|
Gene name - Ultrabithorax Synonyms - bithorax (bx) Cytological map position - 89E1-3 Function - transcription factor Keywords - bithorax complex |
Symbol - Ubx FlyBase ID:FBgn0003944 Genetic map position - 3-58.8 Classification - homeodomain - Antp class Cellular location - nuclear |
| Recent literature | Domsch, K., Carnesecchi, J., Disela, V., Friedrich, J., Trost, N., Ermakova, O., Polychronidou, M. and Lohmann, I. (2019). The Hox transcription factor Ubx stabilizes lineage commitment by suppressing cellular plasticity in Drosophila. Elife 8. PubMed ID: 31050646
Summary: During development cells become restricted in their differentiation potential by repressing alternative cell fates, and the Polycomb complex plays a crucial role in this process. However, how alternative fate genes are lineage-specifically silenced is unclear. This study examined Ultrabithorax (Ubx), a multi-lineage transcription factor of the Hox class, in two tissue lineages using sorted nuclei and interfered with Ubx in mesodermal cells. Depletion of Ubx leads to the de-repression of genes normally expressed in other lineages. Ubx silences expression of alternative fate genes by retaining the Polycomb Group protein Pleiohomeotic at Ubx targeted genomic regions, thereby stabilizing repressive chromatin marks in a lineage-dependent manner. This study demonstrates that Ubx stabilizes lineage choice by suppressing the multipotency encoded in the genome via its interaction with Pho. This mechanism may explain why the Hox code is maintained throughout the lifecycle, since it could set a block to transdifferentiation in adult cells. |
Issa, A. R., Picao-Osorio, J., Rito, N., Chiappe, M. E. and Alonso, C. R. (2019). A single microRNA-Hox gene module controls equivalent movements in biomechanically distinct forms of Drosophila. Curr Biol. PubMed ID: 31327720
Summary: Movement is the main output of the nervous system. It emerges during development to become a highly coordinated physiological process essential to survival and adaptation of the organism to the environment. Similar movements can be observed in morphologically distinct developmental stages of an organism, but it is currently unclear whether or not these movements have a common molecular cellular basis. This study explores this problem in Drosophila, focusing on the roles played by the microRNA (miRNA) locus miR-iab4/8, which has been previously shown to be essential for the normal corrective response displayed by the fruit fly larva when turned upside down (self-righting). This study shows that miR-iab4 is required for normal self-righting across all three Drosophila larval stages. Unexpectedly, it was also discovered that this miRNA is essential for normal self-righting behavior in the adult fly, an organism with different morphology, neural constitution, and biomechanics. Through the combination of gene expression, optical imaging, and quantitative behavioral approaches, evidence is provided that miR-iab4 exerts its effects on adult self-righting behavior in part through repression of the Hox gene Ultrabithorax (Ubx) in a specific set of adult motor neurons, the NB2-3/lin15 neurons. The results show that miRNA controls the function, rather than the morphology, of these neurons and demonstrate that post-developmental changes in Hox gene expression can modulate behavior in the adult. This work reveals that a common miRNA-Hox genetic module can be re-deployed in different neurons to control functionally equivalent movements in biomechanically distinct organisms and describes a novel post-developmental role of the Hox genes in adult neural function. |
Tsai, A., Alves, M. R. and Crocker, J. (2019). Multi-enhancer transcriptional hubs confer phenotypic robustness. Elife 8. PubMed ID: 31294690
Summary: Previous work has shown in Drosophila melanogaster embryos that low-affinity Ultrabithorax (Ubx)-responsive shavenbaby (svb) enhancers drive expression using localized transcriptional environments and that active svb enhancers on different chromosomes tended to colocalize. This study tested the hypothesis that these multi-enhancer 'hubs' improve phenotypic resilience to stress by buffering against decreases in transcription factor concentrations and transcriptional output. Deleting a redundant enhancer from the svb locus led to reduced trichome numbers in embryos raised at elevated temperatures. Using high-resolution fluorescence microscopy, lower Ubx concentration and transcriptional output were observed in this deletion allele. Transcription sites of the full svb cis-regulatory region inserted into a different chromosome colocalized with the svb locus, increasing Ubx concentration, the transcriptional output of svb, and partially rescuing the phenotype. Thus, multiple enhancers could reinforce a local transcriptional hub to buffer against environmental stresses and genetic perturbations, providing a mechanism for phenotypical robustness. |
Delker, R. K., Ranade, V., Loker, R., Voutev, R. and Mann, R. S. (2019). Low affinity binding sites in an activating CRM mediate negative autoregulation of the Drosophila Hox gene Ultrabithorax. PLoS Genet 15(10): e1008444. PubMed ID: 31589607
Summary: Specification of cell identity and the proper functioning of a mature cell depend on precise regulation of gene expression. Both binary ON/OFF regulation of transcription, as well as more fine-tuned control of transcription levels in the ON state, are required to define cell types. The Drosophila melanogaster Hox gene, Ultrabithorax (Ubx), exhibits both of these modes of control during development. While ON/OFF regulation is needed to specify the fate of the developing wing (Ubx OFF) and haltere (Ubx ON), the levels of Ubx within the haltere differ between compartments along the proximal-distal axis. This study identified and molecularly dissected the novel contribution of a previously identified Ubx cis-regulatory module (CRM), anterobithorax (abx), to a negative auto-regulatory loop that decreases Ubx expression in the proximal compartment of the haltere as compared to the distal compartment. Ubx, in complex with the known Hox cofactors, Homothorax (Hth) and Extradenticle (Exd), acts through low-affinity Ubx-Exd binding sites to reduce the levels of Ubx transcription in the proximal compartment. Importantly, Ubx-Exd-binding site mutations sufficient to result in de-repression of abx activity in a transgenic context are not sufficient to de-repress Ubx expression when mutated at the endogenous locus, suggesting the presence of multiple mechanisms through which Ubx-mediated repression occurs. These results underscore the complementary nature of CRM analysis through transgenic reporter assays and genome modification of the endogenous locus; but, they also highlight the increasing need to understand gene regulation within the native context to capture the potential input of multiple genomic elements on gene control. |
Carnesecchi, J., Sigismondo, G., Domsch, K., Baader, C. E. P., Rafiee, M. R., Krijgsveld, J. and Lohmann, I. (2020). Multi-level and lineage-specific interactomes of the Hox transcription factor Ubx contribute to its functional specificity. Nat Commun 11(1): 1388. PubMed ID: 32170121
Summary: Transcription factors (TFs) control cell fates by precisely orchestrating gene expression. However, how individual TFs promote transcriptional diversity remains unclear. This study used the Hox TF Ultrabithorax (Ubx) as a model to explore how a single TF specifies multiple cell types. Using proximity-dependent Biotin IDentification in Drosophila, Ubx interactomes were identifed in three embryonic tissues. Ubx interacts with largely non-overlapping sets of proteins with few having tissue-specific RNA expression. Instead most interactors are active in many cell types, controlling gene expression from chromatin regulation to the initiation of translation. Genetic interaction assays in vivo confirm that they act strictly lineage- and process-specific. Thus, functional specificity of Ubx seems to play out at several regulatory levels and to result from the controlled restriction of the interaction potential by the cellular environment. Thereby, it challenges long-standing assumptions such as differential RNA expression as determinant for protein complexes. |
Duffraisse, M., Paul, R., Carnesecchi, J., Hudry, B., Banreti, A., Reboulet, J., Ajuria, L., Lohmann, I. and Merabet, S. (2020). Role of a versatile peptide motif controlling Hox nuclear export and autophagy in the Drosophila fat body. J Cell Sci 133(18). PubMed ID: 32878938
Summary: Hox proteins are major regulators of embryonic development, acting in the nucleus to regulate the expression of their numerous downstream target genes. By analyzing deletion forms of the Drosophila Hox protein Ultrabithorax (Ubx), the presence of an unconventional nuclear export signal (NES) was identified that overlaps with a highly conserved motif originally described as mediating the interaction with the PBC proteins, a generic and crucial class of Hox transcriptional cofactors that act in development and cancer. This unconventional NES is involved in the interaction with the major exportin protein CRM1 (also known as Embargoed in flies) in vivo and in vitro. This interaction is tightly regulated in the Drosophila fat body to control the autophagy-repressive activity of Ubx during larval development. The role of the PBC interaction motif as part of an unconventional NES was also uncovered in other Drosophila and human Hox proteins, highlighting the evolutionary conservation of this novel function. Together, these results reveal the extreme molecular versatility of a unique short peptide motif for controlling the context-dependent activity of Hox proteins both at transcriptional and non-transcriptional levels. |
Domsch, K., Schroder, J., Janeschik, M., Schaub, C. and Lohmann, I. (2021). The Hox Transcription Factor Ubx Ensures Somatic Myogenesis by Suppressing the Mesodermal Master Regulator Twist. Cell Rep 34(1): 108577. PubMed ID: 33406430
Summary: Early lineage-specific master regulators are essential for the specification of cell types. However, once cells are committed to a specific fate, it is critical to restrict the activity of such factors to enable differentiation. To date, it remains unclear how these factors are silenced. Using the Drosophila mesoderm as a model and a comparative genomic approach, the Hox transcription factor Ultrabithorax (Ubx) was shown to be critical for the repression of the master regulator Twist. Mesoderm-specific Ubx loss-of-function experiments using CRISPR-Cas9 and overexpression studies demonstrate that Ubx majorly impacts twist transcription. A mechanistic analysis reveals that Ubx requires the NK-homeodomain protein Tinman to bind to the twist promoter. Furthermore, these factor interactions were found to be critical for silencing by recruiting the Polycomb DNA binding protein Pleiohomeotic. Altogether, these data reveal that Ubx is a critical player in mediating the silencing of Twist, which is crucial for coordinated muscle differentiation. |
Ponrathnam, T., Saini, R., Banu, S. and Mishra, R. K. (2021). Drosophila Hox genes induce melanized pseudo-tumors when misexpressed in hemocytes. Sci Rep 11(1): 1838. PubMed ID: 33469139
Summary: Hox genes are early determinants of cell identity along the anterior-posterior body axis across bilaterians. Several late non-homeotic functions of Hox genes have emerged in a variety of processes involved in organogenesis in several organisms, including mammals. Several studies have reported the misexpression of Hox genes in a variety of malignancies including acute myeloid leukemia. The Hox genes Dfd, Ubx, abd-A and Abd-B were overexpressed via the UAS-Gal4 system using Cg-Gal4, Lsp2-Gal4, He-Gal4 and HmlD3-Gal4 as specific drivers. Genetic interaction was tested by bringing overexpression lines in heterozygous mutant backgrounds of Polycomb and trithorax group factors. Larvae were visually scored for melanized bodies. Circulating hemocytes were quantified and tested for differentiation. Pupal lethality was assessed. Expression of Dfd, Ubx and abd-A, but not Abd-B in the hematopoietic compartment of Drosophila led to the appearance of circulating melanized bodies, an increase in cell number, cell-autonomous proliferation, and differentiation of hemocytes. Pupal lethality and melanized pseudo-tumors were suppressed in Psc1 and esc2 backgrounds while polycomb group member mutations Pc1 and Su(z)123 and trithorax group member mutation TrlR85 enhanced the phenotype. Dfd, Ubx and abd-A are leukemogenic. Mutations in Polycomb and trithorax group members modulate the leukemogenic phenotype. This RNAseq of Cg-Gal4 > UAS-abd-A hemocytes may contain genes important to Hox gene induced leukemias. |
Kanwal, A., Joshi, P. V., Mandal, S. and Mandal, L. (2021). Ubx-Collier signaling cascade maintains blood progenitors in the posterior lobes of the Drosophila larval lymph gland. PLoS Genet 17(8): e1009709. PubMed ID: 34370733
Summary: Drosophila larval hematopoiesis occurs in a specialized multi-lobed organ called the lymph gland. Extensive characterization of the organ has provided mechanistic insights into events related to developmental hematopoiesis. Spanning from the thoracic to the abdominal segment of the larvae, this organ comprises a pair of primary, secondary, and tertiary lobes. Much understanding arises from the studies on the primary lobe, while the secondary and tertiary lobes have remained mostly unexplored. Previous studies have inferred that these lobes are composed of progenitors that differentiate during pupation; however, the mechanistic basis of this extended progenitor state remains unclear. This study shows that posterior lobe progenitors are maintained by a local signaling center defined by Ubx and Collier in the tertiary lobe. This Ubx zone in the tertiary lobe shares several markers with the niche of the primary lobe. Ubx domain regulates the homeostasis of the posterior lobe progenitors in normal development and an immune-challenged scenario. This study establishes the lymph gland as a model to tease out how the progenitors interface with the dual niches within an organ during development and disorders. |
Loker, R., Sanner, J. E. and Mann, R. S. (2021). Cell-type-specific Hox regulatory strategies orchestrate tissue identity. Curr Biol. PubMed ID: 34358443
Summary: Hox proteins are homeodomain transcription factors that diversify serially homologous segments along the animal body axis, as revealed by the classic bithorax phenotype of Drosophila melanogaster, in which mutations in Ultrabithorax (Ubx) transform the third thoracic segment into the likeness of the second thoracic segment. To specify segment identity, it was shown that Ubx both increases and decreases chromatin accessibility, coinciding with its dual role as both an activator and repressor of transcription. However, the choice of transcriptional activity executed by Ubx is spatially regulated and depends on the availability of cofactors, with Ubx acting as a repressor in some populations and as an activator in others. Ubx-mediated changes to chromatin accessibility positively and negatively affect the binding of Scalloped (Sd), a transcription factor that is required for appendage development in both segments. These findings illustrate how a single Hox protein can modify complex gene regulatory networks to transform the identity of an entire tissue. |
Wei, Y., Du, J. and Zhao, Z. (2021). Integrative Role of 14-3-3epsilon in Sleep Regulation. Int J Mol Sci 22(18). PubMed ID: 34575915
