The Interactive Fly
Zygotically transcribed genes
INDEX
Mechanisms composing Drosophila's clock are conserved within the animal kingdom. To learn how such clocks influence behavioral and physiological rhythms, the complement of circadian transcripts in adult Drosophila heads was determined. High-density oligonucleotide arrays were used to collect data in the form of three 12-point time course experiments spanning a total of 6 days. Analyses of 24 hr Fourier components of the expression patterns revealed significant oscillations for ~400 transcripts. Based on secondary filters and experimental verifications, a subset of 158 genes showed particularly robust cycling and many oscillatory phases. Circadian expression is associated with genes involved in diverse biological processes, including learning and memory/synapse function, vision, olfaction, locomotion, detoxification, and areas of metabolism. Data collected from three different clock mutants (per0, tim01, and ClkJrk), are consistent with both known and novel regulatory mechanisms controlling circadian transcription (Claridge-Chang, 2001).
A genome-wide expression analysis was performed aimed at identifying all transcripts from the fruit fly head that exhibit circadian oscillations in their expression. By taking time points every 4 hr, a data set was obtained that has a high enough sampling rate to reliably extract 24 hr Fourier components. Time course experiments spanning a day of entrainment followed by a day of free-running were performed to take advantage of both the self-sustaining property of circadian patterns and the improved amplitude and synchrony of circadian patterns found during entrainment. 36 RNA isolates from wild-type adult fruit fly heads, representing three 2 day time courses, were analyzed on high-density oligonucleotide arrays. Each array contained 14,010 probe sets (each composed of 14 pairs of oligonucleotide features) including ~13,600 genes annotated from complete sequence determination of the Drosophila genome. To identify different regulatory patterns underlying circadian transcript oscillations, four-point time course data was colleced from three strains of mutant flies with defects in clock genes (per0, tim01, and ClkJrk) during a single day of entrainment. Because all previously known clock-controlled genes cease to oscillate in these mutants but exhibit changes in their average absolute expression levels, the analysis of the mutant data was focused on changes in absolute expression levels rather than on evaluations of periodicity (Claridge-Chang, 2001).
To organize the 158 statistically significant circadian transcripts in a way that was informed by the data, hierarchical clustering was performed. Both the log ratio wild-type data (normalized per experiment) and the log ratios for each of the three clock mutants (normalized to the entire data set) were included to achieve clusters that have both a more or less uniform phase and a uniform pattern of responses to defects in the circadian clock. One of the most interesting clusters generated by this organization is the per cluster. This cluster contains genes that have an expression peak around ZT16 and a tendency to be reduced in expression in the ClkJrk mutant. Strikingly, all genes previously known to show this pattern of oscillation (per, tim, vri) are found in this cluster. In fact, the tim gene, which has multiple representations on the oligonucleotide arrays, has two independent representations in this cluster. Together with the novel oscillator CG5798, per, tim, and vri form a subcluster (average phase ZT14) that shows upregulation in both the per0 and tim01 mutants. The fact that per, tim, and vri all function in the central circadian clock raises the possibility that several other genes from this cluster, including the ubiquitin thiolesterase gene CG5798 and the gene coding for the channel modulator Slowpoke binding protein (Slob) may function in the circadian clock or directly downstream of it (Claridge-Chang, 2001).
The genes in a second cluster (Clock cluster are primarily grouped together based on their peak phase (average phase ZT2). By virtue of the mutant expression data, several subclusters within this phase group can be identified. The known circadian genes Clock and takeout (to) are part of this cluster. Clk is found in a clustered pair with the leucyl aminopeptidase gene CG9285. In terms of chromosomal organization, to, CG11891, and CG10513 map closely together on chromosome 3R. Two additional circadian genes in this chromosomal region (CG11852, CG1055). Interestingly, the Clk cluster contains three pairs of homologous genes with very similar expression patterns: the UDP-glycosyl transferase genes Ugt35a and Ugt35b, the enteropeptidase genes CG9645 and CG9649, and the long-chain fatty acid transporter genes CG6178 and CG11407. In the first two cases, the homologous genes are also directly adjacent to each other on the chromosome. An overview of the map positions of all circadian genes in this study is available as supplemental information online (http://www.neuron.org/cgi/content/full/32/4/657/DC1). Apart from Ugt35a and Ugt35b, several other genes with a predicted function in detoxification are members of the Clk cluster (CG17524, CG8993, CG3174, Cyp6a21). It may also be noteworthy that the genes for three oxidoreductases found in this group [Photoreceptor dehydrogenase (Pdh), CG15093, CG12116] have almost identical phases (ZT3) (Claridge-Chang, 2001).
All genes of the apterous (ap) cluster are defined by both the oscillatory phase of their expression pattern (average phase ZT17) and by a distinct expression profile in the three clock mutants. Although the 6 hr sampling interval in the mutant data makes it difficult to reliably detect oscillations, it seems that the majority of the genes in this cluster shows some degree of periodicity in the three mutant light-dark regime (LD) time courses. Although it cannot be ruled out that there are circadian oscillations independent from the known clock genes, the hypothesis that there may be a light-driven response underlying the observed mutant expression pattern is favored. The genes in this group may, therefore, be regulated not only by the circadian clock, but also by a direct light-dependent mechanism. It should be mentioned that evidence of gene expression patterns that are purely light-driven in wild-type flies was sought, but little indication was found of such regulation. Instead, genes with both a strong light-driven oscillation and a weak circadian component were encountered. apterous (ap) encodes a LIM-homeobox transcription factor, which is known to act both in neural development and in neuropeptide expression. The ap cluster includes the genes for the transcription factor moira, the synaptic regulator syndapin, two septins (Sep1 and CG9699), and two ATP binding cassette (ABC) transporters (CG6162, CG9990). In terms of chromosomal organization, CG6166, the gene adjacent to CG6162 on chromosome 3R is homologous to CG9990 and coregulated with CG6162 and CG9990 (Claridge-Chang, 2001).
The founding member of the fourth cluster, ebony (e), encodes β-alanyl-dopamine synthase and has roles in both cuticle tanning and regulating circadian locomotor behavior. Among the other cluster members are six genes that function in protein cleavage (CG9377, Ser99Da, SP1029, CG7828, CG11531, BcDNA:GH02435), three transcription factor genes (CG15632, CG17257, CG6755), as well as two genes each that act in signal transduction (prune, loco), the cytoskeleton (TpnC47D, Chd64), and lipid metabolism (ATPCL, CG1583) (Claridge-Chang, 2001).
The per0, tim01, and ClkJrk mutations affect genes that are essential for maintaining circadian rhythms and result in both molecular and behavioral arrhythmicity. In addition to abrogating the oscillations of the per, tim, vri, to, and Clk transcripts, these mutations also affect their absolute levels of expression. per0 and tim01 flies have intermediate or somewhat elevated levels of per, tim, vri, and to transcript, and decreased levels of Clk transcript whereas ClkJrk mutants have the opposite effect. Based on these observations, genome-wide expression data was gathered from per0, tim01, and ClkJrk mutant flies in three separate four-point time course experiments. A rank-sum Wilcoxon test was employed to determine if any of the interrogated transcripts were significantly up- or down-regulated in any of the mutants when compared to the total wild-type expression data set (Claridge-Chang, 2001).
Out of the 14010 probe sets on the arrays, 4865 showed up- or down-regulation in one or more of the three mutants with a p value lower than 0.05; 2544 were significantly changed in the tim01 flies; 1810 were significantly different in per0, and 2181 in ClkJrk. It is unclear what proportion of these changes depends on the actual mutations themselves, because (1) the three mutant fly strains have different genetic backgrounds and (2) data was collected for only one population of each mutant strain. Although there are known examples of noncircadian genes whose expression is affected by clock mutations, it was decided that it would be more informative to consider effects of the clock mutations only with respect to the subset of 158 strong oscillators. Among this set, 72 genes were found with one or more significant expression changes in the three clock mutant strains (Claridge-Chang, 2001).
Included in the regulated set are tim (twice independently), vri, to, and Clk, and their patterns agree with previously published observations. The hierarchical clustergram shows four basic patterns of regulation: (type I) increased in per0 and tim01 but decreased in ClkJrk (e.g., vri, CG5798); (type II) decreased in per0 and tim01 but increased in ClkJrk (e.g., Clk, CG15447); (type III) decreased in all three mutants (e.g., Ugt36Bc/CG17932, ea), and (type IV) increased in all three mutants (e.g., Pdh, CG11891). Type I and II match the two known modes of regulation for circadian genes. The behavioral and molecular phenotypes of the per0 and tim01 mutations are almost identical. It may, therefore, be relevant that no circadian genes are found that are significantly upregulated in per0 and significantly downregulated in tim01 or vice versa. Apart from genes that were a priori predicted to have expression patterns of type I (vri, tim, to) or II (Clk), novel genes were found for each of these two expression patterns. The average phases for the type I and type II subclusters are, respectively, ZT12 and ZT7, but there is large variation in phase among the members of each of these subclusters. to is in the type I subcluster and has a phase peak at ZT2, whereas CG15447 is in the type II subcluster and peaks at ZT10. This phenomenon of phase differences among transcripts with a similar response to clock defects has been described previously for type I regulation in the case of to. Here, a similar phenomenon was detected for genes with Clk-like type II regulation. Type III and IV predict a novel and unexpected response to the circadian mutants (Claridge-Chang, 2001).
The promoter sequences of the set of 158 genes was tested for the presence of known and candidate circadian enhancer motifs. The results suggest that in fact this set is enriched in such elements. For example, the frequency of E boxes in the set of oscillators (42 hits in total) is significantly higher than the frequency in random selections of genes. The significance of this result does, however, depend on the inclusion of well-studied genes (per, tim, vri). Some novel oscillators with remarkable frequencies of 'circadian transcription elements' are loco (1 PDP1 site; 3 W boxes; 8 CRE elements; 3 TER elements); trpl (1 E box; 1 PDP1 site; 9 W boxes; 6 CRE elements; 12 TER elements); Rh5 (5 W boxes; 5 CRE elements; 6 TER elements), and Slob (1 E box; 1 PERR element; 2 PDP1 sites; 6 W boxes; 15 TER elements). It is noteworthy that many robustly cycling genes have no known circadian transcription elements in their promoters or first introns (Claridge-Chang, 2001).
The set of 158 circadian genes were organized according to annotated or predicted molecular function. Several of these functional classes may provide insights into pathways influencing rhythmic behavior (Claridge-Chang, 2001).
Synaptic Transmission and Plasticity
An emerging theory of the function of sleep postulates that it is
required for neural plasticity, synaptic maintenance, and remodelling.
Behaviorally defined sleep has been identified in the fly, with
behavioral recordings in LD indicating increased rest during the dark
phase. The assay for circadian expression uncovered a number of genes
known to be involved in synaptic function and synaptic plasticity
(Claridge-Chang, 2001).
Three oscillating trancripts encode synaptic vesicle endocytosis factors: ß-adaptin (Bap), AP-1gamma, and syndapin. The first two are adaptors between the budding membrane and clathrin lattice, while syndapin is thought to connect vesicle endocytosis to actin. Clock modulation of the synaptic vesicle pool is consistent with the idea of modulated synaptic function, although it is unclear what effect raising or lowering endocytotic factors would have on synaptic function (Claridge-Chang, 2001).
The Slob transcript peaks at ZT15 and is downregulated in the ClkJrk mutant, suggesting that CLK acts as an activator of Slob. There is an E box 5.4 kb upstream of the Slob transcriptional start site raising the possibility that CLK acts directly on the Slob promoter (Claridge-Chang, 2001).
If the cycling transcript can be shown to produce oscillating Slob protein, this could indicate a potent mechanism for rhythmic control of synaptic function, including synaptic plasticity: Slob protein binds the calcium-dependent potassium channel Slowpoke (Slo) and has been shown to increase Slo activity and voltage sensitivity. Slob is in turn bound by a second channel regulator, Leonardo. Hypomorphic mutations of leonardo produce defects in learning, and electrophysiological analyses of the larval neuromuscular junction (NMJ) in these mutants show presynaptic function and plasticity is greatly impaired in these animals. In contrast to Slob, Leonardo is a strong inhibitor of Slo, but requires Slob for this interaction. All three proteins colocalize to the presynaptic bouton at larval NMJs. Thus, Slob may contribute to a switching mechanism that ultimately places Slo channel activity under circadian control (Claridge-Chang, 2001).
Slo channels are widely expressed in the adult fly head, including the eye, lamina, medulla, central brain, and mushroom bodies, but it is not known which subset of these areas contain oscillating Slob expression. In situ hybridization was performed with larval brains to localize Slob RNA expression. Prominent staining was observed in a restricted region consistent with placement of the developing mushroom body. The staining also corresponds well with that region of the larval brain receiving PDF-rich projections of the circadian pacemaker cells, the lateral neurons. In future studies, it will be important to determine whether presence of the innervating LNs is required for cycling Slob expression (Claridge-Chang, 2001).
leonardo was initially implicated in presynaptic function by the effect of mutations on learning. Mutations of latheo also cause learning defects, and this protein is also found at larval NMJs. Lowered latheo function has been associated with elevated synaptic transmission and reduced synaptic plasticity. latheo shows cycling expression with peak accumulation at ZT12-15. Rhythmicity was detected in the expression of dunce and Calpain-B genes involved in learning and synaptic long-term potentiation, respectively (Claridge-Chang, 2001).
Amine Neurotransmitter-Related Functions
Two serotonin receptor transcripts, 5-HT2
and 5-HT1A,
were found to oscillate with phases of ZT15 and ZT18, respectively.
Serotonin is known to be involved in a variety of neuronal processes
in animals, including synaptic plasticity, clock entrainment, and
mating behavior. The 104 serotonergic neurons in the adult CNS have
been mapped, but no studies have been done of either 5-HT receptor
localization or receptor mutant phenotypes. Neither of these receptors
are orthologs of the mammalian 5-HT receptor implicated in photic
clock entrainment; this would be represented by theDrosophila 5-HT7
gene. 5-HT1A belongs in a class of receptors that respond to agonists
by decreasing cellular cAMP, while 5-HT2 is homologous to mammalian
receptors whose main mode of action involves activation of
phospholipase C, a function involved in synaptic plasticity
(Claridge-Chang, 2001).
The ebony transcript was found to oscillate, showing a peak of expression around ZT5. ebony oscillation connects with a body of earlier evidence linking ebony to circadian activity rhythms. ebony hypomorphs show severe defects in circadian rhythm including arrhythmicity/aberrant periodicity in the free-running condition, as well as abnormal activity patterns in LD conditions. Ebony is a putative ß-alanyl dopamine synthetase, and hypomorphs show elevated levels of dopamine. Dopamine has been implicated in control of motor behavior, since it induces reflexive locomotion in decapitated flies, and this response is under circadian control. The results suggest that oscillations of ebony contribute to the assembly of rhythmic locomotor behavior. Other evidence points to a role in clock resetting. In addition to impaired entrainment in LD, ebony flies show an abnormal ERG, and ebony is strongly expressed in the lamina and the medulla optic neuropile, a region associated with vision rather than motor control (Claridge-Chang, 2001).
Vision
The Drosophila eye is both a likely target of clock control and partly
responsible for photic input to the central pacemaker. Several genes
found to oscillate by microarray assay are components of visual
processes (Claridge-Chang, 2001).
Photoreceptor cells contain peripheral clocks, suggesting that visual function may be regulated by the clock. In vertebrates, the synthesis of various visual components is known to be under circadian control. In Drosophila, electroretinogram (ERG) measurements of visual sensitivity reveal a 4-fold cycle in sensitivity, with a minimum at ZT4 and a broad peak around lights off (ZT12). This suggests that some of the fly visual components are clock controlled. However, a previous study of five major phototransduction components found no cycling of either mRNA or protein. In this genome-wide assay, the trpl transcript was found to oscillate with peak expression at ZT11. TRPL is one of two ion channels in the visual transduction pathway, along with TRP, a paralog. TRPL and TRP open in response to a G protein-coupled phosphoinositide cascade that is initiated by the isomerization of rhodopsin by light. Their opening produces the light-sensitive conductance in the photoreceptors. Amorphic mutants of each channel show visual defects, while the double null genotype results in a blind fly. A cycling trpl transcript could contribute to the visual sensitivity cycle: (1) the two phenomena are in the same phase with both sensitivity and trpl expression peaking around lights-off; (2) it is estimated that TRPL contributes about half of the wild-type conductance, allowing for a substantial range of modulation by reducing TRPL function. Another possible role for oscillating TRPL function is connected with circadian entrainment. In addition to being blind, trp/trpl double null mutants show reduced circadian behavioral resetting and less TIM degradation in response to light pulses. It is noted that the established entrainment factor CRY is known to cycle and that interactions of CRY and TIM are essential for light-dependent TIM degradation. The oscillating, clock-related protein VIVID has also been shown to regulate photo-entrainment in Neurospora (Claridge-Chang, 2001).
The microarray experiments show that two opsin genes are under circadian control: Rh5 and Rh4. The Rh5 mRNA rhythm peaks at ~ZT 21, while the Rh4 array data show a circadian pattern with a peak 4 hr later, at ZT1. Rh5 is a blue-absorbing rhodopsin expressed in a subset of R8 cells at the base of the retina, while Rh4 is a UV-absorbing pigment expressed in apical R7 cells. Rh5 is never expressed in an R8 cell underlying an Rh4-expressing R7 cell, so in this way all ommatidia would contain one cycling rhodopsin. In terms of regulating sensory receptiveness to light, it is unclear why these two opsins should be targets for clock control. The major blue rhodopsin Rh1 does not cycle so it seems unlikely that an oscillation in these two minor pigments would produce overall tuning of the sensitivity of the fly visual system (Claridge-Chang, 2001).
NinaA is a rhodopsin chaperone and is required to move Rh1 from the endoplasmic reticulum (ER) to the rhabdomeric membrane. ninaA mutants display aberrant accumulation of Rh1 protein in the ER. ninaA mRNA shows cycling expression in fly heads by both array and Northern blot, with peak expression around ZT2. It is tempting to hypothesize that early morning ninaA upregulation would have the effect of releasing a reservoir of Rh1 from the ER, making it available for use in visual transduction. However, a previous assay of NinaA levels in an LD regime revealed no oscillation in protein levels. If true, this would represent a surprising example of a robustly oscillating mRNA producing a constitutive cognate protein. Finally, although its function is still unknown, Photoreceptor dehydrogenase is a robustly oscillating transcript with an extremely high level of expression in the screening pigment cells of the eye (Claridge-Chang, 2001).
Rhythmic Proteases and Accessory Factors
Fifteen of the identified oscillatory genes are implicated in protein
cleavage. CG7828 and BcDNA:GH02435
(peak phase ZT6 and ZT10, respectively) mediate ubiquitination of
protein substrates, thus targeting them for degradation. Conversely,
CG5798 and CG7288 cycle with a peak at respectively, ZT14 and ZT16,
and each produces a ubiquitin specific protease (Ubp;
cleaves ubiquitin from ubiquitin-protein conjugates) that may act to
prevent the degradation of specific protein targets. Two
metalloprotease genes, two aminopeptidase genes, and five serine
peptidases show circadian oscillation. Four of the five serine
peptidase genes cycle with a peak phase between ZT4-7. This profusion
of oscillating proteases suggests that circadian proteolysis may
represent a broad mechanism of clock control, both of clock components
themselves, as well as output factors (Claridge-Chang, 2001).
While circadian transcriptional mechanisms are relatively well understood, less is known about posttranslational mechanisms of circadian regulation. Proteases are known to be involved in circadian control of the degradation of some central clock components, and the clock proteins PER, TIM, CLK, CRY, and VRI are all known to undergo daily cycles of protein accumulation. Degradation of TIM is responsible for photic resetting of the Drosophila clock. This is thought to be mediated by interaction with CRY, followed by ubiquitination and proteasome-dependent loss of TIM. Nothing is known about the factors mediating TIM degradation in the dark, yet patterns of CG5798 expression may be of interest in this regard as this gene encodes a cycling Ubp whose peak expression (ZT14) immediately precedes an interval of rapid TIM accumulation in pacemaker cells (Claridge-Chang, 2001).
A likely clock-related target of one or more proteases is the neuropeptide PDF, whose regulation may be crucial to linking the clock to behavior. While pdf RNA is expressed constitutively, the peptide accumulates rhythmically under indirect control of the clock gene vri. This mechanism has not been further explored, but a clear possibility is that a PDF propeptide is cleaved rhythmically, allowing cyclical release of active PDF. Of the 15 cycling proteases suggested by this study, CG4723 may be of special interest due to its inclusion in a class of proteases known to cleave neuropeptides. The phase of CG4723 expression, ZT4, also coincides with that of PDF immunoreactivity in the Drosophila head (Claridge-Chang, 2001).
Detoxification and Oxidative Stress
One hypothesis of sleep characterizes it as a cellular detoxification
and repair process. Twelve genes were found that have a predicted role
in detoxification. Three additional redox enzymes may also participate
in this process. Toxins are initially modified into reactive species
by reductases/dehydrogenases and cytochrome P450 molecules. Then
glutathione-S-transferases (GSTs) and UDP-glycosyl transferases add
polar groups to the substrates to render them hydrophilic for
elimination by secretion. Four results are noteworthy: (1) following
this pathway, the genes for three circadian dehydrogenases: Pdh,
CG10593, and CG12116, were found.; (2) both morning and night
cytochrome P450 genes (Cyp6a21 and Cyp305a1) were found
to peak early in the day (ZT0 and ZT5), whereas Cyp18a1 and Cyp4d21
peak at approximately the same time late at night (ZT18 and ZT19); (3)
while CG17524 is the only GST gene found in the core set, it was
noticed that two other members of the GST type III gene cluster on
chromosome 2R show 24 hr periodicity (CG17523; CG17527) and (4)
UDP-glycosyl transferase genes are represented in the circadian set by
Ugt35a, Ugt35b, and Ugt36Bc. Of these, Ugt35b
encodes an antennal specific transcript with a potential role in
olfaction, whereas Ugt35a is expressed more uniformly
(Claridge-Chang, 2001).
CG13848 is an alpha-tocopherol transfer protein (alpha-TTP) that is strongly expressed in certain basal cells of the eye, showing peak expression around dawn. Humans with mutations in the homologous alpha-TTP show neurodegenerative ataxia that is associated with a deficiency in alpha-tocopherol (vitamin E) incorporation into lipoprotein particles. Vitamin E acts as an antioxidant, and it is thought that this activity allows it to protect neurons from damage by free radicals. Also from human studies, it is known that photoreceptor cells are particularly susceptible to oxidative damage due to high levels of polyunsaturated fatty acids in the photoreceptor membrane, and their exposure to visible light. Thus, it is proposed that the dawn phase of CG13848 represents an upregulation of alpha-TTP for increased daytime transfer of photoprotective vitamin E into the photoreceptor membrane. Also in the circadian set, Catalase (encoded by Cat) and a thioredoxin (encoded by CG8993) are both involved in neutralizing reactive oxygen species (Claridge-Chang, 2001).
Metabolism
Different aspects of metabolism are represented among the selected set
of oscillating transcripts: lipid metabolism (five genes), amino acid
metabolism (three), carbohydrate metabolism (three), and glycoprotein
biosynthesis (two). Intriguingly, Zw
encodes glucose-6-phosphate 1-dehydrogenase of the pentose-phosphate
pathway (PPP), while CG10611 encodes fructose-bisphosphatase in
gluconeogenesis. The two pathways have antagonizing roles in glucose
metabolism; both genes are key control points in their respective
pathway and are maximally expressed in opposite phases. Zw
transcripts are at their zenith just before dusk (ZT11), whereas
CG10611 transcripts peak at dawn (ZT0). Thus, the antiphase
oscillation of these two genes may produce daily alternation between
glucose anabolism and catabolism. Zw and CG10611 transcript
levels also respond in opposite fashion to clock defects: Zw
levels are significantly decreased in per0 and tim01
mutants, whereas, CG10611 is significantly upregulated in tim01
(Claridge-Chang, 2001).
In more direct relation to the clock itself, Zw is the first committed step in the PPP and generally thought to control flux through this pathway. The PPP is the major pathway of NAD (or NADP) conversion to NAD(P)H. It was recently shown that NAD(P)H can bind homologs of CLK and CYC, promoting their dimerization and DNA binding. Maximal Zw expression at ZT11 -- and therefore presumably NAD(P)H production via the PPP -- is coincident with maximal per and tim transcription by CLK/CYC. This information is consistent with Zw participating in a NAD(P)H-mediated autoregulatory loop of the clockworks (Claridge-Chang, 2001).
Nucleic Acid Metabolism
A subset of 15 genes involved in nucleic acid metabolism were found.
This includes five genes encoding specific RNA polymerase II
transcription factors. Four of these encode parts of the circadian
clock itself (per, tim, vri, and Clk),
whereas the fifth one, apterous (ap), generates a
homeobox transcription factor with a role in neurogenesis and the
expression of neuropeptides. Taf30alpha2 is a subunit of the general
transcription factor TFIID, while moira (mor) is part of the SWI-SNF
chromatin-remodeling complex. One splicing factor gene (DebB)
and one DNA repair gene (CG4049) are also found among this set of
oscillating transcripts (Claridge-Chang, 2001).
Cytoskeleton
Circadian genes with a role in the cytoskeleton include those encoding
two actin binding proteins (Chd64, CG11605), a troponin C (TpnC47D),
and two septins (Sep-1 and CG9699). Chd64 (phase peak ZT4) and TpnC47D
(phase peak ZT7) function specifically in muscle contraction. Another
Drosophila Troponin C, TpnC73F, is found to peak at ZT6
(Claridge-Chang, 2001).
In conclusion, a set of 158 genes expressed with a robust circadian rhythm in the adult Drosophila head was found by microarray screening. These encompass a wide variety of molecular functions, and expression patterns represented essentially all circadian phases. A larger set of genes was identified (393 entries; 293 entries after secondary filters), and the statistical approach again indicated significant circadian rhythmicity for these, but they were characterized by somewhat less robust oscillations than those of the smaller set. Independent verifications indicated substantial enrichment for cycling gene expression in this larger set and beyond. 532 genes passed secondary filters for 24 hr autocorrelation, noise, and the range-to-noise measure. From the frequency of Northern-verified oscillations detected in this larger pool of candidate genes, it is believed that the total complement of circadian genes would include ~400-500 in the adult head (Claridge-Chang, 2001).
There are important factors that might lead to an underestimation of the total complement of circadian genes. The approach that was used would favor genes that are homogeneously expressed in the head. If the same gene is expressed with varied phases in different head tissues, this will lessen the robustness of the apparent oscillation and phase. Similarly, if only a restricted portion of the head generates the cycling gene pattern, but constitutive expression is found elsewhere in the head, amplitude of the signal will be diminished. Differences of this sort might be expected in cases where a cycling gene product produces a limited physiological effect. Regulation of this type might be expected in the antennae, where, for example, electrophysiological responses to odorants vary with a circadian rhythm. It should also be stressed that only the fully differentiated adult head has been sampled. If transient patterns of circadian expression occur during development, these would go unnoticed in the experiment. Given the plethora of tissues housing autonomous circadian clocks, an expanded list of rhythmic genes would probably be derived from any related sampling of the body (e.g., wings, legs, excretory, and digestive tissues), especially as this tissue autonomy may reflect a requirement for tissue-specific pathways of circadian control that lie downstream from a largely uniform clock mechanism (Claridge-Chang, 2001).
How will the many patterns of cycling gene expression be further explored? Molecular tools that reveal the importance of oscillating gene activity have already been applied to a study of several clock genes in Drosophila. In these studies, oscillating patterns of a target gene's expression have been replaced with constitutive activity. Central questions related to vri, per, and tim function have each been explored in this manner. The present study allows an expansion of such work to address the molecular connections between individual behaviors and circadian clocks (Claridge-Chang, 2001).
The transcriptional circuits of circadian clocks control physiological and behavioral rhythms. Light may affect such overt rhythms in two ways: (1) by entraining the clock circuits and (2) via clock-independent molecular pathways. In this study the relationship between autonomous transcript oscillations and light-driven transcript responses were examined. Transcript profiles of wild-type and arrhythmic mutant Drosophila were recorded both in the presence of an environmental photocycle and in constant darkness. Systematic autonomous oscillations in the 12- to 48-h period range were detectable only in wild-type flies and occurred preferentially at the circadian period length. However, an extensive program of light-driven expression was confirmed in arrhythmic mutant flies. Many light-responsive transcripts are preferentially expressed in the compound eyes and the phospholipase C component of phototransduction, NORPA (no receptor potential), is required for their light-dependent regulation. Although there is evidence for the existence of multiple molecular clock circuits in cyanobacteria, protists, plants, and fungi, Drosophila appears to possess only one such system. The sustained photic expression responses identified here are partially coupled to the circadian clock and may reflect a mechanism for flies to modulate functions such as visual sensitivity and synaptic transmission in response to seasonal changes in photoperiod (Wijnen, 2006).
In recent years, five different sets of circadian transcripts have been proposed for the Drosophila head. Unfortunately, the overlap between these transcript sets is very poor (seven transcripts), and it falsely excludes numerous confirmed circadian transcript oscillations. These recent genome-wide surveys for rhythmic transcription have defined groups of circadian transcripts based on empirical ranking and filtering approaches, often using necessarily arbitrary cut-offs. To complement these studies a method was developed for examining periodic expression at the systems level, allowing pursuit of a number of new investigations. This new strategy enabled description of the programs of circadian and light-driven transcription in the adult fly head. Because this method emphasizes uniformity in period length and peak phase while tolerating inter-experimental variability in amplitude, it is particularly successful at measuring oscillatory trends across different independent experiments. Integrative analysis of all available microarray time-series data allowed detection and ranking of oscillatory transcript profiles with improved resolution and revealed a circadian expression program that is much more substantial than the apparent consensus (or lack thereof) between different published studies indicates. Some of the best described and strongest circadian oscillations (per, Clk, Pdp1, cry, and to) were missed in one or more of the previously published studies, but all of these rank high in the current integrative analysis. Although there are relatively few genes (~50) that show the same level of circadian regulation as the oscillating components in the core clock circuits (per, tim, Clk, cry, vri, and Pdp1), the results provide evidence for a substantially broader circadian expression program downstream of the core oscillator. This suggests that the circadian clock is responsible for both the purely circadian expression patterns of a limited set of genes and the partial circadian regulation of a much greater group (Wijnen, 2006).
Whereas many of the genes composing the Drosophila clock are expressed with a circadian rhythm in wild-type flies, all known clock gene oscillations cease if just one of them is lost by mutation. It was reasoned that all of the circadian oscillations in gene expression that were identified in this study should stop in tim01 mutants if these were truly devoid of a circadian clock. Alternatively, rhythmicity could theoretically persist in a subset of the genes if their expression depended on a parallel, novel circadian clock. The distribution analyses allowed addressing of these two alternative possibilities. No alternative systems of oscillatory expression are detectable for the 12-48-h range of period lengths. In the absence of tim-dependent clock circuits, no circadian patterns of gene expression were detected. This latter result, from microarray and Northern analyses, is in agreement with earlier observations, with limited sampling of individual circadian transcripts. Moreover, the absence of detectable molecular circadian rhythms fits well with the abolition of circadian eclosion and locomotor rhythms in tim01 flies. Thus, Drosophila appears to possess only one, tim-dependent, circadian clock. This observation contrasts with results from cyanobacteria, protists, fungi, and plants that suggest the presence of multiple oscillators, sometimes even in the same cell. Although there is no compelling evidence supporting the existence of alternative circadian clocks in Drosophila that are not entrainable to light or independent from transcriptional rhythms, this study does not disprove these possibilities. The results complement and extend previous microarray and differential display analyses using different arrhythmic mutants (per0 or Clkjrk) in which few or apparently no daily transcript oscillations persisted in the mutant context (Wijnen, 2006).
Comparative analysis of data collected from wild-type and arrhythmic mutant flies in the presence or absence of an environmental photocycle allowed identification of a program of light-driven regulation. The tim01 mutant flies used for these experiments do not just have a defective circadian clock, but because TIM degradation is a major mechanism of clock re-setting, they have also lost the main photic input pathway that entrains the clock circuits to light. In a wild-type context, light can directly entrain clock-bearing tissues in a cell-autonomous manner by activating the circadian photoreceptor CRY, or it can entrain the pacemaker neurons in the brain via phototransduction in the visual organs. TIM is the target for CRY's effect on the clock circuits, and it may also play a role in mediating entrainment via the visual organs. In spite of their defective clock circuits and circadian entrainment pathways, tim01 mutant flies retain an extensive set of daily transcript oscillations in the presence of an environmental photocycle. By comparing the light-driven expression signature that was found for tim01 with the microarray analysis for per0 LD and with confirmatory northern analyses, it was established that many light-driven transcripts show the same expression profiles in per0 and tim01 arrhythmic mutants. Moreover, the light-driven expression response found in a combined per0 and tim01 LD microarray dataset is comparable in size to the clock-dependent circadian expression program (Wijnen, 2006).
Light-regulated genes fall into two classes, a clock-independent class, and a group of genes that are also clock-controlled. That there are clock-independent patterns of light-regulated gene expression suggests that coordinate clock- and light-control can be disadvantageous in some circumstances. For example, although the clock carries phase information about the photocycle, it may not be able to carry information about day length and sunlight intensity, and some photoprotective functions might be better linked to acute light activation so that they are delivered only when needed. Such a case might be made for ultraviolet-induced melanogenesis in human skin. In contrast, it is suspected that many genes controlled by light and the clock contribute to processes that require both daily anticipation of changes in light and light responsiveness (Wijnen, 2006).
A survey of published expression studies for the selection of light-regulated genes indicates that many of them are prominently expressed in the adult compound eyes (trpl, CdsA, Pkc53E, dlg1, Slob, CG17352, CG5798, CG7077, CdsA, dlg1, Slob, and trpl). Indeed, comparative transcript profiling studies of wild-type and eya2 mutant flies predict expression in the adult compound eyes for 22 of the 27 light-dependent transcripts (Wijnen, 2006).
Two of the confirmed light-regulated transcripts (trpl and CdsA) encode known regulators of phototransduction. Daily oscillations in the transcript levels have been observed for trpl, which encodes a light-activated calcium channel. Although some effects on light-activated conductance have been observed in a trpl null mutant, the major light-dependent cation channel in Drosophila appears to be encoded by its homolog trp (transient receptor potential). Instead, the TRPL protein may have a specific function in phototransduction during extended illuminations and for adaptation of the light response to dim background light. The effect of TRPL on long-term adaptation is thought to be mediated via light-dependent subcellular translocation of TRPL protein, resulting in a preferred localization at the photoreceptor membranes in the dark and in the cell-bodies in the light. Experiments in the blowfly Calliphora vicina indicate that this translocation does not require regulation at the transcript level, but it is possible that the daily evening peaks of the trpl transcript in Drosophila facilitate efficient accumulation of TRPL protein at the rhabdomeres around dusk. Daily fluctuations are also exhibited by the transcript for CdsA (CDP diglyceride synthetase). The CDSA protein is localized to photoreceptor neurons and catalyzes the synthesis of CDP-diacyl glycerol from phosphatidic acid and CTP071. This enzymatic function helps generate the signaling compound phosphatidyl inositol 4,5-bisphosphate, which is consumed during phototransduction by the phospholipase C NORPA. Studies of CdsA loss-of-function and gain-of-function mutants indicate that by controlling availability of phosphatidyl inositol 4,5-bisphosphate, CDSA expression levels affect the gain of the phototransduction response. Periodic variation of CdsA expression under influence of the environmental photocycle could, therefore, be hypothesized to promote daily variations in visual sensitivity (Wijnen, 2006).
Two other light-driven transcripts, dlg1 and Slob, are associated with the regulation of synaptic transmission. The dlg1 (discs large 1) gene has roles in control of cell growth and differentiation as well as synaptic function. DLG1 spatial expression pattern includes synaptic sites in the adult brain and the outer membrane of photoreceptors, where it localizes Sh (Shaker) potassium channels (Wijnen, 2006).
Slob is negatively regulated by light in a clock-independent manner in addition to being one of the most robustly oscillating circadian transcripts in the adult head. The clock-dependent and light-dependent fluctuations that were uncovered for the Slob transcript are reflected in the SLOB protein levels observed in photoreceptor cells and whole heads. A number of findings point to a possible role for SLOB in mediating overt behavioral rhythms. SLOB protein is thought to bind the SLO and EAG potassium channels, and can directly enhance SLO activity, as well as mediate the inhibitory effect of 14-3-3ζ on SLO. slo mutants have altered potassium channel currents and reported defects in flight, male courtship, and circadian locomotor behavior, whereas mutations of eag display hyperactivity, and affect potassium currents and courtship behavior (Wijnen, 2006).
As mentioned above, circadian rhythms in adult Drosophila can be entrained to a LD cycle via either opsin-mediated photoreception in the light-sensing organs (compound eyes, ocelli, and eyelets) or cell-autonomous activation of the circadian blue-light photoreceptor CRY. Interestingly, the contribution of visual photo-transduction to circadian photo-entrainment is apparently restricted to a few pacemaker neurons in the brain, a situation reminiscent of photo-entrainment of the clock circuits in the mammalian brain via the retina and the retino-hypothalamic tract. In contrast, Drosophila CRY contributes to photo-entrainment in many more clock-bearing tissues, including the visual organs. CRY mediates the light-dependent degradation of TIM, which in turn affects CLK/CYC transcriptional activity in a manner that depends on the phase of the circadian cycle (Wijnen, 2006).
The light-driven transcript responses identified in this study resemble circadian responses in amplitude and duration in the context of a photocycle, and are found for a number of genes with a verified circadian expression profile. It was, therefore, asked whether these light-driven transcript responses depend on the same light sensors as the circadian system. For the most part light-driven regulation was found not to require CRY. Given TIM's status as a target for CRY-mediated light responses, it is perhaps not surprising that light-driven expression responses that do not require TIM function also persist in the absence of CRY. There is one interesting exception to this rule: The light-mediated repression of the Slob transcript apparently requires CRY, but not TIM. If this observation indeed represents a previously unappreciated function for CRY, it may share this role with the phospholipase C enzyme NORPA, since norpA mutants similarly affect the Slob transcript (Wijnen, 2006).
In contrast with CRY, it was found that NORPA phototransduction mediates many if not all of the other clock-independent light responses identified in this study. Based on the overlapping expression of both NORPA and its target transcripts in the adult compound eyes and NORPA's well-documented role in phototransduction, the simplest interpretation of these observations would be that light-driven expression responses are mediated by visual phototransduction. Nevertheless, NORPA is known to be expressed outside of the visual organs, and it has been reported to affect functions unrelated to phototransduction, such as olfaction and temperature-controlled clock gene oscillations. Additionally, norpA loss-of-function mutants show a number of defects in circadian locomotor behavior. Their activity profiles reveal an advanced evening activity peak under LD conditions and a shortened intrinsic period length under DD conditions, and they are slow to adjust their behavior to shifting cycles of light and dark. One possible interpretation of these observations is that NORPA plays a role in seasonal photoperiodic control of locomotor behavior. The norpA mutant phenotype partially mimics the effect of a shortened photoperiod, which also leads to advanced evening activity peaks and shortened period lengths. Recent studies provide further evidence connecting norpA to seasonal control of daily locomotor activity patterns. norpA mutants show abnormally high levels of splicing in the 3' untranslated region of per mRNA. Increased splicing of per transcripts at this site has been shown to contribute to the advanced accumulation of PER protein and the advanced timing of evening locomotor activity that is observed for shorter photoperiods and lower temperatures. Thus, NORPA's effect on splicing of per may be an important determinant of the 'short day' locomotor behavior phenotype of norpA mutants. The sustained photic expression responses that are identified here may reflect yet another mechanism for flies to translate a seasonal environmental signal (photoperiod) into a set of molecular signals. Photoperiodic control of transcripts associated with functions in visual sensitivity (trpl and CdsA) and synaptic transmission (Slob and dlg1) may be relevant to adaptive responses in the visual system and the brain. NORPA's involvement in both regulating per splicing and mediating photoresponses at the transcript level raises questions as to if and how these two molecular functions are connected. One possibility is that both reflect NORPA-dependent selective regulation of mRNA stability that takes place in the compound eyes (and perhaps also the brain). Whether or not NORPA's function in circadian locomotor behavior involves some of the light-dependent expression responses that have been identified could be examined by targeted misexpression studies. The subset of transcripts that have been independently confirmed to exhibit both NORPA-dependent light responses and strong clock-dependent circadian regulation might be particularly relevant to these experiments (Wijnen, 2006).
This paper has reported a new strategy for analyzing oscillatory patterns in microarrray data that allowed answer general questions about oscillatory gene systems in the fly head. By applying this strategy to 17 d of data, it was conclusively demonstrated that there are more than a hundred circadian transcript oscillations in the fly head. Additionally, in a search for rhythmic gene activity over a wide range of periods (from 12 to 48 h), it was established that 24-h periodicity constitutes the only broad program of transcriptional oscillation. It was further found that the tim-dependent clock is the sole transcriptional circadian clock in Drosophila. Thus, the fly appears to differ from cyanobacteria, protists, plants, and fungi, which are thought to possess multiple circadian clocks. Lastly, a novel, light-regulated system of gene regulation was found in Drosophila that is largely dependent on norpA-mediated phototransduction. This system regulates about the same number of genes as the clock, including a number of circadian genes. This study defines three types of transcripts that oscillate in wild-type flies: those from purely clock-regulated genes, those that are purely photocycle-regulated, and those expressed by genes that respond to both inputs (Wijnen, 2006).
In Drosophila, the clock that controls rest-activity rhythms synchronizes with light-dark cycles through either the blue-light sensitive cryptochrome (Cry) located in most clock neurons, or rhodopsin-expressing histaminergic photoreceptors. This study shows that, in the absence of Cry, each of the two histamine receptors Ort and HisCl1 contribute to entrain the clock whereas no entrainment occurs in the absence of the two receptors. In contrast to Ort, HisCl1 does not restore entrainment when expressed in the optic lobe interneurons. Indeed, HisCl1 is expressed in wild-type photoreceptors and entrainment is strongly impaired in flies with photoreceptors mutant for HisCl1. Rescuing HisCl1 expression in the Rh6-expressing photoreceptors restores entrainment but it does not in other photoreceptors, which send histaminergic inputs to Rh6-expressing photoreceptors. These results thus show that Rh6-expressing neurons contribute to circadian entrainment as both photoreceptors and interneurons, recalling the dual function of melanopsin-expressing ganglion cells in the mammalian retina (Alejevski, 2019).
The Drosophila sleep-wake rhythms are controlled by a brain circadian clock that includes about 150 clock neurons. Light synchronizes the clock neuronal network through cell-autonomous and non-cell-autonomous light input pathways. Cry is a blue-light sensitive photoreceptor protein that is expressed in most clock neurons. In the absence of Cry, flies do not phase-shift their behavioral rhythms in response to a short light pulse but still synchronize to light-dark (LD) cycles. Only flies devoid of both Cry and rhodopsin-expressing photoreceptors fail to entrain to LD cycles. Six different rhodopsins (Rhs) have been characterized in the Drosophila photoreceptive structures, which include the compound eye, the Hofbauer-Buchner (H-B) eyelet, and ocelli. The compound eye strongly contributes to circadian photoreception, whereas a modest contribution appears to be brought by the H-B eyelet and the ocelli. A circadian function has been recently associated with the yet poorly characterized rhodopsin 7, although its exact contribution and localization in the brain and/or the eye remains controversial. In addition to entrainment, the visual system controls other features of the clock neuron network by conveying light information to either promote or inhibit the behavioral output of specific clock neuron subsets (Alejevski, 2019).
The compound eye includes about 800-unit eyes (ommatidia), each of which contains eight photoreceptors. The six Rh1-expressing outer photoreceptors (R1-6) are involved in motion detection and project to the lamina neuropile of the optic lobe. The two inner photoreceptors (R7-8) are important for color detection and project to the medulla. They express four different rhodopsins and thus define two types of ommatidia: 'pale' (p) ommatidia (30%) include a Rh3-expressing R7 and a Rh5-expressing R8, whereas 'yellow' (y) ommatidia (70%) include a Rh4-expressing R7 and a Rh6-expressing R8. Each extra-retinal H-B eyelet contains four Rh6-expressing photoreceptors that project to the accessory medulla, in the vicinity of key pacemaker neurons, the ventral lateral neurons (LNvs) that produce the pigment-dispersing factor (PDF) neuropeptide9,20-24. Each of the three ocelli contains about 80 photoreceptors that express Rh225. The Drosophila rhodopsins cover a wide range of wavelengths from 300 nm to 600 nm18,19, with only Rh1 and Rh6 being sensitive to red light (Alejevski, 2019).
Rhodopsin-dependent circadian entrainment involves two downstream signaling pathways, the canonical one that relies on the phospholipase C encoded by the no receptor potential A gene (norpA)2 or an unknown pathway that does not contribute in very low light levels. All but Rh2- and Rh5- expressing photoreceptors support synchronization in very low light, and at least Rh1, Rh5, and Rh6 can signal through the NorpA-independent pathway. Photoreceptors of the compound eye are histaminergic but the H-B eyelet expresses both histamine and acetylcholine. Although the two neurotransmitters might contribute to circadian entrainment, flies devoid of Cry and histidine decarboxylase do not synchronize their rest-activity rhythms with LD cycles. This suggests that besides Cry, there is no histamine-independent pathway to entrain the clock (Alejevski, 2019).
Two genes encoding histamine-gated chloride channels, ora transientless (ort) and Histamine-gated chloride channel subunit 1 (HisCl1), have been identified in Drosophila. The ort-null mutants are visually blind and their electroretinograms have no ON and OFF transients. In contrast, HisCl1 mutants show increased OFF transients, whereas slower responses were observed in the postsynaptic laminar monopolar cells. Based on transcriptional reporters, ort expression in the optic lobes was observed in neurons of both the lamina and medulla/lobula neuropils. Based on reporter gene expression, HisCl1 was localized in glial cells of the lamina. However, recent work reported expression in photoreceptors, in particular in the R7 and R8 inner photoreceptor subtypes. Indeed, Ort and HisCl1 support color opponency between the two subtypes of 'inner' photoreceptors, the ultraviolet (UV)-sensitive R7 and non-UV-sensitive R8, with HisCl1 and Ort mediating direct and indirect inhibition, respectively. The histaminergic pathways that are involved in circadian entrainment are unknown and are the subject of the present study. The results show that both Ort and HisCl1 define two different pathways for circadian entrainment. Whereas Ort contributes through its expression in the interneurons of the optic lobe, HisCl1 mostly contributes through its expression in the Rh6-expressing retinal photoreceptors. The work thus reveals that Rh6-expressing neurons contribute to light-mediated entrainment as both photoreceptors and interneurons (Alejevski, 2019).
This work reveals that the Cryptochrome-independent entrainment of rest-activity rhythms relies on distinct pathways that are contributed by the two histamine receptors Ort and HisCl1. Whereas Ort mediates circadian entrainment through the optic lobe interneurons that are involved in visual functions, HisCl1 defines a new photoreceptive pathway through Rh6-expressing photoreceptors. Although both receptors mediate synchronization with a shifted LD cycle, it seems likely that the two pathways will show differences in specific light conditions. It was not possible to rescue Ort function with HisCl1 expression in the ort-expressing cells, whereas the Ort could replace HisCl1 in Rh6 photoreceptors. It is possible that HisCl1 has a lower affinity for histamine with Rh6 cells receiving more neurotransmitter than optic lobe interneurons. Alternatively, interneurons could sufficiently differ from photoreceptors for their physiology or specific receptor-interacting protein content, preventing HisCl1 from working efficiently. HisCl1 downregulation in Rh6 cells slows down synchronization and flies with HisCl1134 mutant eyes synchronize very poorly with advanced LD cycles, and fail to synchronize with delays. It cannot be excluded that non-photoreceptor cells contribute to HisCl1-dependent entrainment but other pathways appear to have a modest contribution if any (Alejevski, 2019).
HisCl1 is expressed in the H-B eyelet, which could thus contribute to this synchronization pathway. However, the cell-killing experiments indicate that H-B eyelet is not required for HisCl1-mediated synchronization through Rh6 cells. In the recently described color opponency mechanism, retinal R7 cells inhibit R8 and vice versa through HisCl1 expression in the photoreceptors (Schnaitmann, 2018). It is supposed that HisCl1-dependent clock synchronization is also mediated by the hyperpolarization of Rh6-expressing cells. How this hyperpolarization interacts with the light-induced depolarization in Rh6 photoreceptors to result in a synchronization message to the clock neurons remains to be understood. Since only Rh6-expressing R8 and not the other inner photoreceptors contribute to this circadian photoreception pathway, Rh6 cells might have specific connections with downstream interneurons. Such specificity has been described for color vision where each of the four inner photoreceptor subtypes connects to a different type of TmY interneuron in the Medulla. This study shows that HisCl1 expression in Rh6 cells supports synchronization with red light, in the absence of Rh1, indicating that an intra-Rh6-photoreceptor circuit is sufficient. This indicates that Rh6-expressing R8 photoreceptors play a dual photoreceptor/interneuron role in this pathway (Model for the retinal input pathways to the brain clock). Whether the same individual cells have the two roles is unknown, although the HisCl1-dependent color opponency mechanism suggests that it could be the case. It is also unclear whether all Rh6-expressing R8 photoreceptors or only a fraction of them contribute to circadian synchronization. The results imply that, in addition to histaminergic neurotransmission, Rh6-expressing photoreceptors can talk to downstream interneurons through histamine-independent neurotransmission. A recent transcriptomics study indeed revealed the expression of cholinergic markers in R7 and R8 cells, supporting cholinergic transmission in the inner photoreceptors, in addition to histaminergic transmission (Alejevski, 2019).
The data indicate that histaminergic inputs from both outer and inner photoreceptors converge to Rh6 cells to contribute to circadian entrainment. It is possible that some of these inputs rely on Rh7, which seems to be expressed in Rh6-expressing photoreceptors, according to transcriptional reporter data. Putative connections between photoreceptors have been described in Drosophila and other insects. How R1-6 photoreceptors might be connected to Rh6-expressing R8 cells remains difficult to understand, but a few putative contacts between presynaptic outer cells and postsynaptic inner cells have been observed in Musca. The intra-retinal functional connectivity that this study reports in Drosophila is reminiscent to the circuit logic of circadian entrainment in the mammalian retina, where intrinsically photoreceptive retinal ganglion cells express the melanopsin photopigment in addition to receiving inputs from rods and cones. Interestingly, melanopsin appears to share light-sensing properties with the rhabdomeric photoreceptors of invertebrates. It has been shown that the mammalian circadian clock can synchronize with day-night cycles by tracking light color changes in addition to light intensity changes. It will be interesting to investigate the possible contribution of the dual function of Rh6-expressing photoreceptors to integrating different color cues into the retinal information that is sent to the clock (Alejevski, 2019).
Clock (Clk) is a master transcriptional regulator of the circadian clock in Drosophila. To identify Clk direct target genes and address circadian transcriptional regulation in Drosophila, chromatin immunoprecipitation (ChIP) tiling array assays (ChIP-chip) were performed with a number of circadian proteins. Clk binding cycles on at least 800 sites with maximal binding in the early night. The Clk partner protein Cycle (Cyc) is on most of these sites. The Clk/Cyc heterodimer is joined 4-6 h later by the transcriptional repressor Period (Per), indicating that the majority of Clk targets are regulated similarly to core circadian genes. About 30% of target genes also show cycling RNA polymerase II (Pol II) binding. Many of these generate cycling RNAs despite not being documented in prior RNA cycling studies. This is due in part to different RNA isoforms and to fly head tissue heterogeneity. Clk has specific targets in different tissues, implying that important Clk partner proteins and/or mechanisms contribute to gene-specific and tissue-specific regulation (Abruzzi, 2011).
Previous circadian models in Drosophila suggested a transcriptional cascade in which Clk directly controls a limited number of genes, including core clock genes, which then drive the oscillating expression of many different output genes. The results of this study indicate that Clk directly regulates not only the five core clock genes (i.e., pdp1, vri, tim, per, and cwo), but also many output genes, including ~60 additional transcription factors. Some of these are tissue-specific; e.g., lim1 and crp. In addition, Clk direct target gene regulation may impact timekeeping in yet unforeseen ways. For example, Clk, Per, and Cyc bind to three of the four known circadian kinases; i.e., dbt, nmo, and sgg. Although none of these mRNAs have been previously reported to cycle, both dbt and sgg have cycling Pol II, and dbt shows weak oscillations via qRT-PCR. nmo expression is enriched in circadian neurons and has been shown to cycle in l-LNvs. The data, taken together, indicate that this simple ChIP-chip strategy has uncovered important relationships and suggest that the transcriptional regulation of some of these new target genes warrants further investigation (Abruzzi, 2011).
Most of the 1500 Clk direct target genes are also bound by two other circadian transcription factors: Cyc and Per. Because a previous study showed that there is a progressive, ordered recruitment of Clk, Pol II, and Per on per and tim (Menet, 2010), the same basic mechanism is conserved on most Clk direct targets. Clk binding increases in late morning and gives rise to an increase in Pol II signal where detectable (ZT6-ZT10). Clk binding is maximal in the early night (ZT14), and both Clk binding and Pol II occupancy decrease rapidly after the repressor Per is bound to chromatin 4-6 h later, at ZT18. Interestingly, Per binds to nearly all Clk direct targets at the identical Clk/Cyc locations, suggesting Per recruitment via protein-protein interactions (Abruzzi, 2011).
The identical binding sites for Clk, Cyc, and Per suggest that binding is not background binding or 'sterile' binding with no functional consequence. This is because three components of the circadian transcription machinery are present with proper temporal regulation. Pol II cycling on ~30% of cycling Clk targets further supports this interpretation. The Pol II signal is maximal from mid- to late morning (ZT6-ZT10), which slightly anticipates the maximal transcription times of core circadian genes like per and tim. Most Pol II signals are promoter-proximal and may reflect poised Pol II complexes often found on genes that respond quickly to environmental stimuli (Abruzzi, 2011).
To address RNA cycling, ten direct target genes with Pol II cycling were examined. Eight of these genes show oscillating mRNA with >1.5-fold amplitude, suggesting that oscillating Pol II indeed reflects cycling transcription. Because this assay may underestimate cycling transcription due to tissue heterogeneity (i.e., masking by noncycling gene expression elsewhere in the head), ~30% is a minimal estimate of Clk direct targets with cyclical mRNA (Abruzzi, 2011).
Interestingly, previous microarray studies did not detect many of these genes. One possibility is that the alternative start sites that characterize 55% of Clk direct targets are not detectable on microarrays; e.g., moe and mnt. However, several mRNAs that cycle robustly by qRT-PCR are not isoform-specific and are also not consistently identified in microarray studies. A second possibility is that the relatively low cycling amplitude of many target genes -- twofold or less, compared with the much greater amplitudes of core clock genes such as tim, per, and pdp1, assayed in parallel -- may be more difficult to detect on microarrays (Abruzzi, 2011).
Low-amplitude cycling may result from relatively stable mRNA, which will dampen the amplitude of cycling transcription. Alternatively, or in addition, low-amplitude cycling may reflect cycling in one head location and noncycling elsewhere within the head, which will dampen head RNA cycling amplitude. This is likely for many eye-specific Clk targets, which appear expressed elsewhere in the head via a Clk-independent mechanism (Abruzzi, 2011).
A third and arguably more interesting explanation for low-amplitude cycling is that Clk binds on promoters with other transcription factors within single tissues. These could include chromatin modifiers and would function together with Clk in a gene- and tissue-specific fashion. For example, a gene could be constitutively expressed at a basal level by one transcription factor, with temporal Clk binding causing a modest boost to transcription. For example, gol is a Clk target exclusively in the eye, and gol mRNA cycles with a fourfold amplitude. Rather than cycling from 'OFF' (no or very low mRNA levels) to 'ON,' however, gol mRNA levels are quite high even at the trough or lowest time points. This suggests that gol cycles from a substantial basal level in the late night and daytime to an even higher level of expression in the evening and early night. Since mRNA levels decrease by >10-fold in GMR-hid flies, trough transcription levels are not likely from other tissues. Therefore, Clk probably acts on gol and other targets not as an 'ON/OFF switch,' but rather in concert with other factors to boost a basal level of gene expression at a particular time of day and cause low-amplitude cycling within a single tissue (Abruzzi, 2011).
The large number of Clk target genes in fly heads is explained in part by tissue-specific Clk binding. Transcription assays that measure the cycling of mRNA and Pol II binding in one head tissue can be masked by noncycling expression in another. The ChIP assays, in contrast, are not plagued with the same problem. They can identify a gene bound by the cycling circadian transcription machinery even if the same gene is constitutively expressed elsewhere in the head. Surprisingly 44% of Clk direct targets were no longer detected when eyes were ablated with GMR-hid. Because many of these mRNAs are not particularly eye-enriched, it is inferred that their genes are constitutively expressed under the control of other transcription factors elsewhere in the head (Abruzzi, 2011).
The large number of target genes is also explained by the efficiency and sensitivity of the ChIP assay. It is inferred that it can detect Clk binding from a relatively low number of cells within the fly head. Lim1 is one example and is expressed predominantly in a subset of circadian neurons (l-LNvs; enriched more than four times relative to head). Preliminary cell-specific Clk ChIP-chip experiments from LNvs confirm that lim1 is an enriched Clk direct target in these cells, suggesting that this is the source of a large fraction of the binding signal in the head ChIP-chip experiments. Experiments are under way to more clearly define circadian neuron-specific Clk-binding patterns (Abruzzi, 2011).
This tissue specificity also suggests the existence of factors and/or
chromatin modifications that help regulate Clk-mediated gene expression.
They could enable Clk binding to specific genes in one tissue or inhibit
binding in another tissue. These tissue-specific factors are strongly
indicated by the pdp1 and lk6 Clk-binding patterns,
which change so strikingly and specifically in GMR-hid.
Although not unprecedented, tissue-specific factors that enable or
inhibit specific DNA-binding locations are intriguing and warrant
further investigation and identification (Abruzzi, 2011).
Neuropeptides control rhythmic behaviors, but the timing and location of their release within circuits is unknown. Imaging in the brain shows that synaptic neuropeptide release by Drosophila clock neurons is diurnal, peaking at times of day that were not anticipated by prior electrical and Ca(2+) data. Furthermore, hours before peak synaptic neuropeptide release, neuropeptide release occurs at the soma, a neuronal compartment that has not been implicated in peptidergic transmission. The timing disparity between release at the soma and terminals results from independent and compartmentalized mechanisms for daily rhythmic release: consistent with conventional electrical activity-triggered synaptic transmission, terminals require Ca(2+) influx, while somatic neuropeptide release is triggered by the biochemical signal IP(3). Upon disrupting the somatic mechanism, the rhythm of terminal release and locomotor activity period are unaffected, but the number of flies with rhythmic behavior and sleep-wake balance are reduced. These results support the conclusion that somatic neuropeptide release controls specific features of clock neuron-dependent behaviors. Thus, compartment-specific mechanisms within individual clock neurons produce temporally and spatially partitioned neuropeptide release to expand the peptidergic connectome underlying daily rhythmic behaviors (Klose, 2021).
FAP imaging revealed synaptic neuropeptide release from LNv clock neurons that does not conform to predictions from previously used indirect methods. Earlier neuropeptide-content measurements could not resolve whether somatic changes were due to release or traffic and did not detect l-LNv rhythmic neuropeptide release, likely because it is relatively modest and/or obscured by DCV capture that replenishes synaptic neuropeptide stores. Furthermore, Ca2+ measured at the soma was not reflective of release at terminals likely because of somatic IP3 signaling. Thus, presynaptic Ca2+ may be more predictive of release by LNv termini. Finally, somatic electrical recording does not take into account regulation by presynaptic inputs. Thus, direct live imaging of neuropeptide release is essential for monitoring peptidergic transmission in the brain (Klose, 2021).
Indeed, this approach demonstrates that central clock neurons release neuropeptide from terminals and the soma, with each compartment operating with different mechanisms and timing. Release from LNv clock neuron terminals is conventional (i.e., mediated by extracellular Ca2+ influx); because cell specific genetic Ca2+-channel inhibition was not used, the contributions of Ca2+ channels in LNv neurons and their presynaptic inputs was not determined. In contrast, somatic neuropeptide release is triggered by IP3 signaling that operates in the absence of action potential-induced Ca2+ influx. This shows that the two compartments use different mechanisms. It also raises the possibility that release by the two compartments differ in cell autonomy. Most importantly, different release mechanisms allow for multiphasic temporal control of neuropeptide release from separate compartments of the same neuron, each of which releases onto different parts of the clock circuit, thereby providing separate output avenues to independently influence different parameters of behavior (Klose, 2021).
Disruption of the circadian clock, which directs rhythmic expression of numerous output genes, accelerates aging. To enquire how the circadian system protects aging organisms, compare circadian transcriptomes in heads of young and old Drosophila melanogaster were compared. The core clock and most output genes remained robustly rhythmic in old flies, while others lost rhythmicity with age, resulting in constitutive over- or under-expression. Unexpectedly, a subset of genes was identified that adopted increased or de novo rhythmicity during aging, enriched for stress-response functions. These genes, termed late-life cyclers, were also rhythmically induced in young flies by constant exposure to exogenous oxidative stress, and this upregulation is CLOCK-dependent. Age-onset rhythmicity was identified in several putative primary piRNA transcripts overlapping antisense transposons. These results suggest that, as organisms age, the circadian system shifts greater regulatory priority to the mitigation of accumulating cellular stress (Kuintzle, 2017).
Sleep is a fundamental behavior in an animal's life influenced by many internal and external factors, such as aging and diet. Critically, poor sleep quality places people at risk of serious medical conditions. Because aging impairs quality of sleep, measures to improve sleep quality for elderly people are needed. Given that diet can influence many aspects of sleep, this study investigated whether a high-sucrose diet (HSD) affected aging-induced sleep fragmentation using the fruit fly, Drosophila melanogaster. Drosophila is a valuable model for studying sleep due to its genetic tractability and many similarities with mammalian sleep. Total sleep duration, sleep bout numbers (SBN), and average sleep bout length (ABL) were compared between young and old flies on a normal sucrose diet (NSD) or HSD. On the NSD, old flies slept slightly more and showed increased SBN and reduced ABL, indicating increased sleep fragmentation. Short-term maintenance of flies in HSD (up to 8 days), but not long-term maintenance (up to 35 days), suppressed aging-induced sleep fragmentation. This study provides meaningful strategies for preventing the deterioration of sleep quality in the elderly (Lee, 2021).
Many vital processes in the eye are under circadian regulation, and circadian dysfunction has emerged as a potential driver of eye aging. Dietary restriction is one of the most robust lifespan-extending therapies and amplifies circadian rhythms with age. This study demonstrates that dietary restriction extends lifespan in Drosophila melanogaster by promoting circadian homeostatic processes that protect the visual system from age- and light-associated damage. Altering the positive limb core molecular clock transcription factor, CLOCK, or CLOCK-output genes, accelerates visual senescence, induces a systemic immune response, and shortens lifespan. Flies subjected to dietary restriction are protected from the lifespan-shortening effects of photoreceptor activation. Inversely, photoreceptor inactivation, achieved via mutating rhodopsin or housing flies in constant darkness, primarily extends the lifespan of flies reared on a high-nutrient diet. These findings establish the eye as a diet-sensitive modulator of lifespan and indicates that vision is an antagonistically pleiotropic process that contributes to organismal aging (Hodge, 2022).
Progressive declines in circadian rhythms are one of the most common hallmarks of aging observed across most lifeforms. Quantifying the strength, or amplitude, of circadian rhythms is an accurate metric for predicting chronological age. Many cellular processes involved in aging (e.g., metabolism, cellular proliferation, DNA repair mechanisms, etc.) display robust cyclic activities. Both genetic and environmental disruptions to circadian rhythms are associated with accelerated aging and reduced longevity. These observations suggest that circadian rhythms may not merely be a biomarker of aging; rather, declines in circadian rhythms might play a causal role. The observation that DR and DR-memetics, such as calorie restriction and time-restricted feeding, improve biological rhythms suggests that clocks may play a fundamental role in mediating their lifespan-extending benefits (Kato, 2022).
This study identified circadian processes that are selectively amplified by DR. The findings demonstrate that DR amplifies circadian homeostatic processes in the eye, some of which are required for DR to delay visual senescence and improve longevity in Drosophila. Disrupting CLK function within photoreceptors accelerates visual declines and shortens lifespan, while overexpressing wild-type CLK protects against age-associated declines in vision and rescues AL-dependent declines in photoreceptor function. These data also demonstrate that photoreceptor stress has deleterious effects on organismal health; overstimulation of the photoreceptors induced a systemic immune response and reduced longevity (Kato, 2022).
Among the more interesting and unexpecting findings of this study is the observation that the Drosophila eye influences systemic immune responses, as elevated AMP expression was observed in the bodies of flies overexpressing CLK-Δ pan-neuronally and in flies with forced photoreceptor degeneration (ATPα-RNAi). It is possible that GAL4 misexpression may promote inflammatory responses in the fly bodies, although this study found a reduction in systemic inflammation in the rhodopsin-null lines suggesting that this phenomenon can originate at the photoreceptor. These systemic immune responses correlated with lifespan changes (increased body AMP expression is associated with declines in longevity and vice versa), similar to what is observed with chronic inflammation or “inflammaging” in other models. However, it cannot be concluded whether neuronal or eye-mediated increases in systemic inflammation are causal to aging in other tissues. Furthermore, the mechanisms by which the Drosophila eye, and, more specifically, the photoreceptor influence systemic immune responses are unclear. It is speculated that photoreceptor degeneration may disrupt the retinal-blood barrier such that damage signals from the eye propagate through to the hemolymph and activate AMP expression in distal tissues. Future studies are aimed at elucidating this mechanism, and its effect on longevity (Kato, 2022).
The findings of this study establish the eye as a diet-sensitive regulator of lifespan. DR's neuroprotective role in the photoreceptors appears to be mediated via the transcription factor CLK, which promotes the rhythmic oscillation of genes involved in the suppression of phototoxic cell stress. Given that CLK transcriptionally regulates circadian and non-circadian transcripts, future investigations may determine whether the time-of-day regulation of these genes by CLK is germane to promoting eye health with age. These studies may also examine whether the DR-mediated benefits on visual senescence and photoreceptor viability are mediated solely by CLK as a transcription factor (as demonstrated here) or whether circadian clock function (rhythmic output) is required. The findings also support the notion that age-related declines in the visual system impose a high cost on an organism's physiology. Perhaps this provides an alternative hypothesis for why several cave-dwelling animals, whose visual systems have undergone regressive evolution (e.g., cave-dwelling fish and naked-mole rats), are especially long-lived. Failing to develop a visual system may act as a pro-survival mechanism allowing organisms to avoid the damage and inflammation triggered by age-related retinal degeneration. Ultimately, developing a visual system, which is critical for reproduction and survival, may be detrimental to an organism later in life. Thus, vision may be an example of an antagonistically pleiotropic mechanism that shapes lifespan (Kato, 2022).
Lateral ventral neurons (LNvs) in the fly
circadian neural circuit mediate behaviors other than clock resetting, including light-activated acute arousal. Converging sensory inputs often confer functional redundancy. The LNvs have three distinct light input pathways: (1) cell autonomously expressed cryptochrome (CRY), (2) rhodopsin 7 (Rh7), and (3) synaptic inputs from the eyes and other external photoreceptors that express opsins and CRY. This study explored the relative photoelectrical and behavioral input contributions of these three photoreceptor systems to determine their functional impact in flies. Patch-clamp electrophysiology measuring light evoked firing frequency (FF) was performed on large LNvs (l-LNvs) in response to UV (365 nm), violet (405 nm), blue (450 nm), or red (635 nm) LED light stimulation, testing controls versus mutants that lack photoreceptor inputs gl60j, cry-null, rh7-null, and double mutant gl60j-cry-null flies. For UV, violet, and blue short wavelength light inputs, all photoreceptor mutants show significantly attenuated action potential FF responses measured in the l-LNv. In contrast, red light FF responses are only significantly attenuated in double mutant gl60j-cry-null flies. A light-pulse arousal assay was used to compare behavioral responses to UV, violet, blue and red light of control and light input mutants, measuring the awakening arousal response of flies during subjective nighttime at two different intensities to capture potential threshold differences (10 and 400 μW/cm(2)). The light arousal behavioral results are similar to the electrophysiological results, showing significant attenuation of behavioral light responses for mutants compared to control. These results show that the different LNv convergent photoreceptor systems are integrated and together confer functional redundancy for light evoked behavioral arousal (Au, 2023).
The needs fulfilled by sleep are unknown, though the effects of insufficient sleep are manifold. To better understand how the need to sleep is sensed and discharged, much effort has gone into identifying the neural circuits involved in regulating arousal, especially those that promote sleep. In prevailing models, the dorsal fan-shaped body (dFB) plays a central role in this process in the fly brain. This study manipulated various properties of the dFB including its electrical activity, synaptic output, and endogenous gene expression. In each of these experimental contexts it was not possible to identify any effect on sleep that could be unambiguously mapped to the dFB. Furthermore, evidence was found that sleep phenotypes previously attributed to the dFB were caused by genetic manipulations that inadvertently targeted the ventral nerve cord. Expression was examined of two genes whose purported effects have been attributed to functions within a specific subpopulation of dFB neurons. In both cases little to no expression expression was found in the expected cells. Collectively, these results cast doubt on the prevailing hypothesis that the dFB plays a central role in promoting sleep (De, 2023).
While neurotransmitter identity was once considered singular and immutable for mature neurons, it is now appreciated that one neuron can release multiple neuroactive substances (co-transmission) whose identities can even change over time. To explore the mechanisms that tune the suite of transmitters a neuron releases, this study developed transcriptional and translational reporters for cholinergic, glutamatergic, and GABAergic signaling in Drosophila. Many glutamatergic and GABAergic cells also transcribe cholinergic genes, but fail to accumulate cholinergic effector proteins. Suppression of cholinergic signaling involves posttranscriptional regulation of cholinergic transcripts by the microRNA miR-190; chronic loss of miR-190 function allows expression of cholinergic machinery, reducing and fragmenting sleep. Using a "translation-trap" strategy this study shows that neurons in these populations have episodes of transient translation of cholinergic proteins, demonstrating that suppression of co-transmission is actively modulated. Posttranscriptional restriction of fast transmitter co-transmission provides a mechanism allowing reversible tuning of neuronal output. It is concluded that colinergic co-transmission in large populations of glutamatergic and GABAergic neurons in the Drosophila adult brain is controlled by miR-190 (Chen, 2023).
Circadian clocks align various behaviors such as locomotor activity, sleep/wake, feeding, and mating to times of day that are most adaptive. How rhythmic information in pacemaker circuits is translated to neuronal outputs is not well understood. This study used brain-wide, 24-h in vivo calcium imaging in the Drosophila brain and searched for circadian rhythmic activity among identified clusters of dopaminergic (DA) and peptidergic neurosecretory (NS) neurons. Such rhythms were widespread and imposed by the PERIOD-dependent clock activity within the ∼150-cell circadian pacemaker network. The rhythms displayed either a morning (M), evening (E), or mid-day (MD) phase. Different subgroups of circadian pacemakers imposed neural activity rhythms onto different downstream non-clock neurons. Outputs from the canonical M and E pacemakers converged to regulate DA-PPM3 and DA-PAL neurons. E pacemakers regulate the evening-active DA-PPL1 neurons. In addition to these canonical M and E oscillators, evidence is for a third dedicated phase occurring at mid-day: the l-LNv pacemakers present the MD activity peak, and they regulate the MD-active DA-PPM1/2 neurons and three distinct NS cell types. Thus, the Drosophila circadian pacemaker network is a polyphasic rhythm generator. It presents dedicated M, E, and MD phases that are functionally transduced as neuronal outputs to organize diverse daily activity patterns in downstream circuits (Liang, 2023).
The ellipsoid body (EB) is a major structure of the central complex of the Drosophila melanogaster brain. Twenty-two subtypes of EB ring neurons have been identified based on anatomic and morphologic characteristics by light-level microscopy and EM connectomics. A few studies have associated ring neurons with the regulation of sleep homeostasis and structure. However, cell type-specific and population interactions in the regulation of sleep remain unclear. Using an unbiased thermogenetic screen of EB drivers using female flies, the following was found: (1) multiple ring neurons are involved in the modulation of amount of sleep and structure in a synergistic manner; (2) analysis of data for &deltaPdoze/&Delta:Pwake using a mixed Gaussian model detected 5 clusters of GAL4 drivers which had similar effects on sleep pressure and/or depth: lines driving arousal contained R4m neurons, whereas lines that increased sleep pressure had R3m cells; (3) a GLM analysis correlating ring cell subtype and activity-dependent changes in sleep parameters across all lines identified several cell types significantly associated with specific sleep effects: R3p was daytime sleep-promoting, and R4m was nighttime wake-promoting; and (4) R3d cells present in 5HT7-GAL4 and in GAL4 lines, which exclusively affect sleep structure, were found to contribute to fragmentation of sleep during both day and night. Thus, multiple subtypes of ring neurons distinctively control sleep amount and/or structure. The unique highly interconnected structure of the EB suggests a local-network model worth future investigation; understanding EB subtype interactions may provide insight how sleep circuits in general are structured (Yan, 2023).
The circadian system regulates the timing of multiple molecular, physiological, metabolic, and behavioral phenomena. In Drosophila, as in other species, most of the research on how the timekeeping system in the brain controls the timing of behavioral outputs has been conducted in males, or sex has not been included as a biological variable. A critical set of circadian pacemaker neurons in Drosophila release the neuropeptide pigment-dispersing factor (PDF), which functions as a key output factor in the network with complex effects on other clock neurons. Lack of Pdf or its receptor, PdfR, results in most flies displaying arrhythmicity in activity-rest cycles under constant conditions. However, the current results show that female circadian rhythms are less affected by mutations in both Pdf and PdfR. CRISPR-Cas9-mediated mutagenesis of Pdf, specifically in ventral lateral neurons (LNvs), also has a greater effect on male rhythms. The influence of M-cells on the circadian network was tested, and speeding up the molecular clock specifically in M-cells led to sexually dimorphic phenotypes, with a more pronounced effect on male rhythmic behavior. These results suggest that the female circadian system is more resilient to manipulations of M-cells and the PDF pathway, suggesting that circadian timekeeping is more distributed across the clock neuron network in females (Iyer. 2025).
Circadian clocks are autonomous daily timekeeping mechanisms that allow organisms to adapt to environmental rhythms as well as temporally organize biological functions. Clock-controlled timekeeping involves extensive regulation of rhythmic gene expression. To date, relatively few clock-associated promoter elements have been identified and characterized. In an unbiased search of core clock gene promoters from 12 species of Drosophila, a 29-bp consensus sequence was discovered that has been designated as the Clock-Associated Transcriptional Activation Cassette or 'CATAC'. To experimentally address the spatiotemporal expression information associated with this element, constructs were generated with four separate native CATAC elements upstream of a basal promoter driving expression of either the yeast Gal4 or firefly luciferase reporter genes. Reporter assays showed that presence of wild-type, but not mutated CATAC elements, imparted increased expression levels as well as rhythmic regulation. Part of the CATAC consensus sequence resembles the E-box binding site for the core circadian transcription factor CLOCK/CYCLE (CLK/CYC), and CATAC-mediated expression rhythms are lost in the presence of null mutations in either cyc or the gene encoding the CLK/CYC inhibitor, period (per). Nevertheless, the results indicate that CATAC's enhancer function persists in the absence of CLK/CYC. Thus, CATAC represents a novel cis-regulatory element encoding clock-controlled regulation (Sharp, 2017).
The genome of Drosophila melanogaster contains seven rhodopsin genes. Rh1-6 proteins are known to have respective absorption spectra and function as visual pigments in ocelli and compound eyes. In contrast, Rh7 protein was recently revealed to function as a circadian photoreceptor in the brain. However, its molecular properties have not been characterized yet. This study successfully prepared a recombinant protein of Drosophila Rh7 in mammalian cultured cells. Drosophila Rh7 bound both 11-cis-retinal and 11-cis-3-hydroxyretinal to form photo-pigments which can absorb UV light. Irradiation with UV light caused formation of a visible-light absorbing metarhodopsin that activated Gq-type of G protein. This state could be photoconverted back to the original state and, thus Rh7 is a Gq-coupled bistable pigment. Interestingly, Rh7 (lambda max = 350 nm) exhibited an unusual broad spectrum with a longer wavelength tail reaching 500 nm, whose shape is like a composite of spectra of two pigments. In contrast, replacement of lysine at position 90 with glutamic acid caused the formation of a normal-shaped absorption spectrum with maximum at 450 nm. Therefore, Rh7 is a unique photo-sensor that can cover a wide wavelength region by a single pigment to contribute to non-visual photoreception (Sakai, 2017).
Many insects show strong behavioral responses to short wavelength light. Drosophila melanogaster exhibit Cryptochrome- and Hyperkinetic-dependent blue and ultraviolet (UV) light avoidance responses that vary by time-of-day, suggesting that these key sensory behaviors are circadian regulated. This study shows mutant flies lacking core clock genes exhibit defects in both time-of-day responses and valence of UV light avoidance/attraction behavior. Non-genetic environmental disruption of the circadian clock by constant UV light exposure leads to complete loss of rhythmic UV light avoidance/attraction behavior. Flies with ablated or electrically silenced circadian lateral ventral neurons have attenuated avoidance response to UV light. It is concluded that circadian clock proteins and the circadian lateral ventral neurons of the adult brain regulate both the timing and the valence of UV light avoidance/attraction. These results provide mechanistic support for Pittendrigh's "escape from light" hypothesis regarding the co-evolution of phototransduction and circadian systems (Baik, 2018).
The human 1q21.1 deletion of ten genes is associated with increased risk of schizophrenia. This deletion involves the beta-subunit of the AMP-activated protein kinase (AMPK) complex, a key energy sensor in the cell. Although neurons have a high demand for energy and low capacity to store nutrients, the role of AMPK in neuronal physiology is poorly defined. This study shows that AMPK is important in the nervous system for maintaining neuronal integrity and for stress survival and longevity in Drosophila. To understand the impact of this signaling system on behavior and its potential contribution to the 1q21.1 deletion syndrome, this study focused on sleep, an important role of which is proposed to be the reestablishment of neuronal energy levels that are diminished during energy-demanding wakefulness. Sleep disturbances are one of the most common problems affecting individuals with psychiatric disorders. This study shows that AMPK is required for maintenance of proper sleep architecture and for sleep recovery following sleep deprivation. Neuronal AMPKbeta loss specifically leads to sleep fragmentation and causes dysregulation of genes believed to play a role in sleep homeostasis. These data also suggest that AMPKbeta loss may contribute to the increased risk of developing mental disorders and sleep disturbances associated with the human 1q21.1 deletion (Nagy, 2018).
The circadian system, which has a period of about 24 h, is import for organismal health and fitness. The molecular circadian clock consists of feedback loops involving both transcription and translation, and proper function of the circadian system also requires communication among intracellular organelles. As important hubs for signaling in the cell, mitochondria integrate a variety of signals. Mitochondrial dysfunction and disruption of circadian rhythms are observed in neurodegenerative diseases and during aging. However, how mitochondrial dysfunction influences circadian rhythm is largely unknown. This study reports that Drosophila prune (pn), which localizes to the mitochondrial matrix, most likely affects the function of certain clock neurons. Deletion of pn in flies caused decreased expression of mitochondrial transcription factor TFAM and reductions in levels of mitochondrial DNA, which resulted in mitochondrial dysfunction. Loss of pn decreased the amplitude of circadian rhythms. In addition, depletion of mtDNA by overexpression of a mitochondrially targeted restriction enzyme mitoXhoI also decreased the robustness of circadian rhythms. This work demonstrates that pn is important for mitochondrial function thus involved in the regulation of circadian rhythms (Chen. 2019).
Following acute neural injury, severed axons undergo programmed Wallerian degeneration over several following days. While sleep has been linked with synaptic reorganization under other conditions, the role of sleep in responses to neural injuries remains poorly understood. To study the relationship between sleep and neural injury responses, Drosophila melanogaster was examined following the removal of antennae or other sensory tissues. Daytime sleep is elevated after antennal or wing injury, but sleep returns to baseline levels within 24 h after injury. Similar increases in sleep are not observed when olfactory receptor neurons are silenced or when other sensory organs are severed, suggesting that increased sleep after injury is not attributed to sensory deprivation, nociception, or generalized inflammatory responses. Neuroprotective disruptions of the E3 ubiquitin ligase highwire and c-Jun N-terminal kinase basket in olfactory receptor neurons weaken the sleep-promoting effects of antennal injury, suggesting that post-injury sleep may be influenced by the clearance of damaged neurons. Finally, pre-synaptic active zones were shown to be preferentially removed from severed axons within hours after injury, and depriving recently injured flies of sleep slows the removal of both active zones and damaged axons. These data support a bidirectional interaction between sleep and synapse pruning after antennal injury: locally increasing the need to clear neural debris is associated with increased sleep, which is required for efficient active zone removal after injury (Singh, 2020).
Growing evidence indicates that microRNAs play numerous important roles. However, the roles of some microRNAs involved in regulation of circadian rhythm and sleep are still not well understood. This study shows that the miR-276b is essential for maintaining both sleep and circadian rhythm by targeting tim, NPFR and DopR1 genes, with miR-276b deleted mutant flies sleeping more, and vice versa in miR-276b overexpressing flies. Through analysing its promoter, mir-276b was found to be responsive to CLOCK and regulates circadian rhythm through the negative feedback loop of the CLK/CYC-TIM/PER. Furthermore, miR-276b is broadly expressed in the clock neurons and the central complexes such as the mushroom body and the fan-shape body of Drosophila brain, in which up-regulation of miR-276b in tim, npfr1 and DopR1 expressing tissues significantly causes sleep decreases. This study clarifies that the mir-276b is very important for participating in regulation of circadian rhythm and sleep (Zhang, 2020).
The data demonstrate that glia produce Gba1b to non-autonomously control brainsph ingolipids, protein degradation, and neurite remodeling in a circadian circuit. This study identified two specific glial subtypes, ensheathing glia (EG) and perineural glia (PNG), as critical sources of gba1b required for
lysosomal function, proteostasis, circadian behaviors and neurite remodeling. While previous genetic studies in vertebrate models did not determine whether GBA was required in glia or neurons, lysosomal GBA expression in mice and humans is ~5-fold higher in glia and microglia compared to neurons, and gba mice harbor aggregates in astrocytes. Given these expression patterns and the striking similarities in brain phenotypes seen across GBA mutants in flies, fish, and mammals, there appears to be an evolutionarily ancient role for glia in regulating sphingolipid metabolism in the brain (Vaughen, 2022).
Previous work identified the cytoskeletal effector Rho1 and the transcription factor Mef2 as important for sLNv neurite remodeling. The current work demonstrates that these changes must be coordinated with membrane remodeling, particularly sphingolipid degradation and biosynthesis, both of which are required for membrane retraction and growth. In control animals, sLNv neurites are enlarged at dawn, retract across the day to become stunted at dusk, and then re-extend during the night. This cycle coincides with a diurnal sphingolipid cycle in which specific GlcCer and Cer species are elevated during the retraction phase, and reduced during the growth phase. In gba1bΔ mutants, the cycle of sLNv growth and retraction is blocked such that sLNv neurites remain in a stunted state across the day. In parallel, the sphingolipid cycle is also substantially blocked, and many sphingolipids are present at highly elevated levels. Conversely, in animals in which sphingolipid biosynthesis is globally reduced (Pdf>laceRNAi), the cycle of sLNv remodeling is also blocked but sLNv neurites remain aberrantly extended throughout the day. Notably, Lace is bound by the transcription factor Clock and undergoes circadian changes in expression, increasing before dusk and peaking at midnight, the same stage at which RNAi perturbation would be expected to be strongest (given the Pdf promoter. Finally, changing the timing of gba1b expression in sLNv neurites preserved the diurnal cycle of neurite growth and retraction, but inverted its phase such that sLNv neurites were reduced at dawn and enlarged at night. Taken together, these results demonstrate that carefully timed cycles of sphingolipid degradation and biosynthesis are instructive for the diurnal pattern of neurite remodeling. Lipidomics analysis demonstrates that the levels of Cer and GlcCer 14:1/18:1 and 14:1/20:1 species, as well as CerPE14:1/18:0 and CerPe 14:2/18:0, are elevated during the phase of sLNv neurite retraction, suggesting that these specific species might play an important regulatory role. Although sphingolipids are substantially less abundant than phospholipids (with Cer and GlcCer species representing <0.5% of neural membranes), their unique effects on membrane biophysics makes them well-poised to strongly affect membrane remodeling and structural plasticity. Finally, given that GlcT depletion (which blocks GlcCer but not Cer formation) did not increase sLNv volume compared to lace manipulations (which blocks GlcCer, Cer, and CerPE), a model is favored whereby Cer species drive membrane retraction. This hypothesis would be consistent with recent work revealing that diurnal changes in Cer species trigger retraction of microglia processes (Vaughen, 2022).
There is also strong evidence for sphingolipids regulating the cytoskeleton. For example, blocking de novo sphingolipid synthesis in fibroblasts acutely reduced cell area and lamellipodia formation, and altered sphingolipid profiles were associated with shortened neurites and upregulated RhoA following acid ceramidase knockdown in neuroblastoma lines. Moreover, GBA2 (non-lysosomal) knockout mice have cytoskeletal defects, shorter neurites, and dysregulate Rho GTPase localization. Additionally, activity within larval LNv neurites triggers lipid-mediated structural alterations critical for circuit function dependent on Ceramide Synthase. Taken with this work, sphingolipids and their regulatory enzymes may function coordinately or even upstream to Rho1 and attendant cytoskeletal changes in sLNv remodeling, with more precise spatiotemporal control facilitated by glial degradation of remodeled neurite membranes (Vaughen, 2022).
While sLNv neurites are a dramatic example of membrane remodeling, many neurons grow and shrink across circadian cycles and varied environmental conditions. Membrane turnover is likely important during structural plasticity at synapses, and indeed gba1b flies have memory defects. As EG and PNG are broadly distributed throughout the brain, glia-mediated sphingolipid degradation may be central to membrane remodeling in many neural circuits. Interestingly, EG engulf neuronal membranes to clear damage in a sleep- dependent fashion, a cellular response potentially co-opted from a role in the daily remodeling of neurite membranes. Daily neurite remodeling is also a feature of the mouse suprachiasmatic nucleus. Moreover, microglia, which express GBA in mice and humans, locally prune neurites during development and injury, regulate sleep, and are enriched for sphingolipid catabolizing genes. Taken together, this suggests GBA could control many forms of structural plasticity across species. Protein aggregation is dynamically controlled by circadian state and neural activity Our characterization of gba1b mutant animals revealed a surprising circadian cycle of protein aggregate accumulation and removal. This study observed both activity-dependent and direct circadian control of aggregate burden. Protein aggregates have been described in wide-ranging disease models from flies to mice and are a prominent feature of neurodegeneration in humans, including cells derived from GBA mutant mice and patients. Given the observations of this study, it may prove critical to characterize aggregate accumulation (and lipid abundance) with respect to circadian time and neural activity. Indeed, circadian phagocytosis of amyloid-beta, a component of Alzheimer's aggregates, was recently observed in cultured macrophages to be driven by circadian biosynthesis of heparan sulfate proteoglycans (Vaughen, 2022).
These data provide a direct mechanistic connection between the enzymatic activity of gba1b in glia to circadian behavior through sLNv neurite remodeling, as well as broader effects on sleep behavior likely mediated by other circuits. Notably, gba1bΔ mutant animals display deficits in activity and sleep prior to overt accumulation of aberrant protein aggregates. Similarly, many Parkinson's patients have sleep defects prior to development of clinical characteristics typically associated with neuropathological aggregate accumulation. Many genes that impinge on lysosomal function and sphingolipid degradation are linked to Parkinson's disease, and glia are key regulators of Parkinson's and other neurodegenerative diseases. Moreover, precise control of lipid species is central to many neurodegenerative models and is often modulated by neural activity. Recently, long-chain saturated lipids were found to mediate neurotoxicity by reactive astrocytes, and this study also observed increased nighttime long-chain saturated Cer/GlcCer species in gba1bΔ mutants, which coincides with circadian aggregate burden. Based on these observations, mutations in lipid-regulating genes could impair glial remodeling of sleep circuits, and specific lipid species may diurnally drive cyclic aggregates. Sleep has been proposed to drive clearance of aggregates from the brain. Moreover, cell-type specific functions for sphingolipids are known in glia, and regional lipid heterogeneity pervades the human brain\. Unraveling the complicated mechanisms of cell-type specific lipid synthesis and degradation may provide crucial insights into the connections between sleep, circadian rhythms, neurodegenerative diseases, and neuronal membrane dynamics (Vaughen, 2022).
These studies revealed that the brain is sensitive to the level of expression of gba1b when doing rescue experiments, likely reflecting the fact that gba1b itself is under tight transcriptional control and that very low levels of expression are sufficient to rescue most gba1b phenotypes (consistent with scRNA-seq data). A key gap in the field is the absence of tools that afford both cell-type specificity and quantitative control of expression across physiologically relevant levels alongside precise temporal actuation. As such tools develop, further exploring the relative balance of sphingolipid biosynthesis and degradation in shaping neurite growth and retraction would be fascinating. In addition, while analyzing sLNv neurites at specific timepoints has provided key insights, being able to capture the dynamics of lipid turnover in a live imaging preparation would deepen this understanding. This requires the application of fluorescent probes for specific lipids that can be engaged cell-type specifically, combined with a chronic imaging preparation that can span both sleep and wake cycles. Finally, while this study has quantitatively measured changes in membrane lipid (Vaughen, 2022).
The endogenous circadian period of animals and humans is typically very close to 24 h. Individuals with much longer circadian periods have been observed, however, and in the case of humans, these deviations have health implications. Previously, a line of Drosophila was observed with a very long average period of 31.3 h for locomotor activity behavior. Preliminary mapping indicated that the long period did not map to known canonical clock genes but instead mapped to multiple chromosomes. Using RNA-Seq, the whole transcriptome of fly heads from this line was surveyed across time and compared with a wild-type control. A three-way generalized linear model revealed that approximately two-thirds of the genes were expressed differentially among the two genotypes, while only one quarter of the genes varied across time. Using these results, algorithms were applied to search for genes that oscillated over 24 h, identifying genes not previously known to cycle. 166 differentially expressed genes were identified that overlapped with a previous Genome-wide Association Study (GWAS) of circadian behavior, strongly implicating them in the long-period phenotype. Mutations were tested in 45 of these genes for their effect on the circadian period. Mutations in Alk, alph, CG10089, CG42540, CG6034, Kairos (CG6123), CG8768, klg, Lar, sick, and tinc had significant effects on the circadian period, with seven of these mutations increasing the circadian period of locomotor activity behavior. Genetic rescue of mutant Kairos restored the circadian period to wild-type levels, suggesting it has a critical role in determining period length in constant darkness (Kumar, 2020).
The transcription factor CLOCK (CLK) is essential for the development and maintenance of circadian rhythms in Drosophila. However, little is known about how CLK levels are controlled. This study shows that Clk mRNA is strongly regulated post-transcriptionally through its 3' UTR. Flies expressing Clk transgenes without normal 3' UTR exhibit variable CLK-driven transcription and circadian behaviour as well as ectopic expression of CLK-target genes in the brain. In these flies, the number of the key circadian neurons differs stochastically between individuals and within the two hemispheres of the same brain. Moreover, flies carrying Clk transgenes with deletions in the binding sites for the miRNA bantam have stochastic number of pacemaker neurons, suggesting that this miRNA mediates the deterministic expression of CLK. Overall these results demonstrate a key role of Clk post-transcriptional control in stabilizing circadian transcription, which is essential for proper development and maintenance of circadian rhythms in Drosophila (Lerner, 2021).
Rhythmic rest-activity cycles are controlled by an endogenous clock. In Drosophila, this clock resides in ∼150 neurons organized in clusters whose hierarchy changes in response to environmental conditions. The
concerted activity of the circadian network is necessary for the
adaptive responses to synchronizing environmental stimuli. Thus far work
was devoted to unravel the logic of the coordination of different
clusters focusing on neurotransmitters and neuropeptides. This study
further explored communication in the adult male brain through ligands
belonging to the Bone Morphogenetic Protein (BMP) pathway. This study
shows that the Lateral Neurons ventral (LNvs) express the small
morphogen Decapentaplegic
(DPP). DPP expression in the large LNvs triggered a period lengthening
phenotype, while its downregulation caused reduced rhythmicity and
affected anticipation at dawn and dusk, underscoring DPP per se conveys
time-of-day relevant information. Surprisingly, its expression in the
large LNvs impaired circadian remodeling of the small LNv axonal
terminals, likely through local modulation of the guanine nucleotide
exchange factor (GEF) Trio. These findings open the provocative
possibility that the BMP pathway is recruited to strengthen/reduce the
connectivity among specific clusters along the day and thus modulate
their contribution to the circadian network (Polcownuk, 2021).
Full comprehension of circadian clocks
function requires precise understanding of their entrainment to the
environment. The phase of entrained clock is plastic, which depends on
different factors such as the period of endogenous oscillator, the
period of the zeitgeber cycle (T), and the proportion of light and
darkness (day length). This study investigated the importance of the
neuropeptide Pigment-Dispersing Factor (PDF) for entrainment by systematically studying locomotor activity rhythms of Pdf
mutants and wild-type flies under different T-cycles (T22 to T32) and
different day lengths (8, 12, and 16 hour [h]). Furthermore, this study
analysed Period
protein oscillations in selected groups of clock neurons in both
genotypes under T24 and T32 at a day length of 16 h. As expected, it was
found that the phase of Drosophila's evening activity and evening
neurons advanced with increasing T in all the day lengths. This advance
was much larger in Pdf mutants (~7 h) than in wild-type flies causing
(1) pronounced desynchrony between morning and evening neurons and (2)
evening activity to move in the morning instead of the evening. Most
interestingly, it was found that the lights-off transition determines
the phase of evening neurons in both genotypes and that PDF appears
necessary to delay the evening neurons by ~3 h to their wild-type phase (, 2021).
Thus, in T32, PDF first delays the molecular cycling in the evening
neurons, and then, as shown in previous studies, delays their neuronal
firing rhythms to produce a total delay of ~7 h necessary for a
wild-type evening activity phase. It is concluded that PDF is crucial
for appropriate phasing of Drosophila activity rhythm (Vaze, 2021).
Proper timing of rhythmic locomotor behavior is the consequence of integrating environmental conditions and internal time dictated by the circadian clock. Rhythmic environmental input like daily light and temperature changes (called Zeitgeber) reset the molecular clock and entrain it to the environmental time zone the organism lives in. This study attempted to entrain the clock with a Zeitgeber different from the environmental input used for phasing the behavior. 150 clock neurons in the Drosophila melanogaster brain control different aspects of the daily activity rhythms and are organized in various clusters, During regular 12 hr light: 12 hr dark cycles at constant mild temperature (LD 25°C, LD being the Zeitgeber), so called morning oscillator (MO) neurons control the increase of locomotor activity just before lights-on, while evening oscillator (EO) neurons regulate the activity increase at the end of the day, a few hours before lights-off. This study, using 12 h: 12 h 25°C:16°C temperature cycles as Zeitgeber, attempted to look at the impact of light on phasing locomotor behavior. While in constant light and 25°C:16°C temperature cycles (LLTC), flies show an unimodal locomotor activity peak in the evening, during the same temperature cycle, but in the absence of light (DDTC), the phase of the activity peak is shifted to the morning. This study shows that the EO is necessary for synchronized behavior in LLTC but not for entraining the molecular clock of the other clock neuronal groups, while the MO controls synchronized morning activity in DDTC. Interestingly, the data suggest that the influence of the EO on the synchronization increases depending on the length of the photoperiod (constant light vs 12h of light). Hence, these results show that effects of different environmental cues on clock entrainment and activity phase can be separated, allowing to decipher their integration by the circadian clock (Lorber, 2022).
The daily pattern of locomotor behavior is highly plastic and not only depends on the time of day but also on the current environmental condition. How the brain clock integrates Zeitgeber information and various environmental inputs to time locomotor activity is not clear. To address this important question, a specific daily temperature oscillation (25°C-16&frhC) was used as Zeitgeber and light input as the environmental variable. By genetically probing clock- and neuronal function of different parts of the circadian neuronal network in different environmental conditions, it was revealed that the ambient light status determines circadian network balance (Lorber, 2022).
An important question in chronobiology that so far remains unanswered is the mechanism of entrainment by light and temperature when these two environmental parameters are actually not so reliable considering their substantial daily variation. Notably, substantial weather-inflicted variations of temperature and light intensity can occur during the day and these must not lead to circadian clock resetting. Nonetheless, animals behave quite differently depending on the current environmental status regardless of their clock entrainment status. For example, fruit flies lose their two anticipatory activities present in LD 25°C at different constant temperatures. At warm temperature (≥30°C), flies only anticipate the light-on transition while at colder temperatures (≤20°C) they only anticipate the lights-off. On the other hand, when temperature cycles are used as Zeitgeber in constant darkness, the absolute temperatures determine the phase of the activity peak, with TCs of 25°C:16°C and 29°C:20°C inducing activity in the first half or the second half of the thermophase, respectively. In constant light however, both temperature cycles result in an activity peak during the second half of the thermophase (Lorber, 2022).
This study demonstrates that the EO that drives the evening peak in LD25°C is necessary to drive the evening peak in LLTC25°C-16°C, while the MO that drives the morning peak in LD25°C controls the phase in DDTC25°C-16°C. According to this model the ambient environmental condition modulates the clock network balance. Hence, the environmental condition must be taken into account in order to understand the role of each oscillator. Previously, trying to understand how temperature cycles synchronize flies, a similar approach has been performed, although in the absence of light and different absolute temperatures, but with the same amplitude of temperature oscillations (29°C-20°C). Consistent with these findings, ablating both the MO and EO using a cry-gal4 line lead to largely desynchronized behavior. As mentioned above, in DDTC 29°C:20°C conditions flies exhibit an afternoon activity peak, and ablating the PDF neurons does not affect this synchronization. In contrast, the data show that a functional clock in the PDF neurons is important for synchronized morning activity in DDTC 25°C:16°C. Following the model, it is postulated therefore that the DN1p clock neurons also contribute to the behavioral evening activity during warm temperature cycles in constant darkness. The DN1p can drive evening activity during DDTC 29°C:20°C and during low intensity LD cycles when the temperature is constant, while at high light intensity LD cycles they support morning activity. Furthermore, the DN1p receive warm input via the TrpA1 expressing AC neurons. Interestingly however, TrpA1 is not required for temperature entrainment as such, but shapes behavior under warm conditions. Notably, while the loss of TrpA1 function has no effect on the behavioral phase in DDTC 25°C:16°C, TrpA1 mutant flies show an advanced increase of activity and a reduced siesta in DDTC 29°C:20°C compared to controls. Hence, taken together these data re-enforce the model where, depending on the environmental condition, the clock network can change its balance to set behavioral activity phase, bypassing a change of clock entrainment (Lorber, 2022).
The molecular clock in brain clock neurons is entrained by light and temperature. However, these two environmental inputs vary on a day to day basis (e.g., cloudy versus sunny days), and animals change their activity at different times of day in response to the ambient environmental condition. How does the circadian system distinguish between an input that entrains the circadian clock (Zeitgeber) and one that after integration by the clock system, sets the daily activity phase? This study demonstrates that the EO determines the behavioral evening peak (Φ) in the presence of light and 25°C, while the circadian clock is entrained with a 25°C:16°C temperature cycle. In contrast, the MO determines the morning peak in constant darkness and 25°C, during the same temperature cycle. A model is presented explaining how the environment and the circadian clock shape locomotor activity (see Environmental cues act as Zeitgeber and set behavioral activity Phase). On one hand, the on/up and off/down environmental changes that happen once a day entrain the molecular clocks in the system. On the other hand, different environmental input such as light (quality and intensity) and temperature (different levels) are perceived by different oscillators in different manners. Depending on the time of day (the molecular clock status of the system), this will lead to a modification of the network balance and a dominancy of one or several oscillators (blue clocks vs grey ones), resulting in a behavioral activity phase (Φ) according to the internal timing and the current condition. Therefore, to biologically demonstrate this model, a Zeitgeber (in this study TC 25°C-16°C) was fixed, and how light (here presence/absence) modifies the balance was tested. From this basis, it is possible to apply more subtle modifications such as the intensity or the quality of light to change the phase of the behavior and use this to understand the principles underlying neuronal network switches (Lorber, 2022).
Furthermore, the data suggest that the length of the light period (here 12 h vs 24 h) influences the involvement of the EO. Ablating the PDF neurons not only advances the evening peak in LD but it also drastically reduces the synchronization of the flies (S6B and S6C Fig), while they remain strongly synchronized in LLTC. Furthermore, while ablating the EO, even with incomplete penetrance, drastically affected the presence of the synchronized evening peak in LLTC, it only slightly reduced synchronization in LD. Hence, although the clock of the EO can be entrained independently to light , the data indicate that the PDF neurons have a strong influence on the robustness of EO-output in LD25°C but not in LLTC. This suggests that the extent of dark periods increases the influence of the PDF neurons during the day. The evening peak occurs under the same environmental condition (lights-on and 25°C), however with a slight advance in LLTC compared to LD. Therefore, it is proposed that the past experience during the night/cryophase determines the influence of one group toward the other. In summary, these data confirm that the network status varies with the environmental condition, and therefore it is crucial to consider the specific environmental conditions when deciphering the different contributions of circadian network components in regulating behavioural activity. This simple environmental protocol provides a template to further test and extend this model. For example, using temperature cycles as Zeitgeber, it is now possible to modify light quality and intensity to test how the clock network responds to these light variations while the clock is stably entrained (Lorber, 2022).
Synchronization of clock protein oscillations in LPN neurons is preferentially sensitive to temperature. However, it has been observed that these neurons are not necessary for rhythmic behavior under TC conditions in both LL and constant darkness. This study confirms the non-requirement of clock function within the LPN for rhythmic locomotor behavior in LLTC, suggesting that LPN sensitivity for molecular synchronization to TC serves another function unrelated to entrainment. Indeed, a recent study shows that the LPN are activated above 27° and they are important for increased siesta sleep at 30°C. CRY is expressed in about 1/3rd of the clock network in the brain and plays an important role in light entrainment, consistent with the idea that CRY+ neurons are more sensitive to light and CRY- cells are more sensitive to temperature. Notably, isolated brains can still be entrained to LD cycles, and in the absence of visual input all clock neurons are entrained to LD, indicating that the CRY+ neurons synchronize the remaining CRY- neurons non-cell autonomously. However, this study demonstrates an essential role of the CRY+ neurons in controlling rhythmic behavior in LLTC. Nevertheless, even in the absence of all CRY+ neurons, flies exhibit weak synchronized behavior with an evening phase during temperature cycles (in LL and DD), suggesting that CRY- clock neurons (subsets of DN1p and LNd, DN2, and DN3) also play a role in temperature synchronization. Nevertheless, because they are not able to instruct the CRY+ neurons during TC , their role in temperature entrainment is not as prominent as that of CRY+ neurons in LD. Moreover, ablation of PDF+ or all three groups of LN (LNv, LNd, LPN) still allows for weak behavioral evening synchronization to 30°C: 25°C TC in LL. Furthermore, the fact that in the absence of a clock in the Mai179+ cells, after ablation of all CRY+ cells, or in the absence of all LN, molecular oscillations in other clock neurons can be synchronized, supports the idea that multiple and independent pathways contribute to temperature entrainment (Lorber, 2022).
In all organisms with circadian clocks, post-translational modifications of clock proteins control the dynamics of circadian rhythms, with phosphorylation playing a dominant role. All major clock proteins are highly phosphorylated, and many kinases have been described to be responsible. In contrast, it is largely unclear whether and to what extent their counterparts, the phosphatases, play an equally crucial role. To investigate this, a systematic RNAi screen was performed in human cells and protein phosphatase 4 (PPP4) was identified with its regulatory subunit PPP4R2 as critical components of the circadian system in both mammals and Drosophila. Genetic depletion of PPP4 (Pp4-19C in Drosophila) shortens the circadian period, whereas overexpression lengthens it. PPP4 inhibits CLOCK/BMAL1 transactivation activity by binding to BMAL1 (Cycle in Drosophila) and counteracting its phosphorylation. This leads to increased CLOCK/BMAL1 DNA occupancy and decreased transcriptional activity, which counteracts the "kamikaze" properties of CLOCK/BMAL1. Through this mechanism, PPP4 contributes to the critical delay of negative feedback by retarding PER/CRY/CK1Δ-mediated inhibition of CLOCK/BMAL1 (Klemz, 2021).
Sleep is essential for a variety of plastic processes, including learning and memory. However, the consequences of insufficient sleep on circuit connectivity remain poorly understood. To better appreciate the effects of sleep loss on synaptic connectivity across a memory-encoding circuit, changes were examined in the distribution of synaptic markers in the Drosophila mushroom body (MB). Protein-trap tags for active zone components indicate that recent sleep time is inversely correlated with Bruchpilot (BRP) abundance in the MB lobes; sleep loss elevates BRP while sleep induction reduces BRP across the MB. Overnight sleep deprivation also elevated levels of dSyd-1 and Cacophony, but not other pre-synaptic proteins. Cell-type-specific genetic reporters show that MB-intrinsic Kenyon cells (KCs) exhibit increased pre-synaptic BRP throughout the axonal lobes after sleep deprivation; similar increases were not detected in projections from large interneurons or dopaminergic neurons that innervate the MB. These results indicate that pre-synaptic plasticity in KCs is responsible for elevated levels of BRP in the MB lobes of sleep-deprived flies. Because KCs provide synaptic inputs to several classes of post-synaptic partners, a fluorescent reporter for synaptic contacts was used to test whether each class of KC output connections is scaled uniformly by sleep loss. The KC output synapses that were observed in this study can be divided into three classes: KCs to MB interneurons; KCs to dopaminergic neurons, and KCs to MB output neurons. No single class showed uniform scaling across each constituent member, indicating that different rules may govern plasticity during sleep loss across cell types (Weiss, 2021).
This study used genetic reporters to quantify the effects of sleep loss on pre-synaptic active zone markers and putative synaptic contacts in the Drosophila MB lobes. Abundance of Brp, dSyd-1, and Cacophony broadly increase across all MB lobes after overnight sleep deprivation and that acutely increasing sleep for 6 h is sufficient to reduce Brp levels across the α, 'β, γ, and 'β' lobes. KCs strongly contribute to the increase in Brp across each MB lobe following sleep loss, while pre-synapses of other MB cell types are less sensitive to sleep disruption. Because release of Drosophila neuromodulators likely occurs through a combination of classical neurotransmission and extrasynaptic release, these studies do not rule out the possibility that BRP-independent secretion of dopaminergic dense core vesicles might be altered in the MB lobes by sleep loss. The elevated levels of Brp present in KCs of sleep-deprived flies return to control levels within 48 h of ab libitum recovery sleep. While associative learning can recover within only a few hours after sleep deprivation, these studies indicate that some synaptic consequences of prolonged waking may persist for at least 24 h of recovery. These findings parallel those from humans and rodents, suggesting that some measures of cognition and neurophysiology recover rapidly after acute sleep loss while others last much longer, even for several days in some cases. The tractability of Drosophila may provide opportunities for future studies to investigate the processes that mediate recovery from sleep loss and to test whether similar trends in plasticity occur in other neuropil regions across the brain (Weiss, 2021).
Interestingly, sleep deprivation does not seem to increase other active zone components: Rim and Syt1 only show localized changes in some MB lobes, and the primarily post-synaptic marker Dlg shows no significant changes across the MB after sleep loss. Additionally, it was found that the abundance of vesicular proteins Rab3 and nSyb decreases across all MB lobes following overnight sleep deprivation. The varying responses between pre-synaptic components may indicate that sleep deprivation may alter the abundance of some active zone constituents along differing time courses or that active zone release machinery may be regulated differently from synaptic vesicle pools. The varied responses of each synaptic reporter that was observed suggests that Brp, dSyd-1, and Cac levels may underlie the consequences of sleep loss on MB functioning, but the precise physiological consequences of these changes on KC neurotransmitter release are unclear. Previous work finds that increasing BRP gene copy number drives changes in other active zone proteins that recapitulate protein levels observed in short sleeping mutants and also increases sleep in a dose-dependent manner (Weiss, 2021).
It is tempting to speculate that increases in Brp with sleep loss may drive concomitant increases in some core active zone scaffolding components and compensatory decreases in some proteins regulating synaptic vesicle release. Experiments at the Drosophila larval NMJ indicate that elevated Brp levels increase the rate of spontaneous release and enhance facilitation with pairs of stimuli, while other markers of synapse strength, including the amplitudes of evoked and spontaneous junction potentials, remained unchanged (Weiss, 2021).
It is unclear whether acute changes in Brp with sleep loss induce the same physiological changes at MB-output synapses, and additional studies will be required to understand how plastic mechanisms that contribute to memory formation might be altered by the pre-synaptic changes described above. Recent work finds that pan-neuronal knockdown of dSyd-1 can reduce sleep and dampen homeostatic rebound, even in flies with elevated BRP (Weiss, 2021).
Consistent with the idea that dSyd-1 levels may influence sleep pressure, decreased dSyd-1MI05387-GFSTF abundance was found in previously sleep-deprived flies after 48 h of recovery.
While the MB contains several different cell types, pre-synapses in the axons of KCs appear to be uniquely plastic during sleep loss. Use of an activity-dependent fluorescent GRASP reporter of synaptic contacts observed that sleep loss altered synaptic contacts between KCs and distinct post-synaptic partners in different ways (Weiss, 2021).
Among these changes, it was found that GRASP fluorescence reporting contacts from KCs to PPL1 DANs is strongly decreased after sleep loss, indicating a weakening of the KC > PPL1 DAN contacts. Interestingly, these connections may be vital for recurrent activation within MB compartments during learning and could contribute to prediction error signals (Weiss, 2021).
While further studies will be required to examine the contribution of these particular connections to learning deficits after sleep loss, human subjects have been reported to exhibit impaired error prediction and affective evaluation in learning tasks following sleep loss (Weiss, 2021).
Because reduced GRASP signal was observed in KC > PPL1 DAN connections, which mediate aversive reinforcement, and not in KC > PAM DAN connections, which influence appetitive reinforcement,
it is also possible that sleep loss may not equally degrade the encoding of reinforcement signals across all valences or modalities. Recent findings also suggest that not all forms of memory require sleep for consolidation; appetitive olfactory memories can be consolidated without sleep when flies are deprived of food, and sleep-dependent and independent memory traces in these conditions are stored in separate MB zones (Weiss, 2021).
The KC > MBON connections that contribute to sleep-dependent memory (KC > γ2α'1) also show an overall decrease in GRASP signal with sleep loss, while those that are vital for sleep-independent memory (MBON-γ1pedc) show no GRASP change after sleep deprivation. These compartment-specific variations in the effects of sleep on both memory and synaptic distribution further indicate that local MB zones may follow distinct plasticity rules under physiological stressors, including sleep loss (Weiss, 2021).
Additionally, GRASP signal from KCs to APL is significantly elevated following sleep loss, suggesting a strengthening of KC > APL connections. KCs and APL form a negative feedback circuit, where KCs activate APL and APL inhibits KCs: this feedback inhibition maintains sparseness of odor coding and odor specificity of memories (Weiss, 2021).
It is possible that KCs compensate for increased synaptic abundance accumulated during sleep loss by recruiting inhibition from APL. While further experimentation is needed to examine the role of these connections in the regulation of net synaptic strength during sleep loss, sleep deprivation results in increased cortical excitability in humans and rodents, and hyperexcitability is often counteracted by increased synaptic inhibition (Weiss, 2021).
Conversely, sleep loss reduces connectivity between KCs and DPM, a second large interneuron that may facilitate recurrent activity in the MB lobes. The current results also indicate that KC > MBON synaptic contacts exhibit a variety of changes in response to sleep deprivation. The specific KC > MBON connections that show significantly elevated or reduced GRASP signal in this study are not clearly assorted based on valence encoding, contribution to specific associative memory assays, or influence on sleep/wake regulation (Weiss, 2021).
Activity in several MB cell types, including α'/'β' KCs, MBON-γ5'β'2, MBON-γ2 α'1, DPM, and PAM DANs regulate sleep. The observation that KC > MBON-γ5'β'2a labeling is reduced with sleep loss complements previous observations of reduced electrical activity in MBON-γ5'β'2 following sleep deprivation (Weiss, 2021).
Other sleep-promoting MB neurons, however, such as DPM, do not show an overall increase in BRP abundance, suggesting either that other changes in excitability, synaptic drive, or post-synaptic adaptations might drive homeostatic sleep regulation in these cells or that distinct subsets of connections within the populations that were labelled in this study might be sleep regulatory. The compartment-to-compartment variance in KC > MBON responses to sleep loss also parallels previous findings that plasticity rules can vary between MBONs during heterosynaptic plasticity (Weiss, 2021).
While GRASP results suggest diverse changes in putative synaptic contacts with sleep loss, the functional effects of these changes require further study. It is important to note that a significant portion of MB synapses are composed of connections between either pairs or groups of KCs. The genetic strategies that were used in this study have prevented reliable visualization and quantification of these connections. As a result, the effect of sleep loss on KC > KC synapses has not been examined in this study but may comprise a portion of the increase in KC pre-synaptic abundance that was observed in this study. While these studies identify synaptic classes that exhibit altered GRASP labeling across sleep loss, future studies using super resolution imaging and/or physiology could examine the structural and molecular changes that underlie this plasticity. Connections between neurons in the MB may be also influenced by non-neuronal cell types, including astrocytes. Astrocytic contact with KCs can be reduced by sleep loss and astrocytic calcium levels correlate with sleep pressure, which suggests that astrocytic processes could be positioned to mediate sleep-dependent plasticity in the MB (Weiss, 2021).
The broad conservation of release machinery across active zones within and between cell types has simplified examination of pre-synaptic plasticity during sleep loss. Assays of both Hebbian and homeostatic plasticity have also identified a variety of post-synaptic adaptations. Interestingly, post-synaptic densities isolated from rodent cortex show significant reorganization of post-synaptic GluR5 receptors. This depends upon the activity of Homer and sleep-dependent phosphorylation of CaMKII and GluR1, that contribute to consolidation of visual cortex plasticity (Weiss, 2021).
Because MBONs exhibit post-synaptic plasticity during other contexts, including the formation of associative memories, sleep deprivation may also alter post-synaptic organization of MBONs or other cell types in the MB. Although the distribution of Dlg is not significantly changed by sleep loss, the rich variety of post-synaptic receptors for acetylcholine, dopamine, GABA, and other signals in the MB requires development of additional reporters to examine these post-synaptic consequences of insufficient sleep in MB neurons. Additionally, while the data outline changes in pre-synaptic protein abundance and pre-synaptic KC contacts that result from sleep loss, the possibility that these synaptic changes may be accompanied by homeostatic compensation in neuronal excitability or firing patterns remains to be tested. Because sleep-deprived flies can recover the capacity to learn after only a brief nap, homeostatic adjustments in post-synaptic strength and/or excitability may permit MBs to compensate for pre-synaptic changes that appear to persist for at least 24 h after sleep deprivation. Further, recovery sleep or pharmacological sleep enhancement may not simply reverse the effects of sleep loss, and it is unclear how particular subsets of synaptic proteins or connections may be selected for removal during times of elevated sleep (Weiss, 2021).
The consequences of sleep loss on synaptic connectivity are not clearly understood, but previous work has found net changes in synaptic abundance or size across brain regions. This study characterized a diverse array of synaptic responses to sleep loss among different cell types within the same circuit. These findings may suggest that distinct cell types and connections within the MB are governed by heterogeneous plasticity rules during sleep disruption. While previous studies have characterized the synaptic effects of sleep history on individual cell types within plastic circuits, the data provide a more comprehensive understanding of the consequences of sleep loss on MB circuits. While this project outlines the local effects of sleep loss on MB connectivity, it is unclear whether specific neural subsets also drive BRP increases within other neuropil compartments of sleep-deprived brains (Weiss, 2021).
This study found an overall increase in the abundance of reporters for some, but not all, pre-synaptic proteins. These pre-synaptic changes are not distributed equally across all cell types: they are most pronounced in MB-intrinsic KCs. Further, output connections from KCs to different classes of synaptic partners show varying patterns of plasticity in MB sub-circuits that contribute to encoding odor valence, comprise recurrent feedback loops, or relay reinforcement signals. The results indicate that sleep loss may degrade MB-dependent memory by altering several different classes of synapses, but future studies will be required to test the specific roles of changes at individual synapse types and the mechanisms by which prolonged waking reorganizes MB connectivity (Weiss, 2021).
The central complex is one of the major brain regions that control sleep in Drosophila. However, the circuitry details of sleep regulation have not been elucidated yet. This study shows a novel sleep-regulating neuronal circuit in the protocerebral bridge (PB) of the central complex. Activation of the PB interneurons labeled by the R59E08-Gal4 and the PB columnar neurons with R52B10-Gal4 promoted sleep and wakefulness, respectively. A targeted GFP reconstitution across synaptic partners (t-GRASP) analysis demonstrated synaptic contact between these two groups of sleep-regulating PB neurons. Furthermore, it was found that activation of a pair of dopaminergic (DA) neurons projecting to the PB (T1 DA neurons) decreased sleep. The wake-promoting T1 DA neurons and the sleep-promoting PB interneurons formed close associations. Dopamine 2-like receptor (Dop2R) knockdown in the sleep-promoting PB interneurons increased sleep. These results indicated that the neuronal circuit in the PB, regulated by dopamine signaling, mediates sleep-wakefulness (Tomita, 2021).
