by Lewis I. Held, Jr.
purchasing information
| Quirks of Human Anatomy by Lewis I. Held, Jr. | purchasing information |
| back to Quirks index page | |
Figure Legends 6 6.1 * 6.1R * 6.2 * 6.2R * 6.3 * 6.3R figure legends 1 * 2 * 3 * 4 * 5 * 7 * A N.B.: An 'R' suffix denotes reflections (commentaries, annotations, and further references) pertaining to the numbered legend that precedes it. [Select any image to enlarge; use back button to return] Fig. 6.1 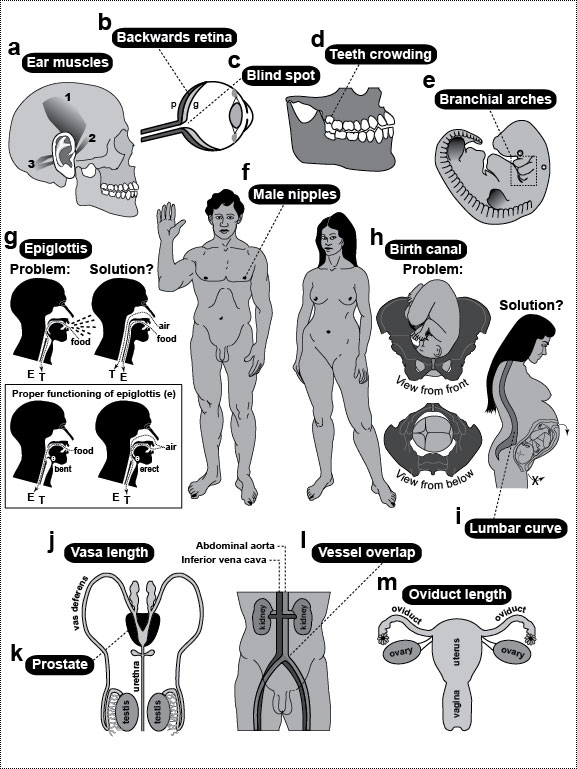 Rogues’ gallery of human anatomical flaws, ranging from the merely silly (a, f, j), to the patently stupid (b, c, d, e, i, k), to the potentially lethal (g, h, l, m). See Table 6.1 for further information. Rogues’ gallery of human anatomical flaws, ranging from the merely silly (a, f, j), to the patently stupid (b, c, d, e, i, k), to the potentially lethal (g, h, l, m). See Table 6.1 for further information.
a. Muscles (dark bands) that permit ear wiggling, a dubious talent that has no apparent function aside from entertaining young children: (1) superior, (2) anterior, and (3) posterior auricularis [1004,1488]. b. Eyeball with optic nerve. The retina is considered to be inside out because its photoreceptors (p) lie behind its ganglion cells (g), an orientation we may have gotten stuck with early in evolution (cf. Fig. 6.2). Since then, the photoreceptors have come to rely on the (outer) layer of pigment cells (not shown) [2098], so it may now be impossible to reverse this order. Thickness of retinal layers is exaggerated. c. Blind spot where ganglion cell axons exit the eye. This suboptimal escape route is due to the inverted geometry shown in b and explained in Fig. 6.2. The hole in the retina is sizeable (~9 full moons in the sky), but we don’t notice it because (1) the blind spots of the two eyes are in separate parts of the visual field and (2) our brain automatically fills in gaps in our visual field by interpolation [2117]. d. The common crowding of human teeth—especially "wisdom" teeth, which erupt last—is traceable to the evolutionary shortening of our jaw. See text. e. Human embryo at 31 days postfertilization. Branchial arches (in dashed box) are a vestige from our fish ancestors. They now serve only as precursors for structures which could develop without such clefts (cf. Ch. 4). f. Nipples are useless in human males (cf. Ch. 5). g. Choking (upper left) is caused by malfunctioning of the epiglottis. The epiglottis is a cartilaginous flap (e) that reflexively covers the trachea (T) (lower box) when we swallow so that food or drink goes to the esophagus (E). The silhouette is a sagittal section with cavities as blank areas [612,889,2502,2737]. The line between E and T represents their adjacent lumenal walls [685]. One way to prevent choking (upper right) would be to have separate pipes for breathing and swallowing. In fact, that configuration already exists in babies, who can suckle and nose breathe at the same time: their larynx is so high that the epiglottis can touch the base of the nasal cavity at the midline [1547], with milk flowing around the juncture (see ref. [1469] for a clear diagram). The same is true for other mammals, who rest their epiglottis on the back edge of the soft palate [889,1546,1547]. The problem with having air bypass the mouth entirely, of course, is that it precludes talking [1931], although toothed whales still manage to "sing" to each other via an asymmetric specialization of their nasal passages [1718,2412]. h. Childbirth involves a tight fit between the baby’s head and the mother’s pelvic opening (cf. Fig. 7.1) [525]. Indeed, the passageway is so confining that the head must turn as shown (long axis transverse) to enter the birth canal, but then must rotate 90 degrees (long axis sagittal) to exit and let the shoulders enter [10,2220]. The process would be safer (and less