Summary: Sleep is a crucial factor for health and survival in all animals. This study found by proteomic analysis that some cancer related proteins were impacted by the circadian clock. The 14-3-3ε protein, expression of which is activated by the circadian transcription factor Clock, regulates adult sleep of Drosophila independent of circadian rhythm. Detailed analysis of the sleep regulatory mechanism shows that 14-3-3ε directly targets the Ultrabithorax (Ubx) gene to activate transcription of the pigment dispersing factor (PDF). The dopamine receptor (Dop1R1) and the octopamine receptor (Oamb), are also involved in the 14-3-3ε pathway, which in 14-3-3ε mutant flies causes increases in the dopR1 and OAMB, while downregulation of the DopR1 and Oamb can restore the sleep phenotype caused by the 14-3-3ε mutation. In conclusion, 14-3-3ε is necessary for sleep regulation in Drosophila. |
Hermann, A., Kosman, D., McGinnis, W. and Tour, E. (2021). The expression of Drosophila melanogaster Hox gene Ultrabithorax is not overtly regulated by the intronic long non-coding RNA lncRNA: PS4 in a wild type genetic background. G3 (Bethesda). PubMed ID: 34791185 Summary: Long non-coding RNAs (lncRNAs) have been implicated in a variety of processes in development, differentiation, and disease. In Drosophila melanogaster, the bithorax Hox cluster (BX-C) contains three Hox genes (Ultrabithorax (Ubx), abdominal-A (abd-A), and Abdominal-B (Abd-B)), along with a number of lncRNAs, most with unknown functions. This study investigated the function of a long non-coding RNA, lncRNA: PS4 that originates in the second intron of Ubx and is transcribed in the antisense orientation to Ubx. The expression pattern of lncRNA: PS4 is complementary to Ubx in the thoracic primordia, and the lncRNA: PS4 coding region overlaps the location of the large insertion that causes the dominant homeotic mutation Contrabithorax-1 (UbxCbx-1), which partially transforms Drosophila wings into halteres via ectopic activation of Ubx. This led the authors to investigate the potential role of this lncRNA in regulation of Ubx expression. The UbxCbx-1 mutation dramatically changes the pattern of lncRNA: PS4, eliminating the expression of most lncRNA: PS4 sequences from parasegment 4 (where Ubx protein is normally absent) and ectopically activating lncRNA: PS4 at high levels in the abdomen (where Ubx is normally expressed). These changes, however, did not lead to changes in the Ubx embryonic transcription pattern. Targeted deletion of the two promoters of lncRNA: PS4 did not result in the change of Ubx expression in the embryos. In the genetic background of a UbxCbx-1 mutation, the lncRNA: PS4 mutation does slightly enhance the ectopic activation of Ubx protein expression in wing discs and also slightly enhances the wing phenotype seen in UbxCbx-1 heterozygotes. |
Togel, M., Pass, G. and Paululat, A. (2021). Wing Hearts in Four-Winged Ultrabithorax-mutant Flies-the role of Hox genes in wing heart specification. Genetics. PubMed ID: 34791231 Summary: Wings are probably the most advanced evolutionary novelty in insects. The development of wings requires the activity of so-called wing hearts located in the scutellum of the thorax. Immediately after the imaginal ecdysis, these accessory circulatory organs remove haemolymph and apoptotic epidermal cells from the premature wing through their pumping action. This clearing process is essential for the formation of functional wing blades. Mutant Drosophila that lack intact wing hearts are flightless and display malformed wings. The embryonic wing heart progenitors originate from two adjacent parasegments corresponding to the later thoracic segments T2 and T3. However, the adult dipterian fly harbors only one pair of wing hearts and also only one pair of wings located in thoracic segment T2. This study shows, that the specification of wing heart progenitors depends on the regulatory activity of the Hox gene Ultrabithorax. Furthermore, the development of four wing hearts was analyzed in the famous four-winged Ultrabithorax (Ubx) mutant, which was first discovered by Ed Lewis in the 1970s. In these flies, the third thoracic segment (T3) is transformed into a second thoracic segment (HT2). This results in a second pair of wings instead of the club-shaped halteres normally formed by T3. This study shows that a second pair of wild-type wing hearts is formed in the four-winged fly and that all wing hearts originate from the wild-type progenitor cells. |
Carnesecchi, J., Boumpas, P., van Nierop, Y. S. P., Domsch, K., Pinto, H. D., Borges Pinto, P. and Lohmann, I. (2022). The Hox transcription factor Ultrabithorax binds RNA and regulates co-transcriptional splicing through an interplay with RNA polymerase II. Nucleic Acids Res 50(2): 763-783. PubMed ID: 34931250
Summary: Transcription factors (TFs) play a pivotal role in cell fate decision by coordinating gene expression programs. Although most TFs act at the DNA layer, few TFs bind RNA and modulate splicing. Yet, the mechanistic cues underlying TFs activity in splicing remain elusive. Focusing on the Drosophila Hox TF Ultrabithorax (Ubx), this work shed light on a novel layer of Ubx function at the RNA level. Transcriptome and genome-wide binding profiles in embryonic mesoderm and Drosophila cells indicate that Ubx regulates mRNA expression and splicing to promote distinct outcomes in defined cellular contexts. These results demonstrate a new RNA-binding ability of Ubx. The N51 amino acid of the DNA-binding Homeodomain is non-essential for RNA interaction in vitro, but is required for RNA interaction in vivo and Ubx splicing activity. Moreover, mutation of the N51 amino acid weakens the interaction between Ubx and active RNA Polymerase II (Pol II). These results reveal that Ubx regulates elongation-coupled splicing, which could be coordinated by a dynamic interplay with active Pol II on chromatin. Overall, this work uncovered a novel role of the Hox TFs at the mRNA regulatory layer. This could be an essential function for other classes of TFs to control cell diversity. |
Folkendt, L., Lohmann, I. and Domsch, K. (2021). An Evolutionary Perspective on Hox Binding Site Preferences in Two Different Tissues. J Dev Biol 9(4). PubMed ID: 34940504
Summary: Transcription factor (TF) networks define the precise development of multicellular organisms. While many studies focused on TFs expressed in specific cell types to elucidate their contribution to cell specification and differentiation, it is less understood how broadly expressed TFs perform their precise functions in the different cellular contexts. To uncover differences that could explain tissue-specific functions of such TFs, this study analyzed genomic chromatin interactions of the broadly expressed Drosophila Hox TF Ultrabithorax (Ubx) in the mesodermal and neuronal tissues using bioinformatics. This investigations showed that Ubx preferentially interacts with multiple yet tissue-specific chromatin sites in putative regulatory regions of genes in both tissues. Importantly, the classical Hox/Ubx DNA binding motif was found to be enriched only among the neuronal Ubx chromatin interactions, whereas a novel Ubx-like motif with rather low predicted Hox affinities was identified among the regions bound by Ubx in the mesoderm. Finally, this analysis revealed that tissues-specific Ubx chromatin sites are also different with regards to the distribution of active and repressive histone marks. Based on these data, it is proposed that the tissue-related differences in Ubx binding behavior could be a result of the emergence of the mesoderm as a new germ layer in triploblastic animals, which might have required the Hox TFs to relax their binding specificity. |
Feng, S. and Mann, R. S. (2022). SpyChIP identifies cell type-specific transcription factor occupancy from complex tissues. Proc Natl Acad Sci U S A 119(25): e2122900119. PubMed ID: 35696584
Summary: Chromatin immunoprecipitation (ChIP) is an important technique for characterizing protein-DNA binding in vivo. One drawback of ChIP-based techniques is the lack of cell type-specificity when profiling complex tissues. To overcome this limitation, this study developed SpyChIP to identify cell type-specific transcription factor (TF) binding sites in native physiological contexts without tissue dissociation or nuclei sorting. SpyChIP takes advantage of a specific covalent isopeptide bond that rapidly forms between the 15-amino acid SpyTag and the 17-kDa protein SpyCatcher. In SpyChIP, the target TF is fused with SpyTag by genome engineering, and an epitope tagged SpyCatcher is expressed in cell populations of interest, where it covalently binds to SpyTag-TF. Cell type-specific ChIP is obtained by immunoprecipitating chromatin prepared from whole tissues using antibodies directed against the epitope-tagged SpyCatcher. Using SpyChIP, the genome-wide binding profiles of the Hox protein Ultrabithorax (Ubx) were obtained in two distinct cell types of the Drosophila haltere imaginal disc. These results revealed extensive region-specific Ubx-DNA binding events, highlighting the significance of cell type-specific ChIP and the limitations of whole-tissue ChIP approaches. Analysis of Ubx::SpyChIP results provided insights into the relationship between chromatin accessibility and Ubx-DNA binding, as well as different mechanisms Ubx employs to regulate its downstream cis-regulatory modules. In addition to SpyChIP, it is suggested that SpyTag-SpyCatcher technology, as well as other protein pairs that form covalent isopeptide bonds, will facilitate many additional in vivo applications that were previously impractical. |
Bu, S., Lau, S. S. Y., Yong, W. L., Zhang, H., Thiagarajan, S., Bashirullah, A. and Yu, F. (2023). Polycomb group genes are required for neuronal pruning in Drosophila. BMC Biol 21(1): 33. PubMed ID: 36793038
Summary: Pruning that selectively eliminates unnecessary or incorrect neurites is required for proper wiring of the mature nervous system. During Drosophila metamorphosis, dendritic arbourization sensory neurons (ddaCs) and mushroom body (MB) γ neurons can selectively prune their larval dendrites and/or axons in response to the steroid hormone ecdysone. An ecdysone-induced transcriptional cascade plays a key role in initiating neuronal pruning. However, how downstream components of ecdysone signalling are induced remains not entirely understood. This study identified that Scm, a component of Polycomb group (PcG) complexes, is required for dendrite pruning of ddaC neurons. Two PcG complexes, PRC1 and PRC2, are important for dendrite pruning. Interestingly, depletion of PRC1 strongly enhances ectopic expression of Abdominal B (Abd-B) and Sex combs reduced, whereas loss of PRC2 causes mild upregulation of Ultrabithorax and Abdominal A in ddaC neurons. Among these Hox genes, overexpression of Abd-B causes the most severe pruning defects, suggesting its dominant effect. Knockdown of the core PRC1 component Polyhomeotic (Ph) or Abd-B overexpression selectively downregulates Mical expression, thereby inhibiting ecdysone signalling. Finally, Ph is also required for axon pruning and Abd-B silencing in MB γ neurons, indicating a conserved function of PRC1 in two types of pruning. This study demonstrates important roles of PcG and Hox genes in regulating ecdysone signalling and neuronal pruning in Drosophila. Moreover, our findings suggest a non-canonical and PRC2-independent role of PRC1 in Hox gene silencing during neuronal pruning. |
Raouf Issa, A., J, A. C. M., Padmanabhan, A. and Alonso, C. R. (2022). A novel post-developmental role of the Hox genes underlies normal adult behavior. Proc Natl Acad Sci U S A 119(49): e2209531119. PubMed ID: 36454751
Summary: The molecular mechanisms underlying the stability of mature neurons and neural circuits are poorly understood. This problem was explored and it was discovered that the Hox genes are a component of the genetic program that maintains normal neural function in adult Drosophila. Post-developmental downregulation of the Hox gene Ultrabithorax (Ubx) in adult neurons leads to substantial anomalies in flight. Mapping the cellular basis of these effects reveals that Ubx is required within a subset of dopaminergic neurons, and cell circuitry analyses in combination with optogenetics allow linking of these dopaminergic neurons to flight control. Functional imaging experiments show that Ubx is necessary for normal dopaminergic activity, and neuron-specific RNA-sequencing defines two previously uncharacterized ion channel-encoding genes as potential mediators of Ubx behavioral roles. This study thus reveals a novel role of the Hox system in controlling adult behavior and neural function. Based on the broad evolutionary conservation of the Hox system across distantly related animal phyla, it is predicedt that the Hox genes might play neurophysiological roles in adult forms of other species, including humans. |
Buffry, A. D., Kittelmann, S. and McGregor, A. P. (2023). Characterisation of the role and regulation of Ultrabithorax in sculpting fine-scale leg morphology. Front Cell Dev Biol 11: 1119221. PubMed ID: 36861038
Summary: Hox genes are expressed during embryogenesis and determine the regional identity of animal bodies along the antero-posterior axis. However, they also function post-embryonically to sculpt fine-scale morphology. To better understand how Hox genes are integrated into post-embryonic gene regulatory networks, this study further analysed the role and regulation of Ultrabithorax (Ubx) during leg development in Drosophila melanogaster. Ubx regulates several aspects of bristle and trichome patterning on the femurs of the second (T2) and third (T3) leg pairs. Repression of trichomes in the proximal posterior region of the T2 femur by Ubx is likely mediated by activation of the expression of microRNA-92a and microRNA-92b by this Hox protein. Furthermore, this study identified a novel enhancer of Ubx that recapitulates the temporal and regional activity of this gene in T2 and T3 legs. Transcription factor (TF) binding motif analysis was used in regions of accessible chromatin in T2 leg cells to predict and functionally test TFs that may regulate the Ubx leg enhancer.The role of the Ubx co-factors Homothorax (Hth) and Extradenticle (Exd) in T2 and T3 femurs was also tested. Several TFs were found that may act upstream or in concert with Ubx to modulate trichome patterning along the proximo-distal axis of developing femurs and that the repression of trichomes also requires Hth and Exd. Taken together our results provide insights into how Ubx is integrated into a post-embryonic gene regulatory network to determine fine-scale leg morphology. |
Khan, S., Pradhan, S. J., Giraud, G., Bleicher, F., Paul, R., Merabet, S. and Shashidhara, L. S. (2023). A Micro-evolutionary Change in Target Binding Sites as a Key Determinant of Ultrabithorax Function in Drosophila. J Mol Evol. PubMed ID: 37341745
Summary: Hox genes encode Homeodomain-containing transcription factors, which specify segmental identities along the anterior-posterior axis. Functional changes in Hox genes have been directly implicated in the evolution of body plans across the metazoan lineage. The Hox protein Ultrabithorax (Ubx) is expressed and required in developing third thoracic (T3) segments in holometabolous insects studied so far, particularly, of the order Coleoptera, Lepidoptera and Diptera. Ubx function is key to specify differential development of the second (T2) and T3 thoracic segments in these insects. While Ubx is expressed in the third thoracic segment in developing larvae of Hymenopteran Apis mellifera, the morphological differences between T2 and T3 are subtle. To identify evolutionary changes that are behind the differential function of Ubx in Drosophila and Apis, which are diverged for more than 350 million years, comparative analyses were performed of genome wide Ubx-binding sites between these two insects. These studies reveal that a motif with a TAAAT core is a preferred binding site for Ubx in Drosophila, but not in Apis. Biochemical and transgenic assays suggest that in Drosophila, the TAAAT core sequence in the Ubx binding sites is required for Ubx-mediated regulation of two of its target genes studied here; CG13222, a gene that is normally upregulated by Ubx and vestigial (vg), whose expression is repressed by Ubx in T3. Interestingly, changing the TAAT site to a TAAAT site was sufficient to bring an otherwise unresponsive enhancer of the vg gene from Apis under the control of Ubx in a Drosophila transgenic assay. Taken together, these results suggest an evolutionary mechanism by which critical wing patterning genes might have come under the regulation of Ubx in the Dipteran lineage. |
