|
Gene name - proboscipedia Synonyms - Cytological map position - 84A4-5 Function - transcription factor Keywords - homeotic, Antennapedia complex |
Symbol - pb FlyBase ID:FBgn0051481 Genetic map position - 3-47.5 Classification - homeodomain - Antp class Cellular location - nuclear |
| Recent literature | Percival-Smith, A. (2016). Non-specificity of transcription factor function in Drosophila melanogaster. Dev Genes Evol [Epub ahead of print]. PubMed ID: 27848019
Summary: A major problem in developmental genetics is how HOX transcription factors, like Proboscipedia (PB) and Ultrabithorax (UBX), regulate distinct programs of gene expression to result in a proboscis versus a haltere, respectively, when the DNA-binding homeodomain (HD) of HOX transcription factors recognizes similar DNA-binding sequences. Indeed, the lack of DNA-binding specificity is a problem for all transcription factors (TFs), as the DNA-binding domains generally recognize small targets of five to six bases in length. Although not the initial intent of the study, extensive non-specificity of TF function were found. Multiple TFs including HOX and HD-containing and non-HD-containing TFs induced both wingless and eyeless phenotypes. The TFs Labial (LAB), Deformed (DFD), Fushi tarazu (FTZ), and Squeeze (SQZ) induced ectopic larval thoracic (T) 1 beard formation in T2 and T3. The TF Doublesex male (DSXM) rescued the reduced maxillary palp pb phenotype. These examples of non-specificity of TF function across classes of TFs, combined with previous observations, compromise the implicit, initial assumption often made that an intrinsic mechanism of TF specificity is important for function. Interestingly, the functional complementation of the pb phenotype may suggest a larger role for regulation of expression of TFs in restriction of function as opposed to an intrinsic specificity of TF function. These observations have major ramifications for analysis of functional conservation in evolution and development. |
Percival-Smith, A., Ponce, G. and Pelling, J. J. (2017). The non-cell autonomous requirement of Proboscipedia for growth and differentiation of the distal maxillary palp during metamorphosis of Drosophila melanogaster. Genet Res Int 2017: 2624170. PubMed ID: 28357140
Summary: The Drosophila maxillary palpus that develops during metamorphosis is composed of two elements: the proximal maxillary socket and distal maxillary palp. The HOX protein, Proboscipedia (PB), was required for development of the proximal maxillary socket and distal maxillary palp. For growth and differentiation of the distal maxillary palp, PB was required in the cells of, or close to, the maxillary socket, as well as the cells of the distal maxillary palp. Therefore, PB is required in cells outside the distal maxillary palp for the expression, by some mechanism, of a growth factor or factors that promote the growth of the distal maxillary palp. Both wingless (wg) and hedgehog (hh) genes were expressed in cells outside the distal maxillary palp in the lancinia and maxillary socket, respectively. Both wg and hh were required for distal maxillary palp growth, and hh was required noncell autonomously for distal maxillary palp growth. However, expression of wg-GAL4 and hh-GAL4 during maxillary palp differentiation did not require PB, ruling out a direct role for PB in the regulation of transcription of these growth factors. |
Miller, S. W. and Posakony, J. W. (2020). Disparate expression specificities coded by a shared Hox-C enhancer. Elife 9. PubMed ID: 32342858
Summary: Can a single regulatory sequence be shared by two genes undergoing functional divergence? This study describes a single promiscuous enhancer within the Drosophila Antennapedia Complex, EO053, that directs aspects of the expression of two adjacent genes, pb (a Hox2 ortholog) and zen2 (a divergent Hox3 paralog), with disparate spatial and temporal expression patterns. It was not possible to separate the pb-like and zen2-like specificities within EO053, and sequences affecting both expression patterns were identified. Importantly, genomic deletion experiments demonstrate that EO053 cooperates with additional pb- and zen2-specific enhancers to regulate the mRNA expression of both genes. Sequence conservation of EO053 was examined within the Schizophora, and patterns of synteny between the Hox2 and Hox3 orthologs in Arthropods are consistent with a shared regulatory relationship extending prior to the Hox3/zen divergence. Thus, EO053 represents an example of two genes having evolved disparate outputs while utilizing this shared regulatory region. |
Proboscipedia is a homeotic protein required for the formation of labial and maxillary palps. It is a member of the Antennapedia Complex (ANTP-C), a linked array of homeodomain proteins. Both Proboscipedia (Pb) and Sex combs reduced (Scr) activities are required for determination of proboscis identity, while Scr determines tarsus identity. Simultaneous removal of Pb and Scr activity results in a proboscis-to-antenna transformation. Previous genetic observations suggest that Pb and Scr activity may interact. Five pieces of evidence support an interaction between Pb and Scr: (1) the proboscis of a null pb mutant is transformed into a pair of tarsi (the terminal segments of the leg), and (2) these alleles also result in reduced maxillary palps, which some investigators have interpreted as a transformation of the maxillary palps into antennae. (3) Ectopic expression of Pb from a heat-shock promoter/pb fusion gene, or in a small clone of cells from a Tubulin a1 (Tub a1) promoter/ pb fusion gene result in the transformation of the antennae into maxillary palps. (4) Ectopic expression of Scr from a heat-shock promoter/Scr fusion gene results in the transformation of the aristae into tarsi. (5) The proboscis of semilethal loss-of-function Scr alleles, and clones of Scr null mutant cells in the proboscis adopt maxillary palp identity (Percival-Smith, 1997 and references).
