|
Gene name - Deformed Synonyms - Cytological map position - 84A4-5 Function - transcription factor Keywords - homeotic, Antennapedia complex |
Symbol - Dfd FlyBase ID:FBgn0000439 Genetic map position - 3-47,5 Classification - homeodomain - Antp class Cellular location - nuclear |
| Recent literature | Anais Tiberghien, M., Lebreton, G., Cribbs, D.,
Benassayag, C. and Suzanne, M. (2015). The
Hox gene Dfd controls organogenesis by shaping territorial border
through regulation of basal DE-Cadherin distribution. Dev Biol
[Epub ahead of print]. PubMed ID: 26206615 Summary: Hox genes are highly conserved selector genes controlling tissue identity and organogenesis. Recent work indicates that Hox genes also control cell segregation and segmental boundary in various species, however the underlying cellular mechanisms involved in this function are poorly understood. In Drosophila, the Hox gene Deformed (Dfd) is required for specification and organogenesis of the adult Maxillary (Mx) palp. This study demonstrates that differential Dfd expression controls Mx morphogenesis through the formation of a physical boundary separating the Mx field and the Peripodial Epithelium (PE). This boundary was shown to rely on DE-cadherin (DE-cad) basal accumulation in Mx cells controlled by differential Dfd expression. Indeed, Dfd controls boundary formation through cell autonomous basal redistribution of DE-cad which leads to subsequent fold at the Dfd expression border. Finally, the loss of Mx DE-cad basal accumulation and hence of Mx-PE folding is sufficient to prevent Mx organogenesis thus revealing the crucial role of boundaries in organ differentiation. Altogether, these results reveal that Hox coordination of tissue morphogenesis relies on boundary fold formation through the modulation of DE-cad positioning. |
Kumar, R., Chotaliya, M., Vuppala, S., Auradkar, A., Palasamudrum, K. and Joshi, R. (2015). Role of Homothorax in region specific regulation of Deformed in embryonic neuroblasts. Mech Dev [Epub ahead of print]. PubMed ID: 26409112
Summary: The expression and regulation of Hox genes in developing central nervous system (CNS) lack important details like specific cell types where Hox genes are expressed and the transcriptional regulatory players involved in these cells. This study has investigated the expression and regulation of Drosophila Hox gene Deformed (Dfd) in specific cell types of embryonic CNS. Using Dfd neural autoregulatory enhancer, it was found that Dfd autoregulates itself in cells of mandibular neuromere. The role was investigated of a Hox cofactor Homothorax (Hth) for its role in regulating Dfd expression in CNS. Hth was found to exhibit a region specific role in controlling the expression of Dfd, but has no direct role in mandibular Dfd neural autoregulatory circuit. These results also suggest that homeodomain of Hth is not required for regulating Dfd expression in embryonic CNS. |
Urbach, R., Jussen, D. and Technau, G.M. (2016). Gene expression profiles uncover individual identities of gnathal neuroblasts and serial homologies in the embryonic CNS of Drosophila. Development 143: 1290-1301. PubMed ID: 27095493 Summary: This study constructed comprehensive neuroblast maps for the three gnathal head segments. Based on the spatiotemporal pattern of neuroblast formation and the expression profiles of 46 marker genes (41 transcription factors), each neuroblast can be uniquely identified. Compared with the thoracic ground state, neuroblast numbers are progressively reduced in labial, maxillary and mandibular segments due to smaller sizes of neuroectodermal anlagen and, partially, to suppression of neuroblast formation and induction of programmed cell death by the Hox gene Deformed. Neuroblast patterns are further influenced by segmental modifications in dorsoventral and proneural gene expression. With the previously published neuroblast maps and those presented here for the gnathal region, all neuroectodermal neuroblasts building the CNS of the fly (ventral nerve cord and brain, except optic lobes) are now individually identified (in total 2×567 neuroblasts). This allows, for the first time, a comparison of the characteristics of segmental populations of stem cells and to screen for serially homologous neuroblasts throughout the CNS. It was found that approximately half of the deutocerebral and all of the tritocerebral (posterior brain) and gnathal neuroblasts, but none of the protocerebral (anterior brain) neuroblasts, display serial homology to neuroblasts in thoracic/abdominal neuromeres. Modifications in the molecular signature of serially homologous neuroblasts are likely to determine the segment-specific characteristics of their lineages. |