Sleep is controlled by homeostatic mechanisms, which drive sleep after wakefulness, and a circadian clock, which confers the 24-h rhythm of sleep. These processes interact with each other to control the timing of sleep in a daily cycle as well as following sleep deprivation. However, the mechanisms by which they interact are poorly understood. These studies show that hugin (+) neurons, previously identified as neurons that function downstream of the clock to regulate rhythms of locomotor activity, are also targets of the sleep homeostat. Sleep deprivation decreases activity of hugin (+) neurons, likely to suppress circadian-driven activity during recovery sleep, and ablation of hugin (+) neurons promotes sleep increases generated by activation of the homeostatic sleep locus, the dorsal fan-shaped body (dFB). Also, mutations in peptides produced by the hugin (+) locus increase recovery sleep following deprivation. Transsynaptic mapping reveals that hugin (+) neurons feed back onto central clock neurons, which also show decreased activity upon sleep loss, in a Hugin peptide-dependent fashion. It is proposed that hugin (+) neurons integrate circadian and sleep signals to modulate circadian circuitry and regulate the timing of sleep (Schwarz, 2021).
Understanding how neural circuits underlie behaviour is challenging even in the connectome era because it requires a combination of anatomical and functional analyses. This is exemplified in the circuit underlying the light avoidance behaviour displayed by Drosophila melanogaster larvae. While this behaviour is robust and the nervous system relatively simple, the circuit is only partially delineated with some contradictions among studies. This study devised trans-Tango MkII, an offshoot of the transsynaptic circuit tracing tool trans-Tango, and implement it in anatomical tracing together with functional analysis. Neuronal inhibition was used to test necessity of particular neuronal types in light avoidance and selective neuronal activation to examine sufficiency in rescuing light avoidance deficiencies exhibited by photoreceptor mutants. These studies reveal a four-order circuit for light avoidance connecting the light-detecting photoreceptors with a pair of neuroendocrine cells via two types of clock neurons. This approach can be readily expanded to studying other circuits (Sorkac, 2022).
The study revealed a circuit consisting of four orders of neurons that connect the Rh5 photoreceptors to PTTH neurons via the 5th-LaN and DN2s. While this circuit mediates the response to bright light, the observation that DN1s are necessary for photophobic response only to low light intensity indicates the existence of an additional pathway for dim light. It is noteworthy that a third, independent system has been reported in which a gustatory receptor mediates photophobic response to high-intensity light in class IV multidendritic neurons. This study has no information as to at which level these pathways might meet, if at all, upstream of the motor neurons (Sorkac, 2022).
The results clarify several earlier studies regarding the role of Pdf-LaNs in light avoidance. In the current experiments, Pdf-LaNs are dispensable for light avoidance, yet their activation is attractive. A potential explanation is that Pdf-LaNs may modulate larval photophobia via inhibition, especially since adult Pdf-LaNs are glycinergic. In addition, the results contradict a previous study reporting that Pdf-LaNs are presynaptic to PTTH neurons. This study relied on a version of GRASP that is in fact not synaptic. Thus, the proposed connection could have been the result of a non-synaptic reconstitution of GFP due to proximity especially since reconstituted GFP was not observed using a synaptic version of GRASP. This result, however, does not rule out a Pdf-LaN-mediated inhibition of the light avoidance circuit from the Rh5 photoreceptors to PTTH neurons. It is conceivable that, alongside their roles in alternative circuits that mediate this behaviour, Pdf-LaNs play inhibitory roles in this circuit as well. Indeed, ablating Pdf-LaNs increases the activity in PTTH neurons as revealed by the GCaMP signal (Sorkac, 2022).
Our analysis of the robust light avoidance response in larvae exemplifies the importance of employing a comprehensive approach combining circuit tracing together with neuronal inhibition and activation to test necessity and sufficiency. Circuit epistasis analysis was made possible by trans-Tango MkII, a new version of trans-Tango that allows researchers to trace and manipulate neural circuits in Drosophila larvae. The combination of a robust and user-friendly genetic tool such as trans-Tango MkII with careful functional analysis constitutes a powerful approach that can be readily expanded to studying other circuits and behaviours (Sorkac, 2022).
Small poikilotherms such as the fruit fly Drosophila depend on absolute temperature measurements to identify external conditions that are above (hot) or below (cold) their preferred range and to react accordingly. Hot and cold temperatures have a different impact on fly activity and sleep, but the circuits and mechanisms that adjust behavior to specific thermal conditions are not well understood. This study use patch-clamp electrophysiology to show that internal thermosensory neurons located within the fly head capsule (the AC neurons(1)) function as a thermometer active in the hot range. ACs exhibit sustained firing rates that scale with absolute temperature-but only for temperatures above the fly's preferred ~25°C (i.e., "hot" temperature). ACs were identified in the fly brain connectome and demonstrate that they target a single class of circadian neurons, the LPNs.(2) LPNs receive excitatory drive from ACs and respond robustly to hot stimuli, but their responses do not exclusively rely on ACs. Instead, LPNs receive independent drive from thermosensory neurons of the fly antenna via a new class of second-order projection neurons (TPN-IV). Finally, silencing LPNs blocks the restructuring of daytime "siesta" sleep, which normally occurs in response to persistent heat. Previous work described a distinct thermometer circuit for cold temperature.(3) Together, the results demonstrate that the fly nervous system separately encodes and relays absolute hot and cold temperature information, show how patterns of sleep and activity can be adapted to specific temperature conditions, and illustrate how persistent drive from sensory pathways can impact behavior on extended temporal scales (Alpert, 2022).
Sleep is required to maintain physiological functions and is widely conserved across species. To understand the sleep-regulatory mechanisms This study reports a novel sleep-regulatory pathway that promotes arousal. This study first focused on FB interneurons and found that cholinergic FB interneurons promoted arousal. The arousal-promoting effect of FB interneurons was confirmed by using more specific drivers. These drivers label FB interneurons which receive input from P-FN neurons (that project from the protocerebral bridge to the ventral FB and the NO) and send output to dFB neurons. There should be other FB interneurons that do not have a connection with P-FN neurons or dFB neurons. It means that FB interneurons labeled by two split drivers are only a part of FB interneurons. Therefore we considered that the weaker effects were due to the smaller number of neurons labeled by split-Gal4 lines than NP2320, not the effect of neurons other than FB interneurons. No aclear sleep rebound was observed after neuronal activation. In the previous study, R52B10-Gal4 was used which is reported to drive sleep rebound in the female fly. A clear sleep rebound was observed in female flies but not in male flies. These results indicated that there is a sex difference in the regulation of sleep rebound, at least, in R52B10 neurons. The current study used only male flies and this could be one of the reasons why no clear sleep rebound was observed. It was next asked about the relationship between FB interneurons and known sleep-regulatory circuits. GRASP and Ca2+ experiments were performed, and FB interneurons and R52B10 neurons which label the output neurons of the PB-FB pathway were shown to be anatomically and functionally connected . Although the possibility of other pathways downstream to R52B10 cannot be excluded, this study demonstrated clearly that FB interneurons are one of the downstream to R52B10. Further study will show the impact size of the connection between them in sleep regulation. To investigate the postsynaptic partners of FB interneurons, a GRASP experiment was conducted. FB interneurons and dFB neurons labeled by R23E10 were found to form close associations. Besides, according to the connectome paper and connectome dataset, vDeltaB, C, D, and hDeltaC receive input from P-FN neurons and send output to FB tangential neurons which arborize in layers 6 and 7. This information also supported the idea that R52B10 neurons including P-FN neurons, FB interneurons like vDeltaB, C, D, and hDeltaC neurons, and dFB neurons that are consisted of FB tangential neurons which arborize in layers 6 and 7 form a neuronal circuit. Future study will clarify their functional connection and the role of this circuit in sleep regulation. Furthermore, a previous study showed that neurons that project to the ventral FB (vFB neurons) promote sleep and mediate consolidation of long-term memory. Since axon terminals and dendrites of FB interneurons arborize in both the dorsal and ventral FB, there would be interactions between dFB and vFB neurons via FB interneurons. Further research will clarify the functional relationship between these neurons (Kato, 2022).
According to previous reports, FB interneurons regulate optomotor behavior and express tachykinin, a neuropeptide that regulates aggression. Additionally, T1 dopamine neurons, which are upstream of R52B10 neurons, regulate aggression as well. Besides, P2 neurons, which include FB interneurons, regulate chronic isolation evoked sleep loss. Moreover, courtship-regulator P1 neurons activate T1 neurons and modulate sleep/courtship balance based on the nutritional status. Taking all the information mentioned above into account, it is considerd that arousal signals related to aggression, courtship, nutrition, and vision converge into the PB-FB pathway including FB interneurons to regulate arousal. Further studies should clarify the role of these arousal signals on the PB-FB pathway and FB interneurons in sleep regulation (Kato, 2022).
In conclusion, the results provided possible sleep-regulatory neurons that may connect with the PB-FB pathway and dFB neurons. It is hypothesized that arousal signals are sent from the PB-FB pathway to FB interneurons, inhibit dFB neurons via inhibitory signals, and regulate sleep (Kato, 2022).
The mushroom body lobes are tiled by discrete anatomic compartments defined by the axons of a specific subset of DANs and the dendrites of one or two mushroom body output neurons (MBONs). This anatomical arrangement positions DANs to strategically convey positive and negative reinforced information by changing the synaptic weight of KC-MBONs in producing aversive and appetitive responses (Driscoll, 2021).
While, the most in-depth analysis of these synapses and distinct DAN-KC-MBON connectivity and behavioral output comes from studies of olfactory conditioning, there is evidence that these synapses play a critical role in innate behaviors like feeding and sleep. Although, role of DA on sleep has been extensively investigated in Drosophila, the commonly used TH-Gal4 driver line labels most dopamine neuron clusters, but is absent from the several PAM clusters that projects to MB (Driscoll, 2021).
This study specifically probed PAM subsets that project to γ5, γ4, and β'2 MB compartments. This study focused on this subset because KCs and MBONs downstream of these PAM neurons can be neuroanatomically resolved and have been shown to be required for wakefulness. Further, KCs and MBONs that form the γ5, γ4, and β'2 synaptic compartments alter their spontaneous neural activity in response to sleep need (induced by mechanical sleep-deprivation). The ability to use cell-specific split-GAL4 tools provides opportunity to resolve the precise circuit mechanisms by which PAM neurons regulate wakefulness (Driscoll, 2021).
GABA signaling also modulates sleep and wake microcircuits within MB. The key source of GABA in the MB is anterior paired lateral neurons, APL and dorsal paired medial neurons (DPM), which are electrically coupled and increase sleep by GABAergic inhibition of wake-promoting KCs. In the context of associative learning, there is strong evidence for interactions between KCs, APL, DPM and DANs but it is not clear if GABA and dopamine signaling represent opposing inputs to the KCs and MBONs in the regulation of sleep. This study found that the excitability of PAM DANs involved in wakefulness is blocked by sleep-promoting GABA signaling and mediated by ionotropic receptor subtype GABAA-Rdl (Driscoll, 2021).
A recent study showed that GABA inhibitory input to the presynaptic terminals of the PAM neurons regulates appetitive memory and that this interaction is mediated by GABA-B3 receptors that are clustered in PAM boutons localized to PAM-γ5 and -α1 compartments. These data are consistent with the findings that PAM-γ5 are GABA responsive and that multiple receptors are critical to this interaction. Since, no role was found for GABA-B3 in PAM mediated sleep regulation, it is likely that PAM γ5, γ4, and β'2 express multiple GABA receptors which are differentially recruited in sleep and learning. How and what regulates the expression of these receptors in PAM subsets presents a potential mechanism of presynaptic gating to MB core circuits. Transcriptomic analysis of PAM neurons reveals extremely high levels of Rdl expression followed by GABA-B3. Among the PAM subsets mean TPM or transcripts per million of Rdl receptor in PAM γ5, γ4, and β'2 are much higher as compared to other PAM subsets (Driscoll, 2021).
Simple connection query search of the recently released hemibrain data85 reveals there is significant bidirectional connectivity between APL, DPM, and PAM neurons. Further, a recent study showed that APL neurons express the inhibitory D2R receptor55. APL mediated GABAergic inhibition of the PAM neurons was recently shown to control the intensity and specificity of olfactory appetitive memory but previous results show that blocking GABA release from APL neurons only modestly affects sleep phenotypes (Driscoll, 2021).
While, the role of APL in GABA signaling to PAM γ5, γ4, and β'2 cannot be completely ruled out, other inputs to wake-regulating PAM DANs could also be GABAergic and critical for promoting sleep. A recent study using EM dataset of a Full Adult Female Fly Brain (FAFB) mapped the inputs and outputs of the PAMγ5 DANs and identified that this cell type is highly heterogenous and in addition to recurrent feedback from MBON01 γ5β'2a, it receives extensive input from other MBONs, sub-esophageal output neurons (SEZONs) and lateral horn output neurons86. The EM data also reveals that octopaminergic neurons synapse onto PAM γ5, γ4, and β'2 DANs. Whether, these inputs play a role in wakefulness is unknown but suggests that the PAMγ5 could serve as a key link between sensory inputs, wake-promoting octopamine signal and core sleep regulating circuitry within the MB. Each of these inputs could modulate PAM-DAN activity and dopamine release in regulating wakefulness via the MB (Driscoll, 2021).
In addition to probing the release and activity of these PAM-DANs the dopamine receptors and their location within the MB in signaling wakefulness were also explored. To this end validated RNAi lines were expressed in subsets of KCs and MBONs; DopR1 and DopR2 were found to be critical in mediating the wakefulness signal via KCs and γ5β'2 MBONs. Knocking down the receptor consistently increased total sleep and bout length. Furthermore, specific manipulations of DopR receptors within the MB did not directly alter locomotor activity as observed by manipulation of these receptors in CX. Although, loss-of-function mutations of D1 dopamine receptor DopR are shown to enhance repetitive air puff startle-induced arousal and increase sleep. Expression and restoration of DopR in the mutant background specifically in the central complex rescues the startle response, while, the sleep phenotype is rescued via a broad MB driver. The current data extends these findings by showing that the DopR receptors regulate sleep via the MB γ5 and β'2 compartment. Although, targeted RNAi experiments show that DopR's are required for sleep regulation by KCs and MBONs, the lack of a sleep phenotype in DopR2 mutant could be a result of global loss of receptor in the mutant as opposed to targeted loss of receptor function within MB. Dopamine signals wakefulness by activation of wake-promoting neurons of MB via DopR1 and DopR2 and within. the central complex, neurons of dFB are inhibited by dopamine via DopR2. Hence, DopR2 has opposing effects within MB and CX (Driscoll, 2021).
In vitro characterization indicates that DopR's signal through distinct G-proteins, with DopR1 via Gαs to stimulate cAMP production and DopR2 coupling to Gαq via increased calcium. These receptors are thought to have differential sensitivity to dopamine and could be potentially recruited by varying DA release or DAN activity. In the context of sleep regulation, this work reveals that both DopR1 and DopR2 induce wakefulness via the γ5 β'2 MB compartment but not γ4 compartment. Although, chronic activation of PAM γ4 induces wakefulness, the glutamatergic MBON γ4 < γ1,2 projects to multiple compartments and could potentially activate or inhibit MBONs and PAMs projecting to γ1 and γ2 compartment. The interaction between compartments is not well understood in the context of sleep and wake regulation and requires further investigation to better understand the role of DopR2 in regulating the γ4 compartment. The neuroanatomical specificity obtained from split-Gal4 lines combined with EM data has paved way for more detailed analysis of the role of dopamine signaling to MB in the context of sleep and other behaviors (Driscoll, 2021).
The sleep-regulating PAM DANs and associated KCs and MBONs identified in this study are also involved in mediating satiety, novelty, caffeine induced arousal, punishment and reward associated experiences suggesting that the activity of these neurons is tuned to several wake and arousal associated behaviors. This is further supported by the EM connectome data showing that MB receives extensive gustatory, auditory and visual input in addition to olfactory input (Driscoll, 2021).
Current models of sleep regulation rely on two main processes, the circadian clock and the sleep homeostat and don't completely account for multiple external and internal factors that influence wakefulness. The ability to sleep, however, is influenced by motivational or cognitive stimuli. It is therefore envisioned that sleep, wakefulness and arousal within MB are not located in distinct circuits, but rather mediated by distinct processes within a common circuit (Driscoll, 2021).
Time-restricted feeding (TRF) has recently gained interest as a potential anti-ageing treatment for organisms from Drosophila to humans. TRF restricts food intake to specific hours of the day. Because TRF controls the timing of feeding, rather than nutrient or caloric content, TRF has been hypothesized to depend on circadian-regulated functions; the underlying molecular mechanisms of its effects remain unclear. To exploit the genetic tools and well-characterized ageing markers of Drosophila, this study developed an intermittent TRF (iTRF) dietary regimen that robustly extended fly lifespan and delayed the onset of ageing markers in the muscles and gut. iTRF enhanced circadian-regulated transcription, and iTRF-mediated lifespan extension required both circadian regulation and autophagy, a conserved longevity pathway. Night-specific induction of autophagy was both necessary and sufficient to extend lifespan on an ad libitum diet and also prevented further iTRF-mediated lifespan extension. By contrast, day-specific induction of autophagy did not extend lifespan. Thus, these results identify circadian-regulated autophagy as a critical contributor to iTRF-mediated health benefits in Drosophila. Because both circadian regulation and autophagy are highly conserved processes in human ageing, this work highlights the possibility that behavioural or pharmaceutical interventions that stimulate circadian-regulated autophagy might provide people with similar health benefits, such as delayed ageing and lifespan extension (Ulgherait, 2021).
Sleep remains a major mystery of biology. In particular, little is known about the mechanisms that account for the drive to sleep. In an unbiased screen of more than 12,000 Drosophila lines, identified a single gene, nemuri (CG31813), that induces sleep. The NEMURI protein is an antimicrobial peptide that can be secreted ectopically to drive prolonged sleep (with resistance to arousal) and to promote survival after infection. Loss of nemuri increased arousability during daily sleep and attenuated the acute increase in sleep induced by sleep deprivation or bacterial infection. Conditions that increase sleep drive induced expression of nemuri in a small number of fly brain neurons and targeted it to the sleep-promoting, dorsal fan-shaped body. It is proposed that NEMURI is a bona fide sleep homeostasis factor that is particularly important under conditions of high sleep need; because these conditions include sickness, these findings provide a link between sleep and immune function (Toda, 2019).
Sleep-like states in diverse organisms can be separated into distinct stages, each with a characteristic arousal threshold. However, the molecular pathways underlying different sleep stages remain unclear. The fruit fly, Drosophila melanogaster, exhibits consolidated sleep during both day and night, with night sleep associated with higher arousal thresholds compared to day sleep. This study identified a role for the neuronal calcium sensor protein Neurocalcin (NCA) in promoting sleep during the night but not the day by suppressing nocturnal arousal and hyperactivity. Both circadian and light-sensing pathways define the temporal window in which NCA promotes sleep. Furthermore, NCA promotes sleep by suppressing synaptic release from a dispersed wake-promoting neural network and the mushroom bodies, a sleep-regulatory center, are a module within this network. These results advance the understanding of how sleep stages are genetically defined (ChenK, 2019).
Sleep is an ancient animal behavior that is regulated similarly in species ranging from flies to humans. Various genes that regulate sleep have been identified in invertebrates, but whether the functions of these genes are conserved in mammals remains poorly explored. Drosophila insomniac (inc) mutants exhibit severely shortened and fragmented sleep. Inc protein physically associates with the Cullin-3 (Cul3) ubiquitin ligase, and neuronal depletion of Inc or Cul3 strongly curtails sleep, suggesting that Inc is a Cul3 adaptor that directs the ubiquitination of neuronal substrates that impact sleep. Three proteins similar to Inc exist in vertebrates-KCTD2, KCTD5, and KCTD17-but are uncharacterized within the nervous system and their functional conservation with Inc has not been addressed. This study shows that Inc and its mouse orthologs exhibit striking biochemical and functional interchangeability within Cul3 complexes. Remarkably, KCTD2 and KCTD5 restore sleep to inc mutants, indicating that they can substitute for Inc in vivo and engage its neuronal targets relevant to sleep. Inc and its orthologs localize similarly within fly and mammalian neurons and can traffic to synapses, suggesting that their substrates may include synaptic proteins. Consistent with such a mechanism, inc mutants exhibit defects in synaptic structure and physiology, indicating that Inc is essential for both sleep and synaptic function. These findings reveal that molecular functions of Inc are conserved through ~600 million years of evolution and support the hypothesis that Inc and its orthologs participate in an evolutionarily conserved ubiquitination pathway that links synaptic function and sleep regulation (Li, 2017).
The presence of sleep states in diverse animals has been suggested to reflect a common purpose for sleep and the conservation of underlying regulatory mechanisms. This study has shown that attributes of the Insomniac protein likely to underlie its impact on sleep in Drosophila-its ability to function as a multimeric Cul3 adaptor and engage neuronal targets that impact sleep-are functionally conserved in its mammalian orthologs. This comparative analysis of Inc family members in vertebrate and invertebrate neurons furthermore reveals that these proteins can traffic to synapses and that Inc itself is essential for normal synaptic structure and excitability. These findings support the hypothesis that Inc family proteins serve as Cul3 adaptors and direct the ubiquitination of conserved neuronal substrates that impact sleep and synaptic function (Li, 2017).
The ability of KCTD2 and KCTD5 to substitute for Inc in the context of sleep is both surprising and notable given the complexity of sleep-wake behavior and the likely functions of these proteins as Cul3 adaptors. Adaptors are multivalent proteins that self-associate, bind Cul3, and recruit substrates, and these interactions are further regulated by additional post-translational mechanisms. The findings indicate that KCTD2 and KCTD5 readily substitute for Inc within oligomeric Inc-Cul3 complexes, and strongly suggest that these proteins recapitulate other aspects of Inc function in vivo including the ability to engage neuronal targets that impact sleep. The simplest explanation for why KCTD2 and KCTD5 have retained the apparent ability to engage Inc targets despite the evolutionary divergence of Drosophila and mammals is that orthologs of Inc targets are themselves conserved in mammals. This inference draws support from manipulations of Drosophila Roadkill/HIB and its mammalian ortholog SPOP, Cul3 adaptors of the MATH-BTB family that regulate the conserved Hedgehog signaling pathway. While the ability of SPOP to substitute for HIB has not been assessed by rescue at an organismal level, clonal analysis in Drosophila indicates that ectopically expressed mouse SPOP can degrade the endogenous HIB substrate Cubitus Interruptus (Ci), and conversely, that HIB can degrade mammalian Gli proteins that are the conserved orthologs of Ci and substrates of SPOP. By analogy, Inc targets that impact sleep are likely to have orthologs in vertebrates that are recruited by KCTD2 and KCTD5 to Cul3 complexes. While the manipulations do not resolve whether KCTD17 can substitute for Inc in vivo, the ability of KCTD17 to assemble with fly Inc and Cul3 suggests that functional divergence among mouse Inc orthologs may arise outside of the BTB domain, and in particular may reflect properties of their C-termini including the ability to recruit substrates (Li, 2017).
The finding that Inc can transit to synapses and is required for normal synaptic function is intriguing in light of hypotheses that invoke synaptic homeostasis as a key function of sleep. While ubiquitin-dependent mechanisms contribute to synaptic function and plasticity and sleep is known to influence synaptic remodeling in both vertebrates and invertebrates, molecular links between ubiquitination, synapses, and sleep remain poorly explored. Other studies in flies have indicated that regulation of RNA metabolism may similarly couple synaptic function and the control of sleep. Alterations in the activity of the Fragile X mental retardation protein (FMR), a regulator of mRNA translation, cause defects in the elaboration of neuronal projections and the formation of synapses as well as changes in sleep duration and consolidation. Loss of Adar, a deaminase that edits RNA, leads to increased sleep through altered glutamatergic synaptic function. Like Inc, these proteins are conserved in mammals, suggesting that further studies in flies may provide insights into diverse mechanisms by which sleep influences synaptic function and conversely, how changes in synapses may impact the regulation of sleep (Li, 2017).
These findings at a model synapse suggest that the impact of Inc on synaptic function may be intimately linked to its influence on sleep but do not yet resolve important aspects of such a mechanism. The synaptic phenotypes of inc mutants-increased synaptic growth, decreased evoked neurotransmitter release, and modest effects on spontaneous neurotransmission-are qualitatively distinct from those of other short sleeping mutants. Shaker (Sh) and Hyperkinetic (Hk) mutations decrease sleep in adults but increase both excitability and synaptic growth at the NMJ, suggesting that synaptic functions of Inc may affect sleep by a mechanism different than broad neuronal hyperexcitability. While a parsimonious model is that Inc directs the ubiquitination of a target critical for synaptic transmission both at the larval NMJ and in neuronal populations that promote sleep, this hypothesis awaits the elucidation of Inc targets, definition of the temporal requirements of Inc activity, and further mapping of the neuronal populations through which Inc impacts sleep. Finally, determining the localization of endogenous Inc within neurons is essential to distinguish possible presynaptic and postsynaptic functions of Inc and whether Inc engages local synaptic proteins or extrasynaptic targets that ultimately influence synaptic function (Li, 2017).
A clear implication of these findings is that neuronal targets and synaptic functions of Inc may be conserved in other animals. While the impact of Inc orthologs on sleep in vertebrates is as yet unknown, findings from C. elegans support the notion that conserved molecular functions of Inc and Cul3 may underlie similar behavioral outputs in diverse organisms. INSO-1/C52B11.2, the only C. elegans ortholog of Inc, interacts with Cul3, and RNAi against Cul3 and INSO-1 reduces the duration of lethargus, a quiescent sleep-like state, suggesting that effects of Cul3- and Inc-dependent ubiquitination on sleep may be evolutionarily conserved. The functions of Inc orthologs and Cul3 in the mammalian nervous system await additional characterization, but emerging data suggest functions relevant to neuronal physiology and disease. Human mutations at the KCTD2/ATP5H locus are associated with Alzheimer's disease, and mutations of KCTD17 with myoclonic dystonia. Cul3 lesions have been associated in several studies with autism spectrum disorders and comorbid sleep disturbances. More generally, autism spectrum disorders are commonly associated with sleep deficits and are thought to arise in many cases from altered synaptic function, but molecular links to sleep remain fragmentary. Studies of Inc family members and their conserved functions in neurons are likely to broaden understanding of how ubiquitination pathways may link synaptic function to the regulation of sleep and other behaviors (Li, 2017).
There is considerable variation in sleep duration, timing and quality in human populations, and sleep dysregulation has been implicated as a risk factor for a range of health problems. Human sleep traits are known to be regulated by genetic factors, but also by an array of environmental and social factors. These uncontrolled, non-genetic effects complicate powerful identification of the loci contributing to sleep directly in humans. The model system, Drosophila melanogaster, exhibits a behavior that shows the hallmarks of mammalian sleep, and this study used a multitiered approach, encompassing high-resolution QTL mapping, expression QTL data, and functional validation with RNAi to investigate the genetic basis of sleep under highly controlled environmental conditions. A battery of sleep phenotypes was measured in >750 genotypes derived from a multiparental mapping panel and identified several, modest-effect QTL contributing to natural variation for sleep. Merging sleep QTL data with a large head transcriptome eQTL mapping dataset from the same population allowed refining the list of plausible candidate causative sleep loci. This set includes genes with previously characterized effects on sleep and circadian rhythms, in addition to novel candidates. Finally, adult, nervous system-specific RNAi was used on the Dopa decarboxylase, dyschronic, and timeless genes, finding significant effects on sleep phenotypes for all three. The genes resolved in this study are strong candidates to harbor causative, regulatory variation contributing to sleep (Smith, 2020).
Most animals restrict their activity to a specific part of the day, being diurnal, nocturnal or crepuscular. The genetic basis underlying diurnal preference is largely unknown. Under laboratory conditions, Drosophila melanogaster is crepuscular, showing a bi-modal activity profile. However, a survey of strains derived from wild populations indicated that high variability among individuals exists, including flies that are nocturnal. Using a highly diverse population, an artificial selection experiment was performed, selecting flies with extreme diurnal or nocturnal preference. After 10 generations, highly diurnal and nocturnal strains were obtained. Whole-genome expression analysis was used to identify differentially expressed genes in diurnal, nocturnal and crepuscular (control) flies. Other than one circadian clock gene (pdp1), most differentially expressed genes were associated with either clock output (pdf, to) or input (Rh3, Rh2, msn). This finding was congruent with behavioural experiments indicating that both light masking and the circadian pacemaker are involved in driving nocturnality. This study demonstrates that genetic variation segregating in wild populations contributes to substantial variation in diurnal preference. Candidate genes associated with diurnality/nocturnality, while data emerging from expression analysis and behavioural experiments suggest that both clock and clock-independent pathways are involved in shaping diurnal preference. The diurnal and nocturnal selection strains provide us with a unique opportunity to understand the genetic architecture of diurnal preference (Pegoraro, 2020).
MiRNAs have attracted more attention in recent years as regulators of sleep and circadian rhythms after their roles in circadian rhythm and sleep were discovered. This study explored the roles of the miR-276a on daily sleep in Drosophila melanogaster, and found a regulatory cycle for the miR-276a pathway, in which miR-276a, regulated by the core CLOCK/CYCLE (CLK/CYC) transcription factor upstream, regulates sleep via suppressing targets TIM and NPFR1. (a) Loss of miR-276a function makes the flies sleep more during both daytime and nighttime, while flies with gain of miR-276a function sleep less; (b) MiR-276a is widely expressed in the mushroom body (MB), the pars intercerebralis (PI) and some clock neurons lateral dorsal neurons (LNds), in which tim neurons is important for sleep regulation; (c) MiR-276a promoter is identified to locate in the 8th fragment (aFrag8) of the pre-miR-276a, and this fragment is directly activated and regulated by CLK/CYC; (4) MiR-276a is rhythmically oscillating in heads of the wild-type w(1118), but this oscillation disappears in the loss of function mutant clk(jrk) ; (5) The neuropeptide F receptor 1 (npfr1) was found to be a downstream target of miR-276a. These results clarify that the miR-276a is a very important factor for sleep regulation (Zhang, 2021).
Sleep is a crucial factor for health and survival in all animals. This study found by proteomic analysis that some cancer related proteins were impacted by the circadian clock. The 14-3-3ε protein, expression of which is activated by the circadian transcription factor Clock, regulates adult sleep of Drosophila independent of circadian rhythm. Detailed analysis of the sleep regulatory mechanism shows that 14-3-3ε directly targets the Ultrabithorax (Ubx) gene to activate transcription of the pigment dispersing factor (PDF). The dopamine receptor (Dop1R1) and the octopamine receptor (Oamb), are also involved in the 14-3-3ε pathway, which in 14-3-3ε mutant flies causes increases in the dopR1 and OAMB, while downregulation of the DopR1 and Oamb can restore the sleep phenotype caused by the 14-3-3ε mutation. In conclusion, 14-3-3ε is necessary for sleep regulation in Drosophila (Wei, 2021).
Circadian transcriptional timekeepers in pacemaker neurons drive profound daily rhythms in sleep and wake. This study revealed a molecular pathway that links core transcriptional oscillators to neuronal and behavioral rhythms. Using two independent genetic screens, mutants of Transport and Golgi organization 10 (Tango10) were identified with poor behavioral rhythmicity. Tango10 expression in pacemaker neurons expressing the neuropeptide PIGMENT-DISPERSING FACTOR (PDF) is required for robust rhythms. Loss of Tango10 results in elevated PDF accumulation in nerve terminals even in mutants lacking a functional core clock. TANGO10 protein itself is rhythmically expressed in PDF terminals. Mass spectrometry of TANGO10 complexes reveals interactions with the E3 ubiquitin ligase CULLIN 3 (CUL3). CUL3 depletion phenocopies Tango10 mutant effects on PDF even in the absence of the core clock gene timeless. Patch clamp electrophysiology in Tango10 mutant neurons demonstrates elevated spontaneous firing potentially due to reduced voltage-gated Shaker-like potassium currents. It is proposed that Tango10/Cul3 transduces molecular oscillations from the core clock to neuropeptide release important for behavioral rhythms (Lee, 2021).
Disrupted circadian rhythms is a prominent feature of multiple neurodegenerative diseases. Yet mechanisms linking Tau (see Drosophila Tau) to rhythmic behavior remain unclear. This study found that expression of a phosphomimetic human Tau mutant (TauE14) in Drosophila circadian pacemaker neurons disrupts free-running rhythmicity. While cell number and oscillations of the core clock protein PERIOD are unaffected in the small LNv (sLNv) neurons important for free running rhythms, a near complete loss of the major LNv neuropeptide pigment dispersing factor (PDF) in the dorsal axonal projections of the sLNvs. This was accompanied by a ~ 50% reduction in the area of the dorsal terminals and a modest decrease in cell body PDF levels. Expression of wild-type Tau also reduced axonal PDF levels but to a lesser extent than TauE14. TauE14 also induces a complete loss of mitochondria from these sLNv projections. However, mitochondria were increased in sLNv cell bodies in TauE14 flies. These results suggest that TauE14 disrupts axonal transport of neuropeptides and mitochondria in circadian pacemaker neurons, providing a mechanism by which Tau can disrupt circadian behavior prior to cell loss (Zhang, 2021).
The mechanisms that generate robust ionic oscillation in circadian pacemaker neurons are under investigation. This study demonstrates critical functions of the mitochondrial cation antiporter leucine zipper-EF-hand-containing transmembrane protein 1 (LETM1), which exchanges K(+)/H(+) in Drosophila and Ca(2+)/H(+) in mammals, in circadian pacemaker neurons. Letm1 knockdown in Drosophila pacemaker neurons reduced circadian cytosolic H(+) rhythms and prolonged nuclear PERIOD/TIMELESS expression rhythms and locomotor activity rhythms. In rat pacemaker neurons in the hypothalamic suprachiasmatic nucleus (SCN), circadian rhythms in cytosolic Ca(2+) and Bmal1 transcription were dampened by Letm1 knockdown. Mitochondrial Ca(2+) uptake peaks late during the day were also observed in rat SCN neurons following photolytic elevation of cytosolic Ca(2+). Since cation transport by LETM1 is coupled to mitochondrial energy synthesis, it is proposed that LETM1 integrates metabolic, ionic, and molecular clock rhythms in the central clock system in both invertebrates and vertebrates (Morioka, 2022).
Statins, HMG Coenzyme A Reductase (HMGCR) inhibitors, are a first-line therapy, used to reduce hypercholesterolemia and the risk for cardiovascular events. While sleep disturbances are recognized as a side-effect of statin treatment, the impact of statins on sleep is under debate. Using Drosophila, this study discovered a novel role for Hmgcr in sleep modulation. Loss of pan-neuronal Hmgcr expression affects fly sleep behavior, causing a decrease in sleep latency and an increase in sleep episode duration. The pars intercerebralis (PI), equivalent to the mammalian hypothalamus, was identified as the region within the fly brain requiring Hmgcr activity for proper sleep maintenance. Lack of Hmgcr expression in the PI insulin-producing cells recapitulates the sleep effects of pan-neuronal Hmgcr knockdown. Conversely, loss of Hmgcr in a different PI subpopulation, the corticotropin releasing factor (CRF) homologue-expressing neurons (DH44 neurons), increases sleep latency and decreases sleep duration. Interestingly, loss of Hmgcr in the PI does not affect circadian rhythm, suggesting that Hmgcr regulates sleep by pathways distinct from the circadian clock. Taken together, these findings suggest that Hmgcr activity in the PI is essential for proper sleep homeostasis in flies (Alsehli, 2022).
The regulation of sleep and metabolism are highly interconnected, and dysregulation of sleep is linked to metabolic diseases that include obesity, diabetes, and heart disease. Furthermore, both acute and long-term changes in diet potently impact sleep duration and quality. To identify novel factors that modulate interactions between sleep and metabolic state, a genetic screen was performed for their roles in regulating sleep duration, starvation resistance, and starvation-dependent modulation of sleep. This screen identified a number of genes with potential roles in regulating sleep, metabolism, or both processes. One such gene encodes the auxiliary ion channel UNC79, which was implicated in both the regulation of sleep and starvation resistance. Genetic knockdown or mutation of unc79 results in flies with increased sleep duration, as well as increased starvation resistance. Previous findings have shown that unc79 is required in pacemaker for 24-hours circadian rhythms. This study found that unc79 functions in the mushroom body, but not pacemaker neurons, to regulate sleep duration and starvation resistance. Together, these findings reveal spatially localized separable functions of unc79 in the regulation of circadian behavior, sleep, and metabolic function (Murakami, 2021).
Cation chloride cotransporters (CCCs) regulate intracellular chloride ion concentration ([Cl(-)](i)) within neurons, which can reverse the direction of the neuronal response to the neurotransmitter GABA. Na(+) K(+) Cl(-) (NKCC) and K(+) Cl(-) (KCC) cotransporters transport Cl(-) into or out of the cell, respectively. When NKCC activity dominates, the resulting high [Cl(-)](i) can lead to an excitatory and depolarizing response of the neuron upon GABA(A) receptor opening, while KCC dominance has the opposite effect. This inhibitory-to-excitatory GABA switch has been linked to seasonal adaption of circadian clock function to changing day length, and its dysregulation is associated with neurodevelopmental disorders such as epilepsy. In Drosophila melanogaster, constant light normally disrupts circadian clock function and leads to arrhythmic behavior. This study demonstrates a function for CCCs in regulating Drosophila locomotor activity and GABA responses in circadian clock neurons because alteration of CCC expression in circadian clock neurons elicits rhythmic behavior in constant light. The same effects were observed after downregulation of the Wnk and Fray kinases, which modulate CCC activity in a [Cl(-)](i)-dependent manner. Patch-clamp recordings from the large LNv clock neurons show that downregulation of KCC results in a more positive GABA reversal potential, while KCC overexpression has the opposite effect. Finally, KCC and NKCC downregulation reduces or increases morning behavioral activity during long photoperiods, respectively. In summary, these results support a model in which the regulation of [Cl(-)](i) by a KCC/NKCC/Wnk/Fray feedback loop determines the response of clock neurons to GABA, which is important for adjusting behavioral activity to constant light and long-day conditions (Eick, 2022).
Sleep disruptions are quite common in psychological disorders, but the underlying mechanism remains obscure. Wolfram syndrome 1 (WS1) is an autosomal recessive disease mainly characterized by diabetes insipidus/mellitus, neurodegeneration and psychological disorders. It is caused by loss-of function mutations of the WOLFRAM SYNDROME 1 (WFS1) gene, which encodes an endoplasmic reticulum (ER)-resident transmembrane protein. Heterozygous mutation carriers do not develop WS1 but exhibit 26-fold higher risk of having psychological disorders. Since WS1 patients display sleep abnormalities, this study aimed to explore the role of WFS1 in sleep regulation so as to help elucidate the cause of sleep disruptions in psychological disorders. It was found in Drosophila that knocking down wfs1 in all neurons and wfs1 mutation lead to reduced sleep and dampened circadian rhythm. These phenotypes are mainly caused by lack of wfs1 in dopamine 2-like receptor (Dop2R) neurons which act to promote wake. Consistently, the influence of wfs1 on sleep is blocked or partially rescued by inhibiting or knocking down the rate-limiting enzyme of dopamine synthesis, suggesting that wfs1 modulates sleep via dopaminergic signaling. Knocking down wfs1 alters the excitability of Dop2R neurons, while genetic interactions reveal that lack of wfs1 reduces sleep via perturbation of ER-mediated calcium homeostasis. Taken together, a role is proposed for wfs1 in modulating the activities of Dop2R neurons by impinging on intracellular calcium homeostasis, and this in turn influences sleep. These findings provide a potential mechanistic insight for pathogenesis of diseases associated with WFS1 mutations (Hao, 2023).
Sleep disruptions are common in individuals with psychiatric disorders, and sleep disturbances are risk factors for future onset of depression. However, the mechanism underlying sleep disruptions in psychiatric disorders are largely unclear. Wolfram Syndrome 1 (WS1) is an autosomal recessive neurodegenerative disease characterized by diabetes insipidus, diabetes mellitus, optic atrophy, deafness and psychiatric abnormalities such as severe depression, psychosis and aggression. It is caused by homozygous (and compound heterozygous) mutation of the WOLFRAM SYNDROME 1 (WFS1) gene, which encodes wolframin, an endoplasmic reticulum (ER) resident protein highly expressed in the heart, brain, and pancreas. On the other hand, heterozygous mutation of WFS1 does not lead to WS1 but increase the risk of depression by 26 fold. A study in mice further confirmed that WFS1 mutation is causative for depression. Consistent with the comorbidity of psychiatric conditions and sleep abnormalities, WS1 patients also experience increased sleep problems compared to individuals with type I diabetes and healthy controls. It has been proposed that sleep symptoms can be used as a biomarker of the disease, especially during relatively early stages, but the mechanisms underlying these sleep disturbances are unclear. Considering that heterozygous WFS1 mutation is present in up to 1% of the population and may be a significant cause of psychiatric disorder in the general population, it was decided to investigate the role of wolframin in sleep regulation so as to probe the mechanism underlying sleep disruptions in psychiatric disorders (Hao, 2023).
Although the wolframin protein does not possess distinct functional domains, a number of ex vivo studies in cultured cells demonstrated a role for it in regulating cellular responses to ER stress and calcium homeostasis, as well as ER-mitochondria cross-talk. Mice that lack Wfs1 in pancreatic β cells develop glucose intolerance and insulin deficiency due to enhanced ER stress and apoptosis. Knocking out Wfs1 in layer 2/3 pyramidal neurons of the medial prefrontal cortex in mice results in increased depression-like behaviors in response to acute restraint stress. This is accompanied by hyperactivation of the hypothalamic-pituitary-adrenal axis and altered accumulation of growth and neurotrophic factors, possibly due to defective ER function. A more recent study in Drosophila found that knocking down wfs1 in the nervous system does not increase ER stress, but enhances the susceptibility to oxidative stress-, endotoxicity- and tauopathy-induced behavioral deficits and neurodegeneration (Sakakibara, 2018). Overall, the physiological function of wolframin in vivo, especially in the brain, remains elusive for the most part. This study identified a role for wolframin in regulating sleep and circadian rhythm in flies. Wfs1 deficiency in the dopamine 2-like receptor (Dop2R) neurons leads to reduced sleep, while inhibiting dopamine synthesis blocks the effect of wfs1 on sleep, implying that wfs1 influences sleep via dopaminergic signaling. It was further found that these Dop2R neurons function to promote wakefulness. Depletion of wfs1 alters neural activity, which leads to increased wakefulness and reduced sleep. Consistent with this, it was found that knocking down the ER calcium channel Ryanodine receptor (RyR) or 1,4,5-trisphosphate receptor (Itpr) rescues while knocking down the sarco(endoplasmic)reticulum ATPase SERCA synergistically enhances the short-sleep phenotype caused by wfs1 deficiency, indicating that wolframin regulates sleep by modulating calcium homeostasis. Taken together, these findings provide a potential mechanism for the sleep disruptions associated with WFS1 mutation, and deepen understanding of the functional role of wolframin in the brain (Hao, 2023).
Sleep problems have been reported in WS1 patients. Their scores on Pediatric Sleep Questionnaire are more than 3 times higher than healthy controls and doubled compared to individuals with type I diabetes, indicating that the sleep issues are not merely due to metabolic disruptions. Indeed, this study suggests that the sleep problems in human patients are of neural origin, specifically in the wake-promoting Dop2R neurons. Given that the rebound sleep is not significantly altered in wfs1 depleted flies, it is believed that lack of wfs1 does not shorten sleep duration by impairing the sleep homeostasis system. Instead, wfs1 deficiency leads to excessive wakefulness which in turn results in decreased sleep. Considering that heterozygous WFS1 mutation is present in up to 1% of the population, it would be interesting to examine whether these heterozygous mutations contribute to sleep disruptions in the general population (Hao, 2023).
In mouse, chick, quail and turtle, Wfs1 has been shown to be expressed in brain regions where dopamine receptor Drd1 is expressed. D1-like dopamine receptor binding is increased while striatal dopamine release is decreased in Wfs1-/- mice. The current results also implicate a role for wolframin in dopamine receptor neurons and that lack of wfs1 impacts dopaminergic signaling, as the effects of wfs1 deficiency on both sleep and mushroom body (MB) calcium concentration is blocked by the tyrosine hydroxylase inhibitor AMPT. Both Dop2RGAL4 and GoαGAL4 exhibit prominent expression in the MB, and to be more specific, in the α and β lobes of MB. Previous studies have shown that dopaminergic neurons innervate wake promoting MB neurons, and this study found Dop2R and Goα+ cells to be wake-promoting as well. Therefore, it is suspected that wolframin functions in MB Dop2R/Goα+ neurons to influence sleep. Taken together, these findings suggest an evolutionarily conserved role of wolframin within the dopaminergic system. As this system is also important for sleep/wake regulation in mammals, it is reasonable to suspect that wolframin functions in mammals to modulate sleep by influencing the dopaminergic tone as well (Hao, 2023).
MB neural activity appears to be enhanced in wfs1 deficient flies based on the results obtained using CaLexA and spH reporters. This elevated activity is consistent with behavioral data, as activation of Dop2R/Goα+ cells reduces sleep, similar to the effects of wfs1 deficiency. Moreover, silencing Dop2R neurons rescues the short-sleep phenotype of wfs1 mutants, while over-expressing wfs1 restores the decreased sleep induced by activation of Dop2R neurons. These findings suggest that wolframin functions to suppress the excitability of MB Dop2R neurons, which in turn reduces wakefulness and promotes sleep. Comparable cellular changes have been observed in SERCA mutant flies. Electric stimulation leads to an initial increase followed by prolonged decrease of calcium concentration in mutant motor nerve terminal compared to the control, while action potential firing is increased in the mutants. This series of results underpin the importance of ER calcium homeostasis in determining membrane excitability and thus neural function (Hao, 2023).
GCaMP6 monitoring reveals that wfs1 deficiency selectively reduces fluorescence signal in the MB both under baseline condition and after dopamine treatment, which should reflect a reduction of cytosolic calcium level that is usually associated with decreased excitability. Previous studies have shown that lack of wolframin leads to increased basal calcium level in neural progenitor cells derived from induced pluripotent stem (iPS) cells of WS1 patients and primary rat cortical neurons, but after stimulation the rise of calcium concentration is smaller in Wfs1 deficient neurons, resulting in reduced calcium level compared to controls. Similarly, evoked calcium increase is also diminished in fibroblasts of WS1 patients and MIN6 insulinoma cells with WFS1 knocked down. Notably, wolframin has been shown to bind to calmodulin (CaM) in rat brain, and is capable of binding with calcium/CaM complex in vitro and in transfected cells. This may undermine the validity of using GCaMP to monitor calcium level in wfs1 deficient animals and cells, and could potentially account for the contradictory data acquired using CaLexA vs GCaMP (Hao, 2023).
It is intriguing that in this study wfs1 deficiency appears to selectively impair the function of Dop2R/Goα+ neurons. It has been shown that in the rodent brain Wfs1 is enriched in layer II/III of the cerebral cortex, CA1 field of the hippocampus, central extended amygdala, striatum, and various sensory and motor nuclei in the brainstem. Wfs1 expression starts to appear during late embryonic development in dorsal striatum and amygdala, and the expression quickly expands to other regions of the brain at birth. It is suspect that in flies wfs1 may be enriched in Dop2R/Goα+ cells during a critical developmental period, and that sufficient level of wolframin is required for their maturation and normal functioning in adults. Another possibility is that these cells are particularly susceptible to calcium dyshomeostasis induced by loss of wfs1. Indeed, this is believed to be an important cause of selective dopaminergic neuron loss in Parkinson's Disease, as dopaminergic neurons are unique in their autonomic excitability and selective dependence on calcium channel rather than sodium channel for action potential generation. It is reasoned that Dop2R/Goα+ neurons may also be more sensitive to abnormal intracellular calcium concentration, making them particularly vulnerable to wfs1 deficiency. The pathogenic mechanism underlying the neurodegeneration of WS1 is quite complex, possibly involving brain-wide neurodegenerative processes and neurodevelopmental dis-regulations. The findings of this study provide some evidence supporting a role for altered dopaminergic system during development. Obviously, much more needs to be done to test these hypotheses (Hao, 2023).
The precise role of wolframin in ER calcium handling is not yet clear. It has been shown in human embryonic kidney (HEK) 293 cells that knocking down WFS1 reduces while over-expressing WFS1 increases ER calcium level. The authors concluded that wolframin upregulates ER calcium concentration by increasing the rate of calcium uptake. Consistently, this study found by genetic interaction that knocking down RyR or Itpr (which act to reduce ER calcium level and thus knocking down either one will increase ER calcium level) rescues the short-sleep phenotype caused by wfs1 mutation, while knocking down SERCA (which acts to increase ER calcium level and thus knocking down this gene will reduce ER calcium level) synergistically enhances the short-sleep phenotype. Based on the results of these genetic interactions, it is proposed that lack of wfs1 increases cytosolic calcium while decreasing ER calcium, leading to hyperexcitability of Dop2R neurons and thus reduced sleep. Knocking down RyR or Itpr decreases cytosolic calcium and increases ER calcium, counteracting the influences of wfs1 deficiency and thus rescuing the short-sleep phenotype. On the other hand, knocking down SERCA further increases cytosolic calcium and decreases ER calcium, rendering an enhancement of the short-sleep phenotype. In line with this, study conducted in neural progenitor cells derived from iPS cells of WS1 patients demonstrated that pharmacological inhibition of RyR can prevent cell death caused by WFS1 mutation. In addition, inhibiting the function of IP3R may mitigate ER stress in wolframin deficient cells. One caveat is that SERCA protein level is increased in primary islets isolated from Wfs1 conditional knock-out mice, as well as in MIN6 cells and neuroblastoma cell line with WFS1 knocked down. It is reasoned that this may be a compensatory increase to make up for the reduced ER calcium level due to wolframin deficiency. It is acknowledged that the hypothesis proposed in in the papert is not supported by GCaMP data, which indicates decreased cytosolic calcium level in Dop2R neurons of wfs1 deficient flies. It is suspected that since the sleep phenotype associated with lack of wfs1 is of developmental origin, it is possible there is an initial increase of cytosolic calcium during critical developmental period in wfs1 deficient flies and this influences the function of Dop2R neurons in adults. Clearly, further characterizations need to be done to fully elucidate this issue, and preferably another calcium indicator independent of the GCaMP system should be employed (Hao, 2023).
In conclusion, this study identified a role for wolframin in the wake-promoting Dop2R neurons. wfs1 depletion in these cells lead to impaired calcium homeostasis and altered neural activity, which in turn leads to enhanced wakefulness and reduced sleep. This study may provide some insights for the mechanisms underlying the sleep disruptions in individuals with WFS1 mutation, as well as for the pathogenesis of WS1 (Hao, 2023).
Homeostatic and circadian processes collaborate to appropriately time and consolidate sleep and wake. To understand how these processes are integrated, brief sleep deprivation was scheduled at different times of day in Drosophila, and elevated morning rebound was compared to evening. These effects depend on discrete morning and evening clock neurons, independent of their roles in circadian locomotor activity. In the R5 ellipsoid body sleep homeostat, this study identified elevated morning expression of activity dependent and presynaptic gene expression as well as the presynaptic protein BRUCHPILOT consistent with regulation by clock circuits. These neurons also display elevated calcium levels in response to sleep loss in the morning, but not the evening consistent with the observed time-dependent sleep rebound. These studies reveal the circuit and molecular mechanisms by which discrete circadian clock neurons program a homeostatic sleep center (Andreani, 2022).
This study describes the neural circuit and molecular mechanisms by which discrete populations of the circadian clock network program the R5 sleep homeostat to control the homeostatic response to sleep loss. A novel protocol was developed to administer brief duration sleep deprivation (SD) and robustly measure homeostatic rebound sleep. Using this strategy, it was demonstrated that homeostatic rebound is significantly higher in the morning than in the evening. Distinct subsets of the circadian clock network and their downstream neural targets were identified that mediate the enhancement and suppression of morning and evening rebound respectively. Using unbiased transcriptomics, very little gene expression significantly altered was observed in response to 2.5 hr sleep deprivation. On the other hand, this study did identify elevated expression of activity-dependent and presynaptic genes in the morning independent of sleep deprivation. Consistent with this finding, elevated levels of the presynaptic protein BRP were observed, that is absent in the absence of Clk. These baseline changes are accompanied by an elevated calcium response to sleep deprivation in the morning mirroring the enhanced behavioral rebound in the morning. Taken together, these data support the model of a circadian regulated homeostat that turns the homeostat up late at night to sustain sleep and down late in the day to sustain wake (Andreani, 2022).
These studies suggest that homeostatic drive in the R5 neurons is stored post-transcriptionally. As part of these studies, a novel protocol using minimal amounts of SD was developed; this protocol could be useful for minimizing mechanical stress effects and isolating underlying molecular processes crucial for sleep homeostasis. Six to 24 hr of SD in Drosophila is commonly used despite the potential stressful or even lethal effects. This study demonstrated that shorter 2.5 hr deprivations not only induce a robust rebound sleep response, but also the percent of sleep lost recovered at ZT0 is close to 100% versus 14-35% seen in 12 hr SD protocols. Using this shorter SD, this study now found that many effects observed in R5 neurons with 12 hr SD (e.g. increased BRP and upregulation of nmdar subunits) are no longer observed with shorter SD, even though the necessity of R5 neurons for rebound is retained after 2.5 hr SD. Previously, translating ribosome affinity purification (TRAP) was used to show upregulation of nmdar subunits following 12 hr SD. FACS and TRAP are distinct methodologies for targeted collection of RNA for sequencing and can yield unique gene lists. One possibility is that upregulation of nmdar subunits is occurring locally in neuronal processes, which are often lost during FACS, and/or is at the level of translation initiation or elongation. Nonetheless, in agreement with previous work, this study observed SD-induced increases in calcium correlated with behavioral rebound in the morning, suggesting that this process is a core feature of the cellular homeostatic response (Andreani, 2022).
Using genetically targeted 'loss-of-function' manipulations, this study has defined small subsets of circadian clock neurons and downstream circuits that are necessary for intact clock modulation of sleep homeostasis. The use of intersectional approaches enabled highly resolved targeting not possible with traditional lesioning experiments in the SCN. Collectively these studies defined a potential Glu+ DN1p-TuBu-R4m circuit important for enhancing morning rebound as well as a discrete group of LNds important for suppressing evening rebound. Importantly, most of these effects on sleep rebound are evident in the absence of substantial changes in baseline activity, despite other studies indicating their necessity for normal circadian behavior. Of note, the proposed roles of the DN1p and LNd clock neurons are sleep and wake promotion consistent with the findings after sleep deprivation. It is hypothesized that by using chronic silencing methods, baseline effects may not be evident due to compensatory changes but that these effects are only revealed when the system is challenged by sleep deprivation. Similar genetic strategies in mammals may be useful in uncovering which SCN neurons are driving circadian regulation of sleep homeostasis given the comparable suppression of sleep rebound in the evening in humans. Nonetheless, the finding of sleep homeostasis phenotypes in the absence of significant baseline effects suggests that a major role of these clock neuron subsets may be to manage homeostatic responses (Andreani, 2022).
These studies suggest that circadian and homeostatic processes do not compete for influence on a downstream neural target but that the circadian clock programs the homeostat itself. Using an unbiased transcriptomic approach, this study discovered time-dependent expression of activity dependent and presynaptic genes, consistent with previous data that the R5 neurons exhibit time-dependent activity. This study observed significant upregulation of several genes involved in synaptic transmission (Syx1a, Rim, nSyb, unc-104, Srpk79D, para, CG5890) evincing a permissive active state for R5 neurons in the morning. This is accompanied by elevated levels of the key presynaptic protein BRP in the morning compared to evening. It is notable that elevated BRP in the morning is the opposite of what would be expected based on a sleep-dependent reduction in BRP proposed by the synaptic homeostasis hypothesis, suggesting a sleep-wake independent mechanism. Previous studies have shown that modulation of BRP levels in the R5 are important for its sleep function, suggesting that changes in BRP levels impact R5 function. It is hypothesized that these baseline transcriptomic changes underlie the differential R5 sensitivity to sleep deprivation is evident as calcium increases in the morning and not the evening. Indeed, trancriptomic and proteomic studies of the mouse forebrain across time and after sleep deprivation are consistent with the model that the circadian clock programs the transcriptome while homeostatic process function post-trranscriptionally, paralleling what was found for R5. It will be of great interest to understand the circuit and molecular mechanisms by which circadian clocks regulate the R5 neuronal calcium and synaptic properties and whether similar circuit architectures underlie daily mammalian sleep-wake (Andreani, 2022).
Circadian behavioral rhythms in Drosophila melanogaster are regulated by about 75 pairs of brain neurons. They all express the core clock genes but have distinct functions and gene expression profiles. To understand the importance of these distinct molecular programs, neuron-specific gene manipulations are essential. Although RNAi based methods are standard to manipulate gene expression in a cell-specific manner, they are often ineffective, especially in assays involving smaller numbers of neurons or weaker Gal4 drivers. A neuron-specific CRISPR-based method has been exploited to mutagenize genes within circadian neurons. This study further explores this approach to mutagenize three well-studied clock genes: the transcription factor gene vrille, the photoreceptor gene Cryptochrome (cry), and the neuropeptide gene Pdf (pigment dispersing factor). The CRISPR-based strategy not only reproduced their known phenotypes but also assigned cry function for different light-mediated phenotypes to discrete, different subsets of clock neurons. Two recently published methods for temporal regulation in adult neurons were tested, inducible Cas9 and the auxin-inducible gene expression system. The results were not identical, but both approaches successfully showed that the adult-specific knockout of the neuropeptide Pdf reproduces the canonical loss-of-function mutant phenotypes. In summary, a CRISPR-based strategy is a highly effective, reliable, and general method to temporally manipulate gene function in specific adult neurons (Richhariya, 2023).
Pigment Dispersing Factor (PDF), is a key signaling molecule coordinating the neuronal network associated with the circadian rhythms in Drosophila. The precursor (proPDF) of the mature PDF (mPDF) consists of two motifs, a larger PDF-associated peptide (PAP) and PDF. Through cleavage and amidation, the proPDF is predicted to produce cleaved-PAP (cPAP) and mPDF. To delve into the in vivo mechanisms underlying proPDF maturation, various mutations were generated that eliminate putative processing sites, and the effect of each mutation on the production of cPAP and mPDF was then analyzed by four different antibodies in both ectopic and endogenous conditions. The knockdown effects of processing enzymes on the proPDF maturation was also studied. At the functional level, circadian phenotypes were measured for all mutants and knockdown lines. As results, the roles of key enzymes and their target residues was confirmed: Amontillado (Amon) for the cleavage at the consensus dibasic KR site, Silver (Svr) for the removal of C-terminal basic residues from the intermediates, PAP-KR and PDF-GK, derived from proPDF, and PHM (Peptidylglycine-α-hydroxylating monooxygenase) for the amidation of PDF. The results suggest that the C-terminal amidation occurs independently of proPDF cleavage. Moreover, PAP domain is important for the proPDF trafficking into the secretory vesicles and a close association between cPAP and mPDF following cleavage seems required for their stability within the vesicles. These studies highlight the biological significance of individual processing steps and the roles of the PAP for the stability and function of mPDF which is essential for the circadian clockworks (Lee, 2023).
Circadian rhythm maintains the sleep-wake cycle in biological systems. Various biological activities are regulated and modulated by the circadian rhythm, disruption of which can result in onset of diseases. Robust rhythms of phosphorylation profiles and abundances of PERIOD (PER) proteins are thought to be the master keys that drive circadian clock functions. The role of casein kinase 2 (CK2) in circadian rhythm via its direct interactions with the PER protein has been extensively studied; however, the exact mechanism by which it affects circadian rhythms at the molecular level is not known. This study proposes an extended circadian rhythm model in Drosophila that incorporates the crosstalk between the PER protein and CK2. The regulatory role of CK2 was studied in the dynamics of PER proteins involved in circadian rhythm using the stochastic simulation algorithm. It was observed that variations in the concentration of CK2 in the circadian rhythm model modulates the PER protein dynamics at different cellular states, namely, active, weakly active, and rhythmic death. These oscillatory states may correspond to distinct pathological cellular states of the living system. Molecular noise was found at the expression level of CK2 to switch normal circadian rhythm to any of the three above-mentioned circadian oscillatory states. The results suggest that the concentration levels of CK2 in the system has a strong impact on its dynamics, which is reflected in the time evolution of PER protein. It is believed that these findings can contribute towards understanding the molecular mechanisms of circadian dysregulation in pathways driven by the PER mutant genes and their pathological states, including cancer, obesity, diabetes, neurodegenerative disorders, and socio-psychological disease (Malik, 2023).
Recent single-cell sequencing of most adult Drosophila circadian neurons indicated notable and unexpected heterogeneity. To address whether other populations are similar, a large subset of adult brain dopaminergic neurons was sequenced. Their gene expression heterogeneity is similar to that of clock neurons, i.e., both populations have two to three cells per neuron group. There was also unexpected cell-specific expression of neuron communication molecule messenger RNAs: G protein-coupled receptor or cell surface molecule (CSM) transcripts alone can define adult brain dopaminergic and circadian neuron cell type. Moreover, the adult expression of the CSM DIP-beta in a small group of clock neurons is important for sleep. It is suggested that the common features of circadian and dopaminergic neurons are general, essential for neuronal identity and connectivity of the adult brain, and that these features underlie the complex behavioral repertoire of Drosophila (Ma, 2023).
PERIOD (PER) and Casein Kinase 1&delta regulate circadian rhythms through a phosphoswitch that controls PER stability and repressive activity in the molecular clock. CK1Δ phosphorylation of the familial advanced sleep phase (FASP) serine cluster embedded within the Casein Kinase 1 binding domain (CK1BD) of mammalian PER1/2 inhibits its activity on phosphodegrons to stabilize PER and extend circadian period. This study shows that the phosphorylated FASP region (pFASP) of PER2 directly interacts with and inhibits CK1δ. Co-crystal structures in conjunction with molecular dynamics simulations reveal how pFASP phosphoserines dock into conserved anion binding sites near the active site of CK1δ. Limiting phosphorylation of the FASP serine cluster reduces product inhibition, decreasing PER2 stability and shortening circadian period in human cells. This study found that Drosophila PER also regulates CK1δ via feedback inhibition through the phosphorylated PER-Short domain, revealing a conserved mechanism by which PER phosphorylation near the CK1BD regulates CK1 kinase activity (Philpott, 2023).
Sleep is essential, but animals may forgo sleep to engage in other critical behaviors, such as feeding and reproduction. Previous studies have shown that female flies exhibit decreased sleep after mating, but understanding of the process is limited. This study reports that postmating nighttime sleep loss is modulated by diet and sleep deprivation, demonstrating a complex interaction among sleep, reproduction, and diet. It was also found that female-specific pC1 neurons and sleep-promoting dorsal fan-shaped body (dFB) neurons are required for postmating sleep plasticity. Activating pC1 neurons leads to sleep suppression on standard fly culture media but has little sleep effect on sucrose-only food. Published connectome data suggest indirect, inhibitory connections among pC1 subtypes. Using calcium imaging, it was shown that activating the pC1e subtype inhibits dFB neurons. It is proposed that pC1 and dFB neurons integrate the mating status, food context, and sleep drive to modulate postmating sleep plasticity (Duhart, 2023).
Sleep disturbances are common in neurodevelopmental disorders, but knowledge of molecular factors that govern sleep in young animals is lacking. Evidence across species, including Drosophila, suggests that juvenile sleep has distinct functions and regulatory mechanisms in comparison to sleep in maturity. In flies, manipulation of most known adult sleep regulatory genes is not associated with sleep phenotypes during early developmental (larval) stages. This study examined the role of the neurodevelopmental disorder-associated gene Neurofibromin 1 (Nf1) in sleep during numerous developmental periods. Mutations in Neurofibromin 1 (Nf1) are associated with sleep and circadian disorders in humans and adult flies. It was found in flies that Nf1 acts to regulate sleep across the lifespan, beginning during larval stages. Nf1 is required in neurons for this function, as is signaling via the Alk pathway. These findings identify Nf1 as one of a small number of genes positioned to regulate sleep across developmental periods (Durkin, 2023).
Sleep behavior is conserved throughout evolution, and sleep disturbances are a frequent comorbidity of neuropsychiatric disorders. However, the molecular basis underlying sleep dysfunctions in neurological diseases remains elusive. Using a model for neurodevelopmental disorders (NDDs), the Drosophila Cytoplasmic FMR1 interacting protein haploinsufficiency (Cyfip(85.1/+)), this study identified a mechanism modulating sleep homeostasis. Increased activity of the sterol regulatory element-binding protein (SREBP) in Cyfip(85.1/+) flies induces an increase in the transcription of wakefulness-associated genes, such as the malic enzyme (Men), causing a disturbance in the daily NADP(+)/NADPH ratio oscillations and reducing sleep pressure at the night-time onset. Reduction in SREBP or Men activity in Cyfip(85.1/+) flies enhances the NADP(+)/NADPH ratio and rescues the sleep deficits, indicating that SREBP and Men are causative for the sleep deficits in Cyfip heterozygous flies. This work suggests modulation of the SREBP metabolic axis as a new avenue worth exploring for its therapeutic potential in sleep disorders (Mariano, 2023).
A single adenosine receptor gene (dAdoR) has been detected in Drosophila melanogaster. However, its function in different cell types of the nervous system is mostly unknown. Therefore, this study overexpressed or silenced the dAdoR gene in eye photoreceptors, all neurons, or glial cells and examined the fitness of flies, the amount and daily pattern of sleep, and the influence of dAdoR silencing on Bruchpilot (BRP) presynaptic protein. Furthermore, the dAdoR and brp gene expression was examined in young and old flies. A higher level of dAdoR was found in the retina photoreceptors, all neurons, and glial cells negatively influenced the survival rate and lifespan of male and female Drosophila in a cell-dependent manner and to a different extent depending on the age of the flies. In old flies, expression of both dAdoR and brp was higher than in young ones. An excess of dAdoR in neurons improved climbing in older individuals. It also influenced sleep by lengthening nighttime sleep and siesta. In turn, silencing of dAdoR decreased the lifespan of flies, although it increased the survival rate of young flies. It hindered the climbing of older males and females, but did not change sleep. Silencing also affected the daily pattern of BRP abundance, especially when dAdoR expression was decreased in glial cells. The obtained results indicate the role of adenosine and dAdoR in the regulation of fitness in flies that is based on communication between neurons and glial cells, and the effect of glial cells on synapses (Bhattacharya, 2023).
While neurotransmitter identity was once considered singular and immutable for mature neurons, it is now appreciated that one neuron can release multiple neuroactive substances (cotransmission) whose identities can even change over time. To explore the mechanisms that tune the suite of transmitters a neuron releases, this study developed transcriptional and translational reporters for cholinergic, glutamatergic, and GABAergic signaling in Drosophila. Many glutamatergic and GABAergic cells also transcribe cholinergic genes, but fail to accumulate cholinergic effector proteins. Suppression of cholinergic signaling involves posttranscriptional regulation of cholinergic transcripts by the microRNA miR-190; chronic loss of miR-190 function allows expression of cholinergic machinery, reducing and fragmenting sleep. Using a "translation-trap" strategy, this study shows that neurons in these populations have episodes of transient translation of cholinergic proteins, demonstrating that suppression of cotransmission is actively modulated. Posttranscriptional restriction of fast transmitter cotransmission provides a mechanism allowing reversible tuning of neuronal output (Chen, 2023).
The neuropeptide pigment-dispersing factor (PDF) plays a pivotal role in the circadian clock of most Ecdysozoa and is additionally involved in the timing of seasonal responses of several photoperiodic species. The pea aphid, Acyrthosiphon pisum, is a paradigmatic photoperiodic species with an annual life cycle tightly coupled to the seasonal changes in day length. Nevertheless, PDF could not be identified in A. pisum so far. The present identified a PDF-coding gene that has undergone significant changes in the otherwise highly conserved insect C-terminal amino acid sequence. A newly generated aphid-specific PDF antibody stained four neurons in each hemisphere of the aphid brain that co-express the clock protein Period and have projections to the pars lateralis that are highly plastic and change their appearance in a daily and seasonal manner, resembling those of the fruit fly PDF neurons. Most intriguingly, the PDF terminals overlap with dendrites of the insulin-like peptide (ILP) positive neurosecretory cells in the pars intercerebralis and with putative terminals of Cryptochrome (CRY) positive clock neurons. Since ILP has been previously shown to be crucial for seasonal adaptations and CRY might serve as a circadian photoreceptor vital for measuring day length, these results suggest that PDF plays a critical role in aphid seasonal timing (Colizzi, 2023).
Sleep is a complex and plastic behavior regulated by multiple brain regions and influenced by numerous internal and external stimuli. Thus, to fully uncover the function(s) of sleep, cellular resolution of sleep-regulating neurons needs to be achieved. Doing so will help to unequivocally assign a role or function to a given neuron or group of neurons in sleep behavior. In the Drosophila brain, neurons projecting to the dorsal fan-shaped body (dFB) have emerged as a key sleep-regulating area. To dissect the contribution of individual dFB neurons to sleep, this study undertook an intersectional Split-GAL4 genetic screen focusing on cells contained within the 23E10-GAL4 driver, the most widely used tool to manipulate dFB neurons. This study demonstrated that 23E10-GAL4 expresses in neurons outside the dFB and in the fly equivalent of the spinal cord, the ventral nerve cord (VNC). Furthermore, it was shown that 2 VNC cholinergic neurons strongly contribute to the sleep-promoting capacity of the 23E10-GAL4 driver under baseline conditions. However, in contrast to other 23E10-GAL4 neurons, silencing these VNC cells does not block sleep homeostasis. Thus, these data demonstrate that the 23E10-GAL4 driver contains at least 2 different types of sleep-regulating neurons controlling distinct aspects of sleep behavior (Jones, 2023).
Animals possess a circadian central clock in the brain, where circadian behavioural rhythms are generated. In the fruit fly (Drosophila melanogaster), the central clock comprises a network of approximately 150 clock neurons, which is important for the maintenance of a coherent and robust rhythm. Several neuropeptides involved in the network have been identified, including Pigment-dispersing factor (PDF) and CCHamide1 (CCHa1) neuropeptides. PDF signals bidirectionally to CCHa1-positive clock neurons; thus, the clock neuron groups expressing PDF and CCHa1 interact reciprocally. However, the role of these interactions in molecular and behavioural rhythms remains elusive. This study,generated Pdf01 and CCHa1SK8) double mutants and examined their locomotor activity-related rhythms. The single mutants of Pdf01 or CCHa1SK8 displayed free-running rhythms under constant dark conditions, whereas approximately 98% of the double mutants were arrhythmic. In light-dark conditions, the evening activity of the double mutants was phase-advanced compared with that of the single mutants. In contrast, both the single and double mutants had diminished morning activity. These results suggest that the effects of the double mutation varied in behavioural parameters. The double and triple mutants of per 01, Pdf01, and CCHa1SK8 further revealed that PDF signalling plays a role in the suppression of activity during the daytime under a clock-less background. These results provide insights into the interactions between PDF and CCHa1 signalling and their roles in activity rhythms (Kuwano, 2023).
Memory formation and sleep regulation are critical for brain functions in animals from invertebrates to humans. Neuropeptides play a pivotal role in regulating physiological behaviors, including memory formation and sleep. However, the detailed mechanisms by which neuropeptides regulate these physiological behaviors remains unclear. This study report sthat neuropeptide diuretic hormone 31 (DH31) positively regulates memory formation and sleep in Drosophila melanogaster. The expression of DH31 in the dorsal and ventral fan-shaped body (dFB and vFB) neurons of the central complex and ventral lateral clock neurons (LNvs) in the brain was responsive to sleep regulation. In addition, the expression of membrane-tethered DH31 in dFB neurons rescued sleep defects in Dh31 mutants, suggesting that DH31 secreted from dFB, vFB, and LNvs acts on the DH31 receptor in the dFB to regulate sleep partly in an autoregulatory feedback loop. Moreover, the expression of DH31 in octopaminergic neurons, but not in the dFB neurons, is involved in forming intermediate-term memory. These results suggest that DH31 regulates memory formation and sleep through distinct neural pathways (Lyu, 2023).
Much is known about environmental influences on metabolism and systemic insulin levels. Less is known about how those influences are translated into molecular mechanisms regulating insulin production. To better understand the molecular mechanisms this study generated marked cells homozygous for a null mutation in the Drosophila TGF-β signal transducer dSmad2 in unmated adult females. Then a side-by-side single cell comparisons was conducted of the pixel intensity of two Drosophila insulin-like peptides (dILP2 and dILP5) in dSmad2- mutant and wild type insulin producing cells (IPCs). The analysis revealed multiple features of dSmad2 regulation of dILPs. In addition, it was discovered that dILP5 is expressed and regulated by dSmad2 in circadian pacemaker cells (CPCs). Outcomes of regulation by dSmad2 differ between dILP2 and dILP5 within IPCs and differ for dILP5 between IPCs and CPCs. Modes of dSmad2 regulation differ between dILP2 and dILP5. dSmad2 antagonism of dILP2 in IPCs is robust but dSmad2 regulation of dILP5 in IPCs and CPCs toggles between antagonism and agonism depending upon dSmad2 dosage. Companion studies of dILP2 and dILP5 in the IPCs of dCORL mutant (fussel in Flybase and SKOR in mammals) and upd2 mutant unmated adult females showed no significant difference from wild type. Taken together, the data suggest that dSmad2 regulates dILP2 and dILP5 via distinct mechanisms in IPCs (antagonist) and CPCs (agonist) and in unmated adult females that dSmad2 acts independently of dCORL and Upd2 (Goldsmith, 2023).
Recent studies revealed behaviorally defined sleep is conserved across broad species from insect to human. For evolutional analysis, it is critical to determine how homologous genes regulate the homologous function among species. Drosophila melanogaster shares numerous sleep related genes with mammals including Sik3, salt-inducible kinase 3, whose mutation caused long sleep both in mouse and fruit fly. The Drosophila rdgB (retinal degeneration B) encodes a membrane-associated phosphatidylinositol transfer protein and its mutation caused light-induced degeneration of photoreceptor cells. rdgB mutation also impaired phototransduction and olfactory behavior, indicating rdgB is involved in the normal neural transmission. Mammalian rdgB homologue, Pitpnm2 (phosphatidylinositol transfer protein membrane-associated 2) was discovered as one of SNIPPs (sleep-need index phosphoproteins), suggesting its role in sleep. This study shows that rdgB is involved in sleep regulation in Drosophila. Pan-neuronal and mushroom body (MB) specific rdgB knockdown decreased nocturnal sleep. MB neurons play a dominant role, since the rescue of rdgB expression only in MB neurons in pan-neuronal knockdown reversed the sleep reducing effect of rdgB knockdown. These results revealed the sleep-related function of rdgB in Drosophila which may be conserved across species (Kobayashi, 2023).
Circadian clocks are composed of transcriptional/translational feedback loops (TTFLs) at the cellular level. In Drosophila TTFLs, the transcription factor Clock (Clk)/Cycle (Cyc) activates clock target gene expression, which is repressed by the physical interaction with Period (Per). This study shows that amino acids (AA) 657-707 of Clk, a region that is homologous to the mouse Clock exon 19-encoded region, is crucial for Per binding and E-box-dependent transactivation in S2 cells. Consistently, in transgenic flies expressing Clk with an AA657-707 deletion in the Clock (Clkout) genetic background (p{Clk-Δ};Clkout), oscillation of core clock genes' mRNAs displayed diminished amplitude compared with control flies, and the highly abundant dCLK&Delta657-707 showed significantly decreased binding to Per. Behaviorally, the p{dClk-Δ};Clkout flies exhibited arrhythmic locomotor behavior in the photic entrainment condition but showed anticipatory activities of temperature transition and improved free-running rhythms in the temperature entrainment condition. Surprisingly, p{dClk-Δ};Clkout flies showed pacemaker-neuron-dependent alterations in molecular rhythms; the abundance of dCLK target clock proteins was reduced in ventral lateral neurons (LNvs) but not in dorsal neurons (DNs) in both entrainment conditions. In p{dClk-Delta};Clkout flies, however, strong but delayed molecular oscillations in temperature cycle-sensitive pacemaker neurons, such as DN1s and DN2s, were correlated with delayed anticipatory activities of temperature transition. Taken together, this study reveals that the LNv molecular clockwork is more sensitive than the clockwork of DNs to dysregulation of dCLK by AA657-707 deletion. Therefore, it is proposed that the dCLK/CYC-controlled TTFL operates differently in subsets of pacemaker neurons, which may contribute to their specific functions (Lee, 2023).
The circadian clock system enables organisms to anticipate the rhythmic environmental changes and to manifest behavior and physiology at advantageous times of day. Transcriptional/translational feedback loop (TTFL) is the basic feature of eukaryotic circadian clock and is based on the rhythmic association of circadian transcriptional activator and repressor. In Drosophila, repression of dCLOCK/CYCLE (dCLK/CYC) mediated transcription by PERIOD (PER) is critical for inducing circadian rhythms of gene expression. Pacemaker neurons in the brain control specific circadian behaviors upon environmental timing cues such as light and temperature cycle. This study shows that amino acids 657-707 of dCLK is important for the transcriptional activation and the association with PER both in vitro and in vivo. Flies expressing dCLK lacking AA657-707 in Clkout genetic background, homologous to the mouse Clock allele where exon 19 region is deleted, display pacemaker-neuron-dependent perturbation of the molecular clockwork. Namely, the molecular rhythms in light-cycle-sensitive pacemaker neurons such as ventral lateral neurons (LNvs) were significantly disrupted but those in temperature-cycle-sensitive pacemaker neurons such as dorsal neurons (DNs) were robust. These results suggest that the dCLK-controlled TTFL diversified in pacemaker-neuron-dependent manner which contribute to specific functions such as different sensitivities to entraining cues (Cho, 2016).
Despite the significant advance in understanding of the molecular basis of light entrainment of the circadian clock in Drosophila, the underlying genetic architecture is still largely unknown. The aim of this study was to identify loci associated with variation in circadian photosensitivity, which are important for the evolution of this trait. Complementary approaches were used that combined quantitative trait loci (QTL) mapping, complementation testing, and transcriptome profiling to dissect this variation. A major QTL was identified on chromosome 2, which was subsequently fine mapped using deficiency complementation mapping into 2 smaller regions spanning 139 genes, some of which are known to be involved in functions that have been previously implicated in light entrainment. Two genes implicated with the clock and located within that interval, timeless and cycle, failed to complement the QTL, indicating that alleles of these genes contribute to the variation in light response. Specifically, the timeless s/ls polymorphism that has been previously shown to constitute a latitudinal cline in Europe is also segregating in the recombinant inbred lines and is contributing to the phenotypic variation in light sensitivity. This study also profiled gene expression in 2 recombinant inbred strains that differ significantly in their photosensitivity and a total of 368 transcripts were identified that showed differential expression (false discovery rate < 0.1). Of 131 transcripts that showed a significant recombinant inbred line by treatment interaction (i.e., putative expression QTL), 4 are located within QTL2 (Adewoye, 2017).
Amino-acid transporters are involved in functions reportedly linked to the sleep/wake cycle: neurotransmitter synthesis and recycling, the regulation of synaptic strength, protein synthesis and energy metabolism. In addition, the existence of bidirectional relationships between extracellular content, transport systems and sleep/wake states is receiving emerging support. Nevertheless, the connection between amino-acid transport and sleep/wake regulation remains elusive. To address this question, this study used Drosophila melanogaster and investigated the role of LAT1 (Large neutral Amino-acid Transporter 1) transporters. This study shows that the two Drosophila LAT1-like transporters: JhI-21 and minidiscs (Mnd) are required in dopaminergic neurons for sleep/wake regulation. Down-regulating either gene in dopaminergic neurons resulted in higher daily sleep and longer sleep bout duration during the night, suggesting a defect in dopaminergic transmission. Since LAT1 transporters can mediate in mammals the uptake of L-DOPA, a precursor of dopamine, amino-acid transport efficiency was assessed by L-DOPA feeding. Downregulation of JhI-21, but not Mnd, reduced the sensitivity to L-DOPA as measured by sleep loss. JhI-21 downregulation also attenuated the sleep loss induced by continuous activation of dopaminergic neurons. Since LAT1 transporters are known to regulate TOR (Target Of Rapamycin) signaling, the role of this amino-acid sensing pathway in dopaminergic neurons was investigated. Consistently, it is reported that TOR activity in dopaminergic neurons modulates sleep/wake states. Altogether, this study provides evidence that LAT1 mediated amino-acid transport in dopaminergic neurons, is playing a significant role in sleep/wake regulation, and is providing several entry points to elucidate the role of nutrients such as amino-acids in sleep/wake regulation (Aboudhiaf, 2018).
Emerging evidence suggests bidirectional relationships between extracellular space content and vigilance states, emphasizing the so far little explored sleep-regulatory role of the transmembrane transport of ions and small molecules. Sleep and wakefulness have a pervasive impact on brain cellular activities linked to neurotransmission, neuronal plasticity, neurotransmitter synthesis, nutrient supply, and waste elimination, relying on the efficient and precise coordination of transport systems. Investigating how transporters and underlying molecular and cellular mechanisms are involved in these mutual interactions requires the targeting of individual genes in specific cell types. This strategy is highly amenable to the Drosophila model. Among the large array of cellular transporters present in the genome and conserved between insects and mammals, the well-characterized large neutral amino acid transporters are particularly relevant given their role in neurotransmitter synthesis and recycling, in the regulation of synaptic strength, in protein synthesis and energy metabolism. In addition, the de novo synthesis of brain monoamines associated with wakefulness and neuromodulation, such as serotonin, dopamine, and noradrenaline, is dependent on large neutral essential amino acids provided by the blood (Aboudhiaf, 2018).
The SLC7A5 (or large neutral amino-acid transporter, light chain or LAT1) and SLC7A8 (LAT2) amino acid transporters are present in most cell types and appear to play a prominent role in the Na+-independent transport of large branched and aromatic neutral amino acids. These transporters belong to the heterodimeric amino acid transporters (HAT) family and require co-expression of the CD98hc /4F2hc (SLC3A2) heavy chain, to which they can be covalently linked by a di-sulfur bridge. The heavy subunit does not appear to confer transport-specific properties, nor to be confined to HAT transporter function. In the mammalian brain, LAT1 is highly expressed in the cells of the blood-brain-barrier and is thought to play a critical role in providing the central nervous system with essential amino acids such as phenylalanine, tyrosine, leucine, and tryptophan, which are nutrients and precursors for monoamine synthesis. The uptake of leucine through LAT1 is a major activator signal for target of rapamycin complex 1 (TORC1), a cellular pathway dependent on the TOR kinase that controls protein synthesis, brain excitability, and plasticity. Reciprocally, inhibition of mTOR by rapamycin has been shown to significantly reduce the activity and the mRNA expression of LAT1. Despite a few pieces of evidence, it remains to be investigated whether LAT1, LAT2, or other SLC7A transporters are also localized in neurons, and whether their function is linked to sleep and wake. In Drosophila, the HAT family of transporters is represented by one heavy chain, CD98hc, and five light chains: juvenile hormone inducible-21 (JHI-21), minidiscs (MND), genderblind (GB), CG9413 protein, and CG1607 protein. The specific activity of these transporters cannot be easily predicted from their sequences and requires functional testing. JHI-21 and MND can transport leucine, are inhibited by BCH (2-aminobicyclo[2.2.1]heptane-2-carboxylic acid), and, at least for JHI-21, require CD98hc to become functional, thus classifying them as LAT1-LAT2 homologs. Both genes are expressed at high levels in the central nervous system and have been shown to play important neurophysiological functions. JHI-21 and GB regulate glutamatergic synaptic strength, primarily through the regulation of extracellular glutamate levels, and are, respectively, required in motor neurons and in glial cells. In a recent report, it has also been shown that MND is required to activate brain insulin-producing neurons in response to circulating leucine. Interestingly those same insulin-producing neurons are connected to wakefulness promoting circuits. This study used the molecular-genetic tools of Drosophila to investigate the potential impact of JhI-21 and Mnd downregulation in neuronal subsets on sleep/wake regulation (Aboudhiaf, 2018).
Evidence is provided that the LAT-1 like transporters JhI-21 and Mnd are required in adult fly dopaminergic neurons to achieve adequate sleep/wake regulation. The results demonstrate that a downregulation of these transporters in dopaminergic neurons results in a decrease in wakefulness, under baseline conditions but also in conditions that increase dopaminergic transmission. This implies that the activity of dopaminergic neurons and/or their ability to release neurotransmitter requires JhI-21 and Mnd. These two amino acid transporters are the closest drosophila LAT1 homologs based on sequence and functional data, suggesting that some amino acid availability plays a critical role in dopaminergic neuronal function. Supporting this hypothesis, this study finds that downregulating the TOR pathway in dopaminergic neurons results in a decrease in wakefulness (Aboudhiaf, 2018).
As in mammals, dopaminergic transmission plays a major role in Drosophila wakefulness and has been suggested to constitute a core ancestral regulator of arousal and sleep entry across invertebrates. Drosophila and mammals share homologs for genes playing a central role in dopamine synthesis, reuptake, and signaling. This includes two D1-like receptors and one D2-like dopamine receptor. Among those, the D1-like receptor Dop1R1 (dDA1) plays a prominent role in sleep/wake regulation. Dop1R1 mutant flies display high daily sleep, longer sleep bout duration, and normal waking activity, a phenotype that closely resembles the one obtained in this studu by downregulating JhI-21 or Mnd in dopaminergic neurons. The effect of Dop1R1 on sleep/wake depends on its expression in the dorsal fan-shaped body (dFB), a key sleep-wake regulatory structure, and on the release of dopamine by a very limited set of dopaminergic neurons projecting to the dFB and located in the PPL1 and PPM3 cluster. Thus, it is possible that Mnd and JhI-21 are required in those specific neurons to achieve normal sleep-wake regulation. Expressing Mnd and JhI-21 UAS-RNAi constructs in PPL1 and PPM3 neurons failed to produce an abnormal sleep/wake phenotype. This lack of effect may be attributable to the limited efficiency of the UAS-RNAi constructs. Alternatively, the inhibition of JhI-21 and Mnd may affect multiple dopamine dependent microcircuits throughout the brain and thus cannot be easily replicated by a more specific manipulation (Aboudhiaf, 2018).
What function could JHI-21 and MND fulfill in dopaminergic neurons? At the glutamatergic neuromuscular junction, JHI-21 appears to regulate the clustering of post-synaptic glutamate receptors. JHI-21 does not appear to trigger directly the release of glutamate in this context, but possibly mediates the entry of amino acids such as leucine to activate molecular pathways controlling glutamatergic physiology. Although abnormal sleep/wake regulation was not observed when JhI-21 was inhibited in glutamatergic neurons, the results indicate that JhI-21 could play a role in dopaminergic physiology similar to the one hypothesized for the neuromuscular junction. The synthesis of brain monoamines depends on the supply of essential amino acids that are provided by food intake, thus requiring efficient cellular transport systems. Although dopaminergic neuron function is impaired when JhI-21 expression is downregulated, no evidence was found of reduced brain dopamine levels under baseline conditions. Dopamine levels were increased in the mutant flies fed with the dopamine precursor L-DOPA, an amino acid that has been shown to be transported by LAT1, further suggesting that JhI-21 expression is not critical for dopamine synthesis. In this experiment, dopamine synthesis could also take place ectopically in dopa-decarboxylase (ddc) expressing serotoninergic neurons, in which the UAS-JhI-21-RNAi construct was not expressed. Dopamine content was still increased, although moderately, in flies in which the UAS-JhI-21-RNAi construct was targeted to both dopaminergic and serotoninergic neurons using the ddc-Gal4 driver. The lack of impact of JhI21 downregulation on L-DOPA-induced increase in dopamine synthesis could be due to functional redundancy between JhI-21 and Mnd, or could be a consequence of the partial inhibition provided by the UAS-RNAi constructs. In contrast to those findings on tissue content, this study clearly observed that JhI-21 downregulation in dopaminergic neurons reduced the sensitivity to L-DOPA as measured with sleep loss, and significantly attenuated the sleep loss phenotype of the TH-GaL4 > UAS-TrpA1 hyperdopaminergic condition. This raises the possibility that JhI-21 mediates the entry of amino acids critical for the physiology of dopaminergic neurons, such as leucine that could activate TOR-dependent processes influencing protein synthesis and energy metabolism, but is not directly involved in dopamine synthesis through the transport of precursors such as tyrosine or phenylalanine. The phenotype observed in flies where both Rheb and JhI-21 are manipulated indicates that JhI-21 could be required for TOR pathway activation. This hypothetical model is conceivable since in mammals enriched intracellular leucine levels via LAT1 transport activate mTORC1. Alternatively, TOR signaling could modulate JHI-21 function and affect LAT1-like-dependent transport mechanisms required for neuronal activity. Elucidating the relationships between TOR signaling and LAT1-like transporters, and understanding how they play a critical role in dopaminergic neurons function will require further work (Aboudhiaf, 2018).
This study showed that the MND and JHI-21 transporters are broadly expressed in the brain. However, targeting the JhI-21 and Mnd UAS-RNAi constructs to most nondopaminergic cell types, including the wake-promoting octopaminergic neurons, failed to affect sleep-wake regulation. This lack of effect may be attributable to the efficiency of genetic tools or alternatively to the more stringent requirement for these transporters in dopaminergic neurons. Further studies are warranted to further evaluate this question. Interestingly, accumulating pieces of evidence support the existence of a dynamic regulation for JhI-21 and Mnd: JhI-21 is modulated during larval development in close correlation with behavioral changes and in adult after long-term memory conditioning, whereas Mnd is differentially expressed after sleep deprivation. The expression of LAT1 transporters in mammalian dopaminergic neurons has not been investigated yet; however, a report using pharmacological methods suggested that such transporters could modulate neuronal activity. The fact that sleep deprivation induces changes in TOR signaling in the brain opens the possibility that LAT1 could be modulated by sleep/wake in mammals. Of note, sleep deprivation could induce changes in LAT1 expression at the blood-brain barrier (Aboudhiaf, 2018).
In conclusion, this study reveals the role of LAT1-like transporters in the function of dopaminergic neurons, adding one more element to the array of cellular and molecular events affecting sleep/wake regulation. Since these transporters are known to be dynamically regulated by physiological cellular states, they provide an entry point to elucidate the role of nutrient in sleep/wake regulation (Aboudhiaf, 2018).
The daily expression of genes and the changes in gene expression after silencing the heme oxygenase (ho) gene were examined in the retina of Drosophila using microarray and SybrGreen qPCR (quantitative polymerase chain reaction) methods. The HO decrease in the morning upregulated 83 genes and downregulated 57 genes. At night, 80 genes were upregulated and 22 were downregulated. The top 20 genes downregulated after ho silencing in the morning modulate phototransduction, immune responses, autophagy, phagocytosis, apoptosis, the carbon monoxide (CO) response, the oxidative stress/UV response, and translation. In turn, the genes that upregulated at night were involved in translation-the response to oxidative stress, DNA damage, and phototransduction. Among the top 20 genes downregulated at night were genes involved in phototransduction, immune responses, and autophagy. For some genes, a low level of HO had an opposite effect in the morning compared to those at night. Silencing ho also changed the expression of circadian clock genes, while the HO decrease during the night enhanced the expression of immune system genes. The results showed that the cyclic expression of HO is important for controlling several processes in the retina, including neuroprotection and those involved in the innate immune system (Damulewicz, 2018).
The aging eye experiences physiological changes that include decreased visual function and increased risk of retinal degeneration. Although there are transcriptomic signatures in the aging retina that correlate with these physiological changes, the gene regulatory mechanisms that contribute to cellular homeostasis during aging remain to be determined. This study integrated ATAC-seq and RNA-seq data to identify 57 transcription factors that showed differential activity in aging Drosophila photoreceptors. These 57 age-regulated transcription factors include two circadian regulators, Clock and Cycle, that showed sustained increased activity during aging. When the Clock:Cycle complex was disrupted by expressing a dominant negative version of Clock (ClkDN) in adult photoreceptors, changes were observed in expression of 15-20% of genes including key components of the phototransduction machinery and many eye-specific transcription factors. Using ATAC-seq, expression of ClkDN in photoreceptors was shown to lead to changes in activity of 37 transcription factors and causes a progressive decrease in global levels of chromatin accessibility in photoreceptors. Supporting a key role for Clock-dependent transcription in the eye, expression of ClkDN in photoreceptors also induced light-dependent retinal degeneration and increased oxidative stress, independent of light exposure. Together, these data suggests that the circadian regulators Clock and Cycle act as neuroprotective factors in the aging eye by directing gene regulatory networks that maintain expression of the phototransduction machinery and counteract oxidative stress (Jauregui-Lozano, 2022).
Drosophila circadian behavior relies on the network of heterogeneous groups of clock neurons. Short- and long-range signaling within the pacemaker circuit coordinates molecular and neural rhythms of clock neurons to generate coherent behavioral output. The neurochemistry of circadian behavior is complex and remains incompletely understood. This study demonstrates that the gaseous messenger nitric oxide (NO) is a signaling molecule linking circadian pacemaker to rhythmic locomotor activity. mutants lacking nitric oxide synthase (NOS) have behavioral arrhythmia in constant darkness, although molecular clocks in the main pacemaker neurons are unaffected. Behavioral phenotypes of mutants are due in part to the malformation of neurites of the main pacemaker neurons, s-LNvs. Using cell-type selective and stage-specific gain- and loss-of-function of NOS, this study also demonstrated that NO secreted from diverse cellular clusters affect behavioral rhythms. Furthermore, perineurial glia, one of the two glial subtypes that form the blood-brain barrier, as the major source of NO that regulates circadian locomotor output. These results reveal for the first time the critical role of NO signaling in the Drosophila circadian system and highlight the importance of neuro-glial interaction in the neural circuit output (Kozlov, 2020).
It is rather surprising that the lack of NOS enzyme is not lethal as NO is part of various developmental processes. NOSΔ mutants are nonetheless strongly arrhythmic in DD and have reduced morning anticipation in LD. The data suggest that both congenital impairments and lack of NO signaling in adulthood contribute to the behavioral phenotype of the mutants. Axonal terminals of the master pacemaker, s-LNvs, in NOS mutants are profoundly disordered, suggesting the wrong or absent synaptic connections with the downstream partners. However, molecular rhythms in the pacemaker neurons are unaffected. During adulthood, NOS activity in the perineurial glia is required for producing free-running locomotor rhythms but not for maintaining PDF rhythms and structure of the s-LNvs. This finding indicates that NO produced in the perineurial glia is necessary for proper the functioning of circadian locomotor output circuits. Taken together, these results demonstrate that NO signaling is essential for establishing and controlling circadian output circuit (Kozlov, 2020).
The functional isoform dNOS1 shows a circadian variation of its RNA levels throughout the day, which suggest that levels of NO could cycle at least in LD. However, dNOS is likely to be regulated by its truncated isoforms in a stage- and cell-type-specific manner, which lays an additional complexity to the regulation of NO production and probably leads to the heterogeneous and context-specific variations of NO. Hyperproduction of NO modulates molecular clockwork, albeit modestly, and is generally detrimental to locomotor rhythms. Therefore, the level and potentially the rhythms of NO production should be tightly controlled in wild-type flies (Kozlov, 2020).
In an NOS RNAi mini screen, two optic lobe-specific drivers, GMR79D04 and GMR85B12, reduced locomotor rhythmicity in DD. This phenotype was observed when NOS was downregulated constitutively in these cells but not when knockdown was restricted to adulthood. These results reinforce the idea that NO is necessary for a proper establishment of neuronal circuits. A low rhythmicity phenotype caused by NOS knockdown with the pan-neuronal driver GMR57C10-GAL4 is congruent with the above findings. Intriguingly, however, in addition to the low rhythmicity, GMR57C10 > NOS RNAi in adulthood resulted in an extended period. What might be the neuronal subsets that produce NO and regulate period length of locomotor activity? A recent study by Aso (2019), NO was shown to act as a co-transmitter in a subset of dopaminergic neurons, specifically in some of the PAMs, PPL1s and PPL2abs. It is thus possible that dopamine signaling modulated by NO is involved in the control of the locomotor activity period. It is also noteworthy that NO-mediated signaling has a profound neuromodulatory effect on spinal motor networks and regulates frequency and amplitude of motor activity in various vertebrate species. NO-mediated regulation of circadian locomotor output in flies might involve a similar mechanism (Kozlov, 2020).
DAR4-M staining showed an enrichment of NO in glial cells, including the surface glia. Targeting glial cells leads to the strongest and most persistent phenotype in locomotor activity both for gain- and loss-of-function of NOS. Among glial subpopulations, the perineurial glia appears to be the major site of NOS activity that regulates locomotor rhythms. The importance of glia in circadian rhythms have been recognized, especially those containing the molecular clocks and exert reciprocal communication with the pacemaker neural circuit. It has been shown that the perineurial glial cells harbor molecular clocks, which drive daily rhythms in the blood-brain barrier permeability but are not required for locomotor activity rhythms. This study is the first to identify NO as a signaling molecule produced in glia and mediates part of the role of glia, independently of their molecular clocks, in Drosophila circadian rhythms (Kozlov, 2020).
It has been shown that in mammals NO mediates light-induced phase-shifts through the cGMP pathway. It is an interesting parallel to note that forced production of NO in the s-LNvs caused phase shift rather than amplitude dampening. Since high levels of NO inhibit E75/UNF dimerization and E75/UNF normally enhances per transcription, it is speculated that NO-induced phase-shift may be partly mediated by the inhibition of E75/UNF (Hormone receptor 51) heterodimerization. It will be interesting to test this hypothesis in future studies. Mammalian clocks contain E75 homologs REV-ERB α/β, which repress Bmal1 transcription. Analogous to the notion in flies, NO is thought to decrease REV-ERB α/β activity. Consistently, in vitro studies in mammalian cell culture showed that excessive presence of NO increases the production of Bmal1 mRNA. These findings altogether point out that NO is an evolutionarily conserved regulator of circadian rhythms (Kozlov, 2020).
In line with recent studies, this research expands the view on the factors that participate in neuronal and molecular mechanisms of circadian rhythmicity. The finding that gaseous messenger NO contributes to the various aspects of circadian rhythmicity emphasizes that non-cell-autonomous, systemic regulation is integral to the circadian circuit operation. These results set a foundation for future studies addressing the mechanism by which NO signaling modulates the state of the pacemaker circuit and its output (Kozlov, 2020).
Mesencephalic Astrocyte-derived Neurotrophic Factor (MANF) is one of a few neurotrophic factors described in Drosophila melanogaster (DmMANF) but its function is still poorly characterized. The present study found that DmMANF is expressed in different clusters of clock neurons. In particular, the PDF-positive large (l-LNv) and small (s-LNv) ventral lateral neurons, the CRYPTOCHROME-positive dorsal lateral neurons (LNd), the group 1 dorsal neurons posterior (DN1p) and different tim-positive cells in the fly's visual system. Importantly, DmMANF expression in the ventral lateral neurons is not controlled by the clock nor it affects its molecular mechanism. However, silencing DmMANF expression in clock neurons affects the rhythm of locomotor activity in light:dark and constant darkness conditions. Such phenotypes correlate with abnormal morphology of the dorsal projections of the s-LNv and with reduced arborizations of the l-LNv in the medulla of the optic lobe. Additionally, it was shown that DmMANF is important for normal morphology of the L2 interneurons in the visual system and for the circadian rhythm in the topology of their dendritic tree. These results indicate that DmMANF is important not only for the development of neurites but also for maintaining circadian plasticity of neurons (Krzeptowski, 2021).
A precise balance between sleep and wakefulness is essential to sustain a good quality of life and optimal brain function. GABA is known to play a key and conserved role in sleep control, and GABAergic tone should, therefore, be tightly controlled in sleep circuits. This study examined the role of the astrocytic GABA transporter (Gat) in sleep regulation using Drosophila melanogaster. A hypomorphic Gat mutation (Gat33-1) increased sleep amount, decreased sleep latency, and increased sleep consolidation at night. Interestingly, sleep defects were suppressed when Gat33-1 was combined with a mutation disrupting wide-awake (wake), a gene that regulates the cell-surface levels of the GABA(A) receptor Resistance to dieldrin (Rdl) in the wake-promoting large ventral lateral neurons (l-LNvs). Moreover, RNAi knockdown of Rdl and its modulators dnlg4 and wake in these circadian neurons also suppressed Gat33-1 sleep phenotypes. Brain immunohistochemistry showed that GAT-expressing astrocytes were located near RDL-positive l-LNv cell bodies and dendritic processes. It is concluded that astrocytic GAT decreases GABAergic tone and RDL activation in arousal-promoting LNvs, thus determining proper sleep amount and quality in Drosophila (Chaturvedi, 2022).
The circadian circuit, a roughly 24 h molecular feedback loop, or clock, is conserved from bacteria to animals and allows for enhanced organismal survival by facilitating the anticipation of the day/night cycle. Recent research has demonstrated that proteins comprising the circadian clock network display a significant amount of intrinsic disorder. This work focussed on the extent of intrinsic disorder in the circadian clock and its potential mechanistic role in circadian timing. The conservation of disorder was highlighted by quantifying the extent of computationally-predicted protein disorder in the core clock of the key eukaryotic circadian model organisms Drosophila melanogaster, Neurospora crassa, and Mus musculus. Previously published work, as well as feature novel experimental evidence, was examined, demonstrating that the core negative arm circadian period drivers FREQUENCY (Neurospora crassa) and PERIOD-2 (PER2) (Mus musculus), possess biochemical characteristics of intrinsically disordered proteins. Finally, the potential contributions are discussed of the inherent biophysical principals of intrinsically disordered proteins that may explain the vital mechanistic roles they play in the clock to drive their broad evolutionary conservation in circadian timekeeping. It is concluded that the pervasive conservation of disorder amongst the clock in the crown eukaryotes suggests that disorder is essential for optimal circadian timing from fungi to animals, providing vital homeostatic cellular maintenance and coordinating organismal physiology across phylogenetic kingdoms (Pelham, 2020).
Starvation caused by adverse feeding stresses or food shortages has been reported to result in sleep loss in animals. However, how the starvation signal interacts with the central nervous system is still unknown. In this study, the adipokinetic hormone (AKH)-Fork head Box-O (FOXO) pathway is shown to respond to energy change and adjust the sleep of Drosophila through remodeling of the s-LNv (small ventral lateral neurons) dorsal projections. The results show that starvation prevents flies from going to sleep after the first light-dark transition. The LNvs are required for starvation-induced sleep loss through extension of the pigment dispersing factor (PDF)-containing s-LNv dorsal projections. Further studies reveal that loss of AKH or AKHR (akh receptor) function blocks starvation-induced extension of s-LNv dorsal projections and rescues sleep suppression during food deprivation. FOXO, which has been reported to regulate synapse plasticity of neurons, acts as starvation response factor downstream of AKH, and down regulation of FOXO level considerably alleviates the influence of starvation on s-LNv dorsal projections and sleep. Taking together, these results outline the transduction pathways between starvation signal and sleep, and reveal a novel functional site for sleep regulation (He, 2020).
While wake duration is a major sleep driver, an important question is if wake quality also contributes to controlling sleep. In particular, this study sought to determine whether changes in sensory stimulation affect sleep in Drosophila. As Drosophila rely heavily on their sense of smell, this study focused on manipulating olfactory input and the olfactory sensory pathway. Sensory deprivation was first performed by removing antennae or applying glue to antennae. Sleep was then measured in response to neural activation, via TRPA1, or inhibition, via KIR2.1, of subpopulations of neurons in the olfactory pathway. Genetically restricting manipulations to adult animals prevented developmental effects. This study found that olfactory deprivation reduces sleep, largely independently of mushroom bodies that integrate olfactory signals for memory consolidation and have previously been implicated in sleep. However, specific neurons in the lateral horn, the other third order target of olfactory input, affect sleep. Also, activation of inhibitory second order projection neurons increases sleep. No single neuronal population in the olfactory processing pathway was found to bidirectionally regulate sleep, and reduced sleep in response to olfactory deprivation may be masked by temperature changes. These findings demonstrate that Drosophila sleep is sensitive to sensory stimulation, and identify novel sleep-regulating neurons in the olfactory circuit. Scaling of signals across the circuit may explain the lack of bidirectional effects when neuronal activity is manipulated. It is proposed that olfactory inputs act through specific circuit components to modulate sleep in flies (Hsu, 2020).
The circadian system produces ~24-hr oscillations in behavioral and physiological processes to ensure that they occur at optimal times of day and in the correct temporal order. At its core, the circadian system is composed of dedicated central clock neurons that keep time through a cell-autonomous molecular clock. To produce rhythmic behaviors, time-of-day information generated by clock neurons must be transmitted across output pathways to regulate the downstream neuronal populations that control the relevant behaviors. An understanding of the manner through which the circadian system enacts behavioral rhythms therefore requires the identification of the cells and molecules that make up the output pathways. To that end, the Drosophila pars intercerebralis (PI) was recently characterized as a major circadian output center that lies downstream of central clock neurons in a circuit controlling rest:activity rhythms. Single-cell RNA sequencing (scRNAseq) was performed to identify potential circadian output genes expressed by PI cells, and cell-specific RNA interference (RNAi) was used to knock down expression of ~40 of these candidate genes selectively within subsets of PI cells. Knockdown of the slowpoke (slo) potassium channel in PI cells reliably decreases circadian rest:activity rhythm strength. Interestingly, slo mutants have previously been shown to have aberrant rest:activity rhythms, in part due to a necessary function of slo within central clock cells. However, rescue of slo in all clock cells does not fully reestablish behavioral rhythms, indicating that expression in non-clock neurons is also necessary. These results demonstrate that slo exerts its effects in multiple components of the circadian circuit, including PI output cells in addition to clock neurons, and it is hypothesized that it does so by contributing to the generation of daily neuronal activity rhythms that allow for the propagation of circadian information throughout output circuits (Ruiz, 2021).
Endogenous clocks enable organisms to adapt cellular processes, physiology, and behavior to daily variation in environmental conditions. Metabolic processes in cyanobacteria to humans are under the influence of the circadian clock, and dysregulation of the circadian clock causes metabolic disorders. In mouse and Drosophila, the circadian clock influences translation of factors involved in ribosome biogenesis and synchronizes protein synthesis. Notably, nutrition signals are mediated by the insulin receptor/target of rapamycin (InR/TOR) pathways to regulate cellular metabolism and growth. However, the role of the circadian clock in Drosophila brain development and the potential impact of clock impairment on neural circuit formation and function is less understood. This study demonstrates that changes in light stimuli or disruption of the molecular circadian clock cause a defect in neural stem cell growth and proliferation. Moreover, this study shows that disturbed cell growth and proliferation are accompanied by reduced nucleolar size indicative of impaired ribosomal biogenesis. Further, this study defines that light and clock independently affect the InR/TOR growth regulatory pathway due to the effect on regulators of protein biosynthesis. Altogether, these data suggest that alterations in InR/TOR signaling induced by changes in light conditions or disruption of the molecular clock have an impact on growth and proliferation properties of neural stem cells in the developing Drosophila brain (Dapergola, 2021).
The Drosophila circadian network dictates the temporal organization of locomotor activity; under light-dark (LD) conditions, flies display a robust bimodal pattern. PDF-positive small ventral lateral neurons (sLNv) have been linked to the generation of the morning activity peak (the "M cells"), whereas the CRY-positive dorsal lateral neurons (LNds) and the PDF-negative sLNv are necessary for the evening activity peak (the "E cells"). While each group directly controls locomotor output pathways, an interplay between them along with a third dorsal cluster (the DN1ps) is necessary for the correct timing of each peak and for adjusting behavior to changes in the environment. M cells set the phase of roughly half of the circadian neurons (including the E cells) through PDF. This study shows the existence of synaptic input provided by the evening oscillator onto the M cells. Both structural and functional approaches revealed that E-to-M cell connectivity changes across the day, with higher excitatory input taking place before the day-to-night transition. Two different neurotransmitters, acetylcholine and glutamate, released by E cells were identified that are relevant for robust circadian output. Indeed, this study shows that acetylcholine is responsible for the excitatory input from E cells to M cells, which show preferential responsiveness to acetylcholine during the evening. These findings provide evidence of an excitatory feedback between circadian clusters and unveil an important plastic remodeling of the E cells' synaptic connections (Duhart, 2020a).
In addition to timed neuronal activation and neuropeptide release, circadian remodeling of neuronal connectivity provides a mechanism to timely restrict circadian neuronal input/output. To explore mechanisms modulating E cell communication, this study sought for changes in presynaptic sites of the E cells by means of expressing an RFP-tagged version of Bruchpilot (BRP), a component of presynaptic active zones. Although the complexity of the E cell arborization pattern precluded a description based on membrane labeling, quantification of BRP particle numbers have proven to uncover circadian synaptic remodeling. The MB122B-splitGal4 and the Mai179Gal4;pdfGal80 drivers were employed to direct expression to E cells. As previously described, E cells showed an extensive arborization of putative presynaptic zones, marked by dense BRPRFP staining, in close proximity to M cell somas, axons, and dorsal terminals; particularly, in the dorsal region, a significantly lower number of active zones were detected at the beginning of the day (ZT2) and higher numbers toward the end of the light phase (at ZT10, 'late-day'). This time-of-day difference was confirmed through a second E cell driver (Mai179Gal4;pdfGal80). Interestingly, the timing of E cell maximum synaptic density coincides with the highest level of neuronal activity, similar to what has previously been described for M cells, suggesting this might be a general mechanism coupling circadian control of structural synaptic output to cellular excitability. E cell presynaptic changes were not detected near the cell bodies or the main axonal tract of the M cells but were paralleled by changes in M cell postsynaptic markers at the dorsal protocerebrum (Gorostiza, 2014), suggesting that these two groups are reciprocally remodeling their terminals in this region (Duhart, 2020a).
To test the possibility that E cells could provide input to M cells, its targets were trans-synaptically tagging with trans-Tango. Out of the two available drivers, the MB122B-splitGal4 was selected because it shows minimal expression beyond this circadian subset, thus providing a more precise tool to map E cell postsynaptic contacts. This approach uncovered a restricted number of yet unidentified neurons (likely including the short neuropeptide F (sNPF)-positive dorsal lateral neurons [LNds], which are postsynaptic to the ion transport peptide (ITP)-positive LNd and 5th small ventral lateral neurons [sLNv]) whose somas and profuse arborizations are located within the dorsolateral region; more importantly, trans-synaptic labeling revealed that pigment-dispersing factor (PDF)+ M cells are postsynaptic to E cells (Duhart, 2020a).
Trans-Tango experiments do not allow any temporal resolution. To gain more insight on the sites and plastic changes of E-to-M connectivity, GFP reconstitution across synaptic partners (GRASP) was employed to direct the expression of a presynaptically tagged GFP fragment to the E cells and describe their degree of contact across the day. Using both the strong Mai179Gal4;pdfGal80 and the specific MB122B-splitGal4 E cell drivers, evidence was found of E-to-M synaptic contacts in the M cell dorsal terminals and axonal tracts. Indeed, GRASP intensity at the dorsal region (but not in the axonal tracts) was higher at dusk compared to dawn, a further indication of strengthened E-to-M connectivity at the dorsal protocerebrum during late day. Of note, GRASP signal was also high toward the end of the night, as it had originally been reported employing a non-targeted split GFP, suggesting that the connection between E and M cells might also be relevant for the night-day transition (Duhart, 2020a).
E cells have the ability to affect the morning activity component, but a functional assessment for E-to-M cell communication has not been reported yet. To test this possibility, E cells were depolarized by means of an ATP perfusion that activates the ectopically expressed P2X2 channel. An excitatory response was uncovered in M cells, revealed by an increase in intracellular calcium levels , indicating a functional excitatory feedback from E to M cells (Duhart, 2020a).
The observation of structural changes in the E cell terminals suggests that the functional E-to-M communication might be subject to daily regulation. Accordingly, it was found that forced depolarization of the E cells led to M cell responses predominantly during the second half of the light period; specifically, at ZT8-10, responses are statistically different from those obtained in the remaining intervals, with the exception of those triggered at ZT6-8. This coincides with the window of maximum E cell presynaptic contacts at the dorsal terminals. Single trace analysis enabled comparison between the individual responses triggered at different times, which highlighted not only the variability among the responses observed at any given time point but also the differential kinetics of M cell responses, which appear to differ at ZT22-0 (faster) compared to ZT8-10 (slower, albeit no statistical analysis of this property was performed (Duhart, 2020a).
Taken together, these findings confirm the E-to-M cell excitatory communication and show daily changes in the functional synaptic connectivity between key components of the Drosophila circadian network (Duhart, 2020a).
Within the circadian network it has been shown that fast neurotransmission from M cells promotes network synchronization; likewise, DN1ps contribute to shape the activity profile under light-dark (LD) cycles through fast neurotransmission. It was hypothesized that fast neurotransmission from E cells would be part of the mechanisms underlying proper circadian outputs and the identity of the putative neurotransmitter(s) were sought by means of expressing specific RNAis to downregulate genes involved in their synthesis, release, or reuptake. Acetylcholine (ACh) and glutamate (Glut) were identified as strong candidates to play a role in circadian timekeeping. To avoid any compensatory or deleterious effect throughout development, RNAi expression was restricted to adulthood by means of the Target system. E cell downregulation of the vesicular acetylcholine transporter (vAChT) or the choline acetyltransferase (ChAT; that catalyzes the biosynthesis of the neurotransmitter acetylcholine) affected the consolidation of the endogenous locomotor activity rhythms, without inducing changes in the clock's speed, confirming the importance of ACh released by the E cells for network synchronization. Interestingly, the evening anticipatory activity was reduced by vAChT and ChAT downregulation in E cells, a behavioral signature of E cell activity under LD cycles (Duhart, 2020a).
Fast neurotransmitter co-release has been described both in vertebrates and invertebrates, and peptidergic/fast neurotransmitter co-transmission is present within the Drosophila circadian network, which led to examining the role of Glut as a second E cell neurotransmitter. Adult-restricted expression of the vesicular glutamate transporter (vGluT) RNAi impaired behavioral rhythmic output, triggering a reduced consolidation of the activity patterns as well as a small change in the circadian period of endogenous rhythms without significantly affecting the LD locomotor patterns. These results confirmed the role of glutamatergic neurotransmission released by E cells, at least under free running conditions. Reduction in rhythm robustness was previously reported by silencing E cells with the inward-rectifying potassium channel Kir2.1, which coincides with the behavioral phenotype uncovered upon cholinergic and glutamatergic downregulation in E cells (Duhart, 2020a).
In addition to classical neurotransmitters, the PDF neuropeptide is crucial for proper coupling among circadian clusters and for modulating the activity of locomotor centers. Interestingly, constant PDF levels are found in flies with defective consolidation of activity rhythms. Thus, it was reasoned that the phenotype associated to prevent neurotransmitter release from E cells could possibly involve alterations in PDF cycling as an output of the network. Along those lines, it was found that impairing cholinergic neurotransmission in E cells abolished PDF oscillations at the sLNv dorsal terminals, with similar levels to those found at night in controls. Likewise, impairing glutamatergic neurotransmission particularly affected PDF levels in the morning, albeit some residual cycling was still observed, suggesting that a balanced neurotransmission from E cells is necessary for proper neuropeptidergic output of the M cells. The idea is favored that reduced amplitude of PDF cycling is the result of an altered balance of sLNv activity, which depends not only on clock-dependent control of ion channel expression but also on direct/indirect input from the circadian network [tested at least for the large LNvs (lLNvs)] (Duhart, 2020a).
Since E cell activation led to M cell depolarization, and ACh from E cells impacted on circadian timekeeping, it was hypothesized that the M cells would be one of the cholinergic targets of the E cells. Bath application of the nicotine antagonist curare prevented M cell excitatory responses triggered by acute depolarization of E cells between ZT6-10. As previously reported, its effects were difficult to wash; however, about 30% of the M cells partially recovered after washing the antagonist (i.e., the fluorescence after washout was at least two times higher than with curare). Taking into account that the M cell response to E cell activation showed an unequivocal time-of-day dependency, and that PDF-positive cells have previously been shown to respond to bath application of Ach, whether M cells exhibit a differential response to ACh was tested at times where E-to-M connectivity differed significantly. Bath application of ACh to brains expressing GCaMP6m in PDF cells led to strong calcium responses in both the lLNv (also PDF-positive cells but not formally part of the M oscillator) and M cells during the evening. However, although ACh application induced equivalent responses in the lLNv during the morning, M cells' activation was highly reduced at this time point, underscoring that the sensitivity of these cells to cholinergic inputs is time-of-day dependent. Cholinergic inputs lead to both Ca2+ and cAMP increases in M cells through the nicotinic ACh receptor (nAChR). Therefore, changes in the levels of the nAChR itself or in the intracellular pathways activated by ACh throughout the day might also contribute to the control of E to M cell cholinergic circuit (Duhart, 2020a).
In addition to the well-studied role of neuropeptides, classical neurotransmitters contribute to the coordination of the multi-oscillator circadian network. Both E cells and DN1ps respond to an inhibitory glycine tone likely released by M cells. Additionally, an inhibitory feedback from the glutamatergic DN1ps to the M and E cells further refines the temporal organization of the activity pattern. The notion of reciprocal regulation between oscillator pairs prompted an investigate of whether E cells feedback onto the M oscillator. The results point toward an excitatory cholinergic feedback from E to M cells during the late day (i.e., around ZT8-10). In M cells, intracellular calcium levels show a strong time-of-day dependence, with a peak shortly before lights-on, declining over the light phase, and rising after lights-off. Thus, increased excitability of E cells and the higher synaptic excitatory input to M cells could provide activating signals which, integrated with other inputs and self-autonomous mechanisms, would ultimately lead to the change in intracellular calcium. Loss of excitatory feedback would then prevent the increase of PDF levels at the dorsal terminals of M cells (through recruitment of PDF-filled dense-core vesicles) and thus affect other timed signaling outputs, leading to network desynchronization and hence, deconsolidated behavioral rhythms. Within the circadian network, E, M, and DN1p clusters express mGluRA RNA, and glutamatergic input from DN1ps impacts both M and E cells. Thus, glutamate released from E cells could impact in any of these clusters, in a time-of-day controlled manner, providing an inhibitory output from E cells into the network. Specifically, the highest connectivity detected through GRASP opens the possibility that glutamatergic input from E to M cells reaches its maximum toward the end of the night, consistent with the inhibitory responses observed during that window, which could directly or indirectly imbalance the active recruitment of PDF dense-core vesicles to the terminals, hence PDF release in the morning. Alternatively, loss of glutamatergic inhibitory input onto glutamatergic DN1ps might result in a persisting increase of the activity of this cluster, which in turn could increase their inhibition onto M cells and also lead to impaired PDF release, a change in the pace of the molecular clock, and hence, deconsolidation of rest-activity patterns (Duhart, 2020a).
