Ultrabithorax
The six Drosophila proteins that belong to the antennapedia-type Homeobox subfamily are
Antennapedia (ANTP), Abdominal-A (ABD-A), Deformed (DFD), Proboscipedia (PB),
Sex combs reduced (SCR) and Ultrabithorax (UBX).
The ExPASy World Wide Web (WWW) molecular biology server of the Geneva University Hospital and
the University of Geneva provides extensive documentation for the 'Homeobox' antennapedia-type protein signature. The four paralogous Hox clusters of mammals contain eight UBX homologs: HoxA6, HoxA7, HoxB6, HoxB7, HoxB8, HoxC6, HoxC8 and HoxD8.
Homeotic (Hox) genes code for principal transcriptional regulators of animal body regionalization. The duplication and divergence of Hox genes, the changes in their regulation, and the changes in the regulation of Hox target genes have all been implicated in the evolution of animal diversity. It is not known whether Hox proteins have also acquired new activities during the evolution of specific lineages. Amino-acid sequences outside the DNA-binding homeodomains of Hox orthologs diverge significantly. These sequence differences may be neutral with respect to protein function, or they could be involved in the functional divergence of Hox proteins and the evolutionary diversification of animals. A transcriptional repression domain has been identified in the carboxy-terminal region of the Drosophila Ultrabithorax (Ubx) protein. This domain is highly conserved among Ubx orthologs in other insects, but is absent from Ubx in other arthropods and onychophorans. The evolution of this domain may have facilitated the greater morphological diversification of posterior thoracic and anterior abdominal segments characteristic of modern insects (Galant, 2002).
Functional comparisons of Hox orthologs have largely focused
on their highly conserved homeodomain sequences and have
demonstrated their functional interchangeability between
species. For example, like Drosophila Ubx (DUbx), ectopic
expression in Drosophila melanogaster of the Ubx protein from an
onychophoran (OUbx) (Onychophora being a sister group to
Arthropoda) induces transformations of the antenna to leg and
the wing to haltere; it also induces ectopic activation of a decapentaplegic embryonic midgut enhancer. This indicates that OUbx
can perform some of the same molecular and developmental
functions as DUbx. However, unlike DUbx, OUbx is unable to
transform segmental identity of the embryonic ectoderm from a
thoracic to an abdominal identity or to repress the DUbx-regulated
target gene Distal-less (Dll). These functional differences between
Ubx orthologs map outside of the homeodomain (Galant, 2002).
The differences between DUbx and OUbx could be due either to
the aggregate divergence of sequences along the length of the
proteins, or to the presence of one or more discrete functional
motifs that arose in the insects or were lost in the onychophorans,
some time after the separation of their lineages from a common
ancestor more than 520 million years (Myr) ago. To better delimit
when during evolution the functional difference among Ubx orthologs
may have arisen, full-length Ubx orthologs were cloned from
two phylogenetically intermediate taxa, the red flour beetle, Tribolium
castaneum (TcUbx), and the butterfly Junonia coenia (JcUbx).
Alignment of their amino-acid sequences with DUbx and OUbx
revealed several domains that were conserved among all four Ubx
orthologs, including the MXSXFE, NGYK and YPWM motifs
amino-terminal to the homeodomain; the homeodomain itself; and
the 'Ubd-A' peptide, a motif also shared with the Abdominal A
protein, which is C-terminal to the homeodomain. It is
surmised that sequences shared by the four Ubx orthologs
probably contribute to functional similarities among them (Galant, 2002).
In contrast, sequences shared by the insect Ubx orthologs but
not by OUbx might account for functional differences between
DUbx and OUbx. Insect-restricted sequences include four regions
N-terminal to the homeodomain (I1-I4), a peptide motif
(QAQAQK), and an extended run of alanine residues C-terminal
to the homeodomain. To determine whether the presence of
these sequences correlates with DUbx functions, the
activity of TcUbx in vivo was analyzed. Ectopic expression of TcUbx throughout
the embryonic ectoderm induces the same phenotypes as those
induced by DUbx: transformation of segmental identity from
thoracic to abdominal, and repression of the activity
of a lacZ reporter gene driven by the Dll304 embryonic limb
enhancer (Dll304-lacZ), an element that is directly regulated by
DUbx in Drosophila. OUbx does not
exhibit either of these activities. This
indicates that the evolution of sequences required for these functions
arose in the Ubx protein before the divergence of Coleoptera
and Diptera, about 200-250 Myr ago, and after their divergence
from Onychophora (Galant, 2002).
To identify protein sequences responsible for the functional
differences between the insect Ubx orthologs and OUbx, chimaeric Ubx proteins were generated between DUbx and OUbx and
they were ectopically expressed in Drosophila. Several chimaeric proteins
in which different OUbx sequences were replaced with those from
DUbx N-terminal to the homeodomain were, like OUbx, completely
unable to transform thoracic larval cuticle to abdominal
identity or to repress Dll304-lacZ.
However, replacement of the short OUbx sequence C-terminal to
the Ubd-A peptide with just the 24-amino-acid sequence C-terminal
to the Ubd-A peptide from DUbx (including the QAQAQK
motif and poly-alanine stretch, QA), resulted in a chimaeric protein
(O/QA) that was competent both to transform thoracic segments to
abdominal identity, and to repress Dll304-lacZ. Importantly, ectopic expression of O/QA did not affect endogenous DUbx expression.
Therefore, the O/QA gain-of-function phenotypes observed are due
solely to the activity of the O/QA chimaeric protein. Furthermore,
simple deletion of the OUbx C terminus had no effect on the in vivo
activity of OUbx, indicating that the OUbx C
terminus does not mask an OUbx repression activity, as is the case in
a crustacean Ubx ortholog (Galant, 2002).
These results indicate that the residues critical in differentiating
DUbx function from that of OUbx are located in the C terminus of
DUbx and are sufficient to impart repression activity on the
otherwise inactive OUbx ortholog (Galant, 2002).
The C terminus of DUbx and the YPWM peptide motif located
N-terminal to the homeodomain have been implicated in mediating
interactions between DUbx and the Hox cofactor Extradenticle
(Exd). Together, the two proteins form a complex with an
increased DNA-binding affinity and regulate several embryonic
enhancers, including repression of Dll304-lacZ. It is possible
that OUbx does not possess C-terminal residues crucial for mediating
an interaction with Drosophila Exd (DExd), thus preventing its
ability to repress Dll. To investigate this possibility, the
DNA- and Exd-interacting ability of Ubx orthologs and chimaeric
proteins were analyzed. As expected, DExd alone does not bind to a DNA probe
containing a Hox/Exd composite site, and neither
DUbx nor OUbx alone binds to the probe very well. However, Drosophila Ubx and Exd together exhibited a
much higher DNA-binding affinity. Significantly,
OUbx and DExd also bind the DNA with a high affinity, indicating that onychophoran
Ubx and DExd indeed interact, as do the O/QA chimaeric protein
and DExd. Thus, the inability of OUbx to repress Dll
is not due to an inability to interact with DExd on target DNA
regulatory elements (Galant, 2002).
Rather, these results and the ability of O/QA to repress Dll304-lacZ suggest that the C-terminal QA domain may be a repression
domain. Poly-alanine-rich and glutamine/alanine-rich sequences
have been found in many repression domains in several homeodomain
proteins as well as other transcription factors. These
domains seem to mediate repression by interacting with the basal
transcriptional machinery. To test whether the DUbx QA domain
has a similar activity, its ability to repress transcription
when fused to the yeast GAL4 DNA-binding domain (GAL4DBD)
or to a chimaeric GAL4 protein that also bears a glutamine-rich
activation domain from the Drosophila Bicoid protein (GAL4DBD-Q) was examined. In transfected Drosophila S2 cells, GAL4DBD mediates a
fourfold increase in the relative activity of a UAS ß-galactosidase
reporter gene. When fused to GAL4DBD, the DUbx QA
domain completely represses activation of reporter gene expression
relative to GAL4DBD alone; the GAL4DBD-Q protein mediates 11-fold relative activation, and the DUbxQA domain, when fused to
GAL4DBD-Q, reduces activation by GAL4DBD-Q to only 1.9-fold
relative reporter activity. The magnitude of repression
mediated by the QA domain is comparable to that observed for
repression domains from other transcriptional repressors. These
assays show that the DUbx C-terminal QA domain is sufficient to
repress transcription in Drosophila S2 cells, and, combined with its
ability to confer repression activity on OUbx in vivo in Drosophila,
demonstrate that the QA domain is a discrete repression domain (Galant, 2002).
It is possible that this repression domain evolved after the split of
the Drosophila and onychophoran lineages, or was present in a
common ancestor, but was subsequently lost by onychophorans. To
address these alternative models, the phylogenetic
distribution of the C-terminal repression domain was examined. Alignment of
part of helix 3 of the homeodomains and C-terminal sequences
from a collection of Ubx orthologs has revealed that the QAQA
peptide motif is shared by all the arthropods except for Artemia. This is consistent with its presence in a common ancestor
of arthropods and its loss in the Artemia lineage. Most notably, the
insect Ubx orthologs share a remarkably conserved poly-alanine
tract, which is absent from onychophoran and other Ubx orthologs,
including that from Collembola, a sister taxon to the insects. The poly-alanine stretch thus seems to have arisen in the
insects, after their divergence from the more basal hexapods and its near-perfect conservation suggests that it is under strong
stabilizing selection. The evolution of the poly-alanine motif in the
Ubx protein in insects may have increased the repression potency of
Ubx or given it a new mode of target gene repression. There must
also be other repression domains within DUbx, because deletion of
the QA domain reduces but does not abolish DUbx repression
activity (Galant, 2002).
The evolution of this repression domain in Ubx demonstrates the
acquisition of a new function within a Hox protein while maintaining
its homeotic role. Additional studies have shown that, the serine/
threonine-rich C terminus of an Artemia Ubx ortholog modulates the repression activity of Artemia Ubx as well as
limb repression by DUbx when inserted in place of the QA domain.
Replacement of Ser/Thr-rich residues with alanine converts Artemia
Ubx to a strong repressor. Taken together, and in light of the view
that crustaceans and insects are sister taxa, these studies suggest that
a C-terminal activity-modulating Ser/Thr domain in the Ubx
protein of a common ancestor of crustaceans and insects was
replaced with the QA repression domain during early insect
evolution, and that this sequence has subsequently remained
under strong selection. Two well-known examples of genes that
evolved from Hox genes are the derivation of fushi tarazu from a
central class Hox gene and the evolution of zerknullt from a
Hox3 predecessor, but in neither case has the protein retained its
homeotic role (Galant, 2002).