Maiti, S., Acharya, B., Boorla, V. S., Manna, B., Ghosh, A. and De, S. (2019). Dynamic studies on intrinsically disordered regions of two paralogous transcription factors reveal rigid segments with important biological functions. J Mol Biol. PubMed ID: 30802457
Summary: Long stretches of intrinsically disordered regions (IDRs) are abundantly present in eukaryotic transcription factors. Although their biological significance is well appreciated, the underlying structural and dynamic mechanisms of their function are still not clear. Using solution NMR spectroscopy, the structural and dynamic features of two paralogous HOX transcription factors, SCR and DFD, from Drosophila were studied. Both proteins have a conserved DNA-binding homeodomain and a long stretch of functionally important IDR. Using NMR dynamics, flexibility of each residue in these proteins was determined. The flexibility of the residues in the disordered region is not uniform. In both proteins, the IDRs have short stretches of consecutive residues with relatively less flexibility, that is, higher rigidity. One such rigid segment is specifically recognized by another co-transcription factor, thus highlighting the importance of these rigid segments in IDR-mediated protein-protein interactions. Using molecular dynamics simulation, it was further showm that the rigid segments sample less conformations compared to the rest of the residues in the disordered region. The restrained conformational sampling of these rigid residues should lower the loss in conformational entropy during their interactions with binding partners resulting in sequence specific binding. This work provides experimental evidence of a "rigid-segment" model of IDRs, where functionally important rigid segments are connected by highly flexible linkers. Furthermore, a comparative study of IDRs in paralogous proteins reveals that in spite of low-sequence conservation, the rigid and flexible segments are sequentially maintained to preserve related functions and regulations of these proteins. |
Ponrathnam, T., Saini, R., Banu, S. and Mishra, R. K. (2021). Drosophila Hox genes induce melanized pseudo-tumors when misexpressed in hemocytes. Sci Rep 11(1): 1838. PubMed ID: 33469139
Summary: Hox genes are early determinants of cell identity along the anterior-posterior body axis across bilaterians. Several late non-homeotic functions of Hox genes have emerged in a variety of processes involved in organogenesis in several organisms, including mammals. Several studies have reported the misexpression of Hox genes in a variety of malignancies including acute myeloid leukemia. The Hox genes Dfd, Ubx, abd-A and Abd-B were overexpressed via the UAS-Gal4 system using Cg-Gal4, Lsp2-Gal4, He-Gal4 and HmlD3-Gal4 as specific drivers. Genetic interaction was tested by bringing overexpression lines in heterozygous mutant backgrounds of Polycomb and trithorax group factors. Larvae were visually scored for melanized bodies. Circulating hemocytes were quantified and tested for differentiation. Pupal lethality was assessed. Expression of Dfd, Ubx and abd-A, but not Abd-B in the hematopoietic compartment of Drosophila led to the appearance of circulating melanized bodies, an increase in cell number, cell-autonomous proliferation, and differentiation of hemocytes. Pupal lethality and melanized pseudo-tumors were suppressed in Psc1 and esc2 backgrounds while polycomb group member mutations Pc1 and Su(z)123 and trithorax group member mutation TrlR85 enhanced the phenotype. Dfd, Ubx and abd-A are leukemogenic. Mutations in Polycomb and trithorax group members modulate the leukemogenic phenotype. This RNAseq of Cg-Gal4 > UAS-abd-A hemocytes may contain genes important to Hox gene induced leukemias. |
Villedieu, A., Alpar, L., Gaugue, I., Joudat, A., Graner, F., Bosveld, F. and Bellaiche, Y. (2023). Homeotic compartment curvature and tension control spatiotemporal folding dynamics. Nat Commun 14(1): 594. PubMed ID: 36737611
