proboscipedia
The six Drosophila proteins that belong to the antennapedia-type Homeobox subfamily are
Antennapedia (ANTP), Abdominal-A (ABD-A), Deformed (DFD), Proboscipedia (PB),
Sex combs reduced (SCR) and Ultrabithorax (UBX).
The ExPASy World Wide Web (WWW) molecular biology server of the Geneva University Hospital and
the University of Geneva provides extensive documentation for the 'Homeobox' antennapedia-type protein signature.
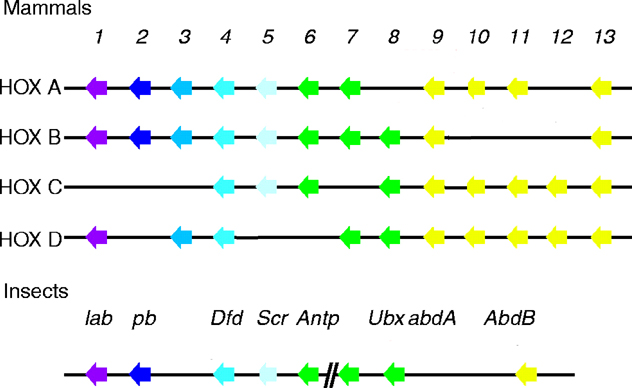
See four paralogous Hox clusters of mammals for homologies of Probosipedia with mammalian Hox cluster proteins.
Genes of the Hox cluster are restricted to the animal kingdom and play a central role in axial patterning in divergent animal
phyla. Despite its evolutionary and developmental significance, the origin of the Hox gene cluster is obscure. The
consensus is that a primordial Hox cluster arose by tandem gene duplication close to animal origins. Several homeobox
genes with high sequence identity to Hox genes are found outside the Hox cluster and are known as 'dispersed' Hox-like
genes; these genes may have been transposed away from an expanding cluster. Three of these dispersed
homeobox genes form a novel gene cluster in the cephalochordate amphioxus.
AmphiCdx, AmphiXlox and AmphiGsx are in respectively the Caudal, Zerknullt, and Gsh (related to Labial and Proboscipedia) families of homeodomain proteins.
The finding that amphioxus Gsx, Xlox and Cdx class genes form a novel homeobox cluster challenges the idea that these homeobox gene classes are 'dispersed' Hox genes.
Instead it is argued that this 'ParaHox' gene cluster is
an ancient paralogue (evolutionary sister) of the Hox gene cluster. ParaHox and Hox gene clusters arose by duplication of a ProtoHox
gene cluster containing Gsx, Xlox and Cdx genes representing an anterior, 'group three' and posterior subfamily. Amphioxus ParaHox genes have co-linear developmental expression patterns in
anterior, middle and posterior tissues. It is proposed that the origin of distinct Hox and ParaHox genes by gene-cluster
duplication facilitated an increase in body complexity during the Cambrian explosion (Brooke, 1998).
Proboscipedia in other insects
The Tribolium castaneum (Tc)
ortholog of the Drosophila homeotic gene Deformed
(Dfd) has been examined and its expression pattern during embryogenesis in this beetle has been determined. Tc Dfd is expressed
in the blastoderm and the condensing germ rudiment
in a region that gives rise to gnathal segments. During
germ band extension Tc Dfd is expressed in the mandibular
and maxillary segments, their appendages, and
the dorsal ridge. Comparison of insect Dfd protein sequences
reveals several highly conserved regions. To determine
whether common molecular features reflect conserved
regulatory functions, the Gal4 system was used to
express the Tribolium protein in Drosophila embryos.
When Tc Dfd is expressed throughout embryonic ectoderm
under the control of P69B, the beetle protein auto-regulates
the endogenous Dfd gene. In addition, the Drosophila
proboscipedia gene (a normal target of Dfd) is
ectopically activated in the antennal and thoracic segments.
The ability of the beetle and fly
proteins to rescue defects in Dfd minus mutants were compared by expressing
each throughout the embryonic during embryogenesis.
Both proteins rescue Dfd minus defects to the same extent in
that they each restore the development of mouth hooks
and cirri, as well as cause gain-of-function abnormalities
of posterior mouth parts. As before, pb is ectopically
activated in the antennal segment. This is the first demonstration
of the ability of a heterologous homeotic selector protein to directly regulate a target gene independent of an endogenous Drosophila autoregulatory loop (Brown, 1999).
The pb gene is located upstream of lab in the
HOM-C and is expressed in insects in the appendages of the maxillary
and labial segments, where it has been shown to specify the posterior
mouthparts in Drosophila and Tribolium castaneum. Oddly, the main expression domain of pb in
Drosophila is not colinear with that of lab and
Dfd. In all of the insects surveyed, the anterior boundary
of Dfd, the next upstream HOM-C gene relative to
pb, is in the mandibular segment, and, based on the
position of pb in the complex, one might expect its anterior
border of expression to be at or anterior to this point. To rationalize this fact, it has been suggested that an ancestral insect pb
gene lost its colinear expression pattern and gained a new,
appendage-specific role in the maxillae and labium. Studies of the
expression patterns of pb in several insect orders reveal
another, albeit weaker, expression domain in the ventral portion of the
intercalary. The significance of this domain is not clear, and in
some groups, including Drosophila, it has been found to be
mesodermal. In the apterygote insect T. domestica, there is
epidermal pb expression in the intercalary, but it is weak,
appears late, and is transient. These latter, more anterior
patches of expression may be remnants of the posited ancestral, more
extensive expression domain (Abzhanov, 1999).
Insects have evolved a large variety of specialized feeding
strategies, with a corresponding variability in mouthpart
morphology. In the Hemiptera, the mandibular and
maxillary segments give rise to two similar pairs of long, thin
stylets. The paired maxillary appendages form
channels for liquid flow and the piercing mandibles lie on
either side. These four interlocked stylets run down a groove
in the long, fused labium, which provides support. Although
in typical mandibulate insects the maxillary and labial
appendages are very similar, in the Hemiptera, it is
the mandibular and maxillary appendages that share a highly
unusual morphology very different from the labium.
These specialized mouthparts represent an important
evolutionary innovation that allows this order of insects to
feed by extracting fluids from other organisms.
There is little understanding of the
developmental mechanisms that underlie mouthpart diversity.
Until recently it was difficult to perform any analysis of
gene function outside of the genetic model insects
Drosophila melanogaster and Tribolium castaneum. In this
paper, the use of dsRNA-mediated interference
(RNAi) is described to dissect gene function in the development of the
milkweed bug Oncopeltus fasciatus, which has specialized
suctorial mouthparts. The Hox genes Deformed (Dfd),
proboscipedia (pb) and Sex combs reduced (Scr) have
previously been shown to be expressed in the gnathal
appendages of this species. Strikingly, the milkweed bug
was found to have an unusual expression pattern of pb.
Here, by analyzing single and combination RNAi
depletions, it has been found that Dfd, pb and Scr are used in the
milkweed bug to specify the identity of the mouthparts. The
exact roles of the genes, however, are different from what
is known in the two genetic model insects. The maxillary
appendages in the bug are determined by the activities of
the genes Dfd and Scr, rather than Dfd and pb as in the fly and beetle. The mandibular appendages are specified by
Dfd, but their unique morphology in Oncopeltus suggests
that Dfd's target genes are different. As in flies and beetles,
the labium is specified by the combined activities of pb and
Scr, but again, the function of pb appears to be different.
Additionally, the regulatory control of pb by the other two
genes seems to be different in the bug than in either of the
other species. These novelties in Hox function, expression
pattern and regulatory relationships may have been
important for the evolution of the unique Hemipteran head (Hughes, 2000).