Hodkinson, L. J., Smith, C., Comstra, H. S., ...., Zeng, H., Schmidt, C. A., Rieder, L. E. (2023). A bioinformatics screen reveals hox and chromatin remodeling factors at the Drosophila histone locus. BMC genomic data, 24(1):54 PubMed ID: 37735352
Summary: Cells orchestrate histone biogenesis with strict temporal and quantitative control. To efficiently regulate histone biogenesis, the repetitive Drosophila melanogaster replication-dependent histone genes are arrayed and clustered at a single locus. Regulatory factors concentrate in a nuclear body known as the histone locus body (HLB), which forms around the locus. Historically, HLB factors are largely discovered by chance, and few are known to interact directly with DNA. It is therefore unclear how the histone genes are specifically targeted for unique and coordinated regulation. RESULTS: To expand the list of known HLB factors, we performed a candidate-based screen by mapping 30 publicly available ChIP datasets of 27 unique factors to the Drosophila histone gene array. We identified novel transcription factor candidates, including the Drosophila Hox proteins Ultrabithorax (Ubx), Abdominal-A (Abd-A), and Abdominal-B (Abd-B), suggesting a new pathway for these factors in influencing body plan morphogenesis. Additionally, six other factors were identified that target the histone gene array: JIL-1, hormone-like receptor 78 (Hr78), the long isoform of female sterile homeotic (1) (fs(1)h) as well as the general transcription factors TBP associated factor 1 (TAF-1), Transcription Factor IIB (TFIIB), and Transcription Factor IIF (TFIIF). This foundational screen provides several candidates for future studies into factors that may influence histone biogenesis. Further, this study emphasizes the powerful reservoir of publicly available datasets, which can be mined as a primary screening technique. |
Liu, B. P., Hua, B. Z. (2024). Distinct roles of the Hox genes Ultrabithorax and abdominal-A in scorpionfly embryonic proleg development. Insect Mol Biol, 33(1):69-80 PubMed ID: 37792400
Summary: The abdominal appendages of larval insects have a complex evolutionary history of gain and loss, but the regulatory mechanisms underlying the abdominal appendage development remain largely unclear. This study investigated the embryogenesis of abdominal prolegs in the scorpionfly Panorpa liui Hua (Mecoptera: Panorpidae) using in situ hybridization and parental RNA interference. The results show that RNAi-mediated knockdown of Ultrabithorax (Ubx) led to a homeotic transformation of the first abdominal segment (A1) into the third thoracic segment (T3) and changed the distributions of the downstream target Distal-less (Dll) expression but did not affect the expression levels of Dll. Knockdown of abdominal-A (abd-A) resulted in malformed segments, abnormal prolegs and disrupted Dll expression. The results demonstrate that the gene Ubx maintains an ancestral role of modulating A1 appendage fate without preventing Dll initiation, and a secondary adaptation of abd-A evolves the ability to specify abdominal segments and proleg identity. It is concluded that changes in abdominal Hox gene expression and their target genes regulate abdominal appendage morphology during the evolutionary course of holometabolous larvae. |
Tendolkar, A., Mazo-Vargas, A., Livraghi, L., Hanly, J. J., Van Horne, K. C., Gilbert, L. E., Martin, A. (2024).. Cis-regulatory modes of Ultrabithorax inactivation in butterfly forewings. Elife, 12 PubMed ID: 38261357
Summary: Hox gene clusters encode transcription factors that drive regional specialization during animal development: for example the Hox factor Ubx is expressed in the insect metathoracic (T3) wing appendages and differentiates them from T2 mesothoracic identities. Hox transcriptional regulation requires silencing activities that prevent spurious activation and regulatory crosstalks in the wrong tissues, but this has seldom been studied in insects other than Drosophila, which shows a derived Hox dislocation into two genomic clusters that disjoined Antennapedia (Antp) and Ultrabithorax (Ubx). This study investigated how Ubx is restricted to the hindwing in butterflies, amidst a contiguous Hox cluster. By analysing Hi-C and ATAC-seq data in the butterfly Junonia coenia, a Topologically Associated Domain (TAD) was shown to maintain a hindwing-enriched profile of chromatin opening around Ubx. This TAD is bordered by a Boundary Element (BE) that separates it from a region of joined wing activity around the Antp locus. CRISPR mutational perturbation of this BE releases ectopic Ubx expression in forewings, inducing homeotic clones with hindwing identities. Further mutational interrogation of two non-coding RNA encoding regions and one putative cis-regulatory module within the Ubx TAD cause rare homeotic transformations in both directions, indicating the presence of both activating and repressing chromatin features. A series of spontaneous forewing homeotic phenotypes obtained in Heliconius butterflies is described, their possible mutational basis is discussed. By leveraging the extensive wing specialization found in butterflies, this initial exploration of Ubx regulation demonstrates the existence of silencing and insulating sequences that prevent its spurious expression in forewings. |
Jiang, Y., Chiu, T. P., Mitra, R., Rohs, R. (2024). Probing the role of the protonation state of a minor groove-linker histidine in Exd-Hox-DNA binding. Biophys J, 123(2):248-259 PubMed ID: 38130056
Summary: DNA recognition and targeting by transcription factors (TFs) through specific binding are fundamental in biological processes. Furthermore, the histidine protonation state at the TF-DNA binding interface can significantly influence the binding mechanism of TF-DNA complexes. Nevertheless, the role of histidine in TF-DNA complexes remains underexplored. This study employed all-atom molecular dynamics simulations using AlphaFold2-modeled complexes based on previously solved co-crystal structures to probe the role of the His-12 residue in the Extradenticle (Exd)-Sex combs reduced (Scr)-DNA complex when binding to Scr and Ultrabithorax (Ubx) target sites. These results demonstrate that the protonation state of histidine notably affected the DNA minor-groove width profile and binding free energy. Examining flanking sequences of various binding affinities derived from SELEX-seq experiments, the relationship between binding affinity and specificity was studied. How histidine protonation leads to increased binding affinity but can lower specificity was uncovered. These findings provide new mechanistic insights into the role of histidine in modulating TF-DNA binding. |
Villarreal-Puente, A., Altamirano-Torres, C., Jimenez-Mejia, G., Hernandez-Bautista, C., Montalvo-Mendez, R., Vazquez, M., Zurita, M., Resendez-Perez, D. (2024). Novel Antennapedia and Ultrabithorax trimeric complexes with TBP and Exd regulate transcription. Hereditas, 161(1):25 PubMed ID: 39080786
Summary: Hox proteins interact with DNA and many other proteins, co-factors, transcriptional factors, chromatin remodeling components, non-coding RNAs and even the extracellular matrix that assembles the Hox complexes. The number of interacting partners continues to grow with diverse components and more transcriptional factors than initially thought. Hox complexes present many activities, but their molecular mechanisms to modulate their target genes remain unsolved. This study showed the protein-protein interaction of Antp with Ubx through the homeodomain using BiFC in Drosophila. Analysis of Antp-deletional mutants showed that AntpHD helixes 1 and 2 are required for the interaction with Ubx. Also, a novel interaction of Ubx with TBP was found, in which the PolyQ domain of TBP is required for the interaction. Moreover, the formation of two new trimeric complexes of Antp with Ubx, TBP and Exd was found using BiFC-FRET; these proteins, however, do not form a trimeric interaction with BIP2 or TFIIEβ. The novel trimeric complexes reduced Antp transcriptional activity, indicating that they could confer specificity for repression. These results increase the number of transcriptional factors in the Antp and Ubx interactomes that form two novel trimeric complexes with TBP and Exd. A new Ubx interaction with TBP was also found. These novel interactions provide important clues of the dynamics of Hox-interacting complexes involved in transcriptional regulation, contributing to better understand Hox function. |
Snir, O., Elgart, M., Gnainsky, Y., Goldsmith, M., Ciabrelli, F., Dagan, S., Aviezer, I., Stoops, E., Cavalli, G., Soen, Y. (2024). Organ transformation by environmental disruption of protein integrity and epigenetic memory in Drosophila. PLoS Biol, 22(5):e3002629 PubMed ID: 38805504
Summary: Despite significant progress in understanding epigenetic reprogramming of cells, the mechanistic basis of "organ reprogramming" by (epi-)gene-environment interactions remained largely obscure. This study used the ether-induced haltere-to-wing transformations in Drosophila as a model for epigenetic "reprogramming" at the whole organism level. The findings support a mechanistic chain of events explaining why and how brief embryonic exposure to ether leads to haltere-to-wing transformations manifested at the larval stage and on. Ether interferes with protein integrity in the egg, leading to altered deployment of Hsp90 and widespread repression of Trithorax-mediated establishment of active H3K4me3 chromatin marks throughout the genome. Despite this global reduction, Ubx targets and wing development genes preferentially retain higher levels of H3K4me3 that predispose these genes for later up-regulation in the larval haltere disc, hence the wing-like outcome. Consistent with compromised protein integrity during the exposure, the penetrance of bithorax transformations increases by genetic or chemical reduction of Hsp90 function. Moreover, joint reduction in Hsp90 and trx gene dosage can cause bithorax transformations without exposure to ether, supporting an underlying epistasis between Hsp90 and trx loss-of-functions. These findings implicate environmental disruption of protein integrity at the onset of histone methylation with altered epigenetic regulation of developmental patterning genes. The emerging picture provides a unique example wherein the alleviation of the Hsp90 "capacitor function" by the environment drives a morphogenetic shift towards an ancestral-like body plan. The morphogenetic impact of chaperone response during a major setup of epigenetic patterns may be a general scheme for organ transformation by environmental cues. |
Sokolov, V., Kyrchanova, O., Klimenko, N., Fedotova, A., Ibragimov, A., Maksimenko, O., Georgiev, P. (2024). New Drosophila promoter-associated architectural protein Mzfp1 interacts with CP190 and is required for housekeeping gene expression and insulator activity. Nucleic Acids Res, 52(12):6886-6905 PubMed ID: 38769058
Summary: In Drosophila, a group of zinc finger architectural proteins recruits the CP190 protein to the chromatin, an interaction that is essential for the functional activity of promoters and insulators. This study describes a new architectural C2H2 protein called Madf and Zinc-Finger Protein 1 (Mzfp1) that interacts with CP190. Mzfp1 has an unusual structure that includes six C2H2 domains organized in a C-terminal cluster and two tandem MADF domains. Mzfp1 predominantly binds to housekeeping gene promoters located in both euchromatin and heterochromatin genome regions. In vivo mutagenesis studies showed that Mzfp1 is an essential protein, and both MADF domains and the CP190 interaction region are required for its functional activity. The C2H2 cluster is sufficient for the specific binding of Mzfp1 to regulatory elements, while the second MADF domain is required for Mzfp1 recruitment to heterochromatin. Mzfp1 binds to the proximal part of the Fub boundary that separates regulatory domains of the Ubx and abd-A genes in the Bithorax complex. Mzfp1 participates in Fub functions in cooperation with the architectural proteins Pita and Su(Hw). Thus, Mzfp1 is a new architectural C2H2 protein involved in the organization of active promoters and insulators in Drosophila. |
Blanco, C., Xiang, W., Boumpas, P., Buvry, E., Scorcelletti, M., Hermon, A. S., Wong, J., Merabet, S., Carnesecchi, J. (2025). Synergistic DNA and RNA binding of the Hox transcription factor Ultrabithorax coordinates splicing and shapes in vivo homeotic functions. Nucleic Acids Res, 53(12) PubMed ID: 40613713
Summary: The dual interaction of many transcription factors (TFs) with both DNA and RNA is an underexplored issue that could fundamentally reshape understanding of gene regulation. This central issue was addressed by investigating the RNA binding activity of the Drosophila Hox TF Ultrabithorax (Ubx) in alternative splicing and morphogenesis. Relying on molecular and genetic interactions, this study uncovered a homodimerisation-dependent mechanism by which Ubx regulates splicing. Notably, this mechanism enables the decoupling of Ubx-DNA and -RNA binding activity in splicing. A critical residue for Ubx-RNA binding was identified, and the essential role of Ubx-RNA binding ability for its homeotic functions was identified. Overall, this study uncovered a unique mechanism for Ubx-mediated splicing and underscore the critical contribution of synergistic DNA/RNA binding for its morphogenetic functions. These findings advance understanding of co-transcriptional regulation and highlight the significance of TF-DNA/RNA synergistic function in shaping gene regulatory networks in living organisms. |
The homeotic genes encoded for by the Antennapedia and bithorax complexes control segment identity in the developing fly. This is developmental decision making for high stakes. Which segment should develop a wing? A leg? How many appendages should there be, and should appendages on different segments be the same or different from one another? Ultrabithorax regulates decisions regarding the number of wings and legs the adult will have.
Normal Drosophila have only one pair of wings, in contrast to the two pair found on butterflies. But Drosophila has the potential to develop a second set of wings. The haltere ( rhymes with "stare"), an external organ involved with balance, develops in place of a wing on the third thoracic segment. Given an appropriate mutation, the haltere could become a second set of wings.
A closer look at the work of Ubx in the determination of wing morphogenesis follows. Mutations of Ubx result in transformation of the dorsal and ventral appendages of the third thoracic segment (the haltere and third leg) into their counterparts on the second thoracic segment. More than a decade ago, it was concluded that normal Ubx expression in the third segment prevents second segment fate. In this manner Ubx mutations have the potential to produce a second set of wings (Struhl, 1982).