Summary: Shape is a conspicuous and fundamental property of biological systems entailing the function of organs and tissues. While much emphasis has been put on how tissue tension and mechanical properties drive shape changes, whether and how a given tissue geometry influences subsequent morphogenesis remains poorly characterized. This study explored how curvature, a key descriptor of tissue geometry, impinges on the dynamics of epithelial tissue invagination. The morphogenesis of the fold separating the adult Drosophila head and thorax segments is driven by the invagination of the Deformed (Dfd) homeotic compartment. Dfd controls invagination by modulating actomyosin organization and in-plane epithelial tension via the Tollo and Dystroglycan receptors. By experimentally introducing curvature heterogeneity within the homeotic compartment, this study established that a curved tissue geometry converts the Dfd-dependent in-plane tension into an inward force driving folding. Accordingly, the interplay between in-plane tension and tissue curvature quantitatively explains the spatiotemporal folding dynamics. Collectively, this work highlights how genetic patterning and tissue geometry provide a simple design principle driving folding morphogenesis during development. |
Coming face to face with a highly magnified fruit fly's head can easily be confused with an extraterrestrial confrontation. Huge compound eyes protrude on either side of this alien head [Images]. Between them, a pair of swollen bulbous antenna lift and arch, covered with a multitude of short spines, as well as antler-like aristae and a sensory pouch, the sacculus. These non-human organs sit in a broad area on the frons, or fly's forehead. Lower down the "face," one finds the clypeus, analogous to a nose only because of anthropocentric attempts to cognitively absorb the strangeness of the fly. The clypeus neither breaths nor smells, nor serves as a perch for glasses. It is part of a segment that includes the labrum, a short tube used for sucking food that emerges to hang down from the bottom of the clypeus. On either side of the labrum are two small bulbs, the maxilla or maxillary palps. These organs are greatly reduced in the fruit fly, when compared to those of other insects. Below the maxilla lies an incredible structure, the labella or labial palps. These large, furrowed bulbs lie at the end of the proboscis, the fly's main feeding organ. The palps serve to gather liquid food, which moves up a series of collecting channels into the straw-like labrum, connecting to the muscular esophagus (Ferris, 1950).
Deformed (Dfd) is responsible for the normal development of the maxillary segment, consisting of those tiny bulbs in front of the labium. Deformed has other developmental responsibilities, as shown by three lethal effects associated with Deformed mutation that show up during metamorphosis: 1) failure to separate the subesophageal ganglion (SEG) from the thoracic ganglion; 2) structural and functional abnormalities of the proboscis and maxillary palps, and 3) failure of the adult salivary glands to extend into the thorax. The SEG is considered a ventral portion of the brain and also as part of the central nervous system (Restifo, 1994).
Broad-Complex is another transcription factor acting late in development. Deformed and Broad-Complex interact in the formation of these adult structures during the pupal stage. Deformed defects are in fact identical to those of Broad-Complex, a locus regulating metamorphosis. The homeotic gene labial is likewise important in the transition from pupal to adult structures (Restifo, 1994). This underscores the significance of homeotic genes in the final stages of pupal development, the transition from pupa to adult. Current interests in the analysis of pupal development and the intricacies of the final stages of wing, leg and head development have been largely side-tracked in this era of ground-breaking molecular genetics, but resurgent interest is only a matter of time.
The reduced size of the maxillary segment in adults, due no doubt to the Drosophila specialization of the proboscis for food gathering, is not as apparent in the larval and pupal stages. Important mouth parts are missing at these stages (Regulski, 1987), indicating that evolutionary change is much more conservative in the pre-adult morphology. The role of homeotics in neural differentiation has not yet been adequately explored. Dfd will very likely have a role in subesophageal ganglion differentiation and in the ventral nervous system, topics that have not yet received sufficient attention (Lou, 1995).
