, activation of both MBON γ3β'1 and γ2α'1 together has an additive effect, which results in a significant increase in preference (Aso et al., 2014b) (Scaplen, 2021).
The FSB and LAL have a well-established structural and functional connectivity. The LAL integrates information from the central complex, including the FSB, and provides a premotor signal to motor centers. However, the behavioral significance of MBON γ3β'1 and γ2α'1 projections to both the FSB and LAL is less clear. Previous work demonstrated that activation of these MBONs while the flies explored an open arena did not significantly affect average speed or angular speed of individual flies. By contrast, this study found that inactivation of the putative downstream LAL neurons significantly increased overall activity of behaving flies in a social context and locomotor assay. Thus, the γ3β'1 and γ2α'1 MBONs may play a modulatory rather than required role in influencing behavioral response to an associated cue (Scaplen, 2021).
Recent work in Drosophila has demonstrated that the DANs that innervate MBON γ2α'1 regulate flight bout durations, and may provide a motivation signal via MBONs to the FSB and LAL to regulate motor activities. The LAL neurons receive multisensory input, and some LAL neurons make direct connections to descending neurons that control movement. Thus, this circuit organization enables integration of sensory signals with punishment or reward to direct the motion of the animal. In contrast, MBON connections with the FSB might play a role in providing context for flexible navigation, goal-directed actions, and memory-based navigation (Scaplen, 2021).
If homology can be defined by shared expression of transcription factors and similar functional roles, the MB-FSB connection may be an appropriate model for understanding functional connections between the hippocampus and striatum and serve as an accessible model for understanding connectivity between more complex brain structures associated with memory. Further, given that the integrative relay role of the LAL is somewhat reminiscent of the vertebrate thalamus, the complex connectivity between the MBONs, FSB, and LAL may also serve as an effective model for predicting and understanding functional connections between the hippocampus, striatum, and thalamus in the context of memory formation and action selection (Scaplen, 2021).
Insects exhibit a great variety of complex behaviors, and significant effort has been devoted to understand the neural circuits that underlie these behaviors. The genetically accessible Drosophila is a great model for studying the interplay between circuit architecture and behavior owing to their complex yet tractable brains. The MB circuits and their role in learning and memory are among the most studied circuits in Drosophila. Although, the majority of these studies have focused on olfactory memory, it is clear that the MB plays a much broader role in insect behavior. In Drosophila, the MB is important for courtship memory, taste aversive memory as well as visual memory. In cockroaches, the MB has a role in place memory and recent data in two different species of ants implicate the MB in spatial navigation to learned locations using visual cues. In mammals, the hippocampus is similarly required for multiple forms of associative memory, including spatial navigation using visual cues. Thus, cross-species similarity in circuit organization and function may exist between the mushroom body and the hippocampus. However, such anatomical and functional cross-species comparisons can also be made between the mushroom body and the cerebellum, suggesting that similar convergent-divergent architecture may be a general principle of structures that encode and update memories (Scaplen, 2021).
In this context, the implementation of trans-Tango to study the MB has high potential in the era of EM reconstruction of the Drosophila brain. Through examination of the circuit connectivity in several individuals, easily afforded by trans-Tango, the value of the EM reconstruction data could be augmented by overlaying on it potential nuanced differences between individuals. In addition, trans-Tango-mediated discoveries in the fly could help illuminate principles of circuit organization in other species. Further, due to the modular design of trans-Tango, it could be readily reconfigured for other types of studies beyond circuit tracing. For example, only minimal modifications are required for implementing a configuration of trans-Tango for identifying the molecular composition of the postsynaptic partners. This strategy could be used to examine the evidence that MBONs stratify the FSB through different classes of peptidergic neurons. Confirmation of these observations would suggest that the MB plays a critical role in regulating modulatory systems of a midbrain region that shares structural and functional commonalities with the vertebrate basal ganglia. Finally, through combining it with new genome editing strategies, trans-Tango could become a useful tool for comparative anatomy in other insects. This would enable the study of synaptic connections in non-model organisms and lead to deeper understanding of biological diversity (Scaplen, 2021).
Understanding how memories are formed, stored, and retrieved necessitates knowledge of the underlying neural circuits. This characterization of the architecture of the neural circuits connecting the MB with downstream central complex structures lays the anatomical foundation for understanding the function of this circuitry.These studies may also provide insight into general circuitry principles for how information is processed to form memories and update them in more complex brains (Scaplen, 2021).
Insects exhibit an elaborate repertoire of behaviors in response to environmental stimuli. The central complex plays a key role in combining various modalities of sensory information with an insect's internal state and past experience to select appropriate responses. Progress has been made in understanding the broad spectrum of outputs from the central complex neuropils and circuits involved in numerous behaviors. Many resident neurons have also been identified. However, the specific roles of these intricate structures and the functional connections between them remain largely obscure. Significant gains rely on obtaining a comprehensive catalogue of the neurons and associated GAL4 lines that arborize within these brain regions, and on mapping neuronal pathways connecting these structures. Toward this end, small populations of neurons in the Drosophila melanogaster central complex were stochastically labeled using the multicolor flip-out technique and a catalogue was created of the neurons, their morphologies, trajectories, relative arrangements and corresponding GAL4 lines. This report focuses on one structure of the central complex, the protocerebral bridge, and identifies just 17 morphologically distinct cell types that arborize in this structure. This work also provides new insights into the anatomical structure of the four components of the central complex and its accessory neuropils that are arborized by PB neurons include the crepine (CRE), rubus (RUB), gall (GA), and lateral accessory lobe (LAL). Most strikingly, the protocerebral bridge was found to contain 18 glomeruli, not 16, as previously believed. Revised wiring diagrams that take into account this updated architectural design are presented. This updated map of the Drosophila central complex will facilitate a deeper behavioral and physiological dissection of this sophisticated set of structures (Wolff, 2014).
The work presented in this study builds on published studies by both defining previously unidentified anatomical features of each of the four components of the central complex as well as updating wiring diagrams to accommodate these new anatomical insights. This paper also reports new cells and new features of previously identified cells and the genetic reporter lines that reveal them, with the prospect that these will form an essential stepping stone both to synaptic studies at the electron microscope level and to functional studies. The most significant new insights from this work are summarized below. As noted earlier, the statements below are drawn from neurons that arborize in the PB (Wolff, 2014).
The most surprising finding of this study is that the Drosophila protocerebral bridge comprises 18 glomeruli. This finding has an important impact on the wiring relationships between the glomeruli and their respective vertical units in the FB, the columns, and in the EB, the wedges and a new volume described in this study, the tiles (Wolff, 2014).
The longstanding belief regarding the correspondence between the PB and FB and PB and EB wedges was that there is a 1:1 relationship between the vertical subdomains of these structures. The finding that there are 18 glomeruli raised the possibility that the FB and EB also exhibit an octodecimal organization. However, compelling evidence is provided that there are just 16 wedges in the EB. and it was further show, that some cells arborize in just half a wedge, indicating the further division of wedges into 32 demi-wedges. The observation that a simple 1:1 correspondence between the PB and EB wedges is lacking and, furthermore, that there are also demi-wedges, has implications for how the system is wired to accommodate this numerical discrepancy (Wolff, 2014).
Another unexpected finding from this work is the existence of a second EB volume that also partitions the EB around its circumference: the tile domain. Tiles are distinct from wedges in that there are half as many tiles as wedges (eight tiles), they are functionally distinct from the wedge (output versus input, respectively), and these two volumes survey different volumes of the EB since they extend to different depths of the EB. Only two PB cell types target the tile domain (Wolff, 2014).
Although a columnar morphology is apparent in layers 1–8 of the FB in nc82-labeled samples, the organization of the cells that populate these layers is not universally columnar. There is a minimum of nine layers in the FB, yet a columnar organization (i.e., vertical stratification) of cell arbors is restricted to layers 1–5 for single column widths (where the columnar organization for layers 4 and 5 is revealed by the PBG1–8.s-FBℓ3,4,5.s.b-rub.b cell); wider, more loosely organized arbors occur in more dorsal layers. With the exception of arbors in layer 1, the column borders are not rigid, as neighboring arbors overlap one another, sometimes extensively. The unique tooth-like structure of layer 1 of the FB definitively shows that there are nine columns in this layer. Due to the overlap of arbors, it is more difficult to count columns in the other layers, but layers 2 and 3, which exhibit a tighter columnar organization than more dorsal layers, likely have 16 columns based on mapping data. This would be consistent with parallel divisions of the EB (wedges) (Wolff, 2014).
Other new anatomical features and subdomains are described in this study. First, it was shown that each of the noduli has subcompartments. The dorsal noduli, NO1, have medial and lateral subcompartments. The medial noduli, NO2, consist of two distinct subcompartments, NO2D and NO2V; no PB cell type arborizes in either of these subcompartments. The ventral noduli, NO3, consist of three distinct subcompartments, NO3A, NO3M, and NO3P. The nubbin is a partial shell on the dorsal, anterior face of the EB, and the "gall tip" is a region at the dorsal tip of the gall. Finally, two undefined regions to which some cells project and that are not clearly demarcated are the dorsal and ventral gall surround (Dga-s and Vga-s) (Wolff, 2014).
Even though the subdomains of the central complex structures can be distinguished from one another, they apparently do not function as isolated subunits. Rather, there is shared communication between most of these subunits. At least for the neurons described in this study (i.e., those that arborize in the PB), both pre- and postsynaptic arbors in the glomeruli, EB tiles, wedges and shells, FB columns, and NO1 (medial and lateral domains) can extend into neighboring domains. This sharing of information is not obvious between NO2D and NO2V, nor between NO3A and NO3M. The boundary between NO3P and NO3M is too obscure to evaluate if arbors in these two domains are completely restricted or shared, although in the examples they appear to be restricted. The frequency and degree to which arbors overlap in the various subunits is cell type-dependent. While some arbors exhibit no or minimal intrusion into adjacent volumes, overlap between neighboring units could serve important circuit functions (Wolff, 2014).
This work identifies 17 unique cell types that arborize in the protocerebral bridge. These fall into four classes: cells that 1) are intrinsic to the PB (n = 2), 2) are intrinsic to the central complex (an additional 6), 3) arborize in the FB, EB, or NO in addition to extra-central complex regions (e.g., the gall; n = 6), and 4) arborize exclusively in the PB and regions outside the central complex (n = 3). Cells that arborize in the PB receive their input from the EB, LAL, PS, IB, and also from within the PB. One cell previously identified in another study (Lin, 2013) was not targeted by any of the ∼35 lines analyzed in this work. To the extent that it is possible to construct wiring diagrams from the images shown in these two studies, it appears that the circuits for these cells are also identical between Drosophila and Schistocerca. In addition, one cell type was identified in this study that was not characterized in Lin (2013) (Wolff, 2014).
The combined total from this work and Lin (2013) brings the current number of identified cell types that arborize in the protocerebral bridge of Drosophila to 18. A potential 19th cell type was seen just twice, and in neither case could the entire cell be traced. Its PB arbor is spiny and very sparse, and while it clearly arborizes in the central brain, it is not clear if it arborizes elsewhere within the central complex. It is also possible that this "cell type" only constitutes a variant. There will likely be additional cells identified that arborize in the PB, although this number is predicted to be small. A complete inventory of all cells in the Drosophila brain awaits a full reconstruction at an electron microscope level (Wolff, 2014).
The wiring diagrams described in this study differ from published reports, in part due to the fact that previous authors were unaware of the existence of 18 glomeruli in the PB and therefore based their models on the historic interpretation that there are 16 glomeruli. This numerical revision and new insights into the anatomical substructure of the central complex components are the primary basis for revisions of existing circuit diagrams (Wolff, 2014).
Although there are 18 glomeruli, no small-field neuron arborizes in all 18 glomeruli. Instead, most cell types arborize in either G1–G8 or G2–G9. Each of these categories adheres to the following basic wiring principle: Cells that arborize in the lateral four glomeruli of each side of the PB stay ipsilateral in the second neuropil (either the FB or EB, depending on the neuron) and cross to the contralateral side at the third neuropil, whereas cells that arborize in the medial four glomeruli cross to the contralateral side in the second neuropil. Consequently, because there are two subsets of PB neurons, the glomerulus that targets a given column, wedge, or tile is shifted by one glomerulus, depending on the subset of cell type. Furthermore, the observation that no small-field neurons arborize in all 18 glomeruli suggests the number of columns in the FB and wedges in the EB would not need to exceed 16 in order to maintain a 1:1 correspondence between the PB and FB/EB (Wolff, 2014).
Arbors from cells that target the PB alternate with one another in the second neuropil such that arbors from the left glomeruli alternate with those from the right glomeruli. The PB wiring diagrams presented in this study differ somewhat from a recent account (Lin, 2013), as follows. The most lateral FB column (or EB wedge) is occupied not by the ipsilateral G9 (or G8 for the G1–G8 cells), as previously described, but instead by the contralateral G2 (or G1 for the G1–G8 cells). This circuit therefore reverses the pattern in the second neuropil (FB or EB) from one in which the most lateral (L) glomeruli project to the most lateral columns (or wedge or tile) on the ipsilateral side to one in which the medial (M) glomeruli from the contralateral half of the PB project to the most lateral columns. In other words, previously published diagrams indicate a pattern of LMLMLM from lateral to medial in the second neuropil, whereas this report shows that pattern to be MLMLML (Wolff, 2014).
The projection map shown in this study for cells that connect the PB to layer 1 of the FB illustrates conclusively the projection pattern between these domains. Obtaining an accurate map between the PB and layers 2 and 3 is difficult given the greater overlap between arbors of cells in these two layers, but the projection patterns observed between the PB and layers 2 and 3 of the FB are consistent with the PB:FBℓ1 map (Wolff, 2014).
As noted above, distribution of information is not always restricted to the subdomains of each central complex structure. When information is shared between neighboring domains (or alternating domains, in the case of the cells that arborize in the dorsal or ventral gall), generally only a small portion of the arbor is shared. The functional significance of these zones of overlap remains to be determined (Wolff, 2014).
Connections between central complex structures are remarkably restricted. For example, of those neurons that arborize in the PB and FB, only FB layers 1, 2, and 3 connect to the noduli, and only to NO2 and NO3. The only link between the PB and NO1 is via the ellipsoid body, so whereas NO2 and NO3 can be considered to work in conjunction with the FB to elicit a behavior based on output from the PB, NO1 cooperates with the EB to elicit a behavior based on output from the PB. In fact, communication is even more specific: layer 3 of the FB communicates directly only with NO2, layer 2 directly only with the anterior subcompartment of NO3, and layer 1 directly only with NO3M and NO3P. Furthermore, the cells in G1 do not communicate directly with the noduli at all—neither via the FB nor via the EB. The absence of direct connections between the PB and upper layers of the FB is also noteworthy. This streamlined and highly segregated network of connections within and between central complex structures suggests a high degree of regional specialization in function for the components of the central complex (Wolff, 2014).
The roles of central complex structures, their subdomains, and related neuropils are poorly understood. While functions remain largely unknown, many circuits described in this study are informative in various other ways. For example, some identify commonalities in function between neuropil subregions, such as FBℓ2 and NO3A, which are both arborized by a common neuron. Other circuits reveal spatial segregation between neuropils. The most intriguing instance is the exclusive relay of information between the ventral gall and even-numbered glomeruli and the dorsal gall and odd-numbered glomeruli, which demonstrates that both information and information flow can be spatially segregated from the glomeruli to the gall. It will be interesting to learn the functional role of the gall and why it segregates a portion of the information it receives and sends, as well as what sort of behavioral response requires this rigidly alternating spatial distribution in the PB. Finally, the absence of connections between neuropils may prove informative in functional studies. For example, G1 is distinct from G2–G8 in that it lacks direct connections with the noduli, and NO1 is distinct from NO2 and NO3 in that it communicates with the PB via the EB rather than the FB, raising the questions of what behaviors G1 does and does not contribute to, and what the differences are in behavioral outputs from NO1 and NO2/NO3 (Wolff, 2014).
The observation that there are 18 glomeruli in the Drosophila PB has significant implications for both the architecture and evolution of the Drosophila brain with respect to the brains of other neopterans. Although the suite of genetic tools available in locusts, bees, beetles, and other insects does not yet include MCFO, improvements in imaging and histology may prove sufficient to reevaluate the number of glomeruli in these species, given that glomeruli can be accurately counted in brains that are labeled only with nc82. Either some or all of these species also have 18 glomeruli, or an extra pair of glomeruli arose in Drosophila. The latter may be unlikely but would raise some intriguing possibilities about how the anatomical correspondence and circuitry between glomeruli in the PB and equivalent vertical partitions in the FB and EB PB circuits may differ between flies and other insects thought to have the same basic cellular composition and organization within the central complex, and how the geometric coordinates would then have had to shift along the axis of the PB to effect accurate behavioral responses (Wolff, 2014).
The central complex (CX) is a midline-situated collection of neuropil compartments in the arthropod central brain, implicated in higher-order processes such as goal-directed navigation. This study provides a systematic genetic-neuroanatomical analysis of the ellipsoid body (EB), a compartment which represents a major afferent portal of the Drosophila CX. The neuropil volume of the EB, along with its prominent input compartment, called the bulb, is subdivided into precisely tessellated domains, distinguishable based on intensity of the global marker DN-cadherin. EB tangential elements (so-called ring neurons), most of which are derived from the DALv2 neuroblast lineage, predominantly interconnect the bulb and EB domains in a topographically organized fashion. Using the DN-cadherin domains as a framework, this connectivity was first characterized by Gal4 driver lines expressed in different DALv2 ring neuron (R-neuron) subclasses. 11 subclasses were identified, 6 of which correspond to previously described projection patterns, and 5 novel patterns. These subclasses both spatially (based on EB innervation pattern) and numerically (cell counts) summate to the total EB volume and R-neuron cell number, suggesting that this compilation of R-neuron subclasses approaches completion. EB columnar elements, as well as non-DALv2 derived extrinsic ring neurons (ExR-neurons), were also incorporated into this anatomical framework. Finally, the connectivity between R-neurons and their targets was addressed, using the anterograde trans-synaptic labeling method, trans-Tango (Omoto, 2018).
The central complex (CX) is an evolutionarily conserved, higher-order neuropil in the arthropod brain thought to integrate sensory and motor information to coordinate and maintain locomotor behavior, thus enabling appropriate navigation. Drosophila mutations that produce structural abnormalities in CX neuropils result in flies with deficiencies in walking and flight. More targeted manipulations, such as silencing of specific CX neuron subclasses, compromise vision-based memories associated with spatial orientation and location. Similar themes emerge from anatomical, electrophysiological, and behavioral studies investigating the CX in other insects. In the cockroach CX, for example, single unit activity correlated with changes in locomotor intensity, turning behavior, or heading direction have been identified. In addition, electrical stimulation of CX neurons in the freely walking cockroach has yielded direct evidence linking CX activity to downstream locomotor output. In other insects, such as locust, cricket, monarch butterfly, and dung beetle, neurons in the CX are tuned to celestial visual cues such as the sun or pattern of polarized skylight. These cues provide the stable environmental signals required to accurately derive relative heading information for short or long range navigations (Omoto, 2018).
The CX consists of four neuropil compartments: the upper (CBU) and lower (CBL) halves of the central body (CB), protocerebral bridge (PB), and paired noduli (NO). In Drosophila, the upper and lower halves of the CB are designated as the fan-shaped body (FB) and ellipsoid body (EB), respectively (see General overview of the ellipsoid body (EB): neuronal interactions and compartmentalization). Recently, the asymmetrical body, a paired neuropil located ventral of the FB and adjacent to the NO, has been proposed as a fifth neuropil compartment of the CX. These neuropil compartments are largely formed by two orthogonally arranged neuronal populations: (1) columnar (small-field) neurons which interconnect the CX compartments along the antero-posterior axis; (2) tangential (large-field) neurons which provide input from lateral brain neuropils to the CX. Terminal arborizations of these neurons define distinct vertical columns and horizontal layers that can be visualized by markers for synaptic or cell adhesion proteins that globally label, but exhibit variable density in, the neuropil. Based on Bruchpilot immunostaining, seven layers were identified in the Drosophila CBU (=FB). The CBL (=EB) also exhibits a layered organization. In Drosophila, this compartment undergoes a morphogenetic transformation during pupal development, whereby the lateral ends of the originally bar-shaped EB primordium bend ventrally to adopt a toroidal arrangement. As a result, tangential neurons of the EB display a circular shape, and hence were called 'ring neurons'. Likewise, layers within the EB are annuli, rather than horizontal slabs. Based on labeling with DN-cadherin, this study has defined five distinct annular domains, termed anterior (EBa), inner and outer central (EBic and EBoc), and inner and outer posterior (EBip and EBop) domains (Omoto, 2018).
Clonal studies in Drosophila show that the neuronal architecture of the CX is organized into lineage-based modules, a ground plan that is likely conserved across insects. A lineage refers to the set of sibling neurons derived from an individual neural progenitor called a neuroblast, and the entire central brain is generated from a fixed number of approximately 100 of such neuroblasts. Four lineages (DM1-4) give rise to the large number of columnar neurons of the CX. The great diversity observed among these neurons is achieved via temporal patterning of molecular determinants in dividing progenitors. Lineages giving rise to the tangential neurons of the CX have been characterized morphologically, but have not yet received much attention experimentally. The most notable exception is lineage DALv2/EBa1 (henceforth called DALv2), that generates ring neurons of the EB. Ring neurons project their axons to distinct annular domains of the EB, and typically possess short globular dendrites ('microglomeruli') in the bulb (BU), a neuropil compartment located laterally adjacent to the EB. The BU encompasses three main partitions [anterior (BUa), superior (BUs), and inferior (BUi) bulb] that are associated with different annular domains of the EB. Furthermore, the BUs and BUi appear to be divisible into anterior (aBUs/aBUi) and posterior (pBUs/pBUi) regions. Input to the BU is provided by neurons of two additional lineages, DALcl1 and DALcl2 (also called AOTUv3 and AOTUv4, respectively). As part of the anterior visual pathway, DALcl1/2 form so-called tubercular-bulbar (TuBu) neurons which project from the anterior optic tubercle to the BU, relaying visual information to ring neurons and thereby the CX as a whole. TuBu neurons form two lineally segregated parallel channels, with DALcl1 establishing connections with ring neurons located in the peripheral domain of the EB via the BUs, and DALcl2 with central ring neurons via the BUi (Omoto, 2018).
Detailed functional studies are beginning to shed light on the circuitry involving ring neurons and their TuBu afferents and columnar efferents. Two-photon calcium imaging has revealed a discrete focus of neural activity, or 'bump,' within a population of columnar neurons ('E-PGs') that interconnect the EB, PB, and gall (GA) of the LAL. E-PG neurons encode an internal compass representation via the activity bump, which dynamically tracks the fly's heading. Additional columnar neuron populations that interconnect the PB, EB, and NO, called P-EN neurons, compute the animals' heading by controlling the movement of the bump in the clockwise or counter-clockwise direction. These findings suggest that the EB may operate as a critical hub in the CX, acting as an interface between neurons that transmit and distribute sensory information (TuBu and ring neurons), and circuits that encode and update a representation of heading direction (E-PG and P-EN neurons). In addition, internal state information is likely integrated into the EB network by additional ring neurons subclasses that signal physiological needs such as sleep and hunger drive (Omoto, 2018).
To make further inroads in understanding how the EB circuitry operates, a comprehensive knowledge of ring neurons and their upstream and downstream connectivity is required. Ultimately, a comprehensive analysis of single cells and their synaptic contacts on the light and electron microscopy level will yield complete coverage of the EB wiring diagram, and certainly inform understanding of how EB-related computations are implemented. However, a current description of subclass-specific projection patterns using genetic driver lines provides a framework to posit inter-class neural interactions that can then be tested physiologically and/or behaviorally, and will assist future efforts for such high-resolution anatomical maps. To this end, this study sought to expand on previous works using this genetic-anatomical approach to more thoroughly describe the EB neuropil. Gal4 driver lines that label ring neuron subclasses were screened and subsequently distinguished from each other based on defined criteria. Many drivers label populations corresponding to previously identified ring neuron subclasses, in addition to several, yet uncharacterized populations. The novel subclasses were given new names per the historical nomenclature system. Columnar elements were also incorporated into this anatomical framework. Based on the domain innervation pattern of each line, putative interactions between elements within the EB network are proposed. Finally, ring neuron drivers were subjected to the anterograde trans-synaptic labeling method, trans-Tango. Ring neurons occupying central domains of the EB commonly display homotypic interactions, such that neurons of a given subclass predominantly form synaptic interactions with other neurons in the same subclass. On the other hand, ring neurons occupying the peripheral domains typically display a larger degree of output into the columnar network. This highlights a fundamental difference in the connectivity, and potentially the functions, of ring neurons in different domains (Omoto, 2018).
This work serves to build upon previous anatomical studies by further clarifying the neuronal architecture of the Drosophila EB. Five definitive DN-cadherin domains constituting the EB neuropil provide fiducial landmarks with which neuron classes can be placed into spatial context. Based on this framework, this study reports several novel ring neuron subclasses and proposes potential interactions between ring, columnar, and neuromodulatory neurons in the EB. Lastly, putative postsynaptic partners of R-neurons were experimentally mapped using trans-Tango, revealing insight into how information may be distributed throughout the EB and the rest of the CX. In addition to the neuroanatomical description of different populations, the identification of driver lines enables genetic access to label or manipulate these populations. This provides an entry point for future studies to probe the functional properties of each class and test the interactions proposed herein. The following summarizes the primary findings, speculates on the functional significance of CX wiring principles, and places this study into a developmental-neuroanatomical context with previous works in Drosophila and homologous structures in other insects (Omoto, 2018).
The CX is viewed as a critical hub for goal-directed navigational behavior in insects. Streams of sensory information from different modalities must converge onto this center of sensorimotor integration to guide navigational decisions based on current trajectory, learned information, and motivational state. Central to this notion was the identification of a stable compass representation that tracks the flies heading in the E-PG neuron population. The robustness of this neural correlate of angular orientation, manifested as a single calcium activity 'bump' that moves around the EB, depends on both visual and proprioceptive cues (Seelig, 2015). Heavily relying upon studies in other insect species as a basis for comparison, recent progress has been made toward identifying the neural pathways that transmit sensory information to the Drosophila CX, with visual input being the most well characterized. The fly CX receives visual information via the anterior visual pathway (AVP), a circuit defined by three successive layers. Information is transmitted from the optic lobe medulla to the anterior optic tubercle, from the tubercle to the bulb (BU), and from there to the EB, via medullo-tubercular (MeTu), tuberculo-bulbar (TuBu), and DALv2 ring neurons (R-neurons), respectively. Parallel ensembles of TuBu neurons terminate in a topographically organized fashion onto the microglomerular dendrites of distinct R-neuron subclasses within the BU. Specific computations are implemented across successive layers in this pathway, such as the integration of recent visual history and self-motion, which may inform downstream behavior. Ring neurons transmit processed visual information concerning features and landmarks to the EB, likely as a stable allothetic reference to guide bump dynamics in E-PG neurons. The interaction between tangential elements of the EB and columnar neurons such as E-PG neurons has been suggested in other insects, and confirmed by GFP reconstitution across synaptic partners (GRASP) in Drosophila. Indeed, this study provides further evidence via trans-Tango that R2 neurons, which are tuned to visual features, provide direct presynaptic input to E-PG neurons. The calcium activity bump in E-PG neurons also shift in total darkness, demonstrating the existence of a proprioceptive input channel that can update the heading representation in the EB in the absence of visual input. It is posited that transmission of idiothetic cues to the CX is mediated in part by R1 and/or ExR4 neurons, as their neurite distribution and polarity suggests feedback from the LAL, a proposed motor signaling center (Omoto, 2018).
Conceivably, the information received by different R-neuron subclasses is transmitted to their ring-shaped neurites, and is processed via connections within the same subclass (homotypic interactions) and/or between subclasses (heterotypic interactions), the extent of which depends on the R-neuron subclass in question. As such, the R-neuron system likely displays recurrent connectivity to enable persistent activity required for memory processes, as has been shown for mushroom body circuits that support courtship memory. Indeed, inner ring neurons (likely R3d and R3p), which comprise a critical nucleus of visual working memory, display prominent homotypic interactions. Future work to define the mechanisms underlying intra-subclass interactions and experiments to perturb them, are required to assess the functional significance of these homotypic interactions (Omoto, 2018).
R-neurons, particularly subclasses of which occupy peripheral EB domains, provide input to several different columnar neuron populations. This study provides novel insight into the nature of subclass-specific, input-output communication between the ring and columnar networks. An important avenue of future work will be to elucidate the tuning properties of each R-neuron subclass and determine the contribution of each input to compass representation. Presumably, R-neuron subclasses that provide prominent, direct input to E-PG neurons, such as R2 or R4m, would exhibit the most influence over compass representation (Omoto, 2018).
Circuit flexibility is likely facilitated by neuromodulatory input on a moment-by-moment basis, which may reconfigure information flow through the network and thus the output of the system. Neuromodulation would likely occur at multiple processing stages, as evidenced by the wide-spread neurites of dopaminergic neurons. For example, a single PPM3 neuron, innervates the GA/LAL, BU, and EBoc/op. It is envisaged that neurite-specific signaling and plasticity may regulate distinct processing nodes, akin to what has been demonstrated for dopaminergic neurons that encode protein hunger. Similarly, 5-HT may also influence R-neuron activity as projections from the serotonergic neurons, ExR3 [corresponding to the posterior medial protocerebrum, dorsal cluster (PMPD)], most prominently innervate EBic. The effect of serotonin may be receptor and circuit specific; distinct 5-HT receptor isoforms are differentially expressed in specific R-neuron subclasses (Omoto, 2018).
For clarity, the five EB domains defined by the global marker DN-cadherin should be reconciled with previously used anatomical terminology of the EB. Frontal sections of the EB at different anteroposterior depths shows that DN-cadherin domains are distinct, annular entities. These domains correspond to 'layers' in other insects, and have sometimes been also referred to as layers in Drosophila as well. Therefore, N-cadherin EB domains are synonymous with layers. Each domain is best represented using a 'dorsal standard view': a horizontal section through the EB containing a lengthwise perspective of the EB canal. From this standard view, the N-cadherin domains are also clearly organized along the anteroposterior axis. Three anteroposterior subdivisions of the EB have been referred to as 'shells,' in line with terminology used for the FB. It is proposef that the anterior most shell encapsulates the anterior domain of the EB (EBa), and therefore consists of only one layer. The intermediate shell encapsulates the inner central (EBic) and outer central (EBoc) domains, and consists of two layers. Finally, the posterior shell encapsulates the inner posterior (EBip) and outer posterior (EBop) domains, and consists of two layers. For example, P-EN neurons occupy the EBop domain, which resides in the posterior EB shell (Omoto, 2018).
Previously, four substructures denoted as 'rings' [EBA (Anterior), EBO (Outer), EBC (Center), EBP (Posterior)], were based on anti-disks large (DLG) immunostaining and roughly correspond to the DN-cadherin domains. Like the DN-cadherin domains, each 'ring' was proposed to contain specific R-neuron subclasses. Based on the ring neuron subclasses to comprise each 'ring', it is inferred that EBA corresponds to EBa and EBic in the current classification system. Furthermore, EBO is EBoc, EBC is EBip, and EBP is EBop (Omoto, 2018).
How does the annular domain structure of the Drosophila EB compare to the lower division of the central body (CBL) described for other insects? Similar to the EB, the CBL represents a multilayered neuropil compartment formed by the neurite contributions of tangential and columnar elements. In insects such as locust (Schistocerca gregaria), which will be used as the primary basis for comparison in the following, the kidney bean or sausage-shaped CBL corresponds to the torus-shaped EB in Drosophila. In locusts, the CBL is effectively located ventrally of the upper division of the central body (CBU), whereas the homologous structures in Drosophila (EB and FB, respectively) are arranged in an antero-posterior fashion. This difference is reflective of a 60° anterior tilt of the locust neuraxis, as evidenced by the peduncle, which extends horizontally in flies but is oriented almost vertically in the locust. In the dung beetle (Scarabaeus lamarcki) and monarch butterfly (Danaus plexippus), the CBL are also sausage-shaped, but the neuraxis orientation is like that of Drosophila. Differences in neuraxis orientation influence the comparison between the internal architecture of the locust CBL and fly EB. The locust CBL is subdivided along the dorso-ventral axis into six horizontal layers (although not stacked seamlessly on top of one another). Based on the expression of global markers, the Drosophila EB is divided into toroidal domains (EBa/ic/oc/ip/op). Considering the tilt in neuraxis, it is posited that dorsal strata (layers 1-2) of the locust CBL roughly correspond to more posterior domains (EBip/op) of the fly EB, whereas ventral strata (layers 3-6) correspond to more anterior EB domains (EBa/ic/oc). Corroborating this notion is the fact that fly P-EN neurons innervate EBop, and the locust homologs (called CL2 neurons) innervate dorsal layers of the CBL (Omoto, 2018).
The EB and its domains, as well as other structures of the CX, are established by the neurite contributions of distinct neuronal populations. How is the neuronal diversity and connectivity of the CX developmentally established? The CX, and brain in general, is organized into structural-genetic modules called lineages; a lineage comprises the set of sibling neurons derived from an individual neural progenitor (neuroblasts). Each neuroblast forms a spatially discrete cluster of neurons with shared wiring properties; sibling neurons extend a limited number of fasciculated axon tract(s) and innervate specific brain compartments. Most brain lineages are 'type I' neuroblast lineages, whose neuroblasts undergo a series of asymmetric divisions each of which renews the neuroblast and produces a ganglion mother cell. Columnar neurons of the CX are generated from four type II lineages which are larger and more complex than type I, with neuroblasts first producing a set of intermediate progenitors which in turn, give rise to ganglion mother cells (Omoto, 2018).
While the columnar neurons contributing to the EB are derived from type II lineages, the tangential elements (R-neurons) are largely derived from a single paired type I neuroblast, forming the lineage DALv2 (also called EBa1). Neurons of the DALv2 lineage have been studied in developmental contexts in a number of previous works. Production of secondary neurons by DALv2 begin around 24 h after hatching. According to Kumar (2009), one of the DALv2 hemilineages undergoes apoptotic cell death, implying that the DALv2 R-neurons forming the adult EB represent a single hemilineage. Cursory heat-shock inducible single-cell clonal analysis carried out in the present study suggests that distinct R-neuron subclasses are born during specific time windows and therefore represent sublineages of DALv2 (Figure 4). Thus, clonal induction shortly after the onset of secondary neuroblast proliferation (20-48 h after hatching) yielded exclusively outer R-neurons of the R4m subclass. At increasingly later time points, these types of clones become rare, and disappeared entirely at induction times after 96 h. The converse is the case for inner ring neurons (R3d/m), which could be induced in increasing numbers with later time points of induction. Given that only a fraction of the overall number of R-neuron subclasses was represented among clones analyzed in this study, additional studies are required to settle the exact birth order of different R-neuron subclasses (Omoto, 2018).
The following provides a brief historical account of ring neuron definitions, attempt to resolve discrepancies in the literature when possible, and provide rationale for naming conventions used in this work (Omoto, 2018).
The R-neuron type corresponds to ring neurons of the DALv2 lineage, with four R-neuron subclasses described in an initial study (R1-4). Two other ring neuron types were designated as 'extrinsic ring neurons' (ExR-neurons), based on large projections outside of the EB; in this study, with this feature were pooled into a single type, the ExR-neurons. The first described type of extrinsic R-neuron (the ExR1 subclass) likely corresponds to helicon cells. The second type (the ExR2 subclass), due to its innervation of the caudal EB, ExR2 may correspond to the EBop-innervating PPM3 dopaminergic neuron. The serotonergic neurons that innervate the EB, corresponding to the PMPD neurons, designate in this study as ExR3. Therefore, ExR1-3 are posteriorly localized ExR-neurons, likely deriving from the DM3-6 lineages. Due to its wide arborization and non-DALv2 based origin, ring neurons of lineage BAmv1, with perikarya in the anterior cortex, were designated in this study a fourth type of ExR-neuron (ExR4); the possibility cannot be excluded that ExR2 from a previous study may correspond to ExR4-neurons, as they too innervate the caudal EB. Furthermore, the 'P'-neurons, described a previous study as having ventrally localized cell bodies and also innervate the caudal EB, likely correspond to what this study designates as ExR4-neurons (Omoto, 2018).
Driver line c105 was found in an earlier study to label R1 neurons, due to their centrifugal arborization pattern, inner ring localization, and extension into the posterior layers of the EB. However, c105-positive R1 neurons exhibit ventrally projecting neurites into the LAL and lack BU microglomeruli, in contrast to what was defined as R1 in a previous study. Due to R1 being the predominant designation this R-neuron subclass thereafter, this classification as R1 is retained in the current study (Omoto, 2018).
In more recent studies, the driver 38H02-Gal4 has been described as labeling R4 (or an R4-subset), in several studies. 38H02-Gal4 does in fact label R4m (based on BUa microglomeruli and centripetal EBoc innervation pattern), but also strongly labels R5. Two other drivers, 15B07-Gal4 and 28D01-Gal4, were used to target EB neurons required for visual-thermal associations in place learning, and were described as labeling 'R1 and R4,' or 'R1 alone,' respectively. Anatomical re-assessment of these drivers reveals that 15B07-Gal4 labels R3d, R3p, and R4d, whereas 28D01-Gal4 labels a neuron subclass indicative of R3m (Omoto, 2018).
In summary, the dorsal view of the EB in conjunction with DN-cadherin immunostaining provide criteria to more definitively identify ring neuron subclasses for future studies. The model organism Drosophila offers unique advantages to examine the circuit motifs that support the broadly relevant computations underlying the processes attributed to the CX; (1) the neurons comprising the CX are spatially and numerically confined, (2) genetic access to label, assess connectivity between, or functionally manipulate, specific neuron types within it, and (3) amenability to electro- or optophysiological recordings, oftentimes in the behaving animal. To fully leverage these advantages, this study provides a systematic description of the ring neuron subclasses comprising the EB, genetic tools to access them, and provide insight into their interactions with other neurons of the CX (Omoto, 2018).
The ellipsoid body (EB) of the Drosophila central complex mediates sensorimotor integration and action selection for adaptive behaviours. Insights into its physiological function are steadily accumulating, however the developmental origin and genetic specification have remained largely elusive. This study identified two stem cells in the embryonic neuroectoderm as precursor cells of neuronal progeny that establish EB circuits in the adult brain. Genetic tracing of embryonic neuroblasts ppd5 and mosaic analysis with a repressible cell marker identified lineage-related progeny as Pox neuro (Poxn)-expressing EB ring neurons, R1-R4. During embryonic brain development, engrailed function is required for the initial formation of Poxn-expressing ppd5-derived progeny. Postembryonic determination of R1-R4 identity depends on lineage-specific Poxn function that separates neuronal subtypes of ppd5-derived progeny into hemi-lineages with projections either terminating in the EB ring neuropil or the superior protocerebrum (SP). Poxn knockdown in ppd5-derived progeny results in identity transformation of engrailed-expressing hemi-lineages from SP to EB-specific circuits. In contrast, lineage-specific knockdown of engrailed leads to reduced numbers of Poxn-expressing ring neurons. These findings establish neuroblasts ppd5-derived ring neurons as lineage-related sister cells that require engrailed and Poxn function for the proper formation of EB circuitry in the adult central complex of Drosophila (Bridi, 2019).
The Drosophila central complex is a composite of midline neuropils that include the protocerebral bridge, the fan-shaped body, the ellipsoid body (EB), the noduli and the lateral accessory lobes. These neuropils are interconnected in a modular way whereby columnar projection neurons leading to and from the central complex connect all its components that are themselves intersected by tangential layers of neural processes, which together form functional modules, each representing a segment of sensory space. Functional studies have identified specific roles for the central complex in higher motor control, courtship and orientation behaviours, visual memory and place learning, as well as sleep, attention, arousal and decision-making (Bridi, 2019).
In contrast to expanding insights into the physiological role of the central complex in regulating behaviour, its developmental origin and genetic specification has largely remained elusive. Earlier work described a primordial central complex at late larval/early pupal stages, which becomes fully formed by 48 h after puparium formation. Genetic studies have identified several alleles of as-yet unidentified genes, as well as orthodenticle, Pax6/eyeless, Pox neuro (Poxn), tay-bridge, roundabout, Pdm3 and semaphorin as genes involved in normal formation of central complex sub-structures (Bridi, 2019).
This study investigate the origin and formation of EB ring neurons R1-R4 in the developing and adult brain of Drosophila. Bilateral symmetric neuroblasts ppd5 were identified in the embryonic procephalic neuroectoderm as founder cells of neuronal progeny that constitute R1-R4 subtypes of tangential ring neurons in the adult EB. Mutant analysis and targeted genetic manipulations reveal a lineage-specific requirement of engrailed (en) and Poxn activity that determines the number and identity of ppd5-derived progeny and their EB ring-specific connectivity pattern in the adult central complex of Drosophila (Bridi, 2019).
Previous studies suggested the Drosophila EB -- as part of the central complex -- develops from precursor cells that differentiate during larval development and during pupal stages generate the EB neuropil. Lineage analysis demonstrates that at least part of its origin can be traced back to the embryonic procephalic neuroectoderm. This study identified Engrailed-expressing neuroblasts ppd5 as embryonic stem cells that give rise to Poxn-expressing progeny, which ultimately differentiate into EB ring neurons. Genetic tracing with en-Gal4 identified R1-R4 ring neurons, suggesting that embryonic neuroblasts ppd5 are the major source of Poxn-expressing progeny leading to EB ring neurons detected in this study. Based on their position, morphology, gene expression patterns and axonal fasciculation, these findings suggest that ppd5-derived larval lineages correspond to previously described larval lineages variously called 'EB-A1/P1', 'DALv2/3', 'MC1' or 'DM'. It was previously demonstrated that these larval lineages express Poxn and give rise to gamma-amino butyric acid (GABA)-ergic ring neurons in the central complex of the adult brain. It therefore is proposed to (re-) name them according to their embryonic origin (Bridi, 2019).
Subclass-specific Gal4 lines together with Poxn expression identifies these lineage-related, ppd5-derived sister cells as R1-R4 ring neurons. Moreover, brain-specific Poxn-Gal4 mediated labelling identifies ring neurons and their axonal projections covering all layers of the EB neuropil, thus suggesting neuroblasts ppd5 give rise to the majority, if not all, of ring neuron subtypes. The ontogenetic relationship between Engrailed-expressing neuroblasts ppd5 and Poxn-expressing EB ring neurons is affirmed by the fact that en-Gal4 and Poxn-Gal4-targeted RNAi-mediated knockdown of Poxn causes similar EB neuropil-specific phenotypes. Together, these data establish that ppd5-derived progeny are clonal units contributing to the EB ring neuron circuitry in the central complex in Drosophila (Bridi, 2019).
How are these units specified? In both insects and mammals, the patterning and specification of neural lineages is regulated by genetic programs from neurogenesis to neuronal differentiation. This study in Drosophila shows that the development and specification of EB-specific circuit elements is likewise dependent on the lineage-specific activity of developmental regulatory genes. Early formation and maintenance of Poxn-expressing ppd5 lineages requires engrailed function as revealed with a deficiency removing both engrailed orthologues, en and invected. Previous studies showed that, engrailed/invected are required for the specification of neuroblast identity in the developing nervous system, suggesting that engrailed is also required for the specification of ppd5. A later, lineage-specific function of engrailed was found in the specification of ring neuron numbers, onsistent with its transient expression in Poxn+ lineages in the embryonic brain but not at later developmental stages nor in adult ring neurons. engrailed codes for a homeodomain transcription factor mediating the activation and suppression of target genes, regulatory interactions that are required for neural lineage formation and specification in the procephalic neuroectoderm. In contrast, no function for Poxn in embryonic brain development has been reported, suggesting that Poxn is only during later stages of development required for lineage and/or neuronal specification in the central brain (Bridi, 2019).
Indeed, experiments identify a postembryonic requirement of Poxn in the specification of ppd5-derived progeny. Previous studies showed that zygotic mutations of Poxn perturb EB neuropil formation, in that presumptive ring neurons are unable to project their axons across the midline and as a consequence, the EB ring neuropil is not formed. In the present study, en-Gal4-targeted knockdown of Poxn reveals Engrailed-expressing cells that project across the midline and form a ring-like neuropil instead of their normal ipsilateral projections to the SP. Significantly, no ppd5-derived GFP-labelled cells were observed that project ipsilaterally towards the SP, neurons that are normally detectable with en-Gal4 targeted GFP expression in the adult brain. Furthermore, en>Poxn-IR-targeted, EB neuron-like projections do not form a torroidal ring but are rather characterised by a ventral cleft. These en>Poxn-IR cells aberrantly retain Engrailed expression even though their axonal projection and connectivity pattern clearly identify them as ring neurons that are normally devoid of Engrailed but instead express Poxn. Together these data suggest that, based on their morphology, Engrailed expression, axogenesis and ring-specific projection patterns, en>GFP cells normally projecting to the SP have been transformed into EB ring neurons in en>mCD8::GFP,Dcr2,Poxn-IR flies (Bridi, 2019).
The resulting additional ring neurons in en>mCD8::GFP,Dcr2,Poxn-IR flies are accompanied with a ventrally open EB ring neuropil. A comparable phenotype is seen in brains of Poxn(757)>Poxn-IR flies which are characterised by an increased number of Poxn(757)-Gal4-targeted ring neurons, suggesting that increasing numbers of EB ring neurons lead to an arch-like neuropil reminiscent of the arch-like EB seen in the majority of arthropods. In support of this notion, previous work has demonstrated that in vivo amplification of ppd5-derived progenitor cells can lead to fully differentiated supernumerary GABAergic ring neurons that form functional connections often characterised by a ventrally open EB ring neuropil. Together, these data identify differential roles of Poxn activity during neuroblast lineage formation, in that Poxn is required for cell identity determination of ppd5-derived progeny, as well as for the specification of cell numbers and terminal neuronal projections of EB ring neurons (Bridi, 2019).
These Poxn functions in ppd5-derived brain lineages are reminiscent of Poxn activity in the peripheral nervous system (PNS) which mediates the specification of sensory organ precursor (SOP) cell lineages giving rise to external sense organs, the tactile and gustatory bristles, respectively. In these SOP lineages, differential Poxn activity determines progeny fate between chemosensory (gustatory) or mechanosensory (tactile) neuronal identities. Furthermore, SOP lineage-specific Poxn function specifies the number of these neurons and their connectivity pattern. The apparent functional commonalities between Poxn-mediated specification of ppd5 neuroblast-derived lineages in the brain and SOP lineages in the PNS, suggest that evolutionarily-conserved mechanisms underlie the development and specification of clonal units as cellular substrates for neural circuit and sensory organ formation (Bridi, 2019).
The cytoarchitecture of both the insect and mammalian brain are characterised by neural lineages generated during development by repeated asymmetric divisions of neural stem and progenitor cells. These ontogenetic clones are thought to constitute building blocks of the insect and mammalian brain. In support of this notion, lineage-related progeny constitutes sets of circuit elements of the mushroom bodies and antennal lobes in Drosophila. Clonal relationship also characterises the lineage-dependent circuit assembly in the mammalian brain, where stem cell-like radial glia give rise to clonally-related neurons that synapse onto each other, as has been shown for cortical columns and GABAergic interneurons in the neocortex and for striatal compartments of the basal ganglia. The current study in Drosophila shows that a pair of bilateral symmetric, engrailed-expressing embryonic stem cells, neuroblasts ppd5, give rise to R1-R4 subtypes of tangential ring neurons that contribute to the layered EB neuropil. Thus, ppd5 neuroblast lineages constitute complete sets of circuit elements intrinsic to the adult central complex in Drosophila (Bridi, 2019).
It has been suggested that clonal expansion of neural lineages contributed to the evolution of complex brains and behaviours. Key to this hypothetical scenario are ancestral circuit elements in the form of genetically encoded stem cell-derived clonal units, like the ones described in the current study. In such a scenario, lineage-related ancestral circuit elements might have been multiplied and co-opted or diversified during the course of evolution. Multiplication and co-option have been suggested for the evolution of the multiple-loop architecture of the basal ganglia that allows processing of cognitive, emotional and motor information. In line with this hypothesis, quantitative control of the transcription factor Prospero is sufficient to cause clonal expansion of ring-neuron circuitry in Drosophila (Shaw, 2018), which has been implicated in cognitive and motor information processing and resembles extensive correspondences to vertebrate basal ganglia, ranging from comparable developmental genetics to behavioural manifestations and disease-related dysfunctions (Bridi, 2019).
In contrast to multiplication and co-option, the diversification of stem cell lineages can equally contribute to neural circuit evolution. The current results identify differential and tightly regulated spatio-temporal functions of engrailed and Poxn that lead to the differentiation of ppd5 progeny into hemi-lineage specific identities in the adult brain. Loss of engrailed affects the formation of precursors cells, whereas its lineage-specific knockdown affects the number of Poxn expressing ring neurons. Correspondingly, en-Gal4-driven lineage-specific knockdown of Poxn results in an identity transformation of Engrailed-expressing neurons in the adult brain in that they no longer project to the SP, but instead reveal an EB ring-neuron identity. These data indicate a binary switch of hemi-lineage identities as the result of a feed-forward mechanism between engrailed and Poxn. engrailed may activate transcription (directly or indirectly) of Poxn, which in turn represses engrailed to permit differentiation of R1-R4 neurons, thereby regulating the specification of neuronal identities in ppd5 hemi-lineages. This hypothesis is consistent with lineage tracing and MARCM experiments, as well as the transient expression of engrailed in embryonic ppd5 lineages but not in adult EB ring neurons. However, further studies are required to elucidate the nature and extend of these putative regulatory interactions between Engrailed and Poxn (Bridi, 2019).
In summary, these findings establish a causal relationship between a pair of bilateral symmetric embryonic stem cells, neuroblasts ppd5 and the lineage-related assembly of their EB ring neuron progeny as structural units of the central complex in Drosophila. Based on these observations it is proposed that amplification and diversification of ontogenetic clones together with the repurposed use or exaptation of resulting circuitries, is a likely mechanism for the evolution of complex brains and behaviours (Bridi, 2019).
The recent discovery of the head-direction (HD) system in fruit flies has provided unprecedented insights into the neural mechanisms of spatial orientation. Despite the progress, the neural substance of global inhibition, an essential component of the HD circuits, remains controversial. Some studies suggested that the ring neurons provide global inhibition, while others suggested the Δ7 neurons. The present study provides evaluations from the theoretical perspective by performing systematic analyses on the computational models based on the ring-neuron (R models) and Δ7-neurons (Delta models) hypotheses with modifications according to the latest connectomic data. Four tests were performed: robustness, persistency, speed, and dynamical characteristics. It was discovered that the two models led to a comparable performance in general, but each excelled in different tests. The R Models were more robust, while the Delta models were better in the persistency test. A hybrid model was tested that combines both inhibitory mechanisms. While the performances of the R and Delta models in each test are highly parameter-dependent, the Hybrid model performed well in all tests with the same set of parameters. These results suggest the possibility of combined inhibitory mechanisms in the HD circuits of fruit flies (Chang, 2023).
The brain perceives visual information and controls behavior depending on its underlying neural circuits. How UV information is represented and processed in the brain remains poorly understood. In Drosophila melanogaster, UV light is detected by the R7 photoreceptor that project exclusively into the medulla layer 6 (M(6)). This study imaged 28,768 single neurons and identified 238 visual projection neurons linking M(6) to the central brain. Based on morphology and connectivity, these visual projection neurons were systematically classified into 94 cell types belonging to 12 families. Three tracts connected M(6) in each optic lobe to the central brain: One dorsal tract linking to the ipsilateral lateral anterior optic tubercle (L-AOTU) and two medial tracts linking to the ipsilateral ventral medial protocerebrum (VMP) and the contralateral VMP. The M(6) information was primarily represented in the L-AOTU. Each L-AOTU consisted of four columns that each contained three glomeruli. Each L-AOTU glomerulus received inputs from M(6) subdomains and gave outputs to a glomerulus within the ellipsoid body dendritic region, suggesting specific processing of spatial information through the dorsal pathway. Furthermore, the middle columns of the L-AOTUs of both hemispheres were connected via the intertubercle tract, suggesting information integration between the two eyes. In contrast, an ascending neuron linked each VMP to all glomeruli in the bulb and the L-AOTU, bilaterally, suggesting general processing of information through the ventral pathway. Altogether, these diverse morphologies of the visual projection neurons suggested multi-dimensional processing of UV information through parallel and bilateral circuits in the Drosophila brain (Tai, 2020).
Animal behavior is guided by the brain. Therefore, adaptations of brain structure and function are essential for animal survival, and each species differs in such adaptations. The brain of one individual may even differ between life stages, for instance, as adaptation to the divergent needs of larval and adult life of holometabolous insects. All such differences emerge during development, but the cellular mechanisms behind the diversification of brains between taxa and life stages remain enigmatic. This study investigated holometabolous insects in which larvae differ dramatically from the adult in both behavior and morphology. As a consequence, the central complex, mainly responsible for spatial orientation, is conserved between species at the adult stage but differs between larvae and adults of one species as well as between larvae of different taxa. This study used genome editing and established transgenic lines to visualize cells expressing the conserved transcription factor Retinal homeobox, thereby marking homologous genetic neural lineages in both the fly Drosophila melanogaster and the beetle Tribolium castaneum. This approach allowed comparison of the development of homologous neural cells between taxa from embryo to the adult. Complex heterochronic changes were found including shifts of developmental events between embryonic and pupal stages. Further, the first example of sequence heterochrony in brain development was found, where certain developmental steps changed their position within the ontogenetic progression. Through this sequence heterochrony, an immature developmental stage of the central complex gains functionality in Tribolium larvae (Farnworth, 2020).
Feeding decisions are fundamental to survival, and decision making is often disrupted in disease. This study shows that neural activity in a small population of neurons projecting to the fan-shaped body higher-order central brain region of Drosophila represents food choice during sensory conflict. Food deprived flies made tradeoffs between appetitive and aversive values of food. An upstream neuropeptidergic and dopaminergic network was identified that relays internal state and other decision-relevant information to a specific subset of fan-shaped body neurons. These neurons were strongly inhibited by the taste of the rejected food choice, suggesting that they encode behavioral food choice. These findings reveal that fan-shaped body taste responses to food choices are determined not only by taste quality, but also by previous experience (including choice outcome) and hunger state, which are integrated in the fan-shaped body to encode the decision before relay to downstream motor circuits for behavioral implementation (Sareen, 2021).
Flexible behaviors over long timescales are thought to engage recurrent neural networks in deep brain regions, which are experimentally challenging to study. In insects, recurrent circuit dynamics in a brain region called the central complex (CX) enable directed locomotion, sleep, and context- and experience-dependent spatial navigation. This study describes the first complete electron-microscopy-based connectome of the Drosophila CX, including all its neurons and circuits at synaptic resolution. This study identified new CX neuron types, novel sensory and motor pathways, and network motifs that likely enable the CX to extract the fly's head-direction, maintain it with attractor dynamics, and combine it with other sensorimotor information to perform vector-based navigational computations. Numerous pathways were identified that may facilitate the selection of CX-driven behavioral patterns by context and internal state. The CX connectome provides a comprehensive blueprint necessary for a detailed understanding of network dynamics underlying sleep, flexible navigation, and state-dependent action selection (Hulse, 2021).
Recent physiological and anatomical studies at the light and EM level have highlighted strong links between circuit structure and function in the adult fly central brain. These links have proven to be valuable both for generating hypotheses and for experimentally testing them. This recent history gives reason to expect that connectomics will continue to accelerate studies of circuit function. CX circuits, in particular, are thought to be involved a wide variety of flexible, context-dependent behaviors. This work provides a detailed description of CX neuron types and circuits, with a particular focus on extracting and examining network motifs from the perspective of what is currently known about CX circuit function and CX-mediated behavior. Many repeating motifs were found, raising the possibility that an understanding of the computational roles of some of these may generalize to others. Some of these motifs match those that have been proposed previously to implement ring attractors for head direction computation. Others seem suitable for gain control in multiple structures. And still others seem to be ideal for vector computations that would be required for robust navigational behaviors. This study found that information from the CX's output neuron types is broadcast through fairly segregated pathways that are distributed across the brain, not just to premotor centers but to sensory regions and, importantly, back into the CX itself. In the sections that follow, some functional implications of these motifs and of other results from this analyses are described. These functional implications are derived not just from connectomic analyses and the historical precedent of structure predicting function in many different neural circuits, but also on published physiological and behavioral studies. Testing the hypotheses are outlined below will require a long series of functional experiments, but the connectome provides an invaluable guide for the design and prioritization of such experiments (Hulse, 2021).
