by Lewis I. Held, Jr.
purchasing information
| Quirks of Human Anatomy by Lewis I. Held, Jr. | purchasing information |
| back to Quirks index page | |
Figure Legends 7 7.1 * 7.1R * A2.1 * A2.1R figure legends 1 * 2 * 3 * 4 * 5 * 6 N.B.: An 'R' suffix denotes reflections (commentaries, annotations, and further references) pertaining to the numbered legend that precedes it. [Select any image to enlarge; use back button to return] Fig. 7.1 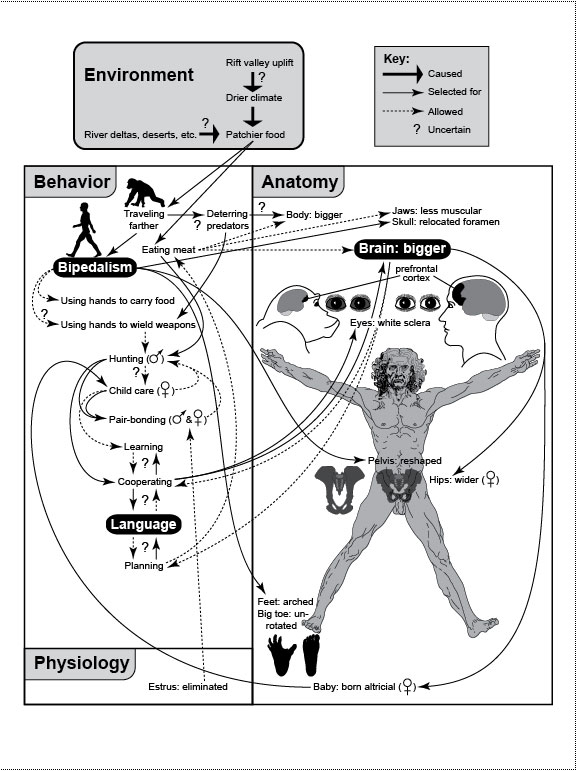 Selective forces that may have led to some important quirks that distinguish us from other apes—notably, bipedalism, language, and big brains. Question marks denote the most speculative links. Divergence of hominins from other apes apparently began when a change in climate nudged a population from an arboreal to a terrestrial lifestyle [2144]. Positive feedback loops may have driven the depicted outcomes [1708,2753]. Comparisons with chimps include our feet [1813], pelvis [786,1866], eyes, and brain [128,259,385]. Key for brain shading: black = prefrontal cortex [253,611]; dark gray = rest of cerebrum; light gray = cerebellum. The enlargement of our prefrontal cortex is important because it is the seat of rational decision making [249,2259] and voluntary action [115,2173,2515] (≈free will? [2413]). Its expansion may have been driven by the fibroblast growth factor FGF17 [449], although the extent of that increase is disputed [611,788,2181,2309], as is the degree to which it arose due to allometry alone [332]. Selective forces that may have led to some important quirks that distinguish us from other apes—notably, bipedalism, language, and big brains. Question marks denote the most speculative links. Divergence of hominins from other apes apparently began when a change in climate nudged a population from an arboreal to a terrestrial lifestyle [2144]. Positive feedback loops may have driven the depicted outcomes [1708,2753]. Comparisons with chimps include our feet [1813], pelvis [786,1866], eyes, and brain [128,259,385]. Key for brain shading: black = prefrontal cortex [253,611]; dark gray = rest of cerebrum; light gray = cerebellum. The enlargement of our prefrontal cortex is important because it is the seat of rational decision making [249,2259] and voluntary action [115,2173,2515] (≈free will? [2413]). Its expansion may have been driven by the fibroblast growth factor FGF17 [449], although the extent of that increase is disputed [611,788,2181,2309], as is the degree to which it arose due to allometry alone [332].Fig. 7.1R The phrase "selected for" (key) does not mean "caused" because the indicated agent is necessary but not sufficient without aptly conducive genetic changes to go along with it. Rather, it means "established a context where natural selection rewarded improvements along this line" [2753]. As for the genetics, how much neural rewiring was needed to convert a quadruped to a biped? Maybe not much [385,2730]: (1) a single mutation in an axon-guidance gene can make a mouse hop like a rabbit [1257] (cf. kangaroo rats [1351]), and (2) a human syndrome (due to one allele?) was found in Turkey where affected adults revert to walking on all fours [2564]! Genes that might have fostered the enlargement of our brain include ASPM, the nonsense alleles of which cause microcephaly [2907]; BF-1, which reduces forebrain size in mice when it is knocked out [59]; plus at least a dozen others [411,776,910,1927,2015]. The "Man the Hunter" scenario, although still viable [1120], has been roundly criticized [204,316,1119,2455], in part because (1) carnivory can involve scavenging instead of hunting [2457], and (2) females are the ones who hunt in bonobo society [609], and bonobos (Pan paniscus) are as close to us as chimps (Pan troglodytes) [608]. References to the depicted links include: Rift valley uplift –> ... patchier food [224,515]; river deltas, deserts, etc. –> patchier food [224,628,2849]; patchier food –> ... bipedalism [786,1522,1578] and carnivory (as a part of omnivory) [1522,1756,2456]; traveling farther (overland) –> ... bigger body [145,2456,2737]; bipedalism –> freed hands [42,607,938], relocated foramen magnum [10,2278], reshaped pelvis [1579], arched foot [1366], and unrotated big toe [956]; manipulation –> hunting [2286,2753]; carnivory –> bigger body [31,1756], bigger brain [59,789,1522], and lighter jaws [2014]; bigger brain <–> cooperating (i.e., society and culture) [255,2017]; bigger brain –> wider pelvis [525,1579,2753] and altricial childbirth [2456,2679,2737]; altricial childbirth –> greater infant dependency [956,1578,2220,2334]; carnivory/hunting cycle [2353,2455,2485] –> cooperating [1716,2170,2457]; cooperating –> white sclera [1398,1410,2515,2613]; division of labor (hunting vs. child care [341]) <–> pair-bonding [204,418]; child care –> learning [1476]; learning <–> cooperating [1476]; cooperating <–> language [689,2461]; language <–> planning [253,1476]; bigger brain –> planning [253]; and loss of estrus –> pair-bonding [316,1578,2002,2463]. Other links (not shown) include bigger body –> bigger brain [770] (via allometry [2181]); bipedalism –> raising our head so high off the ground that our sense of smell waned in value [389,701,2278] –> loss of olfactory genes [1882]; hunting with weapons –> less need for shearing teeth –> reduction in canine size [145,2753]; carnivory –> changes in gut anatomy and enzyme allocation [2310]; and monogamy (pair-bonding) –> larger brain size [2294]. Use of fire by hominins dates back 0.8 MYA or more [1186]. For rebuttals of aspects of this scheme, see refs. [228,406,628,2286,2576]. For skeletal comparisons (Homo vs. Australopithecus vs. Pan), see refs. [267,1522,2915]. For a picture of chimp feet that look hauntingly like human hands, see ref. [10]. Knuckle-walking chimp silhouette was adapted from ref. [1476]. Finally, it is important to dispel a widespread misinterpretation of diagrams like this. Yes, evolution entails problem solving (e.g., the problem of patchier food), but it is neither progressive nor teleological, and it is certainly not Lamarckian. Species do not solve ecological problems, although such wording is sometimes used as a convenient shorthand [2779]. Mutations and recombination do, but only in the same way that you might eventually find the door in a dark room by feeling your way along the wall. Fig. A2.1 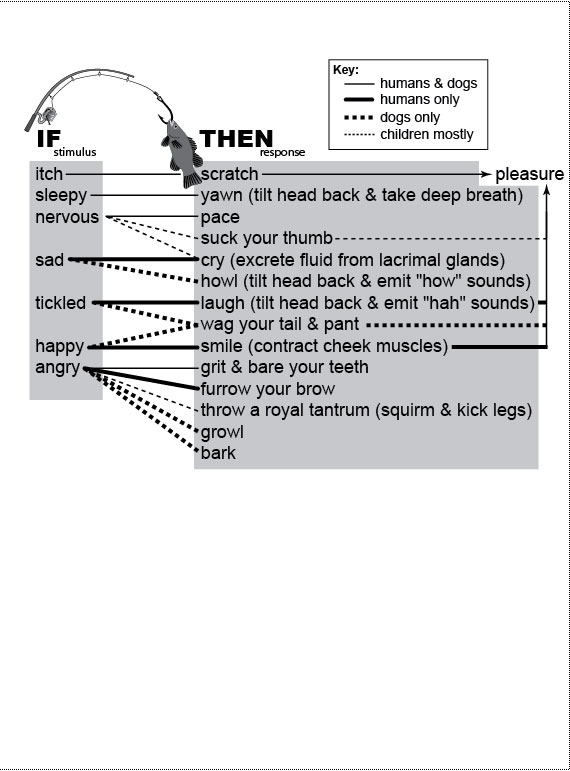 Whimsical sample of hardwired behaviors in humans versus dogs. The involuntary nature of these actions (e.g., laughing, crying, and yawning) is underscored by their retention despite loss of voluntary facial control in Anterior Operculum Syndrome [198,454,1494]. As implied by the fishing icon, many of the responses (right) seem to have been arbitrarily linked (neurally and genetically) to emotions (left), because they make little sense functionally [325,2050,2120]. For example, why should humans and dogs both be ticklish on our bellies [1097]? The only links that do seem reasonable are the baring of teeth and barking in response to anger, which serve as threat displays. Whimsical sample of hardwired behaviors in humans versus dogs. The involuntary nature of these actions (e.g., laughing, crying, and yawning) is underscored by their retention despite loss of voluntary facial control in Anterior Operculum Syndrome [198,454,1494]. As implied by the fishing icon, many of the responses (right) seem to have been arbitrarily linked (neurally and genetically) to emotions (left), because they make little sense functionally [325,2050,2120]. For example, why should humans and dogs both be ticklish on our bellies [1097]? The only links that do seem reasonable are the baring of teeth and barking in response to anger, which serve as threat displays.Fig. A2.1R The aim of this figure is the same as that of Jonathan Swift’s Gulliver’s Travels: to reveal the lunacy of behaviors we take for granted as mundane. By contrasting humans with dogs, we can see the symmetry of the absurdity: they look as silly to us as we must look to them. If they could laugh (and gossip), they might be snickering behind our backs. This "switchboard" is based on Darwin’s Expression of the Emotions in Man and Animals [562], on his Variation of Animals and Plants Under Domestication [560], on reflex physiology [1037,1270,1490], and on child psychology [314,2150,2894]. The schematic is not universal because there are so many deviations from the norm. For example, some people break out in hives when they get angry (not shown), and behavior varies tremendously among dog breeds [1312]. Dogs are an ideal foil for exposing our foibles because we are so familiar with them, and they share so many of our social inclinations [1742]. Tail-wagging exhibits bizarre asymmetries [2104,2357]. Darwin was struck by the vestigial nature of a few dog habits, which will be comically familiar to any dog lover: Dogs, when they wish