|
Gene name - labial Synonyms - Cytological map position - 84A1,2 Function - transcription factor Keywords - homeotic - Antennapedia complex |
Symbol - lab FlyBase ID:FBgn0002522 Genetic map position - 3-[47.5] Classification - homeodomain - Antp class Cellular location - nuclear |
| Recent literature | Billes, V., et al. (2018). Developmentally regulated autophagy is required for eye formation in Drosophila. Autophagy. PubMed ID: 29940806
Summary: This study demonstrates that autophagy, an evolutionarily conserved self-degradation process of eukaryotic cells, is essential for eye development in Drosophila. Autophagic structures accumulate in a specific pattern in the developing eye disc, predominantly in the morphogenetic furrow (MF) and differentiation zone. Silencing of several autophagy genes (Atg) in the eye primordium severely affects the morphology of the adult eye through triggering ectopic cell death. In Atg mutant genetic backgrounds however genetic compensatory mechanisms largely rescue autophagic activity in, and thereby normal morphogenesis of, this organ. The results also show that in the eye disc the expression of a key autophagy gene, Atg8a, is controlled in a complex manner by the anterior Hox paralog lab (labial), a master regulator of early development. Atg8a transcription is repressed in front of, while activated along, the MF by labial. The amount of autophagic structures then remains elevated behind the moving MF. These results indicate that eye development in Drosophila depends on the cell death-suppressing and differentiating effects of the autophagic process. This novel, developmentally regulated function of autophagy in the morphogenesis of the compound eye may shed light on a more fundamental role for cellular self-digestion in differentiation and organ formation than previously thought. |
Singh, N. P., De Kumar, B., Paulson, A., Parrish, M. E., Zhang, Y., Florens, L., Conaway, J. W., Si, K. and Krumlauf, R. (2020). A six-amino-acid motif is a major determinant in functional evolution of HOX1 proteins. Genes Dev. PubMed ID: 33184220
Summary: Gene duplication and divergence is a major driver in the emergence of evolutionary novelties. How variations in amino acid sequences lead to loss of ancestral activity and functional diversification of proteins is poorly understood. This study used cross-species functional analysis of Drosophila Labial and its mouse HOX1 orthologs (HOXA1, HOXB1, and HOXD1) as a paradigm to address this issue. Mouse HOX1 proteins display low (30%) sequence similarity with Drosophila Labial. However, substituting endogenous Labial with the mouse proteins revealed that HOXA1 has retained essential ancestral functions of Labial, while HOXB1 and HOXD1 have diverged. Genome-wide analysis demonstrated similar DNA-binding patterns of HOXA1 and Labial in mouse cells, while HOXB1 binds to distinct targets. Compared with HOXB1, HOXA1 shows an enrichment in co-occupancy with PBX (see Drosophila Extradenticle) proteins on target sites and exists in the same complex with PBX on chromatin. Functional analysis of HOXA1-HOXB1 chimeric proteins uncovered a novel six-amino-acid C-terminal motif (CTM) flanking the homeodomain that serves as a major determinant of ancestral activity. In vitro DNA-binding experiments and structural prediction show that CTM provides an important domain for interaction of HOXA1 proteins with PBX. These findings show that small changes outside of highly conserved DNA-binding regions can lead to profound changes in protein function. |
Labial has two roles in morphogenesis, one in the developing head and a second in the midgut. There is no expression of labial in the labial lobes, suggesting that labial inhibits labial lobe development in the procephalon [Images], an area where labial is expressed. (Diederich, 1991).
labial is one of the five homeotic genes comprising the Antennapedia complex (ANTP-C). Genes in the ANTP-C are spacially ordered in a linkage array, in the same order in which they find expression, anterior to posterior. labial is expressed in the anterior of the fly, and is the first one in the order of genes arrayed in the ANTP-C linkage.