The restricted phylogenetic distribution of the QAQAQK and
poly-alanine repression motifs is especially intriguing in light of its
correlation with the pattern of segmental diversity that evolved in
the insects. Primitive hexapods such as collembolans possess
abdominal limbs, and their posterior thoracic and anterior abdominal
segments are not highly differentiated. More-derived winged
insects, such as Diptera and Lepidoptera, have completely limbless
adult abdomens. Their second and third thoracic segments, which
bear wing appendages and legs, and their anterior abdomen, are
highly differentiated. The evolution of the poly-alanine repression
domain may have facilitated the diversification of these segments by
further potentiating Ubx repression of target genes (Galant, 2002).
The homeobox-complex (HOX-C) programs much of the basic body plan and forms, in most metazoans, one continuous cluster with the order of genes paralleling their order of expression along the body axis. The HOX-C organization of several Drosophila species and of Anopheles gambiae, a representative of lower Dipterans, have been compared. A model was developed to account for the evolution of the HOX-C in the Diptera (Lewis, 2003).
In D. melanogaster, the HOX-C is split into the Antennapedia complex (ANT-C) and the bithorax complex (BX-C). In D. virilis, a split is found between Ubx and abd-A, which corresponds to a separation of approximately 3-4 Mb, based on estimates in D. melanogaster (Lewis, 2003).
Recently, a third species, D. pseudoobscura, has been sequenced and aligned to D. melanogaster. In D. pseudoobscura, the split occurs between Antp and Ubx. Thus, D. pseudoobscura is more similar to D. melanogaster than to D. virilis, which is consistent with the phylogeny of these species. In contrast, D. repleta, like D. virilis, carries the split between Ubx and abd-A, which is consistent with the two species being closely related. This breakdown of the contiguity of the HOX-C in four Drosophila species is presumably a relatively recent event, since in A. gambiae the HOX-C has remained intact (Lewis, 2003).
Sequencing of the Antp and Ubx genes and the intervening region in D. virilis, comprising 308,092 base pairs (bp) (AY333070) and including the Antp gene, confirms that there is no split between Antp and Ubx. However, between Antp and Ubx a gene, CG31217, is inserted immediately downstream of Ubx. In D. melanogaster, CG31217 is located adjacent to Ubx and immediately downstream of CG31217 is the breakpoint of the split (Lewis, 2003).
The length of stretch from the end of the last exon of Ubx to the beginning of the first exon of Antp is 275 kb in A. gambiae and 190 kb in D. virilis. The length of the homologous region in D. melanogaster is 143 kb, excluding the 9.6 Mb represented by the gap. CG31217 is a newly defined gene that was formerly considered to be two separate genes. CG10013 is either absent from A. gambiae or poorly conserved compared to the Drosophila genera (Lewis, 2003).
A. gambiae lacks any obvious protein-coding genes in the interval between Antp and Ubx and the homolog of CG31217 is located distant from the HOX-C on the same chromosome arm, in a 16.2 Mb sequence scaffold (AAAB01008987) (Lewis, 2003).
These results suggest a model for the evolution of the HOX-C of the higher Diptera. At the base of the Dipterans, an ancestor is assumed with the primordial HOX-C, from which two evolutionary branches can be inferred. One branch led to A. gambiae, and the other, after insertion of CG31217, to a postulated ' vir-mel' chromosome, which subsequently led to the chromosome organization in D. virilis and to D. melanogaster (Lewis, 2003).
A hypothetical ancestor of the Diptera contains a colinear HOX-C. The hypothetical chromosome ' vir-mel' has arisen by transposition (Tp) of the gene CG31217 between the Antp and Ubx genes. To account for additional genes present in the Drosophila lineage, ' vir-mel' is postulated to have contained the Sodh gene closely linked to the Cctg gene. The ' pre-vir' chromosome aroses by a postulated inversion, Inv(pre-vir). D. virilis differs from pre-vir in having a Tp of the gene CG10013 between Antp and Ubx. In the D. melanogaster lineage, a precursor chromosome, ' pre-mel' is assumed to have arisen by an inversion, Inv(pre-mel), with breakpoints proximal to CG31217 and between the Sodh and Cctg genes of ' vir-mel' . For the modern D. melanogaster gene arrangement, a second inversion Inv(mel) is postulated that included the BX-C and extended just beyond Cctg (Lewis, 2003).
In the D. virilis lineage, an inversion, Inv(pre-vir), gave rise to a ' pre-vir' chromosome, and insertion of a second gene, CG10013, led to the present configuration in D. virilis. In D. melanogaster, CG10013 is remote from both Antp and Ubx (Lewis, 2003).
In the lineage leading to D. melanogaster, the split between Ubx and Antp is postulated to be an inversion, Inv(pre-mel) that upon an additional inversion Inv(mel) could give rise to the standard gene order. These two events placed Ubx and CG31217 next to a chaperonin-containing gamma gene (Cctg), and Antp next to a sorbitol dehydrogenase I (Sodh) gene (AE001572) (Lewis, 2003).
The simplest assumption is that Sodh and Cctg were closely linked in ' vir-mel' and that the inversion had one breakpoint between them. However, in A. gambiae these well conserved genes are located distant from each other and from the HOX-C (Lewis, 2003).
Interestingly, Inversion(3R)Payne, which is widespread among wild D. melanogaster populations, shows a gene order similar to ' pre-mel' . Moreover, the closely related D. simulans contains the only other known chromosome that is similar to ' pre-mel' . The major cytological difference between D. simulans and D. melanogaster is a large inversion with breakpoints in sections 84B and 92C. The 84B breakpoint is adjacent to, or identical with, the one that separated Antp and Ubx, whereas the 92C breakpoint is distal to the BX-C (Lewis, 2003).
Several hypotheses have been invoked to explain the persistence of the HOX-C as a single cluster. A common view holds that the cis-regulatory regions between adjacent HOX genes are bifunctional. In this scenario, splitting can occur only if preceded or accompanied by a duplication of the enhancer (Lewis, 2003).
The persistence of an intact and colinear HOX-C over hundreds of millions of years suggests that its organization is advantageous. Thus, any rearrangement that splits the HOX-C will generally become fixed only if it confers a selective advantage. Such a rearrangement is evidently a rare event. Possibly it has occurred in Drosophila for a combination of reasons: the extreme fecundity and short life cycle of Drosophila; the fact that paracentric inversions do not reduce fertility when heterozygous (which is due to the elimination of dicentric chromatids that arise from single crossovers within the inverted regions); the rarity of crossing over in the Drosophila male, and, finally, the possibility of a high frequency of transposon-mediated inversions, as has been found for D. buzzatii (Lewis, 2003).
The expression patterns of Hox genes have not previously been comprehensively analyzed in a myriapod. The expression patterns are presented of the ten Hox genes in a centipede, Lithobius atkinsoni, and these results are compared to those from studies in other arthropods. Three major findings are reported. (1) It has been found that Hox gene expression is remarkably dynamic across the arthropods. The expression patterns of the Hox genes in the centipede are in many cases intermediate between those of the chelicerates (spiders) and those of the insects and crustaceans, consistent with the proposed intermediate phylogenetic position of the Myriapoda. (2) Two 'extra' Hox genes were found in the centipede compared with those in Drosophila. Based on its pattern of expression, Hox3 appears to have a typical Hox-like role in the centipede, suggesting that the novel functions of the Hox3 homologs zen and bicoid were adopted somewhere in the crustacean-insect clade. In the centipede, the expression of the gene fushi tarazu suggests that it has both a Hox-like role (as in the mite), as well as a role in segmentation (as in insects). This suggests that this dramatic change in function was achieved via a multifunctional intermediate, a condition maintained in the centipede. (3) It was found that Hox expression correlates with tagmatic boundaries, consistent with the theory that changes in Hox genes had a major role in evolution of the arthropod body plan (Hughes, 2002).
The expression of the Hox genes corresponds roughly with the tagmatic divisions in the centipede. The expression of the genes lab, pb, Hox3 and Dfd is confined to the head, while the trunk is apparently under the control of Antp, Ubx, abd-A and Abd-B. Interestingly, the maxilliped segment has expression of three genes that extend both into the head (Scr and ftz) and into the trunk (Antp). The maxilliped segment is thought to be homologous to the first trunk or thoracic segment of other mandibulate arthropods. The appendages of this segment in the centipede, however, have been highly modified. While their leg-like structure is still evident, they develop to become short and broad fangs, complete with a poison gland. Thus, the first legs of the centipede are modified to become more mouthpart-like, and are used for prey capture and manipulation. This mixed head/trunk identity of the segment seems to be reflected in the Hox code found there. While the segment itself has only a 'trunk' Hox gene (Antp), the appendages have expression of Antp as well as the 'head' genes Scr and ftz, which are also expressed in the maxillary II segment. It remains to be determined how these genes contribute to the development of the centipede fangs. It would also be interesting to know whether the evolution of this novel appendage is correlated with a shift in the expression of these genes. Further studies of Hox expression in other myriapods such as a millipede, or functional studies in the centipede, would be very interesting regarding these issues (Hughes, 2002).
Expression of genes along the centipede trunk is, like the morphology of the trunk, fairly homonomous. Antennapedia extends along the whole trunk in early stages, and later retracts to cover legs one through four. It is not clear whether this later, more restricted domain imparts any developmental difference to these segments, as none is evident morphologically. It is intriguing to note that this restriction to the anteriormost segments of the trunk is reminiscent of a similar restriction of Antp expression in the pleon of malacostracan crustaceans and the thorax of insects. Perhaps the domain of Antp expression was restricted to the anterior portion of the trunk in the myriapod-like mandibulate ancestor, but was only exploited fully in the specialized differentiation of the crustaceans and insects. In the centipede, Ubx and abd-A expression patterns are similarly expressed along the trunk, although Ubx expression fades from the extreme posterior segments. Expression of Abd-B is strongest in the telson, but faint expression extends over the mid-region of leg segments two to seven. Since the genes Ubx, abd-A and Abd-B are likely to have similar roles in patterning the trunks of all mandibulates, it is suggested that the myriapods have developed their unique body plan largely by expanding the number of segments under the control of the 'trunk' genes. This is a similar scenario to that provided by recent findings that snakes seem to have created an elongated body by increasing the numbers of somites under the control of thoracic Hox genes (Hughes, 2002).
Three aspects of UBX mRNA processing have been conserved among D. melanogaster, D. pseudoobscura, D. hydei and
D. virilis, species separated by as much as 60 million years of evolution. The four species exhibit
identical patterns of optional exon use in a region adjacent to the homeodomain. These four
species produce the same family of UBX protein isoforms with identical amino acid sequences in
the optional exons, even though the common amino-proximal region has undergone substantial
divergence. The tissue- and stage-specific patterns of expression of different UBX
isoforms are identical among these Drosophila species (Bomze, 1994).
Hox genes are known to control the identity of
serially repeated structures in arthropods and vertebrates.