The value of EM-level connectomes in understanding the function of neural circuits in small and large brains is widely appreciated. Although recent technical advances have made it possible to acquire larger EM volumes and improvements in machine learning have enabled high-throughput reconstruction of larger neural circuits, the step from acquiring a volume to obtaining a complete connectome still requires considerable human proofreading and tracing effort. As part of this analysis of the CX connectome, it was found that although increased proofreading led to an expected increase in the number of synaptic connections between neurons, it did not necessarily lead to significant changes in the relative weight of connections between different neuron types. While it is important to note that comparisons were made between the hemibrain connectome at fairly advanced stages of proofreading in the CX, the results do suggest that it may be possible to obtain an accurate picture of neural
circuit connectivity from incomplete reconstructions. It may be useful for future large scale
connectomics efforts to incorporate similar validation steps of smaller sample volumes into
reconstruction pipelines to determine appropriate trade-offs between accuracy and cost of
proofreading. Connectivity and neural processing beyond the typical synapse. Although this study provides a detailed description of the CX's hundreds of neuron types, recurrent networks and pathways, there is still more information that could be extracted from the CX connectome. The CX is innervated by a large number of modulatory and peptidergic neurons, many unidentified and almost all of unknown function. These neurons likely significantly modulate the function of recurrent networks in ways that few studies address. Knowing their identities - whether by matching LM images of known neuron types to their EM counterparts in the hemibrain or by advances in machine-learning based identification of neuromodulator/neuropeptide and receptor types - would help guide circuit studies into context- and internal-state-dependent processing in the CX (Hulse, 2021).
A large number of CX neuron types that make T-bar and E-bar synapses in CX structures also send projections to other structures in which they make no such synaptic connections. This study investigated these projections in more detail and consistently found dense core vesicles in these otherwise nearly synapse-free processes. Although the involvement of some of these neuron types, for example PFGs neurons, in sleep-wake circuits suggests a plausible scenario for state-dependent modulation of CX circuits, such explanations are not easily available in all cases (Hulse, 2021).
It is important to note that use of relative weights to assess synaptic strength was informed by observed correlations between synapse counts and the area of synaptic contact in larval Drosophila, and the dependence of synaptic strength on synaptic surface area, at least in the mammalian neocortex. It is expected that relative weights provides only an approximate measure of true functional strength. Further, synapses across the Drosophila brain undergo structural changes depending on the time of day, sleep, activity and the animal's specific experiences; properly accounting for the impact of such factors on connectivity patterns would require comparisons across multiple connectomes. Also, as previously discussed, the hemibrain connectome does not capture glial networks or gap junctions. Despite all these limitations, the identification of chemical synapses between CX neurons and examining their relative weight based on synapse counts allowed extraction of network motifs that make strong predictions about function (Hulse, 2021).
Many flexible, goal-driven behaviors unfold over longer durations than fast reflexive responses and are robust to the temporary loss of sensory cues directly associated with the goal. Desert ants, for example, use path integration to return to their nests after long foraging trips in relatively featureless landscapes, and mammals use working memory to perform delayed match-to-sample tasks. For such behaviors, brains are believed to rely on intermediate representations and neural dynamics that persist or update even in the absence of direct sensory inputs. Such persistent representations have long been believed to be generated, updated and maintained by recurrent attractor networks. These more abstract intermediate representations also enable disparate sensory and self-motion cues of different modalities to be registered to a shared reference frame. A path integrating ant, for example, may use such a representation to register cues from polarized light, visual optic flow and proprioception and all Diptera likely need to register visual and haltere input, as flesh flies do. Ultimately, information in these reference frames must still be dynamically converted to a body-centered reference frame for situation-appropriate action. Decades of experimental work in a variety of species have led theorists to propose gain fields for the implementation of such coordinate transformations, but the predicted neural circuit connectivity has not been directly identified. In addition, for an animal to learn from experience, any past associations of the current context with good or bad outcomes must be recalled and used to modify neural dynamics at the level of such intermediate representations, raising computational questions that have been explored in the field of reinforcement learning. The repertoire of flexible navigational behaviors that insects display suggests that their small brains may solve many of these computational challenges. Further, insect circuits may have evolved solutions to these problems that resemble those proposed by theorists to account for neural response properties in mammalian circuits (Hulse, 2021).
Flies in particular use short-term memory to orient towards the last-known positions of attractive visual beacons that have disappeared. They learn about their body size and use that information when attempting to cross gaps. They learn to avoid heat punishment by using visual patterns around them to orient to safety. Although they are not central place foragers like bees and ants, they are capable of returning to a spot of food even when exploring their surroundings in darkness, and of remembering visual landmarks to navigate to safe spots in an otherwise hostile open space. The CX is thought to be essential for many of these behaviors. In the sections that follow, how the patterns of connectivity revealed by the CX connectome may enable the neural dynamics, coordinate transformations, and learning-induced changes in action selection associated with meeting the computational challenges of some of these behaviors will be discussed (Hulse, 2021).
Head direction representations enable an animal to flexibly rely on a variety of different cues, including self-motion, to orient. Work in Drosophila and other insect species has established that the CX builds a stable head direction representation using information from ring neurons, which convey directional sensory cues, such as polarized light, visual landmarks, and wind direction. In Drosophila visual head direction information reaches the CX via the anterior visual pathway, which appears to convey different visual information in separate, parallel 'channels'). Some 'channels' of this pathway have been characterized functionally, while the function and sensory tuning of other groups of neurons remains elusive. For example, most of the ring neurons (and their inputs) in the superior bulb (BU) are spatiotemporally tuned to visual features with some degree of orientation preference and the pathway through the anterior BU appears to be dedicated to polarization signals. In contrast, little is known about the role of ring neurons that get their inputs in the inferior BU in informing the head direction representation. Wind stimuli reach the compass circuitry through separate input pathway via the LAL and it is unknown whether other sensory modalities are conveyed through this route (Hulse, 2021).
Connectivity-based analysis suggests that there are 20 ring neuron types, 18 of which receive inputs via the anterior visual pathway. In contrast, an anatomical and developmental
characterization of ring neurons found only 11 distinct morphological types. Notably, the current connectome-based typing likely represents a subdivision of the previously suggested types rather than a drastic reorganization. Given that past neurophysiological studies have only tested tuning to a relatively small number of sensory stimuli, it remains to be seen how many functionally distinct input types exist (Hulse, 2021).
The connectome reveals mechanisms by which sensory stimuli are integrated to inform the fly's head direction estimate. The findings suggest that different cues exert differing levels of influence on the ellipsoid body-protocerebral bridge-gall neuron (EPG) neurons that carry the head direction representation. each EPG (compass) neuron arborizes in a single EB sector ('wedge'). A prioritization of certain sensory cues is reflected in the relative locations of synaptic input from different sensory streams onto the EPG dendrites in the EB, in the relative weight of those inputs, in the feedback that some ring neuron pathways receive from the EPG neurons, and in the relative weight of across-type inhibition from some ring neuron types onto others. The implicit hierarchy of ring neuron inputs to the fly compass indicates that the EPG head direction representation preferentially tethers to environmental references that are likely to indicate a global direction (Hulse, 2021).
Bright visual landmarks, for example, may originate from celestial bodies such as the sun, but they could also be generated by local terrestrial objects. By contrast, a polarization pattern in the sky, if available, represents a reliable global reference, which might explain the observed circuit motifs that suggest the preferential use of polarization cues to update the fly's head direction representation. However, the relatively high connection strength between ER4m and EPG neurons may also arise from this fly not being exposed to polarized light stimuli. Such deprivation could have prevented these connections from being subjected to the synaptic depression that other visual pathways may have experienced (but note that there is no evidence yet for long-term structural changes at any of these synapses) (Hulse, 2021).
Particularly when navigating over long distances, skylight cues allow the head direction representation to be tethered to global landmarks such as the sun and to the polarized light patterns of the sky. Indeed, polarized light e-vector information has long been thought to be important for the determination of sky-compass-based head direction in many insects. A dorsal band of the insect eye called the dorsal rim area is structurally specialized for the detection of polarized light e-vectors in the sky. Despite their comparatively small dorsal rim area, flies can also use polarized light cues to determine their heading. Sensory information about the celestial polarization pattern reaches the Drosophila CX1 via a dedicated pathway to the ER4m neurons. Although only 5 ER4m neurons from each hemisphere show strong tuning to e vector orientation, this tuning collectively covers a large part of the 180° range of possible e vector orientations. However, in contrast to the position of the sun, the 180° symmetric polarized light patterns do not immediately provide the ability to distinguish a specific direction from one directly opposite to it (Hulse, 2021).
The CX connectome suggests that the fly's compass may have evolved a solution to this problem. For the polarization-tuned ER4m neurons, it was observed that synapse numbers to EPG neurons varied smoothly along the circumference of the EB, but with mirror-symmetric profiles for ER4m neurons from the left and right hemisphere, respectively. If synapse counts correlated with synaptic strength, this would result in stronger connections from ER4m neurons of the left hemisphere for EB wedges on the right half of the EB and stronger connections from the right hemisphere to the left half of the EB. This structure was even more clearly revealed when the pairwise correlation of EPG neurons according to their ER4m inputs: all EPG neurons on the right side of the EB were positively correlated with each other, while being anticorrelated with those on the left side, and the inverse pattern was observed for the left EPG population. Given that polarized light has a 180° symmetry, this connectivity pattern0 may allow the fly to generate a complete, 360° head direction representation from polarized light input (Hulse, 2021).
One possible mechanism by which this could be achieved hinges on the geometry of the fly's polarization sensors in the dorsal rim area and how it interacts with the natural polarization5 pattern of the sky. The receptive fields of the fly's polarization sensors in the left and right eye face the contralateral celestial hemisphere and tile a small strip along the rostral-caudal axis of the fly. Along this strip tuning to e-vectors varies continuously and covers nearly the full 180° range of possible e-vector orientations. Given the naturalistic celestial polarization pattern, the geometry of the slightly curved receptive field 'strip' might act as a rough 'matched filter', such that neurons in the dorsal rim area on the side of the sun (facing the contralateral sky) are systematically more strongly activated than those on the side facing away from the sun. The all-to-all inhibition between left and right ring neurons in the EB may then systematically select either the left or the right ring neurons to tether the head direction depending on which direction the fly is facing relative to the current position of the sun, thus disambiguating the 180° mirror 06 symmetry in the polarization signal (Hulse, 2021).
In locusts, TL-neurons, homologs of the fly's ring neurons, and (protocerebral bridge) PB neurons have been shown to exhibit matched-filter like tuning to the full-sky polarization patterns generated by the sun. A corollary of these studies is that individual TL neurons in the locust have receptive fields that span large parts of the sky. Indeed, the area of the sky that is sampled by photoreceptors in the dorsal rim area is significantly larger in locusts compared to flies, and it is plausible that further sensory processing along the anterior visual pathway toward the CX differs between species as well, in which case different insects might employ different strategies for disambiguating polarized light stimuli (Hulse, 2021).
The mechanism described above would not require that the sun be directly visible, but it might still be beneficial to have ring neurons that have multimodal tuning to both polarized light and sun-like stimuli. Such cells have been described in other insects. While this has yet to be demonstrated experimentally, multimodal ring neurons tuned to both visual features and polarized light e-vector orientation may also exist in flies. Hardcastle and colleagues report polarization-tuned neurons in the superior bulb (Hardcastle, 2021), where tuning to bright features has also been observed (Hulse, 2021).
Besides visual cues, mechanosensory wind stimuli can drive the fly's head direction system in the EB. Information about wind direction reaches the EB via ring neurons that arborize in the lateral accessory lobe (LAL). Wind tuning has been demonstrated in both ER3a and ER1 neurons, although only ER1 neurons were able to update the head direction estimate. Analysis of the connectome suggests that both the ER1 and ER3a neuron populations consist of multiple types with distinct inputs. Only ER1_b and ER3a_b neurons got strong inputs from cells that are believed to be the wind-sensitive LAL138 WL-L) and WPN neurons. The connectivity of these two ring neuron types onto EPG neurons, with strong connections from ER1_b but no connections from ER3a_b neurons, is consistent with the observation that ER1 but not ER3a neurons can drive the head direction representation. It is also noteworthy that in the EB, ER1_b neurons deviate from the within-type all-to-all inhibition motif that all other ring neurons show in the EB. A possible reason is that an accurate mapping from ER1 neuron activity to a head direction representation requires pooling information from multiple ring neurons at once. This analysis also suggests that ER1_b input to the EPG neurons is suppressed by ER1_a neurons, but it is presently unknown whether ER1_a neurons also encode wind direction or whether these neurons are tuned to a different stimulus. ER1_a and ER1_b inputs in the LAL are distinct and unfortunately little is known about the inputs ER1_a receives (Hulse, 2021).
Whether over short or long distances, olfactory cues are strong indicators of good food sources. Flies are known to fly upwind when they encounter an appetitive odor, a strategy also employed by other insects navigating to an odor source. A robust navigational strategy would allow an insect to maintain the same heading using other cues even if the wind were to transiently die down. Based on the proximity of different ring neuron inputs to the putative spike initiation sites of EPG neurons, the head direction representation is likely to be strongly tethered to wind direction by input from ER1_b neurons (and perhaps also ER1_a neurons, although their function is currently unknown). If visual cues are flexibly mapped onto head direction representation using this wind direction input as a reference, the EPG compass could allow the fly to preserve its heading using those cues even in the absence of wind (Hulse, 2021).
The relative importance of synapse location in determining the cues to which the EPG compass tethers will only be clear with in-depth investigations of EPG neuron biophysics. More broadly, future studies of ring neuron and EPG interactions should provide an implementation-level understanding of a variety of computations related to dynamic multisensory integration and the resolution of conflicts between cues of different reliability (Hulse, 2021).
Recent studies have proposed an important role for fast-timescale, short-term plasticity of synaptic connections between ring and EPG neurons in enabling the EPG compass to quickly adapt to different sensory settings. The connectome suggests that the ring neuron network may also preselect more salient cues for the compass through all-to-all inhibitory connectivity within each type. The precise impact of all-to-all inhibition on the ring neuron network's preprocessing of localizing cues that are used to generate the head direction representation will depend on the timescale of the inhibitory conductance, which is as yet unknown. If the inhibitory conductance is fast, all-to-all inhibition would create winner-take-all dynamics in which a few ring neurons receiving the strongest inputs effectively shut down all other ring neurons. In sensory settings characterized by a single dominant sensory cue, such as the sun or polarized light e-vector orientation in a desert landscape during the day, only a handful of ring neurons with appropriately tuned receptive fields would be active within each type for any particular head direction. Fast all-to-all inhibition in this setting would enhance the activity of the most dominant ring neuron within each type and would minimize the impact of noise from the others, which might otherwise disrupt the stability of the EPG compass (Hulse, 2021).
However, stable and unique heading representations are also generated within scenes with multiple strong cues, such as within a forest or when walking on the branches of a tree, as long as the two-dimensional arrangement of cues allows for a unique determination of heading. In the presence of multiple salient cues, it would be expected that multiple ring neurons would respond with comparable strength for any given heading of the fly and several to respond weakly to any additional visual cues ('clutter') in the scene. Although fast all-to-all inhibition in this scene would still filter out these weaker responses, it could allow multiple, strongly responsive ring neurons to remain active for each heading. A slower inhibitory conductance would, in this situation, induce oscillatory spiking dynamics between these multiple 'winners', a situation that has been referred to as 'winnerless competition', and that has been suggested to be useful for sequential memory (Hulse, 2021).
Ring neuron responses are not determined purely by sensory cues. These neurons appear to be modulated by state, maintain a baseline level of activity, and may be biophysically configured to support oscillatory population activity linked to sleep need. The many additional inputs that many ring neuron types receive in the BU provide clues as to how the activity of these neurons might be modulated by the fly's behavior and its internal state. The visually-tuned superior BU ring neurons primarily receive input from a large interhemispheric anterior optic tubercle (AOTU) neuron, which may mediate dynamic stimulus selection through delayed contralateral inhibition. The same group of ring neurons also shows changes in activity with the fly's behavioral state (flight versus walking) and indeed many of these neurons receive input from the dopaminergic ExR2 neuron that has been linked to changes in the fly's motor activity. A different set of ring neurons that receive their inputs in the inferior BU receives strong inputs from two ExR neurons -ExR1 and ExR3- that have been linked to the control of sleep, and may gate sensory stimuli according to the fly's behavioral state (Hulse, 2021).
The fly's head direction representation tethers to directional sensory cues conveyed by ring neurons, but is also updated by self-motion cues and is maintained across periods of immobility. Strong experimental and theoretical evidence suggests that the representation is maintained by a ring attractor network, which includes at least some of the recurrently connected columnar neurons that link the EB and the PB: the EPG, PEN_a, PEN_b and PEG 24 neuron types. The patterns of connectivity between individual neurons of these types are consistent around the entire EB and across the length of the PB. Similarly, the broad connectivity patterns of individual neurons within these types to tangential neurons -Δ7 neurons in the PB and different types of ring neurons in the EB- are similar across these structures. Notably, however, two distinct classes of neurons (EPGt neurons and the P6-8P9 neurons) only innervate the edges of the network; both innervate the outer glomeruli of the PB, and the EPGt neurons also innervate the corresponding wedges in the EB. These types may help to stitch together what might otherwise be a discontinuity in the ring attractor network. Indeed, the EPGt neurons in the left and right PB arborize in wedges in the EB that lie directly in between the wedges occupied by the EPG neurons on either side of the potential discontinuity (in PB glomeruli 1 and 8). The EPGt neurons may therefore represent angular positions halfway between the edge angles, bridging the gap. It is noted, however, that the Δ7 neurons and the P6-8P9 neurons that output in these outer glomeruli each receive unique input from different sets of EPG neurons, making it hard to assign a clear corresponding EB angle to glomerulus 9 (Hulse, 2021).
The hemibrain connectome further allowed identification of several neuron types and connectivity motifs that are likely involved in the network's function, but whose roles await experimental investigation. Many of these additional types are tangential neurons. Some of them appear to provide additional sources of inhibition, potentially regulating overall network activity. The ER6 neurons, for example, receive input from the EPG and PEG neurons in the GA and send outputs to the PEG, EL and PEN_b neurons in the EB, potentially modulating the EPG-to-PEG-to-PEN_b-to-EPG feedback loop. Furthermore, many of the ExR neurons make connections to and receive input from EB columnar neurons. The PB receives neuromodulatory input from the dopaminergic LPsP neurons and the octopaminergic P1-9 neurons. The LPsP neurons may enable changes in synaptic strength in the PB. Such plasticity in the PB has been suggested to allow flies to calibrate their directed movements to their body size. In sensory brain regions, octopaminergic neurons are known to modulate neuronal conductances based on the fly's behavioral state, and it is an open question whether the P1-9 neurons play a similar role in the PB. Notably, the Δ7 neurons connect recurrently to each other in the PB, but the function of this recurrence is unknown. One possibility is that recurrent Δ7 connections may increase the stability or robustness of the ring attractor network (Hulse, 2021).
Two additional classes of columnar neurons also contact the ring attractor network: EL and IbSpsP. The connectivity pattern of EL neurons in the EB is remarkably similar to that of the EPG neurons, but their function is unknown. In the PB, the IbSpsP neurons bring input into specific glomeruli from regions associated with premotor functions, potentially allowing them to exert an influence on the dynamics of the bump in the PB (Hulse, 2021).
The ring attractor network described above generates a single activity bump in the EB that encodes the fly's head direction. The connectome allowed following this activity bump through the CX as it gets duplicated, reformatted, recombined, and, finally, read out. In the process, network motifs were discovered that seem ideally suited for performing vector computations. These motifs place constraints on the network's computational capacity and inspire conceptual models for how the network might function. First is described how the activity bump is forwarded from the EB to the PB, where it is duplicated and reformatted into a sinusoidal profile. In subsequent sections it is considered how the fan-shaped body (FB) network may recombine these bumps 76 to perform vector computations in support of goal-directed behavior. 77 78 The EPG population divides the EB into 16 'wedges', suggesting that the fly's head direction system samples angular space at 22.5° intervals. Importantly, this does not mean that the system cannot resolve head directions at resolutions higher than 22.5°, since the differential activation of columnar neurons with distinct directional tunings can effectively represent any arbitrary angle within the 360° around the fly. From the EB, EPG neurons convey the hΔbump to both the left and right PB, generating two bumps that are sampled at approximately 45° intervals. Due to the EPG projection pattern, there is a 22.5° shift in the directional tuning between EPG neurons in left and right PB, as recently confirmed by physiological recordings. Importantly, the bumps in the left and right PB still encode the same head direction, but do so using sets of neurons whose sampling of angular space is shifted by 22.5° (Hulse, 2021).
Within the PB, FB columnar neurons inherit a head direction bump directly from EPG neurons and indirectly through Δ7 neurons. The Δ7 populations appear ideally suited to reformat the EB bump into a sinusoidal profile, regardless of its original shape (Hulse, 2021).
Individual Δ7 neurons provide output to 2-3 PB glomeruli spaced ~180° apart (that is, separated by 7 glomeruli). Between these axonal compartments are dendritic segments whose EPG input weight is well fit by a sinusoid, suggesting that individual Δ7 neurons should have a sinusoidal tuning curve. Assuming the Δ7 population uniformly samples angular space (for example, with a 45° sampling interval), this would manifest as two sinusoidal bumps across the PB, one in the left PB and one in the right PB. Furthermore, recurrent Δ7 connections may enforce a sinusoidal activity pattern on the Δ7 population itself, which could improve the ability of Δ7 neurons to reformat the activity bump into a sinusoidal profile before passing it on to PB-FB types. The Δ7 population provides input to ~10 types of PB-FB neurons, effectively duplicating the activity bump in the process. As discussed below, this duplication may allow the FB network to recombine bumps in a way that implements a compact vector calculator (Hulse, 2021).
Why might the Δ7 population reformat the activity bump into a sinusoidal shape? Perhaps because sinusoids are a particularly suitable representation for vector-based computations, since the sum of any two sinusoidal of equal frequency is also a sinusoid. One way of schematizing this process is to use phasor diagrams. Viewed in this way, the sinusoidal activity bumps become vectors whose magnitude reflects bump amplitude and whose angular position indicates bump phase, with each phase mapping to an allocentric direction. Adding vectors is equivalent to adding sinusoidal activity profiles (Hulse, 2021).
Path integration is a canonical vector-based navigation strategy used by a diverse array of both flying and walking animals, potentially including Drosophila. In its most basic form, 2D path integration requires that an animal keep track of its direction and distance relative to a stored goal location, such as a food source or nest, often without the use of external landmarks. The direction and distance to the goal location is thought to be computed through the integration of self-motion signals and stored as a 'home vector'. To return to the goal location, animals are thought to generate appropriate motor commands by comparing their current heading to the stored home vector. While many insects are thought to generate and use visual snapshots of their surroundings to guide return trips, and while such visual homing may involve the MB, this study focused on how a home vector might be constructed and read out in the CX using only a stable head direction signal, a situation that can arise in featureless landscapes or in darkness. Although there is as yet no definitive evidence that the CX is used for path integration, in the next few subsections, it is shown how a network built from FB-inspired circuit motifs could compute a translational velocity vector in an allocentric reference frame whose integration would yield a home vector. It is noted that the framework for vector computations that is describe below is likely to be useful for a much broader array of behaviors involving oriented action selection (Hulse, 2021).
The potential for vector computations in the FB PFN neurons serve as the major columnar input to the FB network. The +/- 45° phase shift that is characteristic of all PFN neuron types implies that activity bumps from the left and right PB would end up ~90° apart in the FB. The amplitude of these activity bumps is likely to be strongly influenced by the different inputs that PFN neurons receive through their lateralized projections in the NO , setting up the possibility of bump-based vector computations in the FB. The PEN_a neurons, which are conjunctively tuned to head direction and angular velocity, perform a similar computation in the EB by providing phase-shifted input to the EPG neurons, thereby updating the position of the EPG bump when the fly turns. Inside the FB, vector computations fed by phase-shifted PFN bumps whose amplitudes are controlled by different conjunctive signals could ultimately drive PFL neuron types to generate appropriate motor commands, an algorithmic idea for which an FB implementation was first proposed (Hulse, 2021).
Although the intra-FB columnar network is highly recurrent, much of it is built from a limited number of circuit motifs. These motifs serve as the backbone of a 2D grid in which activity bumps are constrained to either maintain their phase (that is, maintain their column) while moving across layers, or shift phase by 180° (that is, shift by half the width of the FB). While some pathways directly connect PFN neurons to output pathways, such as those involving PFL neurons, many more pathways run through this 2D grid. Thus, the network has depth, providing multiple layers with which to process activity bumps. In addition, a large number of tangential neuron types selectively innervate different layers of the FB, suggesting that the FB's vector computations are influenced by context and internal state. The sections that follow, draw from published experimental and theoretical work to explore the navigational implications of PB-FB neuron phase shifts. Importantly, for the purposes of discussion, it is assumed that the magnitude of PFN phase shift is precisely 90°, a simplifying assumption about symmetry in the circuit that ignores the type-to-type variability in estimated phase shifts across PFN types, the functional significance of which remains unknown. Similarly, all LNO types are assumed to be excitatory, but the proposed conceptual models could be built from inhibitory LNO types as well. Finally, while the columnar structure of the various dΔand hΔtypes show considerable variability, it is assumed that these neurons can either maintain the phase of an FB bump or shift it by 180° (Hulse, 2021).
Despite the PFN phase shifts, the two 90°-separated activity bumps arising from a single PFN type cannot propagate independently through the FB network, because nearly all single neurons and neuron types that are postsynaptic to PFN neurons sample from left and right populations equally. Instead, each postsynaptic FB type likely sums the bumps from the left and right PFN populations, indicating that each postsynaptic type represents a single, summed bump that will propagate through the FB network (Hulse, 2021).
Much like the PEN neurons, the PFN neurons innervating the left PB project to the right NO and neurons innervating the right PB project to the left NO, where they receive input from various LNO types. One potential function that this differential NO input to the left and right PFN populations could serve is to produce a new, transformed directional representation that could take on angles +/- 45° around the fly's instantaneous head direction. For example a strong excitatory input to the right nodulus would increase the bump amplitude of the left PFN population relative to the right PFN population. In turn, the summation of these two bumps by a postsynaptic neuron type in the FB would result in a new bump that lies closer to that of the left PFN population (Hulse, 2021).
Phasor diagrams of this process are given in the paper. Critically, because these vectors can only take on positive values (firing rates above 0), such differential input could only shift the resulting vector's phase by +/- 45° around the fly's instantaneous head direction. What might this transformed directional representation encode? The answer likely depends on the nature of the input that PFN neurons receive from LNO types. Recent work in Drosophila has shown that some PFN neurons show differential activity in the NO that reflects the fly's turning behavior during flight, but the nature of the rotational velocity signal remains to be determined. Two hypothetical scenarios are highlighted that differ in the specific information carried by PFN neurons (Hulse, 2021).
Directly wiring sensors to actuators in different ways can, in principle, allow a simple agent to display a variety of behaviors. But flies, like most animals, have to deal with an additional complication: some of their sense organs are on body parts that are different from those that enable them to move. PFN phase shifts could enable coordinate transformations, such as converting the allocentric head direction representation into an allocentric body direction representation. Flies make head movements that change their head-body angles by as much as 30° during both flight and walking. In this scenario, LNO neurons that provide input to PFN neurons arborizing in the left and right NO would encode how much the head is rotated to one or the other azimuthal direction of the body's axis, perhaps derived from proprioceptive information. When properly calibrated, such differential input could allow the PFN phase shift (which, at 45°, is sufficient to encode the entire range of head-body angles) to rotate the head direction vector by an angle equal to the head-body angle. This would effectively transform the fly's head direction vector into an allocentric body direction vector. To do so, the network could use gain fields, with an intermediate layer composed of PFN neurons whose head-direction tuning curves are gain-modulated by shared input related to the head-body angle. The neuron types downstream of PFN neurons would complete the coordinate transformation through their structured sampling of PFN neurons with distinct directional tunings. Coordinate transformations such as these may be useful when combining 2 allocentric directional representations with body-centered velocity estimates to estimate direction and distance. This idea is returned to in more detail in subsequent sections on vector computations related to path integration (Hulse, 2021).
A second scenario is presented: if it is assumed that an LNO type carries a motor efference copy of the fly's rotational velocity, as has been shown to exist in the fly visual system, then bump shifts driven by differential input to the left and right PFN populations could function as a forward model that encodes a prediction of the fly's future head direction or body direction. Why might a forward model of head or body direction be useful? Intracellular recordings from neurons in the ring attractor network have revealed that PEN activity is tuned to the fly's rotational velocity, but that this activity lags behavior by ~100 ms. Similar lags may result from propagation delays in neural processing, either along sensory pathways into the EB, or in passing the bump from EB to FB. In situations where flies might rely on the CX to direct their movements - especially in time-critical scenarios - such delays in updating the compass could be costly. One way to overcome this is for the PFN network to compute the fly's approximate future head or body direction so that navigational decisions can effectively be made in real-time, a strategy that dragonflies have been shown to use during rapid prey capture maneuvers. A forward model could also allow the fly to distinguish changes in its head or body direction associated with voluntary movements from those induced by external disturbances, such as changes in wind direction. More generally, matching the predicted head and body direction with the actual direction could enable the fly to fine-tune its movements to produce intended motor outputs. As described below, PFL neuron types have anatomical phase shifts that appear well suited to perform such comparisons (Hulse, 2021).
The PFN neuron types that the computations hypothesized above might involve is as yet unclear. However, the analysis of PFN inputs from LNO types does allow drawing some inferences about the sort of self-motion information that different PFN neuron types might carry. For example, considering that PFL neurons provide selective feedback to the LNO3 neuron type and considering that the PFL3 neurons feed the LCNOp neurons, it is hypothesized that the former may provide its downstream PFN neurons - the PFNv neurons - with efference information related to translational (and potentially forward) movement, and that the latter may provide its PFN targets - several PFNp sub-classes - with efference information related to rotational movements. In addition, many LNO types are downstream of pathways from vPNs, consistent with the use of optic 5flow-based self-motion signals. Furthermore, given that LNO types are the target of multiple input pathways, these neurons could carry combinations of sensory and motor signals to encode self-motion. Physiological recordings in behaving flies would be needed to test such hypotheses (Hulse, 2021).
Phase shifts of a single PFN neuron type could enable the generation of vectors that are within ±45° of the fly's instantaneous head direction. Intriguingly, the FB network appears to be wired to expand this range to allow for computations with vectors of arbitrary angles. he dΔ and hΔ neuron types could, in principle, allow for vector computations across arbitrary azimuthal angles. In this example, this study considered two hypothetical PFN populations (PFN1 and PFN2), both with 45° contralateral phase shifts. On their own, these PFN populations are limited to directional representations spanning 45° around the head direction signal that they inherit. However, an excitatory hΔ (or an inhibitory vΔ) would invert the PFN2 vectors, shifting them by 180°. Thus, if a postsynaptic neuron type were to sum the input from the non-inverted PFN1 neuron population and an inverted PFN2 neuron population, it could form a representation of any arbitrary vector over the full 360° range, even though the PFN1 and PFN2 populations are individually range-limited. A similar inversion could happen at the level of the PB, if one PFN population were to receive excitatory Δ7 input while the other received inhibitory Δ7 input, which is likely how the 180° separation of PEN_a and PEN_b population bumps is generated and maintained in the PB. Together, the non-inverted PFN1 and inverted PFN2 neuron populations form a basis set of four basis vectors, all separated by 90°. As mentioned previously, the requirement for PFN neurons to have a positive firing rate prevents any single PFN population from forming a basis set on its own; instead, forming a basis set requires four independent bumps located at 90° intervals. When this situation is achieved, independent NO input could alter the relative amplitudes of bumps carried by each of the four PFN populations (inverted and non-inverted PFN1 and PFN2 populations), enabling their sum to encode a vector with any angle and could thus represent such independent vectors. Importantly, during navigation, the orientation of this set of four vectors would be dynamically updated with the head direction representation, such that any computations derived from these vectors would be independent of the fly's current head direction (Hulse, 2021).
Could the intra-FB network support the construction of arbitrary vectors, which requires two layers beyond PFN input? A type-to-type network graph suggests that there are many pathways within the FB's 2D grid that could potentially implement a four-vector basis set. Two arbitrary PFN types, PFNd and PFNp_c were chosen, and their connectivity with two downstream dΔand hΔneuron types -vDK and hDA- to illustrate how this might work. It was also shown how a downstream neuron type -PFL3 in this case- could sum the input from the dΔ and hΔ to represent arbitrary vectors determined by independent NO inputs to the left and right PFNd and PFNp_c populations. Physiological investigations will be required to establish which of the FB's many pathways implement such computations, and whether or not the large number of these pathways is an indication of vector computations in different behavioral contexts (Hulse, 2021).
Having established that the FB network of Drosophila could, in principle, compute arbitrary vectors, the potential utility of PB-FB phase shifts and intra-FB connectivity motifs for path integration was explored. A variety of models have been proposed for path integration. These models have several differences, including whether the home vector is stored in an allocentric reference frame or an egocentric reference frame, and whether it is stored using a 'static vectorial basis' or a 'dynamic vectorial basis'. This study focused on models that store the home vector in an allocentric reference frame using a static vectorial basis, which has been shown to have several theoretical advantages and whose implementation is directly suggested by the FB's network architecture. Path integration models can be further divided into two groups according to whether the home vector is stored and read out as independent components or as a single vector (Hulse, 2021).
An example of the first type of path integration model, which stores the home vector as two independent components, was recently put forward by Stone (2017). This work combined anatomical and functional data from bees, including physiological recordings of optic flow-sensitive LNO neurons and EM data, to build an anatomically inspired model of path integration based on the projection and innervation patterns of CX neurons, but without access to their synaptic connectivity. The model utilized PB-FB phase shifts to read out a home vector and, importantly, could also account for holonomic motion, which occurs when animals move in directions that are not aligned with their head/body axis, an issue returned to below. At its core, this model and those derived from it function by modulating the amplitude of left and right PFN bumps according to the insect's motion in the leftward or rightward direction, respectively. Integration of the left and right PFN activities can then store a home vector as two independent components. During readout, a population of PFL neurons is assumed to compare the insect's current head direction to that of directions 45° to the left and to the right to decide which direction is closer to the implicitly stored home vector. This 45° 'functional offset' (phase-shift) was derived from physiological recordings demonstrating that some LNO neurons function as optic flow sensors whose optimal expansion points are 45° to the left and right of the bee (Stone, 2017), a feature returned to below. While conceptually elegant, one major feature of this model is inconsistent with the anatomy and connectivity of the homologous neurons in the Drosophila connectome. In particular, the model requires that the left and right PFN bumps independently propagate to right and left PFL populations, respectively. This operation is unlikely to be supported by the FB columnar network, since every neuron and neuron type postsynaptic to PFN neurons receives input from both the left and right populations (Hulse, 2021).
In the next two sections, use the additional anatomical and connectivity information provided by the CX connectome to propose two conceptual models for computing an allocentric translational velocity (TV) vector whose integration could be stored as a single home vector. The first model builds on the work of Stone (2017) and uses PFN offsets to simplify home vector computation. The second model is more relevant to walking insects and incorporates a head-to-body coordinate transformation to compute the fly's translational velocity vector. In both cases, the key computation performed by the FB network is a coordinate transformation that ensures that egocentric velocity signals and allocentric directional representations are directionally aligned (Hulse, 2021).
Flying insects are thought to perform visual odometry by relying on optic flow sensors to estimate their velocity relative to the ground, consistent with leg-based cues being of little use and motor signals being unreliable in the face of external perturbations, like gusts of wind. In addition, during flight many insects make banked turns involving body rolls that are accompanied by gaze-stabilizing head rotations that keep the head near the horizontal plane. Importantly, flight trajectories often contain a significant sideslip component as well, during which the insect's translational velocity is in a direction that is different from that of its head-body axis (Hulse, 2021).
The FB's recurrent circuitry described above could use self-motion information to compute a flying insect's allocentric translational velocity (TV) vector. One potential model is composed of two PFN neuron types that receive independent input from two hypothetical LNO neuron types, LN1 and LN2, for which there are multiple candidates. It exploits the FB network's ability to form a set of four basis vectors to compute a single TV vector. To do so, it employs the optic flow sensors described by Stone (2017) - with their preferred expansion points spaced at 45° intervals around the fly's head - to modulate the amplitudes of the four basis vectors such that their sum encodes an instantaneous allocentric TV vector. Importantly, this model relies on the fact that the basis vectors and optic flow sensors are directionally aligned. That is, at every moment in time, each bump in the basis set has its amplitude modulated by a velocity input that senses movement in the same direction as encoded by the bump. Much like the model of Stone (2017) this model can account for holonomic motion (that is, an animal's movements in directions not limited to its heading and head direction). Another feature of this model is that it should be insensitive to head movements in the yaw plane, since the optic flow sensors and FB basis vectors are both in head-centered coordinates. A recent study found that, similar to the optic flow sensors described above, some PFN neuron types and their LNO inputs are preferentially tuned to air flow oriented ~45° to left or right of the fly's head, providing for a second potential velocity estimate whose tuning is aligned to PFN basis vector. The next conceptual model explores how this might work in walking insects, when the velocity sensors may be in a body-centered reference frame while the directional representation is in a head centered allocentric reference frame (Hulse, 2021).
Could the FB network compute an instantaneous TV vector in cases where its velocity and directional estimates are in different reference frames? A model explores such a scenario using a hypothetical example of a walking fly whose velocity estimates are computed using cues that operate in an egocentric, body-centered reference frame. These velocity estimates could be derived from motor efference copies or proprioceptive cues, and the existence of estimates for both forward (parallel to the body axis) and sideslip (perpendicular to the body axis) velocity. Computing a TV vector in this scenario is more complicated than in the previous model because the direction of the head and that of the body are not necessarily aligned, which requires a head-to-body coordinate transformation. This model uses the head-body angle to compute the total TV vector as the sum of two components, which represent the distance traveled parallel and perpendicular to the fly's body axis. To compute the parallel and perpendicular components of the TV vector, the model uses two basis sets that receive NO input related to the head-body angle as well as either a forward or sideslip velocity signal (Hulse, 2021).
A circuit for computing the component of the TV vector parallel to the fly's body axis is shown that involves two calculations that occur in parallel. The circuit recruits two independent PFN populations, one to encode movement in the forward direction, and the other for movement in the backward direction. A velocity signal increases the amplitude of the two PFN vectors that point in the direction the fly is moving, resulting in a vector whose amplitude encodes velocity and whose direction is either the fly's head direction, or its reverse head direction, which would be captured by the PFN2 population. At the same time, an input related to the head-body angle transforms the head-centered vector into a body-centered vector. The result is a single vector encoding the component of the fly's movement parallel to the body's axis in either the forward or backward direction. As mentioned above, this sort of computation could employ gain fields, but with the transformed representation (that is, body direction) being scaled by the fly's velocity in the process (Hulse, 2021).
The component of the TV vector that is perpendicular to the fly's body axis could be computed using the same circuitry as above, but with right/left sideslip velocity signals instead of forward/reverse velocity signals. Such a circuit would work regardless of whether the fly is sideslipping right or left or whether its head is to the right or left of the body axis. The output of these two circuits could then be summed tocompute a single vector that encodes the fly's instantaneous translational velocity in an allocentric reference frame (Hulse, 2021).
The conceptual models described above -one for flight and the other for walking- could, in principle, compute an allocentric translational velocity vector whose integration would yield an exact home vector. To accomplish this, the models use coordinate transformations to ensure that allocentric vectors are directionally aligned with the egocentric velocity estimates that control their amplitudes. While these particular models highlight the general utility of such transformations, the FB circuitry could, in principle, accommodate many similar models. In addition, it is possible that animals structure their movements during outbound paths to simplify the computation of the translational velocity vector. For example, if an animal were to only move forward during outbound paths, then circuit components dedicated to encoding backward motions would not be needed by the path integration circuit, a feature explicitly used by the model of Stone (2017). Similarly, it is possible that an exact solution is not always required to perform path integration. For example, if a model generates errors that tend to cancel out during the integration process, the home vector can still effectively guide behavior. The local search behavior of foraging Drosophila, for example, involves relatively short loops that may not require a precise accounting of the goal location. In most situations, the fly should also be able to use local sensory cues in addition to path integration during such search behaviors. Finally, it is possible that egocentric velocity signals could come pre-aligned to FB bumps, assuming that the LAL could implement the trigonometric functions needed to scale velocity signals according to, for example, head-body angle. Taken together, these models highlight the connectome's ability to inspire novel, implementation-level hypotheses about network computation. They also provide a framework for generating many similar models, with specific implementations that likely depend on cell type, species, and behavioral need. Ultimately, evaluating models like these necessarily requires physiological recordings from animals in specific behavioral contexts. Indeed, two contemporaneous studies have discovered direct physiological evidence that FB circuits compute the fly's translational velocity, and have independently proposed theoretical models that are conceptually similar to those described above. Yet, it is currently unclear if the output of this computation encodes the fly's translational velocity vector or just the phase of this vector. Similarly, how the type-to-type variability in PFN phase shift magnitude affects these computations requires future study. Finally, it is also possible that tangential neurons carrying feedback or self-motion signals could scale the magnitude of these vectors within the FB network itself (Hulse, 2021).
Theoretical work has suggested several potential ways to integrate translational velocity vectors. First, translational velocity could be integrated and stored using two separate circuits: a ring attractor that encodes the angle of the home vector and a line attractor that encodes the length of the home vector. Rather than keeping track of the distance traveled in each allocentric direction, this network would shift the columnar location of an activity bump to encode the phase of the home vector. Solutions like these seem unlikely to be implemented by the FB, since they require FB-centered attractors with circuit motifs for shifting the bump, similar to those found in the EB-PB attractor, which no evidence is seen for in the FB network. Second, the model of Stone (2027) employed 18 neurons per FB column and used structured recurrent connections between them to integrate and store a two-component home vector. The hemibrain connectome provides little evidence for such structured recurrent PFN connections, especially in the NO, where some PFN types show all-to-all connectivity. Finally, the conceptual models described above allow for the computation of a single TV vector, suggesting that the FB network could simply integrate the corresponding activity bump and store the resulting home vector directly. In doing so, this integration processes would function by keeping track of the distance traveled in each allocentric direction. How might the network integrate the TV vector and store the resulting home vector (Hulse, 2021)?
Integration and storage could occur through several complementary mechanisms operating at different scales, from changes in synaptic strength or the excitability of individual neurons to persistent activity at the single neuron or network level. In addition, integration and storage mechanisms may vary across species and environmental context depending on the animal's needs. For example, desert ants can maintain multi-day memories of multiple goal vectors to food sites and remember home vectors over 1 to 2 days. This sort of long-term maintenance would favor stable storage mechanisms, such as changes in synaptic strength. In contrast, Drosophila performing relatively brief local searches close to a food source may rely on short-term mechanisms that could involve persistent neural activity. The connectome alone does little to constrain the space of possible storage mechanisms, but it can provide information regarding the likely site of storage and inspire a few conceptual models for how the vector could be stored (Hulse, 2021).
Several considerations narrow the potential site of home vector storage in Drosophila. In the framework of the above conceptual models, the home vector would be stored downstream of the four vector basis sets used to compute the TV vector. The PFL2 and PFL3 neuron types are well positioned to read out the home vector by comparing it to the fly's instantaneous head direction, suggesting that the home vector is perhaps stored by neuron types that provide inputs to the PFL neurons. The PFL neuron types could also store the home vector themselves. Some insects are likely to maintain more than one goal vector, but the PFL neurons could store these different goal vectors through input-synapse-specific presynaptic or postsynaptic plasticity. In addition to direct PFN input, PFL2/3 neurons receive shared input from a handful of hΔtypes, several FC2 types, one or two dΔtypes, and many FB tangential neuron types, each of which could also potentially store a home vector (Hulse, 2021).
Several potential storage mechanisms seem plausible. Many hΔneuron types have within-type recurrent connections, forming small loops that connect pairs of hΔneurons that encode directions 180° apart. If the biophysical properties of these neurons allowed for graded, persistent activity, and hΔneurons have inhibitory connections, each column-pair could encode the direction traveled along one dimension. Alternatively, while the FC neurons providing input to PFL2/3 neurons largely lack within-type recurrent connections, they could maintain a vector in working memory through graded changes in their excitability or activity. Finally, the FB's tangential neurons could potentially store home vectors through column-specific plasticity, as is known to occur between ring neurons and EPG neurons. In general, some recurrent architectures may allow for the storage of home vectors, but an FB ring attractor, if it were to exist, would likely not allow for home vector storage, since these networks have the undesirable property of forming a single activity peak at the expense of the activity in distance columns that would be needed to fully encode the home vector (Hulse, 2021).
Overall, although the connectome can do no more than rule out some circuit implementations of how the home vector might be stored, it should prove useful in prioritizing a search for the likely neural targets for such a function. It is important to note that the entire circuitry described above must function in different modes depending on the animal's behavioral needs - integrating direction and distance traveled to update the home vector when the fly is searching, but switching to reading out the home vector when the fly is attempting to return to a previously visited spot. The likeliest candidates for such behavioral mode switching are the FB's tangential neurons (Hulse, 2021).
Once formed, how might an insect 'read out' the home vector to return to its goal location? In the current formulation, the home vector points from the nest to the insect's current location. Returning home, then, requires that an insect move in a direction opposite to the home vector. To accommodate the other behaviors and computations that these circuits are likely to be involved in, the home vector is referred to as the 'stored vector', which is read out to orient the insect along a 'goal vector'. However, unlike an ant or bee, the fly is not a central place forager. Thus, 'goal' in this context refers only to a spot that the fly is likely to return to during a local search, such as a food source. PFL neurons are generally regarded as the major columnar output of the FB network. Their PB-FB offsets strongly implicate them in reading out stored vectors in ways first proposed by theoretical work and then, at the implementation level, by Stone (2017). In particular, PFL neurons may use their PB-FB phase shifts to compare the fly's instantaneous head direction, which they receive in the PB, to that of the stored vector, which they may receive in the FB, to generate appropriate motor commands to guide the fly to its goal. In doing so, they effectively generate egocentric motorcommands based on allocentric directional variables. Interestingly, each of the three PFL types have characteristic phase shifts that strongly predict their involvement in generating distinct motor commands (Hulse, 2021).
PFL2 neurons may use their 180° phase shift and bilateral LAL projections to increase the fly's forward velocity when its heading is directly away from the stored vector, which in the formulation used in this study is towards the goal location. Unlike the other PFL types, PFL2 neurons receive only a single bump as input in the PB. This suggests that the population cannot make left versus right activity comparisons. In agreement with this, individual PFL2 neurons make bilateral projections to the left and right LAL. Because of their 180° phase shifts, the PFL2 population activity will be largest when the fly is heading directly towards its goal location. The above characteristic suggests that PFL2 neurons are ideally suited to generate a motor command related to forward velocity (Hulse, 2021).
PFL3 neurons may use their 90° phase shifts and lateralized LAL projections to orient the fly towards the goal. Their 90° offset predicts that the left and right PFL3 populations will have their maximum activity when the fly is 90° to the right or left of the goal direction, respectively. If the left PFL3 population generates left turns and the right PFL3 populations generated right turns, then the orienting behavior of the fly will have two equilibrium points: a stable equilibrium that occurs when the fly is oriented towards the goal direction and an unstable equilibrium when the fly is oriented in the opposite direction. This sort of read out would ensure that flies orient directly towards the goal location. It is additionally possible that across-column inhomogeneities in the EPG->PFL synaptic profile and in the PFL->LAL network may provide the fly with a 'default goal' in the absence of any FB input, similar to a hypothesis recently advanced in an independent study. The 45° offset of PFL1 neurons may serve a related function, although they target distinct downstream neurons compared to PFL2/3. One possibility is that the PFL2/3 neurons affect body orientation while the PFL1 population controls a separate variable, such as sideslip or head-body angle. Ultimately, it is also important to remember that brain regions like the LAL and CRE house complex recurrent networks with inter-hemispheric pathways that are likely to be inhibitory. These networks are likely to play a major role in the transformation of PFL population activity into motor commands for the fly, something that these hypotheses do not incorporate (Hulse, 2021).
Summary: vector computations in the FB The discussion above supports the notion that the FB network has the computational capacity to compute, store, and read out vectors in support of goal-directed navigational behaviors. While this study has focused on path integration as a canonical vector-based computation, Drosophila are known to perform several other behaviors that may rely on the formation of 97 goal vectors, including: local search, a path-integration-based foraging strategy; menotaxis, where a constant 99 heading is maintained relative to an arbitrary goal direction to generate straight trajectories that support long-distance dispersal; place learning, which requires associating visual cues with the presence of a cool spot in an otherwise hot 2D environment; and the detour paradigm, where flies orient towards directions associated with attractive landmarks even after they have disappeared (Neuser et al., 2008). In addition, ethologically-based studies in behaving insects have established a range of vector-based4 behaviors, from long distance migrations that require a time-compensated sun compass to the waggle dance that bees use to communicate the distance and direction of a food source. The ability of some insects to store multiple goal vectors and the fact that different insect species may use vector computations to support distinct behaviors has important implications for FB circuits. The FB may have evolved as a general vector calculator that can be co-opted, whether by evolution or in support of distinct behaviors, to support vector-based navigation strategies generally. In support of this idea, FB circuits, neuron types, and motifs are highly conserved across insects. Additionally, the ability of some insects to store multiple goal vectors requires mechanisms for switching between them, a function perhaps mediated by the large class of FB tangential neurons that could convey context and state information to the columnar networks involved in vector operations beyond navigation: the CX as a multifunctional network for context-based action selection. While this study has focused much of the discussion on column-specific computations supporting vector navigation, the CX also receives input from over 150 distinct tangential neuron types. In the sections below, these neurons' role in sensorimotor processing, memory-guided decision making, circadian rhythms, sleep-wake control, and nutrient homeostasis is briefly highlight. Together, these findings suggest that the CX operates as a multifunctional network supporting state- and context-dependent action selection for high-level behavioral control (Hulse, 2021).
Sensorimotor processing Consistent with the CX's involvement in navigation, several studies have implicated FB tangential neurons in sensorimotor processing. For example, ExFl1 neurons, which are likely FB2B_a and/or FB2B_b neurons, are strongly modulated by whether or not the fly is flying and are tuned to progressive optic flow, providing a potential indication of the fly's current sensory and motor state. Similar activity patterns may be expressed by several other FB types as well. In addition, a recent study focused on the LH identified an FB tangential neuron type called PV5k1 (FB2H_a, FB2H_b, and/or FB2I_b) whose activation during closed-loop visual conditions leads to a reduction in the fly's wingbeat frequency. Sensorimotor signals like these are well positioned to influence CX-driven motor commands based on the fly's immediate sensory environment and ongoing motor state (Hulse, 2021).
Memory-guided decision making Flexible behavior also requires animals to respond to their immediate sensory surroundings by evaluating past associations regarding the valence and novelty of available sensory cues. To investigate this, focus was placed on tracing pathways between the MB -the fly's main learning and memory center- and the CX. In agreement with results from a companion manuscript focusing on the MB and trans-Tango-based circuit mapping, extensive pathways were found leading from MBONs to FB tangential neurons. In the context of navigation, the MB is considered a potential source of visual snapshot memory, which may allow insects to base their navigation decisions on remembered panoramic views. Consistent with this general notion, some FB tangential neuron types in FB layers 2 and 8 have been proposed to play a major role in visual learning. In addition, recent studies have implicated MB-to-CX pathways in behaviors other than navigation. For example, MB-to-CX circuits may be important for experience-dependent alcohol preference. In addition, MB-to-CX circuits are involved in consolidating courtship experience into long-term memory (Dag et al., 2019). The sheer number of connections between MBONs and FB tangential neurons suggest this prominent pathway is involved in many behaviors that make use of valence and novelty signals extracted from past associations that the fly has made with 61 its current sensory surroundings (Hulse, 2021).
Circadian influence on the CX Animals also select their actions based on latent environmental variables, such as the time of day, which are predictive of environmental conditions like temperature and humidity. Flies are most active around dawn and dusk, and show consolidated periods of inactivity throughout the night and during a daytime siesta (Dubowy and Sehgal, 2017). This daily rhythm is imposed by outputs from the circadian network and functions to restrict behavior to appropriate times of day. Previous studies have identified a population of anterior-projection DN1 clock neurons that convey circadian information through TuBu neurons to EB ring neurons. Thus, CX circuits are likely to receive circadian information that could be used to select behaviors according to time of day. Whether circadian pathways target other regions of the CX requires further investigation. In addition to receiving circadian inputs that could affect rest-activity rhythms, considerable evidence suggest CX circuits are involved in tracking internal states, such as sleep need and nutritive state, which is discussed next (Hulse, 2021).
Sleep-wake control: While its functions remain largely unknown, sleep is associated with a variety of processes in Drosophila, including synaptic homeostasis, memory formation and consolidation, changes in gene expression, and several metabolic processes. Sleep in flies is behaviorally defined as a reversible state of immobility that is homeostatically regulated and associated with an increased arousal threshold. It is marked by drastic changes in brain-wide activity patterns. The neural circuits involved in tracking sleep need and inducing sleep are thought to partially reside in the CX. In particular, a heterogeneous population of FB tangential neurons labeled by the R23E10 GAL4 line induces sleep when activated and tracks sleep need through changes in baseline firing rate and intrinsic excitability. Similarly, ER5 ring neurons track sleep need, and reciprocal connections between the EB and dFB are hypothesized to form a core circuit for homeostatic control of sleep. Counteracting these sleep-promoting neurons are wake-promoting dopaminergic neurons in the dorsal FB that are thought to promote wakefulness by inhibiting R23E10 neurons. Connectomic analysis revealed extensive reciprocal connections between putative sleep- and wake-promoting populations within the dFB, which could function as a 'flip-flop' switch to ensure that only one population is active at a time. In addition, a large number of previously undescribed pathways were identified leading to and from sleep-wake neuron types whose potential involvement in sleep-wake control requires future investigation, including reciprocal pathways connecting neurons in the EB with those in the dorsal FB (Hulse, 2021).
Several limitations of the hemibrain dataset are notable in the context of sleep: neurons that that show structural changes as a function of the fly's sleep-wake history such as ER5, could have sleep-state-dependent connections different from those described in this study; similarly, at present, the hemibrain connectome does not include reconstructed glia, which are also known to be involved in sleep-wake control; lastly, the hemibrain dataset cannot resolve the presence of gap junctions, which may also be important for sleep-wake control (Hulse, 2021).
Nutrient homeostasis: Recent studies have suggested that the CX is involved in internal state-based action selection beyond sleep-wake control. Within the EB, a population of ring neurons allows flies to assess the nutritive value of sugars, independent of their taste. Similarly, tangential neuron types in the dorsal FB have been implicated in feeding decisions based on the nutritive value of foods, and they may incorporate past experience into these computations. And vΔA_a columnar neurons, which innervate the AB and dFB, show oscillatory dynamics that depend on hemolymph glucose levels, and altering vΔA_a activity levels affects fructose preference. Together, these studies implicate CX circuits in nutrient homeostasis, a process important for successful foraging based on the fly's metabolic needs (Hulse, 2021).
Circuit motifs for high-level behavioral control and action selection: The need for high-level behavior selection may explain the potential interactions of circuits related to navigation, feeding, circadian rhythms, and sleep. Hungry flies, for example, are known to forgo sleep in favor of foraging. Similarly, both sleep and feeding are known to be under circadian control, biasing their occurrence to appropriate times of day. Based on these considerations and the experimental evidence summarized above, it seems likely that the CX operates as a multifunctional network that can be dynamically reconfigured (Marder, 2012) to support a variety of goal-directed behaviors based on immediate sensorimotor variables, learned associations, time of day, sleep need, nutritive state, and other as-yet-unknown inputs. Such a view of the CX is consistent with the variety of neuromodulator and peptides released by FB neurons (Hulse, 2021).
This connectomic analysis identified circuit elements and motifs that may support appropriate action selection. Most notably, many tangential neuron types, including EB ring neurons, form dense recurrent connections, both within neurons of a type and across distinct neuron types. For example, the FB's tangential neurons in Layer 6 that have been implicated in sleep-wake control are highly recurrently connected. It is possible that some of these neurons or other neurons in their layer are involved in decision-making related to feeding. If so, inhibitory interactions between these different tangential neurons may -akin to the interactions of ring neurons for sensory control of the fly's compass- enable the fly to select appropriate actions based on internal need. Related to this, recent studies have reported oscillatory activity in ER5 ring neurons related to sleep-wake control, but how the highly recurrent networks in the EB and FB might support such oscillations remains to be determined. One possibility is that all-to-all inhibition between ring neurons in the EB could, with the appropriate inhibitory conductances, induce such patterns of activity. A different issue raised by the highly recurrent architecture of sleep-wake networks concerns how activity may propagate in these networks. Artificial stimulation of neurons within such potentially self-regulating networks may trigger downstream activity that is never seen in more naturalistic situations, confounding the interpretation of experimental results. Testing such ideas will require a finer-resolution analysis of the role that these neurons play in the action selection process (Hulse, 2021).