Precise gene expression is a fundamental aspect of organismal function and depends on the combinatorial interplay of transcription factors (TFs) with cis-regulatory DNA elements. While much is known about TF function in general, understanding of their cell type-specific activities is still poor. To address how widely expressed transcriptional regulators modulate downstream gene activity with high cellular specificity, binding regions were identified for the Hox TF Deformed (Dfd) in the Drosophila genome. This analysis of architectural features within Hox cis-regulatory response elements (HREs) shows that HRE structure is essential for cell type-specific gene expression. It was also found that Dfd and Ultrabithorax (Ubx), another Hox TF specifying different morphological traits, interact with non-overlapping regions in vivo, despite their similar DNA binding preferences. While Dfd and Ubx HREs exhibit comparable design principles, their motif compositions and motif-pair associations are distinct, explaining the highly selective interaction of these Hox proteins with the regulatory environment. Thus, these results uncover the regulatory code imprinted in Hox enhancers and elucidate the mechanisms underlying functional specificity of TFs in vivo (Sorge, 2012).
In order to quantitatively identify genomic regions bound by the Hox TF Dfd in Drosophila, two complementing approaches were employed: ChIP-seq, which has been successfully applied previously to identify stage- and tissue-specific enhancer activities, and computational detection of clusters of TF binding sequences, which allows the identification of cis-regulatory modules irrespective of temporal and spatial context. To generate genome-wide maps of Dfd binding in vivo, ChIP was performed using stage 10-12 Drosophila embryos and a Dfd-specific antibod. Stage-independent in silico Dfd-specific Hox response elements (HREs) were identified by searching for clusters of conserved Dfd binding motifs, as defined by a position weight matrix (PWM), in the non-coding regions of the genomes of 12 distinct Drosophila species. By applying both approaches, 4526 genomic regions containing clusters of Dfd binding sites and 1079 Dfd ChIP-seq enrichment peaks were identified, including two out of the three well-characterized Dfd-HREs, namely rpr-4S3 and Dfd-EAE. To study the regulatory capacity of novel in silico and ChIP-seq detected HREs, cell culture-based enhancer assays were performed for 11 randomly selected HREs, and it was found that reporter expression driven by the identified genomic regions was in all cases dependent on Dfd binding. In vivo activity was tested of 21 arbitrarily selected enhancers in transgenic reporter lines, revealing that 7 out of 11 ChIP-identified and 5 out of 10 in silico-predicted Dfd-HREs recapitulate the spatio-temporal expression of adjacent genes). Most importantly, it was possible to demonstrate Dfd-dependent regulation of both transgenic reporter expression and endogenous gene expression, suggesting that they are bona fide direct Dfd target genes. Thus, the identified Dfd-HREs represent a data set of biologically relevant regulatory regions and an excellent resource to unravel sequence features within Hox responsive enhancers that might be essential for the highly selective Hox target gene regulation (Sorge, 2012).
Transcriptional regulation in many cases relies on the assembly of regulatory protein complexes mediated by closely spaced TF binding sites within a cis-regulatory module and previous studies have shown that Hox proteins employ this mechanism to control target gene activity in small subsets of cells. The novel HREs were systematically scanned for TF binding motifs appearing in close proximity to Dfd binding sites. Using a statistical test for pair-wise distance distributions, w11 overrepresented DNA motifs for known TFs were found adjoining to Dfd binding sites with 5 of the motifs occurring in both the ChIP-seq and in silico-identified Dfd-HREs. When the expression patterns of six of these transcriptional regulators known to bind to the 11 motifs that were identified were examined, colocalization with Dfd was found in