|
Gene name - Antennapedia Synonyms - Cytological map position - 84B1-2 Function - transcription factor Keywords - Antennapedia complex, homeotic, segment identity, leg selector transcription factor, specifies segmental identity in the pro- and mesothorax |
Symbol - Antp FlyBase ID:FBgn0260642 Genetic map position - 3-47.5 Classification - homeodomain - Antp class Cellular location - nucleus |
| Recent literature | Benmimoun, B., Polesello, C., Haenlin, M. and Waltzer L. (2015). The EBF transcription factor Collier directly promotes Drosophila blood cell progenitor maintenance independently of the niche. Proc Natl Acad Sci 112(29):9052-7. PubMed ID: 26150488 Summary: The maintenance of stem or progenitor cell fate relies on intrinsic factors as well as local cues from the cellular microenvironment and systemic signaling. In the lymph gland, an hematopoietic organ in Drosophila larva, a group of cells called the Posterior Signaling Centre (PSC), whose specification depends on the EBF transcription factor Collier (Col) and the HOX factor Antennapedia(Antp), has been proposed to form a niche required to maintain the pool of hematopoietic progenitors (prohemocytes). In contrast with this model, this study shows that genetic ablation of the PSC does not cause an increase in blood cell differentiation or a loss of blood cell progenitors. Furthermore, although both col and Antp mutant larvae are devoid of PSC, the massive prohemocyte differentiation observed in col mutant is not phenocopied in Antp mutant. Interestingly, beside its expression in the PSC, Col is also expressed at low levels in prohemocytes and it was shown that this expression persists in PSC-ablated and Antp mutant larvae. Moreover, targeted knockdown and rescue experiments indicate that Col expression is required in the prohemocytes to prevent their differentiation. Together, this study shows that the PSC is dispensable for blood cell progenitor maintenance and reveals the key role of the conserved transcription factor Col as an intrinsic regulator of hematopoietic progenitor fate. |
Gabilondo, H., Stratmann, J., Rubio-Ferrera, I., Millán-Crespo, I., Contero-García, P., Bahrampour, S., Thor, S. and Benito-Sipos, J. (2016). Neuronal cell fate specification by the convergence of different spatiotemporal cues on a common terminal selector cascade. PLoS Biol 14: e1002450. PubMed ID: 27148744 Summary: Specification of the myriad of unique neuronal subtypes found in the nervous system depends upon spatiotemporal cues and terminal selector gene cascades, often acting in sequential combinatorial codes to determine final cell fate. However, a specific neuronal cell subtype can often be generated in different parts of the nervous system and at different stages, indicating that different spatiotemporal cues can converge on the same terminal selectors to thereby generate a similar cell fate. However, the regulatory mechanisms underlying such convergence are poorly understood. The Nplp1 neuropeptide neurons in the Drosophila ventral nerve cord can be subdivided into the thoracic-ventral Tv1 neurons and the dorsal-medial dAp neurons. The activation of Nplp1 in Tv1 and dAp neurons depends upon the same terminal selector cascade: col->ap/eya->dimm->Nplp1. However, Tv1 and dAp neurons are generated by different neural progenitors (neuroblasts) with different spatiotemporal appearance. It was found that the same terminal selector cascade is triggered by Kr/pdm->grn in dAp neurons, but by Antp/hth/exd/lbe/cas in Tv1 neurons. Hence, two different spatiotemporal combinations can funnel into a common downstream terminal selector cascade to determine a highly related cell fate. |
Papadopoulos, D. K., Skouloudaki, K., Engstrom, Y., Terenius, L., Rigler, R., Zechner, C., Vukojevic, V. and Tomancak, P. (2019). Control of Hox transcription factor concentration and cell-to-cell variability by an auto-regulatory switch. Development. PubMed ID: 30642837
Summary: The variability in transcription factor concentration among cells is an important developmental determinant, yet how variability is controlled remains poorly understood. Studies of variability have focused predominantly on monitoring mRNA production noise. Little information exists about transcription factor protein variability, since this requires the use of quantitative methods with single-molecule sensitivity. Using Fluorescence Correlation Spectroscopy (FCS), this study characterized the concentration and variability of 14 endogenously tagged TFs in live Drosophila imaginal discs. For the Hox TF Antennapedia this study investigated whether protein variability results from random stochastic events or is developmentally regulated. Antennapedia was found to transition from low concentration/high variability early, to high concentration/low variability later, in development. FCS and temporally resolved genetic studies uncovered that Antennapedia itself is necessary and sufficient to drive a developmental regulatory switch from auto-activation to auto-repression, thereby reducing variability. This switch is controlled by progressive changes in relative concentrations of preferentially activating and repressing Antennapedia isoforms, which bind chromatin with different affinities. Mathematical modelling demonstrated that the experimentally supported auto-regulatory circuit can explain the increase of Antennapedia concentration and suppression of variability over time. |
Zandvakili, A., Uhl, J. D., Campbell, I., Salomone, J., Song, Y. C. and Gebelein, B. (2018). The cis-regulatory logic underlying abdominal Hox-mediated repression versus activation of regulatory elements in Drosophila. Dev Biol. PubMed ID: 30468713
Summary: During development diverse transcription factor inputs are integrated by cis-regulatory modules (CRMs) to yield cell-specific gene expression. Defining how CRMs recruit the appropriate combinations of factors to either activate or repress gene expression remains a challenge. This study compares and contrasts the ability of two CRMs within the Drosophila embryo to recruit functional Hox transcription factor complexes. The Distal-less DCRE CRM recruits Ultrabithorax (Ubx) and Abdominal-A (Abd-A) Hox complexes that include the Extradenticle (Exd) and Homothorax (Hth) transcription factors to repress the Distal-less leg selector gene, whereas the RhoA CRM selectively recruits Abd-A/Exd/Hth complexes to activate rhomboid and stimulate Epidermal Growth Factor secretion in sensory cell precursors. By swapping binding sites between these elements, it was found that the RhoA Exd/Hth/Hox site configuration that mediates Abd-A specific activation can convey transcriptional repression by both Ubx and Abd-A when placed into the DCRE. It was further shown that the orientation and spacing of Hox sites relative to additional binding sites within the RhoA and DCRE is critical to mediate cell- and segment-specific output. These results indicate that the configuration of Exd, Hth, and Hox site within RhoA is neither Abd-A specific nor activation specific. Instead Hox specific output is largely dependent upon the presence of appropriately spaced and oriented binding sites for additional TF inputs. Taken together, these studies provide insight into the cis-regulatory logic used to generate cell-specific outputs via recruiting Hox transcription factor complexes. |
de Mello, L. R., Hamley, I. W., Castelletto, V., Garcia, B. B. M., Han, S. W., de Oliveira, C. L. P. and da Silva, E. R. (2019). Nanoscopic structure of complexes formed between DNA and the cell-penetrating peptide Penetratin. J Phys Chem B. PubMed ID: 31465229
Summary: One of the most remarkable examples of cell-penetrating peptides (CPPs) is Penetratin, a 16-mer fragment derived from the Drosophila Antennapedia homeobox. Understanding the structure of Penetratin/DNA complexes is a key factor for the successful design of new vectors for gene delivery and may assist in optimizing molecular carriers based on CPPs. This srudy presents a comprehensive study on the nanoscale structure of noncovalent complexes formed between Penetratin and DNA. The strong cationic nature of the peptide makes it a very efficient agent for condensing DNA strands via electrostatic attraction, and this study shows that DNA condensation is accompanied by random-to-beta-sheet transitions of Penetratin secondary structure, demonstrating that nucleic acids behave as a structuring agent upon complexation. Nanoscale-resolved spectroscopy was used to provide single-particle infrared data from DNA carriers based on CPPs, and they show that the structures are stabilized by Penetratin beta-sheet cores, whereas larger DNA fractions are preferentially located in the periphery of aggregates. In-solution infrared assays indicate that phosphate diester groups are strongly affected upon DNA condensation, presumably as a consequence of charge delocalization induced by the proximity of cationic amide groups in Penetratin. The morphology is characterized by nanoassemblies with surface fractal features, and short-range order is found in the inner structure of the scaffolds. Interestingly, the formation of beads-on-a-string arrays is found, producing nanoscale architectures that resemble structures observed in early steps of chromatin condensation. A complexation pathway where DNA condensation and peptide pairing into beta-sheets are key steps for organization is proposed. |