to go to sleep on a carpet or other hard surface, generally turn round and round and scratch the ground with their fore-paws in a senseless manner, as if they intended to trample down the grass and scoop out a hollow, as no doubt their wild parents did, when they lived on open grassy plains or in the woods. ... Dogs after voiding their excrement often make with all four feet a few scratches backwards, even on a bare stone pavement, as if for the purpose of covering up their excrement with earth, in nearly the same manner as do cats. [562] (pp. 42ff.) Darwin was also fascinated with the novel quality of dog barking because it has no obvious precedent among wild wolves. The habit of barking, however, which is almost universal with domesticated dogs, forms an exception, as it does not characterize a single natural species of the family. [562] (p. 27) No one knows how barking evolved, except to observe that it arose during domestication [1489,1742,1806]. How odd that by tugging on one "strand" of the genomic "spiderweb" (domestication), we should elicit such a surprising change in a radically different part of the web (barking)! An even more stunning instance of network integration was seen during the recent domestication of foxes [338,1093,2635]: a suite of anatomical traits (floppy ears, short tails, etc.) emerged as a result of the artificial selection for behavior! Such counterintuitive results make one wonder how many of our own quirks might of arisen likewise as side effects of selection for adaptive traits (cf. Fig. 7.1). Stimulus-response links may be easier to revamp than we might think. For example, a recessive mutation puts Doberman pinschers to sleep instantly when they’re excited [2386], and a missense mutation paralyzes goats instantly when they’re startled [159]. In humans, this condition, known as "hyperekplexia" [1110], can be fatal due to its associated apnea. Analogous mutations might have led to the tactic of "playing dead" employed by possums and other animals. Another bizarre link, between a spasmotic trigger and the uttering of curse words [1456], afflicts nearly 1% of us [1263] in the form of Tourette’s Syndrome [1820]. Other mutations that drastically change behavior include (1) miffy, which makes mice hop like rabbits [1257], (2) Hoxb8null, which makes mice groom themselves and their cagemates to the point of bleeding [1006,1010], and (3) a myostatin mutation that enhances the racing ability of whippets and turns them into übermuscular "superdogs" [1510,1816]. In the fly world, the weirdest example may be the ability to turn normal males into homosexuals by overexpressing a pigmentation gene [2908]. By genetically linking a perception, response, or action to the pleasure centers of the brain [1158,1503,2715], natural selection can radically reconfigure the reward-seeking behavior of a species [178,319,1070,1643]. The epitome is orgasm [1488,2068], which links ecstasy to intercourse and compels us to reproduce [1417,2308,2428]. A more mundane example is the itch-scratch-pleasure chain shown here [75,1242,1747,2528], which sadly goes terribly awry in some people [901]. Such "aah that feels good!" reinforcement help explain (1) why cognition is so often overruled by emotion, despite our supposedly exalted intellect [253,667,1328,1643] and (2) why so many people get trapped in self-destructive spirals of obsession, compulsion, and addiction [415,901,1005]. (Just visit a casino in Las Vegas!) Subtler polymorphisms in these links are likely responsible for the inexplicable idiosyncrasies of tastes and temperaments [1127,2677]—for example, the "search images" that guide mate choice (e.g., What does she see in him?) [1827,2165,2236,2537,2791], although some of that imagery may be imprinted during infancy [1363]. A related question concerns the degree to which fear is innately linked to the perception of certain shapes or movements. For example, primates appear to have a hardwired fear of snakes, which makes sense given our arboreal origins [1250], but it begs the deeper question of how evolution was able to program our brains thusly (cf. [2274]). What other old circuits still steer our behavior [1412,1555,2091,2254]? No matter how many there may be, we cannot justifiably claim that we’re just pawns of our genes [1543,1835,2169]. top of page
Lewis I. Held, Jr. is Associate Professor in the Department of Biology at Texas Tech University.
|
 © 2009 Thomas B. Brody, Ph.D. |