Dfd is the sole gene responsible for mandibular identity. This conforms with the expression of Dfd in the mandibular segment. The single depletion of Dfd transforms the mandibular appendages to distal antennal identity, rather than the long thin stylets normally formed. Thus Dfd is necessary for proper mandibular development. In contrast, the depletion of pb leaves the mandibular appendages untouched. Thus pb is not necessary for mandibular development. The phenotype of the Scr depletion is more difficult to interpret, since the mandibular stylets generally fail to grow, and are merely short bristles in a mass of undifferentiated tissue. Since Scr is not expressed in the mandibular segment, it is suspected that this is an indirect effect of the Scr phenotype on other segments of the head. Because head development is integrated to some degree, non-local, indirect effects are often seen in Drosophila Hox mutants, particularly labial and Deformed. In the case of milkweed bug Scr, disruption of the proximal labium may be interfering with the normal development of the adjacent mandibular and maxillary appendages. The Dfd Scr double depletion corroborates this view. The mandibles in the double form distal antennae, indistinguishable from either the Dfd depletion alone or the triple. Therefore it can be inferred that, when present, Scr is not acting in the mandibular appendage to directly specify any identity over that of the default state (Hughes, 2000).
In the maxillary segment, Dfd depletion results in only a partial transformation of the maxilla to antennal identity. However, the Dfd;Scr double depletion results in complete transformation of the maxillae to antennae. Thus it can be concluded that Dfd acts in concert with Scr in the maxillary segment. The curled phenotype of the Dfd depletion maxillary appendage suggests that Scr may be repressing growth of the transformed limb in its posterior domain; this repression is released in the Dfd;Scr double depletion. The phenotypes in the maxillary segment are somewhat in conflict however, regarding the role of pb. Two results suggest that pb is not acting to specify maxillary identity. (1) Depletion of pb alone leaves the maxillary segment unaffected, therefore pb is not necessary for wild-type maxillary development. (2) The phenotype of the Dfd;Scr double is the same as the triple depletion (i.e., antennae), so it can be inferred that pb is not acting to confer any identity over that of the default. There is, however, a subtle difference between the phenotypes of the Dfd single and Dfd;pb double depletion. While the Dfd depletion produces short, curled antenna-like appendages, the Dfd;pb double produces straight antenna-like appendages, often with pretarsal claws. This suggests that in the absence of Dfd, pb can affect the maxillary appendages. Whether this activity derives from its small dorsal maxillary domain of accumulation, or from an expanded domain of pb expression in the Dfd depletion, is not yet clear. Nevertheless, based on the pb depletion, it is concluded that, in wild-type embryos, pb is not active in the specification of the maxillary stylet (Hughes, 2000).
This lack of pb function in determining the maxillary stylet matches the prediction made based on the expression pattern of pb in the bug. Thus far this is the only insect known not to use pb to determine maxillary identity. In Drosophila and Tribolium, pb/mxp mutations cause transformation of the maxillary palps, which become reduced in Drosophila, and transform to legs in Tribolium. In the bug, it would appear that Scr assumes much of this role. This is in contrast to Drosophila or Tribolium, where the Scr/Cx mutation does not affect the maxillary appendage. While Dfd and Scr work together to specify maxillary identity in the bug, it seems that Dfd has the principal role. Perhaps by activating a similar set of target genes as in the mandible, Dfd may induce a similar stylet identity in the maxillary appendage. This contrasts with mandibulate insects, in which the maxillary morphology is most similar to the labium, probably due to the activity of pb in both of those appendages (Hughes, 2000).
By analyzing the depletion phenotypes, it can be inferred that pb and Scr cooperate to pattern the labium. Dfd depletion leaves the labium unaffected, so it can be concluded that Dfd is not necessary for wild-type labial development. Depletion of pb results in a labium that is fused and wild-type basally, but splits distally into a pair of normal T1 legs. From this, it can be inferred that pb is necessary for distal, but not proximal, labial development. Scr depletion results in transformation of the entire appendage to a mixed identity between leg and antenna. From this it can be inferred that Scr is necessary for development of both proximal and distal labium. The pb;Scr double depletion, as well as the triple, transforms the labium to a pair of full-length antennae, which is very different from either single depletion alone. Therefore in the bug as well as the fly and beetle, pb and Scr specify the labium. The Scr depletion phenotype in the labium supports the hypothesis that Scr normally functions to fuse the two labial appendages. This conclusion is derived from the observation that, in wild-type and the pb depletion animals, the transformed labial appendage is basally fused, while the Scr depletion limbs are often well-separated (Hughes, 2000).
By comparing the pb depletion to the pb;Scr double, it can be inferred that the function of Scr in the absence of pb is to induce T1 legs. In light of the Drosophila model, this may seem reasonable, since Scr is also expressed in T1 but, in the case of the milkweed bug, it is actually a bit surprising. Based on its ectodermal pattern of accumulation in a single discrete spot on the T1 leg, Scr would not appear to be capable of conferring overall leg identity to this appendage. Rather, in ventral T1 Scr appears to function solely in the specification of the T1 leg comb. The role of pb in the gnathal appendages is provided by a comparison of the Scr single depletion animals to the pb;Scr doubles. The Scr depletion labium is transformed to a pair of appendages with features of both antenna and leg. This suggests that the activity of pb alone is conferring some leg identity over the default antennal state. This contrasts strongly with the function of pb in flies. In two situations in Drosophila, the presence of pb alone appears to induce maxillary identity: first, in Scr hypomorphic mutant adults, the labium is partially transformed to maxillae; second, when pb+ is ectopically expressed in the antennae, they are transformed to maxillary palps. Since in the bug pb is inducing partial leg identity rather than maxillary identity, this suggests that pb may have a very different function than in the fly. Since both pb and Scr, when acting alone, each seem to be specifying some leg identity, this suggests that pb and Scr have overlapping functions. It is known, however, that the two genes are not entirely redundant, because their depletion phenotypes are different and because when both are present the result is labial identity. Thus the activity of wild-type pb or Scr alone induces leg identity, but the combined synergistic activity of the two together creates a new overriding labium identity. Whether the combinatorial effect of pb and Scr is at the level of transcription of their target genes, or in the activity of their combined target gene products, is not yet clear (Hughes, 2000).
Since the triple depletion animals should lack all Hox activity in the gnathal segments, the default state of these appendages in the absence of Hox function can be inferred. Antennae have been thought to represent the default appendage state in the absence of Hox activity, since a large deletion of much of the Hom-C in the beetle was found to produce a larva bearing a series of antennae instead of mouthparts and legs. Likewise, in the adult fly, the pb and Scr double mutant combination in the adult labium gives rise to antennae. Triple depletion indicates that, in the milkweed bug as well, antennae represent the no-Hox state for the gnathal appendages. In the triple depletion, the maxillary and labial segments form full-length antennae. Since wild-type antennae are the only paired appendages that lack Hox expression, it is reasonable that this morphology is produced in the absence of Hox function. Furthermore, this result indicates that the genetic program that specifies the presence of an appendage is independent of Hox expression, and that the function of the Hox gene products is to confer a specific morphology onto a generic appendage program. In the absence of Hox activity, this generic appendage turns on the antennal differentiation pathway (Hughes, 2000).
In the absence of Hox activity, the mandibular appendages, however, each form only a distal antenna on a short base. This suggests that a portion of the generic appendage is missing. Previous work has suggested that insect mandibles, which lack Distal-less (Dll) expression, are gnathobasic (lack distal portions of the appendage). In Drosophila and Tribolium, Dll is repressed by Dfd in the mandibular segment. In the milkweed bug, therefore, depletion of Dfd is likely associated with Dll derepression. Consistent with this hypothesis, the morphology of the transformed mandibular appendage is primarily to distal antenna. Interestingly, as noted, the basal portions of the appendage are distorted or missing, compared to the maxillary or labial default appendages. This suggests that some aspects of the reduced structure of the insect mandible are independent of the Dfd/Dll pathway (Hughes, 2000).