The expression pattern of the Hox genes Deformed, Sex combs reduced, Antennapedia, and Ultrabithorax/abdominal-A (Ubx/abd-A)
from the honey bee Apis mellifera were analyzed. A
cDNA with the complete coding region of the Antennapedia
gene from Apis was cloned. Comparison with Antp proteins
from other insect species revealed several regions of homology.
The expression patterns of the isolated Hox
genes from Apis show that the original expression patterns
of Dfd, Scr, and Antp appear between late blastoderm
and early germ band stage in a temporal and spatial
sequence. Each of them shows up as a belt, spanning approximately
two segment anlagen; Dfd in the anterior
gnathal region, Scr in the posterior gnathal and anterior
thoracic region, and Antp in the thoracic region. Following
expansion of the Antp domain in the abdomen as a
gradient towards the posterior, Ubx/abd-A expression appears
laterally in the abdomen. During gastrulation and
in the germ band stage the domains of strong expression
do not overlap any more, but touch each other. After gastrulation
the borders of the expression domains partly
correlate with parasegment and partly with segment
boundaries. Laterally, gaps between the domain of each
gene may show no expression of any of the genes examined (Walldorf, 2000).
In the blastoderm stage the expression domains of all
four genes are similar in fruit fly and honey bee embryos.
Certainly these genes play very similar roles in establishing
the general character of that part of the insect
body where they have their main expression domain.
Even in Chelicerata the orthologs show comparable
patterns. However, some differences should be mentioned. In the
fly, the borders of the Dfd domain are segmental. In
the bee, the anterior expression border is initially parasegmental
in the rudiment of the intercalary segment: this changes from segmental to parasegmental during
gastrulation in the ventral part of the maxillary segment. The steady Engrailed pattern shows that this change is not a result of cell movement. The same development of that pattern is reported for Tribolium. This is most
clearly seen ventrally in the parasegmental Scr pattern in
the germ band of the bee, corresponding to segmental in
that area in the fly. While some differences in the Dfd pattern are
consistently observed between the bee and the fly, the
complicated posterior borders of the Scr and the Antp
patterns of young and old embryos of flies and bees are identical. The
same Antp pattern is also found in Thermobia domestica. However, in Manduca sexta the
anterior border of Antp is the compartment border of T1. It may be that some of these differences are only the result of different sensitivity levels of
the stainings. Probably the ventral switch from segmental
to parasegmental is more general in insects but is not
seen very clearly in some cases, such as in the anterior
part of the fruit fly head, due to the subsequent head involution.
The resulting ventrolateral corners of the expression
borders of Dfd, Scr, and Antp correlate fairly
well with the position of the appendage anlagen and may
help to pin down their position (Walldorf, 2000).
The morphological and functional evolution of appendages has played a critical role in animal evolution, but
the developmental genetic mechanisms underlying appendage diversity are not understood. Given that homologous
appendage development is controlled by the same Hox gene in different organisms, and that Hox genes are transcription
factors, diversity may evolve from changes in the regulation of Hox target genes. Two impediments to understanding the
role of Hox genes in morphological evolution have been the limited number of organisms in which Hox gene function can
be studied and the paucity of known Hox-regulated target genes. An analysis was carried out of Hindsight, a butterfly homeotic mutant
in which portions of the ventral hindwing pattern are transformed to ventral forewing identity, and the regulation of target genes by the Ultrabithorax (Ubx) gene product was compared in Lepidopteran and Dipteran hindwings. Ubx gene expression is lost from patches of cells in developing Hindsight hindwings, which correlates
with changes in wing pigmentation, color pattern elements, and scale morphology. This mutant was used to study how regulation of target genes by Ubx protein differs between species. Several Ubx-regulated genes in the Drosophila haltere are not repressed by Ubx in butterfly hindwings, but Distal-less (Dll) expression is regulated by Ubx in a unique manner in butterflies. In Hindsight hindwings, in which patches of cells that lack Ubx protein expression encompass a portion of the eyespot focus, Dll expression clearly increases, as compared with that found in wild-type hindwings. Outside of these patches, where Ubx expression is 'normal' in the eyespot field, Dll is expressed at very low levels in a cell-autonomous fashion. These results suggest that hindwing eyespot size may be controlled by Ubx at two steps in the eyespot developmental pathway: (1) Ubx depresses the production of the focal signal, which is relieved when a portion of the focus loses Ubx expression; (2) Ubx affects the response of genes that are downstream of the focal signal, as for example, Dll. Because the eyespot pattern element has no counterpart in other insect orders, it is deduced that Ubx regulation of eyespot patterning genes must have evolved within the Lepidoptera. It is concluded that the morphological diversification of insect hindwings has involved
the acquisition of different sets of target genes by Ubx in different lineages (Weatherbee, 1999).
Drosophila Serum response factor (blistered), Achaete-Scute Complex, and wingless are repressed in Drosophila halteres. Portions of the expression pattern of Lepidopteran homologs of these genes are not repressed in butterfly hindwings. Unlike the expression patterns of the homologous genes in halteres, butterfly wg is not repressed along the posterior margin in the hindwing, nor is butterfly SRF repressed in intervein regions, and the AS-C homologs are not repressed in cells flanking the dorsal-ventral boundary. These differences in the regulation of wg, SRF and AS-C between Drosophila halteres and butterfly hindwings suggest that these genes became repressed by Ubx when an ancestral hindwing evolved into a haltere in the dipteran lineage, with a concomitant reduction of appendage size, loss of margin bristles, and changes in shape. Two additional exampes of Ubx-regulated differences in gene expression between fly and butterfly flight appendages were found. (1) wg is expressed in two stripes in butterfly forewings that roughly correspond to the future location of the proximal band elements. This protein of the wg pattern is absent from butterfly hindwings and has not counterpart in flies and represents a novel feature regulated by Ubx in butterflies. (2) Dll is expressed along the margin of both butterfly wings and the Drosophila forewing, but this expression is modified in halteres and may be regulated by Ubx.
Changes in Hox-regulated target gene sets are,
in general, likely to underlie the morphological divergence of homologous structures between animals (Weatherbee, 1999).
Developmental processes have been traditionally viewed to be invariant within higher taxa. However,
examples are known whereby closely related species exhibit alterations in early embryogenesis yet
appear very similar as adults. Such developmental changes are thought to occur in response to shifts in
life history. In insects, the regulation of embryonic development has been intensively studied in model
species, such as Drosophila melanogaster. Previous comparative studies suggest that the developmental processes documented in Drosophila adequately describe embryogenesis of advanced, holometabolous, insects generally. There have been few attempts, however, to take into account how life history has
influenced early development of insects or to characterize early development of species with life
histories fundamentally different from flies. A comparison was carried out of early development in two species from
the same family of parasitic wasps that exhibit very different life histories. Bracon hebetor is an
ectoparasite that lays large, yolky eggs on the integument of its host that develop much like the
free-living honeybee and Drosophila. In contrast, Aphidius ervi is an endoparasite that lays small and
apparently yolk-free eggs that develop in the hemocoel of the host. This wasp exhibits a radically
different mode of early development at both the cellular and molecular level from B. hebetor. The
developmental changes in A. ervi reflect functional adaptations to its derived endoparasitic life history and argue
that departures from the fly paradigm may occur commonly among insects whose eggs develop under
conditions different from typical terrestrial species (Grbic, 1998).
To compare patterning events at the molecular
level, B. hebetor and A. ervi embryos were stained with antibodies that recognize conserved
epitopes of Eve, En, and Ubx/Abd-A in different insect species. Eve, a primary
pair-rule gene is expressed in the Drosophila syncytium, forms a characteristic
seven-stripe pattern with double segment periodicity. En, which is regulated by Eve, is a
segment polarity gene that specifies the posterior segmental compartments. Ubx and
Abd-A are Drosophila homeotic proteins that specify the posterior thorax and abdomen.
In B. hebetor, Eve is expressed in a largely conserved fashion when compared to Drosophila and other
long germband insects. Initially, a broad domain of Eve expression splits into broad
pair-rule stripes, followed by a split of the individual pair-rule stripes in rapid
anteroposterior progression to form segmentally iterated stripes. After germband
retraction, Eve localizes in the cells of the dorsolateral mesoderm and
neurons; a pattern conserved in all examined insects. In B. hebetor, En is
expressed in a rapid anteroposterior progression, forming a mature pattern of
segmentally iterated stripes that localize to the posterior segmental compartments.
The antibody against Ubx/Abd-A stains the region from the posterior thorax to the
penultimate abdominal segment. When A. ervi embryos are stained with anti-Eve, neither a pair-rule
nor a segmental pattern is detected. In the extended germband, however, an Eve antigen is detected in
dorsolateral mesoderm and neurons. En stripes appear when embryos
initiate germband extension. These stripes form sequentially as the germband
extends, resulting in a mature pattern of segmentally iterated stripes that localize
to the posterior segmental compartments. Ubx/Abd-A is expressed in the posterior
thorax and abdomen in the retracted germband stage. Thus, despite the divergence of early patterning, late patterning in B. hebetor
and A. ervi includes conserved expression of En and Ubx/Abd-A. This expression pattern in
the germband, the phylotypic stage in insects, suggests conservation of this stage
irrespective of how development begins (Grbic, 1998).
Ultrabithorax is essential for the proper
patterning of the posterior thorax and anterior abdomen in
Drosophila. The Coleoptera and Diptera differ in the organization
and structure of their thorax and anterior abdomen.
Changes in the regulation of Ubx and/or its downstream
target genes are predicted to underlie these altered
morphologies. The feasibility of genetic analysis
in the red flour beetle, Tribolium castaneum, was exploited to examine
the role of its Ubx ortholog in development. Genomic and cDNA clones that predict a polypeptide
with nearly 100% identity with the Drosophila Ubx gene in
the homeodomain and flanking sequences were examined. Southern blot
analysis indicates that these clones represent DNA sequences
within the Homeotic complex (HOM-C) of Tribolium.
Phenotypic analysis of mutant variants of the Ultrathorax
(Utx) gene, and its location within the beetle HOM-C,
strongly supports Utx being the Tribolium ortholog of
Ubx. The embryonic expression pattern of Ubx-homologous
transcripts coincides with the phenotypes associated
with Utx mutations, providing support that the Ubx-homologous
cloned DNA represents the Utx locus. By mid-germ-band
extension Utx transcripts are expressed in a pattern
similar to Ubx in Drosophila. However, during early germ-band
formation Utx transcripts differ in both spatial and
temporal progression. Utx expression is initially detected
in parasegments 4 and 5 (T1p-T3a) because they are established
during early germband formation. This is the first report of
the wild-type parasegmental expression of an insect Ubx
ortholog extending through parasegment 4. The earlier and
more anterior expression in the thorax may underlie the
modification of the Coleopteran thorax (Bennett, 1999).