Postika, N., Schedl, P., Georgiev, P. and Kyrchanova, O.(2021). Mapping of functional elements of the Fab-6 boundary involved in the regulation of the Abd-B hox gene in Drosophila melanogaster. Sci Rep 11(1): 4156. PubMed ID: 33603202
Summary: The autonomy of segment-specific regulatory domains in the Bithorax complex: is conferred by boundary elements and associated Polycomb response elements (PREs). The Fab-6 boundary is located at the junction of the iab-5 and iab-6 domains. Previous studies mapped it to a nuclease hypersensitive region 1 (HS1), while the iab-6 PRE was mapped to a second hypersensitive region HS2 nearly 3 kb away. To analyze the role of HS1 and HS2 in boundary we generated deletions of HS1 or HS1 + HS2 that have attP site for boundary replacement experiments. The 1389 bp HS1 deletion can be rescued by a 529 bp core Fab-6 sequence that includes two CTCF sites. However, Fab-6 HS1 cannot rescue the HS1 + HS2 deletion or substitute for another BX-C boundary - Fab-7. For this it must be combined with a PRE, either Fab-7 HS3, or Fab-6 HS2. These findings suggest that the boundary function of Fab-6 HS1 must be bolstered by a second element that has PRE activity. |
Li, M., Zhao, Q., Belloli, R., Duffy, C. R. and Cai, H. N. (2021). Insulator foci distance correlates with cellular and nuclear morphology in early Drosophila embryos. Dev Biol 476: 189-199. PubMed ID: 33844976
Summary: The three-dimensional (3D) organization of the genome is highly dynamic, changing during development and varying across different tissues and cell types. Recent studies indicate that these changes alter regulatory interactions, leading to changes in gene expression. Despite its importance, the mechanisms that influence genomic organization remain poorly understood. A network of chromatin boundary elements, or insulators, has ben identified in the Drosophila Antennapedia homeotic complex (ANT-C). These genomic elements interact with one another to tether chromatin loops that could block or promote enhancer-promoter interactions. To understand the function of these insulators, their interactions were assessed by measuring their 3D nuclear distance in developing animal tissues. The data suggest that the ANT-C Hox complex might be in a folded or looped configuration rather than in a random or extended form. The architecture of the ANT-C complex, as read out by the pair-wise distance between insulators, undergoes a strong compression during late embryogenesis, coinciding with the reduction of cell and nuclear diameters due to continued cell divisions in post-cleavage cells. These results suggest that genomic architecture and gene regulation may be influenced by cellular morphology and movement during development. |
Fang, C., Xin, Y., Sun, T., Monteiro, A., Ye, Z., Dai, F., Lu, C. and Tong, X. (2022). The Hox gene Antennapedia is essential for wing development in insects. Development 149(2). PubMed ID: 35088829
Summary: A long-standing view in the field of evo-devo is that insect forewings develop without any Hox gene input. The Hox gene Antennapedia (Antp), despite being expressed in the thoracic segments of insects, has no effect on wing development. This view has been obtained from studies in two main model species: Drosophila and Tribolium. This study shows that partial loss of function of Antp resulted in reduced and malformed adult wings in Bombyx, Drosophila and Tribolium. Antp mediates wing growth in Bombyx by directly regulating the ecdysteriod biosynthesis enzyme gene (shade) in the wing tissue, which leads to local production of the growth hormone 20-hydroxyecdysone. Additional targets of Antp are wing cuticular protein genes CPG24, CPH28 and CPG9, which are essential for wing development. It is proposed, therefore, that insect wing development occurs in an Antp-dependent manner. |
Jimenez-Mejia, G., Montalvo-Mendez, R., Hernandez-Bautista, C., Altamirano-Torres, C., Vazquez, M., Zurita, M. and Resendez-Perez, D. (2022). Trimeric complexes of Antp-TBP with TFIIEbeta or Exd modulate transcriptional activity. Hereditas 159(1): 23. PubMed ID: 35637493
Summary: Hox proteins finely coordinate antero-posterior axis during embryonic development and through their action specific target genes are expressed at the right time and space to determine the embryo body plan. This study reports Antennapedia (Antp) Hox protein-protein interaction with the TATA-binding protein (TBP) and the formation of novel trimeric complexes with TFIIEβ and Extradenticle (Exd), as well as its participation in transcriptional regulation. Using Bimolecular Fluorescence Complementation (BiFC), this study detected the interaction of Antp-TBP and, in combination with Forster Resonance Energy Transfer (BiFC-FRET), the formation of the trimeric complex with TFIIEβ and Exd in living cells. Mutational analysis showed that Antp interacts with TBP through their N-terminal polyglutamine-stretches. The trimeric complexes of Antp-TBP with TFIIEβ and Exd were validated using different Antp mutations to disrupt the trimeric complexes. Interestingly, the trimeric complex Antp-TBP-TFIIEβ significantly increased the transcriptional activity of Antp, whereas Exd diminished its transactivation. These findings provide important insights into the Antp interactome with the direct interaction of Antp with TBP and the two new trimeric complexes with TFIIEβ and Exd. These novel interactions open the possibility to analyze promoter function and gene expression to measure transcription factor binding dynamics at target sites throughout the genome. |
Tendolkar, A., Mazo-Vargas, A., Livraghi, L., Hanly, J. J., Van Horne, K. C., Gilbert, L. E., Martin, A. (2024).. Cis-regulatory modes of Ultrabithorax inactivation in butterfly forewings. Elife, 12 PubMed ID: 38261357
Summary: Hox gene clusters encode transcription factors that drive regional specialization during animal development: for example the Hox factor Ubx is expressed in the insect metathoracic (T3) wing appendages and differentiates them from T2 mesothoracic identities. Hox transcriptional regulation requires silencing activities that prevent spurious activation and regulatory crosstalks in the wrong tissues, but this has seldom been studied in insects other than Drosophila, which shows a derived Hox dislocation into two genomic clusters that disjoined Antennapedia (Antp) and Ultrabithorax (Ubx). This study investigated how Ubx is restricted to the hindwing in butterflies, amidst a contiguous Hox cluster. By analysing Hi-C and ATAC-seq data in the butterfly Junonia coenia, a Topologically Associated Domain (TAD) was shown to maintain a hindwing-enriched profile of chromatin opening around Ubx. This TAD is bordered by a Boundary Element (BE) that separates it from a region of joined wing activity around the Antp locus. CRISPR mutational perturbation of this BE releases ectopic Ubx expression in forewings, inducing homeotic clones with hindwing identities. Further mutational interrogation of two non-coding RNA encoding regions and one putative cis-regulatory module within the Ubx TAD cause rare homeotic transformations in both directions, indicating the presence of both activating and repressing chromatin features. A series of spontaneous forewing homeotic phenotypes obtained in Heliconius butterflies is described, their possible mutational basis is discussed. By leveraging the extensive wing specialization found in butterflies, this initial exploration of Ubx regulation demonstrates the existence of silencing and insulating sequences that prevent its spurious expression in forewings. |