In this study, evidence has been found of remodeling in the role of Hox genes in head appendage development. This change in the role of the Hox genes is due to three mechanisms: (1) change in expression patterns; (2) change in regulatory relationships, and (3) change in function. Previous work had shown that pb possesses a unique expression pattern in the milkweed bug. The RNAi analysis confirms that the unusual expression pattern does in fact reflect an unusual domain of activity. Unlike Drosophila, Tribolium and probably most other insects, pb is not necessary for development of the maxillary appendages in the milkweed bug. The pb gene is also regulated differently in the bug, where it appears to be independent of the activity of Dfd and Scr. This contrasts strongly with the regulatory interactions described for the fly and the beetle. Studies of more distant insects are needed, however, before the ancestral state and the direction of change for these regulatory interactions can be confidently inferred. Lastly, evidence was found of novel functions in the bug that probably reflects differences in the suite of target genes activated by the Hox genes. For example, the function of pb is different in the bug, where it induces a mixed leg-like identity rather than a maxillary palp. Even in the mandibular segment, where Dfd is the conserved controller of identity, the resulting mandibular appendage is strikingly different from the mandibles of other insects, indicating a very different developmental module at work downstream of the Hox signal. It is not known, however, whether the repertoire of target genes is different, whether the orthologous target genes themselves have different functions or if both possibilities are correct (Hughes, 2000).
These differences in Hox function are provocative, but a likely scenario for the evolution of the Hemipteran mouthparts cannot yet be described. The exclusion of pb from the maxillary segment may have caused the loss of the maxillary palp -- or the expression of pb may be the result of loss of the palp, which would remove selection to maintain pb in an inactive tissue. Moreover, the evolution of Hemipteran stylate-haustellate mouthparts was clearly a multistep process within the Hemipteroid group: the Psocoptera (booklice) and some Phthiraptera (chewing lice) have pronounced lacinia; the Thysanoptera (thrips), some Phthiraptera (sucking lice) and the Hemiptera have long stylets; and finally, the Hemipterans have lost the maxillary palps entirely. Understanding the changes that accompanied the evolution of these mouthparts will require additional study of the embryology, homologies and gene expression of these various insect orders (Hughes, 2000).
The role of Hox genes in the development of insect gnathal appendages has been examined in three insects: the fruitfly, Drosophila melanogaster; the milkweed bug, Oncopeltus fasciatus; and the red
flour beetle, Tribolium castaneum. In each of these organisms, the identity of the labium depends on the homeotic genes Sex combs reduced (Scr) and proboscipedia (pb). Loss of pb function in each of
the three insects results in homeotic transformation of the labial appendages to legs. In contrast, loss of Scr function yields a different transformation in each species. Interestingly, mutations in Cephalothorax (Cx), the Tribolium ortholog of Scr, transform the labial appendages to antennae, a result seen in the other insects only when both pb and Scr are removed. The Tribolium labial appendages also develop as antennae in double mutants. Further, expression of the Tribolium proboscipedia ortholog maxillopedia (mxp) is greatly reduced or absent in the labium of Cx mutant larvae. Thus, in the wild-type labial segment, Cx function is required (directly or indirectly) for mxp transcription. A similar interaction between Scr and pb during Drosophila embryogenesis has been described recently. Thus, this regulatory paradigm appears to be conserved at least within the Holometabola (DeCamillis, 2001).
The Tribolium Cx single mutant phenotype (transformation of larval labial appendages to antennae) differs from the phenotypes of Drosophila Scr mutant larvae and adults. Understanding the underlying regulatory pathways helps resolve this discrepancy. In both Drosophila and Tribolium embryos, Scr/Cx positively regulates pb/mxp. However, pb lacks an embryonic function in Drosophila, and some other gene is presumably responsible for the residual gnathal identity in the labial segment of Scr mutant larvae. In adult flies, pb functions to specify the maxillary palps. Scr hypomorphs and mitotic clones lacking Scr cause transformation of the adult labial appendages to maxillary palp. However, when clones lack both Scr and pb, cells assume antennal identity. These results suggest that, in contrast to the embryonic paradigm, pb is present and functional during Drosophila adult development even in the absence of Scr. Indeed, expression of pb in the labial imaginal disc does not require Scr function. In fact, to some extent, the reverse appears to be true. Scr expression is greatly reduced in the distal portions of pb null labial imaginal discs (DeCamillis, 2001 and references therein).
Orthologs of the Hox genes Sex combs reduced and proboscipedia are active in the developing labial appendages of all insect species tested. The remarkable variation among insect gnathal structures, particularly in the distal podomeres, suggests two Hox genes may enhance the adaptive potential of gnathal appendage morphology. Functional studies in Drosophila melanogaster, the flour beetle Tribolium castaneum and the milkweed bug Oncopeltus fasciatus show that cooperation between Scr and pb has been generally conserved, but specific mechanisms have been altered during evolution. Cross-regulation of pb by Scr is evident in Drosophila and Tribolium, the more closely related of the three species, but not in Oncopeltus. In all three species, pb function is restricted to the distal podomeres, but details are only known for Drosophila and Oncopeltus, two species exhibiting specialized stylate-haustellate mouthparts. Drosophila pb is required for distal Scr expression, and to repress the appendage patterning genes dachshund and Distal-less. Oncopeltus pb has the novel capacity to specify leg fates. Little is known about distal functions of Tribolium pb. Hypomorphic mutations of the Tribolium pb ortholog maxillopedia can be arranged in a graded phenotypic series of palp to leg transformations along both the proximodistal and dorsoventral axes. Mid-embryonic expression profiles of Tribolium pb, Scr, wingless and Dll genes were examined in maxillopedia hypomorphic and null mutant backgrounds. Levels of pb and Scr are significantly reduced in the distal appendage field. Tribolium pb therefore positively regulates distal Scr expression, a role that it has in common with Drosophila pb. Tribolium wg is normally down-regulated in the distal domain of the embryonic gnathal appendage buds. It becomes activated distally in maxillopedia hypomorphs. Repression of wg by pb has not been reported in the labial imaginal discs of Drosophila. Alterations of Tribolium Scr and wg expression occur in Dll-expressing cells, however, unlike in Drosophila labial imaginal discs, Dll expression appears unaffected in pb hypomorphic backgrounds. It is concluded that the Hox genes Sex combs reduced and proboscipedia control an appendage organizer and cell autonomous fate determination during embryonic labial palp development in Tribolium (DeCamillis, 2003).