It is believed that labial expression marks the intercalary segment, positioned between the procephalic lobe and the gnathocephalon (the "jaw"), consisting of mandibular, maxillary and labial lobes. A majority of labial expressing cells delaminate from the ectoderm surface to incorporate at the base of each brain lobe, a region that corresponds to the intercalary segment. Most of the remaining cells constitute the epidermis of the procephalic lobe and evenually incorporate into the dorsal pouch (frontal sac), the cells of which secrete the dorsal arms of the cephalopharyngeal skeleton (Diederich, 1991).
Labial production in the midgut is coincident with copper cells. These cells may adsorb metal ions from the gut lumin (Diederich, 1989 and Hoppler, 1994). Expression in the midgut is under control of DPP, secreted from visceral mesodermal cells (Staehling-Hampton, 1994).
To gain further insights into homeotic gene action during CNS development, the role of the homeotic genes was characterized in embryonic brain development of Drosophila. Neuroanatomical techniques were used to map the entire anteroposterior order of homeotic gene expression in the Drosophila CNS. This order is virtually identical in the CNS of Drosophila and mammals. All five genes of the Antennapedia Complex are expressed in specific domains of the developing brain. The labial gene has the smallest spatial expression domain; it is only expressed in the posterior part of the tritocerebral anlage. This contrasts with previous reports that lab is expressed throughout the tritocerebral (intercalary) neuromere. The proboscipedia gene has the largest anteroposterior extent of expression, however, in contrast to other homeotic genes, pb is only found in small segmentally repeated groups of 15-20 cells per neuromere. These groups of pb-expressing cells range from the posterior deutocerebrum toward the end of the VNC. Since pb-expressing cells are found anterior to the lab-expressing cells in the brain, this is an exception to the spatial colinearity rule. (Spatial colinearity is conserved in the epidermis, where pb expression is posterior to lab expression). The Deformed gene is expressed in the mandibular neuromere and the anterior half of the maxillary neuromere and the Sex combs reduced gene is expressed in the posterior half of the maxillary neuromere and the anterior half of the labial neuromere. The Antennapedia gene is expressed in a broad domain from the posterior half of the labial neuromere toward the end of the VNC. The three genes of the Bithorax Complex are expressed in the VNC. Ultrabithorax gene expression extends in a broad domain from the posterior half of the T2 neuromere to the anterior half of the A7 neuromere, with highest expression levels in the posterior T3/anterior A1 neuromeres. The abdominal-A gene is expressed from the posterior half of the A1 neuromere to the posterior half of the A7 neuromere. For the above mentioned genes, the anterior border of CNS expression remains stable from stage 11/12 until the end of embryogenesis. In contrast, the anterior border of CNS expression for the Abdominal-B gene shifts at stage 14. Before this stage Abd-B expression extends from the posterior half of neuromere A7 to the end of the VNC; afterwards, it extends from the posterior half of neuromere A5 to the end of the VNC with the most intense expression localized to the terminal neuromeres. With the exception of the Dfd gene, the anterior limit of homeotic gene expression in the CNS is always parasegmental (Hirth, 1998).
A genetic analysis of the labial gene in embryonic brain development was carried out. Loss-of-function mutation and ubiquitous overexpression of labial results in ectopic expression of neighboring regulatory genes. The tritocerebral commissure, which interconnects the brain hemispheres at the level of the tritocerebrum, is completely absent in labial mutants. Moreover, the frontal connectives no longer project into the tritocerebral neuromere, rather they grow ectopically into the more anterior brain neuromeres. This analysis demonstrates that mutational inactivation of labial results in regionalized axonal patterning defects that are due to both cell-autonomous and cell-nonautonomous effects. Thus, in the absence of labial, mutant cells are generated and positioned correctly in the brain, but these cells do not extend axons. Additionally, extending axons of neighboring wild-type neurons stop at the mutant domains or project ectopically, and defective commissural and longitudinal pathways result. Immunocytochemical analysis demonstrates that cells in the mutant domains do not express neuronal markers, indicating a complete lack of neuronal identity. An alternative glial identity is not adopted by these mutant cells. To determine if the defects in lab loss-of-function mutants correlate with altered homeotic gene expression patterns in the tritocerebrum, molecular markers were used to study the expression of the homeotic genes pb, Dfd and Scr in lab null mutants (Hirth, 1998).