Montalvo-Mendez, R. J., Cardenas-Tueme, M., Resendez-Perez, D. (2024). Drosophila in the study of hTBP protein interactions in the development and modeling of SCA17. Gaceta medica de Mexico, 160(1):1-8 PubMed ID: 38753562
Summary: Protein interactions participate in many molecular mechanisms involved in cellular processes. The human TATA box binding protein (hTBP: see drosophila TBP) interacts with Antennapedia (Antp) through its N-terminal region, specifically via its glutamine homopeptides. This PolyQ region acts as a binding site for other transcription factors under normal conditions, but when it expands, it generates spinocerebellar ataxia 17 (SCA17), whose protein aggregates in the brain prevent its correct functioning. To determine whether the hTBP glutamine-rich region is involved in its interaction with homeoproteins and the role it plays in the formation of protein aggregates in SCA17. This study characterized hTBP interaction with other homeoproteins. hTBP interacted with homeoproteins through its glutamine-rich region, and hTBP protein aggregates with expanded glutamines were found to affect the locomotor capacity of flies. The study of hTBP interactions opens the possibility for the search for new therapeutic strategies in neurodegenerative pathologies such as SCA17. |
Villarreal-Puente, A., Altamirano-Torres, C., Jimenez-Mejia, G., Hernandez-Bautista, C., Montalvo-Mendez, R., Vazquez, M., Zurita, M., Resendez-Perez, D. (2024). Novel Antennapedia and Ultrabithorax trimeric complexes with TBP and Exd regulate transcription. Hereditas, 161(1):25 PubMed ID: 39080786
Summary: Hox proteins interact with DNA and many other proteins, co-factors, transcriptional factors, chromatin remodeling components, non-coding RNAs and even the extracellular matrix that assembles the Hox complexes. The number of interacting partners continues to grow with diverse components and more transcriptional factors than initially thought. Hox complexes present many activities, but their molecular mechanisms to modulate their target genes remain unsolved. This study showed the protein-protein interaction of Antp with Ubx through the homeodomain using BiFC in Drosophila. Analysis of Antp-deletional mutants showed that AntpHD helixes 1 and 2 are required for the interaction with Ubx. Also, a novel interaction of Ubx with TBP was found, in which the PolyQ domain of TBP is required for the interaction. Moreover, the formation of two new trimeric complexes of Antp with Ubx, TBP and Exd was found using BiFC-FRET; these proteins, however, do not form a trimeric interaction with BIP2 or TFIIEβ. The novel trimeric complexes reduced Antp transcriptional activity, indicating that they could confer specificity for repression. These results increase the number of transcriptional factors in the Antp and Ubx interactomes that form two novel trimeric complexes with TBP and Exd. A new Ubx interaction with TBP was also found. These novel interactions provide important clues of the dynamics of Hox-interacting complexes involved in transcriptional regulation, contributing to better understand Hox function. |
Antp is the last gene of the Antennapedia Complex (ANTP-C), that is, the most distal from the centromere, and the one expressed in the most posterior locale. Antp is distal to fushi tarazu and even more distant from Sex combs reduced The highest levels of transcription are found in the ventral portion of the second thoracic segment, where Antp is the initiator of a cascade of events that result in the development of an adult leg.
This begs an interesting question: why does a gene that regulates thoracic development get the name Antennapedia? Many of the earliest discovered mutations in Drosophila were dominant. This type of mutation is readily produced and easy to spot, since only one copy of the mutated gene is required in order to see a phenotypic effect. As early as 1949 dominant mutations had been found which converted antenna into the second leg [Image] (the mutant adult having no antenna, and two sets of second legs). The name Antennapedia (antenna-foot) was appropriately descriptive. Such homeotic transformations are exciting to contemplate, and serve as the basis of understanding of gene function in Drosophila (Abbott, 1986).
It is important to understand that adult structures are derived from cells set aside during embryonic development. For example, imaginal discs store the potential for leg development throughout the larval stages. Discs begin to develop in the third instar larval stage, and the metamorphosis to adult takes place during the pupal stage. Therefore, a change in the regulation of the Antennapedia gene, resulting in its expression in the eye-antennal disc, is sufficient to cause an eye to leg transformation that will show up in the adult.
Later discovery of recessive alleles (null alleles) of Antennapedia were found to have an effect just opposite to the one first noted in 1949. The second or mesothoracic leg was transformed into an antenna. Antennapedia was thought to have a positive effect, that is, the promotion of leg fate. Another view posited that Antennapedia acted to repress genes whose expression in the thorax would result in head fate. Similarly, expression of Antp in the head could act to repress head fate, establishing ectopic leg in place of antenna. What these varying results have in common is the transformation of one organ into another, based on the existence of a unified cellular precursor for the whole organ, the imaginal disc. Careful observation reveals that such transformations are rarely complete. They depend on the period during development in which the ectopic expression is carried out and the level of gene expression (Scanga, 1995). Nevertheless, Antennapedia and other homeotic genes can modify the expression of the whole genetic hierarchy of events required for organogenesis.
Because of their widespread effect on cell fate, there is tremendous interest in identifying the downstream targets of homeotic genes. The search has been complicated because of the overlapping actions of the linked array of five segmentally acting genes of the Antennapedia complex. It isn't always clear which gene in the complex is having the noted effect.
One such downstream target for Antennapedia is the gene spalt. spalt is expressed in both anterior and posterior regions of the Drosophila embryo where it promotes head and tail patterns. sal has an added feature: it represses teashirt function. teashirt is known to promote trunk (thoracic) development. Ectopic expression of Antp in the head is likely to repress sal, and thus result in the expression of teashirt (otherwise repressed by sal), and promote trunk identity (Künlein, 1994).
Do Antp null alleles allow for the expression of sal in the leg disc? And if so, is this sufficient to promote a leg-into-antenna transformation? In the case of the spalt,gene, the effects of Antp are known to be recessive. It is not yet known exactly how homeotic genes transform the fate of a complete organ. One clue lies in the lack of specificity of the transformation. Recently it has been shown that ectopic expression of Ultrabithorax, abdominal-A and Abdominal-B cause similar transformations in some of the fruitfly appendages: antennal tissue into leg tissue and wing tissue into haltere tissue. Thus the homeotic requirement to form appendages is, in some cases, non-specific (Casares, 1996).
Antennapedia represses homothorax in leg discs. During the evolution of insects from a millipede-like ancestor, the Hox genes are thought to have promoted the diversification of originally identical body structures. In Drosophila, antennae and legs are homologous structures that differ from each other as a result of the Hox gene Antennapedia (Antp), which promotes leg identities by repressing unknown antennal-determining genes. Four lines of evidence are presented that identify extradenticle (exd) and homothorax (hth) as antennal-determining genes. (1) Removing the function of eitherexd or hth (which is required for the nuclear localization of Exd protein), transforms the antenna into leg; such transformations occur without activation of Antp. (2) In most antennal cells, hth is expressed and Exd is nuclear, whereas both are restricted to proximal cells of the leg. (3) Antp is a repressor of hth. (4) Ectopic expression of Meis1, a murine hth homolog, can trigger antennal development elsewhere in the fly. Taken together, these data indicate that hth is an antennal selector gene, and that Antp promotes leg development by repressing hth, consequently preventing the nuclear transport of Exd (Casares, 1998).