Other invertebrate Proboscipedia-related Hox genes
Expression patterns for five Hox genes were examined by whole-mount in situ hybridization in larvae of Chaetopterus, a
polychaete annelid with a tagmatized axial body plan. The polychaetes are generally thought to be the basal
taxon of the annelids and thus are a good group to sample
for developmental patterns more likely to represent ancestral
states than those of the more phylogenetically derived
leeches. Phylogenetic analysis demonstrates that these genes are orthologs of
the Drosophila genes labial, proboscipedia, zen, Deformed, and Sex combs reduced: they have been termed CH-Hox1, CH-Hox2,
CH-Hox3, CH-Hox4, and CH-Hox5, respectively. Expression studies reveal a biphasic expression pattern. In early larval
stages, well before any indications of segmental organization exist, a novel pattern of expression in bilateral posterior
proliferating cell populations, corresponding to the teloblasts, was detected for each of the genes, with CH-Hox1 and
CH-Hox2 expressed before the remaining three. In middle larval stages, all five genes are expressed in bilateral strips along
the ventral midline, corresponding with the developing ventral nerve cord. In addition, CH-Hox1 and CH-Hox2 show strong
expression at the foregut-midgut boundary. By late larval stages the expression is generally confined to the ventral CNS and
ectoderm of the anterior parapodia. Anterior boundaries of expression are 'colinear', at later larval stages, with CH-Hox2
expressed most rostrally, in the first segment, and anterior expression boundaries for CH-Hox1, CH-Hox3, CH-Hox4, and
CH-Hox5 in segments 2, 3, 4, and 5, respectively. Like vertebrates and spiders, but unlike insects, CH-Hox3 participates in
this colinear axial expression pattern. CH-Hox1 and CH-Hox2 have distinct posterior boundaries of expression in the ninth segment, which corresponds to a major morphological boundary, and may reflect a reorganization of Hox gene regulation
related to the evolutionary reorganization of the Chaetopterus body plan (Irvine, 2000).
The most striking aspect of Hox gene expression found in
this study is the early onset of expression in the posterior of
the larva. This early expression is localized to bilaterally
disposed cell populations located ventrolaterally, just anterior
to the pygidium. CH-Hox1 and CH-Hox2 are the first of
these genes to be expressed at detectable levels, exhibiting
robust in situ hybridization signals from early stage L1 (18
h). Expression of CH-Hox3, CH-Hox4, and CH-Hox5 is not
detected until stage L2 (some 24 h later), with an apparent
temporal gradient of expression onset from CH-Hox1 and 2
to CH-Hox3 and 4, with CH-Hox5 showing the latest
expression. The temporal sequence of Hox gene
expression is not an artifact, since expression of each of
these genes is detectable and persists in a bilaterally symmetrical
fashion in the prepygidial region throughout all
subsequent developmental stages. Although the expression
pattern follows the same sort of temporal sequence seen in
other metazoans, the expression of Hox genes in Chaetopterus
begins well before any indications of segmental organization
appear in the larval body plan (Irvine, 2000).
All five of the Hox genes examined are expressed at
these early stages in the same general region in the posterior
of the animal and may be turning on at nearly the same
times in many of the same cells. Expression initially occurs
in a largely overlapping domain and it is likely that Hox
gene transcription may be loosely regulated in early stages
to be refined later, either by translational regulation or by
differences in the timing of transcriptional downregulation
in a way similar to that seen in planarian flatworms (Irvine, 2000).
The general model of annelid development is based on
the presence of a 'posterior growth zone' just anterior to
the pygidium. The dynamics of cell division and posterior
expansion in this larval region are completely unknown:
various authors have claimed historically that both definitive
ectodermal and mesodermal stem cells reside in this
region and give rise to the various adult tissues of the
posttrochal region. The extreme version
of this type of teloblastic growth is seen in the
clitellates in which highly stereotyped sets of five bilaterally
symmetrical teloblast stem cells can be found and
named and their descendents studied in detail. The condition
in the clitellates is most often interpreted as an
acceleration of the formation of the adult body plan, as
these animals have forsaken larval development to become
direct developers.
The locations of cells expressing Hox gene transcripts
during early development in Chaetopterus coincide with
the predicted location of the prepygidial growth zone, both
in the anterior-posterior level and in the bilateral localization
of expressing cell populations. This largely overlapping
pattern of expression in terminal cell populations at early
stages has not been reported in other taxa, including those
with teloblastic growth, such as molluscs, crustaceans, and
most notably, a related class of annelids, the leeches. In
flies, for example, Antp transcripts are detected at the
cellular blastoderm stage, but are already localized to the
primordia of segments T1 and T2. In contrast,
Chaetopterus transcripts for each of the
genes examined are present in the putative growth
zone before any segmental primordia are formed. The very
early appearance of Hox gene expression in the putative
Chaetopterus growth zone may relate to the possibility of
early specification of segmental fate within teloblast lineages (Irvine, 2000).
Cell transplantation experiments in leeches have
shown that segmental founder cells derived from both
ectodermal and mesodermal teloblasts are determined at
the time of their birth. If
this early fate specification is true in polychaetes, it could
explain the presence of Hox genes, acting in a fate specification
role, in the set of segmental founder cells. The absence of
Hox gene expression in determined segmental founder cells
in leech embryos suggests that Hox gene expression is not
involved in the initial establishment of segmental identity
in annelids, and that the early pattern in polychaetes has
some other function, such as in cell proliferation. If the
early Hox expression detected in Chaetopterus does indeed
have a role in the precocious specification of segmental fate,
the lack of expression in leeches suggests that other, possibly
upstream, genes serve in place of the early phase of Hox
expression seen in Chaetopterus (Irvine, 2000).
cDNA fragments of the homologs of the Drosophila
head homeotic genes labial (lab),
proboscipedia (pb), and
Deformed (Dfd) have been isolated from
the crustacean Porcellio scaber. P. scaber (order Isopoda) belongs to the subclass Malacostraca (higher crustaceans); relative to phylogenetically more basal groups, it is as derived as insects are in its body plan and
tagmatization. Moreover, the interpretations of the expression patterns reported here are based on the
assumptions that: (1) the Insecta is monophyletic, with the order Thysanura a basal group; (2) the Mandibulata are monophyletic with the Crustacea, a sister
group of the Insecta, and (3) the Chelicerata (spiders and mites) is an outgroup in relation to the Insectan-Crustacean clade.
Because the
accumulation domains of the head homeotic complex (Hox) genes had not
been previously reported for crustaceans, the expression
patterns of these genes was examined in P. scaber embryos by using in situ hybridization. The P. scaber lab
homolog is expressed in the developing second antennal segment and
its appendages. This expression domain in crustaceans and in the
homologous intercalary segment of insects suggests that the
lab gene specified this metamere in the last common
ancestor of these two groups. The expression domain of the P.
scaber pb gene is in the posterior part of the second antennal
segment. This domain, in contrast to that in insects, is colinear with
the domains of other head genes in P. scaber; it
differs from the insect pb gene expression domain in the posterior mouthparts, suggesting that the insect and crustacean patterns evolved independently from a broader ancestral domain similar
to that found in modern chelicerates. P. scaber Dfd is expressed in the mandibular segment and paragnaths (a pair of ventral
mouthpart structures associated with the stomodeum) and differs from
insects, where expression is in the mandibular and maxillary segments.
Thus, like pb, Dfd shows a divergent Hox
gene deployment. It is concluded that homologous structures of the
mandibulate head display striking differences in their underlying
developmental programs related to Hox gene expression (Abzhanov, 1999).
To assign the
P. scaber head appendages to specific segments, the monoclonal antibody Mab4D9, which
recognizes EN, was used to indicate the posterior of the segmental borders. The antibody revealed six segments in the
embryonic head of P. scaber: ocular, first antennal, second antennal, mandibular, first maxillary, and second maxillary. The most anterior region of the head, the labrum, develops as a pair of small appendage-like structures that
fuse medially at the stage when development is about 65%-70% complete. In early embryos, the EN stripe of the ocular segment is not
complete and is interrupted on the ventral side by the labrum. The labrum itself does not express EN and, in this respect, appears to be continuous with the
stomodeum. The first antennal segment bears a pair of small uniramose antennae, which are reduced in the adult. The second antennae are the largest pair of
the appendages on the head. The stomodeal opening protrudes at the level of the posterior first antennal (a1) segment and extends to the posterior of the
second antennal (a2) segment: this results in a ventral interruption of the EN stripes of the a1 and a2 segments. The broad mandibular EN band is seen in
the developing posterior mandibular appendages. EN is expressed in a similar fashion in the first and second maxillary segments (Abzhanov, 1999).