This work uncovers that the E cells communicate through fast neurotransmitters with a tight time-dependent control and a versatile output nature. Daily axonal remodeling of neuronal terminals-along with clock-controlled changes in the postsynaptic response to neurotransmitters-leads to plastic functional connectivity and provides novel building blocks to the flexible circadian network. This, in turn, enables fine-tuning reciprocal connections between oscillator pairs, further expanding its ability to adapt to environmental cycles. Thus, rhythmic behavior represents an emerging property of the plastic communication among clock neurons (Duhart, 2020a).
Sleep is a highly conserved state, suggesting that sleep's benefits outweigh the increased vulnerability it brings. Yet, little is known about how sleep fulfills its functions. This study used video tracking in tethered flies to identify a discrete deep sleep stage in Drosophila, termed proboscis extension sleep, that is defined by repeated stereotyped proboscis extensions and retractions. Proboscis extension sleep is accompanied by highly elevated arousal thresholds and decreased brain activity, indicative of a deep sleep state. Preventing proboscis extensions increases injury-related mortality and reduces waste clearance. Sleep deprivation reduces waste clearance and during subsequent rebound sleep, sleep, proboscis extensions, and waste clearance are increased. Together, these results provide evidence of a discrete deep sleep stage that is linked to a specific function and suggest that waste clearance is a core and ancient function of deep sleep (van Alphen, 2021).
The fruit fly, Drosophila melanogaster, has been used as a model organism for the molecular and genetic dissection of sleeping behaviors. However, most previous studies were based on qualitative or semi-quantitative characterizations. This study quantified sleep in flies. An assay was set up to continuously track the activity of flies using infrared camera, which monitored the movement of tens of flies simultaneously with high spatial and temporal resolution. Accurate statistics were obtained regarding the rest and sleep patterns of single flies. Analysis of the data has revealed a general pattern of rest and sleep: the rest statistics obeyed a power law distribution and the sleep statistics obeyed an exponential distribution. Thus, a resting fly would start to move again with a probability that decreased with the time it has rested, whereas a sleeping fly would wake up with a probability independent of how long it had slept. Resting transits to sleeping at time scales of minutes. This method allows quantitative investigations of resting and sleeping behaviors and these results provide insights for mechanisms of falling into and waking up from sleep (Xu, 2021).
Time-restricted feeding (TRF) has recently gained interest as a potential anti-ageing treatment for organisms from Drosophila to humans. TRF restricts food intake to specific hours of the day. Because TRF controls the timing of feeding, rather than nutrient or caloric content, TRF has been hypothesized to depend on circadian-regulated functions; the underlying molecular mechanisms of its effects remain unclear. To exploit the genetic tools and well-characterized ageing markers of Drosophila, this study developed an intermittent TRF (iTRF) dietary regimen that robustly extended fly lifespan and delayed the onset of ageing markers in the muscles and gut. iTRF enhanced circadian-regulated transcription, and iTRF-mediated lifespan extension required both circadian regulation and autophagy, a conserved longevity pathway. Night-specific induction of autophagy was both necessary and sufficient to extend lifespan on an ad libitum diet and also prevented further iTRF-mediated lifespan extension. By contrast, day-specific induction of autophagy did not extend lifespan. Thus, these results identify circadian-regulated autophagy as a critical contributor to iTRF-mediated health benefits in Drosophila. Because both circadian regulation and autophagy are highly conserved processes in human ageing, this work highlights the possibility that behavioural or pharmaceutical interventions that stimulate circadian-regulated autophagy might provide people with similar health benefits, such as delayed ageing and lifespan extension (Ulgherait, 2021).
Balance between sleep, wakefulness and arousal is important for survival of organisms and species as a whole. While, the benefits of sleep both in terms of quantity and quality is widely recognized across species, sleep has a cost for organismal survival and reproduction. here the study focuses on how sleep duration, sleep depth and sleep pressure affect the ability of animals to engage in courtship and egg-laying behaviors critical for reproductive success. Using isogenic lines from the Drosophila Genetic Reference Panel with variable sleep phenotypes this study investigated the relationship between sleep and reproductive behaviors, courtship and oviposition. This study found that three out of five lines with decreased sleep and increased arousal phenotypes, showed increased courtship and decreased latency to court as compared to normal and long sleeping lines. However, the male courtship phenotype is dependent on context and genotype as some but not all long sleeping-low courting lines elevate their courtship in the presence of short sleeping-high courting flies. Sleep phenotypes were less variable and minimally susceptible to social experience. In addition to male courtship, Oviposition was found to be less sensitive to sleep length. Taken together this extensive behavioral analysis shows complex bidirectional interactions between genotype and environment and add to the growing evidence linking sleep duration and sleep-wake switch parameters to behavioral decision making critical to reproductive output (Buchert, 2021).
Circadian disturbances are early features of neurodegenerative diseases, including Huntington's disease (HD). Emerging evidence suggests that circadian decline feeds into neurodegenerative symptoms, exacerbating them. Therefore, it was asked whether known neurotoxic modifiers can suppress circadian dysfunction. A screen was performed of neurotoxicity-modifier genes to suppress circadian behavioural arrhythmicity in a Drosophila circadian HD model. The molecular chaperones Hsp40 and HSP70 emerged as significant suppressors in the circadian context, with Hsp40 being the more potent mitigator. Upon Hsp40 overexpression in the Drosophila circadian ventrolateral neurons (LNv), the behavioural rescue was associated with neuronal rescue of loss of circadian proteins from small LNv soma. Specifically, there was a restoration of the molecular clock protein Period and its oscillations in young flies and a long-lasting rescue of the output neuropeptide Pigment dispersing factor. Significantly, there was a reduction in the expanded Huntingtin inclusion load, concomitant with the appearance of a spot-like Huntingtin form. Thus, this study provided evidence implicating the neuroprotective chaperone Hsp40 in circadian rehabilitation. The involvement of molecular chaperones in circadian maintenance has broader therapeutic implications for neurodegenerative diseases (Prakash, 2022).
Sleep is a very important behavior observed in almost all animals. Importantly, sleep is subject to both circadian and homeostatic regulation. The circadian rhythm determines the daily alternation of the sleep-wake cycle, while homeostasis mediates the rise and dissipation of sleep pressure during the wake and sleep period. As an important kinase, Dbt plays a central role in both circadian rhythms and development. This study investigated the sleep patterns of several ethyl methanesulfonate-induced dbt mutants and discusses the possible reasons why different sleep phenotypes were shown in these mutants. In order to reduce DBT in all neurons in which it is expressed, CRISPR-Cas9 was used to produce flies that expressed GAL4 in frame with the dbt gene at its endogenous locus, and knock-down of DBT with this construct produced elevated sleep during the day and reduced sleep at night. Loss of sleep at night is mediated by dbt loss during the sleep/wake cycle in the adult, while the increased sleep during the day is produced by reductions in dbt during development and not by reductions in the adult. Additionally, using targeted RNA interference, this study uncovered the contribution of dbt on sleep in different subsets of neurons in which dbt is normally expressed. Reduction of dbt in circadian neurons produced less sleep at night, while lower expression of dbt in noncircadian neurons produced increased sleep during the day. Importantly, independently of the types of neurons where dbt affects sleep, this study demonstrated that the PER protein is involved in DBT mediated sleep regulation (Wang, 2022).
The ability of organisms to sense their nutritional environment and adjust their behavior accordingly is critical for survival. Insulin-like peptides (ilps) play major roles in controlling behavior and metabolism; however, the tissues and cells that insulin acts on to regulate these processes are not fully understood. In the fruit fly, Drosophila melanogaster, insulin signaling has been shown to function in the fat body to regulate lipid storage, but whether ilps act on the fly brain to regulate nutrient storage is not known. This study manipulated insulin signaling in defined populations of neurons in Drosophila and measure glycogen and triglyceride storage. Expressing a constitutively active form of the insulin receptor (dInR) in the insulin-producing cells had no effect on glycogen or triglyceride levels. However, activating insulin signaling in the Drosulfakinin (Dsk)-producing neurons led to triglyceride accumulation and increased food consumption. The expression of ilp2, ilp3 and ilp5 was increased in flies with activated insulin signaling in the Dsk neurons, which along with the feeding phenotype, may cause the triglyceride storage phenotypes observed in these flies. In addition, expressing a constitutively active dInR in Dsk neurons resulted in decreased sleep in the fed state and less starvation-induced sleep suppression suggesting a role for insulin signaling in regulating nutrient-responsive behaviors. Together, these data support a role for insulin signaling in the Dsk-producing neurons for regulating behavior and maintaining metabolic homeostasis (Palermo, 2022).
Feeding behavior is essential for growth and survival of animals; however, relatively little is known about its intrinsic mechanisms. This study demonstrates that Gart is expressed in the glia, fat body, and gut and positively regulates feeding behavior via cooperation and coordination. Gart in the gut is crucial for maintaining endogenous feeding rhythms and food intake, while Gart in the glia and fat body regulates energy homeostasis between synthesis and metabolism. These roles of Gart further impact Drosophila lifespan. Importantly, Gart expression is directly regulated by the CLOCK/CYCLE heterodimer via canonical E-box, in which the CLOCKs (CLKs) in the glia, fat body, and gut positively regulate Gart of peripheral tissues, while the core CLK in brain negatively controls Gart of peripheral tissues. This study provides insight into the complex and subtle regulatory mechanisms of feeding and lifespan extension in animals (He, 2023).
Feeding is a necessary behavior for animals to grow and survive, with a characteristic of taking food regularly. The quality and quantity of feeding directly impact the normal growth and development of animals. Time-restricted feeding or fasting is beneficial for preventing obesity, alleviating inflammation, and attenuating cardiac diseases and even has antitumor effects. Metabolic syndrome has become a global health problem. Shift work and meal irregularity disrupt circadian rhythms, with an increased risk of developing metabolic syndrome. The maintenance of the daily feeding rhythm is very important in metabolic homeostasis.Irregular feeding perturbs circadian metabolic rhythms and results in adverse metabolic consequences and chronic diseases (He, 2023).
Most behaviors in animals are synchronized to a ~24 h (circadian) rhythm induced by circadian clocks in both the central nervous system and peripheral tissues. Circadian rhythmic behaviors, such as feeding and locomotion, are involved in complex connections and specific output pathways under the control of the circadian clock. Although the core clock feedback loop has been well established in recent decades, the crucial genes responsible for rhythmic feeding regulation as well as for the interrelation between the core clocks and feeding are still unclear (He, 2023).
To increase the understanding of how the circadian clock regulates feeding and metabolism, this study sought to identify the output genes in the circadian feeding and metabolism control network, in which the model animal Drosophila provides special advantages to explore the mechanistic underpinnings and the complex integration of these primitive responses. Previous studies confirmed that one of juvenile hormone receptors, methoprene tolerance (Met), is important for the control of neurite development and sleep behavior in Drosophila. Many genes related to metabolic regulation have attracted attention in the transcriptome data from Met27, a Met-deficient fly line, in which this study focused on the target genes regulated by CLOCK/CYCLE (CLK/CYC). As a basic Helix-Loop-Helix-Per-ARNT-Sim (bHLH-PAS) transcription factor with a canonical binding site “CACGTG," the CLK/CYC heterodimer is a crucial core oscillator that regulates circadian rhythms (He, 2023).
The Gart trifunctional enzyme, a homologous gene of adenosine-3 in mammals, is a trifunctional polypeptide with the activities of phosphoribosylglycinamide formyltransferase, phosphoribosylglycinamide synthetase, and phosphoribosylaminoimidazole synthetase (Tiong, 1990). Gart in astrocytes of vertebrates plays a role in the lipopolysaccharide-induced release of proinflammatory factors (Zhang, 2014), and Gart expressed in the liver and heart is required for de novo purine synthesis. However, there is no information yet for Gart's functions in feeding rhythm. In this study, Gart was identified as a candidate that was controlled by the core clock gene CLK/CYC heterodimer and was found to be related to feeding behavior in Drosophila. Thus this study focused Gart studies on the role of feeding rhythms and further regulatory mechanisms. This study provides a critical foundation for understanding the complex feeding mechanism. (He, 2023).
In animals, hundreds of genes exhibit daily oscillation under clock regulation, and some of them are involved in metabolic functions. This study identified a CLK/CYC-binding gene, Gart, which is involved in feeding rhythms and energy metabolism independent of locomotor rhythms. Previous research reported that blocking CLK in different tissues yields different phenotypes. This study found that MET, like CYC, can combine with CLK to regulate the transcription of Gart. Knocking down Gart in different tissues exhibits different phenotypes, and Gart in different tissues can rescue the phenotype caused by CLK deletion; thus, the phenomenon caused by CLK deletion is due to the change in Gart (He, 2023).
CLK regulates the feeding rhythms of Drosophila, and its loss can cause disorders of feeding rhythms and abnormal energy storage. Tim01, Cry01, and Per01 mutants have significantly lower levels of truactkglycerides (TAGs). The maintenance of energy homeostasis is achieved by a dynamic balance of energy intake (feeding), storage, and expenditure (metabolic rate), which are crucial factors for longevity and resistance to adverse environments in Drosophila. Additionally, studies have shown that mutations of Timeless and per shorten the adult lifespan of Drosophila. This study further reveals that peripheral CLKs control the oscillation of Gart among different peripheral tissues; however, core CLKs in the brain can negatively regulate Gart expression in peripheral tissues, indicating that a complex and refined network regulatory system exists between CLK and Gart in the brain and in different peripheral tissues to coordinate feeding behavior and energy homeostasis in Drosophila and that it further affects sensitivity to starvation and longevity. These novel findings enrich the network of regulatory mechanisms by the clocks-Gart pathway on feeding, energy homeostasis, and longevity (He, 2023).
Glial cells have vital functions in neuronal development, activity, plasticity, and recovery from injury. This study reveals that flies lacking Gart in glial cells display a significant decline in the viability of Drosophila under starvation, caused by a decrease in energy storage that puts flies under a state of energy deficit. This discovery extends the functions of glial cells in feeding, energy storage, and starvation resistance controlled by Gart (He, 2023).
The fat body is the primary energy tissue for the storage of fuel molecules, such as TAG and glycogen, which play an important role in the regulation of metabolic homeostasis and provide the most energy during starvation. Indeed, functional defects of the fat body increase starvation sensitivity in Drosophila. In this study, flies lacking Gart in the fat body led to decreased energy storage, which reduces the survival rate and the survival time under starvation conditions. However, flies lacking gut Gart still maintain normal energy storage, which is not sensitive to food shortage or starvation. In addition, this study found that although high temperature can stimulate the food intake of Drosophila, which is consistent with previous reports, it does not affect the feeding rhythm (He, 2023).
This study reveals that Gart in the glia and the fat body collectively regulate the homeostasis of energy intake, storage, and expenditure, thereby influencing the viability of flies under starvation stress. Although Gart in the gut strongly influences feeding behavior, it does not play similar functions as the glia and the fat body in adversity resistance. This occurs possibly because the gut has vital roles in digestion and absorption, while the fat body has crucial functions in energy metabolism. In addition, Gart in the glia and the fat body has biased roles in the synthesis of glycogen and TAG, despite having similar functions in energy storage. The biased role of the glia and the fat body may be coordinated to provide effective energy homeostasis. These findings provide new insight into how specific circadian coordination of various tissues modulates adversity resistance and aging (He, 2023).
Such robust findings in Drosophila suggest that a decrease in lifespan and an increase in sensitivity to starvation in Drosophila is a faithful readout of disordered feeding rhythms and abnormal metabolism. Gart effects on metabolism in both glia cells and the fat body indicate the intricacy of the circadian network and energy homeostasis. It is crucial for animals to have a well-organized network to coordinate and ensure that these various tissue regions are in a normal state (He, 2023).
This study has demonstrated that CLK regulates feeding, energy homeostasis, and longevity via Gart. Even though attempts were made to explore more comprehensively how Gart coordinates and regulates the physiological functions in different tissues of D. melanogaster, there are still some limitations. For instance, it is still unclear that how Gart achieves functional differentiation in different tissues, as well as whether Gart regulates lifespan through autophagy and/or bacterial content or not, which are two critical factors related to lifespan. These future studies are of great significance for understanding the relationship between feeding and longevity regulated by Gart (He, 2023).
Circadian clocks generate rhythms of arousal, but the underlying molecular and cellular mechanisms remain unclear. In Drosophila, the clock output molecule WIDE AWAKE (WAKE) labels rhythmic neural networks and cyclically regulates sleep and arousal. This study shows, in a male mouse model, that mWAKE/ANKFN1 labels a subpopulation of dorsomedial hypothalamus (DMH) neurons involved in rhythmic arousal and acts in the DMH to reduce arousal at night. In vivo Ca(2+) imaging reveals elevated DMH(mWAKE) activity during wakefulness and rapid eye movement (REM) sleep, while patch-clamp recordings show that DMH(mWAKE) neurons fire more frequently at night. Chemogenetic manipulations demonstrate that DMH(mWAKE) neurons are necessary and sufficient for arousal. Single-cell profiling coupled with optogenetic activation experiments suggest that GABAergic DMH(mWAKE) neurons promote arousal. Surprisingly, the data suggest that mWAKE acts as a clock-dependent brake on arousal during the night, when mice are normally active. mWAKE levels peak at night under clock control, and loss of mWAKE leads to hyperarousal and greater DMH(mWAKE) neuronal excitability specifically at night. These results suggest that the clock does not solely promote arousal during an animal's active period, but instead uses opposing processes to produce appropriate levels of arousal in a time-dependent manner (Liu, 2023).
Circadian clocks in terrestrial animals are encoded by molecular feedback loops involving the negative regulators PERIOD, TIMELESS or CRYPTOCHROME2 (see Drosophila Cryptochrome) and positive transcription factors CLOCK and BMAL1/CYCLE. The molecular basis of circatidal (~12.4 hour) or other lunar-mediated cycles (~15 day, ~29 day), widely expressed in coastal organisms, is unknown. Pharmacological inhibition of casein kinase 1 (CK1) that targets PERIOD stability in mammals and flies, affects both circadian and circatidal phenotypes in Eurydice pulchra (Ep), the speckled sea-louse. This study shows that these drug inhibitors of CK1 also affect the phosphorylation of EpCLK and EpBMAL1 and disrupt EpCLK-BMAL1-mediated transcription in Drosophila S2 cells, revealing a potential link between these two positive circadian regulators and circatidal behaviour. DsRNAi knockdown of Epbmal1 as well as the major negative regulator in Eurydice, Epcry2 was performed in animals taken from the wild. Epcry2 and Epbmal1 knockdown disrupted Eurydice's circadian phenotypes of chromatophore dispersion, tim mRNA cycling and the circadian modulation of circatidal swimming, as expected. However, circatidal behaviour was particularly sensitive to Epbmal1 knockdown with consistent effects on the power, amplitude and rhythmicity of the circatidal swimming cycle. Thus, three Eurydice negative circadian regulators, EpCRY2, in addition to EpPER and EpTIM (from a previous study), do not appear to be required for the expression of robust circatidal behaviour, in contrast to the positive regulator EpBMAL1. A neurogenetic model is suggested whereby the positive circadian regulators EpBMAL1-CLK are shared between circadian and circatidal mechanisms in Eurydice but circatidal rhythms require a novel, as yet unknown negative regulator (Lin, 2023).
Sleep loss typically imposes negative effects on animal health. However, humans with a rare genetic mutation in the dec2 gene (dec2P384R) present an exception; these individuals sleep less without the usual effects associated with sleep deprivation. Thus, it has been suggested that the dec2P384R mutation activates compensatory mechanisms that allows these individuals to thrive with less sleep. To test this directly, a Drosophila model was used to study the effects of the dec2P384R mutation on animal health. Expression of human dec2P384R in fly sleep neurons was sufficient to mimic the short sleep phenotype and, remarkably, dec2P384R mutants lived significantly longer with improved health despite sleeping less. The improved physiological effects were enabled, in part, by enhanced mitochondrial fitness and upregulation of multiple stress response pathways. Moreover, evidence is provided that upregulation of pro-health pathways also contributes to the short sleep phenotype, and this phenomenon may extend to other pro-longevity models (Pandey, 2023).
Circadian rhythms time physiological and behavioral processes to 24-hour cycles. It is generally assumed that most cells contain self-sustained circadian clocks that drive circadian rhythms in gene expression that ultimately generating circadian rhythms in physiology. While those clocks supposedly act cell autonomously, current work suggests that in Drosophila some of them can be adjusted by the brain circadian pacemaker through neuropeptides, like the Pigment Dispersing Factor (PDF). Despite these findings and the ample knowledge of the molecular clockwork, it is still unknown how circadian gene expression in Drosophila is achieved across the body. This study used single-cell and bulk RNAseq data to identify cells within the fly that express core-clock components. Surprisingly, it was found that less than a third of the cell types in the fly express core-clock genes. Moreover, Lamina wild field (Lawf) and Pox-neuro positive (Poxn) neurons were identified as putative new circadian neurons. In addition, several cell types were found that do not express core clock components but are highly enriched for cyclically expressed mRNAs. Strikingly, these cell types express the PDF receptor (Pdfr), suggesting that PDF drives rhythmic gene expression in many cell types in flies. Other cell types express both core circadian clock components and Pdfr, suggesting that in these cells, PDF regulates the phase of rhythmic gene expression. Together, these data suggest three different mechanisms generate cyclic daily gene expression in cells and tissues: canonical endogenous canonical molecular clock, PDF signaling-driven expression, or a combination of both (Patop, 2023).
All but the simplest phenotypes are believed to result from interactions between two or more genes forming complex networks of gene regulation. Sleep is a complex trait known to depend on the system of feedback loops of the circadian clock, and on many other genes; however, the main components regulating the phenotype and how they interact remain an unsolved puzzle. Genomic and transcriptomic data may well provide part of the answer, but a full account requires a suitable quantitative framework. An artificial selection experiment was conducted for sleep duration with RNA-seq data acquired each generation. The phenotypic results are robust across replicates and previous experiments, and the transcription data provides a high-resolution, time-course data set for the evolution of sleep-related gene expression. In addition to a Hierarchical Generalized Linear Model analysis of differential expression that accounts for experimental replicates a flexible Gaussian Process model was constructed that estimates interactions between genes. 145 gene pairs are found to have interactions that are different from controls. Our method appears to be not only more specific than standard correlation metrics but also more sensitive, finding correlations not significant by other methods. Statistical predictions were compared to experimental data from public databases on gene interactions. Mutations of candidate genes implicated by these results affected night sleep, and gene expression profiles largely met predicted gene-gene interactions (Souto-Maior, 2023).
Genetic variants underlying traits that become either non-adaptive or selectively neutral are expected to have altered evolutionary trajectories. Uncovering genetic signatures associated with phenotypic loss presents the opportunity to discover the molecular basis for the phenotype in populations where it persists. Circalunar clocks were studied in populations of the marine midge Clunio marinus. The circalunar clock synchronizes development to the lunar phase, and it is set by moonlight and tidal cycles of mechanical agitation. Two out of ten studied populations have lost their sensitivity to mechanical agitation while preserving sensitivity to moonlight. Intriguingly, the F1 offspring of the two insensitive populations regained the sensitivity to mechanical entrainment, implying a genetically independent loss of the phenotype. By combining quantitative trait locus mapping and genome-wide screens, this study explored the genetics of this phenotypic loss. QTL analysis suggested an oligogenic origin with one prevalent additive locus in one of the strains. In addition, it confirmed a distinct genetic architecture in the two insensitive populations. Genomic screens further uncovered several candidate genes underlying QTL regions. The strongest signal under the most prominent QTL contains a duplicated STAT1 gene, which has a well-established role in development, and CG022363, an ortholog of the Drosophila melanogaster CG32100 gene, which plays a role in gravitaxis. These results support the notion that adaptive phenotypes have a complex genetic basis with mutations occurring at several loci. By dissecting the most prevalent signals, this study started to reveal the molecular machinery responsible for the entrainment of the circalunar clock (Brisevac, 2023).
Circadian clocks are timing devices that rhythmically adjust organism's behavior, physiology, and metabolism to the 24-h day-night cycle. Eukaryotic circadian clocks rely on several interlocked transcription-translation feedback loops, where protein stability is the key part of the delay between transcription and the appearance of the mature proteins within the feedback loops. In bilaterian animals, including mammals and insects, the circadian clock depends on a homologous set of proteins. Despite mostly conserved clock components among the fruit fly Drosophila and mammals, several lineage-specific differences exist. This study has systematically explored the evolution and sequence variability of insect DBT proteins and their vertebrate homologs casein kinase 1 delta (CKIδ) and epsilon (CKIε), dated the origin and separation of CKIδ from CKIε, and identified at least three additional independent duplications of the CKIδ/ε gene in Petromyzon, Danio, and Xenopus. Conserved regions were identified in DBT specific to Diptera, and a subset of those were functionally tested in D. melanogaster. Replacement of Lysine K224 with acidic residues strongly impacts the free-running period even in heterozygous flies, whereas homozygous mutants are not viable. K224D mutants have a temperature compensation defect with longer free-running periods at higher temperatures, which is exactly the opposite trend of what was reported for corresponding mammalian mutants. All DBTs of dipteran insects contain the NKRQK motif at positions 220-224. The occurrence of this motif perfectly correlates with the presence of BRIDE OF DOUBLETIME, BDBT, in Diptera. BDBT is a non-canonical FK506-binding protein that physically interacts with Drosophila DBT. The phylogeny of FK506-binding proteins suggests that BDBT is either absent or highly modified in non-dipteran insects. In addition to in silico analysis of DBT/CKIδ/ε evolution and diversity, four novel casein kinase 1 genes specific to the Drosophila genus were identified (Thakkar, 2023).
Central pacemaker neurons regulate circadian rhythms and undergo diurnal variation in electrical activity in mammals and flies. Whether and how intracellular chloride regulates circadian rhythms remains controversial. This study demonstrates a signaling role for intracellular chloride in the Drosophila small ventral lateral (sLN(v)) pacemaker neurons. In control flies, intracellular chloride increases in sLN(v)s over the course of the morning. Chloride transport through sodium-potassium-2-chloride (NKCC) and potassium-chloride (KCC) cotransporters is a major determinant of intracellular chloride concentrations. Drosophila melanogaster with loss-of-function mutations in the NKCC encoded by Ncc69 have abnormally low intracellular chloride 6 h after lights on, loss of morning anticipation, and a prolonged circadian period. Loss of kcc, which is expected to increase intracellular chloride, suppresses the long-period phenotype of Ncc69 mutant flies. Activation of a chloride-inhibited kinase cascade, consisting of WNK (with no lysine [K]) kinase and its downstream substrate, Fray, is necessary and sufficient to prolong period length. Fray activation of an inwardly rectifying potassium channel, Irk1, is also required for the long-period phenotype. These results indicate that the NKCC-dependent rise in intracellular chloride in Drosophila sLN(v) pacemakers restrains WNK-Fray signaling and overactivation of an inwardly rectifying potassium channel to maintain normal circadian period length (Schellinger, 2022).
Intracellular chloride oscillates in central pacemaker neurons of the mammalian SCN, but the functional significance of this oscillation has remained unclear. This study demonstrates an NKCC-dependent increase in intracellular chloride in Drosophila sLNv pacemaker neurons over the course of the morning, which constrains activity of the chloride-sensitive WNK kinase; its downstream substrate, Fray; and an inwardly rectifying potassium channel, Irk1, to maintain normal circadian periodicity. Loss of the Ncc69 NKCC in LNv pacemaker neurons results in loss of morning anticipation and lengthening of the circadian period in free-running conditions. These observations are consistent with studies in the SCN implicating NKCC in the determination of intracellular chloride in mammalian pacemaker neurons and connects intracellular chloride to behavioral circadian phenotypes (Schellinger, 2022).
Diurnal variations in intracellular chloride have been proposed to influence the effect of GABAergic neurotransmission on clock neurons in mammals, but a recent study challenged this idea.
Existing data indicate a minor role for sLNv ligand-gated chloride channels in the regulation of circadian period. Rather, this study demonstrated a signaling role for chloride in sLNvs via inhibition of WNK-Fray signaling. As chloride-sensitive kinases, WNKs are poised to interpret changes in intracellular chloride and initiate downstream signal transduction cascades. This has been studied in transepithelial ion transport in Drosophila and mammalian renal epithelia, as well as in the clearance of apoptotic corpses (Schellinger, 2022).
These findings further extend this concept to circadian pacemaker neurons.
KCC and another cation-chloride cotransporter, encoded by the NKCC gene, have been linked to the electrophysiological response to GABA in lLNvs, which express GABAA receptor chloride channels. The transport activity of the NKCC-encoded transporter differs from the Ncc69 NKCC. Expression of Ncc69 and NKCC may also differ. Consistent with this idea, broad clock neuron knockout of NKCC has no effect on morning anticipation or period length but leads to abnormal rhythmicity in constant light, as does NKCC overexpression or loss or gain of kcc. Thus, intracellular chloride likely affects clock neurons both by affecting the driving force for chloride through neurotransmitter-gated chloride channels and via inhibition of WNK-Fray signaling. Interestingly, knocking down WNK and Fray broadly in clock neurons phenocopies loss of kcc and NKCC in the same neurons, suggesting that WNK and Fray act in their usual regulatory roles upstream of the transporters in this case, and further highlighting distinct actions of this pathway in different subpopulations of Drosophila clock neurons (Schellinger, 2022).
Pacemaker neurons in flies and mammals undergo cell-autonomous, molecular clock-controlled circadian variation in electrical activity, and altering the excitability of the LNv pacemakers disrupts circadian rhythms. Two voltage-gated potassium channels have been implicated in LNv neuron electrical oscillations, and a sodium leak current and potassium channels contribute to the day/night cycling of resting membrane potential in Drosophila dorsal clock neurons. Because inwardly rectifying potassium channels play an important role in determining cellular membrane potential,
which is thought to be a determinant of circadian variation in electrical activity, chloride regulation of Irk1 activity could also contribute to the diurnal variation in sLNv excitability. Specifically, Irk1 activation at ZT6 due to low intracellular chloride and activation of the WNK-Fray pathway in Ncc69 mutants may disrupt the usual circadian pattern of membrane depolarization and hyperpolarization in sLNvs, thereby altering period length (Schellinger, 2022).
Could intracellular chloride play a signaling role in SCN pacemaker neurons? NKCC1, KCCs, and WNK3 are expressed in the rat SCN. The repertoire of ion channels regulating pacemaker neuron excitability is complex and incompletely understood, but large-conductance Ca2+-activated potassium channels have been implicated and are regulated by mammalian WNKs. Whether oscillating intracellular chloride observed in SCN neurons regulates these or other ion channels modulating pacemaker neuron electrical properties will be of interest for future investigation (Schellinger, 2022).
Sleep behavior has been observed from non-vertebrates to humans. Sleepy mutation in mice resulted in a notable increase in sleep and was identified as an exon-skipping mutation of the salt-inducible kinase 3 (Sik3) gene (See Drosophila Sik3), conserved among animals. The skipped exon includes a serine residue that is phosphorylated by protein kinase A. Overexpression of a mutant gene with the conversion of this serine into alanine (Sik3-SA) increased sleep in both mice and the fruit fly Drosophila melanogaster. However, the mechanism by which Sik3-SA increases sleep remains unclear. This study found that Sik3-SA overexpression in all neurons increased sleep under both light-dark (LD) conditions and constant dark (DD) conditions in Drosophila. Additionally, overexpression of Sik3-SA only in PDF neurons, which are a cluster of clock neurons regulating the circadian rhythm, increased sleep during subjective daytime while decreasing the amplitude of circadian rhythm. Furthermore, suppressing Sik3-SA overexpression specifically in PDF neurons in flies overexpressing Sik3-SA in all neurons reversed the sleep increase during subjective daytime. These results indicate that Sik3-SA alters the circadian function of PDF neurons and leads to an increase in sleep during subjective daytime under constant dark conditions (Kobayashi, 2023).
Besides the ~24-hour circadian rhythms, ~12-hour ultradian rhythms of gene expression, metabolism and behaviors exist in animals ranging from crustaceans to mammals. Three major hypotheses were proposed on the origin and mechanisms of regulation of ~12-hour rhythms, namely that they are not cell-autonomous and controlled by a combination of the circadian clock and environmental cues, that they are regulated by two anti-phase circadian transcriptional factors in a cell-autonomous manner, or that they are established by a cell-autonomous ~12-hour oscillator. To distinguish among these possibilities, a post-hoc analysis was performed of two high temporal resolution transcriptome dataset in animals and cells lacking the canonical circadian clock. In both the liver of BMAL1 knockout mice and Drosophila S2 cells, robust and prevalent ~12-hour rhythms of gene expression were observed, enriched in fundamental processes of mRNA and protein metabolism that show large convergence with those identified in wild-type mice liver. Bioinformatics analysis further predicted ELF1 (Drosophila Ecdysone-induced protein 74EF) and ATF6B (Drosophila Atf6) as putative transcription factors regulating the ~12-hour rhythms of gene expression independently of the circadian clock in both fly and mice. These findings provide additional evidence to support the existence of an evolutionarily conserved 12-hour oscillator that controls ~12-hour rhythms of gene expression of protein and mRNA metabolism in multiple species (Zhu, 2023).
Chromatin organization plays a crucial role in gene regulation by controlling the accessibility of DNA to transcription machinery. While significant progress has been made in understanding the regulatory role of clock proteins in circadian rhythms, how chromatin organization affects circadian rhythms remains poorly understood. ATAC-seq (Assay for Transposase-Accessible Chromatin with Sequencing) was employed on Drosophila clock neurons to assess genome-wide chromatin accessibility over the circadian cycle. Significant circadian oscillations were observed in chromatin accessibility at promoter and enhancer regions of hundreds of genes, with enhanced accessibility either at dusk or dawn, which correlated with their peak transcriptional activity. Notably, genes with enhanced accessibility at dusk were enriched with E-box motifs, while those more accessible at dawn were enriched with VRI/PDP1-box motifs, indicating that they are regulated by the core circadian feedback loops, PER/CLK and VRI/PDP1, respectively. Further, a complete loss was observed of chromatin accessibility rhythms in per01 null mutants, with chromatin consistently accessible throughout the circadian cycle, underscoring the critical role of Period protein in driving chromatin compaction during the repression phase. Together, this study demonstrates the significant role of chromatin organization in circadian regulation, revealing how the interplay between clock proteins and chromatin structure orchestrates the precise timing of biological processes throughout the day. This work further implies that variations in chromatin accessibility might play a central role in the generation of diverse circadian gene expression patterns in clock neurons (Yuan, 2023).
Royal jelly (RJ) is a biologically active substance secreted by the hypopharyngeal and mandibular glands of worker honeybees. It is widely claimed that RJ reduces oxidative stress. However, the antioxidant activity of RJ has mostly been determined by in vitro chemical detection methods or by external administration drugs that cause oxidative stress. Whether RJ can clear the endogenous production of reactive oxygen species (ROS) in cells remains largely unknown. This study systematically investigated the antioxidant properties of RJ using several endogenous oxidative stress models of Drosophila. RJ was found to enhance sleep quality of aging Drosophila, which is decreased due to an increase of oxidative damage with age. RJ supplementation improved survival and suppressed ROS levels in gut cells of flies upon exposure to hydrogen peroxide or to the neurotoxic agent paraquat. Moreover, RJ supplementation moderated levels of ROS in endogenous gut cells and extended lifespan after exposure of flies to heat stress. Sleep deprivation leads to accumulation of ROS in the gut cells, and RJ attenuated the consequences of oxidative stress caused by sleep loss and prolonged lifespan. Mechanistically, RJ prevented cell oxidative damage caused by heat stress or sleep deprivation, with the antioxidant activity in vivo independent of Keap1/Nrf2 signaling. RJ supplementation activated oxidoreductase activity in the guts of flies, suggesting its ability to inhibit endogenous oxidative stress and maintain health, possibly in humans (Wen, 2024).
Sleeprelates to numerous biological functions, including metabolism. Both dietary conditions and genes related to metabolism are known to affect sleep behavior. Insulin signaling is well conserved across species including the fruit fly and relates to both metabolism and sleep. However, the neural mechanism of sleep regulation by insulin signaling is poorly understood. Here, we report that insulin signaling in specific neurons regulates sleep in Drosophila melanogaster. This study analyzed the sleep behavior of flies with the mutation in insulin-like ligands expressed in the brain and found that three insulin-like ligands participate in sleep regulation with some redundancy. Twenty-one Gal4 drivers were used to express a dominant-negative form of the insulin receptor (InR DN) in various neurons including circadian clock neurons, which express the clock gene, and the pars intercerebralis (PI). Inhibition of insulin signaling in the anterior dorsal neuron group 1 (DN1a) decreased sleep. Additionally, the same manipulation in PI also decreased sleep. Pan-neuronal induced expression of InR DN also decrease4ple, it was demonstrated that H3K27ac and H3K9ac represent universal active marks in promoters, rather than worm-specific marks as previously reported. Second, acting regions of the studied active marks that are common across species (and across a wide range of tissues at different developmental stages) were found to extend beyond the previously defined regions. Thus, it appears that the active histone codes analyzed have a universality that has previously been underappreciated. These results suggested that these universal codes, including those previously considered species-specific, could have an ancient origin, and are important in regulating animal gene expression abundance (Yamaguchi, 2022).
Circadian rhythms and sleep homeostasis constitute the two-process model for daily sleep regulation. However, evidence for circadian control of sleep-wake cycles has been relatively short since clock-less animals often show sleep behaviors quantitatively comparable to wild-type. This study examined Drosophila sleep behaviors under different light-dark regimes and demonstrates that circadian clocks gate light-induced arousal. Genetic excitation of tyrosine decarboxylase 2 (TDC2)-expressing neurons suppressed sleep more evidently at night, causing nocturnal activity. The arousal effects were likely mediated in part by glutamate transmission from the octopaminergic neurons and substantially masked by light. Application of T12 cycles (6-h light: 6-h dark) further showed that the light-sensitive effects of TDC2 neurons depended on the time of the day. In particular, light-sensing via visual input pathway led to strong sleep suppression at subjective night, and such an effect disappeared in clock-less mutants. Transgenic mapping revealed that light-induced arousal and free-running behavioral rhythms require distinct groups of circadian pacemaker neurons. These results provide convincing evidence that circadian control of sleep is mediated by the dedicated clock neurons for light-induced arousal (Lee, 2023).
Liver Kinase B1 (LKB1) is known as a master kinase for 14 kinases related to the adenosine monophosphate-activated protein kinase. Two of them salt inducible kinase 3 and adenosine monophosphate-activated protein kinase α have previously been implicated in sleep regulation. This study generated loss-of-function mutants for Lkb1 in both Drosophila and mice. Sleep, but not circadian rhythms, was reduced in Lkb1-mutant flies and in flies with neuronal deletion of Lkb1. Genetic interactions between Lkb1 and threonine to alanine mutation at residue 184 of adenosine monophosphate-activated protein kinase in Drosophila sleep or those between Lkb1 and Threonine to Glutamic Acid mutation at residue 196 of salt inducible kinase 3 in Drosophila viability have been observed. Sleep was reduced in mice after virally mediated reduction of Lkb1 in the brain. Electroencephalography analysis showed that nonrapid eye movement sleep and sleep need were both reduced in Lkb1-mutant mice. These results indicate that liver kinase B1 plays a physiological role in sleep regulation conserved from flies to mice (Liu, 2022).
Sleep is a universal and extremely complicated function. Sleep is regulated by two systems-sleep homeostasis and circadian rhythms. In a wide range of species, neuropeptides have been found to play a crucial role in the communication and synchronization between different components of both systems. In the fruit fly Drosophila melanogaster, SIFamide (SIFa) is a neuropeptide that has been reported to be expressed in 4 neurons in the pars intercerebralis (PI) area of the brain. Previous work has shown that transgenic ablation of SIFa neurons, mutation of SIFa itself, or knockdown of SIFa receptors reduces sleep, suggesting that SIFa is sleep-promoting. However, those were all constitutive manipulations that could have affected development or resulted in compensation, so the role of SIFa signaling in sleep regulation during adulthood remains unclear. This study, examined the sleep-promoting effect of SIFa through an optogenetic approach, which allowed for neuronal activation with high temporal resolution, while leaving development unaffected. Activation of the red-light sensor Chrimson in SIFa neurons was found to promote sleep in flies in a sexually dimorphic manner, where the magnitude of the sleep effect was greater in females than in males. Because neuropeptidergic neurons often also release other transmitters, RNA interference was used to knock down SIFa while also optogenetically activating SIFa neurons. SIFa knockdown only partially reduced the magnitude of the sleep effect, suggesting that release of other transmitters may contribute to the sleep induction when SIFa neurons are activated. Video-based analysis showed that activation of SIFa neurons for as brief a period as 1 second was able to decrease walking behavior for minutes after the stimulus. Future studies should aim to identify the transmitters that are utilized by SIFa neurons and characterize their upstream activators and downstream targets. It would also be of interest to determine how acute optogenetic activation of SIFa neurons alters other behaviors that have been linked to SIFa, such as mating and feeding (Huang, 2021).
Sex peptide (SP), a seminal fluid protein of Drosophila melanogaster males, has been described as driving a virgin-to-mated switch in females, through eliciting an array of responses including increased egg laying, activity, and food intake and a decreased remating rate. While it is known that SP achieves this, at least in part, by altering neuronal signaling in females, the genetic architecture and temporal dynamics of the female's response to SP remain elusive. This study used a high-resolution time series RNA-sequencing dataset of female heads at 10 time points within the first 24 h after mating to learn about the genetic architecture, at the gene and exon levels, of the female's response to SP. SP was found to be noessential to trigger early aspects of a virgin-to-mated transcriptional switch, which includes changes in a metabolic gene regulatory network. However, SP is needed to maintain and diversify metabolic changes and to trigger changes in a neuronal gene regulatory network. It was further found that SP alters rhythmic gene expression in females and suggests that SP's disruption of the female's circadian rhythm might be key to its widespread effects (Delbare, 2023).
Circadian clocks are self-sustained molecular oscillators controlling daily changes of behavioral activity and physiology. For functional reliability and precision, the frequency of these molecular oscillations must be stable at different environmental temperatures, known as "temperature compensation." Despite being an intrinsic property of all circadian clocks, this phenomenon is not well understood at the molecular level. This study used behavioral and molecular approaches to characterize a novel mutation in the period (per) The circadian rhythm is an evolutionarily conserved mechanism with translational regulation increasingly recognized as pivotal in its modulation. This study found that upstream open reading frames (uORFs) are enriched in Drosophila circadian rhythm genes, with particularly conserved uORFs present in core circadian clock genes. Evidence is presented that the uORFs of the core clock gene, Clock (Clk), rhythmically and substantially attenuate CLK protein translation in Drosophila, with pronounced suppression occurring during daylight hours. Eliminating Clk uORFs leads to increased CLK protein levels during the day and results in a shortened circadian cycle, along with a broad shift in clock gene expression rhythms. Notably, Clk uORF deletion also augments morning sleep by reducing dopaminergic activity. Beyond daily circadian adjustments, Clk uORFs play a role in modulating sleep patterns in response to seasonal daylight variations. Furthermore, the Cl uORFs act as an important regulator to shape the rhythmic expression of a vast array of genes and influence multifaceted physiological outcomes. Collectively, this research sheds light on the intricate ways uORFs dynamically adjust downstream coding sequences to acclimate to environmental shifts (Sun, 2025).
The circadian clock organizes the physiology and behavior of organisms to their daily environmental rhythms. The central circadian timekeeping mechanism in eukaryotic cells is the transcriptional-translational feedback loop (TTFL). In the Drosophila TTFL, the transcription factors CLOCK (CLK) and CYCLE (CYC) play crucial roles in activating expression of core clock genes and clock-controlled genes. Many signaling pathways converge on the CLK/CYC complex and regulate its activity to fine-tune the cellular oscillator to environmental time cues. This study aimed to identify factors that regulate CLK by performing tandem affinity purification (TAP) combined with mass spectrometry (MS) using Drosophila S2 cells that stably express HA/FLAG-tagged CLK and V5-tagged CYC. SNF4Agamma, a homolog of mammalian AMP-activated protein kinase gamma (AMPKgamma), was identified as a factor that co-purified with HA/FLAG-tagged CLK. The AMPK holoenzyme composed of a catalytic subunit AMPKalpha and two regulatory subunits, AMPKbeta and AMPKgamma, directly phosphorylated purified CLK in vitro. Locomotor behavior analysis in Drosophila revealed that knockdown of each AMPK subunit in pacemaker neurons induced arrhythmicity and long periods. Knockdown of AMPKbeta reduced CLK levels in pacemaker neurons, and thereby reduced pre-mRNA and protein levels of CLK downstream core clock genes such as period and vrille. Finally, overexpression of CLK reversed the long-period phenotype that resulted from AMPKbeta knockdown. Thus, it is concluded that AMPK, a central regulator of cellular energy metabolism, regulates the Drosophila circadian clock by stabilizing CLK and activating CLK/CYC-dependent transcription (Cho, 2019).
MicroRNAs are important coordinators of circadian regulation that mediate the fine-tuning of gene expression. The global functional miRNA-mRNA interaction network involved in the circadian system remains poorly understood. This study used CLEAR (Covalent Ligation of Endogenous Argonaute-bound RNAs)-CLIP (Cross-Linking and Immuno-Precipitation) to explore the regulatory functions of miRNAs in the circadian system by comparing the miRNA-mRNA interactions between Drosophila wild-type strain W(1118) and a mutant of the key circadian transcriptional regulator Clock (Clk(jrk)). This experimental approach unambiguously identified tens of thousands of miRNA-mRNA interactions in both the head and body. The miRNA-mRNA interactome showed dramatic changes in the Clk(jrk) flies. Particularly, among ~300 miRNA-mRNA circadian relevant interactions, multiple interactions involving core clock genes pdp1, tim, and vri displayed distinct changes as a result of the Clk mutation. Based on the CLEAR-CLIP analysis, this study found a novel regulation of the circadian rhythm and sleep by the miR-375-timeless interaction. The results indicated that Clk disruption abolished normal rhythmic expression of miR-375 and the functional regulation occurred in the l-LNv neurons, where miR-375 modulated the circadian rhythm and sleep via targeting timeless. This work provides the first global view of miRNA regulation in the circadian rhythm (Xia, 2020).
Circadian clocks regulate ∼24-h oscillations in gene expression, behavior, and physiology. This study has elucidated how spatiotemporal organization and dynamics of core clock proteins and genes affect circadian rhythms in Drosophila clock neurons. Using high-resolution imaging and DNA-fluorescence in situ hybridization techniques, it was demonstrate that Drosophila clock proteins (PERIOD and CLOCK) are organized into a few discrete foci at the nuclear envelope during the circadian repression phase and play an important role in the subnuclear localization of core clock genes to control circadian rhythms. Specifically, this study showed that core clock genes, period and timeless, are positioned close to the nuclear periphery by the PERIOD protein specifically during the repression phase, suggesting that subnuclear localization of core clock genes might play a key role in their rhythmic gene expression. Finally, loss of Lamin B receptor, a nuclear envelope protein, was shown to leads to disruption of PER foci and per gene peripheral localization and results in circadian rhythm defects. These results demonstrate that clock proteins play a hitherto unexpected role in the subnuclear reorganization of core clock genes to control circadian rhythms, revealing how clocks function at the subcellular level. The results further suggest that clock protein foci might regulate dynamic clustering and spatial reorganization of clock-regulated genes over the repression phase to control circadian rhythms in behavior and physiology (Xiao, 2021).
Sleep facilitates memory storage and even brief periods of sleep loss lead to impairments in memory, particularly memories that are hippocampus dependent. Previous studies have shown that the deficit in memory seen after sleep loss is accompanied by deficits in synaptic plasticity. Previous work has also found that sleep deprivation (SD) is associated with reduced levels of cyclic adenosine monophosphate (cAMP) in the hippocampus and that the reduction of cAMP mediates the diminished memory observed in sleep-deprived animals. Based on these findings, it was hypothesized that cAMP acts as a mediator for not only the cognitive deficits caused by sleep deprivation, but also the observed deficits in synaptic plasticity. This study expressed the heterologous Drosophila melanogaster Gαs-protein-coupled octopamine receptor (DmOctβ1R) in mouse hippocampal neurons. This receptor is selectively activated by the systemically injected ligand (octopamine), thus leading to increased cAMP levels in hippocampal neurons during a 5-h sleep deprivation period. The results show that chemogenetic enhancement of cAMP during the period of sleep deprivation prevents deficits in a persistent form of long-term potentiation (LTP) that is induced at the Schaffer collateral synapses in the hippocampal CA1 region. It was found that elevating cAMP levels in either the first or second half of sleep deprivation successfully prevented LTP deficits. These findings reveal that cAMP-dependent signaling pathways are key mediators of sleep deprivation at the synaptic level. Targeting these pathways could be useful in designing strategies to prevent the impact of sleep loss (Walsh, 2023).
Circadian clocks align various behaviors such as locomotor activity, sleep/wake, feeding, and mating to times of day that are most adaptive. How rhythmic information in pacemaker circuits is translated to neuronal outputs is not well understood. This study used brain-wide, 24-h in vivo calcium imaging in the Drosophila brain and searched for circadian rhythmic activity among identified clusters of dopaminergic (DA) and peptidergic neurosecretory (NS) neurons. Such rhythms were widespread and imposed by the PERIOD-dependent clock activity within the ~150-cell circadian pacemaker network. The rhythms displayed either a morning (M), evening (E), or mid-day (MD) phase. Different subgroups of circadian pacemakers imposed neural activity rhythms onto different downstream non-clock neurons. Outputs from the canonical M and E pacemakers converged to regulate DA-PPM3 and DA-PAL neurons. E pacemakers regulate the evening-active DA-PPL1 neurons. In addition to these canonical M and E oscillators, evidence is for a third dedicated phase occurring at mid-day: the l-LNv pacemakers present the MD activity peak, and they regulate the MD-active DA-PPM1/2 neurons and three distinct NS cell types. Thus, the Drosophila circadian pacemaker network is a polyphasic rhythm generator. It presents dedicated M, E, and MD phases that are functionally transduced as neuronal outputs to organize diverse daily activity patterns in downstream circuits (Liang, 2023).
Animals display daily rhythms in a variety of physiological processes and behaviors, such as locomotor activity, sleep/wake, feeding, and mating behaviors.1,2 Many such rhythms are controlled by circadian timing mechanisms, and they exhibit a variety of phases throughout the solar day. Furthermore, the daily spectrum of circadian phases is itself regulated by daily changes in the presentation of environmental zeitgebers, especially light and temperature. Under laboratory conditions, the locomotor activity of the fruit fly Drosophila peaks twice a day, in the morning (M) and in the evening (E). During the M peak, the fly shows a daily peak of feeding behavior3 and mating behavior.4,5 Following each of these two activity peaks, the fly exhibits two separate sleep bouts: one around mid-day (MD) and the other generally throughout the night. These behavioral rhythms are driven by synchronous clock gene oscillations (molecular clocks) in ∼150 circadian pacemaker neurons. How a small population of circadian neurons, sharing a mono-phase molecular clock, regulates all the different phases of behavioral rhythms required for fitness of a species remains poorly understood (Liang, 2023).
Previously, it was reported that molecular clocks generate circadian neural activity rhythms with diverse phases among five major circadian neuron groups. Each group peaks at a specific time of day. Three laterally localized circadian neuron groups: s-LNv, l-LNv, and LNd display spontaneous activity peaks in the M, at MD, and in the E, respectively. Two dorsally localized circadian neuron groups the DN3 and DN1 cells are sequentially active during the nighttime (N1, CT18) and (N2, CT20). Genetic analyses revealed that the molecular clock dictates a default M phase onto the pacemakers, but the polyphasic activity pattern ensues due to the delaying activities of light and neuropeptide modulation within the circadian neuron circuit. This reproducible series of phasic activity periods displayed across the circadian pacemaker network may therefore represent dedicated phasic time points by which different downstream output circuits could achieve temporal order. This general problem—how output circuits are regulated by circadian pacemaker circuits—is a fundamental problem in the field of biological rhythms (Liang, 2023).
It was previously shown that the M and E phases (defined by activity peaks in the s-LNv and LNd/5th s-LNv neurons respectively), drive in biphasic activity patterns in the ring neurons of the ellipsoid body (RNs-EB) and in a subset of the PPM3 dopaminergic (DA) neurons.9 Both the RN-EB and PPM3 neurons are downstream neural circuits responding to clock signals to promote locomotor activity. Thus, the M and E phases of activity in the pacemaker circuit underlie authentic circadian phasic information that shapes premotor output to drive daily rhythmic locomotion (Liang, 2023).
This study asked whether other phasic activity periods presented by the pacemaker circuit-those of the MD (l-LNv group), the early night (N1, DN3 group), and the late night (N2, DN1 group) likewise direct daily rhythmic activity in downstream responsive non-pacemaker circuits. The problem could be approached by testing specific populations known to regulate different physiological and behavioral daily rhythms, as candidate responders of MD, N1, or N2 phasic information. For instance, a recent study10 revealed a candidate neural output circuit that mediates the N1 and N2 phasic information to promote nighttime sleep. However, more such candidate neural output circuits remain to be characterized. As an alternative approach, spontaneous daily activity patterns were measured in vivo across two populations of chemically defined neurons that are known to influence physiology and behavior—DA neurons and peptidergic neurosecretory (NS) neurons. Both populations exhibit daily periods of spontaneous activity whose phases align with either M, E, or MD phases of circadian pacemaker neurons. This study focused on MD phase regulation and present evidence from both experimental manipulations and from normal developmental progression to support the hypothesis that the MD activity phase is dictated by l-LNv pacemaker group activity. Together, these findings extend the hypothesis that polyphasic timing information from the Drosophila pacemaker circuit has broad functional significance. It spreads widely and independently through parallel downstream pathways to generate phase-diverse patterns of physiological and behavioral rhythms (Liang, 2023).
An organism's biological day is characterized by a pattern of anticipatory physiological and behavioral changes that are governed by circadian clocks to align with the 24-h cycling environment. This study used flash electroretinograms (ERGs) and steady-state visually evoked potentials (SSVEPs) to examine how visual responsiveness in wild-type Drosophila melanogaster and the circadian clock mutant ClkJrk varies over circadian time. The ERG parameters of wild-type flies vary over the circadian day, with a higher luminance response during the subjective night. The SSVEP response that assesses contrast sensitivity also showed a time-of-day dependence, including 2 prominent peaks within a 24-h period and a maximal response at the end of the subjective day, indicating a tradeoff between luminance and contrast sensitivity. Moreover, the behaviorally arrhythmic ClkJrk mutants maintained a circadian profile in both luminance and contrast sensitivity, but unlike the wild-types, which show bimodal profiles in their visual response, ClkJrk flies show a weakening of the bimodal character, with visual responsiveness tending to peak once a day. It is concluded that the ClkJrk mutation mainly affects 1 of 2 functionally coupled oscillators and that the visual system is partially separated from the locomotor circadian circuits that drive bouts of morning and evening activity. As light exposure is a major mechanism for entrainment, this work suggests that a detailed temporal analysis of electrophysiological responses is warranted to better identify the time window at which circadian rhythms are most receptive to light-induced phase shifting (Nippe, 2017).
Circadian clocks control daily rhythms in physiology, metabolism, and behavior in most organisms. Proteome-wide analysis of protein oscillations is still lacking in Drosophila. In this study, the total protein and phosphorylated protein in Drosophila heads in a 24-hour daily time-course were assayed by using the iTRAQ (Isobaric Tags for Relative and Absolute Quantitation) method, and 10 and 7 oscillating proteins as well as 19 and 22 oscillating phosphoproteins in the w(1118) wild type and Clk(Jrk) mutant strains were separately identified. Lastly, a mini screen was performed to investigate the functions of some oscillating proteins in circadian locomotion rhythms. This study provides the first proteomic profiling of diurnally oscillating proteins in fly heads, thereby providing a basis for further mechanistic studies of these proteins in circadian rhythm (Du, 2018).
The Drosophila circadian clock keeps time via transcriptional feedback loops. These feedback loops are initiated by CLOCK-CYCLE (CLK-CYC) heterodimers, which activate transcription of genes encoding the feedback repressors PERIOD and TIMELESS. Circadian clocks normally operate in approximately 150 brain pacemaker neurons and in many peripheral tissues in the head and body, but can also be induced by expressing CLK in nonclock cells. These ectopic clocks also require cyc, yet CYC expression is restricted to canonical clock cells despite evidence that cyc mRNA is widely expressed. This study shows that CLK binds to and stabilizes CYC in cell culture and in nonclock cells in vivo. Ectopic clocks also require the blue light photoreceptor CRYPTOCHROME (CRY), which is required for both light entrainment and clock function in peripheral tissues. These experiments define the genetic architecture required to initiate circadian clock function in Drosophila, reveal mechanisms governing circadian activator stability that are conserved in perhaps all eukaryotes, and suggest that Clk, cyc, and cry expression is sufficient to drive clock expression in naive cells (Liu, 2017).
In Drosophila, the clock that controls rest-activity rhythms synchronizes with light-dark cycles through either the blue-light sensitive cryptochrome (Cry) located in most clock neurons, or rhodopsin-expressing histaminergic photoreceptors. This study shows that, in the absence of Cry, each of the two histamine receptors Ort and HisCl1 contribute to entrain the clock whereas no entrainment occurs in the absence of the two receptors. In contrast to Ort, HisCl1 does not restore entrainment when expressed in the optic lobe interneurons. Indeed, HisCl1 is expressed in wild-type photoreceptors and entrainment is strongly impaired in flies with photoreceptors mutant for HisCl1. Rescuing HisCl1 expression in the Rh6-expressing photoreceptors restores entrainment but it does not in other photoreceptors, which send histaminergic inputs to Rh6-expressing photoreceptors. These results thus show that Rh6-expressing neurons contribute to circadian entrainment as both photoreceptors and interneurons, recalling the dual function of melanopsin-expressing ganglion cells in the mammalian retina (Alejevski, 2019).
The Drosophila sleep-wake rhythms are controlled by a brain circadian clock that includes about 150 clock neurons. Light synchronizes the clock neuronal network through cell-autonomous and non-cell-autonomous light input pathways. Cry is a blue-light sensitive photoreceptor protein that is expressed in most clock neurons. In the absence of Cry, flies do not phase-shift their behavioral rhythms in response to a short light pulse but still synchronize to light-dark (LD) cycles. Only flies devoid of both Cry and rhodopsin-expressing photoreceptors fail to entrain to LD cycles. Six different rhodopsins (Rhs) have been characterized in the Drosophila photoreceptive structures, which include the compound eye, the Hofbauer-Buchner (H-B) eyelet, and ocelli. The compound eye strongly contributes to circadian photoreception, whereas a modest contribution appears to be brought by the H-B eyelet and the ocelli. A circadian function has been recently associated with the yet poorly characterized rhodopsin 7, although its exact contribution and localization in the brain and/or the eye remains controversial. In addition to entrainment, the visual system controls other features of the clock neuron network by conveying light information to either promote or inhibit the behavioral output of specific clock neuron subsets (Alejevski, 2019).
The compound eye includes about 800-unit eyes (ommatidia), each of which contains eight photoreceptors. The six Rh1-expressing outer photoreceptors (R1-6) are involved in motion detection and project to the lamina neuropile of the optic lobe. The two inner photoreceptors (R7-8) are important for color detection and project to the medulla. They express four different rhodopsins and thus define two types of ommatidia: 'pale' (p) ommatidia (30%) include a Rh3-expressing R7 and a Rh5-expressing R8, whereas 'yellow' (y) ommatidia (70%) include a Rh4-expressing R7 and a Rh6-expressing R8. Each extra-retinal H-B eyelet contains four Rh6-expressing photoreceptors that project to the accessory medulla, in the vicinity of key pacemaker neurons, the ventral lateral neurons (LNvs) that produce the pigment-dispersing factor (PDF) neuropeptide9,20-24. Each of the three ocelli contains about 80 photoreceptors that express Rh225. The Drosophila rhodopsins cover a wide range of wavelengths from 300 nm to 600 nm18,19, with only Rh1 and Rh6 being sensitive to red light (Alejevski, 2019).
Rhodopsin-dependent circadian entrainment involves two downstream signaling pathways, the canonical one that relies on the phospholipase C encoded by the no receptor potential A gene (norpA)2 or an unknown pathway that does not contribute in very low light levels. All but Rh2- and Rh5- expressing photoreceptors support synchronization in very low light, and at least Rh1, Rh5, and Rh6 can signal through the NorpA-independent pathway. Photoreceptors of the compound eye are histaminergic but the H-B eyelet expresses both histamine and acetylcholine. Although the two neurotransmitters might contribute to circadian entrainment, flies devoid of Cry and histidine decarboxylase do not synchronize their rest-activity rhythms with LD cycles. This suggests that besides Cry, there is no histamine-independent pathway to entrain the clock (Alejevski, 2019).
Two genes encoding histamine-gated chloride channels, ora transientless (ort) and Histamine-gated chloride channel subunit 1 (HisCl1), have been identified in Drosophila. The ort-null mutants are visually blind and their electroretinograms have no ON and OFF transients. In contrast, HisCl1 mutants show increased OFF transients, whereas slower responses were observed in the postsynaptic laminar monopolar cells. Based on transcriptional reporters, ort expression in the optic lobes was observed in neurons of both the lamina and medulla/lobula neuropils. Based on reporter gene expression, HisCl1 was localized in glial cells of the lamina. However, recent work reported expression in photoreceptors, in particular in the R7 and R8 inner photoreceptor subtypes. Indeed, Ort and HisCl1 support color opponency between the two subtypes of 'inner' photoreceptors, the ultraviolet (UV)-sensitive R7 and non-UV-sensitive R8, with HisCl1 and Ort mediating direct and indirect inhibition, respectively. The histaminergic pathways that are involved in circadian entrainment are unknown and are the subject of the present study. The results show that both Ort and HisCl1 define two different pathways for circadian entrainment. Whereas Ort contributes through its expression in the interneurons of the optic lobe, HisCl1 mostly contributes through its expression in the Rh6-expressing retinal photoreceptors. The work thus reveals that Rh6-expressing neurons contribute to light-mediated entrainment as both photoreceptors and interneurons (Alejevski, 2019).
This work reveals that the Cryptochrome-independent entrainment of rest-activity rhythms relies on distinct pathways that are contributed by the two histamine receptors Ort and HisCl1. Whereas Ort mediates circadian entrainment through the optic lobe interneurons that are involved in visual functions, HisCl1 defines a new photoreceptive pathway through Rh6-expressing photoreceptors. Although both receptors mediate synchronization with a shifted LD cycle, it seems likely that the two pathways will show differences in specific light conditions. It was not possible to rescue Ort function with HisCl1 expression in the ort-expressing cells, whereas the Ort could replace HisCl1 in Rh6 photoreceptors. It is possible that HisCl1 has a lower affinity for histamine with Rh6 cells receiving more neurotransmitter than optic lobe interneurons. Alternatively, interneurons could sufficiently differ from photoreceptors for their physiology or specific receptor-interacting protein content, preventing HisCl1 from working efficiently. HisCl1 downregulation in Rh6 cells slows down synchronization and flies with HisCl1134 mutant eyes synchronize very poorly with advanced LD cycles, and fail to synchronize with delays. It cannot be excluded that non-photoreceptor cells contribute to HisCl1-dependent entrainment but other pathways appear to have a modest contribution if any (Alejevski, 2019).
HisCl1 is expressed in the H-B eyelet, which could thus contribute to this synchronization pathway. However, the cell-killing experiments indicate that H-B eyelet is not required for HisCl1-mediated synchronization through Rh6 cells. In the recently described color opponency mechanism, retinal R7 cells inhibit R8 and vice versa through HisCl1 expression in the photoreceptors (Schnaitmann, 2018). It is supposed that HisCl1-dependent clock synchronization is also mediated by the hyperpolarization of Rh6-expressing cells. How this hyperpolarization interacts with the light-induced depolarization in Rh6 photoreceptors to result in a synchronization message to the clock neurons remains to be understood. Since only Rh6-expressing R8 and not the other inner photoreceptors contribute to this circadian photoreception pathway, Rh6 cells might have specific connections with downstream interneurons. Such specificity has been described for color vision where each of the four inner photoreceptor subtypes connects to a different type of TmY interneuron in the Medulla. This study shows that HisCl1 expression in Rh6 cells supports synchronization with red light, in the absence of Rh1, indicating that an intra-Rh6-photoreceptor circuit is sufficient. This indicates that Rh6-expressing R8 photoreceptors play a dual photoreceptor/interneuron role in this pathway (Model for the retinal input pathways to the brain clock). Whether the same individual cells have the two roles is unknown, although the HisCl1-dependent color opponency mechanism suggests that it could be the case. It is also unclear whether all Rh6-expressing R8 photoreceptors or only a fraction of them contribute to circadian synchronization. The results imply that, in addition to histaminergic neurotransmission, Rh6-expressing photoreceptors can talk to downstream interneurons through histamine-independent neurotransmission. A recent transcriptomics study indeed revealed the expression of cholinergic markers in R7 and R8 cells, supporting cholinergic transmission in the inner photoreceptors, in addition to histaminergic transmission (Alejevski, 2019).
The data indicate that histaminergic inputs from both outer and inner photoreceptors converge to Rh6 cells to contribute to circadian entrainment. It is possible that some of these inputs rely on Rh7, which seems to be expressed in Rh6-expressing photoreceptors, according to transcriptional reporter data. Putative connections between photoreceptors have been described in Drosophila and other insects. How R1-6 photoreceptors might be connected to Rh6-expressing R8 cells remains difficult to understand, but a few putative contacts between presynaptic outer cells and postsynaptic inner cells have been observed in Musca. The intra-retinal functional connectivity that this study reports in Drosophila is reminiscent to the circuit logic of circadian entrainment in the mammalian retina, where intrinsically photoreceptive retinal ganglion cells express the melanopsin photopigment in addition to receiving inputs from rods and cones. Interestingly, melanopsin appears to share light-sensing properties with the rhabdomeric photoreceptors of invertebrates. It has been shown that the mammalian circadian clock can synchronize with day-night cycles by tracking light color changes in addition to light intensity changes. It will be interesting to investigate the possible contribution of the dual function of Rh6-expressing photoreceptors to integrating different color cues into the retinal information that is sent to the clock (Alejevski, 2019).
Coupling of autonomous cellular oscillators is an essential aspect of circadian clock function but little is known about its circuit requirements. Functional ablation of the pigment-dispersing factor-expressing lateral ventral subset (LNV) of Drosophila clock neurons abolishes circadian rhythms of locomotor activity. The hypothesis that LNVs synchronize oscillations in downstream clock neurons was tested by rendering the LNVs hyperexcitable via transgenic expression of a low activation threshold voltage-gated sodium channel. When the LNVs are made hyperexcitable, free-running behavioral rhythms decompose into multiple independent superimposed oscillations and the clock protein oscillations in the dorsal neuron 1 and 2 subgroups of clock neurons are phase-shifted. Thus, regulated electrical activity of the LNVs synchronize multiple oscillators in the fly circadian pacemaker circuit (Nitabach, 2006).
Understanding the mechanisms for synchronizing multiple independent neural oscillators in circadian circuits is a key issue in circadian biology. This study provides evidence that the excitability state of the LNV subset of clock neurons plays a critical role in coordinating multiple oscillators in the fly brain. When the LNVs are made electrically hyperexcitable by genetically targeted expression of a voltage-gated sodium channel cloned from a halophilic bacterium, NaChBac, transgenic flies exhibit complex free-running behavioral rhythms with multiple periods along with desynchronization of clock protein cycling throughout the pacemaker circuit and disrupted cycling of PDF levels in the dorsomedial terminal projections of the small LNVs (sLNVs) (Nitabach, 2006).
Anti-PDF immunofluorescence was observed in the dorsomedial terminals of the sLNVs in control flies. However, anti-PDF immunofluorescence in the dorsomedial terminals of the sLNVs of experimental flies expressing NaChBac in the LNVs is maintained at constitutively higher levels. This result is unexpected if PDF release at nerve terminals is the only cellular function influenced by alterations in cellular electrical excitability. Although there remains a formal possibility that NaChBac expression does not cause increased electrical excitability in pacemaker neurons, this is considered highly unlikely because of the robust and opposite effects of NaChBac expression compared with open-rectifier potassium-channel expression on behavior, reciprocal rescue of behavior by coexpression, clock oscillation, and direct electrophysiological recordings of muscle and photoreceptor neurons expressing NaChBac. Furthermore, hyperpolarization of LNv membrane potential after the targeted expression of open-rectifier potassium channels to these cells causes accumulations of PDF in the cell bodies of the LNVs, providing further evidence that membrane potential regulates the rates of synthesis and/or trafficking of PDF as well as release. These results together suggest that regulated electrical excitability of the sLNV plasma membrane underlies cycling PDF levels in the dorsomedial terminals, and that rendering the sLNVs hyperexcitable through NaChBac expression disrupts one or more of the cellular processes (synthesis, trafficking, or release) that determine PDF accumulation in the dorsomedial terminals. It remains unclear whether changes in neuronal membrane excitability directly influences PDF accumulation or whether this is caused by indirect effects via the molecular clock, because PDF accumulation appears to be restricted to pacemaker neurons (Nitabach, 2006).
The behavioral and circuit alterations caused by NaChBac expression in the LNVs may be attributable in part to an altered pattern of PDF release or a yet-unidentified neurotransmitter released by the LNVs, or to complex circuit properties of the pacemaker circuit. Regulated membrane electrical excitability of other neuropeptide-secreting neurons of the insect nervous system is known to be essential for appropriate control of the temporal patterns of peptide release. PDF may act as an intrinsic coupling signal within the circadian clock circuit that synchronizes multiple oscillators that otherwise free-run independently. This interpretation is consistent with a synchronizing role for PDF proposed on the basis of gradual phase dispersal within the sLNV subgroup of Pdf01-null mutant flies in constant darkness. In addition, the results are consistent with the idea that temporally regulated PDF release by the LNVs synchronizes the circuit, and are inconsistent with the hypothesis that PDF plays a purely permissive role (Nitabach, 2006).
Recent electrophysiological evidence in another insect suggests a mechanism for PDF- and GABA-mediated synchronization of multiple oscillators of pacemaker circuits (Schneider, 2005). Extracellular multiunit recordings of the candidate circadian neurons in excised preparations of the cockroach accessory medulla exhibit ultradian oscillatory action potential firing that is synchronized by local application of pressure ejected PDF and GABA through glass micropipettes or bath applied GABA (Schneider, 2005). Similarly, circadian neurons in the fly may fire in PDF-regulated assemblies. Although there is as yet insufficient electrophysiological evidence to allow direct comparison of the results in Drosophila with this recent finding in the cockroach, this raises the interesting possibility that NaChBac expression in the Drosophila LNVs may result in desynchronized firing of pacemaker neurons throughout the circuit, starting with the LNVs themselves. This would be consistent with the biophysical property of NaChBac of low-threshold voltage activation. Interestingly, similar mechanisms for oscillator coupling at the circuit level may also be important in mammals. GABA also modulates phase coupling between the ventral and dorsal oscillators in brain slices prepared from the rat SCN (Nitabach, 2006).
The behavioral results confirm that the Drosophila circadian control circuit contains multiple clocks capable of oscillating independently and capable of independently controlling the pattern, but not the amount, of locomotor activity. They further indicate that properly regulated electrical excitability of the LNVs (and perhaps of particular importance, the LNVs) is required to synchronize these multiple clocks throughout the pacemaker neuronal circuit. The synchronization of multiple oscillators appears to be necessary to generate coherent single-period behavioral rhythms (Nitabach, 2006).
The reciprocal suppression by NaChBac of the arrhythmicity induced by Kir2.1, and by Kir2.1 of the complex rhythmicity induced by NaChBac, strongly supports the interpretation that NaChBac and Kir2.1 have opposite effects on the electrical excitability of the LNVs, with Kir2.1 decreasing excitability and NaChBac increasing excitability. When expressed individually in the LNVs, K+ channels and Na+ channels have opposite behavioral effects: hyperpolarizing K+-channel expression results in arrhythmic behavior, whereas depolarizing Na+-channel expression results in hyper-rhythmic behavior. The coexpression of these two channel types together results in functional reciprocal compensation, yielding nearly normal behavior (Nitabach, 2006).
In a previous studies, LNV membrane potential was manipulated to be hypoexcitable through the targeted expression of modified open-rectifier or inward-rectifier potassium channels (Nitabach, 2002). This caused behavioral arrhythmicity and cell autonomous dampening of the free-running molecular clock in LNV neurons in constant darkness, along with no discernable changes in the cycling of the molecular clock in downstream pacemaker neuronal subgroups at circadian day 2. Those results are consistent with the findings that desynchrony of downstream cell groups does not become apparent in pdf01-null mutant flies until 2 d in constant darkness. In the present study, LNV hyperexcitability induces trans-synaptic changes in the free-running temporal pattern of clock protein accumulation in the dorsal neuron subgroups DN1 and DN2. Thus, the DN neuronal groups appear to be functionally downstream of the LNV neurons in the pacemaker circuit. In negative control flies, the DN1s oscillate in phase with the sLNVs and LNDs, maintaining synchrony on both days 2 and 5 after release into constant darkness from a diurnal 12 h light/dark entraining regime, whereas the DN2s gradually advance from synchrony in 12 h light/dark to a 12 h phase difference by circadian day 5. The DN2s of control flies exhibit peak PDP1 accumulation at CT14 on day 2 in constant darkness and at CT10-CT14 on day 5 in constant darkness. This gradual shift of DN2 PDP1 oscillation from synchrony with the other cell groups in LD to a 12 h phase advance after 5 d in constant darkness is consistent with observations of DN2 PER cycling. In pdf>NaChBac1 flies expressing NaChBac in the LNVs, the DN1s exhibit a PDP1 molecular peak 8 h earlier than control flies on day 2 in constant darkness, and by circadian day 5 this peak has significantly damped and an additional significant peak has appeared at CT22. The DN2s of pdf>NaChBac1 flies exhibit a peak of PDP1 accumulation at CT14 on day 2 in constant darkness, in phase with control flies; by day 5 in constant darkness they peak at CT6, 48 h earlier than in controls. This phase shift suggests that the DN2 molecular oscillator of pdf>NaChBac1 flies is running faster than that of control flies. These differences in the temporal pattern of PDP1 accumulation in the DN1s and DN2s induced by NaChBac expression in the LNVs indicate that properly regulated electrical activity is required for normal patterns of molecular oscillation in these dorsal cell groups (Nitabach, 2006).
The DN2s may be capable of independently driving behavioral outputs, and are possibly the cellular substrate for the ~22 h short-period component of the complex behavioral rhythmicity exhibited by flies expressing NaChBac in the LNVs. The cellular substrates for the ~25.5 h long-period component are likely to reside in other cells within the circuit. In control pdf>TM3 flies, robust free-running PER oscillation is observed in the sLNV,LND, and DN1 neurons after 5 d in constant darkness, with trough levels of PER in the second half of subjective day. The differences in the spatiotemporal pattern of PER accumulation induced by NaCh-Bac expression in the LNVs confirm, as indicated by the effects on PDP1 accumulation, that hyperexcitation of electrical activity in the LNVs causes desynchronization of the coupling and phase of molecular oscillation in dorsal clock neurons (Nitabach, 2006).
Multiple oscillators are distributed throughout the pacemaker circuit in Drosophila. The present study confirms and extends evidence for multiple oscillators in the pacemaker circuit in Drosophila. The independent oscillators driving the multiple period components of the behavioral rhythms that were observed do not appear to correspond directly to the 'morning' and 'evening' oscillators, which have been localized to the LNVs and LNDs, respectively. The current results emphasize that the activity of the LNVs controls the synchronization of independent oscillators throughout the pacemaker circuit. The normal pattern of DN1 and DN2 clock oscillation requires properly regulated electrical excitability of the LNVs. Further, the results suggest that the DN2s, and at least some other cell groups, possess independent output pathways to the downstream locomotor circuitry (Nitabach, 2006).
This study introduces a novel method for inducing electrical hyperexcitability in neurons of interest by the expression of the low-threshold voltage-gated sodium channel NaChBac. This method is likely to be useful for the analysis of other neural circuits. In another study (Luan, 2006), the utility of the NaChBac channel for enhancing excitability in other neurons has also been demonstrated. Targeted expression of ion channel subunits in vivo provides a powerful means for precisely perturbing neuronal membrane excitability to probe the role of activity on neuronal development and function. Initial methods to exogenously regulate electrical excitability in neurons in vivo have used potassium channel expression to electrically silence neurons. Exogenous manipulation of electrical excitability within specific Drosophila neurons can be combined with finer parsing of neural circuits using GAL80 and other genetic approaches (Nitabach, 2006).
This study has shown that aberrations of electrical excitability in Drosophila neurons, either hyperexcitability induced by NaChBac or hypoexcitability induced by Kir2.1, can be rescued by coexpression of an ion channel with an opposite effect on excitability. This provides reason to believe that such an approach to neurological disorders of aberrant electrical activity such as epilepsy might indeed be feasible (Nitabach, 2006).
The molecular mechanisms of circadian rhythms are well known, but how multiple clocks within one organism generate a structured rhythmic output remains a mystery. Many animals show bimodal activity rhythms with morning (M) and evening (E) activity bouts. One long-standing model assumes that two mutually coupled oscillators underlie these bouts and show different sensitivities to light. Three groups of lateral neurons (LN) and three groups of dorsal neurons govern behavioral rhythmicity of Drosophila. Recent data suggest that two groups of the LN (the ventral subset of the small LN cells and the dorsal subset of LN cells) are plausible candidates for the M and E oscillator, respectively. Evidence is provided that these neuronal groups respond differently to light and can be completely desynchronized from one another by constant light, leading to two activity components that free-run with different periods. As expected, a long-period component starts from the E activity bout. However, a short-period component originates not exclusively from the morning peak but more prominently from the evening peak. This reveals an interesting deviation from the original Pittendrigh and Daan (1976) model and suggests that a subgroup of the ventral subset of the small LN acts as 'main' oscillator controlling M and E activity bouts in Drosophila (Rieger, 2006).
Daily biological rhythms are governed by inherent timekeeping mechanisms, called circadian clocks. Such clocks reside in discrete sites of the brain and consist of multiple autonomous single-cell oscillators. Within each neuron, interacting transcriptional and translational molecular feedback loops as well as ionic signaling pathways constitute the oscillatory mechanism of the clock. It is not understood how individual pacemaker neurons interact to drive behavioral rhythmicity. The long-standing model of Pittendrigh and Daan (1976) assumes that the clock consists of two groups of oscillators with different responsiveness to light, one governing the morning (M) and the other the evening (E) activity of the animal. Typical M and E activity bouts are present in animals ranging from insects to mammals and suggest that the two-oscillatory model is generally valid. It has been shown that M and E bouts could be eliminated or reinstated by manipulating different circadian pacemaker neurons in Drosophila. This work has suggested that the ventral (LNv) and dorsal (LNd) subsets of the lateral neurons are the neuronal substrates for the M and E oscillators. It is not known whether these two oscillators respond differently to light (Rieger, 2006).
The particular power of the two-oscillator model is that it explains observed adaptations to seasonal changes in day length. The model predicts that the M oscillator will shorten and the E oscillator will lengthen its period when exposed to extended constant light (LL). As a consequence, the M activity occurs earlier and the E activity occurs later in long summer days, helping day-active animals avoid the midday heat. The model also predicts that the M oscillator will free-run with short period and the E oscillator with long period when animals are placed in constant light. However, such internal desynchronization between oscillators does not occur, because high-intensity constant light usually results in arrhythmicity. In Drosophila, the clock protein Timeless (TIM) is permanently degraded during light-induced interaction with Cryptochrome (CRY), leading finally to the arrest of the clock. Without functional CRY, this does not happen. Indeed, internal desynchronization into two free-running components (one with a short period and the other with a long period) has been described for cryb mutants under constant-light conditions. The present study aims to analyze the molecular state of all clock gene-expressing neurons during behavioral rhythm dissociation to test the PittendrighDaan model and refine the neuronal substrates of the E and M oscillators (Rieger, 2006).
This study supports the notion that the activity rhythm of Drosophila is controlled by at least two sets of neuronal oscillators. Furthermore, the definition of these neuronal substrates of both oscillators were refined more precisely than previously. As proposed by Pittendrigh and Daan (1976), the two oscillators show different responses to light: one is accelerated and the other decelerated by constant light. However, a deviation from the original model was observed. In contrast to previous observations, the current results suggest that the PDF-positive s-LNv cells control not only the M but also the E activity bout. Therefore, the discussion should perhaps not focus of a 'morning' oscillator but rather of an ME or 'main' oscillator (to keep the 'M'), for the following reasons. The PDF-positive s-LNv cells are essential for maintaining activity rhythms after several days under constant conditions, and electrical silencing of the LNv cells severely impairs free-running rhythms. In the present study, the PDF-positive s-LNv cells appear to dominate the rhythms in those flies that did couple E and M components after the first crossing-over on day 11 in LL, because such flies free-ran with short period (Rieger, 2006).
The hypothesis that the PDF-positive LNv cells control not only the M activity but also partly the E activity can also explain other findings. The E activity bout is always the most prominent peak, which persists under constant-dark conditions, whereas the M activity bout is much reduced under such conditions and may even disappear. Thus, mainly the E component constitutes the free-running rhythm, and it seems implausible that the neurons responsible for rhythmicity under these conditions should have no impact on the E component. Indeed, it has been found that the s-LNv show the most robust cycling after extended time under constant conditions. Furthermore, another study emphasizes the importance of the s-LNv cells for the timing of activity peaks under constant conditions (Rieger, 2006 and references therein).
Despite their dominance, the PDF-positive s-LNv cells depend on functional LNd and DN cells to provoke a normal E activity bout under light-dark conditions. Flies with the clock gene PER present only in PDF-positive LNv cells have a prominent M activity bout but lack the E activity bout. It is unclear whether this is attributable to the E activity fusing with the M activity or whether the E activity is suppressed, but these findings show that the output from the PDF cells requires PER in the LNd and DN cells to manifest wild-type activity patterns (Rieger, 2006).
It was found that the PDF-negative 5th s-LNv cell cycles in-phase with the LNd cells under LL and thus may contribute to the E component. Notably, the PDF-negative 5th s-LNv cell shows high-amplitude cycling. Although this is not proof of the involvement of this cell, it suggests that it is an important circadian pacemaker neuron. Little is known about this cell because it could not be distinguished from the other lateral neurons in the former studies in which single-labeled clock protein staining was performed, but the PDF-negative 5th s-LNv cell is the only clock cell beside the PDF-positive s-LNv cells that appears to work from the first larval instar onward. Thus, it might have the same strong impact on the activity rhythm that has been revealed for the PDF-positive s-LNv cells. More work is necessary to reveal the role of the PDF-negative s-LNv cell in more detail (Rieger, 2006).
Additional studies are also necessary to fully reveal the function of the DN cells. The current results suggest that the DN1 and the DN3 cells may contain different subclusters. Indeed, the DN1 cells develop at different times and appear to have distinct projection patterns. It is very likely that some DN1 cells contribute to the M oscillator whereas others supply the E oscillator. Again, there are data that support this hypothesis: if the lateral neurons (s-LNv, l-LNv, and LNd) are absent as a result of mutation or genetic ablation but the dorsal neurons (DN1, DN2, and DN3) are left intact, morning and evening activity bouts are still present under LD conditions, although with reduced amplitude and changed phase. The DN2 cells might play a special role for bimodal activity patterns because, in wild-type flies, they cycle 12 h out-of-phase with the s-LNv and LNd cells under DD conditions. The present study indicates that this is not the case in cryb flies under LL conditions, because the DN2 cells were in-phase with all other neurons on the first day in LL. The same applies for wild-type flies under LD conditions. It has been shown that the DN2 are indeed pacemaker neurons that cycle independently of the s-LNv cells. However, despite their autonomous function, the DN2 cells did not visibly contribute to the activity patterns of the flies under constant darkness. This suggests a minor role of the DN2 cells in the control of the activity rhythm, but the possiblity cannot be exclude that the DN2, together with the other DN groups, may contribute to morning and evening activity bouts under certain conditions (Rieger, 2006).
The blue-light photopigment cryptochrome is regarded as the main photoreceptor of the fruit flies' circadian clock. This study shows that the compound eyes are responsible for period shortening and period lengthening of the molecular oscillations in different subsets of pacemaker neurons (the M and E oscillators) under LL. Their special role may lie in the adaptation of the clock to seasonal changes. This is in line with previous findings showing that the compound eyes are necessary for the adequate timing of M and E activity bouts in long summer days and short winter days. Cryptochrome, conversely, appears to lengthen the period in all clock neurons as can be deduced from the periods of the wild-type flies that showed internal desynchronization under 'moonlight LL.' In such flies, the periods of both components were clearly longer than those of internally desynchronized cryb flies (Rieger, 2006).
The internal desynchronization of activity into long- and short-period components described in this study is reminiscent of previous results for Drosophila mutants with primarily reduced optic lobes or ectopic expression of PDF. Both of these fly strains have ectopic PDF-containing nerve fibers in the dorsal brain that might lead to elevated and/or nonrhythmic secretion of PDF in this brain area and may disturb normal communication between the pacemaker cells. It is unknown whether such a perturbed communication results in internal desynchronization between the s-LNv and the 5th s-LNv and extra LNd as observed in the present study. Dual-oscillator systems have been also described for mammals, but in no case they could be traced to the level of single neurons. Like the circadian pacemaker center of flies, the mammalian pacemaker center, the suprachiasmatic nucleus (SCN), contains a heterogeneous neuronal population. A recent study has shown that internal desynchronization of motor activity into short and long periods similar to the one shown in this study can be provoked in rats by special light schedules. As in Drosophila, both components reflect the separate activities of two oscillators in anatomically defined subdivisions of the SCN. Furthermore, there is some evidence to suggest that the SCN is composed of two oscillating M and E components. These results underline the universality of dual-oscillator systems (Rieger, 2006).
Other studies strongly implicate the PDF-expressing LNv and the LNd cells as the respective neuronal loci for the morning and evening activity bouts. Despite the near 12 h phase difference between the morning and evening locomotor peaks under LD, no obvious molecular phase differences between these pacemakers have been observed that would explain them. Work in mammals suggests that the relationship between molecular phase and locomotion is complex. For example, nocturnal and diurnal rodents show the same phases of PER oscillations. Furthermore, different rat strains that displayed unimodal or multimodal activity patterns, respectively, all exhibited the same unimodal rhythm in melatonin synthesis. Individual Nile grass rats changed their activity patterns from unimodaldiurnal to bimodalnocturnal after introducing a running wheel. Despite this dramatic effect on the activity patterns, the wheel had little effect on the circadian pacemaker, and the spatial and temporal patterns of c-Fos expression in the SCN remained similar. All of these data indicate that the relationship between molecular and behavioral phase is not straightforward. Clearly, a multitude of phase relationships between the molecular rhythm and behavior are possible. Brain regions outside the pacemaker center may be responsible for these different phases as was shown recently for mammals. It appears that the same is true within the circadian system of the fly. The present data show that, during the internally synchronized state, the trough in PER level of all neurons correlates with the main activity bout (the E peak). No second trough appears to correlate with the M peak. However, a second small peak can be seen at closer inspection of the PDF immunoreactivity in the terminals of the s-LNv. This suggests that the unimodal rhythm in clock protein cycling might be converted into a bimodal output already within the neurons (Rieger, 2006).
During the state of behavioral desynchronization under LL conditions, an internal desynchronization was observed simultaneously in PER oscillations among subsets of pacemaker neurons. One interpretation of these data is that constant light causes internal desynchronization between these pacemaker neurons that then in turn drive the behavioral outputs. However, it must be acknowledged that this is only a correlation, and, although the hypothesis is favored that the split molecular rhythms are driving the split locomotor rhythms, it is possible that they are merely tracking or entraining to a split rhythm driven by other pacemakers. For example, the split rhythms might be driven by subsets of dorsal neurons. The hypothesis is preferred that the split behavioral rhythms were driven by the desynchronized PDF-positive LNv and the 5th s-LNv/extra LNd cells for two reasons. First, accumulating evidence points to the lateral neurons (LNv and LNd cells) as major pacemaker cells, whereas the dorsal neurons (the DN1, DN2, and DN3 cells) are not sufficient for locomotor rhythms under constant darkness. Second, in rodents, a similar behavioral desynchronization was correlated with a dissociation of clock gene expression between ventrolateral and dorsomedial subdivisions of the SCN. The established role of this brain center as the circadian clock has led to the uncontroversial conclusion that the split molecular oscillations drive the split behavioral oscillations. It is suggested that the same phenomenon is occurring in main (i.e., small LNv cells) and evening (i.e., 5th s-LNv and extra LNd cells) neuronal oscillators in Drosophila (Rieger, 2006).
Work in Drosophila has defined two populations of circadian brain neurons, morning cells (M-cells) and evening cells (E-cells), both of which keep circadian time and regulate morning and evening activity, respectively. It has long been speculated that a multiple oscillator circadian network in animals underlies the behavioral and physiological pattern variability caused by seasonal fluctuations of photoperiod. This study manipulated separately the circadian photoentrainment pathway within E- and M-cells and shows that E-cells process light information and function as master clocks in the presence of light. M-cells in contrast need darkness to cycle autonomously and dominate the network. The results indicate that the network switches control between these two centers as a function of photoperiod. Together with the different entraining properties of the two clock centers, the results suggest that the functional organization of the network underlies the behavioral adjustment to variations in daylength and season (Stoleru, 2007).
Two populations of circadian brain neurons, morning cells (M-cells) and evening cells (E-cells), have been connected to morning and evening locomotor activity, respectively (Grima, 2004; Stoleru, 2004). Interactions between the two oscillator populations were studied by selectively overexpressing sgg to speed up the clock in only one cell population or the other (Stoleru, 2005). This study has found that sgg overexpression gives rise to LL rhythmicity, which led to a search for the cellular substrates of entrainment. The rhythmicity is predominantly due to sgg overexpression in E-cells, which suggested that this subset of the clock network is particularly important in the light and that Sgg affects the biochemical pathway through which light impacts clock molecules and adjusts phase to the correct time of day. Indeed, strong evidence is presented that Sgg modulates Cry function, which affects in turn the core clock proteins Per and Tim. The separate manipulation of the Sgg/Cry pathway within E- and M-cells also reveals that the E-clocks drive the behavioral rhythm in light, with prominent Per oscillations of nuclear localization. This light dependence of E-cells contrasts with M-cells, which need darkness to cycle autonomously and dominate the activity output pathway. This distinction suggests a simple dual-oscillator model for how the clock adjusts to photoperiod changes, and support for this seasonal model was obtained by examining E- and M-cell cooperation under different photoperiods (Stoleru, 2007).
The free-running pacemaker and entrainment are two important and increasingly understood aspects of circadian rhythms. In contrast, little information exists about seasonal adjustment, namely, how a constant ~24-hr timekeeper accommodates dramatically different photoperiods. This study shows that the previously defined dual oscillator system in Drosophila, M-cells and E-cells, creates different rhythmic patterns by alternating master clock roles. This understanding emerged from restricting Sgg overexpression to E-cells, which allowed the E-oscillator to function and render flies rhythmic in LL. Sgg probably modulates Cry activity and, when overexpressed, provides sufficient Per and Tim to allow E-oscillator function under constant illumination conditions. The E-clocks therefore manifest free-running properties and function as the master pacemakers in LL, analogous to a previous finding that the M-oscillator is the master in DD (Stoleru, 2005). Nonetheless, these constant conditions, and even the perfect standard LD cycles commonly used in the laboratory, are poor approximations of the changing LD environments found in nature. Circadian oscillators and their entrainment mechanisms have adapted to the dramatic seasonal changes in photoperiod. The previous strategy of using oscillators with different speeds, combined with different photoperiods, has led to a model of alternating control between the M-oscillator and E-oscillator (Stoleru, 2007).
Sgg appears to attenuate, rather than inactivate, Cry activity in E-cells. This is because the LL period of timSgg/PdfGAL80 (~23.5 hr) is longer than the intrinsic period of Sgg-expressing E-clocks in DD (~21 hr) (Stoleru, 2005). A longer period in light is compatible with attenuated light perception under high light intensity conditions (1600 lx, which renders wild-type flies completely arrhythmic) and the application of Aschoff's rule to insects [Aschoff, 1979; One of the earliest observations in the study of circadian rhythms was that continuous light (LL) lengthens circadian period in most nocturnal animal species. 'Aschoff's Rule' posits that there is a positive log-linear relationship between the LL intensity and period]. As there is also a prominent effect on Cry stability, Sgg may be the regulator previously predicted to bind to the Cry C terminus (Busza, 2004; Dissel, 2004). Although Cry is favored as the major circadian substrate of Sgg, there may be others, e.g., the serotonin receptor. Biochemical support for GSK3 involvement in mammalian rhythms has recently been obtained (Yin, 2006). Since GSK3 is a proposed therapeutic target of lithium, the relationship between Sgg and Cry reported in this study recalls the intriguing relationship between mood disorders, light sensitivity, and circadian rhythms (Stoleru, 2007).
The cryb genotype markedly affects DD period in some of the rhythmic genotypes described in this study. Although Cry is probably unnecessary for M-cell rhythmicity, this could reflect some redundancy or assay insensitivity. Moreover, the DD period of cryb is slightly shorter than that of wild-type (23.7 versus 24.4), suggesting that 'dark Cry' makes some contribution to pacemaker function in M-cells as well as E-cells. For these reasons, it is suggested that Drosophila Cry is closer to the central pacemaker than previously believed, and therefore closer to the level of importance of its mammalian paralogs in influencing free-running pacemaker activity. Unlike mammalian Cry, however, Drosophila Cry still appears to function predominantly at a posttranslational level. Indeed, the effects of cryb on Sgg overexpression in DD suggest that the proposed effect of Sgg on Tim stability is really an effect of Sgg on Cry followed by an altered Cry-Tim interaction. It is noted that there is a recent proposal (Collins, 2006) that Drosophila Cry, like mammalian Cry, also functions as a transcription factor in peripheral clocks (Stoleru, 2007).
The importance of E-cells in LL rhythmicity is underscored by the staining results of timSgg/PdfGAL80 brains. Only some E-cells and DN2s manifest robust cycling. It has been suspected that E-cells are important in light because they can rescue the output of arrhythmic M-cells in LD, but not in DD (Stoleru, 2004). Indeed, all of these observations make it attractive to view E-cells as autonomous pacemakers. There is, however, evidence that M-cells may not be completely dispensable. Moreover, a synchronizing or stabilization function is compatible with previous observations under different conditions (Stoleru, 2007).
In the timSgg/PdfGAL80 genotype, only Per nuclear localization changes were detectable near the end of LL cycle. The nature of the assay makes it hard to conclude that there were no differences in total Per staining intensity, i.e., no oscillations in Per levels, so the unique nature of the Per nuclear localization cycling is a tentative conclusion. The same caveat applies to the absence of Tim oscillations and nuclear staining, i.e., negative results cannot exclude low-amplitude oscillations; it is noted, however, that Tim cytoplasmic sequestration has been previously observed in cryb flies after several days in LL. Furthermore, the circadian nuclear accumulation of Tim has been shown to respond differently than that of Per to changes in photoperiod. Nonetheless, Tim could be shuttling with a predominant steady-state cytoplasmic localization, nuclear Tim could be rapidly degraded to create a low nuclear pool, or both (Stoleru, 2007).
The importance of E-cells in entrainment is strongly supported by the potent effect of restricted Cry rescue of cryb: E-cell rescue is much more impressive than M-cell rescue. Moreover, the differences between the two rescued phase response curves (PRCs) are striking; E-cell rescue is virtually complete, whereas the M-cell rescue is notably deficient in the delay zone. In addition, flies with Sgg overexpression in E-cells show altered PRCs, whereas flies with Sgg overexpression in M-cells respond normally to light. The results are strikingly different in darkness, as M-cell-restricted expression causes the typical short period determined by Sgg overexpression, whereas E-cell overexpression has no systemic effect (Stoleru, 2007).
The PRC delay zone is the region impacted most strongly by E-cell Sgg overexpression, indicating that the lights-off early night region is most important to E-cell function and light entrainment. Exposure to light in this interval should mimic long days (summer), which, it is speculated, will delay phase by many hours so that evening output of the following day will coincide with the objective evening of the environment. Even the short nights of summer are probably enough time for E-clocks to accumulate sufficient Tim and Per, shuttle them into the nucleus, and reconstitute the rhythmic substrate observed in the Sgg-overexpressing brains in LL. In contrast, M-cells need darkness to cycle robustly. They will become the master clocks and drive the system whenever lights fail to turn on more than 12 hr past lights-off, i.e., during the long nights of winter that mimic the beginning of a DD cycle. Since the intrinsic pacemaker program of M-cells in darkness relies on the changing nature of clock proteins during the night, it is hypothesized that the activity phases under long nights (winter) are locked to lights-off. This suggestion is supported by preliminary data and previous observations showing that per transcription remains locked to lights-off under different entrainment regimes. M-cells are also capable of fully entraining the system in the PRC interval that determines a phase advance (late night). This is consistent with their predicted role in generating an advanced evening output, coincident with the early evenings typical of winter. Otherwise put, long summer days should underlie light primacy as well as long and prominent evening delay zones; both suggest E-cell dominance. Night primacy and M-cells should dominate under winter conditions. This concept endows E- and M-cells with the properties originally envisioned by the Pittendrigh and Daan (1976) dual-oscillator model of entrainment (Stoleru, 2007).
The ability to be synchronized by light-dark cycles is a fundamental property of circadian clocks. Although there are indications that circadian clocks are extremely light-sensitive and that they can be set by the low irradiances that occur at dawn and dusk, this has not been shown on the cellular level. This study demonstrates that a subset of Drosophila's pacemaker neurons responds to nocturnal dim light. At a nighttime illumination comparable to quarter-moonlight intensity, the flies increase activity levels and shift their typical morning and evening activity peaks into the night. In parallel, clock protein levels are reduced, and clock protein rhythms shift in opposed direction in subsets of the previously identified morning and evening pacemaker cells. No effect was observed on the peripheral clock in the eye. These results demonstrate that the neurons driving rhythmic behavior are extremely light-sensitive and capable of shifting activity in response to the very low light intensities that regularly occur in nature. This sensitivity may be instrumental in adaptation to different photoperiods. This adaptation depends on retinal input but is independent of cryptochrome (Bachleitner, 2007).
Nocturnal light provoked an advance of the morning (M) activity and a delay of the evening (E) activity into the night. Simultaneously, the midday trough broadened and the midnight trough diminished, making the flies nocturnal in a cycle of 12 h:12 h. In other words, they switched their temporal niche. Upon transfer to constant conditions, they reverted, with activity in continuous moonlight (MM) always starting from the preceding light phase. A similar switch was observed in night-active white-fronted lemurs (Eulemur fulvus albifrons). These animals switched from night-active to day-active after reduction of the nocturnal illumination below a certain threshold; but on release into constant conditions, free-running activity always started from the preceding dark phase. This switching was thought to be caused by direct effects of light on activity (masking effects) that do not interfere with the circadian clock. In other words, the animals have strong preferences for certain light conditions, and they accordingly avoid being active under both total darkness, because it precludes visual orientation, and in high irradiances, because it may damage their sensitive eyes. Studies on mice (Mus musculus) yield similar results, with nocturnal animals becoming diurnal after mutations, genetic manipulations, or brain lesions that interfere with photoreceptor input to the circadian clock. One possible explanation is that mice with impaired photoreception simply prefer higher irradiances than wild-type mice (Bachleitner, 2007).
This study demonstrates that temporal niche switching may be caused by a change in the phasing of the endogenous clock as induced by dim light levels. Masking effects may still be involved, because the masking that is typically seen in conventional light-dark (LD) is lost under light-'moonlight' (LM) conditions. Instead, the flies' activity appears to be promoted by dim light. Despite a contribution of masking effects, this study shows that Drosophila's circadian pacemaker neurons are highly light-sensitive and respond to nocturnal light levels comparable to moonlight. The peripheral oscillators in the eye fail to do so. This finding is extremely interesting with respect to photoreceptor sensitivity. Whereas photoreceptors for visual image detection should quickly adapt to alterations in light intensity to ensure optimal vision, photoreceptors for the circadian clock should not adapt, at least not in the lower ranges. Otherwise, it would be impossible to measure increasing and decreasing irradiances during dawn and dusk. The observed clock protein oscillations in the photoreceptor cells of the compound eyes might regulate light sensitivity of the circadian system. If true, the clock protein levels and oscillations should not be altered by dim light, and the high sensitivity of the clock during early dawn and late dusk should be preserved. Indeed, no alterations were observed in PER and TIM protein levels under LM conditions. Consistent with this finding, it was found that the compound eyes, not the photoreceptor cryptochrome, mediate the responses to moonlight. This finding is in line with previous observations showing that the compound eyes and, thus, rhodopsins are necessary for adaptation of activity times to long and short days. Interestingly, the insect rhodopsins are closely related to the mammalian melanopsin, which is critically involved in mouse circadian photoreception (Bachleitner, 2007).
But then, what role remains for cryptochrome? Cryptochrome may contribute to the phase delay of the E peak under LM conditions, because cryb mutants show a less dramatic delay of this peak than wild-type flies. However, the E peak was not at all delayed in clieya mutants, suggesting that a phase-delaying effect of cryptochrome is either dependent on the compound eyes or is negligible. Cryptochrome is a blue-light photoreceptor and, thus, most appropriate to detect intensity changes in blue light. Furthermore, the proportion of blue light increases during dawn and decreases during dusk. Thus, cryptochrome appears particularly suited to distinguish dawn or dusk light transitions from moonlight, which does not change its spectrum over time (Bachleitner, 2007).
These results are interpreted as evidence for a differential action of dim light on the pace of M and E oscillators as was originally proposed in the dual-oscillator model for rodents and recently verified for D. melanogaster. Because the M cells phase advance and a subset of the E cells phase delay their clock in response to dim light, these cells are optimally suited to adapt activity rhythms to seasonal changes in day length. In addition, the proposed role of the LNd cells as E cells is not supported by this study. This finding is in line with earlier work demonstrating that the LNd represent a heterogenous group of cells and that, putatively, only one LNd cell behaves as an E oscillator. It is quite possible that this LNd cell also phase delayed in the present study; but without a specific marker, it was not possible to distinguish it from the other cells. In summary, these results are consistent with the involvement of the PDF-positive s-LNv cells and the PDF-negative fifth s-LNv cells on behavioral rhythmicity. The relevance of the two-oscillator model under natural conditions is also shown (Bachleitner, 2007).
Not all animals may be as light-sensitive as are fruit flies, but recent studies showed that even species such as hamsters and humans are more sensitive than supposed. The synchronization of Syrian and Siberian hamsters to different photoperiods was facilitated under dim night illumination (<0.005 lux) as compared with DD. Furthermore, the incidence of bimodal activity patterns and the interval between both components increased under LM conditions. In humans, dawn simulations at low light intensities were found to phase advance the circadian melatonin and the activity rhythm. Together with the results presented here, these studies suggest that clock function in many species is conspicuously altered by nocturnal illumination as experienced under dim moonlight. This finding might go back to the ability of primordial marine animals to synchronize their reproduction to the lunar cycle, an ability that is apparently lost in humans and other terrestrial animals. Presumably, many terrestrial organisms do not use their light sensitivity for moonlight detection, but for timing their clock to the increasing and decreasing irradiances during dusk and dawn. Further studies are necessary to reveal whether these animals hide at night from the moonlight so as not to confound their clocks, whether they switch to nocturnal activity (or become sleepless) during the full moon, or whether they use cryptochrome to distinguish moonlight from dawn and dusk (Bachleitner, 2007).
Large ventral lateral clock neurons (lLNvs) exhibit higher daytime-light-driven spontaneous action-potential firing rates in Drosophila, coinciding with wakefulness and locomotor-activity behavior. To determine whether the lLNvs are involved in arousal and sleep/wake behavior, the effects of altered electrical excitation of the LNvs were examined. LNv-hyperexcited flies reverse the normal day-night firing pattern, showing higher lLNv firing rates at night and pigment-dispersing-factor-mediated enhancement of nocturnal locomotor-activity behavior and reduced quantity and quality of sleep. lLNv hyperexcitation impairs sensory arousal, as shown by physiological and behavioral assays. lLNv-hyperexcited flies lacking sLNvs exhibit robust hyperexcitation-induced increases in nocturnal behavior, suggesting that the sLNvs are not essential for mediation of arousal. It is concluded that light-activated lLNvs modulate behavioral arousal and sleep in Drosophila (Sheeba, 2008b).
The small and large ventral lateral clock neurons (henceforth sLNvs and lLNvs) were among the first cells identified as crucial for normal light entrainment of circadian behavior. Several studies suggest that the sLNvs are responsible for sustained circadian locomotor activity in constant darkness, whereas the lLNvs are less characterized. Recently, it was shown by using whole-cell patch clamp electrophysiology that lLNvs acutely increase their firing rate in response to light in a cryptochrome-dependent fashion (Sheeba, 2008a). Because light is a well-known sensory cue for arousal, as well as circadian entrainment, this study tested whether altered electrical activity of the lLNvs influences locomotor-activity behavior, sleep, and arousal (Sheeba, 2008b).
These studies show that alteration of the balance of day-night neuronal firing by hyperexcitation of lLNvs in D. melanogaster directs behavioral-activity preference toward increased nocturnality and modulates the quantity and quality of nocturnal sleep. Furthermore, other peptidergic neurons encompassed within the c929 expression pattern modulate both day and night wakefulness and sleep. These results, in combination with an earlier detailed electrophysiological analysis (Sheeba, 2008a), suggest that lLNvs constitute a light-activated arousal circuit. Additional support for this is shown by decreased behavioral responsiveness to day onset in PDF-lacking flies and flies with electrically altered LNvs. It has been shown previously that plasticity in temporal day versus night behavioral preference in mammals and other animals can result from changes in environmental sensory time cues or manipulations that alter sensory-input pathways to circadian clocks. In Drosophila, mutants with partial or complete loss of photoreceptors sometimes show greater activity at night. Mutations in a widely expressed putative cation channel DMα1U (narrow abdomen [na]) also results in a switch from diurnal to nocturnal activity (Nash, 2002). Rescue of diurnal activity was achieved by expression of wild-type channel in parts of the circuit that included lLNvs (Sheeba, 2008b and references therein).
In the case of NaChBac-induced hyperexcitation of lLNvs, in which NaChBac, a voltage-gated sodium channel, was expressed in LNvs) the normal pattern of light-driven activity during the day is reversed to a novel pattern of firing rate that favors higher activity in the night. The lLNvs are not likely to be 'nocturnal' neurons. Rather, they appear to drive locomotor activity according to their relative day versus night pattern of excitation. Considering that the wild-type electrophysiological firing properties of lLNvs are so dramatically different from NaChBac-evoked firing and sustained hyperexcitation, it is reasonable to wonder how NaChBac expression in LNvs, and specifically a lLNv subset, yields such a coherent pattern of behavioral activity. The results above show that lLNvs modulate arousal and sleep and that altering the relative pattern of day versus night excitability is sufficient to evoke a temporal change in behavioral output. On the basis of these observations, it is hypothesized that the precise firing pattern or timing of lLNv electrical activity is probably not important, thereby making NaChBac expression an appropriate tool to study these neurons and probably other modulatory circuits for which general changes in the gain of activity rather than the precise pattern of activity dictate functional output (Sheeba, 2008b).
The results reveal that the PDF-expressing peptidergic lLNvs modulate arousal and wakeful behavior as well as sleep stability. Considering these functional studies, lLNvs appear to act as an arousal circuit that is physiologically activated by light and borders with, but is distinct from, the circadian pacemaker and downstream sleep circuits. A number of other recently described modulatory systems in Drosophila influence behavioral locomotion and sleep, including aminergic and GABAergic neurons. As noted, many of the overall features of morning and evening peaks in locomotor activity are retained when lLNv and other peptidergic neuronal subsets are hyperexcited. Considering the naturalistic implications, temporal-niche switching has been observed in a few animals, including the social ant species Camponotus compressus. Among the worker class of these ants, some individuals are diurnal and others nocturnal, and these plastic behavioral differences are associated with differences in their underlying free-running circadian period. On the basis of the current results, it is possible that activity changes in a relatively small number of arousal neurons could influence both short-term temporal-niche switching or long-term evolutionary commitment to a given temporal niche (Sheeba, 2008b).
Circadian clocks synchronize to the solar day by sensing the diurnal changes in light and temperature. In adult Drosophila, the brain clock that controls rest-activity rhythms relies on neurons showing Period oscillations. Nine of these neurons are present in each larval brain hemisphere. They can receive light inputs through Cryptochrome (CRY) and the visual system, but temperature input pathways are unknown. This study investigated how the larval clock network responds to light and temperature. Focus was placed on the CRY-negative dorsal neurons (DN2s), in which light-dark (LD) cycles set molecular oscillations almost in antiphase to all other clock neurons. The phasing of the DN2s in LD depends on the pigment-dispersing factor (PDF) neuropeptide in four lateral neurons (LNs), and on the PDF receptor in the DN2s. In the absence of PDF signaling, these cells appear blind, but still synchronize to temperature cycles. Period oscillations in the DN2s were stronger in thermocycles than in LD, but with a very similar phase. Conversely, the oscillations of LNs were weaker in thermocycles than in LD, and were phase-shifted in synchrony with the DN2s, whereas the phase of the three other clock neurons was advanced by a few hours. In the absence of any other functional clock neurons, the PDF-positive LNs were entrained by LD cycles but not by temperature cycles. These results show that the larval clock neurons respond very differently to light and temperature, and strongly suggest that the CRY-negative DN2s play a prominent role in the temperature entrainment of the network (Picot, 2009).
Although the absence of PDF severely affects Drosophila activity rhythms in DD, the exact function of the neuropeptide in the adult clock neuronal network remains unclear. In LD, PDF is required to produce a morning activity peak and to properly phase the evening peak, but not to entrain the brain clock. The behavioral phenotypes of PDF receptor mutants resemble that of the pdf01 mutant. PDFR is expressed in all clock neurons except the large ventral lateral neurons (l-LNvs), supporting a role of PDF in maintaining phase coherence within the adult clock network in DD. The loss of PER oscillations in the DN2s of pdf01 larvae demonstrates a clear and novel role of PDF in transmitting not only phase information but also a synchronizing signal without which the receiving neurons are not entrained in LD (Picot, 2009).
The current results show that the CRY-less DN2s are 'blind' neurons that perceive light indirectly. The PDF receptor rescue experiments strongly suggest that PDF acts on its receptor on the larval DN2s themselves, which are located in the vicinity of the LN axon terminals. Furthermore, DN2s possess a wide and dense neuritic network that borders on the axons of the LNs over a large fraction of their length. However, it cannot be ruled out that expression of the receptor in the (PDF-negative) fifth LN is involved in synchronizing the DN2s downstream, through PDF-independent mechanisms (Picot, 2009).
The PDF-negative fifth LN is also a CRY-negative clock neuron, but it cycles in phase with the CRY-positive neurons of the larval brain. The visual input to the PDF-expressing LNs appears sufficient to phase them normally even in cryb mutants. It could thus be expected to entrain the CRY-less fifth LN in phase with the other larval LNs, as observed, in contrast to the CRY-less DN2s. A direct input from the visual system to the fifth LN is also consistent with its PDF-independent entrainment by LD cycles. Similarly, light entrainment of the larval DN1s in cryb mutants is consistent with their suggested connection to the visual system. Thus, the CRY-less DN2s would be the only larval clock neurons devoid of such a connection (Picot, 2009).
Adult eclosion rhythms depend on the PDF-expressing LNs and appear to require the PDF-dependent clock that resides in the prothoracic gland. Since the larval DN2s project in the pars intercerebralis, a region of the brain that sends projections to the prothoracic gland, they could play a role in this physiologically important clock function. These results raise the possibility that the damped PER oscillations in the DN2s of the pdf01 mutants participate to their eclosion phenotype (Picot, 2009).
The DN2s are the only larval clock neurons that are phased identically by light and temperature, but their temperature entrainment appears independent of any LN-derived signal. PER oscillations in the DN2s have a larger amplitude in HC cycles than in LD cycles, also suggesting a prominent role of temperature in their entrainment. Conversely, the molecular oscillations of the PDF-positive LNs have a larger amplitude in LD compared with HC cycles. In the latter, the molecular oscillations of the PDF-expressing LNs seem to follow those in the DN2s, with a large phase change compared with LD conditions. The DN1s and the PDF-negative fifth LN, in contrast, share another phase that is slightly advanced. Interestingly, behavioral and transcriptome data in adult flies indicate that HC cycles result in a general phase advance relative to LD cycles. Cooperative synchronization of the clock by light and temperature likely requires temperature changes to act earlier than light changes since changes in temperature always lag behind changes in solar illumination in nature. The very different relative phasing of the larval clock neurons in HC versus LD cycles suggests different ecological constraints on this life stage, spent mostly burrowed in food, in which light may be a weaker Zeitgeber, and in which the lag between temperature and light changes may be quite different (Picot, 2009).
When a functional clock is absent from the DN2s (and the fifth LN), the larval PDF-expressing LNs are unable to entrain to thermocycles, whereas they autonomously entrain to LD cycles. It remains possible that autonomous temperature entrainment of the larval LNs (but not the DN2s) requires per transcriptional regulation, which the GAL4-UAS system is lacking. But the results demonstrate the existence of a control exerted on the LN clock by CRY-negative clock cells when temperature is the synchronizing cue. Although a role of the fifth LN cannot be ruled out, the absence of autonomous photoperception by the DN2s nicely fits with a role in temperature entrainment. The high cycling amplitude of the DN2s in thermocycles and the locking of the phase of the LNs on that of the DN2s in these conditions strongly support their role in the temperature entrainment of the LNs (Picot, 2009).
Additional studies should investigate whether the DN2s communicate with the LNs via fibers that appear to run along the dorsal projection of the LNs. Alternatively, the dense dendritic-like network of the DN2s could ensure reciprocal exchanges between them and the LNs. A model is thus proposed whereby, in the larval brain, the DN2s and the four PDF-positive LNs form a distinct subnetwork, with the LNs entraining the DN2s in LD, whereas the opposite is true in HC). What becomes of their hierarchy in constant conditions, after entrainment stops? Their relative phases appear to change little at least during the first 2 d after entrainment, whether they have been set in antiphase by LD entrainment, or in phase by HC entrainment. This suggests that, whatever the entraining regimen, the LNs and the DN2s run autonomously in constant conditions. However, it cannot be excluded that one of the two groups still dominates but requires more time after the end of entrainment to shift the phase of the other (Picot, 2009).
The rhythmic behavior of the adult flies that emerge from the temperature-entrained larvae is almost in antiphase compared with the one of flies entrained by light during the larval stage. This strongly suggests that the phase of the adult rhythms is set by the antiphasic oscillations of the larval PDF-positive LNs, consistent with these cells being the only neurons in which molecular cycling persists throughout metamorphosis. It is thus believed that the large phase shift of adult activity can be accounted for simply by the large phase shift of molecular oscillations in the PDF-expressing LNs (Picot, 2009).
It is often assumed that temperature affects the molecular clock directly and identically in all clock cells, as opposed to light, which requires dedicated input pathways. However, in the adult, thermocycles phase the brain clock differently from all peripheral clocks, as judged from whole-tissue oscillations of a luciferase reporter enzyme (Glaser, 2005). Recent data suggest that subsets of clock neurons in the Drosophila adult brain may indeed be dedicated to temperature entrainment. In experiments combining LD and HC entrainment, all DN groups, as well as the less studied lateral posterior neurons (LPNs), seem to preferentially follow thermocycles, whereas the other LNs preferentially follow LD cycles (Miyasako, 2007). Although adult PDF+ LNs are able to entrain to thermocycles in the absence of any other functional clock, they do not seem to be required for (and actually slowed down) the temperature entrainment of activity rhythms, whereas the PDF-negative LPNs appear to play a prominent role in such conditions (Picot, 2009).
The current results indicate that a similar specialization toward light or temperature entrainment exists in the larval brain. The DN2s, which appear to be the most temperature-responsive clock neurons, are by themselves completely blind. Conversely, the four PDF-positive LNs, which may be the most light-sensitive clock neurons (with both CRY and the visual system as inputs), appear almost temperature blind, and depend on the DN2s for temperature entrainment. PER-negative DN2s do not allow PER oscillations in the larval LNs, suggesting that entrainment of the latter in HC cycles depends on clock function in the former. The hierarchy of clock neurons thus appears very different during entrainment of the clock network by one or the other Zeitgeber (Picot, 2009).
Light has profound behavioral effects on almost all animals, and nocturnal animals show sensitivity to extremely low light levels. Crepuscular, i.e., dawn/dusk-active animals such as Drosophila melanogaster are thought to show far less sensitivity to light. This study reports that Drosophila respond to extremely low levels of monochromatic blue light. Light levels three to four orders of magnitude lower than previously believed impact circadian entrainment and the light-induced stimulation of locomotion known as positive behavioral masking. GAL4;UAS-mediated rescue of tyrosine hydroxylase (DTH) mutant (ple) flies was used to study the roles of dopamine in these processes. Evidence is presented for two roles of dopamine in circadian behaviors. First, rescue with either a wild-type DTH or a DTH mutant lacking neural expression leads to weak circadian rhythmicity, indicating a role for strictly regulated DTH and dopamine in robust circadian rhythmicity. Second, the DTH rescue strain deficient in neural dopamine selectively shows a defect in circadian entrainment to low light, whereas another response to light, positive masking, has normal light sensitivity. These findings imply separable pathways from light input to the behavioral outputs of masking versus circadian entrainment, with only the latter dependent on dopamine (Hirsch, 2010).