Despite these similarities there are notable differences
in both the spatial and temporal progression of Utx expression
compared with Ubx in Drosophila. Expression
of Ubx is initially detected in PS6, spreading anteriorly
to PS5 and posteriorly to PS12 later in development. In Tribolium, Utx
expression initially extends from PS4 through the posterior
end of the segmented germband (PS16) and is later
restricted to PS5 through PS13 in the ectoderm and mesoderm.
Throughout development Utx is expressed more
posteriorly than its counterpart in Drosophila (with a
posterior limit of PS12). This finding is similar to what
has been observed for the Tribolium abd-A ortholog. The more posterior
expression of both the Ubx and abd-A homologs in
Tribolium relative to Drosophila is correlated with a
change in the expression of the Tribolium Abd-B ortholog.
In Drosophila Abd-B represses Ubx and abd-A transcription
in the posterior abdominal segments. In
Tribolium the anterior limit of Abd-B expression is PS14,
corresponding to the r function found for Abd-B in Drosophila.
The parasegmental expression of a Ubx homolog
throughout PS4 is unique to Tribolium among the insects
studied to date, although in
Drosophila a small group of cells in PS4 expresses Ubx
during late embryogenesis. A PS4 enhancer element has been identified in the
first intron of the Drosophila Ubx locus using an enhancer
trap assay. A P-element insertion
near the Ubx promoter drives lac-Z expression
in PS4 throughout embryogenesis, although lac-Z
expression is identical to the wild-type Ubx pattern in
the imaginal disks. When the Ubx promoter and up-stream
sequences are deleted, the PS4 expression is
detected only through germband extension, resulting in a
pattern that is quite similar to that observed for
the wild-type expression of Utx in Tribolium (Bennett, 1999 and references therein).
The Tribolium orthologs of both hunchback and ftz,
both regulators of Ubx expression in Drosophila, have
been isolated.
Analysis of these orthologs in Tribolium has shown that
they are not expressed in identical domains of their respective
Drosophila counterparts. Thus they may have
different effects on homeotic gene expression. In Tribolium, Hunchback
protein is detected early in the serosal cells and PS2-3;
however, analysis of the later PS4 expression, especially
in relation to the early expression of Utx, has not been
examined. This is a critical point because
Hunchback protein represses Ubx expression anterior to
PS5 in Drosophila. The Tribolium ftz ortholog is expressed
in a pair-rule pattern in Tribolium; however, it is
not essential for Tribolium segmentation. In Drosophila ftz is a positive
regulator of Hox gene expression in even numbered
parasegments. The early expression of Utx in PS4 may
reflect a shift in balance between repression by Hunchback
and activation by Ftz proteins, either through changes
in the Hunchback or Ftz proteins or in the regulatory
regions surrounding the Utx gene (Bennett, 1999 and references therein).
Antibodies were used to examine the expression
patterns of Antennapedia (Antp), Ultrabithorax
(Ubx), Ubx and abdominal-A combined (Ubx/abd-A),
and Distalless (Dll) in the embryos of the moth Manduca
sexta. The spatial and temporal pattern of
Antp expression in Manduca is correlated with the anterior
migration of two patches of epithelium that include
the anterior-most tracheal pits, and with the development
of functional spiracles. Ubx expression shows
an intricate pattern that suggests complex regulation
during development. Throughout Manduca embryogenesis,
the expression of Ubx/Abd-A and Dll is similar to
that reported for other insects. However, there is no
apparent reduction in Ubx/Abd-A expression in the
Manduca abdominal proleg primordium that expresses
Dll. The expression of these four proteins was also examined
in embryos of the Manduca homozygous homeotic mutant Octopod (Octo). The Octo mutation results in the transformation of A1 and A2 in the anterior direction, with homeotic legs appearing on A1 and occasionally
A2. These results suggest that in Octo animals there is a reduction in the level of Ubx protein expression throughout its domain (Zheng, 1999).
Insects show a dramatic diversity in the number and segmental
distribution of abdominal appendages. For example,
among Lepidoptera larvae, the number of abdominal
appendages varies from none to seven pairs. The number of appendages also varies during the course of embryonic development. During early stages
of embryogenesis of Pieris rapae and Bombyx mori ventral appendages
are present on all abdominal segments. Later
some of these appendages regress, leaving prolegs on
A3-A6 and on the terminal segment. In Drosophila both Ubx and Abd-A act to repress Dll and the development of abdominal appendages. However, such repression is absent in the crustacean, indicating that the repressive
function of Ubx and Abd-A evolved in insects. The presence of homeotic legs in Octo Manduca
demonstrates that Manduca Ubx also represses appendage
development. However, in the beetle Tribolium
and the grasshopper Schistocerca an A1 appendage, the
pleuropodia, develops despite high level of Ubx in A1 appendage
primordium. Thus the repressive function of Ubx on A1 appendage development
evolved late in insect evolution, in the Diptera/lepidoptera
lineage. These findings indicate that the repressive function
of Abd-A evolved even later than that of Ubx. The expression
of Dll and the emergence of prolegs in A3-A6 apparently
initiated in the presence of strong Abd-A expression,
suggests that in Manduca the repressive function of
Abd-A on A3-A6 appendages has not yet evolved. This conclusion for Manduca Abd-A differs from that which has been suggested for the butterfly Precis, where Abd-A is believed to suppress appendages development. In order to develop prolegs both Ubx and Abd-A are locally repressed in the proleg primordia (Zheng, 1999).
In an abdominal segment the A1 appendage
developed in Octo is a thoracic leg instead of an abdominal
proleg. This suggests that the level of Ubx/Abd-A
protein determines the type of appendage that develops in
the abdominal region. This observation is consistent with
the following hypothesis: high levels of Ubx and/or Abd-A
protein(s) act to suppress leg development, while moderate
levels of expression, as occur in T3 of wild-type and
A1 and A2 of Octo animals, is a permissive environment
for thoracic limb development. The expression of Abd-A
in the near absence of Ubx protein directs the development
of abdominal prolegs, as in A3-A6. Based on this
hypothesis, the type of A2 homeotic appendage in Octo
should be variable and depend on the level of Ubx expression
on A2. If the expression of Ubx in A2 of Octo is
above a certain threshold level, thoracic legs should develop
on A2. However, if the expression is lower, the A2 appendages
should develop as abdominal prolegs. Consistent
with this is the finding that in Octo animals, the identity
of the A2 homeotic appendage is variable. Some A2
homeotic limbs have characteristics of thoracic legs, while the others possess the characteristics typical of the abdominal prolegs. This model does not explain the absence of prolegs in A7, A8 and their presence in the terminal segment. However, it is quite possible that a proper combination of Abd-A and Abd-B dictates the pattern of appendage development in these more posterior segments (Zheng, 1999).
During the embryogenesis of Drosophila, the homeotic genes are required to
specify proper cell fates along the anterior-posterior axis
of the embryo. Partial cDNAs of homologs
of the Drosophila homeotic gene teashirt and five of the
homeotic-complex (HOM-C) genes were cloned from the thysanuran
insect, Thermobia domestica (the firebrat), and these genes were assayed for their embryonic
expression patterns. The HOM-C genes examined
were labial, Antennapedia, Ultrabithorax, abdominal-A and Abdominal-B. Since the expression pattern of these HOM-C genes is largely conserved among insects
and since Thermobia is a member of a phylogenetically basal
order of insects, the ancestral expression patterns of these genes in insects could be inferred. The expression patterns of the Thermobia HOM-C genes were compared with their
expression in Drosophila and other insects;
the potential roles these genes may have played in insect
evolution are discussed. Interestingly, the teashirt homolog shows
greater variability between Thermobia and Drosophila
than any of the HOM-C genes. In particular, teashirt is
not expressed strongly in the Thermobia abdomen, unlike
Drosophila teashirt. It is proposed that teashirt expression
has expanded posteriorly in Drosophila and contributed to a homogenization of the Drosophila larval thorax and abdomen (Peterson, 1999).
The earliest domain of Ubx expression and
much of its dynamic pattern is conserved among Thermobia,
Drosophila, and Schistocerca and this pattern likely
reflects the ancestral expression pattern. One observed
divergence suggests how changes in the Ubx expression
pattern might be involved in the morphological evolution
of insects. Ubx expression has been best characterized in Drosophila,
where Ubx is first expressed in the anterior compartment
of A1 but expands posteriorly and anteriorly during
embryonic development. Posteriorly, Ubx becomes
expressed in the anterior compartments of the abdominal
segments. Anteriorly Ubx expression appears in a lateral
domain in pT2 and the first half of aT3. Later, the
pT2/partial-aT3 expression expands into the ventral regions
and into pT3.
The early function of Ubx in Drosophila is to initiate
abdominal development and repress thoracic development, particularly appendage
development. Consistent with this function the earliest anterior border of
Ubx expression in grasshoppers and
firebrats is at the T3-A1 segment border, just as it is in
Drosophila. The particular role of the later Ubx expression
in the firebrat and grasshopper thorax is unknown,
but it is likely that it modifies thoracic development leading
to specializations of T2 and T3, as it does in Drosophila. It is possible, however, that Ubx may have
evolved a more efficacious role in pT2-aT3 of higher insects,
since Drosophila Ubx mutations affect pT2-pA1, but
Tribolium Ultrathorax (Utx, or here, Tc-Ubx) mutations appear
only to affect pT3-aA1. It is interesting that the expression of Ubx in T2 and
T3 evolved before the divergence of pterygotes and apterygotes.
One early hypothesis concerning Ubx was that
it evolved in drosophilids to modify the T3 wings into
halteres. This was disproved when it was found that Ubx is expressed in the
T3 imaginal discs of Precis where it is thought to modify
the T3 wings into hindwings. The existence of Ubx expression
in T2 and T3 in firebrat embryos argues that changes in function, not
expression pattern, of Ubx have led to the modification
of the T3 wing, although the relative positions of the potential
wing primordia and Ubx expression have not been
determined (Peterson, 1999).
In contrast, Ubx expression in the T3 leg is not conserved
among insects. In Schistocerca, Ubx accumulates
in the embryonic T3 'jumping' leg, which is much larger
than the other thoracic legs. Similar
expression is seen in the Drosophila T3 imaginal leg
disc. Such expression is not seen in firebrats,
where Ubx accumulates only around the basal periphery
of the structurally similar T2 and T3 limbs. Thus
it is possible that expression of Ubx in the T3 leg is an
expansion of the ancestral Ubx expression pattern and
may be part of a process that modifies the T3 leg, making
it possible to have a unique differentiation or specialization (Peterson, 1999).