painful) if the path mimicked a caesarean section, with the baby exiting through the navel. Drawn on the baby’s head are the fontanels (open sutures) that allow the skull to deform during birth [500,685]. i. Back pain often occurs during pregnancy because of strains in the lumbar region (cf. Fig. 5.1). More serious repercussions of the spine’s sinusoidal shape include herniated disks [10]. j. The vasa deferentia are tubes that conduct sperm from the testes to the urethra. They are much longer than they need to be in humans because of the circuitous route that they took evolutionarily [2384]. k. Routing a tube like the urethra through a solid organ like the prostate runs the risk of strangulation if the organ overgrows—a condition all too common in older men. l. In May-Thurner Syndrome, blood clots typically arise in the left—but not right—leg [196,320,1683]. This peculiar ischemic asymmetry is due to a normal quirk. The abdominal aorta and the inferior vena cava travel side by side to the groin. Just after they split into the two common iliac arteries and the two common iliac veins (one artery and one vein routed to each leg), the right fork of the arteries passes over the left fork of the veins. This overlap compresses the left vein between the spine and the overlying artery, resulting in occasional thrombosis. There is no such crossover on the right side. m. After ovulation, the egg leaves the ovary and enters the flower-shaped opening of the adjacent oviduct. Because fertilization occurs in the oviduct, the embryo can occasionally stick to the oviduct wall before it ever gets to the uterus, resulting in a life-threatening "ectopic" or "tubal" pregnancy [91]. It would have been safer if evolution had kept the length of the oviduct to a bare minimum or done away with it altogether and connected the ovary directly to the uterus. Fig. 6.1R The full-length man and woman are redrawn from a plaque aboard Pioneer 10—the first human-made object ever to leave our solar system [2253]. Launched in 1972, this human-sized spacecraft whipped past Jupiter (picking up speed), crossed Pluto’s orbit, and began coasting toward Aldebaran, which it should reach in ~2 million years [2125]. The plaque was meant as an interstellar "postcard." If any aliens stumble upon it, they will surely chuckle at our quirks (if they can laugh), although turnabout is fair play, and if we were to see their two sexes (or however many they have) au naturel, we might chortle right back at them. The standing humans were drawn by Linda Salzman in collaboration with astronomers Carl Sagan (her then-husband) and Frank Drake [573]. Choking diagrams (g) are adapted from refs. [1866,2331], and childbirth sketches (h) are redrawn from refs. [1579,1786,1866], except for the profile of the gravid woman, which is based on photos of harpist Cheryl Gallagher from her musical compact disc Pregnant Pause. g. As explained in the text, choking became a problem when our lungs arose as a branch of our eating tube. François Jacob ascribed such flaws to the trial-and-error nature of the evolutionary process [1265]: "To make a lung with a piece of esophagus sounds very much like tinkering." There is also a deeper question here: why must people die when deprived of oxygen? Based on the prevalence of hibernation, estivation, and facultative anaerobiosis among animals [76,1675,2505], the answer is unclear [1171]. Sea turtles, for example, can hold their breath for at least three hours [1600]. Evolution, it would seem, could have given us the means to survive episodes of choking, drowning, or suffocation [1316,2229]. Why didn’t it? Presumably, the rarity of asphyxiation among primates (by drowning, etc.) reduced the marginal advantage that any salvational mutations might have had to a negligible level. The greatest irony about oxygen is that it used to be poisonous