Sensitivity to extremely low levels of light is most commonly found in nocturnal animals. These animals, such as nocturnal geckos or insects such as nocturnal hawkmoths, can not only sense extremely low levels of light but can also discern colors at light intensities well below those to which diurnal animals are sensitive. Humans and diurnal vertebrates lose color vision at light intensities comparable to dim moonlight at irradiances of 3-10 nW/cm2. In contrast, nocturnal hawkmoths and geckos can discern colors even at intensities of ~0.01-0.3 nW/cm2 and normally function in starlight, ~0.001 nW/cm2. Extreme light sensitivity in nocturnal insects commonly involves adaptations to their compound eyes to allow summation of photons from many individual ommatidia. These visual system adaptations are not seen in diurnal insects such as the fruit fly Drosophila melanogaster. Accordingly, current data accord Drosophila with rather modest light sensitivity. For light-dependent entrainment of circadian rhythmicity, ~40 nW/cm2 blue light was thought to be required, although subsequent studies show entrainment by 1-5 nW/cm2 white light. Wild-type flies are now thought to entrain at ~0.04 nW/cm2 blue light (C. Helfrich-Forster, personal communication to Hirsch, 2010). An intensity of ~0.5 nW/cm2 white light is reported to cause positive behavioral masking, the largely circadian clock-independent stimulation of locomotion. For comparison, this study found that a dark-adapted human observer loses the ability to perceive the diffuse planar blue light sources used in the present study at intensities of ~0.01-0.03 nW/cm2. This intensity is difficult to compare to published human perception studies, which commonly use short duration flashes of focal light (Hirsch, 2010).
This study found unexpectedly strong light sensitivity for Drosophila melanogaster, with behavioral masking and circadian entrainment at intensities as low as 0.001 nW/cm2 and at least two roles for dopamine in circadian rhythmicity. First, DTH rescue flies showed poor behavioral rhythmicity in constant dark conditions, independent of whether dopamine levels were rescued in the nervous system. Second, it was found that neuronal DTH rescue flies lacking neuronal dopamine showed reduced light sensitivity for circadian entrainment, whereas light sensitivity of behavioral masking was unaffected. Dopamine has several roles in Drosophila neural function, from modulation of locomotor behaviors and arousal states to learning and memory, but a role for dopamine in insect light-dependent circadian behavioral entrainment is novel (Hirsch, 2010).
The two circadian phenotypes likely represent separate roles for dopamine, presumably in different regions of the nervous system, because reduced amplitude of rhythmicity, as seen in DTH rescue lines, is normally associated with higher rather than lower efficacy of reentrainment. The dopaminergic system in Drosophila is highly rhythmic, as evidenced by rhythmicity in responsiveness to dopamine agonists and by the rhythmic transcription of the tyrosine hydroxylase gene ple, which encodes the rate-limiting enzyme in dopamine biosynthesis. The rhythmicity of the ple transcript may explain the poor rhythmicity in ple rescue animals. These animals have near-normal levels of brain dopamine in an apparently normal cellular pattern, but the inclusion of the GAL4 transcription factor into the regulatory cascade will almost certainly interfere with normal temporal cycling of the DTH transcript. Note that significant diurnal variation in levels of brain dopamine in brain extracts have not been detected, but this does not preclude diurnal variation in dopamine neuron subsets (Hirsch, 2010).
Low-light circadian entrainment is disrupted in the brain dopamine-deficient DTHgFS±ple flies. The simplest mechanism for the disruption of low-light circadian entrainment would be due to alterations in the photoreceptive pathway, which could be via cryptochrome (CRY) or visual photoreceptors. There is some support for dopaminergic involvement in the CRY pathway, because Sathyanarayanan (2008) identified ple in a screen for genes that, when targeted by RNA interference, have a strong inhibitory effect on light-dependent degradation of CRY and timeless (TIM) in cultured cells. This could indicate a positive role for dopamine in light-dependent degradation of these molecules, providing a potential mechanism for the reduced light sensitivity for circadian entrainment that was observed in the absence of dopamine (Hirsch, 2010).
Alternatively, it is known that visual photoreceptors are involved in dim-light entrainment because genetic loss of all photoreceptive visual organs results in at least a three-order-of-magnitude reduction in blue light sensitivity for circadian entrainment. Analogous studies in mice show an ~60-fold reduction in dim-light sensitivity for entrainment in animals lacking both rods and cones (Hirsch, 2010).
A role for dopamine in fly visual function has some support in that cyclic AMP (cAMP) can slow the response to light in a preparation of isolated Drosophila photoreceptors (Chyb, 1999), and this effect can be mimicked by application of octopamine or dopamine, an effect interpreted as enhanced adaptation to dark. Dopamine signaling, via cAMP second-messenger pathways, is not currently considered part of the main insect visual transduction pathway. However, dopamine involvement could have been missed if it has an exclusive role in a neural pathway selectively required for circadian entrainment by dim light (Hirsch, 2010).
There is strong support of a role for dopamine functioning in the vertebrate retina, which makes visual involvement of dopamine in the fly all the more likely. The vertebrate retina contains autonomous circadian oscillators that are thought to allow the retina to prepare for the large difference in light intensity between day and night. Central to this rhythmicity are opposing and rhythmic roles for melatonin and dopamine, with release of each modulator inhibiting synthesis and/or release of the other. The best defined role for dopamine in the vertebrate circadian oscillator is in entraining fetal rodents prior to light exposure, a capacity lost in adults. This role of dopamine could be related to the roles that have been uncovered in adult Drosophila (Hirsch, 2010).
The selective effect of neural dopamine on low-light entrainment versus low-light masking behavior implies separable pathways involved in modulating these behaviors, a novel finding because previous studies have only identified circadian components with parallel effects on masking (Mazzoni, 2005). The best defined synaptic connections from eye to circadian neurons are the projections from the Drosophila eyelet, a remnant of the larval photoreceptive Bolwig's organ. This photoreceptive organ makes projections that terminate in close apposition to neurites from the small and large ventral lateral neurons, neurons key to circadian rhythmicity. Connections from the main visual photoreceptors to these circadian neurons must be indirect because the rod-like outer photoreceptor ommatidia terminate in the optic lamina, and the cone-like central ommatidia terminate in the optic medulla. Nonetheless, dopamine could be acting as a neuromodulator in any of these pathways to increase sensitivity to a light-dependent signal. The genetic tools available in Drosophila should prove useful to precisely identify these pathways (Hirsch, 2010).
Cross-species conservation of sleep-like behaviors
predicts the presence of conserved molecular mechanisms underlying
sleep. However, limited experimental evidence of conservation exists.
This prediction is tested directly in this study. During lethargus,
Caenorhabditis elegans spontaneously sleep in short bouts that are
interspersed with bouts of spontaneous locomotion. Twenty-six genes
required for Drosophila melanogaster sleep were identified. Twenty
orthologous C. elegans genes were selected based on similarity. Their
effect on C. elegans sleep and arousal during the last larval
lethargus was assessed. The 20 most similar genes altered both the
quantity of sleep and arousal thresholds. In 18 cases, the direction
of change was concordant with Drosophila studies published previously.
Additionally, a conserved genetic pathway was delineated by which
dopamine regulates sleep and arousal. In C. elegans neurons, G-alpha
S, adenylyl
cyclase, and protein
kinase A act downstream of D1 dopamine receptors to regulate these
behaviors. Finally, a quantitative analysis of genes examined herein
revealed that C. elegans arousal thresholds were directly correlated
with amount of sleep during lethargus. However, bout duration varies
little and was not correlated with arousal thresholds. The comprehensive
analysis presented in this study suggests that conserved genes and
pathways are required for sleep in invertebrates and, likely, across the
entire animal kingdom. The genetic pathway delineated in this study
implicates G-alpha S and previously known genes downstream of dopamine
signaling in sleep. Quantitative analysis of various components of
quiescence suggests that interdependent or identical cellular and
molecular mechanisms are likely to regulate both arousal and sleep entry
(Singh, 2014).
Sleep homeostasis is crucial for sleep regulation. The role of epigenetic regulation in sleep homeostasis is unestablished. Previous studies showed that octopamine is important for sleep homeostasis. However, the regulatory mechanism of octopamine reception in sleep is unknown. This study identified an epigenetic regulatory cascade (Stuxnet-Polycomb-Octβ2R) that modulates the octopamine receptor in Drosophila. stuxnet positively regulates Octβ2R through repression of Polycomb in the ellipsoid body of the adult fly brain and Octβ2R is one of the major receptors mediating octopamine function in sleep homeostasis. In response to octopamine, Octβ2R transcription is inhibited as a result of stuxnet downregulation. This feedback through the Stuxnet-Polycomb-Octβ2R cascade is crucial for sleep homeostasis regulation. This study demonstrates a Stuxnet-Polycomb-Octβ2R-mediated epigenetic regulatory mechanism for octopamine reception, thus providing an example of epigenetic regulation of sleep homeostasis (Zhao, 2021).
Drosophila has been used as a model system to study mechanisms of sleep regulation. The first studies on sleep in Drosophila revealed that they periodically enter a quiescence state that meets a set of criteria for sleep. Drosophila sleep is monitored normally by a Drosophila activity monitoring system (DAMS) and is defined as immobility for 5 min or longer which is a sleep bout. Drosophila sleep mainly happens at night, while a period of siesta is in the mid-day. For example, total sleep time is around 380 min (male) and 250 min (female) during the day time, and 480 min (male) and 490 min (female) during the night time in w1118 (Zhao, 2021).
In Drosophila, central complex structures, especially the ellipsoid body (EB) and fan-shaped body (FSB), are important for sleep homeostasis regulation. Activation of dorsal FSB neurons is sufficient to induce sleep. The dorsal FSB also integrates some sleep inhibiting signals. Both dorsal FSB and EB ring 2 are important in sleep homeostasis. Recently, the helicon cells were found to connect the dorsal FSB and EB Ring 2, indicating that these EB and FSB are connected (Zhao, 2021).
Multiple studies indicate that the epigenetic mechanisms are involved in circadian regulation. However, a direct link between epigenetic regulation and sleep homeostasis is not yet established (Zhao, 2021).
Octopamine (OA) in Drosophila is a counterpart of vertebrate noradrenaline. Previous studies in Drosophila showed that OA is a wake-promoting neurotransmitter and plays an important role in regulating both sleep amount and sleep homeostasis. The mutants of the OA synthesis pathway show an increased total sleep. Activation of OA signaling inhibits sleep homeostasis, while in OA synthesis pathway mutants, an enhanced sleep homeostasis is observed. Study of the neural circuit responsible for the sleep/wake effect of OA showed that octopaminergic ASM neuronsproject to the pars intercerebralis (PI), where OAMB (one of the OA receptors)-expressing insulin-like peptide (ILP)-secreting neurons act as downstream mediators of OA signaling. However, the effects of manipulating ASM neurons or ILP-secreting neurons are much weaker than those observed by manipulating all OA secreting neurons. Moreover, the effect of octopamine is not completely suppressed in the OAMB286 mutant, arguing that another receptor or circuit may participate in this process (Zhao, 2021).
Eight OA receptors are identified to date: OAMB, Octβ1R, Octβ2R, Octβ3R, TAR1, TAR2, TAR3, and Octα2R. Although the expression pattern of OA is identified, the endogenous expression profile of these receptors is lacking. A previous study demonstrated that the mushroom body-expressed OAMB mediates the sleep:wake effect of OA. Recently, Octβ2R was shown to be important for the OA effect on endurance exercise adaptation. How the versatility of OA function is mediated by the diverse array of its receptors needs further study. Moreover, the upstream regulatory mechanisms of OA receptors are still unknown (Zhao, 2021).
A previous study showed that Stuxnet (Stx) is important in mediating Polycomb (Pc) protein degradation in the proteasome (Du, 2016). Stx, which is an ubiquitin like protein, mediates Polycomb (Pc) protein degradation through binding to the proteasome with a UBL domain at its N terminus and to Polycomb through a Pc-binding domain. stx level changes result in a series of homeotic transformation phenotypes. Pc is an epigenetic regulator functioning in Polycomb Group (PcG) Complexes. Although it is reported that PcG component E(Z) is involved in circadian regulation, the role of stx in adult physiological process is unknown (Zhao, 2021).
This study identified the role of the epigenetic regulator stx in sleep regulation. stx positively regulates Octβ2R through regulation of Polycomb in the EB of the adult fly brain. Further study demonstrated that the Stuxnet-Polycomb-Octβ2R cascade plays an important role in sleep regulation. In order to elucidate the role of this Stuxnet-Polycomb-Octβ2R cascade in sleep regulation, the role of various Octβ receptors was systematically identified in sleep regulation. Octβ2R was found to be one of the receptors that mediates OA function in sleep homeostasis. More interestingly, it was found that stx was OA-responsive depending on the Octβ1R. Based on these data, it is proposed that the Stuxnet-Polycomb-Octβ2R cascade provides a feedback mechanism for OA signals to the EB to regulate sleep homeostasis and sleep amount (Zhao, 2021).
This study highlights the importance of epigenetic regulation on sleep. Although epigenetic regulation was intensively studied in adult pathological processes such as cancer, epigenetic factors have been far less studied in other physiological processes such as sleep. This study provides an example of the maintenance role of PcG complex in sleep regulation. Although the core PcG complex component Pc is ubiquitously expressed, its regulator stx is tissue specifically distributed, and this distribution may keep appropriate activity of Pc as well as the PcG complex in a tissue-specific manner. The factors regulating the tissue specificity of stx expression need to be further investigated (Zhao, 2021).
A previous study found that mutation of Octβ2R does not have an obvious sleep phenotype. The current data were compared with the published Octβ2Rf05679 mutant data. Although Octβ2Rf05679 mutant was shown not significantly affected total sleep, this study found that the Octβ2Rf05679 has mild effect on sleep. Other Octβ2R mutants were tested, and it was found that the male flies from these mutations indeed have sleep phenotype (Zhao, 2021).
Published studies have shown that the sleep phenotype of octopamine pathway mutants is different between video-based method and DAM-based method. For example, based on DAM data, the TβH mutant resulted in increased sleep per day, while the same mutant showed decreased sleep based on video data. This study used the video-based method to repeat the TβH mutant phenotype. The results showed that compared with the control flies, the TβH mutant got significantly less sleep. This result is consistent with the previously published data. Through close observation of TβH mutant and control flies, this study found that this mutant has much more frequent grooming behavior than the controls. The TβH mutant and control flies were video recorded for 10 min between ZT3.5 and ZT4.5. The results showed a statistically significant increase of the total number of grooming case. The difference between video-based method and DAM-based method is that these grooming behaviors can be detected in video-based methods, but not in DAM-based methods. Multiple studies have established a positive correlation between octopamine treatment and grooming behavior. Theoretically, TβH allele should result in a decrease in octopamine synthesis. The opposite phenotype may be caused by increased tyramine in TβH mutant or by other feedback regulation. The alleles for Octβ2R receptor used in this study show a similar grooming behavior as the control flies. The previously published octβ2R knockout allele should be a stronger one. The difference of sleep phenotypes between video-based and DAM-based methods may be due to the grooming behavior induced by the massive decrease of octopamine detection. Or other unrelated effects caused by the compensation effect previously reported. One hypothesis is that the significant change of grooming behavior probably masks the sleep behavior. The relationship between grooming and sleep needs to be further clarified. The detection of the sleep phenotype without significant changes in grooming phenotype may be a better strategy to get reliable sleep phenotype. If the increase of grooming in TβH mutant is a side effect caused by the increased tyramine, the identification of the phenotypes of octopamine treatment or collective phenotype of octopamine receptors may be more reliable ways to draw conclusions on the function of octopamine. Furthermore, whether grooming is epistatic to sleep is a problem worthy of further study (Zhao, 2021).
Two aspects of sleep homeostasis need to be further studied. First, this study found that Octβ2R and stx colocalize in a subset of EB neurons. In a previously study, EB R2 neurons were found to be responsible for sleep homeostasis regulation. The relationship of these two groups of EB neurons needs further study. Second, the OA-treated Octβ2R mutant has more sleep recovery than the control. This indicates that OA induces more sleep recovery in the condition of Octβ2R downregulation. It seems that in this condition OA induces certain pathways to counteract its role in sleep homeostasis. One possibility is that Octβ2R negatively regulates Octβ3R which results in increased sleep pressure in the absence of Octβ2R. Further studies are needed to clarify the mechanism (Zhao, 2021).
The results suggest the stx-Pc-Octβ2R regulatory cascade serves as a buffering step for OA function in sleep homeostasis. Two-way regulation of OA on stx leads to reverse changes of stx-the more OA, the less stx and vice versa. Through the function of stx-Pc-Octβ2R regulatory cascade, the Octβ2R transcription is changed accordingly. Variation of Octβ2R transcription could buffer the OA response. As a result, the unfavorable effect of OA causing dramatic decrease of sleep amount and homeostasis could be compensated by its receptor (Zhao, 2021).
Sleep homeostasis is crucial for sleep regulation. The role of epigenetic regulation in sleep homeostasis is unestablished. Previous studies showed that octopamine is important for sleep homeostasis. However, the regulatory mechanism of octopamine reception in sleep is unknown. This study identified an epigenetic regulatory cascade (Stuxnet-Polycomb-Octβ2R) that modulates the octopamine receptor in Drosophila. stuxnet positively regulates Octβ2R through repression of Polycomb in the ellipsoid body of the adult fly brain and Octβ2R is one of the major receptors mediating octopamine function in sleep homeostasis. In response to octopamine, Octβ2R transcription is inhibited as a result of stuxnet downregulation. This feedback through the Stuxnet-Polycomb-Octβ2R cascade is crucial for sleep homeostasis regulation. This study demonstrates a Stuxnet-Polycomb-Octβ2R-mediated epigenetic regulatory mechanism for octopamine reception, thus providing an example of epigenetic regulation of sleep homeostasis (Zhao, 2021).
The Drosophila mushroom
body (MB) is a key associative memory center that has also been
implicated in the control of sleep. However, the identity of MB neurons
underlying homeostatic sleep regulation, as well as the types of sleep
signals generated by specific classes of MB neurons, has remained poorly
understood. Two MB output neuron (MBON) classes whose axons convey sleep
control signals from the MB to converge in the same downstream target
region: a cholinergic sleep-promoting MBON class and a glutamatergic
wake-promoting MBON class have been previously identified. This study
deploys a combination of neurogenetic, behavioral, and physiological
approaches to identify and mechanistically dissect sleep-controlling
circuits of the MB. The existence of two segregated excitatory synaptic
microcircuits that propagate homeostatic sleep information from
different populations of intrinsic MB "Kenyon cells" (KCs) to specific
sleep-regulating MBONs was revealed: sleep-promoting KCs increase sleep
by preferentially activating the cholinergic MBONs, while wake-promoting
KCs decrease sleep by preferentially activating the glutamatergic MBONs.
Importantly, activity of the sleep-promoting MB microcircuit is
increased by sleep deprivation and is necessary for homeostatic rebound
sleep (i.e., the increased sleep that occurs after, and in compensation
for, sleep lost during deprivation). These findings reveal for the first
time specific functional connections between subsets of KCs and
particular MBONs and establish the identity of synaptic microcircuits
underlying transmission of homeostatic sleep signals in the MB
(Sitaraman, 2015a). This study has used a combination of sophisticated cell-specific
genetic manipulations with behavioral sleep analysis and optical
electrophysiology to provide an unprecedented level of detailed
understanding of the propagation of homeostatic sleep signals through
microcircuits of the Drosophila MB. Specifically, two parallel
segregated compartment-specific microcircuits were identified that
regulate sleep: a wake-promoting microcircuit that originates in α'/β'
and γm KCs and converges onto MBON-γ5β'2a/β'2mp/β'2mp_bilateral and a
sleep-promoting microcircuit that originates in γd KCs and converges
onto MBON-γ2α'1. Importantly, it was shown not only that exogenous
activation of these microcircuits is sufficient to regulate sleep, but
also that physiological manipulation of sleep need by sleep deprivation
alters their endogenous neural activity, and propagation of these neural
signals to downstream targets outside the MB is essential for the
generation of homeostatic rebound sleep (Sitaraman, 2015a). Previous studies using broadly expressed traditional GAL4 drivers have
implicated the MB in the control of sleep, but due to lack of
appropriate cell-specific drivers, were unable to resolve specific
sleep-regulating MB cell types, although very recent studies have
specifically implicated α'/β' KCs and MB-MV1/PPL1 dopaminergic MB
neurons in regulating sleep. Moreover, previous studies have not
established a role for the MB in the generation and/or propagation of
homeostatic sleep signals necessary for rebound following sleep
deprivation. While a mutation of the amnesiac gene, which is
expressed in a pair of neurons innervating the MB lobes, was shown to
impair homeostatic sleep rebound, rebound was not found to be strongly
affected by very broad synaptic inactivation of the MB KCs that comprise
the lobes. This report has established a comprehensive catalog of the
KCs and MBONs that control sleep, making use of a novel library of
split-GAL4 lines targeting each of the cell types of the MB. Combining
sophisticated behavioral genetic and optical electrophysiology
approaches has allowed determination of the roles of specific MB cell
types in encoding homeostatic sleep signals under physiological
conditions. Then specific synaptic microcircuits were identified linking
sleep-controlling KCs to sleep-controlling MBONs, revealing the synaptic
mechanisms underlying the propagation of homeostatic sleep signals
through the MB associative network (Sitaraman, 2015a). Based on these results, a detailed mechanistic model is proposed for
homeostatic control of sleep by excitatory microcircuits in the
Drosophila MB. Wake-promoting MBON-γ5β'2a/β'2mp/β'2mp_bilateral and
sleep-promoting MBON-γ2α'1 each receive anatomical inputs from both
wake-promoting γm and α'/β' KCs and sleep-promoting γd KCs. However,
functional segregation of sleep control information into separate
microcircuits is enforced by greater synaptic weights between γm and
α'/β' KCs and MBON-γ5β'2a/β'2mp/β'2mp_bilateral and between γd KCs and
MBON-γ2α'1. Anatomical studies indicate that the axons of
MBON-γ5β'2a/β'2mp/β'2mp_bilateral and MBON-γ2α'1 exit the MB and
terminate convergently in the superior medial protocerebrum (SMP) and
crepine (CRE) neuropils. Intriguingly, dendrites of some central complex
(CX) neurons-a brain region involved in locomotor control and
implicated in sleep and sleep homeostasis-arborize in SMP and CRE. It
is thus speculated that segregated homeostatic sleep-promoting and
wake-promoting signals are generated in the KC-to-MBON microcircuits of
the MB and propagate to the CX, where they are integrated to ultimately
control sleep. Future studies are needed to further refine the
understanding of the neurochemistry and physiology of the MB sleep
control microcircuits, explore the mechanisms by which sleep deprivation
alters microcircuit activity, and elucidate the connections between the
MB and its downstream sleep control targets. In light of the
relationship between sleep, learning, and synaptic homeostasis and the
recent discovery that sleep-regulating MBON-γ5β'2a/β'2mp/β'2mp_bilateral
and MBON-γ2α'1 are also important for some forms of associative
learning, it will be of great interest to determine how activity of the
sleep-controlling MB synaptic microcircuits influences memory formation
and consolidation (Sitaraman, 2015a).
P>
The dynamic nature of sleep in many animals suggests distinct stages that serve different functions. Genetic sleep induction methods in animal models provide a powerful way to disambiguate these stages and functions, although behavioral methods alone are insufficient to accurately identify what kind of sleep is being engaged. In Drosophila, activation of the dorsal fan-shaped body (dFB) promotes sleep, but it remains unclear what kind of sleep this is, how the rest of the fly brain is behaving, or if any specific sleep functions are being achieved. This study developed a method to record calcium activity from thousands of neurons across a volume of the fly brain during spontaneous sleep and compared this to dFB-induced sleep. Spontaneous sleep was found to typically transition from an active "wake-like" stage to a less active stage. In contrast, optogenetic activation of the dFB promotes sustained wake-like levels of neural activity even though flies become unresponsive to mechanical stimuli. When flies were probed with salient visual stimuli, it was found that the activity of visually responsive neurons in the central brain was blocked by transient dFB activation, confirming an acute disconnect from the external environment. Prolonged optogenetic dFB activation nevertheless achieved a key sleep function by correcting visual attention defects brought on by sleep deprivation. These results suggest that dFB activation promotes a distinct form of sleep in Drosophila, where brain activity appears similar to wakefulness, but responsiveness to external sensory stimuli is profoundly suppressed (Tainton-Heap, 2020).
Control of sleep by dopaminergic inputs to the
Drosophila mushroom body The Drosophila mushroom body (MB) is an associative learning network
that is important for the control of sleep. Particular intrinsic MB
Kenyon cell (KC) classes have been identified that regulate sleep
through synaptic activation of particular MB output neurons (MBONs)
whose axons convey sleep control signals out of the MB to downstream
target regions. Specifically, it was found that sleep-promoting KCs
increase sleep by preferentially activating cholinergic
sleep-promoting MBONs, while wake-promoting KCs decrease sleep by
preferentially activating glutamatergic wake-promoting MBONs. By using
a combination of genetic and physiological approaches to identify
wake-promoting dopaminergic neurons (DANs) that innervate the MB, it
was shown that they activate wake-promoting MBONs. These studies
reveal a dopaminergic sleep control mechanism that likely operates by
modulation of KC-MBON microcircuits. This study used a combination of sophisticated cell-specific genetic
manipulations with behavioral sleep analysis and optical
electrophysiology to identify compartment-specific wake-promoting MB
DANs that activate wake-promoting microcircuits. Previous studies have
implicated DANs innervating the central complex (CX) - a brain region
involved in locomotor control - in regulating sleep, and other
non-dopamingeric CX neurons have been implicated in homeostatic
control of sleep. In addition, it has recently been shown that
manipulations of dopamine signaling in the MB alter sleep, although
the precise DANs involved remains unclear. This study has now
identified specific wake-promoting MB DANs and shown that they
innervate lobe compartments also innervated by wake-promoting KCs and
MBONs. Importantly, this study has also shown that dopamine secretion
by DANs innervating a particular MB lobe compartment acts through D1
subtype receptors to activate the wake-promoting microcircuit specific
to that compartment to a much greater extent than it activates the
sleep-promoting microcircuit residing in different compartments. This
provides direct physiological evidence for compartment-specific
dopamine signaling in the regulation of sleep by the MB, and is
consistent with a previous study in the context of learning and memory
(Boto, 2014). Future studies are required to determine additional
cellular and molecular details of how dopamine signals modulate
sleep-regulating microcircuits. On the basis of recently published studies of MB control of sleep and
the results presented in this study, a unified mechanistic model is
proposed for homeostatic control of sleep by excitatory microcircuits
in the Drosophila MB. Wake-promoting MBON-γ5β'2a/β'2mp/β'2mp_bilateral
and sleep-promoting γ2α'1 each receive anatomical inputs from both
wake-promoting γm and α'/β' KCs KCs and sleep-promoting γd KCs.
However, segregation of sleep control information into separate
microcircuits is enforced by greater synaptic weights between γ and γm
and α'/β' KCs and MBON-γ5β'2a/β'2mp/β'2mp_bilateral, and between γm
and α/β' KCs and MBON-γ5β'2a/β'2mp/β'2mp_bilateral, and between γd KCs
and MBON-γ2α'1. Thus it is hypothesize that
compartment-specific dopamine signals from MB DANs could potentially
determine these differences in synaptic weight. Future studies will
test this hypothesis. Interestingly, other fly behaviors have recently been found to be
regulated by sleep-controlling compartment-specific MB microcircuits.
For example, the integration of food odor to suppress innate avoidance
of CO2 is mediated by MBON-γ5β'2a/β'2mp/β'2mp_bilateral and PAM DANs
that innervate the β'2 compartment (Lewis, 2015). Optogenetic
activation experiments reveal that wake-promoting
γ5β'2a/β'2mp/β'2mp_bilateral mediates innate avoidance, while
MBON-γ2α'1 mediates attraction. However, thermogenetic inactivation
studies reveal that both MBON-γ5β'2a/β'2mp/β'2mp_bilateral and
MBON-γ2α'1 are important for various forms of associative memory
formation. These diverse waking behaviors that involve the activity of
sleep-regulating neurons raises the interesting question whether such
roles are independent, or causally linked, which future studies can
address .
Daily sleep cycles in humans are driven by a complex circuit within which GABAergic sleep-promoting neurons oppose arousal. Drosophila sleep has recently been shown to be controlled by GABA, which acts on unknown cells expressing the Rdl GABAA receptor. This study has identified the relevant Rdl-containing cells as PDF-expressing small and large ventral lateral neurons (LNvs) of the circadian clock. LNv activity regulates total sleep as well as the rate of sleep onset; both large and small LNvs are part of the sleep circuit. Flies mutant for pdf or its receptor are hypersomnolent, and PDF acts on the LNvs themselves to control sleep. These features of the Drosophila sleep circuit, GABAergic control of onset and maintenance as well as peptidergic control of arousal, support the idea that features of sleep-circuit architecture as well as the mechanisms governing the behavioral transitions between sleep and wake are conserved between mammals and insects (Parisky, 2008).
Using a variety of mutants and novel genetic strategies to manipulate chronic and acute circuit activity, this study has shown that a small set of circadian clock cells in Drosophila has a critical role in the GABAergic initiation and maintenance of sleep. New genetic tools (dnATPase, ShawRNAi), were developed that allow an increase in the chronic response of neurons to their endogenous inputs. This adds greatly to the arsenal of activity-manipulating tools, most of which suppress firing or neurotransmitter release. Bidirectional manipulation of activity provides much more information about circuit function and dynamics. The utility was demonstrated of a new tool for acute activity manipulation (dTrpA1), which can be used on small numbers of neurons deep within the fly brain. The data suggest a model in which the pdf-GAL4-positive large LNvs (l-LNvs) translate light inputs (and perhaps other arousal signals) into wakefulness. The release of PDF from these cells is required, and l-LNv PDF signals to the smaller s-LNvs. The data demonstrating somnolence after downregulation of PDFR in LNvs indicates that s-LNvs participate in sleep control, although experiments in which they have been ablated suggest that they are not be the only sleep-relevant l-LNv targets. PDF signaling to PDFR-expressing neurons outside the clock that directly control activity is likely to be important. GABA may modulate the ability of LNvs to suppress sleep by acting on either or both s- and l-LNvs (Parisky, 2008).
In mammals, the role of the circadian clock in sleep is not completely understood. It is nonetheless clear that there are genetic (e.g., familial advance sleep phase syndrome) and environmental (e.g., jet-lag, shift work) conditions that disrupt sleep despite primarily affecting the circadian rhythms. The clock has been shown to regulate both when an animal sleeps and how much sleep occurs. The current consensus view is that the mammalian clock is primarily wake-promoting, acting along with the homeostatic sleep drive to shape sleep over the day and night (Parisky, 2008).
The data indicate that in flies PDF and the circadian LNvs more generally regulate both the maintenance of sleep as well as the ability of flies to respond to the wake-promoting effects of light. Although these effects recall the role of the mammalian SCN in sleep regulation, there are few prior links between the Drosophila circadian clock and the regulation of fly sleep. The almost complete elimination of the difference in total sleep between subjective day and subjective night in the pdf01 background adds substantially to this connection, i.e., light regulation of sleep appears to be substantially circadian clock-mediated Therefore, the contribution of the circadian machinery and fly brain clock circuitry to the control of sleep will probably parallel the important role of the mammalian circadian clock and the SCN in sleep regulation (Parisky, 2008).
PDF neurons have been recently shown to be light-responsive, like some neurons of the mammalian SCN. The l-LNvs also act as the dawn photoreceptor for the clock, sending a reset signal each morning to the rest of the clock. There is also good evidence that fly cryptochrome responds directly to light in addition to influencing circadian timekeeping, and a cry mutant substantially decreases the PDF neuron acute light response. Therefore, some of the waking effects described in this study probably reflect a role of PDF cells on acute processes involving light stimulation. Indeed, the phenotypes of flies without PDF or with decreased LNv neuronal excitability resemble some of the acute effects of the loss of orexin/hypocretin in narcoleptic mice. PDF neurons are also regulated by GABAergic inputs, analogous to those from the basal forebrain that regulate orexin/hypocretin neurons (Parisky, 2008).
Despite these similarities, there are also important organizational differences between systems. Most notable is the wide distribution of sleep circuitry in mammals. There are for example many targets of sleep-promoting GABAergic neurons, and the role of the circadian clock may be largely modulatory. The sleep circuitry of flies is almost certainly more circumscribed and simpler. Indeed, the surprisingly large effects of manipulating Rdl in the 16 LNvs argue that they are a principal target of sleep promoting GABAergic neurons and constitute part of the 'core' sleep circuitry. The fact that activation of a subset of these cells, the l-LNvs, has an effect on sleep homeostasis, further suggests that these cells sit at the heart of the sleep circuit. The fly sleep circuitry may therefore have condensed mammalian stimulatory systems (e.g., histaminergic, cholinergic and adrenergic, as well as orexin) into a simpler and more compact region, which may even largely coincide with the sixteen PDF cells of the circadian circuit (Parisky, 2008).
A limited number of other fly brain regions have been proposed to contribute to fly sleep. Manipulations of a broad set of peptidergic (PHM+) cells indicate that peptidergic neurons other than PDF neurons are wake promoting. An attractive hypothesis is that some these other peptidergic cells reside in the pars intercerebralis, a group of neurohumoral cells shown to an important sleep output center. The targets of these cells may even overlap with the targets of LNvs, e.g. the ellipsoid bodies. The PDFR is a class II G-protein coupled receptor and is fairly promiscuous: PDF is the highest affinity ligand, but this receptor is also activated by DH31 and PACAP-38. Since peptidergic modulation may occur by 'volume' transmission instead of by direct synaptic contact, both LNv peptides and peptides from the pars could together affect this motor center to regulate sleep and activity. The role of the pars may be to inform the sleep generation machinery about nutritional and metabolic state, i.e., animals undergoing starvation exhibit hyperlocomotor activity that is believed to be evolutionarily useful as a method for finding food, and alteration of this pars-generated locomotor program affects sleep. The role of l-LNvs is clearly different from that of other PHM+ neurons, and their unique involvement in homeostatic sleep suggests they are central to sleep control (Parisky, 2008).
The only other brain region that has been implicated in Drosophila sleep regulation is the paired structure known as the mushroom bodies. These studies showed that GAL4-driven manipulation of signaling or of neurotransmitter release in this neuropil had complex effects on sleep, not inconsistent with a modulatory role for this sensory integration center. The exact mechanism of these effects is not clear, however, especially since all of the mushroom body GAL4 lines that were examined in this study also express in multiple subsets of clock cells (Parisky, 2008).
The small circuit this study describes presents a tractable model system for understanding the circuit-level control of sleep, the relationship between homeostatic and circadian control as well as the dynamics of sleep-wake transitions; the latter are critical to an understanding of episodic and age-related insomnia (Parisky, 2008).
In the first optic neuropil (lamina) of the fly's visual system, two interneurons, L1 and L2 monopolar cells, and epithelial glial cells show circadian rhythms in morphological plasticity. These rhythms depend on clock gene period (per) and cryptochrome (cry) expression. This study found that rhythms in the lamina of Drosophila may be regulated by circadian clock neurons in the brain since the lamina is invaded by one neurite extending from ventral lateral neurons; the so-called pacemaker neurons. These neurons and the projection to the lamina were visualized by green fluorescent protein (GFP). GFP reporter gene expression was driven by the cry promotor in cry-GAL4/UAS-GFP transgenic lines. It was observed that the neuron projecting to the lamina forms arborizations of varicose fibers in the distal lamina. These varicose fibers do not form synaptic contacts with the lamina cells and are immunoreactive to the antisera raised against a specific region of Schistocerca gregaria ion transport peptide (ITP). ITP released in a paracrine way in the lamina cortex, may regulate the swelling and shrinking rhythms of the lamina monopolar cells and the glia by controlling the transport of ions and fluids across cell membranes at particular times of the day (Damulewicz, 2011).
This study showed a single projection from the pacemaker cells in the brain to the lamina, in which several structural circadian rhythms have been detected. Moreover, this input probably originates from the 5th small LNv. Since the 5th s-LNv does not express PDF, this cell is different from the other LNvs. The possibility that this process originates from other clock cells, for example from the LNds, and extends to the aMe first, and next to the lamina cannot be excluded. A CRY-positive LNd, which is immunoreactive to ITP, could invade the lamina by passing the aMe first. This neuron, however, is also immunoreactive to sNPF, but the projection detected in the lamina is immunoreactive to ITP only. It indicates that this projection originates from the 5th s-LNv, which is immunoreactive to ITP but not to sNPF. This study examined GFP expression driven by cry-GAL4 in thin, 20 microm cryostat sections and thick 100 microm vibratom sections of the Drosophila brain. In most earlier studies on clock neurons and their projections, whole-mount preparations of the Drosophila brain were used, or the lamina was cut-off during preparation. Such procedures from previous studies meant that the very fine projection from the brain to the lamina could not be observed. This study detected the projection by using 20 microm sections and collecting confocal optical sections at a 1 microm interval (Damulewicz, 2011).
In several previous studies, it has been suggested that CRY is present in different types of clock neurons. These results have been obtained using various methods; cry-GAL4 driven GFP expression, cry mRNA in situ hybridization, immunolocalization and cry deletion mutants. Using cry-GAL4 line and 20 microm sections of the D. melanogaster brain, it was found that CRY is located in all s-LNvs, l-LNvs, LNds, DN1s and DN3s but is absent in DN2s and LPNs. These results only partly confirm the results of earlier studies. It has been shown that LNvs but only some DN1, and three or four from the six LNd are CRY - positive, while DN2, DN3 and LPNs are CRY-negative. One study did d not detect CRY in DN2s and DN3s, and in about half of the LNds and DN1, but cry promoter dependent reporter genes and cry mRNA can be detected in these neurons. In this study, all of LNds showed GFP fluorescence in the cry-GAL4 strain, but only 3-4 cells were found to be CRY-immunopositive using antibodies. In turn, using the in situ hybridization method, cry mRNA was not detected in those cells. Since the pattern of cry-GAL4 driven GFP expression depends on the transgene insertion site and whether the first intron of the transgene has been inserted, spatial and circadian regulation of cry was examined. A series of cry-GAL4 transgenes containing different portions of cry upstream and intron 1 sequences was examined. The first intron was shown to drive expression in eyes and antennae, and upstream sequences induce cry expression in brain clock neurons and in peripheral oscillators; in eyes and antennae. In addition, upstream sequences also induce expression of cry, in other non-clock cells in the optic lobe (Damulewicz, 2011).
The results obtained using various methods suggest that in the case of CRY, translation and cry transcription may be specifically regulated. CRY-positive labeling in the 4th LNd was observed in flies kept for 5 days in constant darkness. Flies kept longer in this condition brought on weak staining in one of the DN2 neurons. Thus, the level of CRY in this neuron may be very low, and the CRY level may only be detected after it has accumulated for several days in DD. It is possible, that in some of the LNds, DN1 and DN3 cry expression is very low and protein is undetectable by the immunohistochemistry method, or that cry mRNA is unstable and CRY protein is not synthesized. Among six LNds, three neurons, that show a strong signal of GFP in the brain cryostat sections used in this study, may correspond to CRY-positive cells detected in the studies of other authors. In turn, three LNds with weak GFP in these preparations may correspond to CRY immunonegative cells. These cells had about a 50% lower GFP level than the rest of the LNds at all time points, except at ZT4 when their GFP fluorescence was lower by 20% (Damulewicz, 2011).
Beside neurons, clock genes have also been detected in glial cells. A subpopulation of glial cells in the brain of Drosophila have rhythmic expression of per gene, and they are necessary for maintaining circadian locomotor activity. However, the presence of CRY in glia was not detected in this study. In the optic lobes, GFP driven by cry-GAL4 was observed in many non-clock cells in which the localization pattern was very similar to the distribution of glial cells. But these non-clock cells were not labeled with the antibody against REPO protein, a specific marker for glial cells. The REPO protein is required for glia development and differentiation and has been detected in all types of glia in the adult brain of Drosophila. The analysis of cry-GAL4 driven GFP and REPO immunolabeling showed no co-localization between CRY and REPO. However, in the close vicinity of GFP-positive cells, REPO-positive glial cells were observed. A similar result was obtained using the antibody against the Drosophila vesicular monoamine transporter (DVMAT), which enabled labeling the fenestrated glia in the optic lobe. These results suggest that CRY is present in non-clock neurons in the optic lobe, but not in glial cells (Damulewicz, 2011).
In addition to localization of cry-GAL4 driven GFP in cell bodies of neurons, GFP processes were also detected invading three neuropils in the optic lobe. In the medulla, a dense network of processes originate from DN3s and their terminals seem to form synaptic contacts with not-yet identified target cells. The regular network of processes was also detected in the lobula but their origin is unknown. The most interesting finding is the projection of CRY-positive processes to the lamina. Although the lamina showed robust circadian remodeling of neuron morphology, a circadian input had not been previously detected. In the lamina, per is probably expressed in the epithelial glial cells, however, maintaining the lamina structural rhythms also requires per expression in the retina photoreceptors and in the LNs (Damulewicz, 2011).
Beside PER, CRY is also important for circadian rhythms in the lamina. In an earlier study, it was shown that the circadian rhythm in morphological plasticity of L2 dendritic trees, is not present in per01 mutant while its phase depends on CRY. In cryb mutant, the pattern of daily changes in size of the L2 dendritic tree was different than in wild-type Canton-S flies. In males and females of Canton-S wild-type flies, the largest L2 dendritic tree was found at the beginning of the day. This daily pattern of the structural changes of L2 dendrite resembles the pattern of cry mRNA cycling in Drosophila heads and bodies, and in the 5th s-LNv detected in this study. Although the L2 dendritic tree is the largest at the beginning of the day in the distal lamina, its axon, as well as the axon of L1 monopolar cell, swell at the beginning of both day and night. These changes have been detected in the proximal lamina. Moreover, the α-subunit of the Na+/K+-ATPase and subunits of the V-ATPase also show diurnal changes in abundance in the lamina. Such an occurrence indicates that circadian rhythms in cell structural plasticity are correlated with rhythmic changes in the level of proteins involved in the transport of ions. The rhythm in the α-subunit of the Na+/K+-ATPase level is bimodal with two peaks; in the morning and in the evening. This pattern is changed in the cry0 mutant. It indicates that CRY is not only important for the maintenance of the daily pattern of morphological changes of the L2 dendritic tree but CRY also helps to maintain cycling of the Na+/K+-ATPase in the epithelial glial cells in the lamina (Damulewicz, 2011).
It is uncertain whether there is regulation of lamina rhythms by the brain pacemaker because connections between the pacemaker neurons in the accessory medulla and the lamina have not been observed. It was found, however, that rhythms in axon plasticity of neurons in the lamina are circadian, have two peaks (morning and evening) and are synchronized with locomotor activity. The present results now show, that thin neurite extends from the aMe and arborizes in the distal lamina. In the aMe, the s-LNvs are regarded as the main pacemaker cells maintaining circadian rhythms. The l-LNvs are involved in behavioral arousal and sleep. For these reasons, the LNvs are good candidates as oscillators controlling lamina rhythms. Moreover, all LNvs except the 5th s-LNv, express PDF which may synchronize central oscillators with each other and with peripheral ones. In the housefly, large PDF-immunoreactive neurons, similar to Drosophila's l-LNvs, have terminals in the lamina which show circadian structural changes. Moreover, these neurons cyclically release PDF that affects circadian plasticity in the lamina. In Drosophila, release of PDF from PDF-immunoreactive processes in the medulla, where these processes form a dense network of varicose processes, is also possible. These processes, however, do not extend to the lamina. In the present study, PDF immunolabeling of the newly described Drosophila's CRY-positive terminals in the lamina was negative. This does not exclude PDF action in the lamina, particularly when PDF receptors have been detected in non-neuronal cells between the lamina and the retina. PDF may diffuse in the lamina after release from terminals in the distal medulla (Damulewicz, 2011).
Ion transport peptide (ITP) and short neuropeptide F (sNPF) have been detected in the LNvs. Among the five s-LNvs, ITP was found in the 5th s-LNv, while sNPF was observed in four other s-LNvs which also express PDF. In the present study, ITP-immunoreactive fibers were detected, using the Schgr-ITP antisera, in the distal lamina, co-localized with cry-GAL4 driven GFP. The co-localization with ITP suggests that the projection into the lamina may originate from the 5th s-LNv. Little is known about the function of the 5th s-LNv. It has been suggested, that this neuron, together with LNds and some DN1s, drive the evening peak of D. melanogaster bimodal activity. The finding indicates a possible new function of the 5th s-LNv in regulating circadian structural rhythms in the lamina, since this neuron is immunoreactive to ITP. Like other peptides in the optic lobe, ITP seems to be released from varicose terminals in a paracrine way. This conclusion was reached because no synaptic contacts between ITP-immunoreactive processes and cells in the lamina were detected. This peptide probably diffuses in the distal lamina and may facilitate chloride and/or other ion-dependent swelling and shrinking of the L1 and L2 axons. At least two ion pumps; the V-ATPase and Na+/K+-ATPase, show robust cyclical activity in the epithelial glial cells. The epithelial glial cells swell and shrink in anti-phase to the L1 and L2 interneurons. Preliminary results showed that in a transgenic line carrying RNAi to block ITP expression, the pattern of rhythmic changes in the level of the α-subunit of the Na+/K+-ATPase in the lamina glial cells of Drosophila is different than the pattern in wild-type flies. Thus, not only CRY but also ITP is important for maintaining rhythmic activity changes of the Na+/K+-ATPase (Damulewicz, 2011).
The function of ITP in the nervous system is unknown. In the lamina ITP may play a similar regulatory role as in hindgut of insects, transporting ions and fluids across cell membranes (Damulewicz, 2011).
The functions of sleep remain elusive, but a strong link exists between sleep need and neuronal plasticity. This study tested the hypothesis that plastic processes during wake lead to a net increase in synaptic strength and sleep is necessary for synaptic renormalization. In three Drosophila neuronal circuits it was found that synapse size or number increases after a few hours of wake and decreases only if flies are allowed to sleep. A richer wake experience resulted in both larger synaptic growth and greater sleep need. Finally, it was demonstrated that the gene Fmr1 (fragile X mental retardation 1) plays an important role in sleep-dependent synaptic renormalization (Bushey, 2011).
Sleep is present in every species that has been carefully studied, including Drosophila, but its functions remain elusive. Increasing evidence points to a link between sleep need and neuronal plasticity. A recent hypothesis suggests that a consequence of staying awake is a progressive increase in synaptic strength, as the awake brain learns and adapts to an ever-changing environment mostly through synaptic potentiation. However, such increase would soon become unsustainable, because stronger synapses consume more energy, occupy more space, require more supplies, and cannot be further potentiated, saturating the ability to learn. Thus, according to the synaptic homeostasis hypothesis, sleep may serve an essential function by promoting a homeostatic reduction in synaptic strength down to sustainable levels. Also, the hypothesis predicts that the more one learns and adapts (i.e., the more intense is the wake experience), the more one needs to sleep. Findings in rodents are consistent with this hypothesis. For instance, molecular and electrophysiological markers of synaptic strength are higher after wake and lower after sleep. Moreover, presynaptic terminals of hypocretin neurons in zebrafish larvae undergo both circadian and sleep-wake-dependent structural changes, the latter consistent with sleep-dependent down-regulation. Finally, in the fly brain, overall levels of synaptic proteins increase after wake and decrease after sleep (Gilestro, 2009), and synaptic structural changes have been described after very long sleep deprivation (Donlea, 2009). These results suggest that a role for sleep in synaptic homeostasis may hold in phylogenetically distant species and may thus be of general importance (Bushey, 2011).
The evidence in support of the synaptic homeostasis hypothesis is mainly correlative, and thus it is important to seek direct proof that sleep is necessary for synaptic renormalization and to do so at the level of individual synapses. Moreover, the synaptic homeostasis hypothesis predicts that behavioral paradigms that enhance wake-related plasticity in specific neural circuits should increase synaptic strength in those circuits as well as sleep need, but this prediction has never been tested. Finally, the cellular mechanisms that underlie synaptic and sleep changes remain unexplored. This study exploited the power of Drosophila genetics, combined with confocal microscopy and behavioral analysis, to address these questions (Bushey, 2011).
Changes in synaptic strength are often associated with changes in synaptic structure, including synapse number and size, although the link between structural and functional plasticity is complex. In mammals, the diameter and length of synaptic spines correlate with the size of the postsynaptic density and with the magnitude of electric signals transmitted to the dendritic shaft. Moreover, the induction of synaptic potentiation leads to growth of synapses and spines, whereas synaptic depression causes synapses and spines to retract or shrink. Similarly, in Drosophila, synaptic morphology at the neuromuscular junction changes depending on experience, and these changes correlate with synaptic strength. Previous in vivo experiments in mammals and flies measured overall changes in electrophysiological and molecular markers of synaptic strength, without cellular resolution, and without direct evidence for morphological changes in synaptic terminals. Three specific cell populations in the fly brain were selected, and it was asked whether sleep and wake affect synaptic density and size (Bushey, 2011).
The first cell group studied included the small ventral lateral neurons (LNvs), a subset of circadian oscillator neurons that are part of the wake promoting system and express the neuropeptide pigment dispersing factor (PDF). To visualize changes in presynaptic morphology, a fusion protein between synaptotagmin and enhanced GFP (syt-eGFP) was expressed, whose protein product colocalizes with native synaptic vesicles. PDF expression was also measured, because the latter is another marker of presynaptic boutons in small LNvs. First, adult females (7 days old) collected either during the light period were tested after 7 hours of mainly (>75%) spontaneous wake or during the dark period after 7 hours of mostly sleep (>80%) or sleep deprivation (>90%). Syt-eGFP and PDF staining were both higher in the presynaptic region of sleep-deprived and spontaneously awake flies relative to sleeping flies, whereas no differences were found in the axonal processes extending from the cell bodies to the presynaptic region, suggesting that the changes are independent of circadian time and specific to the presynaptic terminal. Males were then tested because they have less consolidated wake during the day than females. Flies were only tested at night, after sleep or sleep deprivation. Sleep-deprived 3- and 7-day-old males consistently showed higher presynaptic syt-eGFP and PDF staining than sleeping flies. In contrast, 1-day-old flies showed low syt-eGFP and PDF staining after both sleep and sleep deprivation. The lack of PDF staining in very young flies suggests that these neurons are still inactive soon after eclosure. Moreover, because PDF promotes arousal, low PDF staining is consistent with flies being predominantly asleep after eclosure, even if mechanical stimulation was used to try to keep them awake, consistent with high sleep need and elevated arousal threshold in newborn mammals. Syt-eGFP staining did not change in newly eclosed flies, whose PDF levels were very low. Syt-eGFP and PDF expression were also measured in Per01 flies carrying a null mutation of the clock gene Period. Because Per01 mutants have no spontaneous consolidated sleep, flies were collected immediately after 7 hours of sleep deprivation or after 5 additional hours of either recovery sleep or sleep deprivation. Overall, syt-eGFP and PDF staining in presynaptic terminals was reduced in Per01 mutants relative to wild-type (WT) flies but was still high after both 7 and 12 hours of sleep deprivation and low after recovery sleep (Bushey, 2011).
The second cell group analyzed included γ neurons of the mushroom bodies, because they can be targeted by mosaic analysis with a repressible cell marker (MARCM) to visualize single cells, show a relatively simple morphology, and undergo activity-dependent pruning. Moreover, the mushroom bodies are involved in sleep regulation, and mutations altering cyclic adenosine monophosphate-dependent protein kinase signaling or Fmr1 (fragile X mental retardation 1) expression in these brain regions affect both sleep need and experience-dependent structural plasticity . Flies were collected at night after 7 hours of sleep or sleep deprivation, and dissected brains were immunostained for GFP-tagged CD8 to visualize neuronal membranes. It was found that the axonal tips were larger after sleep deprivation than after sleep, consistent with an increase in volume of presynaptic terminals. To confirm this result, fly stocks were generated with γ MARCM clones expressing syt-eGFP, and flies were collected after 7 hours of mostly spontaneous wake, or during the dark period after 7 hours of mostly sleep or sleep deprivation. As expected, syt-eGFP tended to accumulate in puncta along lightly stained processes, in contrast to the diffuse CD8-GFP staining. Syt-eGFP puncta were larger in sleep deprived and spontaneously awake flies relative to sleeping flies (Bushey, 2011).
Next, whether postsynaptic morphological changes also occur as a function of sleep and wake was tested. To do so, focus was placed on the first giant tangential neuron of the lobula plate vertical system (VS). This cell (VS1) is unambiguously recognizable, and its stereotyped dendritic tree shows small actin-enriched protrusions morphologically and functionally similar to mammalian dendritic spines. Flies were compared that were spontaneously awake during the day or that slept or were sleep deprived during the first 7 hours of the night. Single VS1 spines were visualized using an antibody against actin-GFP and counted in one easily identifiable branch. The total number of spines was similar in spontaneously awake and sleeping flies but increased after sleep deprivation relative to both conditions, mainly because of an increase in stubby spines (which were the majority of scored spines). The number of mushroom spines did not change. The increase in spine number after sleep loss was associated with increased branching and lengthening of the dendritic tree, whereas spine density (number of spines divided by branch length) was similar in all conditions. Because sleep-deprived female flies had been mostly awake during the previous light period, this suggests that these postsynaptic changes may need sustained periods of wake. Another possibility, not mutually exclusive, is that changes in VS1 spines require a wake condition richer than that experienced by flies spontaneously awake alone inside small glass tubes. Indeed, sleep-deprived flies were kept awake using vibratory stimuli, resulting in the flies often falling from the top to the bottom of the tubes. Because visually driven responses in VS neurons are stronger during flight than during nonflight, it is possible that these cells were activated by the fall (Bushey, 2011).
To test whether a rich wake experience that engages the VS circuit is sufficient to affect VS1 synaptic morphology, up to 100 flies were housed inside a large lighted chamber ('fly mall') for an entire light period (12 hours). In the mall, flies could fly ad libitum, explore, and interact with each other. Flies were collected immediately after the mall experience and compared with flies that, as usual, had remained awake during the day in single tubes. The enriched experience in the mall had profound morphological effects on the VS1 dendritic tree: Total branch length increased because of the addition of more branches with spines (mainly stubby), resulting in an overall increase in spine number (Bushey, 2011).
Once experience-dependent synaptic changes have occurred, are they stable? If not, is sleep necessary to bring synaptic morphology back to pre-enrichment levels? To answer these questions, two other groups of flies were moved back to single tubes after 12 hours of mall experience; one group was allowed to sleep for 7 hours, whereas the other was kept awake as before using mechanical stimuli. In flies that were sleep-deprived after enrichment, branch length, branch points, and spine number were at levels similar to those seen in flies collected immediately after enrichment. In contrast, in flies that were allowed to sleep after the mall experience, all morphological parameters reverted to the levels observed in awake flies kept in single tubes. Moreover, spine density was negatively correlated with the amount of sleep during the last 7 hours, as well as with the maximal duration of sleep bouts. In another experiment, flies were housed in the mall for 12 hours during the day and then moved back to single tubes to record their sleep. During the 24 hours after the enrichment, flies slept more, both during the day and at. Finally, in the last experiment, flies were housed in the mall for 12 hours during the day, moved back to single tubes and sleep deprived all night (12 hours), and then either collected immediately, allowed to sleep for 6 hours, or kept awake for 6 more hours. Consistent with the previous experiments, decreases in all morphological parameters were seen only in flies that could sleep, and spine density was negatively correlated with the amount of sleep during the last 6 hours, as well as with mean and maximal duration of sleep bouts (Bushey, 2011).
Previous experiments suggest that Fmr1 could mediate at least some of the effects of sleep/wake on synapses. Fmr1 protein product (FMRP) is present in dendritic spines, and loss of FMRP in flies is associated with overgrown dendritic trees, larger synaptic boutons, and defects in developmental and activity-dependent pruning. Notably, Fmr1 overexpression results in the opposite phenotype, with dendritic and axonal underbranching and loss of synapse differentiation. Moreover, Fmr1 expression is reduced by sensory deprivation in flies and increased by sensory stimulation and enrichment in mammals (Bushey, 2011).
It was recently shown that FMRP levels increase in the adult fly brain during wake relative to sleep, independent of time of day or light, suggesting that waking experience is sufficient to affect Fmr1 expression even after the end of development. It has also been shown that Fmr1 overexpression in either the whole brain or in the mushroom bodies is associated with an ~30% decrease in sleep duration, and it is hypothesized that this reduced need for sleep occurs because chronically high Fmr1 levels may allow synaptic pruning to occur at all times, independent of sleep. If so, Fmr1 overexpressing (OE) flies should fail to show increased spine density after prolonged wake. Thus Fmr1 was overexpressed specifically in the vertical and horizontal system of the lobula plate. OE flies were collected at night after 7 hours of either sleep or sleep deprivation and were compared to corresponding sleeping and sleep-deprived WT controls. As expected, Fmr1 expression was concentrated in granules along the VS1 dendritic tree, and overall Fmr1 levels were higher in sleeping and sleep-deprived OE flies than in their corresponding controls, due to larger Fmr1 granules in OE flies. Crucially, in contrast to WT controls, OE flies showed no increase in either spine number, branch length, or branch points after sleep deprivation relative to sleep; all these parameters were similar between the two experimental groups, and their levels were close to those observed in WT flies after sleep. Finally, OE flies slept less than their WT controls during baseline and showed a reduced sleep rebound after 12 hours of sleep deprivation at night. Thus, it seems that Fmr1 overexpression was sufficient to completely abolish the wake-dependent increase in VS1 spine number, whereas the effects on sleep were small. The latter result is not surprising, because sleep need presumably results from the overall amount of synaptic plasticity occurring during wake in many brain areas, whereas Fmr1 overexpression was restricted to a few VS neurons (Bushey, 2011).
Sleep is perhaps the only major behavior still
in search of a function. The results of this study support the
hypothesis that plastic processes during wake lead to a net increase
in synaptic strength in many brain circuits and that sleep is required
for synaptic renormalization. A wake-related increase in synapse
number and strength, if unopposed, would lead to a progressive
increase in energy expenditure and saturation of learning. A
sleep-dependent synaptic homeostasis may explain why sleep is required
to maintain cognitive performance. How sleep would bring about a net
decrease in synaptic strength remains unknown, but in mammals,
potential mechanisms favoring synaptic depression during non-rapid eye
movement sleep may require the repeated sequences of
depolarization/synchronous firing and hyperpolarization/silence at
~1Hz observed in corticothalamic cells, as well as the low levels of
neuromodulators such as noradrenaline and of plasticity-related
molecules such as brain-derived neurotrophic factor. To what extent
such mechanisms may also apply to flies remains to be determined
(Bushey, 2011).
Tetrad synapses are formed between the retina photoreceptor terminals and postsynaptic cells in the first optic neuropil (lamina) of Drosophila. They are remodelled in the course of the day and show distinct functional changes during activity and sleep. These changes result from fast degradation of the presynaptic scaffolding protein Bruchpilot (BRP) by Cryptochrome (CRY) in the morning and depend on BRP-170, one of two BRP isoforms. This process also affects the number of synaptic vesicles, both clear and dense-core, delivered to the presynaptic elements. In cry01 mutants lacking CRY and in brpΔ170, the number of synaptic vesicles is lower in the morning peak of activity than during night-sleep while in wild-type flies the number of synaptic vesicles is similar at these two time points. CRY may also set phase of the circadian rhythm in plasticity of synapses. The process of synapse remodelling stimulates the formation of clear synaptic vesicles in the morning. They carry histamine, a neurotransmitter in tetrad synapses and seem to be formed from glial capitate projections inside the photoreceptor terminals. In turn dense-core vesicles probably carry synaptic proteins building the tetrad presynaptic element (Damulewicz, 2020).
The results confirmed earlier studies that the presynaptic element (T-bar) of tetrad synapses is remodelled during the day and night and this rhythm is regulated by light and circadian clock (Gorska-Andrzejak, 2013; Woznicka, 2015). In the present study, it was found additionally that cyclic changes occur in the T-bar ultrastructure and its volume. Ultrastructural changes in T-bars and synaptic vesicles were possible because of using high resolution electron microscope tomography (EMT). In the present study it was also found that the number of synaptic vesicles cycles during the day, but this rhythm is masked by light. This result indicates that in the case of tetrad synapses, which are sites of fast neurotransmission between photoreceptors and the first order interneurons, intense neurotransmission occurs during the morning peak of locomotor activity, when more synaptic vesicles are attached to the T-bar platform than during sleep (ZT16) and are transported from capitate projections located next to the T-bars. At ZT16, tetrad synapses are ready for neurotransmission, since the total number of synaptic vesicles near tetrad synapses is similar at ZT1 and ZT16, but it is in a standby mode with lower frequency of synapses and vesicles, which are not attached to the T-bar platform and are not delivered from glial cells, respectively. However, transmission can be activated and is efficient because synapses during sleep have larger volume, and synaptic vesicles can be transported to the T-bar platform, if necessary, in response to an unexpected light pulse. Cyclic remodelling of tetrad presynaptic sites depends on BRP, which must be delivered to T-bars and degraded after light exposure after binding to CRY. This fast remodelling of synapses affects the number of vesicles transported to the presynaptic element (Damulewicz, 2020).
The present study was carried out only in light/dark (LD 12:12) conditions because in earlier studies it was found that the rhythm of the changes in BRP abundance in tetrad synapses of D. melanogaster is circadian. The rhythm is maintained in constant darkness (DD) and abolished in the per01 clock null mutant. On the basis of these results, it is assumed that rhythmic changes during the 24 h cycle reported is this paper are also circadian; however, in DD, the morning peak in BRP is not present because it depends on light (Damulewicz, 2020).
Electron microscope tomography (EMT) used in this study showed that synaptic vesicles are attached to the platform of the T-bar with filamentous proteins that are reduced in brpΔ170 and brpΔ190 mutants, which have fewer synaptic vesicles compared with wild-type flies. Two types of vesicles, clear and dense-core were detected. Dense-core synaptic vesicles were less numerous than clear ones. It is known that synaptic vesicles of tetrad synapses contain histamine, while the content of dense-core vesicles is unknown. It is possible that they carry presynaptic proteins to the T-bar. The comparison of ultrastructure of tetrad synapses in the morning peak of activity (ZT1) and during sleep (ZT16) indicates that more vesicles are attached to the T-bar platform and to capitate projections at ZT1, but this pattern is not present in brpΔ170, brpΔ190 and cry01 . This result confirmed an earlier study that both high motor activity in the morning and light exposure increase activity of the visual system. In the morning, there is intense transport of synaptic vesicles to T-bars and delivery of histamine in clear vesicles from the epithelial glial cells through capitate projections. The evidence for a role of capitate projections in neurotransmitter recycling has already been reported and now this study showed, using EMT, that vesicles are produced from capitate projections and directly delivered to T-bars. This intense transport is damaged in all mutants studied, suggesting that both BRP isoforms and CRY are needed for this process. In addition, in the brpΔ190 mutant lacking BRP-190, the T-bar structure is less dense than in the other strains studied. In a previous study, it was found that there are approximately 50% fewer synapses in brpΔ190 than in Canton S and brpΔ170. Another study reported that BRP isoforms are important for the formation of T-bars in neuromuscular junctions, and in brp mutants T-bars are smaller than in controls. T-bar height was reduced in brpΔ190, whereas the widths of pedestal and platform were reduced in both mutants. They also decrease transmission since the active zone was smaller in both mutants and the number of synaptic vesicles was reduced. These ultrastructural changes are correlated with cell physiology since the amplitude of evoked excitatory junctional current was decreased in both mutants with a stronger effect in brpΔ190 (Damulewicz, 2020).
The obtained reconstructions of tetrad T-bars from TEM serial sections of the lamina showed that although there were fewer synapses during the night (ZT16), the volume of the T-bar was larger at that time than in the morning (ZT1), while the total number of synaptic vesicles was similar. In contrast, a day/night difference (ZT1 vs. ZT16) in the number of vesicles was observed in brpΔ170 and cry01 . This suggests that the CRY protein and BRP-170 are responsible for an increase in the number of synaptic vesicles during the morning peak of activity. Since CRY co-localizes with BRP and is involved in BRP degradation in the morning, CRY is probably also involved in the degradation of other proteins of synaptic vesicle organization in the morning since the number of vesicles in cry01 was low in the morning but high at night (ZT16). It seems that in cry01, synaptic vesicles are not delivered to the photoreceptor terminals from capitate projections and tethered to the cytomatrix in the morning. It is also interesting that the daily rhythm in the number of vesicles is not maintained in BRP mutants, which confirms an earlier study, and the BRP N-terminus, which lacks brpΔ190, is necessary to maintain daily remodelling of the T-bar structure. Although both isoforms participate in building the cytomatrix their functions seem to be different in the course of the day. It is also possible that CRY is not only responsible for degradation of synaptic proteins but also as a protein, what is known, affecting the clock. In another cell types, in clock neurons l-LNvs, CRY, except interaction with TIM, is responsible for blue light response and firing of the l-LNvs. In the lamina it was found that in cry01 mutant the daily rhythm in synapse number and their remodelling was delayed in phase and the day/night difference in Canton S increased when peaks in the number of synapses were shift to ZT4 and ZT16. In result the difference between ZT1 and ZT16 was increased in cry01 (Damulewicz, 2020).
BRP is also responsible for rapid remodelling of the presynaptic active zone (AZ), and as reported in Drosophila NMJ, presynaptic homeostatic potentiation increases the number of BRP molecules and other AZ proteins, Unc13A and RBP, within minutes (Damulewicz, 2020).
When synaptic vesicles were counted at two different distances from the T-bar, to 200 nm and above 200 nm, there were differences between clear vesicles containing histamine and dense-core ones located in both areas. More clear vesicles were located near the T-bar and fewer above 200 nm. In the case of dense-core vesicles, their number was similar in both areas. This difference was not so striking in mutants in the case of vesicles located next to the platform, but in brpΔ170 and cry01, there were more dense-core vesicles at ZT16 in the distance above 200 nm than closer to the T-bar. This result indicates that BRP-170 and CRY are important for the distribution of clear synaptic vesicles next to the T-bar as well as dense-core vesicles located above 200 nm from the presynaptic element. It is possible that dense-core vesicles contain T-bar proteins, probably BRP. When transport along the actin cytoskeleton is disrupted, the number of tetrad synapses decreases, and rapid AZ remodelling also fails (Damulewicz, 2020).
The above mentioned ultrastructural changes depend on the level of the presynaptic scaffolding protein BRP, which changes in abundance during the day and night. These changes are controlled by the daily expression of CRY, which seems to have many functions in photoreceptors in addition to being the circadian clock photoreceptor. In an earlier study, it was found that CRY interacts with BRP but only during light exposure and leads to the degradation of BRP during the day/light phase of the 24 h cycle. This seems to be responsible for the decrease in BRP level in the middle of the day after its peak at the beginning of the day. The lack of CRY in cry01 mutants changes the pattern of the tetrad presynaptic profile frequency during the day and the size of the T-bar. However, the rhythm is not completely abolished, which indicates that other proteins are also involved in the daily remodelling of tetrad synapses. Since CRY plays several functions in photoreceptors, changes in the number of tetrad synapse and T-bar size in cry01 may result from different processes and lack of interactions of CRY with TIM and BRP. CRY is a component of the molecular clock and interacts with TIM, and this may affect daily changes in the number and size of T-bars. In addition, light-activated CRY binds BRP and targets it to degradation. Previous work showed that flies with constitutively active CRY have low BRP level. In turn, cry01 mutants show changes in the pattern of BRP expression, with higher BRP level during the day (ZT4), at the time when wild-type flies have minimum of BRP expression. The pattern of BRP expression is similar to the pattern of daily changes in tetrad synapse number, so it is possible that the number of synapses or T-bar size is directly dependent on the amount of BRP which oscillates during the day. However, CRY in the retina photoreceptors binds also actin and is involved in the organization of phototransduction cascade of proteins in rhabdomeres38. This may be also involved in the regulation of T-bar structure. The differences in T-bar size of cry01 are shown at time when in Canton S CRY is active (during the day) or its level increases (ZT16). At the beginning of the night the level of CRY is very low, so there was no effect on T-bar structure and no difference between CS and cry01 was observed (Damulewicz, 2020).
The epithelial glial cells are important for many processes during phototransduction and in recycling neurotransmitters and other compounds during the night. Glia take up histamine and metabolize it to carcinin, which is next delivered to the photoreceptor terminals, and capitate projections are involved in this process. Activity of glial cells is also controlled by the circadian clock. During the night, glial cells seem to be more active than neurons to recycle neurotransmitters, and many proteins, including proteins of ion pumps, are found at higher concentrations at that time. The high number of synaptic vesicles near the tetrad T-bar during the morning peak of activity in Drosophila seems to depend on capitate projections invaginating from the epithelial glia to the photoreceptor terminals in the lamina of Drosophila (Damulewicz, 2020).
Although the presynaptic cytomatrix can be rapidly remodelled with transmission strength, it is also affected by motor and visual system activity, external factors, such as light in the case of the visual system, and the circadian clock, showing plasticity and correlation to changes in behaviour during the day/night cycle. As was shown in the present study synaptic plasticity and synapse remodelling during the day is a complex process which involves presynaptic proteins of the T-bar as well as two types of synaptic vesicles, clear and dense-core, and glial cells. It was also found that fast degradation of proteins involved in transmission is as important as pre- and postsynaptic protein synthesis (Damulewicz, 2020).
Neuropeptide PDF (pigment-dispersing factor)-secreting large ventrolateral neurons (lLNvs) in the Drosophila brain regulate daily patterns of rest and arousal. These bilateral wake-promoting neurons are light responsive and integrate information from the circadian system, sleep circuits, and light environment. To begin to dissect the synaptic circuitry of the circadian neural network, simultaneous dual whole-cell patch-clamp recordings of pairs of lLNvs were performed. Both ipsilateral and contralateral pairs of lLNvs exhibit synchronous rhythmic membrane activity with a periodicity of ~5-10 s. This rhythmic lLNv activity is blocked by TTX, voltage-gated sodium blocker, or α-bungarotoxin, nicotinic acetylcholine receptor antagonist, indicating that action potential-dependent cholinergic synaptic connections are required for rhythmic lLNv activity. Since injecting current into one neuron of the pair had no effect on the membrane activity of the other neuron of the pair, this suggests that the synchrony is attributable to bilateral inputs and not coupling between the pairs of lLNvs. To further elucidate the nature of these synaptic inputs to lLNvs, a variety of neurotransmitter receptors were blocked or activated, and effects on network activity and ionic conductances were measured. These measurements indicate the lLNvs possess excitatory nicotinic ACh receptors, inhibitory ionotropic GABAA receptors, and inhibitory ionotropic GluCl (glutamate-gated chloride) receptors. It was demonstrated that cholinergic input, but not GABAergic input, is required for synchronous membrane activity, whereas GABA can modulate firing patterns. It is concluded that neuropeptidergic lLNvs that control rest and arousal receive synchronous synaptic inputs mediated by ACh (McCarthy, 2011).
Previously studies have shown that neural circuits responsible for generating circadian rhythms and also those neural networks controlling rest and arousal exhibit synchronous membrane activity both in mammals and in insects. Furthermore, neuropeptides, VIP and PDF, in mammals and flies, respectively, and the classical neurotransmitter, GABA, play critical roles in this synchrony. Furthermore, this study found that stereotyped neurons that were positive for a well studied driver exhibited varying degrees of synchrony with lLNv membrane activity. This is consistent with a model in which certain neurons receive some of the same inputs as lLNvs and some unique inputs. Similar to the observations in Drosophila, neurons in some mammalian brain regions outside the suprachiasmatic nucleus (SCN) exhibit synchronized membrane activities with SCN neurons. The data do not, however, preclude the possibility that the synchrony that was observe is attributable to widespread epileptiform or other widespread synchronous brain activity that is not specific to lLNvs. This explanation is not favored because lLNv pairs exhibiting varying degrees of synchrony were observed and also lLNv pairs in which one cell is burst firing while the other is tonically firing, as shown in the paired recording pharmacology experiments. Even if it is the case that the synchronized activity that was seen is attributable to some sort of epileptiform or other widespread synchronous activity, the nature of and mechanisms underlying this activity are still informative, as it provides insight into the connectivity of the network (McCarthy, 2011).
To characterize the nature of synaptic inputs to lLNvs, a combination of agonists and antagonists against neurotransmitter receptors were used in both current-clamp and voltage-clamp mode. Current-clamp mode was used to monitor modulation of membrane activity in the lLNvs in the context of the functional neural network, whereas voltage-clamp mode was used to determine the presence of underlying receptors in the lLNvs themselves. lLNvs were found to receive excitatory cholinergic input through nAChR. Treatment of brains with cholinergic receptor agonists, acetylcholine and nicotine, enhances membrane activity, depolarizing the neurons and increasing action potential firing rate, whereas treatment with ACh receptor antagonists, curare and a-BuTX, inhibits membrane activity. Voltage-clamp recordings in the presence of TTX revealed that ACh- and nicotine-induced currents occur in lLNvs themselves, and these currents reverse near the equilibrium potential for nonselective monovalent cation channels, as expected for currents through nicotinic acetylcholine receptors. In insects, acetylcholine is the primary excitatory neurotransmitter in the CNS, and nAChRs are widely expressed in the Drosophila brain. These receptors are known to mediate fast synaptic transmission in Kenyon cells in the adult mushroom body. A previous study has also shown that dissociated PDF-positive neurons from the larval Drosophila brain, which are developmental precursors of the LNvs, express nAChRs and exhibit both ACh-induced and nicotine-induced increases in intracellular calcium that are dependent on both external sodium and calcium concentrations. The current findings confirm that lLNvs in the adult circadian neural network possess nAChRs and that these receptors mediate excitatory synaptic input and synchrony of rhythmic firing (McCarthy, 2011).
GABA is a major neurotransmitter in the Drosophila CNS, mediating fast inhibitory synaptic transmission through the GABAA receptor. This receptor has been shown to be expressed in LNvs and has been shown genetically to play a major role in the regulation of arousal and sleep by lLNvs specifically (Parisky, 2008; Shang, 2008; Chung, 2009). GABA-induced decreases in intracellular calcium and Cl- currents have been recorded in dissociated PDF neurons from the larval and adult fly brain, respectively, but previous studies have not analyzed effects of GABA on lLNv membrane activity in the context of the intact circadian rest/arousal control network. This study demonstrated that GABA inhibits the membrane activity of lLNvs, whereas the ionotropic GABAR antagonist, picrotoxin, is excitatory. In contrast, another study has shown no rescue of the inhibitory effect of GABA by picrotoxin on PDF+ LNv precursors but does show alleviation of GABA-induced inhibitory responses in these neurons by metabotropic GABABR antagonists. Although the current studies do not exclude a role for GABABRs in lLNvs, the discrepancy of the effect of picrotoxin could be attributable to changes in different GABAR subtype expression at different developmental stages (McCarthy, 2011).
Through an extensive series of voltage-clamp experiments it was determined that GABA induces currents in lLNvs that reverse at the equilibrium potential of Cl-, indicating that these currents are mediated by GABAARs. These data demonstrate that lLNvs express the GABAAR and that lLNvs receive GABAergic inhibitory synaptic input. These data from paired recordings in lLNvs show that GABAergic synaptic input, in conjunction with PDF signaling, plays a critical role in modulating the membrane activity of lLNvs but is not required for the robust synchrony of firing in these neurons, as application of picrotoxin does not abolish synchronous firing. Conversely, in the cockroach, picrotoxin leads to desynchrony within circadian neural networks. In this system, PDF also serves to synchronize these neural populations by inhibiting GABAergic interneurons. This mechanism does not seem to be conserved in Drosophila, but additional experiments are needed to elucidate the effect of PDF on the synchronous electrical activity of the circadian neural circuit (McCarthy, 2011).
Glutamate and its excitatory ionotropic receptors, homologs of the AMPA, kainate, and NMDA receptors in mammals, have been shown to mediate fast excitatory neurotransmission at the neuromuscular junction (NMJ) in Drosophila. Interestingly, the current data demonstrate that treatment of lLNvs with glutamate led to an inhibition of membrane activity, which is opposite to the effect seen at the NMJ. Through voltage-clamp experiments, it was shown that this glutamate-induced current in lLNvs reverses near the equilibrium potential of Cl-. Furthermore, when the Cl- concentration of the external solution was altered and the reversal potential of the current was measured, the experimental value was well predicted by the calculated equilibrium potential for each specific Cl- concentration. These data together indicate that lLNvs possess a glutamate-gated Cl- channel. Members of the GluCl family have been cloned from bothDrosophila and C. elegans but have not been found in vertebrate species. Their functional roles in neural circuits in Drosophila remain enigmatic. The current studies indicate these channels are present in lLNvs, which also express metabotropic glutamate receptors. Their role in synchronous membrane activity between lLNvs remains to be elucidated (McCarthy, 2011).
Given the variety of the synaptic inputs to lLNvs described in this study, the ability of lLNvs to autonomously detect light through the blue light-activated photopigment CRY, and the convergence of the arousal and circadian circuits on lLNvs, these neurons are clearly in a position to integrate complicated signals from all these systems. The data also show that the rhythmic oscillation in membrane activity seen in these neurons is most likely not attributable to intrinsic pacemaking, but instead arises from synchronized synaptic inputs, both excitatory and inhibitory. It remains to be determined where these cholinergic, GABAergic, and glutamatergic synaptic inputs converging on lLNvs originate. Previous studies have demonstrated that the Hofbauer-Büchner adult eyelets, which are derived developmentally from Bolwig's organ in the larvae, send axon bundles to the dendritic region on LNvs. These cholinergic neurons may provide excitatory input to lLNvs via nAChRs. However, it is not considered likely that Hofbauer-Büchner cholinergic inputs to the lLNvs contribute to rhythmic activity in the whole-brain explant. As far as anatomical characterization of the inhibitory inputs into lLNvs, varicosities in the accessory medulla, which abut lLNv dendrites, express glutamic acid decarboxylase, a marker for GABAergic neurons; however, it is not known where cell bodies reside from which these processes originate. In addition, it has been previously shown that other circadian clock neurons are glutamatergic. The axon terminals of these neurons are in close proximity to the dendritic arbors of the lLNvs in the larval optic center and in the accessory medulla of the adult fly. These data, in combination with the current findings, suggest that GluCl within lLNvs may mediate inhibitory synaptic inputs from other clock neurons in the circadian circuit (McCarthy, 2011).
Through the use of whole-cell patch-clamp electrophysiology techniques, this study has demonstrated synchronous membrane activity of lLNvs of the circadian rest/arousal neural network of Drosophila arising from bilateral synchronized synaptic inputs. This synchronous membrane activity is mediated by cholinergic inputs to the lLNvs themselves. However, GABAergic inputs modulate membrane activity of these neurons but are not required for synchrony. The role of glutamatergic signaling in synchronous membrane activity between lLNv pairs remains to be revealed, as agents to pharmacologically inhibit GluCl are not currently available. Building on these findings, future studies are required to elucidate the overlapping neural circuitry of the circadian, rest/arousal, and light input systems, and will discern how these systems are integrated and finely coordinated to generate a robust and complex pattern of behavior (McCarthy, 2011).
Arousal is fundamental to many behaviors, but whether it is unitary or whether there are different types of behavior-specific arousal has not been clear. In Drosophila, dopamine promotes sleep-wake arousal. However, there is conflicting evidence regarding its influence on environmentally stimulated arousal. This study shows that loss-of-function mutations in the D1 dopamine receptor DopR enhance repetitive startle-induced arousal while decreasing sleep-wake arousal (i.e., increasing sleep). These two types of arousal are also inversely influenced by cocaine, whose effects in each case are opposite to, and abrogated by, the DopR mutation. Selective restoration of DopR function in the central complex rescues the enhanced stimulated arousal but not the increased sleep phenotype of DopR mutants. These data provide evidence for at least two different forms of arousal, which are independently regulated by dopamine in opposite directions, via distinct neural circuits (Lebestky, 2009).
'Arousal', a state characterized by increased activity, sensitivity to sensory stimuli, and certain patterns of brain activity, accompanies many different behaviors, including circadian rhythms, escape, aggression, courtship, and emotional responses in higher vertebrates. A key unanswered question is whether arousal is a unidimensional, generalized state. Biogenic amines, such as dopamine (DA), norepinephrine (NE), serotonin (5-HT), and histamine, as well as cholinergic systems, have all been implicated in arousal in numerous behavioral settings. However, it is not clear whether these different neuromodulators act on a common 'generalized arousal' pathway or rather control distinct arousal pathways or circuits that independently regulate different behaviors. Resolving this issue requires identifying the receptors and circuits on which these neuromodulators act, in different behavioral settings of arousal (Lebestky, 2009).
Most studies of arousal in Drosophila have focused on locomotor activity reflecting sleep-wake transitions, a form of 'endogenously generated' arousal. Several lines of evidence point to a role for DA in enhancing this form of arousal in Drosophila. Drug-feeding experiments, as well as genetic silencing of dopaminergic neurons, have indicated that DA promotes waking during the subjective night phase of the circadian cycle. Similar conclusions were drawn from studying mutations in the Drosophila DA transporter (dDAT). Consistent with these data, overexpression of the vesicular monoamine transporter (dVMAT-A), promoted hyperactivity in this species, as did activation of DA neurons in quiescent flies (Lebestky, 2009).
Evidence regarding the nature of DA effects on 'exogenously generated' or environmentally stimulated arousal, such as that elicited by startle, is less consistent. Classical genetic studies and quantitative trait locus (QTL) analyses have suggested that differences in DA levels may underlie genetic variation in startle-induced locomotor activity (see Carbone, 2006 and Jordan, 2006). Fmn (dDAT; Dopamine transporter) mutants displayed hyperactivity in response to mechanical shocks, implying a positive-acting role for DA in controlling environmentally induced arousal (Kume, 2005). In contrast, other data imply a negative-acting role for DA in controlling stimulated arousal. Mutants in Tyr-1, which exhibit a reduction in dopamine levels, show an increase in stimulated but not spontaneous levels of locomotor activity. Genetic inhibition of tyrosine hydroxylase-expressing neurons caused hyperactivity in response to mechanical startle (Friggi-Grelin, 2003). Finally, transient activation of DA neurons in hyperactive flies inhibited locomotion (Lima, 2005). Whether these differing results reflect differences in behavioral assays, the involvement of different types of DA receptors, or an 'inverted U'-like dosage sensitivity to DA (Birman, 2005), is unclear (Lebestky, 2009).
This investigation has developed a novel behavioral paradigm for environmentally stimulated arousal, using repetitive mechanical startle as a stimulus, and a screen was carried out for mutations that potentiate this response. One such mutation is a hypomorphic allele of the D1 receptor ortholog, DopR. This same mutation caused decreased spontaneous activity during the night phase of the circadian cycle, due to increased rest bout duration. In both assays, cocaine influenced behavior in the opposite direction as the DopR mutation, and the effect of cocaine was abolished in DopR mutant flies, supporting the idea that DA inversely regulates these two forms of arousal. Genetic rescue experiments, using Gal4 drivers with restricted CNS expression, indicate that these independent and opposite influences of DopR are exerted in different neural circuits. These data suggest the existence of different types of arousal states mediated by distinct neural circuits in Drosophila, which can be oppositely regulated by DA acting via the same receptor subtype (Lebestky, 2009).
Previous studies of arousal in Drosophila have focused on sleep-wake transitions, a form of 'endogenous' arousal. This study has introduced and characterized a quantitative behavioral assay for repetitive startle-induced hyperactivity, which displays properties consistent with an environmentally triggered ('exogenous') arousal state. A screen was conducted for mutations affecting this behavior, the phenotype of one such mutation (DopR) was analyzed, and the neural substrates of its action was mapped by cell-specific genetic rescue experiments. The results reveal that DopR independently regulates Repetitive Startle-induced Hyperactivity (ReSH) and sleep in opposite directions by acting on distinct neural substrates. Negative regulation of the ReSH response requires DopR function in the ellipsoid body (EB) of the central complex (CC), while positive regulation of waking reflects a function in other populations of neurons, including PDF-expressing circadian pacemaker cells. Both of these functions, moreover, are independent of the function of DopR in learning and memory, which is required in the mushroom body. These data suggest that ReSH behavior and sleep-wake transitions reflect distinct forms of arousal that are genetically, anatomically, and behaviorally separable. This conclusion is consistent with earlier suggestions, based on classical genetic studies, that spontaneous and environmentally stimulated locomotor activity reflect 'distinct behavioral systems' in Drosophila (Lebestky, 2009).
Several lines of evidence suggest that ReSH behavior represents a form of environmentally stimulated arousal. First, hyperactivity is an evolutionarily conserved expression of increased arousal. Although not all arousal is necessarily expressed as hyperactivity, electrophysiological studies indicate that mechanical startle, the type of stimulus used in this study, evokes increases in 20-30 Hz and 80-90 Hz brain activity, which have been suggested to reflect a neural correlate of arousal in flies (Nitz, 2002; van Swinderen, 2004). Second, ReSH does not immediately dissipate following termination of the stimulus, as would be expected for a simple reflexive stimulus-response behavior, but rather persists for an extended period of time, suggesting that it reflects a change in internal state. Third, this state, like arousal, is scalable: more puffs, or more intense puffs, produce a stronger and/or longer-lasting state of hyperactivity. Fourth, this state exhibits sensitization: even after overt locomotor activity has recovered to prepuff levels, flies remain hypersensitive to a single puff for several minutes. Fifth, this sensitization state generalizes to a startle stimulus of at least one other sensory modality (olfactory). In Aplysia, sensitization of the gill/siphon withdrawal reflex has been likened to behavioral arousal. Taken together, these features strongly suggest that ReSH represents an example of environmentally stimulated ('exogenous') arousal in Drosophila (Lebestky, 2009).
DopR mutant flies exhibited longer rest periods during their subjective night phase, suggesting that DopR normally promotes sleep-wake transitions. These data are consistent with earlier studies indicating that DA promotes arousal by inhibiting sleep (Andretic, 2005, Kume, 2005; Wu, 2008). In contrast, prior evidence regarding the role of DA in startle-induced arousal is conflicting. Some studies have suggested that DA negatively regulates locomotor reactivity to environmental stimuli, consistent with the current observations, while others have suggested that it positively regulates this response. Even within the same study, light-stimulated activation of TH+ neurons produced opposite effects on locomotion, depending on the prestimulus level of locomotor activity (Lima, 2005; Lebestky, 2009 and references therein).
This study has found that DA and DopR negatively regulate environmentally stimulated arousal: the DopR mutation enhanced the ReSH response, while cocaine suppressed it. Furthermore, the effect of cocaine in the ReSH assay was eliminated in the DopR mutant but could be rescued by Gal4-driven DopR expression, confirming that the effect of the drug is mediated by DA. Taken together, these results reconcile apparently conflicting data on the role of DA in 'arousal' in Drosophila by identifying two different forms of arousal -- repetitive startle-induced arousal and sleep-wake arousal -- that are regulated by DA in an inverse manner (Lebestky, 2009).
The finding that DopR negatively regulates one form of environmentally stimulated arousal leaves open the question of whether this is true for all types of exogenous arousing stimuli. The 'sign' of the influence of DA on exogenously generated arousal states may vary depending on the type or strength of the stimulus used, the initial state of the system prior to exposure to the arousing stimulus (Birman, 2005; Lima and Miesenbock, 2005), or the precise neural circuitry that is engaged. Future studies using arousing stimuli of different sensory modalities or associated with different behaviors should shed light on this question (Lebestky, 2009).
Several lines of evidence suggest that endogenous DopR likely acts in the ellipsoid body (EB) of the central complex (CC) to regulate repetitive startle-induced arousal. First, multiple Gal4 lines that drive expression in the EB rescued the ReSH phenotype of DopR mutants. Second, endogenous DopR is expressed in EB neurons, including those in which the rescuing Gal4 drivers are expressed. Third, the domain of DopR expression in the EB overlaps the varicosities of TH+ fibers. In an independent study of dopaminergic inputs required for regulating EtOH-stimulated hyperactivity TH+ neurons were identified that are a likely source of these projections to the EB. Fourth, rescue of the ReSH phenotype is associated with re-expression of DopR in EB neurons. Finally, rescue is observed using conditional DopR expression in adults. Taken together, these data argue that rescue of the ReSH phenotype by the Gal4 lines tested reflects their common expression in the EB and that this is a normal site of DopR action in adult flies (Lebestky, 2009).
A requirement for DopR in the EB in regulating ReSH behavior is consistent with the fact that the CC is involved in the control of walking activity. However, the mushroom body has also been implicated in the control of locomotor behavior, and DopR is strongly expressed in this structure as well. Rescue data argue against the MB and in favor of the CC as a neural substrate for the ReSH phenotype of DopR mutants. Unexpectedly, the nocturnal hypoactivity phenotype of DopR mutants was not rescued by restoration of DopR expression to the CC. Thus, not all locomotor activity phenotypes of the DopR mutant necessarily reflect a function for the gene in the CC (Lebestky, 2009).
Interestingly, Gal4 line c547 expresses in R2/R4m neurons of the EB, while lines 189y and c761 express in R3 neurons, yet both rescued the ReSH phenotype of DopR mutants. Similar results have been obtained in experiments to rescue the deficit in ethanol-induced behavior exhibited by the DopR mutant. Double-labeling experiments suggest that endogenous DopR is expressed in all of these EB neuronal subpopulations. Perhaps the receptor functions in parallel or in series in R4m and R3 neurons, so that restoration of DopR expression in either population can rescue the ReSH phenotype. Whether these DopR-expressing EB subpopulations are synaptically interconnected is an interesting question for future investigation (Lebestky, 2009).
Despite its power as a system for studying neural development, function, and behavior, Drosophila has not been extensively used in affective neuroscience, in part due to uncertainty about whether this insect exhibits emotion-like states or behaviors. Increased arousal is a key component of many emotional or affective behaviors. The data presented in this study indicate that Drosophila can express a persistent arousal state in response to repetitive stress. ReSH behavior exhibits several features that distinguish it from simple, reflexive stimulus-response behaviors: scalability, persistence following stimulus termination, and sensitization. In addition, the observation that mechanical trauma promotes release from Drosophila of an odorant that repels other flies suggests that the arousal state underlying ReSH behavior may have a negative 'affective valence' as well. These considerations, taken together with the fact that ReSH is influenced by genetic and pharmacologic manipulations of DA, a biogenic amine implicated in emotional behavior in humans, support the idea that the ReSH response may represent a primitive 'emotion-like' behavior in Drosophila (Lebestky, 2009).
The phenotype of DopR flies is reminiscent of attention-deficit hyperactivity disorder (ADHD), an affective disorder linked to dopamine, whose symptoms include hyper-reactivity to environmental stimuli. If humans, like flies, have distinct circuits for different forms of arousal, then the current data suggest that ADHD may specifically involve dopaminergic dysfunction in those circuits mediating environmentally stimulated, rather than endogenous (sleep-wake), arousal. Given that DA negatively regulates environmentally stimulated arousal circuits in Drosophila, such a view would be consistent with the fact that treatment with drugs that increase synaptic levels of DA, such as methylphenidate (ritalin), can ameliorate symptoms of ADHD (Lebestky, 2009).
In further support of this suggestion, in mammals, dopamine D1 receptors in the prefrontal cortex (PFC) have been proposed to negatively regulate activity, while D1 receptors in the nucleus accumbens are thought to promote sleep-wake transitions. Numerous studies have linked dopaminergic dysfunction in the PFC to ADHD. While most research has focused on the role of the PFC in attention and cognition, rather than in environmentally stimulated arousal per se, dysfunction of PFC circuits mediating phasic DA release has been invoked to explain behavioral hypersensitivity to environmental stimuli in ADHD (Sikstrom, 2007). This view of ADHD as a disorder of circuits mediating environmentally stimulated arousal suggests that further study of such circuits in humans and in vertebrate animal models, as well as in Drosophila, may improve understanding of this disorder and ultimately lead to improved therapeutics (Lebestky, 2009).
An understanding of sleep requires the identification of distinct cellular circuits that mediate the action of specific sleep:wake-regulating molecules, but such analysis has been very limited. This study identifies a circuit that underlies the wake-promoting effects of octopamine in Drosophila. Using MARCM, the ASM cells in the medial protocerebrum were identified as the wake-promoting octopaminergic cells. Octopamine signaling was then blocked in random areas of the fly brain, and the postsynaptic effect was mapped to insulin-secreting neurons of the pars intercerebralis (PI). These PI neurons show altered potassium channel function as well as an increase in cAMP in response to octopamine, and genetic manipulation of their electrical excitability alters sleep:wake behavior. Effects of octopamine on sleep:wake are mediated by the cAMP-dependent isoform of the OAMB receptor. These studies define the cellular and molecular basis of octopamine action and suggest that the PI is a sleep:wake-regulating neuroendocrine structure like the mammalian hypothalamus (Crocker, 2010).
Intercellular signaling is important for accurate circadian rhythms. In Drosophila, the small ventral lateral neurons (s-LNvs) are the dominant pacemaker neurons and set the pace of most other clock neurons in constant darkness. This study shows that two distinct G protein signaling pathways are required in LNvs for 24 hr rhythms. Reducing signaling in LNvs via the G alpha subunit Gs, which signals via cAMP, or via the G alpha subunit Go, which signals via Phospholipase 21c, lengthens the period of behavioral rhythms. In contrast, constitutive Gs or Go signaling makes most flies arrhythmic. Using dissociated LNvs in culture, it was found that Go and the metabotropic GABA(B)-R3 receptor are required for the inhibitory effects of GABA on LNvs and that reduced GABA(B)-R3 expression in vivo lengthens period. Although no clock neurons produce GABA, hyperexciting GABAergic neurons disrupts behavioral rhythms and s-LNv molecular clocks. Therefore, s-LNvs require GABAergic inputs for 24 hr rhythms (Dahdal, 2010).
The long-periods observed with reduced Gs signaling are consistent with four other manipulations of cAMP levels or PKA activity that alter fly circadian behavior. First, long-period rhythms with dnc over-expression complement the short periods of dnc hypomorphs and suggest that the latter are due to loss of dnc from LNvs. dnc mutants also increase phase shifts to light in the early evening. However, this study found no difference in phase delays or advances between Pdf > dnc and control flies, suggesting that altered light-responses of dnc hypomorphs are due to dnc acting in other clock neurons. The period-altering effects seen when manipulating cAMP levels are also consistent with finding stat expressing the cAMP-binding domain of mammalian Epac1 in LNvs lengthens period. This Epac1 domain likely reduces free cAMP levels in LNvs, although presumably not as potently as UAS-dnc. Third, mutations in PKA catalytic or regulatory subunits that affect the whole fly disrupt circadian behavior. Fourth, over-expressing a PKA catalytic subunit in LNvs rescues the period-altering effect of a UAS-shibire transgene that alters vesicle recycling, although the PKA catalytic subunit had no effect by itself. The long periods observed with reduced Gs signaling in LNvs also parallel mammalian studies in which pharmacologically reducing Adenylate cyclase activity lengthened period in SCN explants and mice (Dahdal, 2010).
G-proteins typically transduce extracellular signals. What signals could activate Gs in s-LNvs? PDF is one possibility since PDFR induces cAMP signaling in response to PDF in vitro, indicating that it likely couples to Gs. PDF could signal in an autocrine manner since PDFR is present in LNvs. However, the long-periods observed with reduced Gs signaling differ from the short-period and arrhythmic phenotypes of Pdf and pdfr mutants. The likeliest explanation for these differences is that the altered behavior of Pdf and pdfr mutants results from effects of PDF signaling over the entire circadian circuit, whereas the current manipulations specifically targeted LNvs. Indeed, LNvs are not responsible for the short-period rhythms in Pdf01 null mutant flies. Other possible explanations for the differences between the long-period rhythms with decreased Gs signaling in LNvs and the short-period rhythms of Pdf and pdfr mutants are that additional GPCRs couple to Gs in s-LNvs and influence molecular clock speed and that the current manipulations decrease rather than abolish reception of PDF. In summary, the data shows that Gs signaling via cAMP in s-LNvs modulates period length (Dahdal, 2010).
Go signaling via PLC21C constitutes a novel pathway that regulates the s-LNv molecular clock. This study found that Go and the metabotropic GABAB-R3 receptor are required for the inhibitory effects of GABA on larval LNvs, which develop into adult s-LNvs. The same genetic manipulations that block GABA inhibition of LNvs in culture (expression of Ptx or GABAB-R3-RNAi) lengthened the period of adult locomotor rhythms. Furthermore, the molecular clock in s-LNvs is disrupted when a subset of GABAergic neurons are hyper-excited. Since the LNvs do not produce GABA themselves, s-LNvs require GABAergic inputs to generate 24hr rhythms. Thus s-LNvs are less autonomous for determining period length in DD than previously anticipated (Dahdal, 2010).
Activation of G-proteins can have both short- and long-term effects on a cell. With Go signaling blocked by Ptx, short-term effects on LNv responses were detected in response to excitatory ACh and longer-term effects on the molecular clock. The latter are presumably explained by PLC activation since the behavioral phenotypes of Pdf > GoGTP flies were rescued by reducing Plc21C expression (Dahdal, 2010).
Since s-LNv clocks were unchanged even when the speed of all non-LNv clock neurons were genetically manipulated, it is surprising to find s-LNv clocks altered by signaling from GABAergic non-clock neurons. Why would LNvs need inputs from non-clock neurons to generate 24hr rhythms? One possibility is that LNvs receive multiple inputs which either accelerate or slow down the pace of their molecular clock but overall balance each other to achieve 24hr rhythms in DD. Since reducing signaling by Gs and Go lengthens period, these pathways normally accelerate the molecular clock. According to this model, there are unidentified inputs to LNvs which delay the clock. Identifying additional receptors in LNvs would allow this idea to be tested (Dahdal, 2010).
Previous work showed that GABAergic neurons project to LNvs and that GABAA receptors in l-LNvs regulate sleep. The current data show that constitutive activation of Go signaling dramatically alters behavioral rhythms, suggesting that LNvs normally receive rhythmic GABAergic inputs. But how can s-LNvs integrate temporal information from non clock-containing GABAergic neurons? s-LNvs could respond rhythmically to a constant GABAergic tone by controlling GABAB-R3 activity. Indeed, a recent study found that GABAB-R3 RNA levels in s-LNvs are much higher at ZT12 than at ZT0 (Kula-Eversole, 2010). Strikingly, this rhythm in GABAB-R3 expression is in antiphase to LNv neuronal activity. Thus regulated perception of inhibitory GABAergic inputs could at least partly underlie rhythmic LNv excitability. GABAergic inputs could also help synchronize LNvs as in the cockroach circadian system. Thus GABA's short-term effects on LNv excitability, likely mediated by Gβ/γ, and GABA's longer-term effects on the molecular clock via Go may both contribute to robust rhythms (Dahdal, 2010).
GABAergic signalling is important for normal sleep in humans and flies. This study has advance the current understanding of GABAergic modulation of daily sleep patterns by focusing on the role of slow metabotropic GABAB receptors in Drosophila. It was asked whether GABAB-R2 receptors are regulatory elements in sleep regulation in addition to the already identified fast ionotropic Rdl GABAA receptors. By immunocytochemical and reporter-based techniques it was shown that the pigment dispersing factor (PDF)-positive ventrolateral clock neurons (LNv) express GABAB-R2 receptors. Downregulation of GABAB-R2 receptors in the large PDF neurons (l-LNv) by RNAi reduced sleep maintenance in the second half of the night, whereas sleep latency at the beginning of the night that was previously shown to depend on ionotropic Rdl GABAA receptors remained unaltered. The results confirm the role of the l-LNv neurons as an important part of the sleep circuit in D. melanogaster and also identify the GABAB-R2 receptors as the thus far missing component in GABA-signalling that is essential for sleep maintenance. Despite the significant effects on sleep, no changes were observed changes in circadian behaviour in flies with downregulated GABAB-R2 receptors, indicating that the regulation of sleep maintenance via l-LNv neurons is independent of their function in the circadian clock circuit (Gmeiner, 2013).
The fruit fly has become a well-accepted model for sleep research. As in mammals, it has been shown that the sleep-like state of Drosophila is associated with reduced sensory responsiveness and reduced brain activity, and is subject to both circadian and homeostatic regulation. Similarly to in humans, monaminergic neurons (specifically dopaminergic and octopaminergic neurons) enhance arousal in fruit flies, whereas GABAergic neurons promote sleep (Agosto, 2008). As in humans, GABA advances sleep onset (reduces sleep latency) and prolongs total sleep (increases sleep maintenance). Brain regions possibly implicated in the regulation of sleep in D. melanogaster are the pars intercerebralis, the mushroom bodies and a subgroup of the pigment dispersing factor (PDF)-positive neurons called the l-LNv neurons. The l-LNv belong to the circadian clock neurons, indicating that in flies, as in mammals, the sleep circuit is intimately linked to the circadian clock and that the mechanisms employed to govern sleep in the brain are evolutionarily ancient (Gmeiner, 2013).
The l-LNv are conspicuous clock neurons with wide arborisations in the optic lobe, fibres in the accessory medulla -- the insect clock centre -- and connections between the brain hemispheres. Thus, the l-LNv neurons are anatomically well suited to modulate the activity of many neurons. In addition, their arborisations overlap with those of monaminergic neurons. Several studies show that they indeed receive dopaminergic, octopaminergic and GABAergic input and that they control the flies' arousal and sleep. Furthermore, the l-LNv are directly light sensitive and promote arousal and activity in response to light, especially in the morning (Gmeiner, 2013).
A part of the sleep-promoting effect of GABA on the l-LNv has been shown to be mediated via the fast ionotropic GABAA receptor Rdl (Resistance to dieldrin) (Agosto, 2008). Rdl Cl- channels are expressed in the l-LNv (Agosto, 2008) and, similar to mammalian GABAA receptors, they mediate fast inhibitory neurotransmission. As expected, GABA application reduced the action potential firing rate in the l-LNv, whereas application of picrotoxin, a GABAA receptor antagonist, increased it (McCarthy, 2011). Furthermore, an Rdl receptor mutant with prolonged channel opening and consequently increased channel current significantly decreased sleep latency of the flies after lights-off, whereas the downregulation of the Rdl receptor via RNAi increased it (Agosto, 2008; Gmeiner, 2013 and references therein).
Nevertheless, the manipulation of the Rdl receptor had no effect on sleep maintenance. Because the latter is significantly reduced after silencing the GABAergic neurons (Parisky, 2008), other GABA receptors must be responsible for maintaining sleep. Suitable candidates are slow metabotropic GABAB receptors that are often co-localised with ionotropic GABAA receptors (Enell, 2007). In Drosophila, like in mammals, the metabotropic GABAB receptors are G-protein-coupled seven-transmembrane proteins composed of two subunits, GABAB-R1 and GABAB-R2 (Kaupmann, 1998; Mezler, 2001). The GABAB-R1 is the ligand binding unit and GABAB-R2 is required for translocation to the cell membrane and for stronger coupling to the G-protein (Kaupmann, 1998; Galvez, 2001). This study shows that the l-LNv do express metabotropic GABAB-R2 receptors and that these receptors are relevant for sleep maintenance but not for sleep latency. Thus, metabotropic and ionotropic GABA receptors are cooperating in sleep regulation (Gmeiner, 2013).
This study shows that metabotropic GABAB-R2 receptors are expressed on the PDF-positive clock neurons (LNv neurons), and that their downregulation in the l-LNv by RNAi results in: (1) a higher activity level throughout the day and night and (2) reduced sleep maintenance in the second half of the night. Neither sleep onset nor circadian rhythm parameters were affected by the downregulation. It is concluded that GABA signalling via metabotropic receptors on the l-LNv is essential for sustaining sleep throughout the night and for keeping activity at moderate levels throughout the 24-h day (preventing flies from hyperactivity). A major caveat of RNAi is off-target effects, particularly when Gal4 drivers are expressed in large numbers of non-target neurons. Though GABAB-R2 was downregulated in only eight neurons per brain hemisphere and the behavioural effects of the knockdown experiments were carefully correlated with observation and measures of GABAB-R2 immunostaining in the s-LNv and l-LNv, it is still possible that some effects were due to off-target knockdown of other membrane proteins. Nevertheless, given the fact that no such effects have been reported in the previous paper that used the same GABAB-R2 RNAi line (Root, 2008), it is thought unlikely that the behavioural effects described in this study were due to off-target knockdown of other genes (Gmeiner, 2013).
The results are in line with a former study describing the location of GABAB receptors in D. melanogaster (Hamasaka, 2005). The ionotropic GABAA receptor Rdl has also been identified on the l-LNv neurons and has been shown to regulate sleep, but its downregulation delayed only sleep onset and did not perturb sleep maintenance (Parisky, 2008). In contrast, silencing GABAergic signalling influenced sleep onset and sleep maintenance, indicating that GABA works through the fast Rdl receptor, and also implying a longer-lasting signalling pathway. GABAB receptors are perfect candidates in mediating slow but longer-lasting effects of GABA. Often, GABAA and GABAB receptors cooperate in mediating such fast and slow effects. For example, in the olfactory system, GABAA receptors mediate the primary modulatory responses to odours whereas GABAB receptors are responsible for long-lasting effects (Wilson, 2005; Gmeiner, 2013 and references therein).
In D. melanogaster, GABAB receptors consist of the two subunits GABAB-R1 and GABAB-R2, and only the two units together can efficiently activate the metabotropic GABA signalling cascade (Galvez, 2001; Mezler, 2001). In the current experiments, only GABAB-R2 was downregulated, but this manipulation should also have decreased the amount of functional GABAB-R1/GABAB-R2 heterodimers and, therefore, reduced GABAB signalling in general. Taking into account that sleep maintenance in the second half of the night was already significantly impaired by an ~46% reduction in detectable GABAB-R2 immunostaining intensity in the l-LNv clock neurons, it can be assumed that GABAB receptors account for an even larger portion of the sleep maintenance than detected in these experiments. Thus GABAB receptors play a crucial role in mediating GABAergic signals to the l-LNv neurons, which are needed to sustain sleep throughout the night. This is mainly due to the maintenance of extended sleep bout durations in the second half of the night. When signalling by the GABAB receptor is reduced, sleep bouts during this interval are significantly shortened, leading to less total sleep (Gmeiner, 2013).
Most importantly, this study confirmed the l-LNv as important components in regulation of sleep and arousal (Agosto, 2008; Parisky, 2008; Kula-Eversole, 2010; Shang, 2011). In contrast, the s-LNv seem to be not involved in sleep-arousal regulation but are rather important for maintaining circadian rhythmicity under DD (reviewed by Helfrich-Förster, 2007). One caveat in clearly distinguishing the function of s-LNv and l-LNv is the fact that both cell clusters express the neuropeptide PDF and, as a consequence, Pdf-GAL4 drives expression in both subsets of clock neurons. Though no significant GABAB-R2 knock-down in the s-LNv is seen, it cannot be completely excluded that GABAB-R2 was slightly downregulated in these clock neurons and that this knock-down contributes to the observed alterations in sleep. To restrict the knock-down to the s-LNv the R6-GAL4 line was used that is expressed in the s-LNv but not in the l-LNv. Neither a reduction in GABAB-R2 staining intensity in the s-LNv nor any effects on sleep in the second half of the night was seen. The lack of any visible GABAB-R2 downregulation in the s-LNv with R6-GAL4 is in agreement with observations of Shafer and Taghert (Shafer, 2009), who could completely downregulate PDF in the s-LNv using Pdf-GAL4 but not using R6-GAL4. Thus, R6-GAL4 is a weaker driver than Pdf-GAL4 and is obviously not able to influence GABAB-R2 in the s-LNv. Nevertheless, in the current experiments the R6-Gal4-driven GABAB-R2 RNAi led to flies that had slightly higher diurnal activity levels and less diurnal rest than the control flies. This suggests that GABAB-R2 was downregulated somewhere else. When checking the R6-GAL4 expression more carefully it was found that R6-GAL4 was not restricted to the brain, but was also present in many cells of the thoracic and especially the abdominal ganglia. Given the broad expression of GABAB-R2, a putative knock-down in the ventral nervous system is likely to affect locomotor activity (Gmeiner, 2013).
The results on the l-LNv certainly do not exclude a role of GABA in the circadian clock controlling activity rhythms under DD conditions (here represented by the s-LNv). In mammals, GABA is the most abundant neurotransmitter in the circadian clock centre in the brain -- the suprachiasmatic nucleus. GABA interacts with GABAA and GABAB receptors, producing primarily but not exclusively inhibitory responses through membrane hyperpolarisation. GABA signalling is important for maintaining behavioural circadian rhythmicity, it affects the amplitude of molecular oscillations and might contribute to synchronisation of clock cells within the suprachiasmatic nucleus. The same seems to be true for fruit flies. The s-LNv neurons of adults alter cAMP levels upon GABA application on isolated brains in vitro (Lelito, 2012). Hyperexcitation of GABAergic neurons disrupts the molecular rhythms in the s-LNv and renders the flies arrhythmic (Dahdal, 2010). Thus, GABA signalling affects the circadian clock in the s-LNv. Flies with downregulated GABAB-R2 receptors were found to have slightly longer free-running periods than the control flies, but this turned out to be only significant in comparison with Control 2 and not to Control 1. Dahdal (2010) found similar small effects on period after downregulating GABAB-R2 receptors, but a significant period lengthening after downregulating GABAB-R3 receptors. This indicates that GABA signals via GABAB-R3 receptors to the s-LNv and was confirmed in vitro in the larval Drosophila brain by Ca2+ imaging (Dahdal, 2010). Nevertheless, the study of Dahdal does not rule out that GABA signals via GABAB-R3 plus GABAB-R2 receptors on the adult s-LNv. This study found a rather strong expression of GABAB-R2 receptors in these clock neurons, and were not able to downregulate it significantly by RNAi, although dicer2 was used as amplification. Dahdal did not use dicer2, and they also did not measure the effectiveness of the downregulation of GABAB-R2 by RNAi immunocytochemically directly in the s-LNv. Thus, the exact GABAB receptors that mediate GABA responses in the adult s-LNv need still to be determined (Gmeiner, 2013).
In summary, it is concluded that the l-LNv subgroup of
the PDF-positive clock neurons is a principal target of
sleep-promoting and activity-repressing GABAergic neurons and sits at
the heart of the sleep circuit in D. melanogaster. Thus, the sleep
circuitry of flies is clearly more circumscribed and simpler than that
of mammals. Mammals have many targets of sleep-promoting GABAergic
neurons, and the circadian clock seems to have a mainly modulatory and
less direct influence on sleep (Mistlberger, 2005). The fly sleep
circuitry may therefore have condensed the mammalian arousal and sleep
stimulating systems (e.g., monaminergic, cholinergic, peptidergic and
GABAergic systems) into a simpler and more compact region, which seems
to largely coincide with the eight PDF-positive l-LNv cells of the
circadian circuit (Gmeiner, 2013).
Emerging evidence indicates the role of amino acid metabolism in sleep regulation. This study demonstrates sleep-promoting effects of dietary threonine (SPET) in Drosophila. Dietary threonine markedly increased daily sleep amount and decreased the latency to sleep onset in a dose-dependent manner. High levels of synaptic GABA or pharmacological activation of metabotropic GABA receptors (GABAB-R) suppressed SPET. By contrast, synaptic blockade of GABAergic neurons or transgenic depletion of GABAB-R in the ellipsoid body R2 neurons enhanced sleep drive non-additively with SPET. Dietary threonine reduced GABA levels, weakened metabotropic GABA responses in R2 neurons, and ameliorated memory deficits in plasticity mutants. Moreover, genetic elevation of neuronal threonine levels was sufficient for facilitating sleep onset. Taken together, these data define threonine as a physiologically relevant, sleep-promoting molecule that may intimately link neuronal metabolism of amino acids to GABAergic control of sleep drive via the neuronal substrate of sleep homeostasis (Ki, 2019).
The circadian clock and sleep homeostasis are two key regulators that shape daily sleep behaviors in animals. In stark contrast to the homeostatic nature of sleep, the internal machinery of sleep is vulnerable to external (e.g., environmental change) or internal conditions (e.g., genetic mutation) that lead to adaptive changes in sleep behaviors. Sleep behavior is conserved among mammals, insects, and even lower eukaryotes. Since the identification of the voltage-gated potassium channel Shaker as a sleep-regulatory gene in Drosophila, fruit flies have been one of the most advantageous genetic models to dissect molecular and neural components that are important for sleep homeostasis and plasticity (Ki, 2019).
To date, a number of sleep-regulatory genes and neurotransmitters have been identified in animal models as well as in humans. For instance, the inhibitory neurotransmitter gamma-aminobutyric acid (GABA) is known to have a sleep-promoting role that is conserved in invertebrates and vertebrates. Hypomorphic mutations in mitochondrial GABA-transaminase (GABA-T) elevate GABA levels and lengthen baseline sleep in flies (Chen, 2015). The long sleep phenotype in GABA-T mutants accompanies higher sleep consolidation and shorter latency to sleep onset, consistent with the observations that pharmacological enhancement of GABAergic transmission facilitates sleep in flies and mammals, including humans. In addition, resistance to dieldrin (Rdl), a Drosophila homolog of the ionotropic GABA receptor, suppresses wake-promoting circadian pacemaker neurons in adult flies to exert sleep-promoting effects. Similarly, 4,5,6,7-tetrahydroisoxazolo[5,4 c]pyridin-3-ol (THIP), an agonist of the ionotropic GABA receptor, promotes sleep in insects and mammals (Ki, 2019).
Many sleep medications modulate GABAergic transmission. A prominent side effect of anti-epileptic drugs relevant to GABA is causing drowsiness. Conversely, glycine supplements improve sleep quality in a way distinct from traditional hypnotic drugs, minimizing deleterious cognitive problems or addiction. In fact, glycine or D-serine acts as a co-agonist of N-methyl-D-aspartate receptors (NMDARs) and promotes sleep through the sub-type of ionotropic glutamate receptors. Emerging evidence further supports the roles of amino acid transporters and metabolic enzymes in sleep regulation. In particular, it has been demonstrated that starvation induces the expression of metabolic enzymes for serine biosynthesis in Drosophila brains, and elevates free serine levels to suppress sleep via cholinergic signaling (Sonn, 2018). These observations prompted a hypothesis that other amino acids may also display neuro-modulatory effects on sleep behaviors (Ki, 2019).
The molecular and neural machinery of sleep regulation intimately interacts with external (e.g., light, temperature) and internal sleep cues (e.g., sleep pressure, metabolic state) to adjust the sleep architecture in animals. Using a Drosophila genetic model, this study has investigated whether dietary amino acids could affect sleep behaviors, through this investigation SPET was discovered. Previous studies have demonstrated that the wake-promoting circadian pacemaker neurons are crucial for timing sleep onset after lights-off in LD cycles. In addition, WAKE-dependent silencing of clock neurons and its collaborative function with RDL have been suggested as a key mechanism in the circadian control of sleep onset. However, the current evidence indicates that SPET facilitates sleep onset in a manner independent of circadian clocks. It was further elucidated that SPET operates likely via the down-regulation of metabotropic GABA transmission in R2 EB neurons, a neural locus for generating homeostatic sleep drive (Ki, 2019).
Both food availability and nutritional quality substantially affect sleep behaviors in Drosophila. Sucrose contents in food and their gustatory perception dominate over dietary protein to affect daily sleep. Starvation promotes arousal in a manner dependent on the circadian clock genes Clock and cycle as well as neuropeptide F (NPF), which is a fly ortholog of mammalian neuropeptide Y. On the other hand, protein is one of the nutrients that contribute to the postprandial sleep drive in Drosophila and this observation is possibly relevant to SPET. While Leucokinin (Lk) and Lk receptor (Lkr) play important roles in dietary protein-induced postprandial sleepand in starvation-induced arousal, comparable SPET was observed between hypomorphic mutants of Lk or Lkr and their heterozygous controls. Therefore, SPET and its neural basis reveal a sleep-regulatory mechanism distinct from those involved in sleep plasticity relevant to food intake (Ki, 2019).
What will be the molecular basis of SPET? Given the general implication of GABA in sleep promotion, a simple model will be that a molecular sensor expressed in a subset of GABAergic neurons (i.e., LN) directly responds to an increase in threonine levels, activates GABA transmission, and thereby induces sleep. Several lines of evidence, however, favored the other model that dietary threonine actually down-regulates metabotropic GABA transmission in R2 EB neurons, de-represses the neural locus for generating homeostatic sleep drive, and thereby enhances sleep drive. The latter model does not necessarily conflict with sleep-promoting effects of genetic or pharmacological conditions that generally elevate GABA levels or enhance GABAergic transmission since those effects will be the net outcome of activated GABA transmission via various sub-types of GABA receptors expressed in either wake- or sleep-promoting neurons and their (Ki, 2019).
The structural homology among threonine, GABA, and their metabolic derivatives (e.g., alpha-ketobutyrate and gamma-hydroxybutyrate) led to the hypothesis that these relevant chemicals may act as competitive substrates in enzymatic reactions for their overlapping metabolism. Consequently, dietary threonine may limit the total flux of GABA-glutamate-glutamine cycle possibly through substrate competition, decreases the size of available GABA pool, and thereby down-scales GABA transmission for SPET. This accounts for why genetic or pharmacological elevation of GABA levels rather suppresses SPET. Threonine, GABA, and their derivatives may also act as competitive ligands for metabotropic GABA receptors, explaining weak GABA responses in R2 EB neurons of threonine-fed flies. Biochemical and neural evidence supportive of this hypothesis is quite abundant. It has been previously shown that alpha-ketobutyrate, GABA, and the ketone body beta-hydroxybutyrate act as competitive substrates in common enzymatic reactions. Moreover, functional interactions of beta-hydroxybutyrate or gamma-hydroxybutyrate with GABAergic signaling have been well documented. Finally, threonine and GABA derivatives have anti-convulsive effects, which further support their common structural and functional relevance to GABAergic signaling (Ki, 2019).