Now that it is clear that hth determines antennal fate, it is worthwhile reconsidering the transformation to leg that is produced by hth or exd mutant cells in the antenna. This is the same phenotype seen with dominant Antp mutants, but the leg develops without the activity of Antp, Scr or Ubx. It follows that a leg can be generated without Hox activity, suggesting that the leg pathway is the ground state for ventral appendages. Thus the ground pattern for both larvae and adults is thoracic. Nor does Antp "select" for a specific leg pathway -- it simply represses hth in the leg primordia, thereby blocking antennal development and allowing the development of legs by default. This supports the idea that Antp promotes a ground (mesothoracic) pattern by repressing cephalic genes. This basal pattern is modified by Scr toward prothoracic (first leg) or by Ubx toward metathoracic (third leg) in their respective primordia. The downregulation of hth by Antp explains the phenotype of the dominant Antp mutants is due to homothorax repression. It also explains the ability of other Hox genes such as Ubx, abdominal-A, and Abdominal-B to induce the transformation of antennae into legs. These genes prevent the nuclear translocation of Exd (most likely through hth repression), so the antennal to leg transformations are probable nonspecific and caused by a property that is common to Antp and other Hox proteins (Morata, 1998 and Casares, 1998).
During embryogenesis, in contrast with leg development, Antp selects for a specific developmental pathway. Loss-of-function mutations and experiments to induce ectopic expression show that Antp determines the larval mesothoracic pattern -- a function that is clearly distinct from the other Hox genes. Why legs should be different is not clear, but different Hox genes have similar effects on appendages, possibly because these appendages have no hth activity, without which the Hox genes lack specificity (Morata, 1998 and references).
Hox genes encoding homeodomain transcriptional regulators are known to specify the body plan of multicellular organisms and are able to induce body plan transformations when misexpressed. These findings led to the hypothesis that duplication events and misexpression of Hox genes during evolution have been necessary for generating the observed morphological diversity found in metazoans. It is known that overexpressing Antennapedia in the head induces antenna-to-leg as well as head-to-thorax transformation and eye reduction. At present, little is known about the exact molecular mechanism causing these phenotypes. The aim of this study was to understand the basis of inhibition of eye development. It has been demonstrated that Antp represses the activity of the eye regulatory cascade. By ectopic expression, it has been shown that Antp antagonizes the activity of the eye selector gene eyeless. Using both in vitro and in vivo experiments, it has been demonstrated that this inhibitory mechanism involves direct protein-protein interactions between the DNA-binding domains of Ey and Antp, resulting in mutual inhibition (Plaza, 2001).
If the Antp protein is able to block Ey activity, this mechanism should also function in other tissues. Therefore, ectopic eye formation should also be blocked by Antp. To test this prediction, ectopic eyes were induced on wing, antennae and legs using the UAS-GAL4 system. Results show that the ectopic eye formation induced by ey is completely blocked on co-expressing ey and Antp. Moreover, the Antp induced antenna-to-leg transformation is inhibited by ey. A series of similar tests employing hs-ey and hs-Antp transgenes, singly or in combination, leads to the same conclusions. Furthermore, these tests reveal a specific requirement for the Antp homeodomain (HD), since N-terminal deletions of the Antp protein do not affect its ability to inhibit Ey activity, whereas deletion of the HD results in a protein unable to inhibit ey function. Similarly, using the UAS-GAL4 system, the Antp HD-deleted protein is unable to repress ectopic eye formation. These results made it necessary to demonstrate that both proteins co-localize in the same cells of the discs. Upon examining protein accumulation by confocal microscopy, it was found that both proteins are efficiently co-expressed in these different tissues. Furthermore, immunostaining experiments performed using the ey antibody or the neuronal marker 22C10 confirm that, despite the presence of Ey in the disc, co-expression of Antp leads to inhibition of neuronal differentiation (Plaza, 2001).
In order to test whether the DNA-binding activity of Antp is not required for the inhibition of eye development, an Antp mutant was tested in which the DNA-binding specificity was changed (Q50K). Interestingly, this mutated protein is still able to repress eye development. In addition, mutagenesis experiments were performed to convert Q50 and N51, residues shown to be crucial for DNA contacts, into alanines. This mutant is unable to bind a DNA PS2 probe containing a Hox/Exd/Hth motif, even in the presence of EXD and HTH in the bandshift assay. This A50,A51 mutant protein is still capable of inhibiting eye development when expressed in the eye disc using a strong EyE-GAL4 line, although with a lower activity than the wild-type Antp protein (Plaza, 2001).
Based on these in vivo results it was asked whether Antp and Ey might interact directly and thereby inhibit each others activities. Potential in vitro interactions between Antp and Ey were examined using glutathione S-transferase (GST)-Antp fusion proteins immobilized on glutathione-Sepharose beads. These immobilized proteins were tested for their ability to retain in vitro synthesized 35S-labeled Ey protein. Different portions of the Antp protein were produced and tested separately for their ability to interact with Ey. Only the C-terminal portion of Antp including the HD is able to interact with Ey (Plaza, 2001).
To define the regions within Ey and Antp that are required for the interaction of the two proteins, a set of deletion mutants of each protein was tested for the ability to interact in vitro. Structure-function studies of both proteins have delineated specific domains that contribute to their functions as transcription factors as well as their interactions with other proteins. The Antp HD that mediates DNA binding has also been shown to interact with other HD proteins such as Exd. The Ey protein contains two DNA-binding domains, a paired domain and an HD. The paired domain has been shown to interact with different transcription factors. These findings led to an investigation of whether Ey paired domains and HDs are involved in the interaction with Antp. Deleting either of these domains in the Ey protein results in a partial loss of the interaction with the Antp HD. Furthermore, either the Ey paired domain or the Ey HD alone is still able to interact with Antp. These experiments suggest that since each domain is able to interact with Antp, both domains might cooperate for efficient binding of Ey to Antp. Moreover, deletion of the C-terminal part of the Antp protein results in the loss of binding to both the paired domain and the HD of Ey, confirming that the Antp HD is essential for the interaction with Ey. Since the complexes were formed using Ey paired domain and Antp HD purified from bacteria, the two proteins appear to interact directly through their respective DNA-binding region. It was hypothesized that the DNA interface might be important to stabilize the interaction. Indeed, the interaction between the two proteins requires that one of them binds to DNA (Plaza, 2001).
The question of whether the ability of Antp to repress eye development and to interact with Ey can be extended to other homeotic genes. For this purpose, expression of Scr, Ubx, abdA and AbdB were targeted into the eye disc using dppblink-GAL4. Expression of these different genes also results in inhibition of eye development by inducing apoptosis. Interestingly, these different proteins are also able to interact with Ey in vitro. Deletion of the ABD-A HD region abrogates binding to Ey, suggesting that also for this protein, the HD is required for interaction with Ey (Plaza, 2001).
The transformation of antenna to leg is a classical model for understanding segmental fate decisions in Drosophila. The spineless (ss) gene encodes a bHLH-PAS transcription factor that plays a key role in specifying the identity of distal antennal segments. This report identifies the antennal disc enhancer of ss and then uses enhancer-lacZ reporters to work out how ss antennal expression is regulated. The antennal determinants Distal-less (Dll) and homothorax (hth) are key activators of the antennal enhancer. Dll is required continuously and, when present at elevated levels, can activate the enhancer in regions devoid of hth expression. In contrast, homothorax (hth) is required only transiently both for activation of the enhancer and for specification of the aristal portion of the antenna. The antennal enhancer is repressed by cut, which determines its proximal limit of expression, and by ectopic Antennapedia (Antp). Repression by Antp is not mediated by hth, suggesting that ss may be a direct target of Antp. ss+ is not a purely passive target of its regulators: ss+ partially represses hth in the third antennal segment and lies upstream of Dll in the development of the maxillary palp primordia (Emmons, 2007; full text of article).