How might the CX's columnar architecture support these distinct behaviors? Links between the CX's role in sleep and navigation have begun to be explored both experimentally and computationally, but the CX connectome suggests that the number of pathways and neuron types that connect circuit elements known to be involved in these functions may have been underestimated. For example, the dFB tangential neurons involved in sleep-wake control contact many columnar neuron types. Although it is believed that this columnar structure -and the FB's 2D grid more generally- is convenient for vector computations, why this columnar structure may be needed for sleep-wake control or for feeding- and satiety-related computations remains mysterious. One possibility is that head direction or traveling direction signals may be used as proxies for tracking the quality or quantity of the fly's waking experience, perhaps to estimate sleep and/or nutritional need. Alternatively, the FB's navigational signals may be inherently activity-promoting since they likely drive premotor neurons in the FB. If so, these navigational signals may require suppression to establish a sleep state or to enable a hungry fly to stop on a patch of nutritive food. Another possibility is that tangential neurons may gate incoming sensory information, which could promote sleep or perhaps encourage a hungry fly to continue a feeding bout by ignoring distractors. Ultimately, if the columnar neurons are the main output of the CX, as seems likely, the FB's tangential neurons must impact behavior through them (Hulse, 2021).
Considering that the highest layers of the FB are associated with modulating the fly's activity depending on sleep state and satiety levels, the connectivity pattern within the FB suggests that information about the fly's current navigational state may enter the FB ventrally, that additional processing may happen in the middle layers, which receive considerable input from the MB, and that this processing may then determine the fly's next actions (or inaction) in the dorsal layers. An additional possibility suggested by the flow of bump information from ventral to dorsal layers of the FB, and by the diffusion of columns in the dorsal layers, is that the specificity of actions is organized along the vertical axis of the FB, with oriented actions modulated and signaled by output neurons originating in the middle layers and the fly's overall state of activity modulated in directionally non-specific ways by the highest layers (Hulse, 2021).
Compared to the vector computation models suggested by the CX's columnar structure, deriving connectome-inspired insights into the function of the CX's action selection networks proved more challenging. One reason for this is that most FB tangential neurons receive input from CX-associated regions whose function remains poorly understood, like the SMP/SIP/SLP, making it hard to assign specific circuit functions to these neurons based on their inputs alone. In contrast, the vector computation models relied on a considerable amount of prior experimental data that, when mapped to the connectome, provided physiological hooks for generating novel hypotheses regarding circuit function. In addition, FB tangential types often have extensive reciprocal connections to other tangential types, which, given the absence of functional data, is hard to interpret. Once some of these functions are better understood, it may be possible to derive internal state hierarchies, like those identified for directional sensory cues carried by EB ring neurons, which could suggest how the CX prioritizes different internal states. However, many of these internal states involve variables that evolve over time, such as nutritive state or sleep need, suggesting the underlying CX networks may undergo considerable plasticity that may not be apparent in connectome-level connectivity. The dynamic interaction of different internal state variables is likely also governed by neuropeptidergic signals that bathe the CX, but that this analysis did not capture. Finally, given the limited understanding of the variety of behavior the CX may support, understanding how internal state cues may factor into these behaviors is hard to predict at present. To better understand which behaviors the CX may be involved in, this study used the connectome to identify output pathways, a topic dealt with next (Hulse, 2021).
Directing and modulating movement based on the fly's current state: The outputs of the CX likely modulate the fly's actions in a variety of different behavioral contexts, including voluntary take-offs, negotiating uncertain terrain, feeding, oviposition and fighting. The structure of the FB, in particular, suggests that it could modify the head direction signal to orient the fly with respect to behaviorally specific 'goal' directions. Such goals could be a source of food or safety or, for female flies, a good site for oviposition (Hulse, 2021).
The FB's columnar output types (PFL, PFR, FR, FC and FS neurons) feed relatively independent output subnetworks, which may support, through unknown mechanisms, the maintenance of independent goal locations associated with different behaviors. Alternatively, these subnetworks could control independent sets of behaviors. If true, each subnetwork may carry the potential for the execution of actions towards independent goal locations, each specific for a given behavior and carried by a specific FB columnar type (or set of types). For example, some subnetworks could control behaviors related to goals in front of the animal, such as feeding or gap crossing. Some CX outputs contact a limited number of MBON-associated networks. These connections may allow the CX to modulate some behavioral responses to specific sensory contexts that have been associated with negative or positive valence through the MB. The fact that an oviposition neuron (oviIN) is associated with these MBON networks could mean that CX networks influence spatial decision-making during oviposition, which is known to be informed by several external factors. In contrast, how and why CX signals from the columnar FR1 neurons should directly influence MB neurons themselves (in the case of the FR1 neurons, the MBON30 neurons) is less clear. The variety of different interactions between the MB and CX suggest that investigations of memory-guided orientation and navigation may benefit from a study of both regions acting in concert. Consistent with such an idea, the atypical MBON, MBON27, targets the DNa03 neuron type, which is also targeted by PFL3 40 neurons (Hulse, 2021).
Another axis along which CX-mediated behaviors can be subdivided is the scale of orientation control. From the body to the head and legs, proboscis, abdomen or antenna, all body parts have an orientation relative to the environment. Each of these body parts could benefit from coordinated but independent control and could be individually targeted by CX outputs. The CX could, in the context oviposition, direct abdomen bending for egg laying in a manner that incorporates the fly's internal sense of its body size, posture and orientation relative to its surroundings. Hints for how the CX exerts such directional control may be found in the morphology of its outputs (Hulse, 2021).
Output neurons with bilateral innervation patterns in premotor regions such as the LAL and CRE are likely to modulate symmetric actions (for example, forward walking), while those with unilateral innervations in such regions likely control asymmetric actions (for example, turning). Examples of the former include the PFL2 and FS1-3 neurons, while PFL1, PFL3, PFR, FR, FS4 and 55 FC neurons all show unilateral innervations of premotor regions (Hulse, 2021).
These different output signals could also vary in how directly they control flies' behavior. CX outputs could themselves direct the animal's movements and/or orientation towards a desired location or away from one associated with danger. The PFL2 and PFL3 neurons provide the most direct link from the CX to the motor center, known as the ventral nerve cord (VNC). As the major output channel of the CX, they are prime candidates to guide orientation and/or movements to a CX-specified goal. However, these actions would need to be coordinated with movements of body parts that alter the sensed orientation, most notably head movement. The ExR8 neuron is a candidate to carry out some of those corrections, through connections both to DNs and to the visual system. The remainder of the CX's outputs act more indirectly, and may modulate and gate actions controlled by other brain regions rather than directly controlling them. This is well exemplified by the multiple points of convergence between visual pathways and CX output pathways. Some CX output neuron types (PFL1, ExR7 and most of the FC neurons) and much of their downstream circuitry are completely uncharacterized. THese underexplored brain regions, a more complete connectome, and genetically targeted imaging and perturbation experiments will help to identify the function of these pathways (Hulse, 2021).
Navigation with small networks and with numerical variation in columnar neurons: The remarkable behavioral repertoire of insects is still more remarkable when considering their small brains. The CX connectome suggests that part of the secret behind this wide-ranging repertoire lies in having evolved architectures that are precisely configured for sophisticated behavior, but -physiological and behavioral genetics studies suggest- with weights that are plastic to allow these behaviors to flexibly adapt to context and situational demand. It is likely that the impressive computational power of their brains may also derive from an underexplored aspect of their neurons: their capacity for arbor-specific local computations, possibly even subthreshold computations in which synaptic release does not require spiking, and molecular computations through signal transduction cascades. These issues will require further experiments, but the connectivity observeD in the EB, for example, hints at a rich potential for insights into subcellular computation in the CX (Hulse, 2021).
Regardless of the true computational capacity of single neurons, it is remarkable that the fly can navigate with a head direction system of directionally tuned columns (in the EB) and just a few thousand neurons performing vector computation (in the FB). In principle, such small networks should be exquisitely sensitive to any variations in the number of neurons encoding each direction. However, the CX connectome revealed a striking difference in the number of columnar neurons that innervate each of the 18 PB glomeruli (although these differences are mirror-symmetric). Although several studies have investigated the developmental origins of columnar CX neurons, none have noted or focused on this systematic mirror symmetry. There are indications of numerical variations in some columnar neurons, such as the EPG, PEN and PFL neurons, in the FAFB volume as well, and more complete EM reconstructions of that volume should be able to clarify whether these variations exactly match what is seen in the hemibrain volume (Hulse, 2021).
The functional consequences of the systematic variation of neuron numbers across columns are entirely unknown. It is possible that this variation builds redundancy into a critical navigational system, or that the increased numbers of neurons in specific glomeruli ensure a preferred 'default' location for the bump to occupy within the more central columns of CX structures, and perhaps also a 'default' heading for the fly to adopt, an idea that is similar to a suggestion advanced independently. A different possibility is that such variation is not stereotyped across flies, but is specific to individuals, and that this may account for locomotor biases across the population. Functional experiments with specific perturbations of neuron numbers in different columns may be necessary to further investigate this issue. Regardless of their functional role, how such mirror symmetric numerical variation is achieved may be an intriguing question for future studies of CX development. It is not known if asymmetries and mirror symmetries in columnar neuron numbers are also present in the CX of other insects, but parallel efforts in connectomics should soon make this clear. Some Diptera, including Drosophila, have a closed EB, in contrast with most other insects, whose CBLs have an open, FB-like (CBU-like) structure. The non-uniform distribution of the EPG at the base of the EB, EPGt innervation at that location, and systematic modifications to neuron number across the columnar neuron types may represent an evolutionary adjustment to the closing of this structure (Hulse, 2021).
The CX as a tractable deep recurrent neural network: Technical advances over the past several decades have enabled increasingly large-scale recordings of neural activity from the central brains of a wide range of animals. These recordings have, in turn, enabled high-throughput studies of neural response properties that have focused on relating patterns of neuronal activity to sensory, behavioral and internal state variables. However, the biophysical and circuit mechanisms underlying these response properties have been more challenging to access. Similarly, dramatic progress in the field of machine learning has enabled the creation of sophisticated artificial agents that can solve a variety of different cognitive tasks, including flexible navigation. Some units in these deep networks develop response properties broadly similar to those observed in real brains (Hulse, 2021).
Insights into how such artificial neural networks generate the representations observed in their units - something that could, in principle, guide mechanistic hypotheses for the function of natural neural networks - have been slower to come for progress in uncovering the architectural basis of navigational responses in these networks). In this era of deep learning, a broader question concerns the level of understanding that is appropriate or even possible for the function of large and complex neural networks. What seems achievable is an understanding of learning rules and objective functions that can, in principle, generate networks with realistic population responses for specific cognitive tasks. The conservation of the CX's structure across arthropods perhaps highlights the extent to which the region has, in practice, been shaped by such rules over evolutionary timescales in the service of flexible behavior. But what of an understanding of the actual network implementation itself? Some have argued against the necessity or desirability of such a level of understanding. The fly's relatively brief history in systems neuroscience provides an increasingly compelling counterargument and may eventually offer a roadmap for implementation-level understanding that could scale to much larger brains and more complex cognitive functions (Hulse, 2021).
The fly displays a wide repertoire of flexible behaviors, and some of its recurrent neural circuits show dynamics that have been linked to associative learning and navigation across animals. Its 100,000-neuron brain circuits may appear complex, but they also feature modularity, type-specific connectivity and topography that is genetically pre-specified and has been refined over its evolutionary history. Some of these features apply to much larger brains as well, although there is likely greater flexibility in the wiring of mammalian circuits and greater heterogeneity within cell types in the mammalian brain. It is possible that developmentally-driven organizational features of natural brains may actually make them more tractable than artificial neural networks for an understanding of their function. The connectivity of a small fraction of the fly CX's many neuron types arranged by layers. Taking a single-neuron-resolution view of this subnetwork shows just how densely recurrent it is, even at a small scale. Indeed, if the types and connectivity of these neurons were unknown, extracting network structure from population responses would be a challenge. However, sorting the neurons into types -in this case, inhibitory types- makes the logic of the network clearer. Combining this circuit connectivity with physiological studies has enabled not only the generation of hypotheses for the computations that may be carried out by subnetworks at each layer, but, increasingly, tests of these hypotheses. As a result, it is possible to establish circuit-level mechanisms underlying the generation of different response 80 properties. Importantly, fly circuit connectivity is not always structured, many synaptic connections are plastic, and information from one part of the network often flows to all other parts of it. Nevertheless, the developmentally pre-specified organization of these networks makes them experimentally tractable. Although the computational capacity of the morphologically and biophysically complex neurons in these networks has likely been vastly underestimated, the connectome thus raises the prospects for at least a circuit-level understanding of how the fly's CX generates many of this small animal's flexible behaviors (Hulse, 2021).
Throughout the animal kingdom, GABA is the principal inhibitory neurotransmitter of the nervous system. It is essential for maintaining the homeostatic balance between excitation and inhibition required for the brain to operate normally. Identification of GABAergic neurons and their GABA release sites are thus essential for understanding how the brain regulates the excitability of neurons and the activity of neural circuits responsible for numerous aspects of brain function including information processing, locomotion, learning, memory, and synaptic plasticity, among others. Since the structure and features of GABA synapses are critical to understanding their function within specific neural circuits of interest, this study developed and characterized a conditional marker of GABAergic synaptic vesicles for Drosophila, 9XV5-vGAT. 9XV5-vGAT is validated for conditionality of expression, specificity for localization to synaptic vesicles, specificity for expression in GABAergic neurons, and functionality. Its utility for GABAergic neurotransmitter phenotyping and identification of GABA release sites was verified for ellipsoid body neurons of the central complex. In combination with previously reported conditional SV markers for acetylcholine and glutamate, 9XV5-vGAT was used to demonstrate fast neurotransmitter phenotyping of subesophageal ganglion neurons. This method is an alternative to single cell transcriptomics for neurotransmitter phenotyping and can be applied to any neurons of interest represented by a binary transcription system driver. A conditional GABAergic synaptic vesicle marker has been developed and validated for GABA neurotransmitter phenotyping and subcellular localization of GABAergic synaptic vesicles (Certel, 2022).
Serotonin (5-hydroxytryptamine) acts as a widespread neuromodulator in the nervous system of vertebrates and invertebrates. In insects, it promotes feeding, enhances olfactory sensitivity, modulates aggressive behavior, and, in the central complex of Drosophila, serves a role in sleep homeostasis. In addition to a role in sleep-wake regulation, the central complex has a prominent role in spatial orientation, goal-directed locomotion, and navigation vector memory. To further understand the role of serotonergic signaling in this brain area, this study analyzed the distribution and identity of serotonin-immunoreactive neurons across a wide range of insect species. While one bilateral pair of tangential neurons innervating the central body was present in all species studied, a second type was labeled in all neopterans but not in dragonflies and firebrats. Both cell types show conserved major fiber trajectories but taxon-specific differences in dendritic targets outside the central body and axonal terminals in the central body, noduli, and lateral accessory lobes. In addition, numerous tangential neurons of the protocerebral bridge were labeled in all studied polyneopteran species except for Phasmatodea, but not in Holometabola. Lepidoptera and Diptera showed additional labeling of two bilateral pairs of neurons of a third type. The presence of serotonin in systems of columnar neurons apparently evolved independently in dragonflies and desert locusts. The data suggest distinct evolutionary changes in the composition of serotonin-immunolabeled neurons of the central complex and provides a promising basis for a phylogenetic study in a wider range of arthropod species (Homberg, 2023).
The representation and integration of internal and external cues is crucial for any organism to execute appropriate behaviors. In insects, a highly conserved region of the brain, the central complex (CX), functions in the representation of spatial information and behavioral states, as well as the transformation of this information into desired navigational commands. How does this relatively invariant structure enable the incorporation of information from the diversity of anatomical, behavioral, and ecological niches occupied by insects? This study examined the input channels to the CX in the context of their development and evolution. Insect brains develop from ~100 neuroblasts per hemisphere that divide systematically to form "lineages" of sister neurons, that project to their target neuropils along anatomically characteristic tracts. Overlaying this developmental tract information onto the recently generated Drosophila "hemibrain" connectome and integrating this information with the anatomical and physiological recording of neurons in other species, this study observe neuropil and lineage-specific innervation, connectivity, and activity profiles in CX input channels. It is posited that the proliferative potential of neuroblasts and the lineage-based architecture of information channels enable the modification, over the evolutionary time scale, of neural networks across existing, novel, and deprecated modalities in a species-specific manner, thus forming the substrate for the evolution and diversification of insect navigational circuits (Kandimilla, 2023).
Serial electron microscopic analysis shows that the Drosophila brain at hatching possesses a large fraction of developmentally arrested neurons with a small soma, heterochromatin-rich nucleus, and unbranched axon lacking synapses. All 802 'small undifferentiated' (SU) neurons were digitally reconstructed and assigned to the known brain lineages. By establishing the coordinates and reconstructing trajectories of the SU neuron tracts, a framework is provided of landmarks for the ongoing analyses of the L1 brain circuitry. To address the later fate of SU neurons, focus was placed on the 54 SU neurons belonging to the DM1-DM4 lineages, which generate all columnar neurons of the central complex. Regarding their topologically ordered projection pattern, these neurons form an embryonic nucleus of the fan-shaped body ('FB pioneers'). Fan-shaped body pioneers survive into the adult stage, where they develop into a specific class of bi-columnar elements, the pontine neurons. Later born, unicolumnar DM1-DM4 neurons fasciculate with the fan-shaped body pioneers. Selective ablation of the fan-shaped body pioneers altered the architecture of the larval fan-shaped body primordium but did not result in gross abnormalities of the trajectories of unicolumnar neurons, indicating that axonal pathfinding of the two systems may be controlled independently. This comprehensive spatial and developmental analysis of the SU neurons adds to understanding of the establishment of neuronal circuitry (Andrade, 2019).
The central complex (CX) of the insect brain plays an important role in a variety of different behaviors,
including the fine control of motor movement and spatial orientation. Recent
studies indicate that the CX harbors dynamic neural activity that integrates the animal's external
and internal environment. Anatomical studies have started to reveal the neuronal connectivity that underlies CX function. The
CX is comprised of four major compartments, including (from anterior to posterior) the ellipsoid
body (EB), fan-shaped body (FB) with noduli (NO), and protocerebral bridge (PB). CX circuitry is dominated by an orthogonal scaffold of transversally
oriented ('tangential') widefield neurons, and longitudinally oriented columnar small field neurons.
Tangential neurons, whose fibres are directed parallel to the length axis of the CX neuropils, provide input to the CX from other brain areas. Best understood among these input
neurons are the TL-neurons in locust and their Drosophila counterparts, the
R-neurons, that conduct retinotopically ordered visual information to the ellipsoid body. Columnar neurons, which interconnect the different CX neuropils along the
antero-posterior axis, are characterized by highly localized dendritic and axonal endings in narrow
volumes ('columns') of the respective compartments. Most classes of columnar neurons, within
a given CX neuropil, are confined to a single column ('unicolumnar neurons'). Projections
are characterized by a strict homotopic order, whereby columns within the lateral half of the PB
are connected to columns of the ipsilateral PB and EB, and medial PB columns project to the
contralateral FB and EB. One class of columnar neurons, the so-called pontine neurons of
the fan-shaped body, behave differently. Their
projection is restricted to the FB, interconnecting two FB columns located on either side of the
midline ('bicolumnar neurons') (Andrade, 2019).
Developmental studies provide a valuable approach to unravel the circuitry of the brain, including
the central complex. A hallmark of the Drosophila brain is its composition of invariant neuronal and
glial lineages, originating from stem cells (neuroblasts) that appear in the early embryo. Embryonic
neuroblasts express specific combinations of transcription factors (TFs), which are thought to
provide each lineage with the information needed to shape the connectivity of its neurons. As a result, lineages become structural modules: Neurons of the same lineage generally project together in one or two fiber tracts, and form synapses in specific, spatially restricted brain compartments. Neuron classes of
the CX conform well to the lineage principle. For example, the R-neurons of the ellipsoid body are
derived from one lineage, DALv (also called EBa1). Sublineages of DALv
born at different times further tile the bulb (BU) and EB into discrete layers. The columnar neurons of the CX are produced by four pairs of lineages located in the dorsomedial brain, called DM1/DPMm1, DM2/DPMpm1, DM3/DPMpm2, DM4/CM (called DM1-DM
henceforward). The spatial pattern of these lineages is reflected in the position
at which their corresponding tracts enter and terminate within the CX. In this manner, the four
lineages subdivide the CX neuropils into four evenly sized quadrants (Andrade, 2019).
The brain of Drosophila and other holometabolous insects arises in two distinct phases. During
the first phase, neuroblasts of the embryo produce a relatively small set of primary neurons which
differentiate and form the larval brain. Most neuroblasts then enter a dormant phase that lasts
towards the end of the first larval instar. Subsequently they reactivate and produce secondary,
adult specific neurons. These cells form axon bundles that form a 'blue print' of connections later
established within the adult brain. Differentiation is delayed until metamorphosis, when secondary
neurons, along with re-modeled primary neurons, extend axonal and dendritic branches and
form synapses. In general, primary and secondary neurons of a given lineage show fundamental
structural similarities, whereby projections of secondary neurons follow those of earlier formed
primary neurons. Remarkably, the central complex, as defined anatomically for
the adult, is the one major set of compartments of the fly brain that lacks an obvious counterpart
in the larva. Thus, all tangential and columnar neurons with their highly ordered connections
outlined above are secondary neurons born in the larva, prompting the question of what guidance
mechanisms control CX connectivity, and what part primary neurons play during this process (Andrade, 2019).
Previous work described a set of embryonically born (i.e., primary) neurons
belonging to the DM1-4 lineages. These neurons, visualized by the expression of R45F08-Gal4, form a commissural tract that becomes incorporated into the fan-shaped
body. Along with the emerging tracts and filopodia extended by secondary neurons of DM1-4
the R45F08-Gal4-positive neurons form a 'fan-shaped body primordium' (prFB). This study undertook a detailed analysis of the structure, differentiative fate, and developmental role of the primary neurons that form the fan-shaped body primordium, using serial electron microscopy of the early larval brain in combination with confocal analysis of all stages covering early larva to adult. The neurons of the fan-shaped
body primordium, that will be called fan-shaped body pioneers (FB pioneers) in the following, form
part of a much larger population of early larval brain neurons that are arrested in development,
projecting a simple, thin, unbranched process into the neuropil. The large majority of these neurons
entirely lack synaptic contacts; in a small number of them, a few presynaptic sites are seen.
Aside from their undifferentiated neurite arbor, these neurons differ from regular, mature
neurons by the small size of their cytoplasm and nucleus, and the abundance of heterochromatin.
Virtually every brain lineage possesses a complement of the small undifferentiated (SU) neurons.
The data show further that SU neurons differentiate in the late larva and pupa and give rise to
distinct adult neuron populations; FB pioneers produce the pontine neurons of the fan-shaped
body. Later born secondary neurons of the DM1-4 lineages, destined to form the various classes of
unicolumnar neurons of the central complex, fasciculate with the FB pioneers on their pathway
towards and across the midline. However, selective ablation of FB pioneers did not result in
gross abnormalities of the trajectories of unicolumnar neurons, suggesting that the initial axonal
pathfinding of the two system of columnar neurons may be controlled independently (Andrade, 2019).
This analysis demonstrates that a large fraction of the neurons of the early Drosophila larval brain
does not elaborate a branched neurite arbor and synaptic connections. This finding came as a
surprise; it had been well established that the large number of neurons produced during the
secondary, larval phase of neurogenesis remain undifferentiated until the onset of metamorphosis,
resembling in many ways the SU neurons described in this study, but the same was not assumed for so many
of the embryonically generated primary neurons. Previous studies had shown that in the thoracic
ganglia, a subset of presumptive adult peptidergic neurons and motor
neurons show a SU phenotype, extending a truncated axon into the peripheral
nerve, but failing to form synaptic connections to the musculature. These embryonically born
neurons, which transiently express the transcription factor Broad-Z3, differentiate along with
the larvally born motor neurons and form dendritic and axonal branches and synapses in the
pupa. Due to their delayed differentiation, typical for secondary neurons,
these embryonically born thoracic SU neurons have been termed 'embryonically born secondary neurons'. To avoid confusion, this study will stick to the convention that defines all neurons born during the embryonic period as primary neurons, and call them 'primary SU neurons' (Andrade, 2019).
The existence of SU neurons (primary or secondary) is most likely tied to the holometabolous
life cycle of Drosophila where, in terms of structure and function, the larval body (formed in the
embryo) differs strongly from the adult body (formed in the larva and pupa). The proliferation of
adult-specific cells and organs that takes place in the larva is separated from the differentiated larval
structures, possibly in order to prevent interference between novel growth and organ function. In
case of the musculature, for example, proliferating adult myoblasts form clusters of cells attached
to the peripheral nerves or imaginal discs, outside the larval musculature. Neuroblasts
generating adult specific neurons are part of the larval brain, but their progeny are arrested in
the immature SU state until the onset of metamorphosis, when, under the influence of ecdysone
signals, all neurons start branching and generating synaptic connections. If that were not the case,
that is, if secondary neurons would continuously differentiate according to their birth date (like
regular primary neurons in the embryo), they would constantly and in growing numbers intrude
into existing larval circuits, possibly leading to disruptions in functioning of these circuits. It is
conceivable that the occurrence of primary SU neurons can be explained by the same reasoning.
Based on their (mostly) superficial position, it is assumed that primary SU neurons are born during
the final rounds of embryonic neuroblast divisions. It could be speculated further that there is a
'cut-off' line that limits neurons' ability to commence differentiation, and that this cut-off line falls
before the time interval during which primary SU neurons are born, thereby preventing the latter
from differentiating (Andrade, 2019).
In support of the notion that the presence of SU neurons is an attribute of holometabolous
insects, such cells have not been observed in locusts or other hemitabolans for which neuro-
developmental observations have been made. A good number of central neurons of the brain and
VNC that were followed throughout development show continuous growth and arborization of
their neurite tree. This also includes
the unicolumnar neurons of the central complex which, in Drosophila, are all born as secondary
neurons and undergo a phase of developmental arrest in the larva. In grasshopper, the homologous
neurons mature continuously between mid- and late embryonic stages, to form part of a functional
central complex right after hatching of the embryo (Andrade, 2019).
Drosophila SU neurons described in this paper exhibit structural characteristics that are similar
to those described for neuronal precursors in the developing vertebrate brain. Thus, postmitotic
neuronal precursors of the neocortex or hippocampus, while migrating along radial glia, are small,
electron-dense cells with hetero-chromatin-rich nuclei and scant cytoplasm. Typically, they exhibit
a bipolar shape, extending a leading and trailing process that are in contact with the radial glia. The same phenotype is observed in neuronal precursors ('D-cells') that are generated in the subgranular zone of the hippocampus in adult mammals. As neurons mature, forming dendrites and axons, nuclear
and cytoplasmic size increase, and cells become transcriptionally more active, with a concurrent
reduction in heterochromatin. Experimental studies have shown that a variety of signaling pathways
and receptors for neurotrophic factors become activated by proteins forming part of the complex
cell cycle-controlling molecular machinery. However, the specific mechanism that drives the transition from small, heterochromatin-richneural precursor to differentiated neuron is little understood. In human and mouse, mutations in the MECP2 protein, which encodes a transcriptional repressor, is associated with a reduction or delay of neuronal maturation (Rett Syndrome). The gene network (of which MECP2
may form part) that accompanies neural precursor
maturation has not been established. In Drosophila, this mechanism is embedded into the ecdysone
hormonal cycle that controls larval growth and metamorphosis in general. It has been shown
that different isoforms of the ecdysone receptor (EcR) are expressed and required for different
developmental changes that occur in the nervous system. The EcRB1
and EcRB2 isoforms are expressed in primary neurons that undergo remodeling, including the gamma neurons of the mushroom body, and blocking this receptor will result
in defects of remodeling. In contrast, EcR-A appears to be more dedicated to guide secondary
neurons through their maturation and maintenance. The level of
ecdysone and its receptors are under the control of developmental paracrine signals, such as in
case of the mushroom body cells which produce an activin signal to maintain EcR-B1 levels. In addition, intrinsic determinants expressed sequentially in the dividing neuroblasts
form part of a feed-back mechanism with the ecdysone cycle. SU neurons in
the Drosophila larval brain may present a favorable paradigm to study the process of neuronal
maturation downstream of the ecdysone cycle. SU neurons represent a major population at the
early larval stage (primary SU neurons) and late larval stage (primary and secondary SU neurons),
and can be labeled by specifically expressed factors (e.g., Broad-Z3), which should make them
amenable to FACS sorting and systematic gene expression screens (Andrade, 2019).
Most neuropil compartments of the adult Drosophila brain have a corresponding larval counterpart; outgrowing fibers of secondary neurons,
which form much of the volume of the adult compartments, follow their primary siblings and
establish dendritic and axonal branches around this primary scaffold. This principle does not apply
to the secondary neurons forming the central complex, for which no anatomically defined larval
counterpart exists. Small primordia of the different compartments of the central complex and
associated structures (i.e., the bulb and anterior optic tubercle, which relay input to the central
complex) can be first detected at the late larval stage. These larval primordia of
the central complex compartments are formed by the fiber bundles and associated filopodia of the
secondary lineages which will develop into the central complex of the adult brain. The minute early
larval prFB, formed by the FB pioneers described in the present paper, represent an exceptional
case. Thus, FB pioneers are primary neurons whose axons extend during the late embryonic phase
and gather into a tight commissural bundle located in the center of the crossing fiber masses that
constitute the supraesophageal commissure of the early larval brain. Several aspects of the prFB deserve special comment (Andrade, 2019).
(1) From late embryonic stages onward the FB pioneer axons are enclosed by an exclusive glial
layer formed by the so called interhemispheric ring gli. Several pairs of primary
glia, located close to the brain midline, make up the interhemispheric ring. Processes of these glial
cells assemble into an invariant pattern of sheaths around several individual commissural bundles.
Posteriorly, glial processes form two channels, a ventral one containing the great commissure, and
a dorsal one, dedicated to the prFB. This dorsal channel conducting
the prFB stands out by a pair of glial nuclei attached to its posterior-medial wall; this study could
unequivocally identify a pair of glial nuclei at that position in the serial EM stack. As the larva
grows and secondary tracts of DM1-4 are added to the FB pioneer bundle, the glial channel widens (Andrade, 2019).
This volumetric increase continues throughout metamorphosis, and eventually the interhemispheric
ring glia accommodates the entire fan-shaped body. Similar to other primary neuropil glia, the
interhemispheric ring undergoes apoptotic cell death during mid-pupal stages, and is replaced by a much larger number of small secondary glia that surround
the adult fan-shaped body. Genetic studies indicate that interhemispheric glia does play a role
in the morphogenesis of the fan-shaped body, even though this role may be relatively minor, or
occur late in development. Thus, genetic ablation of glia, or loss of function of
molecular factors expressed specifically in the interhemispheric ring, result in defective shapes of the adult FB and EB. However, the pathways of DM1-
lineages
at the late larval stage did not display major defects (Andrade, 2019).
(2) FB pioneers differentiate into the pontine neurons of the adult central complex. Pontine
neurons differ in their projection from all other columnar neurons, because they connect two
columns on either side of the midline. For example, pontine neurons of the right hemispheric DM4
lineage connect the right lateral column of the FB with its left medial column, thereby crossing the
midline. In contrast, axons of other, unicolumnar neurons of the right DM4 remain ipsilaterally,
connecting only to the right lateral column of the FB. The trajectories of the FB pioneers
reflect this pontine-typical behavior already in the early larva. Thus, the majority of DM4 SU
axons reach the midline and terminate just after crossing it; DM3 axons project slightly further,
followed by DM2 and DM1, which reach more than 20μm into the contralateral hemisphere (Andrade, 2019).
Outgrowing secondary DM1-4
tracts, even though they initially follow the FB pioneers, show their
own characteristic pattern of termination. In particular, secondary axon tracts of DM4
and DM3 do not cross the midline, but form terminal filopodial tufts in the lateral and medial half, respectively, of the ipsilateral prFB (Andrade, 2019).
(3) Even though primary FB pioneers and their secondary follower tracts are in close contact
to each other throughout larval development, ablation of the former does not result in gross
structural abnormalities of the latter. Thus, the characteristic trajectories and branching pattern of
the Neurotactin-positive secondary tracts of DM1-4
in the late larva lacking FB pioneers appeared
indistinguishable from the control. Filopodial tufts of secondary tracts in ablated specimens still
assembled into regularly sized globular structures, representing the forerunners of fan-shaped
body columns. These findings imply that separate guidance systems act on the early
born pontine neurons and later born unicolumnar neurons. Nothing is known about the molecular
nature of global or local signaling systems controlling the highly ordered architecture of the DM1-4
unicolumnar connections within the central complex. Given that neither ablation of glia, nor loss
of primary FB pioneers, causes major changes in this architecture, at least at the initial phase
of axonal pathfinding, it is likely that local interactions among the different DM1-4 lineages and
sublineages plays a predominant role. For example, local repulsion in between neurons of these
lineages could be instrumental in specifying the separate, largely non-overlapping medio-lateral
domains within the the FB and EB where neurons terminate. Similarly, interactions occurring in
between sequentially born sublineages within a given DM lineage could determine the projection
to different territories along the anterior-posterior axis. It is not yet known how the different
classes of DM neurons distinguished by projection (e.g., PB-FB vs PB-EB vs FB-NO etc) relate to
their pedigree, that is, the time they are born, or the sublineage they belong to. It stands to
reason that the different intermediate progenitors born from the type II DM neuroblasts are
responsible to generate structurally different classes of neurons, but this remains to be confirmed
by detailed clonal analysis. If proven correct, one could surmise that intrinsic factors expressed by a
given intermediate progenitor provides its progeny with a specific 'projection identity'. Neurons
descended from a hypothetical intermediate progenitor A might recognize a more posterior territory
within the prFB as their proper destination, whereas neurons formed by a (later born) progenitor B
are repelled by the A neurons, and are forced to terminate in more anterior territory. The former
class would develop into PB-FB neurons, the later into PB-EB neurons. That repulsive interactions
in between sublineages are important has been experimentally proven by a recent analysis of
semaphorin signaling in the ellipsoid body. Here, repulsion among DALv
R-2 neurons, born at different times, is instrumental for the proper central>peripheral projection of
axons within the EB (Andrade, 2019).
(4) It is an open question what, if any, role the primary neurons of DM1-4
(both differerentiated neurons and other [non-prFB] SU neurons) play in the adult central complex. It is quite possible
that these neurons do not contribute at all to this structure; in case of another lineage, DALv2,
that has been shown to be the case: secondary DALv2 neurons form the ellipsoid body of the
adult brain, but primary DALv neurons arborize in the lateral accessory complex (LAL) and inferior
protocerebrum (IPa) of the larva and adult, but do not become part of the ellipsoid body. The same may be true for the primary neurons of DM1-4. On the other hand, at least a
(small) subset of DM4 definitely will be incorporated into the central complex: the dopaminergic
neurons of the PPM3
cluster, which profusely innervate the central complex and its associated
structures (LAL, BU) have been identified as primary neurons of DM. The
arborization pattern and connectivity of primary DM1-4 neurons (as that of primary neurons in
general) will be worked out in the near future, based on the same serial EM stack that served as
the basis for the current work; however, additional markers that remain continuously expressed in
primary neurons from larval to adult stages will help solving the puzzle of how these neurons are
reorganized during metamorphosis and what fate awaits these neurons in the adult brain.
Aside from pioneering the fan-shaped body primordium, SU neurons form part of almost all
lineages of the early larval brain, but it is not yet known what fate awaits these neurons. In
view of the case represented by the FB pioneers, it is surmised that other SU neurons also survive
and differentiate during metamorphosis. The axonal projection of SU neurons of a given lineage
prefigures (generally in a rudimentary way) the pathway formed by later born secondary neurons
of that lineage. Markers similar to the one provided by R45F08-Gal4
are required to establish what type of adult neurons the different SU neurons give rise to. Of particular interest are SU neurons of lineages that, like DM1-4, contribute to the adult central complex. In case of
DALv2, which generates all of the R-neurons of the EB, a single SU neuron exists in each hemisphere. This neuron projects a short axon along the primary tract, but does not reach the EB
primordium described for the late larval stage in previous works. Two
other lineages, DALcl1
and DALcl2, contribute a large number of secondary neurons to the anterior
visual pathway, which provides input to the central complex. Both lineages are
composed of two hemilineages, DALcl1/2d (dorsal) and DALcl1/2v (ventral). DALcl1/2d differentiate
into small neurons whose proximal dendrites innervate the anterior optic tubercle, and distal axons
the bulb, where they target the dendrites of DALv2 neurons.
DALcl1/2 form a relatively large number of SU neurons which for the most part follow the dorsal
pathway, suggesting that they belong to the DALcl1/2d hemilineage. As outlined above, secondary
neurons of DALcl1/2
innervate the anterior optic tubercle and the bulb of the adult brain. Do the
earlier born primary SU neurons form early larval primordia of these compartments, analogous to
the prFB established by SU neurons of DM1-4. Most DALcl1/2
SU neurons extend relatively short axons that follow the differentiated DALcl1/2d neurons, cross the peduncle of the mushroom body,
and terminate shortly thereafter. A few other DALcl1/2
SU neurons project further, but terminate
at different locations along the primary tract. In other words, a spatially restricted territory that
houses all DALcl1/2d SU terminations, and that might therefore be considered a forerunner of the
bulb, does not exist in the early larva. Similarly, no projections of SU neurons are concentrated in a
region that might correspond to the primordium of the anterior optic tubercle. In conclusion, SU
neurons of lineages DM1-4
may represent a rare case where primary neurons establish a blueprint
for an adult-specific brain compartment (Andrade, 2019).
Goal-directed navigation is thought to rely on the activity of head-direction cells, but how this activity guides moment-to-moment actions remains poorly understood. This study characterize how heading neurons in the Drosophila central complex guide moment-to-moment navigational behavior. An innate, heading-neuron-dependent, tethered navigational behavior was established where walking flies maintain a straight trajectory along a specific angular bearing for hundreds of body lengths. While flies perform this task, chemogenetics was used to transiently rotate their neural heading estimate and observe that the flies slow down and turn in a direction that aims to return the heading estimate to the angle it occupied before stimulation. These results support a working model in which the fly brain quantitatively compares an internal estimate of current heading with an internal goal heading and uses the sign and magnitude of the difference to determine which way to turn, how hard to turn and how fast to walk forward (Green, 2019).
Many animals rely on an internal heading representation when navigating in varied environments. How this representation is linked to the sensory cues that define different surroundings is unclear. In the fly brain, heading is represented by 'compass' neurons that innervate a ring-shaped structure known as the ellipsoid body. Each compass neuron receives inputs from 'ring' neurons that are selective for particular visual features; this combination provides an ideal substrate for the extraction of directional information from a visual scene. This study combines two-photon calcium imaging and optogenetics in tethered flying flies with circuit modelling and shows how the correlated activity of compass and visual neurons drives plasticity, which flexibly transforms two-dimensional visual cues into a stable heading representation. This study describes how this plasticity enables the fly to convert a partial heading representation, established from orienting within part of a novel setting, into a complete heading representation. The results provide mechanistic insight into the memory-related computations that are essential for flexible navigation in varied surroundings (Kim, 2019).
Many animals rely on an internal heading representation when navigating in varied environments. How this representation is linked to the sensory cues that define different surroundings is unclear. In the fly brain, heading is represented by 'compass' neurons that innervate a ring-shaped structure known as the ellipsoid body. Each compass neuron receives inputs from 'ring' neurons that are selective for particular visual features; this combination provides an ideal substrate for the extraction of directional information from a visual scene. This study combines two-photon calcium imaging and optogenetics in tethered flying flies with circuit modelling, and shows how the correlated activity of compass and visual neurons drives plasticity, which flexibly transforms two-dimensional visual cues into a stable heading representation. This study also describes how this plasticity enables the fly to convert a partial heading representation, established from orienting within part of a novel setting, into a complete heading representation. These results provide mechanistic insight into the memory-related computations that are essential for flexible navigation in varied surroundings (Kim, 2019).
This study has shown how inhibitory Hebbian plasticity can rapidly transform visual feature information into an attractor-driven internal representation. Angular velocity input to the attractor converts an emerging mapping on the basis of limited views of a scene into a complete and consistent heading representation, a potentially critical function in animal navigation. The induction of inverse maps emphasizes the notable flexibility of the system. A key issue that remains unresolved is the nature of bump dynamics during translation in a two-dimensional environment. Mammalian head-direction cells are unaffected by translation1, but this Drosophila model suggests that the compass circuit tracks the angle between the orientation of the fly and an object in the visual scene without correcting for translation-potentially making it a local compass. However, the plasticity that this study has identified required only a few minutes, and may be even faster under natural conditions when the system can co-opt an existing mapping from ring to compass neurons. In simulations, this timescale prevented nearby objects and transient stimuli-such as neighbouring conspecifics that would not move coherently with the bearing of the fly-from being mapped, but tethered the compass to distant objects that moved coherently with the turns of the fly (Kim, 2019).
The locus of plasticity is likely to be synapses between ring and compass neurons; An accompanying article (Fisher, 2019), presents electrophysiological evidence that is consistent with plasticity altering inhibitory visual inputs to individual compass neurons. At a synaptic and biophysical level, it remains to be seen how the Hebbian mechanism that is proposed in this study relates to, and interacts with, other forms of plasticity such as spike-timing-dependent plasticity, or with plasticity-inducing mechanisms such as nitric oxide signalling in the ellipsoid body, dopaminergic modulation (as seen in the fly mushroom body) or plateau potentials (as seen during remapping of hippocampal place cells) (Kim, 2019).
The results support a model in which plasticity is constantly active to allow rapid adaptation to new settings, enabling the ring attractor to generate a single heading direction even in a complex environment. Such stable sensorimotor representations probably enable animals to overcome transient uncertainties in their surroundings as they pursue diverse behavioural goals (Kim, 2019).
In the Drosophila brain, 'compass' neurons track the orientation of the body and head (the fly's heading) during navigation. In the absence of visual cues, the compass neuron network estimates heading by integrating self-movement signals over time. When a visual cue is present, the estimate of the network is more accurate. Visual inputs to compass neurons are thought to originate from inhibitory neurons called R neurons (also known as ring neurons); the receptive fields of R neurons tile visual space. The axon of each R neuron overlaps with the dendrites of every compass neuron, raising the question of how visual cues are integrated into the compass. Using in vivo whole-cell recordings, this study shows that a visual cue can evoke synaptic inhibition in compass neurons and that R neurons mediate this inhibition. Each compass neuron is inhibited only by specific visual cue positions, indicating that many potential connections from R neurons onto compass neurons are actually weak or silent. It was also shown that the pattern of visually evoked inhibition can reorganize over minutes as the fly explores an altered virtual-reality environment. Using ensemble calcium imaging, it was demonstrated that this reorganization causes persistent changes in the compass coordinate frame. Taken together, these data suggest a model in which correlated pre- and postsynaptic activity triggers associative long-term synaptic depression of visually evoked inhibition in compass neurons. These findings provide evidence for the theoretical proposal that associative plasticity of sensory inputs, when combined with attractor dynamics, can reconcile self-movement information with changing external cues to generate a coherent sense of direction (Fisher, 2019).
The compass neurons in the Drosophila brain exhibit some resemblance to the head-direction cells of the mammalian brain. Visual cues stabilize the tuning preferences of mammalian head-direction cells, and when a visual cue is rotated to a new horizontal position, the preferences of all of the head-direction neurons rotate together. It has been proposed that the mammalian head-direction system represents a ring attractor-a network in which global dynamics exhibit multiple stable states that unfold in a repeated sequence in response to an input. However, it is not known how visual cues anchor the mammalian head-direction system at a mechanistic level. It has been suggested that Hebbian synaptic plasticity of visual inputs enforces the correct mapping between sensory cues and attractor network states (Fisher, 2019).
Similar to mammalian head-direction cells, Drosophila compass neurons (called E-PG neurons) have properties of a ring attractor. Indeed, the dendrites of E-PG neurons are arranged in a ring in the brain. At any point in time, there is one 'bump' of activity in the E-PG ensemble, which rotates as the fly turns. This network receives continuous input from brain regions that track the rotational velocity of the fly via optic flow signals, proprioceptive signals and/or motor efference signals. These rotational velocity inputs push the bump around the circle. Visual cues make the position of the bump more accurate and stable. It is not known whether visual inputs to E-PG neurons are plastic: the offset between the E-PG bump and the visual world is different in different individuals and it can occasionally change unpredictably within an individual; however, network instability alone does not provide evidence for synaptic plasticity (Fisher, 2019).
It is proposed that correlated pre- and postsynaptic activity triggers associative long-term synaptic depression of R-to-E-PG inhibition. This learning rule would explain why visual receptive fields and heading tuning are typically aligned in E-PG neurons. When an individual R neuron is activated by a visual cue, it should push the bump of activity towards the E-PG neurons that it inhibits most weakly. If the full ring attractor network agrees with this outcome, then long-term synaptic depression will occur and those weak R-to-E-PG synapses will become even weaker, further reinforcing this outcome. To ensure network stability, long-term synaptic depression should be balanced by long-term potentiation at R-to-E-PG synapses; the co-existence of long-term synaptic depression and long-term potentiation would also explain why bidirectional changes are found in visual receptive fields after training. These learning rules should produce a doubled pattern of R-to-E-PG synaptic weights after training in a two-cue world, reflecting the twofold symmetry of visuomotor correlations (Fisher, 2019).
The key result of this study-that visual inputs to E-PG neurons are plastic-supports theoretical models that describe how a network can progressively establish a spatial map of the world by incorporating information about consistent sensory cues during exploration. In robotics, this process is called simultaneous localization and mapping. Ther results provide direct experimental evidence for this type of unsupervised learning at the level of synaptic potentials in vivo (Fisher, 2019).
In a simultaneous localization and mapping framework, visual cues are often local, meaning that they can change in size and apparent angle as they are approached; by contrast, this study chose to use visual cues that could not be approached, simplifying the relationship between heading and visual cues. This choice was motivated by the known receptive field properties of R2 or R4d neurons, which seem adapted to detect the position of the Sun (or Moon). Specifically, R2 or R4d neurons have large inhibitory surrounds, meaning that they only respond robustly to isolated visual objects such as the Sun. The Sun is an ideal compass cue because it is effectively at infinity (Fisher, 2019).
It is proposed that plasticity at R-to-E-PG synapses allows the position of the Sun to be flexibly associated with other compass cues, such as the pattern of linearly polarized light in the sky, sky-wide chromatic and intensity gradients, and wind. In other insects, the E-PG network responds to multiple sorts of compass cues, and navigation behaviour can depend on arbitrary learned associations between compass cues. In a companion study, Kim (2019) provides evidence in favour of the idea that plasticity could be used to learn a complex conjunction of visual objects; in the future, to test this idea, it will be interesting to see whether any complex scene can generate a progressively more-stable heading representation (offset) during training. It will also be important to extend the approach that was taken in this study to simulate a more naturalistic virtual world, to study how multiple types of cues influence the behaviour of this network and the organism (Fisher, 2019).
Navigation in many animals involves an internal sense of heading direction. Such a sense of heading is thought to be mediated by neurons that are specifically active when the animal is orienting toward the neuron's preferred direction. These head direction cells reside in multiple brain regions and interact with each other as well as with other neurons, including cells encoding angular velocity (Shiozaki, 2020).
In Drosophila melanogaster, two types of columnar neurons innervating the ellipsoid body (EB) and the protocerebral bridge (PB), subregions of the central complex (CX), encode heading direction. As with head direction cells in mammalian brains, columnar neurons encode heading as the identity of active neurons among the population. This heading representation can be updated by visual and self-motion cues via interactions between specific types of columnar neurons. These cell type-specific analyses have provided support for computational models of heading direction encoding, originally proposed for rodent neurons, where angular information is integrated through recurrent excitation. However, the CX is comprised of four highly interconnected subregions that likely function coordinately, and how information related to heading is represented and further processed in other subregions, especially in downstream areas, remains unexplored (Shiozaki, 2020).
The fan-shaped body (FB), another subregion of the CX, is anatomically considered to be an area downstream of columnar neurons in the EB. A functional connectivity study suggested that columnar neurons in the EB, albeit indirectly, influence the activity of certain FB neurons. In addition, the FB directly receives input from outside the CX and is involved in various aspects of navigation, such as visual memory, locomotor handedness, and processing of optic flow. Therefore, the FB is well positioned to receive heading signals from columnar neurons in the EB and integrate them with other types of signals to guide navigation. However, it remains unclear whether and how neurons in the FB encode information about directional heading in behaving animals (Shiozaki, 2020).
Two-photon calcium imaging in the FB was carried out while flies were flying in visual virtual reality as well as in darkness. Specific types of columnar neurons in the FB show characteristic population dynamics that are prominent during flight but not in quiescence. These dynamics multiplexed information about ongoing turning behavior and heading direction. Activity of these FB neurons was coordinated with that of columnar neurons in the EB, which also encoded turning behavior and heading direction. Despite these similarities, columnar neurons in the FB and EB showed distinct activity in their branches in the noduli (NO), a subregion of the CX, where FB but not EB neurons flipped turn preference depending on the visual environment. These results therefore suggest that the heading direction system of Drosophila is composed of columnar neuron networks spanning the EB and the FB, where heading and angular signals from the EB are combined with information about visual context in FB neurons (Shiozaki, 2020).
This study analyzed neural dynamics in the navigation system of Drosophila by performing cell type-specific calcium imaging during flight. Columnar neurons in the FB and the EB showed coordinated population dynamics that encode ongoing turning behavior and, in short timescales, heading direction. A group of FB neurons flipped the preference for turn direction depending on the visual environment, whereas analogous neurons in the EB showed invariant turn tuning. These data suggest that the heading direction system in Drosophila is composed of multiple interacting circuits that distinctly integrate visual and self-motion information (Shiozaki, 2020).
Silencing of neurons in the FB impairs aspects of navigation, such as memory-guided flight orientation. However, little was known about the information encoded by FB neurons aside from flight-dependent change in visual and baseline calcium signals. The current data show that a population of columnar neurons in the FB encodes flight turning behavior as circular dynamics. The position and velocity of the activity bump in the FB were correlated with a fly's turning behavior. Moreover, the bump position was also correlated with a fly's heading in short timescales. Thus, columnar neurons in the FB multiplex information about two aspects of animal navigation (Shiozaki, 2020).
This population activity in the FB was coordinated with that in the EB, suggesting that they carry similar information. Indeed, columnar neurons in the EB encoded turning behavior and heading as in the FB. Unlike a previous study reporting that the movement of the bump in E-PG neurons is uncorrelated with flight turns in darkness, this study found that turning behavior is correlated with the position and velocity of activity bump in E-PG neurons even in darkness. This apparent discrepancy likely resulted in part from differences in prior experience in closed-loop flight because E-PG neurons did not show turn-related activity in darkness when the flies performed flight with a visual pattern and a bar in advance, as in the previous study. This study also found that the bump position was influenced more by turning behavior than visual cues, unlike in walking flies, where visual but not motor cues dictate the bump position. Together, the activity bump of columnar neurons encodes different types of information depending on experience and behavioral states. On a relevant note, experimental and computational studies have proposed that the association between bump position and visual landmarks is formed through experience-dependent synaptic plasticity. For these proposed models to work, turning behavior must drive the movement of the bump, which was observed in the CX of flying flies as in walking flies (Shiozaki, 2020).
The bump position of columnar neurons in the FB and the EB was correlated with heading only for several seconds during flight with a visual pattern, suggesting that the offset between bump position and heading drifted away over the course of the recordings. Such a strong drift was not observed for E-PG neurons in walking flies. A previous study has shown that the offset can be unstable when the visual scene contains multiple identical objects. Thus, the stronger drift in the current study might stem from the visual pattern, which contained an array of identical bars. Consistent with this idea, the drift was weaker during flight with a single bar. It is likely that heading representations are stable in flies navigating in natural environments where visual scenes contain more features to identify heading than the scenes in the current study (Shiozaki, 2020).
In the FB and EB, the bump position was correlated with turning behavior with a delay of around 1 s. Thus, these turn signals potentially contribute to flight behavior operating on a timescale of seconds. Notably, menotaxis, a behavior that requires columnar neurons in the EB, involves changes in heading on such a timescale. However, it remains to be investigated whether and how the turn signals encoded as the bump position contribute to guiding flight (Shiozaki, 2020).
How are heading circuits organized in the CX? The data show that columnar neurons in the FB and EB coordinately encode heading and turning behavior during flight. It is proposed that this coordination originates in the communication from columnar neurons in the EB to those in the FB for two reasons. First, P-F-R and P-FN neurons have dendrites in the PB, where axons of E-PG neurons reside. Second, E-PG neurons can activate a type of local neuron in the PB, Δ7 neurons, whose activation can influence the activity of P-FN neurons. Therefore, columnar neurons in the FB might inherit heading signals from the EB, where heading is thought to be computed through recurrent connections among columnar neurons (Shiozaki, 2020).
However, information encoded by columnar neurons in the FB and EB are not identical. This study found that P-FN but not P-EN neurons flipped the preference for turning behavior between flight in darkness and with a visual pattern, suggesting that two types of neurons are differentially influenced by visual input. This difference might reflect signals conveyed from visually responsive neurons that have dendrites outside of the CX and axons either in the EB (ring neurons) or in the FB (e.g., ExFl1 neurons). In addition, P-FN and P-EN neurons innervate different compartments in the NO, where axons of different sets of neurons projecting from the lateral accessory lobe terminate. Although the mechanism underlying this difference awaits further investigation, the current results suggest that columnar neurons in the FB likely do not just relay heading signals but integrate them with other inputs. Consistent with this idea, silencing of neurons in the FB and the EB have different effects on navigation (Shiozaki, 2020).
Although P-FN neurons flipped their turn preference in the NO depending on the visual environment, their branches in the FB did not (i.e., the sign of the correlation between turn direction and bump rotation was invariant). This suggests that calcium signals in the NO branches of P-FN neurons are modulated sub-cellularly, as in the EB branches of P-EN neurons (Shiozaki, 2020).
Columnar neurons showed turn-related activity in darkness, indicating that it derives from non-visual cues. One possibility is that the activity represents an efference copy because the CX receives input from putative pre-motor regions, including the lateral accessory lobe and the posterior slope. Alternatively, turn signals might reflect sensory feedback from, for example, mechanosensory neurons in the antennae, which are active during flight. Although the columnar neurons in the EB also encode turns during walking in darkness, its neural underpinnings likely differ from those during flight because the two modes of locomotion involve distinct patterns of neural activation in motor and sensory systems (Shiozaki, 2020).
Activity of P-F-R neurons increased during flight as in other types of CX neurons. This activity might be inherited from neurons in the optic lobe, whose activity increases during flight. Alternatively, this phenomenon might be mediated by octopaminergic neurons innervating the CX because octopamine regulates flight-dependent neural modulation in the optic lobe. Beyond mechanisms, it will be important to determine whether and how flight-dependent neural modulation contributes to sensory processing and behavior (Shiozaki, 2020).
Networks of columnar neurons in the FB are upstream of the motor system; a type of columnar neurons in the FB, PF-LCre neurons, send axons to the lateral accessory lobe, where dendrites of descending neurons reside. Because PF-LCre neurons are considered to be a primary output channel of the CX, the FB likely plays a unique role in modulating locomotion. In fact, suppressing the activity of columnar neurons in the FB influences walking behaviors in Drosophila, and electrical stimulation of the FB modifies walking in cockroaches. Because E-PG neurons are necessary for adopting arbitrary heading relative to a visual landmark but are dispensable for phototaxis, the FB might modulate elementary motor programs based on spatial information computed within the CX (Shiozaki, 2020).