Also studied was the expression of the empty spiracles (ems) gene, which in the wild-type brain is expressed in a large domain anterior to lab . In lab loss-of-function mutants, the ems gene is expressed ectopically in the tritocerebral domain in which lab is normally expressed; this ectopic ems expression occurs with 100% penetrance and ranges from 5-7 cells per hemisegment. The expression of pb disappears in the deutocerebrum and tritocerebrum of lab loss-of-function mutants but not in more posterior neuromeres. In contrast, the expression patterns of Dfd and Scr remain unaltered. Thus, in the tritocerebral domain in which lab is normally expressed, two changes in regulatory gene expression occur: activation of ems and inactivation of pb. To determine if ubiquitous overexpression of labial also alters regulatory gene expression patterns in the tritocerebral domain, transgenic flies were used carrying the lab gene under control of a heat-inducible promoter. In these mutants, ubiquitous overexpression of lab following heat-shock results in ectopic expression of the posteriorly active Dfd gene in the posterior tritocerebrum; this occurred with 100% penetrance when the heat shock was given around embryonic stage 10/11 (Hirth, 1998).
Cell-autonomous and cell-nonautonomous axogenesis defects occur in lab mutants. This suggests that mutation of the lab gene leads to cell-autonomous as well as cell-nonautonomous defects in brain axogenesis. Evidence for cell-autonomous defects in axogenesis is provided by combining anti-tubulin immunocytochemistry together with lab-lacZ reporter gene expression. In the wild type, anti-tubulin immunostaining reveals the axons of the normal tritocerebral commissure that extend from the lab-lacZ-expressing neurons of the tritocerebrum. Anti-tubulin immunostaining also shows longitudinal axon fascicles in the tritocerebral domain. In the lab null mutant, there are no labeled commissural axons projecting from the lab-lacZ-expressing neurons of the tritocerebrum; the tritocerebral commissure is deleted. Moreover, there are no labeled longitudinal axon fascicles in the mutant tritocerebral domain. It is concluded that the cells in the mutant domain of the tritocerebrum do not generate commissural or longitudinal axons. Although there are no axons associated with the mutant domain, some `microspike'-like tubulin labeling does appear there. Evidence for cell-nonautonomous defects in axogenesis comes from the analysis of Fasciclin II-expressing descending and ascending axons from other parts of the brain. In the wild type, these axons project through the lab domain of the tritocerebrum in well-formed fascicles. In lab mutants, the fasciclin axons do not project through the mutant domain. They either stop at the border of the mutant domain or they avoid this domain and extend ectopically. The fact that the lab mutant cells do not extend axons suggests that these cells might not differentiate into neurons (Hirth, 1998).
It was determined that cells with the characteristics of neuroblasts and ganglion mother cells (GMCs) are present in the mutant tritocerebral domain (labeled by lab-lacZ reporter gene expression). Cells that have the characteristic position and morphology of wild-type lab-expressing neuroblasts, and which also express the neuroblast markers hunchback and asense, are present in the mutant tritocerebral domain. Similarly, cells that have the characteristic position and morphology of wild-type lab-expressing GMCs, and which also express the GMC marker prospero, are also present in the mutant tritocerebral domain (Hirth, 1998).
The expression of several neuronal molecular markers were studied in the posterior tritocerebral domain of wild-type and lab mutant embryonic brains. In these experiments, the position of the mutant tritocerebral domain was always determined by lab-lacZ reporter gene expression. The neuron-specific RNA-binding protein ELAV is expressed exclusively in neurons of the CNS beginning from the time of birth of the first neurons. In the wild type, anti-ELAV immunostaining reveals all of the neurons in the embryonic brain including the neurons in the tritocerebral lab domain. Anti-ELAV immunostaining is found in neuronal cell bodies and, in contrast to anti-HRP immunolabeling, does not stain the neuropil. In lab mutants, anti-ELAV immunostaining is no longer seen in any of the mutant cells in the posterior tritocerebral domain, but continues to be expressed in all of the other neuronal cells of the embryonic brain. The cell adhesion molecule Fasciclin II is expressed in the wild type by a subset of the Labial-positive neurons in the tritocerebrum and by the axons that these neurons project into the tritocerebral commissure. In lab mutants, none of the cells in the corresponding posterior tritocerebral domain and none of the axons in that region express Fasciclin II (Hirth, 1998).