Can the repressive effects of Ubx be used to explain some of the difference between flies and butterflies? Three different studies both suggest the answer is no. Ubx is expressed in posterior thoracic segments in all insects, including butterflies and other modern four winged insects. There is, however, a lack of specificity homeotic transformation. Recently it has been shown that ectopic expression of Ultrabithorax, abdominal-A and Abdominal-B cause similar transformations of the fruitfly appendages: antennal tissue into leg tissue and wing tissue into haltere tissue. Thus the homeotic requirement to form appendages is, in some cases, non-specific (Casares, 1996). It is concluded that instead of there being a single evolutionary switch in regulation, there must have been multiple changes in many genes responsible for wing development regulation by Ubx, or for that matter, other homeotics (Warren, 1994 and Carroll, 1995).
However, regulation of larval abdominal prolegs does fit the expected result. Butterflies and Drosophila have diverged in the abdominal expression of Ubx (as opposed to thoracic expression). Early Ubx expression in the abdomen of butterflies is downregulated (shut off), allowing for the derepression of distal-less and the specification of prolegs; in Drosophila, Ubx expression remains high, inhibiting abdominal limb development (Warren, 1994). Thus, in the specification of prolegs, a single genetic change (Ubx expression) in the abdomen can explain the difference between flies and butterflies.
To complicate an already complicated picture, consider splice variants. UBX has a total of six different protein species (isozymes), generated from mRNA splice variants that are found in various Drosophila tissues. What possible function(s) could all these variants serve? For most multiply spliced RNAs the answer is not apparent, but for Ubx, a provocative answer is found.
One of the areas of developmental and molecular biology that has captured much attention and efforts to build a better understanding, is post-transcriptional processing of RNA. Genes are transcribed into high molecular weight nuclear RNA species that contain both the transcribed exons and the discarded introns. At a later time, this RNA is processed; the introns are removed and the exons are spliced together. It is this processed mRNA that serves to code for proteins.
Splicing factors are pivotal to the success of this process. These factors can recognize specific sequences in RNA, and based on sequence recognition, specifically remove unwanted introns and splice together the desired exons.
In RNAs subjected to alternative splicing (UBX provides a good example), different introns and exons are removed depending on particular tissue types. The result is different tissue specific splice variants, a total of six different protein isozymes coded for by the different splice variants of UBX. The UBX splice variants differ in the distance between the homeodomain and a domain responsible for interaction with Extradenticle, required as a coactivator on UBX target genes. It has been concluded that different UBX isozymes function effectively with EXD on different target genes dependent on the distance between the homeodomain and the interactive domain (Johnson, 1995).
This is a remarkable result. It means that the tissue specificity and developmental role of UBX is determined by splicing. What is true for UBX must be true for other transcription factors. The splicing factors available in a particular cell regulate the developmental fate of that cell by generating different splice variants of transcription factors. The arcane world of post-transcriptional regulation moves from a biochemical curiosity to the center ring of the developmental biology circus.
The differentiation of the Drosophila haltere from the wing through the action of the Ultrabithorax (Ubx) gene is a classic example of Hox regulation of serial homology. This study reveals several features of the control of haltere development by Ubx which, in principle, are likely to apply to the Hox-regulated differential development of other serially homologous structures in other animals. Specifically, it has been shown that Ubx acts: (1) at many levels of regulatory hierarchies, on long-range signaling proteins and their target genes, as well as genes further downstream; (2) selectively on a subset of downstream target genes of signals common to both wing and haltere, and (3) independently on these diverse targets. This information is presented in terms of the effects of Ubx on gene expression in the three axes of appendage formation, since these axes are to a large extent independently regulated and independent gene regulation in the axes serves to structure the entire wing (Weatherbee, 1998).
In the anterior posterior axis, Ubx represses selected Dpp target genes. The expression pattern of en is essentially the same in the haltere disc as in the wing disc, indicating that Ubx is not regulating haltere identity by altering the expression of this compartmental selector gene. Similarly, the expression of dpp in the developing haltere on the anterior side of the AP compartment boundary resembles that in the wing disc. Because these discs give rise to very different appendages, there may be genes downstream of the Dpp signal that are regulated by Ubx. To identify these, an examination was carried out of how a number of genes involved in the development of specific wing characters are expressed and regulated in the developing haltere (Weatherbee, 1998).
From its cellular site of secretion, Dpp acts as a morphogen to organize wing growth, AP pattern, and to activate target gene expression over a distance. The optomotor blind (omb), spalt (sal), and spalt related (salr) genes are expressed in nested patterns centered on the Dpp stripe and are necessary for proper development of the central wing region, including veins II-IV. The expression of these Dpp target genes was examined in the haltere disc: although omb is expressed in the developing haltere pouch (straddling the Dpp stripe as it does in the wing disc), salr and sal are not expressed in the haltere pouch. These results show that the Dpp signal transduction machinery operates in the haltere disc but that selected wing target genes are not activated by the Dpp signal. To determine whether Ubx represses salr expression in the haltere disc, homozygous Ubx clones were generated. Indeed, salr is derepressed in Ubx clones in the anterior compartment of the haltere disc. As in the wing disc, salr expression in these clones depends on their distance from the Dpp source. To determine whether Ubx is sufficient to repress salr, salr expression was examined in CbxM1/+ wing discs in which Ubx is ectopically expressed along part of the DV boundary. In these wing discs salr expression is repressed in a cell autonomous fashion. Because sal/salr are required for the induction of vein development, the selective repression of salr by Ubx suppresses part of the Dpp-mediated AP wing patterning program in the haltere. As with the spatial patterning of wing veins, the pattern of intervein tissue is also determined by specific regulatory genes and critical for morphogenesis. The Drosophila Serum Response Factor (DSRF, or blistered) gene is expressed in future intervein tissue and required for the adhesion of the dorsal and ventral surfaces of the flat wing. The haltere, however, is more balloon-like; interestingly, DSRF expression is absent from the haltere pouch except for two crescents at the extreme dorsal and ventral edges of the anterior compartment. This difference is caused by Ubx regulation, because in Ubx clones in the haltere disc, repression of DSRF is relieved and a pattern of DSRF expression homologous to that in the wing forms within the boundaries of the clone. Conversely, ectopic expression of Ubx in wing discs extinguishes DSRF expression in a cell-autonomous manner (Weatherbee, 1998).
In the dorsoventral axis, Ultrabithorax represses Wingless in the posterior compartment and selectively represses genes along the dorsal-ventral boundary. It has long been assumed that the global coordinate systems in homologous appendages are the same and, indeed, the apterous selector gene is expressed in the dorsal compartment of the haltere disc as in the wing. However, it was found that Wg, which is expressed along both the anterior and posterior extent of the DV boundary in the wing disc, is not expressed in the posterior compartment of the haltere disc. Because Wg function along the DV boundary is required for growth and patterning of the wing disc, the absence of Wg in the posterior haltere disc probably contributes to its disproportionately smaller size in comparison to the anterior compartment. In posterior Ubx clones in the haltere disc, Wg is expressed along the DV boundary, suggesting that Ubx represses the posterior portion of the Wg expression pattern. The activation of Wg along the DV boundary occurs via the Notch receptor signaling pathway. This pathway also activates the "boundary" enhancer of the vestigial gene, which is activated along the entire anterior and posterior extent of the DV boundary in the haltere. These results demonstrate that the Notch pathway is active along the entire DV boundary but that Ubx selectively prevents Wg activation by this pathway in the posterior compartment. Wg is expressed in the anterior compartment of the haltere disc, yet its phenotypic effects are markedly different from those in the anterior of the wing disc (Weatherbee, 1998).
The most conspicuous difference is that in the wing, Wg activity along the DV boundary induces the formation of the prominent triple and double rows of bristles along the wing margin, whereas in the haltere, it does not. The formation of margin bristles is regulated by Wg via the induction of the proneural achaete (ac) and scute (sc) target genes and also requires the Cut transcription factor. In the haltere disc, Cut is expressed along the anterior DV boundary, whereas ac and sc are not induced. To determine if Ubx represses ac/sc activation by Wg, Ubx clones were examined. In the haltere disc, sc expression is derepressed in clones that touch or cross the anterior portion of the DV boundary. Conversely, sc expression is lost in anterior wing disc cells that ectopically express Ubx. This repression by Ubx is sensitive to the dosage of Ubx activity, since ectopic ac/sc expression is observed in Ubx/+ haltere discs. This ectopic expression corresponds with ectopic bristles found on the halteres of Ubx/+ adults. Further reductions of Ubx function in haltere discs cause greater derepression of sc on the DV boundary and a corresponding emergence of triple row bristles on the adult haltere. The haltere has several types of sense organs, including the proximally located pedicellular sensillae, which are not present on the wing. Correspondingly, sc is expressed in the presumptive pedicellar portion of the haltere disc but not in the equivalent part of the wing disc. In Ubx clones in this region of the haltere disc, sc expression is lost. Therefore, Ubx is required to positively regulate sc in this unique pattern in the haltere disc. Together with the repression of sc along the DV boundary of the haltere, these observations suggest that Ubx acts on two independent domains of the sc expression pattern, presumably via specific cis-regulatory elements controlling each aspect of sc gene expression (Weatherbee, 1998).
In the proximodistal axis, Ubx selectively represses one enhancer of the vestigial gene. vg is expressed and required in the cells that will give rise to the distal appendage fields of the wing and haltere imaginal discs. vg expression in the wing field is regulated by two distinct enhancers that are activated by different signaling pathways. vg expression is first activated along the DV boundary of the wing disc by the Notch pathway, through the boundary enhancer; later it is activated in the growing wing pouch by the Dpp and Wg signals, through the "quadrant" enhancer. Similarly, the boundary enhancer is activated in both the wing and haltere discs, however, the quadrant enhancer is silent in the haltere field. The repression of the quadrant enhancer in the haltere is sensitive to the dosage of Ubx and is partially derepressed in Ubx/+ haltere discs. More importantly, in Ubx clones in the haltere disc, the quadrant enhancer is fully activated. These results show that Ubx selectively represses a portion of the native vg wing expression pattern in the haltere disc through the quadrant enhancer (Weatherbee, 1998).
In these experiments five genes have been identified whose function is necessary for the formation or patterning of various wing characters but whose expression is negatively regulated by Ubx in the haltere disc. For each gene, their repression in the haltere disc correlates with the absence of, or difference between, haltere characters and those in the serially homologous wing. One means by which to test the significance of the repression of these genes in the haltere disc is to determine what effects their derepression might have upon haltere morphology. It is crucial to recognize that the effects of expressing target genes in the haltere does not depend only on the sufficiency of a given gene to induce a phenotype in the wing or at an ectopic site (legs, eyes, etc.). Another crucial factor is the architecture of the Ubx-regulated gene hierarchies in the haltere. There are three possible outcomes and interpretations for the ectopic expression of a differentially expressed gene:
(1) Ectopic expression of individual genes in the haltere could be sufficient to induce a wing character. This result would show that the regulation of this gene by Ubx is the key event to determine the difference of that character in the wing and haltere.
(2) There could be no effect on haltere morphology. Given that these genes are sufficient to induce ectopic phenotypes in the wing or elsewhere, this result could occur if downstream genes are independently regulated by Ubx and therefore prevented from being activated even when upstream activators are present.
(3) One could induce haltere characters or structures with intermediate identity. This would imply that Ubx modifies the morphology of characters through other genes in addition to the ectopically expressed gene (Weatherbee, 1998).
The effects of ectopic expression of the vestigial gene was examined in the haltere and other tissues under the control of the GAL4/UAS system. Whereas vg expression in all other appendages and tissues causes wing-like outgrowths, in the haltere no significant change in adult appendage size or morphology was observed. However, striking differences between the morphology of the outgrowths formed on the second and third thoracic legs were observed. The former had clear wing-like morphology, whereas the latter had haltere-like morphology. The failure of ectopic vg expression to significantly alter haltere morphology and the distinct haltere-like character of the outgrowths formed in third thoracic legs suggests that Ubx acts on genes that are downstream of or parallel to vg in the genetic hierarchy. To test whether Ubx regulates genes downstream of vg, a search was carried out for candidate genes whose expression depends on Vg. The spalt and DSRF genes that are normally not expressed in leg imaginal discs are ectopically induced in first and second thoracic leg imaginal discs as a response to targeted expression of Vg, and may thus be activated in the developing wing through some mechanism that is dependent on Vg. The patterns of ectopic induction of sal and DSRF in T1 and T2 leg discs are reminiscent of their normal expression patterns in wing discs. In contrast to T3 leg discs, which also express Ubx, the central domains of ectopic induction of Sal and DSRF expression are suppressed. These results demonstrate that downstream targets of Vg are also regulated by Ubx, independent of the Ubx regulation of Vg itself. The repression of these and other targets by Ubx would then suggest why the deregulation of Vg expression in the developing haltere is insufficient to reprogram haltere development toward wing development and to alter the morphology of the adult haltere. Similarly, ectopic expression of the DSRF or Sal (M. Averof, personal communication to Weatherbee, 1998) transcription factors also does not alter haltere size, shape, or cell morphology. These results imply that there are genes downstream of DSRF and Sal whose expressions are necessary for the realization of a phenotype but which are repressed by Ubx in the haltere disc (Weatherbee, 1998)
Ectopic expression of the scute gene in the developing haltere is sufficient to induce ectopic sensory organs. Interestingly, near the DV boundary, large bristles resembling those of the wing margin are induced, whereas in more proximal regions, sense organs form that are characteristic of the haltere. This result suggests that the repression of sensory organ formation by Ubx at the DV boundary is largely at the level of the sc gene, whereas the character of the proximal sense organs is modified by Ubx action downstream of or parallel to scute. Thus, all three outcomes outlined above are obtained in these ectopic expression experiments, which reveal that Ubx acts independently on the five genes identified as well as on genes further downstream of or parallel to these regulators in the wing patterning hierarchy (Weatherbee, 1998).
The Drosophila Hox gene Ultrabithorax (Ubx) produces a family of protein isoforms through alternative splicing. Isoforms differ from one another by the presence of optional segments-encoded by individual exons-that modify the distance between the homeodomain and a cofactor-interaction module termed the 'YPWM' motif. To investigate the functional implications of Ubx alternative splicing, this study analyzed the in vivo effects of the individual Ubx isoforms on the activation of a natural Ubx molecular target, the decapentaplegic (dpp) gene, within the embryonic mesoderm. These experiments show that the Ubx isoforms differ in their abilities to activate dpp in mesodermal tissues during embryogenesis. Furthermore, using a Ubx mutant that reduces the full Ubx protein repertoire to just one single isoform, specific anomalies were obtained affecting the patterning of anterior abdominal muscles, demonstrating that Ubx isoforms are not functionally interchangeable during embryonic mesoderm development. Finally, a series of experiments in vitro reveals that Ubx isoforms also vary in their capacity to bind DNA in presence of the cofactor Extradenticle (Exd). Altogether, results indicate that the structural changes produced by alternative splicing have functional implications for Ubx protein function in vivo and in vitro. Since other Hox genes also produce splicing isoforms affecting similar protein domains, it is suggested that alternative splicing may represent an underestimated regulatory system modulating Hox gene specificity during fly development (Reed, 2010).
