by Lewis I. Held, Jr.
purchasing information
| Quirks of Human Anatomy by Lewis I. Held, Jr. | purchasing information |
| back to Quirks index page | |
Figure Legends 2 2.1 * 2.1R * 2.2 * 2.2R * 2.3 * 2.3R * 2.4 * 2.4R figure legends 1 * 3 * 4 * 5 * 6 * 7 * A N.B.: An 'R' suffix denotes reflections (commentaries, annotations, and further references) pertaining to the numbered legend that precedes it. Fig. 2.1 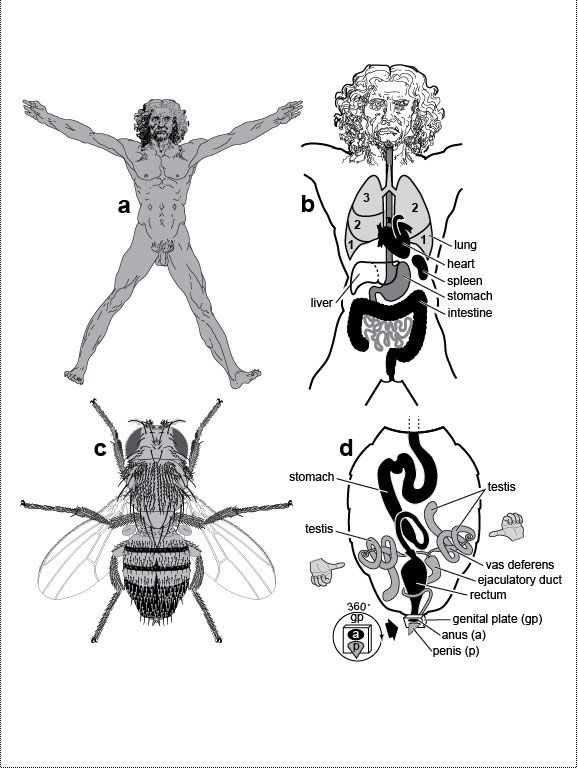 The paradox of external symmetry versus internal asymmetry, as exemplified by a human and a fly. The paradox of external symmetry versus internal asymmetry, as exemplified by a human and a fly.a. Vitruvian Man (by Leonardo da Vinci, ca 1492), a member of the phylum Chordata. b. Our right lung has three lobes but our left lung only two [256]. Our heart is shifted to left [308], our spleen is located on the left [322,2000], and our stomach bulges to the left, whereas our liver is shifted to the right [376,2311]. Our colon curls into a question mark, although its exact path can vary from person to person [187,1866,2828]. c. Vitruvian Fly (with apologies to Lenny). Male fruit fly (Drosophila melanogaster, ~2 mm long). Its phylum (Arthropoda) is only distantly related to ours (Chordata), but we share a common ancestor (cf. Fig. 1.3). d. The fly’s long stomach has constant loops (as does our gut) [528,1746]. What is really weird, though, are its testes, which form spirals. At first glance, the spirals look as if they might be symmetric, and in most fly species they are [2467], but in D. melanogaster, they both coil around their vas deferens like the fingers of a left hand curl around its thumb [1745,2467]. In other words, they are not mirror images! Rather, they’re like a man with two left hands. No one knows how this asymmetry is generated or why it exists. Another quirk is a 360-degree rotation of the genitals during development, which twists the ejaculatory duct sinistrally around the rectum [919,1216]. (See text for discussion.) Males can still mate as long as their genitals are not >20 degrees off kilter [250]. Accessory organs (paragonia and sperm pump) have been omitted (cf. Fig. 5.2). Fig. 2.1R Mutations can completely reverse the asymmetry of the viscera to a create a "situs inversus" phenotype (not shown) in both humans [321,397,1258,1711,2441] and flies [1216,2429]. Ambiguous intermediate states are also commonly found [1430,1482]. Drawings of human viscera are adapted from refs. [353,1646] and those of fly viscera are simplified from refs. [1745,1746]. Although fly and human guts appear quite different anatomically, they turn out to use the same genetic circuitry and stem-cell gadgetry to replenish