different sub-populations of cells in all cases. Colocalization was already known for two TFs, whose binding sites were coupled to Dfd motifs, including Extradenticle (Exd) , which is known to cooperatively bind with Hox proteins to DNA and thereby increase Hox DNA-binding selectivity. It was next asked whether the short-distance arrangements in Dfd-HREs are of biological relevance and translated into the regulation of similar classes of target genes. To this end, the overrepresentation was statistically tested of expression and biological terms of genes associated with HREs harbouring specific combinations of Dfd and close-by motifs. This analysis revealed that only those Dfd-HREs with short distance intervals between the Dfd and adjacent motifs were coupled to similar gene classes, while random distance intervals did not show any correlation. Strikingly, genes associated with specific short-distance HREs had similar expression and functional annotations as the TFs interacting with the Hox adjoining motifs, suggesting that time and place of Hox action is dictated by spatio-temporally restricted co-regulators. Support for this hypothesis stems from the observation that one of the close-distance partners, Optix, regulates similar processes as Dfd, since Dfd and Optix mutants displayed comparable morphological defects in the head region, such as the absence of mouth hooks, a maxillary segment-derived structure known to be specified by Dfd. In addition, one of the genes associated with a Dfd-Optix HRE, the known Dfd target gene reaper (rpr), is expressed in the ventral epidermis primordium as predicted by its HRE architecture, and regulated by Dfd and Optix in ventral-maxillary cells, which also express these factors. A cell-culture assay using the well-established Dfd responsive module responsible for rpr expression in a few anterior-maxillary cells, the rpr-4S3 Dfd-HRE, with wild-type or mutated Dfd binding sites or reduction of Dfd levels by RNAi confirmed the requirement for simultaneous activity of Dfd and Optix on the rpr-4S3 Dfd-HRE for strong reporter gene induction. Optix binding to the rpr-4S3 Dfd-HRE was additionally confirmed by electrophoretic mobility shift assay (EMSA) experiments. Furthermore, transgenic reporter expression induced by the rpr-4S3 Dfd-HRE was lost in Optix mutant embryos or when the Optix binding sites were mutated. These results demonstrate that Optix, one of the newly identified factors, is a Dfd co-regulator required for proper regulation of the important Hox target gene rpr (Sorge, 2012).
Whether The precise spacing between Hox and adjacent binding sites plays a role for enhancer activity was explored. The rpr-4S3 Dfd HRE, which induces gene expression in a few anterior-maxillary cells, has previously been shown to be under the control of Dfd and Glial cells missing (Gcm), a Dfd co-regulator also identified in this study. Dfd and Gcm as well as Optix binding sites within the rpr-4S3 HRE are directly adjacent to each other, thus a 5- and 10-bp spacer was introduced to interfere with potential interactions of the proteins on the enhancer. In all cases, reporter gene expression was strongly reduced or completely abolished, showing that the close-distance arrangements between Dfd and Gcm as well as Dfd and Optix are required for the in vivo activity of the rpr-4S3 enhancer (Sorge, 2012).
While the results regarding the close-distance arrangement of Dfd and Gcm binding sites suggested the formation of a Dfd-Gcm protein complex, like in the case of Dfd and Exd, only independent binding of the two proteins to the rpr-4S3 enhancer was observed in EMSA experiments , supporting the idea of Hox proteins collaborating with other TFs on target HREs in the absence of physical contact. It has been shown before that Hox proteins together with other TFs that bind in the immediate vicinity recruit non-DNA binding cofactors to HREs. To test if such factors could interact with Dfd and the newly identified short distance binding TFs, the modENCODE data set was scanned and it was found that dCBP/Nej, a member of the CBP/p300 family of transcriptional co-activators bearing acetyltransferase