The experiments described in this study indicate that the generation of structural differences among Ubx proteins by alternative splicing is relevant for the functional specificity of Ubx in vivo. These structural features modulate essential biochemical properties of Ubx proteins such as their DNA-binding profiles in the presence of a cofactor (Reed, 2010).
The differential effects of Ubx in vivo are apparent in the posterior visceral mesoderm, but not in the anterior. To understand this it must first be noted that the mechanism of dpp674 visceral mesoderm enhancer repression is different anterior and posterior to PS7. In the anterior visceral mesoderm, repression requires Exd, since in Exd null mutants, dpp674 is ectopically expressed anterior to PS7. But Hox genes appear to play no role in the normal repression of dpp in this region. The same Exd-dependent mechanism may also be acting in the posterior, but it is not necessary, for no posterior ectopic expression is observed in Exd null mutants. The posterior repression depends instead on the Hox protein Abd-A, which is presumably able to repress dpp674 in the absence of Exd as a cofactor (Reed, 2010).
All Ubx protein isoforms are able to induce dpp ectopically in the anterior, suggesting that they can all override the normal repression mediated by Exd. However, they exert differential effects in the posterior, where Abd-A is the controlling repressor. Abd-A has a very similar DNA-binding specificity to that of Ubx. It binds to multiple sites in the dpp674 enhancer, including those to which Ubx binds. Through some of these sites it serves as an activator, but through others it acts as a repressor. With Ubx form Ia, the repressing effect by Abd-A is dominant. The results suggest that Ubx form IVa is either able to compete more effectively as an activator with the repressing action of Abd-A bound at other sites or able to displace the binding of Abd-A at the sites where it mediates repression (Reed, 2010).
Ubx form IVa also overrides the normal specificity of the dpp-lacZ enhancer for the visceral mesoderm, activating it ectopically in the somatic mesoderm of most trunk segments. It is not known whether the activity of this enhancer is normally restricted to the visceral mesoderm by a required cofactor that is present only in the visceral mesoderm, by repressors present in other tissues, or by both mechanisms. However, UbxIVa at high levels seems able to override this normal tissue specificity. The more efficient DNA binding of this isoform (in the presence of Exd) may bypass the requirement for a cofactor or displace a repressor more effectively (Reed, 2010).
A quantitatively controlled study showed that the levels of Ubx protein are very important to determine the functional outcome of Ubx in vivo. In this context, the results show that in spite of significant variation across expression, levels of UbxIVa protein in different transgenic lines, this isoform is consistently able to produce a similar output in terms of dpp target activation in posterior regions. This suggests that for UbxIVa dpp target activation and protein concentration may relate to one another in the form of a sigmoidal function with a narrow protein concentration interval acting as a threshold that is crossed by all UbxIVa lines tested in this study. Given that UbxIa lines achieving comparable protein expression levels to UbxIVa lines, it was possible to activate dpp in posterior regions of the embryo, it is concluded that qualitative differences in Ubx protein structure as determined by alternative splicing are causal to the observed differential behavior in target activation (Reed, 2010).
The tissue-specific effects of Ubx isoform ectopic expression emphasize the likely role that the splice isoforms play in mediating specific Ubx functions in different tissues. Indeed, the isoforms have different tissue distributions in embryogenesis with UbxIa expressed predominantly in epidermis, mesoderm, and peripheral nervous system whereas UbxIVa appears to be exclusively expressed in the central nervous system. The analysis of the UbxMX17 mutation, which retains the full Ubx expression pattern but generates only isoform UbxIVa, supports the endogenous relevance of splice isoforms for tissue-specific Hox function. Although in UbxMX17 embryos UbxIVa can replace the function of Ubx isoforms I and II in the epidermis with the generation of a normal cuticle pattern, the peripheral nervous system is affected, and in this study clear defects were found in the segmental specification of somatic muscles. Tissue-specific isoform functions may be mediated by effects on cofactor interaction or may also involve effects on collaborative regulatory interactions between Hox proteins and tissue-specific regulators (Reed, 2010).
The results of in vitro binding studies of the Ubx/Exd element show that Ubx isoforms differ in their capacity to interact with DNA in the presence of the cofactor Exd. A possibility that emerges from these results is that different Ubx isoforms display differential levels of interaction with Exd (in the presence of target DNA) and, accordingly, could perform specific functions in vivo as a consequence to the distinct levels of nuclear Exd available in different regions of the embryo (Reed, 2010).
in vitro studies also show that ablation of the YPWM motif has little effect on the ability of Ubx form Ia to form a complex with Exd, but it significantly reduces complex formation by Ubx form IVa (Reed, 2010).
The observation that mutated forms of the Ubx protein lacking the hexapeptide interact with Exd at all is at first sight surprising, especially in view of the structural studies showing that the YPWM motif provides the major contact between Hox and Exd proteins bound to DNA. However, this finding is not inconsistent with earlier work. The article that originally described cooperative interactions between Ubx and Exd used Ubx proteins deleted of all sequences located amino-terminal to the homeodomain. These proteins therefore lacked the hexapeptide motif entirely. Interactions with Exd that do not require the hexapeptide have also been reported in a study focused on Ubx functions independent from Exd and Hth proteins. In addition, alanine replacement of the hexapeptide does not affect the way another Hox protein (i.e., Abd-A) interacts with Exd. Furthermore, a recent study suggests that Ubx-Exd recruitment may rely not on a single, but on several different mechanisms, some of which require a short evolutionarily conserved motif originally termed UbdA. Thus, the hexapeptide is unlikely to be the only Hox protein motif that interacts with Exd (Reed, 2010).
Crystallographic studies show that the Ubx and Exd homeodomains are closely adjacent, almost touching each other, the extent of this proximity being revealed by a significant reduction in solvent-accessible surface area within the area of putative contact. In addition, the distance between the Ubx recognition helix C terminus and the Exd recognition helix N terminus is just 9 Å, with one particular residue of Ubx (Lys58) well positioned to form hydrogen bonds with a residue of Exd (Ser48). Thus, regions within the Ubx homeodomain may be responsible for additional interactions with Exd. Sequences elsewhere in the proteins may also contribute to these interactions. A fine combination of protein mapping and crystallographic studies may be required to reveal the structural details of such interactions (Reed, 2010).
These experiments would be consistent with the possibility that the Ubx linker region itself could be a previously uncharacterized region of Ubx that makes contacts with Exd. This would be supported by experiments of protein-protein interaction in yeast, which also suggest that the Ubx linker region could affect the interaction between Ubx and Exd, and by the high evolutionary conservation of these sequences across phylogenetically distant species of flies. The results could also be accommodated in a model in which the Ubx linker region induces a conformational change elsewhere in the Ubx protein, such that the degree of interaction with Exd is affected. Alternatively, the regulatory potential of the Ubx protein could be affected. In particular, linker-dependent conformational changes may affect the behavior of critical regulatory motifs of Ubx, such as the recently studied QA motif involved in the modulation of Ubx activities in a tissue-specific manner (Hittinger, 2005) and the SSYF motif (Tour, 2005), an evolutionarily conserved motif close to the N terminus of the Ubx protein that is involved in transcriptional activation (Reed, 2010).
Other reports have also emphasized that the Hox linker regions are not just passive spacers. One such study shows that mutation of the short linker region in the Hox protein Abd-A specifically disables its capacity to activate wingless, a natural target gene, without affecting its ability to repress dpp. When comparing cofactor and DNA-binding properties of this Abd-A mutant protein with those of the wild-type Abd-A on a repressor element of the Distalless gene (DllR), no differences were seen in the interaction between the mutant form of Abd-A and Exd protein. (It is perhaps worth noting that these in vitro experiments required the presence of Homothorax protein; on this particular DNA target (DllR), the formation of complexes with Abd-A and Exd alone was below the limit of detection of the assay. Another report described Ubx isoform-specific functions for the repression of the Dll gene. These studies suggest that sequences within the Hox linker regions may modify the trans-regulatory potential of Hox proteins without necessarily affecting the DNA-binding properties of these proteins. The results extend these studies, showing that linker regions may at times affect target gene regulation and the DNA binding/cofactor interaction abilities of a Hox protein. The binding results are consistent with a recent study that integrates DNA-binding tests with computational predictions of ordered and disordered segments of the Ubx protein (Liu, 2008), proposing that sequences outside the homeodomain can reduce the DNA-binding activities of the Ubx protein by twofold (Reed, 2010).
Beyond the effects that alternative splicing may have on modulating Hox interactions with cofactors, other possibilities must also be considered. Given that in these experiments a very unstable binding of the Ubx proteins to target DNA was observed in the absence of Exd, it is difficult to advance arguments regarding the affinities of the individual Ubx isoforms for this molecular target on firm grounds. In spite of this, given that in various physiological contexts the binding of Hox proteins to target sites in the absence of cofactors has been demonstrated, it is pertinent to suggest that alternative splicing might also be influencing Hox DNA binding independently from cofactor interactions, at the level of changing DNA-binding affinities of the individual isoforms for their target DNAs. This possibility would be suitable for experimental investigation in the physiological context of Drosophila adult appendage development, as Ubx is known to act independently from cofactors during the development of these structures (Reed, 2010).
Examples from other Drosophila Hox genes further support a more general role of differential splicing in the diversification of Hox function during development, affecting protein modules outside the Ubx linker region discussed above. For instance, genetic dissection of the Abd-B gene demonstrated the existence of two distinct gene functions, originally termed morphogenetic (m) and regulatory (r). The spectrum of mRNAs derived from the Abd-B locus have been cloned, and a family of Abd-B transcripts generated by differential promoter use has been revealed , that, in turn, leads to different splicing variants affecting 5' sequences of the gene. Two proteins products, called m and r, are produced from the Abd-B transcripts; m differs from r in that it encodes an additional large glutamine-rich amino-terminal domain. Furthermore, ectopic expression of each Abd-B protein class leads to specific effects on the larval cuticle, suggesting that the isoforms have specific developmental functions (Reed, 2010).
Given that in Drosophila several Hox proteins possess alternatively spliced modules modifying the distance between the hexapeptide and the homeodomain, while others produce functionally different isoforms via differential promoter use coupled to alternative splicing, alternative splicing may truly represent a very important, yet underexplored regulatory mechanism modulating the functional specificity of Hox proteins during development (Reed, 2010).
In Drosophila, differential development of wing and haltere, which differ in cell size, number and morphology, is dependent on the function of Hox gene Ultrabithorax (Ubx). This paper reports studies on Ubx-mediated regulation of the Fat/Hippo and IIS/dAkt pathways, which control cell number and cell size during development. Over-expression of Yki or down regulation of negative components of the Fat/Hippo pathway, such as expanded, caused considerable increase in haltere size, mainly due to increase in cell number. These phenotypes were also associated with the activation of Akt pathways in developing haltere. Although activation of Akt alone did not affect the cell size or the organ size, dramatic increase was observed in haltere size when Akt was activated in the background where expanded is down regulated. This was associated with the increase in both cell size and cell number. The organ appeared flatter than wildtype haltere and the trichome morphology and spacing resembled that of wing suggesting homeotic transformations. Thus, these results suggest a link between cellular growth and pattern formation and the final differentiated state of the organ (Singh, 2015).
Wing and haltere are the dorsal appendages of second and third thoracic segments, respectively, of adult Drosophila. They are homologous structures, although differ greatly in their morphology. The homeotic gene Ultrabithorax (Ubx), which is required and sufficient to confer haltere fate to epithelial cells, is known to regulate many wing patterning genes to specify haltere, but the mechanism is still poorly understood (Singh, 2015).
There are a number of differences between wing and haltere at the cellular and organ levels. Wing is a large, flat and thin structure, while haltere is a small globular structure, although both are made up of 2-layered sheet of epithelial cells. Space between the two layers of cells in haltere is filled with haemocytes. Cuticle area of each wing cell is 8 fold more than a haltere cell. Haltere has smaller and fewer cells than the wing. Trichomes of wing cells are long and thin, while haltere trichomes are short and stout in morphology. The ratio of anterior to posterior compartment size in the haltere (~2.5:1) is much different from that in the wing (~1.2:1). Haltere also lacks wing-type vein and sensory bristles. Haltere cells are more cuboidal compared to flatter wing cells (Roch, 2000). Thus, cell number, size and shape all add to the differences in the size and shape of the two organs (Singh, 2015).
However, cells of the third instar larval wing and haltere discs are similar in size and shape (Makhijani, 2007). The difference between cell size and shape becomes evident at late pupal stages (Roch, 2000). Wing cells become much larger, compared to haltere cells. At pupal stages, they also exhibit differences in the organization of actin cytoskeleton elements viz. F-actin levels are much higher in haltere cells compared to wing cells (Roch, 2000) (Singh, 2015).
In the context of final shape of wings and halteres, one needs to understand the mechanism by which Ubx influences cell size, shape and arrangement. It is possible that Ubx regulates overall shape of the haltere by regulating either cell size and/or shape. The current understanding of mechanisms by which wing and haltere differ at cellular, tissue and organ level is ambiguous (Sanchez-Herrero, 2013). For example, while removal of Ubx from the entire haltere, or at least from one entire compartment, leads to haltere to wing transformation with increased growth of Ubx minus tissues, mitotic clones of Ubx (using the null allele Ubx6.28) show similar sized twin spot in small clones (Crickmore, 2006, De Navas, 2006; Makhijani, 2007). Only when very large clones of Ubx6.28Ubx6.28 are generated, one can see increased growth compared to their twin spots (Crickmore, 2006). This suggests that unless a certain threshold level of growth factors is de-repressed, the haltere does not show any overgrowth phenotype (Singh, 2015).