By gathering the knowledge acquired in various insects, recent work proposed a computational model of the CX network that is capable of performing path integration. In this model, E-PG and P-FN neurons have distinct functions; E-PG neurons encode heading, whereas P-FN neurons integrate heading and speed signals over time to calculate the direction and distance an animal has traveled. Although this study has analyzed just one of a few types of P-FN neurons, the results suggest that P-FN neurons encode heading and turning behavior in a manner similar to E-PG neurons. Monitoring the activity of various types of P-FN neurons in flies engaged in behaviors that require path integration would provide data for direct tests of the model (Shiozaki, 2020).
Neurons innervating the dorsal part of the FB promote sleep, during which locomotion is suppressed. In the current study, neurons in this part did not show circular dynamics, suggesting that these sleep-promoting neurons do not encode spatial information in the same way as FB columnar neurons. It is tempting to speculate that the sleep circuit might rather inhibit locomotion by downregulating the activity of FB columnar neurons; for example, through the EB. Similar processing may shape other behaviors requiring the FB, such as memory-guided flight orientatio, courtship memory, protein seeking, nociceptive avoidance, and aggression, because locomotion is a key building block of these behaviors. The FB might mediate the interaction between spatial representations and the internal state to dictate when and in which direction an animal moves (Shiozaki, 2020).
Object-based attention describes the brain's capacity to prioritize one set of stimuli while ignoring others. Human research suggests that the binding of diverse stimuli into one attended percept requires phase-locked oscillatory activity in the brain. Even insects display oscillatory brain activity during visual attention tasks, but it is unclear if neural oscillations in insects are selectively correlated to different features of attended objects. This question was addressed by recording local field potentials in the Drosophila central complex, a brain structure involved in visual navigation and decision making. Attention was found to selectively increase the neural gain of visual features associated with attended objects; attention could be redirected to unattended objects by activation of a reward circuit. Attention was associated with increased beta (20to 30-Hz) oscillations that selectively locked onto temporal features of the attended visual objects. These results suggest a conserved function for the beta frequency range in regulating selective attention to salient visual features (Grabowska, 2020).
Selective attention refers to the brain's capacity to focus on a subset of stimuli while ignoring others. While subjectively intuitive in humans, selective attention has also been documented in a wide variety of animals, such as other primates, birds, and even insects. What is attended to depends on stimulus salience (e.g., loudness or brightness), as well as on the perceived value of a stimulus and the motivational state of the animal. What is attended to also depends on what is perceived as a singular object. Object-based attention refers to the capacity to direct attention to a conjunction of different features linked as part of the same object. Attending to one feature of a given object would thus enhance not only the neural representation of that particular feature, but also other features that are associated with the object. How this form of generalization works is not entirely understood but seems to require some form of feature binding to first determine which stimuli belong together as a unified object and then to link the object to some inherent value, or valence. Hence, feature binding appears to be essential for object-based attention, as neural gain has to be allocated to specific features first in order to perceive an object as a whole. At the same time, distinct stimulus features can become unbound from an attended object if they are selectively ignored (Grabowska, 2020).
In the mammalian brain, feature binding and object-based attention have been proposed to be associated mechanisms, both of which seem to be facilitated by synchronized activity of neuronal assemblies, which can be detected as phase-locked neural oscillations. In particular, oscillations in the range of 13 to 30 Hz (beta) and 30 to 80 Hz (gamma) seem to reflect this form of binding based on their strong synchronization at various time points following visual or auditory stimulation, with distinct oscillatory processes potentially reflecting different levels of perception. For example, early (<100-ms) stimulus-evoked synchronization in the gamma range has been suggested to represent rapid integration of unconscious sensory processes, whereas later (200to 400-ms) synchronization in both the beta and gamma range is hypothesized to reflect feature binding and conscious perception. Stimulus-evoked beta and gamma oscillations would thus represent a phase reset of ongoing neuronal activity associated with enhancing attentional gain for specific features, by facilitating information transfer or binding among different brain regions (Grabowska, 2020).
While there is neural evidence for object-based attention in nonhuman primates, it is unknown if the smallest animal brains, such as those of insects, combine diverse sensory stimuli into unified percepts, or if they even have a subjective awareness. Behavioral studies in honeybees suggest that some insects can detect illusory contours as single objects and can group distinct stimuli into abstract concepts such as 'sameness' or 'difference', which could indicate a form of categorization through object-based attention. Similarly, visual learning paradigms for Drosophila melanogaster have uncovered a capacity for context generalization, where flies perceived visual objects as the same despite changes in color, suggesting they were attending to the object shape feature and ignoring color cues. There is growing evidence for attention-like processes in insects, such as during visual fixation, decision making, and novelty detection in Drosophila flies, as well as multiple object tracking in dragonflies. The latter electrophysiological study uncovered motion-detecting neurons in dragonflies that selectively lock onto the timing or phase of salient objects, which was shown by 'tagging' competing objects with distinct flicker frequencies. However, it is unknown how such selective neural processes are controlled in the insect brain or whether these neural measures are relevant to behavioral decision making (Grabowska, 2020).
It is possible that the insect brain, like the mammalian brain, employs oscillatory activity and stimulus-evoked phase locking to prioritize and bind stimulus features, and to enhance attentional gain. Indeed, earlier electrophysiological studies revealed oscillatory activity in the 20 to 30-Hz range in the Drosophila brain that was associated with detecting salience effects, such as visual novelty, suggesting that these endogenously generated oscillations might be more broadly involved in regulating attention-like processes in the fly brain. However, it remained unclear which neurons might be generating these oscillations. One likely neuropil is the central complex (CX), a heterogeneous structure in the central brain that has been associated with visual pattern learning. Recent studies in behaving Drosophila also identified the CX as a key brain region for visual navigation. This suggests a broader role for the CX in directing attention-like processes, which could also reflect ring attractor dynamics within CX circuits. While its role in visual perception is increasingly evident, whether the CX produces neural oscillations relevant to visual attention and feature binding is unknown. To address this question requires not only measuring electrical activity in the CX of behaving flies, but also correlating any endogenous brain activity to distinct neural signatures associated with competing visual stimuli or stimulus features (Grabowska, 2020).
In tethered virtual reality experiments, flies tend to fixate on large objects and avoid small objects, whether they are flying or walking. This innate visual dichotomy was explored to examine mechanisms underlying visual selective attention in Drosophila. To disambiguate between the attractive and aversive stimuli in the fly brain, and to relate neural activity to ongoing behavioral choices, local field potentials (LFPs) were recorded from the CX, and the competing visual stimuli were made to flicker at distinct frequencies, thereby evoking steady-state visually evoked potentials (SSVEPs) in the fly brain. It was first shown that the SSVEPs varied in amplitude depending on the visual objects being fixated upon, allowing an investigation of how attention guided the binding of different visual features, such as object size, brightness, and flicker frequency. By calculating phase-locking strength between the distinct SSVEPs and endogenous brain activity, how oscillations in the CX interacted with one another was then examined. Frequency-specific phase locking between endogenous oscillations was found in the 20 to 30-Hz frequency range and the object features that the fly paid attention to, suggesting that beta-like oscillations could be employed for object-based attention in the insect brain (Grabowska, 2020).
The brain's ability to link complex patterns of sensory input into coherent objects has been termed the binding problem, with the 'problem' being that it remains unclear how diverse sensory streams are unified into a single conscious percept. Human subjective experience of the world is of discretely bound units rather than segregated sensory streams, and this capacity of the human brain is probably adaptive as sensory cues are often correlated, such as voices with faces or fruits with colors. The adaptive advantage of perceiving the world in such a unitary fashion raises the question of whether other brains do this and if so, whether the selective attention mechanisms observed in simpler animals such as insects facilitate a form of feature binding (Grabowska, 2020).
In humans, the modulation of endogenous beta (15 to 30-Hz) oscillations is associated with the perception and integration of visual stimuli, as well as decision making, among other cognitive functions. Intriguingly, beta oscillations have also been associated with task-related engagement and reward processing as well as stimulus-locked attentional load effects. However, a full understanding of how beta oscillations are deployed to achieve these functions is lacking, and there remains debate regarding their functional role. Finding beta-like oscillations involved in object-based attention in the insect brain lends support to the view that these oscillations perform a conserved function relevant to perception, as it seems unlikely that a completely different neuroanatomy (an insect brain) would have preserved a neural epiphenomenon. Consistent with a causal role for oscillations in the insect brain, this study found that the neuropeptide F (dNPF) circuit activation in Drosophila increased 20 to 30-Hz activity in the CX, which promoted phase locking to attended visual stimuli. Interestingly, in open-loop conditions dNPF activation seemed to produce a valence reversal, suggesting that attention was redirected covertly to the smaller, aversive object. Why the smaller object should have higher value in this specific open-loop context remains unclear. An alternative interpretation of this result is that salience for the smaller object was increased, rather than it having been rendered more attractive. Thus, the dNPF circuit might be more involved with regulating salience rather than valence, and the salience of the aversive small bar could thus have been magnified by dNPF activation in open-loop conditions, when the fly is not in control. Interestingly, the increased salience assigned to the competing small bar was associated with increased walking speed, suggesting a motivation to respond behaviorally (Grabowska, 2020).
Electroencephalography (EEG) studies have found that when humans have no control over an array of emotionally laden visual images, these images evoke a higher SSVEP response compared with emotionally neutral images; however, strongly aversive images evoked the greatest SSVEP responses of all. Perhaps similarly in the fly brain, an uncontrollable aversive object becomes much more salient upon dNPF activation. This highlights the importance of accounting for behavioral control in any understanding of brain functions underlying perception, including flies in open vs. closed-loop experiments (Grabowska, 2020).
Beta-like oscillations have been observed previously in the insect brain. For example, recordings in the locust have identified 20 to 30-Hz oscillations associated with processing of olfactory stimuli, and comparable oscillations have also been associated with visual attention in flies. Additionally, there is increasing evidence that insect brains employ a variety of oscillations, comparable in range with the mammalian brain. These include 7 to 12 Hz (alpha), 20 to 50 Hz (beta and gamma), and even 1 Hz (delta). These oscillations have been shown to be involved in processes such as olfaction, vision, and sleep, suggesting conserved functions that might transcend the differences in brain architecture between insects and mammals. Whether any of these oscillations are functionally comparable remains to be seen. Nevertheless, the current findings suggest that beta-like oscillations might be employed by the insect brain to bind different stimulus features into unified percepts that guide the animal's attention. Although this study did not investigate nonvisual stimulus modalities in this study, previous work has demonstrated that odors modulate the amplitude of visually evoked 20 to 30-Hz activity, suggesting these oscillations might govern cross-modal binding as well. Whether endogenous 20to 30-Hz activity in the fly brain is performing a similar function to beta oscillations in the human brain remains an open question. It is, however, possible that oscillatory processes are supported by different brain architectures that have conserved circuit timing relationships through evolution. Such conservation might be expected if these oscillations were performing a key function for a variety of adaptive behaviors, such as navigation, finding food, or avoiding predators. This study suggests that oscillations in the beta range (20 to 30 Hz) are indeed performing an important phase-locking function to choreograph meaningful information and thereby, guide selective attention. Although mammalian and fly brains are obviously different, they share some organizational principles that could support the preservation of such oscillatory functions (Grabowska, 2020).
To determine whether the significant phase-amplitude correlations observed were due to a physiologically relevant shift between SSVEP phases and endogenous 20to 30-Hz oscillations, rather than just due to increases in stimulation frequency amplitudes, a simulation was performed where the amplitudes of only the SSVEPs were artificially increased while keeping other frequencies constant. In the simulation, no effect of LFP amplitudes on ESC were found. The simulation was then increased, with a specific increase in 20 to 30-Hz and 30 to 40-Hz amplitudes, and saw no correlation to the SSVEPs was seen. This indicates that the phase correlations observed in real fly brain activity are functionally relevant and not a by-product of multiple superimposed oscillations of varying amplitudes (Grabowska, 2020).
Although 20 to 30-Hz activity stood out as relevant for phase locking to attended objects, other endogenous frequencies showed significant changes upon visual stimulation. In open-loop conditions, visual stimulation alone (without NPF activation) led to an increase in phase-amplitude coupling between SSVEPs and endogenous frequencies in the gamma range (30 to 50 Hz). In the mammalian brain, gamma oscillations have been proposed to provide different functions in sensory processing, depending on the frequency range and timing poststimulus induction. For example, EEG activity in the lower gamma range (30 to 40 Hz) can be elicited by brief and steady visual stimuli, and an increase in oscillatory power for this frequency range can be observed up to 100 ms after stimulation. One idea is that these stimulus-locked gamma oscillations might be relevant for rapid (i.e., unconscious) integration processes that might not necessarily be stimulus relevant. Nevertheless, gamma oscillations in humans can also be significantly modulated by attention and stimulus saliency. In contrast, a nonstimulus-locked component in the gamma range, occurring around 250 to 350 ms after stimulus presentation, has been proposed to be more relevant for object representation. Intriguingly, a similar frequency shift is seen in the fly brain. This study observed an increase of 30 to 50-Hz phase locking when visual stimuli were presented, while 20 to 30-Hz phase locking predominated upon NPF circuit activation. In humans, it has been shown that synchronized oscillations in the gamma and beta ranges have a high degree of interdependence, showing a so-called 'gamma-to-beta' transition in response to novel auditory stimuli, for example. Whether a gamma-to-beta transition is also occurring in the insect brain, associated with visual perception, remains difficult to address because any evidence for perception in flies must ultimately depend on behavior, which occurs on a slower timescale than stimulus-evoked neural oscillations (Grabowska, 2020).
By grounding this study on innate visual preferences, it could be, however, infered what the flies were most likely paying attention to. An innately attractive visual object was found to evokes a greater response in the fly brain than an aversive object and that this effect is preserved even under open-loop conditions, when flies are not in control. This suggests a neural correlate of object-based attention or in other words, a brain signal that correctly identifies what a fly is paying attention to-even in the absence of correlated behavior. Although this remains speculative, future experiments tapping directly from this brain signal in closed-loop paradigms should be able to test if it indeed provides a level of cognitive control (Grabowska, 2020).
One hallmark of the visual system is a strict retinotopic organization from the periphery toward the central brain, where functional imaging in Drosophila revealed a spatially accurate representation of visual cues in the central complex. This raised the question how, on a circuit level, the topographic features are implemented, as the majority of visual neurons enter the central brain converge in optic glomeruli. This study discovered a spatial segregation of topographic versus nontopographic projections of distinct classes of medullo-tubercular (MeTu) neurons into a specific visual glomerulus, the anterior optic tubercle (AOTU). These parallel channels synapse onto different tubercular-bulbar (TuBu) neurons, which in turn relay visual information onto specific central complex ring neurons in the bulb neuropil. Hence, these results provide the circuit basis for spatially accurate representation of visual information and highlight the AOTU's role as a prominent relay station for spatial information from the retina to the central brain (Timaeus, 2020).
Like various other sensory modalities for which spatial information is critical, neural circuits in the visual system of many animals are organized in a topographic fashion to maintain the neighboring relationship of adjacent pixels detected by photoreceptors in the periphery, along the visual pathways into the central brain. The topographic representation of different kinds of sensory information within the central brain of Drosophila is currently being investigated using molecular genetic tools in combination with cell-type-specific driver lines. Although it is well known that spatially patterned visual stimuli induce coherent activity bumps in the Drosophila CX, the pathway translating peripheral visual information into central activity patterns remains poorly understood (Timaeus, 2020).
This study has shown that medulla inputs to the AOTU fall into two morphological types regarding their arborization patterns: broad innervation versus spatially restricted axon terminals. In both cases, only a single domain within the AOTU is targeted. Although the topographic representation from the lobula neuropil is mostly lost in the broad innervation pattern of converging and intermingling LC projection neurons onto the majority of optic glomeruli, a unique spatial organization could be identified for the output channel from the medulla. Topographic representation of the medulla (at least its dorsal half, where most driver lines used in this study are expressed) is maintained in the SU of the AOTU, which is spatially separated from lobula representation within the AOTU (the LU). Interestingly, a strict topographic correlation only exists between the a-p position of the dendritic fields of MeTu projection neurons in the medulla and their restricted axon termination along the d-v axis within distinct domains of the SU in the AOTU. No such topography exists along the d-v axis in the medulla. These neurons are therefore well suited for filtering out specific visual information (such as landmarks or celestial bodies) for guiding heading decisions during visually guided navigation (Timaeus, 2020).
Based on their morphology, as well as their molecular identity, three principle types of MeTu neurons provide input into the AOTU, with overlapping dendritic fields within the medulla but segregated axon terminals to distinct AOTU (sub-)domains. MeTu-l and -c classes have a similar neuronal morphology with dendrite arborization restricted to a single medulla layer (M6) and spatially narrow axon termination areas in four separate AOTU subdomains (SU-la, -lp, -ca, and -cp), thereby building several pathways arranged in parallel. The nomenclature of the SU subdomain organization differs slightly from previous studies, because it is now based on the expression patterns of different cell surface molecules, which might reflect the functional organization of these structures. Because of this new classification, both lateral and central domains (but not the medial domain) of the SU become further subdivided into anterior (SU-la and SU-cp) and posterior halves (SU-lp and SU-cp). Nevertheless, it should be noted that the total number of subdomains remains the same in both nomenclatures, with the major difference being the posterior-lateral subdomain ('lp'), which has been attributed to the central domain (SU-cp) in this study, as part of the Connectin-positive central neuropil. Based on the connectome reconstruction of the hemibrain dataset, which reports in a total number of 347 MeTu neurons ('MC61-type'), it is estimated ∼60 MeTu neurons per topographic class (and twice that for MeTu-m cells), assuming an equal innervation of SU subdomains of similar volume. Because 8-12 TuBu neurons were counted from three independent expression lines, a convergence ratio from MeTu to TuBu neurons of about 8:1 to 5:1. Only the organization of the MeTu-l and MeTu-c neurons clearly enables a spatial projection of visual information from the columnar organization in the medulla to the corresponding AOTU domains, which seems well suited to relay topographic information along one spatial axis toward the central brain (Timaeus, 2020).
The borders of the SU compartments are respected by molecularly defined populations of TuBu neurons, thereby defining the next synaptic elements in the parallel pathways toward the bulb neuropil. Although this neuropil with its afferent (TuBu) and efferent (R neurons) channels has been intensively studied in recent years, there still remains a gap in knowledge concerning how precise synaptic connections convey topographic information to the central complex. Four major findings of the TuBu->EB circuit are revealed by study. First, the topographic position of TuBu dendrites in the SU is not translated into a defined position within the bulb but instead exhibits a targeting plasticity within a restricted bulb area. Secondly, although the recent dissection of the AOTU->EB pathways described the bulb as a tripartite structure including both afferent and efferent neurons, this picture can now be refined by highlighting that although this analysis of TuBu-neurons is mainly restricted to only two representative TuBu classes (one in the SU-lp and the other in the SU-ca domain), both these classes target to areas within the superior bulb (BUs). More broadly expressed driver lines revealed exclusive TuBu neuron innervation of the BUs, indicating that additional TuBu classes target to this bulb area. Thus, at least four different classes of TuBu neurons are expected to exclusively innervate the BUs (TuBu-la, TuBu-lp, TuBu-ca, and TuBu-cs), each of them connecting to a different set of output neurons, indicating an even more complex organization of the bulb, in particular the BUs. Thirdly, TuBu classes project onto dendritic areas of R neuron classes (so called “sectors”) within the bulb, and specific connections are formed between TuBu neurons and R neuron classes. Although it was not possible to identify three R neuron classes within the BUs, there probably exists a much higher diversity of connections within this small area of the bulb, reaching beyond the scope of this study. For instance, the postsynaptic partners of one subset of TuBu-ca neurons as well as neurons contacted by R2 and R5 dendrites remain to be identified. Additional postsynaptic partners other than R neurons are contacted by TuBu neurons, like contralaterally projecting neurons described in the locust and the bumblebee, which connect the AOTU units of both hemispheres (TuTu neurons) (Timaeus, 2020).
It appears therefore that topography is conserved within the AOTU output neuron projections toward the bulb and ring neurons, which is in good agreement with their physiological responses to visual stimuli, like bright objects. All ring neurons of the same type occupy the same ring layer within the ellipsoid body, raising the question of how topographic information is integrated within central complex neuropils. Interestingly, different MeTu neuron types with similar receptive fields may innervate different AOTU domains and thereby connect to different TuBu neuron populations forming parallel channels that then diverge within the bulb regions, where SU-lp and SU-ca efferents were found mapping onto separate ring neurons (R4d versus R2). Hence, at least two distinct topographic MeTu channels into the central brain could be defined. Although functional differences between the BUi and BUs have been described, functional studies have not yet compared the physiological responses of different TuBu classes or the responses of R neurons within the BUs. Based on the data presented in this study, it would be expected that retinotopic information in the BUs remains represented in the respective sector that is associated with their TuBu class (Timaeus, 2020).
A morphologically distinct class of MeTu cells is formed by MeTu-m cells. One distinguishing feature in respect to other MeTu cell types is that many cells arborize broadly in their respective AOTU domain. Axon terminals of single MeTu-m neurons invariably spread across the a-p axis of their SU-domain, whereas in the d-v axis they either covered their domain completely or partially-the former case being reminiscent of the afferent organization of LC neurons from the lobula within optic glomeruli in the PVLP regions, whereas the latter case is similarly described for lobula neurons innervating the AOTU's large unit (LU), where the topography of LC10 neurons in the LU has been analyzed, resulting in the distinction of four different LC10-classes. It remains to be seen whether MeTu-m neurons also could be divided into such classes. Those cells innervating the complete SU-m are well suited to form a nontopographic channel to the central brain. Interestingly, although topographic MeTu-l and -c neurons form dendritic fields within a single medulla layer, MeTu-m neurons integrate from three different medulla layers, reminiscent and in fact similar to some lobular LC neuron types, the main afferents of the AOTU large unit, for which a comparable rough topography along the dorsoventral axis has previously been found. Furthermore, only MeTu-m neurons form a collateral arborization in the lobula, indicating that this pathway could directly integrate visual information from both the medulla and lobula. The observation that MeTu-m neurons contact a population of TuBu neurons that projects into the inferior bulb area (Bui) separated from other TuBu neurons further suggests a different role for this pathway. A contralateral inhibition mediated by the Bui has been described, supporting a model in which the SU-m pathway is involved in suppressing ipsilateral stimuli with the expense of reduced spatial resolution (Timaeus, 2020).
Taken together, topographic and nontopographic afferents generate an interesting assembly of adjacent domains within the AOTU, from exclusively topographic medulla input in SU-l and SU-c domains, nontopographic medullar (and potentially also lobular) input in SU-m, and another large area of nontopographic input exclusively from the lobula in the LU. Thus, this study has identified multiple parallel topographic pathways separated from a parallel nontopographic channel (Timaeus, 2020).
This principle visual pathway involving the AOTU as a central relay station between medullar/lobular inputs and the central brain is widely shared among different insect taxa, where homologous structures can be found, e.g., orthopterans, hymenopterans, and beetles. The stimuli conveyed by this 'anterior visual pathway' have been addressed in only a few insect species so far. Most prominently, the AOTU has been associated with celestial orientation using polarized skylight in several species or in chromatic processing. Dorsal rim ommatidia harboring polarization-sensitive photoreceptors for polarized light vision are crucial for the sky-compass orientation and exist in most insects analyzed, such as locusts. However, it remains unknown whether MeTu neurons receive direct or indirect input from modality-specific cell types located in the DRA. In addition, processing of chromatic information was also shown to be accomplished via the AOTU in several insects. This study has now identified inputs to this pathway, by identifying direct connections between MeTu cells and UV-sensitive R7 photoreceptor cells in medulla layer M6 (Timaeus, 2020).
Furthermore, the molecular markers used in this study can serve as future tools to reveal the molecular mechanisms that underlie the formation of the LC-optic glomeruli network across species. Because Drosophila is among the smallest species for which the AOTU has been characterized and is believed to be a behavioral generalist, even more sophisticated architectures of the SU-homologue could exist in other insect taxa. On the anatomical and functional level, optic glomeruli share many features with the synaptic neuropil within the antennal lobe, which led to the postulation that the glomerular organization in the protocerebrum (optic glomeruli) and the deutocerebrum (olfactory glomeruli) are in fact homologous structures. Indeed, molecular characteristics in the PVLP and AOTU were found that resemble the combinatorial code of cell-surface proteins in the olfactory system (e.g. expression patterns of Ten-m, Con, Caps, and Sema1a in both systems). However, future developmental studies of mutant LC and MeTu neurons are needed to test to what extent common mechanisms of glomerular circuit assembly exist in both sensory systems. Although the idea of a serial homology of glomerular organized neural system is far from being resolved, it will be intriguing for further studies to analyze the developmental mechanisms that underlie the circuit formation of these parallel AOTU pathways and optic glomeruli circuits as well as to compare them with known molecular functions during olfactory system maturation (Timaeus, 2020).
The fact cannot be excluded that the SU of the AOTU might consist of additional functional units that so far have not been identified and that neurons were missed in this analysis due to the lack of expression lines to visualize them. Populations of neurons that were classified as a single type might turn out to be different enough (by morphology and/or synapse partners) to justify the establishment of further pathways, and these cell types might have been missed in the single cell labeling experiment, as this method involves random events where scarcer neurons can easily remain unnoticed. In vivo experiments measuring neuronal activity and responses to visual stimuli were beyond the scope of ths study but will be an essential part for understanding the functional features of the circuit. The wealth of genetic tools and their manifold combinations in Drosophila certainly provide capabilities of detailed analyses. As the driver lines used for this study to unravel the components of the visual pathway are publicly available and could be used to measure and manipulate neuronal activity, it is hoped that this study paves the way for future studies of components of this visual circuit (Timaeus, 2020).
The insect central complex (CX) is thought to underlie goal-oriented navigation but its functional organization is not fully understood. This study recorded from genetically-identified CX cell types in Drosophila and presented directional visual, olfactory, and airflow cues known to elicit orienting behavior. A group of neurons targeting the ventral fan-shaped body (ventral P-FNs) was found to be robustly tuned for airflow direction. Ventral P-FNs did not generate a 'map' of airflow direction. Instead, cells in each hemisphere were tuned to 45° ipsilateral, forming a pair of orthogonal bases. Imaging experiments suggest that ventral P-FNs inherit their airflow tuning from neurons that provide input from the lateral accessory lobe (LAL) to the noduli (NO). Silencing ventral P-FNs prevented flies from selecting appropriate corrective turns following changes in airflow direction. These results identify a group of CX neurons that robustly encode airflow direction and are required for proper orientation to this stimulus (Currier, 2020).
Autism spectrum disorder (ASD) is a neurodevelopmental disorder characterized by impairments in social interaction and repetitive stereotyped behaviors. Previous studies have reported an association of serotonin or 5-hydroxytryptamine (5-HT) with ASD, but the specific receptors and neurons by which serotonin modulates autistic behaviors have not been fully elucidated. RNAi-mediated knockdown was done to destroy the function of tryptophan hydroxylase (Trh) and all the five serotonin receptors. Given that ubiquitous knockdown of 5-HT2B showed significant defects in social behaviors, the CRISPR/Cas9 system was used to knock out the 5-HT2B receptor gene. Social space assays and grooming assays were the major methods used to understand the role of serotonin and related specific receptors in autism-like behaviors of Drosophila melanogaster. A close relationship was identified between serotonin and autism-like behaviors reflected by increased social space distance and high-frequency repetitive behavior in Drosophila. The binary expression system was further utilized to knock down all the five 5-HT receptors; the 5-HT2B receptor was observed to act as the main receptor responsible for the normal social space and repetitive behavior in Drosophila for the specific serotonin receptors underlying the regulation of these two behaviors. These data also showed that neurons in the dorsal fan-shaped body (dFB), which expressed 5-HT2B, were functionally essential for the social behaviors of Drosophila. Collectively, these data suggest that serotonin levels and the 5-HT2B receptor are closely related to the social interaction and repetitive behavior of Drosophila. Of all the 5 serotonin receptors, 5-HT2B receptor in dFB neurons is mainly responsible for serotonin-mediated regulation of autism-like behaviors (Cao, 2022).
LRP1, the low-density lipoprotein receptor 1, would be a novel candidate gene of epilepsy according to bioinformatic results and the animal study. This study explored the role of LRP1 in epilepsy and whether beta-hydroxybutyrate, the principal ketone body of the ketogenic diet, can treat epilepsy caused by LRP1 deficiency in drosophila. UAS/GAL4 system was used to establish different genotype models. Flies were given standard, high-sucrose, and ketone body food randomly. The bang-sensitive test was performed on flies and seizure-like behavior was assessed. In morphologic experiments, it was found that LRP1 deficiency caused partial loss of the ellipsoidal body and partial destruction of the fan-shaped body. Whole-body and glia LRP1 defect flies had a higher seizure rate compared to the control group. Ketone body decreased the seizure rate in behavior test in all LRP1 defect flies, compared to standard and high sucrose diet. Overexpression of glutamate transporter gene Eaat1 could mimic the ketone body effect on LRP1 deficiency flies. This study demonstrated that LRP1 defect globally or in glial cells or neurons could induce epilepsy in drosophila. The ketone body efficaciously rescued epilepsy caused by LRP1 knockdown. The results support screening for LRP1 mutations as discriminating conduct for individuals who require clinical attention and further clarify the mechanism of the ketogenic diet in epilepsy, which could help epilepsy patients make a precise treatment case by case (Zhang, 2022).
The FoxP family of transcription factors is necessary for operant self-learning, an evolutionary conserved form of motor learning. The expression pattern, molecular function and mechanisms of action of the Drosophila FoxP orthologue remain to be elucidated. By editing the genomic locus of FoxP with CRISPR/Cas9, this study found that the three different FoxP isoforms are expressed in neurons, but not in glia and that not all neurons express all isoforms. Furthermore, FoxP expression was found in the protocerebral bridge, the fan-shaped body and in motor neurons, but not in the mushroom bodies. Finally, this study discovered that FoxP expression during development, but not adulthood, is required for normal locomotion and landmark fixation in walking flies. While FoxP expression in the protocerebral bridge and motor neurons is involved in locomotion and landmark fixation, the FoxP gene can be excised from dorsal cluster neurons and mushroom-body Kenyon cells without affecting these behaviours (Palazzo, 2020).
The genomic locus of the Drosophila FoxP gene was edited in order to better understand the expression patterns of the FoxP isoforms and their involvement in behaviour. The isoforms differ with respect to their expression in neuronal tissue. For instance, isoform B (FoxP-iB) expression was found in neuropil areas such as the superior medial protocerebrum, the protocerebral bridge, the noduli, the vest, the saddle, the gnathal ganglia and the medulla, while areas such as the antennal lobes, the fan-shaped body, the lobula and a glomerulus of the posterior ventrolateral protocerebrum contain other FoxP isoforms but not isoform B. Previous results that FoxP is expressed in a large variety of neuronal cell types was corroborated. Genomic manipulations created several new alleles of the FoxP gene which had a number of behavioural consequences that mimicked other, previously published alleles. Specifically, it was found that constitutive knock-out of either FoxP-IB alone or of all FoxP isoforms affects several parameters of locomotor behaviour, such as walking speed, the straightness of walking trajectories or landmark fixation. Mutating the FoxP gene only in particular neurons can have different effects. For instance, knocking FoxP out in neurons of the dorsal cluster (where FoxP is expressed) or in MB Kenyon cells (where no FoxP expression was detected) had no effect in Buridan's paradigm, despite these neurons being required for normal locomotion in Buridan's paradigm. By contrast, without FoxP in the protocerebral bridge or motor neurons, flies show similar locomotor impairments as flies with constitutive knock-outs. These impairments appear to be due to developmental action of the FoxP gene during larval development, as no such effects can be found if the gene is knocked out in all cells in the early pupal or adult stages (Palazzo, 2020).
The exact expression pattern of FoxP has been under debate for quite some time now. Initial work combined traditional reporter gene expression with immunohistochemistry. A previous study created a FoxP-Gal4 line where a 1.5 kb fragment of genomic DNA upstream of the FoxP coding region was used to drive Gal4 expression. These the resulting expression pattern was validated with the staining of a commercial polyclonal antibody against FoxP. The same antibody was used in the current work and observed perfect co-expression with the reporter. The previous description of the FoxP expression pattern as a small number of neurons distributed in various areas of the brain, particularly in the protocerebral bridge, matches the current results (Palazzo, 2020).
Subsequent reports on FoxP expression patterns also used putative FoxP promoter fragments to direct the expression of Gal4. One study used a 1.4 kb sequence upstream of the FoxP transcription start site, while another used 1.9 kb. The larger fragment contained the sequences of the two previously used fragments. The latest study reporting on FoxP expression in Drosophila avoided the problematic promoter fragment method and instead tagged FoxP within a genomic segment contained in a fosmid, intended to ensure expression of GFP-tagged FoxP under the control of its own, endogenous regulatory elements. This study was the first to circumvent the potential for artifacts created either by selection of the wrong promoter fragment or by choosing an inappropriate basal promoter with the fragment. However, since they also used insertion of a transgene, their expression pattern, analogous to that of a promoter fragment Gal4 line, may potentially be subject to local effects where the fosmid with the tagged FoxP was inserted (Palazzo, 2020).
In an attempt to eliminate, the last source of error for determining the expression pattern of FoxP in Drosophila, CRISPR/Cas9 with homology-directed repair was used to tag FoxP in situ, avoiding both the potential local insertion effects of the previous approaches and without disrupting the complex regulation that may occur from more distant parts in the genome. For instance, in human cells, there are at least 18 different genomic regions that are in physical contact with the FOXP2 promoter, some of which act as enhancers. The effects of these regions may be disrupted even if the entire genomic FoxP locus were inserted in a different genomic region as in. Interestingly, the first promoter fragment approach and the fosmid approach agree both with the most artefact-avoiding genome editing approach and the immunohistochemistry with an antibody validated by at least three different FoxP-KO approaches. This converging evidence from four different methods used in three different laboratories suggests that FoxP is expressed in about 1800 neurons in the fly nervous system, of which about 500 are located in the ventral nerve cord. Expression in the brain is widespread with both localized clusters and individual neurons across a variety of neuronal cell types. Notably, the four methods also agree that there is no detectable FoxP expression in the adult or larval MBs. By contrast, in honey bees, there is converging evidence of FoxP expression in the MBs (Palazzo, 2020).
This comparison of the current data with the literature prompts the question why two different promoter fragment approaches suggested FoxP expression in the MBs (confirmed by a ribosome-based approach) when there is no FoxP protein detectable there (Palazzo, 2020).
A first observation used the classic hsp70-based pGaTB vector to create a Gal4 line, while two other studies used the more modern Drosophila synthetic core promoter (DSCP)-based pBPGUw vector. The two vectors differ with regard to their effects on gene expression. In addition to carrying two different basal promoters, the modern pBPGUw sports a 3'UTR that is designed to increase the longevity and stability of the mRNA over the pGaTB vector, which can result in twofold higher Gal4 levels (Palazzo, 2020).
This observation is complemented by single-cell transcriptome data. FoxP RNA can be detected in more than 4100 brain cells, likely overcounting the actual FoxP expression more than threefold. For instance, FoxP RNA is detected in over 1000 glial cells where none of the published studies has ever detected any FoxP expression (Palazzo, 2020).
Taking these two observations together, it becomes plausible that there may be transient, low-level FoxP transcription in some MB neurons (and likely thousands of other cells as well), which in wild-type animals rarely leads to any physiologically relevant FoxP protein levels in these cells. Only when gene expression is enhanced by combining some arbitrary promoter fragments with genetically engineered constructs designed to maximize Gal4 yield such as the pBPGUw vector, such transient, low-abundance mRNAs may be amplified to a detectable level (Palazzo, 2020).
These considerations may also help explain why the ribosome-based method of was able to detect FoxP RNA in MB Kenyon cells: the transcript that was detected may have been present and occupied by ribosomes, but ribosomal occupancy does not automatically entail translation. It remains unexplained, however, how a previous study failed to detect all those much more strongly expressing and numerous neurons outside of the MBs. All of the above is consistent with other insect species showing FoxP expression on the protein level in their MBs, as only limited genetic alterations would be needed for such minor changes in gene expression (Palazzo, 2020).
The stochasticity of gene expression is a well-known fact and known to arise from the transcription machinery. Post-transcriptional gene regulation is similarly well-known. It is thus not surprising if it is observed that many cells often express transcripts that rarely, if ever, are translated into proteins. The final arbiter of gene expression must therefore remain the protein level, which is why this study validated the expression analysis with the appropriate antibody. On this decisive level, FoxP has not been detected in the MBs at this point (Palazzo, 2020).
The genome editing approach allowed distinguishing of differences in the expression patterns of different FoxP isoforms. The isoform specifically involved in operant self-learning, FoxP-iB, is only expressed in about 65% of all FoxP-positive neurons. The remainder express either FoxP-iA or FoxP-iIR or both. Neurons expressing only non-iB isoforms are localized in the antennal lobes, the fan-shaped body, the lobula and a glomerulus of the posterior ventrolateral protocerebrum. Combined with all three isoforms differing in their DNA-binding FH box, the different expression patterns for the different isoforms adds to the emerging picture that the different isoforms may serve very different functions (Palazzo, 2020).
Alterations of FoxP family genes universally result in various motor deficits on a broad scale in humans and mice for both learned and innate behaviours. Also in flies, manipulations of the FoxP locus by mutation or RNA interference have revealed that FoxP is involved in flight performance and other, presumably inborn, locomotor behaviours as well as in motor learning tasks (Palazzo, 2020).
The locomotor phenotypes described so far largely concerned the temporal aspects of locomotion, such as initiation, speed or duration of locomotor behaviours. Using Buridan's paradigm, this study reports that manipulations of FoxP can also alter spatial aspects of locomotion, such as landmark fixation or the straightness of trajectories. The results further exemplify the old insight that coarse assaults on gene function such as constitutive knock-outs of entire genes or isoforms very rarely yield useful, specific phenotypes. Rather, it is often the most delicate of manipulations that reveal the involvement of a particular gene in a specific behaviour. This fact is likely most often due to the pleiotropy of genes, often paired with differential dominance which renders coarse neurogenetic approaches useless in most instances, as so many different behaviours are affected that the specific contribution of a gene to a behavioural phenotype becomes impossible to dissect (Palazzo, 2020).
In the case of FoxP, it was already known, for instance, that the different isoforms affect flight performance to differing degrees and that a variety of different FoxP manipulations affected general locomotor activity. This study shows that a complete knock-out of either FoxP-iB or all isoforms affected both spatial and temporal parameters of locomotion, but the insertion mutation FoxP3955 did not alter stripe fixation. Remarkably, despite the ubiquitous and substantial locomotor impairments after nearly any kind of FoxP manipulation be it genomic or via RNAi reported in the published literature failed to detect the locomotor defects of these flies (Palazzo, 2020).
While some of the manipulations used in this study did not affect locomotion significantly (e.g. knock-out in MBs or DCNs), most of them affected both spatial and temporal locomotion parameters, despite these parameters commonly not co-varying. Thus, while one would expect these behaviours to be biologically separable, the manipulations carried out in this study did not succeed in this separation (Palazzo, 2020).
Taken together, the results available to-date reveal FoxP to be a highly pleiotropic gene with phenotypes that span both temporal and spatial domains of locomotion in several behavioural modalities, lifespan, motor learning, social behaviour and habituation. It is straightforward to conclude that only precise, cell-type-specific FoxP manipulations of specific isoforms will be capable of elucidating the function this gene serves in each phenotype. With RNAi generally yielding varying levels of knock-down and, specifically, with currently available FoxP RNAi lines showing only little, if any, detectable knock-down with RT-qPCR, CRISPR/Cas9-mediated genome editing lends itself as the method of choice for this task. Practical considerations when designing multi-target gRNAs for FoxP prompted testing of the CRISPR/Cas9 system as an alternative to RNAi with an isoform-unspecific approach first, keeping the isoform-specific approach for a time when more experience in this technique has been collected. In a first proof-of-principle, CRISPR/Cas9 was used to remove FoxP from MB Kenyon cells, DCNs, motor neurons and the protocerebral bridge (Palazzo, 2020).
MBs have been shown to affect both spatial and temporal aspects of locomotion reported a subtle structural phenotype in a subset of MB Kenyon cells that did not express FoxP. As detailed above, two groups have reported FoxP expression in the MBs and it appears that some transcript can be found in MB Kenyon cells. With a substantial walking defect both in FoxP3955 mutant flies (which primarily affects FoxP-iB expression) and in flies without any FoxP, together with the MBs being critical for normal walking behaviour, the MBs were a straightforward candidate for a cell-type-specific FoxP-KO. However, flies without FoxP in the MBs walk perfectly normally. There are two possible reasons for this lack of an effect of this manipulation: either FoxP protein is not present in MBs or it is not important in MBs for walking. While at this point it is not possible to decide between these two options, the expression data concurring with those from previous studies suggest the former explanation may be the more likely one. Remarkably, a publication that did report FoxP expression in the MBs did not detect the walking deficits in FoxP3955 mutant flies despite testing for such effects. Motor aberrations as those described here and in other FoxP manipulations constitute a potential alternative to the decision-making impairments ascribed to these flies (Palazzo, 2020).
DCNs were recently shown to be involved in the spatial component (landmark fixation) of walking in Buridan's paradigm, but removing FoxP from DCNs showed no effect, despite abundant FoxP expression in DCNs. It is possible that a potential effect in stripe fixation may have been masked by already somewhat low fixation in both control strains. On the other hand, even at such control fixation levels, significant increases in stripe deviation can be obtained. Before this is resolved, one explanation is that FoxP is not required in these neurons for landmark fixation in Buridan's paradigm, while the neurons themselves are required (Palazzo, 2020).
Motor neurons are involved in all aspects of behaviour and have been shown to be important for operant self-learning. With abundant expression of FoxP in motor neurons, these neurons are considered a prime candidate for a clear FoxP-cKO phenotype. Indeed, removing FoxP specifically from motor neurons only, mimicked the effects of removing the gene constitutively from all cells. It is noteworthy that this manipulation alone was sufficient to affect both temporal and spatial parameters, albeit only one of the two driver lines showed clear-cut results. One would not necessarily expect motor neurons to affect purportedly 'higher-order' functions such as landmark fixation. It is possible that the higher tortuosity in the trajectories of the flies where D42 was used to drive the UAS-gRNA construct is largely responsible for the greater angular deviation from the landmarks in these flies and that this tortuosity, in turn is caused by the missing FoxP in motor neurons. Alternatively, D42 is also driving in non-motor neurons where FoxP is responsible for landmark fixation. The driver line C380 showed similar trends, albeit not quite statistically significant at an alpha value of 0.5%, suggesting that potentially the increased meander parameter may be caused by motor neurons lacking FoxP (Palazzo, 2020).
The protocerebral bridge is not only the arguably most conspicuous FoxP-positive neuropil, it has also been reported to be involved in temporal aspects of walking. Moreover, the protocerebral bridge provides input to other components of the central complex involved in angular orientation. Similar to the results in motor neurons, removing FoxP from a small group of brain neurons innervating the protocerebral bridge, phenocopies constitutive FoxP mutants (Palazzo, 2020).
Taken together, the motor neuron and protocerebral bridge results suggest that both sets of neurons serve their locomotor function in sequence. At this point, it is unclear which set of neurons precedes the other in this sequence (Palazzo, 2020).
There is ample evidence that the FoxP family of transcription factors acts during development in a variety of tissues. What is less well known is if adult FoxP expression serves any specific function. A recent study in transgenic mice in operant conditioning and motor learning tasks showed postnatal knock-out of FOXP2 in cerebellar and striatal neurons affected leverpressing and cerebellar knock-out also affected motor-learning. At least for these tasks in mammals, a FoxP family member does serve a postnatal function that is independent of brain development (brain morphology was unaltered in these experiments). Also in birds, evidence has been accumulating that adult FoxP expression serves a song plasticity function. The temporally controlled experiments in this study suggest that at least locomotion in Buridan's paradigm can function normally in the absence of FoxP expression in the adult, as long as FoxP expression remains unaltered during larval development. Future research on the role of FoxP in locomotion and landmark fixation hence needs to focus on the larval development before pupation (Palazzo, 2020).
Many insects use patterns of polarized light in the sky to orient and navigate. This study functionally characterized neural circuitry in the fruit fly, Drosophila melanogaster, that conveys polarized light signals from the eye to the central complex, a brain region essential for the fly's sense of direction. Neurons tuned to the angle of polarization of ultraviolet light are found throughout the anterior visual pathway, connecting the optic lobes with the central complex via the anterior optic tubercle and bulb, in a homologous organization to the 'sky compass' pathways described in other insects. This study detailed how a consistent, map-like organization of neural tunings in the peripheral visual system is transformed into a reduced representation suited to flexible processing in the central brain. This study identifies computational motifs of the transformation, enabling mechanistic comparisons of multisensory integration and central processing for navigation in the brains of insects (Hardcastle, 2021).
A critical challenge of active locomotion is knowing the right way to go. Sensorimotor reflexes can influence momentary changes in direction to hold a course or to avoid looming threats, but goal-directed behaviors, such as returning to a previous location from unfamiliar surroundings, require additional information and processing. External sensory cues must be transformed into an internal representation of position and orientation within the environment, Yhis can also be modified by past experience. In Dipteran flies, as in other invertebrates, a collection of neuropils known as the central complex (CX) is believed to coordinate such behaviors and plays a role in spatial memory, object memory, and action selection, in addition to homeostatic processes including hunger and sleep (Hardcastle, 2021).
Recent studies in Drosophila have revealed that activity in a network of CX neurons encodes and maintains a representation of the animal's angular heading relative to its environment, with similarity to head-direction cells in vertebrates. This neural representation of heading can be updated by internal, proprioceptive estimates of self-motion during locomotion, and by external cues, such as moving visual patterns and directional airflow. In other insects, including locusts, crickets, bees, butterflies, and beetles, the functional organization of the CX has frequently been studied in the context of navigation via celestial cues, particularly polarized light. The nearly ever-present pattern of polarization in the sky, formed by scattering of light in the atmosphere, offers an indicator of orientation to organisms able to detect and interpret it, and may be more stable than terrestrial landmarks. In these non-Dipteran insects, a multimodal neural circuit transmits polarization signals from the eyes to the central complex. This circuit is known as the 'sky compass' pathway for its proposed role in processing skylight polarization patterns and information about the position of the sun to bestow an animal with a sense of direction. In Drosophila, the anterior visual pathway (AVP), which comprises neurons connecting the medulla, anterior optic tubercle, bulb, and ellipsoid body, has been postulated to represent the homologue of the sky compass pathway. Visual processing in the AVP appears to be segregated into three topographically organized, parallel streams, of which two have been shown to encode distinct small-field, unpolarized stimuli. The neurons involved in polarization processing in Drosophila have not been identified beyond peripheral circuits of the dorsal rim area, a specialized region of the eye for detecting skylight polarization (Hardcastle, 2021).
A detailed mapping of the relevant polarization-sensitive neurons would allow the exquisite genetic tools and connectomic studies available in Drosophila to be leveraged to understand the workings of the CX and its integration of multiple sensory modalities. Behavioral experiments have demonstrated that Drosophila orient relative to polarization patterns while walking and in tethered-flight. A comparative approach would therefore provide insight into the processing strategies employed across taxa as well as species-specific adaptations. Furthermore, it may be possible to reconcile the existing evidence of a common, fixed representation of polarization patterns in the CX of non-Dipteran insects with the emerging model of a flexible representation of both visual information and heading direction in the Drosophila CX. Alternatively, fundamental differences in the organization and processing of polarized light signals between species may reflect specialized navigational requirements (Hardcastle, 2021).
This study set out to test the hypothesis that the anterior visual pathway conveys polarized light signals from the eye to the central complex in Drosophila. Neurogenetic tracing techniques and in vivo calcium imaging were used to characterize the organization of the neurons at each stage and their coding and transformation of visual features. Parallel circuitry in the medulla conducts polarization signals from photoreceptors in the dorsal rim area to a stereotyped domain of the anterior optic tubercle. From there, a postsynaptic population of neurons projecting to the anterior bulb relays polarization signals to ring neurons of the ellipsoid body, and in turn, the 'compass neurons' of the central complex. The superior bulb multiplexes polarized and unpolarized light signals, while the inferior bulb does not appear to be involved in polarization processing. Finally, population responses in the central complex were examined, and hallmarks were found of a flexible encoding of a single angle of polarization which could be used to direct motor output for navigation behavior (Hardcastle, 2021).
This study demonstrated that each section of the Drosophila anterior visual pathway (AVP) contains polarization-tuned neurons. Together, they provide a circuit to convey polarized light signals from the specialized dorsal rim area of the eye to the compass neurons of the central complex, via the anterior optic tubercle and bulb. This study shows that the pathway also conveys information about unpolarized visual features. The encoding of multiple visual modalities, the similarities in the constituent neurons, and the organization of the neuropils which accommodate them, support the view that the AVP in Drosophila is homologous to the sky compass pathway described in locusts, bees, butterflies, and beetles, among other insects (Hardcastle, 2021).
The approach of this study to investigating the neural processing of polarization vision offered a number of advantages over traditional intracellular electrophysiology. Firstly, it allowed simultaneous recording from whole populations of neurons, which would otherwise be technically challenging. This study investigates the spatial organization of polarization responses in an individual animal. This may be key in understanding the central complex, where dynamic responses reflect circuit plasticity and depend on numerous factors, such as proprioceptive inputs, internal states and goal-direction. Next, targeted expression of calcium indicators allowed isolation of specific anatomical groups of neurons, such as specific TuBu or ring neuron populations, greatly increasing the repeatability of functional characterizations. Crucially, the identification of corresponding genetic drivers will enable silencing experiments, optogenetic stimulation, and multi-population recordings to probe circuit function in the future. Imaging of calcium indicators also facilitated the characterization of neurons whose axons are prohibitively thin for recording intracellularly. MeTu-like neurons, for example, have long been assumed to deliver polarization signals from the medulla to the anterior optic tubercle, and this study was able to confirm this by direct observation for the first time (Hardcastle, 2021).
Since each detector for polarized light in the dorsal rim area (DRA) essentially has a different field of view, the success of this approach depended on the ability to stimulate a sizable number of DRA ommatidia. Surprisingly, almost the full extent of the DRA was stimulated by polarized light originating from a single point in the visual field with a common angle of polarization. A wide range of polarization tunings was subsequently revealed in downstream neurons, supporting the idea that the Drosophila medulla dorsal rim area (MEDRA) analyzes the overall pattern of polarized light in the sky and extracts a predominant angle of polarization (AoP), rather than performing many local AoP estimates. During the morning and evening when D. melanogaster are most active, the pattern of polarization in the sky can be well approximated by a single, predominant AoP. DmDRA1 neurons appear to spatially integrate polarization signals from multiple columns of the MEDRA, and individual neurons heavily overlap each other. This could provide an additional robustness to occlusions of the sky or of the DRA itself and average out inconsistencies in the available light (Hardcastle, 2021).
The parallel circuitry between DRA R7, DmDRA1, and MeTu neurons in MEDRA columns, resembles the color-processing pathway found in non-DRA columns involving R7, Dm8, and Tm5c. MeTu neurons in the MEDRA may also integrate color signals, as their dendritic fields extend into the non-DRA medulla, indicating that color and polarization processing are compatible. Parallel circuits may support antagonistic processing of the color and polarization pathways downstream, potentially providing a means to selectively process polarization cues for navigation from the anti-solar hemisphere where they are strongest. The responses of DmDRA2 neurons that contact R8 in this study have not been functionally described, and these neurons may be differently integrated with color processing. Both parallel functions will likely need to be incorporated to build a complete conceptual model of skylight polarization processing in the medulla (Hardcastle, 2021).
In the anterior optic tubercle (AOTU), polarization-sensitive neuron populations were found entering and leaving the tubercle via the intermediate-lateral domain. Polarization responses in the lateral domain were also observed, although it is unclear whether this is a result of separate polarization-sensitive MeTu types projecting from the MEDRA to different AOTU domains. Alternatively, since MeTu neurons are also postsynaptic in the AOTU, signals from a single polarization input channel could be redistributed to different regions of the AOTU for integration with other visual modalities or bilateral interactions. The AOTU in Drosophila is also likely to be a site for modulation of signals depending on time or internal states, and a capacity to modify responses may explain why multiple polarotopic organizations were observed in a MeTu neuron population in the AOTU. However, there may also be multiple functional subtypes within the population that more tailored experiments may be able to distinguish (Hardcastle, 2021).
Intriguingly, none of the polarotopies found in presynaptic MeTu neurons matched the polarotopy of postsynaptic TuBu dendrites in the AOTU, which was extremely consistent across animals. The findings suggest that TuBu neurons extract a processed form of the signals in the AOTU, encoding visual features within fewer neurons than the MeTu populations. TuBu neurons appear to divide signals into functional groups, and the anterior bulb-projecting TuBua group in every fly contained a set of around six tunings covering -90° to +90° of polarization space in approximately 30° steps, tightly packed in a micro-glomerular structure with no apparent polarotopy. The question remains open as to whether a sun position system and skylight polarization system are independent in the bulb. Unlike the TuLAL neurons in locusts (homologous to TuBu), where there is convergence on the dendrites of postsynaptic neurons, TuBu neurons appear to form one-to-one contact with individual ring neurons. Hence, it is posited that the site of integration of celestial cues is not at the synapse between TuBu and ring neurons. Although evidence was found that angles of polarization are represented in the superior bulb, where unpolarized cues are also known to be represented, the populations that were recorded contained a limited range of tunings and resembled a system for detecting visual features with a particular polarization signature, such as horizontally polarized light reflected from surfaces like water, rather than a system for accurate estimation of orientation. Such responses would likely be mediated by more ventral regions of the eye than the DRA. It should be noted that the polarized light stimulus broadly illuminated the eye from a dorsal position and, although attempts were made to minimize reflections, whether reflected polarized light fell on the ventral eye during these experiments was not measured (Hardcastle, 2021).
By recording the ensemble response of a population of ER4m ring neurons, both in the anterior bulb and ellipsoid body (EB), it was determined that they do not simply relay the responses of presynaptic TuBua neurons to the EB. Instead, they appear to deliver a subset of signals more prominently than others, bestowing the population with an ensemble response tuned to a specific angle of polarization. Furthermore, this study found that this population tuning conveys a different angle of polarization in individual animals, and one exciting possibility is that this represents a flexible heading signal relative to polarized light cues, which could direct behavior). A question to address in future work is whether the preferred angle of polarization of an individual ring neuron is itself fixed, in which case this study may have observed the result of a winner-take-all competition among the ER4m population in the EB, or if the whole population flexibly re-tunes to preferentially respond to a common AoP. Recordings from individual neurons will be required to resolve this (Hardcastle, 2021).
It is clear that among ER4m and E-PG neurons, polarization tunings are not represented with a retinotopic map in the EB or PB that is common between individual animals. This is in contrast with the consistent polarotopic organizations found upstream in the MEDRA or AOTU, but in agreement with a previous study which showed that the azimuthal position of unpolarized visual stimuli is also not represented retinotopically in E-PG neurons. The lack of organization in E-PG responses also matches previous findings in the corresponding CL1a neurons in locusts, but contrasts with the polarotopic organization found in other columnar neurons in the locust CX, such as CPU1, and the tangential TB1 neurons. A potential explanation for the lack of consistent polarotopy in CL1a, or indeed E-PG neurons, was offered in a previous study: at least two of each neuron type innervates an individual glomerulus in the PB. Could each of these have differential responses to polarized light to enable different configurations across the PB? Intriguingly, the TB1-like Δ7 neurons in the Drosophila PB appear to synapse onto only a subset of the E-PG neurons in a single glomerulus, perhaps indicating independent functional groups. Therefore, a polarotopic organization of responses in the Drosophila CX might yet be found. Alternatively, such an organization may reflect a common, genetically pre-programmed directional goal to facilitate migration, which flies may lack, instead using polarization cues to follow a fixed course and disperse along idiosyncratic heading (Hardcastle, 2021).
The current data suggests that in a given fly, E-PG neurons may respond to one of two approximately orthogonal angles of polarization, effectively dividing the population into two groups. Interestingly, when data from locust CPU1 neurons (likely homologues of P-F-R neurons in Drosophila) were pooled with tunings obtained from a number of other polarization-sensitive columnar CX neuron types, including CL1b (P-EG), CL2 (P-EN), CPU2, and CPU4 (P-FN), the organization of tunings in the locust PB could be interpreted as clustering around two orthogonal preferred angles. A binary system such as this would be well suited to influence downstream processes in a motor-centered coordinate frame. For example, the eventual output of the compass network may be a command signal to activate one descending neuron of a bilateral pair to initiate a turn to either the left or right, and thus maintain a heading specified by polarization patterns in the sky (Hardcastle, 2021).