The Hox genes have been implicated as central to the evolution of animal body plan diversity. Regulatory changes both in Hox
expression domains and in Hox-regulated gene networks have arisen during the evolution of related taxa, but there is little knowledge of
whether functional changes in Hox proteins have also contributed to morphological evolution. For example, the evolution of greater
numbers of differentiated segments and body parts in insects, as compared with the simpler body plans of arthropod ancestors, may have
involved an increase in the spectrum of biochemical interactions of individual Hox proteins. The in vivo functions of
orthologous Ultrabithorax (Ubx) proteins from the insect Drosophila melanogaster and from an onychophoran, a member of a sister phylum with a more primitive
and homonomous body plan are compared. These Ubx proteins, which have been diverging in sequence for over 540 million years, can generate many of the same gain-of-function
tissue transformations and can activate and repress many of the same target genes when expressed during Drosophila development. However, the onychophora
Ubx (OUbx) protein does not transform the segmental identity of the embryonic ectoderm or repress the Distal-less target gene. This functional divergence is due to
sequence changes outside the conserved homeodomain region. The inability of OUbx to function like Drosophila Ubx (DUbx) in the embryonic ectoderm indicates
that the Ubx protein may have acquired new cofactors or activity modifiers since the divergence of the onychophoran and insect lineages (Grenier, 2000).
A novel single-sided specific polymerase chain reaction (PCR) strategy inspired by ligation-mediated PCR has been used to clone
fragments of divergent homeobox genes from a flatworm, the planarian Polycelis nigra. Eight homeobox-containing fragments were
amplified, belonging to the Hox, msh, NK-1 and NK-2 classes. Together with the results obtained from several genomes of
platyhelminths, this screening shows the presence of the same array of homeodomain developmental regulators in planarians,
traditionally regarded as primitive metazoans in terms of body plan, as in coelomate organisms. However, the presence of a
Ubx/abd-A homolog may indicate that platyhelminths are more closely related to protostomes than to deuterostomes and supports the
idea that flatworms have inherited an elaborate HOX cluster (seven or eight genes) from their ancestor. Likely homologs of the fly
genes tinman, bagpipe and S59 suggest that the mesoderm might be patterned by the same genes in all bilaterally symmetrical animals.
Finally, a msh-like gene, a family known to be involved in inductive mechanisms in vertebrates, has been found. These results support
the hypothesis that the tremendous diversity of metazoan body plans is specified by a largely conserved array of homeobox-containing
developmental genes (Balavoine, 1996).
Changes in the expression of the Hox genes Ultrabithorax and AbdominalA in different crustaceans correlate well with the modification of their anterior thoracic limbs into feeding appendages (maxillipeds). In branchiopod crustaceans (such as Artemia), which do not have maxillipeds, Ubx and abdA are expressed throughout the thoracic region. In peracarids, the first, and sometimes second, of the eight thoracic segments bear limbs that have acquired several characteristics of feeding appendages. The modification of these segments correlates with the repression of Ubx and abdA in these segments. Uniform early expression becomes modulated within individual metameres during later development. Decapods are generally described as having three pairs of maxillipeds and five pairs of walking limbs in their thorax. In Periclimenes Ubx and abdA expression is excluded from the first three thoracic parasegments and limbs, is weaker in T4, and stronger in more posterior segments. In Homarus, only the T1 and T2 limbs appear to be distinctly reduced at hatching. Ubx and abdA staining is absent from the first two thoracic parasegments and strong in T3 and more posterior segments. Thus, the anterior boundary of embryonic expression of Ubx and abdA in Homarus appears to be shifted backwards by two metameric units corresponding to the morphological transition in thoracic limbs seen at hatching. It is suggested that spatially modulated distribution of Ubx and abdA expression and temporal changes in the expression of Hox genes are responsible for different decisions on regional identity. In some limbs identity could be determined as a mosaic, with some parts of a segment retaining a thoracic identity and others becoming homeotically transformed to a gnathal fate (Averof, 1997).
Representatives of the Insecta and the Malacostraca
(higher crustaceans) have highly derived body plans
subdivided into several tagma (groups of segments united
by or fused into a common function and/or morphology). The
tagmatization of segments in the trunk, the part of the body
between head and telson, in both lineages is thought to have
evolved independently from ancestors with a distinct head
but a homonomous, undifferentiated trunk. In the
branchiopod crustacean, Artemia franciscana, the trunk
Hox genes are expressed in broad overlapping domains
suggesting a conserved ancestral state. In comparison, in
insects, the Antennapedia-class genes of the homeotic
clusters are more regionally deployed into distinct domains
where they serve to control the morphology of the different
trunk segments.
In Drosophila Antp is expressed in
and required for the specification of the three-segmented
locomotory thorax. Both Ubx and abd-A are involved in the
development of the legless abdomen. Ubx is also expressed in the posterior thorax where it is known to be involved in the development of the modified hind
wings, the halteres. Thus an originally Artemia-like pattern of
homeotic gene expression has apparently been modified in
the insect lineage associated with and perhaps facilitating
the observed pattern of tagmatization. Since insects are the
only arthropods with a derived trunk tagmosis tested to
date, the expression patterns of the Hox genes
Antp, Ubx and abd-A were examined in the malacostracan crustacean
Porcellio scaber (Oniscidae, Isopoda). Unlike
the pattern seen in Artemia, these genes are expressed in
well-defined discrete domains coinciding with tagmatic
boundaries that are distinct from those of the insects. These
observations suggest that, during the independent
tagmatization in insects and malacostracan crustaceans,
the homologous 'trunk' genes evolved to perform different
developmental functions. It is also proposed that, in each
lineage, the changes in Hox gene expression pattern may
have been important in trunk tagmatization (Abzhanov, 2000).
Contemporary molecular and morphological phylogenies of
the Crustacea indicate that this group comprises a
monophyletic assembly with some classes such as the
Remipidia and Branchiopoda at a basal position and the
Malacostraca as a crown group. Additionally, according to recent phylogenies, the
Crustacea are placed as the sister group to the Insecta in the
subphylum Mandibulata. Alternatively, some studies suggest
that crustaceans may be paraphyletic with regard to the Insecta
with the Malacostraca as the closest sister group to insects. The
Mandibulata also includes the more distantly related
Myriapoda. The Chelicerates are generally regarded as a sister
group to the Mandibulata (Abzhanov, 2000 and references therein).
In insects, Ubx possesses a well-conserved expression
domain in the abdomen. In Drosophila the earliest expression is seen first in A1
and more posterior abdominal segments.
This early domain of Ubx expression is invariant amongst all
the classes studied.
In these insects, later in development, Ubx expands into the
posterior thorax. The exact extent of this shift is different from
order to order, e.g., in Drosophila, Ubx moves into the hind
wing and hind leg primordia, whereas, in the firebrat, it is
restricted to the posterior portion of the segments at the periphery
of the T2 and T3 legs. The two distinct
temporal domains likely reflect the two different
developmental functions of Ubx: the early function of Ubx is
to specify abdominal identity whereas later it plays a
modifying role in specialization and differentiation of existing
thoracic structures (Abzhanov, 2000).
In Artemia Ubx is expressed throughout the trunk with the
anterior boundary in the first trunk segment. Interestingly, the anterior boundary of Ubx, as
detected with the FP6.87 antibody, has been modified several
times during crustacean evolution. This change correlates
with the transformation of the anterior legs into maxillipeds
that have lost Ubx expression. In
isopod malacostracans, including P. scaber, this is seen in the
first pair of trunk legs, which are transformed into maxillipeds
during development and never express Ubx. The posterior
boundary of PsUbx expression falls at the pereon/pleon border.
Note, that unlike PsAntp whose domain reflects embryonic
morphological junctions, PsUbx demarcates the future adult
cephalon/pereon and pereon/pleon borders. A comparison with
the domain of Ubx in insects reveals that PsUbx exhibits a
surprising dissimilarity, since this gene is expressed in the
abdomen of insects but in the pereon (analogous to the insect
thorax) of Porcellio (Abzhanov, 2000).
In summary, the expression domains of the trunk genes in P. scaber are distinct from both insects and branchiopod
crustaceans. They are better defined than the broadly
overlapping domains in A. franciscana and despite a superficial
resemblance to the discrete domains of their insect
homologs, the anterior and posterior expression domain
boundaries are quite different from those in insects. These
observations suggest that the trunk genes were co-expressed
and performed redundant roles in the homonomous trunk in the
last ancestor of insects and higher crustaceans, and that the
trunk of the ancestor has independently differentiated into the
thorax/abdomen of insects and pereon/pleon of
malacostracans via specialization in the deployment and
function of the Hox genes. This being the case, it is likely that
the homologous Hox trunk genes have evolved to acquire
different developmental functions in the closely related classes
Insecta and Malacostraca (Abzhanov, 2000 and references therein).
The regulatory function of the mouse introns for Hoxa-4 and Hoxb-4 (homologs of Drosophila Deformed) was analyzed in Drosophila. Introns of both these genes contain a cluster of three homeodomain binding sites called the HB1 element, which is also found in the introns of other Hox genes ranging from fish to humans as well as in the Ultrabithorax and decapentaplegic genes of Drosophila. The enhancer of the Hoxa-4 intron responds to several homeobox genes by activating transcription in cells of Drosophila embryonic epidermis. The response is strong to Deformed and Ubx. The enhancer activity is similar to previously described autoregulatory elements of Deformed, but additional expression is observed in more posterior segments activated by Ubx and repressed by abdominal-A. Point mutations in the homeodomain binding sites in HB1 abolish the HB1 enhancer activity. A second site suppression experiment shows that UBX interacts directly with the HB1 element. When the HB1 element in the Hoxa-4 intron is replaced by that of the mesodermal enhancer of dpp, which is under contol of Ubx, Ubx-dependent activation is retained, but repression by abd-A is lost. The same result is obtained when the third binding site of HB1 is altered, suggesting that this site is reponsible of abd-A-dependent repression. Deletion of potential cofactor binding sites flanking the HB1 element reveals that they are important for enhancer function in Drosophila and that the Dfd-dependent and the Ubx-dependent expression requires different sites. The cofactor sites are likely to render cell specific expression. The Hox-b4 intron is not functional in Drosophila (Haerry, 1997).
Insects are easily distinguishable by the absence of legs on the adult abdomen. Studies performed on the Dipteran, Drosophila melanogaster, indicate that this is because of the repressive effects of the homeotic genes Ultrabithorax and abdominal-A on the limb promoting gene Distal-less during embryonic development. However, in many species appendage-like
structures are present on abdominal segments in embryonic and juvenile stages. By using classical genetics and double-stranded
RNA-mediated gene silencing in the red flour beetle, Tribolium castaneum, a species that develops an appendage on the first
abdominal segment, it was possible to examine the roles of Ubx and Abd-A in abdominal limb development. In Tribolium, Abd-A, but not Ubx, represses early
expression of Dll in the embryonic abdomen. Ubx appears to modify the A1 appendage. This difference in the activities of Abd-A and Ubx is critical for proper
development of this appendage. It is suggested that an ancestral role of Abd-A in insect abdominal appendage development was in the repression of Dll initiation and
that an ancestral role of Ubx was in modulation of abdominal appendage morphology (Lewis, 2000).