There have been several efforts to identify functional and molecular mechanisms by which Ubx regulates genes/pathways to provide haltere its distinct morphology. Various approaches have been used to identify targets of Ubx that are expected to differentially express between wing and haltere, e.g., loss-of-function genetics, deficiency screens, enhancer-trap screening and genome wide approaches such as microarray analysis and chromatin immunoprecipitation (ChIP). Targets include genes involved in diverse cellular functions like components of the cuticle and extracellular matrix, genes involved in cell specification, cell proliferation, cell survival, cell adhesion, or cell differentiation, structural components of actin and microtubule filaments, and accessory proteins controlling filament dynamics (reviewed in Sanchez-Herrero, 2013; Singh, 2015).
Decapentaplegic (Dpp), Wingless (Wg), and Epidermal growth factor receptor (EGFR) are some of the major growth and pattern regulating pathways that are repressed by Ubx in the haltere (Weatherbee, 1998, Shashidhara, 1999; Prasad, 2003; Mohit, 2006; Crickmore, 2006, Pallavi, 2006; De Navas, 2006; Makhijani, 2007). However, over-expression of Dpp, Wg, Vestigial (Vg) or Vein (Vn) provides only marginal growth advantage to haltere compared to the wildtype. In this context, additional growth regulating pathways amongst the targets of Ubx were examined. Genome wide studies have identified many components of Fat/Hippo and Insulin-insulin like/dAkt signalling (IIS/dAkt) pathways as potential targets of Ubx. The Fat/Hippo pathway is a crucial determinant of organ size in both Drosophila and mammals. It regulates cell proliferation, cell death, and cell fate decisions and coordinates these events to specify organ size. In contrast, the IIS/dAkt pathway is known to regulate cell size (Singh, 2015).
Recent studies have revealed that the Fat/Hippo pathway networks with other signalling pathways. For example, during wing development, Fat/Hippo pathway activities are dependent on Four-jointed (Fj) and Dachous (Ds) gradients, which are influenced by Dpp, Notch, Wg and Vg. Glypicans, which play a prominent role in morphogen signalling, are regulated by Fat/Hippo signalling (Baena-Lopez, 2008). EGFR activates Yorkie (Yki; effector of Fat/Hippo pathway) through its EGFR-RAS-MAPK signalling by promoting the phosphorylation of Ajuba family protein WTIP (Reddy, 2013). However, EGFR negatively regulates events downstream of Yki (Herranz, 2012). The Fat/Hippo pathway is also known to inhibit EGFR signalling, which makes the interaction between the two pathways very complex and context-dependent. IIS/dAkt pathway is also known to activate Yki signalling and vice-versa. Thus, Fat/Hippo pathway may specify organ size by regulating both cell number (directly) and cell size (via regulating IIS/dAkt pathway) (Singh, 2015).
This study reports studies on the functional implication of regulation of Fat/Hippo and IIS/dAkt pathways by Ubx in specifying haltere development. Over-expression of Yki or down regulation of negative components of the Fat/Hippo pathway, such as expanded (ex), induced considerable increase in haltere size, mainly due to increase in cell number. Although activation of dAkt alone did not affect the organ size or the cell size, activation of Yki or down regulation of ex in the background of over-expressed dAkt caused dramatic increase in haltere size, much severe than Yki or ex alone. In this background, increase was observed in both cell size and cell number. The resulted haltere appeared flatter than wildtype haltere and the morphology of trichomes and their spacing resembled that of wing suggesting homeotic transformations. Thus, these results suggest a link between cellular growth and pattern formation and the final differentiated state of the organ (Singh, 2015).
The findings suggest that, downstream of Ubx, the Fat/Hippo pathway is critical for haltere specification. It is required for Ubx-mediated specification of organ size, sensory bristle repression, trichome morphology and arrangement. The Fat/Hippo pathway cooperates with the IIS/dAkt pathway, which is also a target of Ubx, in specifying cell size and compartment size in developing haltere. The fact that over-expression of Yki or downregulation of ex show haltere-to-wing transformations at the levels of organ size and shape, and trichome morphology and arrangement, suggest that regulation of the Fat/Hippo pathway by Ubx is central to the modification of wing identity to that of the haltere (Singh, 2015).
The observations made in this study pose new questions and suggest various interesting possibilities to study the Fat/Hippo pathway with a new perspective.
(1) It was observed that while Yki is nuclear in haltere discs, it appears to be non-functional. Yki is a transcriptional co-activator protein, which requires other DNA-binding partners for its activity. In this context, understanding the precise relationship between Yki and Ubx may provide an insight into mechanism of haltere specification (Singh, 2015).
(2) The Fat/Hippo pathway (along with the IIS/dAkt pathway) may be involved in the specification of cell size, trichome morphology and their arrangement, all of which are important parameters in determining organ morphology. Recent studies indicate that the Fat/Hippo pathway regulates cellular architecture and the mechanical properties of cells in response to the environment. It would be interesting to study the role of the Fat/Hippo pathway in regulating the cytoskeleton of epithelial cells during development. Haltere cells at pupal stages exhibit higher levels of F-actin than wing cells. One possible mechanism that is currently being investigated is lowering of F-actin levels in transformed haltere cells due to over-expression of Yki or down regulation of ex. This may cause flattening of cells during morphogenesis leading to larger organ size (Singh, 2015).
(3) Reversing cell size and number was sufficient to induce homeotic transformations at the level of haltere morphology. This suggests the importance of negative regulation of genetic mechanisms that determine cell size and number, in specifying an organ size and shape. As a corollary, Ubx-mediated regulation of Fat/Hippo and IIS/dAkt pathways provides an opportunity to study cooperative repression of cell number and cell size during organ specification (Singh, 2015).
(4) Certain genetic backgrounds investigated in this study showed severe effect on cell proliferation in haltere discs than in wing discs. This could be due to the fact that, the wing disc has already attained a specific size by the third instar larval stage (the developmental stage examined in this study), which is controlled by several pathways. Any change to this size may need more drastic alteration to the controlling mechanisms. As Ubx specifies haltere by modulating various wing-patterning events, there may still exist a certain degree of plasticity in mechanisms that determine the size of the haltere. However, in absolute terms, the haltere is also resistant to changes in growth control due to regulation by Ubx at multiple levels. Thus, differential development of wing and haltere provides a very good assay system to study not only growth control, but also to dissect out function of important growth regulators (tumour suppressor pathways) such as the Fat/Hippo pathway using various genome-wide approaches (Singh, 2015).
Drosophila bithorax complex (BX-C) is one of the best model systems for studying the role of boundaries (insulators) in gene regulation. Expression of three homeotic genes, Ubx, abd-A, and Abd-B, is orchestrated by nine parasegment-specific regulatory domains. These domains are flanked by boundary elements, which function to block crosstalk between adjacent domains, ensuring that they can act autonomously. Paradoxically, seven of the BX-C regulatory domains are separated from their gene target by at least one boundary, and must 'jump over' the intervening boundaries. To understand the jumping mechanism, the Mcp boundary was replaced with Fab-7 and Fab-8. Mcp is located between the iab-4 and iab-5 domains, and defines the border between the set of regulatory domains controlling abd-A and Abd-B. When Mcp is replaced by Fab-7 or Fab-8, they direct the iab-4 domain (which regulates abd-A) to inappropriately activate Abd-B in abdominal segment A4. For the Fab-8 replacement, ectopic induction was only observed when it was inserted in the same orientation as the endogenous Fab-8 boundary. A similar orientation dependence for bypass activity was observed when Fab-7 was replaced by Fab-8. Thus, boundaries perform two opposite functions in the context of BX-C-they block crosstalk between neighboring regulatory domains, but at the same time actively facilitate long distance communication between the regulatory domains and their respective target genes (Postaka, 2018).
The three homeotic (HOX) genes in the Drosophila Bithorax complex (BX-C), Ultrabithorax (Ubx), abdominal-A (abd-A) and Abdominal-B (Abd-B), are responsible for specifying cell identity in parasegments (PS) 5-14, which form the posterior half of the thorax and all of the abdominal segments of the adult fly. Parasegment identity is determined by the precise expression pattern of the relevant HOX gene and this depends upon a large cis-regulatory region that spans 300 kb and is subdivided into nine PS domains that are aligned in the same order as the body segments in which they operate. Ubx expression in PS5 and PS6 is directed by abx/bx and bxd/pbx, while abd-A expression in PS7, PS8, and PS9 is controlled by iab-2, iab-3, and iab-4. Abd-B is regulated by four domains, iab-5, iab-6, iab-7 and iab-8, which control expression in PS10, PS11, PS12 and PS13 respectively (Postaka, 2018).
Each regulatory domain contains an initiator element, a set of tissue-specific enhancers and Polycomb Response Elements (PREs) and is flanked by boundary/insulator elements. BX-C regulation is divided into two phases, initiation and maintenance. During the initiation phase, a combination of gap and pair-rule proteins interact with initiation elements in each regulatory domain, setting the domain in the on or off state. In PS10, for example, the iab-5 domain, which regulates Abd-B, is activated by its initiator element, while the more distal Abd-B domains, iab-6 to iab-8 are set in the off state. In PS11, the iab-6 initiator activates the domain, while the adjacent iab-7 and iab-8 domains are set in the off state. Once the gap and pair-rule gene proteins disappear during gastrulation, the on and off states of the regulatory domains are maintained by Trithorax (Trx) and Polycomb (PcG) group proteins, respectively (Postaka, 2018).
In order to select and then maintain their activity states independent of outside influence, adjacent regulatory domains are separated from each other by boundary elements or insulators. Mutations that impair boundary function permit crosstalk between positive and negative regulatory elements in adjacent domains and this leads to the misspecification of parasegment identity. This has been observed for deletions that remove five of the BX-C boundaries (Front-ultraabdominal (Fub), Miscadestral pigmentation (Mcp), Frontadominal-6 (Fab-6), Frontadominal-7 (Fab-7), and Frontadominal-8 (Fab-8)) (Postaka, 2018).
While these findings indicate that boundaries are needed to ensure the functional autonomy of the regulatory domains, their presence also poses a paradox. Seven of the nine BX-C regulatory domains are separated from their target HOX gene by at least one intervening boundary element. For example, the iab-6 regulatory domain must 'jump over' or 'bypass' Fab-7 and Fab-8 in order to interact with the Abd-B promoter. That the blocking function of boundaries could pose a significant problem has been demonstrated by experiments in which Fab-7 is replaced by heterologous elements such as scs, gypsy or multimerized binding sites for the architectural proteins dCTCF, Pita or Su(Hw). In these replacements, the heterologous boundary blocked crosstalk between iab-6 and iab-7 just like the endogenous boundary, Fab-7. However, the boundaries were not permissive for bypass, preventing iab-6 from regulating Abd-B (Postaka, 2018).
A number of models have been proposed to account for this paradox. One is that BX-C boundaries must have unique properties that distinguish them from generic fly boundaries. Since they function to block crosstalk between enhancers and silencers in adjacent domains, an appealing idea is that they would be permissive for enhancer/silencer interactions with promoters. However, several findings argue against this notion. For one, BX-C boundaries resemble those elsewhere in the genome in that they contain binding sites for architectural proteins such as Pita, dCTCF, and Su(Hw). Consistent with their utilization of these generic architectural proteins, when placed between enhancers (or silencers) and a reporter gene, BX-C boundaries block regulatory interactions just like boundaries from elsewhere in the genome. Similarly, there is no indication in these transgene assays that the blocking activity of BX-C boundaries are subject to parasegmental regulation. Also arguing against the idea that BX-C boundaries have unique properties, the Mcp boundary, which is located between iab-4 and iab-5, is unable to replace Fab-7. Like the heterologous boundaries, it blocks crosstalk, but it is not permissive for bypass. A second model is that there are special sequences, called promoter targeting sequence (PTS), located in each regulatory domain that actively mediate bypass. While the PTS sequences that have been identified in iab-6 and iab-7 enable enhancers to 'jump over' an intervening boundary in transgene assays, they do not have a required function in the context of BX-C and are completely dispensable for Abd-B regulation (Postaka, 2018).
A third model is suggested by transgene 'insulator bypass' assays. In one version of this assay, two boundaries instead of one are placed in between an enhancer and the reporter. When the two boundaries pair with each other, the enhancer is brought in close proximity to the reporter, thereby activating rather than blocking expression. Consistent with a possible role in BX-C bypass, these pairing interactions can occur over large distances and even skip over many intervening boundaries. The transgene assays point to two important features of boundary pairing interactions that are likely to be relevant in BX-C. First, pairing interactions are specific. For this reason boundaries must be properly matched with their neighborhood in order to function appropriately. A requirement for matching is illustrated in transgene bypass experiments in which multimerized binding sites for specific architectural proteins are paired with themselves or with each other. Bypass was observed when multimerized dCTCF, Zw5 or Su(Hw) binding sites were paired with themselves; however, heterologous combinations (e.g., dCTCF sites with Su(Hw) sites) did not support bypass. A second feature is that pairing interactions between boundaries are typically orientation dependent For example, scs pairs with itself head-to-head, not head-to-tail (Postaka, 2018).
If both blocking and bypass activities are intrinsic properties of fly boundaries then the BX-C boundaries themselves may facilitate contacts between the regulatory domains and their target genes. Moreover, the fact that both blocking and bypass activity are non-autonomous (in that they depend on partner pairing) could potentially explain why heterologous Fab-7 replacements like gypsy and Mcp behave anomalously while Fab-8 functions appropriately. Several observations fit with this idea. There is an extensive region upstream of the Abd-B promoter that has been implicated in tethering the Abd-B regulatory domains to the promoter and this region could play an important role in mediating bypass by boundaries associated with the distal Abd-B regulatory domains (iab-5, iab-6, iab-7). Included in this region is a promoter tethering element (PTE) that facilitates interactions between iab enhancers and the Abd-B promoter in transgene assays. Just beyond the PTE is a boundary-like element, AB-I. In transgene assays AB-I mediates bypass when combined with either Fab-7 or Fab-8. In contrast, a combination between AB-I and Mcp fails to support bypass. The ability of both Fab-7 and Fab-8 to pair with AB-I is recapitulated in Fab-7 replacement experiments. Unlike Mcp, Fab-8 has both blocking and bypass activity when inserted in place of Fab-7. Moreover, its bypass but not blocking activity is orientation-dependent. When inserted in the same orientation as the endogenous Fab-8 boundary, it mediates blocking and bypass, while it does not support bypass when inserted in the opposite orientation (Postaka, 2018).