An important next step will be to understand how polarized light influences the activity bump in columnar neurons and whether the activity of columnar neurons reciprocally influences the tunings of ER4m neurons. No activity bump was observed in E-PG neurons in the PB, likely due to the open-loop stimulus presentation as well as recordings performed in immobilized animals, although evidence was seen of flexible encoding of polarization information. According to these mappings of E-PG responses in the PB, the influence of a rotating polarized light stimulus might be to move the activity bump discontinuously between two positions, not dissimilar to observations in a recent investigation of the influence of airflow on the bump in E-PG neurons (Okubo, 2020). However, a limitation of the polarization stimulus used in this study is that the intensity gradient and position of the light source did not change as the angle of polarization rotated, as they would be seen by an animal turning under a natural sky. If the ambiguity between 0/180° polarization cues is resolved by integrating light intensity information, then the stimulus used in this study presented contradictory, unnatural changes (Hardcastle, 2021).
Behavioral studies in ants and dung beetles have demonstrated that skylight polarization cues can have a greater influence than other visual features in guidance and navigation behaviors. In Drosophila, intensity gradients have been shown to have a greater behavioral significance than polarized light, yet recent connectome analysis of the Drosophila CX highlights the polarization-sensitive ring neurons that were identified in this study as potentially being at the top of a hierarchy of sensory inputs. Furthermore, the unique pattern of asymmetrical connectivity between the ER4m populations from each brain hemisphere and the E-PG network hints at an attractively simple system for obtaining 360° heading information from ambiguous 0/180° polarization cues, by using signals from one population or the other depending on which side of the animal the sun is on. A key challenge for future studies will be to uncover such mechanisms for integrating and selecting from the multiple sensory modalities and visual qualities represented in the central brain in order to navigate complex environments (Hardcastle, 2021).
Spatial orientation memory plays a crucial role in animal navigation.
Recent studies of tethered Drosophila melanogaster (fruit fly) in a
virtual reality setting showed that the head direction is encoded in the
form of an activity bump, i.e., localized neural activity, in the
torus-shaped ellipsoid body (EB).
However, how this system is involved in orientation working memory is
not well understood. This study investigated this question using free
moving flies (D. melanogaster) in a spatial orientation memory task by
manipulating two EB subsystems, C and P circuits, which are hypothesized
for stabilizing and updating the activity bump, respectively. To this
end, two types of inhibitory ring neurons (EIP and P) which innervate
EB were suppressed or activated, and it was discovered that manipulating
the two inhibitory neuron types produced distinct behavioral deficits,
suggesting specific roles of the inhibitory neurons in coordinating the
stabilization and updating functions of the EB circuits. The neural
mechanisms underlying such control circuits were further elucidated
using a connectome-constrained spiking neural network model (Han, 2021).
Maintaining spatial orientation is a crucial cognitive capability required for animal navigation, and understanding the detailed neural mechanisms of spatial orientation is of great interest to researchers in the fields of neurobiology or neuromorphic engineering. In recent years, significant progress has been made in identifying the neural circuits that support spatial orientation in the central complex of Drosophila melanogaster. The central complex has long been associated with short-term spatial memory, visual pattern memory, and motor control. The recent discoveries of head-direction selectivity and localized neural activity in two central complex neuropils, the ellipsoid body (EB) and the protocerebral bridge (PB), have also linked the central complex to the function of spatial orientation. These studies suggested that the head orientation is encoded by localized neural activity, called activity bump, and the bump location in EB shifts in accordance with changes of heading during movement. The function of the EB neurons resemble that of a compass and is, therefore, termed 'neural compass' (Han, 2021).
In light of these empirical observations, several neural circuit models of the central complex have been proposed to elucidate the neural circuit mechanisms of head-direction selectivity or other functions associated with the central complex. Some models focused on the stability of the activity bump or on the differences in the circuit dynamics between locus and fruit fly. Other models studied the plasticity involved in the flexible retinotopic mapping but used simpler firing rate models or schematic models. A large-scale firing-rate neural network model that covered the entire central complex was able to reproduce the steering and homing behavior of bees, but the EB circuits were rather simple with minimal details (Han, 2021).
Recently, a spiking-neuron model of the EB-PB circuits was proposed. The model used a more realistic spiking-neuron model and synaptic dynamics to elucidate how the circuits can maintain a stable activity bump when fruit flies switch between forward movement and rotation states in the absence of landmarks. The model suggested the involvement of two subcircuits: one forms an attractor network and maintains (or stabilizes) an activity bump; the other forms a shifter network and shifts (or updates) the bump position in accordance with changes in body orientation. The model successfully demonstrated the angular errors when a fly moved in darkness
and predicted the asymmetric activity in the PB during rotation (Han, 2021).
The model made an important and unique prediction: the function of spatial orientation working memory requires coordinated activation of the bump-maintaining (or stabilizing) and bump-shifting (or updating) circuits that are controlled by the upstream ring neurons (Han, 2021).
However, most of the experimental studies used tethered flies in a virtual reality setting and focused on how manipulation of neurons affects the bump activity. It is not clear how these neurons, in particular those involved in stabilizing and updating the activity bump, play roles in cognition-relevant behavior such as spatial orientation memory in free-moving flies with a more realistic behavioral setting. This study aimed to address these questions and designed a behavioral task of spatial orientation working memory based on the classic Buridan's paradigm. Specifically, two types of GABAergic ring neurons were manipulated that are hypothesized to control these neurons. Ring neurons project their axons into EB and inhibit neurons including those display the activity bump. Previous studies have reported the roles of the ring neurons in visually-guided behavior, ethanol sensitivity, sleep regulation, olfactory memory and mating behavior. However, their roles in the working memory of spatial orientation in the presence or absence of visual cues remain unclear. In addition to the neural functional experiments, computer simulations were performed using the EB-PB model, which produced neural activities that were consistent with the behavioral changes observed in the fruit flies with different experimental conditions. The present study provides a detailed picture on how coordinated activation between the neural processes of stabilization and update plays a crucial role in spatial orientation working memory (Han, 2021).
It was hypothesized that the C circuit and P circuit in the EB circuits stabilize and update the orientation-encoding activity bump and they are regulated by corresponding GABAergic ring neurons. We tested this hypothesis by manipulating two types of GABAergic ring neurons in a spatial orientation working memory task with free moving fruit flies, and discovered manipulating each ring neuron type led to different behavioral abnormality. By performing computer simulations on a previously proposed EB-PB neural circuit model, it was possible to explain the results of the experiments and provided a picture of the neural circuit mechanism underlying spatial orientation working memory: the orientation-encoding bump is maintained through two alternately activated neural processes: one that stabilizes the position of the activity bump and one that updates the position of the bump. The former is activated when a fruit fly maintains a steady head direction and the latter is activated when the fly rotates its body. The control of this process is performed through specific GABAergic ring neurons. Therefore, overactivating or suppressing the ring neurons disrupts the alternation of the two processes and leads to incorrect or even loss of orientation memory (Han, 2021).
There are a few more interesting discoveries worth discussing. Performing fixation toward previous landmark directions requires two things to be remembered: the earlier event of the landmark presentation (what) and the directions of the landmarks (where). Flies that fail to remember the former would not exhibit the fixation behavior at all, while flies that forget the latter would still perform the fixation but toward incorrect directions. Discoveries of strong fixation but with large deviation from the true directions of the landmarks for flies with EIP-ring neuron suppression during the third stage may imply the segregation of the neural mechanisms of orientation memory regarding the 'where' and 'what' of a landmark (Han, 2021).
One interesting finding of the present study is a long duration of spatial orientation working memory during the poststimulus stage. Previous studies reported the occurrence of poststimulus fixation behavior that lasted only for a few seconds immediately following the offset of the landmarks. Indeed, this study observed that the flies tended to stop their movement a few seconds after the sudden disappearance of the landmarks in the third stage. But they usually resumed the movement in a few seconds. This might be the reason why earlier studies only claimed a few seconds of fixation if their analyses did not include the resumed movement. Further studies are needed to investigate this issue (Han, 2021).
A couple issues regarding the choice of molecular tools should be discussed. In the present study tub-GAL80ts was used in combination with UAS-Kir2.1 or UAS-TNT to suppress targeted ring neurons. The method involves raising the temperature 1 d before the behavioral experiments and therefore taking effects on a much longer time scale than using optogenetic tools. This long-term suppression may induce other effects at the cellular or circuit levels, which are beyond what the model can simulate. Further study may be required to carefully examine the long-term effects. An ideal solution is to transiently suppress the ring neurons using optogenetic tools such as UAS-NpHR (peak sensitivity wavelength ~589 nm) or GtACRs (peak sensitivity wavelength ~ 527 nm for GtACR1 and ~457 nm for GtACR2). However, the wavelength of the required activation light is within the visible range of the fruit flies. Preliminary tests on UAS-NpHR showed that the onset of the activation light seriously disrupted the fixation pattern of wild-type flies. A new optogenetic tool or a carefully re-designed optical system is required to transiently suppress targeted ring neurons while not interfering the visual experiments. The second issue is related to the GAL4 lines. In the present study only two most specific lines, c105-GAL4 and VT5404-GAL4, were used to target the EIP-ring and P-ring neurons, respectively. There are several other less specific GAL4 lines available for the two types of neurons. It is necessary to conduct the same experiments using these overlapping lines to further confirm the results presented in this study (Han, 2021).
Several other important questions remain to be addressed. Previous studies showed that EB does not maintain a fixed retinotopic map and a bump can start from a random location in the beginning of a trial (Seelig, 2015). For the sake of modeling simplicity, this study did not model the random starting point feature in the model. But this feature is easy to implement and does not affect the conclusion of this study. A global excitation needs to be applied to the entire EB to reset the system. The excitation will induce strong competition between the EIP neurons and a new bump will start at a random location through the winner-take-all dynamics. Following this issue, random regeneration of an activity bump also needs to be discussed. The model showed that photoactivation of either EIP-ring or P-ring neurons during the poststimulus stage permanently abolished the activity bump. However, based on the observation of spontaneous generation of activity bump in other studies (Seelig, 2015), the bump is likely to be regenerated at a random location after the offset of photoactivation. The regeneration can be easily implemented in the current model using the same mechanism described above. Since the regenerated bump starts from a random location, the fruit flies lose the reference to the landmark locations. Thus, adding a spontaneous bump or not both lead to the same conclusion: the files fail to fixate on the previous landmark locations. Although not affecting the conclusion of the present study, the spontaneous bump feature may be crucial in future studies that involve modeling of the steering mechanism (Han, 2021).
Another issue is that a couple experimental and modeling studies suggested that ring neurons provides the mechanisms underlying flexible retinotopic mapping in EPG (or EIP) neurons rather than the simple suppression/activation mechanism as hypothesized in the present study. However, the ring neurons (R2 and R4d) tested in one study are of different types from what was tested (R1 and R6) in this study. Additional experiments that measure the activities of R2 and R4d using the setups described in previous studies are required to clarify this issue (Han, 2021).
It is important to compare and discuss differences between computational models of the central complex in terms of the functions investigated in the present study. However, most models focused on different aspects of the compass circuit functions. The major difference from previous models is that they proposed that the PB intrinsic neurons as the main source of inhibition that regulates the attractor dynamics, while in the current model this function is conducted by the EIP ring neurons with two additional ring neuron types (C-ring and P-ring neurons) modulating different subcircuits of the system. An in-depth model comparison and experimental manipulation of PB intrinsic neurons and ring neurons under the present behavioral task may be able to clarify this issue (Han, 2021).
A final issue is related to the function of the activity bump which is commonly thought to represent the fly's sense of orientation in a manner similar to that of the head-direction system found in rodents. However, as aforementioned 'what' and 'where' mechanisms, performing the fixation behavior as an indication of orientation working memory may require several serial or parallel neural components beyond EB and PB. For example, how is this innate fixation behavior initiated (motivation)? When a fly stops fixating, it is not clear whether the fly forgets the landmark directions or simply enters a different behavior state (but still remembers the landmark directions). It also remains unclear whether the memory is stored in another neural circuit and the EB merely provides a reference frame for orientation, or whether the activity bump in the EB represents the actual memory of the landmarks. The current experimental setup is not able to address this issue. A novel task that can disassociate these two components is required for further investigation (Han, 2021).
The present study concludes the following. First, the experiment indicated that long-term suppression of EIP-ring neurons reduced the accuracy of orientation working memory (fixation with an increased deviation angle), whereas long-term suppression of P-ring neurons abolished the memory completely (no fixation). Similarly, transiently activating either ring neuron types in the absence of landmark immediately abolished the memory. Second, the experimental observation can be explained by the EB-PB neural circuit model in which the EIP-ring neurons are responsible for controlling the width of the bump and the P-ring neurons are responsible for shifting (updating) the position of the bump. Third, put the experiment and the theory together, the present study suggests that coordinated activation of the two ring neuron types which control the downstream EB-PB subcircuits is crucial for spatial orientation working memory (Han, 2021).
Insect neural systems are a promising source of inspiration for new navigation algorithms, especially on low size, weight, and power platforms. There have been unprecedented recent neuroscience breakthroughs with Drosophila in behavioral and neural imaging experiments as well as the mapping of detailed connectivity of neural structures. General mechanisms for learning orientation in the central complex (CX) of Drosophila have been investigated previously; however, it is unclear how these underlying mechanisms extend to cases where there is translation through an environment (beyond only rotation), which is critical for navigation in robotic systems. This study developed a CX neural connectivity-constrained model that performs sensor fusion, as well as unsupervised learning of visual features for path integration; the viability of this circuit was demonstrated for use in robotic systems in simulated and physical environments. Furthermore, a theoretical understanding is demonstrated of how distributed online unsupervised network weight modification can be leveraged for learning in a trajectory through an environment by minimizing orientation estimation error. Overall, these results may enable a new class of CX-derived low power robotic navigation algorithms and lead to testable predictions to inform future neuroscience experiments (Robinson, 2022).
Many behavioural tasks require the manipulation of mathematical vectors, but, outside of computational models, it is not known how brains perform vector operations. This study shows how the Drosophila central complex, a region implicated in goal-directed navigation, performs vector arithmetic. First, a neural signal in the fan-shaped body is described that explicitly tracks the allocentric travelling angle of a fly, that is, the travelling angle in reference to external cues. Past work has identified neurons in Drosophila and mammals that track the heading angle of an animal referenced to external cues (for example, head direction cells), but this new signal illuminates how the sense of space is properly updated when travelling and heading angles differ (for example, when walking sideways). A neuronal circuit was characterized that performs an egocentric-to-allocentric (that is, body-centred to world-centred) coordinate transformation and vector addition to compute the allocentric travelling direction. This circuit operates by mapping two-dimensional vectors onto sinusoidal patterns of activity across distinct neuronal populations, with the amplitude of the sinusoid representing the length of the vector and its phase representing the angle of the vector. The principles of this circuit may generalize to other brains and to domains beyond navigation where vector operations or reference-frame transformations are required (Lyu 2022).
Whether mammalian brains have neurons that are tuned to the allocentric travelling direction of an animal as in Drosophila is still unknown. Although a defined population of neurons tuned to travelling direction has yet to be highlighted in mammals, such cells could have been missed because their activity would loosely resemble that of the head-direction cells outside a task in which the animal is required to sidestep or walk backwards (Lyu 2022).
Neurons are often modelled as summing their synaptic inputs, but the heading inputs that PFN cells receive from the EPG system appear to be multiplied by the self-motion (for example, optic flow) input, resulting in an amplitude or gain modulation. Multiplicative or gain-modulated responses appear in classic computational models for how neurons in area 7a of the primate parietal cortex might implement a coordinate transformation, alongside similar proposals in mammalian navigation. The Drosophila circuit described in this study strongly resembles aspects of the classic models of the parietal cortex. Units that multiply their inputs are also at the core of the 'attention' mechanism used, for example, in machine-based language processing. The experimental evidence for input multiplication in a biological network may indicate that real neural circuits have greater potential for computation than is generally appreciated (Lyu 2022).
This study describes a travelling direction signal and how it is built; related results and conclusions appear in a parallel study. The mechanisms described for calculating the travelling direction are robust to left-right rotations of the head and to the possibility of the allocentric projection vectors being non-orthogonal. It is possible that the travelling signal of hΔB cells is compared with a goal-travelling direction to drive turns that keep a fly along a desired trajectory. Augmented with an appropriate speed signal (or if the fly generally travels forward relative to its body), the hΔB signal could also be integrated over time to form a spatial-vector memory via path integration. There are hundreds more PFN cells beyond the 40 PFNd and 20 PFNv cells examined in this study, and thus the central complex could readily convert other angular variables from egocentric to allocentric coordinates via the algorithm described in this study. Because many sensory, motor and cognitive processes can be formalized in the language of linear algebra and vector spaces, defining a neuronal circuit for vector computation may open the door to better understanding of several previously enigmatic circuits and neuronal activity patterns across multiple nervous systems (Lyu 2022).
Complex behaviors depend on the precise developmental specification of neuronal circuits, but the relationship between genetic programs for neural development, circuit structure, and behavioral output is often unclear. The central complex (CX) is a conserved sensory-motor integration center in insects that governs many higher order behaviors and largely derives from a small number of Type II neural stem cells. This study shows that Imp, a conserved IGF-II mRNA-binding protein expressed in Type II neural stem cells, specifies components of CX olfactory navigation circuitry. This study shows: (1) that multiple components of olfactory navigation circuitry arise from Type II neural stem cells and manipulating Imp expression in Type II neural stem cells alters the number and morphology of many of these circuit elements, with the most potent effects on neurons targeting the ventral layers of the fan-shaped body. (2) Imp regulates the specification of Tachykinin expressing ventral fan-shaped body input neurons. (3) Imp in Type II neural stem cells alters the morphology of the CX neuropil structures. (4) Loss of Imp in Type II neural stem cells abolishes upwind orientation to attractive odor while leaving locomotion and odor-evoked regulation of movement intact. Taken together, this work establishes that a single temporally expressed gene can regulate the expression of a complex behavior through the developmental specification of multiple circuit components and provides a first step towards a developmental dissection of the CX and its roles in behavior (Hamid, 2023).
How the brain perceives sensory information and generates meaningful behavior depends critically on its underlying circuitry. The protocerebral bridge (PB) is a major part of the insect central complex (CX), a premotor center that may be analogous to the human basal ganglia. By deconstructing hundreds of PB single neurons and reconstructing them into a common three-dimensional framework, this study has constructed a comprehensive map of PB circuits with labeled polarity and predicted directions of information flow. The analysis reveals a highly ordered information processing system that involves directed information flow among CX subunits through 194 distinct PB neuron types. Circuitry properties such as mirroring, convergence, divergence, tiling, reverberation, and parallel signal propagation were observed; their functional and evolutional significance is discussed. This layout of PB neuronal circuitry may provide guidelines for further investigations on transformation of sensory (e.g., visual) input into locomotor commands in fly brains (Lin, 2014: PubMed).
Ring attractors are a class of recurrent networks hypothesized to underlie the representation of heading direction. Such network structures, schematized as a ring of neurons whose connectivity depends on their heading preferences, can sustain a bump-like activity pattern whose location can be updated by continuous shifts along either turn direction. A population of fly neurons in the ellipsoid body has been shown to represent the animal's heading via bump-like activity dynamics (see Bump attractors and spontaneous pattern formation). This study combined two-photon calcium imaging in head-fixed flying flies with optogenetics to overwrite the existing population representation with an artificial one, which was then maintained by the circuit with naturalistic dynamics. A network with local excitation and global inhibition enforces this unique and persistent heading representation. Ring attractor networks have long been invoked in theoretical work; this study provides physiological evidence of their existence and functional architecture (Kim 2017).
Studies of neural circuits near the sensory periphery have produced deep mechanistic insights into circuit functions. However, it has been more challenging to understand circuit functions in central brain regions dominated by recurrent networks, which often produce complex neural activity patterns. These dynamics play a major role in shaping cognitive functions, such as the maintenance of heading information during navigation. A heading representation must be unique (because an animal can face only one direction at a given time) and persistent (to allow an animal to keep its bearings in darkness), yet must allow updating that matches the magnitude and speed of heading changes expected from the animal's movements. Theoretically, this can be accomplished by ring attractor networks (see, for example, Balanced neural architecture and the idling brain), wherein the position of a localized subset of active neurons in a topological ring represents the animal's heading direction. However, whether the brain uses these hypothesized networks is still unknown. A recent study reported that a population of neurons, called E-PG neurons [to signify their predominantly spiny (and, thus, putatively post-synaptic) projections within the ellipsoid body ('E-') and their predominantly bouton-like projections within the protocerebral bridge ('-P') and the gall ('G')], in the Drosophila melanogaster ellipsoid body (EB) appears to use bump-like neural activity dynamics to represent the animal's heading in visual environments and in darkness. This study establishes essential properties of the network that enables this representation (Kim 2017).
Whether the E-PG population activity bump tracks the fly's heading direction relative to its visual surroundings during tethered flight was determined first. Two-photon imaging with the genetically encoded calcium indicator GCaMP6f was performed to record dendritic calcium activity of the entire E-PG population in the EB while the fly was flying in a virtual-reality LED arena. The azimuthal velocity of the visual scene was proportional to the fly's yaw velocity. As with walking flies, E-PG population activity during flight was organized into a single bump, whether the visual scene contained a single bar or a more complex pattern. The activity bump closely tracked the fly's heading in flight and persisted in darkness. However, unlike in walking, the activity bump seldom tracked the fly's motor actions in darkness, potentially because tethering deprives the fly of normal sensory feedback about its rotational movements from its halteres. Although the location of the activity bump eventually drifted in some flies, the bump's movement was, on average, uncorrelated to the animal's turning movements in darkness. These findings suggest that the representation of heading in the E-PG population has intact, visually driven dynamics as well as persistence, but is largely uncoupled from updating by self-motion cues during tethered flight (Kim 2017).
To test whether the fly's compass network enforces a unique bump within the EB, advantage was taken of the relative persistence of the visually evoked activity bump in darkness, and asked whether this bump could coexist with an 'artificial' bump of activity. Localized optogenetic stimulation was used to create artificial activity bumps in different locations within the E-PG population. Using a transgenic fly line in which E-PG neurons coexpressed CsChrimsonand GCaMP6f, alternating two-photon laser scan lines of excitation (higher laser intensity) and imaging (normal laser intensity) were used to monitor changes in E-PG population dynamics in response to an optogenetically created spot of local activity. By varying the intensity of stimulation light delivered to the target location, bumps were created of increased calcium activity. As the new bump formed, activity at the previous location began to decline and eventually disappeared without significantly perturbing the fly's behavior. When the optogenetic excitation was terminated, the amplitude of the artificially created bump settled at levels typically evoked by sensory stimuli and did not disappear; it either stayed in the induced location for several seconds or slowly drifted away (Kim 2017).
The bump's uniqueness may arise through either recurrent mutual suppression or an indirect mechanism whereby strong bump activity in the EB functionally inhibits feedforward sensory inputs to other E-PG neurons. To discriminate between these alternatives, two locations on the EB ring were simultaneously excited. A reference location was excited at a fixed laser power, and a second, spatially offset location was excited at increasing levels of laser power. The reference bump could always be suppress by increasing laser power at the second location above a certain threshold, consistent with mutual suppression (Kim 2017).
Recurrent suppression can ensure a unique activity bump through a simple winner-take-all (WTA) circuit. However, an animal's representation of its angular orientation should favor more continuous updates based on turning actions. Such gradual, ordered drift to nearby locations would be more consistent with continuous, or ring, attractor models. This study therefore examined changes in the location of an artificially created bump after the stabilization of its peak activity at the 'natural' level. The experiments were performed in darkness to untether the bump from any potentially lingering visual input. If EB dynamics were driven by a WTA network, bumps would be expected to disappear at times and to jump to random distant locations. In contrast, the bump drifted gradually around the EB; this finding suggests that the fly's heading representation is updated through functionally excitatory interactions between neighboring E-PG neurons, consistent with a ring attractor model. These observations together rule out the possibility that network dynamics in darkness result purely from cell-intrinsic mechanisms or slowly decaying visual input. Most important, direct manipulation of E-PG neuron activity changed the network state, which implies that E-PG neurons do not merely mirror dynamics occurring in a different circuit, but are themselves an important component of the ring attractor (Kim 2017).
The next area of focus was the effective connectivity pattern underlying ring attractor dynamics in the E-PG population. A wide range of network structures can, in principle, implement ring attractors. This study focused efforts to a model space between two extreme network architectures that are analytically solvable: (1) a 'global model' based on global cosine-shaped interactions and (ii) a 'local model' based on relatively local excitatory interactions. Under constraints of a fixed bump width of 90° to match physiological observations and an assumption of effectively excitatory visual input without any negative bias, both models could explain the basic properties of bump dynamics, including its uniqueness and its persistence in darkness. The network's response to more artificial conditions, such as abrupt visual stimulus shifts, was therefore probed (Kim 2017).
How the E-PG population responded to unnatural, abrupt visual shifts was examined experimentally first. Depending on the distance of the shift, the E-PG bump either 'flowed' continuously (shorter shift distances) or 'jumped' to the new location (longer shift distances). In simulations, both models predicted a mixture of jump and flow responses, depending on the strength and width of the abruptly shifting visual input. For example, weak wide input induced flows and strong narrow input evoked jumps. However, the jump-flow balance predicted by the two models differed and was more consistent with the local model in several aspects. First, the visual input strength inferred from normal conditions was much weaker than required by the global model for bump jumps. Second, the global model required a much-wider-than-normal range of visual input strengths to explain jumps at multiple distances. Third, using parameters consistent with the rest of the findings, it was possible to reproduce the jump-flow ratio that was observed with the local model but not with the global model (Kim 2017).
To obtain more concrete evidence, model predictions were compared to experimentally observed bump dynamics, under conditions in which input strength, polarity, and shift distance were controlled through optogenetic stimulation. To simulate moderate and large input shift distances, two small regions were sequentially stimulated in the EB-each with an angular width of 22.5°-separated by either 90° or 180°. The stimulation laser power was varied to detect the threshold required for the bump to jump. The laser power required to elicit a jump was not significantly different between the two different shift distances, favoring the local model. The strength of input to the network was then inferred by comparing the amplitude of the optogenetically evoked bump to natural bump amplitudes in darkness. The optogenetic input strength required to induce jumps was smaller than the global model's prediction but matched that of the local model and the range of the inferred visual input strength under normal conditions. Finally, intermediate models that lie between the extremes of the local and global models were then test; any model that exhibited the observed jumps in response to a weak 22.5°-wide input had narrow connectivity profiles was then found. All these observations were once again consistent with the local model (Kim 2017).
In mammals, heading representations are thought to be distributed across multiple neural populations and multiple brain areas. In Drosophila as well, the compass system likely involves multiple cell types, including neurons in the protocerebral bridge (PB). Further, occasional changes observed in the dynamics suggest network modulation by other factors not yet known. For example, sometimes sudden changes were observed in E-PG dynamics, as when the amplitude of the sensory-evoked activity bump changed depending on whether or not the tethered fly was flying and, occasionally, during flight. Nonetheless, the E-PG population provides a powerful physiological handle on the internal representation of heading: a single activity bump moving through topographically arranged neurons. The experimental approach this enabled provides one avenue for investigating which of multiple populations are key circuit components of a computation and which simply read out the results of that computation. It was found that the artificial bump created by directly manipulating E-PG population activity displays natural dynamics, which indicates that these neurons are a key component of the heading circuit (Kim 2017).
The finding that the uniqueness of the E-PG activity bump is ensured via global competition strengthens the conclusion that this population encodes an abstract internal representation of the fly's heading direction. Such abstract representations permit an animal to untether its actions from the grasp of its immediate sensory environment and thereby confer flexibility in both time and behavioral use. Combining an analysis of artificially induced bump dynamics with theoretical modeling allowed interrogation of this recurrent circuit architecture. It was found that the effective network connectivity profile was consistent with ring attractor models characterized by narrow local excitation and flat long-range inhibition. This neural circuit motif of local excitation and long-range inhibition is ubiquitous across many brain areas and across animal taxa. Such observations support the idea that common circuit motifs might be evolutionarily adapted to serve as crucial building blocks of cognitive function (Kim 2017).
Neural representations of head direction (HD) have been discovered in many species. Theoretical work has proposed that the dynamics associated with these representations are generated, maintained, and updated by recurrent network structures called ring attractors. This theorized structure-function relationship was evaluated by performing electron-microscopy-based circuit reconstruction and RNA profiling of identified cell types in the HD system of Drosophila melanogaster. Motifs were identified that have been hypothesized to maintain the HD representation in darkness, update it when the animal turns, and tether it to visual cues. Functional studies provided support for the proposed roles of individual excitatory or inhibitory circuit elements in shaping activity. This study also discovered recurrent connections between neuronal arbors with mixed pre- and postsynaptic specializations. These results confirm that the Drosophila HD network contains the core components of a ring attractor while also revealing unpredicted structural features that might enhance the network's computational power (Turner-Evans, 2020).
Mammalian head direction (HD) cells provide one of the clearest examples of an internal representation of an animal's relationship to its surroundings. The HD representation uses visual cues in the environment as a reference but persists in darkness, where it is updated by self-motion cues. This internal representation likely guides navigation behaviors. Indeed, perturbations to the system in rats induce errors in path integration. Theoretical studies have proposed that the neuronal population dynamics associated with HD representations are maintained by network structures called ring attractors. These recurrent networks are often schematized as a ring of neurons whose connectivity depends on their directional tuning preferences. In most models, neurons with similar directional tuning excite each other and those with different tuning inhibit each other, thereby enabling the generation of a stable pattern of localized activity in any part of the network. Recurrent loops with a shift move the activity 'bump' around the ring as the animal turns, and the compass-like representation uses visual inputs as a reference. Experimental support for this general theoretical formulation has come from analyses of HD cell population activity under a variety of different conditions. However, different network implementations of this general formulation make distinct assumptions about the connectivity of their constituent neurons. Importantly, such assumptions, which dictate exactly how the circuit functions, have been difficult to test in the large mammalian brain. Mammalian HD cells are distributed across many brain regions and are as yet not well classified into subtypes, making it challenging to identify and target them reliably (Turner-Evans, 2020).
An internal representation similar to the mammalian HD representation has also been discovered in the insect brain (Seelig, 2015; Schematic of fly central brain and CX: ellipsoid body (EB), fan-shaped body (FB), protocerebral bridge (PB), paired noduli (NO), lateral accessory lobe (LAL) and gall (Gall). MB: mushroom body). Further, there is strong evidence that this HD representation is implemented by a ring attractor. A ring attractor model that assumes recurrent connectivity between heading neurons and neurons that encode both angular velocity and heading maintains an accurate heading representation when driven by realistic velocity inputs. This model replicates the fly HD network's dynamics in darkness. Other models have invoked plasticity between visual inputs and heading neurons to show how visual and angular velocity information might update the representation in a mutually consistent manner (Turner-Evans, 2020).
Importantly, however, all fly ring attractor models have assumed the circuit's connectivity based on relatively indirect evidence. For example, the location of pre- and postsynaptic arbors has been inferred from whether neural processes visible in light-microscopic images seem spiny or bouton-like in specific substructures. The hypothesized connectivity of the circuit has then been derived from light-level overlap between the putatively pre- and postsynaptic arbors of neurons, in some cases further supported by GFP reconstitution across synaptic partners (GRASP) and trans-Tango experiments, although the reliability and accuracy of these methods to estimate pairwise connectivity is known to be limited. Similarly, measurements of functional connectivity by optogenetic stimulation of a population of one type of neurons and calcium imaging of another can be difficult to interpret within recurrent networks (Turner-Evans, 2020 and references therein).
In the current study, reconstructions were used based on serial transmission electron microscopy (EM). to determine synaptic connectivity within the neural network underlying the fly's HD representation. Neural connectivity matrices were compared to those extracted from the recently released fly hemibrain connectome, which was obtained by using focused ion beam scanning electron microscopy (FIBSEM). Cell-type-specific RNA sequencing (RNA-seq) and fluorescence in situ hybridization (FISH) allowed characterization of the expression profiles of the key cellular components of the ring attractor network. This integrated information was used to assess the role of each of the constituent cell types in the ring attractor's dynamics. It was found that the fly HD network contains motifs similar to those proposed in theoretical models. These motifs were hypothesized to maintain HD activity and update it both in the dark and when visual features are present. These ideas were tested using targeted two-photon calcium imaging and thermogenetic perturbations of the constituent neuron types in behaving, head-fixed Drosophila. It was also found that many neurons have mixed pre- and postsynaptic specializations within their innervations to single brain structures, creating 'hyper-local' recurrent loops that may allow local computations to supplement the role of recurrence at the network level. Moreover, although many ring attractor models rely on distinct units that provide local excitation and long-range inhibition to shape activity into one stable bump, consistent with the biological results, apparent redundancy was found in these structural elements. Taken together, these results provide new structural and functional insights into how a small biological ring attractor network allows an animal to maintain an accurate internal sense of direction (Turner-Evans, 2020).
Efforts to model the dynamics of HD networks have long focused on ring attractors. Although this conceptual framework has been very influential, testing the validity of the structural assumptions and functional predictions made by ring attractor models in biological circuits has been challenging. This study has bridged this gap by characterizing the neurons that compose the fly HD circuit and examining how they together produce ring attractor dynamics. A previous study proposed a ring attractor model that captured the observed dynamics of the fly HD network (Turner-Evans, 2017). The weights of this model were tuned without any knowledge of the actual synaptic connections. If the synapse counts measured in this work are extrapolated to the entire biological network, the theoretical and the experimentally derived connectivity matrices are remarkably similar (Turner-Evans, 2020).
Some of these results will require follow-up experimental and theoretical work. The long-range inhibition that has been invoked to form a stable bump of activity appears to be split across multiple brain regions and multiple classes of neurons. In addition to the Δ7s, GABAergic R4d (and other) visual ring neurons also inhibit much of the ellipsoid body-protocerebral bridge-gall neuron (E-PG) population in the presence of visual stimuli. Further, the gall-EB ring neuron population provides a source of inhibition onto the E-PGs. Indeed, the cumulative inhibition from multiple classes of ring neurons may account for some of the mutual suppression between E-PGs (Turner-Evans, 2020).
Local excitation also appears to be implemented through multiple classes of neurons. The redundancy of local excitation and long-range inhibition may allow the network to maintain HD activity in the absence of external input. These network features may also provide a means to stabilize the heading representation in the presence of noisy inputs and inhomogeneous synaptic weights, which can disrupt the function of continuous attractor networks (Turner-Evans, 2020).
Synapses between visual ring neurons were also observed, consistent with previous observations from EM in locusts and trans-Tango experiments in flies. Visual scenes are often complex and dynamic, and the observed inter-ring neuron connections may either form a winner-take-all network that leads one visual feature to dominate, preventing the bump from moving erratically as the scene shifts over time, or provide a mechanism for gain control that normalizes the total level of inhibition from those ring neurons onto the E-PGs (Turner-Evans, 2020).
At a methodological level, the recently completed FIBSEM-based connectome reinforces the conclusions from FAFB. Although there are sometimes more connections detected between specific types in one or the other dataset, only rarely are connections in one dataset not seen in the other (Turner-Evans, 2020).
Finally, the results highlight the challenges of linking structure to function. Connectomics is often perceived as providing strong constraints on models of circuit function. However, although this was partly true for the fly HD network, the results also identified unexpected computational mechanisms that the circuit may employ. For example, the profusion of mixed pre- and postsynaptic specializations within single compartments of individual neurons creates many more locally recurrent loops between similar and different neuron classes, and a functional account for such dense recurrence cannot yet be provide. Future theoretical and experimental work to probe their function may well reveal that fly brains are more powerful than their numerical simplicity might suggest (Turner-Evans, 2020).
Relative motion breaks a camouflaged target from a same-textured background, thus eliciting discrimination of a motion-defined object. Ring (R) neurons are critical components in the Drosophila central complex, which has been implicated in multiple visually guided behaviors. Using two-photon calcium imaging with female flies, this study demonstrated that a specific population of R neurons that innervate the superior domain of bulb neuropil, termed superior R neurons, encoded a motion-defined bar with high spatial frequency contents. Upstream superior tuberculo-bulbar (TuBu) neurons transmitted visual signals by releasing acetylcholine within synapses connected with superior R neurons. Blocking TuBu or R neurons impaired tracking performance of the bar, which reveals their importance in motion-defined feature encoding. Additionally, the presentation of a low spatial frequency luminance-defined bar evoked consistent excitation in R neurons of the superior bulb, whereas either excited or inhibited responses were evoked in the inferior bulb. The distinct properties of the responses to the two bar stimuli indicate there is a functional division between the bulb subdomains. Moreover, physiological and behavioral tests with restricted lines suggest that R4d neurons play a vital role in tracking motion-defined bars. It is concluded that the central complex receives the motion-defined features via a visual pathway from superior TuBu to R neurons and might encode different visual features via distinct response patterns at the population level, thereby driving visually guided behaviors (Duan, 2023).
Many animals navigate using a combination of visual landmarks and path integration. In mammalian brains, head direction cells integrate these two streams of information by representing an animal's heading relative to landmarks, yet maintaining their directional tuning in darkness based on self-motion cues. This study used two-photon calcium imaging in head-fixed Drosophila melanogaster walking on a ball in a virtual reality arena to demonstrate that landmark-based orientation and angular path integration are combined in the population responses of neurons whose dendrites tile the ellipsoid body, a toroidal structure in the centre of the fly brain. The neural population encodes the fly's azimuth relative to its environment, tracking visual landmarks when available and relying on self-motion cues in darkness. When both visual and self-motion cues are absent, a representation of the animal's orientation is maintained in this network through persistent activity, a potential substrate for short-term memory. Several features of the population dynamics of these neurons and their circular anatomical arrangement are suggestive of ring attractors, network structures that have been proposed to support the function of navigational brain circuits (Seelig, 2015).
Visual landmarks can provide animals with a reliable indicator of their whereabouts. In the absence of such cues, many animals track their position relative to a reference point by continuously monitoring their own motion, a process called path integration. Estimates of position based purely on self-motion cues, however, can accumulate error over time. Successful navigation then, requires animals to flexibly combine these distinct sources of information. In mammalian brains this process of integration is evident in head direction cells, which are neurons sensitive to an animal's heading relative to visual cues in its surroundings that maintain their representation of heading in total darkness using self-motion cues. With their smaller brains and identifiable neurons, insects offer tractable systems to examine the integrative neural computations underlying navigation. Indeed, many insects (for example, desert ants and honeybees) are known to navigate using landmarks and path integration. Experiments in a variety of insects indicate the involvement of the central complex (CX)-a brain region conserved across insects-in such behaviour. In the fruitfly, behavioural genetics experiments have suggested that the CX is required for several components of navigation, including memory for visual landmarks, patterns and places, and directional motor control. Electrophysiological recordings in immobilized locusts and butterflies have revealed a map-like representation for the orientation of electric field vectors of polarized light, which may enable sun-compass navigation. Extracellular recordings from CX neurons in tethered walking cockroaches have shown encodings of turning direction and of wide-field optic flow, a potential cue for self-motion. However, previous studies of visual responses in the CX were conducted under conditions in which insects passively viewed visual stimuli. This study sought to uncover integrative neural processes relevant to navigation in the CX by allowing a tethered fly to control and respond to visual stimuli while simultaneously recording its neural activity and behaviour (Seelig, 2015).
Two-photon imaging was used with the genetically encoded calcium indicator GCaMP6f to monitor neural responses in the CX while a head-fixed fly walked on an air-supported ball within a light-emitting-diode (LED) arena. In previous experiments, a subset of neurons was identified with projections to the CX, and specifically to rings of the ellipsoid body (EB), that show strong tuning to localized visual features including vertical stripes, a class of stimuli that also induce innate fixational responses in flies. To probe how such visual information might be used within the CX this study focused on a class of columnar neurons of the CX, each of which sends dendrites to a specific wedge of the EB. These neurons are termed EBw.s neurons. The dendritic responses of the entire EBw.s population were monitored in the EB during walking, both under closed-loop virtual reality conditions in which the rotation of visual patterns was driven by the fly's turning movement on the ball and in darkness (Seelig, 2015).
This network was found to use information from both landmark-based and angular path integration systems to create a compass-like representation of the animal's orientation in the environment. Previous studies have described static visual maps in the CX. Such maps may allow navigating insects to maintain a sun-compass-based heading direction. This study found that EBw.s neurons track the fly's orientation relative to visual landmarks in a variety of different visual environments, suggesting that the CX dynamically adapts to estimate the fly's orientation within its visual surroundings. Subsets of ring neurons are likely to bring information about spatially localized visual features to specific rings of the ellipsoid body. It is not yet clear how this information is converted into an abstract and flexible representation of the animal's orientation relative to landmarks, but EBw.s responses in a symmetric environment with two indistinguishable cues hint at an underlying winner-take-all process for landmark selection. Combining landmark orientation with information about the animal's movement effectively creates an internal reference frame for the animal in its surroundings. Many of the proposed functions of the CX in directed locomotion, visual place learning, and action-selection, may rely on this internal reference. Although the EBw.s population tracks the fly's rotational movements in darkness, it is not yet known where and how translational motion, an important component of a complete navigational system, is incorporated. Additionally, although the calcium sensor that was chosen for imaging experiments has the temporal resolution necessary to capture EBw.s representations of the fly's angular rotation, it lacks the precision necessary for determination of whether EBw.s activity represents the fly's predicted future orientation or its estimate of current orientation (Seelig, 2015).
The observation that EBw.s activity was maintained in the absence of self-motion suggests that internal dynamics play a significant role in shaping neural activity in the fly brain, much as they do in the brains of larger animals. Persistent activity in the CX can maintain compass information when the fly is standing in darkness for 30 s - two orders of magnitude longer than might be explained by calcium sensor decay kinetics. Persistent activity has been shown to support maintenance of eye position in the goldfish and has been proposed to underlie working memory in mammals. In the CX, this activity may allow the fly to retain a short-term orientation memory even when landmarks are temporarily out of sight. Consistent with this notion, the EBw.s activity bump largely remained tethered to the position of one landmark even in the presence of another identical landmark in front of the fly. The bump also did not always shift instantaneously following an abrupt displacement of visual landmarks, as if temporarily retaining the original orientation reference before locking on to its new position (Seelig, 2015).
Several models have been proposed to explain how visual landmark and self-motion cues are integrated at the level of head direction cell activity in mammals. Most rely on circuits organized as ring attractors: neurons are schematized as being arranged in a circle based on their preferred directions, with connection strengths that depend on their angular separation. With initial sensory input and an appropriate balance of recurrent excitation and inhibition, such a circuit can generate and sustain a localized activity bump. The bump's position on the circle corresponds to the animal's heading which is then updated by directional drive from self-motion signalling neurons. Direct experimental evidence in support of these models has been difficult to obtain in mammals owing to the distributed nature of the underlying circuits. Although the functional connectivity between EBw.s neurons is not yet known, several of the expected features of ring attractor models were observed in the dynamics of this population of CX neurons: organization of activity into a localized bump, movement of the bump to neighbouring wedges based on self-motion, drift in bump location in darkness, persistent activity, and both abrupt jumps and gradual transitions of the activity bump when triggered by strong visual input. Cell-intrinsic mechanisms could also underlie some of these features, including, for example, persistent activity. The genetic tools available in Drosophila to target and manipulate the activity of identified cell types should allow different models for visually guided orientation and angular path integration to be discriminated at the level of synaptic, cellular and network mechanism (Seelig, 2015).
Recent studies of the Central Complex in the brain of the fruit fly have identified neurons with activity that tracks the animal's heading direction. These neurons are part of a neuronal circuit with dynamics resembling those of a ring attractor. The homologous circuit in other insects has similar topographic structure but with significant structural and connectivity differences. This study modeled the connectivity patterns of two insect species to investigate the effect of these differences on the dynamics of the circuit. The circuit found in locusts can also operate as a ring attractor but differences in the inhibition pattern enable the fruit fly circuit to respond faster to heading changes while additional recurrent connections render the locust circuit more tolerant to noise. These findings demonstrate that subtle differences in neuronal projection patterns can have a significant effect on circuit performance and illustrate the need for a comparative approach in neuroscience (Pisokas, 2020).
The availability of tools for the study of insect brains at the single neuron level has opened the way to deciphering the neuronal organisation and principles of the underlying circuit's behaviour. However, even where there is progress towards a complete connectome, the lack of data on synaptic strengths, neurotransmitter identity, neuronal conductances, etc. leave many parameters of the circuit unspecified. Exploring these parameters via computational modelling can help to illuminate the functional significance of identified neural elements. This study has applied this approach to gain greater insight into the nature of the heading encoding circuit in the insect central complex (CX), including the consequences of differences in circuit connectivity across two insect species (Pisokas, 2020).
This study has focused on a subset of neurons in the PB and EB which have been hypothesised to operate as a ring attractor, with a 'bump' of neuronal activity moving across columns consistently with the changing heading direction of the animal. The neuronal projection patterns and columnar organisation differ between the two insect species that were analysed, the fruit fly and the locust. There are additional morphological columns in the PB of flies (9 vs. 8), resulting in a different number of functional units that could influence the symmetry of the underlying neural circuits. Also, the EB in the fruit fly forms a physical ring, while the homologous region in the locust is an open structure. This analysis of the connectivity as a directed graph has revealed, surprisingly, that the circuits are nevertheless equivalent in their effective structure, forming a closed ring attractor in both species with an identical functional role for each neuron class. The preservation of this circuit across 400 million years of evolutionary divergence suggests that it is an essential, potentially fundamental, part of the insect brain (Pisokas, 2020).
It is worth noting that an essential part of the circuit, namely the functionally closed ring that was found in both species, is achieved with two different solutions. In the fruit fly, the torus-shaped EB provides an anatomical solution to the closure of the ring via overlapping projections from E-PG neurons innervating the innermost and outermost PB glomeruli. In contrast, in the locust the midline spanning output fibers of the E-PG neurons in the medial protocerebral bridge (PB) glomeruli serve this function in combination with a slightly different projection pattern that results in the P-EN (protocerebral bridge-ellipsoid body-noduli neurons) forming reciprocal connections back to E-PG neurons in the same octant. In this context, it is interesting to note that neither solution to this problem is possible for insects of a different order, the lepidoptera (moths and butterflies). These insects have an almost straight EB, their PB is split along the midline, and right-left connections between the two halves are realised by a neuropil-free fiber bundle. Neither midline crossing E-PG fibers within the PB, nor local connections around the ring of the EB are therefore morphologically possible, suggesting that a functional closure of the heading direction circuit is either not required or achieved via other means in these species. The notion that there are many solutions to the same problem is further highlighted by data from bumblebees showing the existence of a ninth E-PG neuron that connects the medialmost PB glomerulus to the outermost ipsilateral EB wedge, closing the ring in yet another way. Exploring these different solutions across many species could provide key insights into the evolution of this circuit under a multitude of evolutionary history constraints (Pisokas, 2020).
In combination, these findings underline that the large-scale anatomical differences at the level of neuropils and projection patterns do not necessarily affect the core functions of the circuit. Rather, the functional constraints appear significant enough that even in those parts of the circuit that clearly differ between species convergent solutions have evolved that solve similar problems, albeit in slightly different ways
Surprisingly, more subtle differences in the morphology between the two species have significant effects on the dynamical response of the heading direction circuit. First, the shape of the dendritic arborizations of one type of CX neuron determines how quickly the model circuit tracks rotational movements. Second, a difference in the overlap of neuronal projections in the EB results in an extra feedback loop between the P-EN and E-PG neurons in the locust circuit that makes it more robust to synaptic noise (Pisokas, 2020).
It is suggested that the effects of these differences are consistent with the behavioural ecology of the two species. On the one hand, the faster response of the ring attractor circuit in the fruit fly accommodates the fast body saccades that fruit flies are known to perform. On the other hand, the locust is a migratory species, so its behaviour demands maintenance of a defined heading for a long period of time. This requirement for heading stability might have provided the selective pressure needed to drive the evolution of a more noise resilient head direction circuit (Pisokas, 2020).
As any model, the circuits are necessarily condensed and simplified versions of the real circuits in the insect brain. In comparison to previous models, the work presented in this study has been more precisely constrained by the latest anatomical evidence. The models were additionally constrained to use plausible values for the biophysical properties of neurons (membrane conductance and capacitance) as well as spiking rates (background activity) supported by electrophysiological evidence. Furthermore, in building these models it was not assumed that the underlying circuits must be ring attractors, but rather whether, given the available connectivity data, they can be was asked and investigated. This was especially the case for the locust model since this work represents the first model of this circuit to date. Nevertheless, it is important to outline those areas where the assumptions cannot be fully justified from the existing data and to identify the potential consequences for the modelling results (Pisokas, 2020).
In the model of the fruit fly heading tracking circuit, a uniform distribution was assumed of dendrites across the Delta7 neurons. Imaging of these neurons suggests that there might be a subtle variation of the dendritic density along their length. However, it is unclear how this subtle variation might be related to synaptic density and efficacy. Therefore, the simplifying assumption was initially made that these neurons have uniform synaptic efficacy across the PB. However, this study also explored the effect of varying the degree of synaptic uniformity, showing that there is a range of synaptic efficacy distributions that still can produce the fly-like rapidity in the circuit response (Pisokas, 2020).
In general, arborization trees of neurons in the CX can be very complex, as they are not only confined to specific slices, but also to one or several layers, especially within the EB. In Drosophila, the spiny terminal arbors of E-PG neurons extend to the width of single wedges in the EB, occupying both the posterior and medial layers. In contrast, P-EG and P-EN neurons arborize in tiles, hence innervating only the posterior surface volume of the EB. Therefore, it is assumed that presynaptic terminals of P-EG and P-EN neurons form synapses with E-PG postsynaptic terminals in the posterior layer of the EB. In locusts, the E-PG arborizations are more complex, as these cells innervate a single wedge of the anterior and medial EB layers, but extend at least twice this width to either side in the posterior layer that provides overlap with the P-EN neurons. Additionally, the wider fibres have a different morphological appearance. P-EG neurons in this species innervate all layers evenly. Although these detailed differences likely have consequences for connectivity, these arborizations were simplified to their most essential components, aiding the extraction of the core features. With the advance of comparative connectomics, these aspects will become accessible for investigation (Pisokas, 2020).
Several assumptions were made while deriving the neuronal connectivity in these models. Well delineated borders of synaptic domains were assumed, which is clearly not always the case. Especially in the EB, some overlapping of neighbouring synaptic domains due to stray terminals is to be expected. The circumferential extent of arbors in wedges and tiles may affect the integrity of the resulting circuit and its properties. However, due to lack of adequate data about the extent of such overlap it is not possible to currently model this aspect in a sensible way (Pisokas, 2020).
Furthermore, neuronal connectivity was mostly inferred by co-location of neuronal arbors, that is, projection patterns. A functional connectivity study has reported that stimulation of E-PG neurons triggered significant responses to Delta7 neurons but no columnar neurons. However, as that study noted, the lack of response might be due to the limitations of the method used. Alternatively, such connections might be mediated by interneurons instead of being monosynaptic. Future work using electron microscopy data will elucidate which of the overlapping arborizations correspond to functional connections and allow augmentation of these models (Pisokas, 2020).
Further assumptions were made about neuronal polarity, type of synapses and synaptic efficacies. One study hsd characterise the EB arbor of E-PG neurons in Drosophila as having both presynaptic and postsynaptic domains; however, another study reports that using anti-synaptotagamin is inconclusive for presynaptic terminals. In the models presented in this studyfor both the fruit fly and the locust it was thus assumed that E-PG neurons are purely postsynaptic in the EB, following the most parsimonious polarity estimate. Connectomics data from a recent preprint demonstrate that in Drosophila synapses exist that directly link Delta7 to E-PG neurons in the PB (Turner-Evans, 2019). These synapses are most likely inhibitory and would thus inhibit the distal portion of the ring and thus would not alter the location of the activity 'bump'. For simplicity and because they do not affect the functional layout of the circuit, these synapses were not included in the model (Pisokas, 2020).
Furthermore, the Delta7 neurons are assumed to have inhibitory effect on their postsynaptic neurons, as a previous study proposed. However, there is evidence that Delta7 neurons make both inhibitory and excitatory synapses to other neurons. Indeed, these cells were recently shown to be glutamatergic, enabling both inhibitory and excitatory effects on postsynaptic cells via different glutamate receptors (Turner-Evans, 2019). As the downstream neurons with demonstrated excitatory responses (P-FN neurons) are not part of the current model, the simplifying assumption was made that Delta7 neurons have exclusively inhibitory effect on their postsynaptic neurons, both in flies and locusts. It is also possible that there are other sources of inhibition in the circuit, for example mediated by the GABAergic ring neurons originating in the bulbs, as suggested by one study, or via GABAergic Gall-EB ring neurons (Turner-Evans, 2019). This study did not explore this possibility (Pisokas, 2020).
It was additionally assumed that the synaptic strengths of all synapses of each class are identical. This might not be the case in the actual animals, especially considering that one of the EB tiles (T1) is innervated by twice as many neurons as other tiles in fruit flies (see Projection patterns of the excitatory portion of the fruit fly circuit). Neurons innervating this tile might have reduced synaptic efficacy in order to maintain the radial symmetry of the circuit intact. Similarly, the synaptic strengths of the neurons closing the ring in locusts would be expected to be different than those of other synapses if the ring does not have a functional 'seam'. Such a synaptic efficacy variation is suggested by the fact that the arborization density of E-PG neurons innervating the two medial PB glomeruli (G9 and G1) is not the same in both of them. There is certainly space for further exploration of the effect of synaptic efficacy in those segments of the ring in both species. Finally, synaptic strength variation might exist for the two Delta7 neurons that have presynaptic terminals in three glomeruli instead of two (Pisokas, 2020).
All types of neurons in these models were assumed to have the same nominal biophysical properties even though anatomical evidence has shown that their morphology, somata size and main neurite thickness differ. To relax this assumption, the effect of heterogeneity in the biophysical properties of the neuronal population was explored. The conclusions were corroborated using both rate-based and Leaky Integrate and Fire neurons with refractory period. This allowed highlighting of the significance of the neuronal connectivity on the circuit dynamics. The point spiking neuron model was sufficient for investigating the performance characteristics, spike timing dynamics and potential spike synchronisation effects in the ring attractors when exposed to neuronal noise, but clearly is highly abstracted in comparison to real neurons. However, most of the necessary detail is lacking to constrain more complex neural models. One caveat is that intrinsic properties of neurons could provide short-term memory that would radically alter the circuit response. It is not possible to explore this possibility with the models that were used, but it can be concluded that such properties do not appear to be necessary for generating basic ring attractor dynamics. Furthermore, it will be interesting to study how differences in the biophysical properties of neurons between the two species might be affecting performance. This possibility was not explored in this study (Pisokas, 2020).
This work has compared the hypothetical heading tracking circuit of two evolutionary distant species. The study beyond mere simulation of neuronal projection data by analysing and deriving the effective underlying circuit structure of the two ring attractors. This analysis and derivation of the complete effective neuronal circuits revealed not only differences in dynamics but also the construction principles of these circuits. This approach allowed identification of elements that differ in several ways from the 'canonical' ring attractor described in earlier theoretical models (Pisokas, 2020).
For example, the circuit found in the two insect species combines two functionalities in the P-EN neurons that are typically assigned to separate neural populations in computational models of ring attractors. Such computational models use one set of neurons to provide the lateral excitation to nearest neighbours and a different set of neurons that receive angular velocity input to drive the left-right rotation of the heading signal. In the insect circuit, the P-EN cells are part of the lateral excitation circuit, providing excitation to their two nearest neighbours, but they also receive angular velocity input. This difference is suggestive of a more efficient use of neuronal resources than the typical computational models of ring attractors. Another novel element found in the insect ring attractors is the presence of local feedback loops within each octant of the circuit structure (P-EG to E-PG and P-EN to E-PG). Both of these feedback loops increase the tolerance of the ring attractors to noise (Pisokas, 2020).
Another unique aspect of this modelling work is the comparison of related, but not identical, circuits found in two species. Indeed, using computational modelling allows investigation of 'hybrid' circuits, combining features of each, in order to try to understand the functional significance of each observed difference independently. Nevertheless, some differences between these circuits are not explained by the current model, and may require additional work to fully explicate (Pisokas, 2020).