This study used lacZ reporters to identify the enhancers responsible for most aspects of ss expression during embryonic and imaginal development. Antennal expression is driven by two large fragments from the ss 5' region, B6.9 and EX8.2. Both of these fragments drive expression in the antennal segment of the embryo and in the distal portion of the pupal antenna. B6.9 is also expressed in the antennal disc through most or all larval development. Dissection of B6.9 allowed localization of the larval antennal enhancer to a fragment of 522 bp. The B6.9 and 522 reporters were used as a proxy for ss expression in experiments to determine the effects of potential upstream regulators of ss. This strategy has its strengths and weaknesses, but has been made necessary by an inability to generate antisera against Ss. A major strength of the approach is that it was possible to assess the effects of regulators on individual enhancers. It is likely that monitoring endogenous ss expression would give results that are less clear cut since both the antennal and tarsal enhancers of ss are active within the antenna. A potential weakness is that the reporters may not faithfully reproduce the normal expression of ss. However, as far as is possible to tell, the antennal reporters reproduce ss expression very well. The expression of B6.9 and EX8.2 in the embryonic antennal segment and the pupal antenna corresponds very closely to that of endogenous ss. Expression of B6.9 and 522 in the larval antennal disc appears very similar or identical to that of ss+, and the transient requirement for hth+ in the activation of these reporters corresponds well to the transient requirement for hth+ in aristal specification. The tarsal enhancer P732 likely also reproduces the spatial pattern of ss+ expression as its tarsal expression domain corresponds well to the region deleted in ss− mutants (Emmons, 2007).
The results of this dissection of the B6.9 fragment were surprising. Removal of the left-hand 2 kb of B6.9 to produce S4.9 resulted in the loss of antennal specificity; S4.9 reporters are expressed in both antennal and leg discs. The E2.0 subfragment of S4.9 shows a similar expression pattern, and expression of this fragment in both leg and antennal discs is independent of Hth, but requires Dll continuously. On further subdivision of the E2.0 fragment, it was found that antennal and leg expression are separable; the 522 fragment is largely specific for the antenna, whereas the 531 fragment drives expression primarily in leg discs. To summarize, antennal specificity is present in B6.9, lost in S4.9 and E2.0 and regained in 522. How can sense be made of this? The region deleted from B6.9 to produce S4.9 clearly plays an important role in enforcing antennal specificity. Since this region contains a PRE, one might suspect that it functions in larval stages to maintain repression of the enhancer outside of the antennal segment. However, that the E2.0 fragment has lost the requirement for Hth in both the antenna and leg (S4.9 has not been tested) suggests that the PRE-containing region might function in both locations. One possibility is that this region represses the enhancer in both antennal and leg discs. In the antenna, this repression can be overcome by the combined action of Hth and Dll, while in the leg Dll alone is not sufficient for activation. When the PRE-containing region is deleted, repression is absent or reduced, so that Dll can activate the enhancer without assistance from Hth, and expression is seen in both antennal and leg discs. Why then is antennal specificity restored in the 522 subfragment? Perhaps this fragment is lacking a subset of Dll interaction sites so that it can no longer be activated by Dll alone, but requires combined activation by Hth and Dll. Although this model is consistent with many of the results, it does not provide a ready explanation for the leg specificity of the 531 fragment (Emmons, 2007).
In addition to activation by combined Hth and Dll, the ss antennal disc enhancer is repressed by Cut and by ectopic Antp. Each of these regulators will be discussed separately. It was found that hth+ is required only transiently for activation of the B6.9 reporter. hth− clones induced in the embryo or first instar lose expression of B6.9 autonomously in both A3 and the aristal primordia. However, some time in the second of early third instar. Regulatory instar expression of B6.9 becomes independent of hth. Consistent with this transient requirement, it is shown that hth+ is required only early in larval development for specification of the arista. hth− clones induced in the first and second instars show a transformation of the entire antenna to a leg-like appendage. However, clones induced after this time show normal aristal development. These temporal requirements are reflected in the expression pattern of hth: hth is expressed throughout the antennal primordium early in development, but in the second or early third instar is repressed in the central domain, which will produce the arista (Emmons, 2007).
The stable activation of B6.9 by Hth suggests that this fragment contains a 'cellular memory module'. The presence of a PRE within B6.9 is consistent with this idea. The ss locus binds Polycomb protein in salivary gland chromosomes and was recently shown to contain PREs by chromatin immunoprecipitation. In the latter work, ss PREs were localized to within the E1.6 subfragment of B6.9 as well as the EX8.2 fragment, both of which showed pairing dependent suppression in this work. PREs are generally thought of as functioning to stably repress genes. However, PREs can also be associated with activating elements to form memory modules that mediate stable activation. It seems likely that B6.9 contains such a module that responds to Hth. Like a memory module from the hedgehog gene, activity of the ss module is set sometime around the second instar. Surprisingly, it was found that activation of the 522 reporter by Hth can also be persistent, although not as stable as for B6.9. The 522 fragment does not appear to contain a PRE, suggesting that Hth may directly recruit factors to the 522 element that cause semi-stable transcriptional activation (Emmons, 2007).
ss is not a completely passive target of hth; ss partially represses hth in antennal discs, which causes hth to be expressed at a lower level in A3 than in A2. This repression appears to be important for normal development as ectopic expression of Hth can delete A3. Moreover, clones ectopically expressing Hth are largely blocked from entering A3 from the proximal (A2) side, suggesting that the different levels of Hth present in A2 and A3 cause a difference in cell affinities between these segments. Hth-expressing clones are similarly restricted to the two most proximal segments in leg discs, although here there is no endogenous expression of hth more distally (Emmons, 2007).
In contrast to hth, Dll is required continuously for expression of both B6.9 and 522 as Dll− clones induced even very late in development lose expression of these reporters. This continuous requirement for Dll indicates that stable activation of the B6.9 memory module by Hth does not by itself commit the reporter to expression; rather, activation by Hth appears to render B6.9 open to interaction with Dll and perhaps other positive factors (Emmons, 2007).
Three lines of evidence suggest that Dll is the primary activator of the ss antennal enhancer. (1) It was found that expression of B6.9 and 522 is sensitive to the dosage of Dll+. Expression of both reporters is reduced in animals carrying only one dose of Dll+, and for 522, expression is enhanced in clones having extra doses of Dll+. This dose sensitivity suggests that ss is a direct target of Dll. (2) It was found that expression of both reporters is often induced within clones expressing ectopic Dll, even in the apparent absence of Hth expression. Such activation is seen in clones in the distal leg, wing and elsewhere. (3) It was found that the embryonic antennal enhancer carried by B6.9 is absolutely dependent upon Dll+, but independent of hth. Taken together, these observations suggest that Dll is a primary activator of the ss antennal enhancers. Hth may provide antennal specificity by boosting the level of activation by Dll in the antennal disc (Emmons, 2007).