activity, binds to the rpr-4S3 enhancer in vivo. As nej has been previously reported to genetically interact with Dfd, its function was examined in Dfd/Gcm-mediated transcriptional activation. Both factors, Dfd and Gcm, are required for transcriptional activation, since expression of Gcm in Drosophila D.Mel-2 cells, which have basal levels of Dfd activity, resulted in strong induction of reporter gene expression, while abolishing Dfd binding to the rpr-4S3 HRE by mutating all Dfd binding sites or by reducing Dfd protein levels in D.Mel-2 cells using RNAi, strongly reduced reporter gene expression in the presence of Gcm. Strikingly, Dfd- and Gcm-mediated reporter gene expression was strongly reduced in nej dsRNA-treated cells, whereas inhibition of protein deacetylation by Trichostatin A (TSA0) restored reporter gene expression. Consistently, rpr expression was abolished in nej mutant embryos. These results demonstrate that dCBP/Nej-mediated protein acetylation/histone modification is important for the combined activity of Dfd and Gcm on the rpr-4S3 HRE. While it was not possible to demonstrate that nej physically interacts with Dfd protein using various assays, EMSA experiments show that nej interacts with Gcm. Furthermore, acetylation of transiently transfected Gcm was detected in cultured Drosophila cells. Acetylation of Gcm is dependent on Nej, as it was reduced upon RNAi-mediated downregulation of nej. These results are consistent with published work demonstrating that in human cells CBP interacts with Gcma, resulting in its acetylation and stimulation of its transcriptional activity. Since about 10% of all Dfd and nej in vivo genomic binding events during embryonic stages 10-12 overlap, the functional interaction of Dfd and nej observed at the rpr locus does not seem an exception. This finding suggests that the interaction of co-activators (and co-repressors) with Hox proteins and close distance binding TFs on enhancer modules could be a commonly used mechanism to achieve highly specific spatio-temporal control of target gene activity. In this scenario, Hox proteins would control downstream genes by direct transcriptional and/or epigenetic regulation depending on HRE composition and thus cofactor identity and recruitment (Sorge, 2012)
Despite very similar DNA binding behaviour in vitro, Hox proteins regulate distinct morphological features along the anterior-posterior body axis in animal systems. To elucidate the mechanistic basis for the differences in their regulatory properties, Dfd-HREs identified in this study were compared to genomic regions bound by the Hox TF Ultrabithorax (Ubx) at identical developmental stages, as identified by the modENCODE consortium. Searching for overrepresented DNA motifs in both enriched ChIP regions, it was found that Dfd and Ubx bind to identical DNA sequences in vivo, reminiscent to in vitro systems. However, individual binding motifs seem to play only a minor role for Hox binding site selection in vivo, since this analysis revealed that Dfd and Ubx exclusively interact with non-overlapping genomic regions in embryonic stages 9-12. Consequently, Dfd- and Ubx-HREs were found to be associated with distinct classes of genes, revealing that genes with roles in the epidermis are primarily under the control of Dfd at the analysed embryonic stages while genes with mesoderm-related functions are predominantly regulated by Ubx. Consistently, it was found that the expression of tartan (trn), one of the genes associated with a Dfd-HRE, is regulated exclusively by Dfd, but not by Ubx, in epidermal cells, while parcas (pcs), one of the genes linked to a Ubx-HRE is under the selective control of Ubx in mesodermal cells. Furthermore, only Ubx-HREs were found to substantially overlap with cis-regulatory elements stage specifically bound by the mesoderm-specifying TFs Myocyte enhancer factor 2 (Mef2), Twist and Tinman. In contrast, the common ability of both Dfd and Ubx to regulate genes involved in nervous system development was underlined by comparable representations of binding motifs for the neuronal-specifying TFs Asense, Deadpan and Snail in Dfd- and Ubx-HREs (Sorge, 2012).