The pb expression domain of P. scaber was found
to be quite dissimilar from the insect pattern. In this crustacean,
pb accumulation is restricted to the posterior part of the
a2 segment and includes neither the mx1 nor the mx2 segments or
appendages. The first and
second maxillae of crustaceans are homologous to the maxillae and
labium of insects, respectively. Since pb is required for
maxillary and labial appendage development in insects, it clearly
cannot be performing a similar function in the P. scaber
embryo. At this point, it is difficult to discern a possible
developmental function for pb in P. scaber embryos (Abzhanov, 1999).
A
comparison of the crustacean/insect and chelicerate (spiders and mites) patterns of Hox
gene expression is made difficult by the uncertainty of segmental
homologies between the two groups. The more traditional view, based on
anatomy and patterns of innervation, concludes that the segment
associated with the deuterocerebral ganglion of the central nervous
system is greatly reduced or absent in modern chelicerates. If
this conclusion is correct, the more anterior ocular/protocerebral
segment would be homologous to the same segment in insects and
crustaceans, whereas the next posterior segment in the spider would
correspond to the intercalary/tritocerebral segment of insects and to
the second antennal segment of crustaceans. Thus, the homolog of the
insect antennal and the crustacean first antennal segment would be
absent in chelicerates. More recently, this question of
head-segment homology has been revisited by using the patterns of Hox
gene expression as a basis for determining the presence or absence of
the deuterocerebral segment in chelicerates. Using the spider Cupiennius salei, the mite
Archegozetes longisetosus, and several head Hox genes as
probes, it has been concluded that this segment is present.In chelicerates, lab
is expressed anteriorly in the developing pedipalps (homologous to the
appendages of the intercalary segment). This expression extends
posteriorly, to the fourth pair of walking limbs: this location corresponds
to the first pair of the thoracic appendages in insects and crustaceans
(maxillipeds in P. scaber). Therefore, the anterior
boundary of lab expression appears to be conserved among
chelicerates, crustaceans, and insects. It is notable, however, that
the posterior limit of expression is not conserved. The integumentary
expression of lab is limited to the intercalary or second
antennal segment in insects and crustaceans, respectively, whereas in
chelicerates, expression extends posteriorly a further four full
segments. In A. longisetosus, pb is expressed in a broad
domain from the pedipalps to the third pair of walking legs, where it
is accumulated in the appendages. This boundary is colinear with
both lab and Dfd, and is thus similar to the
relative expression domains of these genes in vertebrates, annelids,
and Porcellio; but it is dissimilar to that seen in
insects. Thus, the anterior boundary of pb expression in
the a2 segment of P. scaber appears to resemble that seen in
chelicerates rather than that in insects. However, it should also be
noted that, as for lab, the expression domain of chelicerate
pb extends further to the posterior than that in insects and
crustaceans (Abzhanov, 1999).
Comparison of the expression patterns of the crustacean and insect Hox
genes demonstrates that there is conservation of segment affinity
(e.g., lab) and spatial colinearity (e.g., lab,
pb, Dfd, and Scr) of expression. In addition, the anterior boundaries
of the lab and Dfd genes appear to be conserved
in insects, crustaceans, and chelicerates. However, there is also
divergence of the observed expression domains (e.g., pb and
Dfd). Consequently, substantial
variation in the deployment of the Hox genes, and presumably in the
developmental processes regulated by them, can be seen in homologous
and morphologically similar crustacean and insect head structures.
Genes involved in the development of mandibles and posterior mouthparts
in insects are expressed in novel, though still colinear domains. For
example, in insects, the maxillary and labial mouthparts express
pb, whereas in P. scaber, the homologous
appendages both express and probably depend on Scr, a
different head homeotic gene (Abzhanov, 1999).
It is hypothesized that the mandibulate head evolved prior to the
establishment of the defined head Hox gene expression domains which
have been recruited independently to their current regions and developmental functions in crustaceans and insects. This model involves an intermediate hypothetical
mandibulate ancestor that did not have segment-specific expression
domains and probably resembled the pattern of expression seen in modern
chelicerates. The specification of individual segments and mouthparts
in such an animal would depend on the redundant and/or fractional
functions of multiple Hox genes, and would be facilitated by the
subsequent evolution of more distinct expression domains. That is, the head Hox genes would have functioned in a manner analogous to the
genes of the D. melanogaster Bithorax complex. To test this model and to better understand the evolution of the Hox genes and head
structures, further studies across different crustacean and myriapod
groups will be required (Abzhanov, 1999).
The expression patterns of Hox genes have not previously been comprehensively analyzed in a myriapod. The expression patterns are presented of the ten Hox genes in a centipede, Lithobius atkinsoni, and these results are compared to those from studies in other arthropods. Three major findings are reported. (1) It has been found that Hox gene expression is remarkably dynamic across the arthropods. The expression patterns of the Hox genes in the centipede are in many cases intermediate between those of the chelicerates (spiders) and those of the insects and crustaceans, consistent with the proposed intermediate phylogenetic position of the Myriapoda. (2) Two 'extra' Hox genes were found in the centipede compared with those in Drosophila. Based on its pattern of expression, Hox3 appears to have a typical Hox-like role in the centipede, suggesting that the novel functions of the Hox3 homologs zen and bicoid were adopted somewhere in the crustacean-insect clade. In the centipede, the expression of the gene fushi tarazu suggests that it has both a Hox-like role (as in the mite), as well as a role in segmentation (as in insects). This suggests that this dramatic change in function was achieved via a multifunctional intermediate, a condition maintained in the centipede. (3) It was found that Hox expression correlates with tagmatic boundaries, consistent with the theory that changes in Hox genes had a major role in evolution of the arthropod body plan (Hughes, 2002).
The expression of the Hox genes corresponds roughly with the tagmatic divisions in the centipede. The expression of the genes lab, pb, Hox3 and Dfd is confined to the head, while the trunk is apparently under the control of Antp, Ubx, abd-A and Abd-B. Interestingly, the maxilliped segment has expression of three genes that extend both into the head (Scr and ftz) and into the trunk (Antp). The maxilliped segment is thought to be homologous to the first trunk or thoracic segment of other mandibulate arthropods. The appendages of this segment in the centipede, however, have been highly modified. While their leg-like structure is still evident, they develop to become short and broad fangs, complete with a poison gland. Thus, the first legs of the centipede are modified to become more mouthpart-like, and are used for prey capture and manipulation. This mixed head/trunk identity of the segment seems to be reflected in the Hox code found there. While the segment itself has only a 'trunk' Hox gene (Antp), the appendages have expression of Antp as well as the 'head' genes Scr and ftz, which are also expressed in the maxillary II segment. It remains to be determined how these genes contribute to the development of the centipede fangs. It would also be interesting to know whether the evolution of this novel appendage is correlated with a shift in the expression of these genes. Further studies of Hox expression in other myriapods such as a millipede, or functional studies in the centipede, would be very interesting regarding these issues (Hughes, 2002).