By examining TcDll and TcEn expression in TcUbx and
Tcabd-A mutant embryos, a better
understanding of the role of each in suppressing and modifying limb
programs in the beetle abdomen was obtained. In TcUbx mutant embryos,
TcDll expression in the abdomen remains restricted to anterior A1, whereas, in Tcabd-A, mutant embryo
TcDll is ectopically expressed in each abdominal segment,
resulting in abdominal appendage development. These results clearly
support the role for TcAbd-A as a primary TcDll
repressor (and therefore appendage repressor) in the
Tribolium abdomen. The role of TcUbx in
regulating Dll expression appears to be more complex. Although
TcDll and TcUbx are initially coexpressed during
early pleuropod development, later TcUbx is absent in the
TcDll-expressing cells, leaving open the possibility that
TcUbx represses TcDll late in development. Whether or not late expression of TcUbx represses
TcDll expression in these cells, it is evident from mutant
analysis that TcUbx is required for the proper
differentiation of these cells. In TcUbx mutants, the nuclei
of TcDll-expressing cells in the pleuropod never become
morphologically distinct as they do in the wild type. It is therefore
believed that TcUbx acts as a modifier rather than a
repressor of abdominal appendage development (Lewis 2000).
The dynamic relationship between TcUbx and TcDll
expression in the pleuropod and the effect of TcUbx
expression on the differentiation of TcDll-expressing cells
suggests that TcUbx acts to modify the way cells in the
anterior A1 compartment interpret signaling cues. In the absence of
TcUbx, cells respond to signaling cues as if they were no
longer pleuropodial. The failure of the appendage to invaginate and the
presence of the subterminal tarsal claw in TcUbx mutant
larvae support this view. In addition, the position of the subterminal
tarsal claw appears to correspond to the boundary of TcEn
expression and the cluster of TcDll-expressing cells in the
developing appendage of the embryo. This is interpreted as evidence that
these cells now respond to signaling cues as if they were leg, with the
distal-most tip, the tarsal claw in the leg, at the intersection of the
anterior-posterior boundary (Lewis 2000).
Differences in the manner in which TcUbx-expressing
cells respond to signaling cues could be because of TcUbx
acting directly on signaling pathway components or their targets.
Studies performed on Ubx control of wing vs. haltere development in
Drosophila have indeed shown that Ubx can act at multiple
levels of a genetic hierarchy. In the case of pleuropod
development, the levels of Ubx and/or the presence of Hox cofactors
are likely to be responsible for pleuropod-specific gene expression. The former explanation is favored since very high levels of TcUbx
are found in the pleuropod, compared with the levels found in other
regions of the embryo. The levels of TcUbx expression may be
important to outcompete other proteins expressed in these cells, such
as Antennapedia, which normally promotes leg patterning. In
addition, it has been shown that TcUbx levels are decreased
in TcEn-expressing cells of the thorax and abdomen in
wild-type embryos. Differences in TcUbx levels in these
compartments may also explain why, in Tcabd-A mutants, only
the cells in the anterior compartment of the abdominal segments are
able to differentiate as pleuropodial cells, whereas the
TcEn-expressing cells in the posterior compartment differentiate as leg cells. The possible effect of Ubx levels on
pleuropod patterning is consistent with data obtained in Drosophila on the effects of Ubx levels on patterning ps6 in
the embryo and bristles on the T2 leg in the adult (Lewis 2000).
Comparing the data obtained in this study on beetle abdominal
appendage development with that obtained from other holometabolous insects, it is suggested that abdominal limb repression
through direct Abd-A repression of Dll expression evolved at the latest
in the last common ancestor of the holometabola. This is the most
parsimonious interpretation, given that the repressive activity of Abd-A
is evident in species from all of the holometabolous orders examined.
However, one holometabolous insect species, the Lepidopteran
Manduca sexta, appears to be an exception. In the
developing abdominal prolegs in this species, Dll is expressed despite
the coexpression of Ubx/Abd-A. It is interesting to note that the
ability to express Dll in developing prolegs has arisen using at least
two different mechanisms within the Lepidoptera. In the butterfly
Precis coenia, activation of Dll expression in the abdomen
is correlated with regional repression of Ubx/Abd-A, whereas, in
the moth Manduca sexta, Dll expression occurs through a
different mechanism, presumably involving the escape of Dll from the
repressive effects of Abd-A. These data suggest that the release
of the repressive effect of Abd-A on abdominal limbs in higher
holometabolous insects occurred convergently through changes at
different levels of the limb regulatory hierarchy. Alternatively, it is
possible, although less likely, that the regional
repression/expression of Ubx/Abd-A has no causative effect on
proleg outgrowth, leaving open the possibility that the presence of
prolegs in these two Lepidopteran species is not convergent (Lewis 2000).
In higher holometabolous insect species, such as those found in the
orders Diptera and Lepidoptera, Ubx can act as a primary repressor of
Dll expression in the abdomen, whereas, in the more basal species such
as Tribolium, Ubx acts instead as a modifier of abdominal
limb development. Both the modifier role of Ubx in the anterior A1
compartment and the repressive role of Abd-A in the posterior
compartment are required for proper pleuropod development in
Tribolium. Because pleuropodia develop in the A1 segment of most insect orders, it is believed limb modification rather than limb
repression is a more ancient property of Ubx. Given the
conserved expression patterns of Ubx and Abd-A in the insect abdomen,
it will be of interest to examine how the functions of these genes in
regulating abdominal appendage development have changed during the
course of insect evolution (Lewis 2000).
A fascinating question in biology is how molecular changes in developmental
pathways led to macroevolutionary changes in morphology. Mutations in homeotic
(Hox) genes have long been suggested as potential causes of morphological
evolution, and there is abundant evidence that some changes in Hox expression
patterns correlate with transitions in animal axial pattern. A major
morphological transition in metazoans occurred about 400 million years ago, when
six-legged insects diverged from crustacean-like arthropod ancestors with
multiple limbs. In Drosophila melanogaster and other insects, the Ultrabithorax
(Ubx) and abdominal A (AbdA) Hox proteins are expressed largely in
the abdominal segments, where they can suppress thoracic leg development during
embryogenesis. In a branchiopod crustacean, Ubx/AbdA proteins are expressed in
both thorax and abdomen, including the limb primordia, but do not repress limbs.
It has been proposed that gain and loss of transcriptional
activation and repression functions in Hox proteins is a plausible mechanism to
diversify morphology during animal evolution. This study shows that naturally
selected alteration of the Ubx protein is linked to the evolutionary transition
to hexapod limb pattern (Ronshaugen, 2002).
It has been proposed that the hexapod body plan evolved
from crustacean-like ancestors in two phases: (1) mutations
restricted Ubx/AbdA expression to the proto-abdominal region; (2) mutations in Ubx/AbdA pathways resulted in suppression of thoracic-type limbs in the proto-abdomen. The mutations in this second 'limb suppression' phase could have
occurred in Ubx/AbdA coding sequences, in regulatory or coding
sequences for genes downstream of Ubx/AbdA, in regulatory or
coding sequences for Hox cofactors, or in a combination of these.
In embryos of Drosophila melanogaster, ectopic expression of the
Ubx protein in the thorax suppresses nearly all limb development;
thus the cofactors required for limb repression are present in both
thorax and abdomen. This ectopic expression assay can be used
to test whether a Ubx protein from crustaceans or other arthropods
can repress limb development. Since there is evidence that
branchiopod crustaceans and hexapod insects are sister groups,the Ubx protein from the crustacean Artemia franciscana was tested
for a limb-suppressing function in Drosophila embryos (Ronshaugen, 2002).
The Ubx protein sequence from Artemia was compared with Ubx
sequences from Drosophila, a hexapod mosquito (Anopheles gambiae)
and an onychophoran (A. kaputensis). There are large blocks of
amino-acid sequence present in Drosophila Ubx that are absent
from Artemia Ubx and vice versa. Within the DNA-binding
homeodomain, the Artemia Ubx protein has an identical
sequence when compared to the two other arthropod Ubx proteins except for a
single Ala-to-Ser change. All of the arthropod and the
onychophoran Ubx amino-acid sequences share six blocks of
homology, but there are an additional six blocks
of homology shared between the two hexapod Ubx sequences (Ronshaugen, 2002).
Transgenic Drosophila lines that ectopically produce
Artemia or Drosophila versions of Ubx were tested with or without
hemagglutinin antigen (HA) fused to their carboxy termini. The
HA epitope was used to show protein pattern and abundance of the
ectopically expressed proteins, and to distinguish them from
endogenous Ubx. No detectable differences were found between the
phenotypes induced by HA-tagged Drosophila or Artemia Ubx
proteins and those induced by wild-type proteins, and neither
Drosophila nor Artemia proteins nor their variants induced ectopic
transcription of the endogenous Ubx or AbdA genes (Ronshaugen, 2002).
When either Drosophila or Artemia Ubx-HA is expressed in the
embryonic thorax at levels equivalent to those of endogenous
Ubx in the abdomen, the ectopic proteins partially transformed thoracic denticle belts toward abdominal-like identities. The Drosophila and Artemia
proteins are also similar in suppressing the first thoracic (T1)
denticle 'beard', suppressing the formation of normal head structures,
and promoting the development of abdominal denticles in head segments. The Drosophila Ubx-HA protein
produces stronger versions of these phenotypes than does Artemia
Ubx-HA. However, it is clear that the Artemia Ubx protein
produced in fly embryos is functional, and capable of ectopically
inducing some aspects of abdominal identity in a manner similar to
Drosophila Ubx (Ronshaugen, 2002).
The Ubx homologs from these two species show striking
differences in their abilities to suppress thoracic embryonic limbs
(Keilin's organs): Drosophila Ubx-HA suppresses all of the limbs
whereas Artemia Ubx-HA suppresses only 15%. Distal-less (Dll) is an important limb-promoting gene in most or all
arthropods, and Drosophila Dll transcription is directly repressed
by the binding of Ubx protein to an upstream enhancer called
Dll304. As expected, Drosophila Ubx-HA strongly
represses Dll transcripts and Dll304 reporter transcripts in embryonic limb primordia; however, Artemia Ubx-HA has only a modest
repressive effect on Dll transcripts and Dll304 reporter levels. The inability of the Artemia protein to strongly repress Dll is not
due to the absence of a general repressive function, because
embryonic transcripts from the Antennapedia (Antp) P1 promoter
are completely repressed by Artemia Ubx-HA, similar to Drosophila
Ubx-HA (Ronshaugen, 2002).