One question is what is the role, if any, of the ninth PB glomeruli found so far only in Drosophila? In particular, the existence of the innermost glomeruli that are not innervated by the P-EN neurons seems perplexing. The same signals from tile 1 of the EB are sent to both ends of each hemisphere of the PB (glomeruli 1 and 9) and from there action potentials propagate along the Delta7 neurons along the PB length. It is speculated that this may be a mechanism to reduce the distance and time these signals have to travel to cover the full PB, that is, the maximum distance any signal must travel is only half of the distance it would need to propagate from one end of the PB to the other as in other species, such as the locust. If this is the case, it would constitute one more specialisation in Drosophila that reduces the response time of the ring attractor. It therefore seems that several specialisations have been orchestrated in minimising the response delays in fruit flies. Testing this idea would require multi-compartmental models to capture the action potential transmission time along neurites; as argued above, this may be contingent upon first obtaining detailed biophysical characterisation of the Delta7 neurons (Pisokas, 2020).
Another remaining question is what is the role of the closed ring-shaped EB in D. melanogaster. One possibility is that such a closed ring topology would allow local reciprocal connections between P-EN and E-PG neurons all around the EB ring, as reported in Turner-Evans (2019). This would allow direct propagation of signals between these neurons within the EB instead of requiring them to travel via the PB, as in the current model, again increasing the speed with which the heading direction can be tracked and allowing smoother transition between neighbouring tiles. Note that such direct reciprocal connections within the EB can only span the full ring with a closed ring anatomy and would not be possible between the two ends of the EB in the locust. To investigate the potential effect of such hypothetical reciprocal connections within the EB, further studies are required. Possibly blocking signal transmission via the PB to isolate functional connectivity within the EB would allow comparison of signal transmission time measurements within the EB versus via the PB. Such measurements would determine how different and hence significant those two pathways might be in the ring attractor performance (Pisokas, 2020).
A further hypothesis relates to the evolutionary lineage of these two features in the Drosophila CX. It will be of interest to study whether the ring shaped EB appeared before or after the appearance of the ninth glomeruli. One possibility is that the EB evolved into a ring shape after the appearance of the ninth glomeruli in the PB, allowing connections from one common tile to both glomeruli 1 and 9 and hence providing such a common driving signal. Alternatively, a pre-existing ring-shaped EB might have allowed the evolution of usable ninth glomeruli that resulted in faster propagation. Similarly, the P-EN to E-PG recurrency found only in the locust might be an acquired adaptation of the locust that increases robustness to noise, or an ancestral feature that has been lost in fruit flies (Pisokas, 2020).
Comparison of different species could potentially elucidate such questions. Individual species would be expected to have a selective subset of the specialisations that wer found, endowing them with brain circuits supporting the behavioural repertoire suiting their ecological niche. It will, therefore, be informative to analyse the effective heading direction circuit of other species, spanning evolutionary history, in order to get insights into how such adaptations relate to and accommodate behaviour. The results emphasise the importance of comparative studies if it is possible to derive general principles about neuronal processing, even in systems that appear highly conserved such as the CX head direction circuit in insects. Many of the circuit properties observed in Drosophila appear to reflect specific evolutionary adaptations related to tracking rapid flight manoeuvres. Despite the many strengths of Drosophila as an experimental model, it therefore remains important to ground conclusions about the insect brain in comparison with other species (Pisokas, 2020).
The Drosophila central brain consists of developmental-structural units of macrocircuitry formed by the sibling neurons of single neuroblasts. Lineage guides the connectivity and function of neurons, providing input to the central complex, a collection of neuropil compartments important for visually guided behaviors. The ellipsoid body (EB) is formed largely by the axons of ring (R) neurons, all of which are generated by a single lineage, DALv2. Two further lineages, DALcl1 and DALcl2, produce neurons that connect the anterior optic tubercle, a central brain visual center, with R neurons. Finally, DALcl1/2 receive input from visual projection neurons of the optic lobe medulla, completing a three-legged circuit that is called the anterior visual pathway (AVP). The AVP bears a fundamental resemblance to the sky-compass pathway, a visual navigation circuit described in other insects. DALcl1 and DALcl2 form two parallel channels, establishing connections with R neurons located in the peripheral and central domains of the EB, respectively. Although neurons of both lineages preferentially respond to bright objects, DALcl1 neurons have small ipsilateral, retinotopically ordered receptive fields, whereas DALcl2 neurons share a large excitatory receptive field in the contralateral hemifield. DALcl2 neurons become inhibited when the object enters the ipsilateral hemifield and display an additional excitation after the object leaves the field of view. Thus, the spatial position of a bright feature, such as a celestial body, may be encoded within this pathway (Omoto, 2017).
Although anatomy is often the first step in assigning functions to neural structures, it is not always clear whether architecturally distinct regions of the brain correspond to operational units. Whereas neuroarchitecture remains relatively static, functional connectivity may change almost instantaneously according to behavioral context. This study imaged panneuronal responses to visual stimuli in a highly conserved central brain region in the fruit fly, Drosophila, during flight. In one substructure, the fan-shaped body, automated analysis reveals three layers that are unresponsive in quiescent flies but become responsive to visual stimuli when the animal is flying. The responses of these regions to a broad suite of visual stimuli suggest that they are involved in the regulation of flight heading. To identify the cell types that underlie these responses, activity was imaged in sets of genetically defined neurons with arborizations in the targeted layers. The responses of this collection during flight also segregate into three sets, confirming the existence of three layers, and they collectively account for the panneuronal activity. These results provide an atlas of flight-gated visual responses in a central brain circuit (Weir, 2015).
Orientation and navigation in a complex environment requires path planning and recall to exert goal-driven behavior. Walking Drosophila flies possess a visual orientation memory for attractive targets which is localized in the central complex of the adult brain. This study shows that this type of working memory requires the cGMP-dependent protein kinase encoded by the foraging gene in just one type of ellipsoid-body ring neurons. Moreover, genetic and epistatic interaction studies provide evidence that Foraging functions upstream of the Ignorant Ribosomal-S6 Kinase 2, thus revealing a novel neuronal signaling pathway necessary for this type of memory in Drosophila (Kuntz, 2012).
For signaling has previously been implicated in different types of memories; however, in contrast to the working memory in the detour paradigm, these memories require a longer time frame to be established. In mammals, nitric oxide, the initiating molecule of the cGMP/PKG-pathway, is thought to act as a retrograde messenger during the induction of long-term potentiation (LTP). A LTP enhancement has been reported after adding PKG activators and a long-term depression after the addition of PKG inhibitors. Mice carrying a knock-out for the Pkg gene show reduced ability of motor learning due to a loss of synaptic plasticity in the cerebellum. Furthermore, mice lacking Pkg in the amygdala exhibit an impairment in fear conditioning and cGMP/PKG signaling in the hippocampus is required for novel object recognition. In insects, For is involved in different types of food searching behavior and associative memories in which establishing the learning traces takes at least seconds. In contrast, the orientation memory observed in the detour paradigm presented in this study represents a form of working memory which has to be updated continuously in fractions of seconds. Whereas the phosphorylation and activation of For and Ignorant might be the mechanism by which these kinases affect longer-lasting memories, it is thought unlikely that this mechanism is involved in the constantly and rapidly changing orientation memory. Both kinases would have to be activated or inactivated in an online fashion during every turn of the fly. On the other hand, RSK2 has been implicated in multiple cellular processes and transcriptional control. It is therefore speculated that the biochemical pathway both kinases work in is necessary to endow the ring neurons with the capacity to efficiently change signaling rapidly to encode orientation. For instance, ring neurons might need a higher density of synaptic release sites and/or dendritic neurotransmitter receptors to exert their specific function (Kuntz, 2012).
All organisms wishing to survive and reproduce must be able to respond adaptively to a complex, changing world. Yet the computational power available is constrained by biology and evolution, favouring mechanisms that are parsimonious yet robust. This study investigated the information carried in small populations of visually responsive neurons in Drosophila melanogaster. These so-called 'ring neurons', projecting to the ellipsoid body of the central complex, are reported to be necessary for complex visual tasks such as pattern recognition and visual navigation. Recently the receptive fields of these neurons have been mapped, allowing an investigation of how well they can support such behaviours. For instance, in a simulation of classic pattern discrimination experiments, this study shows that the pattern of output from the ring neurons matches observed fly behaviour. However, performance of the neurons (as with flies) is not perfect and can be easily improved with the addition of extra neurons, suggesting the neurons' receptive fields are not optimised for recognising abstract shapes, a conclusion which casts doubt on cognitive explanations of fly behaviour in pattern recognition assays. Using artificial neural networks, it was then assessed how easy it is to decode more general information about stimulus shape from the ring neuron population codes. These neurons were shown to be well suited for encoding information about size, position and orientation, which are more relevant behavioural parameters for a fly than abstract pattern properties. This leads to a suggestion that in order to understand the properties of neural systems, one must consider how perceptual circuits put information at the service of behaviour (Dewar, 2017).
Chemotaxis, the ability to direct movements according to chemical cues in the environment, is important for the survival of most organisms. The vinegar fly, Drosophila melanogaster, displays robust olfactory aversion and attraction, but how these behaviors are executed via changes in locomotion remains poorly understood. In particular, it is not clear whether aversion and attraction bidirectionally modulate a shared circuit or recruit distinct circuits for execution. Using a quantitative behavioral assay, this study has determined that both aversive and attractive odorants modulate the initiation and direction of turns but display distinct kinematics. Using genetic tools to perturb these behaviors, specific populations of neurons were identified that are required for aversion, but not for attraction. Inactivation of these populations of cells affects the completion of aversive turns, but not their initiation. Optogenetic activation of the same populations of cells triggers a locomotion pattern resembling aversive turns. Perturbations in both the ellipsoid body and the ventral nerve cord, two regions involved in motor control, were shown to result in defects in aversion. It is concluded that aversive chemotaxis in vinegar flies triggers ethologically appropriate kinematics distinct from those of attractive chemotaxis and requires specific motor-related neurons (Gao, 2013).
This study is the first direct demonstration that odorants modulate turn initiation and direction in freely walking insects. Moreover, aversive and attractive turns involve distinct kinematics. Intuitively, these quantitative analyses reveal that flies speed up and follow straighter trajectories after turning away from a noxious smell, which should shorten their exposure to potential harm. Such a strategy is not employed for attraction. Chemotaxis has been studied in tethered adult flies, paradigms in which mimicking the olfactory inputs a freely moving fly would encounter proved challenging. For example, in one group of studies, a 'fly-on-the-ball' paradigm, aversion was not triggered even using a strong repellent. In another study, flying flies responded symmetrically to aversive and attractive odorants. The current more naturalistic approach provides new insights into the relationship between aversive and attractive chemotaxes (Gao, 2013).
In bacteria, aversion and attraction are achieved through bidirectional modulation of the same mechanism. Similarly, in C. elegans, aversion and attraction are thought to utilize a push-pull mechanism on one set of antagonizing command neurons (Faumont, 2012). Genetic inactivation experiments suggest that, in flies, aversion is executed through specific neurons distinct from attraction. In this study. two candidate circuit components, the EB and a subset of the VNC neurons, were identified that appear redundantly necessary for aversive chemotaxis. The EB is part of the central complex, defects in which are associated with uncoordinated walking. In grasshoppers and cockroaches, activating central complex neurons induces specific kinematics. In the VNC, dTdc2+ neurons are prominent candidates for mediating aversion, although other neurons might be involved. These othere neurons are homologous to dorsal/ventral unpaired median neurons in other insects because they are octopaminergic and show similar projection patterns]. The activity in these neurons is correlated with specific aspects of locomotion in locusts, crickets, and moths. Given that 441 > shits1 flies display defects in aversive turn completion but not initiation, it is postulated that the current genetic manipulation does not interfere with the perception, processing, decision-making, or even initiation steps of aversive chemotaxis, but rather the execution of motor programs specifically necessary for this behavior. These discoveries bridge the extensive investigation of olfactory processing in insects such as honeybees and moths with studies focused on motor control mechanisms in species such as cockroaches and stick insects (Gao, 2013).
It is intriguing that part of the aversion-specific circuit resides in the VNC, and can be artificially activated to generate a pattern similar to aversive turns. Although the larval VNC is sufficient for substrate exploration, VNC autonomy in motor pattern generation in adult flies has only been established for escape flight and courtship song, both highly specialized for certain ethological functions. When a fly continuously explores the environment and updates its walking pattern, the division of labor between the brain and the VNC is less clear. Conceptually, one possibility is that circuit modules in the VNC only encode basic elements of locomotion. For example, right turns may always involve the same VNC circuit, and the only difference is their embedding within different sequences of actions based on the combination of descending signals from the brain. Alternatively, VNC circuit modules could be task-specific; once a descending signal specifies the task, the details of the motor output will unfold according to a pre-wired VNC circuit. The current findings support the latter possibility in the context of aversive chemotaxis. In both vertebrates and invertebrates, artificial activation of neurons in the spinal cord or the VNC generates specific motor outputs, but rarely have these neurons been demonstrated to be necessary for specific sensory-driven tasks. It would be interesting to test the generality of having autonomous motor-related circuits specifically responsive to certain sensory triggers (Gao, 2013).
In Drosophila, aversive olfactory memory is believed to be stored in a prominent brain structure, the mushroom body (MB), and two pairs of MB intrinsic neurons, the dorsal paired medial (DPM) and the anterior paired lateral (APL) neurons, are found to regulate the consolidation of middle-term memory (MTM). This study reports that another prominent brain structure, the ellipsoid body (EB), is also involved in the modulation of olfactory MTM. Activating EB R2/R4m neurons does not affect the learning index, but specifically eliminates anesthesia-sensitive memory (ASM), the labile component of olfactory MTM. It was further demonstrated that approximately two-thirds of these EB neurons are GABAergic and are responsible for the suppression of ASM. Using GRASP (GFP reconstitution across synaptic partners), potential synaptic connections were revealed between the EB and MB in regions covering both the presynaptic and postsynaptic sites of EB neurons, suggesting the presence of bidirectional connections between these two important brain structures. These findings suggest the existence of direct connections between the MB and EB, and provide new insights into the neural circuit basis for olfactory labile memory in Drosophila (Zhang, 2014).
Previous studies have shown that the EB plays an essential role in visual pattern memory, orientation memory and place learning (Pan, 2009; Ofstad, 2011; Kuntz, 2012), and thus it is usually considered to be a center of visual learning and mem- ory. Interestingly, one study on NMDA receptors reported that the EB is required for olfactory long-term memory (LTM) consolidation; however, the underlying neural circuits remain uninvestigated. The current results reveal that a group of EB neurons, the c819-labeled R2/R4m neurons, plays an inhibitory role in the modulation of MTM but not the immediate memory. This points to a new function of the EB in olfactory cognition and further demonstrates that the EB could be involved in the process of olfactory aversive learning and memory from an earlier stage than previously thought (Zhang, 2014).
The presence of dense GABA-like immunoreactivity has been demonstrated in the EB ring and RF tract suggesting that the bulk of EB neurons are GABAergic. Although this finding has been confirmed by several other studies, the function of these GABAergic neurons in cognition is still unclear. The current results further reveal that approximately two-thirds of the c819-EB neurons are GABAergic, and they play an inhibitory role in ASM modulation. The GABAA receptor, resistant to dieldrin (RDL), has been shown to be highly expressed in the MB lobes and the EB. It is thus possible that these EB GABAergic neurons function through RDL receptors (Zhang, 2014).
As a component of MTM, ASM has been suggested to be stored in the MB and to be modulated by MB intrinsic APL and DPM neurons, of which the neural terminals are restricted to the MB. This study reports the EB, a brain structure separate from the MB, is also involved in the modulation of ASM. WEB neurons may be both presynaptic and postsynaptic to MB neurons, suggesting that they may suppress 3 h ASM via putative direct connections between the EB and MB. That the MB and EB are two discrete but possibly interconnected and interacting brain regions, suggests that it is important to study the process of learning and memory over a more widely distributed neural network. However, the interaction between neurons from different structures may endow the network with greater capacity for more complex activities (Zhang, 2014).
It is also interesting to discover that activating c819-EB GABAergic neurons during training impaired 3 h ASM instead of immediate learning performance. Recently it has been reported that blocking two pairs of dopaminergic neurons during intertrial intervals in spaced training suppresses the formation of 24 h LTM by interfering with the slow oscillations in these dopaminergic neurons. Since this study has shown that bidirectional connections may exist between the EB and MB, it is proposed that there may be a small feedback circuit between the EB and MB, which may have prolonged oscillations and therefore affect 3 h ASM consolidation. Further functional imaging studies may provide more clues on how this neural circuit functions (Zhang, 2014).
CREB (cAMP response element-binding protein) is an evolutionarily conserved transcription factor, playing key roles in synaptic plasticity, intrinsic excitability and long-term memory (LTM) formation. The Drosophila homologue of mammalian CREB, dCREB2, is also important for LTM. However, the spatio-temporal nature of dCREB2 activity during memory consolidation is poorly understood. Using an in vivo reporter system, this study examined dCREB2 activity continuously in specific brain regions during LTM processing. Two brain regions that have been shown to be important for Drosophila LTM are the ellipsoid body (EB) and the mushroom body (MB). dCREB2 reporter activity is persistently elevated in EB R2/R4m neurons, but not neighboring R3/R4d neurons, following LTM-inducing training. In multiple subsets of MB neurons, dCREB2 reporter activity is suppressed immediately following LTM-specific training, and elevated during late windows. In addition, heterogeneous responses were observed across different subsets of neurons in MB αβ lobe during LTM processing. All of these changes suggest that dCREB2 functions in both the EB and MB for LTM formation, and that this activity contributes to the process of systems consolidation (Zhang, 2014).
Intracellular Ca(2+) is a widely used neuronal activity indicator. This study describes a transcriptional reporter of intracellular Ca(2+) (TRIC) in Drosophila that uses a binary expression system to report Ca(2+)-dependent interactions between calmodulin and its target peptide. In vitro assays predicted in vivo properties of TRIC. TRIC signals in sensory systems were show to depend on neuronal activity. TRIC was able to quantitatively monitor neuronal responses that changed slowly, such as those of neuropeptide F-expressing neurons to sexual deprivation and neuroendocrine pars intercerebralis (PI) cells to food and arousal. Furthermore, TRIC-induced expression of a neuronal silencer in nutrient-activated cells enhanced stress resistance, providing a proof of principle that TRIC can be used for circuit manipulation. Thus, TRIC facilitates the monitoring and manipulation of neuronal activity, especially those reflecting slow changes in physiological states that are poorly captured by existing methods. TRIC's modular design should enable optimization and adaptation to other organisms (Gao, 2015).
Using cultured cells and multiple in vivo assays, this study found that TRIC reports changes in Ca2+ levels under diverse conditions in visual, olfactory and neuromodulatory systems. The results provide quantitative assessments for choosing TRIC variants with appropriate sensitivity and stringency, and proof of principle that TRIC can be used to express a circuit manipulator. Thus, TRIC acts as a useful complement to functional Ca2+ imaging by integrating changes in activity over long periods of time and offering genetic access to neurons on the basis of their activity (Gao, 2015).
Vertebrate immediate-early-gene (IEGs) which evolved to be expressed in a high signal-to-baseline ratio in response to neuronal activation, are widely used to report neuronal activity. However, as they rely on endogenous signaling networks, their response properties and cell-type biases are difficult to modify. TRIC can be considered a rationally designed IEG, by exogenously introducing a protein-peptide interaction to detect Ca2+. The modular design of TRIC renders it more amenable to optimization. TRIC reports a rise in nuclear Ca2+ levels, which have previously been used to monitor pan-neuronal activity in C. elegans, and also accompanies neuronal activation in mammalian neurons likely shuttled by Ca2+-binding proteins. The current experiments indicate that nuclear Ca2+ correlates with activity in diverse neuronal classes in flies. It is likely that not all cell types have the same efficiency in converting cytoplasmic Ca2+ signal to nuclear Ca2+ signal. Thus, TRIC efficiency and optimization may differ for different neuronal types (Gao, 2015).
While this manuscript was in review, a Ca2+ integrator (CaMPARI) was reported in which the ultraviolet conversion of emission spectrum of a fluorescent protein was engineered to be contingent on Ca2+ concentration. CaMPARI can capture neuronal activity on a shorter time scale than TRIC or IEG. However, access of neurons to ultraviolet may limit the use of CaMPARI in deep tissues, at least in large animals, whereas TRIC and IEG report neuronal activity in the entire nervous system non-invasively. Notably, unlike CaMPARI or IEG, TRIC offers genetic access to active neurons, allowing activity-based circuit manipulation (Gao, 2015).
The results underscore the importance of optimizing TRIC for specific neuronal types. In this study, TRIC was optimized for multiple cell types, and many variants were described that can help users in other cells. It is recommended that users begin with CaM/MKII-mediated TRIC in their neurons of interest. If TRIC signal is detected, the users can attempt QA-mediated or FLP-mediated regulation of the timing of TRIC onset. The signal-to-baseline ratio can be further improved by titrating expression of TRIC using QA, choosing reporters with different stabilities, or switching to nlsLexADBDo or the MKIIK11A variant. Stoichiometry can also be leveraged to boost TRIC signal (Gao, 2015).
With the current version of TRIC, the signal accumulates and decays over many hours. To detect shorter periods of neuronal activity, an important future goal is to increase signal strength while avoiding saturation by basal Ca2+ concentrations. One solution to this problem would be to restrict TRIC to a narrower time window than that offered by the QA- or the FLP-mediated strategy. For example, TRIC could be split into DBD-X, Y-target peptide and CaM-AD, where X and Y are two interacting modules controlled by light. One could then synchronize TRIC with a specific manipulation, or even trigger TRIC repetitively with specific behavioral features using feedback from automated tracking. To preserve phasic information about neuronal activity, reporters with faster decays than CD8::GFP could be used or the TRIC components could be destabilized with tags for protein degradation. Given that the current TRIC was able to interact with endogenous CaM and its target peptides, another important direction is to 'isolate' TRIC by co-engineering the CaM and MKII components to lose binding to their endogenous partners, but maintain their mutual interaction. Future TRIC optimization could be achieved using high throughput screens in cultured cells, which can predict in vivo performance (Gao, 2015).
Previous studies used Ilp2 immunostaining, epitope-tagged Ilp2 or a secreted GFP as indirect indicators of PI activity. The major conclusions of these studies were validated using TRIC. After enhancing the dynamic range of TRIC, additional insight was gained into how PI activity is regulated. In particular, given that PI cells affect diverse processes, how do these cells determine their output according to all relevant inputs? For example, an animal may encounter conflicting metabolic needs, such as conserving energy versus defending territory in an impoverished environment. Nutrient and OA comparison could be viewed as a minimal model of such a dilemma, as OA contributes to arousal and is necessary for 'fight or flight' in insects. PI cells exhibited graded, yet more readily saturated, responses to such events. In contrast, the linear PI response to nutrients extended over a wider range. These distinctions, as well as the additive interaction between yeast and OA, point to the independent operation of these two categories of inputs. To further survey the input landscape, one could genetically manipulate candidate receptors autonomously or candidate upstream neurons non-autonomously while monitoring PI activity using TRIC (Gao, 2015).
The physiological states of flies can change over hours to days and can be accompanied by changes in the activities of neurons expressing modulatory neurotransmitters or neuropeptides. Although previous work has focused on the targets of modulatory neurotransmitters, inputs to these cells remain largely unknown. In addition, there are ~75 predicted neuropeptides in flies, only a small subset of which have been examined. TRIC can be applied to neurons expressing specific transmitters or neuropeptides and tested in different physiological states (for example, the NPF neurons). It is noted that the current TRIC variants might not fit the dynamic range of all neuronal types, and it might be necessary to test other AD/DBD ratios or other MKII mutants following the examples in this paper of optimization for PI cells (Gao, 2015).
Finally, TRIC can report a rise of intracellular Ca2+ that accompanies any cellular, developmental or physiological processes in flies and can be adapted for similar use in other model organisms. TRIC modules can be introduced as transgenes or by viral vectors, and specific stoichiometry can be achieved by specifying the number of AD and DBD sequences in multi-cistronic constructs. TRIC expression can be made contingent on recombinase or other binary systems in model organisms, such as mice, where many Cre lines are available for spatiotemporal control, which can help refine activity monitoring and circuit manipulation in specific cell types (Gao, 2015).
Metabolic homeostasis is regulated by the brain, but whether this regulation involves learning and memory of metabolic information remains unexplored. This study use a calorie-based, taste-independent learning/memory paradigm to show that Drosophila form metabolic memories that help in balancing food choice with caloric intake; however, this metabolic learning or memory is lost under chronic high-calorie feeding. Loss of individual learning/memory-regulating genes causes a metabolic learning defect, leading to elevated trehalose and lipid levels. Importantly, this function of metabolic learning requires not only the mushroom body but also the hypothalamus-like pars intercerebralis, while NF-κB activation in the pars intercerebralis mimics chronic overnutrition in that it causes metabolic learning impairment and disorders. Finally, this study evaluated this concept of metabolic learning/memory in mice, suggesting that the hypothalamus is involved in a form of nutritional learning and memory, which is critical for determining resistance or susceptibility to obesity. In conclusion, these data indicate that the brain, and potentially the hypothalamus, direct metabolic learning and the formation of memories, which contribute to the control of systemic metabolic homeostasis (Zhang, 2015).
This work consists of a series of studies in Drosophila: these animals were found to temporarily develop metabolic learning to balance food choice with caloric intake. In Drosophila research, sugar has often been used for studying the appetitive reward value of food taste. Of interest, recent research has suggested that fruit flies can distinguish caloric values from the taste property of food. Using tasteless sorbitol as a carbohydrate source to generate an environmental condition that contained NC versus HC food, this study revealed that Drosophila can develop a form of metabolic learning and memory independently of taste, by which flies are guided to have a preference for normal caloric environment rather than high-caloric environment. However, this form of metabolic memory does not seem robust, as it is vulnerably diminished under genetic or environmental influences. It is postulated that this vulnerability to overnutrition is particularly prominent for mammals (such as C57BL/6J mice), and overnutritional reward-induced excess in caloric intake can quickly become dominant. This effect can be consistently induced in Drosophila when learning/memory-regulating genes are inhibited in the brain or the hypothalamus-like PI region. It was observed that each of these genetic disruptions led to impaired metabolic learning, resulting in increased caloric intake and, on a chronic basis, the development of lipid excess and diabetes-like phenotype. Indeed, it has been documented that chronic high-sugar feeding is sufficient to cause insulin resistance, obesity and diabetes in Drosophila. It is yet unclear whether this metabolic learning can induce an appetitive memory of normal caloric environment or an aversive memory of high-caloric environment. Regardless, the findings in this work have provoked a stimulating question, that is, whether this form of metabolic learning and memory is present in the mammals and, if so, whether this mechanism can be consolidated to improve the control of metabolic physiology and prevent against diseases. These mouse studies may provide an initial support to this concept and strategy, but clearly, in-depth future research is much needed (Zhang, 2015).
In light of the underlying neural basis for this metabolic learning, this study indicates that multiple brain regions are required, including the hypothalamus-like PI region in addition to the MB (equivalent to the hippocampus in mammals), which is classically needed for learning and memory formation. Anatomically, the PI region is located in the unpaired anteromedial domain of the protocerebral cortex, which is near the calyces of the MB and the dorsal part of the central complex (another brain region for regulating learning and memory). Functionally, the PI region has been demonstrated to coordinate with the MB in regulating various physiological activities in Drosophila. Thus, it is very possible that some PI neurons present nutritional information to the MB and thus induce metabolic learning and memory formation. However, the underlying detailed mechanism is still unknown, especially if this process involves a role of dilps, which represent the prototypical neuropeptides produced by the PI neurons. Considering that the PI region in flies is equivalent to the mammalian hypothalamus, this study was extended to mouse models by comparatively analysing A/J versus C57BL/6J mice—which are known to have different diet preference as well as different susceptibilities to obesity development. While A/J mice showed a learning process of distinguishing NC versus HC food, C57BL/6J mice failed to do so. It is particularly notable that this difference of learning and memory between these two strains is associated with differential expression profiles of learning/memory genes in the hypothalamus rather than the hippocampus. This finding, in conjunction with the Drosophila study, highlights a potential that the hypothalamus has a unique role in mediating metabolic learning and memory formation. Although the mouse experiments cannot exclude the impacts from the taste/smell properties of the studied food, the results demonstrated that there is a form of nutritional memory, which seems dissociable from the memory of overnutritional reward. These initial observations in mice lend an agreement with findings in Drosophila, suggesting that the brain and potentially the hypothalamus can link nutritional environment to a form of metabolic learning and memory homeostasis (Zhang, 2015).
From a disease perspective, metabolic learning in Drosophila is impaired under chronic overnutrition, and the mouse study was in line with this understanding. This response to overnutrition is useful when famine is outstanding; however, it is a dilemma when metabolic disease is of concern, much like the scenario pertaining to leptin resistance under chronic overnutrition, whereas an increase in leptin sensitivity is demanded to reduce obesity. Recently, it was established that NF-κB-dependent hypothalamic inflammation links chronic overnutrition to the central dysregulation of metabolic balance. This study showed that activation of the NF-κB pathway in the PI region weakened the function of metabolic learning and, conversely, NF-κB inhibition in this region provided a protective effect against chronic overnutrition-impaired metabolic learning. These findings are in alignment with the literature, for example, pan-neuronal NF-κB inhibition was shown to improve activity-dependent synaptic signalling and cognitive function including learning and memory formation, and persistent NF-κB activation inhibits neuronal survival and the function of learning and memory formation. Hence, overnutrition-induced neural NF-κB activation has a negative impact on metabolic learning and memory formation in regulating metabolic homeostasis homeostasis (Zhang, 2015).
To summarize the findings in this work, a series of behavioural studies was performed revealing that Drosophila have a form of metabolic learning and memory, through which the flies are directed to balance food choice with caloric intake in relevant environments. Several learning/memory-regulating genes including rut, dnc and tequila are involved in this function, and brain regions including the PI in addition to the MB are required to induce this mechanism. On the other hand, metabolic learning is impaired under chronic overnutrition through NF-κB activation, leading to excess exposure to calorie-enriched environment, which causes metabolic disorders. Overall, metabolic learning and memory formation by the brain and potentially the hypothalamus play a role in controlling metabolic homeostasis homeostasis (Zhang, 2015).
Prolonged wakefulness leads to an increased pressure for sleep, but how this homeostatic drive is generated and subsequently persists is unclear. From a neural circuit screen in Drosophila, this study identified a subset of ellipsoid body (EB) neurons whose activation generates sleep drive. Patch-clamp analysis indicates these EB neurons are highly sensitive to sleep loss, switching from spiking to burst-firing modes. Functional imaging and translational profiling experiments reveal that elevated sleep need triggers reversible increases in cytosolic Ca(2+) levels, NMDA receptor expression, and structural markers of synaptic strength, suggesting these EB neurons undergo 'sleep-need'-dependent plasticity. Strikingly, the synaptic plasticity of these EB neurons is both necessary and sufficient for generating sleep drive, indicating that sleep pressure is encoded by plastic changes within this circuit. These studies define an integrator circuit for sleep homeostasis and provide a mechanism explaining the generation and persistence of sleep drive (Liu, 2016).
In Drosophila, a crucial component of the machinery for sleep homeostasis is a cluster of neurons innervating the dorsal fan-shaped body (dFB) of the central complex. dFB neurons in sleep-deprived flies tend to be electrically active, with high input resistances and long membrane time constants, while neurons in rested flies tend to be electrically silent. This study demonstrates state switching by dFB neurons, identifies dopamine as a neuromodulator that operates the switch, and delineates the switching mechanism. Arousing dopamine causes transient hyperpolarization of dFB neurons within tens of milliseconds and lasting excitability suppression within minutes. Both effects are transduced by Dop1R2 receptors and mediated by potassium conductances. The switch to electrical silence involves the downregulation of voltage-gated A-type currents carried by Shaker and Shab, and the upregulation of voltage-independent leak currents through a two-pore-domain potassium channel that was termed Sandman. Sandman is encoded by the CG8713 gene and translocates to the plasma membrane in response to dopamine. dFB-restricted interference with the expression of Shaker or Sandman decreases or increases sleep, respectively, by slowing the repetitive discharge of dFB neurons in the ON state or blocking their entry into the OFF state. Biophysical changes in a small population of neurons are thus linked to the control of sleep-wake state (Pimentel, 2016).
Recordings were made from dFB neurons (which were marked by R23E10-GAL4 or R23E10-lexA-driven green fluorescent protein (GFP) expression) while head-fixed flies walked or rested on a spherical treadmill. Because inactivity is a necessary correlate but insufficient proof of sleep, the analysis was restricted to awakening, which is defined as a locomotor bout after >5 min of rest, during which the recorded dFB neuron had been persistently spiking. To deliver wake-promoting signals, the optogenetic actuator CsChrimson was expressed under TH-GAL4 control in the majority of dopaminergic neurons, including the PPL1 and PPM3 clusters, whose fan-shaped body (FB)-projecting members have been implicated in sleep control. Illumination at 630 nm, sustained for 1.5 s to release a bolus of dopamine, effectively stimulated locomotion. dFB neurons paused in successful (but not in unsuccessful) trials, and their membrane potentials dipped by 2-13 mV below the baseline during tonic activity. When flies bearing an undriven CsChrimson transgene were photostimulated, neither physiological nor behavioural changes were apparent. The tight correlation between the suppression of dFB neuron spiking and the initiation of movement might, however, merely mirror a causal dopamine effect elsewhere, as TH-GAL4 labels dopaminergic neurons throughout the brain. Because localized dopamine applications to dFB neuron dendrites similarly caused awakening, this possibility is considered remote (Pimentel, 2016).
Flies with enhanced dopaminergic transmission exhibit a short-sleeping phenotype that requires the presence of a D1-like receptor in dFB neurons, suggesting that dopamine acts directly on these cells. dFB-restricted RNA interference (RNAi) confirmed this notion and pinpointed Dop1R2 as the responsible receptor, a conclusion reinforced by analysis of the mutant Dop1R2MI08664 allele. Previous evidence that Dop1R1, a receptor not involved in regulating baseline sleep, confers responsiveness to dopamine when expressed in the dFB indicates that either D1-like receptor can fulfill the role normally played by Dop1R2. Loss of Dop1R2 increased sleep during the day and the late hours of the night, by prolonging sleep bouts without affecting their frequency. This sleep pattern is consistent with reduced sensitivity to a dopaminergic arousal signal (Pimentel, 2016).
To confirm the identity of the effective transmitter, avoid dopamine release outside the dFB, and reduce the transgene load for subsequent experiments, optogenetic manipulations of the dopaminergic system were replaced with pressure ejections of dopamine onto dFB neuron dendrites. Like optogenetically stimulated secretion, focal application of dopamine hyperpolarized the cells and suppressed their spiking. The inhibitory responses could be blocked at several nodes of an intracellular signalling pathway that connects the activation of dopamine receptors to the opening of potassium conductances: by RNAi-mediated knockdown of Dop1R2; by the inclusion in the patch pipette of pertussis toxin (PTX), which inactivates heterotrimeric G proteins of the Gi/o family; and by replacing intracellular potassium with caesium, which obstructs the pores of G-protein-coupled inward-rectifier channels. Elevating the chloride reversal potential above resting potential left the polarity of the responses unchanged, corroborating that potassium conductances mediate the bulk of dopaminergic inhibition (Pimentel, 2016).
Coupling of Dop1R2 to Gi/o, although documented in a heterologous system, represents a sufficiently unusual transduction mechanism for a predicted D1-like receptor to prompt verification of its behavioural relevance. Like the loss of Dop1R2, temperature-inducible expression of PTX in dFB neurons increased overall sleep time by extending sleep bout length (Pimentel, 2016).
While a single pulse of dopamine transiently hyperpolarized dFB neurons and inhibited their spiking, prolonged dopamine applications (50 ms pulses at 10 Hz, or 20 Hz optogenetic stimulation, both sustained for 2-10 min) switched the cells from electrical excitability (ON) to quiescence (OFF). The switching process required dopamine as well as Dop1R2, but once the switch had been actuated the cells remained in the OFF state-and flies, awake-without a steady supply of transmitter. Input resistances and membrane time constants dropped to 53.3 ± 1.8 and 24.0 ± 1.3% of their initial values (means ± s.e.m.), and depolarizing currents no longer elicited action potentials (15 out of 15 cells). The biophysical properties of single dFB neurons, recorded in the same individual before and after operating the dopamine switch, varied as widely as those in sleep-deprived and rested flies (Pimentel, 2016).
Dopamine-induced changes in input resistance and membrane time constant occurred from similar baselines in all genotypes and followed single-exponential kinetics with time constants of 1.07-1.10 min. The speed of conversion points to post-translational modification and/or translocation of ion channels between intracellular pools and the plasma membrane as the underlying mechanism(s). In 7 out of 15 cases, recordings were held long enough to observe the spontaneous recommencement of spiking, which was accompanied by a rise to baseline of input resistance and membrane time constant, after 7-60 min of quiescence (mean ± s.e.m. = 25.86 ± 7.61 min). The temporary suspension of electrical output is thus part of the normal activity cycle of dFB neurons and not a dead end brought on by the experimental conditions (Pimentel, 2016).
dFB neurons in the ON state expressed two types of potassium current: voltage-dependent A-type (rapidly inactivating) and voltage-independent non-A-type currents. The current-voltage (I-V) relation of iA resembled that of Shaker, the prototypical A-type channel: no current flowed below -50 mV, the approximate voltage threshold of Shaker; above -40 mV, peak currents increased steeply with voltage and inactivated with a time constant of 7.5 ± 2.1 ms (mean ± s.e.m.). Non-A-type currents showed weak outward rectification with a reversal potential of -80 mV, consistent with potassium as the permeant ion, and no inactivation (Pimentel, 2016).
Switching the neurons OFF changed both types of potassium current. iA diminished by one-third, whereas inon-A nearly quadrupled when quantified between resting potential and spike threshold. The weak rectification of inon-A in the ON state vanished in the OFF state, giving way to the linear I-V relationship of an ideal leak conductance. dFB neurons thus upregulate iA in the sleep-promoting ON state. When dopamine switches the cells OFF, voltage-dependent currents are attenuated and leak currents augmented. This seesaw form of regulation should be sensitive to perturbations of the neurons' ion channel inventory: depletion of voltage-gated A-type (KV) channels (which predominate in the ON state) should tip the cells towards the OFF state; conversely, loss of leak channels (which predominate in the OFF state) should favour the ON state. To test these predictions, sleep was examined in flies carrying R23E10-GAL4-driven RNAi transgenes for dFB-restricted interference with individual potassium channel transcripts (Pimentel, 2016).
RNAi-mediated knockdown of two of the five KV channel types of Drosophila (Shaker and Shab) reduced sleep relative to parental controls, while knockdown of the remaining three types had no effect. Biasing the potassium channel repertoire of dFB neurons against A-type conductances thus tilts the neurons' excitable state towards quiescence, causing insomnia, but leaves transient and sustained dopamine responses unaffected. The seemingly counterintuitive conclusion that reducing a potassium current would decrease, not increase, action potential discharge is explained by a requirement for A-type channels in generating repetitive activity of the kind displayed by dFB neurons during sleep. Depleting Shaker from dFB neurons shifted the interspike interval distribution towards longer values, as would be expected if KV channels with slow inactivation kinetics replaced rapidly inactivating Shaker as the principal force opposing the generation of the next spike. These findings identify a potential mechanism for the short-sleeping phenotypes caused by mutations in Shaker, its β subunit Hyperkinetic, or its regulator sleepless (Pimentel, 2016).
Leak conductances are typically formed by two-pore-domain potassium (K2P) channels. dFB-restricted RNAi of one member of the 11-strong family of Drosophila K2P channels, encoded by the CG8713 gene, increased sleep relative to parental controls; interference with the remaining 10 K2P channels had no effect. Recordings from dFB neurons after knockdown of the CG8713 gene product, which this study termed Sandman, revealed undiminished non-A-type currents in the ON state and intact responses to a single pulse of dopamine but a defective OFF switch: during prolonged dopamine applications, inon-A failed to rise, input resistances and membrane time constants remained at their elevated levels, and the neurons continued to fire action potentials (7 out of 7 cells). Blocking vesicle exocytosis in the recorded cell with botulinum neurotoxin C (BoNT/C) similarly disabled the OFF switch. This, combined with the absence of detectable Sandman currents in the ON state, suggests that Sandman is internalized in electrically active cells and recycled to the plasma membrane when dopamine switches the neurons OFF (Pimentel, 2016).
Because dFB neurons lacking Sandman spike persistently even after prolonged dopamine exposure, voltage-gated sodium channels remain functional in the OFF state. The difficulty of driving control cells to action potential threshold in this state must therefore be due to a lengthening of electrotonic distance between sites of current injection and spike generation. This lengthening is an expected consequence of a current leak, which may uncouple the axonal spike generator from somatodendritic synaptic inputs or pacemaker currents when sleep need is low (Pimentel, 2016).
The two kinetically and mechanistically distinct actions of dopamine on dFB neurons-instant, but transient, hyperpolarization and a delayed, but lasting, switch in excitable state-ensure that transitions to vigilance can be both immediate and sustained, providing speedy alarm responses and stable homeostatic control. The key to stability lies in the switching behaviour of dFB neurons, which is driven by dopaminergic input accumulated over time. Unlike bistable neurons, in which two activity regimes coexist for the same set of conductances, dFB neurons switch regimes only when their membrane current densities change. This analysis of how dopamine effects such a change, from activity to silence, has uncovered elements familiar from other modulated systems: simultaneous, antagonistic regulation of multiple conductances; reduction of iA; and modulation of leak currents. Currently little is known about the reverse transition, from silence to activity, except that mutating the Rho-GTPase-activating protein Crossveinless-c locks dFB neurons in the OFF state, resulting in severe insomnia and an inability to correct sleep deficits. Discovering the signals and processes that switch sleep-promoting neurons back ON will hold important clues to the vital function of sleep (Pimentel, 2016).
Implicit knowledge of peripersonal space in humans is first acquired during infancy but will be continuously updated throughout life. In contrast, body size of holometabolous insects does not change after metamorphosis; nevertheless, they do have to learn their body reaches at least once. The body size of Drosophila imagines can vary by about 15% depending on environmental factors like food quality and temperature. To investigate how flies acquire knowledge about and memorize their body size, their decisions to either refrain from or initiate climbing over gaps exceeding their body size were studied. Naive (dark-reared) flies overestimate their size and have to learn it from the parallax motion of the retinal images of objects in their environment while walking. Naive flies can be trained in a striped arena and manipulated to underestimate their size, but once consolidated, this memory seems to last for a lifetime. Consolidation of this memory is stress sensitive only in the first 2 h after training but cannot be retrieved for the next 12 h. This study has identified a set of intrinsic, lateral neurons of the protocerebral bridge of the central complex that depend on dCREB2 transcriptional activity for long-term memory consolidation and maintenance (Krause, 2019).
Emerging evidence indicates the role of amino acid metabolism in sleep regulation. This study demonstrates sleep-promoting effects of dietary threonine (SPET) in Drosophila. Dietary threonine markedly increased daily sleep amount and decreased the latency to sleep onset in a dose-dependent manner. High levels of synaptic GABA or pharmacological activation of metabotropic GABA receptors (GABAB-R) suppressed SPET. By contrast, synaptic blockade of GABAergic neurons or transgenic depletion of GABAB-R in the ellipsoid body R2 neurons enhanced sleep drive non-additively with SPET. Dietary threonine reduced GABA levels, weakened metabotropic GABA responses in R2 neurons, and ameliorated memory deficits in plasticity mutants. Moreover, genetic elevation of neuronal threonine levels was sufficient for facilitating sleep onset. Taken together, these data define threonine as a physiologically relevant, sleep-promoting molecule that may intimately link neuronal metabolism of amino acids to GABAergic control of sleep drive via the neuronal substrate of sleep homeostasis (Ki, 2019).
The circadian clock and sleep homeostasis are two key regulators that shape daily sleep behaviors in animals. In stark contrast to the homeostatic nature of sleep, the internal machinery of sleep is vulnerable to external (e.g., environmental change) or internal conditions (e.g., genetic mutation) that lead to adaptive changes in sleep behaviors. Sleep behavior is conserved among mammals, insects, and even lower eukaryotes. Since the identification of the voltage-gated potassium channel Shaker as a sleep-regulatory gene in Drosophila, fruit flies have been one of the most advantageous genetic models to dissect molecular and neural components that are important for sleep homeostasis and plasticity (Ki, 2019).
To date, a number of sleep-regulatory genes and neurotransmitters have been identified in animal models as well as in humans. For instance, the inhibitory neurotransmitter gamma-aminobutyric acid (GABA) is known to have a sleep-promoting role that is conserved in invertebrates and vertebrates. Hypomorphic mutations in mitochondrial GABA-transaminase (GABA-T) elevate GABA levels and lengthen baseline sleep in flies (Chen, 2015). The long sleep phenotype in GABA-T mutants accompanies higher sleep consolidation and shorter latency to sleep onset, consistent with the observations that pharmacological enhancement of GABAergic transmission facilitates sleep in flies and mammals, including humans. In addition, resistance to dieldrin (Rdl), a Drosophila homolog of the ionotropic GABA receptor, suppresses wake-promoting circadian pacemaker neurons in adult flies to exert sleep-promoting effects. Similarly, 4,5,6,7-tetrahydroisoxazolo[5,4 c]pyridin-3-ol (THIP), an agonist of the ionotropic GABA receptor, promotes sleep in insects and mammals (Ki, 2019).
Many sleep medications modulate GABAergic transmission. A prominent side effect of anti-epileptic drugs relevant to GABA is causing drowsiness. Conversely, glycine supplements improve sleep quality in a way distinct from traditional hypnotic drugs, minimizing deleterious cognitive problems or addiction. In fact, glycine or D-serine acts as a co-agonist of N-methyl-D-aspartate receptors (NMDARs) and promotes sleep through the sub-type of ionotropic glutamate receptors. Emerging evidence further supports the roles of amino acid transporters and metabolic enzymes in sleep regulation. In particular, it has been demonstrated that starvation induces the expression of metabolic enzymes for serine biosynthesis in Drosophila brains, and elevates free serine levels to suppress sleep via cholinergic signaling (Sonn, 2018). These observations prompted a hypothesis that other amino acids may also display neuro-modulatory effects on sleep behaviors (Ki, 2019).
The molecular and neural machinery of sleep regulation intimately interacts with external (e.g., light, temperature) and internal sleep cues (e.g., sleep pressure, metabolic state) to adjust the sleep architecture in animals. Using a Drosophila genetic model, this study has investigated whether dietary amino acids could affect sleep behaviors, through this investigation SPET was discovered. Previous studies have demonstrated that the wake-promoting circadian pacemaker neurons are crucial for timing sleep onset after lights-off in LD cycles. In addition, WAKE-dependent silencing of clock neurons and its collaborative function with RDL have been suggested as a key mechanism in the circadian control of sleep onset. However, the current evidence indicates that SPET facilitates sleep onset in a manner independent of circadian clocks. It was further elucidated that SPET operates likely via the down-regulation of metabotropic GABA transmission in R2 EB neurons, a neural locus for generating homeostatic sleep drive (Ki, 2019).
Both food availability and nutritional quality substantially affect sleep behaviors in Drosophila. Sucrose contents in food and their gustatory perception dominate over dietary protein to affect daily sleep. Starvation promotes arousal in a manner dependent on the circadian clock genes Clock and cycle as well as neuropeptide F (NPF), which is a fly ortholog of mammalian neuropeptide Y. On the other hand, protein is one of the nutrients that contribute to the postprandial sleep drive in Drosophila and this observation is possibly relevant to SPET. While Leucokinin (Lk) and Lk receptor (Lkr) play important roles in dietary protein-induced postprandial sleepand in starvation-induced arousal, comparable SPET was observed between hypomorphic mutants of Lk or Lkr and their heterozygous controls. Therefore, SPET and its neural basis reveal a sleep-regulatory mechanism distinct from those involved in sleep plasticity relevant to food intake (Ki, 2019).
What will be the molecular basis of SPET? Given the general implication of GABA in sleep promotion, a simple model will be that a molecular sensor expressed in a subset of GABAergic neurons (i.e., LN) directly responds to an increase in threonine levels, activates GABA transmission, and thereby induces sleep. Several lines of evidence, however, favored the other model that dietary threonine actually down-regulates metabotropic GABA transmission in R2 EB neurons, de-represses the neural locus for generating homeostatic sleep drive, and thereby enhances sleep drive. The latter model does not necessarily conflict with sleep-promoting effects of genetic or pharmacological conditions that generally elevate GABA levels or enhance GABAergic transmission since those effects will be the net outcome of activated GABA transmission via various sub-types of GABA receptors expressed in either wake- or sleep-promoting neurons and their (Ki, 2019).
The structural homology among threonine, GABA, and their metabolic derivatives (e.g., alpha-ketobutyrate and gamma-hydroxybutyrate) led to the hypothesis that these relevant chemicals may act as competitive substrates in enzymatic reactions for their overlapping metabolism. Consequently, dietary threonine may limit the total flux of GABA-glutamate-glutamine cycle possibly through substrate competition, decreases the size of available GABA pool, and thereby down-scales GABA transmission for SPET. This accounts for why genetic or pharmacological elevation of GABA levels rather suppresses SPET. Threonine, GABA, and their derivatives may also act as competitive ligands for metabotropic GABA receptors, explaining weak GABA responses in R2 EB neurons of threonine-fed flies. Biochemical and neural evidence supportive of this hypothesis is quite abundant. It has been previously shown that alpha-ketobutyrate, GABA, and the ketone body beta-hydroxybutyrate act as competitive substrates in common enzymatic reactions. Moreover, functional interactions of beta-hydroxybutyrate or gamma-hydroxybutyrate with GABAergic signaling have been well documented. Finally, threonine and GABA derivatives have anti-convulsive effects, which further support their common structural and functional relevance to GABAergic signaling (Ki, 2019).
The removal of the amino group is the initial step for amino acid metabolism, and various transaminases mediate its transfer between amino acids and alpha-keto acids. On the other hand, a group of amino acids (i.e., glutamate, glycine, serine, and threonine) has their own deaminases that can selectively remove the amino group. The presence of these specific deaminases is indicative of active mechanisms that individually fine-tune the baseline levels of these amino acids in metabolism, and possibly in the context of other physiological processes as well. This idea is further supported by the conserved roles of glutamate, glycine, and serine as neurotransmitters or neuromodulators important for brain function, including sleep regulation. In fact, serine, glycine, and threonine constitute a common metabolic pathway, and threonine may contribute indirectly to glycine- or serine-dependent activation of sleep-promoting NMDAR. Nonetheless, this study found that sleep-modulatory effects of dietary glycine were distinct from SPET and thus, it is speculated that threonine may act as an independent neuromodulator, similar to other amino acids with their dedicated deaminases (Ki, 2019).
While several lines of the data support that threonine is likely to be an endogenous sleep driver in fed conditions, it wa recently demonstrated that starvation induces serine biosynthesis in the brain and neuronal serine subsequently suppresses sleep via cholinergic signaling (Sonn, 2018). These two pieces of relevant works establish a compelling model that the metabolic pathway of serine-glycine-threonine functions as a key sleep-regulatory module in response to metabolic sleep cues (e.g., food ingredients and dietary stress). It is further hypothesized that the adaptive control of sleep behaviors by select amino acids and their conserved metabolic pathway suggests an ancestral nature of their sleep regulation. Future studies should address if the serine-glycine-threonine metabolic pathway constitutes the sleep homeostat that can sense and respond to different types of sleep needs. In addition, it will be interesting to determine if this metabolic regulation of sleep is conserved among other animals, including humans (Ki, 2019).
Both the structure and the amount of sleep are important for brain function. Entry into deep, restorative stages of sleep is time dependent; short sleep bouts selectively eliminate these states. Fragmentation-induced cognitive dysfunction is a feature of many common human sleep pathologies. Whether sleep structure is normally regulated independent of the amount of sleep is unknown. This study shows that in Drosophila melanogaster, activation of a subset of serotonergic neurons fragments sleep without major changes in the total amount of sleep, dramatically reducing long episodes that may correspond to deep sleep states. Disruption of sleep structure results in learning deficits that can be rescued by pharmacologically or genetically consolidating sleep. Two reciprocally connected sets of ellipsoid body neurons were identified that form the heart of a serotonin-modulated circuit that controls sleep architecture. Taken together, these findings define a circuit essential for controlling the structure of sleep independent of its amount (Liu, 2019).
This study describes a circuit that regulates of sleep structure without affecting the total amount of sleep. 5HT acts to enhance the response of 5HT7-GAL4+ neurons to basally active excitatory inputs. 5HT-dependent calcium signals are blocked by TTX, while its ability to increase cAMP is not, supporting the existence of these active excitatory inputs to 5HT7-GAL4+ cells. In contrast, VT-GAL4+ cells do not have basally active excitatory inputs. 5HT modulation of the circuit likely occurs primarily via inputs to 5HT7-GAL4+ neurons since the response of VT038828-GAL4 (VT-GAL4)+ neurons is weaker and lower affinity. Whether there are other, perhaps situationally active, inputs to this circuit is currently unknown (Liu, 2019).
Within the ellipsoid body (EB) the circuit is complex. VT-GAL4+ neurons are functionally connected with the 5HT7-GAL4+ group. VT-GAL4+ neurons provide feedback inhibition to a subset of 5HT7-GAL4+ neurons, which enhances fragmentation, likely via output to non-central complex regions. How inhibition of a subset of the 5HT7-GAL4+ cells acts to modulate the behavioral output of the rest of the population is not yet clear, but it is noted that many of the 5HT7-GAL4+ cells are GABAergic. While all the details of the circuit's complex dynamics remain to be discovered, it is clear that this circuit has a profound and selective effect on sleep architecture (Liu, 2019).
The circuit described in this study is modulated by 5HT, a neurochemical known to be important for regulation of behavioral states in many species. While 5HT in mammals is important in a wide variety of contexts, it was controversial for nearly half a century whether it promoted sleep or wakefulness. In Drosophila, 5HT has only been thought to promote sleep. The current data show that upregulation of serotonergic signaling can also induce sleep fragmentation, suggesting that 5HT's role in sleep in flies exhibits a complexity similar to that of its roles in mammals. The genesis of this apparently conserved complexity may be the extensive involvement of 5HT in non-sleep processes. For an animal in the wild, sleep has inherent risks: predation and loss of opportunities for mating or feeding are just a few. Sleep/wake systems in the brain must control arousal state in collaboration with systems that assess competing needs. 5HT, because it is central to so many critical behavioral circuits, is ideally poised to be an integration point for sleep and the general state of the animal. The diverse, circuit-specific, roles in sleep that 5HT exhibits across phyla may be a result of its ubiquity (Liu, 2019).
The role this study has uncovered for 5HT as a regulator of sleep architecture aligns well with this idea. The daily neuronal activity profile reported by Tric-LUC, a calcium sensor that drives luciferase expression in response to neuronal activity, in sleep fragmentation-generating neurons maps to dawn and dusk, when crepuscular organisms such as fruit flies are most active. Fragmentation of sleep at these times would presumably be beneficial since flies would not enter into deep sleep states at times when they should be feeding and mating. Interestingly, the circuit described in this study accomplishes this feat by increasing P(doze), the probability of falling asleep from a wake state, leaving the scaling of P(wake), a parameter associated with dopamine and arousal, free to be modulated by other factors (e.g., danger from predation, appearance of potential mates). The fact that long sleep bouts can be prevented without putting the animal into a hyperaroused state is advantageous, allowing flexible responsiveness to changing conditions. The involvement of P(Doze), a parameter associated with sleep drive, is also congruent with the sleep-promoting role of 5HT in other brain regions (Liu, 2019).
While controlled sleep fragmentation appears to assist in active period behavior, there is also a need for consolidated sleep. In both mammals and Drosophila, sleep has electrophysiologically distinct substrates with progressively higher arousal thresholds that appear in an ordered fashion during a sleep episode. The deeper sleep stages in mammals, REM and slow wave sleep, are strongly associated with maintenance of cognitive function. Fragmentation of sleep, because it truncates sleep episodes before deeper stages are reached, can result in a selective deprivation of deep sleep stages even when total sleep is not changed. In this study, it was demonstrated that decreasing sleep consolidation, without changing the amount of sleep, can disrupt associative learning. These results suggest that in Drosophila, like in mammals, there are time-dependent changes in the depth of sleep that are important for its beneficial effects. This idea is also supported by modeling and analysis of the structure of fly sleep, which indicate that there are time-dependent changes in the probability of sleep-wake transitions consistent with the existence of deep sleep stages that are only accessed in long sleep episodes (Liu, 2019).