Comparing the expression of the centipede Hox genes with those of other arthropods reveals significant variability in the observed patterns. For example, in the chelicerate head the Hox expression domains broadly overlap. These same genes are expressed in much more restricted domains in the head segments of crustaceans and insects. Interestingly, the expression domains of these genes in the centipede are intermediate between these two extremes. For example, the gene lab is expressed over five segments in the spider, two segments in the centipede, and only a single segment in the crustaceans and insects. Likewise, the three-segment expression domain of centipede Dfd is intermediate between the four-segment domain in the spider and mite, and the two-segment domains of the crustaceans and insects. Most striking is the comparison between expression of pb among the four groups. In the spider, pb is expressed over five segments, from the pedipalps through the fourth walking leg. In the centipede, the expression domain covers four segments, from the intercalary to the maxillary II. In the crustaceans, the expression is restricted to the antennal II segment, which is homologous to the intercalary segment. In the insects, however, the expression of pb is more posterior, limited mainly to the appendages of the maxillary and labial segments (homologous to the maxillary I and II segments of the centipede). These expression patterns suggest that the centipede may retain some Hox expression domains in an intermediate state of their evolution, from the broad domains of the chelicerates to the more-restricted, less overlapping patterns of the crustaceans and insects. Moreover, the expression domain of pb apparently became differently subdivided in different lineages -- towards the anterior in the crustaceans, and towards the posterior in the insects (Hughes, 2002).
Domain structure of Proboscipedia-related Hox genes
Hox proteins control developmental patterns and cell differentiation in vertebrates by acting as positive or negative regulators of still unidentified downstream target genes. The homeodomain and other small accessory sequences encode the DNA-protein and protein-protein interaction functions that ultimately dictate target recognition and functional specificity in vivo. The effector domains responsible for either positive or negative interactions with the cell transcriptional machinery are unknown for most Hox proteins, largely due to a lack of physiological targets on which to carry out functional analyses. The transcriptional activation domains have been identified for three human Hox proteins, HOXB1, HOXB3, and HOXD9, that interact in vivo with the autoregulatory and cross-regulatory enhancers of the murine Hoxb-1 and human HOXD9 genes. Activation domains have been defined both in a homologous context, i.e., within a HOX protein binding as a monomer or as a HOX-PBX heterodimer to the specific target, and in a heterologous context, after translocation to the yeast Gal4 DNA-binding domain. Transfection analysis indicates that activation domains can be identified in different regions of the three HOX proteins depending on the context in which they interact with the DNA target. These results suggest that Hox proteins may be multifunctional transcriptional regulators, interacting with different cofactors and/or components of the transcriptional machinery, depending on the structure of their target regulatory elements (Vigano, 1998).
A conventional deletion analysis
on the 270-aa N terminus of HOXD9 shows that the first 75 residues contain a potential
transcriptional activator when tested in the context of a Gal4 chimeric protein. In contrast, this region is
dispensable when the activity of the protein is tested on the HCR (Hox control region, a
90-bp, ATTA-rich autoregulatory element identified in the HOXD9 locus), a context in which most of the
activating function appears to be located within residues 76 to 264. The regions identified by the two
alternative assays share no obvious characteristics with canonical eukaryotic activator domains and are
only loosely conserved among different vertebrate species. The activation domain of another posteriorly expressed Hox protein, HOXD8, can be localized to a
similar sub-N-terminal region. HOXD8 and HOXD9 bind the multiple ATTA-containing sites
within the HCR as monomers in a noncooperative fashion, while Gal4-DBD chimeras bind the
Gal4-responsive element (UAS) as a homodimer, a context that could force the HOXD9 N-terminal
region to assume a different structural conformation and unmask a potential activating function in the
N-terminal 75 residues. For the HOXB1-PBX heterodimer, the analysis carried out on the natural autoregulatory element
identifies a transcriptional activation domain in a Ser-Pro-rich, 52-residue sub-N-terminal region. This
region also contains most of the HOXB1 transcriptional activity when tested as a Gal4-DBD chimera,
a possible indication that the 52-residue region assumes a similar conformation or activates transcription
by a similar mechanism, either in the context of a homodimer or in that of a HOX-PBX heterodimer.
The activity of HOXB3 was tested in three different contexts, i.e., upon binding DNA as a monomer to
an ATTA-containing element: as a HOX-PBX heterodimer to a bipartite HOX-PBX core element, and
as a Gal4-DBD chimeric homodimer to the Gal4-responsive element. Although in the context of a
monomer the transcriptional activity is spread over the entire protein sequence, only the C terminus
contains a potent activator domain in the context of a Gal4 homodimer or of a PBX heterodimer. The
71-residue C terminus is relatively highly conserved in the mammalian group 3 Hox proteins (Vigano, 1998).
Proboscipedia related Hox genes and lateral line development
As a further investigation of vertebrate head morphogenesis, expression patterns of several homeobox-containing genes were examined using whole-mount in situ hybridization in a sensory system considered to be primitive for the vertebrate subphylum: the axolotl (class: Amphibia, order: Urodela) lateral lines and the placodes from which they develop. The lateral line system develops from the ectodermal placodes. The lateral line placodes develop in a dorsolateral series parallel to the main body axis; it has been hypothesized that the dorsolateral and ventrolateral placode series may be patterned by a mechanism similar to the Hox code described for the head and branchial regions of amniote embryos. Axolotl Msx-2 and Dlx-3 are expressed in all of the lateral line placodes. Both genes are expressed throughout development of the lateral line system and their expression continues in the fully developed neuromasts. Expression within support cells is highly polarized. In contrast to most other observations of Msx genes in vertebrate organogenesis, expression of Msx-2 in developing lateral line organs is exclusively epithelial and is not associated with epithelial-mesenchymal interactions. A Hox-complex gene, Hoxb-3, is shown to be expressed in the embryonic hindbrain and in a lateral line placode at the same rostrocaudal level, but not in other placodes nor in mature lateral line organs. A Hox gene of a separate paralog group, Hoxa-4, is expressed in a more posterior hindbrain domain in the embryo, but is not expressed in the lateral line placode at that rostrocaudal level. These data provide the first test of the hypothesis that the neurogenic placodes develop in two rostrocaudal series aligned with the rhombomeric segments and are patterned by combinations of Hox genes in parallel with the central nervous system (Metscher, 1997).
Mutation in Proboscipedia-related Hox genes
Mice with a disruption in the hoxb-2 locus (homolog: Drosophila proboscipedia) were generated by gene targeting. 75% of the
hoxb-2 mutant homozygotes died within 24 hours of birth. While a majority of these mice have
severe sternal defects that compromise their ability to breathe, some have relatively normal
sternum morphology, suggesting that one or more additional factor(s) contribute to
neonatal lethality. At 3-3.5 weeks of age, half of the remaining hoxb-2 homozygotes become
weak and subsequently die. All of the mutants that survive to 3 weeks of age show
marked facial paralysis similar to, but more severe than, that reported for hoxb-1 mutant
homozygotes (homolog: Drosophila labial). As for the hoxb-1 mutations, the facial paralysis observed in
mice homozygous for the hoxb-2 mutation results from a failure to form the somatic motor
component of the VIIth (facial) nerve which controls the muscles of facial expression.
Features of this phenotype closely resemble the clinical signs associated with Bell's Palsy
and Moebius Syndrome in humans. The sternal defects seen in hoxb-2 mutant mice are
similar to those previously reported for hoxb-4 mutant mice (homolog: Drosophila Deformed). The above results
suggest that the hoxb-2 mutant phenotype may result in part from effects of the hoxb-2
mutation on the expression of both hoxb-1 and hoxb-4. Consistent with this proposal, it was
found that the hoxb-2 mutation disrupts the expression of hoxb-1 in cis. In addition, the
hoxb-2 mutation changes the expression of hoxb-4 and the hoxb-4 mutation, in turn, alters
the pattern of hoxb-2 expression. hoxb-2 and hoxb-4 appear to function together to mediate
proper closure of the ventral thoracic body wall. Failure in this closure results in severe defects of the sternum (Barrow, 1996).