In sum, full-length Artemia Ubx provides an 'abdominalizing'
function in the Drosophila embryonic epidermis, but has little
repressive effect on thoracic limb development in Drosophila
embryos. Further, the limb-suppressing difference between Drosophila
and Artemia Ubx is at least partly mediated by their different
abilities to transcriptionally repress the Dll gene. Although the distinction between the two proteins is referred to as a difference in limb-repression
function, it is not meant that this repression function is
solely directed to limb-promoting genes (Ronshaugen, 2002).
To map the Ubx limb-repression domain(s) that Drosophila
apparently possesses and Artemia lacks, a series of
hybrid and mutant proteins were constructed. The Ubx hybrid consisting of
the amino-terminal 356 amino acids of Drosophila and only the C-terminal
29 residues of Artemia has lost nearly all limb-repressing ability. Conversely, when the Drosophila Ubx C-terminal 26
residues replace the C terminus of Artemia Ubx (Art250Dros), the hybrid protein gains limb-repressing ability (Ronshaugen, 2002).
These results are interpreted to mean that the Drosophila Ubx
protein has a limb-repression domain in its C-terminal 26 amino
acids, whereas C-terminal sequences from Artemia are not sufficient
for limb repression. Another interpretation is that Artemia C-terminal
sequences may regulate (inhibit) a limb-repression
domain present elsewhere in both the Artemia and Drosophila
Ubx proteins. This latter function would be consistent with previous
studies indicating that the C terminus of Drosophila Ubx can
be deleted with little or no effect on its embryonic limb-repression
function (Ronshaugen, 2002).
To help distinguish between these possibilities, an
Artemia Ubx-HA mutant protein in which the C terminus was
deleted was tested. This mutant protein is a strong limb repressor. A variant of Drosophila Ubx was tested as was a variant of the
Art250Dros hybrid in which a notable block of conserved sequence
consisting of glutamines and alanines was
deleted. Both of the QA-deleted constructs still possess potent
embryonic limb-repression functions. This indicates that
the C terminus, and specifically the QA motif, are not required for
the full repressive activities of Drosophila Ubx or Artemia/Drosophila
Ubx hybrids, and that the C-terminal 29 amino acids of
Artemia Ubx are inhibiting a limb-repression domain elsewhere in
that protein (Ronshaugen, 2002).
In these assays, the C-terminal 45 amino acids of Drosophila Ubx
had a largely permissive role in Artemia/Drosophila chimaeric
proteins, failing to inhibit a limb-repression domain elsewhere in
Drosophila Ubx or Artemia Ubx. However, some positive repression
function may be encoded in the highly conserved QA motif, since the
repression of Keilin's organs is reduced by about 20% when this
motif is deleted. This is consistent with results
indicating that sequences that include the Drosophila
Ubx C-terminal QA domain are sufficient to provide a limb-repressive
function in an onychophoran/Drosophila hybrid protein
in embryos, and are also sufficient to supply transcriptional repressive
function in tissue-culture transfection assays (Ronshaugen, 2002).
Because the C-terminal regions of Ubx from a crustacean can
exert an inhibitory effect on the limb-repressive function of proteins
from the fruit fly or the brine shrimp, Ubx C-terminal
sequences were surveyed from a variety of insects and other arthropods
for potentially informative patterns of amino-acid conservation.
Notably, all of the Ubx proteins that are known or believed to
lack a limb-repressive function have multiple serine and/or threonine
amino acids as part of consensus phosphorylation sites in their
C-terminal domains. In Artemia Ubx, the most C-terminal
Ser is part of a casein kinase II (CKII) consensus phosphorylation
site, which after phosphorylation would generate additional CKII
and GSK-3 consensus sites. None of the insect Ubx
proteins have Ser or Thr residues in their C-terminal domains. This correlation is of great potential interest because Ser/Thr residues in the Antp Hox protein have been shown to modulate its function in embryos. Replacement of Ser or Thr by Ala residues in four CKII consensus sites of Antp results in a Hox protein that is a potent repressor of limb development and Dll transcription. One of these CKII sites, just downstream of the homeodomain, is highly conserved in Antp-like
Hox proteins in mammals. This suggests that the inability of the Ubx proteins from
Artemia and other multi-limbed arthropods to repress limbs might
reside in Ser/Thr phosphorylation sites that inhibit a covert limb-repression
domain in arthropod Ubx proteins (Ronshaugen, 2002).
To test this, mutant versions of Artemia Ubx were generated in
which C-terminal Ser/Thr residues were mutated to Ala. In the first
such mutant (Art Ubx S/T to A 1-5), the first five Ser and Thr
residues in the C-terminus are changed to Ala. This mutant Ubx has
little limb-repression function, similar to wild-type Artemia Ubx. However, the mutation of one additional Ser in a CKII
consensus site (Art Ubx S/T to A 1-5 and 7) results in a Ubx that
strongly represses embryonic limbs (Ronshaugen, 2002).
On the basis of these results, it is proposed that Ubx proteins in
some crustacean/insect ancestors uncovered a limb-repression
function by the mutation of C-terminal Ser/Thr phosphorylation
sites. Together with the restriction of Ubx expression to the
posterior trunk and expansion of a QA-rich domain, the loss of
these sites would have contributed to the evolution of the hexapod
body plan. The putative phosphorylation-mediated regulation of
transcriptional repression function in arthropod Ubx proteins may
occur by a similar mechanism to that recently described for the
Drosophila Even-skipped protein. In both cases, such a mechanism
would provide for the mediation by signal transduction of the
control of transcriptional activation and repression functions of
homeobox genes (Ronshaugen, 2002).
This is the first experimental evidence that
links naturally selected alterations of a specific protein sequence to a
major morphological transition in evolution. There are at least two
major reasons why the mutation of mutiple Ser/Thr residues that
inhibit a repression function might be advantageous from an
evolutionary aspect: (1) mutating the residues would give dominant
phenotypes, eliminating the need to fix two recessive
mutations in a morphologically evolving lineage; (2) the
successive removal of Ser/Thr residues might quantitatively influence
repression function and morphology, allowing viable micro-evolutionary
steps toward 'hopeful monsters' (Ronshaugen, 2002).
Retinoid signaling is required for the establishment of a zone of polarizing activity (ZPA) and for the expression of
Hoxb-8, a mediator of ZPA formation in the chick. Retinoid receptor antagonists applied to the presumptive wing region block
the formation of a ZPA. Although Hoxd-13 is still expressed after treatment with anti-retinoids, both sonic hedgehog and bmp-2 expression are absent in limb bud mesenchyme of wing regions treated with anti-retinoid activity. This suggests a direct relationship
between retinoid signaling and the establishment of the ZPA. The
Hox gene Hoxb-8 is a direct target of retinoid signaling since exogenously applied RA
rapidly induces this gene in the absence of protein synthesis; moreover, retinoid
receptor antagonists down-regulate Hoxb-8 expression. In addition in the
lateral plate mesoderm, the domains of Hoxb-8 expression and of polarizing activity are
coextensive. Taken together, these findings support the hypothesis that retinoids are
required for the establishment of a ZPA, and that retinoids act, at least in part, through
Hoxb-8, a gene associated with ZPA formation. Apparently there is only a brief temporal overlap between Hoxb-8 and shh expression. Furthermore, only a subset of Hoxb-8 expressing cells eventually express shh. It is thought that the RA-induced ectodermal factor is required for the formation of the ZPA. It is thought that shh is expressed in those Hoxb-8 positive cells that are in proximity to the AER and thus receive the FGF signal (Lu, H.-C., 1997).
Repertoires of grooming behaviors critical to survival are exhibited by most animal species, including humans. Mice with disruptions of Hoxb8 show, with 100% penetrance, excessive grooming leading to hair removal and lesions. Additionally, these mice excessively groom normal cagemates. No skin or PNS abnormalities have been detected in Hoxb8 mutants. These observations suggest that the excessive, pathological grooming exhibited by these mice results from CNS abnormalities. Consistent with this interpretation, Hoxb8 expression has been detected in regions of the adult mouse CNS previously implicated in the control of grooming. The aberrant behavior observed in Hoxb8 mutants is not unlike that of humans suffering from the obsessive compulsive- (OC)-spectrum disorder, trichotillomania. Interestingly, Hoxb8 is expressed in regions of the CNS known as the 'OCD-circuit' (Greer, 2002).
The excessive grooming behavior associated with the loss of Hoxb8 function in mice is of potential clinical interest because of its possible relation to the etiology of human trichotillomania. The mouse and human pathologies are remarkably similar. There is controversy as to whether trichotillomania should be classified as an obsessive-compulsive disorder (OCD) or as an impulse-control disorder. Based on the expression pattern of Hoxb8 in the CNS and the behavior analysis of affected animals, this work would favor classification of this disease as an OC-spectrum disorder. Functional imaging analysis has led to the description of an OCD circuit involving the basal ganglia and components of the neocortex. The basal ganglia are connected to the neocortex by parallel loops of cortico-basal ganglia connections. Different sets of cortico-basal ganglia loops are believed to be associated with specialized functions depending on the cortical areas involved in the circuit. The OCD circuit links the orbitofrontal and anterior cingulate cortex to multiple sites in the striatum and limbic system. Additionally, the anterior cingulate cortex is connected to the motor cortex allowing for selected output of action in response to perceived stimuli. These cortico-basal ganglia circuits are believed to form a neural system that is required for habit learning, routine performance of these habits, and the acquisition of stereotyped behaviors. Abnormal function of this circuitry has been linked to the abnormal repetitive behaviors exhibited by patients afflicted with OCD and OC-spectrum disorders. Expression of Hoxb8 has been detected in the principle components of the OCD circuit, which include the orbitofrontal cortex, the anterior cingulate cortex, the striatum, and the limbic system, as well as in other regions of the CNS associated with rodent grooming behavior. It is thus suggested that trichotillomania may arise from a misregulation of an innate autogrooming behavior resulting in excessive repetition of this behavior and its associated pathology (Greer, 2002).
The vertebrate Hox genes have been shown to confer regional identity along the anteroposterior axis
of the developing embryo, especially within the central nervous system (CNS) and the paraxial
mesoderm. The notochord has been shown to play vital roles in patterning adjacent tissues along both
the dorsoventral and mediolateral axes. However, the notochord's role in imparting anteroposterior
information to adjacent structures is less well understood, especially since the notochord shows no
morphological distinctions along the anteroposterior axis and is not generally described as a segmental
or compartmentalized structure. Four zebrafish hox genes (hoxb1, hoxb5, hoxc6
and hoxc8) are regionally expressed along the anteroposterior extent of the developing notochord.
Notochord expression for each gene is transient, but maintains a definite, gene-specific anterior limit
throughout its duration. The hox gene expression in the zebrafish notochord is spatially colinear with
those genes lying most 3' in the hox clusters having the most anterior limits. The expression patterns of
these hox cluster genes in the zebrafish are the most direct molecular evidence for a system of
anteroposterior regionalization of the notochord in any vertebrate studied to date (Prince, 1998).