The removal of the amino group is the initial step for amino acid metabolism, and various transaminases mediate its transfer between amino acids and alpha-keto acids. On the other hand, a group of amino acids (i.e., glutamate, glycine, serine, and threonine) has their own deaminases that can selectively remove the amino group. The presence of these specific deaminases is indicative of active mechanisms that individually fine-tune the baseline levels of these amino acids in metabolism, and possibly in the context of other physiological processes as well. This idea is further supported by the conserved roles of glutamate, glycine, and serine as neurotransmitters or neuromodulators important for brain function, including sleep regulation. In fact, serine, glycine, and threonine constitute a common metabolic pathway, and threonine may contribute indirectly to glycine- or serine-dependent activation of sleep-promoting NMDAR. Nonetheless, this study found that sleep-modulatory effects of dietary glycine were distinct from SPET and thus, it is speculated that threonine may act as an independent neuromodulator, similar to other amino acids with their dedicated deaminases (Ki, 2019).
While several lines of the data support that threonine is likely to be an endogenous sleep driver in fed conditions, it wa recently demonstrated that starvation induces serine biosynthesis in the brain and neuronal serine subsequently suppresses sleep via cholinergic signaling (Sonn, 2018). These two pieces of relevant works establish a compelling model that the metabolic pathway of serine-glycine-threonine functions as a key sleep-regulatory module in response to metabolic sleep cues (e.g., food ingredients and dietary stress). It is further hypothesized that the adaptive control of sleep behaviors by select amino acids and their conserved metabolic pathway suggests an ancestral nature of their sleep regulation. Future studies should address if the serine-glycine-threonine metabolic pathway constitutes the sleep homeostat that can sense and respond to different types of sleep needs. In addition, it will be interesting to determine if this metabolic regulation of sleep is conserved among other animals, including humans (Ki, 2019).
Feeding and sleep are highly conserved, interconnected behaviors essential for survival. Starvation has been shown to potently suppress sleep across species; however, whether satiety promotes sleep is still unclear. This study used the fruit fly, Drosophila melanogaster, as a model organism to address the interaction between feeding and sleep. The sleep of flies that had been starved for 24 h was monitored, and sleep amount was found to increase in the first 4 h after flies were given food. Increased sleep after starvation was due to an increase in sleep bout number and average sleep bout length. Mutants of translin or adipokinetic hormone, which fail to suppress sleep during starvation, still exhibited a sleep increase after starvation, suggesting that sleep increase after starvation is not a consequence of sleep loss during starvation. It was also found that feeding activity and food consumption were higher in the first 10-30 min after starvation. Restricting food consumption in starved flies to 30 min was sufficient to increase sleep for 1 h. Although flies ingested a comparable amount of food at differing sucrose concentrations, sleep increase after starvation on a lower sucrose concentration was undetectable. Taken together, these results suggest that increased food intake after starvation enhances sleep and reveals a novel relationship between feeding and sleep (Regalado, 2017).
Most animals sleep or exhibit a sleep-like state, yet the adaptive significance of this phenomenon remains unclear. Although reproductive deficits are associated with lifestyle induced sleep deficiencies, how sleep loss affects reproductive physiology is poorly understood, even in model organisms. This study aimed to bridge this mechanistic gap by impairing sleep in female fruit flies and testing its effect on egg output. Sleep deprivation by feeding caffeine or by mechanical perturbation was shown to result in decreased egg output. Transient activation of wake-promoting dopaminergic neurons decreases egg output in addition to sleep levels, thus demonstrating a direct negative impact of sleep deficit on reproductive output. Similarly, loss-of-function mutation in dopamine transporter fumin (fmn) leads to both significant sleep loss and lowered fecundity. This demonstration of a direct relationship between sleep and reproductive fitness indicates a strong driving force for the evolution of sleep (Potdar, 2018a).
Neurotransmitters play an important role in regulating the physiological activity of the animal, especially in emotion and sleep. While nucleotides are involved in almost all cellular processes. However, the characteristics of sleep-related neurochemicals under different life cycles and environment remain poorly understood. A rapid and sensitive analytical method was established with LC-MS/MS to determine eight endogenous neurochemicals in Drosophila melanogaster and the levels of neurochemicals in the different developmental stages of Drosophila melanogaster were evaluated. The results indicated that there were significant discrepancies among different stages, especially from pupal stage to adult. The levels of these compounds in caffeine-induced insomnia model of Drosophila melanogaster were investigated. Compared with normal group the eight endogenous metabolites did not fluctuate significantly in insomnia Drosophila melanogaster, which may be due to the mechanism of caffeine-induced insomnia through other pathways, such as adenosine. The results provide a reference for decoding of neurochemicals involved in the development of the full cycle of mammalian life and exploration of insomnia even other mental diseases induced by exogenous substances in the future (Sun, 2022).
Circadian clocks modulate
timing of sleep/wake cycles in animals; however, the underlying
mechanisms remain poorly understood. In Drosophila melanogaster, large
ventral lateral neurons (l-LNv) are known to promote wakefulness through
the action of the neuropeptide pigment
dispersing factor (PDF), but the downstream targets of PDF
signalling remain elusive. In a screen using downregulation or
overexpression (OEX) of the gene encoding PDF receptor (pdfr),
this study found that a subset of dopaminergic neurons responds
to PDF to promote wakefulness during the day. Moreover, this study found
that small LNv (sLNv) and dopaminergic neurons form synaptic contacts,
and PDFR signalling inhibited dopaminergic neurons specifically during
day time. It is proposed that these dopaminergic neurons that respond to
PDFR signalling are sleep-promoting and that during the day when PDF
levels are high, they are inhibited, thereby promoting wakefulness.
Thus, this study has identified a novel circadian clock pathway that
mediates wake promotion specifically during day time (Potdar, 2018b).
Daily cycles in several environmental factors synchronize endogenous circadian clocks which drive rhythmic sleep/wake patterns in many organisms. Homeostatic mechanisms modulate the amount and depth of sleep, and also allow animals to recover from any sleep deprivation they may have incurred. Together, these processes control the timing and occurrence of sleep and wake states, thereby modulating sleep/wake cycles. Since the discovery that sleep behavior of Drosophila melanogaster is similar to mammalian sleep in several aspects, many pathways and neuronal circuits involving sleep homeostat and circadian clocks have been uncovered. Genes such as minisleep (mns) and hyperkinetic (hk) encoding subunits of Shaker potassium channel function in the sleep homeostat. More recently, central complex structures such as dorsal fan-shaped body (FB) and the ellipsoid body (EB) have been shown to function as effector and modulator of the sleep homeostat, respectively. Meanwhile, mutations in core circadian clock genes such as Clock (clk) and Cycle (cyc) have been shown to cause impaired timing of sleep as they tend to become nocturnal. The circadian neuropeptide pigment dispersing factor (PDF) and its receptor (PDFR) are involved in relaying wake-promoting signals from the circadian pacemaker ventral lateral neurons (LNvs) in response to light input as well as dopamine. While it has been suggested that the EB may be the downstream target of this wake-promoting PDF/PDFR signaling, the evidence in favor of the same is limited (Potdar, 2018b).
In the recent past, in the quest to uncover output pathways of the circadian clocks that help in timing of sleep/wake cycles, a few dedicated circuits have been mapped. Most notably, timing of sleep onset at the beginning of night is a function of increased inhibition of wake-promoting large LNv (l-LNv) by GABA. In contrast, sleep is suppressed at the end of night by the action of PDF on the PDFR+ dorsal neuron 1 (DN1) group of the circadian network that in turn secretes the wake-promoting neuropeptide diuretic hormone 31 (DH31). Furthermore, yet another group showed that DN1s through glutamate modulate day-time siesta and night-time sleep by inhibiting the morning (small LNv; s-LNv) and evening (dorsal lateral neurons; LNds) activity controlling circadian neurons. Yet, none of the studies so far have shed light on how circadian neurons may induce wakefulness during the day (Potdar, 2018b).
This question was addressed by screening for putative downstream targets of PDFR signaling by altering the levels of pdfr expression in several subsets of neurons - namely, circadian neurons that are known to express pdfr, subsets of mushroom body (MB) neurons that are sleep- or wake-promoting, wake-promoting pars intercerebralis (PI), sleep homeostat EB, and sleep-promoting FB neurons as well as aminergic neuronal groups, most of which are reported to be wake-promoting. Strikingly, this study found that a subset of dopaminergic neurons responds to changes in pdfr expression by changing the levels of day-time sleep, increasing pdfr levels decreases day-time sleep and vice versa. Moreover, PDF+ and dopaminergic neurons were found to form synaptic contacts with one another, along with the possibility of the former inhibiting the latter. Thus, these results uncover a dedicated pathway involving signaling from the PDF+ neurons perhaps to the PPM3 dopaminergic neurons in the regulation of wakefulness during the day (Potdar, 2018b).
Dopamine is primarily involved in promoting wakefulness and is known to act on l-LNv as well as inhibit sleep-promoting dFB to carry out its wake-promoting function. This study has revealed that certain dopamine neurons are in fact sleep-promoting and through the inhibitory action of PDFR signaling, wakefulness gets promoted specifically during the day. additional experiments that use optogenetic techniques can shed more light on whether these dopaminergic neurons promote sleep directly, or indirectly by preventing wakefulness either through a gating mechanism or by a permissive role. Interestingly, a previous study has found that dopamine acts on l-LNv to promote wakefulness and this study found that PDFR signaling acts on dopamine neurons, suggesting a feed-forward pathway for wake promotion, where dopamine acting on l-LNv promotes the inhibition of sleep-promoting dopaminergic neurons by PDFR signaling. The identity of dopamine neurons acting on l-LNv and those responding to PDFR signaling may differ which can be uncovered with additional experiments (Potdar, 2018b).
The role of s-LNv in modulating sleep and wake has been explored in some detail in the recent years. s-LNv have also been shown to promote sleep via short NPF (sNPF) as well as myoinhibitory peptide (MiP) by inhibiting the wake-promoting l-LNv. This study shows that PDF+ s-LNv make synaptic contacts with dopaminergic neurons and that PDFR signaling inhibits the downstream dopaminergic neurons to promote wakefulness during the day. Moreover, this study has shown a secondary role for s-LNv in modulating wake-promoting effects of l-LNv. Yet, how this wake-promoting signal which originates in the l-LNv gets relayed to the s-LNv is not understood. Furthermore, from the screen it is clear that this function is not mediated via PDFR signaling among the LNv, as downregulating and overexpressing pdfr in s-LNv (Clk 9M GAL4 and Pdf GAL4) do not result in any sleep defects. Thus, l-LNv to s-LNv wake-promoting signal is independent of PDF while s-LNv to dopamine wake-promoting signal requires PDFR signaling (Potdar, 2018b).
PDFR being a class B1 GPCR utilizes cAMP as its second messenger, although there is evidence for Ca2+ also acting as the second messenger. For most of the functions of PDF including stabilizing core clock proteins such as TIMELESS and PERIOD in different target neurons such as DN1s and s-LNv, cAMP is the major secondary messenger. Moreover, it is thought that different actions of PDF of slowing and speeding up of morning and evening clock neurons is also mediated by different components of cAMP signaling mechanism. However, this study shows that for the function of regulating wake levels during the day time, PDFR signaling changes levels of intracellular Ca2+ in dopamine neurons with negligible role for cAMP signaling, suggesting a mechanism by which a neuropeptide that has diverse effects on its downstream targets can modulate different functions independently. This study therefore identified a unique subset of downstream targets for PDFR signaling among the dopamine neurons that promote wakefulness depending on time of day (Potdar, 2018b).
Interestingly, in this screen it is noted that there are several driver lines which there are significant changes in day-time sleep but with only one type of manipulation of pdfr levels (Clk 4.1M, 30y, 104y, 121y GAL4). This may be due to ineffective downregulation of pdfr achieved through the Pdfr RNAi line with these particular drivers. Given that PDF is a neuropeptide which can have long-range non-synaptic effects, even misexpressing it (104y and 121y GAL4) in different substrates has resulted in altered day-time sleep levels. Because DH31 can also respond to PDFR, it is possible that these effects could be mediated by DH31 binding to misexpressed PDFR. However, this may not be the case as downregulating DH31-receptor in these regions does not cause changes in sleep levels. Thus, it can be concluded that in regions previously not known to express pdfr, misexpression of pdfr can cause sleep level deficits suggesting that PDF can act in regions which are not direct targets yet may lie in the vicinity of LNv projections (Potdar, 2018b).
The role of PDF/PDFR signaling is well-known in synchronizing the free-running molecular rhythms in neurons across the circadian network. PDFR signaling in the 'evening' neurons (LNd and 5th s-LNv) is important for appropriate phasing of the evening bout of activity in light/dark cycles. While the role of PDF as a wake-signal has been known, this study demonstrates that a subset of dopaminergic neurons is downstream of the PDF/PDFR signaling. While the PDFR expression is not conclusive, it is shown that perhaps one PPM3 neuron per hemisphere may express the PDFR. Additional experiments that more directly test the functional connectivity between dopaminergic neurons and PDF+ neurons, as well as responsiveness of dopaminergic neurons to PDF may result in a clearer picture. Downregulating pdfr in these neurons results in increase of day-time sleep, which is a phenocopy of the sleep behavior of loss-of-function pdfr whole-body mutants. On the other hand, overexpressing pdfr in these neurons leads to decrease of day-time sleep specifically. It was further shown that PDF and dopaminergic neurons make synaptic contacts with each other at the site of the axonal projection of s-LNv. Moreover, the effect of PDFR signaling on the PPM3 neurons appears to be inhibitory, suggesting that the PDFR+ PPM3 neurons promote sleep. Taken together, it is concluded that wake-promoting LNv make synaptic connections with sleep-promoting dopaminergic neurons and promote wakefulness specifically during the day time through inhibitory PDFR signaling (Potdar, 2018b).
Each day and in conjunction with ambient daylight conditions, neuropeptide PDF regulates the phase and amplitude of locomotor activity rhythms in Drosophila through its receptor, PDFR, a Family B G protein-coupled receptor (GPCR). The in vivo process was studied by which PDFR signaling turns off, by converting as many as half of the 28 potential sites of phosphorylation in its C terminal tail to a non-phosphorylatable residue (alanine). Many such sites are conserved evolutionarily, and their conversion creates a specific behavioral syndrome opposite to loss-of-function phenotypes previously described for pdfr. That syndrome includes increases in the amplitudes of both Morning and Evening behavioral peaks, as well as multi-hour delays of the Evening phase. The precise behavioral effects were dependent on day-length, and most effects mapped to conversion of only a few, specific serine residues near the very end of the protein and specific to its A isoform. Behavioral phase delays of the Evening activity under entraining conditions predicted the phase of activity cycles under constant darkness. The behavioral phenotypes produced by the most severe PDFR variant were ligand-dependent in vivo, and not a consequence of changes to their pharmacological properties, nor of changes in their surface expression, as measured in vitro. The mechanisms underlying termination of PDFR signaling are complex, subject to regulation that is modified by season, and central to a better understanding of the peptidergic modulation of behavior (Li, 2022).
In Drosophila, neuropeptide PDF signaling helps pattern the output of the fly circadian pacemaker network that controls rhythmic daily locomotor activity. Its functions have been compared to those of neuropeptide VIP in regulating mammalian circadian physiology. Historically PDF was first isolated as an active principle (Pigment Dispersing Hormone) that mediates light-dependent dispersion of pigment granules in diverse chromatophores of crustacea. In the insect circadian system, PDF acts for a specified period within each 24 hr cycle, and works in conjunction with environmental light to set phase and amplitude for locomotor activity rhythms that normally occur around dawn and dusk. Each day, the precise times of dusk and of dawn change, which alters the time interval between them. These facts require that the time of PDF signaling, the point when it starts and the point when it stops, must also be adjusted each day to appropriately follow and reflect these daily variations in the light: dark transitions. PDF signaling starts following its release by specific pacemaker neurons, whose period of activity in vivo tracks the dawn in a variety of photoperiodic conditions. A comparable understanding of how PDF receptor signaling normally stops is lacking: this work addresses that mechanism (Li, 2022).
The PDF receptor (PDFR) is a member of the Family B (secretin receptor-like) GPCR group: it is Gs-coupled and its activation elevates cAMP levels in vivo. It regulates different adenylate cyclases (AC) in diverse target pacemakers (Duval, 2012; Duval, 2013), which, through PKA activation, ultimately regulate the pace of the molecular clock through regulation of Timeless (Seluzicki, 2014). PDFR autoreceptor signaling promotes dramatic, daily morphological changes in the axonal terminals of sLNv pacemakers. In addition to its effects on the pace of the molecular pacemaker, PDFR activation also regulates calcium dynamics in subsets of pacemaker neuron groups to help dictate their group-specific, daily phases of activation (in the sLNv, in the 5th sLNv and in subsets of LNd, and DN3 groups). Such target cell-specific delays of PER-dependent neuronal activity illustrate the basis by which the circadian network produces a daily series of staggered phasic, neuronal outputs. Finally, PDF/PDFR signaling is long-lasting: its depression of basal calcium levels in target neurons persists without abatement over many hours. These observations raise fundamental questions regarding the mechanism and the time course by which PDFR signaling diminishes in anticipation of the next day's cycle of signaling (Li, 2022).
The canonical model of GPCR phosphorylation and homologous desensitization features G protein-coupled receptor kinases (GRKs) which associate with activated GPCRs and phosphorylate cytosolic segments, thereby recruiting βarrestin via its β-arrestins uncouple the receptors from G proteins. or enhance receptor endocytosis; they can also serve as signal transducers by recruiting distinct signaling molecules. A second major regulatory mechanism to reduce GPCR signaling is heterologous desensitization, whereby second-messenger-dependent kinases (PKA or PKC) phosphorylate GPCRs. Thus, experiments were to modify evolutionarily-conserved residues in the C terminal tail of PDFR that could conceivably serve as substrates for phosphorylation and subsequent signal termination. The working hypothesis was that, by their actions in vivo, such modified PDF receptors would reveal extended lifetimes of activation (Li, 2022).
Little is known about the mechanisms that underlie normal termination of PDFR signaling. In the small LNv, pdfr levels are higher late in the day than in early morning. Sensitivity to PDF in vivo peaks in the early day and is regulated by the PER-dependent clock, via post-transcriptional mechanisms: the EC50 for PDF responses in identified neurons varies systematically 5-10 fold: as a consequence of RalA action, as a function of time of day, and as a function of seasonality. PDFR signaling is long-lasting: it persists for many hours in vivo, for as long as free peptide ligand is available in the bath. In addition, β-arrestin2-GFP is not efficiently recruited to activated PDFR when the receptor is functionally-expressed in hEK-293T cells. In contrast, each of 13 other Drosophila neuropeptide GPCRs (including two Family B GPCRs, CG8422 and CG17415) efficiently recruit β-arrestin2-GFP, when they are activated by their cognate ligands in that cellular environment (Li, 2022).
This study reports (1) that PDFR is normally phosphorylated in vivo at conserved C terminal residues; (2) that loss of conserved PDFR phosphorylatable sites leads to a behavioral syndrome opposite to loss-of-function pdf and pdfr phenotypes, with effects on both the amplitude and phase of the daily locomotor peaks. This 'gain of function' approach reveals a multi-hour range of potential phases for both the Morning and Evening activity peaks, within which neuropeptide PDF:PDFR signaling normally specifies rhythmic behavior, according to season. In addition, using a structure-function approach, this work identifies specific PDFR sequence elements that are major points at which the duration of receptor activity is regulated, and through which behavior is modulated in season-specific fashion (Li, 2022).
A gain-of-function approach was used to measure neuropeptide GPCR function in vivo. In doing so, attempts were made to learn about the maximal consequences of PDFR GPCR signaling and also learn something about the phosphorylation events that normally control the duration of PDFR's signaling lifetime. Expression of a PDF receptor variant, containing numerous Ala-substitutions of conserved phosphorylatable residues in the C terminal domain (1-7A), fundamentally altered rhythmic locomotor behavior in Drosophila. The changes included increases in the amplitudes of the Morning and/or Evening activity peaks, and delays in the Phase of the Evening peak. Such results are generally consistent with a prediction whereby non-phosphorylatable PDFR variants-those with a potential to increase the duration of PDFR signaling-would produce behavioral actions opposite to those seen in loss-of-function pdf, or pdfr mutant stocks. Accordingly, it is speculated that phosphorylation of the PDFR at these conserved residues normally defines the duration of its active signaling state each day. The longer duration that follows the substitutions of Ala may allow receptor signal strength to also increase by accrual: such an effect could explain the increased amplitude of the Morning and Evening peaks, as seen with expression of some of the variants. In this context, it is acknowledged that modification of GPCRs by phosphorylation does not exclusively lead to signal termination. GRK2-dependent phosphorylation of the smoothened (smo) GPCR follows reception of the Hh signal and helps mediate its signal: phosphorylation leads to Smo activation in both Drosophila and mammalian systems. Thus modification of putative phosphorylation sites on GPCRs (to preclude phosphorylation) will not exclusively promote extended signaling. Therefore the interpretations must correspondingly consider outcomes without a priori assumption of mechanisms (Li, 2022).
PDFR belongs to the Secretin Receptor Family (Family B) of neuropeptide receptors: there is no clear consensus regarding mechanisms of desensitization and internalization for this receptor family. For VPAC2 receptors, phosphorylation and internalization is mediated exclusively by GRK2. Likewise, Drosophila orthologues of Cortocotrophin Releasing Factor receptors (CG8422, DH44-R1) and Calcitonin receptors (CG17415, DH31-R1) are internalized in hEK cells following recruitment of β-arrestin2. In contrast, PDFR, which is also related to the mammalian Calcitonin receptor, is not internalized following exposure to PDF. VPAC2 receptors are also regulated by heterologous receptor signaling: M3 cholinergic receptors via PKC signaling can block VPAC2 phosphorylation, desensitization and internalization. Furthermore, secretin receptors and VPAC1 receptors undergo phosphorylation by GRKs and β-arrestin2-dependent desensitization, but these are not sufficient to facilitate or mediate internalization. Finally, GLP2-Receptor associates with β-arrestin2 via its distal C terminal sequences, but that receptor domain is required neither for GLP2-R desensitization nor its internalization. Thus kinases other than GRKs and effectors other than β-arrestin2 may regulate internalization of diverse Family B receptors (Li, 2022).
Following activation of rhodopsin in the mammalian retina, visual arrestin is recruited with a time constant of < 80 ms; for many Drosophila neuropeptide receptors, β-arrestin2 is recruited within a minute of exposure to ligand. The PDFR GPCR signals over a time base of many hours: could phosphorylation/de-phosphorylation also regulate its activity? The mammalian blue-light sensitive GPCR melanopsin (OPN4) mediates intrinsic light sensitivity in certain classes of retinal ganglion cells (RGCs): melanopsin signaling is distinguished by long latencies, graded responses and sustained RGC depolarization that can outlast the duration of the light stimulus. Melanopsin-expressing RGCs produce exceptionally long time-course integration because that GPCR behaves differently from other opsins in two fundamental ways. Firstly, it undergoes photoequilibration between signaling and silent states: a property that maintains the availability of pigment molecules for activation, termed 'tristability'. Secondly, phosphorylation and β-arrestin recruitment do contribute to the kinetics of melanopsin signaling, but over a long time-course. It has been proposed that the distal portion of the melanopsin C terminal tail creates a steric blockade covering a cluster of specific serine residues situated in more proximal regions. The time required for relief of that blockade dictates the time course of GPCR phosphorylation and therefore delays subsequent desensitization (Li, 2022).
The 1-7A PDFR variant modified the greatest number of phosphorylatable residues in the series that was tested, and typically produced the strongest behavioral effects. Notably, the 6A variant-which modified only two specific Ser residues-produced effects that were nearly identical to those exhibited by flies expressing the 1-7A form. This suggests that much PDFR post-translational modification, that which is capable of restricting its signaling time-course, may be directed to specific phosphorylation sites near the extreme C terminal end. There are two broadly divergent hypotheses to describe the mechanisms by which GPCR phosphorylation promotes desensitization. The first proposes that modification of specific residues have the greatest significance for downstream desensitizing mechanisms: such a mechanism can explain the effects seen with the highly limited 6A PDFR variant. The second hypothesis invokes triggering of a termination processes by the aggregate negative charge accumulated with bulk phosphorylation, regardless of where it might occur along a GPCR's intracellular sequences. For some GPCRs (e.g., OPN4), the evidence supports both models. The current data concerning PDFR desensitization also support both viewpoints and a model is presented (see A model predicting the effects of phosphorylating different sites on the PDFR CT on downstream signaling.). In particular CL6 and CL7 are considered to be the specific residues especially critical in terminating the time course of PDFR signaling. Notably, converting just the single pair of CL6 AAs, or the single pair of CL7 AAs, is enough to generate hours-long delays in the peaks of locomotor rhythms under Light:Dark conditions. It is noted that the splicing event that distinguishes the PDFR-A and PDFR-D isoforms occurs just prior to the position of sequences encoding CL6 and 7, such that the D form lacks these two highly conserved domains (flybase.org/download/sequence/FBgn0260753/FBpp). Other clusters (like CL2-3) also appear to have potential for 'specific' contributions to PDFR regulation. Together, these results speak to the potency of the inferred specific termination mechanisms for PDFR GPCR signaling. In contrast, the evidence for the bulk phosphorylation hypothesis comes from comparison of the 1-4A, versus the 1-5A, 1-6A and 1-7A variants. This series inactivates increasing numbers of phosphorylatable residues, and with it increasingly delayed evening phases in short day conditions are observed. The tandem mass-spectroscopy results, indicating endogenous phosphorylation of certain PDFR residues demonstrates such post-translational modifications can occur. However, it is cautioned that these phospho-peptide measurements derive from whole head extracts and may not provide a complete or relevant accounting of those sites that are modified in critical E pacemaker neurons (Li, 2022).
The results also point to what is proposed as 'context-dependent' effects of modifying specific GPCR residues-ones that may produce opposing behavioral effects, depending on the phosphorylation status of neighboring sites. In particular, opposing results were observed with Ala-variants of the CL5 site. In comparing results from the series 1-4A, 1-5A, 1-6A and 1-7A, it was found that under Short Day conditions, the 1-4A had only mild effects on the evening peak phase compared to over-expressing WT PDFR. Whereas 1-5A, 1-6A and 1-7A all produced significant delays. The difference between the 1-$A and 1-5A variants is a change in a single S residue at CL5 (S633A): Those observations suggest modification of the CL5 residue normally promotes desensitization of PDFR and termination of PDFR signaling. However, the PDFR variant 5-7A was constructed with the expectation that it would be as effective as 1-5A, 1-6A or 1-7A in delaying the evening phase. Instead it proved only weakly effective. This same mis-match of expected behavioral effects for 5-7A was also seen in other photoperiodic conditions. Likewise, Langlet (2005) reported that effects on mutating Serine residues in the
terminus of VPAC1 did not produce additive effects. It is proposed that phosphorylation of CL5 will have either positive or negative consequences on PDFR desensitization depending on which other neighboring residues are also modified. Thus it is speculated that CL5 takes on outsize importance in determining desensitization rates for PDFR and so may itself be subject to exceptional regulation. Such complex interactions between phosphorylation sites is reminiscent of interactions documented between diverse phosphorylation sites in the circadian clock protein PERIOD. Better resolution of these two paradoxical mechanisms [(1) specific phosphorylation versus bulk negative charge, and (2) context-dependent effects of CL5 site phosphorylation] awaits precise molecular definition of where and when PDFR is modified in key pacemaker neurons in vivo, and by which post-translational modifications (Li, 2022).
The actions of PDFR variants that were described, following substitutions of Ala for various Ser/Thr or Tyr residues, are not easily explained by a hypothesis invoking neomorphic or constitutive GPCR properties. The evidence for this conclusion is three-fold. First, the effects on both amplitude and phase of activity peaks by these variants are not of a random assortment: rather they are all strictly opposite to that of loss-of-function models for pdf and pdfr. Second, the actions of PDFR variants display strong dependence on wild type pdf gene function: this strongly argues that the actions of PDFR variants reflect responses to normal, endogenous PDF signaling. Third, even the strongest actions of PDFR variants on the phases and amplitudes of locomotor activity (that of the 1-7A variant) reflected PDFR sequence variation, and not the properties of the GFP C-terminal fusion. Together these observations support the hypothesis that phosphorylation of some or all of the C terminal residues that were studied are normally modified to attenuate the strength and duration of PDFR signaling during the 24-hr day (Li, 2022).
PDF neuropeptide actions were discovered in the context of physiological adaptions to daylight in crustacea. PDF expression was discovered within a defined subset of the insect Drosophila circadian pacemaker network. Pdf has been shown to make a fundamental contribution by setting the normal behavioral phase of Evening activity in Drosophila during light:dark entrainment and promoting normal rhythmicity during constant darkness. PDF signaling coordinates with that driven by light: It works in parallel to direct photosensitivity along with the CRY blue light photoreceptor. Flies doubly-mutant for cry and pdf display pronounced deficits of locomotor rhythmicity, and can be as severe as measured in clock-deficient flies. PDF and environmental light also work coordinately at the level of neuronal activity patterns: in the case of the evening pacemakers (LNd and the 5th s-LNv), both light and PDF signaling promote a delay in their PER-dependent activation period, and together help align it to a phase just prior to dusk. Based on the close association of PDF signaling and photoperiodic signaling, it is not surprising that the duration of the photoperiod strongly influenced the effects of PDFR sequence variants on the Morning versus the Evening activity peaks. Delayed Evening activity peak produced by PDFR gain-of-function variants under Short Day conditions has been reported: that delay appears strongly inhibited by light durations > 8 hr. It remains unclear at what molecular level light exerts such an inhibitory effect, however, a similarity is noted of these observations with effects recently reported from loss-of-function states for the phosphatase PRL-1. Like the most active PDFR variants this study described (e.g., 6A, 7A, 1-5A 1-6A and 1-7A), PRL-1 mutants produce a 3-4 h delay in the phase of the evening activity peak, but only under winter-like (short day) conditions, and not under equinox or summer-like (long day) conditions. It has been shown that TIM phosphorylation is affected by PRL-1 activity and suggest the seasonal action of PRL-1 to advance the evening locomotor activity phase is mediated by modification of TIM levels within the transcription-translation feedback loop. The extensive similarity of these two sets of behavioral phenotypes reveals either serial or parallel pathways to effect comparable outcomes on locomotor activity. If serial, then according to the simplest model, PRL1 acts downstream of PDFR, and the combined results to-date suggest PRL-1 would be inhibited by PDFR activation. Further genetic and biochemical experiments are needed to evaluate if and how these pathways converge (Li, 2022).
Using an in vitro assay for cyclic AMP generation, this study found that modifying phosphorylation properties of PDFR does not affect the strength of signaling. That conclusion suggests that in vivo other GPCR features are normally affected to regulate the extent/duration of PDFR signaling according to season. This speculation corresponds to findings described with other Family B GPCRs like VPAC1: Mutation of all the Ser and Thr residues of the C terminal tail, and of Ser250 to Ala, led to a receptor with binding properties and adenylate cyclase activity similar to the wild type receptor; however that variant receptor was neither phosphorylated nor internalized. It is proposed that the PDFR variants this study has described do not modify locomotor behavior by virtue of greater second messenger signaling. Rather they do so by generating cAMP (and perhaps other second messengers) over time periods longer than that normal sustained by a WT receptor over a portion of each 24-hr cycle (Li, 2022).
Is PDFR modulation of diurnal phase independent of its modulation of circadian period? A conventional interpretation of the phenotypes that have been described is that PDFR signaling affects the pace of the molecular clock (re-setting the period) and consequently affects behavioral phase indirectly, which is downstream of the clock. However, PDFR variants reliably delayed the Evening activity phase under winter-like conditions, but these variants did not significantly lengthen circadian period. All effects of the 1-7A variant on circadian period were derived from the GFP fusion. Therefore, an alternative hypothesis cannot be ruled out: that the duration of PDF>PDFR signaling to target pacemakers in the Drosophila brain is modulated each day to directly delay Evening pacemaker neuronal activity independent of its effects on entrainment of the molecular clock. In fact, the pdfr han mutant flies exhibited quasi-normal tau values in this report, although they more traditionally exhibit shortened ones. Evening locomotor activity derives in large part from the activity of the primary Evening oscillators, termed E cells, the LNd and the 5th small LNv. However, numerous network interactions are also known to provide critical contributions to the accuracy, precision and adaptability of that timing system. The Evening oscillator group itself is known to be heterogeneous and to constitute at least three separate, functionally distinct oscillators, only some of which express PDFR-A and respond to PDF. It has been reported that the phase of Period protein accumulation in E neurons is phase-locked to the previous lights-off transition and also sensitive to PDF signaling. It was concluded that the peak of PER accumulation is key to determining the phase of the evening activity peak, but also note that the correlation between the Period-clock timing cue and the Evening activity peak is not perfect. It has been suggested the Evening behavioral phase is better described by also factoring in the delay in E cell neuronal activity driven by PDF neuromodulation. It is speculated that it is the combination of two separate PDFR signaling effects that is essential for proper rhythmic behavioral outcomes across seasons. If valid, the hypothesis predicts a bifurcation of signaling pathways downstream of PDFR, to independently regulate diurnal phase and also circadian period within E pacemaker neurons (Li, 2022).
Sleep during development is involved in refining brain circuitry, but a role for sleep in the earliest periods of nervous system elaboration, when neurons are first being born, has not been explored. This study has identified a sleep state in Drosophila larvae that coincides with a major wave of neurogenesis. Mechanisms controlling larval sleep are partially distinct from adult sleep: octopamine, the Drosophila analog of mammalian norepinephrine, is the major arousal neuromodulator in larvae, but dopamine is not required. Using real-time behavioral monitoring in a closed-loop sleep deprivation system, sleep loss in larvae was found to impair cell division of neural progenitors. This work establishes a system uniquely suited for studying sleep during nascent periods, and demonstrates that sleep in early life regulates neural stem cell proliferation (Szuperak, 2018).
Sleep and metabolism are physiologically and behaviorally intertwined; however, the molecular basis for their interaction remains poorly understood. This study identified a serine metabolic pathway as a key mediator for starvation-induced sleep suppression. Transcriptome analyses revealed that enzymes involved in serine biosynthesis were induced upon starvation in Drosophila melanogaster brains. Genetic mutants of astray (aay), a fly homolog of the rate-limiting phosphoserine phosphatase in serine biosynthesis, displayed reduced starvation-induced sleep suppression. In contrast, a hypomorphic mutation in a serine/threonine-metabolizing enzyme, serine/threonine dehydratase (stdh), exaggerated starvation-induced sleep suppression. Analyses of double mutants indicated that aay and stdh act on the same genetic pathway to titrate serine levels in the head as well as to adjust starvation-induced sleep behaviors. RNA interference-mediated depletion of aay expression in neurons, using cholinergic Gal4 drivers, phenocopied aay mutants, while a nicotinic acetylcholine receptor antagonist selectively rescued the exaggerated starvation-induced sleep suppression in stdh mutants. Taken together, these data demonstrate that neural serine metabolism controls sleep during starvation, possibly via cholinergic signaling. It is proposed that animals have evolved a sleep-regulatory mechanism that reprograms amino acid metabolism for adaptive sleep behaviors in response to metabolic needs (Sonn, 2018).
Salt (sodium chloride) is an essential dietary requirement, but excessive consumption has long-term adverse consequences. A high-salt diet (HSD) increases the risk of chronic diseases such as cardiovascular conditions and diabetes and is also associated with poor sleep quality. Little is known, however, about the neural circuit mechanisms that mediate HSD-induced sleep changes. This study sought to identify the effects of HSD on the sleep and related neural circuit mechanisms of Drosophila. Strikingly, it was found that HSD causes young Drosophila to exhibit a fragmented sleep phenotype similar to that of normal aging individuals. Importantly, it was further shown that HSD slightly impairs circadian rhythms and that the HSD-induced sleep changes are dependent on the circadian rhythm system. In addition, it was demonstrated that HSD-induced sleep changes are dopaminergic-system dependent. Together, these results provide insight into how elevated salt in the diet can affect sleep quality (Xie, 2019).
This study has shown that R8 photoreceptors split visual perception and circadian photoentrainment by co-transmitting two neurotransmitters. This visual segregation is further supported by postsynaptic circuitry in the medulla, such that each unicolumnar neuron mainly receives histaminergic inputs from a single R8 photoreceptor (thus transmitting the fine retinotopic signal), whereas each multicolumnar accessory medulla-innervating, multicolumnar and arcuate neuron (AMA neuron) integrates cholinergic inputs from up to 100 R8 photoreceptor cells (thus integrating the irradiance visual signal). These AMA neurons directly excite downstream clock neurons, forming a shallow three-node circuit for circadian photoentrainment. Thus, this clock-entrainment circuit integrates irradiance signals directly from conventional photoreceptors, bypassing the downstream image-forming processing circuit to avoid a less efficient reconstruction of irradiance signals from the already highly processed signals in the contrast-encoding visual pathways. Notably, the AMA neurons overlap with recently discovered xCEOs that sustain circadian timekeeping of free-running circadian clocks (Tang, 2022). Together, these findings reveal that the AMA neurons and xCEOs play crucial roles in circadian timekeeping, resetting circadian clocks to synchronize their endogenous rhythms to local time by integrating irradiance signals from retinal photoreceptors under LD cycles and sustaining free-running circadian timekeeping through their intrinsic rhythmic electrical oscillations under constant darkness (Xiao, 2023).
The observation that the compound eye-driven electrical responses in clock neurons were reduced by half in the absence of histamine receptors suggests that conventional photoreceptors other than R8 photoreceptors (for example, R1-R6) can use histamine-mediated circuitry pathways to excite clock neurons for circadian photoentrainment. The downstream circuits of R1-R6 photoreceptors might reconstruct irradiance signals from image-forming signals via yet-to-be-uncovered mechanisms as mammalian rod and cone pathways do, indicating the mechanisms underlying irradiance encoding for circadian photoentrainment by conventional photoreceptor pathways might have evolved convergently (Xiao, 2023).
This study also identified an unexpected crosstalk between the image-forming vision and circadian photoentrainment. The chloride channel HisCl1 mediates negative feedback of histamine in R8 photoreceptor cells, thus dynamically reducing photoreceptor depolarization during long light stimulation. This feedback regulation tunes ACh release to avoid its local depletion so that the irradiance signal can be continuously transmitted from R8 photoreceptors to clock neurons during the entire light phase (Xiao, 2023).
This work demonstrates that visual perception and circadian photoentrainment can be segregated as early as the first-order synapses in the visual system, providing a simple yet robust mechanism to enact distinct sensory functions. Furthermore, although cotransmission or co-release of neurotransmitters is an emerging principle in brain research, its behavioural significance remains largely unknown. These finding that cotransmission from the same photoreceptor cells enables segregation and translation of distinct visual features into different behaviours also paves the way for understanding this key, indispensable aspect of the nervous system (Xiao, 2023).
Circadian rhythms offer an excellent opportunity to dissect the neural circuits underlying innate behavior because the genes and neurons involved are relatively well understood. This study sought to understand how Drosophila clock neurons interact in the simple circuit that generates circadian rhythms in larval light avoidance. Genetics was used to manipulate two groups of clock neurons, increasing or reducing excitability, stopping their molecular clocks, and blocking neurotransmitter release and reception. The results revealed that lateral neurons (LNvs) promote and dorsal clock neurons (DN1s) inhibit light avoidance, these neurons probably signal at different times of day, and both signals are required for rhythmic behavior. Similar principles apply in the more complex adult circadian circuit that generates locomotor rhythms. Thus, the changing balance in activity between clock neurons with opposing behavioral effects generates robust circadian behavior and probably helps organisms transition between discrete behavioral states, such as sleep and wakefulness (Collins, 2012).
This study identified some of the network logic that helps generate a simple rhythmic behavior through precise genetic manipulations of the larval circadian circuit and extended these findings to the more complex adult circadian network. Previous studies have shown that intercellular signaling in clock neuron networks promotes molecular clock synchrony and can strengthen genetically weak molecular clocks. This study increases the importance of circadian neural networks by finding that non-LNv clock neurons are as important as the 'master' pacemaker LNv clock neurons for rhythmic behavior both in larvae and adult flies. However, LNvs can still be considered pacemakers in DD because most manipulations to non-LNv clock neurons do not affect period length (Collins, 2012).
Non-LNv signals appear to gate pacemaker neuron activity. Why is this necessary when LNvs have their own intrinsic excitability rhythms? It is proposed that the interaction of two oscillators with opposite signs helps reduce the time when LNvs signal. Without signaling from non-LNvs, adult locomotor activity rhythms are weak and activity is distributed throughout the day and night as in tim-Gal4; Pdf-Gal80 > dORKΔC flies. In contrast, in tim-Gal4; Pdf-Gal80 > NaChBac flies, the timing of locomotor activity is narrowed. Thus, the gating of LNv activity by non-LNvs may help turn gradual changes in the excitability of each neuronal group into thresholds that promote a switch in overall output and allow flies to abruptly transition from inactivity to activity (Collins, 2012).
This gating system can only function if LNvs
and non-LNvs have differently phased neuronal activity.
However, most Drosophila clock neurons have similarly phased
molecular clocks. It is proposed that molecular clocks in different
clock neurons regulate divergent sets of output genes to generate
distinct phases of neuronal excitability. This would be analogous to
the mammalian circadian system, in which molecular clocks in different
tissues drive tissue-specific outputs. In summary, this genetic
dissection of a circadian neural circuit reveals an unexpected and
essential role for inhibitory signals from non-LNvs (E
cells) in shaping activity profiles at dawn and a mechanism for how
clock neurons couple together to promote robust rhythms (Collins,
2012).
The circadian system is comprised three components: a network of core clock cells in the brain that keeps time, input pathways that entrain clock cells to the environment, and output pathways that use this information to ensure appropriate timing of physiological and behavioral processes throughout the day. Core clock cells can be divided into molecularly distinct populations that likely make unique functional contributions. This study clarifies the role of the dorsal neuron 1 (DN1) population of clock neurons in the transmission of circadian information by the Drosophila core clock network. Using an intersectional genetic approach that allowed selective and comprehensive targeting of DN1 cells, this study shows the suppressing DN1 neuronal activity alters the magnitude of daily activity and sleep without affecting overt rhythmicity. This suggests that DN1 cells are dispensable for both the generation of circadian information and the propagation of this information across output circuits (Nettnin, 2021).
Animals display a body body temperature rhythm (BTR). Little is known about the mechanisms by which a rhythmic pattern of BTR is regulated and how body temperature is set at different times of the day. As small ectotherms, Drosophila exhibit a daily temperature preference rhythm (TPR), which generates BTR. This study demonstrates dorsal clock networks that play essential roles in TPR. Dorsal neurons 2 (DN2s) are the main clock for TPR. It was found that DN2s and posterior DN1s (DN1ps) contact and the extent of contacts increases during the day and that the silencing of DN2s or DN1ps leads to a lower temperature preference. The data suggest that temporal control of the microcircuit from DN2s to DN1ps contributes to TPR regulation. This study also identified anterior DN1s (DN1as) as another important clock for TPR. Thus, this study shows that the DN networks predominantly control TPR and determine both a rhythmic pattern and preferred temperatures (Chen, 2022).
A newly formed memory is initially unstable. However, if it is consolidated into the brain, the consolidated memory is stored as stable long-term memory (LTM). Despite the recent progress, the molecular and cellular mechanisms of LTM have not yet been fully elucidated. The fruitfly Drosophila melanogaster, for which various genetic tools are available, has been used to clarify the molecular mechanisms of LTM. Using the Drosophila courtship-conditioning assay as a memory paradigm, it was previously identified that the circadian clock gene period (per) plays a vital role in consolidating LTM, suggesting that per-expressing clock neurons are critically involved in LTM. However, it is still incompletely understood which clock neurons are essential for LTM. This study shows that dorsal-lateral clock neurons (LNds) play a crucial role in LTM. Using an LNd-specific split-GAL4 line, it wa confirmed that disruption of synaptic transmission in LNds impaired LTM maintenance. On the other hand, induction of per RNAi or the dominant-negative transgene of Per in LNds impaired LTM consolidation. These results reveal that transmitter release and Per function in LNds are involved in courtship memory processing (Suzuki, 2022).
Drosophila's lateral posterior neurons (LPNs) belong to a small group of circadian clock neurons that is so far not characterized in detail. Thanks to a new highly specific split-Gal4 line, this study describes LPNs' morphology in fine detail, their synaptic connections, daily bimodal expression of neuropeptides, and a putative role of this cluster in controlling daily activity and sleep patterns is proposed. The three LPNs were found to be heterogeneous. Two of the neurons with similar morphology arborize in the superior medial and lateral protocerebrum and most likely promote sleep. One unique, possibly wakefulness-promoting, neuron with wider arborizations extends from the superior lateral protocerebrum toward the anterior optic tubercle. Both LPN types exhibit manifold connections with the other circadian clock neurons, especially with those that control the flies' morning and evening activity (M- and E-neurons, respectively). In addition, they form synaptic connections with neurons of the mushroom bodies, the fan-shaped body, and with many additional still unidentified neurons. Both LPN types rhythmically express three neuropeptides, Allostatin A, Allostatin C, and Diuretic Hormone 31 with maxima in the morning and the evening. The three LPN neuropeptides may, furthermore, signal to the insect hormonal center in the pars intercerebralis and contribute to rhythmic modulation of metabolism, feeding, and reproduction. These findings are discussed in the light of anatomical details gained by the recently published hemibrain of a single female fly on the electron microscopic level and of previous functional studies concerning the LPN (Reinhard, 2021).
Though much is known about the cellular and molecular components of the circadian clock, output pathways that couple clock cells to overt behaviors have not been identified. A screen was conducted for circadian-relevant neurons in the Drosophila brain, and this study reports that cells of the pars intercerebralis (PI), a functional homolog of the mammalian hypothalamus, comprise an important component of the circadian output pathway for rest:activity rhythms. GFP reconstitution across synaptic partners (GRASP) analysis demonstrates that PI cells are connected to the clock through a polysynaptic circuit extending from pacemaker cells to PI neurons. Molecular profiling of relevant PI cells identified the corticotropin-releasing factor (CRF) homolog, DH44, as a circadian output molecule that is specifically expressed by PI neurons and is required for normal rest:activity rhythms. Notably, selective activation or ablation of just six DH44+ PI cells causes arrhythmicity. These findings delineate a circuit through which clock cells can modulate locomotor rhythms (Cavanaugh, 2014).
Given its location near the axonal projections of several groups of clock neurons and its function in metabolic, locomotor, and sleep processes, the PI has been proposed as a possible component of the output pathway in Drosophila, but direct evidence supporting a contribution to behavioral or physiological rhythms has been lacking. This study used a combined genetic, anatomical, and molecular approach to unequivocally identify specific subsets of PI cells as comprising part of the circadian output circuit for rest:activity rhythms. Ectopic activation of PI neurons is sufficient to induce behavioral arrythmicity, and similarly, ablation of small subsets of PI neurons results in loss of rest:activity rhythms. This latter result is consistent with previous studies showing that surgical destruction of the PI in both crickets and cockroaches results in loss of locomotor rhythms. It was further shown that manipulations of the PI that result in behavioral arrhythmicity do not affect the underlying molecular clock in s-LNvs, thus demonstrating that the PI exerts its effects downstream of clock neurons (Cavanaugh, 2014).
Importantly, this study has uncovered a segregation of different behavioral and physiological outputs by specific neurons of the PI. Thus, kurs58-GAL4+ PI neurons function to modulate locomotor behavior, whereas insulin-like peptide-producing PI cells, which constitute a nonoverlapping subset, influence metabolic processes. It will be of interest to determine whether Dilp2+ cells are also modulated by the clock, because such a result would suggest that the PI is a common relay for multiple circadian output circuits that couple to unique physiological functions, each subserved by discrete subpopulations of PI neurons. Furthermore, within kurs58-GAL4+ cells, there appear to be at least two subsets of neurons that contribute to rest:activity cycles. Interestingly, ablation of the SIFa-GAL4+ subset results in reduced rhythmicity, accompanied by decreases in sleep, whereas ablation of the DH44VT-GAL4+ subset also results in reduced rhythmicity, but in this case, the effect on sleep, if any, is an increase. Thus, it is possible that these two molecularly distinct populations control behavioral rhythms through opposing effects on locomotion and/or sleep, and thus, that the contribution of a particular subset predominates depending on time of day (Cavanaugh, 2014).
In conjunction with behavioral studies, GRASP analysis was used to trace neuronal connections emanating from the clock network. It was found that s-LNvs, which function as master pacemakers, make limited connections within the clock cell network and do not appear to directly access output cells of the PI. Instead, PI output cells receive time-of-day information through inputs from DN1 clock cells, as demonstrated by the fact that presynaptic components of DN1 cells adjoin dendrites of PI neurons, in the same brain region where GRASP analysis reveals cellular contacts between these two cell groups. Several studies corroborate a function of DN1 neurons downstream of s-LNvs to mediate rest:activity rhythms. Dorsal neurons are responsive to bath application of PDF, and restoration of the PDF receptor selectively in these neurons of pdfr mutant flies is sufficient to rescue multiple aspects of circadian locomotor rhythms. Furthermore, speeding up the molecular clock in s-LNvs causes concomitant acceleration of molecular cycling in several groups of dorsal neurons, including DN1s. These experiments, along with the current study, argue that DN1 neurons serve an important output function within the clock network and likely reside downstream of s-LNvs in the output circuit for rest:activity rhythms. The data are therefore consistent with a very simple circadian output circuit, in which time-of-day information from the clock network, which is generated by master pacemaker cells (s-LNvs and possibly LNds), passes through dorsal clock neurons (including DN1s) before accessing downstream output neurons of the PI, which then integrate these signals to modulate locomotor rhythms. Whether the PI also lies downstream of other groups of dorsal clock neurons, in addition to DN1s, or whether all time-of-day signals received by the PI pass through DN1 cells remains to be determined (Cavanaugh, 2014).
Within the brain, projections from the PI primarily terminate in the dorsal tritocerebrum; however, more diffuse termination patterns throughout the central brain and optic lobes have been observed for SIFa+ PI neurons. The PI also accesses neurohemal organs via the esophageal canal, as well as directly releasing peptides into the hemolymph. Thus, signals released from the PI could either act within neuronal tissue or systemically via release of peptide neurotransmitters and other hormones. The latter possibility is consistent with studies that showed that transplantation of pers brains into the abdomen of per mutant flies rescued locomotor rhythms, demonstrating that release of a secreted factor underlies brain control of rest:activity rhythms in flies. Similarly, abdominal transplantation of PI cells is sufficient to alter sexually dimorphic locomotor patterns, indicating that the PI can modulate locomotor behavior in a neuroendocrine manner (Cavanaugh, 2014).
Through single-cell transcriptome analysis, the CRF-like peptide, DH44, was identified as a candidate molecule through which PI neurons might influence locomotor behavior. Consistent with this possibility, RNAi-mediated knockdown, or genetic antagonism, of DH44 resulted in altered locomotor behavior and weakened rest:activity rhythms. In addition, selective activation or destruction of DH44+ PI neurons also substantially weakened rest:activity rhythms. In flies, DH44 acts as a diuretic hormone, which stimulates fluid secretion from Malpighian tubules through a cyclic AMP (cAMP) pathway. Its role as a stress molecule is less clear, but DH44 receptor has also been localized to corazonin+ cells of the lateral protocerebrum, which may be involved in the stress response of the fly. Notably, manipulations of neuronal excitability in corazonin+ cells alter stress-induced locomotor activity. In mammals, stress hormones, such as glucocorticoids, show diurnal cycles of secretion and serve as entrainment signals for peripheral clocks. Thus, stress hormones may play a conserved role in circadian regulation of behavioral and physiological processes (Cavanaugh, 2014).
How to build and maintain a reliable yet flexible circuit is a fundamental question in neurobiology. The nervous system has the capacity for undergoing modifications to adapt to the changing environment while maintaining its stability through compensatory mechanisms, such as synaptic homeostasis. This study describes findings in the Drosophila larval visual system, where the variation of sensory inputs induces substantial structural plasticity in dendritic arbors of the postsynaptic neuron and concomitant changes to its physiological output. Furthermore, a genetic analysis has identified the cyclic adenosine monophosphate (cAMP) pathway and a previously uncharacterized cell surface molecule as critical components in regulating experience-dependent modification of the postsynaptic dendrite morphology in Drosophila (Yuan, 2011).
Proper functions of neuronal circuits rely on their fidelity, as well as plasticity, in responding to experience or changing environment, including the Hebbian form of plasticity, such as long-term potentiation, and the homeostatic plasticity important for stabilizing the circuit. Activity-dependent modification of neuronal circuits helps to establish and refine the nervous system and provides the cellular correlate for cognitive functions, such as learning and memory. Multiple studies have examined synaptic strength regulation by neuronal activity, whereas to what extent and how the dendritic morphology may be modified by neuronal activity remain open questions (Yuan, 2011).
In Drosophila larvae, Bolwig's organ (BO) senses light, and its likely postsynaptic targets are LN(v)s. As the major circadian pacemaker, LN(v)s are important for the entrainment to environmental light-dark cycles and larval light avoidance behavior. In the larval brain, Bolwig's nerve (BN), the axonal projection from BO, terminates in an area overlapping the dendritic field of LN(v)s. Using the FRT-FLP system [in which DNA sequences flanked by flippase recognition targets (FRT) are snipped out by flippase (FLP)] along with three-dimensional (3D) tracing, the dendritic arbor of individual LN(v) neurons were labeled and analyzed. Then potential synaptic connections were demonstrated between BN and LN(v)s using the GRASP [green fluorescent protein (GFP) reconstitution across synaptic partners] technique to drive expression of one-half of the split GFP in the BN by means of Gal4/UAS and expression of the other half of the split GFP in LN(v)s via LexA/LexAop. The proximity of putative synaptic connections between BN and LN(v)s' dendrites reconstituted GFP fluorescence for photoreceptors expressing either rhodopsin 5 (Rh5) or rhodopsin 6 (Rh6) in BO, which suggested that both groups of photoreceptors may have synaptic connections with LN(v)s (Yuan, 2011).
To test whether LN(v)s can be activated by BN inputs through light stimulation, calcium imaging was performed using GCaMP3 transgenic flies with the larval brain-eye preparation, which included BO, BN, developing eye disks, the larval brain, and ventral nerve cord. Because BO senses blue and green light, the confocal laser at 488 nm (blue) and 543 nm (green) were used to stimulate these larval photoreceptors. LN(v)s' axon terminals displayed a relatively stable baseline of GCaMP3 fluorescence and, upon light stimulation, yielded large calcium responses, which increased with the greater intensity and longer duration of the light pulses (Yuan, 2011).
Recent studies suggest that Cryptochrome (CRY) in adult large LN(v)s senses light and elicits neuronal firing. In larvae, however, severing BN abolished light-induced calcium responses in LN(v)s. The loss-of-function mutation of NorpA (no-receptor-potential A), encoding a phospholipase C crucial for phototransduction, also eliminated these calcium responses, which indicated that light-elicited responses in LN(v)s are generated via phototransduction in larval photoreceptors rather than as a direct response to light by LN(v)s (Yuan, 2011).
In animals with BO genetically ablated, the dendritic field of LN(v) is absent. To test whether BO is required for LN(v)s' dendrite maintenance, the expression of cell death genes rpr and hid was induced in BO after synapse formation, and the LN(v) dendrite length was also found to be greatly reduced. Whereas physical contacts with BN or growth-promoting factors released from presynaptic axons could be important for LN(v)s' dendrite maintenance, it is also possible that synaptic activity from BN promotes LN(v) dendrite growth, as suggested by previous studies. To explore the latter scenario, newly hatched larvae were provided with different visual experiences through various light regimes-including the standard 12 hours of light and 12 hours of dark daily cycle (LD); constant darkness (DD) for sensory deprivation; constant light (LL) for enhanced light input; 16-hour light and 8-hour dark cycle, mimicking a long day; and 8-hour light and 16-hour dark cycle, mimicking a short day. The dendrite morphology of LN(v)s of late third instar larvae was examined. Whereas different light exposure had no detectable effects on larval developmental timing, increasing light exposure reduced the total dendrite length of individual LN(v) neurons, with the longest dendrite in constant darkness and the shortest dendrite length in constant light condition. Thus, not only is the LN(v) dendrite dependent on the presence of presynaptic nerve fibers, its length is modified by the sensory experience in a compensatory fashion, whereby an increase in sensory inputs causes a reduction in the dendrite length and vice versa (Yuan, 2011).
Whereas adult LN(v)s alter their axon terminal structures in a circadian cycle-controlled fashion, no difference was found in dendrite morphology of LN(v)s from larvae collected at four different time points around the clock, which indicated that circadian regulation is not involved in the light-induced modification of LN(v) dendrites. Under regular light-dark conditions, LN(v) dendrites expanded as the larval brain size increased from the second to the third instar stage. However, the dendrite length of the LL group increased at a significantly slower rate than the DD group. It thus appears that light exposure retards the growth of LN(v) dendrites throughout the larval development (Yuan, 2011).
To test the contribution of different light-sensing pathways, loss-of-function mutations of Cry (cry01) or NorpA (norpA36) and of double mutants lacking both Rh5 and Rh6 (rh52;rh61) were examined. Although wild-type and cry01 larvae displayed differences in their dendrite length when exposed to constant darkness versus constant light, such light-induced changes were absent in the rh52;rh61 double mutant and the norpA36 mutant. Thus, similar to the calcium response to light, light-induced modification of LN(v) dendritic structure requires visual transduction mediated by rhodopsin and NorpA in BO but not Cry function in LN(v)s (Yuan, 2011).
To manipulate the level of synaptic activity, the BO excitability was weither increased by expressing the heat-activated Drosophila transient-receptor-potential A1 (dTrpA) channel, or transmitter release from BN was reduced through a temperature-sensitive form of the dominant-negative dynamin, Shibirets (Shits). These manipulations eliminated light-induced modification of LN(v) dendrites at 29°C. Reducing BO activity by means of Shits caused dendrite expansion, as if the animal detected no light, whereas increasing BO activity by means of the dTrpA channel resulted in reduction of LN(v) dendrites, a process reminiscent of constant light exposure (Yuan, 2011).
Whether intrinsic LN(v) neuronal activity drives modification of its dendrite morphology was further tested by expression of either the sodium channel NaChBac to increase excitability or the potassium channel Kir2.1 to reduce excitability. LN(v)s expressing Kir2.1 showed reduced or no calcium responses upon light stimulation. In contrast, LN(v)s expressing NaChBac displayed numerous peaks in GCaMP3 signals in the presence or absence of light stimulation, indicative of elevated spontaneous activities. Upon examining LN(v) dendrites, it was found that neuronal excitability of the LN(v) was inversely proportional to its dendrite length (Yuan, 2011).
These results obtained using genetic approaches agreed with findings in experiments with different environmental light conditions. They suggested that LN(v)'s dendritic structures are modified according to its neuronal activity, which varies with light-induced synaptic inputs (Yuan, 2011).
To test whether synaptic contacts of BN on LN(v)s are modified by light, synapses formed by BN with EGFP (enhanced green fluorescent protein)-tagged Cacophony (Cac-EGFP) were marked, because Cacophony is a calcium channel localized at presynaptic terminals and its distribution correlates with the number of synapses. Close association was found of Cac-EGFP-expressing structures with LN(v)s' dendritic arbors. Compared with regular light-dark conditions, constant darkness increased, whereas constant light reduced, the total intensity of Cac-EGFP, which suggested that light modified not only dendritic arbors of LN(v)s but also the number of synaptic contacts impinging on LN(v) dendrites (Yuan, 2011).
Next, using calcium imaging, whether there are light-induced functional modifications of LN(v)s was examined. Increased light exposure caused LN(v)s to be less responsive. Conversely, sensory deprivation in constant darkness increased LN(v)s' sensitivity to light. Thus, in contrast to stable synaptic responses observed in synaptic homeostasis, light-induced responses of central neurons postsynaptic to photoreceptors in the Drosophila larval visual circuit have a dynamic range, modifiable by sensory experiences and positively correlated to the dendrite length (Yuan, 2011).
In dunce1, a loss-of-function mutant of the fly homolog of 3'5'-cyclic nucleotide phosphodiesterase, the LN(v)s' dendrite length was comparable among LD, LL, and DD groups. Reducing dunce gene expression specifically in LN(v)s through RNA interference (dncIR) resulted in a similar indifference of LN(v)s' dendrite size to the light exposure, which implicated a cell-autonomous action of dunce in LN(v) neurons (Yuan, 2011).
To explore the possibility that the elevated cAMP level caused by the dunce mutation interfered with dendrite plasticity, tests were performed for the involvement of downstream components of the cAMP pathway, including the catalytic subunit of protein kinase A (PKAmc), which up-regulates cAMP signaling, and a dominant-negative form of the cAMP response element-binding protein (CREBdn), which inhibits cAMP-induced transcription activation. Expression of either transgene specifically in LN(v)s obliterated their ability to adjust dendrite length under different light-dark conditions. Calcium imaging further revealed that the expression of PKAmc or CREBdn eliminated changes of LN(v)s' light responses produced by different light-dark conditions. Thus, the cAMP pathway regulates both structural and functional plasticity of LN(v)s (Yuan, 2011).
The screen for mutants with defective LN(v) dendritic plasticity also identified babos-1, a mutant with a P-element insertion near the transcriptional start site of CG3624, a previously uncharacterized immunoglobulin domain-containing cell surface protein. The LN(v) dendrite length of babos-1 mutant larvae was comparable to controls in LD and LL but has no compensatory increase in DD. Similar phenotypes were found in larvae expressing an RNAi transgene targeting CG3624 in LN(v)s. Moreover, flies carrying a hypomorphic allele of CG3624, CG3624[KG05061], also showed defective light-induced dendritic plasticity, which was fully rescued by expressing the UAS-CG3624 transgene specifically in LN(v)s. Thus, the function of this immunoglobulin domain-containing protein in LN(v)s is important for the dendrite expansion in constant darkness (Yuan, 2011).
Bioinformatic analyses suggest that CG3624 is a cell surface protein containing an N-terminal signal peptide, extracellular immunoglobulin domains followed by a transmembrane helix, and a short C-terminal cytoplasmic tail. CG3624 is widely expressed in the nervous system throughout development. Its specific requirement for the adjustment of LN(v)s' dendrite length in constant darkness suggests that elevation or reduction of sensory inputs likely invokes separate mechanisms for compensatory modifications of central neuronal dendrites (Yuan, 2011).
A functioning nervous system must have the capacity for adaptive modifications while maintaining circuit stability. This study of the Drosophila larval visual circuit reveals large-scale, bidirectional structural adaptations in dendritic arbors invoked by different sensory exposure. Whereas the circuit remains functional with modified outputs, this type of homeostatic compensation may modify larval light sensitivity according to its exposure during development and could facilitate adaption of fly larvae toward altered light conditions, such as seasonal changes. The observations also suggest shared molecular machinery between homeostasis and the Hebbian plasticity with respect to the cAMP pathway and indicate the feasibility of genetic studies of experience-dependent neuronal plasticity in Drosophila (Yuan, 2011).
The incidence of reproductive diapause is a critical aspect of life history in overwintering insects from temperate regions. Much has been learned about the timing, physiology and genetics of diapause in a range of insects, but how the multiple changes involved in this and other photoperiodically regulated traits are interrelated is not well understood. This study performed quasinatural selection on reproduction under short photoperiods in a northern fly species, Drosophila montana, to trace the effects of photoperiodic selection on traits regulated by the photoperiodic timer and / or by a circadian clock system. Selection changed several traits associated with reproductive diapause, including the critical day length for diapause (CDL), the frequency of diapausing females under photoperiods that deviate from daily 24 h cycles and cold tolerance, towards the phenotypes typical of lower latitudes. However, selection had no effect on the period of free-running locomotor activity rhythm regulated by the circadian clock in fly brain. At a genomic level, selection induced extensive divergence between the selection and control line replicates in 16 gene clusters involved in signal transduction, membrane properties, immunologlobulins and development. These changes resembled ones detected between latitudinally divergent D. montana populations in the wild and involved SNP divergence associated with several genes linked with diapause induction. Overall, this study shows that photoperiodic selection for reproduction under short photoperiods affects diapause-associated traits without disrupting the central clock network generating circadian rhythms in fly locomotor activity (Kauranen, 2019).
Photoperiodic reproductive diapause is an essential part of female life cycle in several insect species living on high latitudes, where overwintering in reproductive stage involves high risks for survival and progeny production. The sensitive period (SP), during which photoperiodic cues can trigger the switch from direct development to diapause, can last from a few hours or days after emergence to the entire life span of females. Moreover, in some species, sexually mature females can enter post-reproductive diapause as a response to decreasing day length and/or temperature. The duration of SP for diapause induction and the females' ability to enter post-reproductive diapause at short day lengths in Drosophila montana strains was assessed from different latitudes in Europe, North America, and Japan. This study shows that the females of this species have a life-long SP and that they retain an ability to switch between reproduction and diapause as a response to back-and-forth changes in day length for at least 3 months. D. montana strains from different latitudes showed high variation in females' ability to enter post-reproductive diapause; females of the southern strains generally requiring longer time and/or lower temperature to enter this stage than those of the northern strains. Moreover, the proportion of females that switched to post-reproductive diapause in 3 weeks in short day conditions at 16 °C showed positive correlation with the critical day length (CDL) for diapause induction and the latitudinal and continental origin of the strains. Life-long SP increases females' flexibility to respond to short-term changes in environmental conditions and enables reproducing females to switch to post-reproductive diapause when the days get shorter and colder toward the autumn. This ability can play a major role in species phenology and should be taken into account in theoretical and empirical studies on insect adaptation to seasonal variation (Lankinen, 2022).
Insect adaptation to climatic conditions at different latitudes has required changes in life-history traits linked with survival and reproduction. Several species, including Drosophila montana, show robust latitudinal variation in the critical day length (CDL), below which more than half of the emerging females enter reproductive diapause at a given temperature. This study used a novel approach to find out whether D. montana also shows latitudinal variation in the critical temperature (CTemp), above which the photoperiodic regulation of diapause is disturbed so that the females develop ovaries in daylengths that are far below their CDL. CTemp was estimated for 53 strains from different latitudes on 3 continents after measuring their diapause proportions at a range of temperatures in 12 h daylength (for 29 of the strains also in continuous darkness). In 12 h daylength, CTemp increased towards high latitudes alongside an increase in CDL, and in 3 high-latitude strains diapause proportion exceeded 50% in all temperatures. In continuous darkness, the diapause proportion was above 50% in the lowest temperature(s) in only 9 strains, all of which came from high latitudes. In the second part of the study, changes were measured in CTemp and CDL in a selection experiment favouring reproduction in short daylength (photoperiodic selection) and by exercising selection for females that reproduce in LD12:12 at low temperature (photoperiodic and temperature selection). In both experiments selection induced parallel changes in CDL and CTemp, confirming correlations seen between these traits along latitudinal clines. Overall, these findings suggest that selection towards strong photoperiodic diapause and long CDL at high latitudes has decreased the dependency of D. montana diapause on environmental temperature. Accordingly, the prevalence and timing of the diapause of D. montana is likely to be less vulnerable to climate warming in high- than low-latitude populations (Lankinen, 2023)
Circadian clocks orchestrate a variety of physiological and behavioural functions within the 24-h day. These timekeeping systems have also been implicated in developmental and reproductive processes that span more (or less) than 24 h. Whether natural alleles of cardinal clock genes affect entire sets of life-history traits (i.e., reproductive arrest, developmental time, fecundity), thus providing a wider substrate for seasonal adaptation, remains unclear. This study shows that natural alleles of the timeless (tim) gene of Drosophila melanogaster, previously shown to modulate flies' propensity to enter reproductive dormancy, differentially affect correlated traits such as early-life fecundity and developmental time. Homozygous flies expressing the shorter TIM isoform (encoded by the s-tim allele) not only show a lower dormancy incidence compared to those homozygous for ls-tim (which produce both the short and an N-terminal additional 23-residues longer TIM isoform), but also higher fecundity in the first 12 days of adult life. Moreover, s-tim homozygous flies develop faster than ls-tim homozygous flies at both warm (25°C) and cold (15°C) temperatures, with the gap being larger at 15°C. In summary, this phenotypic analysis shows that natural variants of tim affect a set of life-history traits associated with reproductive dormancy in Drosophila. It is speculated that this provides further adaptive advantage in temperate regions (with seasonal changes); it is proposed that the underlying mechanisms might not be exclusively dependent on photoperiod, as previously suggested (Andreatta, 2022).
Circadian rhythms in adult eclosion of Drosophila are postulated to be regulated by a pair of coupled oscillators: one is the master clock that is light sensitive and temperature compensated and the other that is a slave oscillator whose period is temperature sensitive and whose phase is reflected in the overt behavior. Within this framework, it was reasoned that in populations of Drosophila melanogaster that have been artificially selected for highly divergent phases of eclosion rhythm, there may be changes in this network of the master-slave oscillator system, via changes in the temperature-sensitive oscillator and/or the coupling of the light- and temperature-sensitive oscillators. Light/dark cycles were used in conjunction with different constant ambient temperatures and two different amplitudes of temperature cycles in an overall cool or warm temperature and analyzed phases, gate width, and normalized amplitude of the rhythms in each of these conditions. The populations selected for eclosion in the morning (early flies) do not vary their phases with change in temperature regimes, whereas the populations selected for eclosion in the evening (late flies) show phase lability of up to ~5 h. These results imply a genetic correlation between timing of behavior and temperature sensitivity of the circadian clock (Abhilash, 2019).
One hypothesis for the function of sleep is that it serves as a mechanism to conserve energy. Recent studies have suggested that increased sleep can be an adaptive mechanism to improve survival under food deprivation in Drosophila melanogaster. To test the generality of this hypothesis, Sleep and its plastic response to starvation was compared in a temperate and tropical population of Drosophila melanogaster. Flies from the temperate population were found to be more starvation resistant, and it was hypothesized that they would engage in behaviors that are considered to conserve energy, including increased sleep and reduced movement. Surprisingly, temperate flies slept less and moved more when they were awake compared to tropical flies, both under fed and starved conditions, therefore sleep did not correlate with population-level differences in starvation resistance. In contrast, total sleep and percent change in sleep when starved were strongly positively correlated with starvation resistance within the tropical population, but not within the temperate population. Thus, unexpectedly complex relationships between starvation and sleep were observed that vary both within and across populations. These observations falsify the simple hypothesis of a straightforward relationship between sleep and energy conservation. The hypothesis that starvation is correlated with metabolic phenotypes was tested by investigating stored lipid and carbohydrate levels; stored metabolites were found to partially contributed towards variation starvation resistance. These findings demonstrate that the function of sleep under starvation can rapidly evolve on short timescales and raise new questions about the physiological correlates of sleep and the extent to which variation in sleep is shaped by natural selection (Sarikaya, 2020).
Lately, Drosophila has been favored as a model in sleep and circadian rhythm research due to its conserved mechanism and easily manageable operation. These studies have revealed the sophisticated parameters in whole-day sleep profiles of Drosophila, drawing connections between Drosophila sleep and human sleep. This study tested several sleep deprivation protocols (mechanical shakes and light interruptions) on Drosophila and delineated their influences on Drosophila sleep. A daytime light-deprivation protocol (DD) was applied mimicking jet-lag to screen drugs that alleviate sleep deprivation. Characteristically, classical sleep-aid compounds exhibited different forms of influence: phenobarbital and pentobarbital modified total sleep time, while melatonin only shortened the latency to sleep. Such results construct the basis for further research on sleep benefits in other treatments in Drosophila. Seven herb extracts were screened, and very diverse results were found regarding their effect on sleep regulation. For instance, Panax notoginseng and Withania somnifera extracts displayed potent influence on total sleep time, while Melissa officinalis increased the number of sleep episodes. By comparing these treatments, it was possible to rank drug potency in different aspects of sleep regulation. Notably, the presence of sleep difficulties in a Drosophila Alzheimer's disease (AD) model was confirmed with an overexpression of human Aβ, and clear differences were recognized between the portfolios of drug screening effects in AD flies and in the control group. Overall, potential drug candidates and receipts for sleep problems can be identified separately for normal and AD Drosophila populations, outlining Drosophila's potential in drug screening tests in other populations if combined with the use of other genetic disease tools (Wang, 2020).
Social isolation and loneliness have potent effects on public health. Research in social psychology suggests that compromised sleep quality is a key factor that links persistent loneliness to adverse health conditions. Although experimental manipulations have been widely applied to studying the control of sleep and wakefulness in animal models, how normal sleep is perturbed by social isolation is unknown. This study reports that chronic, but not acute, social isolation reduces sleep in Drosophila. Quantitative behavioural analysis and transcriptome profiling were used to differentiate between brain states associated with acute and chronic social isolation. Although the flies had uninterrupted access to food, chronic social isolation altered the expression of metabolic genes and induced a brain state that signals starvation. Chronically isolated animals exhibit sleep loss accompanied by overconsumption of food. This resonates with anecdotal findings of loneliness-associated hyperphagia in humans. Chronic social isolation reduces sleep and promotes feeding through neural activities in the peptidergic fan-shaped body columnar neurons of the fly. Artificial activation of these neurons causes misperception of acute social isolation as chronic social isolation and thereby results in sleep loss and increased feeding. These results present a mechanistic link between chronic social isolation, metabolism, and sleep, addressing a long-standing call for animal models focused on loneliness (Li, 2021).
Fruit flies are social animals, and exhibit dynamic social network structures and collective behaviours, that contribute to environmental sensing, foraging, feeding, fighting, mating, oviposition, circadian time setting and even the existence of 'culture'. These important aspects of social interactions imply that insects can provide suitable models for studying how the objective absence of social relationships is perceived and represented in the brain (Li, 2021).
Social experience affects sleep need in Drosophila. This study revisited this finding by exploring how social isolation affects sleep in flies that have prior social experience. Sleep behaviour was tested after maintaining 1, 2, 5, 25 or 100 male flies in a food vial for 7 days. Group-housed flies, regardless of group size (2, 5, 25 or 100), exhibited similar sleep profiles. By contrast, flies housed in isolation displayed a significant loss of sleep, mainly distributed during the daytime (Li, 2021).
The duration of social isolation was manipulated: flies were either isolated or housed in a group of 25 flies for 1, 3, 5, or 7 days, before sleep was measured in a Drosophila activity monitor (DAM). Sleep profiles, which display the proportion of time spent sleeping in consecutive 30-min segments over 24 h, showed that chronic social isolation (5 or 7 days) changed sleep architecture primarily during the daytime and especially during an interval of several hours following dawn (lights on). Although short durations of social isolation (1 or 3 days) did not induce sleep loss, chronic social isolation (5 or 7 days) significantly reduced daily total sleep, daytime sleep and sleep between Zeitgeber time (ZT) 0 and ZT4 (corresponding to the first 4 h after lights-on in a light-dark (LD) cycle) (Li, 2021).
To assess how social isolation alters daytime sleep, all daytime sleep bouts were pooled from all animals tested for a given condition and their distributions were plotted as cumulative relative fractions for bout lengths. Acute social isolation (1 day) produced sleep bout distributions that were indistinguishable from those of 1-day group-housed flies. Flies that were socially isolated for 3 days showed slightly different sleep bout distributions from their group-housed counterparts. However, there was no deficit in total daily sleep, daytime sleep or ZT0-4 sleep in these flies. Flies that were isolated for longer periods (5 or 7 days) had sleep distributions that were significantly different from those of their group-housed counterparts. Cumulative relative frequency curves of daytime sleep bouts from chronically isolated flies climbed faster than those of their group-housed counterparts as shorter sleep bouts accumulated (5 or 7 days). Over seven days of isolation, daily total sleep, daytime sleep and ZT0-4 sleep all decreased progressively (Li, 2021).
Age-matched flies were used to rule out the possibility that chronic social isolation induced sleep loss because the flies were older. Chronic social isolation (7 days) induced sleep loss consistently in various isogenic strains, in aged (4-week-old) wild-type flies, and in sleep inbred panel (SIP) strains with different baseline levels of sleep (Li, 2021).
RNA sequencing (RNA-seq) libraries were prepared for three conditions: socially enriched flies (group treated, Grp), chronically isolated flies (isolated for 7 days, Iso_7D) and acutely isolated flies (isolated for 1 day, Iso_1D). Raster plots demonstrate sleep bouts of individual animals during a 24-h period measured immediately after group enrichment or social isolation. Daytime sleep was reduced and much more fragmented in chronically isolated flies than in group-housed or acutely isolated flies. Fly heads were collected between ZT0.5 and ZT2, a window of time within ZT0-4 that immediately preceded significant loss of daytime sleep in chronically isolated flies. Using differential gene expression analyses, intersectional and clustering strategies, candidate genes were identified for the sleep loss induced by chronic social isolation. These 214 candidate genes showed differences in expression in chronically isolated flies compared with both acutely isolated and group-housed flies and underwent unidirectional changes during the process of chronic social isolation. Gene ontology enrichment analysis suggested that these 214 genes are enriched for biological pathways associated with oxidation-reduction processes, one-carbon metabolic processes and carbohydrate metabolic processes. The rest of the gene ontology of biological pathways showed a strong preference for metabolic functions, such as fatty acid, pyruvate, glucose and amino acid metabolic processes. Consistent with the sleep loss phenotype, sleep was also among the most overrepresented gene ontologies for biological pathways (Li, 2021).
Among the top 20 genes in this list, two genes stood out: Limostatin (Lst, CG8317), expression of which increased 1.67-fold after chronic isolation, and Drosulfakinin (Dsk), expression of which decreased 2.03-fold after chronic isolation. Limostatin is a decretin hormone that is induced by starvation and suppresses insulin release. Drosulfakinin, a satiety-inducing cholecystokinin-like peptide, is secreted when the animal is fed. As a signal of satiety, drosulfakinin is depleted under starvation conditions. A third gene, target of brain insulin (tobi), also showed significantly increased expression (1.76-fold) during chronic social isolation. tobi encodes an α-glucosidase that is regulated by Drosophila insulin and glucagon analogues. In addition, 7 of these top 20 genes and 32 of the total 214 candidate genes were previously identified as being regulated in Drosophila heads after 24 h of starvation. Thus, from a transcriptomic perspective, the brain of a fly maintained in chronic social isolation closely resembles the brain of a starving fly, despite continuous access to food. It was reasoned that such a 'starvation brain state' might broadly affect gene expression associated with metabolic processes. Massive changes in mitochondrial functions and oxidation-reduction processes could be direct consequences of starvation and/or elevated feeding (Li, 2021).
The activity recording capillary feeder (ARC) assay, a video recording capillary feeder (CAFE) assay that monitors sleep and feeding behaviours simultaneously and continuously in individual Drosophila, was used. ARC assays validated the isolation-induced sleep loss phenotype previously observed with DAM assays: daily total sleep, daytime sleep and ZT0-4 sleep were reduced significantly after 7 days of social isolation. In addition, nighttime sleep was also reduced, probably owing to higher sensitivity in detecting movements using the positional tracking method, or differences in chamber shape and food source between the ARC and DAM systems. As predicted from the gene expression profiling results, increased feeding was observed in socially isolated animals compared to their group-treated counterparts. Flies isolated for 7 days showed significant increases in total food consumption, daytime food consumption, nighttime food consumption and ZT0-4 food consumption in comparison to flies that were group-housed for 7 days. Thus, chronic social isolation induces a starvation state in Drosophila at the levels of both gene expression in the brain and behaviour (Li, 2021).
The altered feeding pattern produced by chronic social isolation is not merely a consequence of sleep loss, because several classic sleep mutants all exhibited normal feeding behaviour. In addition, acutely isolated flies did not show a strong increase in food consumption (Li, 2021).
The candidate gene Lst is normally induced by nutrient restriction in endocrine neurons in the corpora cardiaca. However, the RNA profiling experiment suggested that there could be a previously unknown brain source for LST production. A resource of high-resolution transcriptomes of 100 GAL4 driver lines suggested that cells labelled by the driver line NPF-GAL4 (NPF, neuropeptide F [the fly homologue of neuropeptide Y)] are likely to express LST. Using a monoclonal antibody against LST, co-localized LST immunoreactivity and NPF-GAL4-driven GFP signals were detected. Among six known neuronal clusters that express NPF, LST immunoreactivity appeared to be co-localized with NPF-GAL4-driven GFP signals at the dorsal stratum of the fan-shaped body (dorsal fan-shaped body, dFB) and in a cluster of small cell bodies in the dorsal brain. Neurons without known function that comprise this cluster of NPF cells were previously named P2. A recent study used a split-GAL4 driver, SS0020-split-GAL4 (abbreviated as P2-GAL4 below), to strongly label the majority of P2 neurons that showed positive immunoreactivity for LST and NPF22 (Li, 2021).
Notably, the projections of the P2 neurons overlapped with the axonal projections of the dFB neurons labelled by R23E10-GAL4, which suggests that P2 neurons might signal to sleep-promoting dFB neurons. At the cell body level, P2 neurons differ from the R23E10-GAL4 labelled cells. A MultiColor FlpOut (MCFO) approach was used to stochastically decorate individual neurons labelled by P2-GAL4, and it was found that they are fan-shaped body columnar neurons. The hemibrain connectome allowed determination that P2 neurons include, as a dominant constituent, the hDeltaK cell type—a columnar cell class, where each neuron has a stereotypical dendritic input in the ellipsoid body (EB) in addition to the FB innervation. hDeltaK cells exhibit extensive synaptic connections with a known subset of R23E10-GAL4-labelled sleep-promoting dFB neurons27 . On the basis of the above connectome data and existing evidence that NPF/NPY is involved in animal metabolism and stress responses, focus was placed on P2 neurons (Li, 2021).
To test whether P2 neurons contribute to sleep loss induced by chronic social isolation, these neurons were chronically silenced by expressing the inward-rectifying potassium channel Kir2.1 under the control of P2-GAL4. In flies carrying both P2-GAL4 and UAS-Kir2.1, chronic isolation no longer induced an altered sleep profile when compared to their group-housed counterparts. The cumulative relative frequency curve of daytime sleep bouts for socially isolated animals no longer climbed faster than that of group-reared animals. Raster plots of sleep bouts in individual flies showed little difference between chronically isolated and group-housed flies. No difference was found between isolated and group-housed animals for daily total sleep, daytime sleep, or ZT0-4 sleep. By contrast, in heterozygous parental control animals carrying either the P2-GAL4 or the UAS-Kir2.1 transgene, chronic social isolation robustly induced sleep loss. Temporally silencing P2 neurons using UAS-shibirets1 during group enrichment or social isolation did not block social isolation-induced sleep loss. Although isolated flies carrying both P2-GAL4 and UAS-Kir2.1 still showed some overconsumption of food, they no longer showed excessive food consumption for ZT0-4, and the total increase in daytime food consumption was much smaller than in parental controls (Li, 2021).
Using [Ca2+] imaging, it was found that the activity of P2 neurons was correlated with locomotor activity in both group-housed and isolated flies. One might expect that P2 neurons would be tonically more active in isolated flies than in group-housed flies, but this effect in baseline [Ca2+] levels could not be detected. Alternatively, it can be hypothesize that locomotion drives more P2 neuron total activity during 7 days of isolation than during 1 day of isolation (Li, 2021).
Therefore whether boosting activity in P2 neurons during acute social isolation (1 day) is sufficient to promote behavioural changes that resemble the effects of chronic social isolation (7 days) was measured. To activate P2 neurons, a Drosophila warmth-gated cation channel, UAS-dTRPA1, was expressed with P2-GAL4. The P2-GAL4-labelled neurons were activated by treating the flies at 28 °C during acute social isolation or group enrichment (1 day). Control experiments, using flies of the same genotype, were conducted by treating the flies at 22 °C during acute social isolation or group enrichment (1 day). Following these treatments, all flies were subsequently maintained at 22 °C for measurement of sleep or feeding behaviour. In animals carrying both P2-GAL4 and UAS-dTRPA1, there were significant interactions between temperature treatment and isolation status for total, daytime, and ZT0-4 sleep and food consumption: activation of P2 neurons during acute social isolation promoted significant sleep loss and excessive feeding, whereas activation of P2 neurons in group housing did not alter either sleep or feeding behaviour. In control experiments, no evidence was found for interactions between temperature treatment and isolation status in the heterozygous parental flies (Li, 2021).
Notably, P2 neurons are connected to the dFB neurons that are known to regulate sleep homeostasis and couple energy metabolism to sleep. Artificial activation of P2 neurons can produce a behavioural state that resembles the effects of chronic isolation after social isolation for a single day; however, activation of P2 neurons failed to produce these behaviours in the complete absence of social isolation. This indicates that both activity in P2 neurons and a status of being socially isolated are required to induce reduced sleep and increased feeding. Social isolation might be sensed by P2 neurons or elsewhere in the brain, but in either case appears to cause the activity of P2 neurons to be interpreted differently and thereby to generate novel behaviours. Downregulation of a secreted cytokine in a non-neural tissue, the fat body, suppresses sleep and promotes feeding in Drosophila. It would be interesting to determine whether these behavioural responses also depend on P2 neuronal activity (Li, 2021).
Modifications of feeding circuits appear to be crucial for the evolution of complex social behaviours. For example, in C. elegans, a single-residue difference in the neuropeptide Y receptors of naturally occurring strains determines whether the strains exhibit solitary or social feeding behaviour. As antibodies to neuropeptide F, the fly homologue of neuropeptide Y, label P2 neurons, future work may ascertain whether Drosophila's P2 neurons influence social patterns of feeding behaviour as well as mediating feeding and sleep responses to social isolation (Li, 2021).
In humans, social isolation promotes new emotional states that intensify with the passage of time.
Sleep loss in Drosophila is a faithful readout of the duration of social isolation, and this allowed identification of specific patterns of gene and behavioural states that emerge as social isolation becomes chronic. This unexpected association between social isolation, sleep and metabolism in an insect model is reminiscent of the connection observed by social psychologists between loneliness, sleep difficulties and hyperphagia. Such robust findings in Drosophila suggest that studies of animal models might identify conserved brain states, genes, and neural circuits that are associated with social isolation (Li, 2021).
The cellular consequences of sleep loss are poorly characterized. In the pyramidal neurons of mouse frontal cortex this study found that mitochondria and secondary lysosomes occupy a larger proportion of the cytoplasm after chronic sleep restriction compared to sleep, consistent with increased cellular burden due to extended wake. For each morphological parameter the within-animal variance was high, suggesting that the effects of sleep and sleep loss vary greatly among neurons. However, the analysis was based on 4-5 mice/group and a single section/cell. this study applied serial block-face scanning electron microscopy to identify signatures of sleep and sleep loss in the Drosophila brain. Stacks of images were acquired and used to obtain full 3D reconstructions of the cytoplasm and nucleus of 263 Kenyon cells from adult flies collected after a night of sleep (S) or after 11 hours (SD11) or 35 hours (SD35) of sleep deprivation (9 flies/group). Relative to S flies, SD35 flies showed increased density of dark clusters of chromatin and of Golgi apparata and a trend increase in the percent of cell volume occupied by mitochondria, consistent with increased need for energy and protein supply during extended wake. Logistic regression models could assign each neuron to the correct experimental group with good accuracy, but in each cell nuclear and cytoplasmic changes were poorly correlated, and within-fly variance was substantial in all experimental groups. Together, these results support the presence of ultrastructural signatures of sleep and sleep loss but underscore the complexity of their effects at the single-cell level (Flores, 2022).
Sleep is conserved across species, and it is believed that a fixed amount of sleep is needed for normal neurobiological functions. Sleep rebound follows sleep deprivation; however, continuous sleep deprivation for longer durations is believed to be detrimental to the animal's wellbeing. Under some physiologically demanding situations, such as migration in birds, the birth of new offspring in cetaceans, and sexual interactions in pectoral sandpipers, animals are known to forgo sleep. The mechanisms by which animals forgo sleep without having any obvious negative impact on the proper functioning of their neurobiological processes are yet unknown. Therefore, a simple assay is needed to study how animals forgo sleep. The assay should be ecologically relevant so it can offer insights into the physiology of the organisms. Equally important is that the organism should be genetically amenable, which helps in understanding the cellular and molecular processes that govern such behaviors. This paper presents a simple method of sociosexual interaction to understand the process by which animals forgo sleep. In the case of Drosophila melanogaster, when males and females are in proximity, they are highly active and lose a significant amount of sleep. In addition, there is no sleep rebound afterward, and instead, males engaged in sexual interactions continue to show normal sleep. Thus, sexual drive in the fruit flies is a robust assay to understand the underlying mechanism by which animals forgo sleep (Mishra, 2023).
Animals increase their locomotion activity and reduce sleep duration under starved conditions. This suggests that sleep and metabolic status are closely interconnected. The nutrient and hunger sensors in the Drosophila brain, including diuretic hormone 44 (DH44)-, CN-, and cupcake-expressing neurons, detect circulating glucose levels in the internal milieu, regulate the insulin and glucagon secretion and promote food consumption. Food deprivation is known to reduce sleep duration, but a potential role mediated by the nutrient and hunger sensors in regulating sleep and locomotion activity remains unclear. This study shows that DH44 neurons are involved in regulating starvation-induced sleep suppression, but CN neurons or cupcake neurons may not be involved in regulating starvation-induced sleep suppression or baseline sleep patterns. Inactivation of DH44 neurons resulted in normal daily sleep durations and patterns under fed conditions, whereas it ablated sleep reduction under starved conditions. Inactivation of CN neurons or cupcake neurons, which were proposed to be nutrient and hunger sensors in the fly brain, did not affect sleep patterns under both fed and starved conditions. It is proposed that the glucose-sensing DH44 neurons play an important role in mediating starvation-induced sleep reduction (Oh, 2023).
Recent studies have shown that gut microorganisms can modulate host lifespan and activities, including sleep quality and motor performance. However, the role of gut microbial genetic variation in regulating host phenotypes remains unclear. This study investigated the links between gut microbial genetic variation and host phenotypes using Saccharomyces cerevisiae and Drosophila melanogaster as research models. The results suggested a novel role for peroxisome-related genes in yeast in regulating host lifespan and activities by modulating gut oxidative stress. Specifically, it was found that deficiency in catalase A (CTA1) in yeast reduced both the sleep duration and lifespan of fruit flies significantly. Furthermore, this research also expanded understanding of the relationship between sleep and longevity. Using a large sample size and excluding individual genetic background differences, this study found that lifespan is associated with sleep duration, but not sleep fragmentation or motor performance. Overall, this study provides novel insights into the role of gut microbial genetic variation in regulating host phenotypes and offers potential new avenues for improving health and longevity (Li, 2023).
Modern lifestyle is often at odds with endogenously driven rhythmicity, which can lead to circadian disruption and metabolic syndrome. One signature for circadian disruption is a reduced or altered metabolite cycling in the circulating tissue reflecting the current metabolic status. Drosophila is a well-established model in chronobiology, but day-time dependent variations of transport metabolites in the fly circulation are poorly characterized. This study sampled fly hemolymph throughout the day and analysed diacylglycerols (DGs), phosphoethanolamines (PEs) and phosphocholines (PCs) using LC-MS. In wildtype flies kept on sugar-only medium under a light-dark cycle, all transport lipid species showed a synchronized bimodal oscillation pattern with maxima at the beginning and end of the light phase which were impaired in period01 clock mutants. In wildtype flies under constant dark conditions, the oscillation became monophasic with a maximum in the middle of the subjective day. In strong support of clock-driven oscillations, levels of DGs, PEs and PCs peaked once in the middle of the light phase under time-restricted feeding independent of the time of food intake. When wildtype flies were reared on full standard medium, the rhythmic alterations of hemolymph lipid levels were greatly attenuated. These data suggest that the circadian clock aligns daily oscillations of DGs, PEs and PCs in the hemolymph to the anabolic siesta phase, with a strong influence of light on phase and modality. This finding raises the question of whether and to what extent the circadian regulation of transport lipid levels in the hemolymph contributes to the health of the fly (Amatobi, 2023).
Commensal microbes influence various aspects of vertebrate and invertebrate brain function. It has been previously reported that Lactiplantibacillus plantarum SBT2227 promotes sleep in the fruit fly, Drosophila melanogaster. However, how widely the sleep-promoting effects are conserved in gut bacterial species remains unknown. This study orally administered human intestinal and food-associated bacterial species (39 in total) to flies and investigated their effects on sleep. Six species of bacteria were found to have significant sleep-promoting effects. Of these, Bifidobacterium adolescentis, which had the greatest sleep-promoting effect, was further investigated and it was found that the strength of the sleep effect varied among strains of the same bacterial species. The B. adolescentis strains BA2786 and BA003 showed strong and weak effects on sleep, respectively. Transcriptome characteristics compared between the heads of flies treated with BA2786 or BA003 revealed that the gene expression of the insulin-like receptor (InR) was increased in BA2786-fed flies. Furthermore, a heterozygous mutation in InR suppressed the sleep-promoting effect of BA2786. These results suggest that orally administered sleep-promoting bacteria (at least BA2786), may act on insulin signaling to modulate brain function for sleep (Ko, 2023).
Light exposure impacts several aspects of Drosophila development including the establishment of circadian rhythms, neuroendocrine regulation, life-history traits, etc. Introduction of artificial lights in the environment has caused almost all animals to develop ecological and physiological adaptations. White light which comprises different lights of differing wavelengths shortens the lifespan in fruit flies Drosophila melanogaster. The wavelength-specific effects of white light on Drosophila development remains poorly understood. This study shows that different wavelengths of white light differentially modulate Drosophila development in all its concomitant stages when maintained in a 12-h light: 12-h dark photoperiod. It was observed that exposure to different monochromatic lights significantly alters larval behaviours such as feeding rate and phototaxis that influence pre-adult development. Larvae grown under shorter wavelengths of light experienced an altered feeding rate. Similarly, larvae were found to avoid shorter wavelengths of light but were highly attracted to the longer wavelengths of light. Most of the developmental processes were greatly accelerated under the green light regime while in other light regimes, the effects were highly varied. Interestingly, pre-adult survivorship remained unaltered across all light regimes but light exposure was found to show its impact on sex determination. This study for the first time reveals how different wavelengths of white light modulate Drosophila development which in the future might help in developing non-invasive therapies and effective pest measures (Ramakrishnan, 2022).
Alcohol tolerance is a simple form of behavioral and neural plasticity that occurs with the first drink. Neural plasticity in tolerance is likely a substrate for longer term adaptations that can lead to alcohol use disorder. Drosophila develop tolerance with characteristics similar to vertebrates, and it is useful model for determining the molecular and circuit encoding mechanisms in detail. Rapid tolerance, measured after the first alcohol exposure is completely metabolized, is localized to specific brain regions that are not interconnected in an obvious way. A forward neuroanatomical screen was used to identify three new neural sites for rapid tolerance encoding. One of these was comprised of two groups of neurons, the DN1a and DN1p glutamatergic neurons, that are part of the Drosophila circadian clock. Rapid tolerance was localized to the two DN1a neurons that regulate arousal by light at night, temperature-dependent sleep timing, and night-time sleep. Two clock neurons that regulate evening activity, LNd6 and the 5th LNv, are postsynaptic to the DN1as and they promote rapid tolerance via the metabotropic glutamate receptor. Thus, rapid tolerance to alcohol overlaps with sleep regulatory neural circuitry, suggesting a mechanistic link (Lang, 2023).
Sleep is conserved in the animal kingdom and plays a pivotal role in the adaptation of species. Sleep in Drosophila melanogaster is defined as any continuous 5 min of quiescence, shows a prominent siesta, and consolidated nighttime sleep. This study analyzed the sleep of two other species D. malerkotliana (DMK) and D. ananassae (DA), and compared it with D. melanogaster (DM). The DMK males and females have siesta like DM. However, unlike DM, flies continue to sleep beyond siesta till the evening. DA has a less prominent siesta compared to DM and DMK. In the morning, DA took a longer time to respond to the lights ON and continued to sleep for at least half an hour. The nighttime sleep of the DA flies is higher than the other two species. Average length of sleep episode is three times more than that of DM and DMK with few wake episodes. Thus, the nighttime sleep of DA males and females is deep and needs exposure to more potent stimuli to wake up relative to the other two species. DA males and females show higher sleep rebound than the other two species, suggesting the robustness of sleep homeostasis. Although total sleep of DMK and DA is similar, DA is a day-active species with highly consolidated night sleep. DMK, like DM, is a crepuscular species with a midday siesta. Thus, these results suggest that temporal partitioning of sleep, in sympatric species may contribute to temporal segregation (Mishra, 2023).
Genome-wide association studies (GWAS) in humans have identified loci robustly associated with several heritable diseases or traits, yet little is known about the functional roles of the underlying causal variants in regulating sleep duration or quality. This study applied an ATAC-seq/promoter focused Capture C strategy in human iPSC-derived neural progenitors to carry out a "variant-to-gene" mapping campaign that identified 88 candidate sleep effector genes connected to relevant GWAS signals. To functionally validate the role of the implicated effector genes in sleep regulation, a neuron-specific RNA interference screen was performed in the fruit fly, Drosophila melanogaster, followed by validation in zebrafish. This approach identified a number of genes that regulate sleep including a critical role for glycosylphosphatidylinositol (GPI)-anchor biosynthesis. These results provide the first physical variant-to-gene mapping of human sleep genes followed by a model organism-based prioritization, revealing a conserved role for GPI-anchor biosynthesis in sleep regulation (Palermo, 2023).
The mechanisms by which the genotype interacts with nutrition during development to contribute to the variation of complex behaviors and brain morphology of adults are not well understood. This stusy used the Drosophila Genetic Reference Panel to identify genes and pathways underlying these interactions in sleep behavior and mushroom body morphology. Early-life nutritional restriction effects on sleep behavior and brain morphology was shown to depend on the genotype. Genes associated with sleep sensitivity to early-life nutrition were enriched for protein-protein interactions responsible for translation, endocytosis regulation, ubiquitination, lipid metabolism, and neural development. By manipulating the expression of candidate genes in the mushroom bodies and all neurons, it was confirmed that genes regulating neural development, translation and insulin signaling contribute to the variable response of sleep and brain morphology to early-life nutrition. The interaction between differential expression of candidate genes with nutritional restriction in early life resides in the mushroom bodies or other neurons, and these effects are sex specific. Natural variation in genes that control the systemic response to nutrition and brain development and function interact with early-life nutrition in different types of neurons to contribute to the variation of brain morphology and adult sleep behavior (Olivares, 2023).
Sleep in mammals is broadly classified into two different categories: rapid eye movement (REM) sleep and slow wave sleep (SWS), and accordingly REM and SWS are thought to achieve a different set of functions. The fruit fly Drosophila melanogaster is increasingly being used as a model to understand sleep functions, although it remains unclear if the fly brain also engages in different kinds of sleep as well. This study compared two commonly used approaches for studying sleep experimentally in Drosophila : optogenetic activation of sleep-promoting neurons and provision of a sleep-promoting drug, Gaboxadol. These different sleep-induction methods were found to have similar effects on increasing sleep duration, but divergent effects on brain activity. Transcriptomic analysis reveals that drug-induced deep sleep ('quiet' sleep) mostly downregulates metabolism genes, whereas optogenetic 'active' sleep upregulates a wide range of genes relevant to normal waking functions. This suggests that optogenetics and pharmacological induction of sleep in Drosophila promote different features of sleep, which engage different sets of genes to achieve their respective functions (Anthoney, 2023).
Sleep is a fundamental physiological function and is essential for all animals. Sleep is affected by diet compositions including protein (P) and carbohydrates (C), but there has not been a systematic investigation on the effect of dietary macronutrient balance on sleep. This study used the nutritional geometry framework (NGF) to explore the interactive effects on sleep of protein (P) and carbohydrates (C) in the model organism Drosophila. Both female and male flies were fed various diets containing seven ratios of protein-to-carbohydrates at different energetic levels for 5 days and sleep was monitored by the Drosophila Activity Monitor (DAM) system. The results showed that the combination of low protein and high carbohydrates (LPHC) prolonged sleep time and sleep quality, with fewer sleep episodes and longer sleep duration. It was further found that the effects of macronutrients on sleep mirrored levels of hemolymph glucose and whole-body glycogen. Moreover, transcriptomic analyses revealed that a high-protein, low-carbohydrate (HPLC) diet significantly elevated the gene expression of metabolic pathways when compared to the LPHC diet, with the glycine, serine, and threonine metabolism pathway being most strongly elevated. Further studies confirmed that the contents of glycine, serine, and threonine affected sleep. These results demonstrate that sleep is affected by the dietary balance of protein and carbohydrates possibly mediated by the change in glucose, glycogen, glycine, serine, and threonine (Lai, 2023).
Sleep is vital for most animals, yet its mechanism and funwction remain unclear. This study found that permeability of the BBB (blood-brain barrier)-the organ required for the maintenance of homeostatic levels of nutrients, ions, and other molecules in the brain-is modulated by sleep deprivation (SD) and can cell-autonomously effect sleep changes. Increased BBB permeability was observed in known sleep mutants as well as in acutely sleep-deprived animals. In addition to molecular tracers, SD-induced BBB changes also increased the penetration of drugs used in the treatment of brain pathologies. After chronic/genetic or acute SD, rebound sleep or administration of the sleeping aid gaboxadol normalized BBB permeability, showing that SD effects on the BBB are reversible. Along with BBB permeability, RNA levels of the BBB master regulator moody are modulated by sleep. Conversely, altering BBB permeability alone through glia-specific modulation of moody, g&slpha;o, loco, lachesin, or i>neuroglian -each a well-studied regulator of BBB function-was sufficient to induce robust sleep phenotypes. These studies demonstrate a tight link between BBB permeability and sleep and indicate a unique role for the BBB in the regulation of sleep (Axelrod, 2023).
This study establish the BBB as a sleep regulatory center in Drosophila. It was observed that the BBB dynamically and rapidly—both functionally and molecularly—responds to acute as well as chronic-genetic SD and that altering BBB function alone, through mutations of the moody signaling pathway, can affect sleep, suggesting that the BBB senses sleep need and responds to it by altering its permeability. The correct BBB permeability state seems to be required for normal sleep, as disrupting BBB function causes sleep loss (Axelrod, 2023).
In normal physiology, the BBB controls influx and efflux of molecules in and out of the brain, including ions, amino acids, neurotransmitters, vitamins, proteins, carbohydrates, lipids, and hormones. Transport is facilitated via various mechanisms which include active transporters, transcytosis, and passive diffusion. The function of the BBB is not static but is affected by circadian rhythms, neuronal activity, and the metabolism. the data add sleep as an additional parameter modulating BBB function, as has been suggested by studies in mammals, and this study found that the relationship between the BBB and sleep is reciprocal. The results show that the BBB rapidly and dynamically responds to SD by increasing its permeability and that permeability increases conversely cause sleep loss. It is conceivable that BBB permeability changes allow the passage of one or multiple molecules that affect sleep into or out of the brain. Defective xenobiotic exclusion of steroid hormones from the brain via the BBB has been shown to affect sleep in Drosophila, demonstrating that molecules outside the CNS can affect sleep. Future studies may determine whether the BBB mediates a rapid global switch from a sleeping to an awake brain, for example, by controlling ionic diffusion and potassium concentrations, which could broadly raise neuronal activation thresholds and lead to the observed extracellular ionic changes in the sleeping mammalian brain (Axelrod, 2023).
The BBB creates a highly controlled microenvironment in the brain, perturbations of which affect neuronal function, as evidenced by neuropathologies at least in part caused by BBB disruption including stroke, Alzheimer's, Multiple Sclerosis, epilepsy and brain tumor metastasis. BBB disruption also occurs during normal aging, under chronic stress and during systemic inflammation and infection; however, whether the relationship between BBB disruption and these pathologies is causal or merely correlative is less clear. The magnitude of BBB changes in disease states can be an order of magnitude larger than the observed changes after sleep loss described in this study, illustrating the physiological relevance of these experiments. All of the pathologies listed above are also marked by sleep disruption, which in some instances increases the risk for developing the disease in the first place and the severity of which correlates with disease severity. The fly ortholog of a human autism gene has been reported to act in the BBB and elicit sleep fragmentation via developmental hyperserotonemia. However, a direct involvement of the BBB in sleep regulation had not been shown until this present study (Axelrod, 2023).
BBB and sleep disruptions have been, so far independently, associated with causing illness, and this study provides additional evidence for the notion that the two processes are linked. If BBB permeability and sleep regulation are fundamentally connected as part of the same system, it will be interesting to study how they relate to disease risk, development, and prognosis, and whether interventions that affect one also augment the other (Axelrod, 2023).
Adequate drug delivery to the brain is a problem plaguing the treatment of many neuropathologies. This puts strong importance on finding possible regulatory sites for controlling the BBB as a means of finding better ways to deliver drugs to the brain. The BBB has distinct mechanisms to exclude molecules from the brain including xenobiotic transporters in the cell membrane that efflux most small hydrophobic molecules taken up transcellularly through the membrane, and the paracellular barrier formed by junctions between the BBB forming cells. The latter creates a diffusion barrier for hydrophilic molecules in a size-dependent manner with a maximum passage size of about 0.5 kDa. Due to these mechanisms, the BBB poses a major obstacle to treatment of brain pathologies, because 100% of large molecules (>0.5 kDa), like the brain cancer drug vinblastine (molecular weight: 1 kDa), and 98% of small molecule drugs (<0.5 kDa), like the non-steroidal anti-inflammatory drug paracetamol, cannot penetrate it in therapeutic amounts. Current solutions to this problem include pharmacologically increasing the diffusion barrier permeability, implantation of ultrasonic devices to temporarily permeabilize the diffusion barrier, fusing molecules to receptor ligands to engage the transcellular route, or intracranial administration via injection. Provided the observed effects are conserved in humans, a patient's sleep and nighttime administration of drugs could have a significant effect on drug penetration into the CNS and possibly treatment outcomes (Axelrod, 2023).
In Drosophila, the BBB's subperineurial glia form a layer of polarized epithelial cells, with septate junctions between them creating a seal for water-soluble molecules over 500 Da. The observation that SD rapidly induces increases in BBB permeability raises the question: What are the molecular and cellular mechanisms that underlie the remodeling of septate junctions on the scale of hours? The Moody GPCR pathway has been shown to play a key role in the development of the epithelial barrier, as well as its maintenance in adulthood. Moody activates a signaling cascade which, via PKA activation, controls the developmental assembly of the septate junction belt in a highly coordinated spatiotemporal manner that involves rearrangements of the actin cytoskeleton and vesicular trafficking. This study has shown that RNAi-based suppression of moody pathway genes causes increased BBB permeability. Strong downregulation of moody was observed in every genetic sleep mutant tested. These findings suggest that altered moody pathway function may be primarily responsible for the leaky barriers of these mutants (Axelrod, 2023).
During SD and recovery, it is conceivable that similar processes occur. Indeed, it has been shown that perturbation of endocytosis at the Drosophila BBB alters sleep, possibly interfering with Moody-activated remodeling during sleep/wake cycles. Scute SD has been shown to rapidly depress moody expression in wild-type Drosophila, which is accompanied by increased BBB permeability. Moody is expressed on the neuronal-facing apical cell surface of the subperineurial glia. While its ligand is not known, this topology could permit neuronal states related to sleep to be communicated to glia via this GPCR pathway. As many known sleep genes in Drosophila have been shown to be expressed in neurons and this study observed BBB changes upon sleep-suppressing neuronal activation, neuron-glia signaling seems likely to be part of the process, and it will be interesting to elucidate the involved signaling molecules including Moody's ligand (Axelrod, 2023).
The two-step model of sleep regulation posits that circadian rhythm and sleep homeostasis control the timing of sleep. Sleep homeostasis is a process that controls sleep duration to meet an organism's daily sleep need. Rebound sleep after SD is interpreted as proof of the existence of sleep homeostasis. The physiological and cellular nature of the sleep homeostat is-like the nature of sleep itself-the subject of current research. Sleep homeostasis has been shown to be affected by oxidative stress, neuronal, glial, and microglial activity, neuropeptide signaling, infection, the circadian clock, temperature, and starvation. Synaptic downscaling, glymphatic clearance of toxins, peripheral glucose homeostasis, reactive oxygen species clearance, and inhibition of motor function have been proposed as processes occurring during sleep homeostasis, constituting functions of sleep. As at least some of these processes must involve yet to be identified body-brain signals, future studies will reveal how the BBB changes shown here affect known or novel functions of sleep (Axelrod, 2023).
Together the data indicate a system where an increase of sleep need is measured by the BBB, resulting in an adjustment of Moody levels, an opening of the BBB, and the initiation of rebound sleep. This process is likely to involve neuronal to BBB signaling (dopaminergic SD also alters the BBB, and many of the sleep mutants are expressed in neurons) as well as yet-to-be-characterized molecular exchange across the BBB (Axelrod, 2023).
The master circadian clock generates 24-hour rhythms to orchestrate daily behavior, even running freely under constant conditions. Traditionally, the master clock is considered self-sufficient in sustaining free-running timekeeping via its cell-autonomous molecular clocks and interneuronal communications within the circadian neural network. This study found a set of bona fide ultradian oscillators in the Drosophila brain that support free-running timekeeping, despite being located outside the master clock circuit and lacking clock gene expression. These extra-clock electrical oscillators (xCEOs) generate cell-autonomous ultradian bursts, pacing widespread burst firing and promoting rhythmic resting membrane potentials in clock neurons via parallel monosynaptic connections. Silencing xCEOs disrupts daily electrical rhythms in clock neurons and impairs cycling of neuropeptide pigment dispersing factor, leading to the loss of free-running locomotor rhythms. Together, it is concluded that the master clock is not self-sufficient to sustain free-running behavior rhythms but requires additional endogenous inputs to the clock from the extra-clock ultradian brain oscillators (Tang, 2022).
Circadian clocks are highly conserved transcriptional regulators that control ~24 hr oscillations in gene expression, physiological function, and behavior. Circadian clocks exist in almost every tissue and are thought to control tissue-specific gene expression and function, synchronized by the brain clock. Many disease states are associated with loss of circadian regulation. How and when circadian clocks fail during pathogenesis remains largely unknown because it is currently difficult to monitor tissue-specific clock function in intact organisms. This study developed a method to directly measure the transcriptional oscillation of distinct neuronal and peripheral clocks in live, intact Drosophila; the method was termed Locally Activatable BioLuminescence, or LABL. Using this method, it was observed that specific neuronal and peripheral clocks exhibit distinct transcriptional properties. Loss of the receptor for PDF, a circadian neurotransmitter critical for the function of the brain clock, disrupts circadian locomotor activity but not all tissue-specific circadian clocks. While peripheral clocks in non-neuronal tissues were less stable after the loss of PDF signaling, they continued to oscillate. It was also demonstrated that distinct clocks exhibit differences in their loss of oscillatory amplitude or their change in period, depending on their anatomical location, mutation, or fly age. These results demonstrate that LABL is an effective tool that allows rapid, affordable, and direct real-time monitoring of individual clocks in vivo (Johnstone, 2022).
Photic history, including the relative duration of day versus night in a 24-hour cycle, is known to influence subsequent circadian responses to light in mammals. Whether such modulation is present in Drosophila is currently unknown. This study constructed the first high-resolution Drosophila seasonal atlas for light-induced circadian phase-resetting. Testing the light responses of over 4,000 Drosophila at 120 timepoints across 5 seasonally-relevant rectangular photoperiods (i.e., LD 8:16, 10:14, 12:12, 14:10, and 16:8; 24 hourly intervals surveyed in each), it was determined that many aspects of the fly circadian PRC waveform are conserved with increasing daylength. Surprisingly though, irrespective of LD schedule, the start of the PRCs always remained anchored to the timing of subjective sunset, creating a differential overlap of the advance zone with the morning hours after subjective sunrise that was maximized under summer photoperiods and minimized under winter photoperiods. These data suggest that there may be differences in flies versus mammals as to how the photoperiod modulates the waveform and amplitude of the circadian PRC to light. On the other hand, they support the possibility that the lights-off transition determines the phase-positioning of photic PRCs across seasons and across species. More work is necessary to test this claim and whether it might factor into the timing of seasonal light responses in humans (Dollish, 2022).
Rapid technology development, exposure to gadgets, and artificial lights (with different monochromes) have disturbed lifestyles and the circadian clock, which otherwise confers better regulation of behavioral patterns and sleep/wake cycles in most organisms including Drosophila melanogaster. This study assayed the effect of LD12:12 hr (light: dark) monochromatic lights (violet, blue, green, yellow, orange, and red) on the lifespan, activity, and sleep of the D. melanogaster. A shortened lifespan was observed under 12h of violet, blue, green, and yellow lights, while significantly reduced activity levels under the light phase of blue and green light as compared to their dark phase is observed. Significant increase in the evening anticipation index of flies under blue and green light alongside increased and decreased sleep depth during the day and night respectively suggests the light avoidance, while there is no effect of colored light on the waking time, daily active time, and sleep time. Thus, this study shows short and long-term exposure to certain colored lights in terms of reduced lifespan and locomotor activity, which cause qualitative as well as quantitative changes in the sleep of flies; probably as a sign of aversion towards a specific light (Krittika, 2022).
The role of the circadian system in memory formation is an important question in neurobiology. Despite this hypothesis being intuitively appealing, the existing data is confusing. Recent work in Drosophila has helped to clarify certain aspects of the problem, but the emerging sense is that the likely mechanisms are more complex than originally conceptualized. This report, identifies a post-training window of time (during consolidation) when the circadian clock and its components are involved in memory formation. In the broader context, the data suggest that circadian biology might have multiple roles during memory formation. Testing for its roles at multiple timepoints, and in different cells, will be necessary to resolve some of the conflicting data (Yin, 2023).
Resveratrol (RES), a natural polyphenolic phytochemical, has been suggested as a putative anti-aging molecule for the prevention and treatment of Alzheimer's disease (AD) by the activation of sirtuin 1 (Sirt1/Sir2). This study, tested the effects of RES and Sirt1/Sir2 on sleep and courtship memory in a Drosophila model by overexpression of amyloid precursor protein (APP), whose duplications and mutations cause familial AD. A mild but significant transcriptional increase of Drosophila Sir2 (dSir2) was found by RES supplementation for up to 17 days in APP flies, but not for 7 days. RES and dSir2 almost completely reversed the sleep and memory deficits in APP flies. It was further demonstrated that dSir2 acts as a sleep promotor in Drosophila neurons. Interestingly, RES increased sleep in the absence of dSir2 in dSir2-null mutants, and RES further enhanced sleep when dSir2 was either overexpressed or knocked down in APP flies. Finally, this study showed that A&betal aggregates in APP flies were reduced by RES and dSir2, probably via inhibiting Drosophila β-secretase (dBACE). These data suggest that RES rescues the APP-induced behavioral deficits and Aβ burden largely, but not exclusively, via dSir2 (Hao, 2023).
During sleep, behavioral responsiveness to external stimuli is decreased. This classical definition of sleep has been applied effectively across the animal kingdom to identify this common behavioral state in a growing list of creatures, from mammals to invertebrates. Yet, it remains unclear whether decreased behavioral responsiveness during sleep is necessarily associated with decreased responsiveness in brain activity, especially in insects. Long-term multichannel electrophysiology was performed in tethered Drosophila melanogaster flies exposed continuously to repetitive visual stimuli. Flies were still able to sleep under these visual stimulation conditions, as determined by traditional immobility duration criteria for the field. Interestingly, this study did not find any difference between responses to repetitive visual stimuli during sleep compared with wake when local field potentials (LFPs) across a transect of the fly brain from optic lobes to the central brain was recorded. However, LFP responses were found to be altered when visual stimuli were variable and of lower probability, especially in the central brain. Central brain responses to less predictable or 'deviant' stimuli were lower during the deepest stage of sleep, a time of quiescence characterized by more regular proboscis extensions. This shows that the sleeping fly brain processes low-probability visual stimuli in a different way from more repeated stimuli, and presents Drosophila as a promising model for studying the potential role of sleep in regulating predictive processing (Van De Poll, 2025).
Circadian rhythms in physiology and behaviour have near 24 h periodicities that must adjust to the exact 24 h geophysical cycles on earth to ensure adaptive daily timing. Such adjustment is called entrainment. One major mode of entrainment is via the continuous modulation of circadian period by the prolonged presence of light. Although Drosophila melanogaster is a prominent insect model of chronobiology, there is little evidence for such continuous effects of light in the species. This study demonstrates that prolonged light exposure at specific times of the day shapes the daily timing of activity in flies. It was also established that continuous UV- and blue-blocked light lengthens the circadian period of Drosophila, and evidence is provided that this is produced by the combined action of multiple photoreceptors which, includes the cell-autonomous photoreceptor Cryptochrome. Finally, ramped light cycles were introducted as an entrainment paradigm that produces light entrainment that lacks the large light-driven startle responses typically displayed by flies and requires multiple days for entrainment to shifted cycles. These features are reminiscent of entrainment in mammalian models systems and make possible new experimental approaches to understanding the mechanisms underlying entrainment in the fly (Abhilash, 2023).
Animal behavior spans many timescales, from short, seconds-scale actions to circadian rhythms over many hours to life-long changes during aging. Most quantitative behavior studies have focused on short-timescale behaviors such as locomotion and grooming. Analysis of these data suggests there exists a hierarchy of timescales; however, the limited duration of these experiments prevents the investigation of the full temporal structure. To access longer timescales of behavior, individual Drosophila melanogaster were continuously recorded at 100 frames per second for up to 7 days at a time in featureless arenas on sucrose-agarose media. The deep learning framework SLEAP was used to produce a full-body postural data set for 47 individuals resulting in nearly 2 billion pose instances. Stereotyped behaviors were identified such as grooming, proboscis extension, and locomotion, and the resulting ethograms were used to explore how the flies' behavior varies across time of day and days in the experiment. Distinct circadian patterns were observed all of the stereotyped behavior, and also changes were seen in behavior over the course of the experiment as the flies weaken and die (McKenzie-Smith, 2023).