Strikingly, the basic design principles of Dfd- and Ubx-HREs were found to be similar: like in Dfd-HREs, six binding motifs for known TFs were located adjacent to Ubx binding sites and colocalization studies showed that they are expressed in subsets of Ubx-positive cells. Again, Ubx binding sites and motifs for potential co-regulators occurred most frequently in specific short intervals and only those Ubx-HREs with the preferred distance were associated with specific gene classes. This analysis also revealed that four of the six short-distance motifs were specific for Ubx-HREs, which is consistent with the data showing that Hox proteins interact with different and spatially restricted co-regulators to control target gene expression in selected cells. Importantly, in the cases of the close-distance motifs detected in both HREs, namely the binding sites for the TFs Ladybird early (Lbe) and Cut (Ct), the associated target genes were also expressed in non-overlapping tissues. This raised the question of how different Hox proteins can act on distinct target genes, even when their target HREs exhibit similar binding site compositions including short-distance arrangements. Since Lbe is active in both mesodermal and epidermal cells, one Dfd-Lbe and one Ubx-Lbe HRE was exemplarily analysed, and binding of Lbe protein was confirmed to both HREs by EMSAs. As predicted by the presence of Lbe binding sequences. Complex formation between the Hox protein and Lbe was observed in the case of Ubx and Lbe while Dfd and Lbe interact independently with the Dfd-Lbe HRE, indicating that the two Hox proteins employ different mechanisms for binding to the selected HREs. Lbe interaction with the Dfd-Lbe and Ubx-Lbe HREs is essential for in vivo activity, since in both cases ectopic reporter gene expression was observed when Lbe binding sites were mutated. Even more important, reporter gene expression was specifically changed only in segments in which either Dfd or Ubx is active, meaning in the case of the Dfd-Lbe HRE in maxillary cells and in the case of the Ubx-Lbe HRE in abdominal segments A1-A7. Taken together, these results demonstrate that the combined activity of Lbe and the Hox proteins Dfd or Ubx on selected HREs is critical for the precise spatiotemporal and segment-specific control of HRE activity. It was next asked whether additional (DNA- and non-DNA-binding) factors contribute to the predicted cell type-specific expression of the Dfd-Lbe and Ubx-Lbe HREs. Using the Drosophila Interactions Database (DroID; Murali, 2011) and published genome-wide DNA binding studies a search was carried out for unique Dfd-lbe and Ubx-lbe interactors. It was discovered that almost 20% of all Ubx-Lbe HREs but none of the Dfd-Lbe HREs were found to interact with the mesoderm-specifying factor Mef2 in vivo, while H3K9me3 histone marks, which are mediated by one of the unique Dfd-lbe interactors, Enhancer of zeste E(z), are enriched only within Dfd-Lbe HREs. Interestingly, E(z) modifies chromatin also by trimethylating H3K27 residues, a histone mark highly enriched at the genomic region spanning the ChIP-detected Dfd-Lbe HRE. Consistent with the repressive function of this histone modification, loss of Lbe binding to the Dfd-Lbe HRE results in ectopic reporter gene expression, suggesting that Lbe (and Dfd) recruits E(z) to the Dfd-Lbe HRE for cell type-specific target gene repression (Sorge, 2012).
Taken together, these results demonstrate that Hox proteins interact with different regulatory proteins on HREs, which allows them to differentially regulate their target genes despite their similar DNA binding properties. The fact that these interactions occur only in a few cells for a short period of time is very likely one of the major reasons why the identification of factors conferring regulatory precision and specificity to Hox function has met with little success so far (Sorge, 2012).
labial is the most proximal member of the homeobox genes of the Antennapedia Complex. It is followed in the distal direction by proboscopedia, zerknüllt, bicoid, Deformed, Sex combs reduced, fushi tarazu and Antennapedia (Kaufman, 1990). Genomic length - 11 kb
cDNA clone length - 1758 bp
Bases in 5' UTR - 491
Exons - five
Bases in 3' UTR - 491
The DFD protein has two N-terminal glycine rich regions with a histidine rich region between them. There is a central acidic domain, a homeodomain, and a C-terminal polyglutamine and polyasparagine domain (Regulksi, 1987).
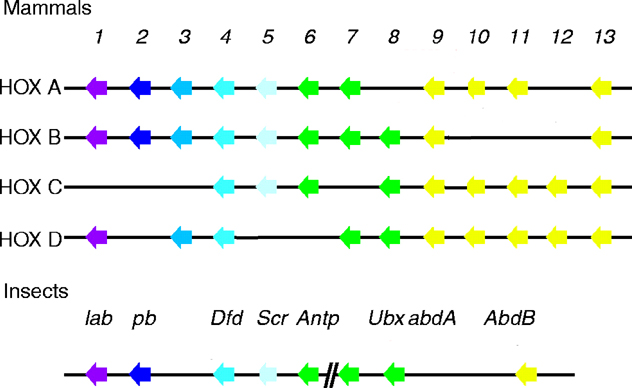
See four paralogous Hox clusters of mammals for homologies of Deformed with mammalian Hox proteins.
date revised: 25 September 2023
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
{kind=link}