Boundaries flanking the Abd-B regulatory domains must block crosstalk between adjacent regulatory domains but at the same time allow more distal domains to jump over one or more intervening boundaries and activate Abd-B expression. While several models have been advanced to account for these two paradoxical activities, replacement experiments argued that both must be intrinsic properties of the Abd-B boundaries. Thus Fab-7 and Fab-8 have blocking and bypass activities in Fab-7 replacement experiments, while heterologous boundaries including multimerized dCTCF sites and Mcp from BX-C do not. One idea is that Fab-7 and Fab-8 are simply 'permissive' for bypass. They allow bypass to occur, while boundaries like multimerized dCTCF or Mcp are not permissive in the context of Fab-7. Another is that they actively facilitate bypass by directing the distal Abd-B regulatory domains to the Abd-B promoter. Potentially consistent with an 'active' mechanism that involves boundary pairing interactions, the bypass activity of Fab-8 and to a lesser extent Fab-7 is orientation dependent (Postaka, 2018).
This study has tested these two models further. For this purpose the Mcp boundary was used for in situ replacement experiments. Mcp defines the border between the regulatory domains that control expression of abd-A and Abd-B. In this location, it is required to block crosstalk between the flanking domains iab-4 and iab-5, but it does not need to mediate bypass. In this respect, it differs from the boundaries that are located within the set of regulatory domains that control either abd-A or Abd-B, as these boundaries must have both activities. If bypass were simply passive, insertion of a 'permissive' Fab-7 or Fab-8 boundary in either orientation in place of Mcp would be no different from insertion of a generic 'non-permissive' boundary such as multimerized dCTCF sites. Assuming that Fab-7 and Fab-8 can block crosstalk out of context, they should fully substitute for Mcp. In contrast, if bypass in the normal context involves an active mechanism in which more distal regulatory domains are brought to the Abd-B promoter, then Fab-7 and Fab-8 replacements might also be able to bring iab-4 to the Abd-B promoter in a configuration that activates transcription. If they do so, then this process would be expected to show the same orientation dependence as is observed for bypass of the Abd-B regulatory domains in Fab-7 replacements (Postaka, 2018).
Consistent with the idea that a boundary located at the border between the domains that regulate abd-A and Abd-B need not have bypass activity, this study found that multimerized binding sites for the dCTCF protein fully substitute for Mcp. Like the multimerized dCTCF sites, Fab-7 and Fab-8 are also able to block crosstalk between iab-4 and iab-5. Blocking activity of Fab-7 is incomplete and there are small clones of cells in which the mini-y reporter is activated in A4. In contrast, the blocking activity of Fab-8 is comparable to the multimerized dCTCF sites and the mini-y reporter is off throughout A4. One plausible reason for this difference is that Mcp and the boundaries flanking Mcp (Fab-4 and Fab-6) utilize dCTCF as does Fab-8, while this architectural protein does not bind to Fab-7(Postaka, 2018).
Importantly, in spite of their normal (or near normal) ability to block crosstalk, both boundaries still perturb Abd-B regulation. In the case of Fab-8, the misregulation of Abd-B is orientation dependent just like the bypass activity of this boundary when it is used to replace Fab-7. When inserted in the reverse orientation, Fab-8 behaves like multimerized dCTCF sites and it fully rescues the Mcp deletion. In contrast, when inserted in the forward orientation, Fab-8 induces the expression of Abd-B in A4 (PS9), and the misspecification of this parasegment. Unlike classical Mcp deletions or the McpPRE replacement described here, expression of the Abd-B gene in PS9 is driven by iab-4, not iab-5. This conclusion is supported by two lines of evidence. First, the mini-y reporter inserted in iab-5 is off in PS9 cells indicating that iab-5 is silenced by PcG factors as it should be in this parasegment. Second, the ectopic expression of Abd-B is eliminated when the iab-4 regulatory domain is inactivated (Postaka, 2018).
These results, taken together with previous studies, support a model in which the chromatin loops formed by Fab-8 inserted at Mcp in the forward orientation brings the enhancers in the iab-4 regulatory domain in close proximity to the Abd-B promoter, leading to the activation of Abd-B in A4 (PS9). In contrast, when inserted in the opposite orientation, the topology of the chromatin loops formed by the ectopic Fab-8 boundary are not compatible with productive interactions between iab-4 and the Abd-B promoter. Moreover, it would appear that boundary bypass for the regulatory domains that control Abd-B expression is not a passive process in which the boundaries are simply permissive for interactions between the regulatory domains and the Abd-B promoter. Instead, it seems to be an active process in which the boundaries are responsible for bringing the regulatory domains into contact with the Abd-B gene. It also seems likely that bypass activity of Fab-8 (and also Fab-7) may have a predisposed preference, namely it is targeted for interactions with the Abd-B gene. This idea would fit with transgene bypass experiments, which showed that both Fab-7 and Fab-8 interacted with an insulator like element upstream of the Abd-B promoter, AB-I, while the Mcp boundary didn't (Postaka, 2018).
Similar conclusions can be drawn from the induction of Abd-B expression in A4 (PS9) when Fab-7 is inserted in place of Mcp. Like Fab-8, this boundary inappropriately targets the iab-4 regulatory domain to Abd-B. Unlike Fab-8, Abd-B is ectopically activated when Fab-7 is inserted in both the forward and reverse orientations. While the effects are milder in the reverse orientation, the lack of pronounced orientation dependence is consistent with experiments in which Fab-7 was inserted at its endogenous location in the reverse orientation. Unlike Fab-8 only very minor iab-6 bypass defects were observed. In addition to the activation of Abd-B in A4 (PS9) the Fab-7 Mcp replacements also alter the pattern of Abd-B regulation in more posterior segments. In the forward orientation, A4 and A5 are transformed towards an A6 identity, while A6 is also misspecified. Similar though somewhat less severe effects are observed in these segments when Fab-7 is inserted in the reverse orientation. At this point the mechanisms responsible for these novel phenotypic effects are uncertain. One possibility is that pairing interactions between the Fab-7 insert and the endogenous Fab-7 boundary disrupt the normal topological organization of the regulatory domains in a manner similar to that seen in boundary competition transgene assays. An alternative possibility is that Fab-7 targets iab-4 to the Abd-B promoter not only in A4 (PS9) but also in cells in A5 (PS10) and A6 (PS11). In this model, Abd-B would be regulated not only by the domain that normally specifies the identity of the parasegment (e.g., iab-5 in PS10), but also by interactions with iab-4. This dual regulation would increase the levels of Abd-B, giving the weak GOF phenotypes. Potentially consistent with this second model, inactivating iab-4 in the McpF8 replacement not only rescues the A4 (PS9) GOF phenotypes but also suppresses the loss of anterior trichomes in the A6 tergite (Postaka, 2018).
In metazoans, the pausing of RNA polymerase II at the promoter (paused Pol II) has emerged as a widespread and conserved mechanism in the regulation of gene transcription. While critical in recruiting Pol II to the promoter, the role transcription factors play in transitioning paused Pol II into productive Pol II is, however, little known. By studying how Drosophila Hox transcription factors control transcription, this study uncovered a molecular mechanism that increases productive transcription. The Hox proteins AbdA and Ubx target gene promoters previously bound by the transcription pausing factor M1BP, containing paused Pol II and enriched with promoter-proximal Polycomb Group (PcG) proteins, yet lacking the classical H3K27me3 PcG signature. AbdA binding to M1BP-regulated genes results in reduction in PcG binding, the release of paused Pol II, increases in promoter H3K4me3 histone marks and increased gene transcription. Linking transcription factors, PcG proteins and paused Pol II states, these data identify a two-step mechanism of Hox-driven transcription, with M1BP binding leading to Pol II recruitment followed by AbdA targeting, which results in a change in the chromatin landscape and enhanced transcription (Zouaz, 2017).
Understanding Hox transcriptional networks is central to understanding their wide repertoire of functions, yet observing where they bind in the genome does not explain why they bind there. In using a homogenous cell-based system devoid of endogenous Hox expression to conditionally express the Hox protein Ubx or AbdA, this study has demonstrated that Drosophila Hox proteins target proximal promoters genome-wide, which is conserved (for Ubx at least) in developing embryos. While studies into Hox genomic binding have historically focussed on enhancer elements in spatially and temporarily controlling individual gene expression, genome-wide promoter enrichment of Hox proteins is known to occur for mouse HoxB4 in hematopoietic stem cells, mouse Hoxa2 in the second branchial arches and for zebrafish Hoxb1a in early embryogenesis. However, why Hox proteins target the promoter-proximal region has been little explored. A major advantage of the Drosophila S2 cell system is that the conditional Hox expression system allows studying in fine detail the sequence of events occurring upon promoter binding and the impact on gene expression (Zouaz, 2017).
The promoters targeted by both AbdA and Ubx in Drosophila are essentially promoters containing either GAF or M1BP. GAF controls mainly development and morphogenic genes, whereas M1BP controls genes mainly involved in basic cellular processes (Li, 2013), and this distinction in gene ontology is reflected in the genes whose promoters are targeted by AbdA. As AbdA and Ubx also target enhancer regions, it cannot be ruled out that the observed promoter binding is the result of enhancer~promoter interaction. However, given that the majority of genes controlled by M1BP do not have distal enhancers (Zabidi, 2015), it is unlikely that this is the case for M1BP-targeted promoters. Both GAF and M1BP are important and distinct Drosophila Pol II pausing factors, a role that proved important in understanding the nature of promoters targeted by AbdA and Ubx, since the majority of all promoters targeted by the Hox proteins contained poised Pol II. GAF binding sites have previously been shown enriched at Ubx targets, although a link between Hox and GAF in regulating gene transcription was not demonstrated. Similarly, in S2-AbdA cells, AbdA binding at GAF-regulated promoters has little clear-cut effect on poised Pol II status, although the amount of elongating Pol II and gene transcription appears reduced. It was at M1BP-bound promoters where AbdA was found to have an effect on Pol II pausing, whereby AbdA binding results in a reduction in poised Pol II giving rise to increased productive transcription. Taken together with the findings that both Ubx and AbdA target nearly identical promoters, that AbdA and M1BP synergise in reporter gene expression, that both Ubx and AbdA interact with M1BP in embryos and AbdA colocalises with M1BP on polytene chromosomes, these data suggest functional cooperation between M1BP and AbdA/Ubx. To this end, demonstrating that M1BP expression is essential in inhibiting autophagy in the larval fat body, an Exd-independent cellular function shared by all Drosophila Hox proteins where the loss of expression of all Hox genes is essential for autophagy induction, suggests that M1BP may function with Hox proteins in their generic function of autophagy inhibition (Zouaz, 2017).
Similar to the distinct mechanisms at play in pausing Pol II at M1BP- and GAF-controlled promoters, these data suggest that distinct mechanisms of release of paused Pol II exist at the two classes of poised Pol II promoters: additional factors that are not present in S2 cells are likely required to permit Hox-induced productive transcription at GAF-controlled promoters since little evidence is foundthat AbdA binding affects Pol II pausing, whereas at M1BP-controlled genes, AbdA binding is sufficient to increase gene transcription through the release of paused Pol II (Zouaz, 2017).
Testing the association of AbdA ChIP peaks in S2-AbdA cells with those of numerous publicly available histone-modifying proteins and histone marks in S2 cells, this study found that the M1BP- and GAF-poised Pol II promoters targeted by AbdA were enriched for PcG proteins and H3K4me3. The finding that AbdA-enhanced transcription at M1BP promoters was more consistently concomitant with a loss of promoter PcG protein binding than at GAF-controlled promoter, suggests that the emerging role for PcG proteins in maintaining a poised Pol II state can be perturbed by Hox binding. Indeed, it is noteworthy that of the PcG proteins tested here, it is promoter-bound dRing that is most affected upon AbdA binding, suggesting that, like in vertebrates where Ring1 plays a major role in restraining the poised Pol II at promoters, dRing may play a major role in tethering the poised Pol II state in Drosophila. Given that no clear effect on PcG binding occurs at GAF-controlled promoters, even when these genes are repressed upon AbdA binding, it reinforces the notion that contrary to M1BP targets, the control of gene expression by AbdA at GAF genes is unlikely to occur through the regulation of poised Pol II status (Zouaz, 2017).
Where PcG proteins are linked to maintaining gene repression, trithorax group proteins (trxG) are the PcG antagonists, responsible for maintaining gene expression. As a transcription factor, GAF has been traditionally classified as a trxG protein although it displays repressive activity and can recruit PcG complexes. As such, GAF can be classified as a member of the growing family of genes that display both PcG and trxG phenotypes, the so-called enhancers of trithorax and polycomb (ETP) family. This study shows that M1BP colocalises with PcG proteins at promoters in S2 cells and phenotypically enhances the PcG homeotic phenotype of extra male sex combs on the second and third pairs of legs. Indeed, M1BP-/Pc- transheterozygous males have an average of 5.3 legs displaying sex combs, which is more than most combinations of PcG mutant transheterozygotes, demonstrating the large increase in penetrance of the Pc phenotype upon M1BP mutation. As such, M1BP would be genetically classified as a PcG gene. However, PcG genes, by definition, display homeotic phenotypes due to the derepression of Hox genes when mutated and so since this study observed neither increased derepression of the upstream Hox gene responsible for sex comb development, Scr, upon M1BP mutation nor Hox expression in fat body cells following RNAi, M1BP cannot thus be classified as a PcG gene. Given that M1BP is a transcription factor involved in gene expression (Li, 2013), the hypothesis is therefore favoured that, like GAF, M1BP is likely to be a member of the ETP family. How ETP proteins can enhance the phenotypes of both repressors (PcG) and activators (trxG) has long remained a mystery. Demonstrating here that GAF and M1BP colocalise with PcG at poised Pol II promoters with the loss of PcG at those genes displaying increased expression upon AbdA binding, may go a long way to better understand how transcription factors and transcriptional repressors intricately cooperate to regulate gene transcription (Zouaz, 2017).