Fragmentation of sleep induced by activation of 5HT inputs to the EB also produced an increase in sleep after the activation was terminated. Excess sleep in the recovery day after a perturbation is a hallmark of a homeostatic process. Homeostatic regulation of total sleep time has been previously demonstrated in Drosophila, but the data suggest that there is also homeostatic regulation of sleep quality. In mammals, individual sleep substates have been demonstrated to be homeostatically regulated- selective deprivation of REM or slow wave sleep, in the absence of loss of total sleep time, drive rebound increases of the deprived stage and mechanical sleep fragmentation has been shown to lead to an increase in total sleep. The ability of the EB circuit in Drosophila to selectively modulate sleep structure, without changing the total amount of sleep, has allowed for the first time the selective probing of the cognitive importance of long sleep bouts and deep sleep stages in the fly. The fact that fragmentation triggers rebound sleep implies that these long sleep bouts may also be important for the general health benefits of sleep (Liu, 2019).
The dynamic nature of sleep in many animals suggests distinct stages that serve different functions. Genetic sleep induction methods in animal models provide a powerful way to disambiguate these stages and functions, although behavioral methods alone are insufficient to accurately identify what kind of sleep is being engaged. In Drosophila, activation of the dorsal fan-shaped body (dFB) promotes sleep, but it remains unclear what kind of sleep this is, how the rest of the fly brain is behaving, or if any specific sleep functions are being achieved. This study developed a method to record calcium activity from thousands of neurons across a volume of the fly brain during spontaneous sleep and compared this to dFB-induced sleep. Spontaneous sleep was found to typically transition from an active "wake-like" stage to a less active stage. In contrast, optogenetic activation of the dFB promotes sustained wake-like levels of neural activity even though flies become unresponsive to mechanical stimuli. When flies were probed with salient visual stimuli, it was found that the activity of visually responsive neurons in the central brain was blocked by transient dFB activation, confirming an acute disconnect from the external environment. Prolonged optogenetic dFB activation nevertheless achieved a key sleep function by correcting visual attention defects brought on by sleep deprivation. These results suggest that dFB activation promotes a distinct form of sleep in Drosophila, where brain activity appears similar to wakefulness, but responsiveness to external sensory stimuli is profoundly suppressed (Tainton-Heap, 2020).
The central complex is one of the major brain regions that control sleep in Drosophila. However, the circuitry details of sleep regulation have not been elucidated yet. This study shows a novel sleep-regulating neuronal circuit in the protocerebral bridge (PB) of the central complex. Activation of the PB interneurons labeled by the R59E08-Gal4 and the PB columnar neurons with R52B10-Gal4 promoted sleep and wakefulness, respectively. A targeted GFP reconstitution across synaptic partners (t-GRASP) analysis demonstrated synaptic contact between these two groups of sleep-regulating PB neurons. Furthermore, it was found that activation of a pair of dopaminergic (DA) neurons projecting to the PB (T1 DA neurons) decreased sleep. The wake-promoting T1 DA neurons and the sleep-promoting PB interneurons formed close associations. Dopamine 2-like receptor (Dop2R) knockdown in the sleep-promoting PB interneurons increased sleep. These results indicated that the neuronal circuit in the PB, regulated by dopamine signaling, mediates sleep-wakefulness (Tomita, 2021).
Sleep is controlled by homeostatic mechanisms, which drive sleep after wakefulness, and a circadian clock, which confers the 24-h rhythm of sleep. These processes interact with each other to control the timing of sleep in a daily cycle as well as following sleep deprivation. However, the mechanisms by which they interact are poorly understood. These studies show that hugin (+) neurons, previously identified as neurons that function downstream of the clock to regulate rhythms of locomotor activity, are also targets of the sleep homeostat. Sleep deprivation decreases activity of hugin (+) neurons, likely to suppress circadian-driven activity during recovery sleep, and ablation of hugin (+) neurons promotes sleep increases generated by activation of the homeostatic sleep locus, the dorsal fan-shaped body (dFB). Also, mutations in peptides produced by the hugin (+) locus increase recovery sleep following deprivation. Transsynaptic mapping reveals that hugin (+) neurons feed back onto central clock neurons, which also show decreased activity upon sleep loss, in a Hugin peptide-dependent fashion. It is proposed that hugin (+) neurons integrate circadian and sleep signals to modulate circadian circuitry and regulate the timing of sleep (Schwarz, 2021).
During sleep, the brain undergoes dynamic and structural changes. In Drosophila, such changes have been observed in the central complex, a brain area important for sleep control and navigation. The connectivity of the central complex raises the question about how navigation, and specifically the head direction system, can operate in the face of sleep related plasticity. To address this question, this study develop a model that integrates sleep homeostasis and head direction. By introducing plasticity, the head direction system was shown to function in a stable way by balancing plasticity in connected circuits that encode sleep pressure. With increasing sleep pressure, the head direction system nevertheless becomes unstable and a sleep phase with a different plasticity mechanism is introduced to reset network connectivity. The proposed integration of sleep homeostasis and head direction circuits captures features of their neural dynamics observed in flies and mice (Flores-Valle, 2021).
Two forms of associative learning-delay conditioning and trace conditioning-have been widely investigated in humans and higher-order mammals. In delay conditioning, an unconditioned stimulus (for example, an electric shock) is introduced in the final moments of a conditioned stimulus (for example, a tone), with both ending at the same time. In trace conditioning, a 'trace' interval separates the conditioned stimulus and the unconditioned stimulus. Trace conditioning therefore relies on maintaining a neural representation of the conditioned stimulus after its termination (hence making distraction possible), to learn the conditioned stimulus-unconditioned stimulus contingency; this makes it more cognitively demanding than delay conditioning. By combining virtual-reality behaviour with neurogenetic manipulations and in vivo two-photon brain imaging, this study shows that visual trace conditioning and delay conditioning in Drosophila mobilize R2 and R4m ring neurons in the ellipsoid body. In trace conditioning, calcium transients during the trace interval show increased oscillations and slower declines over repeated training, and both of these effects are sensitive to distractions. Dopaminergic activity accompanies signal persistence in ring neurons, and this is decreased by distractions solely during trace conditioning. Finally, dopamine D1-like and D2-like receptor signalling in ring neurons have different roles in delay and trace conditioning; dopamine D1-like receptor 1 mediates both forms of conditioning, whereas the dopamine D2-like receptor is involved exclusively in sustaining ring neuron activity during the trace interval of trace conditioning (Grover, 2022).
Homeostatic and circadian processes collaborate to appropriately time and consolidate sleep and wake. To understand how these processes are integrated, brief sleep deprivation was scheduled at different times of day in Drosophila, and elevated morning rebound was compared to evening. These effects depend on discrete morning and evening clock neurons, independent of their roles in circadian locomotor activity. In the R5 ellipsoid body sleep homeostat, this study identified elevated morning expression of activity dependent and presynaptic gene expression as well as the presynaptic protein BRUCHPILOT consistent with regulation by clock circuits. These neurons also display elevated calcium levels in response to sleep loss in the morning, but not the evening consistent with the observed time-dependent sleep rebound. These studies reveal the circuit and molecular mechanisms by which discrete circadian clock neurons program a homeostatic sleep center (Andreani, 2022).
Sleep is characterized by quiescence, increased arousal thresholds, changes in neuronal activity, and circadian and homeostatic regulation. Flies display each of these hallmarks and have simple, well-characterized circadian and sleep neural networks. About 150 central pacemaker neurons that express molecular clocks. Of these, four small ventral lateral neurons (sLNvs) (per hemisphere) that express pigment dispersing factor (PDF) are necessary for driving morning activity in anticipation of lights on and exhibit peak levels of calcium around dawn (~ZT0). The dorsal lateral neurons (LNds) and a 5th PDF- sLNv are necessary for evening anticipation of lights off and show a corresponding evening calcium peak (ZT8-ZT10). The posterior DN1 (DN1ps) consist of glutamate-positive (Glu+) subsets necessary for morning anticipation and Glu- necessary for evening anticipation under low light conditions. Lateral posterior neurons (LPN) are not necessary for anticipation but are uniquely sensitive to temperature cycling. Specific pacemaker subsets have been linked to wake promotion [PDF+ large LNv, diuretic hormone 31 (DH31+) DN1ps] and sleep promotion (Glu+ DN1ps, Allostatin A+ LPNs), independently of their clock functions. How these neurons regulate homeostatic sleep drive itself remains unsettled (Andreani, 2022).
Timed signaling from these clock neurons converges on the neuropil of the ellipsoid body (EB). The sLNvs and LNds appear to communicate to R5 EB neurons through an intermediate set of dopaminergic PPM3 neurons based largely on correlated calcium oscillations. The anterior projecting subset of DN1ps provide sleep promoting input to other EB neurons (R2/R4M) via tubercular bulbar (TuBu) interneurons. Activation of a subset of these TuBu neurons synchronizes the activity of the R5 neurons which is important for sleep maintenance. Critically, the R5 neurons are at the core of sleep homeostasis in Drosophila. R5 neuronal activity is both necessary and sufficient for sleep rebound. Extended sleep deprivation (12-24 hr) elevates calcium, the critical presynaptic protein BRUCHPILOT (BRP), and action potential firing rates in R5 neurons. The changes in BRP in this region not only reflect increased sleep drive following sleep deprivation (SD) but also knockdown (KD) of brp in R5 decreases rebound suggesting it functions directly in regulating sleep homeostasis. R5 neurons stimulate downstream neurons in the dorsal fan-shaped body (dFB), which are sufficient to produce sleep. Yet how the activity of key clock neurons are integrated with signals from the R5 homeostat to determine sleep drive remains unclear (Andreani, 2022).
This study dissect the link between the circadian and homeostatic drives by examining which clock neural circuits regulate sleep rebound at different times of day in Drosophila. Akin to the forced desynchrony protocols, wakefulness was enforced at different times of day and sleep rebound was assessed. Flies were exposed to 7 hr cycles of sleep deprivation and recovery, enabling assessment of homeostasis at every hour of the day. It was found that rebound is suppressed in the evening in a Clk-dependent manner. Time-dependent rebound was demonstrated to be is mediated by specific subsets of pacemaker neurons, independently of their effects on locomotor activity. Moreover, homeostatic R5 EB neurons integrate circadian timing and homeostatic drive; it was demonstrated that activity dependent and presynaptic gene expression, BRP expression, neuronal output, and wake sensitive calcium levels are all elevated in the morning compared to the evening, providing an underlying mechanism for clock programming of time-of-day dependent homeostasis (Andreani, 2022).
This study describes the neural circuit and molecular mechanisms by which discrete populations of the circadian clock network program the R5 sleep homeostat to control the homeostatic response to sleep loss. A novel protocol was developed to administer brief duration SD and robustly measure homeostatic rebound sleep. Using this strategy, it was demonstrated that homeostatic rebound is significantly higher in the morning than in the evening. Distinct subsets were identified of the circadian clock network and their downstream neural targets that mediate the enhancement and suppression of morning and evening rebound respectively. Using unbiased transcriptomics very little gene expression was observed that was significantly altered in response to the 2.5 hr sleep deprivation. On the other hand,elevated expression of activity-dependent and presynaptic genes were identified in the morning, independent of sleep deprivation. Consistent with this finding, elevated levels of the presynaptic protein BRP were observed that was absent in the absence of Clk. These baseline changes are accompanied by an elevated calcium response to sleep deprivation in the morning mirroring the enhanced behavioral rebound in the morning. Taken together, these data support the model of a circadian regulated homeostat that turns the homeostat up late at night to sustain sleep and down late in the day to sustain wake (Andreani, 2022).
These studies suggest that homeostatic drive in the R5 neurons is stored post-transcriptionally. As part of these studies, a novel protocol was developed using minimal amounts of SD which could be useful for minimizing mechanical stress effects and isolating underlying molecular processes crucial for sleep homeostasis. Six to 24 hr of SD in Drosophila is commonly used despite the potential stressful or even lethal effects. This study demonstrates that shorter 2.5 hr deprivations not only induce a robust rebound sleep response, but also the percent of sleep lost recovered at ZT0 is close to 100% versus 14-35% seen in 12 hr SD protocols. Using this shorter SD, it was found that many effects observed in R5 neurons with 12 hr SD (e.g. increased BRP and upregulation of Nmdar subunits) are no longer observed with shorter SD, even though the necessity of R5 neurons for rebound is retained after 2.5 hr SD. Previously, translating ribosome affinity purification (TRAP) was used to show upregulation of nmdar subunits following 12 hr SD.FACS and TRAP are distinct methodologies for targeted collection of RNA for sequencing and can yield unique gene lists. One possibility is that upregulation of nmdar subunits is occurring locally in neuronal processes, which are often lost during FACS, and/or is at the level of translation initiation or elongation. Nonetheless, in agreement with previous work, this study observed SD-induced increases in calcium correlated with behavioral rebound in the morning, suggesting that this process is a core feature of the cellular homeostatic response (Andreani, 2022).
Using genetically targeted 'loss-of-function' manipulations, this study has defined small subsets of circadian clock neurons and downstream circuits that are necessary for intact clock modulation of sleep homeostasis. The use of intersectional approaches enabled highly resolved targeting not possible with traditional lesioning experiments in the SCN. Collectively these studies defined a potential Glu+ DN1p-TuBu-R4m circuit important for enhancing morning rebound as well as a discrete group of LNds important for suppressing evening rebound. Importantly, most of these effects on sleep rebound are evident in the absence of substantial changes in baseline activity, despite other studies indicating their necessity for normal circadian behavior. Of note, the proposed roles of the DN1p and LNd clock neurons are sleep and wake promotion consistent with the current findings after sleep deprivation. It is hypothesized that by using chronic silencing methods, baseline effects may not be evident due to compensatory changes but that these effects are only revealed when the system is challenged by sleep deprivation. Similar genetic strategies in mammals may be useful in uncovering which SCN neurons are driving circadian regulation of sleep homeostasis given the comparable suppression of sleep rebound in the evening in humans. Nonetheless, the finding of sleep homeostasis phenotypes in the absence of significant baseline effects suggests that a major role of these clock neuron subsets may be to manage homeostatic responses (Andreani, 2022).
These studies suggest that circadian and homeostatic processes do not compete for influence on a downstream neural target but that the circadian clock programs the homeostat itself. Using an unbiased transcriptomic approach, this study discovered time-dependent expression of activity dependent and presynaptic genes, consistent with previous data that the R5 neurons exhibit time-dependent activity. Significant upregulation was observed of several genes involved in synaptic transmission (Syx1a, Rim, nSyb, unc-104, Srpk79D, para, CG5890) evincing a permissive active state for R5 neurons in the morning. This is accompanied by elevated levels of the key presynaptic protein BRP in the morning compared to evening. It is notable that elevated BRP in the morning is the opposite of what would be expected based on a sleep-dependent reduction in BRP proposed by the synaptic homeostasis hypothesis, suggesting a sleep-wake independent mechanism. Previous studies have shown that modulation of BRP levels in the R5 are important for its sleep function, suggesting that changes in BRP levels impact R5 function. It is hypothesized that these baseline transcriptomic changes underlie the differential R5 sensitivity to sleep deprivation is evident as calcium increases in the morning and not the evening. Indeed, trancriptomic and proteomic studies of the mouse forebrain across time and after sleep deprivation are consistent with the model that the circadian clock programs the transcriptome while homeostatic process function post-trranscriptionally , paralleling what this study has found for R5. It will be of great interest to understand the circuit and molecular mechanisms by which circadian clocks regulate the R5 neuronal calcium and synaptic properties and whether similar circuit architectures underlie daily mammalian sleep-wake (Andreani, 2022).
Sensory perception modulates aging, yet little is known about how. An understanding of the neuronal mechanisms through which animals orchestrate biological responses to relevant sensory inputs would provide insight into the control systems that may be important for modulating lifespan. This study provides new awareness into how the perception of dead conspecifics, or death perception, which elicits behavioral and physiological effects in many different species, affects lifespan in the fruit fly, Drosophila melanogaster. Previous work demonstrated that cohousing Drosophila with dead conspecifics decreases fat stores, reduces starvation resistance, and accelerates aging in a manner that requires both sight and the serotonin receptor 5-HT2A. This study demonstrated that a discrete, 5-HT2A-expressing neural population in the ellipsoid body (EB) of the Drosophila central complex, identified as R2/R4 neurons, acts as a rheostat and plays an important role in transducing sensory information about the presence of dead individuals to modulate lifespan. Expression of the insulin-responsive transcription factor foxo in R2/R4 neurons and insulin-like peptides dilp3 and dilp5, but not dilp2, are required, with the latter likely altered in median neurosecretory cells (MNCs) after R2/R4 neuronal activation. These data generate new insights into the neural underpinnings of how perceptive events may impact aging and physiology across taxa (Gendron, 2023).
The ellipsoid body (EB) is a major structure of the central complex of the Drosophila melanogaster brain. Twenty-two subtypes of EB ring neurons have been identified based on anatomic and morphologic characteristics by light-level microscopy and EM connectomics. A few studies have associated ring neurons with the regulation of sleep homeostasis and structure. However, cell type-specific and population interactions in the regulation of sleep remain unclear. Using an unbiased thermogenetic screen of EB drivers using female flies, the following was found the: (1) multiple ring neurons are involved in the modulation of amount of sleep and structure in a synergistic manner; (2) analysis of data for &deltaPdoze/&Delta:Pwake using a mixed Gaussian model detected 5 clusters of GAL4 drivers which had similar effects on sleep pressure and/or depth: lines driving arousal contained R4m neurons, whereas lines that increased sleep pressure had R3m cells; (3) a GLM analysis correlating ring cell subtype and activity-dependent changes in sleep parameters across all lines identified several cell types significantly associated with specific sleep effects: R3p was daytime sleep-promoting, and R4m was nighttime wake-promoting; and (4) R3d cells present in 5HT7-GAL4 and in GAL4 lines, which exclusively affect sleep structure, were found to contribute to fragmentation of sleep during both day and night. Thus, multiple subtypes of ring neurons distinctively control sleep amount and/or structure. The unique highly interconnected structure of the EB suggests a local-network model worth future investigation; understanding EB subtype interactions may provide insight how sleep circuits in general are structured (Yan, 2023).
Sleep is essential, but animals may forgo sleep to engage in other critical behaviors, such as feeding and reproduction. Previous studies have shown that female flies exhibit decreased sleep after mating, but understanding of the process is limited. This study reports that postmating nighttime sleep loss is modulated by diet and sleep deprivation, demonstrating a complex interaction among sleep, reproduction, and diet. It was also found that female-specific pC1 neurons and sleep-promoting dorsal fan-shaped body (dFB) neurons are required for postmating sleep plasticity. Activating pC1 neurons leads to sleep suppression on standard fly culture media but has little sleep effect on sucrose-only food. Published connectome data suggest indirect, inhibitory connections among pC1 subtypes. Using calcium imaging, it was shown that activating the pC1e subtype inhibits dFB neurons. It is proposed that pC1 and dFB neurons integrate the mating status, food context, and sleep drive to modulate postmating sleep plasticity (Duhart, 2023).
The needs fulfilled by sleep are unknown, though the effects of insufficient sleep are manifold. To better understand how the need to sleep is sensed and discharged, much effort has gone into identifying the neural circuits involved in regulating arousal, especially those that promote sleep. In prevailing models, the dorsal fan-shaped body (dFB) plays a central role in this process in the fly brain. This study manipulated various properties of the dFB including its electrical activity, synaptic output, and endogenous gene expression. In each of these experimental contexts it was not possible to identify any effect on sleep that could be unambiguously mapped to the dFB. Furthermore, evidence was found that sleep phenotypes previously attributed to the dFB were caused by genetic manipulations that inadvertently targeted the ventral nerve cord. Expression was examined of two genes whose purported effects have been attributed to functions within a specific subpopulation of dFB neurons. In both cases little to no expression expression was found in the expected cells. Collectively, these results cast doubt on the prevailing hypothesis that the dFB plays a central role in promoting sleep (De, 2023).
Sleep is a complex and plastic behavior regulated by multiple brain regions and influenced by numerous internal and external stimuli. Thus, to fully uncover the function(s) of sleep, cellular resolution of sleep-regulating neurons needs to be achieved. Doing so will help to unequivocally assign a role or function to a given neuron or group of neurons in sleep behavior. In the Drosophila brain, neurons projecting to the dorsal fan-shaped body (dFB) have emerged as a key sleep-regulating area. To dissect the contribution of individual dFB neurons to sleep, this study undertook an intersectional Split-GAL4 genetic screen focusing on cells contained within the 23E10-GAL4 driver, the most widely used tool to manipulate dFB neurons. This study demonstrated that 23E10-GAL4 expresses in neurons outside the dFB and in the fly equivalent of the spinal cord, the ventral nerve cord (VNC). Furthermore, it was shown that 2 VNC cholinergic neurons strongly contribute to the sleep-promoting capacity of the 23E10-GAL4 driver under baseline conditions. However, in contrast to other 23E10-GAL4 neurons, silencing these VNC cells does not block sleep homeostasis. Thus, these data demonstrate that the 23E10-GAL4 driver contains at least 2 different types of sleep-regulating neurons controlling distinct aspects of sleep behavior (Jones, 2023).
Memory formation and sleep regulation are critical for brain functions in animals from invertebrates to humans. Neuropeptides play a pivotal role in regulating physiological behaviors, including memory formation and sleep. However, the detailed mechanisms by which neuropeptides regulate these physiological behaviors remains unclear. This study report sthat neuropeptide diuretic hormone 31 (DH31) positively regulates memory formation and sleep in Drosophila melanogaster. The expression of DH31 in the dorsal and ventral fan-shaped body (dFB and vFB) neurons of the central complex and ventral lateral clock neurons (LNvs) in the brain was responsive to sleep regulation. In addition, the expression of membrane-tethered DH31 in dFB neurons rescued sleep defects in Dh31 mutants, suggesting that DH31 secreted from dFB, vFB, and LNvs acts on the DH31 receptor in the dFB to regulate sleep partly in an autoregulatory feedback loop. Moreover, the expression of DH31 in octopaminergic neurons, but not in the dFB neurons, is involved in forming intermediate-term memory. These results suggest that DH31 regulates memory formation and sleep through distinct neural pathways (Lyu, 2023).
Sleep is required to maintain physiological functions and is widely conserved across species. To understand the sleep-regulatory mechanisms This study reports a novel sleep-regulatory pathway that promotes arousal. This study first focused on FB interneurons and found that cholinergic FB interneurons promoted arousal. The arousal-promoting effect of FB interneurons was confirmed by using more specific drivers. These drivers label FB interneurons which receive input from P-FN neurons (that project from the protocerebral bridge to the ventral FB and the NO) and send output to dFB neurons. There should be other FB interneurons that do not have a connection with P-FN neurons or dFB neurons. It means that FB interneurons labeled by two split drivers are only a part of FB interneurons. Therefore we considered that the weaker effects were due to the smaller number of neurons labeled by split-Gal4 lines than NP2320, not the effect of neurons other than FB interneurons. No aclear sleep rebound was observed after neuronal activation. In the previous study, R52B10-Gal4 was used which is reported to drive sleep rebound in the female fly. A clear sleep rebound was observed in female flies but not in male flies. These results indicated that there is a sex difference in the regulation of sleep rebound, at least, in R52B10 neurons. The current study used only male flies and this could be one of the reasons why no clear sleep rebound was observed. It was next asked about the relationship between FB interneurons and known sleep-regulatory circuits. GRASP and Ca2+ experiments were performed, and FB interneurons and R52B10 neurons which label the output neurons of the PB-FB pathway were shown to be anatomically and functionally connected . Although the possibility of other pathways downstream to R52B10 cannot be excluded, this study demonstrated clearly that FB interneurons are one of the downstream to R52B10. Further study will show the impact size of the connection between them in sleep regulation. To investigate the postsynaptic partners of FB interneurons, a GRASP experiment was conducted. FB interneurons and dFB neurons labeled by R23E10 were found to form close associations. Besides, according to the connectome paper and connectome dataset, vDeltaB, C, D, and hDeltaC receive input from P-FN neurons and send output to FB tangential neurons which arborize in layers 6 and 7. This information also supported the idea that R52B10 neurons including P-FN neurons, FB interneurons like vDeltaB, C, D, and hDeltaC neurons, and dFB neurons that are consisted of FB tangential neurons which arborize in layers 6 and 7 form a neuronal circuit. Future study will clarify their functional connection and the role of this circuit in sleep regulation. Furthermore, a previous study showed that neurons that project to the ventral FB (vFB neurons) promote sleep and mediate consolidation of long-term memory. Since axon terminals and dendrites of FB interneurons arborize in both the dorsal and ventral FB, there would be interactions between dFB and vFB neurons via FB interneurons. Further research will clarify the functional relationship between these neurons (Kato, 2022).
According to previous reports, FB interneurons regulate optomotor behavior and express tachykinin, a neuropeptide that regulates aggression. Additionally, T1 dopamine neurons, which are upstream of R52B10 neurons, regulate aggression as well. Besides, P2 neurons, which include FB interneurons, regulate chronic isolation evoked sleep loss. Moreover, courtship-regulator P1 neurons activate T1 neurons and modulate sleep/courtship balance based on the nutritional status. Taking all the information mentioned above into account, it is considerd that arousal signals related to aggression, courtship, nutrition, and vision converge into the PB-FB pathway including FB interneurons to regulate arousal. Further studies should clarify the role of these arousal signals on the PB-FB pathway and FB interneurons in sleep regulation (Kato, 2022).
In conclusion, the results provided possible sleep-regulatory neurons that may connect with the PB-FB pathway and dFB neurons. It is hypothesized that arousal signals are sent from the PB-FB pathway to FB interneurons, inhibit dFB neurons via inhibitory signals, and regulate sleep (Kato, 2022).
The insect central complex (CX) has been implicated in a wide range of behaviours. Recent experimental evidence from Drosophila and the cockroach (Blaberus discoidalis) has demonstrated the existence of neural activity corresponding to the animal's orientation within a virtual arena (a neural 'compass'), and this provides an insight into one component of the CX structure. There are two key features of the compass activity: an offset between the angle represented by the compass and the true angular position of visual features in the arena, and the remapping of the 270 degrees visual arena onto an entire circle of neurons in the compass. This study presents a computational model which can reproduce this experimental evidence in detail, and predicts the computational mechanisms that underlie the data. It is predicted that both the offset and remapping of the fly's orientation onto the neural compass can be explained by plasticity in the synaptic weights between segments of the visual field and the neurons representing orientation. Furthermore, it is predicted that this learning is reliant on the existence of neural pathways that detect rotational motion across the whole visual field and uses this rotation signal to drive the rotation of activity in a neural ring attractor. This model also reproduces the 'transitioning' between visual landmarks seen when rotationally symmetric landmarks are presented. This model can provide the basis for further investigation into the role of the central complex, which promises to be a key structure for understanding insect behaviour, as well as suggesting approaches towards creating fully autonomous robotic agents (Cope, 2017).
Many animals maintain an internal representation of their heading as they move through their surroundings. Such a compass representation was recently discovered in a neural population in the Drosophila melanogaster central complex (see Anatomy suggests a potential circuit mechanism to update a compass representation), a brain region implicated in spatial navigation. This study used two-photon calcium imaging and electrophysiology in head-fixed walking flies to identify a different neural population that conjunctively encodes heading and angular velocity, and is excited selectively by turns in either the clockwise or counterclockwise direction. These mirror-symmetric turn responses combine with the neurons' connectivity to the compass neurons to create an elegant mechanism for updating the fly's heading representation when the animal turns in darkness. This mechanism, which employs recurrent loops with an angular shift, bears a resemblance to those proposed in theoretical models for rodent head direction cells. These results provide a striking example of structure matching function for a broadly relevant computation (Turner-Evans, 2017).
A stable internal representation of heading is fundamental to successful navigation. Neurons that maintain such a representation in darkness have been reported across various species. Several computational models have been proposed to explain how a population representation of heading might be updated using angular velocity signals from different neural populations, but identifying connections between neurons that carry and integrate these disparate signals has been challenging in mammals. This study took advantage of the small size, strong topography and well-described anatomy and cell types of the fly central complex to identify a candidate neuron population, P-ENs, which carry angular velocity signals. Cell-type-specific genetic tools were used to perform electrophysiological recordings from single P-EN neurons and two-photon calcium imaging from entire populations of both P-ENs and the previously described 'compass neurons' (E-PGs) in head-fixed walking flies to demonstrate how these neurons together create an elegant circuit mechanism to update a heading representation when the fly turns in darkness. The circuit motif underlying this mechanism shares some characteristics with past conceptual models of head-direction cell function (Turner-Evans, 2017).
The rate model that was implemented in this study was able to capture the essence of the observed network activity, reproducing physiological activity in response to an input that is specific to one side of the protocerebral bridge, but uniform otherwise. This suggests a level of control over moving the activity bump that is quite simple to implement in neural circuitry. In addition, the model is agnostic to the type of input that is needed to rotate the bump. It does, however, require inputs that are activated when the fly turns, with a strength proportional to the strength of the turn, and that such inputs preferentially innervate one hemisphere to create a mirror-symmetry in the system. This description anatomically matches at least one known cell type: PBG1/2-9.b-SPSi.s (Wolff, 2015). The model also requires inhibition to maintain a stationary bump and linear velocity integration. The widely arborizing and glutamatergic PB18.s-GxΔ7Gy.b neurons may provide such large-scale inhibition onto the P-EN neurons (Turner-Evans, 2017).
Some discrepancies remain between the proposed model and the experimental evidence presented in this study. The model assumes only one P-EN neuron per protocerebral bridge glomerulus (Wolff, 2015), which puts a strong constraint on the angular velocity integration properties of the circuit. In particular, although the circuit displays linear velocity integration within the typical range of angular velocities, the activity bump gets 'stuck' at individual P-EN neurons for small turns. That is, when the fly turns slowly, the corresponding small inputs to the circuit do not trigger bump movements. No such bump dynamics were observed in the imaging experiments, indicating that other, unexplored factors may help smooth bump movement in the actual circuit. Noise in the circuit, potential gap junctions and dendro-dendritic connections within and between E-PG and P-EN neurons, as well as the activity of other cell types in the circuit, such as the PBG1-8.s-EBt.b-D/Vgall.b neurons (Wolff, 2015), may all play a role in smoothing bump movement. These factors may also contribute to differences in bump shape and width between the model and experimental data. Further, the model suggests that E-PG activity is directly passed to the P-EN neurons in the protocerebral bridge, possibly with some anatomical offset and modulation through inhibition. Indeed, almost coincident bumps of activity were observed in the bridge for the two cell types. However, while functional connectivity showed a clear connection from the P-EN neurons to the E-PG neurons, the connectivity, electrophysiology, and imaging results suggested that the E-PG to P-EN connection might be more indirect and also recruit inhibition. In the functional connectivity experiments, very strong activation of the E-PG population reliably excited the P-EN neurons, but weaker excitation evoked a variety of responses. Electrophysiological recordings also revealed an unanticipated complexity in the tuning of the P-ENs' membrane potential. Membrane potential tuning curves generally showed a peak at the same heading as the spike rate tuning curves, but also a pronounced trough about 150° distant from that peak. That trough, likely a result of inhibition in the circuit, was not always evident in the spike rate tuning. Finally, in two-color imaging, offsets were observed of up to one glomerulus between the E-PG and P-EN activity on the ipsilateral side of the bridge, and unexpected P-EN activity on the contralateral side, also offset from the E-PG activity. These results were consistent for both color indicator pairings, as well as in experiments involving a second driver line, suggesting that the effects are not merely an artifact of indicator kinetics or co-expression in another population of neurons. It was noted that during slow rotations, when P-EN activity is low and the E-PG bump is weak, these offsets decreased and, depending on the driver line used, also differed between the ipsi- and contralateral side during a turn. These ae taken as indications that the connectivity between the E-PG and P-EN neurons in the protocerebral bridge may be partly indirect. Future studies will address how excitatory and inhibitory connectivity between these populations and others shape the circuit's compass function (Turner-Evans, 2017).
Still uncertain is whether an activity bump can be independently sustained in the P-EN and E-PG populations, or in the left vs. right P-EN subpopulations. The connections from P-ENs to E-PGs may be the substrate that sustains the maintenance of E-PG bump position in the ellipsoid body in the absence of both visual and self-motion cues (Seelig, 2015), as in the current model. The significant reduction seen in E-PG bump amplitude and PVA (population vector average) strength when synaptic transmission from P-ENs was blocked is supportive of such an idea. Whether E-PG input is similarly essential to the maintenance of P-EN bump strength is less clear, but P-EN heading tuning hints at a dependence on E-PG input. On the other hand, appropriate local connections between nearby neurons either in the ellipsoid body or in the protocerebral bridge may allow bumps of activity to be independently sustained in the E-PG neurons. Signs of such internal connections come, for example, from evidence of presynaptic specializations of E-PGs in the ellipsoid body. Bump persistence could also be achieved through long time-scale cellular biophysics. Future experiments and electron microscopy-based circuit reconstruction efforts should provide stronger constraints on the space of possible models, and clarify the functional and behavioral relevance of the actual circuit structure (Turner-Evans, 2017).
For a circuit mechanism in which phase relationships and conjunctive coding are important, calcium imaging may seem an unreliable arbiter of truth. Somatic single cell recordings, on the other hand, can be hard to interpret given the intricate projection patterns of fly neurons and the compartmentalization of information processing that this can produce. However, the results from calcium imaging and electrophysiology experiments in P-EN neurons were found to be in broad agreement. The electrical signature of P-EN responses to angular and forward velocity mirrored seem with calcium imaging in the noduli. The measured width of a single P-EN neuron's receptive field (~60°) was lower than that observed with calcium imaging (~110°), but this may arise from the slow decay kinetics of calcium indicators. One inconsistency between results, however, related to the imaging of neural activity in the protocerebral bridge. Based on imaging in the noduli and electrophysiology, it was expected that turns in one direction would evoke a steady decrease in activity on the other (contralateral) side of the bridge with increasing rotational velocity. Instead, imaging in the bridge showed a mild increase in activity at higher velocities, albeit while preserving the expected asymmetry between the ipsi- and contralateral side. It is hypothesized that this calcium signal might represent synaptic inputs to the P-ENs more than their spiking activity (Turner-Evans, 2017).
Blocking P-EN output using shiTS had two effects on the E-PG bump: Its amplitude was reduced and its position sometimes changed dramatically during small turns (visible as an increase in variability and low R2 for the correlation of changes in heading versus PVA. The bump amplitude decrease in shiTS flies at high temperature can be readily explained by the reduction in synaptic input to the E-PGs -- indeed, in the firing rate model P-EN input is essential to the maintenance of the E-PG bump. Several factors may explain why the E-PG bump did not completely disappear during this manipulation. First, cell-intrinsic properties of the E-PG neurons may contribute to the persistence of activity in those neurons even in the absence of external input. Second, the shiTS block may have been incomplete, meaning that there was sufficient P-EN drive even at high temperatures to keep the E-PG bump alive. Third, it the possibility of gap junctions between P-EN and E-PG neurons, which the experiments would not block, cannot be ruled out. Finally, other neuron types may also provide synaptic input to the E-PG population in the ellipsoid body. Some of these possibilities have been suggested in a recent study that used the anatomy of protocerebral bridge neurons to create a spiking model that generates ring attractor dynamics (Kakaria, 2017; Turner-Evans, 2017 and references therein).
Further, the conceptual and firing rate models would imply that if the P-EN to E-PG connections were entirely removed, E-PG activity would be unable to follow the fly's turns. However, the E-PG activity does still track the fly's turns at high temperature, when the P-EN synaptic output should be blocked. This may, once again, be the result of an incomplete block. It is speculated that one reason that the E-PG bump makes large movements across the ellipsoid body even during small turns is that a reduction in bump amplitude destabilizes the compass representation. Thus, fluctuations in the activity of the E-PGs elsewhere in the ellipsoid body may exert a greater influence on the movements of the bump than under normal conditions, when activity in distant E-PGs is likely to be suppressed. Yet another possibility that could explain the bump's movements is raised by a parallel study, which provides further evidence for P-ENs serving a role in angular integration and describes a second subtype of P-EN neurons that likely also influences the position of the E-PG bump (Green, 2017; Turner-Evans, 2017 and references therein).
The coordinated activity of the E-PG population and its control by the P-EN population when the fly turns are strongly evocative of a compass. The animal could, in principle, use such a neural compass to tether its actions to local landmarks or other sensory cues during navigation, and maintain its bearings in the temporary absence of such cues . Consistent with this idea, the PVA computed with E-PG population activity tracks the fly's heading quite accurately even in darkness. However, it is not yet known how downstream circuits read out E-PG population activity. Thus, although the PVA metric is a useful representation of E-PG compass-like activity, whether downstream circuits perform similar computations to extract the fly's heading is unclear. Further, the PVA was derived by combining the strength and angular position of activity in the E-PG population. Although both these features of E-PG activity likely influence downstream neurons, their specific influence on such neurons will depend on the precise connectivity of the circuit, something that a combination of functional connectivity studies and electron microscopy may reveal in time. Although there is considerable evidence across insects suggesting that CX neurons influence action initiation and turning movements , the connection of E-PG and P-EN neurons to the largely unidentified class of CX neurons that drive behavioral decisions is as yet unclear (Turner-Evans, 2017).
This study has focused on the effects of self-motion cues on bump movement, to which end most of the experiments were conducted with flies walking in the dark. However, E-PG activity is strongly influenced by visual cues, as evidenced by the fact that cue jumps can reset the bump position (Seelig, 2015; Kim, 2017). The angular velocity representation of P-EN neurons, by contrast, seemed unaffected by the presence of closed loop visual feedback. Thus, while a circuit mechanism was suggested for updating heading representation in the dark using self-motion signals, it is anticipated that strong sensory inputs, including those from visual cues, control updating in other circumstances. For example, it was previously observed that the ring neurons retinotopically respond to visual cues (Seelig, 2013). As the putative ring neuron axons arborize in the ellipsoid body along with the E-PG dendrites, it may be possible for them to convey visual information to the E-PG neurons, influencing the movement of the bump of activity. Further, it was suggested above that the E-PG to P-EN connection in the protocerebral bridge may be indirect and recruit sources of inhibition. There exist a few classes of bridge interneurons which may serve as intermediaries in E-PG to P-EN connections. Future studies should help clarify their role in the compass network (Turner-Evans, 2017).
Fly E-PG neurons share several characteristics with mammalian head direction cells. Both head direction cells and E-PG neurons maintain one stable bump of activity and both track the animal's heading in darkness, a feature that is well described by appropriately wired ring attractor models. Rodents that are deprived of proprioceptive and motor efference signals, as in passive transport experiments, show impaired heading representation. To update their heading in darkness, head direction cells in rodent thalamic nuclei and post-subiculum are thought to depend on angular velocity input from the vestibular system, mediated by the dorsal tegmental nucleus. Although 75% of neurons in this region were found to encode angular head velocity, only about a third of those did so in the mirror-symmetric, turn-direction-selective fashion of Drosophila P-EN neurons that were describe in this study (Turner-Evans, 2017).
Individual P-EN neurons were deterministic in their left-right mirror-symmetric rotation tuning, but diverse in the range of rotational velocities that their tuning curves spanned. Indeed, the measured bandwidth of individual P-ENs ranged anywhere between 30° and 270°/s. This diversity may reflect the diversity of tuning of the three to four P-EN neurons that innervate each protocerebral bridge glomerulus (estimated from cell body counts. Such a range of sensitivities and bandwidths would permit a more precise tracking of the flies' turns across a wide range of rotational velocities (Turner-Evans, 2017).
The origins of angular velocity responses in P-ENs are as yet unclear, but these responses show a latency relative to the fly's turning movements that are estimated to be ~150 ms, suggesting that they arise from proprioception rather than motor efference. Anatomically, both the two halves of the protocerebral bridge as well as the two noduli are mirror-symmetric structures innervated by a number of neuron types in a lateralized manner, making them likely candidates for receiving such rotation-tuned input. In the cockroach, neurons encoding angular as well as forward velocity have been recorded in the fan-shaped body, a substructure of the central complex that is evolutionarily conserved in flies. Of note, only one of the forty turn responsive neurons in the latter study showed bidirectional modulation, with excitation for turns in the preferred direction and inhibition for turns the other way, a hallmark of the P-EN neurons. These studies, which relied on extracellular recordings and did not identify cell types, found that changes in spike rate regularly preceded locomotor changes instead of tracking them as was found for the fly P-ENs. If it is assumed that neurons of the type recorded in the cockroach also exist in the fly, it is not yet clear whether the P-EN/E-PG compass network that is described in this article exploit advance information about expected changes in angular velocity (Turner-Evans, 2017).
A striking aspect of the fly compass system is its structural symmetry. Mirror symmetry is a prominent feature of the anatomical layout of the protocerebral bridge. The developmental origins of the anatomical positions of central complex neurons have been the focus of numerous studies. However, although the two sides of the protocerebral bridge and the noduli are tuned to rotations in opposite directions, maintaining symmetry at the large scale, the activity of the E-PGs and P-ENs at the scale of bridge glomeruli breaks this symmetry. During a turn, bumps of activity propagate through the left and right sides of the bridge in parallel, in a manner reminiscent of windshield wipers, rather than obeying mirror symmetry. This pattern of activity, together with the connectivity of protocerebral bridge glomeruli and ellipsoid body sectors ensures that the E-PG bump moves smoothly around the ellipsoid body when the fly turns (Turner-Evans, 2017).
More broadly, topographical organization is a striking feature of many sensory circuits, but structure often follows computational function in neural circuits in the central brain as well. The feedforward pathways to and from the Mauthner cell make clear these neurons role in rapid escape behavior, and the parallel delay loops of the barn owl auditory system and the electric fish point to their comparative roles in localizing prey. The anatomical shift of the P-EN neurons with respect to the E-PG neurons provided an immediate clue to a potential structure/function relationship, that of a mechanism for shifting the bump of E-PG activity to update their internal representation of heading. The fact that topography often matches topology in the small fly brain makes the system ideal for the identification of circuit mechanisms underlying complex computations. Only time -- and perhaps large scale circuit reconstruction efforts -- will tell whether such network motifs are also present, but perhaps better hidden, in the more distributed circuits of much larger brains (Turner-Evans, 2017).
Neural networks regulate brain functions by routing signals. Therefore, investigating the detailed organization of a neural circuit at the cellular levels is a crucial step toward understanding the neural mechanisms of brain functions. To study how a complicated neural circuit is organized, this study analyzed recently published data on the neural circuit of the Drosophila central complex, a brain structure associated with a variety of functions including sensory integration and coordination of locomotion. Except for a small number of 'atypical' neuron types, it was found that the network structure formed by the identified 194 neuron types can be described by only a few simple mathematical rules. Specifically, the topological mapping formed by these neurons can be reconstructed by applying a generation matrix on a small set of initial neurons. By analyzing how information flows propagate with or without the atypical neurons, it was found that while the general pattern of signal propagation in the central complex follows the simple topological mapping formed by the 'typical' neurons, some atypical neurons can substantially re-route the signal pathways, implying specific roles of these neurons in sensory signal integration. The present study provides insights into the organization principle and signal integration in the central complex (Chang, 2017).
Boyan, G., Liu, Y., Khalsa, S. K. and Hartenstein, V. (2017). Dev Genes Evol 227(4): 253-269. PubMed ID: 28752327
The conserved nature of fan-shaped body neuroarchitecture in insects such as Drosophila and the grasshopper makes it likely that there is also a high degree of correspondence among their commissural fascicles. The current study now enables the development of the fan-shaped body to be understood at the level of individually identified commissural fascicles, and so provides the basis for interspecific comparisons of central complex development involving Drosophila, Tenebrio and the sightless dipluran Campodea where similar patterns of axon decussation are found. Equally, mutant analyses in Drosophila may allow the pattern of decussation present in the grasshopper to be understood with greater precision. For example, the topographic decussation of axons at stereotypic locations in both species suggests the presence of choice points across the midbrain similar to that reported for the ventral nerve cord. Although the mechanism has yet to be identified in the brain, a dysregulation of cell surface adhesion/recognition molecules in a graded manner across the midbrain represents one possibility. In the peripheral nervous system of Drosophila mutant for the cell surface molecule fasciclin III, for example, axons switch fascicles to an incorrect branch of the segmental nerve and so project to inappropriate body wall muscles, while in the visual system, relative expression levels of adhesion molecules have been found to regulate the wiring of neurite fascicles. Glia have also been shown to direct neuronal axogenesis in the CNS and midline glia are present in the both the grasshopper and Drosophila brain during commissure formation. In the karussell mutant (mutation affecting β-spectrin), for example, dysregulation of midline glia belonging to the pointed group results in commissural axons of the ventral nerve cord decussating between anterior and posterior fascicles, a neuroarchitecture not found in the wild type (Boyan, 2017).
The central complex comprises an elaborate system of modular neuropils which mediate spatial orientation and sensory-motor integration. The neuroarchitecture of the largest of these modules, the fan-shaped body, is characterized by its stereotypic set of decussating fiber bundles. These are generated during development by axons from four homologous protocerebral lineages which enter the commissural system and subsequently decussate at stereotypic locations across the brain midline. It is not clear how the decussating bundles relate to individual lineages, or if the projection pattern is conserved across species. This study traced the axonal projections from the homologous central complex lineages into the commissural system of the embryonic and larval brains of both the grasshopper and Drosophila. Projections into the primordial commissures of both species are found to be lineage-specific and allow putatively equivalent fascicles to be identified. Comparison of the projection pattern before and after the commencement of axon decussation in both species reveals that equivalent commissural fascicles are involved in generating the columnar neuroarchitecture of the fan-shaped body. Further, the tract-specific columns in both the grasshopper and Drosophila can be shown to contain axons from identical combinations of central complex lineages, suggesting that this columnar neuroarchitecture is also conserved (Boyan, 2017).
In both the grasshopper S. gregaria and Drosophila, the fan-shaped body with its prominent columnar neuroarchitecture (see Wiring of the central complex subserves information processing) comprises the largest module of the adult central complex. In the grasshopper, this columnar neuroarchitecture develops from an initially orthogonal primary axon scaffold during the second half of embryogenesis and is functional at the time of hatching. The neuroarchitecture is generated when subsets of axons from four lineages (termed W, X, Y, Z) in each protocerebral hemisphere innervate the existing commissural system but then decussate from anterior to more posterior lying fascicles at stereotypic locations across the central brain in a process known as 'fascicle switching'. In Drosophila, decussation of axons from four putatively equivalent lineages to those of the grasshopper also occurs , but during the larval to pupal transition, so that the resulting neuroarchitecture is essentially an adult feature. Species comparisons reveal that fascicle switching is present at some stage of development in the central brain of all arthropods and so may be considered a conserved mode of axogenesis (Boyan, 2017).
A major drawback in understanding of axon decussation in the insect brain has been the lack of a systematic identification of the embryonic commissural fascicles involved. In the grasshopper, for example, although a map of all commissures for the adult brain has been available for some time, the embryonic commissures have to date only been superficially allocated into anterior (ac) and posterior (pc) subsets, in keeping with the nomenclature for the early ventral nerve cord. Further, while the pioneers of the w, x, y, and z tracts from each protocerebral hemisphere have been shown to project into the primary commissural fascicle of the brain just after its formation early in embryogenesis and then to fasciculate with its pioneers, the early axons from the W, X, Y, and Z lineages remain within the commissural fascicles they originally pioneer. Later, in growing axons from these same lineages, however, subsequently decussate but the commissural fascicles involved have remained undescribed. This study reconstructed the axon projections from representative lineages of the central complex into the commissural system of the brain at various developmental stages in both the grasshopper and Drosophila. At the same time, the commissural organization itself was analyzed at these stages using the nomenclature applicable to the grasshopperand Drosophila. This analysis leads to the conclusion that in setting up the columnar neuroarchitecture of the fan-shaped body, comparable choices are being made by subsets of axons from equivalent lineages in both the grasshopper and Drosophila, consistent with a conserved wiring plan for this brain region (Boyan, 2017).
In both Drosophila and the grasshopper, the axon scaffold of the embryonic brain comprises an orthogonal system of axonal projections around the stomodeum. Anterior to the stomodeum, this scaffold in Drosophila had earlier been resolved to the level of grouped anterior or posterior commissures, but not individual fascicles. Recent studies, using specific Gal-4 lines, on the other hand, have documented a large number of single projections from larval/pupal neurons of the protocerebrum to the protocerebral bridge and then to the fan-shaped body, ellipsoid body and noduli, many of a commissural nature. Such commissural elements may now be integrated into a plan equivalent to that developed for the grasshopper (Boyan, 2017).
Multiple brain regions respond to harmful nociceptive stimuli. However, it remains unclear as to whether behavioral avoidance of such stimuli can be modulated within the same or distinct brain networks. This study found subgroups of neurons localized within a well-defined brain region capable of mediating both innate and conditioned nociceptive avoidance in Drosophila. Neurons in the ventral, but not the dorsal, of the multiple-layer organized fan-shaped body (FB) are responsive to electric shock (ES). Silencing ES-responsive neurons, but not non-responsive neurons, leads to reduced avoidance of harmful stimuli, including ES and heat shock. Activating these neurons consistently triggers avoidance and can serve as an unconditional stimulus in an aversive classical conditioning task. Among the three groups of responsive neurons identified, two also have reduced activity in ES-conditioned odor avoidance. These results demonstrate that both innate and conditioned nociceptive avoidance might be represented within neurons confined to a single brain region (Hu, 2018).
The current work investigated the roles of a well-defined fly brain region, the FB, and several groups of identifiable FB neurons in nociceptive stimuli-induced innate avoidance and conditioned avoidance. To ensure genetic manipulation specifically targeted the FB neurons, four gal4 lines, R49H02, R89E07, R38E07, and C205, which have relatively specific expression patterns, were used to label FB neurons of interest. In addition, MB-Gal80 was included in relevant transgenic flies to exclude the potential expression of transgenes that could lead to alteration in neuronal activity in the mushroom body (MB) neurons, which are important for aversive olfactory conditioning. Moreover, assays confirmed that locomotion and aversive odor sensitivity, two basic behaviors necessary for the tasks involved, remained normal with targeted neuronal activity manipulation through either temperature shifts or light (Hu, 2018).
The results presented above demonstrate that FB neurons are capable of mediating both innate and conditioned avoidance. First, at least three subgroups of large-field FB neurons, labeled by R49H02, R89E07, and R38E07 Gal4 lines and distributed within layers 1, 2, 4, and 5, that responded to ES. Second, it was demonstrated that these three groups of responsive FB neurons contributed to nociceptive stimuli-induced innate avoidance as follows: inhibition of activity of these neurons attenuated both ES and heat-induced innate avoidance, whereas activation of these neurons alone drove innate avoidance as well as served as a harmful US in an aversive conditioning task. These neurons appear to be specific to nociceptive stimuli-induced avoidance, but not to avoidance induced by non-harmful sensory stimuli (e.g., aversive odors). Third, three subgroups of FB neurons were identified, labeled by the R49H02, R89E07, and C205 Gal4 lines, which mediated conditioned avoidance in the ES- and odor-coupled classical conditioning tasks. Although odor avoidance is not mediated through these neurons, ES-conditioned odor avoidance was significantly reduced when activity of these neurons was inhibited. Such reduction was specific to nociceptive stimuli-conditioned odor avoidance, but not sugar-conditioned attraction. All lines of evidence presented are highly consistent in their support of the conclusion that the FB region is involved in nociceptive stimuli-induced innate and conditioned avoidance (Hu, 2018).
In the current study, ventral FB layers, including L1, L2, L4, and L5, are highly responsive to ES stimuli. However, silencing different FB layers shows different behavioral phenotypes in ES avoidance. Intensity of ES-induced calcium response and behavioral effect of ES avoidance are not in a simple linear relationship. Outputs of L1 and L4 neurons (R49H02) are required in avoiding both weak and strong ES stimuli. Interestingly, silencing L2 (R89E07) or L5 neurons (R38E07) specifically decrease behavioral avoidance of weak or strong ES stimuli, respectively. A possible explanation of these experimental data might be that behavioral avoidance of ES stimuli with different intensity is determined by combination of different ES-responsive FB layers. Such combination may allow animals to better react to nociceptive stimuli with different intensity. Consistent with this explanation, silencing one or two ES-responsive FB layers only partially affects ES avoidance behavior. It would be important to verify this idea by testing whether all ventral layers (i.e., L1-L5) of FB neurons are either inhibited or activated. An attempt to make a combination of R49H02, R89E07, and R38E07 failed because they are located too close to one another genetically (Hu, 2018).
The subgroups of the FB neurons labeled by the four Gal4 lines, R49H02, R89E07, R38E07, and C205, may be subdivided into three functional groups: type 1, conditioned avoidance only; type 2, innate avoidance only; or type 3, both innate and conditioned avoidance. Type 1 is represented by layer 6 (L6) neurons labeled by C205, which is a highly specific Gal4 line. These neurons do not respond to ES and have no influence on ES- or heat-induced acute innate avoidance but attenuate ES-conditioned odor avoidance. Type 2 is represented by layer 5 (L5) neurons labeled by R38E07. R38E07-labeled neurons (L5, L8, and L9) affect only ES- and heat-induced innate avoidance and play no role in conditioned avoidance. Because L8 and L9 do not respond to ES, it is likely that L5 neurons are responsible for the observed innate avoidance. Type 3 is representk are reported to sense heat in the larval stage. Specific heat nociceptors have yet to be reported in adult flies. In the central brain, how nociceptive stimuli, including ES and heat, are detected and processed are still unknown. This study showed that these two nociceptive stimuli share the same FB region in processing and in guiding avoidance. These data imply that this specific brain region may take part in general nociceptive processing (Hu, 2018).
As one of the distinct parts of the CX in the fly brain, the FB is important for multiple functions, including locomotion control, visual features recognition and processing, courtship maintenance, and quiescence regulation. It is interesting to note that two major functions of the FB, visual features recognition and quiescence regulation, largely rely on the dorsal layers of the FB, whereas nociceptive functions are localized mainly in the ventral and middle layers. The different layers of the FB may serve as hubs for different types of behavioral information (Hu, 2018).
Insects are ectothermal animals that are constrained in their survival and reproduction by external temperature fluctuations which require either active avoidance of or movement towards a given heat source. In Drosophila, different thermoreceptors and neurons have been identified that mediate temperature sensation to maintain the animal's thermal preference. However, less is known how thermosensory information is integrated to gate thermoresponsive motor behavior. This study used transsynaptic tracing together with calcium imaging, electrophysiology and thermogenetic manipulations in freely moving Drosophila exposed to elevated temperature and identify different functions of ellipsoid body ring neurons, R1-R4, in thermoresponsive motor behavior. The results show that warming of the external surroundings elicits calcium influx specifically in R2-R4 but not in R1, which evokes threshold-dependent neural activity in the outer layer ring neurons. In contrast to R2, R3 and R4d neurons, thermogenetic inactivation of R4m and R1 neurons expressing the temperature-sensitive mutant allele of dynamin, shibire(TS), results in impaired thermoresponsive motor behavior at elevated 31 °C. trans-Tango mediated transsynaptic tracing together with physiological and behavioral analyses indicate that integrated sensory information of warming is registered by neural activity of R4m as input layer of the ellipsoid body ring neuropil and relayed on to R1 output neurons that gate an adaptive motor response. Together these findings imply that segregated activities of central complex ring neurons mediate sensory-motor transformation of external temperature changes and gate thermoresponsive motor behavior in Drosophila (Buhl, 2021).
In flies, neuronal sensors detect prandial changes in circulating fructose levels and either sustain or terminate feeding, depending on internal state. This study describes a three-part neural circuit that imparts satiety-dependent modulation of fructose sensing. Dorsal fan-shaped body neurons display oscillatory calcium activity when hemolymph glucose is high, and these oscillations require glutamatergic input from SLP-AB or 'Janus' neurons projecting from the protocerebrum to the asymmetric body. Suppression of activity in this circuit, either by starvation or by genetic silencing, promotes specific drive for fructose ingestion. This is achieved through neuropeptidergic signaling by tachykinin, which is released from the fan-shaped body when glycemia is high. Tachykinin, in turn, signals to Gr43a-positive fructose sensors to modulate their response to fructose. Together, these results demonstrate how a three-layer neural circuit links the detection of two sugars to produce precise satiety-dependent control of feeding behavior (Musso, 2021).
Regulation of energy intake is a complex process involving food search, an animal's internal state, and the sensory qualities of food. In flies, fructose, either consumed directly or rapidly metabolized from precursors, promotes feeding through activation of a brain fructose sensor called Gr43a. This study describes how a neuronal network composed of neurons in the FB and asymmetric body contributes to energy homeostasis by detecting satiety-dependent changes in hemolymph glucose and modulating fructose drive (Musso, 2021).