Severe sleep deprivation (SD) has been highly associated with systemic energy wasting, such as lipid loss and glycogen depletion. Despite immune dysregulation and neurotoxicity observed in SD animals, whether and how the gut-secreted hormones participate in SD-induced disruption of energy homeostasis remains largely unknown. Using Drosophila as a conserved model organism, this study characterize that production of intestinal Allatostatin A (AstA), a major gut-peptide hormone, is robustly increased in adult flies bearing severe SD. Interestingly, the removal of AstA production in the gut using specific drivers significantly improves lipid loss and glycogen depletion in SD flies without affecting sleep homeostasis. The molecular mechanisms were revealed whereby gut AstA promotes the release of an adipokinetic hormone (Akh), an insulin counter-regulatory hormone functionally equivalent to mammalian glucagon, to mobilize systemic energy reserves by remotely targeting its receptor AstA-R2 in Akh-producing cells. Similar regulation of glucagon secretion and energy wasting by AstA/galanin is also observed in SD mice. Further, integrating single-cell RNA sequencing and genetic validation, this study uncovered that severe SD results in ROS accumulation in the gut to augment AstA production via TrpA1. Altogether, these results demonstrate the essential roles of the gut-peptide hormone AstA in mediating SD-associated energy wasting (Li, 2023).
The Unified Theory suggests that sleep is a process that developed in eukaryotic animals from a relationship with an endosymbiotic bacterium. Over evolutionary time the bacterium evolved into the modern mitochondrion that continues to exert an effect on sleep patterns, e.g. the bacterium Wolbachia establishes an endosymbiotic relationship with Drosophila and many other species of insects and is able to change the host's behaviour by making it sleep. The hypothesis is supported by other host-parasite relationships, e.g., Trypanosoma brucei which causes day-time sleepiness and night-time insomnia in humans and cattle. For eukaryotes such as Monocercomonoids that don't contain mitochondria, no evidence was found of them sleeping. Mitochondria produce the neurotransmitter gamma aminobutyric acid (GABA), and ornithine a precursor of the neurotransmitter GABA, together with substances such as 3,4dihydroxy phenylalanine (DOPA) a precursor for the neurotransmitter dopamine: These substances have been shown to affect the sleep/wake cycles in animals such as Drosophilia and Hydra. Eukaryote animals have traded the very positive side of having mitochondria providing aerobic respiration for them with the negative side of having to sleep. NREM (Quiet sleep) is the process endosymbionts have imposed upon their host eukaryotes and REM (Active sleep) is the push-back adaptation of eukaryotes with brains, returning to wakefulness (Adams, 2023).
Prolonged cellular activity may overload cell function, leading to high rates of protein synthesis and accumulation of misfolded or unassembled proteins, which cause endoplasmic reticulum (ER) stress and activate the unfolded protein response (UPR) to re-establish normal protein homeostasis. Previous molecular work has demonstrated that sleep deprivation (SD) leads to ER stress in neurons, with a number of ER-specific proteins being upregulated to maintain optimal cellular proteostasis. This study investigated the transcriptional and ultrastructural ER and mitochondrial modifications induced by sleep loss. Gene expression analysis in mouse forebrains was used to show that SD was associated with significant transcriptional modifications of genes involved in ER stress but also in ER-mitochondria interaction, calcium homeostasis, and mitochondrial respiratory activity. Using electron microscopy, it was also shown that SD was associated with a general increase in the density of ER cisternae in pyramidal neurons of the motor cortex. Moreover, ER cisternae established new contact sites with mitochondria, the so-called mitochondria associated membranes (MAMs), important hubs for molecule shuttling, such as calcium and lipids, and for the modulation of ATP production and redox state. Finally, it was demonstrated that Drosophila male mutant flies (elav > linker), in which the number of MAMs had been genetically increased, showed a reduction in the amount and consolidation of sleep without alterations in the homeostatic sleep response to SD. This study has provided evidence that sleep loss induces ER stress characterized by increased crosstalk between ER and mitochondria. MAMs formation associated with SD could represent a key phenomenon for the modulation of multiple cellular processes that ensure appropriate responses to increased cell metabolism. In addition, MAMs establishment may play a role in the regulation of sleep under baseline conditions (El Alaoui, 2023).
After mating, the physiology of Drosophila females undergo several important changes, some of which are reflected in their rest-activity cycles. To explore the hypothesis that mating modifies the temporal organization of locomotor activity patterns, this study recorded fly activity by a video tracking method. Monitoring rest-activity patterns under light/dark (LD) cycles indicated that mated females lose their ability to anticipate the night-day transition, in stark contrast to males and virgins. This postmating response is mediated by the activation of the sex peptide receptor (SPR) mainly on pickpocket (ppk) expressing neurons, since reducing expression of this receptor in these neurons restores the ability to anticipate the LD transition in mated females. Furthermore, evidence is provided of connectivity between ppk+ neurons and the pigment-dispersing factor (PDF)-positive ventral lateral neurons (sLNv), which play a central role in the temporal organization of daily activity. Since PDF has been associated to the generation of the morning activity peak, it is hypothesized that the mating signal could modulate PDF levels. Indeed, it was confirmed that mated females have reduced PDF levels at the dorsal protocerebrum; moreover, SPR downregulation in ppk+ neurons mimics PDF levels observed in males. In sum, these results are consistent with a model whereby mating-triggered signals reach clock neurons in the fly central nervous system to modulate the temporal organization of circadian behavior according to the needs of the new status (Riva, 2022).
Virtually every single living organism on Earth shows a circadian (i.e. "approximately a day") internal rhythm that is coordinated with planet rotation (i.e. 24 hours). External cues synchronize the central clock of the organism. Consequences of biological rhythm disruptions have been extensively studied on cancer. Still, mechanisms underlying these alterations, and how they favor tumor development remain largely unknown. This study shows that glioblastoma-induced neurodegeneration also causes circadian alterations in Drosophila. Preventing neurodegeneration in all neurons by genetic means reestablishes normal biological rhythms. Interestingly, in early stages of tumor development, the central pacemaker lengthens its period, whereas in later stages this is severely disrupted. The re-adjustment of the external light:dark period to longer glioblastoma-induced internal rhythms delays glioblastoma progression and ameliorates associated deleterious effects, even after the tumor onset (Jarabo, 2022).
Previous investigations in humans and rodent animal models have assessed the interplay of sleep in the circadian system's phase responses to nighttime light exposure. The resulting data have been mixed, but generally support a modulatory role for sleep in circadian photic resetting (not an absolute requirement). Drosophila have been historically used to provide important insights in the sleep and circadian sciences. However, no experiments to date have evaluated how immediate sleep need or recent sleep history affects their pacemaker's phase readjustments to light. We did so in the current study by (1) forcing separate groups of animals to stay awake for 1 or 4 h after they were shown a broadspectrum pulse (15 min during the first half of the night, 950 lux), or (2) placing them on a restricted sleep schedule for a week before light presentation without any subsequent sleep disruption. Forced sleep restriction, whether acute or chronic, did not alter the size of light-induced phase shifts. These data are consistent with observations made in other diurnal animals and raise the possibility, more broadly, that phototherapies applied during sleep-such as may be necessary during the winter months-may still be efficacious in individuals experiencing sleep-continuity problems such as insomnia (Negelspach, 2022).
It is well established that pheromones are used by insects to transmit information between individuals. However, research has revealed that individual insects can be both the sender and the receiver of some pheromonal signals. It is therefore interesting to consider whether the pheromonal state of an individual insect can exert an effect on itself. This study monitored the sleep activity of single flies exhibiting a mutation that leads to pheromonal deficiency and found that cuticular hydrocarbons (CHs) exerted self-regulatory effects on the amount of sleep experienced by these flies. To identify the physiological significance of this mechanism, the amounts of sleep were compared in individual young flies and individual old flies (flies are known to sleep less as they get older), and this data was compared with young and old flies exhibiting mutations that lead to CH reception defects. The differences in the amount of sleep experienced by young and old mutant flies were significantly lower than those of the control flies. These data show that hydrocarbon signals produced by the cuticle in Drosophila can be self-perceived and regulate the amount of sleep acquired in a maturation-dependent manner (Wu, 2022).
People tend to fall asleep when gently rocked or vibrated. Experimental studies have shown that rocking promotes sleep in humans and mice. However, the mechanisms underlying the phenomenon are not well understood. A habituation model proposes that habituation, a form of non-associative learning, mediates sleep induction by monotonous stimulation. This study showed that gentle vibration promotes sleep in Drosophila in part through habituation. Vibration-induced sleep (VIS) leads to increased homeostatic sleep credit and reduced arousability, and can be suppressed by heightened arousal or reduced GABA signaling. Multiple mechanosensory organs mediate VIS, and the magnitude of VIS depends on vibration frequency and genetic background. Sleep induction improves over successive blocks of vibration. Furthermore, training with continuous vibration does not generalize to intermittent vibration, demonstrating stimulus specificity, a characteristic of habituation. These findings suggest that habituation plays a significant role in sleep induction by vibration (Ozturk-Colak, 2020).
The importance of sleep in maintaining cognitive functions such as learning and memory has been reported in both vertebrates and invertebrates. Previous studies demonstrated that sleep deprivation impaired the olfactory memory retention of fruit flies as described in the classical conditioning paradigm. This study shows that sleep deprivation leads to a preference for the odours of the rearing environment in Drosophila melanogaster. Flies whose sleep had been disturbed with periodic rotation stimuli during night-time preferred apple cider vinegar (ACV) to broth, while this preference was lower in flies without sleep deprivation and those rotated during daytime. Experiments using single odours showed an increase in responses to ACV due to sleep deprivation. These results suggest that sleep functions in food odour preference. Flies grown on medium supplemented with ACV showed greater preference for ACV, and those grown with broth supplementation showed a greater preference for broth under sleep-deprived conditions. These results suggest that flies with night-time sleep deprivation become attached to the environment on which they have developed, and that sleep contributes to preference for novel food odours. This study offers an approach to investigating the interaction between sleep and neural disorders concerning cognitive deficits towards novel stimuli (Tanizawa, 20021).
Photoperiod is one of the most reliable seasonal cues that organisms can use to prepare for upcoming environmental changes. Evidence suggests that exposure to different photoperiod can activate plastic responses in stress resistance traits, while there is limited evidence on the plastic response induced by daily progressive cumulative changes in photoperiod. This study assayed the effect of within generation daily uni-directional and cumulative changes in photoperiod on stress resistance and life history traits in four Drosophila species. It was predicted that daily increasing photoperiod, mimicking upcoming summer conditions, should lead to an increase in heat resistance and establish trade-offs with other fitness related traits. On the other hand, it was predicted that daily decreasing photoperiod should reflect upcoming winter conditions leading to an increase in cold resistance. It was found that within genreation changes in photoperiod had a significant effect on life history and stress resistance traits in the four Drosophila species. The observed response was different across species, with D. melanogaster showing five out of six studied traits affected, while in D. mercatorum only one trait was significantly affected. The exposure to changing photoperiod led to an increased upper thermal resistance in D. melanogaster and D. mercatorum and a decreased lower thermal resistance in D. melanogaster and D. simulans, as well as a decreased starvation and desiccation resistance in D. virilis. The developmental time was shorter when flies were exposed to the two photoperiod regimes compared to constant day length control in D. melanogaster and D. simulans. A limited effect was observed on egg-to-adult-viability and desiccation resistance. The results of this study show that daily change in photoperiod induced a plastic response in different traits of drosophilids, suggesting that this environmental parameter needs to be carefully considered in evolutionary studies (Manenti, 2021).
With the rapid development of science and technology, 5G technology will be widely used, and biosafety concerns about the effects of 5G radiofrequency radiation on health have been raised. Drosophila melanogaster was selected as the model organism for this study, in which a 3.5 GHz radiofrequency radiation (RF-EMR) environment was simulated at intensities of 0.1 W/m(2), 1 W/m(2), and 10 W/m(2). The activity of parent male and offspring (F1) male flies was measured using a Drosophila activity monitoring system under short-term and long-term 3.5 GHz RF-EMR exposure. Core genes associated with heat stress, the circadian clock and neurotransmitters were detected by QRT-PCR technology, and the contents of GABA and glutamate were detected by UPLC-MS. The results show that short-term RF-EMR exposure increased the activity level and reduced the sleep duration while long-term RF-EMR exposure reduced the activity level and increased the sleep duration of F1 male flies. Under long-term RF-EMR, the expression of heat stress response-related hsp22, hsp26 and hsp70 genes was increased, the expression of circadian clock-related per, cyc, clk, cry, and tim genes was altered, the content of GABA and glutamate was reduced, and the expression levels of synthesis, transport and receptor genes were altered. In conclusion, long-term RF-EMR exposure enhances the heat stress response of offspring flies and then affects the expression of circadian clock and neurotransmitter genes, which leads to decreased activity, prolonged sleep duration, and improved sleep quality (Wang, 2021).
Sleep is plastic and is influenced by ecological factors and environmental changes. The mechanisms underlying sleep plasticity are not well understood. This study shows that manipulations that impair flight in Drosophila increase sleep as a form of sleep plasticity. Flight was disrupted by blocking the wing-expansion program, genetically disrupting flight, and by mechanical wing perturbations. A new sleep regulatory circuit is defined starting with specific wing sensory neurons, their target projection neurons in the ventral nerve cord, and the neurons they connect to in the central brain. In addition, a critical neuropeptide (Burs) and its receptor (Rickets) were identified that link wing expansion and sleep. Disrupting flight activates these sleep-promoting projection neurons, as indicated by increased cytosolic calcium levels, and stably increases the number of synapses in their axonal projections. These data reveal an unexpected role for flight in regulating sleep and provide new insight into how sensory processing controls sleep need (Melnattur, 2020).
Circadian rhythms regulate physiology and behavior through
transcriptional feedback loops of clock genes running within specific
pacemaker cells. In Drosophila, molecular oscillations in the small
ventral lateral neurons (sLNvs) command rhythmic behavior under
free-running conditions releasing the neuropeptide Pigment Dispersing
Factor (PDF) in a circadian fashion. Electrical activity in the sLNvs
is also required for behavioral rhythmicity. Yet, how temporal
information is transduced into behavior remains unclear. This study
developed a new tool for temporal control of gene expression to obtain
adult-restricted electrical silencing of the PDF circuit, which led to
reversible behavioral arrhythmicity. Remarkably, Period (Per)
oscillations during the silenced phase remained unaltered, indicating
that arrhythmicity is a direct consequence of the silenced activity.
Accordingly, circadian axonal remodeling and PDF accumulation were
severely affected during the silenced phase. It is concluded that
although electrical activity of the sLNvs is not a clock component, it
coordinates circuit outputs leading to rhythmic behavior
(Depetris-Chauvin, 2011). Work from many laboratories has shaped the current view of the
molecular clockworks. Although the relative contribution of specific
molecular mechanisms is still a matter of debate, it is clear that a
transcriptional and translational negative feedback loop is key to
give rise to and sustain molecular oscillations. Years ago it was
proposed that circadian oscillations arise from interactions between
ion transport systems across the cell membrane and the resulting ion
concentration gradients. In fact, in support of such possibility,
electrical silencing of a key pacemaker circuit in Drosophila stopped
the free-running clock both in the larval and adult brains, leading to
the proposition that active ionic conductances are an essential
component of this cellular mechanism. One potential caveat of those
experiments is that they rely on the long-term expression of ion
channels from early circuit development, which could not only trigger
compensatory mechanisms to avoid net changes in excitability but also
trigger cell death (Depetris-Chauvin, 2011). To more precisely examine the connection between the membrane and the
molecular clock, expression of an inward rectifier K+ channel (KIR)
was restricted to adult stages. Such genetic manipulation rendered the
flies as behaviorally arrhythmic as those expressing the channel from
early circuit development and prevented action potential firing to a
similar extent. Interestingly, however, no effects were observed in
the pace of the molecular oscillations after several days under
free-running conditions (i.e., on DD4, and even in DD9), which, along
with the reversibility observed once kir 2.1 expression was turned off
in several affected outputs (free-running locomotor behavior, PDF
immunoreactivity), strongly support the notion of an unaltered
molecular clock during the silenced phase. In favor of an alternative
interpretation of the original observations, a rundown in the
molecular oscillations - and even no oscillations whatsoever - was
noticed after prolonged KIR expression, opening the possibility that
long-term changes on intrinsic properties of the neurons, likely
through the alteration of second messenger cascades, as it has been
shown in a different but also extreme condition, ultimately impinge
upon cell viability and thus indirectly result in abnormal clock
function. In fact, adult-restricted silencing of the PDF circuit
triggered morphological changes in second order processes, giving rise
to a less complex arborization pattern; it follows that a more severe
treatment, such as long-term KIR expression, could result in stronger
structural phenotypes indicative of defective cell physiology
(Depetris-Chauvin, 2011). In addition, constantly low PDF levels could potentially account for
the progressive run-down in molecular oscillations. Along this line it
has been shown that, in the absence of PDF, the sLNvs eventually
desynchronize, becoming evident by DD6. Because acute electrical
silencing of PDF neurons clamps the neuropeptide to trough levels that
are insufficient to sustain synchronicity in dorsal oscillators,
affecting excitability for longer terms could eventually result in
reduced amplitude oscillations and internal desynchronization in
central pacemakers. In the mammalian SCN, evidence from different
laboratories has lent support to the notion that membrane excitability
or, more precisely, a certain degree of depolarization and activation
of Ca2+ and cAMP second messenger cascades, may be required for
sustained molecular oscillations. These observations underscore that
intercellular communication is important to reinforce high amplitude
molecular oscillations through synchronization of independent cellular
oscillators, as opposed to being an essential component within the
mechanism responsible for the generation of the molecular
oscillations. Interestingly, it has been reported that, in a subset of
SCN neurons, molecular oscillations of a circadian reporter still take
place even in the absence of synaptic connectivity, highlighting the
autonomy of the molecular oscillator (Depetris-Chauvin, 2011). Adult-restricted silencing of the PDF circuit impairs locomotor
behavior to a similar extent compared to constitutively silencing
them, demonstrating that regardless of the overall levels of KIR
achieved through the inducible system, short-term expression
effectively prevents communication with other neuronal targets. Such a
scenario offers the possibility to identify the direct consequences of
reducing the excitability of the PDF circuit in a defined temporal
window. Surprisingly, despite kir expression being limited to the
adult brain, it correlated with axonal arbors of reduced complexity
throughout the day in the dorsal protocerebrum, even though the
circadian remodeling phenomenon continued to take place. The latter
lends further support to the notion that no effect on the pace of the
molecular oscillator became evident during the acutely silenced phase
(Depetris-Chauvin, 2011). In addition, adult-restricted silencing correlated with noncycling
PDF levels. PDF is transported along the axonal tract in large dense
core vesicles (DCV), which apparently are released outside of the
chemical synapse. Although no precise information is available on PDF,
it is expected for neuropeptides to be released after high frequency
stimulation, suggesting that during the silenced phase, the DCV would
accumulate in the axonal terminals. It has been proposed that the
trough of PDF accumulation at dusk might represent the depletion of
the PDF readily releasable pool, and it correlates with the time of
day when the sLNvs are most hyperpolarized. Interestingly, despite no
release expected to occur while KIR is expressed, PDF intensity at the
axonal terminals stayed at trough levels throughout the day,
underscoring that reduced excitability affected additional steps such
as peptide synthesis, processing, or transport. In favor of this
possibility, hyperexcitation of the PDF circuit correlates with
constantly high (daytime) PDF levels at the dorsal protocerebrum.
Moreover, once kir expression was turned off, PDF levels resume to
cycle, indicating a direct modulatory effect of membrane excitability
on this specific output. In line with a defective output from the
sLNvs, desynchronization of dorsal oscillators (i.e., the DN1s) became
evident as early as in DD4. Gaining more insight into the mechanisms
of communication within the circadian network, as well as those
connecting the cell membrane with the molecular clock, will provide a
better understanding on how these components interact to sustain
temporal and spatial order to shape rhythmic overt behavior
(Depetris-Chauvin, 2011). Taken together, these results confirm that in
Drosophila, altering membrane excitability mainly affects the output
of pacemaker cells and thus intercellular communication, as is the
case in the eye of the mollusk Bulla and the rodent SCN, highlighting
the degree of conservation in the mechanisms underlying the biological
clock in distant organisms (Depetris-Chauvin, 2011). A daily body temperature rhythm (BTR) is critical for the maintenance
of homeostasis in mammals. Whereas mammals use internal energy to
regulate body temperature, ectotherms typically regulate body
temperature behaviorally. Some ectotherms maintain homeostasis via a
daily temperature preference rhythm (TPR), but the underlying
mechanisms are largely unknown. This study shows that Drosophila
exhibit a daily circadian clock-dependent TPR that resembles mammalian
BTR. Pacemaker neurons critical for locomotor activity are not
necessary for TPR; instead, the dorsal neuron 2 s (DN2s), whose
function was previously unknown, is sufficient. This indicates that
TPR, like BTR, is controlled independently from locomotor activity.
Therefore, the mechanisms controlling temperature fluctuations in fly
TPR and mammalian BTR may share parallel features. Taken together,
these results reveal the existence of a novel DN2-based circadian
neural circuit that specifically regulates TPR; thus, understanding
the mechanisms of TPR will shed new light on the function and neural
control of circadian rhythms (Kaneko, 2012). Drosophila exhibit a daily TPR-low in the morning, high in the
evening-that follows a similar pattern as in humans. This study is not
only the first demonstration of fly TPR, but also the first systematic
analysis of the molecular and neural mechanisms underlying TPR. TPR is
controlled by the DN2s, which might explain why temperature preference
remains rhythmic in LL. The DN2s do not express CRYPTOCHROME (CRY), a
blue-light sensor crucial for circadian photoreception. Arrhythmicity
in LL is caused by constant activation of CRY and thus constant
degradation of Tim. Therefore, CRY-negative DN2 neurons may maintain
residual rhythms in LL for a longer period of time than CRY-positive
circadian neurons. To explore this possibility, immunostaining of
brains were performed with Tim-antibody, and the staining levels of
DN2 cells were analyzed in LL 4 days. Although Tim signal was found to
be weakly rhythmic in DN2 neurons, these oscillations were not
statistically significant. Further studies will thus be needed to
verify that DN2 neurons maintain residual rhythm in LL. Because
locomotor activity is controlled by CRY-positive circadian neurons and
rapidly becomes arrhythmic in LL, the maintenance of TPR rhythms in LL
also supports the conclusion that locomotor activity and TPR are
controlled by independent circadian neural pathways (Kaneko, 2012). The data reveal striking parallel features between fly TPR and
mammalian BTR, although the modes of heat production are not the same.
Flies exhibit robust temperature increases during the daytime, which
is the same phenomenon as diurnal mammalian BTR. Furthermore, ablation
studies in rats show that BTR is controlled by SCN neurons that target
a different subset of subparaventricular zone neurons than those that
control locomotor activity. Thus, both fly TPR and mammalian BTR
exhibit circadian clock dependent temperature fluctuations,
independently regulated from locomotor activity. Taken together, these
data raises the possibility that mammalian BTR and fly TPR are
evolutionally conserved, which may be because temperature fluctuation
in an animal's body is fundamental for maintenance of its homeostasis
(Kaneko, 2012). Why do flies exhibit TPR? Flies probably exhibit TPR primarily to
maintain homeostasis, like mammals. Mammalian BTR has been shown to
have a clear interaction with sleep, and it has been reported that
mechanisms controlling fly sleep are analogous to those controlling
mammalian sleep. Therefore, fly TPR may also have a relationship with
sleep. Intriguingly, it was observed that PER expression limited to
pdf neurons can generate a weak TPR with an abnormal phase in per01
mutants, suggesting that pdf neurons may have a role in the TPR
circuits. pdf-positive (sLNv and lLNv) neurons regulate sleep and
sLNvs are known to project near the DN2s. Therefore, pdf neurons may
be able to modulate DN2 activity even when these neurons are
arrhythmic and may represent the neural basis for an interaction
between TPR and sleep (Kaneko, 2012). Additionally, TPR may provide feedback to
circadian pacemakers. Ambient temperature fluctuations can entrain not
only peripheral clocks in mammals and flies but also circadian
pacemaker neurons in the fly brain, which contribute to morning and
evening locomotor activity. Because TPR can generate temperature
fluctuations in the fly body, the output of TPR may thus reinforce or
refine circadian rhythm entrainment. For circadian locomotor behavior,
the DN2s could actually participate in the reinforcement, because in
the larval brain the DN2s help the sLNvs entraining to temperature
cycles. Therefore, by further exploring this newly discovered
circadian rhythm, Drosophila TPR might not only help understanding the
mechanisms underlying body temperature control in animals but also
contribute to a greater understanding of circadian rhythm's mammalian
CSLs plasticity (Kaneko, 2012).
Circadian clocks organize biological processes to occur at optimized times of day and thereby contribute to overall fitness. While the regular daily changes of environmental light and temperature synchronize circadian clocks, extreme external conditions can bypass the temporal constraints dictated by the clock. Despite advanced knowledge about how the daily light-dark changes synchronize the clock, relatively little is known with regard to how the daily temperature changes influence daily timing and how temperature and light signals are integrated. In Drosophila, a network of approximately 150 brain clock neurons exhibit 24-hr oscillations of clock gene expression to regulate daily activity and sleep. This study shows that a temperature input pathway from peripheral sensory organs, which depends on the gene nocte, targets specific subsets of these clock neurons to synchronize molecular and behavioral rhythms to temperature cycles. Strikingly, while nocte1 mutant flies synchronize normally to light-dark cycles at constant temperatures, the combined presence of light-dark and temperature cycles inhibits synchronization. nocte1 flies exhibit altered siesta sleep, suggesting that the sleep-regulating clock neurons are an important target for nocte-dependent temperature input, which dominates a parallel light input into these cells. In conclusion, this study reveals a nocte-dependent temperature input pathway to central clock neurons and shows that this pathway and its target neurons are important for the integration of sensory light and temperature information in order to temporally regulate activity and sleep during daily light and temperature cycles (Chen, 2018).
This study reveals a nocte-dependent and presumably chordotonal organ (Cho)-dependent temperature input to LNd and DN subsets of the Drosophila clock circuit. While it has previously been shown that nocte and Cho contribute to temperature entrainment of the circadian clock, it was not known which of the clock neurons are targeted by this peripheral sensory input pathway. The neuronal targets that have been identified in this study are largely consistent with previous studies showing that most of the clock neurons not expressing CRY are more responsive to fluctuating temperature signals compared to CRY+ cells. The lack of normal synchronized TIM expression in the DN1-DN3 and LNds of nocte1 flies shows that nocte-dependent temperature input serves as the main thermal entrainment signal for these cells. Interestingly, loss of the variant glutamate receptor IR25a, which impairs entrainment to temperature cycles (TCs), also strongly affected the DN1s and LNds, highlighting the importance of these neuronal groups for temperature entrainment. In fact, blocking synaptic transmission from the DN1 via expression of tetanus toxin interfered with behavioral synchronization to TCs. The low-amplitude oscillations of TIM in the LNv groups are either caused by alternative temperature inputs into these cells or result from the initial exposure to LD cycles before the flies were transferred to TCs (Chen, 2018).
In addition to uncovering the neuronal targets of the NOCTE-Cho temperature pathway, this study revealed that its function is also important during more natural environmental situations, i.e., the combined presence of in-phase or conflicting TCs and LD cycles. This is very surprising, considering that nocte mutants synchronize normally to LD cycles at constant temperatures. During combined LD and TC conditions, the DN1-DN3 and LNd groups were most severely affected as evidenced by the lack (DN2) or severe amplitude reduction of synchronized TIM and PER-LUC oscillations. In contrast to the situation in DD and TC, all three exclusively CRY-positive LNv groups now exhibit synchronized TIM oscillations, which can be explained by CRY-mediated light resetting within these neurons. Only about 50% of the DN1s and LNds express CRY, so that the non-significant, low-amplitude TIM oscillations within these groups could potentially be explained by light synchronization within the CRY-positive neurons. However, this idea is not favored, because in the absence of LD cycles, imilar non-significant TIM oscillations were observed within these neuronal groups. Moreover, the CRY-negative DN3s exhibit the same low-amplitude oscillations in the presence of combined LD and TCs. Based on the observation that low-amplitude TIM oscillations are abolished in non-entrained and increased in weakly entrained nocte1 flies, it is argued that the lack of full penetrance contributes to the shallow TIM oscillations observed when both nocte1 phenotypic classes are averaged (Chen, 2018).
Over the last few years, the DN1s have been shown to be a versatile subset of the clock neuronal circuit. They receive and integrate multiple environmental sensory inputs to regulate accurate timing of locomotor activity and sleep. Avery recent study elegantly shows that the DN1s are activated by very brief (<20 s) temperature reductions from 23°C to 16°C in a nocte- and Cho-dependent manner. Because nocte1 mutants or reducing nocte function in ChO impairs temperature entrainment, these rapid DN1 temperature responses most likely also contribute to the detection of long-term temperature fluctuations relevant for daily temperature entrainment (Chen, 2018).
DN1s support morning activity at warmer (25°C) and evening activity at colder (20°C) temperatures, while at low light levels, they also support evening activity at 25°C, presumably revealing a light and temperature integration function of the DN1s. This study shows that the DN1s are among the clock neurons that are strongly affected by the lack of nocte function, both during 16°C:25°C TC and combined TCs and LD cycles, supporting a role for this neuronal group in light and temperature integration. The strong nocte1 effects on the CRY-positive LNvs during combined LD and TCs raise the possibility that they are coupled to the temperature-input-receiving LNd and DN groups. This would also explain the 'dominant-negative' effects of nocte1 on synchronization during combined LD and TCs. It is proposed that, in the presence of TCs, nocte prevents inappropriate signaling from the DN and LNd groups to the LNvs, thereby contributing to the correct integration of light and temperature (Chen, 2018).
With regard to sleep regulation, temperature-dependent functions of the DN1 are more complex. In general, warmer temperatures increase daytime sleep and decrease sleep at night. The daytime increase in sleep depends on a functional clock and PER expression in the DN1s, showing that a subset of this heterogeneous neuronal group can have sleep-promoting functions. This DN1 subset presumably represses the activity-promoting LNvs and LNds via glutamate (Chen, 2018).
This study shows that nocte1 flies exhibit significantly reduced siesta sleep (ZT3-ZT9) during the warm phase of combined TC and LD cycles-a condition disrupting molecular synchronization in the DN1. Although this study did not directly address a role for nocte in regulating DN1 function in sleep, these results are in line with a sleep-promoting function for the DN1s below 30°C. In the absence of light cues (TCs in DD), wild-type flies gradually initiate a period of inactivity and sleep during the warm phase starting after ZT6 and reaching peak inactivity and sleep levels toward the end of the warm phase at ZT10. During this interval (ZT6-ZT10) nocte1 flies show the exact opposite behavior and drastically decrease their sleep levels. This points again to a sleep-supporting role for the DN1s, in line with the results obtained for combined LD and TC synchronization and results by Yadlapalli (2018) showing that the DN1s promote sleep during ramped TCs. It is proposed that, in nocte1 flies, DN1 activity is altered during TCs, resulting in impaired behavioral and molecular synchronization, as well as in a reduced ability to repress activity promoting clock neurons (Chen, 2018).
Human body temperature increases during wakefulness and decreases during sleep. The body temperature rhythm (BTR) is a robust output of the circadian clock and is fundamental for maintaining homeostasis, such as generating metabolic energy and sleep, as well as entraining peripheral clocks in mammals. However, the mechanisms that regulate BTR are largely unknown. Drosophila are ectotherms, and their body temperatures are close to ambient temperature; therefore, flies select a preferred environmental temperature to set their body temperature. This study identified a novel circadian output, the temperature preference rhythm (TPR), in which the preferred temperature in flies increases during the day and decreases at night. TPR, thereby, produces a daily BTR. Fly TPR shares many features with mammalian BTR. Diuretic hormone 31 receptor (DH31R) was found to mediates Drosophila TPR, and the closest mouse homolog of DH31R, calcitonin receptor (Calcr), is essential for mice BTR. Importantly, both TPR and BTR are regulated in a distinct manner from locomotor activity rhythms, and neither DH31R nor Calcr regulates locomotor activity rhythms. These findings suggest that DH31R/Calcr is an ancient and specific mediator of BTR. Thus, understanding fly TPR will provide fundamental insights into the molecular and neural mechanisms that control BTR in mammals (Goda, 2019).
Circadian clocks are characterized by three properties: they run in constant conditions with a period of ∼24 h, synchronize to the environmental cycles of light and temperature, and are temperature compensated, meaning they do not speed up with temperature. Central brain clocks regulate daily activity rhythms, whereas peripheral clocks are dispersed throughout the body of insects and vertebrates. Using a set of luciferase reporter genes, this study shows that Drosophila peripheral clocks are self-sustained but over-compensated, i.e., they slow down with increasing temperature. In contrast, central clock neurons in the fly brain, both in intact flies and in cultured brains, show accurate temperature compensation. Although this suggests that neural network properties contribute to temperature compensation, the circadian neuropeptide Pigment Dispersing Factor (PDF) is not required for temperature-compensated oscillations in brain clock neurons. These findings reveal a fundamental difference between central and peripheral clocks, which likely also applies for vertebrate clocks (Versteven, 2020).
The Drosophila circadian pacemaker consists of transcriptional feedback loops subjected to post-transcriptional and post-translational regulation. While post-translational regulatory mechanisms have been studied in detail, much less is known about circadian post-transcriptional control. Thus, this study targeted 364 RNA binding and RNA associated proteins with RNA interference. Among the 43 hits that were identified was the alternative splicing regulator P-element somatic inhibitor (PSI). PSI regulates the thermosensitive alternative splicing of timeless (tim), promoting splicing events favored at warm temperature over those increased at cold temperature. Psi downregulation shortens the period of circadian rhythms and advances the phase of circadian behavior under temperature cycle. Interestingly, both phenotypes were suppressed in flies that could produce TIM proteins only from a transgene that cannot form the thermosensitive splicing isoforms. Therefore, it is concluded that PSI regulates the period of Drosophila circadian rhythms and circadian behavior phase during temperature cycling through its modulation of the tim splicing pattern (Foley, 2019).
Increasing evidence indicates that post-transcriptional mechanisms controlling gene expression are also critical for the proper function of circadian clocks in many organisms. In Drosophila, the post-transcriptional regulation of per mRNA has been best studied. per mRNA stability changes as a function of time. In addition, per contains an intron in its 3'UTR (dmpi8) that is alternatively spliced depending on temperature and lighting conditions. On cold days, the spliced variant is favored, causing an advance in the accumulation of per transcript levels as well as an advance of the evening activity peak. This behavioral shift means that the fly is more active during the day when the temperature would be most tolerable in their natural environment. The temperature sensitivity of dmpi8 is due to the presence of weak non-canonical splice sites. However, the efficiency of the underlying baseline splicing is affected by four single nucleotide polymorphisms (SNPs) in the per 3'UTR that vary in natural populations and form two distinct haplotypes. Also, while this splicing is temperature-sensitive in two Drosophila species that followed human migration, two species that remained in Africa lack temperature sensitivity of dmpi8 splicing. Furthermore, it has been recently demonstrated that the the trans-acting splicing factor B52 enhances dmpi8 splicing efficiency, and this effect is stronger with one of the two haplotypes. per is also regulated post-transcriptionally by the TWENTYFOUR-ATAXIN2 translational activation complex. This complex works by binding to per mRNA as well as the cap-binding complex and poly-A binding protein. This may enable more efficient translation by promoting circularization of the transcript. Interestingly, this mechanism appears to be required only in the circadian pacemaker neurons. Non-canonical translation initiation has also been implicated in the control of PER translation. Regulation of PER protein translation has also been studied in mammals, with RBM4 being a critical regulator of mPER1 expression. In flies however, the homolog of RBM4, LARK, regulates the translation of DBT, a PER kinase. miRNAs have emerged as important critical regulators of circadian rhythms in Drosophila and mammals, affecting the circadian pacemaker itself, as well as input and output pathways controlling rhythmic behavioral and physiological processes (Foley, 2019).
RNA-associated proteins (RAPs) include proteins that either bind directly or indirectly to RNAs. They mediate post-transcriptional regulation at every level. Many of these regulated events - including alternative splicing, splicing efficiency, mRNA stability, and translation - have been shown to function in molecular clocks. Thus, to obtain a broad view of the Drosophila circadian RAP landscape and its mechanism of action, an RNAi screen was performed targeting 364 of these proteins. This led to the discovery of a role for the splicing factor P-element somatic inhibitor (PSI) in regulating the pace of the molecular clock through alternative splicing of tim (Foley, 2019).
The results identify a novel post-transcriptional regulator of the circadian clock: PSI. PSI is required for the proper pace of both brain and body clock, and for proper phase-relationship with ambient temperature cycles. When Psi is downregulated, the circadian pacemaker speeds up and behavior phase under temperature cycles is advanced by 3 hr, and these phenotypes appear to be predominantly caused by an abnormal tim splicing pattern. Indeed, the circadian period and behavior phase of flies that can only produce functional TIM protein from a transgene missing most introns is insensitive to Psi downregulation. It is noted however that cwo's splicing pattern is also affected by Psi downregulation, and sgg splicing pattern was not studied, although it might also be controlled by PSI. It is therefore not possible to exclude a small contribution of non-tim splicing events to PSI downregulation phenotypes, or that in specific tissues these other splicing events play a greater role than in the brain (Foley, 2019).
Interestingly, Psi downregulation results in an increase in intron inclusion events that are favored under cold conditions (tim-sc and tim-cold), while an intron inclusion event favored under warm conditions is decreased (tim-M). However, the ability of tim splicing to respond to temperature changes is not abolished when Psi is downregulated. This could imply that an as yet unknown factor specifically promotes or represses tim splicing events in a temperature-dependent manner. Another possibility is that the strength of splice sites or tim's pre-mRNA structure impacts splicing efficiency in a temperature-dependent manner. For example, suboptimal per splicing signals explain the lower efficiency of per's most 3' splicing event at warm temperature (Foley, 2019).
How would the patterns of tim splicing affect the pace of the circadian clock, or advance the phase of circadian behavior under temperature cycles? In all splicing events that were studied, intron retention results in a truncated TIM protein. It is therefore possible that the balance of full length and truncated TIM proteins, which may function as endogenous dominant-negatives, determines circadian period. For example, truncated TIM might be less efficient at protecting PER from degradation, thus accelerating the pacemaker, or affecting its phase. Consistent with this idea, overexpression of the shorter cold-favored tim isoform (tim-sc) shortens period. Strikingly, Psi downregulation increases this isoform's levels and also results in a short phenotype. It has been proposed that production of tim-M transcripts (called tim-tiny in their study) delays the rate of TIM accumulation. Such a mechanism could also contribute to the short period observed when Psi is downregulated, since this reduces tim-M levels, which may accelerate TIM accumulation. Another interesting question is how PSI differentially affects specific splice isoforms of tim. One possibility is that the execution of a specific tim splicing event negatively influences the probability of the occurrence of other splicing events. For example, PSI could downregulate tim-sc and tim-cold by enhancing splicing and removal of the introns whose retention is necessary for production of these isoforms. This could indirectly reduce splicing of the intron that is retained in the warm tim-M isoform and result in tim-M upregulation. Conversely, PSI could directly promote tim-M intron retention and indirectly downregulate production of tim-sc and tim-cold (Foley, 2019).
Other splicing factors have been shown to be involved in the control of circadian rhythms in Drosophila. SRm160 contributes to the amplitude of circadian rhythms by promoting per expression, while B52/SMp55 and PRMT5 regulate per's most 3' splicing, which is temperature sensitive. Loss of PRMT5 results in essentially arrhythmic behavior, but this is unlikely to be explained by its effect on per's thermosensitive splicing. B52/SMp55 knockdown flies show a reduced siesta, which is controlled by the same per splicing. With the identification of Psi, this study has uncover a key regulator of tim alternative splicing pattern and shows that this pattern determines circadian period length, while per alternative splicing regulates the timing and amplitude of the daytime siesta. Interestingly, a recent study identified PRP4 kinase and other members of tri-snRNP complexes as regulators of circadian rhythms. Downregulation of prp4 caused excessive retention of the tim-M intron. PSI and PRP4 might thus have complementary functions in tim mRNA splicing regulation, working together to maintain the proper balance of tim isoform expression (Foley, 2019).
An unexpected finding is the role played by both PDF neurons and other circadian neurons in the short period phenotype observed with circadian locomotor rhythms when Psi was knocked-down. Indeed, it is quite clear from multiple studies that under constant darkness, the PDF-positive sLNvs dictate the pace of circadian behavior. Why, in the case of Psi downregulation, do PDF negative neurons also play a role in period determination? The explanation might be that PSI alters the hierarchy between circadian neurons, promoting the role of PDF negative neurons. This could be achieved by weakening PDF/PDFR signaling, for example (Foley, 2019).
While this study focused on PSI, several other interesting candidates were identified in the screen. The presence of a large number of splicing factors is noted. This adds to the emerging notion that alternative splicing plays a critical role in the control of circadian rhythms. Several per splicing regulators have been mentioned that can impact circadian behavior. In addition, a recent study demonstrated that specific classes of circadian neurons express specific alternative splicing variants, and rhythmic alternative splicing is widespread in these neurons. Interestingly, in this study, the splicing regulator barc, which was identified in the screen and which has been shown to causes intron retention in specific mRNAs, was found to be rhythmically expressed in LNds. Moreover, in mammals, alternative splicing appears to be very sensitive to temperature, and could explain how body temperature rhythms synchronize peripheral clocks. Another intriguing candidate is cg42458, which was found to be enriched in circadian neurons (LNvs and Dorsal Neurons 1). In addition to emphasizing the role of splicing, the screen suggests that regulation of polyA tail length is important for circadian rhythmicity, since several members of the CCR4-NOT complex and deadenylation-dependent decapping enzymes were identified. Future work will be required to determine whether these factors directly target mRNAs encoding for core clock components, or whether their effect on circadian period is indirect. Interestingly, the POP2 deadenylase, which is part of the CCR4-NOT complex, was recently shown to regulate tim mRNA levels post-transcriptionally. It should be noted that while the screen targeted 364 proteins binding or associated with RNA, it did not include all of them. For example, LSM12, which was recently shown to be a part of the ATXN2/TYF complex, was not included in the screen because it had not been annotated as a potential RAP when the screen was initiated (Foley, 2019).
In summary, this work provides an important resource for identifying RNA associated proteins regulating circadian rhythms in Drosophila. It identifies PSI is an important regulator of circadian period and circadian phase in response to thermal cycles, and points at additional candidates and processes that determine the periodicity of circadian rhythms (Foley, 2019).
Mosquitoes exhibit activity rhythms, crucial for the transmission of pathogens, under the control of a circadian clock. Aedes aegypti is one of the world's leading vectors. For decades, several studies have linked the rise in ambient temperature with the increase in their activity. This study identified candidate genes whose expression is influenced by temperature cycles and may affect Aedes locomotor activity. timeless completely lost its rhythmic expression in light/dark, with out-of-phase temperature cycles, and by RNAi mediated knockdown of nocte, an important gene for Drosophila circadian synchronization by temperature cycles. Thus, timeless and nocte are important genes for synchronization by temperature cycles in Aedes aegypti. To reinforce these findings, the gradual temperature fluctuations that were as close as possible to daily temperature variations in Brazil were simulated in the laboratory. It was observed that the activity and the expression of the molecular circadian clock of Ae. aegypti differs significantly from that of mosquitoes subjected to constant or rectangular abrupt changes in temperature. It is suggested that for understanding the circadian behavior of Aedes with possible implications for intervention strategies, the seminatural paradigm needs to replace the traditional laboratory study (Teles-de-Freitas, 2020).
Circadian rhythms are based on biochemical oscillations generated by clock genes/proteins, which independently evolved in animals, fungi, plants, and cyanobacteria. Temperature compensation of the oscillation speed is a common feature of the circadian clocks, but the evolutionary-conserved mechanism has been unclear. This study shows that Na(+)/Ca(2+) exchanger (NCX) mediates cold-responsive Ca(2+) signaling important for the temperature-compensated oscillation in mammalian cells. In response to temperature decrease, NCX elevates intracellular Ca(2+), which activates Ca(2+)/calmodulin-dependent protein kinase II and accelerates transcriptional oscillations of clock genes. The cold-responsive Ca(2+) signaling is conserved among mice, Drosophila, and Arabidopsis. The mammalian cellular rhythms and Drosophila behavioral rhythms were severely attenuated by NCX inhibition, indicating essential roles of NCX in both temperature compensation and autonomous oscillation. NCX also contributes to the temperature-compensated transcriptional rhythms in cyanobacterial clock. These results suggest that NCX-mediated Ca(2+) signaling is a common mechanism underlying temperature-compensated circadian rhythms both in eukaryotes and prokaryotes (Kon, 2021).
Circadian TTFLs are an elaborate system that drives a wide range of overt rhythms with various phase angles and amplitudes. The oscillation speed of the TTFLs is temperature compensated, although many of the biochemical reactions in TTFLs are slowed down by decreasing temperature. This study demonstrates that the temperature compensation of the TTFL in mammalian cells was compromised when Ca2+-dependent phosphorylation signaling was inhibited. An important role was found for NCX-CaMKII activity as the state variable of the circadian oscillator. This present study and a series of preceding works demonstrate that the Ca2+ oscillator plays essential roles in the circadian oscillation mechanism. Functional studies clearly demonstrated essential roles of NCX-dependent Ca2+ signaling in the three important properties of the circadian clock, i.e., cell-autonomous oscillation, temperature compensation, and entrainment. The circadian Ca2+ oscillation is observed in mice lacking Bmal1 or Cry1/Cry2, implicating that the Ca2+ oscillator is an upstream regulator of the TTFL in mammals (Kon, 2021).
The effects of NCX2 and NCX3 deficiencies on the regulation of mouse behavioral rhythms suggest involvement of Na+/Ca2+ exchanging activity in the Ca2+ dynamics of the SCN. Previous studies showed that L-type Ca2+ channel (LTCC) and voltage-gated Na+ channel (VGSC) are required for high-amplitude Ca2+ rhythms in the SCN. Because NCX activities are regulated by local concentrations of Na+/Ca2+ and the membrane potential, cooperative actions of LTCC, VGSC, and NCX seem to play important roles in generation mechanism of the robust Ca2+ oscillations in the SCN (Kon, 2021).
It should be emphasized that the role of Ca2+/calmodulin-dependent protein kinases is conserved among clockworks in insects, fungi, and plants, suggesting that the Ca2+ oscillator might be a core timekeeping mechanism in their common ancestor (see Involvement of ancient Ca2+ signaling for temperature-compensated circadian rhythms). After divergence of each lineage, a subset of clock genes should have independently evolved in association with the Ca2+ oscillator. It is noteworthy that NCX is also required for temperature compensation of PTO-based cyanobacterial clock. Because intracellular Ca2+ in cyanobacteria is elevated in response to temperature decrease, YrbG-mediated Ca2+ signaling may regulate the PTO in vivo. Conservation of NCX among eukaryotes, eubacteria, and archaea suggests that NCX-dependent temperature signaling is essential for adaptation of a wide variety of organisms to environment. Further studies on NCX-regulated Ca2+ flux will provide evolutionary insights into the origin of the circadian clocks (Kon, 2021).
Circadian rhythms optimize health by coordinating the timing of physiological processes to match predictable daily environmental challenges. The circadian rhythm of body temperature is thought to be an important modulator of molecular clocks in peripheral tissues, but how daily temperature cycles impact physiological function is unclear. This study examined the effect of constant (25°C, T(CON)) and cycling (28°C/22°C during light/dark, T(CYC)) temperature paradigms on lifespan of Drosophila melanogaster, and the expression of clock genes, Heat shock protein 83 (Hsp83), Frost (Fst), and Senescence-associated protein 30 (smp-30). Male and female Drosophila housed at T(CYC) had longer median lifespans than those housed at T(CON) T(CYC) induced robust Hsp83 rhythms and rescued the age-related decrease in smp-30 expression that was observed in flies at T(CON), potentially indicating an increased capacity to cope with age-related cellular stress. Ageing under T(CON) led to a decrease in the amplitude of expression of all clock genes in the bodies of male flies, except for cyc, which was non-rhythmic, and for per and cry in female flies. Strikingly, housing under T(CYC) conditions rescued the age-related decrease in amplitude of all clock genes, and generated rhythmicity in cyc expression, in the male flies, but not the female flies. The results suggest that ambient temperature rhythms modulate Drosophila lifespan, and that the amplitude of clock gene expression in peripheral body clocks may be a potential link between temperature rhythms and longevity in male Drosophila Longevity due to T(CYC) appeared predominantly independent of clock gene amplitude in female Drosophila (Goh, 2021).
Sleep-wake stability is imbalanced with natural aging, and microRNAs (miRNAs) play important roles in cell proliferation, apoptosis, and aging; however, the biological functions of miRNAs in regulating aging-related sleep-wake behavior remain unexplored. This study varied the expression pattern of dmiR-283 in Drosophila and the result showed that the aging decline in sleep-wake behavior was caused by the accumulation of brain dmiR-283 expression, whereas the core clock genes cwo and Notch signaling pathway might be suppressed, which regulate the aging process. In addition, to identify exercise intervention programs of Drosophila that promote healthy aging, mir-283SP/+ (mir-23SP referes to mir-283sponge) and Pdf > mir-283SP flies were driven to perform endurance exercise for a duration of 3 weeks starting at 10 and 30 days, respectively. The results showed that exercise starting in youth leads to an enhanced amplitude of sleep-wake rhythms, stable periods, increased activity frequency upon awakening, and the suppression of aging brain dmiR-283 expression in mir-283SP/+ middle-aged flies. Conversely, exercise performed when the brain dmiR-283 reached a certain accumulation level showed ineffective or negative effects. In conclusion, the accumulation of dmiR-283 expression in the brain induced an age-dependent decline in sleep-wake behavior. Endurance exercise commencing in youth counteracts the increase in dmiR-283 in the aging brain, which ameliorates the deterioration of sleep-wake behavior during aging (Li, 2023).
The brain as a central regulator of stress integration determines what is threatening, stores memories, and regulates physiological adaptations across the aging trajectory. While sleep homeostasis seems to be linked to brain resilience, how age-associated changes intersect to adapt brain resilience to life history remains enigmatic. This study provides evidence that a brain-wide form of presynaptic active zone plasticity ("PreScale"), characterized by increases of active zone scaffold proteins and synaptic vesicle release factors, integrates resilience by coupling sleep, longevity, and memory during early aging of Drosophila. PreScale increased over the brain until mid-age, to then decreased again, and promoted the age-typical adaption of sleep patterns as well as extended longevity, while at the same time it reduced the ability of forming new memories. Genetic induction of PreScale also mimicked early aging-associated adaption of sleep patterns and the neuronal activity/excitability of sleep control neurons. Spermidine supplementation, previously shown to suppress early aging-associated PreScale, also attenuated the age-typical sleep pattern changes. Pharmacological induction of sleep for 2 days in mid-age flies also reset PreScale, restored memory formation, and rejuvenated sleep patterns. The data suggest that early along the aging trajectory, PreScale acts as an acute, brain-wide form of presynaptic plasticity to steer trade-offs between longevity, sleep, and memory formation in a still plastic phase of early brain aging (Huang, 2022).
As a normal physiological phenomenon, aging has a significant impact on sleep. Aging leads to sleep impairment, including sleep loss, fragmented sleep, and a lower arousal threshold, leading to various diseases. Because sleep regulates memory consolidation, age-dependent sleep impairment also affects memory. However, the mechanisms underlying age-related sleep dysregulation and its impact on memory remain unclear. Using male and female Drosophila, which possesses sleep characteristics similar to those of mammals and exhibits age-dependent sleep impairment as a model, small-molecule screening was performed to identify novel regulators of age-dependent decline in sleep. The screening identified 3,3´-difluorobenzaldazine (DFB), a positive allosteric modulator of the metabotropic glutamate receptor (mGluR) 5, as a novel sleep-promoting compound in aged flies. The study found that mutant flies of mGluR, a single mGluR gene in Drosophila, and decreased mGluR expression had significant impairment in sleep and memory due to olfactory conditioning. The decreased sleep phenotype in the mGluR mutants was not promoted by DFB, suggesting that the effects of DFB on age-dependent sleep impairment are dependent on mGluR. Although aging decreases the expression of mGluR and the binding scaffold proteins Homer and Shank, the transient overexpression of mGluR in neurons improves sleep in both young and aged flies. Overall, these findings indicate that age-dependent decreased expression or function of mGluR impairs sleep and memory in flies, which could lead to age-related sleep and memory impairment (Hou, 2023).
Circadian clocks control many self-sustained rhythms in physiology and behavior with approximately 24-hour periodicity. This study examined the effects of clock disruptions on locomotor aging and longevity in Drosophila. Lifespan was found to be similarly reduced in three arrhythmic mutants (ClkAR, cyc0 and tim0) and in wild-type flies under constant light, which stops the clock. In contrast, ClkAR mutants showed significantly faster age-related locomotor deficits (as monitored by startle-induced climbing) than cyc0 and tim0, or than control flies under constant light. Clk, but not Cyc, inactivation by RNA interference in the pigment-dispersing factor (PDF)-expressing central pacemaker neurons led to similar loss of climbing performance as ClkAR. Conversely, restoring Clk function in these cells was sufficient to rescue the ClkAR locomotor phenotype, independently of behavioral rhythmicity. Accelerated locomotor decline of the ClkAR mutant required expression of the PDF receptor and correlated to an apparent loss of dopaminergic neurons in the posterior protocerebral lateral 1 (PPL1) clusters. This neuronal loss was rescued when the ClkAR mutation was placed in an apoptosis-deficient background. Impairing dopamine synthesis in a single pair of PPL1 neurons that innervate the mushroom bodies accelerated locomotor decline in otherwise wild-type flies. These results therefore reveal a novel circadian-independent requirement for Clk in brain circadian neurons to maintain a subset of dopaminergic cells and avoid premature locomotor aging in Drosophila (Vaccaro, 2023).
Circadian clocks may mediate lifespan extension by caloric or dietary restriction (DR). This study found that the core clock transcription factor Clock is crucial for a robust longevity and fecundity response to DR in Drosophila. To identify clock-controlled mediators, RNA-sequencing was performed from abdominal fat bodies across the 24 h day after just 5 days under control or DR diets. In contrast to more chronic DR regimens, no significant changes were detected in the rhythmic expression of core clock genes. Yet it was discovered that DR induced de novo rhythmicity or increased expression of rhythmic clock output genes. Network analysis revealed that DR increased network connectivity in one module comprised of genes encoding proteasome subunits. Adult, fat body specific RNAi knockdown demonstrated that proteasome subunits contribute to DR-mediated lifespan extension. Thus, clock control of output links DR-mediated changes in rhythmic transcription to lifespan extension (Hwangbo, 2023).
Sleep is an essential and evolutionarily conserved behavior that is modulated by many environmental factors. Ambient temperature shifting usually occurs during climatic or seasonal change or travel from high-latitude area to low-latitude area that affects animal physiology. Increasing ambient temperature modulates sleep in both humans and Drosophila. Although several thermosensory molecules and neurons have been identified, the neural mechanisms that integrate temperature sensation into the sleep neural circuit remain poorly understood. This study reveals that prolonged increasing of ambient temperature induces a reversible sleep reduction and impaired sleep consolidation in Drosophila via activating the internal thermosensory anterior cells (ACs). ACs form synaptic contacts with a subset of posterior dorsal neuron 1 (DN1p) neurons and release acetylcholine to promote wakefulness. Furthermore, this study identified that this subset of DN1ps promotes wakefulness by releasing CNMamide (CNMa) neuropeptides to inhibit the Dh44-positive pars intercerebralis (PI) neurons through CNMa receptors. This study demonstrates that the AC-DN1p-PI neural circuit is responsible for integrating thermosensory inputs into the sleep neural circuit. Moreover, the CNMa signaling pathway was identified as a newly recognized wakefulness-promoting DN1 pathway (Jin, 2021).
Sleep is an essential and evolutionarily conserved behavior across species and it is critical for synaptic plasticity, learning and memory, neuronal development, and metabolite clearance in both vertebrates and invertebrates. Sleep is a reversible status characterized by quiescence and an increased arousal threshold which is modulated by many environmental factors. Among these, temperature shifting is a common and essential factor that is associated with climatic or seasonal change or travel from high-latitude area to low-latitude area to affect the physiology of all animals. In humans, increased ambient temperature influences sleep patterns and correlates with reductions in rapid eye movement sleep. In Drosophila, numerous studies have shown that increasing ambient temperature modifies sleep architecture. However, how thermosensory input integrates into the sleep neural circuit remains unclear (Jin, 2021).
Drosophila melanogaster is sensitive to temperature, and several thermosensory molecules with different temperature sensitivities have been identified, including TRPA1, Gr28b, Pyrexia, and Painless. Moreover, internal and external thermosensory neurons have also been identified in the brain and in the antenna. These thermosensory pathways enable Drosophila to adapt behavioral responses to attractive or noxious temperatures. Drosophila also exhibits sleep-like behavior and have been used to dissect the neural mechanism that regulates sleep as well as the neurobiological function of sleep. Dorsal neuron 1 (DN1) neurons include several subsets with distinct anatomical projection patterns that play distinct roles in waking and sleep behaviors. Recent studies have shown that DN1 neurons receive environmental temperature inputs. Thus, DN1 neurons might be a critical site to integrate various thermosensory inputs into the sleep neural circuit (Jin, 2021).
This study shows that thermal-sensing anterior cell (AC) neurons monitor ambient temperature shifting and release acetylcholine (ACh) to promote wakefulness. This study demonstrates that the AC-DN1p-pars intercerebralis (PI) neural circuit integrates thermosensory inputs to promote wakefulness. Moreover, this study identified the CNMamide (CNMa) signaling as a newly recognized wakefulness-promoting signaling (Jin, 2021).
Sleep is a reversible status characterized by quiescence and an increased arousal threshold. Thus, animals exhibit reduced alertness to risks during sleep. The increasing of ambient temperature usually occurs during climatic or seasonal change, but the excessively high temperature is fatal to animals. Increasing ambient temperature modifies sleep architecture, prolongs the latency for sleep onset, and reduces nighttime sleep. Warmth-induced wakefulness ensures that Drosophila increases their alertness to avoid fatal harm during ambient temperature shifting. It has been shown that increasing ambient temperature prolonged daytime sleep onset and increased total day sleep time and characterized the mechanism of warmth-induced morning wakefulness. This study characterized the effects of increased ambient temperature on nighttime sleep reduction and identified the circuit and molecular mechanisms of warmth-induced nighttime sleep reduction (Jin, 2021).
Internal thermosensory AC neurons, expressing TRPA1 channels, have been shown to set preferred temperature before dawn and to regulate prolonged morning wakefulness. Moreover, loss of TRPA1 function affects siesta behavior, but not synchronization to temperature cycles. In addition, increasing ambient temperature with different protocols may induce different effects on the flies. Consistent with a the previous report, this study showed that both the strong loss-of-function mutant and depletion of TRPA1 in ACs exhibited the suppression in warmth-induced wakefulness (Jin, 2021).
Although TrpA1ins-GAL4 is able to label TRPA1-expressing AC neurons, it also labels some other neurons that project to the same brain region as AC neurons. This study identified the NP5130-GAL4 driver that labels AC neurons and a pair of neurons in SOG. Previous GRASP analysis has shown that AC neurons form synaptic connections with DN1ps by using TrpA1ins-GAL4 and Clk4.1M-lexA lines. However, blocking the synaptic transmission of Clk4.1M+ DN1ps failed to suppress warmth-induced wakefulness. GRASP analysis narrowed down the neurons that synaptically contact with AC neurons by using NP5130-GAL4 and R18H11-lexA lines. Supportive to this connectivity, calcium-imaging studies and genetic and behavioral analysis further support that ACs make synaptic contact with a subset of DN1ps and promote wakefulness via ACh signaling. Consistent with the previous studies, this study reveals that R18H11+, but not Clk4.1M+ or R18H11+/Clk4.1M−, DN1ps promote warmth-induced wakefulness. These results suggest that R18H11+/Clk4.1M+ DN1ps are essential for warmth-induced wakefulness. The potential roles of R18H11−/Clk4.1M+ DN1 neurons in warmth-induced wakefulness need to be further investigated (Jin, 2021).
DN1 neurons have been shown to integrate circadian, light, and temperature inputs to influence locomotor rhythms. Consistent with previous reports, consecutive calcium imaging reveals that acute heating rapidly inhibits CNMa+ DN1ps, whereas prolonged heating slowly increases Ca2+ level in CNMa+ DN1ps. These observations suggest that DN1ps receive multiple thermosensory inputs through different pathways. This study further reveals that CNMa signaling promotes warmth-induced wakefulness and demonstrates that DN1ps promote wakefulness via releasing the CNMa neuropeptide to inhibit Dh44+ PI neurons. Moreover, this study reveals that CNMa is involved in daytime sleep regulation and DN1 neurons also receive light inputs to influence locomotor rhythms. The mechanism of CNMa that regulates daytime sleep needs to be further investigated (Jin, 2021).
To sum up, the AC-DN1p-PI neural circuit integrates thermosensory inputs to promote wakefulness. These newly identified circuits initiate a further road map to understand integrative mechanisms at the circuit level (Jin, 2021).
Recent studies have shown that alternative splicing (AS) plays an important role in regulating circadian rhythm. However, it is not clear whether clock neuron-specific AS is circadian rhythm dependent and what genetic and environmental factors mediate the circadian control of AS. By genome-wide RNA sequencing, SRP54 was identified is one of the Clock (Clk) dependent alternative splicing factors. Genetic interaction between Clock and SRP54 alleles showed that the enhancement of the circadian phenotype increased with temperature, being strongest at 29 °C and weakest at 18 °C. The alternative splicing and differential gene expression profile of Clock and SRP54 overlapped with the circadian-related gene profiles identified in various genome-wide studies, indicating that SRP54 is involved in circadian rhythm. By analyzing of the RNA-seq results at different temperatures, it was found that the roles of Clock and SRP54 are temperature dependent. Multiple novel temperature-dependent transcripts not documented in current databases were also found (Li, 2022).
Ambient temperature varies constantly. However, the period of circadian pacemakers is remarkably stable over a wide-range of ecologically- and physiologically-relevant temperatures, even though the kinetics of most biochemical reactions accelerates as temperature rises. This thermal buffering phenomenon, called temperature compensation, is a critical feature of circadian rhythms, but how it is achieved remains elusive. This study uncovered the important role played by the Drosophila PERIOD (PER) phosphodegron in temperature compensation. This phosphorylation hotspot is crucial for PER proteasomal degradation and is the functional homolog of mammalian PER2 S478 phosphodegron, which also impacts temperature compensation. Using CRISPR-Cas9, a series of mutations was introduced that altered three Serines of the PER phosphodegron. While all three Serine to Alanine substitutions lengthened period at all temperatures tested, temperature compensation was differentially affected. S44A and S45A substitutions caused undercompensation, while S47A resulted in overcompensation. These results thus reveal unexpected functional heterogeneity of phosphodegron residues in thermal compensation. Furthermore, mutations impairing phosphorylation of the pers phosphocluster showed undercompensation, consistent with its inhibitory role on S47 phosphorylation. It was observed that S47A substitution caused increased accumulation of hyper-phosphorylated PER at warmer temperatures. This finding was corroborated by cell culture assays in which S47A slowed down phosphorylation-dependent PER degradation at high temperatures, causing PER degradation to be excessively temperature-compensated. Thus, these results point to a novel role of the PER phosphodegron in temperature compensation through temperature-dependent modulation of the abundance of hyper-phosphorylated PER. This work reveals interesting mechanistic convergences and differences between mammalian and Drosophila temperature compensation of the circadian clock (Joshi, 2022).
Despite the fact that sleep deprivation substantially affects the way animals regulate their body temperature, the specific mechanisms behind this phenomenon are not well understood. In both mammals and flies, neural circuits regulating sleep and thermoregulation overlap, suggesting an interdependence that may be relevant for sleep function. To investigate this relationship further, flies were exposed to 12 h of sleep deprivation, or 48 h of sleep fragmentation, and temperature preference was evaluated in a thermal gradient. Flies exposed to 12 h of sleep deprivation chose warmer temperatures after sleep deprivation. Importantly, sleep fragmentation, which prevents flies from entering deeper stages of sleep, but does not activate sleep homeostatic mechanisms nor induce impairments in short-term memory also resulted in flies choosing warmer temperatures. To identify the underlying neuronal circuits, RNAi was used to knock down the receptor for Pigment dispersing factor, a peptide that influences circadian rhythms, temperature preference and sleep. Expressing UAS-Pdfr(RNAi) in subsets of clock neurons prevented sleep fragmentation from increasing temperature preference. Finally, temperature preference was evaluated after flies had undergone a social jet lag protocol which is known to disrupt clock neurons. In this protocol, flies experience a 3 h light phase delay on Friday followed by a 3 h light advance on Sunday evening. Flies exposed to social jet lag exhibited an increase in temperature preference which persisted for several days. These findings identify specific clock neurons that are modulated by sleep disruption to increase temperature preference. Moreover, the data indicate that temperature preference may be a more sensitive indicator of sleep disruption than learning and memory (Roach, 2023).
Organisms adapt to seasonal changes in photoperiod and temperature to survive; however, the mechanisms by which these signals are integrated in the brain to alter seasonal biology are poorly understood. It has been reported previously that EYES ABSENT (EYA) shows higher levels in cold temperature or short photoperiod and promotes winter physiology in Drosophila. Nevertheless, how EYA senses seasonal cues is unclear. Pigment-dispersing factor (PDF) is a neuropeptide important for regulating circadian output rhythms. Interestingly, PDF has also been shown to regulate seasonality, suggesting that it may mediate the function of the circadian clock in modulating seasonal physiology. This study investigated the role of EYA in mediating the function of PDF on seasonal biology. It was observed that PDF abundance is lower on cold and short days as compared with warm and long days, contrary to what was previously observed for EYA. Manipulating PDF signaling in eya+ fly brain neurons, where EYA and PDF receptor are co-expressed, modulates seasonal adaptations in daily activity rhythm and ovary development via EYA-dependent and EYA-independent mechanisms. At the molecular level, altering PDF signaling impacted EYA protein abundance. Specifically, it was shown that protein kinase A (PKA), an effector of PDF signaling, phosphorylates EYA promoting its degradation, thus explaining the opposite responses of PDF and EYA abundance to changes in seasonal cues. In summary, these results support a model in which PDF signaling negatively modulates EYA levels to regulate seasonal physiology, linking the circadian clock to the modulation of seasonal adaptations (Hidalgo, 2023).
Seasonal plasticity in insects is often triggered by temperature and photoperiod changes. When climatic conditions become sub-optimal, insects might undergo reproductive diapause, a form of seasonal plasticity delaying the development of reproductive organs and activities. During the reproductive diapause, the cuticular hydrocarbon (CHC) profile, which covers the insect body surface, might also change to protect insects from desiccation and cold temperature. However, CHCs are often important cues and signals for mate recognition and changes in CHC composition might affect mate recognition. This study investigated the CHC profile composition and the mating success of Drosophila suzukii in 1- and 5-day-old males and females of summer and winter morphs. CHC compositions differed with age and morphs. However, no significant differences were found between the sexes of the same age and morph. The results of the behavioral assays show that summer morph pairs start to mate earlier in their life, have a shorter mating duration, and have more offspring compared to winter morph pairs. It is hypothesized that CHC profiles of winter morphs are adapted to survive winter conditions, potentially at the cost of reduced mate recognition cues (Karpati, 2023).
The circadian clock regulates animal physiological activities. How temperature reorganizes circadian-dependent physiological activities remains elusive. Using in-vivo two-photon imaging with the temperature control device, this study investigated the response of the Drosophila central circadian circuit to temperature variation and identified that DN1as serves as the most sensitive temperature-sensing neurons. The circadian clock gate DN1a's (for information on DN1a neurons, go to Google AI and query "DN1a neurons Drosophila") diurnal temperature response. Trans-synaptic tracing, connectome analysis, and functional imaging data reveal that DN1as bidirectionally targets two circadian neuronal subsets: activity-related E cells and sleep-promoting DN3s (see Temperature fluctuation is integrated into the DN1a-DN3 circuit to dynamically control nocturnal locomotion). Specifically, behavioral data demonstrate that the DN1a-E cell circuit modulates the evening locomotion peak in response to cold temperature, while the DN1a-DN3 circuit controls the warm temperature-induced nocturnal sleep reduction. These findings systematically and comprehensively illustrate how the central circadian circuit dynamically integrates temperature and light signals to effectively coordinate wakefulness and sleep at different times of the day, shedding light on the conserved neural mechanisms underlying temperature-regulated circadian physiology in animals (Li, 2024).
In nature, organisms have developed a sophisticated neuronal mechanism that allows them to finely adjust their metabolism and behavior in response to fluctuations in environmental temperature, occurring either daily or seasonally. In order to unravel the intricate encoding and regulation of temperature-dependent physiological activity changes in the central brain, a highly precise temperature-controlled Peltier device was devised that was seamlessly integrated with an in vivo two-photon calcium imaging system. This innovative setup enabled capturing and analysis of the intricate responses of the circadian circuit to temperature variations with greater precision. Through this comprehensive investigations, a groundbreaking discovery was made: the existence of a distinct circuit encompassing DN1as, LNds (Dorsal Lateral Neurons), and DN3s, which acts as an integrator of both circadian and temperature information, ultimately facilitating the adaptation of fly behaviors to ambient temperature fluctuations. Notably, the pivotal role of DN1a in regulating the activity of the locomotor-promoting LNds during the evening was unveiled, as well as the sleep-promoting DN3s during the night. This exquisite regulation ensures the promotion of locomotion during cold periods and the suppression of sleep during hot nights, thereby orchestrating an optimal behavioral response to temperature changes (Li, 2024).
This comprehensive study extends previous findings on thermal response features in Drosophila central circadian neurons, as it was found that DN1as respond to both increases and decreases in temperature in a circadian rhythm-dependent manner. This observation aligns with the connectome of DN1as, as their upstream neurons comprise not only TPN-IIs that convey low-temperature information but also TPN-IVs (for information on TPN-IV neurons, go to In these observations, a response to temperature changes was evident in a subset of DN1 and DN2 (for information on DM2 neurons, go to The DN1a-DN3 circuit's role in temperature-dependent sleep regulation is a crucial area of research. DN3s, an unidentified subpopulation representing approximately 35%-40% of circadian neurons, are involved in sleep regulation. Using DN3-specific Split-GAL4 lines, it was discovered that DN3s (SMP232), located downstream of DN1a, regulate temperature-dependent nocturnal sleep. Interestingly, DN3s respond to both light and high temperature, aligning with their coupling during the day. In contrast, darkness and low temperature at night release DN3s from inhibition and gradually increase their activity, consistent with previous findings showing DN3 activity peaking early in the night. DN3s are implicated in regulating the offset of the E peak and initiating nighttime sleep2. Additionally, DN3-secreted AstC inhibits LNds through AstC-R2, modulating the timing of evening locomotor activity. Different DN3 subpopulations exhibit varying responsiveness to temperature, potentially due to the expression of specific receptors. DN3s downstream of DN1as rely on the glutamate-GluCl pathway, while DN3s downstream of other temperature-sensing neurons may be activated by acetylcholine or NMDAR receptors. Ablation of specific DN3 groups resulted in a delayed E peak and reduced nighttime sleep, similar to high temperature conditions. These findings demonstrate that DN3s integrate temperature, light, and circadian information to regulate physiological behaviors, such as locomotion and sleep, in fruit flies (Li, 2024).
While this study has identified the DN1a-LNd and DN1a-DN3 circuits that regulate physiological activities cycles in response to temperature changes in flies, the intricate molecular mechanisms through which circadian proteins exert their influence on the temperature response of circadian neurons has not been studied. It is proposed that circadian proteins may exert their regulatory effects through multiple pathways. One plausible mechanism involves the regulation of transcription in downstream genes, such as GABA and acetylcholine receptors, to potentially enhance the sensitivity of DN1a to temperature cues mediated by GABAergic or cholinergic TPNs during nocturnal periods. Alternatively, the circadian clock may modulate diurnal variations in neuronal membrane excitability and firing patterns, as well as internal endoplasmic reticulum calcium stores, thereby influencing the diurnal calcium level and temperature response of DN1as. These data show that knockdown of SERCA, the sarco/endoplasmic reticulum calcium pump, not only reduces the amplitude of calcium rhythms in DN1a, but also enhances its diurnal responsiveness to temperature changes, Nevertheless, further investigations are warranted to elucidate the precise mechanisms by which the circadian clock modulates SERCA expression levels and endoplasmic reticulum calcium levels, and to elucidate their role in regulating circadian differences in the temperature response of circadian neurons (Li, 2024).
Most cyclic biological processes are under control of
a circadian molecular timing system that synchronizes these phenomena
to the 24-h day. One generic property of circadian-controlled
processes is that they operate within a specific temperature range,
below which the manifestation of rhythm ceases. Little is known about
the evolutionary relevance of the lower temperature limit of
rhythmicity or about the mechanism underlying the loss of overt
circadian behavior below this lower limit, especially in one model
organism of chronobiology, Drosophila melanogaster. Natural
populations of Drosophila are evolving under divergent selection
pressures and so provide a source of diversity necessary to address
these issues. Using lines derived from African populations, this study
found that there is natural variation in the expression of rhythmic
behavior under low-temperature conditions. Evidence was found that
this variability is evolutionarily relevant at extremely low
temperature (12 degrees C) because high-altitude populations exhibit
selection for locally adapted genomes that contribute to rhythmic
behavior. Lines resistant to 15 degrees C show an additional layer of
diversity in their response to temperature extremes because some lines
are resistant to low temperature (15 degrees C) only, whereas others
are cross-resistant to high and low temperature (15 degrees C and 30
degrees C). Genetic analysis of one cold-resistant circadian line at
15 degrees C reveals that the phenotype maps to the X-chromosome but
not to the core clock genes, per and sgg. Analysis of
the central clock cells of this line reveals that maintenance of
rhythm is associated with robust clock function, which is compromised
in a standard laboratory strain. These data indicate that the
cold-resistant circadian phenotype is clock based. This study
highlights the importance of using natural populations to inform us of
the basic features of circadian traits, especially those that might be
under temperature-based selection (Maguire, 2014). Circadian clocks are endogenous timers adjusting behaviour and
physiology with the solar day. Synchronized circadian clocks improve
fitness and are crucial for physical and mental well-being. Visual and
non-visual photoreceptors are responsible for synchronizing circadian
clocks to light, but clock-resetting is also achieved by alternating
day and night temperatures with only 2-4oC difference. This
temperature sensitivity is remarkable considering that the circadian
clock period (~24 h) is largely independent of surrounding ambient
temperatures. This study shows that Drosophila Ionotropic Receptor 25a
(IR25a) is required for behavioural synchronization to low-amplitude
temperature cycles. This channel is expressed in sensory neurons of
internal stretch receptors previously implicated in temperature
synchronization of the circadian clock. IR25a is required for
temperature-synchronized clock protein oscillations in subsets of
central clock neurons. Extracellular leg nerve recordings reveal
temperature- and IR25a-dependent sensory responses, and IR25a
misexpression confers temperature-dependent firing of heterologous
neurons. It is proposed that IR25a is part of an input pathway to the
circadian clock that detects small temperature differences. This
pathway operates in the absence of known 'hot' and 'cold' sensors in
the Drosophila antenna, revealing the existence of novel
periphery-to-brain temperature signalling channels (Chen, 2015). In Drosophila, daily activity rhythms are controlled by a network of
~150 clock neurons expressing the clock genes period (per) and
timeless (tim). These encode repressor proteins that negatively
feedback on their own promoters resulting in 24 h oscillations of
clock molecules. Temperature cycles (TC) synchronize molecular clocks
present in peripheral appendages in a tissue-autonomous manner,
whereas synchronization of clock neurons in the brain mainly depends
on peripheral temperature receptors located in the chordotonal organs
(ChO) and the ChO-expressed gene nocte (Chen, 2015). To discover novel factors involved in temperature entrainment, this
study identified NOCTE-interacting proteins by co-immunoprecipitation
and mass-spectrometry. Focus was placed on IR25a, a member of a
divergent subfamily of ionotropic glutamate receptors, and the
interaction was verified by co-immunoprecipitation after
overexpressing IR25a and NOCTE in all clock cells using tim-gal4
. IR25a is expressed in different populations of sensory neurons,
including those in the antenna and labellum. In the olfactory system
IR25a acts as a co-receptor with different odour-sensing IRs (Chen,
2015). To investigate if IR25a is co-expressed with nocte in
ChO, IR25a expression was analyzed in femur and antennal ChO
using an IR25a-gal4 line (Chen, 2015). IR25a-gal4-driven mCD8-GFP labelled subsets of ChO
neurons in the femur, overlapped substantially with nompC-QF
driven QUAS-Tomato signals (using the QF binary
transcriptional activation system). nompC-QF is expressed in
larval ChO18 and in the adult femur ChO. Comparison of IR25a-driven
mCD8-GFP and nuclear DsRed signals with those of other
ChO neuron drivers (F-gal4 and nocte-gal4 suggests that
IR25a is expressed in a subset of femur ChO neurons and
Johnston's Organ (JO) neurons. To determine if IR25a-gal4 ChO
signals reflect endogenous IR25a expression, the presence of IR25a
mRNA was confirmed in the femur and leg and the co-localization of
anti-IR25a immunofluorescence signals in femur ChO neurons. IR25a was
detected in ChO neuron cell bodies and ciliated dendrites, as was an
mCherry-IR25a fusion protein expressed in these cells (Chen, 2015). As nocte mutants do not synchronize to temperature cycles in
constant light (LL), IR25a-/- mutants were analyzed
under these conditions. Unlike nocte, the IR25a-/-
flies synchronized well to this regime and similar results were
obtained at warmer temperature cycles. To test whether IR25a
is specifically required for synchronization to small temperature
intervals, IR25a-/- flies were subjected to various
temperature cycles with an amplitude of only 2oC.
Surprisingly, and in contrast to wild-type, IR25a-/-
mutants did not synchronize to any of the shallow temperature cycles
in LL or constant darkness (DD). In LL, wild-type and IR25
rescue flies showed a clear activity peak in the second part of the
warm period before and after the 6 h shift of the temperature cycle.
By contrast, IR25a-/- mutants were constantly
active throughout the temperature cycle, apart from a short period of
reduced activity at the beginning of the warm phase of TC1. In DD,
control flies slowly advanced (or delayed) their evening activity peak
during phase-advanced (or delayed) temperature cycles. The phase of
this activity peak was maintained in the subsequent free-running
conditions (DD, constant 25°C) indicating stable re-entrainment of the
circadian clock. By contrast, IR25a mutants did not shift
their evening peak during the temperature cycle, keeping their
original phase throughout the experiment (Chen, 2015). To quantify entrainment in LL, the 'entrainment index' (EI) was
determined, whereas for most DD experiments the phase difference of
the main activity peak was calculated upon release into constant
conditions between IR25a mutants and controls. In all 2 Temperature receptors located in fly antennae and arista are not
required for temperature-synchronized behaviour. As expected, it was
found that antennal IR25a function is not required for
temperature entrainment. To reveal the importance of IR25a expression
in ChO neurons, tissue-specific IR25a RNA interference (RNAi)
was performed using validated transgenes. IR25a RNAi in all or
subsets of ChO neurons. By contrast, IR25a RNAi in
multidendritic, TRPA1-expressing or clock neurons did not impair
temperature entrainment. These findings are consistent with the
absence of IR25a expression in clock neurons and the brain and
show that IR25a functions in ChO neurons for temperature
entrainment to 25°C:27°C temperature cycles in LL (Chen, 2015). To identify the neural substrates underlying the lack of behavioural
synchronization,clock protein levels was quantified in wild-type, IR25a-/-,
and IR25a-/- rescue flies exposed to a shallow
temperature cycle in LL. Although TIM expression was robustly rhythmic
and synchronized in all clock neuronal groups in controls, TIM was
barely detectable in the Dorsal Neuron 1 (DN1) and DN2 of IR25a-/-
flies. Moreover, in the small and large ventral lateral neurons (s-LNv
and l-LNv), TIM expression exhibited an additional peak during the
warm phase. In the DN3, TIM declined earlier compared to controls and
there was no effect on the dorsal lateral neurons (LNd). In
temperature cycles and DD, TIM levels in DN1 were also blunted but
oscillations in the DN2 and DN3 were similar to controls. In contrast
to LL, TIM did not oscillate in any of the LN groups and was at
constantly low levels. The alterations of TIM expression are
temperature specific, as normal oscillations were observed in LD
cycles at 25°C. An increase of the temperature cycle amplitude to 4°C
also restored normal TIM expression in IR25a-/-
flies, in agreement with the behavioural rescue. In summary, in
low-amplitude temperature cycles, IR25a is required for normally
synchronized TIM oscillations in DN1-3 and LNv in LL and in DN1 and LN
clock neurons in DD (Chen, 2015). Tests were performed to see if the clock neurons affected by the lack
of IR25a are indeed involved in regulating behavioural synchronization
to shallow temperature cycles by blocking synaptic transmission using
tetanus-toxin (TNT). Indeed, TNT-expression in DN1 and DN2 blocked
synchronization in LL, whereas in DD only DN1 blockage interfered with
temperature entrainment. Consistent with the differential effect on
TIM oscillations in LL and DD these results strongly suggest that
IR25a is required for the synchronized output of the DN1 (LL and DD)
and DN2 (LL) to control temperature-entrained behaviour (Chen, 2015).
Next, it was asked if ChO might directly sense temperature in an
IR25a-dependent manner. Leg nerve activity was measured in restrained
preparations and ChO units were identified in the compound signal. In
both wild-type and -/- flies, spontaneous leg movement
changed as a function of temperature along with motor and sensory
activity. Additionally, presumed ChO activity of wild-type flies also
increased during periods without movement. This temperature-induced
but movement-independent, ChO activity was absent in -/-
flies, showing that temperature is sensed in the legs in an
IR25a-dependent manner. To test if IR25a contributes directly to
temperature-sensing, this channel was expressed in the physiologically
well-characterized, IR25a-negative, l-LNv. As a positive control, the
temperature-sensitive Drosophila TRPA1 channel was also expressed in
the l-LNv. Isolated brains were exposed to a temperature ramp, and
spike frequency of individual l-LNv was recorded. Control l-LNv did
not show a significant temperature-dependent change in neural
activity. As expected, the firing rate of TRPA1 expressing neurons
drastically increased linearly with temperature, as did other cellular
parameters. IR25a expression resulted in a linear and
reversible temperature-dependent increase in action potential firing
frequency, whereas other cellular parameters showed no difference.
Increasing the temperature by only 2-3°C also lead to a reversible
increase in firing frequency in IR25a expressing l-LNv. By
contrast, expression of the related, but olfactory-specific
co-receptor IR8a (which is not required for temperature entrainment)
did not confer temperature-sensitivity. These observations suggest
that IR25a is at least part of a thermosensory receptor required for
temperature entrainment (Chen, 2015). These data indicating that IR25a contributes to
temperature sensing within ChO extend the roles of IR's beyond
chemoreception, reminiscent of the requirement for the 'gustatory
receptor' Gr28b in warmth-avoidance. Although this study shows that
IR25a-expressing leg neurons are capable of sensing temperature and
mediating temperature entrainment, it is possible that this receptor
has a similar role elsewhere in the peripheral nervous system. IR25a
responds to small temperature changes and it is proposed that the fly
continuously integrates temperature signals received from multiple ChO
across the whole body for synchronization of the clock. This potential
reliance on weakly responding temperature receptors might explain why
the Drosophila circadian clock is insensitive to brief temperature
pulses, which could help maintain synchronized clock function in
natural conditions of rapid and large temperature fluctuations (Chen,
2015).
Harbison, S. T., Serrano Negron, Y. L., Hansen, N. F. and Lobell, A. S. (2017). Selection for long and short sleep duration in Drosophila melanogaster reveals the complex genetic network underlying natural variation in sleep. PLoS Genet 13(12): e1007098. PubMed ID: 29240764
Why do some individuals need more sleep than others? Forward mutagenesis screens in flies using engineered mutations have established a clear genetic component to sleep duration, revealing mutants that convey very long or short sleep. Whether such extreme long or short sleep could exist in natural populations was unknown. This study applied artificial selection for high and low night sleep duration to an outbred population of Drosophila melanogaster for 13 generations. Night sleep duration diverged by 9.97 hours in the long and short sleeper populations, and 24-hour sleep was reduced to 3.3 hours in the short sleepers. Whole genome sequence data revealed several hundred thousand changes in allele frequencies at polymorphic loci across the genome. Combining the data from long and short sleeper populations across generations in a logistic regression implicated 126 polymorphisms in 80 candidate genes, and three of these genes and a larger genomic region were confirmed with mutant and chromosomal deficiency tests, respectively. Many of these genes could be connected in a single network based on previously known physical and genetic interactions. Candidate genes have known roles in several classic, highly conserved developmental and signaling pathways-EGFR, Wnt, Hippo, and MAPK. The involvement of highly pleiotropic pathway genes suggests that sleep duration in natural populations can be influenced by a wide variety of biological processes, which may be why the purpose of sleep has been so elusive (Harbison, 2017).
Circadian clocks have evolved to synchronize
physiology, metabolism and behaviour to the 24-h geophysical cycles of
the Earth. Drosophila melanogaster's rhythmic locomotor behaviour
provides the main phenotype for the identification of higher
eukaryotic clock genes. Under laboratory light-dark cycles, flies show
enhanced activity before lights on and off signals, and these
anticipatory responses have defined the neuronal sites of the
corresponding morning (M) and evening (E) oscillators. However, the
natural environment provides much richer cycling environmental stimuli
than the laboratory, so this study sought to examine fly locomotor
rhythms in the wild. Several key laboratory-based assumptions about
circadian behaviour are not supported by natural observations. These
include the anticipation of light transitions, the midday 'siesta',
the fly's crepuscular activity, its nocturnal behaviour under
moonlight, and the dominance of light stimuli over temperature. Also,
a third major locomotor component in addition to M and E, which was
termed 'A' (afternoon). Furthermore, it was shown that these natural
rhythm phenotypes can be observed in the laboratory by using realistic
temperature and light cycle simulations. The results suggest that a
comprehensive re-examination of circadian behaviour and its molecular
readouts under simulated natural conditions will provide a more
authentic interpretation of the adaptive significance of this
important rhythmic phenotype. Such studies should also help to clarify
the underlying molecular and neuroanatomical substrates of the clock
under natural protocols (Vanin, 2012). Sleep is required to maintain physiological functions, including
memory, and is regulated by monoamines across species. Enhancement of
dopamine signals by a mutation in the dopamine transporter (DAT)
decreases sleep, but the underlying dopamine circuit responsible for
this remains unknown. This study found that the D1 dopamine receptor
(DA1, also known as DopR) in the dorsal fan-shaped body (dFSB)
mediates the arousal effect of dopamine in Drosophila. The short sleep
phenotype of the DAT mutant was completely rescued by an additional
mutation in the DA1 gene, but expression of wild-type DA1 in the dFSB
restored the short sleep phenotype. Anatomical and physiological
connections were found between dopamine neurons and the dFSB neuron.
Finally, mosaic analysis with a repressive marker found that a single
dopamine neuron projecting to the FSB activates arousal. These results
suggest that a local dopamine pathway regulates sleep (Ueno, 2012). Neurons in the dFSB are involved in dopaminergic sleep regulation in
Drosophila an the PPM3-FSB dopamine pathway, which is distinct
from that required for memory formation, regulates arousal. A previous
study found that the rescue of DA1 mutants outside of the
mushroom body using a pan-neuronal GAL4 driver, elav-GAL4,
coupled with the mushroom body suppressor MB-GAL80 in DA1dumb1
mutants can recover methamphetamine sensitivity. This suggests that
dopamine regulates arousal outside of the mushroom body. Previous
findings have shown that DA1 in the PDF neurons (lateral ventral
neurons) regulates sleep-wake arousal and that DA1 in the ellipsoid
body regulates stress-induced arousal (Ueno, 2012). A previous study identified PDF neurons that mediate the buffering
effects of light on dopamine-induced arousal. However, the current
ablation experiments showed that dopamine can elicit strong arousal
effects without PDF neurons. The previous report used heterozygous DA1
mutant flies in a wild-type background for most experiments. However,
this study used homozygous (null) DA1 mutants, as we found
that the heterozygous DA1 mutants crossed with DATfmn
showed an almost equivalent short sleep phenotype. Thus, one possible
explanation for the difference between the previous study and this one
is that heterozygous expression of DA1 in the FSB is
sufficient to elicit the arousal effects of dopamine, which is also
regulated partly through PDF neurons. Given that DA1
expression in the dFSB neurons alone is sufficient for the majority of
the wake-promoting effects of dopamine, the arousal regulating
dopamine pathway appears to converge at the FSB (Ueno, 2012). Activation of the dopamine neurons required for aversive memory
formation had little effect on sleep. On the other hand, TrpA1
stimulation in a single FSB-projecting PPM3 dopamine neuron was able
to reduce sleep. As the magnitude of the decrease in sleep by the
activation of the single PPM3 neuron was smaller than that of most
dopamine neurons with TH-GAL4, it is possible that other
dopamine neurons, which were not labeled in MARCM screening, also
affect arousal. For example, in the PPL1 cluster, at least five types
of projections to the mushroom body and FSB-projecting neurons have
been described (Ueno, 2012). However, in a MARCM experiment, many of the labeled neurons in the
PPL1 cluster were seen to project to the alpha lobe of the mushroom
body; it is possible that the contribution of other PPL1 neurons were
not fully determine. Alternatively, combinatorial activation of the
FSB-projecting PPM3 neurons and other dopamine neurons may have a
synergistic effect. Further MARCM experiments using various clone
induction protocols may help to formulate a more comprehensive
characterization of single dopamine neurons. In addition, it is also
possible that dopamine neurons that are not labeled by TH-GAL4
also have an arousal effect. It was noticed that TH-GAL4induced
GFP expression and tyrosine hydroxylase staining do not overlap
completely, and some clusters, such as the PAM cluster, are covered
only partially by TH-Gal4 (Ueno, 2012). The association between sleep and memory in Drosophila has
recently been described in various reports. Other short sleep flies,
hyperkinetic and calcineurin knockdown, also suffer from impaired
memory. Sleep deprivation has been shown to impair aversive olfactory
memory and learned phototaxis suppression. Conversely, memory
formation increased sleep duration in normal flies, and this has not
been observed in learning-deficient mutants. In the Drosophila
brain, the expression of synaptic component proteins decreased during
sleep and increased during waking, suggesting that sleep is required
for the maintenance of synapses. In contrast, the current data imply a
functional dissociation between sleep and memory circuits by different
dopamine neurons. These results will provide clues to uncover a
possible physiological relationship between them, for example, by the
activation of only one to examine the causal relationship (Ueno,
2012). The FSB has been reported to have a role in visual
memory processing, but its involvement in other behaviors is not yet
fully understood. A recent report showed that activation of dFSB
neurons induces sleep. This suggests that the dopamine signal
regulates sleep via control of the neural properties of the dFSB
neurons through DA1. In mammals, dopamine signaling via the D1-type
dopamine receptor is thought to increase firing probability. However,
dopamine signaling via the D1-like receptor DA1 in flies results in
the inhibition of neural activity. Although the previous report
suggested that the inhibition is independent of protein kinase A,
activation of adenylyl cyclase with Gs* in
dFSB decreased sleep levels. Taken together, these findings indicate
that DA1 activation in the dFSB inhibits neural activity and results
in the promotion of wakefulness. It has been reported that sleep
induction through the activation of the dFSB neurons promotes memory
consolidation after courtship conditioning. However, this study found
that DA1 expression in the FSB itself had little effect on aversive
olfactory memory. The contradiction between these findings might be a
result of the difference in the memory tasks. It is also possible that
functional interaction between sleep and memory is implemented
downstream of the FSB. Further studies are required to elucidate the
causal relationship between sleep and memory (Ueno, 2012). To advance the understanding of sleep regulation, a
screen was performed in Drosophila for sleep-promoting cells, and
neurons expressing neuropeptide Y-like short
neuropeptide F (sNPF) were identifed. Sleep induction by sNPF
meets all relevant criteria. Rebound sleep following sleep deprivation
is reduced by activation of sNPF neurons, and flies experience negative
sleep rebound upon cessation of sNPF neuronal stimulation, indicating
that sNPF provides an important signal to the sleep homeostat. Only a
subset of sNPF-expressing neurons, which includes the small
ventrolateral clock neurons, is sleep promoting. Their release of sNPF
increases sleep consolidation in part by suppressing the activity of
wake-promoting large ventrolateral clock neurons, and suppression of
neuronal firing may be the general response to sNPF receptor activation.
sNPF acutely increases sleep without altering feeding behavior, which it
affects only on a much longer time scale. The profound effect of sNPF on
sleep indicates that it is an important sleep-promoting molecule (Shang,
2013). This study presents several independent lines of evidence indicating
that sNPF acutely increases sleep and alters sleep homeostasis. This is
because release of animals from sNPF neuron activation after several
days of hypersomnolence resulted in a transient decrease in sleep or
negative sleep rebound. Moreover, activation of sNPF neurons during
mechanical sleep deprivation blunted the rebound sleep following the
deprivation. This suggests that sNPF might alter the internal perception
of sleep state during the deprivation despite an apparently behaviorally
awake state. It also suggests that sNPF might directly modulate the
sleep homeostat (Shang, 2013). The most potent in vivo manipulations of sNPF function, mutation of
the sNPF gene and strong activation of sNPF neurons with dTRPA1, affect
daytime as well as nighttime sleep levels. These manipulations also
strongly alter sleep bout duration, a measure of consolidation, in the
opposite direction to the sleep duration effects. More limited
manipulations of sNPF signaling (cell-specific downregulation of sNPF
levels or of sNPF signaling) indicate that sNPF is most important for
promoting sleep at night. It also affects the structure of daytime
sleep, a function of sNPF circuitry normally suppressed during the day
by wake-promoting GABAergic neurons, acting via GABAA receptors.
Suppression of excitability with Kir2.1 likely mimics this daytime
GABAergic function. These results in aggregate suggest that sNPF action
differs depending on the time of day, a result that supports the idea
that daytime and nighttime sleep may be regulated by different
circuitries (Shang, 2013). The role of sNPF in promoting more consolidated sleep is consistent
with a general antiarousal function. As in mammals, Drosophila arousal
can be measured electrophysiologically, but the most straightforward
measure of arousal state is behavioral, and sleep fragmentation is
indicative of a less stable, more easily aroused state. The main
neurochemical previously implicated in fly arousal is DA, and l-LNvs
play a prominent role in the arousal circuitry (Shang, 2013). Although the imaging assays indicated that sNPF alone did not lead to
significant cAMP changes in the l-LNvs, it mildly suppressed the
activation effect of DA on the l-LNvs. As one subset of clock neurons in
the sleep circuit releases sNPF and promotes sleep at night and an
adjacent subset responds to sNPF and suppresses nighttime sleep, sNPF
may be used by the s-LNv-to-l-LNv pathway to coordinate the timing of
sleep with other circadian behaviors. Indeed, sNPF mRNA is a potent
cycling mRNA in s-LNvs (Kula-Eversole, 2010). Importantly, the
electrophysiological assays in larval central neurons suggest that
inhibition of neuronal firing may be a general feature of sNPF function
and relevant to other sleep centers in addition to the clock neurons
(Shang, 2013). sNPF and other sleep-relevant neuromodulators like DA are likely to
act at multiple sites in the brain given the major state change effected
by the sleep/wake transition. This expectation also reflects the modest
effects of sNPFR manipulation within l-LNvs on total sleep time.
Moreover, fan-shaped body neurons have recently been shown to be
important for DA-mediated arousal (Liu, 2012; Ueno, 2012). The ability
of these neuromodulators to act on many circuits may allow for more
flexible integration of sleep with other behaviors and with other
external and internal factors (Shang, 2013) An important influence on sleep is metabolic state. Indeed, sNPF
facilitated the OA-to-DILP circuit, which may reflect its role in
sleep/wake, feeding and/or metabolic regulation. However, the
wake-promoting effect of activating the DILP pathway is
context-dependent, occurring only in LD. Moreover, acute activation of
octopaminergic neurons by dTRPA1 only mildly affects sleep and also in a
condition-dependent manner, and feeding animals with octopamine only
significantly suppresses total sleep after 2-3 days of exposure.
Although long-term activation of octopaminergic neurons leads to
long-lasting increases in food dwelling, these effects contrast sharply
with the rapid and condition-independent effects seen with acute
increases in dopamine signaling (Shang, 2011). Dopaminergic neurons have
also been shown to be a critical part of NPF-regulated changes in
satiety and response to food, and activation of these neurons indeed led
to an immediate onset of food dwelling, which reversed rapidly upon
dTRPA1 inactivation. As expected, tracker analysis shows that these
food-dwelling flies also sleep very little, indicating that dopamine
affects both sleep and feeding rapidly. These effects contrast with the
slow effects on food dwelling by sNPF neuronal activation (Shang, 2013)
The simplest interpretation of this slow food-dwelling response is
that it is secondary to a more primary effect of sNPF on sleep. Indeed,
a slow buildup in hunger or even starvation as a consequence of too much
sleep is a simple explanation consistent with most if not all of these
data. Behavioral effects as a secondary consequence of some other more
direct effect is also an interpretation of many of the sleep effects of
activation of peptidergic neurons seen in this study, in which only sNPF
robustly increased sleep, i.e., under both LD and DD conditions. It is
therefore suggested that a necessary condition for serious consideration
of a molecule as behavior-relevant is a rapid response, which is also
relatively condition independent. Dopamine as a wake-promoting molecule
and now sNPF as a sleep-promoting molecule meet these criteria (Shang,
2013).
This study reports a new protein involved in the
homeostatic regulation of sleep in Drosophila. A forward genetic
screen was conducted of chemically mutagenized flies to identify
short-sleeping mutants, and one, redeye
(rye; nicotinic Acetylcholine Receptor α4), was found that shows a
severe reduction of sleep length. Cloning of rye reveals that it
encodes a nicotinic acetylcholine receptor alpha subunit required for
Drosophila sleep. Levels of RYE oscillate in light-dark cycles and peak
at times of daily sleep. Cycling of RYE is independent of a functional
circadian clock, but rather depends upon the sleep homeostat, as protein
levels are up-regulated in short-sleeping mutants and also in wild type
animals following sleep deprivation. It is proposed that the homeostatic
drive to sleep increases levels of RYE, which responds to this drive by
promoting sleep (Shi, 2014). The molecular mechanism of sleep homeostasis is a mystery and a subject
of intense research in the sleep field. In addition to the investigation
of mechanisms underlying sleep drive, considerable effort is being put
into identifying biomarkers of sleep need. Based on what is known about
the so-called sleep homeostat, which consists of increasing sleep
pressure during wakefulness and dissipation of such pressure following
sleep, it is suggested that a component or direct output of the
homeostat should satisfy three criteria: (1) the gene product should
regulate the sleep:wake cycle (i.e., genetic alleles of this gene should
have some sleep phenotype); (2) expression levels or activity of the
gene product should go up during wakefulness or during sleep deprivation
and (3) expression levels or activity should decrease after sleep. The
function of RYE and the molecular kinetics of the RYE protein largely
satisfy these criteria. However, while RYE builds up during sleep
deprivation, it does not accumulate gradually over the wake period in a
daily cycle. Rather, it displays a marked increase close to the time of
sleep onset, suggesting that it is not a central component of the
homeostat, but responds to an upstream homeostatic signal, perhaps when
that signal reaches a certain threshold. The fact that over-expression
of RYE does not promote sleep also supports the idea that it is not the
sleep-inducing homeostatic signal. Nevertheless, RYE is not simply a
sleep output gene or sleep biomarker. It is required for implementation
of signals from the homeostat and it functions to maintain sleep. Thus,
it is proposed that rye is a sleep-regulating gene immediately
downstream of the homeostat (Shi, 2014). It is suggested that RYE represents a molecular correlate of delta
power, a characteristic of an electroencephalogram (EEG) that reflects
sleep drive. Recently, a few other molecules were reported to change
with sleep drive, but the effects were at the level of the mRNA, the
magnitude of the increase was less than we report here for RYE and loss
of the molecules did not affect baseline sleep duration. In addition,
only one is expressed cyclically, indicating that others reflect sleep
drive only under pathological conditions of sleep deprivation. RYE
levels oscillate robustly in a daily cycle, although the phase is not as
coherent as seen for circadian clock proteins. The timing of the peak
varies within a temporal range, such that there is almost always a
daytime peak and a night-time peak but not necessarily at the exact same
time. It is suggested that RYE cycles under control of the sleep
homeostat, which may not time behavior as precisely as the circadian
clock, perhaps because sleep can be influenced by many factors. The
variability in RYE cycling is particularly pronounced in short-sleeping
mutants and in the ClkJrk circadian clock
mutant, suggesting that the clock does influence RYE expression although
it is not required for its cycling in an LD cycle. Interestingly, RYE
cycles exclusively at the level of the protein, indicating translational
or post-translational mechanisms. It is worth noting that a recently
identified sleep regulator, Insomniac,
is a component of specific protein degradation pathways in the cell.
Although this study indicates that RYE cycling does not require
Insomniac, it is possible that it is regulated by other protein
turn-over machinery. Thus, translational/posttranslational regulation
appears to be part of the mechanism of sleep homeostasis (Shi, 2014). This study shows that RYE not only reflects sleep drive, but is also
required for sleep maintenance. Given that RYE is induced by sleep
deprivation and it promotes sleep, one might expect over-expression of
the protein to increase sleep. However, transgenic expression of rye
in a wild type background does not increase sleep, suggesting that while
rye is necessary, it is not sufficient for sleep onset. The
possibility cannot be excluded that RYE functions together with other
signals as part of the sleep-inducing homeostatic drive. On the other
hand, it is also possible that transgenic expression does not produce
adequate amounts of RYE protein in relevant cells. This might be the
case if RYE is tightly regulated at the level of protein stability. For
the moment, though, the parsimonious explanation noted above is
preferred, that RYE is not part of the homeostat, but immediately
downstream of it (Shi, 2014). Acetylcholine signaling has been long proposed as an arousal factor, as
the nAChR complex is a cation channel that normally promotes neuronal
activity and ACh is released during wakefulness in mammals. In contrast,
this study indicates that at least one nAChR subunit (RYE) promotes
sleep in the fly. There are more than 10 paralogs of nAChR subunits in
the fly genome. One possibility is that RYE is expressed specifically in
sleep promoting neurons, while other subunits of AChRs are in
wake-promoting cells. An increase in ACh during wakefulness may
contribute to the accumulation of sleep drive and to the increase in
RYE. Alternatively, sleep drive may increase RYE independently of ACh,
but in either scenario, RYE then promotes sleep. The precise site of RYE
action is currently not known. rye-gal4 driven GFP is
expressed widely in the brain, but it is not certain that endogenous RYE
is as widespread, as an antibody was not effective in
immunohistochemistry experiments, and GAL4 drivers are often quite
promiscuous (Shi, 2014). Sleepless
(SSS) was previously identified as a sleep promoting factor,
essential for maintaining baseline sleep and for homeostatic rebound. An
interaction between rye and sss is therefore not
surprising. What is surprising is that overexpression of SSS promotes
wakefulness in ryeT227M heterozygotes. SSS
is a GPI-anchored protein that functions as a neuronal modulator.
Previous studies indicate that SSS promotes activity of the
voltage-gated potassium channel, Shaker. This study reports that SSS
acts like a brake on nAChR (RYE) activity, as does Lynx-1, a SSS-like
molecule in mammals. Although the data shown for Drosophila
receptors used only the RYE α subunit, it is likely that SSS also
inhibits activity of other Drosophila nAChR receptors. As both
sssP1 (a null mutation) and sssP2
(a hypomorphic allele) are short-sleeping mutants, it is proposed that
the overall effect of SSS is to promote sleep. The reduced sleep in sss
mutants probably results from an increase of neuronal excitability,
through inactivation of potassium channels (Shaker), or from
hyperactivity of nAChR channels in wake-promoting neurons. Thus,
typically the sleep-inhibiting effect of SSS, mediated through RYE, is
masked by these other more dominant influences. However, in a sensitized
background (i.e., rye/+), this effect is evident. RYE promotes
sleep, and so loss of RYE results in a decrease in sleep, which is
further impacted by SSS overexpression (Shi, 2014). It is noted that there are some caveats to these data. For instance,
the ryeT227M allele could confer a
neomorphic function that accounts for the interaction with sss.
Likewise, the effects in oocytes could be non-physiological, not
necessarily reflecting what happens in the fly brain. However, given
that interactions were observed in these two very different types of
assays, and both assays indicate repression of nAchR function by SSS,
which is the effect predicted from the role of the mammalian SSS-like
protein, Lynx1, it is believed SSS does indeed regulate nAchRs such as
RYE. Interactions between SSS and nicotinic acetylcholine receptors are
also confirmed by other recent unpublished studies (Shi, 2014). It is interesting that genes identified through independent genetic
screens in Drosophila are turning out to interact with one
another. SSS and Shaker were isolated independently as sleep-regulating
genes, and subsequently shown to interact, and now it turns out that RYE
interacts with SSS. Given that each of these genes represents a
relatively infrequent hit in an unbiased screen, the interactions
suggest that genetic approaches are converging upon specific
sleep-regulating pathways. Interestingly, a recent Genome-wide
association study for sleep-altering loci in humans identified
significant effects of SNPs in an nAchR subunit as well as in a
regulatory subunit of Shaker, suggesting that these mechanisms are also
conserved across species (Shi, 2014).
How the circadian clock regulates the timing of sleep is poorly
understood. This study identifies a Drosophila mutant, wide awake
(wake), that exhibits a marked delay in sleep onset at dusk.
Loss of Wake in a set of arousal-promoting clock neurons, the large
ventrolateral neurons (l-LNvs), impairs sleep onset. Wake levels
cycle, peaking near dusk, and the expression of Wake in l-LNvs is
Clock dependent. Strikingly, Clock and cycle mutants
also exhibit a profound delay in sleep onset, which can be rescued by
restoring Wake expression in LNvs. Wake interacts with the GABAA
receptor Resistant to Dieldrin (Rdl), upregulating its levels and
promoting its localization to the plasma membrane. In wake
mutant l-LNvs, GABA sensitivity is decreased and excitability is
increased at dusk. It is proposed that Wake acts as a clock output
molecule specifically for sleep, inhibiting LNvs at dusk to promote
the transition from wake to sleep (Liu, 2014). The molecular pathways by which the circadian clock modulates the
timing of sleep are unknown. This study identified a molecule, Wide
Awake, that promotes sleep and is required for circadian timing of
sleep onset. The data argue for a direct role for the circadian
oscillator in regulating sleep and support a model whereby Wake acts
as a molecular intermediary between the circadian clock and sleep. In
this model, Wake transmits timing information from the circadian clock
to inhibit arousal circuits at dusk, thus facilitating the transition
from wake to sleep. wake is transcriptionally upregulated by
Clk activity, specifically in LNv clock neurons. Wake levels in l-LNvs
rise during the day and peak at the early night, near the wake/sleep
transition. This increase in Wake levels upregulates Rdl in l-LNvs,
enhancing their sensitivity to GABA signaling and serving to inhibit
the l-LNv arousal circuit. In this manner, cycling of Wake promotes
cycling of the excitability of l-LNv cells. In wake mutants,
l-LNvs lose this circadian electrical cycling; the higher firing rate
of these cells at dusk leads to increased release of Pdf, which would
act on Pdfr on downstream neurons to inhibit sleep onset. The identity
of the GABAergic neurons signaling to the l-LNvs is currently unknown,
but if they serve to convey information about sleep pressure from
homeostatic circuits, the l-LNvs could serve as a site of integration
for homeostatic and circadian sleep regulatory signals (Liu, 2014). Although Wake is expressed in clock neurons and its levels vary
throughout the day, Wake itself is not a core clock molecule, since
period length and activity rhythm strength are intact in wake mutants
in constant darkness. The effects of Wake on sleep latency are not
attributable to alterations in core clock function. In addition,
because locomotor rhythm strength is intact in wake mutants,
Wake is not a clock output molecule for locomotor rhythms. Rather,
Wake is the first clock output molecule shown to specifically regulate
sleep timing (Liu, 2014). Previous studies have demonstrated that Rdl in LNvs regulates sleep
in Drosophila. This work further implicates Rdl as a key factor in the
circadian modulation of sleep. In mammals, the localization and
function of GABAA receptors are regulated by a variety of cytosolic
accessory proteins, some of which are associated with the plasma
membrane and cytoskeletal elements. The data suggest that Wake acts as
an accessory protein for Rdl, upregulating its levels and promoting
its targeting to the plasma membrane. Rdl is broadly expressed
throughout the adult Drosophila brain, whereas Wake appears more
spatially restricted. It is likely that Rdl is regulated by Wake in
specific cells (e.g., Wake+ cells), while in other cells that express
Rdl but not Wake, other factors are involved. Together, these data
suggest a model in which increased GABA sensitivity is required in
specific arousal circuits to facilitate rapid and complete switching
between sleep/wake states at the appropriate circadian time (Liu,
2014). Intriguingly, the data, as well as data from the Allen
Brain Atlas, suggest that the putative mouse homolog of Wake (ANKFN1)
is enriched in the mouse SCN, the master circadian pacemaker in mammals.
Specifically, ANKFN1 is expressed in the 'core' region of the SCN, which
is analogous to the large LNvs in flies, in that it receives light input
and its molecular oscillator does not cycle or cycles weakly in DD.
These observations support a potential conservation of Wake function in
regulating clock-dependent timing of sleep onset, which will be
evaluated by ongoing genetic analysis in mice. The pronounced difficulty
of wake flies to fall asleep at lights off is reminiscent of
sleep-onset insomnia in humans. Moreover, the most widely used
medications for the treatment of insomnia are GABA agonists. Thus, the
identification of a molecule that mediates circadian timing of sleep
onset by promoting GABA signaling may lead to a deeper understanding of
mechanisms underlying insomnia and its potential therapies (Liu, 2014).
Neurons use two main schemes to encode information: rate coding (frequency of firing) and temporal coding (timing or pattern of firing). While the importance of rate coding is well established, it remains controversial whether temporal codes alone are sufficient for controlling behavior. Moreover, the molecular mechanisms underlying the generation of specific temporal codes are enigmatic. This study shows in Drosophila clock neurons that distinct temporal spike patterns, dissociated from changes in firing rate, encode time-dependent arousal and regulate sleep. From a large-scale genetic screen, this study identified the molecular pathways mediating the circadian-dependent changes in ionic flux and spike morphology that rhythmically modulate spike timing. Remarkably, the daytime spiking pattern alone is sufficient to drive plasticity in downstream arousal neurons, leading to increased firing of these cells. These findings demonstrate a causal role for temporal coding in behavior and define a form of synaptic plasticity triggered solely by temporal spike patterns (Tabuchi, 2018).
Understanding how the brain represents and processes information is a fundamental goal of neuroscience research. For the first half of the 20th century, the dominant neural coding model postulated that simple action potential (spike) counts in the relevant time window encode information about the environment or internal states (i.e., the rate-coding model). However, it has long been recognized that neural coding schemes using temporal codes (timing and/or pattern of spiking) would be computationally more powerful than traditional rate codes. In contrast to rate coding, which comprises a singular mechanism, temporal coding encompasses a diverse repertoire of coding schemes in individual or groups of neurons, ranging from latency to first spike to synchronization of oscillatory activity between spatially segregated neuronal populations (Tabuchi, 2018).
A wide variety of temporal codes have now been observed to correlate with specific external stimuli in different settings, including sensory systems, hippocampal place cells, and neocortical circuits. However, the assessment of whether and how temporal codes embody neurobiologically relevant information is complicated by multiple factors, such as the concomitant presence of changes in firing rate, integration of spatial with temporal information, and the requirement for multiple interacting brain regions in the regulation of behavior. The rigorous demonstration of a causal role for temporal codes in representing biological information requires the fulfillment of three criteria. First, the temporal code should exist under native conditions or be elicited by naturalistic stimuli. Second, as articulated in the 'reader-actuator' model, a meaningful neural code should trigger a distinct response in the downstream neural circuit. Third, the temporal code should have physiological significance and be utilized by the brain to inform behavioral choices (Tabuchi, 2018).
While many studies have identified temporal codes occurring in response to naturally occurring sensory stimuli, relatively few studies have shown that induction of specific temporal codes alter the firing of target neural circuits or affect behavior. For instance, in olfactory sensory neurons in mammals, varying the timing of firing relative to other neurons or the sniff cycle impacts firing of downstream neurons. However, other studies have found that temporal coding in olfactory, visual, and somatosensory systems had no effect on the activity of target neurons or behavioral readouts. Thus, the functional relevance for temporal coding alone to represent information about the environment and internal states remains controversial. Moreover, the molecular mechanisms that underlie the generation of different temporal codes within a neural circuit are largely unknown (Tabuchi, 2018).
This study, in the clock neuron network in Drosophila, demonstrates the presence of naturally occurring temporal spiking patterns associated with daytime versus nighttime; the cycling of these patterns was found to depend on the core clock and wide awake (wake), a recently identified clock output gene required for circadian regulation of sleep. Using optogenetic approaches in vivo, this study shows that these distinct patterns of clock neuron firing, in the absence of changes in firing rate, serve as a temporal code to signify time-dependent arousal and directly impact sleep behavior. From a large-scale forward genetic screen, this study identified the molecular mechanisms underlying the generation of these clock-dependent temporal codes. Electrophysiological and computational analyses was used to delineate the biophysical processes that rhythmically shape spike morphology and tune the firing patterns of these clock neurons. Remarkably, the temporal spiking pattern alone was found to drive neural plastic changes, which mediate the transformation of temporal codes in clock neurons to increased firing of a downstream arousal circuit. Together, these data demonstrate a causal role for temporal coding in behavior that is mediated by a distinct form of synaptic plasticity specifically triggered by the pattern of neural spiking (Tabuchi, 2018).
The molecular mechanisms underlying the generation of different temporal codes are largely unknown. This study shows that the circadian clock drives distinct temporal spiking patterns, as defined by the second-order temporal structure of interspike intervals, by adjusting ionic flux in clock neurons in a time-dependent manner. These changes are mediated by the clock output molecule WAKE, which controls the membrane targeting of SLOB and a Na+/K+ ATPase β subunit. This dynamic regulation of ionic flux leads to cycling of specific aspects of spike waveforms, which in turn induces the temporal spiking patterns seen during the day versus the night (Tabuchi, 2018).
From a broader perspective, this work addresses a central issue in neuroscience: the functional importance of temporal codes in encoding information and impacting behavior. One challenge in demonstrating a causal role of temporal coding is identifying systems with a defined neural circuit where changes in the pattern or timing of spiking occur naturally, lead to measurable effects in target neurons, and regulate a specific behavior. An additional confounding factor is that information can be coded in a multiplexed manner with concurrent spatial, temporal, and rate-related features. This study shows that the Drosophila clock network fulfills these criteria and finds that time is encoded unidimensionally by the spiking patterns of these neurons in the absence of changes in firing rate or network timing (due to synchronization of neural firing within a cluster). Moreover, using computational, in vivo optogenetic, and electrophysiological approaches in these clock neurons, it was demonstrated that this temporal coding has functional consequences on the firing of a target arousal circuit and on sleep behavior. While the findings suggest that the irregular second-order spiking pattern is critical for this process, it is also possible that the temporal code consists of brief periods of faster spiking that are repeated over a >40-s time frame (Tabuchi, 2018).
Previous work has demonstrated that WAKE is critical for clock-dependent regulation of sleep onset at dusk and that it upregulates and properly targets GABAA receptor to mediate this process by markedly suppressing the firing rate of clock neurons (Liu, 2014). Why would multiple neural coding mechanisms (rate coding changes at dusk and temporal coding at mid-day and mid-night) evolve to underlie circadian clock regulation of sleep at different times? One possibility relates to the dynamics of sleep onset versus sleep quality. Transitions between sleep and wake are major changes in brain state occurring on a relatively short timescale and hence may require dramatic changes in firing rate (i.e., rate coding) that are energetically costly. In contrast, maintenance of sleep quality occurs over hours; thus, it may be more energetically favorable for the relevant neurons to alter the pattern, instead of the rate, of their firing. Because of these potential energy savings, it is speculated that the use of temporal spiking patterns to encode information could be a broadly used mechanism for representing persistent internal states, such as hunger or emotion (Tabuchi, 2018).
Finally, this study demonstrate that changes in the pattern of spike firing in the DN1ps, independent of changes in firing rate, triggers NMDA-receptor-dependent postsynaptic plasticity in the Dilp2+ PI neurons. Importantly, these data suggest a specific mechanism for inducing synaptic plasticity distinct from previously described processes that are dependent on changes in rate coding (e.g., long-term potentiation) or relative timing of individual spike events (e.g., STDP). These data represent one of the first examples of synaptic plasticity being induced specifically by the intrinsic temporal pattern of spiking, expanding the repertoire by which neural codes can generate plasticity. Together, these findings suggest that temporal patterns of spike firing are a crucial mechanism for driving neural plastic changes that mediate how internal states modulate behavior (Tabuchi, 2018).
Daily cycles of rest and activity are a common example of circadian
control of physiology. In Drosophila, rhythmic locomotor cycles rely
on the activity of 150-200 neurons grouped in seven clusters. Work
from many laboratories points to the small ventral lateral neurons
(sLNvs) as essential for circadian control of locomotor rhythmicity.
sLNv neurons undergo circadian remodeling of their axonal projections,
opening the possibility for a circadian control of connectivity of
these relevant circadian pacemakers. This study shows that circadian
plasticity of the sLNv axonal projections has further implications
than mere structural changes. First, it was found that the degree of
daily structural plasticity exceeds that originally described,
underscoring that changes in the degree of fasciculation as well as
extension or pruning of axonal terminals could be involved.