their epithelial cell cohorts [2041,2558]. This homology adds to the growing list of shared bilaterian devices uncovered by evo-devo research [884] (cf. "Study Aids" in The Interactive Fly). About 1 in 10 of us are left-handed [1715,1808,2022], a frequency that has not changed in at least 10,000 years [522,750]. Curiously, handedness varies independently of visceral asymmetry: only ~10% of people with situs inversus are left-handed [1713]. Hence, these traits must be under separate genetic control [511,512,1630]. Indeed, the circuitry for left-right asymmetry must have multiple branchpoints [208,441,732,1528]. b. Mouse lungs are even more asymmetric than ours: they have four lobes on the right and only one on the left [1181]. Other human asymmetries (not shown) include the sinistral coiling of our umbilical cord vessels and the dextral coiling of the ducts of our gall bladder and sweat glands [510,2024]. (Do Schwann cells wrap axons with the same chirality, regardless of whether they’re on the left or right?) In some other mammals (e.g., elk and lemurs) the colon forms a perfect spiral with 6 or more gyres [2806]. Our most dangerous asymmetry is inside the heart where we make only one, unilateral pacemaker [1201]. We would be better off if we had a "backup" pacemaker on the other side that could take over in the event of a heart attack. The evolution of the vertebrate heart is an epic story that we are only now beginning to unravel morphologically [777,2062,2402] and genetically [438,481,582,1933,2280]. d. For clarity, the tip and coils of each testis are drawn on the same side of the abdomen (they really lie on opposite sides), the gyres of coils are spaced apart, and other organs (crop, Malpighian tubules, accessory glands, and ejaculatory bulb) are omitted. See Fig. 5.2 for a fuller rendering of the fly’s reproductive system. The discovery that myosin (vs. tubulin) is the key agent in fruit fly left-right asymmetry [1216,2429] offers hope that the upstream circuitry will yield its secrets [528]. However, our grasp of dipteran phylogeny is pathetically weak compared with what we know of chordates [1016], so it will be hard to decipher the order in which the genetic tinkering occurred. Fig. 2.2 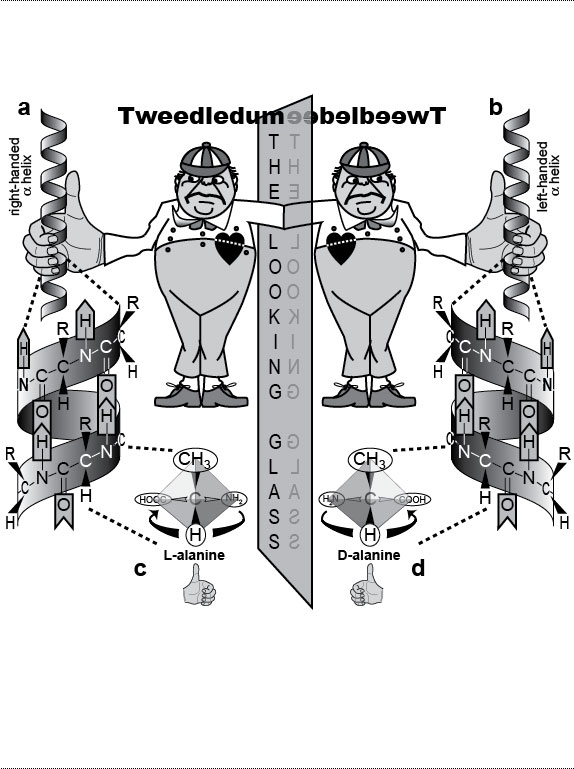 The geometric basis of molecular chirality. On the left of the mirror are molecules as they naturally occur in living things [495,1682]. On the right are their mirror images, which are rare in living things [827]. The geometric basis of molecular chirality. On the left of the mirror are molecules as they naturally occur in living things [495,1682]. On the right are their mirror images, which are rare in living things [827].