In Hoxa-2(-/- ) embryos, the normal skeletal elements of the second branchial arch are replaced by a duplicated set of first arch elements. Hoxa-2 is here shown to direct proper skeletal formation in the second arch by preventing chondrogenesis and
intramembranous ossification. In normal embryos, Hoxa-2 is expressed throughout the second arch mesenchyme, but is excluded from the chondrogenic condensations. In the absence of Hoxa-2, chondrogenesis is activated ectopically within the rostral Hoxa-2
expression domain to form the mutant set of cartilages. In Hoxa-2(-/- ) embryos the Sox9 expression domain is shifted into the normal
Hoxa-2 domain. Misexpression of Sox9 in this area produces a phenotype resembling that of the Hoxa-2 mutants. These results
indicate that Hoxa-2 acts at early stages of the chondrogenic pathway, upstream of Sox9 induction. Hoxa-2 is also shown to inhibit
dermal bone formation when misexpressed in its precursors. Furthermore, molecular analyses indicate that Cbfa1 is upregulated in the
second branchial arches of Hoxa-2 mutant embryos, suggesting that prevention of Cbfa1 induction might mediate Hoxa-2 inhibition of
dermal bone formation during normal second arch development. Cbfa1/Osf2, is a member of the Runt-related family of transcription factors that plays a critical role
during osteoblast differentiation (Kanzler, 1998).
Hox genes appear to have a central role in specifying A-P positional information to the cranial neural crest up to rhombomere 3 and therefore may regulate the morphogenetic programs of the hyoid (branchial arch 2) and more posterior brachial arches. Mice lacking Hoxa-2 form skeletal structures in branchial arch 2 that resemble proximal first arch elements. Thus in B2, Hoxa-2 and perhaps Hoxb-2 may affect the expression of genes involved in P-D patterning (e.g., Distal-less homologs Dlx-1 and -2). This would be similar to the role of Drosophila Deformed regulating Distal-less during development of the maxillary segment. Otx-2 could regulate Dlx genes anterior to branchial arch 2 (Qui, 1997).
Segmentation plays an important role in neuronal diversification and organization in the developing hindbrain. For instance, cranial nerve branchiomotor nuclei are organized segmentally within the basal plates of successive pairs of rhombomeres. To reach their targets, motor axons follow highly stereotyped pathways exiting the hindbrain only via specific exit points in the even-numbered rhombomeres. Hox genes are good candidates for controlling this pathfinding, since they are segmentally expressed and involved in rhombomeric patterning. In Hoxa-2(-/-) embryos, the segmental identities of rhombomere (r) 2 and r3 are altered molecularly as well as anatomically. Cellular analysis by retrograde dye labelling reveals that r2 and r3 trigeminal motor axons turn caudally and exit the hindbrain from the r4 facial nerve exit point and not from their normal exit point in
r2. Dorsal r2-r3 patterning is also affected, with loss of cochlear nuclei and enlargement of the lateral part of the cerebellum. These results point to a novel role for Hoxa-2 in the control of r2-r3
motor axon guidance, and also suggest that its absence may lead to homeotic changes in the alar plates of these rhombomeres (Gavalas, 1997).
Three different alleles of the Hoxb4 locus were generated by gene targeting in mice. Two alleles contained insertions of a selectable marker in the first exon in either orientation, and, in the third, the selectable marker was removed, resulting in premature termination of the protein. Presence and orientation of the selectable marker correlate with the severity of the phenotype, indicating that the selectable marker induces cis effects on neighboring genes that influence the phenotype. Homozygous mutants of all alleles had cervical skeletal defects similar to those reported for Hoxb4 mutant mice. In the most severe allele, Hoxb4PolII, homozygous mutants died either in utero at approximately E15.5 or immediately after birth, with a severe defect in ventral body wall formation. Analysis of embryos showed thinning of the primary ventral body wall in mutants relative to control animals at E11.5, before secondary body wall formation. Prior to this defect, both Alx3 and Alx4 were specifically down regulated in the most ventral part of the primary body wall in Hoxb4PolII mutants. Hoxb4loxp mutants in which the neo gene has been removed did not have body wall or sternum defects. In contrast, both the Hoxb4PolII and the previously described Hoxb2PolII alleles that have body wall defects have been shown to disrupt the expression of both Hoxb2 and Hoxb4 in cell types that contribute to body wall formation. These results are consistent with a model in which defects in ventral body wall formation require the simultaneous loss of at least Hoxb2 and Hoxb4, and may involve Alx3 and Alx4 (Manley, 2001).
Hoxa2 is required for a variety of developmental processes in the branchial arches and in the hindbrain. A Hoxa2 allele has been created
that is about 45% as active in transcription as its wild-type counterpart. This allele, together with the Hoxa2 null and wild-type alleles,
allows the generation of embryos to develop in the presence of different levels of Hoxa2 activity. Analysis of these embryos indicates that
in general the hindbrain is more resistant to Hoxa2 deficiencies than the second branchial arch. Also, within the second arch, proximo-caudal areas are more sensitive than the rostro-distal. In the hindbrain, basic segmentation and patterning processes seem to occur normally at Hoxa2 levels as low as 20% of the normal. In addition, specific neuronal markers along the dorso-ventral axis of the hindbrain seem differentially affected by reduced Hoxa2 levels. These results provide new clues to understand the role of Hoxa2 in the different embryonic areas where it is required (Ohnemus, 2001).
Although the general R-C and D-V developmental plans
seemed preserved at very low Hoxa2 levels, different sensitivities
to Hoxa2 deficiencies were observed in markers for
specific neuronal subtypes. In particular, Evx1, which stains
a subset of ventral interneurons and whose expression is compromised in the absence of Hoxa2, is still expressed in the r2
and r3 areas of Hoxa2tetO/delta1
embryos. Conversely, Phox2b, which is dorsally restricted to a specific column of differentiating neurons, was almost absent
from the same embryos. This suggests that different mechanisms
might mediate the effects of Hoxa2 on the expression
of different neuronal markers. Phox2b, which lies within the
Hoxa2 expression domain, could be under direct Hoxa2
regulation, and therefore more sensitive to changes in this
gene's levels. This might explain its apparent independence
from other D-V patterning processes. In the case of other
more ventral markers, like Evx1, which are not included in
the Hoxa2 high expression column, dependence on Hoxa2
expression might be indirect, secondary to proper D-V cell
segregation in the hindbrain. Therefore, they are affected
only when Hoxa2 levels are low enough to interfere with
D-V patterning processes. This is consistent with the close
association of the expression patterns of Pax6 and Evx1 in
embryos with different Hoxa2 genotypic combinations (Ohnemus, 2001).
Hox genes are known key regulators of embryonic segmental identity, but little is known about the mechanisms of their action. To address this issue, how Hoxa2 specifies segmental identity in the second branchial arch was analyzed. Using a subtraction approach, it was found that Ptx1 (see Drosophila Ptx1) is upregulated in the second arch mesenchyme of Hoxa2 mutants. This upregulation has functional significance because, in Hoxa2-/-;Ptx1-/- embryos, the Hoxa2-/- phenotype is partially reversed. Hoxa2 interferes with the Ptx1 activating process, which is dependent on Fgf signals from the epithelium. Consistently, Lhx6, another target of Fgf8 signaling, is also upregulated in the Hoxa2-/- second arch mesenchyme. These findings have important implications for the understanding of developmental processes in the branchial area and suggest a novel mechanism for mesenchymal patterning by Hox genes that acts to define the competence of mesenchymal cells to respond to skeletogenic signals (Bobola, 2003).