In summary, this work identifies a novel mechanism for Pol II pausing release mediated by AbdA: at genes bound by M1BP, targeting of AbdA results in the specific loss of PcG proteins, the release of poised Pol II and increases in H3K4me3 histone marks, which results in promoting productive transcription. Identified in S2 cells where Hox PBC-class cofactors are absent, this mechanism may more generally apply to Hox-generic functions that are independent of PBC-class cofactors, such as the repression of autophagy in the Drosophila fat body or sex comb development in Drosophila males. It may also apply to PBC-dependent Hox target gene regulatien by cooperating with Hox PBC-bound genomic regions located remote from the promoter. Further work aimed at studying Hox PBC-bound enhancers together with poised Pol II status, and promoter-proximal Hox and PcG binding, should provide further insight into how enhancer-bound protein complexes influence the basic mechanisms of transcription regulated through poised Pol II. Uncovering such a Hox-driven mechanism of gene regulation by sequence-specific transcription factors, PcG proteins and poised Pol II in the developing animal would have been fraught with difficulties, not least of which is the quagmire of PcG proteins being essential global repressors of all Hox genes (Zouaz, 2017).
Mutations in the Ultrabithorax (Ubx) gene cause homeotic transformation of the normally two-winged Drosophila into a four-winged mutant fly. Ubx encodes a HOX family transcription factor that specifies segment identity, including transformation of the second set of wings into rudimentary halteres. Ubx is known to control the expression of many genes that regulate tissue growth and patterning, but how it regulates tissue morphogenesis to reshape the wing into a haltere is still unclear. This study shows that Ubx acts by repressing the expression of two genes in the haltere, Stubble and Notopleural, both of which encode transmembrane proteases that remodel the apical extracellular matrix to promote wing morphogenesis. In addition, Ubx induces expression of the Tissue inhibitor of metalloproteases in the haltere, which prevents the basal extracellular matrix remodelling necessary for wing morphogenesis. These results provide a long-awaited explanation for how Ubx controls morphogenetic transformation (Diaz-de-la-Loza, 2020).
The results reveal how Ubx – a homeotic gene that encodes the founding member of the HOX-family of transcription factors – regulates apical and basal matrix remodelling to control epithelial morphogenesis (see Ubx controls apical and basal ECM degradation to regulate morphogenesis). Ubx strongly represses two genes encoding apical matrix proteases (Np and Sb), as well as partially repressing two genes encoding basal matrix metalloproteases (Mmp1 and Mmp2), while inducing an inhibitor of Mmp1/2 (Timp) in the haltere. In this way, Ubx prevents both apical and basal matrix remodelling in the haltere, a key event in the homeotic wing-to-haltere transformation. In addition to regulating morphogenesis, Ubx controls many other genes affecting wing growth and pattern. Together, the combined repression of morphogenesis, growth and patterning by Ubx is responsible for the full transformation of wing to haltere (Diaz-de-la-Loza, 2020).
Ubx controls apical and basal ECM degradation to regulate morphogenesis. Schematic of Ubx expression and function in Drosophila and a hypothetical four-winged ancestor. Ubx controls organ shape via regulation of aECM and bECM proteases, in addition to its known functions in regulating organ growth and patterning. These target genes have presumably evolved to be specifically regulated in the Drosophila wing and/or haltere, and must be insensitive to Ubx in four-winged ancestors (Diaz-de-la-Loza, 2020).
These findings also support the general view that transcriptional control of matrix synthesis and degradation is a conserved mechanism by which information encoded in the genome is deployed to govern the shape of tissues and organs in animals. Although this concept is broadly appreciated for the regulation of the bECM, the notion that the aECM is also developmentally regulated during tissue morphogenesis needs further investigation, particularly in mammals. Beyond animals, morphogenesis of plants, fungi and bacteria is also known to be fundamentally dependent on patterned synthesis and degradation of the cell wall, a type of ECM. Thus, genetic control of the matrix appears to be a general principle that shapes all life forms (Diaz-de-la-Loza, 2020).
In eukaryotes, members of transcription factor families often exhibit similar DNA binding properties in vitro, yet orchestrate paralog-specific gene regulatory networks in vivo. The serially homologous first (T1) and third (T3) thoracic legs of Drosophila, which are specified by the Hox proteins Scr and Ubx, respectively, offer a unique opportunity to address this paradox in vivo. Genome-wide analyses using epitope-tagged alleles of both Hox loci in the T1 and T3 leg imaginal discs, the precursors to the adult legs and ventral body regions, show that ~8% of Hox binding is paralog-specific. Binding specificity is mediated by interactions with distinct cofactors in different domains: the Hox cofactor Exd acts in the proximal domain and is necessary for Scr to bind many of its paralog-specific targets, while in the distal leg domain, the homeodomain protein Distal-less (Dll) enhances Scr binding to a different subset of loci. These findings reveal how Hox paralogs, and perhaps paralogs of other transcription factor families, orchestrate alternative downstream gene regulatory networks with the help of multiple, context-specific cofactors (Feng, 2022).
This study used a combination of whole-genome and mechanistic approaches to understand how serially homologous appendages, such as the fly T1 and T3 legs, obtain their unique morphologies due to the activities of parallel Hox gene networks. The very similar transcriptomes in the three pairs of leg discs suggest that the different morphologies are largely a consequence of changing the expression patterns of the same sets of genes. By comparing the genome-wide DNA-binding profiles of the two relevant Hox paralogs, Scr and Ubx, in their native physiological contexts, this study found hundreds of paralog-specific Hox targets, accounting for ~8% of all binding events for these two Hox proteins. Next, differences in chromatin accessibility and Hox monomer binding preferences were shown to be unlikely to account for paralog-specific binding. Instead, it was demonstrated that interaction with the Hox cofactor Exd explains a large fraction of Scr's paralog-specific binding events. Finally, this study identified Dll as a Hox cofactor in the complementary distal domain of the leg disc. Results from RNA-seq, CBP ChIP, and reporter assays suggest that about 1/3 of the paralog-specific Scr-binding events are functional and lead to tissue-specific gene regulation. Thus, paralog-specific Hox-DNA binding, which is mediated by multiple cofactors including Exd and Dll, contribute significantly to paralog-specific Hox gene networks (Feng, 2022).
Previous in vitro studies provided compelling evidence that the DNA-binding specificities of different Hox-Exd dimers are more divergent from each other than those of Hox monomers, a phenomenon termed latent specificity. There have also been several in vivo examples in which paralog-specific Hox-DNA binding and target regulation was shown to depend on an interaction with Exd. This study shows that, on a genome-wide scale, the interaction with Exd explains a significant fraction of paralog-specific Hox binding, which often leads to paralog-specific gene regulation (Feng, 2022).
Earlier work also suggested that there is a tradeoff between specificity and affinity for Hox-Exd-binding motifs, where high affinity binding motifs are more likely to have low specificity for different Hox-Exd heterodimers. The paralog-specific, Exd-dependent CRMs characterized in this study (ac-1, h-1, and fj<-1), have higher affinity Hox-Exd-binding motifs than those previously described in the shavenbaby (svb) gene: the major Scr-Exd motif in the fj-1 CRM has an affinity of about 0.06 relative to the optimal motif in the genome, while the motifs in ac-1-1 and h-1 have even higher relative affinities of nearly 0.15 and 0.2, respectively. In contrast, the Ubx-Exd-binding motifs in CRMs from svb have a relative affinity of <0.0118. One possible explanation for this difference is that the svb CRMs are active in embryos, which have many different cell types, while the CRMs characterized in this study are active in leg discs, which have significantly less cell-type complexity. Embryonic CRMs may require especially low-affinity binding motifs to distinguish their activities in a context with many cell types. Consistent with this idea, the fkh250 CRM, which is also active in embryos, uses an Scr-Exd-binding motif with a low relative affinity of 0.01718,36. Notably, the relative affinities for the Scr-Exd-binding motifs in ac-1, h-1, and fj-1 are at least eightfold higher than for Ubx-Exd. Manual inspection of other intergenic and intronic loci with ScrT1 > UbxT3 binding suggests that there are many other CRMs that follow this same rule. Thus, for specificity to occur, the most relevant feature may be that the affinity for the 'correct' TFs, in this case Scr-Exd, must be significantly greater compared to the affinity for other 'incorrect' TFs that are co-expressed in the same or homologous cells (Feng, 2022).
Because Exd is only nuclear in a subset of cells during Drosophila development, such as the proximal domain of the leg disc, it was unlikely that Exd was the only Hox cofactor. In fact, CRMs that are directly regulated by Ubx have been described in cells where Exd is not available to be a cofactor. However, it has remained an unresolved question whether non-Exd cofactors are used in these examples. More generally for the leg imaginal disc, the entire distal domain, extending from the trochanter to the tarsus, is without nuclear Exd, yet has Hox-dependent segment-specific morphological characteristics, such as the sex combs on the male T1 leg. Although several candidate TFs have been proposed to be Hox cofactors, none have been confirmed. This study provides evidence that Dll is a distally acting Hox cofactor in leg discs (Feng, 2022).
There are many differences between how Exd and Dll interact with Hox proteins when bound to DNA. The Scr-Exd-binding motif is comprised of two partially overlapping half-sites, while the Scr-Dll motif consists of two HD-binding motifs separated by a spacer of several base pairs. Another difference is that the amount of cooperativity observed for Hox-Exd is far greater than that observed for Hox-Dll. The overlapping nature of the Hox- and Exd-binding motifs may be important for latent specificity, which for Scr requires an Exd-induced conformational change of the homeodomain. In contrast, there is no evidence that latent specificity occurs as a consequence of Hox-Dll binding. Instead, the modest cooperativity observed for the Scr-Dll heterodimer is likely a consequence of increasing Scr-DNA-binding affinity via a protein-protein interaction and closely spaced Dll and Scr-binding motifs (Feng, 2022).
More generally, it is suggested that mode of DNA binding exhibited by Hox-Exd, which is highly cooperative and reveals latent specificity, may be the exception rather than the rule for TF-TF interactions within CRMs, and that the Scr-Dll example, with weak cooperativity between TFs stemming from a protein-protein interaction, may be the more common mode of interaction to distinguish the binding of paralogous TFs. In support of this notion, a systematic in vitro study identified 315 TF-TF interactions, only five of which exhibited latent specificity (Feng, 2022).
The TALE homeodomain proteins, which include Exd and Hth, are very ancient TFs that were present before the split of plants and animals, and TALE-mediated nuclear localization analogous to the Hth-Exd example in flies has been described in plants. In contrast, the Hox gene family is only present in metazoans, and Dll is specific to bilaterians. Moreover, it has been proposed that Dll initially functioned in the CNS, and was later co-opted to pattern the distal appendage. Based on these observations, it is plausible that the Hox-Dll interaction evolved more recently than the Hox-Exd interaction, accounting for why Exd interacts with all Hox paralogs, while Dll may be a more limited Hox cofactor. This is supported by the results from a small-scale bimolecular fluorescence complementation (BiFC) screen that revealed Dll interacts with some Hox proteins, but not others (Feng, 2022).
Notably, the combined activities of Exd and Dll still do not account for all ScrT1 > UbxT3-binding events genome-wide and reporter analysis suggests the presence of additional, yet to be identified Hox cofactors that have the capacity to promote Scr-specific binding. It is suggested that the Hox-Dll mode of binding uncovered in this study may be representative of additional TFs that also have the ability to promote paralog-specific Hox binding and activity at specific CRMs. Further, it is noted that the differentiation of the T1 and T3 leg fates is a continuous developmental process and that the observations described in this study are limited to the late 3rd instar stage. Nevertheless, it is expected that the principles governing Hox paralog specificity uncovered in this study will likely extend to other developmental stages and tissues. Finally, although this study focused on the role of paralog-specific TF-DNA binding, there may be additional mechanisms that do not depend on differences in DNA binding between paralogous TFs that also contribute to their specific functions (Feng, 2022).
There are two transcripts that differ in their 3'UTR length as determined by two termination signals. UBX is coded for by the following four or five elements: a 5' exon containing the 5'UTR and the N-terminal coding sequence, two micro-exons that are incorporated in certain splice variants, an optional B element that is made part of the C terminal exon in some splice variants, and a C terminal exon containing both the homeodomain and the 3'UTR. There are a total of six specific splice variants: two expressed in epidermis, mesoderm and the peripheral nervous system; two in the central nervous system, epidermis, mesoderm and peripheral nervous system, and two in the central nervous system only. The micro-exons each code for 17 amino acids; the B element codes for 9 amino acids. Therefore the smallest protein species contains 43 fewer amino acids than the largest (O'Connor, 1988, Kornfield, 1989 and Subramanian, 1994).
genomic DNA length - 76 kb
cDNA clone length - 3.2 and 4.6 kb
Bases in 5' UTR - 964
Bases in 3' UTR - 1580 and 2212
Alternative splicing of UBX transcript generates a family of five proteins (UBX isoforms) that function as transcription factors. All isoforms contain a homeodomain within a common 99 aa C-terminal region (C-constant region) joined to a common 247 aa N-terminal (N-constant) region by different combinations of three small optional elements. UBX isoforms expressed in D. melanogaster cells are phosphorylated on serine and threonine residues, located primarily within a 53 aa region near the middle of the N-constant region, to form at least five phosphorylated states per isoform. Temporal developmental profiles of UBX isoforms parallel those for the respective mRNAs; all isoforms are similarly phosphorylated throughout embryogenesis (Gavis, 1991).
The homeodomain is C-terminal. There is a 15 residue N-terminal extention to the homeodomain that includes a WPWM motif. This motif determines interaction with Extradenticle. The size and sequence of the region between the WPWM element and the homeodomain differ among the UBX isoforms. This variable region affects interaction with EXD (Johnson, 1995).
See four paralogous Hox clusters of mammals for homologies of Ultrabithorax with mammalian Hox proteins.
date revised: 25 March 2026
Home page: The Interactive Fly © 2026 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
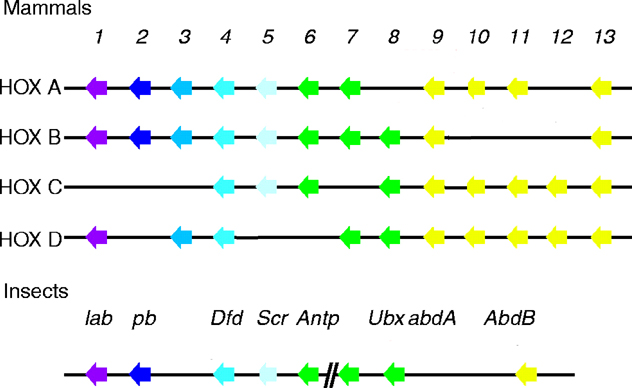{kind=link}