a, b. Proteins typically fold into right-handed α-helices (a), not left-handed ones (b) [852]. Handedness is conventionally defined by whichever thumb matches the ribbon direction traced by the curl of the fingers. Magnified section below shows the peptide backbone (N-C-C-N-C-C) and hydrogen bonds (between C=O and N-H groups) that bridge the gyres. Abbreviations: C = carbon; H = hydrogen; N = nitrogen; O = oxygen; R = reactive side group that varies among amino acids. c, d. Mirror-image stereoisomers of the amino acid alanine (R = CH3 group). Bonds are denoted by lines out of (black triangles) or into (hollow triangles) the page. The central carbon binds four constituents (vertices of tetrahedron) that can be arranged in either one of two ways [1682]. One isomer rotates polarized light to the left (L-alanine = levorotatory), the other to the right (D-alanine = dextrorotatory). Fig. 2.2R a, b. Other molecules that have a characteristic chirality, regardless of whether they are on the left or the right side of the body include DNA [1130], helicoid chitin fibers in the cuticle of insects [1870,1871], and helicoid collagen fibers in the corneal stroma of vertebrates [2629], and the same is true at a higher level for sister chromatids [254]. c, d. Living things use L-amino acids but D-sugars [495]. How this bias arose at the dawn of life has been much debated but never resolved [114,483]. If Earth’s history were rebooted, those early events could conceivably have culminated in humans with D-amino acids and L-sugars instead [865,1681]. Thus, it is possible, in the spirit of Lewis Carroll, to imagine a counterpart of Alice from that mirror-image universe. Let’s call her "Ecila." If Ecila were to encounter Alice, would they look and act alike? Perhaps. One thing is certain: they could not share a meal. The reason is clear. Enzymes catalyze reactions through a hand-in-glove fit with their substrates [287,1431,1731], so Alice’s food would not suit Ecila, nor vice versa. Indeed, Carroll addressed this problem in Through the Looking Glass: Alice opined to her cat that looking-glass milk might not be good to drink [380]. Fig. 2.3 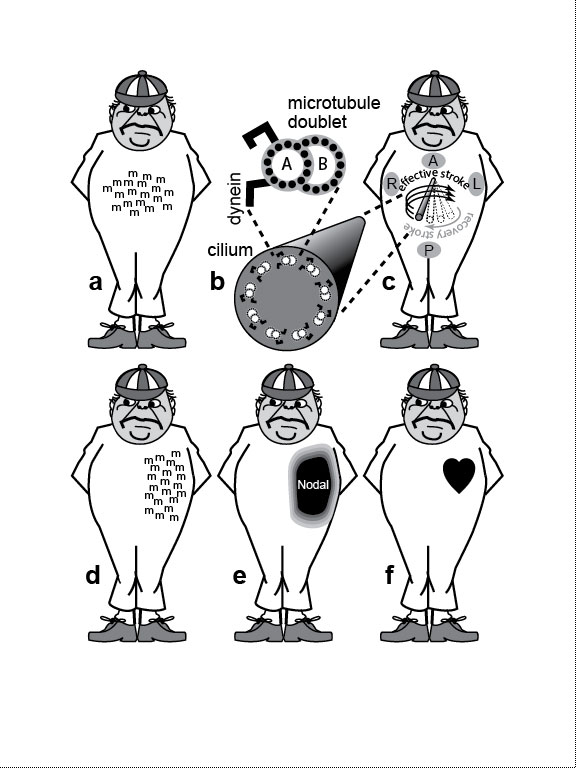 How molecular chirality dictates visceral asymmetry in mammals [1924]. Tweedledum represents a human embryo at the gastrula stage. How molecular chirality dictates visceral asymmetry in mammals [1924]. Tweedledum represents a human embryo at the gastrula stage.a. Signaling molecules termed "morphogens" (m) are secreted near the midline in an initially even distribution, left versus right. (Candidates for the actual signal, which is not yet known [1898,2569], include Sonic hedgehog [1966] and retinoic acid [2754].) In this same area is a lawn of ~250 hairlike cilia [1164,2376], although only one representative cilium is depicted henceforth. b. In cross-section, each "9 + 0" cilium displays the 9 microtubule doublets (AB) typical of standard "9 + 2" cilia but lacks the central pair of singlet microtubules. Small black circles are protofilaments of the protein tubulin. (B tubules can have 10 or 11 protofilaments [1837,1892,2524].) The A tubule bears two dynein motor proteins that walk along the B tubule neighbor of the adjacent doublet [2140]. Slippage of adjacent doublets moves the cilium [1875]. c. Each 9 + 0 cilium (enlarged) spins like a propeller (vs. beating like a metronome) [1253], and other 9 + 2 and 9 + 4 cilia here do also [756]. A, P, L, and R denote anterior, posterior, left and right. Because each cilium is tilted ~40 degrees from the perpendicular [1924], the fluid fails to move (because of surface adhesion) as the cilium swings close to the surface (recovery stoke), so it only flows (right to left) during the effective stroke [394]. The leftward flow is thus caused by (1) the clockwise spin of each cilium [773,2529], which is due to (2) the handedness of the tubulin and dynein proteins, which ultimately comes from (3) the chirality of their L-amino acids (cf. Fig. 2.2). d. Gradually, "m" molecules intensify on the left [1898,2569]. e. Eventually, they activate the gene nodal [2257,2691], whose protein product (Nodal) then suffuses the area. f. The heart and other organs develop asymmetrically [308,1201] because of Nodal’s influence on their cellular processes [2119]. Fig. 2.3R b. Single-celled protists called "ciliates" use cilia for propulsion and put more of these "oars" into the water than a Roman trireme. For them, ciliary