The central complex, which is composed of the FB, the protocerebral bridge (PB), the ellipsoid body, and the noduli, is regarded as a center for sensorimotor integration that functions in goal-directed behavior. The FB is organized in nine horizontal layers and nine vertical columns. FB large field neurons of layers 1 to 3, and inputs to these layers from the PB, encode flight direction and general sensory orientation. FB layers 6 and 7 are well known to regulate sleep and arousal, locomotor control, courtship, visual memory, and decision-making related to taste. Layer 6 also plays a role in avoiding conditioned odors, while layers 1, 2, 4, and 5 respond to electric stimuli and are required for innate odor avoidance. However, the function of the most dorsal FB layers (8 and 9), mostly innervated local tangential neurons and AB-FBl8 (or vΔA_a), remained poorly understood. The results demonstrate a role for these layers in feeding regulation (Musso, 2021).
dFB oscillations were found to be require glutamatergic input from Janus neuron projections to the asymmetric body. Described for the first time in 2004, very little is known about AB function; 92.4% of flies display asymmetry in the AB, with the body present only in the right hemisphere, while 7.6% also have a body on the left side. It is noted that oscillations in the dFB display a tendency to be faster on the right side, with clearly asynchronous activity between the two sides that may reflect their asymmetric input from Janus neurons. The small proportion of flies displaying symmetry in the AB have defects in LTM, a process that is known to require energy. It is speculated that these symmetric flies may have a dysfunctional Janus neurons-to-dFB connection, resulting in impaired Tk release. This could affect LTM either directly or through changes in feeding. A role for TK in memory has been demonstrated in honeybees and mammals, and TkR86C appears to be expressed in serotonergic paired neurons known to interact with MB-MP1 neurons required for LTM formation. Tk also acts through TkR99D to modulate activity in neurons producing insulin-like peptides, which affect LTM formation (Musso, 2021).
Modulation of dFB oscillations by Janus neurons requires glutamatergic signaling through a group of glutamate receptors including KaiR1D, NmdaR1, NmdaR2, and GluClα, but not AMPA receptors. Both KaiR1D receptors, which are homomeric, and N-methyl-D-aspartate (NMDA) receptors, which are heteromeric complexes between subunits 1 and 2, pass Ca2+ current. NMDA receptors (NMDAR) are well known for their role in mediating synaptic plasticity and can also trigger oscillatory activity. NMDAR function as molecular coincidence detectors, requiring simultaneous ligand binding and membrane depolarization for activation. It is possible that dFB neuron oscillations are triggered by the coincident detection of glutamate from Janus neurons and glucose from the hemolymph; however, because the FB are receiving many inputs from other brain region, it is suspect that dFB oscillations require additional inputs as well. The chloride channel GluClα is also required for dFB oscillations. GluClα has been previously implicated in on/off responses of the visual system of flies and memory retention in honeybees, demonstrating a role in regulating cell excitability. Perhaps, GluClα functions in repolarization of the dFB neurons between calcium bursts. Further study will be required to fully understand how the suite of glutamate receptors function together to drive oscillations, along with the source of input to Janus neurons in the protocerebrum (Musso, 2021).
Because glucose is the primary circulating energy source, one might intuitively expect that enhancing feeding in response to postingestive glucose detection would be the most efficient means of optimizing energy uptake. However, using elevation of hemolymph glucose as a signal to continue feeding is problematic because glucose levels are tightly regulated and elevated glucose serves as a signal of satiety. On the other hand, internal fructose can vary widely in response to ingestion and can therefore be a more reliable indicator of recent sugar intake. Thus, the separation of glucose as a satiety indicator and fructose as marker of sugar consumption removes the potential ambiguity of each as a signal. Moreover, fructose typically coexists with other nutritive sugars in common food sources. Therefore, it may not be the case that flies specifically benefit from fructose intake but rather that fructose serves as an effective proxy for general carbohydrate ingestion. By using fructose and the narrowly tuned Gr43a fructose receptor to survey sugar consumption, flies can effectively benefit from both a fructose-mediated positive feedback loop and glucose-mediated negative feedback to co-operatively ensure appropriate energy intake (Musso, 2021).
The finding that dFB glucose sensing modulates fructose feeding via Gr43a brain neurons fits with the established model of Gr43a brain neurons as central fructose sensors. For this mechanism to effectively sustain feeding on a rich sugar source, ingested sugars must rapidly increase fructose signaling to Gr43a brain neurons, which then must acutely promote feeding. While the precise kinetics of internal fructose elevation after sugar consumption have not been quantified, fructose levels in the head rapidly increase 10-fold after fructose feeding and then return to baseline. The role of direct fructose sensing by Gr43a brain neurons is highlighted by the observation that Gr43a knockdown in those neurons results in markedly lower relative intake of fructose compared to glucose. Unexpectedly, knockdown of Gr64a, another sugar receptor expressed in the same neurons, produced the opposite effect. This could be because Gr64a contributes to modulation of Gr43a brain neurons by other sugar cues, and the absence of this activity makes Gr43a-mediated fructose responses more pronounced. Alternatively, Gr43a may be expressed more strongly after Gr64a knockdown, leading to an increased fructose response (Musso, 2021).
Little is known about the mechanisms downstream of Gr43a brain neurons that promote feeding. All Gr43a brain neurons express the peptide Crz, but knockdown of Crz expression produced no significant effect on fructose preference over glucose. This suggests an important functional role for another neurotransmitter, although it is also possible that the RNAi knockdown was not effective. Irrespective of mechanism, two experiments support the idea that activation of Gr43a neurons acutely enhances feeding. First, silencing of dFB neurons by genetic manipulation or prolonged starvation produces Gr43a-dependent fructose preference within the first 10 min of a flyPAD assay. Second, closed-loop optogenetic activation of Gr43a brain neurons was sufficient to produce a strong positive preference within 10 min in the STROBE (Musso, 2021).
The separable functions of glucose and fructose sensing in flies bear notable resemblance to the differential effects of these two sugars in the mammalian hypothalamus. In particular, AMPK expression in the arcuate nucleus of the hypothalamus is known to link energy levels to food drive. When glycemia is low, AMPK is activated and thereby promotes feeding through orexigenic AgRP/NPY neuron activity. Glucose administration suppresses activity in these peptidergic neurons, while fructose can have the opposite effect and promote further feeding. The first description of fly Gr43a neurons noted their orexinegenic activity and suggested a potential functional homology with the hypothalamus. In the present study, a multilayered neural system centered on a brain energy sensor (dFB), was uncovered whose activation by glucose leads to anorexigenic behavior through inhibition of the brain fructose sensor Gr43a. Thus, the results are consistent with at least partial functional homology between the mammalian hypothalamus and brain Gr43a neurons of the fly (Musso, 2021).
Many animals orient using visual cues, but how a single cue is selected from among many is poorly understood. This study shows that Drosophila ring neurons-central brain neurons implicated in navigation-display visual stimulus selection. Using in vivo two-color two-photon imaging with genetically encoded calcium indicators, this study demonstrates that individual ring neurons inherit simple-cell-like receptive fields from their upstream partners. Stimuli in the contralateral visual field suppressed responses to ipsilateral stimuli in both populations. Suppression strength depended on when and where the contralateral stimulus was presented, an effect stronger in ring neurons than in their upstream inputs. This history-dependent effect on the temporal structure of visual responses, which was well modeled by a simple biphasic filter, may determine how visual references are selected for the fly's internal compass. This approach highlights how two-color calcium imaging can help identify and localize the origins of sensory transformations across synaptically connected neural populations (Sun, 2017).
The central complex is a highly conserved insect brain region composed of morphologically stereotyped neurons that arborize in distinctively shaped substructures. The region is implicated in a wide range of behaviors and several modeling studies have explored its circuit computations. Most studies have relied on assumptions about connectivity between neurons based on their overlap in light microscopy images. This study presents an extensive functional connectome of Drosophila melanogaster's central complex at cell-type resolution. Using simultaneous optogenetic stimulation, calcium imaging and pharmacology, the connectivity was tested between 70 presynaptic-to-postsynaptic cell-type pairs. Numerous inputs to the central complex were identified, but only a small number of output channels (see The central complex neuropiles and the hypothesized flow of information based on overlap of arbors in light-microscopy images). Additionally, the connectivity of this highly recurrent circuit appears to be sparser than anticipated from light microscopy images. Finally, the connectivity matrix highlights the potentially critical role of a class of bottleneck interneurons. All data is provided for interactive exploration on a website (Franconville, 2018).
Despite their small brains, insects can navigate over long distances by orienting using visual landmarks, skylight polarization, and sun position. Although Drosophila are not generally renowned for their navigational abilities, mark-and-recapture experiments in Death Valley revealed that they can fly nearly 15 km in a single evening. To accomplish such feats on available energy reserves, flies would have to maintain relatively straight headings, relying on celestial cues. Cues such as sun position and polarized light are likely integrated throughout the sensory-motor pathway, including the highly conserved central complex. Recently, a group of Drosophila central complex cells (E-PG neurons) have been shown to function as an internal compass, similar to mammalian head-direction cells. Using an array of genetic tools, this study set out to test whether flies can navigate using the sun and to identify the role of E-PG cells in this behavior. Using a flight simulator, it was found that Drosophila adopt arbitrary headings with respect to a simulated sun, thus performing menotaxis, and individuals remember their heading preference between successive flights-even over several hours. Imaging experiments performed on flying animals revealed that the E-PG cells track sun stimulus motion. When these neurons are silenced, flies no longer adopt and maintain arbitrary headings relative to the sun stimulus but instead exhibit frontal phototaxis. Thus, without the compass system, flies lose the ability to execute menotaxis and revert to a simpler, reflexive behavior (Giraldo, 2018).
In the absence of normal E-PG function, flies might directly orient toward the sun, because they lack the ability to compare their instantaneous heading to a stored value of their directional preference. Such a loss of function in the compass network might unmask a simpler reflexive behavior, such as phototaxis, that does not require the elaborate circuitry of the central complex. Consistent with this hypothesis, stripe fixation was not different between control and experimental animals. This interpretation is compatible with a recent model that showed that frontal object fixation could result from a simple circuit involving two asymmetric wide-field motion integrators, without the need for the central complex (Giraldo, 2018).
The findings are consistent with an emerging model of a navigational circuit involving the central complex. E-PG cells have an excitatory relationship with another cell class in the central complex (protocerebral bridge to ellipsoid body and noduli, or P-EN, neurons), creating an angular velocity integrator that allows a fly to maintain its heading in the absence of visual landmarks. Furthermore, the E-PG neurons are homologous to the CL1 neurons described in locusts, monarchs, dung beetle, and bees and likely serve similar functions across taxa. Extracellular recordings from the central complex in cockroaches revealed neurons that act as head-direction cells relative to, or in the absence of, visual landmarks, although precise cell types were not identified. Inputs to E-PG neurons likely occur via the anterior visual pathway from the medulla to the anterior optic tubercle and on to the bulb. From there, tubercle-bulb neurons, one class of which is responsive to the azimuth and elevation of small bright spots, synapse onto ring neurons that project to the ellipsoid body, thus bringing visual information into the compass network. In a recent model of path integration in bees, CL1 neurons are part of a columnar circuit that provides instantaneous heading information to an array of self-excitatory networks that also receive convergent optic flow information, thereby storing a memory of distance traveled in each direction (Stone, 2017). This information is then retrieved as an animal returns home, by driving appropriate steering commands in another class of central complex neurons. The putative memory cells suggested by this model, CPU4 cells, could be homologous to protocerebral bridge-fan-shaped-body noduli (P-FN) neurons described for Drosophila. Furthermore, cells responsive to progressive optic flow are found throughout the central complex of flies, including neuropil in the fan-shaped body containing the P-FN cells. In addition to their role in path integration, the CPU4 network might also function to store the desired heading during sun navigation. Although the results do not directly test this model, they are consistent with the role of CL1 neurons in providing heading direction to circuits that generate steering commands toward an arbitrary orientation whose memory is stored in the network of CPU4 (P-FN) neurons (Giraldo, 2018).
Stripe fixation and sun navigation behaviors may represent two different flight modes in Drosophila. Stripe fixation is thought to be a short-range behavioral reflex to orient toward near objects, which, in free flight, is quickly terminated by collision avoidance or landing behaviors. In contrast, navigation using the sun is likely a component of long-distance dispersal behavior that could be used in conjunction with polarization vision either in a hierarchical or integrative manner. Individuals could differ in where they lie on the continuum of long-range dispersal to local search, which could explain the inter-individual variation observed in heading fidelity during sun orientation experiments. In general, dispersal is a condition-dependent behavior that is known to vary with hunger or other internal factors. Given the architectural similarity of the central complex among species, the celestial compass identified in Drosophila is likely one module within a conserved behavioral toolkit, allowing orientation and flight over long distances by integrating skylight polarization, the position of the sun or moon, and other visual cues. An independent study has recently found that the E-PG compass neurons are also necessary in walking flies for maintaining arbitrary headings relative to a small bright object. The expanding array of genetic tools developed for flies and the rapid growth in understanding of the neural circuitry involved in rientation and flight make this a promising system for exploring such essential and highly conserved behaviors (Giraldo, 2018).
Successful navigation depends on an animal's ability to perceive its spatial orientation relative to visual surroundings. Heading direction in insects is represented in the central complex (CX), a navigation center in the brain, to generate steering commands. In insects that navigate relative to sky compass signals, CX neurons are tuned to celestial cues indicating the location of the sun. The desert locust CX contains a compass-like representation of two related celestial cues: the direction of unpolarized direct sunlight and the pattern of polarized light, which depends on the sun position. Whether congruent tuning to these two compass cues emerges within the CX network or is inherited from CX input neurons is unclear. To address this question, this study intracellularly recorded from GABA-immunoreactive TL neurons, input elements to the locust CX (corresponding to R neurons in Drosophila), while applying visual stimuli simulating unpolarized sunlight and polarized light across the hemisphere above the animal. TL neurons were shown to have large receptive fields for both types of stimuli. However, faithful integration of polarization angles across the dorsal hemisphere, or matched-filter ability to encode particular sun positions, was found in only two out of 22 recordings. Those two neurons also showed a good match in sun position coding through polarized and unpolarized light signaling, whereas 20 neurons showed substantial mismatch in signaling of the two compass cues. The data, therefore, suggest that considerable refinement of azimuth coding based on sky compass signals occurs at the synapses from TL neurons to postsynaptic CX compass neurons (Takahashi, 2022).
The central complex, a set of neuropils in the center of the insect brain, plays a crucial role in spatial aspects of sensory integration and motor control. Stereotyped neurons interconnect these neuropils with one another and with accessory structures. Over 5000 Drosophila melanogaster GAL4 lines were screened for expression in two neuropils, the noduli (NO) of the central complex and the asymmetrical body (AB), and multicolor stochastic labelling was used to analyze the morphology, polarity and organization of individual cells in a subset of the GAL4 lines that showed expression in these neuropils. Nine NO and three AB cell types were identified and are described in this study. The morphology of the NO neurons suggests that they receive input primarily in the lateral accessory lobe and send output to each of the six paired noduli. The AB is demonstrated to be a bilateral structure which exhibits asymmetry in size between the left and right bodies. The AB neurons are shown to directly connect the AB to the central complex and accessory neuropils, that they target both the left and right ABs, and that one cell type preferentially innervates the right AB. It is proposed that the AB be considered a central complex neuropil in Drosophila. Finally, highly restricted GAL4 lines are presented for most identified protocerebral bridge, NO and AB cell types. These lines, generated using the split-GAL4 method, will facilitate anatomical studies, behavioral assays, and physiological experiments (Wolff, 2018).
Located at the center of the insect brain, the central complex is a set of highly
interconnected neuropils that processes complex, multisensory information from
the environment, integrates it with information about the insect's internal state
and past experiences, and guides motor outputs that drive appropriate behavioral
responses (Wolff, 2018).
One of the most studied roles of the insect central complex is the integration of
sensory information, predominantly from visual input. The output of this sensory
processing encompasses diverse motor and behavioral responses. In this
capacity, the central complex regulates locomotor behaviors such as
handedness, turn direction, initiation and termination of walking. The central complex is thought to play a key role in migration, navigation and orientation using input such as celestial cues and displays responses to looming stimuli suggestive of
an involvement in generating escape responses in the locust and fly. The central complex has
been suggested to contain a ring attractor network that maintains a representation of
the fly's heading direction that may be useful for navigation and orientation in
visual conditions as well as in darkness. The central complex is also involved in the formation and recall of short- and long-term visual memories, in the processing of olfactory and gustatory inputs and in maintaining information about the fly's satiety state (Wolff, 2018).
Understanding the core principles of operation of the central complex has been
greatly enabled by the dissection of behavior at a single neuron level and the
neuron-by-neuron assembly of circuits. A comprehensive anatomical atlas and
genetic lines that enable manipulation of individual cell types are invaluable tools
for this strategy. This study describes the neuronal composition of the NO
and the AB, neither of which has been extensively studied in Drosophila. An
understanding of the function of the noduli in behavior lags far behind
that of the other central complex structures: the protocerebral bridge (PB), fan-
shaped body (FB) and ellipsoid body (EB). The only documented roles for the
NO in Drosophila are in the
time course of walking activity and in influencing
handedness during locomotion. The locust neurons that
connect the PB, EB and NO and the PB, FB and NO are sensitive to polarized
light. Most recently, recordings from optic-flow-sensitive neurons that connect the lateral accessory lobe (LAL) to the NO and other neurons that link the
NO to the FB in the bee have demonstrated the NO are involved in path
integration. Finally, the fact that this structure appears to be
present only in the subclass of winged insects has led to the speculation that the
noduli may regulate flight (Wolff, 2018).
Structural conservation of the central complex across insect species is strong,
but not absolute. The discussion that follows focuses on the anatomy of the
Drosophila neuropils. The PB, FB, EB and NO
are midline structures and exhibit a stratified organization. The PB is a handlebar-
shaped structure in the posterior dorsal brain. The EB is shaped like
a torus and is tilted
on its dorso-ventral axis such that its ventral border defines
the anterior margin of the central complex. The FB lies between the
PB and EB and represents the largest of the four central complex structures. The bilateral noduli, historically called ventral tubercles, are the most ventral neuropil of the central complex and are nestled beneath the FB. There are three pairs of noduli neatly stacked on top of one
another from dorsal to ventral; each pair is bisected by the midline. The dorsal nodulus (NO
1) displays some hint of a transverse division whereas the medial (NO2)
and ventral (NO3) noduli exhibit longitudinal segmentation. NO2 is divided into
dorsal and ventral subdomains (NO2D and NO2V) and NO3 has three subdomains (Wolff, 2018).
The neuropils considered to be components of the central complex have evolved
over time. Power's
1943 description of the central complex includes the FB, EB, and 'ventral tubercles', or NO. By the mid-1970s, the modern view of the central complex had emerged: Williams included the PB within the locust
central complex, alongside the FB, EB and NO. The asymmetrical body (AB) is a
relatively inconspicuous structure located at the
midline, adjacent to the ventral FB. It was first described
in the fly as a round, almost exclusively right-hemisphere structure. The AB was observed in both hemispheres in just 7.6% of 2,250 brains
immunolabeled with an antibody against the Fasciclin II (Fas II) protein, which is
expressed in this structure. Flies with bilateral ABs were reported to have
disrupted long-term memory, leading to the suggestion that asymmetry of this
structure is important for long-term, but not short-term, memory. Although thousands of GAL4 lines that drive expression in small subsets
of neurons in the larval and adult fly brains have been examined, the AB is the only reported instance of an asymmetric structure in the adult fly brain (Wolff, 2018).
Elements with likely homology to the Drosophila AB have also been described in
the grey flesh fly Neobellieria bullata and the blowfly Calliphora
erythrocephala. In both species, these bodies occur
bilaterally, and one of the two is consistently smaller and less densely innervated
than the other. In addition, the smaller of the two appears fragmented.
A previous study identified five GAL4 lines that show asymmetric innervation of
the AB. That analysis revealed lines ranging from a strong right hemisphere bias
in innervation to those with asymmetric but bilateral expression, with more
conspicuous expression in the right AB. This study builds on the previous study
by providing a systematic characterization of the neurons that target the AB,
and leads to a proposal that the AB be added as the fifth neuropil of the
Drosophila central complex (Wolff, 2018).
In this work, a characterization is presented of cell types of the NO and AB,
including morphology, presumed polarity and population size. A set of split-GAL4 lines for NO and AB cell types, reagents was also generated
and characterized that will greatly facilitate functional studies. In addition, since publication of a description of the neurons that arborize in the PB, this study has gained several new insights into the PB neurons. These include: 1) one new PB neuron family has been identified; 2) a neuron
identified by in a previous study has since been found in the GAL4 collection and is
characterized in this study using the multicolor flip-out technique (MCFO);
and 3) a set of split-GAL4 lines was generated
for PB cell types (Wolff, 2018).
The asymmetrical body: the fifth central complex structure
The neuropils considered to be constituent components of the central complex
have changed over the decades. The central body), included what
became known as the ellipsoid body and the fan-shaped body. The noduli were known
as the ventral tubercles. More recently, the central complex has been defined as
'a group of modular neuropils across the midline of the insect brain',
'...interconnected neuropils and nuclei that populate the
midline of the forebrain-midbrain boundary region', and
'a system of interconnected neuropils lying at, or about, the midline of the
protocerebrum'. Although the modular architecture of the central
complex structures is conspicuous (e.g. the glomeruli of the PB and the trajectory
patterns of neurons that project to, from, and within the central complex
structures), it is the assigned boundaries that encompass the central complex
that seem to be the feature that defines these structures as members of the
central complex (Wolff, 2018).
This study has illustrated that the Drosophila AB, which appears to be a structure that is
distinct from the FB, meets the criteria outlined above for central complex
neuropils: It is a midline neuropil; it falls within the boundaries of the central complex; and it is interconnected (to the FB and SLP) by a network of previously undocumented (with one exception) neurons (Wolff, 2018).
Since the AB meets all the criteria previously used to define neuropils as
components of the central complex, it is proposed that the AB be added as a fifth
neuropil of the central complex of Drosophila.
The AB is not unique to Drosophila. A previous study
describe the presence of likely homologous bilateral, asymmetrically sized ABs in
N. bullata and C. erythrocephala. That work also identified a tangential FB
neuron that bears a resemblance to the SLP-AB-FBl8 neuron described in this study (Wolff, 2018).
It remains to be determined if the AB is more widely represented in other insect
orders. The right AB is significantly larger than the left. At a minimum, this difference is
likely due to a combination of smaller arbors in the left AB and the lower
frequency with which the left AB is targeted: only the ipsilateral-contralateral-
projecting form of the SLP-AB neuron, which arborizes in both the left and right
ABs, targets the left AB, whereas the ipsilateral and contralateral-projecting
forms of the SLP-AB neuron target exclusively the right AB. Thus, the right AB
appears to receive a disproportionately larger share of information from the SLP,
although the right and left hemispheres appear to be equally represented as
sources of input. Notably, this left-right bias is restricted to the AB, as a parallel
preference is not shown for the SLP. The availability of genetic lines that target
AB-specific cell types will enable experiments aimed at revealing the relevance of
this left-right bias (Wolff, 2018).
Three unusual features distinguish the five most commonly seen NO neurons
described from other central complex neurons. First, in contrast to the
majority of PB neurons described to date, the projections of four of these five NO
neurons are ipsilateral. Second, while anatomical features identify distinct input
and output neuronal populations in other central complex neuropils, the noduli
appear to be sites for receiving primarily input from other neuropils (boutons
appear to be the predominant anatomical feature in the noduli in confocal micrographs
of NO, PB-FB-NO and PB-EB-NO neurons). Golgi preparations and
data from the likely locust equivalent of the PB-FB-NO neuron (the CPU4
neuron), however, indicate these NO arbors are mixed; perhaps the intensity of the dense
populations of boutons masks the presence of spines in confocal preparations.
Third, although the noduli do receive input from central complex neuropils (e.g.
via the PB-FB-NO and PB-EB-NO neurons, from FB tangential neurons, etc.),
the majority of direct input for this new set of neurons is provided
by just one neuropil, the LAL. Such a restricted thoroughfare of communication is in stark
contrast to the PB neurons, for example, which have a much broader and more
diverse network of direct communication. The LAL.s-CREc.s-NO3
Pc.b cell type is distinct from the other four common NO
neurons in that it delivers contralateral, rather than ipsilateral, input
from the LAL and CRE to NO3P. The posterior compartment of NO3 is therefore unique in that
it is the only nodulus subcompartment to communicate directly with the
contralateral hemisphere. Given that NO3P (and NO3M) also receives ipsilateral
terminals from the LAL via LAL.s-CREi.s-NO3P/Mi.b, this subcompartment may
act as a limited integration center between the fly's left and right sensory fields (Wolff, 2018).
Physiological data from two neurons in the sweat bee offer insight into a likely
role for the NO neurons described in this study. The TN1 and TN2 neurons ('noduli tangential neurons') share a high degree of anatomical homology with the LAL-
NO neurons: TN1 and TN2 are ipsilateral neurons with input branches in the
lateral central brain and blebbed branches in the noduli.
Recordings from these two cells reveal they fire in response to simulated
backward and forward flight, respectively, and that the rate of firing is dependent
on the stimulus velocity, suggesting these neurons encode speed using optic-
flow and can thereby track the distance traveled by the bee. Similar physiological
features and path integration functions would not be unexpected for the apparent
homologous Drosophila neurons.
The LAL is the primary source of input for the NO neurons described in this study and its
activity may provide additional insight into the roles of the LAL-NO neurons. It is
a large, bilateral neuropil that is highly interconnected with neuropils of the
central complex. Functionally, the LAL is considered to be a sensorimotor
integration center, based on several lines of evidence in various insect species.
For example, in crickets and moths, activity in LAL neurons is associated with
walking. In the locust, assorted LAL neurons exhibit changes in activity in response to various
aspects of flight, implicating this brain region in flight control. In
Drosophila, LAL neurons involved in walking backwards have been documented (Wolff, 2018).
It has been suggested that the noduli are involved in walking and motor control in
Drosophila. The neurons implicated in left-right turning bias in locomotion are the PB-FB-NO neurons, which have presumed input (fine terminals) in the PB,
and presumed output (boutons) in the FB and NO. It has been speculated that
the bias to turn in one direction or the other is influenced by an interplay between
the nodulus subdomains that are targeted by the different PB-FB-NO cell types (Wolff, 2018).
Direct communication between the PB-FB-NO neurons and the LAL-NO neurons
is not unexpected, as a previous study has shown synaptic contacts between
the bumblebee equivalents of these two cells, the CPU4 and TN cells,
respectively. Considering the sensorimotor contribution made by the LAL in
various types of movement, the LAL-NO neurons described in this study are strong
candidates to contribute to the circuits involved in turning (Wolff, 2018).
The catalog of NO neurons described in this study is incomplete. Analyses of other
GAL4 lines has identified several large-field FB neurons that also arborize in the
noduli, as well as other brain regions that are currently being characterized; some
of these neurons are illustrated in the Golgi stains of a previous study.
Other cell types with arbors in the noduli that
have so far eluded identification. Finally,
it seems likely that there would be output neurons from the
NO, although such neurons have not been identified. Electron microscopic-level analysis should provide a path to identifying these neurons (Wolff, 2018).
The debate continues to swirl over what constitutes a distinct cell type.
Morphology and function have long been accepted as reliable criteria to
distinguish cell types. While morphology is a straightforward and easy means of
classifying cell types, it can be misleading in that cells that appear identical may
have functional differences. For example, a previous study describe clearly
distinct physiological roles for two PB neurons that appear to have
indistinguishable morphology at the light level (Wolff, 2018).
Morphological features evident
with light microscope-level resolution will therefore likely be insufficient to
distinguish all cell types, so knowledge of some combination of synaptic
connectivity, functional properties and the genetic programs used to specify
these attributes will be necessary to fully define cell types. Similar limitations confound the assignment of neuropil boundaries and sub-compartments (Wolff, 2018).
Synaptic density varies considerably across brain regions and
this variation has provided landmarks used to define the neuropils of the fly brain. While the boundaries of some structures are unambiguous (e.g.,
the PB and EB), neuropil margins are not universally so clear-cut, with many
neuropils appearing to meld seamlessly with adjacent neuropils. The opportunity
to map the domains of arbors within neuropils identifies distinct regions that are
not revealed by differences in synaptic density (e.g. wedge and tile domains in
the EB). For example, the mushroom body lobes can be divided into a series of
non-overlapping compartments with distinct functions by the extent of the arbors
of dopaminergic input neurons and mushroom body output neurons. The LAL provides an example of one
neuropil that may have functionally distinct subregions. It is a large neuropil with
no obvious boundaries revealed by anti-Brp staining, yet the arbors of many
neurons that target this neuropil exhibit strong regional preferences. Mapping the
domains of these arbors may identify regions that are functionally distinct (Wolff, 2018).
Three major efforts aimed at cataloging all the neurons in the Drosophila brain
are in progress. One, typified by this and others' work, characterizes
one structure at a time using light microscopy in combination with the generation
and analysis of highly specific GAL4 driver lines. The second method is a
modern implementation of the Golgi approach of randomly labeling small
numbers of neurons in order to describe their morphology.
And the third, which is now becoming practical at the required scale, involves
reconstruction of neuronal morphology and circuits through analysis of image
volumes collected using electron microscopy. It is believed that such light and electron
microscopic-level analyses will be highly synergistic. Light microscopy, with
genetically marked cells, provides the ability to observe the morphology of
hundreds of individual cells of the same cell type in many different individuals,
providing insights on stereotypy. However, its dependence on GAL4 drivers
means that completeness of coverage cannot be assured. Conversely, electron
microscopic analysis, while usually limited to a single sample, not only ensures
completeness but also enables visualization and quantification of synaptic
connectivity. Moreover, since EM samples do not carry transgenes expressing
ectopic membrane proteins that can interfere with development, wiring errors
may be less likely. While only electron microscopy is likely to provide the
complete wiring diagram of a circuit, light level analysis of genetic driver lines will
be needed to provide the critical bridge between circuit maps and the tools
required to precisely manipulate the activity of their individual components (Wolff, 2018).
Sleep is essential but incompatible with other behaviors, and thus sleep drive competes with other motivations. Previous work has shown Drosophila males balance sleep and courtship via octopaminergic neurons that act upstream of courtship-regulating P1 neurons. This study shows that nutrition modulates the sleep-courtship balance and identify sleep-regulatory neurons downstream of P1 neurons. Yeast-deprived males exhibited attenuated female-induced nighttime sleep loss yet normal daytime courtship, which suggests male flies consider nutritional status in deciding whether the potential benefit of pursuing female partners outweighs the cost of losing sleep. Trans-synaptic tracing and calcium imaging identified dopaminergic neurons projecting to the protocerebral bridge (DA-PB) as postsynaptic partners of P1 neurons. Activation of DA-PB neurons led to reduced sleep in normally fed but not yeast-deprived males. Additional PB-projecting neurons regulated male sleep, suggesting several groups of PB-projecting neurons act downstream of P1 neurons to mediate nutritional modulation of the sleep-courtship balance ( , 2020).
Andrade, I. V., Riebli, N., Nguyen, B. M., Omoto, J. J., Cardona, A. and Hartenstein, V. (2019). Developmentally arrested precursors of pontine neurons establish an embryonic blueprint of the Drosophila central complex. Curr Biol 29(3): 412-425. PubMed ID: 30661802
Andreani, T., Rosensweig, C., Sisobhan, S., Ogunlana, E., Kath, W. and Allada, R. (2022). Circadian programming of the ellipsoid body sleep homeostat in Drosophila. Elife 11. PubMed ID: 35735904
Boyan, G., Liu, Y., Khalsa, S. K. and Hartenstein, V. (2017). A conserved plan for wiring up the fan-shaped body in the grasshopper and Drosophila. Dev Genes Evol 227(4): 253-269. PubMed ID: 28752327
Bridi, J. C., Ludlow, Z. N. and Hirth, F. (2019). Lineage-specific determination of ring neuron circuitry in the central complex of Drosophila. Biol Open 8(7). PubMed ID: 31285267
Buhl, E., Kottler, B., Hodge, J. J. L. and Hirth, F. (2021). Thermoresponsive motor behavior is mediated by ring neuron circuits in the central complex of Drosophila. JSci Rep 11(1): 155. PubMed ID: 33420240
Cao, H., Tang, J., Liu, Q., Huang, J. and Xu, R. (2022). Autism-like behaviors regulated by the serotonin receptor 5-HT2B in the dorsal fan-shaped body neurons of Drosophila melanogaster. Eur J Med Res 27(1): 203. PubMed ID: 36253869
Certel, S. J., McCabe, B. D. and Stowers, R. S. (2022). A conditional GABAergic synaptic vesicle marker for Drosophila. J Neurosci Methods 372: 109540. PubMed ID: 35219770
Chang, N., Huang, H. P. and Lo, C. C. (2023). Global inhibition in head-direction neural circuits: a systematic comparison between connectome-based spiking neural circuit models. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. PubMed ID: 36781446
Chang, P. Y., Su, T. S., Shih, C. T. and Lo, C. C. (2017). The topographical mapping in Drosophila central complex network and its signal routing. Front Neuroinform 11: 26. PubMed ID: 28443014
Chen, W. F., Maguire, S., Sowcik, M., Luo, W., Koh, K. and Sehgal, A. (2015). A neuron-glia interaction involving GABA transaminase contributes to sleep loss in sleepless mutants. Mol Psychiatry 20(2): 240-251. PubMed ID: 24637426
Cope, A. J., Sabo, C., Vasilaki, E., Barron, A. B. and Marshall, J. A. (2017). A computational model of the integration of landmarks and motion in the insect central complex. PLoS One 12(2): e0172325. PubMed ID: 28241061
Currier, T. A., Matheson, A. M. and Nagel, K. I. (2020). Encoding and control of orientation to airflow by a set of Drosophila fan-shaped body neurons. Elife 9. PubMed ID: 33377868
De, J., Wu, M., Lambatan, V., Hua, Y. and Joiner, W. J. (2023). Re-examining the role of the dorsal fan-shaped body in promoting sleep in Drosophila. Curr Biol. PubMed ID: 37552985
Dewar, A. D. M., Wystrach, A., Philippides, A. and Graham, P. (2017). Neural coding in the visual system of Drosophila melanogaster: How do small neural populations support visually guided behaviours?. PLoS Comput Biol 13(10): e1005735. PubMed ID: 29016606
Duan, W., Zhang, Y., Zhang, X., Yang, J., Shan, H., Liu, L. and Wei, H. (2023). A Visual Pathway into Central Complex for High-Frequency Motion-Defined Bars in Drosophila. J Neurosci 43(26): 4821-4836. PubMed ID: 37290936
Duhart, J. M., Baccini, V., Zhang, Y., Machado, D. R. and Koh, K. (2020). Modulation of sleep-courtship balance by nutritional status in Drosophila. Elife 9. PubMed ID: 33084567
Duhart, J. M., Buchler, J. R., Inami, S., Kennedy, K. J., Jenny, B. P., Afonso, D. J. S. and Koh, K. (2023). Modulation and neural correlates of postmating sleep plasticity in Drosophila females. Curr Biol 33(13): 2702-2716. PubMed ID: 37352854
Farnworth, M. S., Eckermann, K. N. and Bucher, G. (2020). Sequence heterochrony led to a gain of functionality in an immature stage of the central complex: A fly-beetle insight. PLoS Biol 18(10): e3000881. PubMed ID: 33104689
Faumont, S., Lindsay, T. H. and Lockery, S. R. (2012). Neuronal microcircuits for decision making in C. elegans. Curr Opin Neurobiol 22: 580-591. PubMed ID: 22699037
Fisher, Y. E., Lu, J., D'Alessandro, I. and Wilson, R. I. (2019). Sensorimotor experience remaps visual input to a heading-direction network. Nature. PubMed ID: 31748749
Flores-Valle, A., Goncalves, P. J. and Seelig, J. D. (2021). Integration of sleep homeostasis and navigation in Drosophila. PLoS Comput Biol 17(7): e1009088. PubMed ID: 34252086
Gao, X. J., Potter, C. J., Gohl, D. M., Silies, M., Katsov, A. Y., Clandinin, T. R. and Luo, L. (2013). Specific kinematics and motor-related neurons for aversive chemotaxis in Drosophila. Curr Biol 23: 1163-1172. PubMed ID: 23770185
Gao, X. J., Riabinina, O., Li, J., Potter, C. J., Clandinin, T. R. and Luo, L. (2015). A transcriptional reporter of intracellular Ca(2+) in Drosophila. Nat Neurosci 18: 917-925. PubMed ID: 25961791
Gendron, C. M., Chakraborty, T. S., Duran, C., Dono, T. and Pletcher, S. D. (2023). Ring neurons in the Drosophila central complex act as a rheostat for sensory modulation of aging. PLoS Biol 21(6): e3002149. PubMed ID: 37310911
Giraldo, Y. M., Leitch, K. J., Ros, I. G., Warren, T. L., Weir, P. T. and Dickinson, M. H. (2018). Sun navigation requires compass neurons in Drosophila. Curr Biol 28(17):2845-2852. PubMed ID: 30174187
Grabowska, M. J., Jeans, R., Steeves, J. and van Swinderen, B. (2020). Oscillations in the central brain of Drosophila are phase locked to attended visual features. Proc Natl Acad Sci U S A. PubMed ID: 33177231
Green, J., Adachi, A., Shah, K. K., Hirokawa, J. D., Magani, P. S. and Maimon, G. (2017). A neural circuit architecture for angular integration in Drosophila. Nature 546(7656): 101-106. PubMed ID: 28538731
Green, J., Vijayan, V., Mussells Pires, P., Adachi, A. and Maimon, G. (2019). A neural heading estimate is compared with an internal goal to guide oriented navigation. Nat Neurosci. PubMed ID: 31332373
Grover, D., Chen, J. Y., Xie, J., Li, J., Changeux, J. P. and Greenspan, R. J. (2022). Differential mechanisms underlie trace and delay conditioning in Drosophila. Nature 603(7900): 302-308. PubMed ID: 35173333
Hamid, A., Gattuso, H., Caglar, A. N., Pillai, M., Steele, T., Gonzalez, A., Nagel, K. and Syed, M. H. (2023). The RNA-binding protein, Imp specifies olfactory navigation circuitry and behavior in Drosophila. bioRxiv. PubMed ID: 37398350
Han, R., Huang, H. P., Chuang, C. L., Yen, H. H., Kao, W. T., Chang, H. Y. and Lo, C. C. (2021). Coordination through Inhibition: Control of Stabilizing and Updating Circuits in Spatial Orientation Working Memory. eNeuro 8(5). PubMed ID: 34385152
Hardcastle, B. J., Omoto, J. J., Kandimalla, P., Nguyen, B. M., Keleş, M. F., Boyd, N. K., Hartenstein, V. and Frye, M. A. (2021). A visual pathway for skylight polarization processing in Drosophila. Elife 10. PubMed ID: 33755020
Homberg, U., Kirchner, M., Kowalewski, K., Pitz, V., Kinoshita, M., Kern, M. and Seyfarth, J. (2023). Comparative morphology of serotonin-immunoreactive neurons innervating the central complex in the brain of dicondylian insects. J Comp Neurol 531(14): 1482-1508. PubMed ID: 37478205
Hu, W., Peng, Y., Sun, J., Zhang, F., Zhang, X., Wang, L., Li, Q. and Zhong, Y. (2018). Fan-shaped body neurons in the Drosophila brain regulate both innate and conditioned nociceptive avoidance. Cell Rep 24(6): 1573-1584. PubMed ID: 30089267
Hulse, B. K., Haberkern, H., Franconville, R., Turner-Evans, D. B., Takemura, S. Y., Wolff, T., Noorman, M., Dreher, M., Dan, C., Parekh, R., Hermundstad, A. M., Rubin, G. M. and Jayaraman, V. (2021). A connectome of the Drosophila central complex reveals network motifs suitable for flexible navigation and context-dependent action selection. Elife 10. PubMed ID: 34696823
Hwang, H. J. and Rulifson, E. (2011). Serial specification of diverse neuroblast identities from a neurogenic placode by Notch and Egfr signaling. Development 138(14): 2883-2893. PubMed ID: 21653613
Franconville, R., Beron, C. and Jayaraman, V. (2018). Building a functional connectome of the Drosophila central complex. Elife 7. PubMed ID: 30124430
Jones, J. D., Holder, B. L., Eiken, K. R., Vogt, A., Velarde, A. I., Elder, A. J., McEllin, J. A. and Dissel, S. (2023). Regulation of sleep by cholinergic neurons located outside the central brain in Drosophila. PLoS Biol 21(3): e3002012. PubMed ID: 36862736
Kakaria, K. S. and de Bivort, B. L. (2017). Ring Attractor Dynamics Emerge from a Spiking Model of the Entire Protocerebral Bridge. Front Behav Neurosci 11: 8. PubMed ID: 28261066
Kandimalla, P., Omoto, J. J., Hong, E. J. and Hartenstein, V. (2023). Lineages to circuits: the developmental and evolutionary architecture of information channels into the central complex. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. PubMed ID: 36932234
Kato, Y. S., Tomita, J. and Kume, K. (2022). Interneurons of fan-shaped body promote arousal in Drosophila. PLoS One 17(11): e0277918. PubMed ID: 36409701
Ki, Y. and Lim, C. (2019). Sleep-promoting effects of threonine link amino acid metabolism in Drosophila neuron to GABAergic control of sleep drive. Elife 8. PubMed ID: 31313987
Kim, S. S., Rouault, H., Druckmann, S. and Jayaraman, V. (2017). Ring attractor dynamics in the Drosophila central brain. Science 356(6340): 849-853. PubMed ID: 28473639
Kim, S. S., Hermundstad, A. M., Romani, S., Abbott, L. F. and Jayaraman, V. (2019). Generation of stable heading representations in diverse visual scenes. Nature 576(7785): 126-131. PubMed ID: 31748750
Krause, T., Spindler, L., Poeck, B. and Strauss, R. (2019). Drosophila acquires a long-lasting body-size memory from visual feedback. Curr Biol. PubMed ID: 31104933
Kumar, A., Bello, B. and Reichert, H. (2009). Lineage-specific cell death in postembryonic brain development of Drosophila. Development 136(20): 3433-3442. PubMed ID: 19762424
Kuntz, S., Poeck, B., Sokolowski, M. B. and Strauss, R. (2012). The visual orientation memory of Drosophila requires Foraging (PKG) upstream of Ignorant (RSK2) in ring neurons of the central complex. Learn Mem 19: 337-340. PubMed ID: 22815538
Lin, C. Y., Chuang, C. C., Hua, T. E., Chen, C. C., Dickson, B. J., Greenspan, R. J. and Chiang, A. S. (2013). A comprehensive wiring diagram of the protocerebral bridge for visual information processing in the Drosophila brain. Cell Rep 3: 1739-1753. PubMed ID: 23707064
Liu, C., Meng, Z., Wiggin, T. D., Yu, J., Reed, M. L., Guo, F., Zhang, Y., Rosbash, M. and Griffith, L. C. (2019). A serotonin-modulated circuit controls sleep architecture to regulate cognitive function independent of total sleep in Drosophila. Curr Biol 29(21): 3635-3646. PubMed ID: 31668619
Liu, S., Liu, Q., Tabuchi, M. and Wu, M. N. (2016). Sleep drive is encoded by neural plastic changes in a dedicated circuit. Cell 165: 1347-1360. PubMed ID: 27212237
Lovick, J. K., Omoto, J. J., Ngo, K. T. and Hartenstein, V. (2017). Development of the anterior visual input pathway to the Drosophila central complex. J Comp Neurol. PubMed ID: 28675433
Lyu, C., Abbott, L. F. and Maimon, G. (2022). Building an allocentric travelling direction signal via vector computation. Nature 601(7891): 92-97. PubMed ID: 34912112
Lyu, S., Terao, N., Nakashima, H., Itoh, M. and Tonoki, A. (2023). Neuropeptide diuretic hormone 31 mediates memory and sleep via distinct neural pathways in Drosophila. Neurosci Res. PubMed ID: 36780946
Musso, P. Y., Junca, P. and Gordon, M. D. (2021). A neural circuit linking two sugar sensors regulates satiety-dependent fructose drive in Drosophila. Sci Adv 7(49): eabj0186. PubMed ID: 34851668
Ofstad, T. A., Zuker, C. S. and Reiser, M. B. (2011). Visual place learning in Drosophila melanogaster. Nature 474: 204-207. PubMed ID: 21654803
Okubo, T. S., Patella, P., D'Alessandro, I. and Wilson, R. I. (2020). A neural network for wind-guided compass navigation. Neuron. PubMed ID: 32681825
Omoto, J. J., Keles, M. F., Nguyen, B. M., Bolanos, C., Lovick, J. K., Frye, M. A. and Hartenstein, V. (2017). Visual input to the Drosophila central complex by developmentally and functionally distinct neuronal populations. Curr Biol 27(8): 1098-1110. PubMed ID: 28366740
Omoto, J. J., Nguyen, B. M., Kandimalla, P., Lovick, J. K., Donlea, J. M. and Hartenstein, V. (2018). Neuronal constituents and putative interactions within the Drosophila ellipsoid body neuropil. Front Neural Circuits 12: 103. PubMed ID: 30546298
Palazzo, O., Rass, M. and Brembs, B. (2020). Identification of FoxP circuits involved in locomotion and object fixation in Drosophila. Open Biol 10(12): 200295. PubMed ID: 33321059
Pan, Y., Zhou, Y., Guo, C., Gong, H., Gong, Z. and Liu, L. (2009). Differential roles of the fan-shaped body and the ellipsoid body in Drosophila visual pattern memory. Learn Mem 16: 289-295. PubMed ID: 19389914
Pimentel, D., Donlea, J.M., Talbot, C.B., Song, S.M., Thurston, A.J. and Miesenbock, G. (2016). Operation of a homeostatic sleep switch. Nature 536(7616):333-7. PubMed ID: 27487216
Pisokas, I., Heinze, S. and Webb, B. (2020). The head direction circuit of two insect species. Elife 9. PubMed ID: 32628112
Robinson, B. S., Norman-Tenazas, R., Cervantes, M., Symonette, D., Johnson, E. C., Joyce, J., Rivlin, P. K., Hwang, G. M., Zhang, K. and Gray-Roncal, W. (2022). Online learning for orientation estimation during translation in an insect ring attractor network. Sci Rep 12(1): 3210. PubMed ID: 35217679
Sareen, P. F., McCurdy, L. Y. and Nitabach, M. N. (2021). A neuronal ensemble encoding adaptive choice during sensory conflict in Drosophila. Nat Commun. 12(1):4131. PubMed ID: 34226544
Scaplen, K. M., Talay, M., Fisher, J. D., Cohn, R., Sorkac, A., Aso, Y., Barnea, G. and Kaun, K. R. (2021). Transsynaptic mapping of Drosophila mushroom body output neurons. Elife 10. PubMed ID: 33570489
Schwarz, J. E., King, A. N., Hsu, C. T., Barber, A. F. and Sehgal, A. (2021). Hugin (+) neurons provide a link between sleep homeostat and circadian clock neurons. Proc Natl Acad Sci U S A 118(47). PubMed ID: 34782479
Seelig, J. D. and Jayaraman, V. (2013). Feature detection and orientation tuning in the Drosophila central complex. Nature 503(7475): 262-266. PubMed ID: 24107996
Seelig, J. D. and Jayaraman, V. (2015). Neural dynamics for landmark orientation and angular path integration. Nature 521(7551): 186-191. PubMed ID: 25971509
Shiozaki, H. M., Ohta, K. and Kazama, H. (2020). A multi-regional network encoding heading and steering maneuvers in Drosophila. Neuron 106(1):126-141. PubMed ID: 32023429
Sonn, J. Y., Lee, J., Sung, M. K., Ri, H., Choi, J. K., Lim, C. and Choe, J. (2018). Serine metabolism in the brain regulates starvation-induced sleep suppression in Drosophila melanogaster. Proc Natl Acad Sci U S A 115(27): 7129-7134. PubMed ID: 29915051
Stone, T., Webb, B., Adden, A., Weddig, N. B., Honkanen, A., Templin, R., Wcislo, W., Scimeca, L., Warrant, E. and Heinze, S. (2017). An anatomically constrained model for path integration in the bee brain. Curr Biol 27(20): 3069-3085 e3011. PubMed ID: 28988858
Sun, Y., Nern, A., Franconville, R., Dana, H., Schreiter, E. R., Looger, L. L., Svoboda, K., Kim, D. S., Hermundstad, A. M. and Jayaraman, V. (2017). Neural signatures of dynamic stimulus selection in Drosophila. Nat Neurosci [Epub ahead of print]. PubMed ID: 28604683
Tai, C. Y., Chin, A. L. and Chiang, A. S. (2020). A Comprehensive Map of Visual Projection Neurons for Processing Ultraviolet Information in the Drosophila Brain. J Comp Neurol. PubMed ID: 33174208
Tainton-Heap, L. A. L., Kirszenblat, L. C., Notaras, E. T., Grabowska, M. J., Jeans, R., Feng, K., Shaw, P. J. and van Swinderen, B. (2020). A Paradoxical Kind of Sleep in Drosophila melanogaster. Curr Biol. PubMed ID: 33238155
Takahashi, N., Zittrell, F., Hensgen, R. and Homberg, U. (2022). Receptive field structures for two celestial compass cues at the input stage of the central complex in the locust brain. J Exp Biol. PubMed ID: 35048987
Timaeus, L., Geid, L., Sancer, G., Wernet, M. F. and Hummel, T. (2020). Parallel visual pathways with topographic versus nontopographic organization connect the Drosophila eyes to the central brain. iScience 23(10): 101590. PubMed ID: 33205011
Tomita, J., Ban, G., Kato, Y. S. and Kume, K. (2021). Protocerebral Bridge Neurons That Regulate Sleep in Drosophila melanogaster. Front Neurosci 15: 647117. PubMed ID: 34720844
Turner-Evans, D., Wegener, S., Rouault, H., Franconville, R., Wolff, T., Seelig, J. D., Druckmann, S. and Jayaraman, V. (2017). Angular velocity integration in a fly heading circuit. Elife 6. PubMed ID: 28530551
Turner-Evans, D. B., Jensen, K. T., Ali, S., Paterson, T., Sheridan, A., Ray, R. P., Wolff, T., Lauritzen, J. S., Rubin, G. M., Bock, D. D. and Jayaraman, V. (2020). The neuroanatomical ultrastructure and function of a biological ring attractor. Neuron. PubMed ID: 32916090
Walsh, K. T. and Doe, C. Q. (2017). Drosophila embryonic type II neuroblasts: origin, temporal patterning, and contribution to the adult central complex. Development 144: 4552-4562. PubMed ID: 29158446
Weir, P.T. and Dickinson, M.H. (2015). Functional divisions for visual processing in the central brain of flying Drosophila. Proc Natl Acad Sci U S A 112(40):E5523-32. PubMed ID: 26324910
Wolff, T., Iyer, N. A. and Rubin, G. M. (2014). Neuroarchitecture and neuroanatomy of the Drosophila central complex: A GAL4-based dissection of protocerebral bridge neurons and circuits. J Comp Neurol 523(7):997-1037. PubMed ID: 25380328
Wolff, T. and Rubin, G. M. (2018). Neuroarchitecture of the Drosophila central complex: A catalog of nodulus and asymmetrical body neurons and a revision of the protocerebral bridge catalog. J Comp Neurol. PubMed ID: 30084503
Yan, W., Lin, H., Yu, J., Wiggin, T. D., Wu, L., Meng, Z., Liu, C. and Griffith, L. C. (2023). Subtype-Specific Roles of Ellipsoid Body Ring Neurons in Sleep Regulation in Drosophila. J Neurosci 43(5): 764-786. PubMed ID: 36535771
Zhang, J., Tanenhaus, A. K., Davis, J. C., Hanlon, B. M. and Yin, J. C. (2014). Spatio-temporal in vivo recording of dCREB2 dynamics in Drosophila long-term memory processing. Neurobiol Learn Mem 118C: 80-88. PubMed ID: 25460038
Zhang, J. M., Chen, M. J., He, J. H., Li, Y. P., Li, Z. C., Ye, Z. J., Bao, Y. H., Huang, B. J., Zhang, W. J., Kwan, P., Mao, Y. L. and Qiao, J. D. (2022). Ketone Body Rescued Seizure Behavior of LRP1 Deficiency in Drosophila by Modulating Glutamate Transport. J Mol Neurosci. PubMed ID: 35668313
Zhang, Y., Liu, G., Yan, J., Zhang, Y., Li, B. and Cai, D. (2015). Metabolic learning and memory formation by the brain influence systemic metabolic homeostasis. Nat Commun 6: 6704. PubMed ID: 25848677
Zhang, Z., Li, X., Guo, J., Li, Y. and Guo, A. (2013). Two clusters of GABAergic ellipsoid body neurons modulate olfactory labile memory in Drosophila. J Neurosci 33: 5175-5181. PubMed ID: 23516283
genes expressed in brain morphogenesis Genes involved in organ development
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
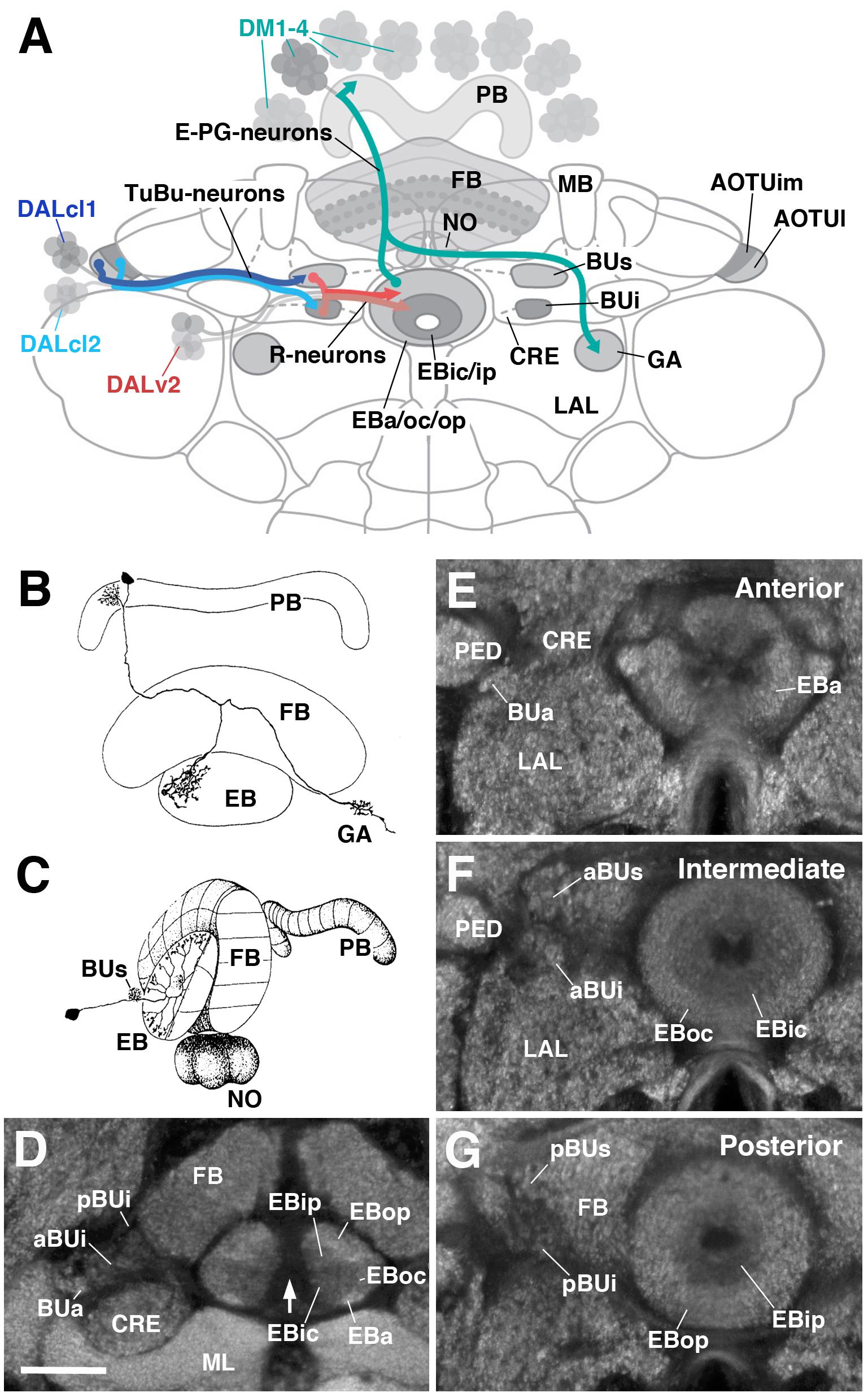{kind=link}
{kind=link}