Surprisingly, it was found that the regulatory relationship between ss and Dll is reversed in the maxillary palp. Here, ss is expressed prior to Dll and is required for the normal initiation of Dll expression. Although some Dll expression ultimately takes place in the palp primordium in ss− animals, this expression is weak and occurs in only a few cells. It has not been worked out how ss is activated in the palp. However, it seems likely that dpp plays a role as the 531 subfragment of B6.9 drives expression in a stripe in the region of the palp that roughly coincides with a stripe of dpp expression. The positioning of ss upstream of Dll in the palp may explain why the region ventral to the antenna is so sensitive to ectopic expression of Ss. Strong activation of Dll here by ectopic Ss combined with endogenous expression of hth might be expected to cause frequent induction of ectopic antennae, as is observed. Since ss is normally expressed in the palp, why should earlier ectopic Ss cause the palp primordium to develop as antenna? It seems likely that timing is key, but level of Ss expression could also be important (Emmons, 2007).
The reciprocal regulatory roles of ss and Dll in the antenna and palp suggest a particularly close relationship between these genes. This relationship is reinforced by the finding that ss is required for the development of bracts in the femur, as is Dll (Emmons, 2007).
The finding that Dll and Hth are both activators of the ss antennal reporters is consistent with the proposal that antennal identity is defined by the combined activity of these regulators. However, the results indicate that this model is an oversimplification. Examination of clones expressing Dll, Hth, or both proteins together revealed little correlation between activation of the B6.9 and 522 antennal reporters and combined expression of Dll and Hth. Strikingly, Dll-expressing clones often activate the reporters ectopically without any apparent concomitant expression of Hth, and clones expressing both proteins usually do not activate the reporters. These experiments also reveal strong context dependence. Examples include the leg, where Dll-expressing clones can activate the reporters distally, but not proximally (where endogenous hth expression occurs) and the wing disc, where clones expressing Dll or both Dll and Hth activate the reporters in the wing pouch, but not at all in the notum. The level of expression of both proteins also appears to be key as high levels of Dll can activate the reporters in the leg in the absence of Hth and elevated levels of Hth can repress expression in the normal antennal domain. Previous results have shown that antennal structures can be induced by ectopic expression of Dll in the wing hinge region or proximal leg (which express hth endogenously) or by combined expression of Dll and Hth elsewhere. While this is true, the results indicate highly variable effects in such ectopic expression experiments and fail to detect the strongly synergistic activation of antennal identity by combined Hth and Dll implied by the model. The results indicate that Dll is the primary activator of the ss antennal reporters, that Hth serves to promote this activity and that activation by Dll and Hth is highly context-dependent (Emmons, 2007).
Consistent with direct control of the antennal reporters by Dll and Hth, two highly conserved regions within the 522 fragment contain apparent binding sites for Dll, Hth, and the Hth dimerization partner Extradenticle. The functional importance of these binding sites is currently being tested (Emmons, 2007).
This study has show that the proximal boundary of B6.9 and 522 expression is defined by repression by cut. This repression likely explains why ectopic Cut causes a transformation of arista to tarsus. cut has been shown to define the proximal expression limit of distal antenna (dan) and distal antenna related (danr); since ss lies upstream of these genes, it seems very likely that their regulation by cut is indirect. The mechanism of action of Cut is not well understood, since only one direct target has been characterized in Drosophila (Emmons, 2007).
Ectopic expression of Antp in the antenna represses the B6.9 and 522 reporters. This finding was expected, since it is well known that expression of Antp or other Hox genes in the antenna causes a transformation to leg. The conventional view is that this transformation results from the repression of hth by ectopic Hox proteins. Repression of hth early in development would be expected to lead secondarily to loss of ss expression and loss of distal antennal identity. However, it was found that clones expressing Antp repress the B6.9 and 522 reporters even when these clones are induced very late in development, long after the requirement for activation by hth has passed. Late repression of the antennal reporters by Antp must therefore occur independently of hth and could be direct. One possibility, currently being tested, is that Antp might compete with Dll for binding to the 522 enhancer. Late repression of the ss antennal enhancer by Antp is consistent with the effects of Antp-expressing clones on antennal identity: such clones induced in the mid to late third instar cause transformations of distal antenna to leg (Emmons, 2007).
Clones induced late that ectopically express Antp in a sustained fashion were examined. In contrast, previous work studied the effects of pulses of Antp expression induced by one-hour heat shocks in a heat shock/Antp line. It had been found that transformations of arista to tarsus were induced by such pulses only when they were administered at the end of the second instar. Why do pulses of Antp at this time cause a stable, heritable transformation of the distal antenna? The current results suggest an explanation. The period sensitive to Antp pulses coincides roughly with when the ss antennal enhancer becomes independent of hth. This correlation suggests that pulses of Antp in the second instar cause heritable transformations by interfering with the stable activation of ss by Hth. Recently, it has been reported that ectopic Antp does not repress hth in the antenna early in larval development. This observation suggests that Antp might act directly on the ss antennal enhancer to prevent its stable activation by Hth (Emmons, 2007).
The regulation of ss by ectopic Antp suggests that Antp may normally play a significant role in repressing ss antennal enhancer activity in the legs. Although this idea has not been tested directly, it seems unlikely that Antp is primarily responsible for keeping the ss antennal enhancers inactive in the leg. Antp null clones do cause activation of the ss target gene dan in leg discs, implying ectopic activation of ss. However, this activation occurs only proximally, with the distal leg appearing to develop independently of Antp. Expression of Antp in the proximal leg may account for why Dll-expressing clones fail to activate B6.9 or 522 in this location. Ectopic activation of the ss antennal enhancers in the leg primordia of the embryo is not seen in an Antp null mutant (Emmons, 2007).
These studies suggest that antennal structures are specified in a combinatorial fashion by Hth, Dll, Ss and probably other factors. In A3, all three proteins are required for normal antennal identity. In ss− antennae, hth continues to be expressed in A3 (although at elevated levels), as does Dll. Despite this continued expression of hth and Dll, A3 develops without antennal characteristics and produces only naked cuticle. Thus, Hth and Dll are unable to specify A3 characters in the absence of Ss. Conversely, assuming that ss is stably activated in the antenna by Hth, as is B6.9, then hth− clones induced late would show persistent expression of both ss and Dll in A3. Such clones are transformed to leg, implying that Ss and Dll have no ability to direct A3 identity in the absence of Hth. Taken together, these observations suggest that Hth, Dll and Ss must act together to specify A3 identity. This requirement for combined action accounts for why ectopic expression of Ss does not induce A3 tissue in the medial leg, since hth is not normally expressed here. The view of combinatorial control suggests that many A3-specific target enhancers might be identifiable in genome searches as regions that contain clustered binding sites for Hth, Dll and Ss; tests of this prediction will be presented elsewhere (Emmons, 2007).
In contrast to A3, the aristal primordium appears to be specified by ss and Dll acting together in the absence of hth expression. hth is expressed in the aristal region early in development, where it functions to establish ss expression, but it is soon repressed here. Therefore, for most of development, the arista is specified by Ss and Dll acting without input from Hth. Consistent with this picture, the arista adopts leg identity in ss null mutants, and ectopic expression of ss causes the distal tip of the leg to develop as arista (Emmons, 2007).
In ss− mutants, the distal antenna is terminated by a single tarsal segment (the fifth). In contrast, in ss mutants that lack only antennal enhancer activity (e.g. the breakpoint mutations ssD114.3 and ssD114.7, the distal antenna develops with a near complete set of tarsal segments. This difference likely reflects the activity of the tarsal enhancer in the antenna. In support of this view, the ss tarsal enhancer drives expression in the segmented base of the arista, a region known as the basal cylinder. This region transforms to tarsal segments 2-4 in Antp-induced transformations of antenna to leg. However, the question arises as to why normal antennal expression of ss causes the proximal arista to develop as basal cylinder, whereas ss expression driven by the tarsal enhancer alone causes this same region to develop as tarsal segments. Likely, the key difference is that expression driven by the tarsal enhancer is transient, whereas expression driven by the antennal enhancer is sustained. Perhaps transient expression of ss allows growth and subsegmentation to produce a full set of tarsal segments, whereas sustained expression inhibits growth, producing the basal cylinder. Consistent with this idea, sustained expression of ss driven by the GAL4 method can cause deletion of tarsi in the legs. The levels of expression driven by the tarsal and antennal enhancers may also be important as flies having only one dose of ss show a partial transformation of the basal cylinder to tarsus. The ss tarsal enhancer drives weak expression in A3 as well as in the basal cylinder, likely accounting for the presence of some specialization of A3 in ss mutants lacking the antennal enhancers (Emmons, 2007).