for living things before ~2 billion years ago [468,774], but eukaryotes evolved ways to detoxify and harness it to our metabolism so that it has become vital for us [12,152,2128]. This abrupt reversal of fortune was as dramatic as any Shakespearean plot. h. While we are decrying the pain of childbirth, we should also pity the poor kiwi, whose egg comprises 25% of her body weight [915], for she, too, labors mightily [2549]. Some quirk of ratite history (as yet unknown [342,343,2071]) doomed those birds to that lunacy [969]. The depicted solution for humans (i.e., birth through the navel) may seem farfetched, but the spotted hyena has rerouted its birth canal just as drastically: hyena pups must make a ~180-degree turn when they reach the mother’s pelvis and then exit through her clitoris (pseudopenis) [803]! The problem with this path is that the diameter of the clitoral meatus is too narrow (~2.2 cm) to pass the pup’s head (~6.5 cm), so the pain must be excruciating, which makes one wonder why hyenas laugh at all! Darwin made an interesting observation about our fontanels. He noted how lucky we are that these hinges were already in place (because of how skull bones grow) before they acquired the function of allowing our skull to deform during the tight squeeze of the birth process. In other words, mammalian sutures were "co-opted" as hominin hinges: The sutures in the skulls of young mammals have been advanced as a beautiful adaptation for aiding parturition ... but as sutures occur in the skulls of young birds and reptiles, which have only to escape from a broken egg, we may infer that this structure has arisen from the laws of growth, and has been taken advantage of in the parturition of the higher animals. [559] (p. 197) Fig. 6.2 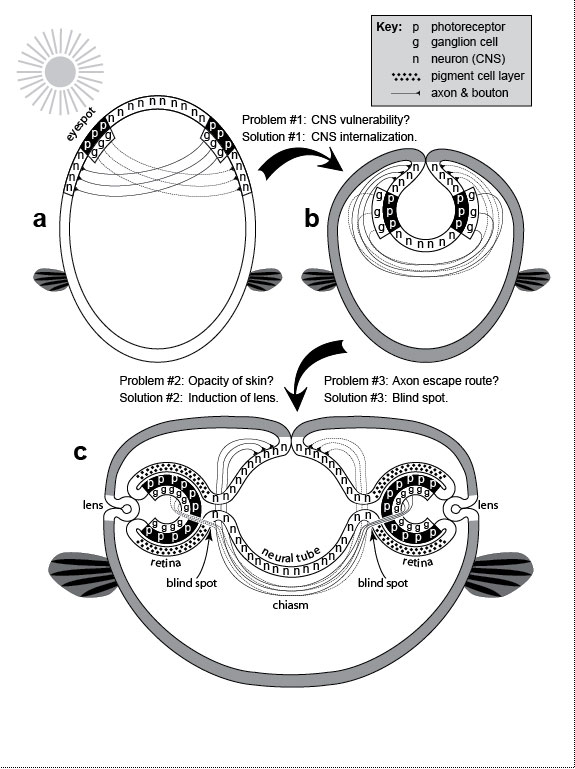 Hypothetical (bislagiatt) explanation for two flaws of human eyes: our backward retina and our blind spot. ("Bislagiatt" stands for "But it seemed like a good idea at the time.") Hypothetical (bislagiatt) explanation for two flaws of human eyes: our backward retina and our blind spot. ("Bislagiatt" stands for "But it seemed like a good idea at the time.")This diagram incorporates the inversion scheme of Balfour (1881) [2736], the axonal inferences of Polyak (1957) [2061] and Sarnat and Netsky (1981) [2278], and the opacity conjecture of Walls (1942) [539,2736], all of which are based on the tendency of chordate development to recapitulate its evolution [956]. Cartoons are transverse sections. Fish fins (which evolved later) are added merely to orient the reader (dorsal up, ventral down). Notwithstanding the symbols in the key, photoreceptors (p) and ganglion cells (g) are also neurons (n), albeit part of the peripheral versus central nervous system (CNS). Cell size is greatly exaggerated. a. Our bilaterian (protochordate?) ancestor is thought to have had (1) a superficial nerve net [1458,1584,2579], (2) eyespots made of photoreceptors and ganglion cells [1461], and (3) criss-crossed wiring where ganglion cells projected axons to motor neurons (subset of n) on the opposite flank [2278]. The advantage of this contralateral wiring was that it allowed each eyespot to turn the body reflexively away from potential harm whenever it detected the shadow of a possible predator [2061,2278]. The problem with having neurons on the surface, of course, is that they could be easily damaged. To put it bluntly, it is not a good idea to "wear your brain on your sleeve"! b. One solution to this Vulnerability Problem was for the CNS to move inside. The internalization trick that evolved in our chordate ancestors was "neurulation" [1038]—an involution of the dorsal surface to form a tube [1778,2312]—seen as a circle here in cross-section. Our neural tube ultimately forms our brain and spinal cord. During the CNS invagination of our protochordate