Best known as epigenetic repressors of developmental Hox gene transcription, Polycomb complexes alter chromatin structure by means of post-translational modification of histone tails. Depending on the cellular context, Polycomb complexes of diverse composition and function exhibit cooperative interaction or hierarchical interdependency at target loci. The present study interrogated the genetic, biochemical and molecular interaction of BMI1 [Drosophila homologs Psc and Su(z)2] and EED (Drosophila homolog; Esc), pivotal constituents of heterologous Polycomb complexes, in the regulation of vertebral identity during mouse development. Despite a significant overlap in dosage-sensitive homeotic phenotypes and co-repression of a similar set of Hox genes, genetic analysis implicated eed and Bmi1 in parallel pathways, which converge at the level of Hox gene regulation. Whereas EED and BMI1 formed separate biochemical entities with EzH2 and Ring1B, respectively, in mid-gestation embryos, YY1 engaged in both Polycomb complexes. Strikingly, methylated lysine 27 of histone H3 (H3-K27), a mediator of Polycomb complex recruitment to target genes, stably associated with the EED complex during the maintenance phase of Hox gene repression. Juxtaposed EED and BMI1 complexes, along with YY1 and methylated H3-K27, were detected in upstream regulatory regions of Hoxc8 and Hoxa5. The combined data suggest a model wherein epigenetic and genetic elements cooperatively recruit and retain juxtaposed Polycomb complexes in mammalian Hox gene clusters toward co-regulation of vertebral identity (Kim, 2006).
At least two PcG complexes with diverse composition and function in chromatin remodeling have been identified in mammals. The Polycomb repressive complex 1 (PRC1) involves the paralogous PcG proteins BMI1/MEL18, M33/PC2, RAE28, and RING1A. Evidence for PRC1-mediated chromatin modification derived from ubiquitylation at lysine 119 of histone H2A (H2A-K119). A second PcG complex, PRC2, encompasses EED, the histone methyltransferase EZH2, the zinc finger protein SUZ12, the histone-binding proteins RBAP46/RBAP48, and the histone deacetylase HDAC1. Several EED isoforms, generated by alternate translation start site usage of eed mRNA, differentially engage in PRC2-related complexes (PRC2/3/4), targeting the histone methyltransferase activity of EZH2 to H3-K27 or H1-K26. PcG complexes bind to cis-acting Polycomb response elements (PREs), which encompass several hundred base pairs and are necessary and sufficient for PcG-mediated repression of target genes. Whereas the function of several PREs has been delineated in Drosophila, similar elements await characterization in mammals (Kim, 2006 and references therein).
An antibody raised against residues 123-140 of the EED amino terminus
precipitated three distinct isoforms of approximately 50 and 75 kDA from E12.5
trunk, representing three of the four EED isoforms previously reported in 293 cells. In
addition to EZH2 and YY1, dimethylated H3-K27 co-immunoprecipitated with EED. Immunoprecipitation identified three BMI1 isoforms of approximately 39-41 kDA. BMI1 was found in a complex with RING1B, but not dimethylated H3-K27. Similar to the EED complex,
the BMI1 complex also contained YY1. It should be emphasized that all (co-)immunoprecipitating bands were detected by at least two antibodies against different epitopes. Strikingly, while dimethylated H3-K27 engaged in the EED complex,
trimethylated H3-K27 did not appear to associate with either the EED or the
BMI1 complex. Importantly, reciprocal co-immunoprecipitation detected EED and
BMI1 in separate protein complexes (Kim, 2006).
Ectopic expression in mutant embryos revealed Hoxc8 and
Hoxa5 as downstream targets of EED and BMI1 function. ChIP detected EED and
BMI1 binding immediately upstream of the Hoxc8 transcribed region
near putative promoter elements. The binding sites could not be separated, indicating close proximity of the complexes. EED and BMI1 binding also
clustered within a small fragment 1.5 kb upstream of the Hoxc8
transcription start site, suggesting long-range juxtaposition of heterologous PcG
complexes. Similar to EED and BMI1, YY1 localized to both regions. In support
of YY1 binding to Hox regulatory regions, inspection of the mouse genome
sequence revealed clusters of putative YY1 binding sites in
both regions a and b, including TGTCCATTAG and
CCCCCATTCC (region a), as well as ACACCATGGC,
TTTCCATTAG and TCCCCATAAA (region b). CCAT represents
the core of the YY1 consensus binding site, while flanking sequences exhibited
significant tolerance for multiple nucleotides. EED,
BMI1 and YY1 also co-localized approximately 1.5 kb upstream of the
transcription start site of Hoxa5. In addition to PcG binding, ChIP detected trimethylated H3-K27 throughout the regulatory regions of Hoxc8 and Hoxa5. Furthermore, dimethylated H3-K27 localized to region b of Hoxc8 (Kim, 2006).
Spatial regulation of EED and BMI1 binding to Hox regulatory regions was
evident from ChIP analysis of dissected anterior and posterior regions of
E12.5 trunk. In agreement with transcriptional silencing of Hoxc8 and
Hoxa5, EED and BMI1 binding was detected upstream of these loci in
anterior regions of the trunk. By contrast, EED and BMI1 binding was absent from posterior regions of the trunk, where Hoxc8 and Hoxa5 are transcribed.
These findings implicate PcG complexes in Hox gene repression in anterior
regions of the AP axis (Kim, 2006).
The combined interpretation of the co-immunoprecipitation and ChiP results
indicates that trimethylated H3-K27 did not form a complex with EED or BMI1,
despite co-localization of the three proteins in Hox regulatory regions. By
contrast, co-immunoprecipitation demonstrated physical association of the EED
complex with dimethylated H3-K27. In aggregate, the results support a model in
which EED- and BMI1-containing chromatin remodeling complexes exist as
separate, but juxtaposed, biochemical entities at Hox target loci (Kim, 2006).
During lymphatic development, Prox1 plays central roles in the differentiation of blood vascular endothelial cells (BECs) into lymphatic endothelial cells (LECs), and subsequently in the maturation and maintenance of lymphatic vessels. However, the molecular mechanisms by which Prox1 elicits these functions remain to be elucidated. This study identified FoxC2 and angiopoietin-2 (Ang2), which play important roles in the maturation of lymphatic vessels, as novel targets of Prox1 in mouse embryonic-stem-cell-derived endothelial cells (MESECs). Furthermore, expression of HoxD8 was found to be significantly induced by Prox1 in MESECs, a finding confirmed in human umbilical vein endothelial cells (HUVECs) and human dermal LECs (HDLECs). In mouse embryos, HoxD8 expression was significantly higher in LECs than in BECs. In a model of inflammatory lymphangiogenesis, diameters of lymphatic vessels of the diaphragm were increased by adenovirally transduced HoxD8. It was also found that HoxD8 induces Ang2 expression in HDLECs and HUVECs. Moreover, HoxD8 induces Prox1 expression in HUVECs and knockdown of HoxD8 reduces this expression in HDLECs, suggesting that Prox1 expression in LECs is maintained by HoxD8. These findings indicate that transcriptional networks of Prox1 and HoxD8 play important roles in the maturation and maintenance of lymphatic vessels (Harada, 2009).
Retinoic acid (RA) signaling plays an important role in determining the anterior boundary of Hox gene expression in the neural tube during embryogenesis. In particular, RA signaling is implicated in a rostral expansion of the neural expression domain of 5' Hoxb genes (Hoxb9-Hoxb5) in mice. However, underlying mechanisms for this gene regulation have remained elusive due to the lack of RA responsive element (RARE) in the 5' half of the HoxB cluster. To identify cis-regulatory elements required for the rostral expansion, a recombineering technology was developed to serially label multiple genes with different reporters in a single bacterial artificial chromosome (BAC) vector containing the mouse HoxB cluster. This allowed simultaneous monitoring of the expression of multiple genes. In contrast to plasmid-based reporters, transgenic BAC reporters faithfully recapitulated endogenous gene expression patterns of the Hoxb genes including the rostral expansion. Combined inactivation of two RAREs, DE-RARE and ENE-RARE, in the BAC completely abolished the rostral expansion of the 5' Hoxb genes. Knock-out of endogenous DE-RARE lead to significantly reduced expression of multiple Hoxb genes and attenuated Hox gene response to exogenous RA treatment in utero. Regulatory potential of DE-RARE was further demonstrated by its ability to anteriorize 5' Hoxa gene expression in the neural tube when inserted into a HoxA BAC reporter. These data demonstrate that multiple RAREs cooperate to remotely regulate 5' Hoxb genes during CNS development, providing a new insight into the mechanisms for gene regulation within the Hox clusters (Ahn, 2014).
Ultrabithorax (Ubx) is a member of the Hox gene group involved in cell fate decisions, cell proliferation and organ identity. Its function has been extensively researched in Drosophila melanogaster but little is known about it in Lepidoptera. To uncover the function of Ubx in the development of lepidopterans, this study constructed the Ubx overexpression (Ubx(OE)) strain based on the Nistari strain of Bombyx mori. The Ubx(OE) strain showed a small body size, transparent intersegmental membrane and abnormal posterior silk gland (PSG). The current study focused on the effect of Ubx overexpression on the posterior silk gland. As the major protein product of PSG, the mRNA expression of fibroin heavy chain (Fib-H) and fibroin light chain (Fib-L) was upregulated three fold in Ubx(OE), but the protein expression of Fib-H and Fib-L was not significantly different. It is speculated that the overexpression of Ubx downregulated the expression of Myc and further caused abnormal synthesis of the spliceosome and ribosome. Abnormalities of the spliceosome and ribosome affected the synthesis of protein in the PSG and changed its morphology (Li, 2023).
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Ultrabithorax:
Biological Overview
| Transcriptional Regulation
| Targets of activity
| Protein Interactions
| Posttranscriptional regulation
| Developmental Biology
| Effects of Mutation
| References
Society for Developmental Biology's Web server.
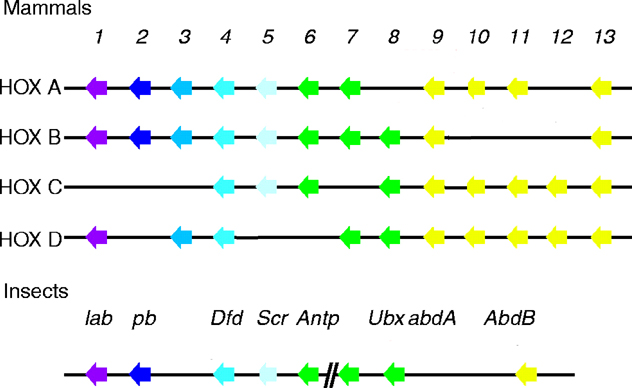{kind=link}