Proboscipedia related Hox interact with TALE class homeodomain proteins
Pbx/exd proteins modulate the DNA binding affinities and specificities of Hox proteins and contribute to the execution of
Hox-dependent developmental programs in arthropods and vertebrates. Pbx proteins also stably heterodimerize and bind DNA with Meis and Pknox1-Prep1 (see Drosophila Homothorax), additional members of the TALE (three-amino-acid loop extension) superclass of homeodomain proteins that function on common genetic pathways with a subset of Hox proteins. Pbx and Meis bind DNA as heterotrimeric complexes with Hoxb1 on a genetically defined Hoxb2 enhancer, r4, which mediates the cross-regulatory transcriptional
effects of Hoxb1 in vivo. The DNA binding specificity of the heterotrimeric complex for r4 is mediated by a Pbx-Hox site in conjunction with a distal Meis site, which is required for ternary complex formation and Meis-enhanced transcription. Formation of heterotrimeric complexes in which all three homeodomains bind their cognate DNA sites is topologically facilitated by the ability of Pbx and Meis to interact through their amino termini and bind DNA without stringent half-site
orientation and spacing requirements. Furthermore, Meis site mutation in the Hoxb2 enhancer phenocopies Pbx-Hox site mutation to abrogate enhancer-directed expression of a reporter transgene in the murine embryonic hindbrain, demonstrating that DNA binding by all three proteins is required for trimer function in vivo. These data provide in vitro and in vivo evidence for the combinatorial regulation of Hox and TALE protein functions that are mediated, in part, by their interdependent DNA binding activities as ternary complexes. As a consequence, Hoxb1 employs Pbx and Meis-related proteins, as a pair of essential cofactors in a higher-order molecular complex, to mediate its transcriptional effects on an endogenous Hox response element (Jacobs, 1999).
The observation that Meis contributes to the DNA binding requirements of ternary complexes differs from previous observations and raises the possibility that its
contributions may vary with different enhancers or under different cellular conditions. The most compelling evidence that DNA binding by a Meis-related protein is required for the in vivo function of ternary complexes is provided by the analysis of the
requirements for function of the Hoxb2 r4 enhancer in rhombomere 4 of the developing hindbrain. Elegant genetic studies have demonstrated that this enhancer
directs the appropriate expression of the Hoxb2 gene in response to Hoxb1 cross-regulation in rhombomere 4 at approximately 8.5 to 10 days of hindbrain
development. Extensive mapping has shown that a consensus Pbx-Hox site is essential for r4 enhancer-mediated expression in rhombomere 4, but not in
rhombomeres 3 and 5. These earlier studies also indicate that the Pbx-Hox site is sufficient for
r4-directed expression, a conclusion that conflicts with the current findings that mutation of the flanking Meis site phenocopies Pbx-Hox site mutation. This disparity
may be accounted for by the fact that the previous studies employed synthetic elements that were not in a natural configuration and that contained iterated copies of
Pbx-Hox sites. Since Meis crossbinds to Pbx-Hox consensus sites, synthetic elements containing them in tandem resemble the natural tripartite
Meis-Pbx-Hox elements identified here and, in fact, weakly support DNA binding by ternary Hoxb1 complexes in vitro. This may also account for
the Meis-mediated enhancement of the expression of reporter genes containing similar multimerized configurations of the ARE r3 site (Jacobs, 1999 and references).
The current studies demonstrate a
consistent requirement for the Meis site in vitro and in vivo, but it is not yet clear which of the various Meis-Prep1 family members may be directly responsible for r4
enhancer function in the developing hindbrain. Western blot analyses show that both Meis and Prep1 proteins are expressed in the hindbrain at embryonic day 9.5. Since Meis genes display dynamic expression profiles during embryonic development, a more precise determination of the in vivo roles of individual
Meis-related proteins in r4 functions will require studies with mice that are nullizygous for one or more of the Meis genes.
In summary, these studies provide support at the molecular level for previous observations that each component of the TALE heterodimer interacts and functions on
common genetic pathways with a subset of Hox proteins. Although its generality for Hox function remains to be determined, a trimeric model invoking a higher-order
assembly of Hox and TALE proteins provides a molecular framework for integrating the functions of these developmentally important proteins (Jacobs, 1999).
Proboscipedia related Hox genes and heart development
The anteroposterior (A-P) patterning of the developing heart underlies atrial and ventricular lineage specification and heart
chamber morphogenesis. The posteriorization of cardiomyogenic phenotype with retinoic acid (RA) treatment of primitive
streak stage chicken embryos is suggestive of a role for the clustered homeobox (Hox) genes in early heart patterning. A screen for Hox genes expressed in chick heart
primordia and primitive heart led to the isolation of anterior genes of the Hox clusters expressed during cardiogenesis. Specific
hoxd-3, hoxa-4, and hoxd-4 transcripts are detected at the early stages of heart formation and full-length cDNA clones have been
isolated. Expression of hoxd-3 is detected in the heart forming region of embryos prior to heart tube formation. Expression
of hoxa-4, hoxd-3, and hoxb-5 is increased in cardiogenic tissue treated with RA in culture conditions that also produced
changes in positionally restricted cardiomyogenic phenotypes. Hox genes expressed in cardiac explants exhibit distinct
sensitivities to RA and ouabain treatment, when compared to genes, such as nkx-2.5, that are involved in cardiac commitment
and differentiation. These studies support a role for Hox genes in early heart patterning and suggest that positional information
in the cardiogenic region is established by regulatory mechanisms distinct from early heart lineage specification (Searcy, 1998).
Divergent Hox genes related to Proboscipedia
A novel human homeobox gene, HB9, was isolated from a cDNA library prepared from in vitro
stimulated human tonsil B lymphocytes and from a human genomic library. The HB9 gene is
composed of 3 exons spread over 6 kilobases of DNA. The HB9 homeodomain is most similar to that of the proboscipedia. Transcripts are detected in RNA prepared from tonsil B cells: in situ hybridization studies localize them in the germinal center region of adult tonsil. These findings suggest the involvement of HB9 in regulating gene transcription in lymphoid and pancreatic tissues (Harrison, 1994).
gax is a relatively divergent Antp class homeobox gene with a homeodomain nearly identical to that of Mox-1, a recently described homeobox gene restricted to mesoderm and mesodermally derived tissue. Gax is less homologous to either proboscipedia (66% identity in the homeodomain) or to Deformed (61% identity in the homeodomain. gax is expressed in vascular smooth muscle cells (VSMCs) and
is down-regulated in vitro by mitogen stimulation and in vivo in response to vascular
injury that leads to cellular proliferation. Recombinant Gax protein microinjected into
VSMCs and fibroblasts inhibited the mitogen-induced entry into S-phase when
introduced either during quiescence or early stages of G1. Overexpression of gax with
a replication-defective adenovirus vector results in G0/G1 cell cycle arrest of
VSMCs and fibroblasts. The gax-induced growth inhibition correlates with a
p53-independent up-regulation of the cyclin-dependent kinase inhibitor p21. Gax
overexpression also leads to an association of p21 with cdk2 complexes and a decrease
in cdk2 activity. Fibroblasts deficient in p21 are not susceptible to a reduction in cdk2
activity or growth inhibition by gax overexpression. Localized delivery of the virus to
denuded rat carotid arteries significantly reduces neointima formation and luminal
narrowing. These data indicate that gax overexpression can inhibit cell proliferation in
a p21-dependent manner and can modulate injury-induced changes in vessel wall
morphology that result from excessive cellular proliferation (Smith, 1997).
Transcriptional regulation of Proboscipedia homologs
Continued: proboscipedia Evolutionary homologs part 2/3
| part 3/3
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
proboscipedia:
Biological Overview
| Regulation
| Developmental Biology
| Effects of Mutation
| References
Society for Developmental Biology's Web server.
{kind=link}