forebear, the eyespots must have been dragged along and hence turned inside out. Ever since, our clade has, sadly, been saddled with a backward retina. Although this reversal turned out to be a mistake in hindsight, the eyespots were still just shadow detectors (not image detectors) at that time, so lower resolution was not yet a problem. This optical system would have worked quite well as long as the skin stayed transparent. However, any skin darkening would have severely reduced the effectiveness of the eyespots, and selective pressure would thereby have compelled (1) the eyespots to move nearer the surface and (2) the skin at the contact site to become transparent [2736]. c. To solve this Opacity Problem (presumably), vertebrates evolved a transparent lens where the prospective retina contacts the surface [1308]. During our development, the retinal outgrowths are initially shaped like balloons [2155] but then collapse, as if burst, into two-layered cups [18]. The outer layer facing the CNS becomes a pigmented epithelium. At some point in our evolutionary history, the lens must have blocked the escape route of ganglion cell axons. To solve this Entrapment Problem, the axons exited through the back wall, hence burdening us with a blind spot in perpetuity. Only a few axons are sketched here to indicate the routing. Fig. 6.2R These stages suggest how easy it was for evolution to build an eye step by step [591,1478] because any upgrade in imaging would have conferred a significant advantage [1308,2384]. Darwin need not have fretted about the apparent challenge that the eye posed for his gradualistic theory [876]. For a schematic that nicely traces the layering of our retina all the way back to amphioxus, see ref. [59]. a. Surprisingly, all metazoans use virtually the same sensor in their photoreceptors [539,1478]. This gadget first evolved in prokaryotes [2449]. It relies on (1) the vitamin A derivative "retinal" (or a variant [932]) to absorb photons [1019,1723], (2) an "opsin" protein to monitor shape changes in retinal [176,1993,2288], and (3) a signal-relay chain of downstream effectors [787,1466,1971]. The evolutionary riddles posed by this universality are (1) how long did it take for prokaryotic genomes, by random mutation, to stumble upon an opsin-like protein that could cradle a chromophore and transduce its twitching [621,876,2044,2241]?, (2) why was 11-cis retinal recruited instead of some other photoactive agent [932,1847,2709]?, and (3) how did it get linked to a particular transduction pathway [763,2521]? Believe it or not, the following creatures actually exist [876,2628]: (1) a jellyfish larva with bona fide photoreceptors but no nervous system to process their output [1896], (2) a fish that uses chlorophyll as a chromophore [675], and (3) a single-celled dinoflagellate that sports a humanoid "eye," complete with lens, photosensitive membranes, and a pigment shield [2013]! This odd menagerie prompted one author to propose that the eye evolved before the brain [876]! b. In arthropods the nervous system arose evolutionarily from a superficial sheet of ectoderm [1779], as did ours [1463]. Strangely, however, arthropods are not all alike in how they solve the CNS Vulnerability Problem. Two classes, chelicerates and myiapods, undergo a variant version of neurulation from the ventral (vs. dorsal) surface [2494], but two other classes, insects and crustaceans, develop entirely differently [1778] (cf. other phyla [1678]). Instead of ectodermal origami, they "percolate" cells inside one by one to form their nerve cord [1104,1165,2495]. c. Fusions have been omitted to preserve the geometry of (1) neural invagination at the dorsal midline to form a tube and (2) lens invagination on the flank surface to form a sphere. The origami of eye development is different in other phyla [1478], which explains why, for