Interestingly, the quantity of active zones changes along the day,
lending support to the attractive hypothesis that new synapses are
formed while others are dismantled between late night and the
following morning. More remarkably, taking full advantage of the GFP
reconstitution across synaptic partners (GRASP) technique, this study
showed that, in addition to new synapses being added or removed, sLNv
neurons contact different synaptic partners at different times along
the day. These results lead to a proposal that the circadian network,
and in particular the sLNv neurons, orchestrates some of the
physiological and behavioral differences between day and night by
changing the path through which information travels (Gorostaza, 2014).
Circadian remodeling of the small ventral lateral neuron (sLNv)
dorsal terminals was first described at the peak and trough levels of
pigment-dispersing factor (PDF) immunoreactivity, that is at zeitgeber
time 2 (ZT2) and ZT14 (2 hr after lights ON and lights OFF,
respectively), as well as their counterparts under constant darkness
(DD) (circadian time 2 [CT2] and CT14). For a more precise examination
of the extent of structural remodeling, a time course was carried out.
An inducible GAL4 version termed GeneSwitch restricted to PDF neurons
(pdf-GS) combined with a membrane-tethered version of GFP (mCD8GFP)
was used as control. As expected from the original observations, a
significant reduction in complexity of the axonal arbor-measured as
total axonal crosses-could be seen between CT2 and CT14 and between
CT18 and CT22, which remained unchanged at nighttime. However, toward
the end of the subjective night (CT22), the primary processes appeared
to be shorter. To more precisely describe this additional form of
plasticity, the length of the maximum projection was measure from the
lateral horn toward the midbrain. This analysis revealed that toward
the end of the subjective night (CT22), PDF projections are
significantly shorter than at the beginning of the day (CT2). These
observations imply that mechanisms other than the proposed changes in
the degree of fasciculation are recruited during circadian plasticity.
To get a deeper insight into the nature of the phenomena, the changes
were monitored in brain explants kept in culture for 48 hr after
dissection. Transgenic pdf-GAL4; UAS-mCD8RFP flies (referred to as
pdf>RFP) were dissected under safe red light, and brains were
maintained under DD. Imaging of individual brains at two different
time points highlighted three types of changes experienced by axonal
terminals: (1) changes in the degree of fasciculation/defasciculation,
more common in primary branches, (2) the addition/retraction of new
processes, mostly affecting those of secondary or tertiary order, and
(3) positional changes of minor terminals, thus confirming and
extending previous observations. Altogether, these results indicate
that a rather complex remodeling process takes place on daily basis in
the axonal terminals of PDF neurons (Gorostaza, 2014). The level of structural remodeling occurring at the dorsal terminals
suggested that synapses themselves could undergo changes in a
time-dependent fashion. The presynaptic protein Synaptotagmin (SYT)
was examined at different times across the day as an indicator of
vesicle accumulation. A GFP-tagged version of SYT was expressed in PDF
neurons (pdf >sytGFP), and both the number and area span
by SYT+ puncta (most likely describing the accumulation of several
dense core vesicles) were analyzed separately at the sLNv dorsal
terminals. No statistical differences were observed in the number of
SYT+ puncta (although there is a tendency for higher numbers in the
early morning), perhaps as a result of the nature of the signal, which
is too diffuse for precise identification of individual spots. On the
other hand, SYT+ puncta were larger and, as a result, the area covered
by SYT+ immunoreactivity was significantly different at CT2 compared
to CT14, but not between CT22 and CT2, perhaps reflecting that
vesicles started to accumulate at the end of the day in preparation
for the most dramatic membrane change taking place between CT22 and
the beginning of the following morning (Gorostaza, 2014). The observation that a more complex structure correlated with a
larger area covered by presynaptic vesicles reinforced the notion that
indeed the number of synapses could be changing throughout the day and
prompted analysis of Bruchpilot (BRP), a well-established indicator of
active zones. Expressing a tagged version of BRP in PDF neurons, the
number of BRP+ puncta was quantitated as a proxy for active zones at
times when the most dramatic changes in structure had been detected
(i.e., CT2, CT14, and CT22). Interestingly, the number of active zones
was significantly larger at CT2 than at CT14 or CT22; in fact, no
statistical differences were observed between the last two time
points, underscoring that axonal remodeling can occur (i.e., pruning
of major projections taking place toward the end of the night) without
significantly affecting overall connectivity. Thus, circadian
structural plasticity is accompanied by changes in the number of
synapses. Not only are more vesicles recruited toward CT2, but also a
higher number of active zones are being established (Gorostaza, 2014).
Circadian changes in the abundance of the presynaptic active zone BRP
have also been shown in the first optic neuropil of the fly brain,
although BRP abundance in the lamina increases in the early night
under DD conditions, in contrast to the oscillations in BRP levels
observed at the dorsal protocerebrum that peak in the early subjective
day just described. In addition, rhythmic changes in the number of
synapses have also been described in the terminals of adult motor
neurons in Drosophila examined through transmission electron
microscopy, as well as BRP+ light confocal microscopy, underscoring
the validity of the approach employed herein. Interestingly, in
different brain areas, the level of presynaptic markers (such as BRPRFP
or SYTGFP) also changes in response to the sleep/wake
'state,' being high when the animals are awake and lower during sleep;
this observation led to the proposal that sleep could be involved in
maintaining synaptic homeostasis altered during the awaking state.
This trend coincides with observation of higher levels during the
subjective morning and lower levels at the beginning of the subjective
night; however, no changes were detected through the night, suggesting
that, at least in clock neurons, there is a circadian rather than a
homeostatic control of synaptic activity. Given that clock outputs are
predominantly regulated at the transcriptional level and that there is
circadian regulation of MEF2, a transcription factor that turns on a
program involved in structural remodeling, this correlation opens the
provocative possibility that the circadian clock is controlling the
ability of assembling novel synapses in particularly plastic neurons,
which might become recruited and/or stabilized, or otherwise pruned
(disassembled), toward the end of the day (Gorostaza, 2014). Adult-specific electrical silencing of PDF neurons reduces the
complexity of dorsal arborizations, although a certain degree of
circadian remodeling of the axonal terminals still takes place. To
examine whether electrical alterations could affect circadian changes
in the number of active zones, either Kir2.1 or NaChBac was expressed
(to hyperpolarize or depolarize PDF neurons, respectively). To avoid
any undesired developmental defects, pdf-GS was used to drive
expression of the channels only during adulthood. Interestingly,
Kir2.1 expression abrogated circadian changes in the number of active
zones. In fact, PDF neurons displayed a reduced number of active zones
compared to controls at CT2 and remained at similar levels throughout
the day, indistinguishable from nighttime controls. On the other hand,
when neurons were depolarized through NaChBac expression, the number
of active zones did not change along the day and was maintained at
daytime levels even at CT14 and CT22 (Gorostaza, 2014). It has recently been shown that MEF2, a transcription factor involved
in activity-dependent neuronal plasticity and morphology in mammals,
is circadianly regulated and mediates some of the remodeling of PDF
dorsal terminals through the regulation of Fasciclin2. In contrast,
adult-specific silencing (and depolarization) of PDF neurons abolishes
cycling in the number of BRP+ active zones, despite the fact that it
does not completely obliterate the remodeling of the axonal terminals,
suggesting that some of the mechanisms underlying structural
plasticity are clearly activity independent and are most likely the
result of additional clock-controlled output pathways still to be
identified (Gorostaza, 2014). Since structural remodeling of PDF neurons results in the formation
and disappearance of new synapses on daily basis, it was anticipated
that not only the number but also the postsynaptic partners of these
contacts could concomitantly be changing. To shed light on this
possibility, GFP reconstitution across synaptic partners (GRASP),
which labels contacts between adjacent membranes, was used. In brief,
two complementary fragments of GFP tethered to the membrane are
expressed in different cells. If those cells are in contact, GFP is
reconstituted and becomes fluorescent. GRASP has previously been
employed to monitor synapses in adult flies. Given the complex
arborization at the dorsal protocerebrum, it was asked whether
specific subsets of circadian neurons projecting toward that area
could be contacting across the day. Perhaps not surprisingly, an
extensive reconstituted GFP signal could be observed between the sLNv
dorsal projections and those of the posterior dorsal neuron 1 cells
(DN1ps, lighted up by the dClk4.1-GAL4 line, suggesting contacts along
the entire area, which are detectable across all time points analyzed
(ZT2, ZT14, and ZT22). Consistent with these observations, extensive
physical contact between the sLNv projections and those of the DN1p
neurons has just been reported at the dorsal protocerebrum with no
clear indication of the time of day examined. Next the study examined
whether a subset of dorsal LNs (LNds), projecting toward both the
accessory medulla and the dorsal protocerebrum (through the combined
expression of Mai179-GAL4; pdf-GAL80), could also contact the profuse
dorsal arborization of sLNv neurons; this genetic combination enables
expression of split-GFP in a restricted number of circadian cells
(which are part of the evening oscillator, i.e., up to four LNds,
including at least a CRYPTOCHROME-positive one, and the fifth sLNv),
as well as others located within the pars intercerebralis (PI), a
neurosecretory structure recently identified as part of the output
pathway relevant in the control of locomotor behavior. In contrast to
the extensive connections between DN1p and sLNv clusters, only very
discreet reconstituted puncta were detected. Quite strikingly, the
degree of connectivity appeared to change across the day, reaching a
maximum (when almost every brain exhibited reconstituted signal) at
ZT22. However, due to the nature of the signal, no quantitation of its
intensity was attempted. Although a more detailed analysis is required
to define the identity (i.e., whether it is one or several LNds, the
fifth sLNv, or both groups that directly contact the sLNvs), this
finding highlights a potentially direct contact between the neuronal
substrates of the morning and evening oscillators. In sum, through
GRASP analysis, this study has begun to map the connectivity within
the circadian network; commensurate with a hierarchical role, the
sLNvs appear to differentially contact specific subsets in a
distinctive fashion (Gorostaza, 2014). To address the possibility that PDF neurons could be contacting
noncircadian targets at different times across the day, an enhancer
trap screen was carried out employing a subset of GAL4 enhancers
selected on the basis of their expression pattern in the adult brain,
i.e., known to drive expression in the dorsal protocerebrum, and an
additional requirement imposed was that none of the selected GAL4
lines could direct expression to the sLNv neurons to avoid internal
GFP reconstitution. Reconstitution of the GFP signal at the sLNv
dorsal terminals by recognition through specific antibodies was
assessed at three different time points for each independent GAL4 line
(ZT2, ZT14, and ZT22). Some of the GAL4 lines showed reconstituted GFP
signal at every time point analyzed, suggesting that those neuronal
projections are indeed in close contact across the day and might
represent stable synaptic contacts. No GFP signal was detected in the
negative parental controls. Despite the fact that several GAL4 drivers
directed expression to the proximity of the PDF dorsal terminals, some
of the selected lines did not result in reconstituted GFP signal
(Gorostaza, 2014). Quite remarkably, a proportion of the GAL4 lines showed GFP+ signal
only at a specific time point. One such example is line 3-86, where
reconstitution was detected in most of the brains analyzed at ZT2, but
not at nighttime. Being able to identify putative postsynaptic
contacts to the sLNvs in the early morning is consistent with the
observation of a higher number of BRP+ active zones in the early day.
This enhancer trap spans different neuropils, such as the mushroom
body (MB) lobes and lateral horn, and directs expression to
particularly high levels in the PI, a structure that has recently been
implicated in the rhythmic control of locomotor activity. In fact,
some yet unidentified somas in the PI appear to arborize profusely
near the PDF dorsal terminals, underscoring a potential link between
the two neuronal groups. These direct contacts are unlikely to be the
ones reported by Mai179-GAL4; pdf-GAL80 since those connect to the
sLNv neurons preferentially at night. Interestingly, a subset of
neurons in the PI is relevant in mediating the arousal promoting
signal from octopamine; in addition, sleep promoting signals are also
derived from a different subset of neurons in the PI, opening the
attractive possibility that both centers could be under circadian
modulation (Gorostaza, 2014). GRASP analysis also uncovered a different neuronal cluster (4-59)
that contacts PDF neurons preferentially during the early night
(ZT14), which is in itself striking, since this time point corresponds
to that with fewer arborizations and an overall decrease in the number
of synapses. This enhancer trap is expressed in the MBs, subesophagic
ganglion, antennal lobes, and accessory medulla. Among those
structures, the MBs are important for higher-order sensory integration
and learning in insects. Interestingly, circadian modulation of
short-term memory and memory retrieval after sleep deprivation has
been reported; short-term memory was found to peak around ZT15-ZT17,
coinciding with the window of GFP reconstitution, thus providing a
functional connection to the synaptic plasticity observed. To
corroborate whether there is a direct contact between the two neuronal
clusters, the extensively used GAL4 driver OK107, which is expressed
in the α'/β'and the γ lobes of the MBs and to a lower extent in the
PI, was employed for GRASP analysis. Surprisingly, reconstituted GFP
signal could be observed at every time point analyzed, suggesting that
MB lobes contact PDF neurons throughout the day but that specific
clusters (for example those highlighted by the 4-59 line) establish
plastic, time-of-day-dependent physical contact with PDF neurons
(Gorostaza, 2014). It was next asked whether these prospective postsynaptic targets of
PDF neurons could play a role in the output pathway controlling
rhythmic locomotor activity. To address this possibility, the impact
of adult-specific alteration of excitability of distinct neuronal
groups was examined through expression of TRPA1. Interestingly,
adult-specific depolarization of specific neuronal populations
triggered a clear deconsolidation of the rhythmic pattern of activity,
which resulted in less-rhythmic flies accompanied by a significant
decrease in the strength of the underlying rhythm. These results lend
support to the notion that the underlying neuronal clusters are
relevant in the control of rest/activity cycles (Gorostaza, 2014). Over the years, it has become increasingly clear
that the circadian clock modulates structural properties of different
cells. In fact, a number of years ago, it was reported that the
projections of a subset of core pacemaker fly PDF+ and mammalian VIP+
neurons undergo structural remodeling on daily basis. The work
presented in this study lends support to the original hypothesis that
circadian plasticity represents a means of encoding time-of-day
information. By changing their connectivity, PDF neurons could drive
time-specific physiological processes. As new synapses assemble while
others are dismantled, the information flux changes, allowing PDF
neurons to promote or inhibit different processes at the same time.
This type of plasticity adds a new level to the complex information
encoded in neural circuits, where PDF neurons could not only modulate
the strength in the connectivity between different partners, but also
define which neuronal groups could be part of the circadian network
along the day. Although further analysis of the underlying process is
ensured, evidence so far supports the claim that structural plasticity
is an important circadian output (Gorostaza, 2014). Aging has been linked with decreased neural
plasticity and memory formation in humans and in laboratory model
species such as the fruit fly, Drosophila melanogaster. This study
examined plastic responses following social experience in Drosophila
as a high-throughput method to identify interventions that prevent
these impairments. Young (5-day old) or aged (20-day old) adult female
Drosophila were housed in socially enriched or isolated environments,
then assayed for changes in sleep and for structural markers of
synaptic terminal growth in the ventral lateral neurons (LNVs) of the
circadian clock. When young flies are housed in a socially enriched
environment, they exhibit synaptic elaboration within a component of
the circadian circuitry, the LNVs, which is followed by increased
sleep. Aged flies, however, no longer exhibit either of these plastic
changes. Because of the tight correlation between neural plasticity
and ensuing increases in sleep, sleep after enrichment was used as a
high-throughput marker for neural plasticity to identify interventions
that prolong youthful plasticity in aged flies. To validate this
strategy, three independent genetic manipulations were used that delay
age-related losses in plasticity: (1) elevation of dopaminergic
signaling, (2) over-expression of the serum response factor
transcription factor blistered
(bs) in the LNVs, and (3) reduction of the Imd
immune signaling pathway. These findings provide proof-of-principle
evidence that measuring changes in sleep in flies after social
enrichment may provide a highly scalable assay for the study of
age-related deficits in synaptic plasticity. These studies demonstrate
that Drosophila provides a promising model for the study of age-related
loss of neural plasticity and begin to identify genes that might be
manipulated to delay the onset of functional senescence (Donlea, 2014a).
All organisms have to adapt to acute as well as to
regularly occurring changes in the environment. To deal with these
major challenges organisms evolved two fundamental mechanisms: the p38
mitogen-activated protein kinase (MAPK) pathway, a major stress
pathway for signaling stressful events, and circadian clocks to prepare
for the daily environmental changes. Both systems respond sensitively to
light. Recent studies in vertebrates and fungi indicate that p38 is
involved in light-signaling to the circadian clock providing an
interesting link between stress-induced and regularly rhythmic
adaptations of animals to the environment, but the molecular and
cellular mechanisms remained largely unknown. This study demonstrates by
immunocytochemical means that p38 is expressed in Drosophila
melanogaster's clock neurons and that it is activated in a
clock-dependent manner. Surprisingly, it was found that p38 is most
active under darkness and, besides its circadian activation,
additionally gets inactivated by light. Moreover, locomotor activity
recordings revealed that p38 is essential for a wild-type timing of
evening activity and for maintaining approximately 24 h behavioral
rhythms under constant darkness: flies with reduced p38 activity in
clock neurons, delayed evening activity and lengthened the period of
their free-running rhythms. Furthermore, nuclear translocation of the
clock protein Period was significantly delayed on the expression of a
dominant-negative form of p38b in Drosophila's most important clock
neurons. Western Blots revealed that p38 affects the phosphorylation
degree of Period, what is likely the reason for its effects on nuclear
entry of Period. In vitro kinase assays confirmed the Western Blot
results and point to p38 as a potential 'clock kinase' phosphorylating
Period. Taken together, these findings indicate that the p38 MAP Kinase
is an integral component of the core circadian clock of Drosophila in
addition to playing a role in stress-input pathways (Dusik, 2014 - Open
access: 25144774).
Class IIa histone deacetylases (HDACs) regulate the
activity of many transcription factors to influence liver
gluconeogenesis and the development of specialized cells including
muscle, neurons and lymphocytes. This study describes a conserved role
for class IIa HDACs in sustaining robust circadian behavioral rhythms
in Drosophila and cellular rhythms in mammalian cells. In mouse
fibroblasts, over-expression of HDAC5 severely disrupts
transcriptional rhythms of core clock genes. HDAC5 over-expression
decreases BMAL1 acetylation on Lys537 and pharmacological inhibition
of Class IIa HDACs increases BMAL1 acetylation. Furthermore, cyclical
nucleocytoplasmic shuttling of HDAC5 was observed in mouse fibroblasts
that is characteristically circadian. Mutation of the Drosophila
homolog Circadian clock and chromatin-remodeling complexes are tightly intertwined systems that regulate rhythmic gene expression. The circadian clock promotes rhythmic expression, timely recruitment, and/or activation of chromatin remodelers, while chromatin remodelers regulate accessibility of clock transcription factors to the DNA to influence expression of clock genes. It has been previously reported that the BRAHMA (BRM) chromatin-remodeling complex promotes the repression of circadian gene expression in Drosophila. This study investigated the mechanisms by which the circadian clock feeds back to modulate daily BRM activity. Using chromatin immunoprecipitation, rhythmic BRM binding to clock gene promoters was observed despite constitutive BRM protein expression, suggesting that factors other than protein abundance are responsible for rhythmic BRM occupancy at clock-controlled loci. Since it was previously reported that BRM interacts with two key clock proteins, CLOCK (CLK) and TIMELESS (TIM), their effect on BRM occupancy to the period (per) promoter was examined. Reduced BRM binding to the DNA was observed in clk null flies, suggesting that CLK is involved in enhancing BRM occupancy to initiate transcriptional repression at the conclusion of the activation phase. Additionally, reduced BRM binding to the per promoter was observed in flies overexpressing TIM, suggesting that TIM promotes BRM removal from DNA. These conclusions are further supported by elevated BRM binding to the per promoter in flies subjected to constant light and experiments in Drosophila tissue culture in which the levels of CLK and TIM are manipulated. In summary, this study provides new insights into the reciprocal regulation between the circadian clock and the BRM chromatin-remodeling complex (Tabuloc, 2023).
Circadian clocks keep time via ~ 24 h transcriptional feedback loops. In Drosophila, CLOCK-CYCLE (CLK-CYC) activators and PERIOD-TIMELESS (PER-TIM) repressors are feedback loop components whose transcriptional status varies over a circadian cycle. This study used mass spectrometry to identify proteins that interact with GFP-tagged CLK (GFP-CLK) in fly heads at different times of day. Many expected and novel interacting proteins were detected, of which several interacted rhythmically and were potential regulators of protein levels, activity or transcriptional output. Genes encoding these proteins were tested to determine if they altered circadian behavior via RNAi knockdown in clock cells. The NIPPED-A protein, a scaffold for the SAGA and Tip60 histone modifying complexes, interacts with GFP-CLK as transcription is activated, and reducing Nipped-A expression lengthens circadian period. RNAi analysis of other SAGA complex components shows that the SAGA histone deubiquitination (DUB; see Non-stop) module lengthened period similarly to Nipped-A RNAi knockdown and weakened rhythmicity, whereas reducing Tip60 HAT expression drastically weakened rhythmicity. These results suggest that CLK-CYC binds NIPPED-A early in the day to promote transcription through SAGA DUB and Tip60 HAT activity (Mahesh, 2020).
It is known that endocytosis
and/or vesicle recycling mechanisms are essential in adult Drosophila
glial
cells for the neuronal control of circadian locomotor activity.
The goal of this study was to identify specific glial vesicle
trafficking, recycling, or release factors that are required for
rhythmic behavior. From a glia-specific, RNAi-based genetic screen,
eight glial factors were identified and were found to be required for
normally robust circadian rhythms in either a light-dark cycle or in
constant dark conditions. In particular, it was shown that conditional
knockdown of the ROP
vesicle release factor in adult glial cells results in arrhythmic
behavior. Immunostaining for ROP reveals reduced protein in glial cell
processes and an accumulation of the Par
Domain Protein 1ε (PDP1ε) clock output protein in the small
lateral clock neurons. These results suggest that glia modulate rhythmic
circadian behavior by secretion of factors that act on clock neurons to
regulate a clock output factor (Ng, 2015).
Nutrient sensing pathways adjust metabolism and
physiological functions in response to food intake. For example, sugar
feeding promotes lipogenesis by activating glycolytic and lipogenic
genes through the Mondo/ChREBP- This study establishes the Drosophila klf10 ortholog, cbt,
as a repressive effector of the sugar sensing transcriptional network.
Specifically, (1) cbt expression is activated by dietary sugars
in mlx-dependent manner; (2) cbt is a direct target of Mlx; (3)
many key metabolic genes are rapidly repressed by CBT upon sugar
feeding; (4) CBT binds to the proximity of pepck genes; (5) pepck1
is dispensable for viability, but essential for glucose and glycerol
homeostasis; (6) deregulation of pepck1 underlies lethality of mlx
mutants, and (7) CBT modulates the circadian system by controlling the
cycling of a subset of circadian output genes. Based on these findings,
a model is proposed in which CBT serves as a repressive downstream
effector of the Mondo-Mlx-mediated sugar sensing, which contributes to
diet-induced physiological readjustment, including flux of central
carbon metabolism and cycling of metabolic circadian clock targets
(Bartok, 2015). By uncovering the CBT-mediated repression of pepck isoforms
downstream of Mondo-Mlx, This study provides a mechanistic explanation
to the regulation of cataplerosis in response to intracellular sugar
sensing. Drosophila Mondo-Mlx is known to drive activation of
glycolysis, for example, by promoting the expression of phosphofructokinase
2. Placing the rate-limiting enzymes of gluconeogenesis
downstream, the same sensor mechanism that activates glycolysis provides
an elegant mechanism to adjust the direction of flux of glucose
metabolism in response to sugar input. Such simple network topology
provides a robust safeguard against loss of energy in futile cycles
caused by simultaneous high activity of glycolysis and gluconeogenesis
(Bartok, 2015). Mondo-Mlx also activates the expression of lipogenic gene expression
(e.g., FAS and ACC) in order to promote conversion of
excess sugars into triglycerides. In addition to fatty acid moieties,
which are built by FAS and ACC, triglyceride biosynthesis requires
glycerol-3-phosphate. Substrate-labeling studies in mammals have shown
that a significant portion of glycerol in triglycerides is in fact
derived from the PEPCK-dependent glyceroneogenesis pathway. This is
supported by the current findings showing significantly lower
circulating glycerol levels in well-fed pepck1-mutant larvae.
The impact of glyceroneogenesis on triglyceride homeostasis is also
likely reflected in the reduced triglyceride levels in pepck1
mutant flies. Similarly, mammalian studies have shown that elevated
expression of PEPCK-C in adipose tissue increases fat mass, whereas
reduced PEPCK-C expression leads to lower fat content. Moreover, in
humans, adipose tissue expression of PEPCK-C positively correlates with
adiposity and plasma triacylglycerol levels. Control of pepck
through CBT places both branches of triglyceride biosynthesis under
Mondo-Mlx. Inhibition of PEPCK-mediated cataplerosis upon high sugar
intake allows maximal conversion of excess glucose-6-phosphate into the
glycerol moieties of triglycerides through the glycolytic route of
glycerol-3-phosphate synthesis. Simultaneous impairment of de novo
lipogenesis and failure to suppress glyceroneogenesis likely leads to
the breakage of glycerol homeostasis and massive accumulation of
circulating glycerol, as observed in mlx mutants. Interestingly,
a recent study showed that fasting serum levels of glycerol predicted
development of hyperglycemia and type 2 diabetes. It will be interesting
to learn whether this diagnostic marker is associated with deregulation
of pepck isoforms and the activity of ChREBP/MondoA-Mlx and
Klf10-dependent transcriptional network (Bartok, 2015). According to the current view, Mondo/ChREBP and Mlx act mainly in
nutrient sensing and metabolic regulation. In contrast, CBT and its
human ortholog Klf10 are multifunctional transcription factors. In
Drosophila, cbt was originally identified as a developmental
regulator with an essential function in dorsal closure early in
development. Moreover, cbt is a direct transcriptional target of
the circadian transcription factor CLK, and this study establishes it is
deeply involved in the control of the circadian transcriptional network.
While CBT overexpression leads to strong behavioral abnormalities, they
are not accompanied by noticeable changes in the oscillation of the core
clock components in the fly heads. This suggests that it reflects a
specific effect on circadian output. If the behavioral patterns were due
to an effect in the general health of the animal, deregulation of core
clock components would be expected. Despite the null effect of cbt
overexpression in the expression of core clock genes, this study
observed that cbt modulates the expression of an important
subset of CLK and circadian-controlled genes, most of which are involved
in metabolic functions. Strong effects of cbt downregulation
were observed in circadian oscillation of metabolic genes, establishing
CBT as a new regulator of the circadian transcriptome. Interestingly,
downregulation of CBT in circadian cells decreases the amplitude of
oscillation of a large number of circadian-controlled genes, providing a
direct link between food intake, circadian gene expression, and
behavior. Given the established link between feeding time, metabolism,
and the circadian system in Drosophila, it will be interesting to
further analyze the importance of CBT in this coordination (Bartok,
2015). Although the functional analysis in this study largely focused on the
metabolic role of pepck regulation by Mondo-Mlx-CBT network,
microarray and RNA-seq analyses revealed other interesting CBT
transcriptional targets including bmm. This gene is an ortholog
of the human adipocyte triglyceride lipase gene, and it is an
essential regulator of triglyceride stores in Drosophila. bmm
expression is positively regulated by the Forkhead transcription factor
FoxO, which is activated during starvation through inhibition of
insulin-like signaling. The sugar-dependent repression of bmm
expression by CBT is in perfect agreement with the lipogenic role of
Mondo-Mlx (Bartok, 2015). It has been proposed that CBT mammalian ortholog Klf10 acts as a
negative feedback regulator for ChREBP-activated genes, including
lipogenic genes FAS and ACC. This conclusion was based on suppression of
ChREBP-activated transcription upon Klf10 overexpression in primary
liver cells. This model was tested by analyzing the expression of
Mondo-Mlx targets FAS and ACC, but no effect was
observed by cbtRNAi. In contrast, genome-wide expression analysis of CBT
loss-of-function flies revealed that the CBT-dependent branch of the
sugar sensing transcriptional network mediates rapid repression of gene
expression. It is interesting to note that while most metabolic targets
of CBT are rapidly and persistently downregulated, cbt
expression is rapidly attenuated during prolonged sugar feeding. This is
likely due to the negative autoregulation demonstrated earlier and
supported by ChIP data. The finding that most of the identified
CBT-dependent mRNAs are stably repressed for many hours after cbt
levels have significantly attenuated suggests that CBT-mediated
repression might involve regulation at the chromatin level. This is in
agreement with the possible involvement of Sin3A in CBT-mediated
repression. Through such persistent regulatory marks, sugar feeding may
have a long-lasting influence on metabolic homeostasis, which is a topic
that certainly deserves to be more thoroughly analyzed in the future
(Bartok, 2015). In sum, this work provides a mechanistic explanation for the
transcriptional repression upon Mondo-Mlx-mediated intracellular sugar
sensing through the transcription factor CBT. The CBT-mediated
repressive branch of the sugar sensing network is involved in securing
the mutually exclusive activity of glycolysis and gluconeogenesis and
coordination of fatty acid and glycerol biosynthesis with respect to
dietary sugar intake. This study also establishes a mechanism for
nutrient input into the circadian gene expression. As intracellular
sugar-sensing and circadian regulation are highly conserved processes,
the insight achieved in this study in the genetically tractable
Drosophila model should provide a new conceptual framework for
forthcoming studies in human subjects and mammalian model systems
(Bartok, 2015).
Nocturnin (NOCT) is a rhythmically expressed protein that regulates metabolism under the control of circadian clock. It has been proposed that NOCT deadenylates and regulates metabolic enzyme mRNAs. However, in contrast to other deadenylases, purified NOCT lacks the deadenylase activity. To identify the substrate of NOCT, a mass spectrometry screen was conducted, and NOCT was found to specifically and directly convert the dinucleotide NADP(+) into NAD(+) and NADPH into NADH. Further, it was demonstrated that the Drosophila NOCT ortholog, Curled, has the same enzymatic activity. The 2.7 A crystal structure of the human NOCT*NADPH complex, revealed that NOCT recognizes the chemically unique ribose-phosphate backbone of the metabolite, placing the 2'-terminal phosphate productively for removal. Evidence is provided for NOCT targeting to mitochondria and it is proposed that NADP(H) regulation, which takes place at least in part in mitochondria, establishes the molecular link between circadian clock and metabolism (Estrella, 2019).
Metabolic state is a potent modulator of sleep and circadian behavior and animals acutely modulate their sleep in accordance with internal energy stores and food availability. Growing evidence suggests the fat body is a critical regulator of complex behaviors, but little is known about the genes that function within the fat body to regulate sleep. To identify molecular factors functioning in non-neuronal tissues to regulate sleep, an RNAi screen selectively knocking down genes in the fat body. Knockdown was performed of Phosphoribosylformylglycinamidine synthase/Pfas(Ade2), a highly conserved gene involved the biosynthesis of purines, sleep regulation and energy stores. Flies heterozygous for multiple Ade2 mutations are also short sleepers and this effect is partially rescued by restoring Ade2 to the Drosophila fat body. Targeted knockdown of Ade2 in the fat body does not alter arousal threshold or the homeostatic response to sleep deprivation, suggesting a specific role in modulating baseline sleep duration. Together, these findings suggest Ade2 functions within the fat body to promote both sleep and energy storage, providing a functional link between these processes (Yurgel, 2018).
Post-translational modification of histones, such as histone methylation controlled by specific methyltransferases and demethylases, play critical roles in modulating chromatin dynamics and transcription in eukaryotes. Misregulation of histone methylation can lead to aberrant gene expression, thereby contributing to abnormal development and diseases such as cancer. As such, the mammalian lysine-specific demethylase 2 (KDM2) homologs, KDM2A and KDM2B, are either oncogenic or tumor suppressive depending on specific pathological contexts. However, the role of KDM2 proteins during development remains poorly understood. Unlike vertebrates, Drosophila has only one KDM2 homolog (dKDM2), but its functions in vivo remain elusive due to the complexities of the existing mutant alleles. To address this problem, two dKdm2 null alleles were generated using the CRISPR/Cas9 technique. These dKdm2 homozygous mutants are fully viable and fertile, with no developmental defects observed under laboratory conditions. However, the dKdm2 null mutant adults display defects in circadian rhythms. Most of the dKdm2 mutants become arrhythmic under constant darkness, while the circadian period of the rhythmic mutant flies is approximately 1 h shorter than the control. Interestingly, lengthened circadian periods are observed when dKdm2 is overexpressed in circadian pacemaker neurons. Taken together, these results demonstrate that dKdm2 is not essential for viability; instead, dKDM2 protein plays important roles in regulating circadian rhythms in Drosophila. Further analyses of the molecular mechanisms of dKDM2 and its orthologs in vertebrates regarding the regulation of circadian rhythms will advance understanding of the epigenetic regulations of circadian clocks (Zheng, 2018).
The environmental light-dark cycle is the dominant cue
that maintains 24-h biological rhythms. In Drosophila, light
entrainment is mediated by the photosensitive protein Circadian clocks regulate membrane excitability in
master pacemaker neurons to control daily rhythms of sleep and wake.
This study found that two distinctly timed electrical drives
collaborate to impose rhythmicity on Drosophila clock neurons. In the
morning, a voltage-independent sodium conductance via the NA/NALCN ion
channel Narrow abdomen depolarizes these neurons. This current is
driven by the rhythmic expression of NCA
localization factor-1 (CG10420), linking the molecular clock to
ion channel function. In the evening, basal potassium currents peak to
silence clock neurons. Remarkably, daily antiphase cycles of sodium and
potassium currents also drive mouse clock neuron rhythms. Thus, this
study reveals an evolutionarily ancient strategy for the neural
mechanisms that govern daily sleep and wake (Flourakis, 2015). The hypothesis that circadian clocks confer adaptive advantage to
organisms has been proposed based on its ubiquity across almost all
levels of complexity and organization of life-forms. This thought has
received considerable attention, and studies employing diverse
strategies have attempted to investigate it. However, only a handful
of them have examined how selection for circadian clock controlled
rhythmic behaviors influences life-history traits which are known to
influence Darwinian fitness. The 'early' and 'late' chronotypes are
amongst the most widely studied circadian phenotypes; however,
life-history traits associated with these chronotypes, and their
consequences on Darwinian fitness remain largely unexplored, primarily
due to the lack of a suitable model system. Several life-history
traits of Drosophila melanogaster populations were studied that were
subjected to laboratory selection for morning (early) and evening
(late) emergence. This paper reports that the late eclosion
chronotypes evolved longer pre-adult duration as compared to the early
eclosion chronotypes both under light/dark (LD) and constant dark (DD)
conditions, and these differences appear to be mediated by both clock
dependent and independent mechanisms. Furthermore, longer pre-adult
duration in the late chronotypes does not lead to higher body-mass at
pupariation or eclosion, but the late females were significantly more
fecund and lived significantly shorter as compared to the early
females. It is concluded that coevolution of multiple life-history
traits in response to selection on timing of eclosion highlights
correlations of the genetic architecture governing timing of eclosion
with that of fitness components which suggests that timing
ecologically relevant behaviors at specific time of the day might
confer adaptive advantage (Nikhil, 2016).
Even though the rhythm in adult emergence and rhythm in locomotor activity are two different rhythmic phenomena that occur at distinct life-stages of the fly life cycle, previous studies have hinted at similarities in certain aspects of the organisation of the circadian clock driving these two rhythms. For instance, the period gene plays an important regulatory role in both rhythms. Previous work showed that selection on timing of adult emergence behaviour in populations of Drosophila melanogaster leads to the co-evolution of temperature sensitivity of circadian clocks driving eclosion. In this study, it was asked if temperature sensitivity of the locomotor activity rhythm has evolved in the selected populations with divergent timing of adult emergence rhythm, with the goal of understanding the extent of similarity (or lack of it) in circadian organisation between the two rhythms. In response to simulated jetlag with temperature cycles, late chronotypes (populations selected for predominant emergence during dusk) indeed re-entrain faster than early chronotypes (populations selected for predominant emergence during dawn) to 6-h phase-delays, thereby indicating enhanced sensitivity of the activity/rest clock to temperature cues in these stocks (entrainment is the synchronisation of internal rhythms to cyclic environmental time-cues). Additionally, it was found that late chronotypes show higher plasticity of phases across regimes, day-to-day stability in phases and amplitude of entrainment, all indicative of enhanced temperature sensitive activity/rest rhythms. These results highlight remarkably similar organisation principles between emergence and activity/rest rhythms (Abhilash, 2020).
It is a common notion that phases-of-entrainment of circadian rhythms are adaptive, in that they enable organisms to time their behavior to specific times of the day to enhance their fitness. Previous studies have shown that selection for morning and evening phasing of adult emergence in Drosophila melanogaster populations leads to divergent coevolution of free-running periods of both adult emergence and activity/rest rhythms, such that early (morning) and late (evening) adult emergence chronotypes have shorter and longer circadian periods, respectively. However, there is little evidence to support the notion that phases-of-entrainment in these fly stocks is indeed driven by non-parametric mechanisms. Extending from a previous hypothesis based on anecdotal evidence for parametric mechanisms being in play, this study explored the extent of non-parametric and parametric effects of light on circadian clocks of early and late chronotypes. Predictions of the non-parametric model of entrainment were systematically tested, the Circadian Integrated Response Characteristic (CIRC) of the stocks , the effect of light pulses on amplitude of the behavior and the effect of duration of light pulse on phase-shifts of the clock were assessed were sketched. In addition to the differences in clock period, divergent CIRCs contribute to entrainment of the activity/rest rhythm. The differences in CIRC could be explained by differential transient amplitude responses and duration responses of the clock's phase between the early and late chronotypes. This study thus highlights the role of amplitude responses and phase-shifts due to long durations of light in entrainment of circadian rhythms of D. melanogaster (Abhilash, 2020).
Light is the primary signal that calibrates circadian neural circuits and thus coordinates daily physiological and behavioral rhythms with solar entrainment cues. Drosophila and mammalian circadian circuits consist of diverse populations of cellular oscillators that exhibit a wide range of dynamic light responses, periods, phases, and degrees of synchrony. How heterogeneous circadian circuits can generate robust physiological rhythms while remaining flexible enough to respond to synchronizing stimuli has long remained enigmatic. Cryptochrome is a short-wavelength photoreceptor that is endogenously expressed in approximately half of Drosophila circadian neurons. This study applied analysis of real-time bioluminescence experimental data to show detailed dynamic ensemble representations of whole circadian circuit light entrainment at single neuron resolution. Organotypic whole-brain explants were either maintained in constant darkness (DD) for 6 days or exposed to a phase-advancing light pulse on the second day. Stronger circadian oscillators were found to support robust overall circuit rhythmicity in DD, whereas weaker oscillators can be pushed toward transient desynchrony and damped amplitude to facilitate a new state of phase-shifted network synchrony. Additionally, mathematical modeling was used to examine how a network composed of distinct oscillator types can give rise to complex dynamic signatures in DD conditions and in response to simulated light pulses. Simulations suggest that complementary coupling mechanisms and a combination of strong and weak oscillators may enable a robust yet flexible circadian network that promotes both synchrony and entrainment. A more complete understanding of how the properties of oscillators and their signaling mechanisms facilitate their distinct roles in light entrainment may allow direction and augmentation of the circadian system to speed recovery from jet lag, shift work, and seasonal affective disorder (Roberts, 2016).
Robustness, the ability to maintain stable biological phenotypes across environments, is thought to be of adaptive value. Previous studies have reported higher intrinsic activity levels and power of rhythm in Drosophila populations (stocks) reared in constant darkness (DD stocks) as compared to those reared in constant light (LL stocks) and 12:12-h light-dark cycles (LD stocks) for over 19 years (approximately 330 generations). The current study intended to examine whether the enhanced levels of activity observed in DD stocks persist under various environments such as photoperiods, ambient temperatures, non-24-h light-dark (LD) cycles, and semi-natural conditions (SN). DD stocks largely retain their phenotype of enhanced activity levels across most of the above-mentioned environments suggesting the evolution of robust circadian clocks in DD stocks. Furthermore, the change in peak activity levels upon entrainment was not significantly different across the three stocks for any of the examined environmental conditions. This suggests that the enhancement of activity levels in DD stocks is not due to differential sensitivity to environment. Thus, these results suggest that rearing in constant darkness (DD) leads to evolution of robust circadian clocks suggesting a possible adaptive value of possessing such rhythms under constant dark environments (Shindey, 2016).
The level of rescue of clock function in genetically arrhythmic Drosophila melanogaster hosts using interspecific clock gene transformation was used to study the putative intermolecular coevolution between interacting clock proteins. Among them PER and TIM are the two important negative regulators of the circadian clock feedback loop. This study transformed either the D. pseudoobscura per or tim transgenes into the corresponding arrhythmic D. melanogaster mutant (per01 or tim01) and observed >50% rhythmicity but the period of activity rhythm was either longer (D. pseudoobscura-per) or shorter than 24h (D. pseudoobscura-tim) compared to controls. By introducing both transgenes simultaneously into double mutants, it was observed that the period of the activity rhythm was rescued by the pair of hemizygous transgenes (~24h). These flies also showed a more optimal level of temperature compensation for the period. Under LD 12:12 these flies have a D. pseudoobscura like activity profile with the absence of morning anticipation as well as a very prominent earlier evening peak of activity rhythm. These observation are consistent with the view that TIM and PER form a heterospecific coevolved module at least for the circadian period of activity rhythms. However the strength of rhythmicity was reduced by having both transgenes present, so while evidence for a coevolution between PER and TIM is observed for some characters it is not for others (Noreen, 2018).
Most organisms possess time-keeping devices called circadian clocks. At the molecular level, circadian clocks consist of transcription-translation feedback loops (TTFLs). Although some components of the negative TTFL are conserved across the animals, important differences exist between typical models, such as mouse and the fruit fly. In Drosophila, the key components are PERIOD (PER) and TIMELESS (TIM-d) proteins, whereas the mammalian clock relies on PER and CRYPTOCHROME (CRY-m). Importantly, how the clock has maintained functionality during evolutionary transitions between different states remains elusive. Therefore, this study systematically described the circadian clock gene setup in major bilaterian lineages and identified marked lineage-specific differences in their clock constitution. Then a thorough functional analysis was performed of the linden bug Pyrrhocoris apterus, an insect species comprising features characteristic of both the Drosophila and the mammalian clocks. Unexpectedly, the knockout of timeless-d, a gene essential for the clock ticking in Drosophila, did not compromise rhythmicity in P. apterus, it only accelerated its pace. Furthermore, silencing timeless-m, the ancestral timeless type ubiquitously present across animals, resulted in a mild gradual loss of rhythmicity, supporting its possible participation in the linden bug clock, which is consistent with timeless-m role suggested by research on mammalian models. The dispensability of timeless-d in P. apterus allows drawing a scenario in which the clock has remained functional at each step of transition from an ancestral state to the TIM-d-independent PER + CRY-m system operating in extant vertebrates, including humans (Kotwica-Rolinska, 2022).
Circadian clocks help animals to be active at the optimal time of the day whereby for most species the daily light-dark cycle is the most important zeitgeber for their circadian clock. In this respect, long arctic summer days are particularly challenging as light is present almost 24 h per day, and continuous light makes the circadian clocks of many animals arrhythmic. This is especially true for the fruit fly, Drosophila melanogaster, which possesses a very light-sensitive clock. The blue-light photoreceptor Cryptochrome (CRY) and the clock protein Timeless (TIM) are the light-sensitive components of the circadian clock and are responsible for constant light-induced arrhythmicity even at very low light intensities. Nevertheless, D. melanogaster was able to spread from its tropical origin and invade northern latitudes. This study tested whether a natural polymorphism at the timeless (tim) locus, s-tim and ls-tim, helped adaptation to very long photoperiods. The recently evolved natural allele, ls-tim, encodes a longer, less light sensitive form of TIM (L-TIM) in addition to the shorter (S-TIM) form, the only form encoded by the ancient s-tim allele. ls-tim has evolved in southeastern Italy and slowly spread to higher latitudes. L-TIM is known to interact less efficiently with CRY as compared with S-TIM. The locomotor activity patterns of ~40 wild s-tim and ls-tim isofemale lines caught at different latitudes was measured under simulated high-latitude summer light conditions (continuous light or long photoperiods with 20-h daily light). It was found that the ls-tim lines were significantly more rhythmic under continuous light than the s-tim lines. Importantly, the ls-tim lines can delay their evening activity under long photoperiods, a behavioral adaptation that appears to be optimal under high-latitude conditions. These observations suggest that the functional gain associated with ls-tim may drive the northern spread of this allele by directional selection (Deppisch, 2022).
Light is one of the strongest environmental time cues for entraining
endogenous circadian rhythms. Emerging evidence indicates that CREB-regulated transcription co-activator 1 (CRTC1) is a key player in this pathway, stimulating light-induced Period1
(Per1) transcription in mammalian clocks. This study demonstrates
a light-independent role of Drosophila CRTC in sustaining circadian behaviors. Genomic deletion of the crtc locus causes long but poor
locomotor rhythms in constant darkness. Overexpression or RNA
interference-mediated depletion of CRTC in circadian pacemaker neurons
similarly impairs the free-running behavioral rhythms, implying that Drosophila
clocks are sensitive to the dosage of CRTC. The crtc null
mutation delays the overall phase of circadian gene expression yet it
remarkably dampens light-independent oscillations of TIMELESS
(TIM) proteins in the clock neurons. In fact, CRTC overexpression enhances
CLOCK/CYCLE (CLK/CYC)-activated
transcription from tim but not per
promoter in clock-less S2 cells whereas CRTC depletion suppresses it.
Consistently, TIM overexpression partially but significantly rescues the
behavioral rhythms in crtc mutants. Taken together, these data
suggest that CRTC is a novel co-activator for the CLK/CYC-activated tim
transcription to coordinate molecular rhythms with circadian behaviors
over a 24-hour time-scale. The study proposes that CRTC-dependent clock mechanisms have co-evolved with selective clock genes among different species (Kim, 2016).
CREB-dependent transcription has long been implicated in different aspects of circadian gene expression. In mammalian clocks, light exposure triggers intracellular signaling pathways that activate CREB-dependent Per1 transcription, thereby adjusting the circadian phase of master circadian pacemaker neurons in the suprachiasmatic nucleus (SCN). The phase-resetting process involves the specific CREB coactivator CRTC1 and its negative regulator SIK1, constituting a negative feedback in the photic entrainment via a CREB pathway (Sakamoto, 2013; Jagannath, 2013). This report demonstrates a novel role of Drosophila CRTC that serves to coordinate circadian gene expression with 24-hour locomotor rhythms even in the absence of light. CRTC may regulate several clock-relevant genes, including those clock output genes that might be involved in the rhythmic arborizations and PDF cycling of the circadian pacemaker neurons. However, tim transcription was identified as one of the primary targets of Drosophila CRTC to sustain circadian rhythms in the free-running conditions, thus defining its light-independent clock function (Kim, 2016).
CREB could employ another transcriptional coactivator CBP (CREB-binding protein) to activate CRE-dependent transcription. In fact, CBP is a rather general coactivator recruited to gene promoters by other DNA-binding transcription factors. Previous studies have shown that Drosophila CBP associates with CLK, titrating its transcriptional activity. Mammalian CBP and the closely related coactivator p300 also form a complex with CLOCK-BMAL1, a homolog of the Drosophila CLK-CYC heterodimer, to stimulate their transcriptional activity. One possible explanation for CRTC-activated tim transcription is that Drosophila CRTC may analogously target the CLK-CYC heterodimer to stimulate CLK-CYC-tivate CRE-dependent transcription. Under these circumstances, a circadian role of light-sensitive TIM might have degenerated, while-induced clock genes. Moreover, CRTC associates with the bZIP domain in CREB protein, whereas CBP/p300 binds CREB through the phosphorylated KID domain, indicating that they might not necessarily target the same transcription factors apart from CREB. Finally, a protein complex of CLK and CRTC could not be detected in Drosophila S2 cells. Thus, it is likely that CRTC and CBP/p300 play unique roles in circadian transcription through their interactions with different DNA-binding transcription factors (Kim, 2016).
If CRTC augments CLK-CYC-dependent tim transcription indirectly, then why do crtc effects require CLK? A recent study suggested that mammalian CLOCK-BMAL1 may regulate the rhythmic access of other DNA-binding transcription factors to their target promoters in the context of chromatin, acting as a pioneer-like transcription factor. Given the structural and functional homology between Drosophila CLK-CYC and mammalian CLOCK-BMAL1, the presence of CLK-CYC in the tim promoter might allow the recruitment of additional transcription factors (e.g., CREB) and their co-activators including CRTC for maximal tim transcription. The transcriptional context of tim promoter might thus define its sensitivity to crtc effects among other clock promoters. In addition, the differential assembly of transcription factors on the tim promoter could explain tissue-specific effects of crtc on TIM oscillations (i.e., circadian pacemaker neurons versus peripheral clock tissues). Interestingly, chromatin immunoprecipitation with V5-tagged CLK protein revealed that CLK-CYC heterodimers associate with both tim and Sik2 gene promoters in fly heads. In LD cycles, however, their rhythmic binding to the Sik2 promoter is phase-delayed by ~4.5 hours compared with that to the tim promoter. These modes of transcriptional regulation may gate crtc effects on tim transcription in a clock-dependent manner, particularly in the increasing phase of tim transcription (Kim, 2016).
Transcription from CREB-responsive reporter genes shows daily oscillations, both in Drosophila and mammals, implicating this transcriptional strategy in the evolution of molecular clocks. In fact, cAMP signaling and CRE-dependent transcription constitute the integral components of core molecular clocks, serving to regulate daily rhythmic transcription of circadian clock genes. For instance, reciprocal regulation of dCREB2 and per at the transcription level has been reported to sustain free-running circadian rhythms in Drosophila. During fasting in mammals, a transcriptional program for hepatic gluconeogenesis is induced by CREB phosphorylation and CRTC2 dephosphorylation. Fasting-activated CREB-CRTC2 then stimulates Bmal1 expression61, whereas CLOCK-BMAL1-induced CRY rhythmically gates CREB activity in this process by modulating G protein-coupled receptor activity and inhibiting cAMP-induced CREB phosphorylation62. This molecular feedback circuit thus mutually links mammalian clocks and energy metabolism in terms of CREB-dependent transcription (Kim, 2016).
On the basis of these observations, a model is proposed for the evolution of CRTC-dependent clocks to explain the distinctive circadian roles of CRTC homologs (see A model for the evolution of CRTC-dependent clocks). CRTC is a transcriptional effector that integrates various cellular signals (Altarejos, 2011). It was reasoned that ancestral clocks may have employed CREB-CRTC-mediated transcription to sense extracellular time cues cell-autonomously and integrate this timing information directly into the earliest transcription-translation feedback loop (TTFL). This strategy would have generated simple but efficient molecular clocks to tune free-running molecular rhythms in direct response to environmental zeitgebers, such as light and the availability of nutrients. A circadian role of CRTC then has differentially evolved along with a selective set of clock targets. In poikilothermic Drosophila, light is accessible directly to circadian pacemaker neurons in the adult fly brain. Therefore, TIM degradation by the blue-light photoreceptor CRY plays a major role in the light entrainment of Drosophila clocks, although the photic induction of CLK/CYC-dependent tim transcription has been reported specifically at lower temperatures. Accordingly, Drosophila CRTC retained a constitutive co-activator function from the ancestral TTFL to support CLK/CYC-activated tim transcription and sustain free-running circadian behaviors. In homeothermic mammals, light input to the SCN is indirectly mediated by neurotransmitter release from presynaptic termini of the retinohypothalamic tract (RHT). Intracellular signaling relays in the SCN converge on the dephosphorylation and nuclear translocation of CRTC1 to activate CRE-dependent transcription. Under these circumstances, a circadian role of light-sensitive TIM might have degenerated, while per took over a role in the light-entrainment pathway by retaining CREB-CRTC1-dependent transcriptional regulation from the primitive TTFL. Consequently, mammalian clocks have lost a homolog of the Drosophila-like cry gene family, but instead evolved CRY homologs of the vertebrate-like cry gene family with transcriptional repressor activities in CLOCK-BMAL1-dependent transcription (Kim, 2016).
Regulation of metabolism and stress responses by neuronal CREB-CRTC-SIK pathways has been well documented in Drosophila. Given the demonstration of a circadian role of CRTC in the pacemaker neurons, it is possible that CRTC might sense metabolic cues in the context of circadian neural circuits to entrain molecular clocks cell-autonomously. Alternatively, but not exclusively, CRTC could participate in the regulation of clock-relevant metabolism as clock outputs from pacemaker neurons. These hypotheses remain to be validated in future studies (Kim, 2016).
Circadian clocks enable organisms to anticipate daily
changes in the environment and coordinate temporal rhythms in
physiology and behavior with the 24-hour day-night cycle. The robust
cycling of circadian gene expression is critical for proper
timekeeping, and is regulated by transcription factor binding, RNA
polymerase II (RNAPII) recruitment and elongation, and
post-transcriptional mechanisms. Recently, it has become clear that
dynamic alterations in chromatin
landscape at the level of histone posttranslational modification
and nucleosome density facilitate rhythms in transcription factor
recruitment and RNAPII activity, and are essential for progression
through activating and repressive phases of circadian transcription.
This study discusses the characterization of the Brahma
(Brm) chromatin-remodeling protein in Drosophila in the context of
circadian clock regulation. By dissecting its catalytic vs.
non-catalytic activities, a model is proposed in which the non-catalytic
activity of Brm functions to recruit repressive factors to limit the
transcriptional output of Clock (Clk) during the active phase of
circadian transcription, while the primary function of the ATP-dependent
catalytic activity is to tune and prevent over-recruitment of negative
regulators by increasing nucleosome density. Finally, ongoing efforts
and investigative directions towards a deeper mechanistic understanding
of transcriptional regulation of circadian gene expression at the
chromatin level are described (Kwok, 2016).
The timing of behavior under natural light-dark conditions is a function of circadian clocks and photic input pathways, but a mechanistic understanding of how these pathways collaborate in animals is lacking. This study demonstrates in Drosophila that the Phosphatase of Regenerating Liver-1 (PRL-1) sets period length and behavioral phase gated by photic signals. PRL-1 knockdown in PDF clock neurons dramatically lengthens circadian period. PRL-1 mutants exhibit allele-specific interactions with the light and clock-regulated gene timeless (tim). Moreover, this study shows that PRL-1 promotes TIM accumulation and dephosphorylation. Interestingly, the PRL-1 mutant period lengthening is suppressed in constant light, and PRL-1 mutants display a delayed phase under short, but not long, photoperiod conditions. Thus, these studies reveal that PRL-1-dependent dephosphorylation of TIM is a core mechanism of the clock that sets period length and phase in darkness, enabling the behavioral adjustment to change day-night cycles (Kula-Eversole, 2020).
This study has identified a novel circadian clock gene PRL-1 that mediates the behavioral response to changes in photoperiod. Circadian regulation of this phosphatase provides a mechanism for mediating rhythmic phosphorylation, a widely conserved feature of circadian clocks. Reduction or loss-of-function in PRL-1 can induce potent effects on circadian period length even larger than those caused by the reduction of canonical clock genes. These effects are widely conserved and are evident even in human cells. PRL-1 selectively dephosphorylates and suppresses the accumulation of the light-sensitive clock component TIM. PRL-1 affects behavioral phase in short but not long photoperiods, revealing a novel mechanism for appropriate timing across ecologically relevant photoperiods (Kula-Eversole, 2020).
The data suggest PRL-1 plays an especially important role in setting period length comparable to or exceeding those of the canonical core clock components. RNAi knockdown of PRL-1 with two independent RNAi lines results in among the strongest RNAi-induced period-altering phenotypes seen in Drosophila with period lengths exceeding 28 h. Interestingly, the period effects observed through PRL-1 knockdown in the PDF-positive LNv (pdfGAL4) are larger than those observed with a broader circadian driver (timGAL4) or in the putative null allele of PRL-1, suggesting that PRL-1 may have distinct and even antagonistic period setting roles in different clock neurons. The other clock genes with RNAi effects comparable to PRL-1 are the regulatory beta subunit of the protein kinase CK2 and the E3 ubiquitin ligase circadian trip, further highlighting the role of post-translation modifications in determining period length (Kula-Eversole, 2020).
Several lines of evidence support the model that TIM is an in vivo target of PRL-1. PRL-1 mutants exhibit non-additive effects with a tim allele, timS1, suggesting PRL-1 and tim function within the same pathway. Non-additive genetic interactions with specific alleles can reflect functional, even direct biochemical, interactions such as between per and tim78 and between per and Dbt. Simple additive effects on period length with timUL flies probably reflect the fact that TIMUL is stable and exhibits prolonged nuclear localization; therefore, stabilizing effects of timUL are not counteracted by the destabilizing effects of PRL-1 mutation during the rising phase of TIM accumulation. Loss of PRL-1 results in substantially reduced and delayed nuclear accumulation of TIM during the first day of DD. These effects are much larger than those simply predicted based on the lengthened period and much larger than effects on PER, suggesting a selective effect on TIM in vivo. Using PRL-1 overexpression and knockdown in clock-less Drosophila S2 cells, it was found that PRL-1 dephosphorylates TIM but not PER. Given that PRL-1 mRNA expression peaks around dusk, it is proposed that PRL-1 accumulation during the early night results in TIM dephosphorylation, stabilization, and nuclear localization (Kula-Eversole, 2020).
Strikingly, period lengthening is evident even after siRNA knockdown of all three highly conserved human orthologs of PRL-1, PTP4a-1, -2, and -3 in human U2OS cells, suggesting a widely conserved role in period determination. Like fly PRL-1, mammalian orthologs of PRL-1 also undergo rhythmic expression in many tissues. PTPs may also function via TIM. Mammalian TIM interacts with CRY1 as well as PER1/2, is involved in CLOCK-BMAL1 repression of CLOCK-BMAL1 activation, and sets period length. Mutation of human TIMELESS also results in familial advanced sleep phase syndrome perhaps via altered light responses. Thus, PTP4 action may similarly function to regulate TIM in mammals (Kula-Eversole, 2020).
The connection between PRL-1 and photosensitive TIM suggested that PRL-1 may function to mediate clock responses to light. PRL-1 mutants exhibit a light-dependent and CRY-independent period phenotype. To examine rhythms in LL, PRL-1 mutant period length was examined in a cry mutant background in which flies exhibit free-running rhythms. These flies retain their long periods in DD. On the other hand, in LL they exhibit period lengths comparable to their PRL-1+ controls. Thus, light can act independently of CRY photoreception to compensate for the PRL-1 mutant period lengthening. It is hypothesized that signaling through the visual system could accomplish this (Kula-Eversole, 2020).
The finding of LL-dependent period phenotypes led to an examination of the role of PRL-1 in mediating behavioral adaptations to seasonal changes in photoperiod. PRL-1 mutants display delayed morning and evening activity phases under short (6:18) photoperiods. In contrast, they have no alteration of evening activity phase under long (18:6) photoperiods. These photoperiods approximate those that would be experienced by wild Drosophila melanogaster at more extreme latitudes. It is hypothesized that PRL-1 defines a molecular pathway through which the clock adjusts behavioral phase in response to different photoperiods (Kula-Eversole, 2020).
The phenomenon of photoperiod-dependent network hierarchy suggests that M and E cells differ in their core clock properties, responses to light, and/or their network connectivity. It is hypothesized that PRL-1 represents such a specialization of M cell clocks that enables the appropriate behavioral adjustments to different photoperiods. PRL-1 exhibits robust PDF neuron specific cycling, and PRL-1 knockdown selectively in PDF neurons substantially lengthens period. The finding of period phenotypes in DD and not LL is also consistent with M cell specific function. The findings of photoperiod-dependent effects in PRL-1 mutants could reflect enhanced coupling between LNv and CRY-negative DN1p in short photoperiods, consistent with prior findings of light intensity-dependent sLNv-DN1p coupling. Under this model, selective PRL-1-dependent changes in the M cell phase are propagated to E cells in a photoperiod-dependent manner as observed for other M cell-specific changes. Of note, other core clock components have been identified with differential functions between M and E cells, including those involved in TIM phosphorylation, suggesting a wider role for specialized core clock components in M and E cells. It will be of interest to determine if M or E cell-specific mechanisms also function to control behavioral adaptations to different photoperiods and, if so, how they work with respect to PRL-1 (Kula-Eversole, 2020).
Sensitivity of the circadian clock to light/dark cycles ensures that biological rhythms maintain optimal phase relationships with the external day. In animals, the circadian clock neuron network (CCNN) driving sleep/activity rhythms receives light input from multiple photoreceptors, but how these photoreceptors modulate CCNN components is not well understood. This study shows that the Hofbauer-Buchner eyelets, located between the retina and the medulla in the fly optic lobes, differentially modulate two classes of ventral lateral neurons (LNvs) within the Drosophila CCNN. The eyelets antagonize Cryptochrome (CRY)- and compound-eye-based photoreception in the large LNvs while synergizing CRY-mediated photoreception in the small LNvs. Furthermore, it was shown that the large LNvs interact with subsets of 'evening cells' to adjust the timing of the evening peak of activity in a day length-dependent manner. This work identifies a peptidergic connection between the large LNvs and a group of evening cells that is critical for the seasonal adjustment of circadian rhythms (Schlichting, 2016).
Circadian clocks create an endogenous sense of time that is used to produce daily rhythms in physiology and behavior. A defining characteristic of a circadian clock is a modest deviation of its endogenous period from the 24.0 h period of daily environmental change. For example, the average human clock has an endogenous period of 24 h and 11 min. Thus, to maintain a consistent phase relationship with the environment, the human clock must be sped up by 11 min every day. A sensitivity of the circadian clock to environmental time cues (zeitgebers) ensures that circadian clocks are adjusted daily to match the period of environmental change. This process, called entrainment, is fundamental to the proper daily timing of behavior and physiology. For most organisms, daily light/dark (LD) cycles are the most salient zeitgeber (Schlichting, 2016).
Although most tissues express molecular circadian clocks in animals, the clock is required in small islands of neural tissue for the presence of sleep/activity rhythms and many other daily rhythms in physiology. Within these islands, a circadian clock neuron network (CCNN) functions as the master circadian clock. Subsets of neurons within the CCNN receive resetting signals from photoreceptors, and physiological connections between these neurons and their clock neuron targets ensure light entrainment of the CCNN as a whole (Schlichting, 2016).
In both mammals and insects, the CCNN receives light input from multiple photoreceptor types. In Drosophila, the CCNN is entrained by photoreceptors in the compound eye, the ocelli, the Hofbauer-Buchner (HB) eyelets, and by subsets of clock neurons that express the blue light photoreceptor Cryptochrome (CRY). Understanding how multiple light input pathways modulate the CCNN to ensure entrainment to the environmental LD cycle is critical for understanding of the circadian system and its dysfunction when exposed to the unnatural light regimens accompanying much of modern life (Schlichting, 2016).
This study investigates the physiological basis and circadian role of a long-suspected circadian light input pathway in Drosophila: the HB eyelets. These simple accessory eyes contain four photoreceptors located at the posterior edges of the compound eyes and project directly to the accessory medullae (AMe), neuropils that support circadian timekeeping in insects. In Drosophila, the AMe contain projections from ventral lateral neurons (LNvs), important components of the CCNN that express the neuropeptide pigment dispersing factor (PDF), an output required for robust circadian rhythms in locomotor activity. The axon terminals of the HB eyelets terminate near PDF-positive LNv projections and analysis of visual system and cry mutants reveals a role for the HB eyelet in the entrainment of locomotor rhythms to LD cycles, but how the eyelets influence the CCNN to support light entrainment is not well understood (Schlichting, 2016).
This study presents evidence that this circadian light input pathway excites the small LNvs (s-LNvs) and acts to phase-dependently advance free-running rhythms in sleep/activity while inhibiting the large LNvs (l-LNvs). This work reveals that input from external photoreceptors differentially affects specific centers within the fly CCNN. Furthermore, it was shown that, under long summer-like days, the l-LNvs act to modulate subsets of so-called evening cells to delay the onset of evening activity. These results reveal a neural network underlying the photoperiodic adjustment of sleep and activity (Schlichting, 2016).
The experiments described in this study lead to two unexpected findings regarding the network properties of circadian entrainment in Drosophila. First, the l-LNvs govern the phase of evening peak of activity through PdfR-dependent effects on evening cells that bypass the s-LNvs. Although previous work has implicated the l-LNvs in the control of evening peak phase, the current results are the first to provide evidence that there is a direct connection between the l-LNvs and evening cells within the AMe and that this connection mediates the photoperiodic adjustment of sleep and activity in the fly. Second, the HB eyelets light input pathways, long implicated in circadian entrainment, have opposing effects on the l-LNvs and s-LNvs, inhibiting the former and exciting the latter. These results reveal not only a differential effect of a light input pathway on specific nodes of the CCNN but also establish that light from extraretinal photoreceptors can have synergistic or antagonistic effects on CRY- and compound eye-mediated light responses, depending on the clock neuron target in question (Schlichting, 2016).
Both the l-LNvs and s-LNvs express the blue light circadian photoreceptor CRY, the expression of which renders neurons directly excitable by light entering the brain through the cuticle. How such CRY-mediated light input interacts with input from external photoreceptors is not well understood, although it is known that each system alone is sufficient for the entrainment of locomotor rhythms. Genetic evidence suggests that the HB eyelets have relatively weak effects on circadian entrainment: flies with functional eyelets that lack compound eyes, ocelli, and CRY entrain relatively poorly to LD cycles relative to flies with functional eyes or CRY. The small phase responses of locomotor rhythms to HB eyelet excitation further supports a relatively weak effect of the eyelet on free-running locomotor rhythms (Schlichting, 2016).
The LNvs are critical nodes in the CCNN and are closely associated with input pathways linking the central brain to external photoreceptors. Work on the LNvs has provided evidence for a division of labor among the l-LNvs and s-LNvs: the l-LNvs are wake-promoting neurons that acutely govern arousal and sleep independently of the s-LNvs, whereas the s-LNvs act as key coordinators of the CCNN to support robust circadian timekeeping. Anatomical and genetic evidence has long supported the notion that the dorsal projections of the s-LNvs represent the key connection between the LNvs and the remaining components of the CCNN. However, a smaller body of work has suggested that the l-LNvs also contribute to the entrainment of sleep/activity rhythms under LD cycles. The Pdf knockdown and PdfR rescue experiments under long day conditions indicate that, as the day grows longer, the l-LNvs play a greater role in the timing of the evening peak. Moreover, the effects of PDF released from the l-LNvs are mediated not by the PDF receptive s-LNvs but rather by the fifth s-LNvs and a subset of the LNds, the NPF and ITP coexpressing LNds in particular (with some influence of the other PDF-receptor positive LNds). These same neurons were recently identified as evening cells that are physiologically responsive to PDF but relatively weakly coupled to LNv clocks under conditions of constant darkness. The results suggest that the l-LNvs differentially modulate the NPF/ITP-positive evening oscillators as a function of day length, producing stronger PDF-dependent delays under long day conditions through increased release of PDF from the l-LNvs, thereby delaying the evening activity peak. Thus, the l-LNvs mediate their effects on the evening peak of activity through their action on the NPF/ITP-positive subset of evening oscillators. The proposed PDF release from the l-LNvs under long days requires their activation via CRY and/or the compound eyes via ACh release from lamina L2 interneurons. It is hypothesized that the inhibitory influence of the HB eyelets ceases under long days allowing the compound eyes and CRY to maximally excite the l-LNvs. Indeed, previous work has established that the compound eyes are especially important for adapting fly evening activity to long days. Furthermore, several studies have suggested that the compound eyes signal to the l-LNvs leading to enhanced PDF release and a slowing-down of the evening oscillators. A recent paper measuring Ca2+ rhythms in the different clock neurons in vivo supports this view (Liang, 2016): Ca2+ rhythms in the l-LNvs peak in the middle of the day, unlike the s-LNvs, which display Ca2+ peaks in the late night/early morning. It is suggested that this phasing is produced by the inhibition of l-LNvs by the eyelets in the morning, followed by the excitation of the l-LNvs by the compound eyes and CRY. Interestingly, the only other clock neuron classes to display Ca2+ increases during the day are the LNds and fifth-sLNv, which phase lag the l-LNvs by ~2.5 h and display peak Ca2+ levels in the late afternoon, a time that coincides with the evening peak of activity (Liang, 2016). It is proposed that the relative coordination of Ca2+ rhythms between the l-LNvs and the LNds/fifth-sLNv is produced by the connection this study has identified between these neurons and the action of the eyelet and visual system on the l-LNvs (Schlichting, 2016).
Recent work has revealed that evening activity is promoted directly by the evening oscillator neurons and that the mid-day siesta is produced by the daily inhibition of evening oscillators by a group of dorsal clock neurons (Guo, 2016). It is proposed that the connections described in this study govern the timing of the evening peak of activity through the PDF-dependent modulation of the molecular clocks within the evening oscillator neurons, although PDF modulation likely results in the excitation of target neurons, which would promote evening activity. The results reveal new and unexpected network properties underlying the entrainment of the circadian clock neuron network to LD cycles. Excitatory effects of light on the LNvs are differentially modulated by the HB eyelets via cholinergic excitation of the s-LNvs and histaminergic inhibition of the l-LNvs. The work further reveals PDF-dependent modulatory connections in the AMe between the l-LNvs and the s-LNvs and, most surprisingly, between the l-LNvs and a small subset of evening oscillators. This work indicates that the latter connection is critical for the adjustment of evening activity phase during long, summer-like days. This network model of entrainment reveals not only how CRY and external photoreceptors interact within specific nodes of the CCNN, but also how photoreception is likely to drive changes in CCNN output in the face of changing day length (Schlichting, 2016).
The light-input factor Quasimodo (Qsm) regulates rhythmic electrical excitability of clock neurons, presumably via an Na+, K+, Cl- cotransporter (NKCC) and the Shaw K+ channel (dKV3.1). Because of light-dependent degradation of the clock protein Timeless (Tim), constant illumination (LL) leads to a breakdown of molecular and behavioral rhythms. Both overexpression (OX) and knockdown (RNAi) of qsm, NKCC, or Shaw led to robust LL rhythmicity. Whole-cell recordings of the large ventral lateral neurons (l-LNv) showed that altering Qsm levels reduced the daily variation in neuronal activity: qsmOX led to a constitutive less active, night-like state, and qsmRNAi led to a more active, day-like state. Qsm also affected daily changes in K+ currents and the GABA reversal potential, suggesting a role in modifying membrane currents and GABA responses in a daily fashion, potentially modulating light arousal and input to the clock. When directly challenged with blue light, wild-type l-LNvs responded with increased firing at night and no net response during the day, whereas altering Qsm, NKKC, or Shaw levels abolished these day/night differences. Finally, coexpression of ShawOX and NKCCRNAi in a qsm mutant background restored LL-induced behavioral arrhythmicity and wild-type neuronal activity patterns, suggesting that the three genes operate in the same pathway. It is proposed that Qsm affects both daily and acute light effects in l-LNvs probably acting on Shaw and NKCC (Buhl, 2016).
All organisms are subject to predictable but drastic daily environmental changes caused by the earth's rotation around the sun. It is critical for the fitness and well-being of an individual to anticipate these changes, and this anticipation is done by circadian timekeeping systems (clocks). These regulate changes in behavior, physiology, and metabolism to ensure they occur at certain times during the day, thereby adapting the organism to its environment. The circadian system consists of three elements: the circadian clock to keep time, inputs that allow entrainment, and outputs that influence physiology and behavior. Like a normal clock, circadian clocks run at a steady pace (24 h) and can be reset. In nature this environmental synchronization is done via daily light and temperature cycles, food intake, and social interactions (Buhl, 2016). In Drosophila the central clock comprises 75 neuron pairs grouped into identifiable clusters that subserve different circadian functions. The molecular basis of the circadian clock is remarkably conserved from Drosophila to mammals. This intracellular molecular clock drives clock neurons to express circadian rhythms in electrical excitability, including variation in membrane potential and spike firing. Clock neurons are depolarized and fire more during the day than at night, and circadian changes in the expression of clock-controlled genes encoding membrane proteins such as ion channels and transporters likely contribute to these rhythms. Such cyclical variations in activity play a critical role in synchronizing different clock neurons and conveying circadian signals to other parts of the nervous system and body. Furthermore, they provide positive feedback to the molecular clock, which can dampen rapidly without such feedback (Buhl, 2016).
Light resets the circadian clock every morning to synchronize the clock to the environment via Timeless (Tim) degradation after activation of the blue-light photoreceptor Cryptochrome (Cry), Quasimodo (Qsm), and potentially also visual photoreceptors. Qsm acts either independently or downstream of Cry and also is able to affect clock protein stability in Qsm-negative neurons by an unknown non-cell-autonomous mechanism (Chen, 2011). Recently Cry has been shown to regulate clock neuron excitability via the redox sensor of the Hyperkinetic voltage-gated potassium (KV)-β subunit (Hk) (Fogle, 2015), and this study asked if Qsm affects the clock neurons in a similar way (Buhl, 2016).
Membrane potential is important for control of circadian behavior, and manipulation of Shaw and the Narrow Abdomen (NA) channels, both of which are expressed and function within clock neurons influence neuronal electrical activity, the circadian clock, and clock-controlled behavior in both flies and mice. The firing rate is a key component in mammalian circadian rhythmicity and can be regulated by regional and circadian expression of the sodium potassium chloride cotransporter NKCC, which switches the effects of GABA from inhibitory to excitatory across the day (Buhl, 2016).
This study shows that down-regulation or overexpression of the three membrane proteins encoded by the genes qsm, Shaw, and NKCC leads to rhythmicity in constant illumination (LL) and that these genes interact. All three genes are expressed in the well-characterized pigment-dispersing factor (Pdf)- and Cry-positive large ventral lateral neurons (l-LNv), which are important for arousal and light input to the clock. Whole-cell recordings of l-LNvs were used to characterize their physiological properties and acute light effects across the day, and Qsm was found to help set the circadian state of clock neurons and modify their response to light, possibly by acting via Shaw and NKCC (Buhl, 2016).
Light is the dominant circadian zeitgeber that resets the molecular clock. This study determined how light affects membrane excitability via the membrane proteins Qsm, Shaw, and NKCC. Previously it was shown that Qsm contributes to circadian clock light input with down-regulation in all clock neurons (tim-gal4) resulting in robust rhythmic behavior in LL (Chen, 2011). This study shows that overexpression (qsmOX) also results in robust LL rhythmicity, but with predominantly ~13-h periods, suggesting a more robust morning oscillator that is normally weakened in DD conditions. Manipulating the expression levels of both the potassium channel Shaw and the ion cotransporter NKCC also resulted in LL rhythmicity, whereas several visual system mutants behaved like wild-type flies and became arrhythmic. These experiments show that the membrane proteins encoded by qsm, Shaw, and NKCC control rhythmic behavior in LL. Furthermore, the rescue of wild-type behavior and neurophysiological properties by reciprocal changes of Qsm and Shaw and by the simultaneous reduction of Qsm and NKCC suggests that Qsm interacts genetically and perhaps directly with Shaw and NKCC (Buhl, 2016).
Clock neurons are more depolarized and fire more during the daytime, and circadian changes in the expression of clock-controlled genes such as ion channels and transporters are likely to play a part. Contributing to this rhythm is a sodium leak current mediated by NA that recently has been shown to depolarize Drosophila clock neurons. This study shows that Shaw, NKCC, and Qsm also contribute to daily electrical activity rhythms: overexpression and RNAi knock-down of qsm and Shaw compared with NKCC resulted in opposing phenotypes. Interestingly, qsmOX or ShawOX and NKCCRNAi promote the less active nighttime state, whereas qsmRNAi, ShawRNAi, and NKCCOX push the neurons into the more depolarized daytime state, eliminating the acute day/night differences in all cases. Previous work has shown that Shaw regulates circadian behavior and that, in agreement with the current findings, Shaw regulates membrane potential and firing in Drosophila motoneurons. NKCC activity is electrically neutral but increases the intracellular Cl- concentration so that the GABAA receptor opens in response to GABA but, as a consequence, Cl- presumably exits the cell down its electrochemical gradient, thereby depolarizing the membrane potential so that GABA effectively becomes an excitatory neurotransmitter. The current data show that in Drosophila a similar mechanism occurs, which is consistent with potential NKCC enrichment in l-LNv at dawn. The mechanism setting the neuronal state to either daytime or nighttime via Qsm, Shaw, and NKCC is likely to be predominantly cell-autonomous, because all components have been shown to act or to be expressed in the l-LNv. Although in an earlier study using qsm-gal4 lines qsm expression was not detected in the l-LNv, these lines may not report expression faithfully in all qsm cells. Now, the finding that qsm RNA is enriched in the l-LNv, combined with the strong effects of two qsm-RNAi lines on l-LNv electrical properties presented in this study, indicates that qsm is endogenously expressed in these neurons (Buhl, 2016).
Physiological studies are limited to the Pdf-expressing l-LNv neurons. These neurons are unlikely candidates for driving behavioral rhythms in LL, and previous work has shown that qsm knockdown in Pdf neurons (s-LNv and l-LNv) does not result in robust LL rhythmicity. Therefore the effects of light on the electrical properties of l-LNv reported here do not necessarily explain the LL rhythmicity observed after manipulating qsm, Shaw, and NKCC in all clock neurons. However, the electrophysiological results using tim-gal4 show that Qsm, Shaw, and NKCC could fulfill similar functions in other clock neurons, including those crucial for LL rhythmicity (e.g., LNd and DN1). Additionally, or alternatively, the manipulated l-LNv could generate signals interfering with normal network function, resulting in the observed rhythmic LL behavior (Buhl, 2016).
Although qsm is a clock-controlled gene, the acute blue-light effects that were observed are too fast to be mediated by transcriptional changes. Therefore, a more direct membrane-localized mechanism is favored in which rapid light-dependent posttranslational changes of Qsm alter the activity of Shaw and NKCC. Because (i) Cry is required for light-dependent Tim degradation in l-LNv, (ii) changing the Qsm level has no effect on Cry levels, and (iii) qsmOX triggers Tim degradation in the absence of Cry, the most likely explanation for the results reported in this study is that, in addition to activating Hk, Cry acts upstream of Qsm, which in turn regulates the activity of Shaw and NKCC. It is assumed that Qsm is activated by light because a light pulse at night rapidly increases protein levels. Qsm is an extracellular zona-pellucida (ZP) membrane-anchored protein, and it is hypothesized that after light exposure the extracellular ZP domain is cleaved at a conserved furin protease cleavage site, a form of posttranslational processing typical for ZP-domain proteins. It is also possible that Qsm signals to Shaw and NKCC in both membrane-bound and cleaved forms. For example, at night membrane-bound Qsm could block NKCC, whereas light-induced cleavage could release this block, and the freed extracellular part could inactivate Shaw. This mechanism is reminiscent to the mechanism by which the GPI-anchored extracellular protein Sleepless increases Shaker (KV1 channel) activity for regulating Drosophila sleep (Buhl, 2016).
How Qsm-induced changes in clock neuron activity influence the molecular clock remains an open question. Recent work shows that, in addition to the canonical degradation via Cry and Jetlag, Tim is also degraded via a Cul-3 and neuronal activity-dependent pathway in DD that has been implicated in mediating phase delays in the circadian clock. In contrast to this activity-dependent Cul-3 pathway, the light responses in the current study depend on Cry. Therefore a model is favored in which the combined functions of Qsm, Shaw, and NKCC contribute to the canonical Cry- and Jetlag-dependent Tim-degradation pathway (Buhl, 2016).
In conclusion, this study demonstrates that Qsm affects both daily and acute light responses of l-LNvs, and therefore (Qsm) presumably contributes to light-input to the Drosophila circadian clock. Qsm possibly signals downstream of Cry and acts on Shaw and NKCC to change clock neuronal activity in response to light (Buhl, 2016).
Light at night disrupts the circadian clock and causes serious health problems in the modern world. This study shows that newly developed four-package light-emitting diodes (LEDs) can provide harmless lighting at night. To quantify the effects of light on the circadian clock, the concept of circadian illuminance (CIL) was employed. CIL represents the amount of light weighted toward the wavelengths to which the circadian clock is most sensitive, whereas visual illuminance (VIL) represents the total amount of visible light. Exposure to 12 h:12 h cycles of white LED light with high and low CIL values but a constant VIL value (conditions hereafter referred to as CH/CL) can entrain behavioral and molecular circadian rhythms in flies. Moreover, flies re-entrain to phase shift in the CH/CL cycle. Core-clock proteins are required for the rhythmic behaviors seen with this LED lighting scheme. Taken together, this study provides a guide for designing healthful white LED lights for use at night, and proposes the use of the CIL value for estimating the harmful effects of any light source on organismal health (Cho, 2016).
In many animals the circadian rhythm of locomotor activity is controlled by an endogenous circadian clock. Using custom made housing and video tracking software in order to obtain high spatial and temporal resolution, wthe statistical properties of the locomotor activity of wild type and two clock mutants of Drosophila melanogaster were studied. This study showed that the distributions of activity and quiescence bouts for the clock mutants in light-dark conditions (LD) are very different from the distributions obtained when there are no external cues from the environment (DD). In the wild type these distributions are very similar, showing that the clock controls this aspect of behavior in both regimes (LD and DD). Furthermore, the distributions are very similar to those reported for Wistar rats. For the timing of events important differences were observed, quantified by how the event rate distributions scale for increasing time windows. For the wild type these distributions can be rescaled by the same function in DD as in LD. Interestingly, the same function has been shown to rescale the rate distributions in Wistar rats. On the other hand, for the clock mutants it is not possible to rescale the rate distributions, which might indicate that the extent of circadian control depends on the statistical properties of activity and quiescence (Cascallares, 2018).
Behavioral responsiveness to external stimulation is shaped by context. This study examined how sensory information can be contextualized, by examining light-evoked locomotor responsiveness of Drosophila relative to time of day. Light elicits an acute increase in locomotion (startle) that is modulated in a time-of-day-dependent manner: startle is potentiated during the nighttime, when light is unexpected, but is suppressed during the daytime. The internal daytime-nighttime context is generated by two interconnected and functionally opposing populations of circadian neurons-LNvs generating the daytime state and DN1 as generating the nighttime state. Switching between the two states requires daily remodeling of LNv and DN1a axons such that the maximum presynaptic area in one population coincides with the minimum in the other. It is proposed that a dynamic model of environmental light resides in the shifting connectivities of the LNv-DN1a circuit, which helps animals evaluate ongoing conditions and choose a behavioral response (Song, 2021).
Alterations of neuronal activity, rather than morphology, are usually considered the cause of cognitive flexibility. The mechanism that this study describes relies on physical cellular restructuring. What are the advantages of a system like this? While near-instantaneous electrical activity is the basic language of neurons, many behaviors and internal states occur on much longer time scales. Morphological remodeling is a slow process, aligning with functions that change over the course of hours. In support of this view, changes in neuronal morphology have been found to underlie appetite, sexual experience, and foraging history. Although seemingly wasteful, physical remodeling may be particularly useful for encoding relatively stable states, due to presumably high energetic barrier (Song, 2021).
Understanding the mechanisms of circuit state transitions may help clarify the etiology of mood disorders like depression or bipolar disorder. These disorders are characterized by excessive or insufficient transitions between extreme states and, as such, may reflect a collapse of organizational principles that permit flexible circuit function. It is generally unknown how behavioral states can be stable across long time scales while also being able to undergo flexible transitions. Motifs from the LNv-DN1a circuit illustrate one solution to this apparent contradiction. LNvs and DN1as are arranged in a mutually inhibitory system, which may help ensure stability, consistency, and accuracy over long time scales. In the absence of external influences, reciprocal inhibition can stabilize a winner-take-all steady state. Structural plasticity is a potential way to overcome rigidity by providing a molecular mechanism to surmount electrical inhibition (Song, 2021).
In this model, the molecular oscillations of the circadian clock direct oscillations in cell shape through molecular effectors such as Rho1, ultimately leading to changes in behavior. Rho1 overexpression experiments suggest that neuronal morphology causally influences behavior. However, it cannot be ruled out that high levels of Rho1 cause off-target effects that have not been accounted for. Previous work used conditional methods to determine that Rho1 regulates daily LNv remodeling rhythms, but this study used constitutive Rho1 overexpression (to avoid using high temperatures required for the conditional experiment), and thus cannot exclude the possibility of developmental confounds (Song, 2021).
It is possible, if not likely, that other circadian neuronal populations also contribute to generating predictions about light. In this study, the optogenetic silencing phenotypes of LNvs and DN1as together recapitulate the phenotype produced by network-wide loss of clock function. However, the native circuit signal may be built by the cooperative action of multiple subpopulations with overlapping tuning. Hints of subpopulation cooperativity are apparent in the data: there was a stronger phenotype when PDF was knocked down in both small and large LNvs, compared to small LNvs alone. Glycine was recently found to be a fast, inhibitory neurotransmitter that is co-released from s-LNvs in addition to the neuropeptide PDF. While a necessary and sufficient role for the neuropeptide PDF in the behavioral assay, it cannot be ruled out that glycine instead mediates the fast inhibition seen during ex vivo calcium imaging. Furthermore, this study focused on two time points, but other populations may be more influential during other times of day. In support of this theory, it was recently found that lateral dorsal neurons also show axon remodeling rhythms centered around the evening, a time point that this study did not address. Because the other clock neuron subpopulations are active at different times, and recurrent inhibition is prevalent in the fly circadian network, it is likely that LNv-DN1a interactions are necessary, but not sufficient, for circadian network operations in the context described in this study (Song, 2021).
What this study calls 'startle' is commonly referred to as 'masking' because acute reactivity to lights-on and lights-off can distort measurements of circadian rhythmicity. For this reason, most assays of circadian function are done in unchanging environmental conditions (e.g., constant darkness). This study showed that startle to lights-on can also be an informative indicator of internal timekeeping. The conditional experiments using optogenetics suggest that clock neuron activity contextualizes light on an ongoing basis. However, because the experimental animals were exposed to daytime light during the habituation period, the possibility that LNv and DN1a activity is also required during entrainment cannot be excluded. Another consideration is that light resets the molecular clock. One hour of nighttime light can advance or delay circadian rhythms, which is apparent in the timing of rest and activity on subsequent days. It is unknown if this phase-shifting phenomenon affects the acute reactivity observed in this study (Song, 2021).
Circadian clocks have evolved in the context of consistent light schedules, and this predictability has been relatively unchallenged across evolutionary history. Ubiquitous artificial lighting introduces new strains on the circadian system. Misalignment between internal rhythms and the external world can have profound consequences on health and cognition. The feeling of jetlag demonstrates the acute physical and mental burden of when internal clocks are in conflict with the external world. Chronic misalignment, such as in the case of night-shift workers, causes increased rates of cancer. The use of electronic devices before bedtime has been linked to delays in sleep onset and reductions in sleep quality. The system this study described in Drosophila presents a model to understand the acute consequences of circadian misalignment (Song, 2021).
A predictive nervous system enables continual evaluation of reality relative to context. One result of this is that a fixed stimulus can evoke a multitude of behaviors depending on an animal's history, needs, and external context. This study shows how the Drosophila circadian system creates a dynamic internal reference for what environmental conditions should be. Many of the motifs observed are conserved: Mammalian circadian clock neuron subpopulations are also active at different times and are linked by recurrent inhibition, suggesting that mammalian temporal estimation may operate using similar principles. The paradigm that this study developed offers opportunities to understand the interface between internal models and sensory evidence. Circadian neurons are sensitive to environmental inputs: can they autonomously compute prediction error? They communicate with downstream dopaminergic populations: are those analogous to mammalian midbrain dopaminergic neurons whose activities reflect prediction error? It is proposed that flies assign valence to experienced environmental conditions, a computation that uses an internal model generated through circuit remodeling (Song, 2021).
Neuropeptides play pivotal roles in modulating circadian rhythms. Pigment-dispersing factor (PDF) is critical to the circadian rhythms in Drosophila locomotor activity. This study demonstrates that diuretic hormone 31 (DH31) complements PDF function in regulating free-running rhythmicity using male flies. It was determined that Dh31 loss-of-function mutants (Dh31#51) showed normal rhythmicity, whereas Dh31(#51);Pdf01 double mutants exhibited a severe arrhythmic phenotype compared to Pdf-null mutants (Pdf01). The expression of tethered-PDF or tethered-DH31 in clock cells, posterior dorsal neurons 1 (DN1ps), overcomes the severe arrhythmicity of Dh31(#51);Pdf01double mutants, suggesting that DH31 and PDF may act on DN1ps to regulate free-running rhythmicity in a hierarchical manner. Unexpectedly, the molecular oscillations in Dh31(#51);Pdf01 mutants were similar to those in Pdf01 mutants in DN1ps, indicating that DH31 does not contribute to molecular oscillations. Furthermore, a reduction in Dh31 receptor (Dh31r) expression resulted in normal locomotor activity and did not enhance the arrhythmic phenotype caused by the Pdf receptor (Pdfr) mutation, suggesting that PDFR, but not DH31R, in DN1ps mainly regulates free-running rhythmicity. Taken together, this study identifies a novel role of DH31, in which DH31 and PDF hierarchically regulate free-running rhythmicity through DN1ps (Goda, 2019).
This study has demonstrated a novel function of DH31 in regulating Drosophila locomotor activity rhythms. Dh31#51 mutants maintained a robust free-running rhythm, whereas Dh31#51;Pdf01 double-mutant flies exhibited a severe disruption of their free-running rhythm compared to Pdf01 mutants. These findings suggest that Dh31#51 mutants maintain a robust free-running rhythm because the primary factor, PDF, can sustain a strong rhythm. ~40% of Pdf01 single-mutant flies exhibited a preserved rhythmic state, which is because DH31 can partially support free-running rhythmicity. Thus, the severe disruptions of free-running rhythm in Pdf01 and Dh31#51 double-mutant flies is likely caused by the loss of both pathways (Goda, 2019).
PDF is secreted from the main circadian neurons, LNvs, and acts on other clock cells through PDFR to synchronize and maintain robust molecular rhythms. PDF expression from LNvs in Dh31#51;Pdf01 mutants restored rhythmicity, in contrast to tethered-PDF (t-PDF) expression in LNvs, indicating that an autoreceptor of PDF signals in LNvs is not sufficient to maintain rhythmicity. Instead, t-PDF expression in DN1ps restored rhythmicity, suggesting that PDF signaling in DN1ps is sufficient to maintain robust free-running rhythmicity. Recently, the responsiveness to PDF was shown to be strongly altered for 24 h via RalA GTPase in sLNvs28. Therefore, it is expected that the continuous activation of PDFR by t-PDF generates rhythmic downstream signaling in PDFR-expressing neurons (Goda, 2019).
Molecular oscillations in DN1s were strongly dampened in Pdf01 mutants compared with WT flies. These data are consistent with previous studies in which the molecular oscillations of PER in Pdf01 mutants held under DD conditions were dampened in DN1s10 and the genetic manipulation of the circadian clocks in PDF-positive cells altered the molecular rhythms in DN1ps. Furthermore, Pdfr expression in DN1ps has been reported to prevent the arrhythmic phenotype in Pdfr5304 mutants. These findings support the idea that PDF is secreted from LNvs and acts on DN1ps to regulate free-running rhythmicity (Goda, 2019).
Furthermore, it was shown that t-DH31 expression in DN1ps rescued the Pdf01 and Dh31#51 double-mutant phenotypes, which suggests that DH31 acts on DN1ps to regulate rhythmicity. Although it has been suggested that DH31 release might increase at dawn and that DH31-mRNA expression levels oscillate for 24 h, how t-DH31 expression causes rhythmic behavioral output remains unclear. Because DH31 can modestly activate PDFR in vitro, it cannot be excluded that t-DH31 overexpression might simply activate PDFR in DN1ps instead of the intrinsic PDF signals, thereby restoring locomotor activity rhythms in the flies. However, the rhythmicity of Dh31#51;Pdf01 mutants overexpressing t-DH31 in tim-Gal4-expressing neurons or R18H11-Gal4-expressing DN1ps only reached levels similar to that of the Pdf01 single-mutant flies. Therefore, DH31 likely acts on DN1ps separately from the PDF pathway (Goda, 2019).
Although it has been shown that DH31 is expressed in a subset of DN1ps, DH31 expression using R18H11-Gal4 did not rescue the Pdf01 and Dh31#51 double-mutant phenotypes, suggesting that DH31 expression in R18H11-Gal4-expressing neurons is insufficient to maintain rhythmicity. Instead, DH31 is expressed in DN1as and DH31 expression in tim-Gal4-expressing neurons rescued the phenotype, which suggests that DH31 expression in clock neurons maintains rhythmicity. That said, given that DH31 is expressed in nonclock neurons and that tim-Gal4 is expressed in nonclock cells, it cannot be excluded that DH31 expression in nonclock neurons might play a role in rescuing the severe phenotype of Dh31#51;Pdf01 mutants. Alternatively, although DH31 expression in LNvs was not detectable via anti-DH31 antibody staining, a recent RNA-seq analysis detected Dh31 gene expression in both LNvs and DN1s. Therefore, DH31 expression from LNvs may potentially act on DN1s to support locomotor activity rhythms (Goda, 2019).
In summary, it is proposed that PDF and DH31 regulate free-running rhythms in a hierarchical fashion in DN1ps. As t-DH31 or t-PDF expression in DN1ps resulted in a similar level of rhythmicity as that observed in flies expressing t-DH31 or t-PDF, respectively, in tim-Gal4-expressing neurons, DN1ps are at least one of the important clock cells that regulate free-running rhythmicity (Goda, 2019).
Given that Dh31#51;Pdf01 mutants exhibited severe arrhythmicity in free-running rhythm, it is speculated that the severe arrhythmic phenotype might be a result of abnormal molecular oscillations. However, the molecular oscillations of Dh31#51;Pdf01 mutants were similar to those of Pdf01 mutants. Therefore, the molecular mechanisms by which DH31 regulates free-running rhythms still remain unclear. Importantly, the peak of VRI expression in LNds in Dh31#51 was at ZT 19, which was delayed compared with those of WT flies and the other mutants. The data suggested that DH31 is involved in the regulation of molecular oscillations in LNds. Because LNds are the evening pacemaker, the delayed VRI oscillations in LNds might be associated with the longer period of free-running rhythm in Dh31#51 (Goda, 2019).
Recently, the intracellular calcium rhythms in each clock cell were reported to be nonsynchronous and associated with morning and evening peaks in locomotor activity. DH31 signaling may possibly contribute to the downstream output that controls molecular rhythms in pacemaker processes, such as intracellular calcium rhythms. Given that PDF from sLNvs regulates strong molecular rhythms in DN1ps and generates robust free-running rhythms under constant conditions, DH31 may help maintain vigorous output signals downstream of the molecular clocks in DN1ps (Goda, 2019).
Recently work has shown that both Dh31r1/Df mutants and flies undergoing Dh31r knockdown in their neurons showed normal rhythmicity in the locomotor activity rhythm25. In contrast to Dh31#51;Pdf01 double mutants, Pdfr5304;Dh31r1/Df double mutants did not enhance the arrhythmicity observed in Pdfr single mutants, which suggests that Dh31r does not complement PDFR function; thus, Dh31r does not function as a receptor for DH31 in this context. Given that Dh31r1/Df flies showed a strong abnormality in the TPR phenotype25, it is more likely that Dh31r does not play an important role in locomotor activity rhythms. However, Dh31r1/Df4 mutants are not null25, and it cannot be excluded that a small amount of residual Dh31r might drive robust locomotor activity rhythms with the PDF pathway (Goda, 2019).
Which receptors might function with DH31 to regulate free-running rhythmicity? Given that DH31 can activate PDFR in vitro, bath applications of DH31 can activate LNvs via PDFR19 and DH31 can function as a ligand of PDFR in TPR at the onset of night, PDFR may function as a receptors for both DH31 and PDF in the regulation of free-running rhythmicity. However, because the arrhythmicity of Pdfr5304 mutants was not as severe as that of Dh31#51;Pdf01 mutants, PDFR does not appear to act as a receptor for DH31 in this context (Goda, 2019).
Both Dh31r and PDFR are class II G-protein coupled receptors (GPCRs), which also include Hector and Diuretic hormone 44 receptors 1 and 2 (DH44R1 and DH44R2, respectively). Interestingly, the DH44R1 and DH44R2 ligand DH44 has been implicated in circadian output circuits. Therefore, although there is no evidence from in vitro or in vivo experiments, these receptors might nevertheless function as receptors for DH31 to regulate free-running rhythmicity (Goda, 2019).
Orchestration of neuropeptides regulates locomotor activity rhythms in species ranging from flies to mammals
The orchestration of neuropeptides is critical for regulating circadian clock functions in species that range from flies to mammals. In mammals, several neuropeptides, including vasoactive intestinal polypeptide (VIP), arginine vasopressin (AVP) and neuromedin S (NMS), are expressed in the SCN, which is the center for circadian clock control. The hierarchy of neuropeptide signaling contributes to circadian function in the SCN. Several recent studies in Drosophila have identified the neuropeptides, including ion transport peptide (ITP), neuropeptide F (NPF), allatostatin A, short neuropeptide F, leucokinin and DH44, that regulate locomotor activity and sleep. However, given that DH31 complements the function of PDF in regulating free-running rhythmicity in the same clock cells, DH31 not only serves as one of the neuropeptides that regulates circadian rhythms but also might selectively influence PDF function in the regulation of free-running rhythms. Thus, these findings shed new light on the next steps required to improve understanding of the core neuropeptide regulatory mechanisms involved in the circadian rhythm (Goda, 2019).
Circadian clocks regulate much of behavior and physiology, but the mechanisms by which they do so remain poorly understood. While cyclic gene expression is thought to underlie metabolic rhythms, little is known about cycles in cellular physiology. This study found that Drosophila insulin-producing cells (IPCs), which are located in the pars intercerebralis and lack an autonomous circadian clock, are functionally connected to the central circadian clock circuit via DN1 neurons. Insulin mediates circadian output by regulating the rhythmic expression of a metabolic gene (sxe2) in the fat body. Patch clamp electrophysiology reveals that IPCs display circadian clock-regulated daily rhythms in firing event frequency and bursting proportion under light:dark conditions. The activity of IPCs and the rhythmic expression of sxe2 are additionally regulated by feeding, as demonstrated by night feeding-induced changes in IPC firing characteristics and sxe2 levels in the fat body. These findings indicate circuit-level regulation of metabolism by clock cells in Drosophila and support a role for the pars intercerebralis in integrating circadian control of behavior and physiology (Barber, 2016).
A 24-h rhythm of feeding behavior, or synchronized feeding/fasting episodes during the day, is crucial for survival. Internal clocks and light input regulate rhythmic behaviors, but how they generate feeding rhythms is not fully understood. This study aimed to dissect the molecular pathways that generate daily feeding patterns. By measuring the semidiurnal amount of food ingested by single flies,it was demonstrated that the generation of feeding rhythms under light:dark conditions requires quasimodo (qsm) but not molecular clocks. Under constant darkness, rhythmic feeding patterns consist of two components: CLOCK (CLK) in digestive/metabolic tissues generating feeding/fasting episodes, and the molecular clock in neurons synchronizing them to subjective daytime. Although CLK is a part of the molecular clock, the generation of feeding/fasting episodes by CLK in metabolic tissues was independent of molecular clock machinery. These results revealed novel functions of qsm and CLK in feeding rhythms in Drosophila (Maruko, 2023).
Many organisms, including insects, fish, birds, rodents, and primates, show particular feeding patterns during the day. This study investigated the roles of molecular clock genes in food intake in Drosophila. In LD, light stimuli were sufficient to induce feeding via the QSM-mediated pathway, while feeding rhythms in DD can be dissected into two components: generation of feeding/fasting episodes by Clk/cyc in metabolic tissues and their synchronization by molecular clocks in neurons. These results suggest novel roles of qsm and clock molecules regulating feeding behavior (Maruko, 2023).
There has been a significant advance in understanding the circuits and neurotransmitters that mediate central clocks in the brain that control feeding behavior. As for the peripheral clocks, their functions in feeding behavior seem more diverse than those in the central clocks. In addition to contributing to the regulation of 24-h feeding rhythms, it was also reported that peripheral clocks negatively regulate food intake amount and feeding rhythm strength. However, it was elucidated whether the increase in food intake amount is caused by the disruption of the molecular clock machinery or other functions of CLK/CYC. The results suggest that fat body CLK regulates food intake amount independent of molecular clocks. Since flies with Clk/cyc disruption with various genotypes consistently show a reduction in fluctuation in food intake compared to their control (i.e., ClkJrk, cycout, cyc02, to>dnClk, cyc02, to > cyc), it is less likely that fluctuation in food intake is due to differences in the body size or other background effects. The suppression of CLK function in the fat body reduced fluctuations of feeding amount in per01 background, indicating that CLK/CYC in the metabolic tissues contributes to rhythmic feeding behavior in addition to its function as components of the molecular clock. It explains the different effects of clock genes on patterns of feeding rhythms: per- and tim-null mutants showed a shift in the peak of the feeding timing, and Clk- and cyc-null mutants ingest food constantly without peak of the feeding timing (Maruko, 2023).
The results suggest that the effect of CLK/CYC in the metabolic tissues to reduce feeding rhythm strength is likely to be mediated by suppression of feeding during the fasting period. Several factors that act on suppression of food intake in peripheral tissues have been reported, and expression of some of these genes, such as allatostatin A and its receptor, are regulated by peripheral CLK/CYC. In addition, rhythmic expression of the allatostatin A receptor-2 gene is affected by the disruption of the fat body clock. CLK is also associated with cAMP-responsive element binding protein (CREB), which is involved in the energy homeostasis of insects and mammals. Blocking CREB activity in the fat body increases food intake in flies, and Nejire, a homolog of CREB-binding protein (CBP)/p300, has been reported as a regulator of CLK/CYC-dependent transcription. Further studies on these pathways may reveal a novel signaling axis that constitute the feeding/fasting cycle (Maruko, 2023).
This study found the novel role of qsm in feeding behavior in LD. It has been reported that cry-null mutants do not display the morning peak in LD. It was also observed that cry-null mutants do not display the early morning peak in LD, while these flies still showed the synchronized feeding/fasting episodes in LD. In addition, the feeding/fasting episodes in DD were observed without the morning peak. Thus, the morning peak is not associated with the daytime feeding pattern that was the focus of this study. The results revealed that qsm is indispensable in the daytime feeding pattern. qsm encodes a ZP (Zona Pellucida) domain and constitutes part of CRY-independent light input to the circadian clock. QSM is expressed in many cells in the immediate proximity of clock neurons, and it is not clear in which cells QSM influences the feeding pattern for now. Further study to understand where and how the QSM regulates feeding rhythms in LD would help understand light-induced regulation of feeding behavior (Maruko, 2023).
In summary, these results revealed novel pathways that regulate the formation of feeding rhythms in Drosophila. Feeding/fasting rhythms coordinate metabolism and affect aging and life span. Further studies of these axes may contribute to human health (Maruko, 2023).
This study has successfully dissected the molecular pathways that regulate feeding patterns. However, these analyses are limited to several key components, and their upstream and downstream molecules remain to be elucidated. It was found that qsm regulates feeding rhythms under light:dark conditions, while its downstream signaling is unknown. This study also found that, under constant darkness conditions, CLK/CYC in digestive/metabolic tissues generates feeding/fasting episodes, and the molecular clock in neurons synchronizes them. However, molecular mechanisms that link those two components remain to be elucidated. Further studies are needed to understand the entire picture of the molecular pathways that generate feeding patterns (Maruko, 2023).
Many behaviors exhibit ~24-h oscillations under control of an endogenous circadian timing system.Most circadian research in Drosophila has focused on the generation of locomotor activity rhythms, but a fundamental question is how the circadian clock orchestrates multiple distinct behavioral outputs. This study has investigated the cells and circuits mediating circadian control of feeding behavior. This study shows that the presence of feeding rhythms requires molecular clock function in the ventrolateral clock neurons of the central brain. This study further demonstrate that the speed of molecular clock oscillations in these neurons dictates the free-running period length of feeding rhythms. In contrast to the effects observed with central clock cell manipulations, This study shows that genetic abrogation of the molecular clock in the fat body, a peripheral metabolic tissue, is without effect on feeding behavior. Under these conditions, the period of feeding rhythms tracks with molecular oscillations in central brain clock cells, consistent with a primary role of the brain clock in dictating the timing of feeding behavior. Finally, despite a lack of effect of fat body selective manipulations, this study found that flies with simultaneous disruption of molecular clocks in multiple peripheral tissues (but with intact central clocks) exhibit decreased feeding rhythm strength and reduced overall food intake. This study concluded that both central and peripheral clocks contribute to the regulation of feeding rhythms, with a particularly dominant, pacemaker role for specific populations of central brain clock cells (Fulgham, 2021).
The link between the biological clock and reproduction is evident in most metazoans. The fruit fly Drosophila melanogaster, a key model organism in the field of chronobiology because of its well-defined networks of molecular clock genes and pacemaker neurons in the brain, shows a pronounced diurnal rhythmicity in oogenesis. Still, it is unclear how the circadian clock generates this reproductive rhythm. A subset of the group of neurons designated "posterior dorsal neuron 1" (DN1p), which are among the ~150 pacemaker neurons in the fly brain, produces the neuropeptide allatostatin C (AstC-DN1p). This study reports that six pairs of AstC-DN1p neurons send inhibitory inputs to the brain insulin-producing cells, which express two AstC receptors, star1 and AICR2. Consistent with the roles of insulin/insulin-like signaling in oogenesis, activation of AstC-DN1p suppresses oogenesis through the insulin-producing cells. This study shows evidence that AstC-DN1p activity plays a role in generating an oogenesis rhythm by regulating juvenile hormone and vitellogenesis indirectly via insulin/insulin-like signaling. AstC is orthologous to the vertebrate neuropeptide somatostatin (SST). Like AstC, SST inhibits gonadotrophin secretion indirectly through gonadotropin-releasing hormone neurons in the hypothalamus. The functional and structural conservation linking the AstC and SST systems suggest an ancient origin for the neural substrates that generate reproductive rhythms (Zhang, 2021).
Six pairs of DN1p neurons were discovered that are part of the circadian pacemaker neuron network in the brain and make functional inhibitory connections to the brain IPCs. The IPCs are endocrine sensors that link the organism's nutritional status with anabolic processes, such as those associated with growth in developmental stages and with reproduction in adults. In juvenile stages, activation of insulin and insulin-like growth factor (IGF) signaling (IIS) through the InR results in larger flies, whereas inhibition of this pathway produces smaller flies. Consistent with this, it was also found that forced activation of the AstC-DN1p (i.e., CNMa-Gal4/UAS-NaChBac) during development resulted in 12% smaller adults, confirming their role as a negative regulator of the IPCs. In adults, the IPCs are associated with many physiological and behavioral processes, such as feeding, glycaemic homeostasis, sleep, lifespan, and stress resistance. As such, the IPCs receive a variety of modulatory inputs from both central and peripheral sources, such as sNPF, corazonin, tachykinins, limostatin, allatostatin A, adipokinetic hormone, GABA, serotonin, and octopamine. Regarding reproduction, IIS directed by the IPCs stimulates GSC proliferation and vitellogenesis. The results also indicate that AstC from AstC-DN1p suppresses the secretory activity of the IPCs and juvenile hormone (JH)-dependent oocyte development (i.e., vitellogenesis). Indeed, it was found that the JH mimic methoprene can rescue the suppression of oogenesis induced by AstC-DN1p activation. From these results it is concluded that IPCs are inhibited by AstC released by AstC-DN1p. A similar link between IIS and the circadian clock has also been reported in mammals, but the mechanism remains unclear (Zhang, 2021).
Although the genetic evidence supporting the inhibitory action of AstC-DN1p on IPCs is compelling, it is also puzzling because a previous study found forced activation of 8 to 10 pairs of DN1p neurons (i.e., Clk4.1-LexA+ neurons) induced Ca2+ transients in IPCs. This study also found that, under LD 12:12 conditions, the IPCs showed electrical activity early in the morning when DN1p neurons are also active. The same study, however, reported that, under DD conditions, the IPCs showed no bursting activity in the morning (i.e., CT0 to -4). Instead, they showed bursting activity in the late afternoon (i.e., CT8 to -12) when DN1 activity falls. Furthermore, DN1p activation evokes varying levels of Ca2+ transients from individual IPCs, some of which produce no detectable Ca2+ transient. Thus, like mammalian pancreatic β-cells, the IPCs in Drosophila seem to comprise a heterogeneous cell population. It is noted that individual IPCs show highly variable AstC-R1 expression, which would also lead to individual IPCs showing variable responses to AstC (Zhang, 2021).
In D. melanogaster, the LD cycle generates an egg-laying rhythm by influencing oogenesis and oviposition. Oviposition depends on light cues, whereas oogenesis cycles with the circadian rhythm that itself continues to run in DD conditions. In live-brain Ca2+ imaging experiments, DN1 neurons show a circadian Ca2+ activity rhythm that peaks around CT19 and reaches its lowest point between CT6 and CT8. This DN1 activity rhythm correlates well with the rhythm of vitellogenesis initiation observed in this study. In this model, the lowest point in DN1 Ca2+ activity between CT6 and CT8 leads to a significant attenuation of AstC secretion. This leads to a derepression of IPC activity, which eventually induces JH biosynthesis and vitellogenesis initiation. The 6-h delay required for previtellogenic stage 7 follicles to develop into vitellogenic stage 8 follicles would result in a peak in the number of stage 8 follicles between CT12 and CT14. Notably, the ovaries of the AstC-deficient mutant showed similar numbers of stage 8 oocytes at all examined circadian time points, indicating that any other JH- or vitellogenesis-regulating factors play only minor roles in producing the circadian vitellogenesis rhythm (Zhang, 2021).
Like the IPCs, the DN1p cluster is also heterogeneous. A subset of the DN1p neurons is most active at dawn and promotes wakefulness. Another subset of the DN1p cluster (also known as, spl-gDN1) promotes sleep. The DN1p cluster comprises two morphologically distinct subpopulations, a-DN1p and vc-DN1p. The a-DN1p subcluster promotes wakefulness by inhibiting sleep promoting neurons, whereas the vc-DN1p subcluster resembles the sleep-promoting spl-gDN1. These results indicate AstC-DN1p are a-DN1p neurons that project to the anterior optic tubercle (AOTU. Although the possibility cannot be ruled out that AstC-DN1p is also heterogeneous and includes some vc-DN1p neurons, the wake-promoting role of a-DN1p aligns well with the circadian vitellogenesis rhythm that requires the secretory activity of AstC-DN1p to be lowest in the afternoon and highest at dawn. Furthermore, AstC-DN1p neurons express Dh31. Dh31-expressing DN1 clock neurons are intrinsically wake-promoting and Dh31-DN1p activity in the late night or early morning suppresses sleep. Again, this is consistent with the observation that AstC-DN1p are also wake-promoting a-DN1p. It is speculated Dh31 plays a limited role in oogenesis regulation, because unlike AstC, RNAi-mediated knockdown of Dh31 had a negligible impact on female fecundity (Zhang, 2021).
Besides AstC-DN1p, the female brain has many additional AstC neurons. However, it seems unlikely that other AstC neurons contribute to the circadian vitellogenesis rhythm. This is because restoring AstC expression specifically in AstC-DN1p almost completely restored the vitellogenesis rhythm in AstC-deficient mutants. It is feasible, however, that other AstC neurons contribute to different aspects of female reproduction. Indeed, a sizable difference was noted in the final oogenesis outcome between AstC-Gal4 neuron activation and brain-specific AstC-Gal4 neuron activation. This suggests AstC cells outside of the brain also regulate oogenesis probably in other physiological contexts, such as the postmating responses (Zhang, 2021).
AstC receptors are orthologous to mammalian SST receptors (sstr1-5). SST is a brain neuropeptide that was originally identified as an inhibitor of growth hormone (GH) secretion in the anterior pituitary. Thus, the observation that AstC inhibits IIS from IPCs, a major endocrine signal that promotes growth in Drosophila, suggests remarkable structural and functional conservation between the invertebrate AstC and vertebrate SST systems. In addition, SST inhibits the hypothalamic neuropeptide GnRH, which stimulates the anterior pituitary's production of follicle-stimulating hormone (FSH) and luteinizing hormone (LH). FSH stimulates clutches of immature follicles to initiate follicular development, while LH stimulates ovulation. Thus, both AstC and SST regulate the secretion of gonadotropins (JH in insects, FSH and LH in mammals) indirectly through the IPCs in insects and through the hypothalamic GnRH neurons in mammals. This functional conservation between AstC and SST is also evident in the immune system. AstC inhibits the innate immune system in insects, while SST inhibits inflammation in mammals (Zhang, 2021).
In many seasonal breeders, the changing photoperiod as the seasons progress acts as an environmental cue for the biological clock system, which would then direct any necessary physiological changes. During the winter, Drosophila females enter a form of reproductive dormancy characterized by a pronounced suppression of vitellogenesis. A winter-like condition (i.e., short-day length, low temperature, and food shortage) down-regulates neural activity in the IPCs. But the IPCs are not equipped with a cell-autonomous clock, so they must receive seasonal information from the brain clock neuron network. Indeed, two clock related neuropeptides~pigment dispersing factor and short neuropeptide F~from circadian morning pacemaker or M-cells have been implicated in regulating reproductive dormancy. Intriguingly, AstC-DN1p neurons are the DN1p subset that receives pigment dispersing factor signals from these M-cells. Furthermore, DN1p can process light and temperature information for the circadian regulation of behavior. Finally, the finding that AstC-DN1p generates the circadian vitellogenesis rhythm via the IPCs makes AstC-DN1p neurons the prime candidates for integrating the seasonal cues that control the entrance, maintenance, or exit from reproductive dormancy. Considering the functional and structural conservation between the AstC and SST systems, the SST system may also link the brain clock, GnRH, and/or its downstream reproductive pathways in controlling seasonal reproductive patterns in vertebrates (Zhang, 2021).
Twenty-four hour rhythms in behavior are organized by
a network of circadian pacemaker neurons. Rhythmic activity in this
network is generated by intrinsic rhythms in clock neuron physiology
and communication between clock neurons. However, it is poorly
understood how the activity of a small number of pacemaker neurons is
translated into rhythmic behavior of the whole animal. To understand
this, a screen was carried out for signals that could identify
circadian output circuits in Drosophila melanogaster. Leucokinin
neuropeptide (LK) and its receptor (LK-R)
were found to be required for normal behavioral rhythms. This LK/LK-R
circuit connects pacemaker neurons to brain areas that regulate
locomotor activity and sleep. These experiments revealed that pacemaker
neurons impose rhythmic activity and excitability on LK- and
LK-R-expressing neurons. Pacemaker neuron-dependent activity rhythms
were also found in a second circadian output pathway controlled by DH44
neuropeptide-expressing neurons. It is concluded that rhythmic clock
neuron activity propagates to multiple downstream circuits to
orchestrate behavioral rhythms (Cavey, 2016). The mammalian circadian clock influences most aspects of
physiology and behavior through the transcriptional control of a wide
variety of genes, mostly in a tissue-specific manner. About 20
clock-controlled genes (CCGs) oscillate in virtually all mammalian
tissues and are generally considered as core clock components. One of
them is Ubiquitin-Specific
Protease 2 (Usp2), whose status remains controversial, as it may
be a cogwheel regulating the stability or activity of core cogwheels or
an output effector. This study reports that Usp2 is a clock output
effector related to bodily Ca2+ homeostasis, a feature that is conserved
across evolution. Drosophila with a whole-body knockdown of the
orthologue of Usp2, (Ubiquitin
specific protease 2) predominantly die during pupation but are
rescued by dietary Ca2+ supplementation. Usp2-KO mice show
hyperabsorption of dietary Ca2+ in small intestine, likely due to strong
overexpression of the membrane scaffold protein NHERF4,
a regulator of the Ca2+ channel TRPV6 mediating dietary Ca2+ uptake. In
this tissue, USP2-45 is found in membrane fractions and negatively
regulates NHERF4 protein abundance in a rhythmic manner at the protein
level. In clock mutant animals (Cry1/Cry2-dKO; see Drosophila Cryptochrome),
rhythmic USP2-45 expression is lost, as well as the one of NHERF4,
confirming the inverse relationship between USP2-45 and NHERF4 protein
levels. Finally, USP2-45 interacts in vitro with NHERF4 and endogenous
Clathrin Heavy Chain (see Drosophila Clathrin
Heavy Chain). Taken together these data has led the authors to
define USP2-45 as the first clock output effector acting at the
post-translational level at cell membranes and possibly regulating
membrane permeability of Ca2+ (Pouly, 2016). This study used a laboratory selection approach
to raise populations of Drosophila melanogaster that emerge in the
morning (early) or in the evening (late), and over 14 years of
continued selection, clear divergence of their circadian phenotypes is
reported. The molecular correlates of early and late emergence
chronotypes were also assessed, and significant divergence is reported
in transcriptional regulation, including the mean phase, amplitude and
levels of period
(per), timeless
(tim), clock
(clk) and vrille
(vri) messenger RNA (mRNA) expression. Corroborating some of
the previously reported light-sensitivity and oscillator network
coupling differences between the early and the late populations,
differences are also reported in mRNA expression of the circadian
photoreceptor cryptochrome
(cry) and in the mean phase, amplitude and levels of the
neuropeptide pigment-dispersing
factor (PDF). These results provide the first-ever direct
evidence for divergent evolution of molecular circadian clocks in
response to selection imposed on an overt rhythmic behavior and
highlight early and late populations as potential models for chronotype
studies by providing a preliminary groundwork for further exploration of
molecular-genetic correlates underlying circadian clock-chronotype
association (Nikhil, 2016). Circadian
clocks are endogenous timers adjusting behaviour and physiology
with the solar day. Visual and non-visual photoreceptors are responsible
for synchronizing circadian clocks to light, but clock-resetting is also
achieved by alternating day and night temperatures with only 2-4 degrees
C difference. This study shows that Drosophila Ionotropic
Receptor 25a (IR25a) is required for behavioural synchronization
to low-amplitude temperature cycles. This channel is expressed in
sensory neurons of internal stretch receptors previously implicated in
temperature synchronization of the circadian clock. IR25a is required
for temperature-synchronized clock protein oscillations in subsets of
central clock neurons. Extracellular leg nerve recordings reveal
temperature- and IR25a-dependent sensory responses, and IR25a
misexpression confers temperature-dependent firing of heterologous
neurons. It is proposed that IR25a is part of an input pathway to the
circadian clock that detects small temperature differences. This pathway
operates in the absence of known 'hot' and 'cold' sensors in the
Drosophila antenna, revealing the existence of novel periphery-to-brain
temperature signalling channels. Sleep, a reversible quiescent state found in both
invertebrate and vertebrate animals, disconnects animals from their
environment and is highly regulated for coordination with wakeful
activities, such as reproduction. The fruit fly, Drosophila
melanogaster, has proven to be a valuable model for studying the
regulation of sleep by circadian clock and homeostatic mechanisms.