That both Pb and Scr activities are required for determination of proboscis identity, and that individual expression of Pb and Scr activities determines maxillary palp and tarsus identities, respectively, suggests a simple model for determination of four developmental identities. It is proposed that the expression patterns of Pb and Scr determine antenna, maxillary palp, tarsus and proboscis identities. Specifically, the absence of Pb and Scr expression, the default state, leads to antennal identity, expression of only Pb activity leads to maxillary palp identity, expression of only Scr activity leads to tarsus identity, and expression of both Pb and Scr activities leads to proboscis identity. A prediction of this simple model is that a proboscis primordial cell that is unable to express either Pb or Scr will adopt antennal identity (Percival-Smith, 1997).
Two mechanisms for the role of Pb and Scr in proboscis determination may be proposed. In both models, Pb regulates a set of Pb-regulated genes which, when expressed in isolation, determine maxillary palp identity. Similarly, Scr regulates a set of Scr-regulated genes that, when expressed in isolation, determine tarsal identity. In one model, expression of both sets of Pb-regulated genes and Scr-regulated genes in the same cell determines proboscis identity. In a second model, expression of Pb and Scr proteins in the same cell leads to formation of a Pb-Scr-containing, heteromeric, protein complex that regulates a novel set of genes that determines proboscis identity, the Pb-Scr-regulated genes. If the second model is correct, it should be possible to design dominant negative Pb and Scr molecules that will inhibit one another's activity (Percival-Smith, 1997).
In choosing the mutations used for the designed dominant negative Pb and Scr molecules, the properties of previously described change of DNA-binding specificity mutants made them ideal candidates. Both Pb and Scr have a glutamine at position 50 of the homeodomain (HD): pb and Scr genes have been created where this glutamine has been substituted for a lysine. This change is expected to change the DNA-binding specificity of Pb and Scr from Antennapedia class DNA-binding sites to Bicoid class DNA-binding sites, as has been extensively documented for other HDs. The result of this change would be that the Pb Q50K and Scr Q50K molecules, as well the Pb Q50K Scr and Pb-Scr Q50K -containing complexes, would not only have diminished affinity for their normal interaction site, but would also have an increased affinity for another set of sites, dragging away from their normal site of interaction the Pb Q50K and Scr Q50K molecules, as well as the Pb Q50K Scr and Pb-Scr Q50K -containing complexes (Percival-Smith, 1997).
Dominant negative Pb molecules inhibit the activity of Scr indicating that Pb and Scr interact in a multimeric protein complex in determination of proboscis identity. These data suggest that the expression pattern of Pb and Scr and the ability of Pb and Scr to interact in a multimeric complex control the determination of four adult structures (see above: antenna, maxillary palp, tarsus and proboscis). However, the Pb-Scr interaction is not detectable in vitro and is not detectable genetically in the head region during embryogenesis, indicating the Pb-Scr interaction may be regulated and indirect (for example, an additional factor binding to both proteins). This regulation may also explain why ectopic expression of Scr(Q50K) and Scr does not result in the expected transformation of the maxillary palp to an antennae and proboscis, respectively. Previous analysis of the requirements of Scr activity for adult pattern formation has shown that ectopic expression of Scr results in an antenna-to-tarsus transformation, but removal of Scr activity in a clone of cells does not result in a tarsus-to-arista transformation. In five independent assays the reason for this apparent contradictory requirement of Scr activity in tarsus determination is shown. Scr activity is required cell nonautonomously for tarsus determination. Specifically, it is proposed that Scr activity is required in the mesodermal adepithelial cells of all leg imaginal discs at late second/early third instar larval stage for the synthesis of a mesoderm-specific, tarsus-inducing, signaling factor, which after secretion from the adepithelial cells acts on the overlaying ectodermal cells determining tarsus identity (Percival-Smith, 1997).