example, the retinas of octopi and squid are not backward [1732]. Also omitted are (1) the cornea, which is relatively trivial histologically [2103]; (2) the neurons (horizontal, bipolar, and amacrine) that intervene between photoreceptors and ganglion cells [2597,2736]; and (3) the later stages of lens development wherein it abandons its hollow state to simulate a solid glass camera lens [548]. For decades the lens was thought to be induced in a one-step process by the optic cup [1692,2432], but we now know that the optic cup plays more of supporting role along with other actors [670,994,1487,2627]. The optic cups of one unfortunate frog must have taken a wrong turn because its eyes (lenses and all) wound up in the roof of its mouth [2211]: the frog could only see when it yawned! Inexplicably, our iris is the only muscle in the body that comes from ectoderm instead of mesoderm [191,584]. (How on earth did the ectoderm get the password to unlock the "muscle vault" in the genome?) Thus, when you gaze into your lover’s eyes (the iris in particular), you’re actually seeing a colorful part of his or her brain [120]. Even stranger is the fact that the iris, which normally develops independently of the lens, can regenerate a lens in newts if the lens is artificially removed [1125,1273,2639]. This ability must be a spandrel of how eye parts are wired in the genome, but we have no clue about how it evolved [344,1056]. One final quirk worth noting is that the iris is intrinsically photosensitive—that is, it constricts on its own [2641]. How it does so is unclear, although we do know that it detects light using some sort of cryptochrome rather than an opsin pigment. Fig. 6.3 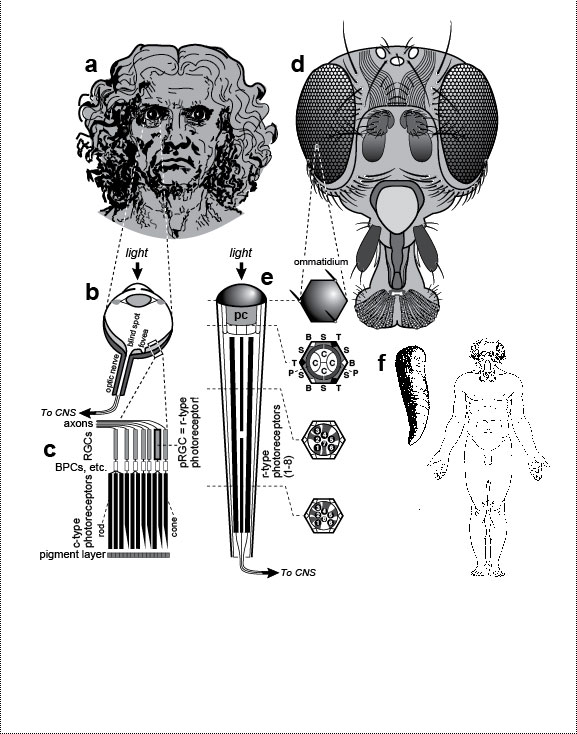 Eye anatomy in humans (a-c) versus flies (d, e). Eye anatomy in humans (a-c) versus flies (d, e). We have a simple eye; flies have a compound one. Human photoreceptors are mainly c-type (ciliary), whereas fly receptors are exclusively r-type (rhabdomeric). Recently (see text) it has been discovered that humans also have an r-type receptor in the guise of what looks like a garden-variety neuron (c). a. It would seem that humans have only two eyes, but a third one (or what’s left of it) resides inside our head as the pineal gland (cf. Ch. 3). b. Our eye works like a digital camera. Light is focused by the lens onto a screen (the retina), where it gets pixelated and converted into electrical signals. Those signals, in turn, are processed through various layers of the retina [2788] and sent to the brain via the optic nerve [2042,2098]. Acuity is greatest in the fovea. Axons exit nearby at a spot where we are blind. Abbreviation: CNS = central nervous system. c. Schematic of retinal stratification (layers not to scale). There are five main types of neurons in the retina [1679] among ~55 types total [1352]. Only three are shown: RGCs (retinal ganglion cells), BPCs (bipolar cells), and photoreceptors. (Horizontal and amacrine cells are omitted.) A small subset (~3%) of RGCs are photosensitive (pRGC) [552,798]. They are r-type receptors, but they lack the stacked membrane system that we associate with bona fide photoreceptors [89]. The pigment layer absorbs photons to prevent back-scatter [1478]. Note that multiple rods converge onto single RGCs (the ratio is actually ~20 to 1), whereas every cone typically has its own dedicated RGC [2098]. This difference is one reason our cone-rich fovea has such high acuity. d. Each of the fly’s compound eyes has ~750 simple eyes called "ommatidia" [1137]. Three separate simple eyes (white spots) are visible on top of the head: these "ocelli" mainly detect shadows [1776,1777]. e. The cuticular hexagonal dome serves as a cornea, whereas the pseudocone (pc) acts as a lens. The latter is a gelatinous cylinder secreted by four cone (C) cells (no relation to vertebrate