chirality poses enough puzzles of polarity, asymmetry, and geometry to fill a book [851,1861], and Joseph Frankel has indeed written extensively about their looking-glass world [804-806]. f. Other classes of vertebrates use different tricks to create a nodal bias [1528]. Recent evidence indicates that protostomes may set up left-right asymmetry using nodal as well [1156]. The "Nodal Cilia" Model illustrated here is named for Hensen’s "node" (cf. Fig. 3.2a) [166,569]. The fluid flow actually occurs in the posterior notochordal region [1164,1924], which is on the dorsal side in a human gastrula (not the ventral side depicted here for convenience). A variant of this model invokes mechanical (vs. chemical) signals to activate leftness genes [1705,2548,2885]. Evidence that seems inconsistent with both versions includes (1) the failure of inv mutant mice, which show 100% situs inversus [2882], to exhibit reversed flow [511,1923] and (2) a prior asymmetric bias in the expression of Lefty1 (a component of the nodal circuit) in the mouse blastocyst [2557]. Other models are still debated [2101]. Fig. 2.4  External asymmetries of three vertebrate species and what we would look like if we evolved like them. External asymmetries of three vertebrate species and what we would look like if we evolved like them.a. The narwhal (a whale), Monodon monoceros [1876]. Male narwhals have a tusk up to 2.6 m long (possibly used for jousting [2399]) that is a modified upper left incisor. It is cone-shaped with helical grooves that always twist sinistrally [1388]. Rarely, the right incisor grows out to form a second tusk (not shown), and in every such case its grooves are also sinistral [2146]—a situation as strange as a man with two left hands (cf. Fig. 2.1d legend). Another asymmetry in narwhals (both sexes) is a blowhole left of center [1718,1987], which evolved from fused nostrils with differing roles: respiration (left) versus vocalization (right) [532,1155]. b. Face of the barn owl, Tyto alba, with most feathers removed to reveal preaural skin flaps (gray areas flanking the eyes) that cover the ear openings (D-shaped holes; adapted from ref. [1408]). The ears’ height disparity allows localization of prey in a vertical plane [1893,2703]. The system is fine-tuned visually during adolescence [1409], and it is aided by the barn owl’s having the longest cochlea of any bird [1421]. (Insects use other tricks for directional hearing [2194].) N.B.: Ear asymmetry occurs in a third of all owl species. In the great gray owl, the asymmetry is opposite (i.e., right ear higher), and the shift is seen not just in the skin but also in the skull [1612]. c. Starry flounder, Platichthys stellatus, redrawn from ref. [2057]. Juvenile flatfish look normal, but then one eye migrates across the midline so that both eyes look up when the adult lies flat [2250]. The trajectory (left vs. right) varies among species and is independent of visceral asymmetry [1111]. Goldschmidt cited flatfish to support his "hopeful monster" idea, because it was admittedly hard to imagine any utility for a partial migration [930]. Nevertheless, fossils of just such missing links (early flatfish with partly shifted eyes) have now been found [822,1276]. Having both eyes on one side should make the fish dizzy when swimming [2315], but evolution also rewired the vestibular system to compensate [988]. Fig. 2.4R All of the depicted traits are "directionally asymmetric" insofar as they appear consistently on one side only. In contrast, traits that are "randomly asymmetric" (a.k.a. "antisymmetric") appear as often on the left as on the right [243,929,1871,1976,1980] (cf. "fluctuating asymmetry" [992,1521,1787]). Another bizarre dental bias (aside from the narwhal) exists in a snake species that eats dextrally coiled snails: it has ~25 teeth on the right side of each jaw but only ~17 teeth on the left [2143]! An oddity as baffling as any of those shown here (OK, excluding the Picasso fish) occurs in the long-legged fly, Erebomyia exalloptera [2242]. In males of this species, the right wing is always concave at its tip, whereas the left is always convex—the most extreme wing asymmetry of any flying animal, although the males can still fly! Why such a lopsided trait when symmetry is typically seen as ideal [996]? Consider that flies in general are notorious troubadours (second only to birds), who use their wings as musical instruments [2438]. E. exalloptera are no