The view that antennal identity is specified by the combined action of Hth, Dll and Ss contradicts the now prevalent view that antennal identity is determined solely by hth. The major evidence supporting the latter view is that early hth− clones transform the entire antenna to leg, and ectopic expression of Hth can induce ectopic antennal structures in the anal plates. Moreover, Dll shows little antennal specificity, being expressed in the distal portions of all of the ventral appendages, and ss expression in the antenna is dependent upon hth+. Should hth be viewed as the antennal 'selector' gene? hth does not seem to be a selector in the same sense as the Hox genes; it is expressed very broadly in the embryo and in other imaginal discs and plays no role in activating ss in the antennal segment of the embryo. Moreover, the ability of ectopic Hth to induce antennal structures is very limited: transformations of anal plate to distal antenna have been reported following ectopic expression of Hth or Meis1, a mammalian homolog. However, others have been unable to reproduce this effect by ectopic expression of Hth, matching the results of this study. That anal plates are susceptible to transformation at all is likely due to the fact that Dll and ss are coexpressed here in normal development. A further dissimilarity is that hth acts only as an establishment regulator of ss in the antennal disc, unlike the continuous requirements usually seen for the Hox genes. Ultimately, assessment of the importance of hth will depend on whether its function in the antenna is conserved. The expression pattern of hth in the antenna does appear to be conserved in the milkweed bug Oncopeltus. However, localization of nuclear Exd (a proxy for Hth expression) indicates that Hth is not differentially expressed in the antenna and leg of the cricket. Expression of hth in the crustacean Porcellio also appears to be identical in the second antenna and the legs. Characterization of hth, Dll and ss expression and function in additional arthropods will be required to assess properly the importance of these genes in antennal specification (Emmons, 2007).
Homeodomains (HDs) increase their DNA-binding specificity by interacting with additional cofactors outlining a Hox interactome with a multiplicity of protein-protein interactions. In Drosophila, the first link of functional contact with a general transcription factor (GTF) was found between Antennapedia (Antp) and BIP2 (TFIID complex). Hox proteins also interact with other components of Pol II machinery such as the subunit Med19 from Mediator (MED) complex, TFIIEbeta and transcription-pausing factor M1BP. This paper focused on the Antp-TFIIEbeta protein-protein interface to establish the specific contacts as well as its functional role. TFIIEbeta was found to interact with Antp through the HD independently of the YPWM motif and the direct physical interaction is at helix 2, specifically amino acidic positions I32 and H36 of Antp. These two positions in helix 2 are crucial for Antp homeotic function in head involution, and thoracic and antenna-to tarsus transformations. Interestingly, overexpression of Antp and TFIIEbeta in the antennal disc showed that this interaction is required for the antenna-to-tarsus transformation. These results open the possibility to more broadly analyze Antp-TFIIEbeta interaction on the transcriptional control for the activation and/or repression of target genes in the Hox interactome during Drosophila development (Altamirano-Torres, 2018).
To analyze the interplay between Hox and the general transcription machinery, this study focused on Antp-TFIIEβ protein-protein interface to establish the specific contacts, as well as the functional role of this interaction. The results showed a direct physical interaction of TFIIEβ with the 32 and 36 positions of helix 2 Antp HD in cell culture and in vivo. These two positions on helix 2 HD are required for interaction with TFIIEβ, and this interaction is necessary for homeotic transformation (Altamirano-Torres, 2018).
The results demonstrate that Antp HD was necessary for maintaining the interaction with TFIIEβ. Previous studies have confirmed that the HD is sufficient for interaction with GTFs. For example, it has been found that the AbdA HD was sufficient for TFIIEβ interaction and that when the DNA-binding of the HD is mutated, the interaction is diminished but not abolished. Another example used Bimolecular fluorescence complementation (BiFC) in vivo to demonstrate that the Ubx HD and AbdA HD are sufficient for direct interaction with Med19. In addition of the conserved HD affinities to DNA and RNA, several protein-protein interactions also relied on the HD, such as dimerization of Scr, and Antp interaction with Eyeless (Altamirano-Torres, 2018).
Although this study found that Antp-TFIIEβ interaction is YPWM-independent in BiFC cell culture and the presence of an intact YPWM motif in the helix 2 Antp mutant showed neither interaction by BiFC nor functional activity, co-expression of the YPWM mutant and TFIIEβ reduced the signal interaction in embryos. A similar result in embryos was found in a earlier study where YPWM Antp mutant showed a reduction but not an abolition of TFIIEβ interaction on Drosophila embryos, that could be attributable to the presence of helix 2 in the mutant. Altogether, this suggest that interactions of Antp with TFIIEβ could change from one tissue to another with complex formation in different tissues using various interfaces (YPWM and/or HD), contributing to the plasticity of Hox interaction properties (Altamirano-Torres, 2018).
Deletional analysis of Antp HD suggested interaction of TFIIEβ through the helix 2 of Antp HD. Based on the reported 3D-structure of Antp HD DNA complex, in which helix 2 is on the opposite side of the HD-DNA binding, this study selected the conserved residues 32 and 36, which are exposed and physically available, as candidates for TFIIEβ interaction. To perform a molecular dissection on the Antp-TFIIEβ interaction, the residues I32 and H36 of helix 2, either individually or together, were studied by site-directed mutagenesis in cell culture. BiFC results show a drastic reduction of the interaction by mutation of these two residues, indicating that they are directly involved on Antp-TFIIEβ interaction. It has been demonstrated that AntpHD is internalized to the nuclei, through the residues 43-58 of the third helix. Therefore, since the mutations examined in this study are present on helix 2, the Antp NLS were not affected. To confirm that, immunostaining of Antp helix 2 mutants on cells and embryos showing very clearly the nuclear localization of Antp helix 2 single mutants and double mutant Antp. These results indicated that Antp helix 2 mutants include NLSs for their localization into the nucleus. Moreover, it was also demonstrated that helix 2 mutant keeps its transactivation activity and is capable to interact with EXD in cells and embryos confirming that mutation of these amino acids did not alter DNA binding affinity and the protein conformation to perform essential activities required for in vivo transformation (Altamirano-Torres, 2018).
Since both substitutions by alanines or structurally similar residues affected Antp-TFIIEβ interaction in cell culture in the same manner, I32A-H36A HD mutant was selected for the in vivo analysis in Drosophila. In concordance with BiFC cell culture assay, the results showed no interaction in embryos or in imaginal discs with Antp mutant I32A-H36A. Therefore, residues 32 and 36 of Antp helix 2 are crucial for the interaction with TFIIEβ in BiFC assays in Drosophila embryos and imaginal discs. This is relevant because residues 32 and 36 on Antp helix 2 are identical and highly conserved within Drosophila Hox proteins and can be extrapolated for the interaction with TFIIEβ to another homeoproteins due to the high Hox conservation (Altamirano-Torres, 2018).