Immunocytochemical analysis shows that a number of other molecular labels are present in the tritocerebral lab domain of the wild type, but are absent in the corresponding tritocerebral domain of lab mutants. Among these are the LIM-homeodomain gene islet, the neuron-specific NAC epitope recognized by the anti-HRP antibody, the axon-specific BP102 epitope and the segment polarity gene engrailed. Taken together, these findings imply that the lab mutant cells have not acquired a neuronal identity. This suggests that the lab mutant cells either fail to differentiate into neurons or adopt another cell fate such as that of glial cells. To determine if these cells do differentiate into glia, the expression of the glial-specific repo gene was studied in the lab mutant tritocerebral domain. The pattern and number of repo-expressing cells in the tritocerebral labial domain are similar in the wild type and in the labial null mutant. This indicates that the lab mutant cells have not acquired a glial identity. It also suggests that gliogenesis is not affected in the lab mutant domain. It is therefore postulated that the lab gene is necessary for the establishment of correct neuronal cell fate, but not glial cell fate, in the part of the developing brain in which it is normally expressed (Hirth, 1998).
Comparable effects to those seen in labial mutants are seen in Deformed mutants but not in other homeotic gene mutants. In Dfd mutants the longitudinal pathways that normally project through the mandibular neuromere are missing or reduced. The mandibular commissure, which interconnects the mandibular hemineuromeres, is completely absent. The cells in the mandibular neuromere are generated in Dfd mutants. Homeotic gene expression patterns are altered in Dfd mutants; in the null mutant, lab is ectopically expressed in the mandibular segment in which Dfd is normally expressed. Ubiquitous overexpression of Dfd results in ectopic expression of the posteriorly active Scr gene in the Dfd domain. Cell-autonomous and cell-nonautonomous axogenesis defects occur in the Dfd mutants; mutant cells do not project axons, and descending and ascending axons from other parts of the brain do not project through the mutant domain. The Dfd mutant cells of the mandibular neuromere show a loss of neuronal markers such as ELAV, whereas the pattern and number of repo-expressing glial cells in the mutant domain is unaffected. These findings demonstrate that the action of the homeotic genes labial and Deformed are required for neuronal differentiation in the developing brain of Drosophila (Hirth, 1998).
The labial gene is the most proximal (first in order) of the Antennapedia complex
Bases in 5' UTR - 238
Exons - three
Bases in 3' UTR - 701
Labial has an antennapedia-type homeodomain, and an opa activation domain (Diederich, 1989).
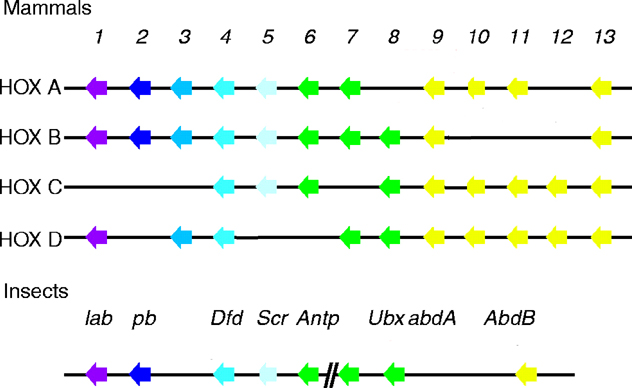
See four paralogous Hox clusters of mammals for homologies of Labial with mammalian Hox cluster proteins.
date revised: 7 July 98
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
{kind=link}