It is suggested that the Drosophila leg is made up of two developmental fields: the tarsus and the proximal leg. These two developmental fields may correlate with the nuclear (proximal) versus cytoplasmic (distal) intracellular localization of Extradenticle, and the distal expression of Distalless. It is also proposed that there are four genetic pathways working in leg determination. The first pathway is the cell nonautonomous Scr-dependent, tarsus-inducing, signal pathway, and this lays down the plan for the basic unmodified tarsus. The second pathway is the relatively cell autonomous proximal leg pathway, which can be activated by the expression of Scr, Antp or Ubx and which lays out the basic plan for the proximal leg. The third and fourth pathways are cell autonomous pathways that Scr and Ubx control. A basic leg plan results in second leg identity, but expression of Scr or Ubx in both the proximal and distal portions of this basic plan brings about modifications resulting in first or third leg identity, respectively (Percival-Smith, 1997 and references).
When pb is mutated, labial palps are transformed to prothoracic legs and maxillary palps are small and malformed. pb has proven an ideal gene to study the role of homeotic proteins in regional determination and regulation of other homeotic genes. Two examples are given below.
In the first example, a particular deletion mutation in the ANTP-C results in diminished function of proboscipedia, and consequently in a defective head phenotype. This mutant also has a dominant thoracic defect related to diminished expression of the homeotic Antennapedia (Antp) gene. The reduced expression of Antp is a consequence of ectopic pb expression in the thorax. In the mutant the pb gene has come under the control of a second promoter, Antp P1, in addition to pb's own promoter. Ectopic PB protein expression occurs under Antp P1 control and results in diminished head transcription of pb and diminished accumulation of ANTP protein in the imaginal disc cells where Antp P1 is normally expressed. Thus PB protein is capable of participating in the negative regulation of a more posteriorly expressed homeotic gene (Antp), as well as serving a homeotic "selector" function in the head (Cribbs, 1992a).
pb is found between labial, the most proximal gene of the ANTP-C, and the rogue homeobox gene zerknült (Pultz, 1988).
Using alternatuive splicing, coding sequences across a 34 kb interval yield four identified mRNA forms that differ immediately upstream of the homeobox. As a consequence, the homeodomain is expected to reside in four different contexts in the predicted protein isoforms (Cribbs, 1992b).
Base pairs in 5' UTR - 1170
Exons - nine
Base pairs in 3' UTR - 360
A collection of pb mutant alleles were examined using antisera directed against either the N-terminal region, the center or the C-terminal region of the protein. Surprisingly, several partial loss-of-function pb alleles appear to generate partially functional proteins truncated at their C-termini. This suggests that a significant portion of the protein contributes quantitatively to PB function, but is partially dispensable (Cribbs, 1992b).
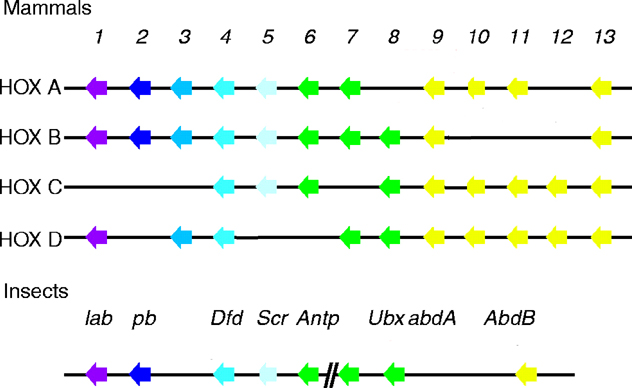
See four paralogous Hox clusters of mammals for homologies of Probosipedia with mammalian Hox cluster proteins.
date revised: 2 February 98
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
{kind=link}