cones). All but two of the eight photoreceptors (R cells) span the length of the ommatidium. As shown by cross-sections, R7 is above R8, an arrangement that enables R7 and R8 (which have orthogonal rhabdomere gratings) to detect polarized light [1090,2634]. Instead of using a pigment backscreen, flies wrap their photoreceptors in pigment tubes [1478] that are assembled like jigsaw puzzles from primary (P), secondary (S), and tertiary (T) pigment cells [1137]. Three of the six vertices of each hexagon are occupied by a bristle (B) instead. f. Reciprocal human-fly chimeras in a woodcut (1793) by William Blake (1757-1827). This drawing was published by Claudio Stern [2472] from the private collection of Blake biographer Geoffrey Keynes (C. D. Stern, personal communication). It may have been an informal study. Blake did use the chrysalis with a baby’s face in the frontispiece (captioned "What is Man!") for his book Gates of Passion (1793), but he never published the man with a fly’s head [199] (cf. blakearchive.org). Fig. 6.3R With the juxtaposition of these two faces we have, in essence, a family portrait. Humans and flies are basically twins separated at birth over half a billion years ago. In many ways we have come to look overtly different—for example, (1) we have an endoskeleton, whereas they have an exoskeleton; (2) we have four limbs, whereas they have six (not counting their wings); and (3) our eyes are simple, whereas theirs are compound [388,2923]. Yet despite these obvious disparities in gross anatomy, the same genetic gadgetry sculpts each of our respective structures during development [195,392], an abiding legacy of our common ancestry. Our bilaterian ancestor was considerably more primitive than either of these descendants [119,195,886,2663]. The fly portion of the diagram was adapted from ref. [1137]. c. Identities of the various retinal cell types are specified by a hierarchical combinatorial code [1084,1658,1819] that is time-dependent [616,1800,1920]. The regular spacing of the retinal ganglion cell (RGC) tiling pattern is achieved by ~80% apoptotic attrition [1550], homotypic lateral inhibition [705], and small-scale cell rearrangements [2145], without any need for homotypic contact [1568]. Nevertheless, homotypic contacts do play a role in RGC axon guidance [2042]. Instead of an absorptive pigment layer, cats and other nocturnal animals have a reflective tapetum that sends undetected photons back through the photoreceptors a second time [1478]. e. Despite what appears to be a solid-state dioptric apparatus, flies can adjust the amount of light reaching the photoreceptors by means of a myosin-operated "pupil" [2279]. For a primer on how fly eyes work, see refs. [1091,1815,1947]. (N.B. for fly aficionados: the upper cross-section is not the minimal "repeat unit" of standard textbook diagrams [2139]; instead, it shows the entire ring of cells encircling the photoreceptor core.) f. William Blake’s fascination with the human-fly metaphor was not confined to this woodcut. In Songs of Innocence and of Experience (1794), he had a poem called "The Fly": "Little fly, / thy summer’s play / my thoughtless hand / has brushed away. // Am not I / a fly like thee? / Or art not thou / a man like me? // For I dance / and drink & sing, / till some blind hand / shall brush my wing. // If thought is life / and strength & breath, / and the want / of thought is death, // then am I / a happy fly, / if I live, / or if I die." Reciprocal human-fly chimeras were also central to the classic 1958 horror/science-fiction movie The Fly starring Vincent Price. In the 1986 remake with Jeff Goldblum, a human-fly hybrid was featured instead. (Don’t let your kids see either movie!) top of page
Lewis I. Held, Jr. is Associate Professor in the Department of Biology at Texas Tech University.
|
 © 2009 Thomas B. Brody, Ph.D. |