exception: the males serenade females by fanning their wings, so their odd scalloping might possibly be retuning their love songs [898]. Other fly species have negligible wing asymmetries [1401], and repeated attempts to select for directional asymmetry in various traits have failed in Drosophila [706,1688,1978]. One bias that has been figured out occurs in manatees, which lack hindlimbs but retain pelvic remnants that are larger on the left side [2344]. They lost their hindlegs because of reduced Pitx1 (which sets hindlimb identity [2193]; cf. Fig. 4.7), but they still make Pitx2 on their left side (as part of the nodal circuit; see text). Thus, Pitx2 is evidently compensating for the loss of its Pitx1 paralog [2344] as in sticklebacks [174,175]. Darwin cited manatees in Origin (p. 454) because they have silly vestiges of fingernails on their flippers! Other notable asymmetries, some of which were pondered by Darwin [1979], include unilateral lungs in snakes [16], unilateral ovaries in the platypus [966] and in birds [1028,1254,2053,2082] (except kiwis for some unknown reason [969]), lopsided syrinxes (vocal cords) [399,2532], twisted beaks in crossbills [180,1438], corkscrew mega-penises in waterfowl [273], skewed nasal passages in whales [188,1335,2085], helical partitions ("spiral valves") in shark intestines [16,2417], analogous partitions in the rabbit appendix [2806], asymmetric claws in crustaceans [985,2004,2871], twisted abdomens in hermit crabs [281,1107,1264,1712] (a trait utterly undone by king crabs [544]), and gnarled genitals in male water bugs [1236], not to mention another whole world of spiral and helical traits in plants [674,1112,1301,2592] (cf. reviews [510,1871,1974,2024]). Perhaps the strangest asymmetry ever described is a flatworm larva whose eyes rely on different types of photoreceptors—rhabdomeric type for the right eye versus a combination of rhabdomeric and ciliary types for the left one (cf. Ch. 6) [695]. Aristotle was especially intrigued with claw asymmetries {PoA:4:8:684a26ff.} [1941,1975], which are common in fiddler crabs [2871,2902], hermit crabs [1107,1264,1712], and shrimps [1725,2004]. In lobsters, as he noted, either the left or the right claw can dominate. Which claw becomes the larger crusher (vs. smaller cutter) is fixed before adulthood based on usage [987,1564]: both claws become cutters if they are underused, but an unknown neural circuit prevents them both from becoming crushers [985,986]. The crusher-cutter asymmetry in crustaceans is eerily similar to our molar-incisor dichotomy, and the convergence gets even spookier in the molarlike "teeth" that line their crusher. Both gadgets (claws and jaws) are lovely illustrations of physical constraints because they evolved as simple levers and are subject to the same formulae as scissors and pliers. The only asymmetry that has been investigated genetically to any useful extent is helical coiling in snail shells [97,1156,2297]. Dextral shells are the norm and sinistral ones are rare in most gastropod species [976,984], although a few whole species are sinistral [97,2196,2297]. Why dextrality prevails is unknown [538,850,1305,2684]. Mating between mirror-image snails can occur [102,1210], and chirality turns out to be due to a single gene that acts before fertilization [586,1031,2196,2520,2658]! How the maternal gene product (a cytoplasmic RNA or protein) biases the spiral cleavage of the zygote [813,2363,2741] is unclear [1085,1156]. Our annoying inability to use both hands with equal grace can be blamed on our hemispheric lateralization [79,369,872]. Brain lateralities of some kind [79,261,574,872,920] must also dictate left-right behavioral preferences in fish [603,1203,1554,1714], frogs [207,1630,1845], snakes [2142], birds [659,933,1229,2666], rats [1474], humans [112,519,520,750,2879], pre-human hominins [856,914], and nonhuman primates [361,1068,1117,1517,1803]. Indeed, lateralized brain function has even been found in honeybees [1526]! It remains a mystery [369,790,1945,2001]. Charles Dodgson (a.k.a. Lewis Carroll)’s fascination with asymmetry, which is so apparent in his book Through the Looking-Glass, may have been sparked by his own asymmetric peculiarities: one shoulder was higher, his eyes were at different levels, and his smile was crooked [380]. top of page
Lewis I. Held, Jr. is Associate Professor in the Department of Biology at Texas Tech University.
| |
 © 2009 Thomas B. Brody, Ph.D. |