Although the results very clearly show Antp-TFIIEβ interaction through positions 32 and 36 of helix 2, this does not exclude the possibility of another amino acid positions, either at helix 2 or the intervening loop, that could be involved to a minor extent on the interaction. For example, position 30 and 33, in addition to the helix 2 amino acids 32 and 36, have also been reported in human POU proteins Oct-1 and Oct-2 interaction with VP16 transactivator factor of Herpes Simplex Virus (Altamirano-Torres, 2018).
Because the precise molecular mechanisms of Antp in transcriptional regulation remains unclear, attempts were made to shed light on these by determining whether I32 and H36 are important for Antp function. When Antp is ectopically expressed on embryos it causes inhibition of head-involution and transformation of prothoracic segment T1 into T2 and antennae into mesothoracic (T2) legs. Antp ectopic expression exhibits that residues 32 and 36 of HD helix 2 are essential for its function in embryo head involution and homeotic transformations of thorax and antenna. Lack of homeotic transformations of AntpI32A-H36A double mutant expression indicates that residues 32 and 36 of HD helix 2 are absolutely required for the Antp ectopic homeotic function in Drosophila. Likewise, Antp mutated in the YPWM motif is not capable of transforming the antenna, and a single exposed residue on helix 1 of Scr HD is necessary for its homeotic function, showing that beside the HD DNA-binding, exposed positions on the HD are crucial for Hox functional activity (Altamirano-Torres, 2018).
To determine the functional relevance of the Antp-TFIIEβ interaction, co-expression of TFIIEβ and double mutant AntpI32A-H36A was directed to the antenna, showing a drastic reduction of the antenna transformation. These findings clearly demonstrate that Antp-TFIIEβ interaction (visualized by BiFC in live larvae) is necessary for the Antp homeotic function with a very strong transformation of the antenna into T2 mesothoracic leg. Together, these results imply that very subtle changes of two amino acids in the Antp HD helix 2 can have dramatic effects on protein-protein interaction with TFIIEβ, affecting transcriptional control and the functional properties of antenna-to-tarsus transformation (Altamirano-Torres, 2018).
These results show that the interaction between TFIIEβ and Antp HD contributes to transcriptional regulation and functional activities of Antennapedia. In the Pol II PIC formation, TFIIE is a heterodimer with α and β subunits, regulating TFIIH activities such as kinase on RNA Pol II CTD, ATPase and DNA helicase. TFIIEβ binds to both TFIIB and TFIIF in important activities needed for promoter melting and stabilization as well as for the transition to elongation. Thus, Antp-TFIIEβ interaction may represent a key control point for modulation of transcription factors involved in activation or repression functions. Repression activity of Antp-TFIIEβ interaction may imply destabilization of the PIC complex or the inhibition of TFIIEβ functions modulating TFIIH ATPase, CTD kinase or helicase activities. For example, it has been determined by in vitro transcription and co-immunoprecipitation assays that the zinc-finger TF Kruppel (Kr), a Drosophila segmentation protein for late embryonic development, interacts in a dimeric way with TFIIEβ and this interaction represses transcription. If it is considered that Antp dictates leg fate by repressing the activity of antenna-determining genes such as Hth and Dll in the leg imaginal discs, it could be reasonable that Antp-TFIIEβ can be involved in repression. Co-expression of Antp with TFIIEβ resulted in a reduction to 47% of the expression of Luciferase compared with of Antp alone, however further experiments need to be done to evaluate the precise molecular mechanism of this interaction. It could also be possible that Antp facilitates the arrival of TFIIEβ to the PIC and subsequently the recruitment and/or activation of TFIIH, allowing an efficient transcription elongation. For example, mutation of Med19 on haltere imaginal discs shows that Med19 is required for Ubx target gene activation. Another example would be that Kr binds to TFIIB in a monomeric way, and this interaction activates transcription in vitro. Thus, further experiments are needed to determine the fine molecular mechanism of how interaction between Antp and TFIIEβ contribute to transcriptional regulation by activation or repression activities, or even both (Altamirano-Torres, 2018).
This study has presented a clear interaction of TFIIEβ with two amino acid positions of Antp HD that are important for Antp homeotic function, and this interplay is essential to the Antp antenna-to-tarsus transformation. In conclusion, amino acids 32 and 36 of Antp HD helix 2 play a very important role in determining the specificity of the TFIIEβ interaction. Altogether, these results provide insights into the molecular interface of Antp HD with TFIIEβ to evaluate the extent to which these molecular contacts translate into functional properties in activation or repression of target genes. The role of residues 32 and 36 on Antp helix 2 can be extrapolated for the interaction of TFIIEβ with other homeoproteins, for example Scr, Ubx and AbdA, due to the highly Hox conservation. In addition, Antp-TFIIEB interaction open the possibility to more broadly explore the interplay between Antp and additional transcription factors in the Hox interactome for the genetic control of development in Drosophila (Altamirano-Torres, 2018).
Antp has two promoters, P1 and P2. The resulting ANTP mRNAs contain 1512-nucleotide (P1) and 1727-nucleotide (P2) 5'-noncoding regions, composed of exons A, B, D, and E (P1) or exons C, D, and E (P2), respectively. Multiple AUG (translational start) codons are present in exons A, B, and C. 252-nucleotide exon D, common to mRNAs from both transcription units and devoid of AUG codons, can mediate initiation of translation by internal ribosome binding in cultured cells. Many mRNAs in Drosophila contain long 5'-noncoding regions with apparently unused AUG codons, suggesting that internal ribosome binding may be a common mechanism of translational initiation, and possibly its regulation (Oh, 1992).
cDNA clone length - 5.0 kb and 3.5 kb depending on 3'UTR length
Bases in 5' UTR - P1 - 1512; P2 - 1727
Exons - P1 - seven; P2 - six
Bases in 3' UTR - 533. There are additional downstream termination signals.
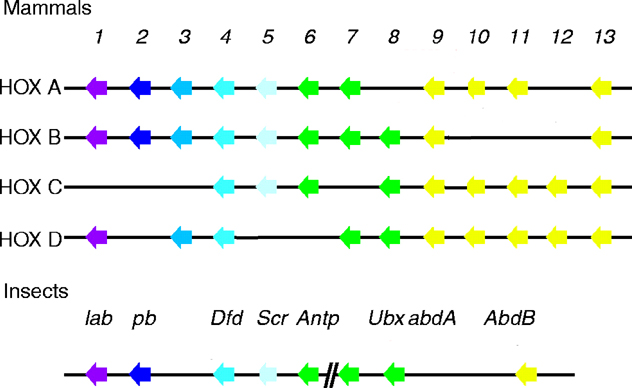
The six Drosophila proteins that belong to the antennapedia-type Homeobox subfamily are Antennapedia (ANTP), Abdominal-A (ABD-A), Deformed (DFD), Proboscipedia (PB), Sex combs reduced (SCR) and Ultrabithorax (UBX). The ExPASy World Wide Web (WWW) molecular biology server of the Geneva University Hospital and the University of Geneva provides extensive documentation for the 'Homeobox' antennapedia-type protein signature. The four paralogous Hox clusters of mammals contain eight ANTP homologs: HoxA6, HoxA7, HoxB6, HoxB7, HoxB8, HoxC6, HoxC8 and HoxD8.
The ANTP homeodomain is located near the C-terminal end. The protein is rich in prolines and glutamines. A glutamine rich region is found one third of the way into the protein (Stroeber, 1986 and Laughon, 1986).
See four paralogous Hox clusters of mammals for homologies of Antennapedia with mammalian hox proteins.
date revised: 25 August 2025
Home page: The Interactive Fly © 2025 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
{kind=link}