This study demonstrates that the Sex
peptide receptor (SPR) of Drosophila, known for its role in female
reproduction, is also important in stabilizing sleep in both males and
females. Mutants lacking either the SPR or its central ligand, myoinhibitory
peptide (MIP), fall asleep normally, but have difficulty in
maintaining a sleep-like state. This analyses have mapped the SPR sleep
function to pigment
dispersing factor (pdf) neurons, an arousal center in the insect
brain. MIP downregulates intracellular cAMP levels in pdf neurons
through the SPR. MIP is released centrally before and during night-time
sleep, when the sleep drive is elevated. Sleep deprivation during the
night facilitates MIP secretion from specific brain neurons innervating
pdf neurons. Moreover, flies lacking either SPR or MIP cannot recover
sleep after the night-time sleep deprivation. These results delineate a
central neuropeptide circuit that stabilizes the sleep state by feeding
a slow-acting inhibitory input into the arousal system and plays an
important role in sleep homeostasis (Oh, 2014: PubMed).
Body temperature exhibits rhythmic fluctuations over a 24 h period and decreases during the night, which is associated with sleep initiation. However, the underlying mechanism of this temperature decrease is largely unknown. Previous work has shown that Drosophila exhibit a daily temperature preference rhythm (TPR), in which their preferred temperatures increase during the daytime and then decrease at the transition from day to night (night-onset). Because Drosophila are small ectotherms, their body temperature is very close to that of the ambient temperature, suggesting that their TPR generates their body temperature rhythm. This study demonstrates that the neuropeptide diuretic hormone 31 (DH31) and pigment-dispersing factor receptor (PDFR) contribute to regulate the preferred temperature decrease at night-onset. PDFR and tethered-DH31 expression in dorsal neurons 2 (DN2s) restore the preferred temperature decrease at night-onset, suggesting that DH31 acts on PDFR in DN2s. Notably, it was previously shown that the molecular clock in DN2s is important for TPR. Although PDF (another ligand of PDFR) is a critical factor for locomotor activity rhythms, Pdf mutants exhibit normal preferred temperature decreases at night-onset. This suggests that DH31-PDFR signaling specifically regulates a preferred temperature decrease at night-onset. Thus, it is proposed that night-onset TPR and locomotor activity rhythms are differentially controlled not only by clock neurons but also by neuropeptide signaling in the brain (Goda, 2016).
Body temperature rhythm (BTR) is fundamental for maintaining homeostasis, such as in generating metabolic energy and sleep. BTR is one of the most robust circadian outputs and can affect the peripheral clocks of mammals. The rhythmic patterns of BTR and locomotor activity rhythms are analogous. For instance, in diurnal mammals, both body temperature and locomotor activity increase during the daytime and decrease at night. Nonetheless, BTR and locomotor activity rhythms are regulated by different subsets of subparaventricular zone (SPZ) neurons, suggesting that these rhythms are controlled independently (Goda, 2016).
Mammals are not the only examples of this phenomenon. Previous work has shown that Drosophila exhibit a daily temperature preference rhythm (TPR), in which their preferred temperatures increase during the daytime and then decrease at the transition from day to night (night-onset) (Kaneko, 2012). Because Drosophila are small ectotherms, their body temperature is very close to that of the ambient temperature, suggesting that their TPR generates their BTR. In Drosophila, TPR and locomotor activity rhythms are regulated by different subsets of clock neurons (Kaneko, 2012). There are ~150 central pacemaker cells in the fly brain, which are functionally homologous to mammalian suprachiasmatic nucleus (SCN) neurons. These pacemaker cells are approximately divided into lateral neurons [LNs; small ventral LNs (s-LNvs), large ventral LNs (l-LNvs), and dorsal lateral neurons (LNds)] and dorsal neurons (DNs; DN1, DN2, and DN3) based on their location and size. The clocks in DN2s are critical for regulating TPR during the daytime (Kaneko, 2012), but not for regulating locomotor activity rhythms (Goda, 2016 and references therein).
Neuropeptides and their receptors have important roles in synchronizing circadian clocks. A class II G-protein-coupled receptor, pigment-dispersing factor receptor (PDFR), and its ligand (PDF) play important roles in synchronizing circadian clocks and are required for robust circadian locomotor activity in Drosophila. Notably, PDF and PDFR function in a similar manner to vasoactive intestinal peptide (VIP) and its receptor VPAC2 in mammals, both of which play important roles in the ability of clock neurons to regulate the rhythmicity and synchrony of both locomotor activity rhythms and BTRs (Goda, 2016).
Recent reports have suggested that, in addition to PDF, diuretic hormone 31 (DH31) also activates PDFR based on in vitro experiments and a study that used brain imaging with bath-applied DH31. Moreover, it has been shown that DH31 is expressed in the posterior dorsal neurons 1 (DN1ps) and that it modulates sleep as a wake-promoting signal before dawn but does not affect locomotor activity rhythms in Drosophila. DH31 is a functional homolog of mammalian calcitonin gene-related peptide (CGRP), which mediates thermosensation and thermoregulation. However, it is unknown whether CGRP is involved in the regulation of BTR in mammals (Goda, 2016).
This study demonstrates that DH31 and PDFR play important roles for TPR at night-onset. DN2s are the main clock cells for TPR, and the data suggest that DH31 binding to PDFR in DN2s regulates temperature preference decreases at night-onset, which is the first in vivo evidence that DH31 could function as a ligand of PDFR. Therefore, it is proposed that circadian locomotor activity and night-onset TPR are regulated by different neuropeptides that use the same receptor expressed in different clock cells (Goda, 2016).
Both Dh31 and Pdfr mutants exhibited abnormal night-onset TPR, and t-DH31 and PDFR expression in DN2s are sufficient to control night-onset TPR, suggesting that DH31 could be a ligand of PDFR in DN2s. Unexpectedly, PDF, an important neuropeptide for locomotor activity, is not required for night-onset TPR. It suggests that PDF and DH31 appear to act on different subsets of PDFR-expressing clock cells to regulate locomotor activity rhythms and night-onset TPR, respectively. Together, the data suggest that locomotor activity and night-onset TPR are regulated through PDFR by different subsets of clock neurons using different neuropeptides. Nonetheless, because all Dh31, Pdf, and Pdfr mutants still maintain normal daytime TPR, the underlying molecular mechanisms of daytime TPR are still obscure (Goda, 2016).
In humans, body temperature dramatically decreases at night, which is associated with sleep initiation. Although it suggests a relationship between BTR and sleep-wake cycles, the underlying mechanisms are largely unclear. A recent study suggested that DH31 mediates sleep-wake cycles and that DH31 secretion functions as a wake-promoting signal before dawn. Given that it was found that DH31 is required for night-onset TPR, DH31 could regulate both sleep-wake cycles and TPR with different timing (i.e., at night-onset for TPR and before dawn for sleep) (Goda, 2016).
While t-DH31 expression in DN2s rescues the Dh31#51 phenotype, t-PDF expression in DN2s slightly rescues in comparison with UAS controls. This shows that t-DH31 in DN2s rescues more efficiently than t-PDF in DN2s. However, DH31 stimulates PDFR less efficiently than PDF in vitro. This suggests that either DH31 may more efficiently bind PDFR in DN2s than PDF or that DN2s may express another receptor of DH31 that also regulates night-onset TPR. Notably, although the DH31 receptor (DH31R) is a known receptor for DH31 in vitro, this study observed that DH31R is not expressed in DN2s and that the Dh31r mutant exhibited normal night-onset TPR. Therefore, it would be interesting to further explore whether additional receptors for DH31 in DN2s are involved in night-onset TPR. It has been previously shown that flies in the light prefer a 1°C higher temperature than in the dark (light-dependent temperature preference [LDTP]), thus demonstrating that light influences temperature preference behavior. Importantly, while LDTP is only affected by light, night-onset TPR is affected by both light and time (ZT10-ZT12 and ZT13-ZT15). Therefore, they are not controlled by the same pathway. In fact, PDFR expression using Clk4.5F-Gal4 rescues the abnormal LDTP phenotype of Pdfr5304 mutants, but it did not rescue night-onset TPR (Goda, 2016).
DH31 is a functional homolog of mammalian CGRP. Importantly, CGRP is related to many physiological functions, such as temperature sensation, migraines, and chronic pain in mammals. CGRP has recently attracted attention for its role in the treatment of migraine attacks, which are associated with body-temperature fluctuations. Additionally, CGRP is expressed in the SCN, but its function in circadian rhythms is not entirely clear. Thus, the findings raise the possibility that CGRP may be involved in BTR regulation and may mediate the connection between the circadian clock and other physiological functions. Moreover, this research will not only expand current understanding of neuropeptidergic regulation, but may also ultimately facilitate a discovery of the fundamental mechanisms of BTR (Goda, 2016).
To maintain homeostasis, animals must ingest appropriate quantities, determined by their internal nutritional state, of suitable nutrients. In the fruit fly Drosophila melanogaster, an amino acid deficit induces a specific appetite for amino acids and thus results in their increased consumption. Although multiple processes of physiology, metabolism, and behavior are under circadian control in many organisms, it is unclear whether the circadian clock also modulates such motivated behavior driven by an internal need. Differences in levels of amino acid consumption by flies between the light and dark phases of the day:night cycle were examined using a capillary feeder assay following amino acid deprivation. Female flies exhibited increased consumption of amino acids during the dark phase compared with the light phase. Investigation of mutants lacking a functional period gene (per0), a well-characterized clock gene in Drosophila, found no difference between the light and dark phases in amino acid consumption by per0 flies. Furthermore, increased consumption of amino acids during the dark phase was observed in mated but not in virgin females, which strongly suggested that mating is involved in the rhythmic modulation of amino acid intake. Egg production, which is induced by mating, did not affect the rhythmic change in amino acid consumption, although egg-laying behavior showed a per0-dependent change in rhythm. Elevated consumption of amino acids during the dark phase was partly induced by the action of a seminal protein, sex peptide (SP), on the sex peptide receptor (SPR) in females. Moreover, the increased consumption of amino acids during the dark phase is induced in mated females independently of their internal level of amino acids. These results suggest that a post-mating SP/SPR signal elevates amino acid consumption during the dark phase via the circadian clock (Uchizono, 2017).
The dopamine ontogeny hypothesis for schizophrenia proposes that transient dysregulation of the dopaminergic system during brain development increases the likelihood of this disorder in adulthood. To test this hypothesis in a high-throughput animal model, this study transiently manipulated dopamine signaling in the developing fruit fly Drosophila melanogaster and examined behavioral responsiveness in adult flies. Either a transient increase of dopamine neuron activity or a transient decrease of dopamine receptor expression during fly brain development permanently impairs behavioral responsiveness in adults. A screen for impaired responsiveness revealed sleep-promoting neurons in the central brain as likely postsynaptic dopamine targets modulating these behavioral effects. Transient dopamine receptor knockdown during development in a restricted set of ~20 sleep-promoting neurons recapitulated the dopamine ontogeny phenotype, by permanently reducing responsiveness in adult animals. This suggests that disorders involving impaired behavioral responsiveness might result from defective ontogeny of sleep/wake circuits (Ferguson, 2017).
Circadian coordination of metabolism, physiology, and behaviour is found in all living kingdoms. Clock genes are transcriptional regulators, and their rhythmic activities generate daily rhythms in clock-controlled genes which result in cellular and organismal rhythms. Insects provide numerous examples of rhythms in behaviour and reproduction, but less is known about control of metabolic processes by circadian clocks in insects. Recent data suggest that several pathways involved in protecting cells from oxidative stress may be modulated by the circadian system, including genes involved in glutathione (GSH) biosynthesis. Specifically, rhythmic expression of the gene encoding the catalytic subunit (Gclc) of the rate-limiting GSH biosynthetic enzyme was detected in Drosophila melanogaster heads. The aim of this study was to determine which clocks in the fly multi-oscillatory circadian system are responsible for Gclc rhythms. Genetic disruption of tissue-specific clocks in D. melanogaster revealed that transcriptional rhythms in Gclc mRNA levels occur independently of the central pacemaker neurons, because these rhythms persisted in heads of behaviourally arrhythmic flies with a disabled central clock but intact peripheral clocks. Disrupting the clock specifically in glial cells abolished rhythmic expression of Gclc, suggesting that glia play an important role in Gclc transcriptional regulation, which may contribute to maintaining homeostasis in the fly nervous system (Chow, 2017).
Sleep is under homeostatic control, whereby increasing wakefulness generates sleep need and triggers sleep drive. However, the molecular and cellular pathways by which sleep need is encoded are poorly understood. In addition, the mechanisms underlying both how and when sleep need is transformed to sleep drive are unknown. Using ex vivo and in vivo imaging, this study shows in Drosophila that astroglial Ca(2+) signaling increases with sleep need. This signaling is dependent on a specific L-type Ca(2+) channel and is necessary for homeostatic sleep rebound. Thermogenetically increasing Ca(2+) in astrocytes induces persistent sleep behavior, and this phenotype was exploited to conduct a genetic screen for genes required for the homeostatic regulation of sleep. From this large-scale screen, TyrRII, a monoaminergic receptor required in astrocytes for sleep homeostasis, was identifed. TyrRII levels rise following sleep deprivation in a Ca(2+)-dependent manner, promoting further increases in astrocytic Ca(2+) and resulting in a positive-feedback loop. Moreover, these findings suggest that astrocytes then transmit this sleep need to a sleep drive circuit by upregulating and releasing the interleukin-1 analog Spätzle, which then acts on Toll receptors on R5 neurons. These findings define astroglial Ca(2+) signaling mechanisms encoding sleep need and reveal dynamic properties of the sleep homeostatic control system (Blum, 2020).
Although astrocytes have been implicated in the homeostatic regulation of sleep, their specific role and the underlying mechanisms are unresolved. The current data support a role for astrocytes as sensors of sleep need and define signaling mechanisms within these cells that mediate the integration and transmission of this information to a downstream homeostatic sleep circuit. In this model, neural activity is sensed by astrocytic processes, leading to an increase in Ca2+ levels, which depends at least in part on specific L-type voltage-gated Ca2+ channels (VGCC). Although astrocytes have been shown to exhibit hyperpolarized membrane potentials with small depolarizations, this particular subtype of L-type VGCC can be activated at substantially lower membrane potentials than other members of this channel family. Interestingly, two recent studies in mice found that intracellular Ca2+ levels in astrocytes vary across sleep/wake states and that Ca2+ signaling in these cells is required for normal sleep architecture and responses to sleep deprivation. These observations suggest a conserved role for astroglial Ca2+ signaling in sleep homeostasis (Blum, 2020).
The current model further suggests that, as the increased neural activity persists, Ca2+-mediated transcription of TyrRII is induced in astrocytes. TyrRII is relatively unstudied, but in vitro data suggest that it responds non-specifically to multiple monoamines. Thus, its upregulation in astrocytes should sensitize these cells to signaling via monoamines, which are intimately associated with wakefulness. The requirement for monoamines in this pathway may provide a logic gate for the system, imparting specificity to the signaling mechanism acting downstream of neural activity, whose semantic properties may be too broad. TyrRII itself is required for further Ca2+ elevations, forming a positive-feedback loop (Blum, 2020).
The data suggest that this amplification of astrocytic Ca2+ signals results in transcriptional upregulation of spz, the fly analog of IL-1. There is an accumulating body of evidence implicating IL-1 in sleep homeostasis in mammals, and the current findings demonstrating a functional role for astrocytic Spz in sleep homeostasis demonstrate that these mechanisms are conserved from invertebrates to vertebrates. In this model, Spz is released from astrocytes under conditions of strong sleep need and transmits this information by signaling to a central sleep drive circuit (the R5 neurons) to promote homeostatic sleep rebound. It is worth noting that fly astrocytes likely possess multiple output mechanisms to regulate sleep, as they not only activate sleep-promoting neurons (R5 neurons) but also inhibit arousal-promoting neurons (l-LNvs) (Blum, 2020).
From a broader perspective, this model draws attention to a fundamental, yet poorly understood, aspect of sleep homeostasis-how a highly dynamic input (i.e., neural activity operating on the millisecond timescale) is integrated and transformed to generate a sleep homeostatic force that functions on a significantly slower timescale. Although the precise identity of the signals embodying sleep need remain unclear, there is substantial experimental and conceptual support for the notion that neural activity increases with wakefulness and is a key trigger for this process. Yet the dynamic mechanisms by which this neural activity and, by extension, sleep need are transformed to sleep drive are unknown. The homeostatic regulation of processes and behaviors involving bistable states, such as sleep versus wakefulness, requires a prominent delay between the detection of the disturbance and the generation of the response. In addition, the stability and switching between such bistable states can be facilitated by positive-feedback loops. It is speculated that the transcription/translation of TyrRII, coupled with the generation of a positive-feedback loop, provide a timing delay followed by a more rapid elevation in astroglial Ca2+ after reaching a set threshold, thus enabling a non-linear response to the continual sampling of sleep need. The transcriptional/translation upregulation of Spz could represent an additional layer of delay (Blum, 2020).
Flies can form sleep-dependent and sleep-independent memory in an appetitive conditioning paradigm, but how they switch between these distinct forms of memory consolidation remains unexplored. This study found that Gr64f GRNs-mediated sweet taste signal is essential for sleep-dependent memory. Sweet taste activates a specific subset of PAM DANs, PAM-β'2mp, for the consolidation of sleep-dependent memory. In contrast, in the absence of sweet stimulus, sleep becomes dispensable for memory consolidation and PAM-α1 DANs are recruited to form long-term memory. These findings indicate that the presence of reward can modulate the role of sleep in memory consolidation (Chouhan, 2022).
Taste serves as an important modality in feeding decisions in Drosophila as perceived sweetness is used to estimate the carbohydrate content of food. Flies prefer sweet sugars over substances with a higher nutritive value indicating that taste determines initial feeding preferences. Sweetness drives sugar ingestion during odor-reward pairing that is essential to form appetitive memories. Also, sweet taste tempers food-seeking behavior as flies on a sweet but non-nutritive medium show lower activity than starved flies. Therefore, the presence of sweet taste modifies behavior by providing a sensation of food (Chouhan, 2022).
Flies kept on food vials require sleep to form appetitive memories (Chouhan, 2021). As in the case of the other behaviors mentioned above, sweet taste appears to be sufficient to signal the presence of food and thereby trigger sleep-dependent memory formation. Flies kept on sucralose, a sweet but non-nutritive sugar, show an enhancement in post-training sleep and require sleep to consolidate long-term memory. In contrast, flies on sorbitol, a tasteless but nutritive substance, form sleep-independent memory. Also, disrupting the activity of sweet-sensing Gr64f+ neurons results in the formation of sleep-independent memory in fed flies, while stimulating Gr64f+ neurons in starved flies induces a switch to sleep-dependent memory consolidation. While sweet taste may induce sleep-dependent memory, it is also possible that processes driving sleep-dependent and sleep-independent memory consolidation interact so as to be mutually exclusive. For instance, as starvation suppresses sleep, it may trigger sleep-independent memory, while sweet taste could drive sleep-dependent memory consolidation by releasing starvation-induced suppression of sleep. Neural connections between the two relevant circuits (described below) could mediate such interactions (Chouhan, 2022).
Sleep is typically thought to be induced under conditions of higher energy consumption; however, the data indicate that, at least for sleep-dependent memory consolidation, sweet taste rather than caloric intake serves as a signal for sleep. Sweet taste is not an accurate reporter for the nutritional value of food. Indeed, flies switch their preference from non-nutritive sweeteners to tasteless nutritive sugars over time in a two-choice assay. Therefore, sweet taste can only provide a short-term impression of the presence of food, which might be sufficient to induce sleep-dependent consolidation as flies form sleep-dependent or sleep-independent memory in the first 4 h after training (Chouhan, 2022).
The reward signal from PAM DANs during training is essential to form odor-reward associations. MB neurons are tiled by individual DAN terminals and the dendrites of corresponding MBONs to form 15 anatomically and functionally distinct compartments. PAM DANs largely innervate distinct compartments of the horizontal MB lobes. As per a previous study, a recurrent circuit that couples PAM-α1 DANs with the corresponding MBON in the α1 MB compartment mediates appetitive memory consolidation in starved flies. However, the current study found that PAM-α1 DANs are dispensable for memory consolidation in fed flies. Instead, PAM DANs that innervate the β'2mp compartment of the MB are essential for long-term memory in fed flies. Consistent with the findings that distinct PAM DANs mediate memory consolidation in fed versus starved states, the activity of PAM-β'2mp DANs was higher in trained and fed flies, while calcium in PAM-α1 DANs was significantly higher in trained and starved flies. These results complement previous work in rats, humans, and flies demonstrating that discrete neural circuits mediate sleep-dependent and sleep-independent memory consolidation (Chouhan, 2022).
PAM-β'2mp DANs signal reward to form 'safety-memory' of the unpaired odor during spaced aversive conditioning. Intriguingly, these neurons connect to MBONs implicated in appetitive memory consolidation under fed and starved conditions. PAM-β'2mp DANs receive input from MBON-γ2α'1 neurons, which are required for memory under fed conditions, and through axo-axonic projections they connect to the MBON-γ1Pedc neurons that are essential for memory under starved conditions. It is proposed that MBON-γ2α'1 neurons engage PAM-β'2mp DAN-mediated reward signaling to establish a sleep-dependent memory trace. By contrast, an axo-axonal connection from MBON-γ1Pedc neurons may modulate neurotransmission in PAM-β'2mp DANs such that it allows PAM-α1 DANs to form sleep-independent memory. Future work can investigate how these discrete neural circuits interact to form sleep-dependent or sleep-independent memories (Chouhan, 2022).
Sweet taste is perceived by a set of seven transmembrane receptors on the proboscis and tarsal segments of the leg. The taste information is transmitted by GRNs that terminate in the suboesophageal ganglion. Second-order neurons of the gustatory system send projections from the suboesophageal ganglion to the central brain for higher-order processing of taste information. A subset of these projections terminates in the superior protocerebrum that also contains processes from DANs. Consistent with a connection between sweet-sensing neurons and DANs, an exposure to sucrose evokes a robust calcium response in PAM DANs. Therefore, sweet taste may signal reward by stimulating PAM DANs. Consistently, a significant calcium/GFP increase in PAM-β'2mp DANs was found in fed flies compared with starved flies. Also, flies kept on arabinose or sucralose after training demonstrate a significant increase in PAM-β'2mp DAN activity compared with flies moved to sorbitol following training. It is suggested that the Gr64f+ GRNs mediated sweet taste signal acts in conjunction with the feedback from MBON-γ2α'1 neurons to recruit PAM-β'2mp DANs to form sleep-dependent memory (Chouhan, 2022).
Rewarding experiences modulate behavior to enhance survival. These experiences are preferentially replayed during sleep for better retention. Indeed, memory consolidation involves neuronal reactivation in the mesolimbic dopaminergic reward system during sleep. Conversely, sleep loss modifies reactivity in the reward centers of the brain such that it induces reward-seeking behavior. This work suggests that the processing of sweet taste reward signals determines the necessity for sleep in memory consolidation. A general role for reward in determining the need for sleep is open to investigation (Chouhan, 2022).
Starvation causes a motivational state that facilitates diverse behaviors such as feeding, walking, and search. Starved Drosophila can form odor/feeding-time associations but the role of starvation in encoding of "time" is poorly understood. This study shows that the extent of starvation is correlated with the fly's ability to establish odor/feeding-time memories. Prolonged starvation promotes odor/feeding-time associations after just a single cycle of reciprocal training. Starvation is also show to be required for acquisition but is dispensable for retrieval of odor/feeding-time memory. Finally, even with extended starvation, a functional circadian oscillator is indispensable for establishing odor/feeding-time memories (Chouhan, 2017).
∂
The small ventral lateral neurons (sLNvs) constitute a central circadian pacemaker in the Drosophila brain. They organize
daily locomotor activity, partly through the release of the neuropeptide pigment-dispersing factor (PDF), coordinating the action of the
remaining clusters required for network synchronization. Despite extensive efforts, the basic
principles underlying communication among circadian clusters remain obscure. This study identified
classical neurotransmitters released by sLNvs through disruption of specific transporters.
Adult-specific RNAi-mediated downregulation of the glycine transporter or
impairment of glycine synthesis in LNv neurons increased period length by nearly an hour without
affecting rhythmicity of locomotor activity. Electrophysiological recordings showed that glycine
reduces spiking frequency in circadian neurons. Interestingly, downregulation of glycine receptor
subunits in specific sLNv targets impaired rhythmicity, revealing involvement of glycine in
information processing within the network. These data identify glycinergic inhibition of specific
targets as a cue that contributes to the synchronization of the circadian network (Frenkel, 2017).
Anatomical and functional evidence indicates that classical neurotransmitters participate along with PDF in the output of the LNvs. In an attempt to define their classical neurotransmitter, this study manipulated the level of membrane or vesicular transporters of known neurotransmitter systems exclusively in this neuronal group. Downregulation of CG5549, the glycine transporter (dGlyT) responsible for recycling this transmitter from the extracellular space, triggered a consistent change in the endogenous period. This effect is specific, as downregulation of List (CG15088, the lithium-inducible SLC6 transporter) did not change any circadian parameter. On the other hand, downregulation of Drosophila Serine hydroxymethyltransferase CG3011 (dShmt; glycine hydroxymethyltransferase activity) also gives rise to period lengthening, consistent with an effect on the circadian clock as opposed to a general effect on LNv viability, which results in progressive loss of rhythmicity. Interestingly, it has been shown that expression of a tethered version of PDF in the LNvs in a condition where neurotransmitters cannot be released increases in almost an hour the endogenous circadian period, suggesting that neurotransmission contributes to rhythm acceleration, lending further support the findings showing that glycine plays such a role in the adult brain (Frenkel, 2017).
Glycinergic inhibitory transmission plays a role in nociception and motor control in the brainstem and spinal cord. Glycine is also a modulator of neuronal excitation mediated by NMDA receptors at glutamatergic synapses in the central brain. This study found glycine in the Drosophila CNS (specifically, in a subset of circadian pacemakers) and found that it stops action potential firing in a postsynaptic target. Interestingly, at least some neurons of the suprachiasmatic nucleus (SCN) are inhibited in the presence of glycine, highlighting another layer of conservation among circadian clocks (Frenkel, 2017).
Glycine receptors are part of the complex cys-loop receptor family of pentameric ligand-gated ion channels. Part of the complexity resides in their ability to assemble specific receptors depending on the subunits recruited, thus leading to both excitatory and inhibitory responses. Little is known about the glycinergic ones, particularly in the fruit fly. Initially, a single gene was reported as the putative glycine receptor GRD. To identify additional glycine subunits, an in silico approach was used. Three putative glycine receptor subunits share conserved features present in ligand-gated chloride channel ones. Interestingly, combined downregulation of those genes in DN1ps revealed they mediate, in part, the response to glycine. Additional subunits are also required to form other types of glycine receptors. GRD was shown to assemble functional GABA receptors in Xenopus oocytes only in the context of Lcch3, highlighting the underlying complexity. Grd is also involved in GABAergic transmission, as its downregulation in specific patterns attenuates the sleep-promoting effects of a GABA-A-R agonist. Thus, depending on the collection of subunits expressed in a particular neuron, GRD takes part of receptors that respond to different neurotransmitters (Frenkel, 2017).
Several scenarios are envisioned accounting for the mismatch between depleting glycine in PDF neurons and the inability to respond to it in circadian targets. If other circadian neurons communicate time-related information through this fast neurotransmitter, downregulating glycine availability in LNvs would surely give rise to a different behavioral output than chronic downregulation of a subset of GlyR subunits in the entire circadian network. Additional non-circadian neurons could also be relevant in defining the properties of locomotor behavior, and the resulting unbalance (derived from inhibition of certain circadian clusters while the non-circadian remain active) would be the cause for the desynchronization; candidate neurons would be the pars intercerebralis. Alternatively, since the three subunits analyzed in this study do not assemble all functional GlyRs, a partial impairment of glycinergic transmission is ensured. In addition, neurons could express different spliced variants of each subunit hampering the efficiency of RNAi. In sum, modifying the stoichiometry among different receptor subunits in discrete neuronal subsets may alter the properties of native glycine receptors in an unpredictable manner (i.e., ligand affinity, ion conductance, and channel kinetics), influencing neuronal responsiveness to glycine and ultimately impinging on neuronal firing. Thus, behavioral complexity ultimately reflects the combination of functional changes in individual neuronal clusters and the orchestration resulting from network interactions (Frenkel, 2017).
Drastically altering neuronal excitability triggered shortening or lengthening of the circadian period, depending on the cluster. Likewise, disrupting glycinergic transmission led to cluster-associated changes in the free-running period. Interestingly, downregulation of single receptor subunits in the DN1ps lengthened the period, further supporting their relevance and their ability to feed back onto the sLNvs. On the other hand, altering glycine transmission onto the LNds+5thsLNv gave rise to a shorter period, a phenotype also observed upon downregulation of all three subunits, in either the E-oscillator or the whole circadian network. Interestingly, a short period phenotype accompanied by deconsolidation of rhythmic activity is a hallmark of pdf01 mutants, in which the LNds are uncoupled from the sLNvs and run at a faster pace; these similarities open the provocative possibility that both PDF and glycine released from the sLNvs play a similar role onto the LNds. Altogether, these results support the notion that each cluster has a differential contribution to the dynamic operation of the circadian network (Frenkel, 2017).
It has been suggested that electrical activity integrates phase information from endogenous oscillators within different regions of the SCN. In that model, the ventral SCN can shift the dorsal SCN and cause it to resynchronize to the new phase, and GABA is required for coupling those two regions. The fact that the sLNvs release an inhibitory neurotransmitter onto dorsal clusters is reminiscent of GABA’s role. In the SCN, Cl- reversal potential changes during the day in different circadian clusters, giving rise to either excitatory or inhibitory responses to GABA; if this were true in Drosophila, glycinergic responses could also change in a time-of-day fashion (Frenkel, 2017).
It is not necessarily envisioned that glycine operates as a synchronizing cue to the molecular clocks, a role already shown to depend on PDF, but instead, it might coordinate the activity of independent clusters to provide coherence to the circadian network. Under this scenario, it is proposed the that sLNvs are acting as an orchestra conductor that relies on at least two batons: one fast inhibiting signal (glycine) and a slower excitatory one (PDF). Thus, the sLNv could operate as a time-of-day switch that rapidly turns off specific targets to keep the circadian network synchronized (Frenkel, 2017).
Antibiotics have been identified as obesogens contributing to the prevalence of obesity. Moreover, their environmental toxicity shows sex dependence, which might also explain the sex-dependent obesity observed. Yet, the direct evidence for such a connection and the underlying mechanisms remain to be explored. In this study, the effects of tetracycline, which is a representative antibiotic found in both environmental and food samples, on Drosophila melanogaster were studied with consideration of both sex and circadian rhythms (represented by the eclosion rhythm). Results showed that in morning-eclosed adults, tetracycline significantly stimulated the body weight of females (AM females) at 0.1, 1.0, 10.0 and 100.0 μg/L, while tetracycline only stimulated the body weight of males (AM males) at 1.0 μg/L. In the afternoon-eclosed adults, tetracycline significantly stimulated the body weight of females (PM females) at 0.1, 1.0 and 100.0 μg/L, while it showed more significant stimulation in males (PM males) at all concentrations. Notably, the stimulation levels were the greatest in PM males among all the adults. The results showed the clear sex dependence of the obesogenic effects, which was diminished by dysrhythmia. Further biochemical assays and clustering analysis suggested that the sex- and rhythm-dependent obesogenic effects resulted from the bias toward lipogenesis against lipolysis. Moreover, they were closely related to the preference for the energy storage forms of lactate and glucose and also to the presence of excessive insulin, with the involvement of glucolipid metabolism. Such relationships indicated potential bridges between the obesogenic effects of pollutants and other diseases, e.g., cancer and diabetes (Guo, 2023).
The physiology and behavior of many organisms are subject to daily cycles. In Drosophila melanogaster the daily locomotion patterns of single flies are characterized by bursts of activity at dawn and dusk. Two distinct clusters of clock neurons-morning oscillators (M cells) and evening oscillators (E cells)-are largely responsible for these activity bursts. In contrast, male-female pairs of flies follow a distinct pattern, most notably characterized by an activity trough at dusk followed by a high level of male courtship during the night. This male sex drive rhythm (MSDR) is mediated by the M cells along with DN1 neurons, a cluster of clock neurons located in the dorsal posterior region of the brain. This study reports that males lacking Salt-inducible kinase 3 (SIK3) expression in M cells exhibit a short period of MSDR but a long period of single-fly locomotor rhythm (SLR). Moreover, lack of Sik3 in M cells decreases the amplitude of Period (Per) cycling in DN1 neurons, suggesting that SIK3 non-cell-autonomously regulates DN1 neurons' molecular clock. This study also shows that Sik3 reduction interferes with circadian nucleocytoplasmic shuttling of Histone deacetylase 4 (HDAC4), a SIK3 phosphorylation target, in clock neurons and that constitutive HDAC4 localization in the nucleus shortens the period of MSDR. Taking these findings together, it is concluded that SIK3-HDAC4 signaling in M cells regulates MSDR by regulating the molecular oscillation in DN1 neurons (Fujii, 2017).
The physiology and behavior of most animals undergo daily oscillations, which are controlled by a small set of clock neurons in the brain. In mammals, a heterodimeric complex between CLOCK (CLK) and BMAL1 activates transcription of Period (Per1 and Per2) and Cryptochrome (Cry1 and Cry2) genes, and their protein products in turn inhibit the activity of CLK/BMAL1. Likewise, Drosophila heterodimeric complexes between CLK and CYCLE (CYC) activate the genes period (per) and timeless (tim), and their respective protein products repress CLK/CYC. These conserved negative-feedback loops, which include several kinases, produce rhythmic transcription profiles in numerous genes (Fujii, 2017).
The Drosophila brain contains ∼150 clock neurons which are divided into seven clusters based on their anatomical locations and functional characteristics: the small and large ventral lateral neurons (sLNvs and lLNvs), the dorsal lateral neurons (LNds), the lateral posterior neurons (LPNs), and three dorsal neuron clusters (DN1-3). Some of these clock neurons have distinct functions in circadian locomotor behavior. Specifically, four sLNvs, also referred to as 'morning' (M) cells, express the neuropeptide pigment-dispersing factor (PDF) and control the timing of morning locomotor activity during light:dark (LD) cycles; these neurons are also the key pacemaker neurons in constant darkness (DD). The fifth, PDF−, sLNv and the LNds, referred to as 'evening' (E) cells, are required for the generation of the evening activity peak in LD cycles. Communication between various groups of cells within this interconnected neural network enhances the synchrony of molecular oscillation in each neuron (Fujii, 2017).
Ventral lateral neuron (LNv)-derived PDF plays a critical role in regulating the molecular clock. Specifically, PDF participates in synchronization of clock neurons by up-regulating cAMP, which activates PKA, which in turn regulates the stability of PER and TIM in PDF receptor (PDFR)-expressing target neurons. Thus, ion status and is regu neurosecretory signaling: In well-fed flies, SIK3 is thought to be indirectly activated by insulin-likeavioral rhythms even when flies are kept in DD (Fujii, 2017).
Locomotor activity is the best-characterized circadian behavior in Drosophila, but numerous other behaviors, such as courtship and mating, sleep, and feeding, are under strong circadian influence. Previously work has shown that male-female pairs of flies exhibit activity patterns strikingly distinct from those of singly kept males or females (i.e., single-fly locomotor rhythm or SLR) or same-sex pairs of flies. The activity pattern of male-female pairs, which is referred to as 'male sex-drive rhythm' (MSDR), is characterized by a trough at subjective dusk, followed by a sharp increase in male-driven courtship activity (especially 'following' behavior) that peaks during the subjective night. A functional molecular clock in both Pdf+ LNvs and DN1 neurons is necessary and sufficient for proper MSDR. However, few other cellular and molecular components contributing to MSDR have been identified to date. Specifically, information is lacking about both the molecular and cellular identity of downstream effectors of the main clock components that are important for MSDR (Fujii, 2017).
This study reports the identification of two downstream effectors of the molecular clock that play distinct roles in MSDR and SLR. Using an RNAi screen for kinases, this study shows that Salt-inducible kinase 3 (SIK3) is a critical component for circadian behavior. Sik3 knockdown in subsets of clock neurons (DN1 neurons or Pdf+ LNvs) causes a short period of MSDR, whereas the period length of SLR is slightly shortened with Sik3 knockdown in DN1 neurons and is slightly elongated with Sik3 knockdown in sLNvs. This study also found that transcriptional activity of Histone deacetylase 4 (HDAC4) is regulated by SIK3 in a circadian manner. Finally, Sik3 reduction in Pdf+ LNvs reduces the amplitude of PER oscillation in DN1 neurons and shortens the length of the MSDR period, suggesting that SIK3-HDAC4 signaling plays an important role in the determination of MSDR period by modulating the intercellular communication between clock neurons (Fujii, 2017).
SIK-HDAC (class IIa) signaling is evolutionarily conserved from worm to mammals, operating in a number of tissues, including the nervous system, liver, and muscle. In mice, SIK1-HDAC signaling is important for muscle integrity by regulating the activity of the transcription factor MEF2 (Stewart, 2013). In the fly, SIK3-HDAC4 signaling was shown to control the expression of lipolytic and gluconeogenic genes in the fat body (Choi, 2015). Furthermore, both Drosophila HDAC4 and MEF2 have been implicated in circadian rhythm, as has the related HDAC5 gene in mice. This paper has established a critical role for SIK3 in two circadian behaviors, single-fly locomotor activity and male sex drive, respectively (Fujii, 2017).
MSDR is mediated through the activity of Pdf+ LNvs and DN1 neurons (Fujii, 2010). This study specifically targeted SIK3 in either group of circadian neurons using RNAi. Strikingly, SIK3 knockdown in LNvs shortened the period length of MSDR but slightly yet reproducibly lengthened that of SLR. The loss of PER rhythm amplitude observed specifically in the DN1 neurons and its apparent phase advance on the second or third day of constant conditions would fit with these observations. The advance would be symptomatic of DN1 neurons free-running with a short period and thus presumably explains the short-period MSDR. The loss of amplitude could indicate that a small subset of DN1 neurons runs at a different pace, perhaps explaining the slightly long period of the SLR. Indeed, both SLR and MSDR depend on sLNvs driving DN1 neurons. The broader M peak might also be an early sign that DN1 neurons are not as coherent, even under LD, because the DN1 neurons function downstream of the sLNvs to control morning anticipatory activity. The same short-period DN1 neurons might drive the M peak and MSDR. Unfortunately, the amplitude of the M peak in DD was too low to be able to determine whether it free-runs with a short period (Fujii, 2017).
Knockdown of SIK3 in DN1 neurons shortened the period length of MSDR that is well correlated with shortened PER oscillations in DN1 neurons, and these flies show subtle but significantly shortened period length in SLR. Together, these findings suggest that SIK3 is a key component in molecular oscillator coupling between sLNvs and DN1 neurons and that its role is especially important for maintaining an appropriate MSDR period length. However, the possibility cannot be excluded that SIK3 also influences the period of the circadian pacemaker in a neuron-specific manner (i.e., in the DN1 neurons), as was proposed for SGG and CKII.
It was also demonstrated that HDAC4 cycles in a SIK3-dependent fashion between the cytoplasm and the nucleus in the M cells (Fujii, 2017).
Because M-cell restricted overexpression of phosphorylation-defective, constitutively nuclear-located HDAC43A, but not wild-type HDAC4, mimics the phenotype of flies lacking SIK3 in these cells, it is suggested that HDAC4 is a critical component for the transduction of the circadian intercellular signal from M cells to DN1 neurons. However, the function of SIK3 in oscillator coupling is unlikely to be mediated by HDAC4 in DN1 neurons, because most of these neurons do not express HDAC4. Another potential SIK3 phosphorylation target such as CREB or CRTC, which are implicated in cAMP-mediated signaling and the circadian clock, may play a role in the regulation of oscillator coupling in DN1 neurons for MSDR (Fujii, 2017).
SIK3-dependent circadian shuttling of HDAC4 in sLNvs implies that the activity of SIK3 is under circadian control. How is SIK3 activity regulated in sLNvs? In fat cells (and rat adipocytes) SIK3 activity is dependent on nutrition status and is regulated indirectly through neurosecretory signaling: In well-fed flies, SIK3 is thought to be indirectly activated by insulin-like peptides (ILPs), whereas in starved flies, it is inhibited by adipokinetic hormone (AKH). SIK3 activity itself is regulated via phosphorylation by AKT1 (activated by ILPs) and cAMP-dependent protein kinase A (PKA) (activated by AKH). These kinases target distinct but overlapping sets of serine and threonine residues, and thus it appears that SIK3 activity is dependent on the particular phosphorylation pattern at these sites. Intriguingly, it has been reported that PDF stabilizes PER by increasing cAMP levels and PKA activity in Pdfr+ clock neurons (including M cells) at dawn, a time when HDAC4 is activated and translocated into the nucleus. PDF thus could be an indirect circadian regulator of SIK3 activity via PKA. However, the reduction of PDF in M cells did not shorten the MSDR period length, suggesting that PDF signaling probably does not regulate SIK3. Moreover, RNAi-mediated knockdown of MEF2, which regulates SIK3-HDAC in the mouse, had no effect on MSDR. Future experiments will be needed to investigate how SIK3 activity is regulated and how HDAC4 controls intercellular communications between M cells and DN1 neurons (Fujii, 2017).
How does the lack of SIK3 in M cells (i.e., sLNvs) alter the robustness of PER cycling in some (DN1 neurons) but not other (sLNvs and LNds) PDFR-expressing clock cells? One possibility might be the manner by which sLNvs communicate with other clock cells. Functional and anatomical studies including GFP Reconstitution Across Synaptic Partners strongly suggest that at least some DN1 neurons are direct downstream targets of sLNvs, and hence accurately timed communication between these neurons likely occurs through synapses, which is proposed to rely on SIK3 function in LNvs. In contrast, autocrine (sLNvs) and paracrine (LNds) communication likely occurs via untargeted release of PDF, a process that is suggested not to be dependent on SIK3. The projections of sLNvs to DN1 neurons, in addition to PDF-containing dense core vesicles, harbor small clear vesicles that house classical neurotransmitters, raising the possibility that communication between sLNvs and DN1 neurons pertinent to the robust amplitude of PER oscillation in DN1 neurons is mediated by an as yet unidentified HDAC4-dependent signal. Moreover, DN1 neurons are probably heterogeneous in function, and thus it is quite likely that only some of these cells respond to the sLNv-derived and SIK3-HDAC4-dependent signal, whereas another either overlapping or entirely distinct group of DN1 neurons is responsive to the sLNv-derived PDF. In this context, it is worth noting that sLNvs also express the small neuropeptide F (sNPF). Moreover, a discrete requirement for both PDF-mediated and classical neurotransmitter signaling has been proposed for distinct aspects of SLR, and glycine in sLNvs was recently proposed to coordinate locomotor behavior and appears either to accelerate or to slow down circadian oscillators in specific neuronal groups. Future studies will be necessary to identify the LNv-derived signal that maintains the appropriate amplitude and speed of the clock in DN1 neurons to coordinate MSDR and SLR (Fujii, 2017).
It is surprising that the loss of SIK3 in sLNvs results in a long SLR and a short MSDR, whereas the loss of SIK3 in the DN1 neurons shortens both SLR and MSDR, because in either case it appears that the DN1 neurons are disconnected from the sLNvs. One explanation could be that SIK3 is differentially modulated in different subpopulations of DN1 neurons by the sLNv synchronizing cue, thus resulting in DN1 desynchronization in flies lacking SIK3 in the sLNvs. However, when SIK3 is missing in DN1 neurons, they all adopt a short period by default (Fujii, 2017).
In summary, this work unexpectedly reveals the existence of a SIK3-HDAC4 regulatory pathway that allows the M cells -- the critical circadian pacemaker neurons of the fly brain -- to control specific circadian neurons and behaviors. This pathway could prove particularly important in explaining how circadian behaviors can be differentially modulated in response to environmental conditions or internal states. Indeed, the ability to tune and prioritize specific behaviors in a daily manner to minimize energy expenditure and to maximize fitness and reproductive output is critical for animals. Given the strong neural and molecular homologies between the circadian system of fruit flies and mammals, it will be particularly interesting to determine whether the SIK-HDAC pathway is also active in VIP (vasoactive intestinal polypeptide-expressing) neurons of the mammalian suprachiasmatic nucleus and, if so, whether it also controls specific circadian behaviors (Fujii, 2017).
This study investigated the function of Drosophila miR-210 in circadian behaviour by misexpressing it within circadian clock cells. Manipulation of miR-210 expression levels in the PDF (pigment dispersing factor) positive neurons affected the phase of locomotor activity, under both light-dark conditions and constant darkness. PER cyclical expression was not affected in clock neurons, however, when miR-210 was up-regulated, a dramatic alteration in the morphology of PDF ventral lateral neuron (LNv) arborisations was observed. A transcriptomic analysis revealed that miR-210 overexpression affects the expression of several genes belonging to pathways related to circadian processes, neuronal development, GTPases signal transduction and photoreception. Collectively, these data reveal the role of miR-210 in modulating circadian outputs in flies and guiding/remodelling PDF positive LNv arborisations and indicate that miR-210 may have pleiotropic effects on the clock, light perception and neuronal development (Cusumano, 2018)
This study examined the possibility to distinguish four extreme chronotypes among fruit flies and the possibility of the differential response of such chronotypes to light and heat stressors. Circadian rhythms of locomotor activity and sleep-wake pattern were tested in constant darkness, and four strains of fruit flies originating from three wild populations of Africa, Europe, and the USA were selected to represent four distinct chronotypes: "larks" (early morning and evening activity peaks), "owls" (late morning and evening peaks), "swifts" (early morning and late evening peaks), and "woodcocks" (late morning and early evening peaks). The circadian rhythms and sleep efficiency of the selected chronotypes were further tested under such extreme conditions as either long day (LD20:4 at 20 ° C) or a combination of LD20:4 with hot temperature (29 ° C). Despite the identity of such experimental conditions for four chronotypes, their circadian rhythms and sleep timing showed significantly distinct patterns of response to exposure to heat and/or long days. All two-way repeated measures analysis of variances yielded a significant interaction between chronotype and time of the day. It is concluded that an experimental study of heritable chronotypes in the fruit fly can facilitate a search for genetic underpinnings of individual variation in vulnerability to circadian misalignment, maladaptive sleep-wake behavior, and sleep disorders (Zakharenko, 2018).
Seven neuropeptides are expressed within the Drosophila brain circadian network. Previous mRNA profiling suggested that Allatostatin-C (AstC) is an eighth neuropeptide and specifically expressed in dorsal clock neurons (DN1s). The results of this study show that AstC is, indeed, expressed in DN1s, where it oscillates. AstC is also expressed in two less well-characterized circadian neuronal clusters, the DN3s and lateral-posterior neurons (LPNs). Behavioral experiments indicate that clock-neuron-derived AstC is required to mediate evening locomotor activity under short (winter-like) and long (summer-like) photoperiods. The AstC-Receptor 2 (AstC-R2) is expressed in LNds, the clock neurons that drive evening locomotor activity, and AstC-R2 is required in these neurons to modulate the same short photoperiod evening phenotype. Ex vivo calcium imaging indicates that AstC directly inhibits a single LNd. The results suggest that a novel AstC/AstC-R2 signaling pathway, from dorsal circadian neurons to an LNd, regulates the evening phase in Drosophila (Diaz, 2019).
To learn more about how the ~150 clock neurons within the adult fly brain communicate, RNA-sequencing data were examined from the LNds, LNvs, and DN1s for neuropeptides not yet associated with this circuitry. AstC was a promising candidate, because mRNAs encoding both the peptide and one of its receptors (AstC-R2) were identified within the three clock neuron clusters; these data suggested a novel intra-clock circuitry signaling pathway. AstC transcripts as well as the neuropeptide are, indeed, well expressed in the DN1s, and the neuropeptide signal undergoes strong cycling in DD as well as LD conditions. Moreover, AstC is also expressed in two other circadian neuron subgroups, the DN3s and the LPNs, and it is the first neuropeptide identified in these circadian clusters. Behavioral data after RNAi knockdown indicate that the AstC binds to AstC-R2 expressed in E-cells to modulate the timing of evening locomotor activity. Ex vivo calcium imaging indicates that AstC directly inhibits a single LNd (Diaz, 2019).
AstC is required in the clock neurons to regulate the evening locomotor activity phase in short and long photoperiods, suggesting that 'masking' effects in 12:12 LD obscured a phenotypic effect. This shift in the timing of the E-peak occurs when AstC is reduced in all circadian neurons (tim-GAL4 driver) (Diaz, 2019).
AstC cycles in the DN1s, not only under standard 12:12 LD conditions but also under DD conditions and the short photoperiod condition of 6:18 LD. In all cases, AstC staining intensity in the DN1 soma is lowest during the early day. It was hypothesizef that the AstC staining is reduced at this time, because the peptide is being transported from the soma to the dendritic arbors for secretion. Indeed, this early-day timing correlates with DN1 firing, as DN calcium and firing frequency are highest at this time and, thus, may be associated with activity-dependent peptide release. This temporal regulation could coincide with the timing of the AstC effect on the phase of the evening locomotor peak. If this is the case, however, residual AstC remaining after knockdown in the DN1s with the clk4.1M-GAL4 driver must still be sufficient to promote a wild-type phenotype (Diaz, 2019).
In addition, the LPNs are probably not a key circadian source of AstC: their AstC levels were also dramatically reduced in the clk856-GAL4-mediated knockdown without any phenotypic effect. All AstC-expressing DN3s are targeted by the tim-GAL4 driver, yet most of these DN3s are not included in the clk856-GAL4 driver. Although a tim-GAL4 source from outside the circadian network cannot be excluded, these results suggest that the DN3s are the key source of AstC. This tentative conclusion is based on negative data, and the lack of a DN3-specific GAL4 driver makes it impossible to test this model directly. Therefore, three possible models are proposed: (1) the DN3s are the primary source of AstC within the circadian circuit; (2) the DN1s, DN3s, and LPNs, or some combination, are functionally redundant; or (3) a small amount of residual AstC within DN1s is sufficient for its behavioral role in the evening activity peak assay. Although neuron-specific deletion of AstC might contribute to distinguishing between these three possibilities, no accurate and efficient CRISPR-based strategy is available for achieving temporal and spatial specificity (Diaz, 2019).
Once released by dorsal circadian neurons, AstC signals to the LNds via binding to its receptor, AstC-R2. This is because AstC-R2 knockdown in the LNds and knockdown of AstC in the entire circadian circuit give rise to the identical delayed E-peak phenotype. Moreover, the DNs and the LNds both extend projections to the dorsal protocerebrum region (near the pars intercerebralis), where they come in close proximity. In summary, this study adds a new player to the neuronal circuitry governing E-peak modulation. It is suggested that the LNd neuronal activity is modulated not only by signaling from M-cells but also by AstC signaling from the dorsal region of the brain (Diaz, 2019).
The functional imaging strongly indicates that AstC binding to LNd-localized AstC-R2 leads to neuronal inhibition. The effect is consistent with several previous electrophysiology experiments and contributes to an emerging theme of LNd inhibition. It is also one of the first pieces of evidence indicating that the DNs can be a source of inhibition onto the LNds. Interestingly, only a single LNd is directly AstC sensitive, further attesting to LNd heterogeneity and suggesting that behavioral regulation of the evening phase arises from this signaling to a single LNd. It is suggested that this response is communicated directly to the rest of the LNds - for example, via gap junctions - but a more indirect and circuitous route of communication cannot be excluded (Diaz, 2019).
It is noted that there are several caveats to the current model. The AstC and AstC-R2 knockdown experiments were conducted with only a single RNAi line each, due to the lack of additional functional RNAi lines. However, the two experiments show essentially identical phenotypes, suggesting that off-target effects are unlikely to pose a problem. It was not possible to rule out that the AstC knockdown phenotypes are not due to a developmental requirement. Experiments to address this point are challenging, because when the temperature is raised to reduce tubgal80ts repression and allow for an adult-only knockdown, the heat itself dramatically changes the E-peak timing. Lastly, lack of LPN- and DN3-specific drivers precluded addressing whether DN3-derived AstC is required for this evening activity peak modulation and whether the DN3s can directly inhibit the LNds (Diaz, 2019).
Although the AstC peptide sequence is highly conserved among insect species, only the AstC-R2 receptor has a mammalian homolog, the somatostatin-galanin-opioid receptor family. Inhibitory somatostatin (SST) interneurons are present in the mammalian equivalent of a central core clock, the suprachiasmatic nucleus (SCN). SST interneurons are also known to affect sleep and circadian behaviors. Interestingly, SST is associated with proper adaptation under photoperiod conditions for both diurnal and nocturnal mammals, suggesting a highly conserved function with AstC/AstC-R2 for adaptation under different equinox environments. It will be interesting to see whether the SCN-resident SST interneurons are important for this adaptation, like the AstC-containing clock neurons described in this study (Diaz, 2019).
Sleep is nearly universal among animals, yet remains poorly understood. Recent work has leveraged simple model organisms, such as Caenorhabditis elegans and Drosophila melanogaster larvae, to investigate the genetic and neural bases of sleep. However, manual methods of recording sleep behavior in these systems are labor intensive and low in throughput. To address these limitations, this study developed methods for quantitative imaging of individual animals cultivated in custom microfabricated multiwell substrates, and used them to elucidate molecular mechanisms underlying sleep. This paper describes the steps necessary to design, produce, and image these plates, as well as analyze the resulting behavioral data. Approaches are described for experimentally manipulating sleep. Following these procedures, after ~2 h of experimental preparation, it is possible to simultaneously image 24 C. elegans from the second larval stage to adult stages or 20 Drosophila larvae during the second instar life stage at a spatial resolution of 10 or 27 microm, respectively. Although this system has been optimized to measure activity and quiescence in Caenorhabditis larvae and adults and in Drosophila larvae, it can also be used to assess other behaviors over short or long periods. Moreover, with minor modifications, it can be adapted for the behavioral monitoring of a wide range of small animals (Churgin, 2019).
The stability of circadian clock mechanisms under cyclic environments contributes to increased Darwinian fitness by accurately timing daily behavior and physiology. Earlier studies on biological clocks speculated that the timing of behavior and its accuracy are determined by the intrinsic period (tau) of the circadian clock under constant conditions, its stability, the period of the external cycle (T), and resetting of the clock by environmental time cues. However, most of these previous studies suffered from certain limitations, the major ones being a narrow range of examined tau values and a non-uniformity in the genetic background across the individuals tested. The data in this study rigorously test the following hypotheses by employing Drosophila melanogaster fruit flies with tau ranging from 17 to 30 h in a uniform genetic background. Tests were performed to see whether 1) precision (day-to-day stability of tau) is greater for clocks with tau close to 24 h; 2) accuracy (i.e., day-to-day stability of the phase relationship (psi), where psi is the duration between a phase of the rhythm and a phase of the external cycle) is greater for clocks with tau close to 24 h; 3) Psi is delayed with an increase in tau; and 4) Psi becomes more advanced with an increase in length of zeitgeber cycle (T). This study shows that precision is not always maximum for ~24-h clocks, but that accuracy is greatest when tau approximates T. Further, flies exhibit a delayed phase relationship with increasing tau and an advanced phase relationship under long T-cycles as compared with shorter T-cycles. Relationships between activity and rest durations are described and how these observations fit predictions from models of circadian entrainment. Overall, it is confirmed that accuracy and phase of entrained rhythm are governed by both intrinsic clock period and the length of the external cycle; however, it was found that the relationship between intrinsic period and precision does not fit previous predictions (Srivastava, 2019).
Recent work suggests that the circadian pacemaker responds optimally to millisecond flashes of light, not continuous light exposure as has been historically believed. It is unclear whether these responses are influenced by the physical characteristics of the pulsing. In the present study, Drosophila (n = 2199) were stimulated with 8, 16 or 120 ms flashes. For each duration, the energy content of the exposure was systematically varied by changing the pulse irradiance and the number of stimuli delivered over a fixed 15 min administration window (64 protocols surveyed in all). Results showed that per microjoule invested, 8 ms flashes were more effective at resetting the circadian activity rhythm than 16- and 120 ms flashes (i.e. left shift of the dose-response curve, as well as a higher estimated maximal response). These data suggest that the circadian pacemaker's photosensitivity declines within milliseconds of light contact. Further introduction of light beyond a floor of (at least) 8 ms leads to diminishing returns on phase-shifting (Kaladchibachi, 2019).
Drosophila Myc (dMyc) is highly conserved and functions as a transcription factor similar to mammalian Myc. Previous work has found that oncogenic Myc disrupts the molecular clock in cancer cells. This study demonstrates that misregulation of dMyc expression affects Drosophila circadian behavior. dMyc overexpression results in a high percentage of arrhythmic flies, concomitant with increases in the expression of clock genes cyc, tim, cry, and cwo. Conversely, flies with hypomorphic mutations in dMyc exhibit considerable arrhythmia, which can be rescued by loss of dMnt, a suppressor of dMyc activity. Metabolic profiling of fly heads revealed that loss of dMyc and its overexpression alter steady-state metabolite levels and have opposing effects on histidine, the histamine precursor, which is rescued in dMyc mutants by ablation of dMnt and could contribute to effects of dMyc on locomotor behavior. These results demonstrate a role of dMyc in modulating Drosophila circadian clock, behavior, and metabolism (Hsieh, 2019).
Nearly all organisms evolved endogenous self-sustained timekeeping mechanisms to track and anticipate cyclic changes in the environment. Circadian clocks, with a periodicity of about 24 h, allow animals to adapt to day-night cycles. Biological clocks are highly adaptive, but strong behavioral rhythms might be a disadvantage for adaptation to weakly rhythmic environments such as polar areas. Several high-latitude species, including Drosophila species, were found to be highly arrhythmic under constant conditions. Furthermore, Drosophila species from subarctic regions can extend evening activity until dusk under long days. These traits depend on the clock network neurochemistry, and it has been previously proposed that high-latitude Drosophila species evolved specific clock adaptations to colonize polar regions. This study broadened this analysis to 3 species of the Chymomyza genus, which diverged circa 5 million years before the Drosophila radiation and colonized both low and high latitudes. C. costata, pararufithorax, and procnemis, independently of their latitude of origin, possess the clock neuronal network of low-latitude Drosophila species, and their locomotor activity does not track dusk under long photoperiods. Nevertheless, the high-latitude C. costata becomes arrhythmic under constant darkness (DD), whereas the two low-latitude species remain rhythmic. Different mechanisms are behind the arrhythmicity in DD of C. costata and the high-latitude Drosophila ezoana, suggesting that the ability to maintain behavioral rhythms has been lost more than once during drosophilid evolution and that it might indeed be an evolutionary adaptation for life at high latitudes (Bertolini, 2019).
Organisms possess an endogenous molecular clock which enables them to adapt to environmental rhythms and to synchronize their metabolism and behavior accordingly. Circadian rhythms govern daily oscillations in numerous physiological processes. Drosophila larvae have relatively simple nervous system compared to their adult counterparts, yet they both share a homologous molecular clock with mammals, governed by interlocking transcriptional feedback loops with highly conserved constituents. Larvae exhibit a robust light avoidance behavior, presumably enabling them to avoid predators and desiccation, and DNA-damage by exposure to ultraviolet light, hence are crucial for survival. Circadian rhythm has been shown to alter light-dark preference, however it remains unclear how distinct behavioral strategies are modulated by circadian time. To address this question, this study investigate the larval visual navigation at different time-points of the day employing a computer-based tracking system, which allows detailed evaluation of distinct navigation strategies. The results show that due to circadian modulation specific to light information processing, larvae avoid light most efficiently at dawn, and a functioning clock mechanism at both molecular and neuro-signaling level is necessary to conduct this modulation (Asirim, 2020).
Natural genetic variation affects circadian rhythms across the evolutionary tree, but the underlying molecular mechanisms are poorly understood. This study investigated population-level, molecular circadian clock variation by generating >700 tissue-specific transcriptomes of Drosophila melanogaster (w(1118)) and 141 Drosophila Genetic Reference Panel (DGRP) lines. This comprehensive circadian gene expression atlas contains >1700 cycling genes including previously unknown central circadian clock components and tissue-specific regulators. Furthermore, >30% of DGRP lines exhibited aberrant circadian gene expression, revealing abundant genetic variation-mediated, intertissue circadian expression desynchrony. Genetic analysis of one line with the strongest deviating circadian expression uncovered a novel cry mutation that, as shown by protein structural modeling and brain immunohistochemistry, disrupts the light-driven flavin adenine dinucleotide cofactor photoreduction, providing in vivo support for the importance of this conserved photoentrainment mechanism. Together, this study revealed pervasive tissue-specific circadian expression variation with genetic variants acting upon tissue-specific regulatory networks to generate local gene expression oscillations (Litovchenko, 2021).
Sleep is vital for survival. Yet under environmentally challenging conditions, such as starvation, animals suppress their need for sleep. Interestingly, starvation-induced sleep loss does not evoke a subsequent sleep rebound. Little is known about how starvation-induced sleep deprivation differs from other types of sleep loss, or why some sleep functions become dispensable during starvation. This study demonstrate that down-regulation of the secreted cytokine unpaired 2 (upd2) in Drosophila flies may mimic a starved-like state. A genetic knockdown strategy to investigate the consequences of upd2 on visual attention and sleep in otherwise well-fed flies, thereby sidestepping the negative side effects of undernourishment. Knockdown of upd2 in the fat body (FB) is sufficient to suppress sleep and promote feeding-related behaviors while also improving selective visual attention. Furthermore, this study shows that this peripheral signal is integrated in the fly brain via insulin-expressing cells. Together, these findings identify a role for peripheral tissue-to-brain interactions in the simultaneous regulation of sleep quality and attention, to potentially promote adaptive behaviors necessary for survival in hungry animals (Ertekin, 2020).
Sleep deprivation has been shown to negatively impact health outcomes, leading to decreased immune responses, memory loss, increased activity of stress and inflammatory pathways, weight gain, and even behavioral changes. These observations suggest that sleep deprivation substantially interferes with important physiological functions, including metabolic pathways of energy utilization. Many of those phenotypes are correlated with age, suggesting that disrupted sleep may interfere with the aging process. However, little is known about how sleep disruption affects aging and longevity. This study investigated this relationship using eight representative fruit fly lines from the Sleep Inbred Panel (SIP). The SIP consists of 39 inbred lines that display extreme short- and long-sleep patterns, and constitutes a crucial Drosophila community resource for investigating the mechanisms of sleep regulation. The data show that flies with short-sleep periods have ∼16% longer life span, as well as reduced aging rate, compared to flies with long-sleep. In contrast, disrupting normal circadian rhythm reduces fly longevity. Short-sleep SIP flies moreover show slight metabolic differences to long-sleep lines, and to flies with disrupted circadian rhythm. These data suggest that the inbred SIP lines engage sleep mechanisms that are distinct from the circadian clock system (Thompson, 2020).
Globally, neonicotinoids are the most used insecticides, despite their well-documented sub-lethal effects on beneficial insects. Neonicotinoids are nicotinic acetylcholine receptor agonists. Memory, circadian rhythmicity and sleep are essential for efficient foraging and pollination and require nicotinic acetylcholine receptor signalling. The effect of field-relevant concentrations of the European Union-banned neonicotinoids: imidacloprid, clothianidin, thiamethoxam and thiacloprid were tested on Drosophila memory, circadian rhythms and sleep. Field-relevant concentrations of imidacloprid, clothianidin and thiamethoxam disrupted learning, behavioural rhythmicity and sleep whilst thiacloprid exposure only affected sleep. Exposure to imidacloprid and clothianidin prevented the day/night remodelling and accumulation of pigment dispersing factor (PDF) neuropeptide in the dorsal terminals of clock neurons. Knockdown of the neonicotinoid susceptible Dα1 and D&beta2 nicotinic acetylcholine receptor subunits in the mushroom bodies or clock neurons recapitulated the neonicotinoid like deficits in memory or sleep/circadian behaviour respectively. Disruption of learning, circadian rhythmicity and sleep are likely to have far-reaching detrimental effects on beneficial insects in the field (Tasman, 2021).
The adaptive significance of adjusting behavioral activities to the right time of the day seems obvious. Laboratory studies implicated an important role of circadian clocks in behavioral timing and rhythmicity. Yet, recent studies on clock-mutant animals questioned this importance under more naturalistic settings, as various clock mutants showed nearly normal diel activity rhythms under seminatural zeitgeber conditions. This study reports evidence that proper timing of eclosion, a vital behavior of the fruit fly Drosophila melanogaster, requires a functional molecular clock under quasi-natural conditions. In contrast to wild-type flies, period(01) mutants with a defective molecular clock showed impaired rhythmicity and gating in a temperate environment even in the presence of a full complement of abiotic zeitgebers. Although period(01) mutants still eclosed during a certain time window during the day, this time window was much broader and loosely defined, and rhythmicity was lower or lost as classified by various statistical measures. Moreover, peak eclosion time became more susceptible to variable day-to-day changes of light. In contrast, flies with impaired peptidergic interclock signaling (Pdf(01) and han5304 PDF receptor mutants) eclosed mostly rhythmically with normal gate sizes, similar to wild-type controls. The results suggest that the presence of natural zeitgebers is not sufficient, and a functional molecular clock is required to induce stable temporal eclosion patterns in flies under temperate conditions with considerable day-to-day variation in light intensity and temperature. Temperate zeitgebers are, however, sufficient to functionally rescue a loss of PDF-mediated clock-internal and -output signaling (Ruf, 2021).
Memory is initially labile but can be consolidated into stable long-term memory (LTM) that is stored in the brain for extended periods. Despite recent progress, the molecular and cellular mechanisms underlying the intriguing neurobiological processes of LTM remain incompletely understood. Using the Drosophila courtship conditioning assay as a memory paradigm, this study showed that the LIM homeodomain (LIM-HD) transcription factor Apterous (Ap), which is known to regulate various developmental events, is required for both the consolidation and maintenance of LTM. Interestingly, Ap is involved in these 2 memory processes through distinct mechanisms in different neuronal subsets in the adult brain. Ap and its cofactor Chip (Chi) are indispensable for LTM maintenance in the Drosophila memory center, the mushroom bodies (MBs). On the other hand, Ap plays a crucial role in memory consolidation in a Chi-independent manner in pigment dispersing factor (Pdf)-containing large ventral-lateral clock neurons (l-LNvs) that modulate behavioral arousal and sleep. Since disrupted neurotransmission and electrical silencing in clock neurons impair memory consolidation, Ap is suggested to contribute to the stabilization of memory by ensuring the excitability of l-LNvs. Indeed, ex vivo imaging revealed that a reduced function of Ap, but not Chi, results in exaggerated Cl- responses to the inhibitory neurotransmitter gamma-aminobutyric acid (GABA) in l-LNvs, indicating that wild-type (WT) Ap maintains high l-LNv excitability by suppressing the GABA response. Consistently, enhancing the excitability of l-LNvs by knocking down GABAA receptors compensates for the impaired memory consolidation in ap null mutants. Overall, these results revealed unique dual functions of the developmental regulator Ap for LTM consolidation in clock neurons and LTM maintenance in MBs (Inami, 2021).
Sleep is a universally conserved physiological state which contributes toward basic organismal functions, including cognitive operations such as learning and memory. Intriguingly, organisms can sometimes form memory even without sleep, such that Drosophila display sleep-dependent and sleep-independent memory in an olfactory appetitive training paradigm. Sleep-dependent memory can be elicited by the perception of sweet taste, and this study now shows that a mixed-sex population of flies maintained on sorbitol, a tasteless but nutritive substance, do not require sleep for memory consolidation. Consistent with this, silencing sugar-sensing gustatory receptor neurons in fed flies triggers a switch to sleep-independent memory consolidation, whereas activating sugar-sensing gustatory receptor neurons results in the formation of sleep-dependent memory in starved flies. Sleep-dependent and sleep-independent memory relies on distinct subsets of reward signaling protocerebral anterior medial dopaminergic neurons (PAM DANs) such that PAM-β'2mp DANs mediate memory in fed flies whereas PAM-α1 DANs are required in starved flies. Correspondingly, a feeding-dependent calcium increase was observed in PAM-β'2mp DANs, but not in PAM-α1 DANs. Following training, the presence of sweet sugars recruits PAM-β'2mp DANs, whereas tasteless medium increases calcium in PAM-α1 DANs. Together, this work identifies mechanistic underpinnings of sleep-dependent memory consolidation, in particular demonstrating a role for the processing of sweet taste reward signals (Chouhan, 2022).
Flies can form sleep-dependent and sleep-independent memory in an appetitive conditioning paradigm, but how they switch between these distinct forms of memory consolidation remains unexplored. This study found that Gr64f GRNs-mediated sweet taste signal is essential for sleep-dependent memory. Sweet taste activates a specific subset of PAM DANs, PAM-β'2mp, for the consolidation of sleep-dependent memory. In contrast, in the absence of sweet stimulus, sleep becomes dispensable for memory consolidation and PAM-α1 DANs are recruited to form long-term memory. These findings indicate that the presence of reward can modulate the role of sleep in memory consolidation (Chouhan, 2022).
Taste serves as an important modality in feeding decisions in Drosophila as perceived sweetness is used to estimate the carbohydrate content of food. Flies prefer sweet sugars over substances with a higher nutritive value indicating that taste determines initial feeding preferences. Sweetness drives sugar ingestion during odor-reward pairing that is essential to form appetitive memories. Also, sweet taste tempers food-seeking behavior as flies on a sweet but non-nutritive medium show lower activity than starved flies. Therefore, the presence of sweet taste modifies behavior by providing a sensation of food (Chouhan, 2022).
Flies kept on food vials require sleep to form appetitive memories (Chouhan, 2021). As in the case of the other behaviors mentioned above, sweet taste appears to be sufficient to signal the presence of food and thereby trigger sleep-dependent memory formation. Flies kept on sucralose, a sweet but non-nutritive sugar, show an enhancement in post-training sleep and require sleep to consolidate long-term memory. In contrast, flies on sorbitol, a tasteless but nutritive substance, form sleep-independent memory. Also, disrupting the activity of sweet-sensing Gr64f+ neurons results in the formation of sleep-independent memory in fed flies, while stimulating Gr64f+ neurons in starved flies induces a switch to sleep-dependent memory consolidation. While sweet taste may induce sleep-dependent memory, it is also possible that processes driving sleep-dependent and sleep-independent memory consolidation interact so as to be mutually exclusive. For instance, as starvation suppresses sleep, it may trigger sleep-independent memory, while sweet taste could drive sleep-dependent memory consolidation by releasing starvation-induced suppression of sleep. Neural connections between the two relevant circuits (described below) could mediate such interactions (Chouhan, 2022).
Sleep is typically thought to be induced under conditions of higher energy consumption; however, the data indicate that, at least for sleep-dependent memory consolidation, sweet taste rather than caloric intake serves as a signal for sleep. Sweet taste is not an accurate reporter for the nutritional value of food. Indeed, flies switch their preference from non-nutritive sweeteners to tasteless nutritive sugars over time in a two-choice assay. Therefore, sweet taste can only provide a short-term impression of the presence of food, which might be sufficient to induce sleep-dependent consolidation as flies form sleep-dependent or sleep-independent memory in the first 4 h after training (Chouhan, 2022).
The reward signal from PAM DANs during training is essential to form odor-reward associations. MB neurons are tiled by individual DAN terminals and the dendrites of corresponding MBONs to form 15 anatomically and functionally distinct compartments. PAM DANs largely innervate distinct compartments of the horizontal MB lobes. As per a previous study, a recurrent circuit that couples PAM-α1 DANs with the corresponding MBON in the α1 MB compartment mediates appetitive memory consolidation in starved flies. However, the current study found that PAM-α1 DANs are dispensable for memory consolidation in fed flies. Instead, PAM DANs that innervate the β'2mp compartment of the MB are essential for long-term memory in fed flies. Consistent with the findings that distinct PAM DANs mediate memory consolidation in fed versus starved states, the activity of PAM-β'2mp DANs was higher in trained and fed flies, while calcium in PAM-α1 DANs was significantly higher in trained and starved flies. These results complement previous work in rats, humans, and flies demonstrating that discrete neural circuits mediate sleep-dependent and sleep-independent memory consolidation (Chouhan, 2022).
PAM-β'2mp DANs signal reward to form 'safety-memory' of the unpaired odor during spaced aversive conditioning. Intriguingly, these neurons connect to MBONs implicated in appetitive memory consolidation under fed and starved conditions. PAM-β'2mp DANs receive input from MBON-γ2α'1 neurons, which are required for memory under fed conditions, and through axo-axonic projections they connect to the MBON-γ1Pedc neurons that are essential for memory under starved conditions. It is proposed that MBON-γ2α'1 neurons engage PAM-β'2mp DAN-mediated reward signaling to establish a sleep-dependent memory trace. By contrast, an axo-axonal connection from MBON-γ1Pedc neurons may modulate neurotransmission in PAM-β'2mp DANs such that it allows PAM-α1 DANs to form sleep-independent memory. Future work can investigate how these discrete neural circuits interact to form sleep-dependent or sleep-independent memories (Chouhan, 2022).
Sweet taste is perceived by a set of seven transmembrane receptors on the proboscis and tarsal segments of the leg. The taste information is transmitted by GRNs that terminate in the suboesophageal ganglion. Second-order neurons of the gustatory system send projections from the suboesophageal ganglion to the central brain for higher-order processing of taste information. A subset of these projections terminates in the superior protocerebrum that also contains processes from DANs. Consistent with a connection between sweet-sensing neurons and DANs, an exposure to sucrose evokes a robust calcium response in PAM DANs. Therefore, sweet taste may signal reward by stimulating PAM DANs. Consistently, a significant calcium/GFP increase in PAM-β'2mp DANs was found in fed flies compared with starved flies. Also, flies kept on arabinose or sucralose after training demonstrate a significant increase in PAM-β'2mp DAN activity compared with flies moved to sorbitol following training. It is suggested that the Gr64f+ GRNs mediated sweet taste signal acts in conjunction with the feedback from MBON-γ2α'1 neurons to recruit PAM-β'2mp DANs to form sleep-dependent memory (Chouhan, 2022).
Rewarding experiences modulate behavior to enhance survival. These experiences are preferentially replayed during sleep for better retention. Indeed, memory consolidation involves neuronal reactivation in the mesolimbic dopaminergic reward system during sleep. Conversely, sleep loss modifies reactivity in the reward centers of the brain such that it induces reward-seeking behavior. This work suggests that the processing of sweet taste reward signals determines the necessity for sleep in memory consolidation. A general role for reward in determining the need for sleep is open to investigation (Chouhan, 2022).
In Drosophila, in vivo functional imaging studies revealed that associative memory formation is coupled to a cascade of neural plasticity events in distinct compartments of the mushroom body (MB). In-depth investigation of the circuit dynamics, however, will require an ex vivo model that faithfully mirrors these events to allow direct manipulations of circuit elements that are inaccessible in the intact fly. The current ex vivo models have been able to reproduce the fundamental plasticity of aversive short-term memory, a potentiation of the MB intrinsic neuron (Kenyon cells [KCs]) responses after artificial learning ex vivo However, this potentiation showed different localization and encoding properties from those reported in vivo and failed to generate the previously reported suppression plasticity in the MB output neurons (MBONs). This study developed an ex vivo model using the female Drosophila brain that recapitulates behaviorally evoked plasticity in the KCs and MBONs. This plasticity accurately localizes to the MB α'3 compartment and is encoded by a coincidence between KC activation and dopaminergic input. The formed plasticity is input-specific, requiring pairing of the conditioned stimulus and unconditioned stimulus pathways; hence, it was named pairing-dependent plasticity. Pairing-dependent plasticity formation requires an intact CaMKII gene and is blocked by previous-night sleep deprivation but is rescued by rebound sleep. In conclusion, this study showed that the ex vivo preparation recapitulates behavioral and imaging results from intact animals and can provide new insights into mechanisms of memory formation at the level of molecules, circuits, and brain state (Adel, 2022).
A neutral experience (conditioned stimulus [CS]) can be remembered as positive or negative if closely followed by rewarding or punishing reinforcement (unconditioned stimulus [US]). The ability to form this type of 'associative' memory is phylogenetically conserved; Drosophila form robust associative memories, most of which are encoded and stored in the mushroom body (MB). The MB is a higher brain structure made of 15 distinct compartments. Each compartment is built on a scaffold of axons of one of the three main types of Kenyon cells (KCs; αβ, α'β', and γ). The KCs connect to MB output neurons (MBONs), which project out of the MB to bias behavior. The KC->MBON synapses are modulated by dopaminergic neurons (Adel, 2022).
During aversive olfactory associative learning, an odor (the CS) activates a sparse group of KCs, such that this odor identity is represented across all MB compartments. Simultaneously, dopaminergic neurons from the protocerebral posterior lateral (PPL1) cluster are activated by the US, encoding negative prediction errors in MB compartments. When KC activation and the dopaminergic signal coincide within a compartment, the KC->MBON synapses in that compartment are depressed, biasing the circuit output to aversion (Adel, 2022).
Many studies have investigated the properties of this circuitry using in vivo calcium imaging in intact animals. In contrast, explanted brains have been used mostly for establishing connectivity between neurons or interrogating a specific biochemical pathway; only a few studies have attempted to understand memory circuit logic ex vivo. In the best-developed paradigm, a previous study observed a potentiation of KC responses in the tips of the MB vertical lobes which they termed 'long-term enhancement' (LTE). This laid the groundwork for developing ex vivo models of this circuit, but there were major differences between LTE and associative memory observed in intact animals. The most significant were that the plasticity was not specific to the α'β' KCs and that dopamine release by the US was not observed; it was only seen after CS+US coincidence (Ueno et al., 2013, 2017) (Adel, 2022).
This study establish an ex vivo paradigm that resolves these discrepancies and exhibits the cardinal features of associative learning. Pairing odor and punishment pathway activation in dissected brains results in a localized potentiation of the α'β' KCs and suppression of their postsynaptic MBONs in the α'3 compartment. Because both KC potentiation and MBON suppression are strictly dependent on temporal coincidence of the CS and US, this paradigm was termed 'pairing-dependent plasticity' (PDP). Like the CS specificity of associative memories, PDP is specific to the subset of odor-representing projection neurons activated during the artificial training. Evidence is also provided that dopamine is released by activation of the US pathway and does not require CS+US coincidence (Adel, 2022).
This ex vivo paradigm can be used for obtaining new mechanistic insight into memory formation at the molecular and circuit levels. Data is presented indicating that the 3'UTR of the CaMKII gene is critical for short-term memory (STM) formation and that the primacy of α' compartment plasticity in learning is because of differences in input/response relationships between α and α'. Finally, it was demonstrated that the ability of the ex vivo brain to be plastic can be influenced by prior in vivo experience, as this study reports that brains of sleep-deprived flies fail to form PDP, but as little as 2 h of recovery sleep rescues this learning impairment (Adel, 2022).
Drosophila neural circuits are traditionally studied by relating in vivo genetic and chemical manipulations with their consequent behavioral outcomes, from which circuit information can then be inferred. More recently, the advent of in vivo calcium imaging allowed for tracing neural activity in actively behaving flies. Over more than a decade of such in vivo studies, the general circuit mechanisms of associative memory have been discovered, but there are limitations imposed by imaging the brain of an active intact fly. These include the relatively low signal-to-noise ratio, the inaccessibility of multiple brain regions because of restrictions on imaging angles, the difficulty of doing acute pharmacological studies, and the possible confounds of studying the brain of a movement-restricted fly experiencing ongoing stress. Taking inspiration from the way the LTP hippocampal slice model revolutionized understanding of mammalian memory, this study provides an ex vivo model of Drosophila memory which can overcome these limitations and offer a powerful preparation for studying Drosophila memory circuits. Importantly, this model provides a framework for investigating the dynamics of neural circuits in the fly brain (Adel, 2022).
Most of the previous studies investigating the associative learning circuit ex vivo have focused on mapping connectivity or characterizing a specific biochemical pathway. Only a few ex vivo studies have focused on understanding MB circuit logic. In the LTE model, pairing a stimulation of the CS and US pathways induced a potentiation of KC responses in the tips of the MB vertical lobes, but LTE did not fully recapitulate other characteristics of associative memory observed in intact flies. This study developed a modified ex vivo model that resolves these discrepancies, showing that the paired activation of odor and punishment pathways induces appropriate plasticity at multiple nodes in the circuit: potentiation of KCs and suppression of MBONs. Several mechanisms for encoding those opposite forms of the plasticity have been proposed, including spike timing-dependent plasticity and activation of distinct dopaminergic receptors. Spike timing-dependent plasticity mechanisms appear less likely as MBON suppression was shown to not require MBON spiking. Perhaps the strongest model so far comes a previous demonstration that differences in the order of KCs activation and dopaminergic input activate distinct dopaminergic receptors, DopR1 or DopR2, which encode MBON suppression or potentiation, respectively. It is important to note that previous work studied the plasticity in MB medial lobes, while the current study focused on MB vertical lobes, so this paradigm may be useful in gaining greater mechanistic insight into this sign transformation in the vertical lobes (Adel, 2022).
PDP is localized to the MB α'3 compartment and not in α3, in alignment with most imaging studies in intact flies. Importantly, in this ex vivo preparation, punishment information is relayed to the MB through dopaminergic release from the PPL1 subset. Bath application of dopamine in the current preparation does not interfere with the specificity of associative learning since PDP is exclusively formed in the cells that were active during the dopamine application. These data settle several inconsistencies between previous ex vivo studies and the majority of in vivo reports. It is suggest that the genesis of the discrepancies was not because of any inherent difference between intact and ex vivo brains but was rather a consequence of technical considerations, including stimulation strength, dopamine concentration, and the sensor tools used (Adel, 2022).
An ex vivo preparation that recapitulates the cardinal features of the circuits underlying associative memory formation should be useful for mechanistic studies at the molecular, cellular, and systems levels. This model was used to ask a new question about the innerworkings of the circuit at each of these levels. At the molecular level, the importance of normal levels of CaMKII was demonstrated by manipulating the 3'UTR of CaMKII mRNA. Deletion of this region of the CaMKII gene drastically reduces the amount of CaMKII protein in synaptic regions and blunts the ability to form STM and to generate a potentiation PDP in KC axons. The data argue that the role of this molecule is downstream of the CS+US coincidence detector, as a much weaker PDP was observed in CaMKIIUdel flies. Whether the behavioral defect is due solely to the KC PDP defect is not completely clear since CaMKII likely has active roles at other circuit nodes (Adel, 2022).
At the cellular level, it was asked why STM and PDP form in the α'3 but not the nearby α3 compartment when both compartments respond to odors and AL stimulation, and both receive dopaminergic input from the same PPL1 cluster. Previous work found that real odors cause activity in only 5%-12% of KCs and elicit a much higher spike rate in the α'β' KCs than in the αβ KCs. This study found that low-intensity AL stimulation (100 μAmps) elicits a stronger response in the α'3 than in the α3 compartment, while high-intensity AL-stimulation (200 μAmps) causes strong responses in the α3 compartment and recruits it to the learning circuit. Coupling this with the observation of lower dopamine release in α3 suggests a model in which odor presentation during associative learning causes subthreshold responses in αβ cells such that the CS+US coincidence detector is not triggered, while the stronger responses in the α'β' cells bypass this threshold, allowing plasticity in the α'β' cells only. This notion is in agreement with the previous finding that α'β' cells have a lower firing threshold than αβ cells. Further, It is possible that long-term memory and the enhancement memory trace in the αβ KCs after repetitive space training require a gradual potentiation of the αβ KC responses with every training session such that the responses bypass the coincidence detection threshold after several training sessions. Whether repetition of AL+DA pairings recruits PDP in the α3 compartment remains unclear. It is also yet to be determined whether shortcutting the circuit and recruiting αβ cells in the first training session reduces the need for multiple spaced training sessions in long-term memory formation (Adel, 2022).
In conclusion, this study looked at the ability of the effects of prior experience, or brain state, on the memory circuit to be retained in the ex vivo preparation. Excitingly, it was found that sleep-deprived flies could not form PDP, but that as little as 2 h of rest before dissection allowed the brain to recover PDP formation. The complete abolition of PDP in sleep-deprived flies at first and the gradual recovery in plasticity afterward suggest that sleep converges on the memory circuit upstream of the CS+US coincidence detector. Whether this involves regulation of dopamine receptors in the MB during sleep remains to be determined. The ability to retain in some functional way the internal state of the brain will allow this preparation to be used to understand how memory formation is altered by global system alterations (Adel, 2022).
Although many genes are known to influence sleep, when and how they impact sleep-regulatory circuits remain ill-defined. This study shows that insomniac (inc), a conserved adaptor for the autism-associated Cul3 ubiquitin ligase, acts in a restricted period of neuronal development to impact sleep in adult Drosophila. The loss of inc causes structural and functional alterations within the mushroom body (MB), a center for sensory integration, associative learning, and sleep regulation. In inc mutants, MB neurons are produced in excess, develop anatomical defects that impede circuit assembly, and are unable to promote sleep when activated in adulthood. These findings link neurogenesis and postmitotic development of sleep-regulatory neurons to their adult function and suggest that developmental perturbations of circuits that couple sensory inputs and sleep may underlie sleep dysfunction in neurodevelopmental disorders (Li, 2021).
During sleep, the brain undergoes dynamic and structural changes. In Drosophila, such changes have been observed in the central complex, a brain area important for sleep control and navigation. The connectivity of the central complex raises the question about how navigation, and specifically the head direction system, can operate in the face of sleep related plasticity. To address this question, this study develop a model that integrates sleep homeostasis and head direction. By introducing plasticity, the head direction system was shown to function in a stable way by balancing plasticity in connected circuits that encode sleep pressure. With increasing sleep pressure, the head direction system nevertheless becomes unstable and a sleep phase with a different plasticity mechanism is introduced to reset network connectivity. The proposed integration of sleep homeostasis and head direction circuits captures features of their neural dynamics observed in flies and mice (Flores-Valle, 2021).
In metazoan organisms, circadian (∼24 h) rhythms are regulated by pacemaker neurons organized in a master-slave hierarchy. Although it is widely accepted that master pacemakers and slave oscillators generate rhythms via an identical negative feedback loop of transcription factor CLOCK (CLK) and repressor PERIOD (PER), their different roles imply heterogeneity in their molecular clockworks. Indeed, in Drosophila, defective binding between CLK and PER disrupts molecular rhythms in the master pacemakers, small ventral lateral neurons (sLN(v)s), but not in the slave oscillator, posterior dorsal neuron 1s (DN1ps). This study developed a systematic and expandable approach that unbiasedly searches the source of the heterogeneity in molecular clockworks from time-series data. In combination with in vivo experiments, it was found that sLNvs exhibit higher synthesis and turnover of PER and lower CLK levels than DN1ps. Importantly, light shift analysis reveals that due to such a distinct molecular clockwork, sLNvs can obtain paradoxical characteristics as the master pacemaker, generating strong rhythms that are also flexibly adjustable to environmental changes. These results identify the different characteristics of molecular clockworks of pacemaker neurons that underlie hierarchical multi-oscillator structure to ensure the rhythmic fitness of the organism (Jeong, 2022).
The circadian clock enables organisms to manifest about 24-h (circadian) rhythms of behavior and physiology coordinated with rhythmic environmental changes. The generic model of the circadian clock is composed of input, oscillator, and output, wherein the oscillator entrains to time cues (zeitgeber) and regulates the output rhythms. This system operates as a network in which the master pacemaker and slave oscillator are organized in a hierarchical manner. The master pacemaker receives the light signal, the prominent zeitgeber, and then drives the slave oscillator that regulates distinct outputs such as sleep, feeding, metabolic homeostasis, etc. In this hierarchy system, the master pacemaker can generate strong rhythms to yield clear signals to the slave oscillator while still being able to flexibly adjust their phase in response to changes in environmental lighting conditions. However, the molecular mechanisms underlying these somewhat paradoxical characteristics of the master pacemaker are poorly understood (Jeong, 2022).
In Drosophila, small ventral lateral neurons (sLNvs) act as the master pacemaker. That is, sLNvs maintain free-running rhythms under constant darkness. On the other hand, posterior dorsal neuron 1s (DN1ps) act as the slave oscillator receiving neuropeptide pigment-dispersing factor (PDF) from sLNvs, which is critical to maintain their rhythms. Without PDF signaling from sLNvs, DN1ps rapidly lose molecular oscillation, and DN1ps follow the speed of genetically modified sLNvs. Furthermore, DN1ps harbor connections with output centers such as premotor, sleep, and neuroendocrine centers. Taken together, the circadian clock of Drosophila has a hierarchical organization, with sLNvs being the master pacemaker receiving light signals and DN1ps being the slave oscillator releasing output signals, although the organization can be potentially changed in the presence of environmental or genetic perturbations (Jeong, 2022).
Despite the different roles of these pacemaker neurons, they share common molecular mechanisms to generate circadian rhythms that are well conserved in all life-forms: the interlocked multiple transcriptional-translational feedback loops (TTFLs) composed of core clock proteins. In the Drosophila core TTFL, CLK, and CYCLE (CYC) activate the transcription of per and timeless (tim); PER and TIM proteins, in turn, repress their own transcription in which PER is the core repressor. This core TTFL regulates the 24-h period rhythmic expression of clock genes and other clock-controlled genes. Although sLNvs and DN1ps generate circadian rhythms via identical TTFLs, unexpectedly, it was previously found that their rhythms are altered in a different way in p{Clk-Δ};Clkout flies (here referred to as Clk-Δ flies), which express CLK defective in PER binding. Specifically, in Clk-Δ flies, PER oscillation was significantly dampened in sLNvs but quasinormal in DN1ps, demonstrating heterogeneity in their molecular clocks (Jeong, 2022).
This study took advantage of Clk-Δ flies to understand pacemaker neuron–specific molecular clockworks. Specifically, the neuron-specific alteration of a time series of PER in Clk-Δ flies was analyzed by developing a systematic modeling approach. This allowed systematic investigation of all possible molecular differences in the core TTFL between sLNvs and DN1ps with their mathematical models developed in this study. With a combination of in vivo experiments, essential differences were found in the molecular clockworks of sLNvs and DN1ps to reproduce their different rhythm alterations by Clk-Δ: CLK levels are higher in DN1ps than in sLNvs, the synthesis of PER is more efficient, and the degradation of PER is faster in sLNvs than in DN1ps. Furthermore, it was found that such distinct molecular mechanisms of the core TTFL in sLNvs are critical for its ability to act as the master pacemaker, generating strong rhythms while flexibly adapting its phase to environmental changes (e.g., jet lag) via in silico experiments. In conclusion, this study presents pacemaker neuron–specific molecular clockworks that underlie the hierarchical organization of the circadian clock to ensure the rhythmic fitness of the organism (Jeong, 2022).
In the Drosophila circadian clock, some molecular differences in pacemaker neurons have been reported in addition to their different repertoire of transcripts. However, it has been poorly understood whether and why key molecular mechanisms for generating circadian rhythms differ among pacemaker neurons. This study found differences between the core TTFL of the circadian clock in sLNvs and DN1ps by using a combination of theoretical and experimental approaches. Furthermore, it was found that such distinct characteristics of the core TTFL in sLNvs enables them to generate strong rhythms while flexibly adapting their phase upon changes to the environmental lighting conditions (Jeong, 2022).
To understand why PER rhythms are altered differently by the same Clk-Δ mutation between sLNvs and DN1ps, a systematic modeling approach was developed. That is, all possible differences in the core TTFL of sLNvs and DN1ps were investigated to identify key differences that explain their different alterations by the Clk-Δ mutation. This allowed identification of the parameter sets of mathematical models that can reproduce different time series of PER between sLNvs and DN1ps. Then, by analyzing the common patterns of the parameter sets, it was possible to identify key differences in molecular clockworks between sLNvs and DN1ps: higher synthesis and turnover of PER and lower CLK levels in sLNvs than DN1ps. While it was assumed that the dissociation constants are the same to avoid the identifiability issue of the parameter estimation, the dissociation constants could also differ in molecular clockworks between sLNvs and DN1ps (Jeong, 2022).
The patterns of parameter sets rather than a single best-fit parameter set to avoid overfitting, because the best-fit parameter may not yield the most meaningful parameters when models contain a large number of parameters. Such a systematic approach has also been successfully used to resolve the unexpected dynamics in biological systems. While these previous studies focused on identification of hidden regulation underlying a single system, this study investigated the difference between two systems, sLNvs and DN1ps. Thus, the regularization cost, penalizing the difference in the values of parameters between the LN and DN models, was used to avoid unnecessary differences. This systematic modeling framework is expandable to identify heterogeneity of other systems (Jeong, 2022).
The predicted lower CLK levels in sLNvs than in DN1ps were confirmed by in vivo experiments. Given the lower amount of the transcription factor CLK, one can imagine that de novo–synthesized nascent per transcript would be lower, producing a lower amount of PER in sLNvs than in DN1ps. But PER is more rapidly synthesized in sLNvs than in DN1ps, indicating that the production of PER in sLNvs might be enhanced by posttranscriptional mechanisms. Intriguingly, the translation activation complex of ATX2 and TYF posttranscriptionally regulates per mRNA in an LN-specific manner. miRNAs are also important posttranscriptional regulators of gene expression that degrade target mRNA and/or inhibit its translation. Numerous miRNAs regulate the circadian rhythm by affecting Clk, clockwork orange, tim, or output genes. While no microRNA (miRNA) targeted toward per mRNA has been identified so far, the different repertoire of miRNA in pacemaker neurons might be responsible for the different kinetics of PER accumulation (Jeong, 2022).
In addition, the predicted higher turnover rate of PER in sLNvs than in DN1ps was also confirmed by in vivo experiments. What could cause these differences in the degradation rate of PER between sLNvs and DN1ps? Throughout the day, PER is progressively phosphorylated by several kinases including DBT (casein kinase I ortholog in flies), casein kinase 2 (CK2), NEMO, and Shaggy. Hyperphosphorylated PER is degraded by the ubiquitin-proteasome system via recognition of Ser47 phosphorylation by the ubiquitin ligase Supernumerary Limbs (SLIMB). Thus, different repertoires of kinase activity in each group of pacemaker neurons might result in different kinetics of PER hyperphosphorylation leading to Ser47 phosphorylation, which is the PER degradation mark. For instance, Ser47 phosphorylation and PER degradation is delayed by NEMO-dependent phosphorylation of the middle part of PER. Intriguingly, NEMO is expressed in sLNvs, large ventral lateral neurons, dorsal lateral neurons, and DN1s but not in DN2s or DN3s. Furthermore, CK2 is expressed only in LNvs. Collectively, it was reasoned that a unique repertoire of kinases cooperates to make PER more susceptible to degradation in sLNvs than in DN1ps. Of course, given that the phosphorylation status of a protein is regulated by phosphatases, the phosphatase repertoire and/or expression level could be another important determinant of degradation rate. Indeed, PP1 and PP2A affected PER phosphorylation and thus its stability (Jeong, 2022).
The mathematical model predicted that the CLK-Δ mutation induced different rhythm alterations in sLNvs and DN1ps due to the differences in molecular clockworks between sLNvs and DN1ps. Specifically, when PER levels are higher than CLK levels in the nucleus, the majority of CLK is sequestered and, thus, transcription of per mRNA is suppressed. As a result, only when PER levels are lower than CLK levels in the nucleus is the transcription of per mRNA promoted (i.e., the activation phase). However, as the binding between CLK and PER is disrupted due to the CLK-Δ mutation, even when PER levels are higher than CLK levels, free CLK is available, and thus, the transcription of per mRNA is weakly promoted. This weak transcription of per mRNA dramatically increases PER levels compared to CLK levels due to the high synthesis rate of PER and the low CLK levels in the LN-Δ model. As a result, the transcription of per mRNA cannot be fully promoted. This disruption of the transition from the suppression phase to the activation phase dampens circadian rhythms in the LN-Δ model. On the other hand, the weak transcription of per mRNA has little effect on the ratio between PER levels and CLK levels due to the high CLK levels and low synthesis rate of PER in the DN-Δ model. Thus, even in the presence of the CLK-Δ mutation, the transition between the activation and suppression phases occurs, leading to quasinormal circadian rhythms in the DN-Δ model. Taken together, the LN model shows a greater sensitivity of the transcription in response to the change in the level of PER than the DN model to generate stronger rhythms. However, due to such greater sensitivity, when the system is perturbed (i.e., mutation), the LN model shows a more sensitive response compared to the DN model, leading to the loss of rhythms. As the greater sensitivity is cooperatively generated by the high synthesis rate of PER and low level of CLK, the LN-Δ model predicts that changing a single parameter alone (i.e., synthesis rate of PER) will not rescue the rhythms disrupted by the CLK-Δ mutation in sLNvs. It would be interesting in future work to investigate whether the disrupted rhythms can be rescued by simultaneously changing multiple parameters of the molecular clockworks in sLNvs (Jeong, 2022).
Why, then, do sLNvs have such different molecular properties from DN1ps? Due to the fast synthesis and turnover rates of PER, sLNvs can generate rhythms with high amplitudes, which is critical to yield clear signals to slave oscillators. Unexpectedly, although typically strong oscillators with high amplitudes have difficulty in adapting to environmental changes (e.g., jet lag), this study found that sLNvs can be reentrained to the new LD cycle as rapidly as DN1ps. Interestingly, before the reentrainment, unlike DN1ps, the phases of sLNvs are greatly dispersed. This in silico study proposes that the phase dispersion stems from the distinct property of TTFL in sLNvs from DN1ps due to its spike-like and sensitive transcription yielding strong oscillation. Another study also showed that the sensitive response of per mRNA in response to the environmental change, yielding phase plasticity, is the feature of robust oscillators. That is, the sensitive change of per mRNA, and thus sensitive phase shifts under environmental change, counterbalance the change of the other reaction rates and lead to period robustness. Taken together, due to the distinct molecular clockworks of sLNvs compared to those of DN1ps, sLNvs could obtain both robustness (i.e., high amplitude and period robustness) and plasticity (i.e., fast entrainment and a wide range of entrainment), which are critical characteristics for a master pacemaker (Jeong, 2022).
Blue light activation of the photoreceptor
Cryptochrome (Cry) evokes rapid depolarization and increased action
potential firing in a subset of circadian
and arousal neurons in Drosophila melanogaster. This study shows
that acute arousal behavioral responses to blue light significantly
differ in mutants lacking Cry, as well as mutants with disrupted
opsin-based phototransduction. Light-activated Cry couples to membrane
depolarization via a well conserved redox sensor of the voltage-gated
potassium (K+) channel β-subunit (Kvβ) Hyperkinetic
(Hk). The neuronal light response is almost completely absent in hk-/-
mutants, but is functionally rescued by genetically targeted
neuronal expression of WT Hk, but not by Hk point mutations that disable
Hk redox sensor function. Multiple K+ channel α-subunits that coassemble
with Hk, including Shaker, Ether-a-go-go, and Ether-a-go-go-related
gene, are ion conducting channels for Cry/Hk-coupled light response.
Light activation of Cry is transduced to membrane depolarization,
increased firing rate, and acute behavioral responses by the Kvβ subunit
redox sensor (Fogle, 2015). Acute behavioral arousal to blue light is significantly attenuated in
CRY mutants. This study identified a redox signaling couple between blue
light-activated CRY and rapid membrane depolarization via the redox
sensor of Kvβ channel subunits coassembled with Kvα channel subunits.
Additional unknown factors may act as intermediates between CRY and Hk.
This finding provides in vivo validation of a very longstanding
hypothesis that the highly conserved redox sensor of Kvβ subunit
functionally senses cellular redox events to physiological changes in
membrane electrical potential. Genetic loss of any single component
functionally disrupts the CRY-mediated blue light response, which is
functionally rescued by LNv restricted expression of their WT genes in
the null backgrounds. Although little is known about the structural
contacts between Kvβ and EAG subunits, Kvβ subunits make extensive
physical contacts in a fourfold symmetric fashion in 1:1 stoichiometry
with the T1 assembly domain of other coassembled tetrameric Kvα subunits
that form the complete functional channel (Fogle, 2015). Application of Kvβ redox chemical substrate modulates voltage-evoked
channel peak current, steady-state current, and inactivation in
heterologously expressed α-β channels, which are reversed by fresh
NADPH. These results indicate that measurements of channel biophysical
properties can reflect the redox enzymatic cycle of Kvβ as these channel
modulatory effects are absent in preparations that lack the expression
of WT Kvβ subunits or express redox sensor mutant Kvβ subunits. Whether
direct chemical redox reactions occur between CRY and Hk is unclear. For
CRY, light or chemical reduction induces one-electron reduction of the
FAD cofactor of CRY, whereas the reductive catalytic mechanism of AKRs
(such as Hk) requires a hydride ion transferred from NADPH to a
substrate carbonyl, then a solvent-donated proton reduces the substrate
carbonyl to an alcohol. These differences in redox chemistry between CRY
and Hk suggest that other intermediates, such as oxygen, are possibly
required for redox coupling (Fogle, 2015). Spectroscopic analysis of animal and plant CRYs suggest that light
activation causes reduction of the FAD oxidized base state. Light
activation of Drosophila CRY also evokes conformational changes in the C
terminus of CRY that clearly promotes CRY C-terminal access to
proteolytic degradation and subsequent interactions with the Timeless
clock protein, thus signaling degradation and circadian entrainment.
However, all existing evidence suggests that light activated
CRY-mediated circadian entrainment and membrane electrical
phototransduction operate under different mechanisms, including their
different activation thresholds and relative dependence on the C
terminus of CRY. Further distinguishing the distinct mechanisms of the
downstream effects of light-activated CRY, the light-induced
conformational changes that couple CRY to ubiquitin ligase binding (thus
causing circadian entrainment) occur in oxidized and reduced states of
CRY and are unaffected in CRY tryptophan mutants that presumably are
responsible for intraprotein electron transfer reactions following
light-evoked reduction of the FAD cofactor. Another recent study shows
that light- or chemical-evoked reduction of Drosophila CRY FAD is
coupled to conformational changes of the CRY C terminus, along with
reporting a surprising negative result that DPI has no effect on the
reoxidation of the reduced anionic semiquinone of purified Drosophila
CRY. DPI could hypothetically influence the electrophysiological light
response by blocking the pentose phosphate pathway, which produces the
Hk redox cofactor NADPH, but this does not explain the light dependence
for DPI blocking the electrophysiological light response herein. The
available evidence indicates that CRY-mediated light evoked membrane
depolarization occurs independently of conformational changes in the CRY
C-terminal domain but depends on redox changes in CRY, whereas
CRY-mediated light evoked circadian entrainment depends on
conformational changes in the CRY C-terminal domain and may or may not
depend on CRY redox state (Fogle, 2015). Light-activated CRY evokes rapid membrane depolarization through the
redox sensor of the Kvβ subunit Hk. A general role for circadian
regulation of redox state coupled to membrane excitability has been
described recently in mammalian suprachiasmatic neurons. Redox
modulation of circadian neural excitability may be a well-conserved
feature (Fogle, 2015).
Drosophila's circadian clock can be perturbed by magnetic fields, as well as by lithium administration. Cryptochromes are critical for the circadian clock. Further, the radical pairs in cryptochrome also can explain magnetoreception in animals. Based on a simple radical pair mechanism model of the animal magnetic compass, this study shows that both magnetic fields and lithium can influence the spin dynamics of the naturally occurring radical pairs and hence modulate the circadian clock's rhythms. Using a simple chemical oscillator model for the circadian clock, this study showed that the spin dynamics influence a rate in the chemical oscillator model, which translates into a change in the circadian period. This model can reproduce the results of two independent experiments, magnetic field and lithium effects on the circadian clock. The model predicts that stronger magnetic fields would shorten the clock's period. It is also predicted that lithium influences the clock in an isotope-dependent manner. Furthermore, the model also predicts that magnetic fields and hyperfine interactions modulate oxidative stress. The findings of this work suggest that the quantum nature of radical pairs might play roles in the brain, as another piece of evidence in addition to recent results on xenon anesthesia and lithium effects on hyperactivity (Zadeh-Haghighi, 2022).
Although electric fields (EF) exert beneficial effects on animal wound healing, differentiation, cancers and rheumatoid arthritis, the molecular mechanisms of these effects have remained unclear about a half century. This study aimed to elucidate the molecular mechanisms underlying EF effects in Drosophila melanogaster as a genetic animal model. This study shows that the sleep quality of wild type (WT) flies was improved by exposure to a 50-Hz (35 kV/m) constant electric field during the day time, but not during the night time. The effect was undetectable in cryptochrome mutant (cryb) flies. Exposure to a 50-Hz electric field under low nutrient conditions elongated the lifespan of male and female WT flies by ~18%, but not of several cry mutants and cry RNAi strains. Metabolome analysis indicated that the adenosine triphosphate (ATP) content was higher in intact WT than cry gene mutant strains exposed to an electric field. A putative magnetoreceptor protein and UV-A/blue light photoreceptor, CRYPTOCHROME (CRY) is involved in electric field (EF) receptors in animals. The present findings constitute hitherto unknown genetic evidence of a CRY-based system that is electric field sensitive in animals (Kawasaki, 2021).
Cryptochrome (CRY) is a conserved protein associated with the circadian clock in a broad range of organisms, including plants, insects, and mammals. In Drosophila, cry is a pleiotropic gene that encodes a blue light-dedicated circadian photoreceptor, as well as an electromagnetic field sensor and a geotaxis behavior regulator. This study has generated a panel of nearly-isogenic strains that originated from various wild populations and which carry different natural alleles of cry. Sequencing of these alleles revealed substantial polymorphism, the functional role of which was elusive. To link this natural molecular diversity to gene function, association mapping was used. Such analysis revealed two major haplogroups consisting of six linked nucleotides associated with circadian phase (haplotypes All1/All2). A maximum-likelihood gene-tree that uncovered an additional pair of haplogroups (B1/B2). Behavioral analysis of the different haplotypes indicated significant effect on circadian phase and period, as well on the amount of activity and sleep. The data also suggested substantial epistasis between the All and B haplogroups. Intriguingly, circadian photosensitivity, assessed by light-pulse experiments, did not differ between the genotypes. Using CRISPR-mediated transgenic flies, the effect of B1/B2 polymorphism on circadian phase was verified. The transgenic flies also exhibited substantially different levels of cry transcription.The geographical distribution of the B1/B2 haplotypes was analyzed, focusing on a 12 bp insertion/deletion polymorphism that differentiates the two haplotypes. Analysis of cry sequences in wild populations across Europe revealed a geographical cline of B1/B2 indel frequency, which correlated with seasonal bioclimatic variables. This spatial distribution of cry polymorphism reinforces the functional importance of these haplotypes in the circadian system and local adaptation (Pegoraro, 2022).
Sleep circuitry evolved to have both dedicated and context-dependent modulatory elements. Identifying modulatory subcircuits and understanding their molecular machinery is a major challenge for the sleep field. Previously, 25 sleep-regulating microRNAs were identified in Drosophila melanogaster, including the developmentally important microRNA bantam. This study shows that bantam acts in the adult to promote early nighttime sleep through a population of glutamatergic neurons that is intimately involved in applying contextual information to behaviors, the γ5β'2a/β'2mp/β'2mp_bilateral Mushroom Body Output Neurons (MBONs). Calcium imaging revealed that bantam inhibits the activity of these cells during the early night, but not the day. Blocking synaptic transmission in these MBONs rescued the effect of bantam knockdown. This suggests bantam promotes early night sleep via inhibition of the γ5β'2a/β'2mp/β'2mp_bilateral MBONs. RNAseq identifies Kelch and CCHamide-2 receptor as possible mediators, establishing a new role for bantam as an active regulator of sleep and neural activity in the adult fly (Hobin, 2022).
Cryptochrome (CRY) is a short-wavelength light-sensitive photoreceptor expressed in a subset of circadian neurons and eyes in Drosophila that regulates light-evoked circadian clock resetting. Acutely, light evokes rapid electrical excitation of the ventral lateral subset of circadian neurons and confers circadian-modulated avoidance behavioral responses to short-wavelength light. Recent work shows dramatically different avoidance versus attraction behavioral responses to short-wavelength light in day-active versus night-active mosquitoes. To determine whether CRY1s mediate species-specific coding for behavioral and electrophysiological light responses,an "empty neuron" approach was used and transgenically expressed diurnal Aedes aegypti (AeCRY1) versus nocturnal Anopheles gambiae (AgCRY1) in a cry-null Drosophila background. AeCRY1 is much less light sensitive than either AgCRY1 or DmCRY as shown by partial behavioral rhythmicity following constant light exposure. Remarkably, expression of nocturnal AgCRY1 confers low survival to constant white light as does expression of AeCRY1 to a lesser extent. AgCRY1 mediates significantly stronger electrophysiological cell-autonomous responses to 365 nm ultraviolet (UV) light relative to AeCRY1. AgCRY1 expression mediates electrophysiological sensitivity to 635 nm red light, whereas AeCRY1 does not, consistent with species-specific mosquito red light responses. AgCRY1 and DmCRY mediate intensity-dependent avoidance behavior to UV light at different light intensity thresholds, whereas AeCRY1 does not, thus mimicking mosquito and fly behaviors. These findings highlight CRY as a key non-image-forming visual photoreceptor that mediates physiological and behavioral light responses in a species-specific fashion (Au, 2022).
The moon provides highly reliable time information to organisms. Whereas sunlight is known to set daily animal timing systems, mechanistic insight into the impact of moonlight on such systems remains scarce. This study establish that the marine bristleworm Platynereis dumerilii times the precise hours of mass spawning by integrating lunar light information into a plastic daily timing system able to run with circadian (~24 h) or circalunidian (~24.8 h) periodicity. The correct interpretation of moonlight is mediated by the interplay of two light sensors: a cryptochrome and a melanopsin ortholog provide information on light valence and moonrise time, respectively. Besides its ecological relevance, this work provides a plausible explanation for long-standing observations of light intensity-dependent differences in circadian clock periods (Zurl, 2022).
This study has uncover an ~24-h endogenous oscillator in marine broadcast-spawning worms that exhibits marked, moonlight-dependent plasticity in its period length upon extended moonlight illumination, as it occurs during FM periods. The clock is also adjusted by moonrises during the waning moon phase. It is currently unclear if the latter adjustment also relies on a change in period length or by phase resetting. Furthermore, during nights when swarming time is fine-tuned by the light of the moon, the plastic circadian/circalunidian clock (PCC) is not reset by the following solar light (sunrise) (Zurl, 2022).
The modulation of the worms' ~24-h oscillator by naturalistic moonlight provides a plausible model for how worms synchronize their nuptial dance, targeting a specific hour during the dark portion of moonlit nights. Restricting swarming behavior to the dark portion of the night might be advantageous to avoid predators that hunt during moonlight. On a mechanistic level, it is suggested that this PCC clock shares elements with the conventional core circadian oscillator and reveal two highly sensitive light receptors, r-Opsin1 and L-Cry, that are critical to sense and interpret naturalistic moonlight (Zurl, 2022).
Sensitivity to moonlight is directly relevant for a broad panel of marine broadcast spawners. The challenge of tagging nocturnal light information with the correct valence, however, likely extends beyond this specific ecological context. The classical categorization of organisms into nocturnal versus diurnal species typically neglects the aspect of moonlight. Any animal entraining its ~24-h clock to light will need to correctly interpret the occurrence of nocturnal light. Even though it has been shown that the circadian system of many species is sensitive to light levels as low as moonlight intensity, such as in flies and mice, chronobiological studies have so far spent relatively little effort in dissecting how animal clocks prevent potential disturbance by moonlight and interpret naturalistic light regimes that combine both sunlight and moonlight (Zurl, 2022).
The data presented in this study provide possible mechanistic explanations for the ability of the PCC clock to decode a combined sunlight and moonlight regime. A first tier is connected to the specific properties of cryptochrome: whereas under naturalistic moonlight, Platynereis L-Cry protein levels remain elevated, comparable to dark conditions, and are predominantly localized to the nucleus, the onset of sunlight causes a rapid degradation, with residual L-Cry protein found in the cytoplasm. On the biochemical level, L-Cry is highly sensitive to naturalistic moonlight. Moonlight evokes a different state in L-Cry than sunlight. Based on these detailed biochemical studies, including multiangle light scattering and size exclusion chromatography analyses, it is hypothesized that Pdu-L-Cry functions as a dimer with different light sensitivities of each of the two monomers, potentially arising from different quantum yields for FAD photoreduction. This would allow for the differential response to high-intensity sunlight versus very low intensity moonlight, as moonlight can only photoreduce the flavin cofactor in the low-light sensitive L-Cry monomer, whereas sunlight can photoreduce both. Dimers as part of the photoresponse mechanism have been well documented for Cryptochromes in plants. These Cryptochromes form dimers or even multimers upon illumination, which are essential for the light response. Even though plant Cry2 members are frequently called low-light receptors, their responses have only been analyzed under light conditions that still mimic sunlight, which are magnitudes away from moonlight. Thus, the mechanism by which Pdu-L-Cry responds to the very dim moonlight has to be more efficient even compared to the low-light sensors of the plant Cry2 family, while L-Cry also still retains the ability to detect high-intensity sunlight. The dimer model of L-Cry fulfills this requirement. In contrast to L-Cry, its direct Drosophila ortholog dCRY does not form dimers. This difference is consistent with the lack of photoreduction of purified dCRY under naturalistic moonlight that is reported in this study (Zurl, 2022).
Taken together, the data are consistent with the idea that-besides the canonical strong-light induced degradation-based signaling pathway for cytoplasmic Cryptochrome-L-Cry, but not dCRY, possesses a second, dim-light-induced, nuclear mode of signaling. Its presence in the nucleus and absence of degradation under moonlight makes it tempting to speculate that L-Cry could function as a transcriptional repressor. Notably, Drosophila Cryptochrome has been proposed to function as transcriptional repressor in the flies' peripheral tissues under darkness. However, whereas the worms' vertebrate/animalCry2 ortholog tr-Cry showed transcriptional repressor activity in a heterologous S2 cell assay, Platynereis L-Cry (Pdu-L-Cry) did not under the same conditions. Future work may determine if (worm) cell type-specific factors could be required for L-Cry to exert transcriptional activity or if it could rather regulate posttranscriptional responses (Zurl, 2022).
A second lead on how moonlight can impact on ~24-h timing is provided by identification of r-Opsin1 as a second moonlight sensor. Whereas it remains to be uncovered how the r-Opsin1-dependent signals tie in with the different signaling states of L-Cry, the existence of two distinct sensors already opens up the possibility for a combinatorial setup, in which the (nuclear) presence of L-Cry allows an Opsin-dependent signal to be interpreted as moonlight, whereas the reduction of L-Cry upon sunlight illumination allows an incoming Opsin-dependent signal to be interpreted as sunlight. Just the combination of the two signals alone would be sufficient for the light distinction. This combinatorial model could also explain the situation of Drosophila, where purified dCRY biochemically does not respond to naturalistic moonlight but is genetically required to shield the Drosophila circadian clock from moonlight impact. Opsin-based light input also plays a role for circadian entrainment, including the detection of light at moonlight intensity, and together with the information from dCRY, the natural source of light could be discriminated. For a diurnal/crepuscular species such as D. melanogaster, the main importance of the discrimination between sunlight and moonlight is to reduce the impact of moonlight on its ~24-h clock and not to tune it to moonlight. Therefore, the moonlight sensitivity of the cryptochrome component itself is a dispensable feature for the fruitfly's dCRY. In contrast, the nocturnal bristle worm fine-tunes its plastic PCC with moonlight. A direct sensitivity of L-Cry to moonlight is likely beneficial for this moonlight-responding system and hence exists biochemically for L-Cry (Zurl, 2022).
Evidence for plasticity of the conventional circadian clock has started to emerge from other marine systems: work on the circatidal oscillators of oysters maintained under controlled laboratory conditions revealed that core circadian clock genes exhibit ~12.4-h cycles under constant darkness, whereas the transcripts of the same genes cycle with an ~24-h oscillation under light/dark conditions. This provides evidence for the ability of the canonical clock to alternate between circadian (~24 h) and (semi)circalunidian (~12.4 h/~24.8 h) periodicities. Of note, switches between circadian and circalunidian cycles might also occur in humans. For instance, mood switches of bipolar patients correlate with a period lengthening of their body temperature cycles that looks as if the circadian timing system can be intermittently entrained to a 24.8-h rhythm. Moreover, already classical chronobiological studies documented changes of the ~24-h clock periodicity under dim light in various organisms, including birds, mice, hamsters, and humans, as well as the fruit fly D. melanogaster. Whereas the meaning of these results had remained enigmatic, they could well be explained by the conceptual framework of combined solar and lunar light cues that is presented in this study. It is anticipated that research on organisms for which lunar impact is of known biological relevance will be key to disentangle the interplay of solar and lunar timing cues (Zurl, 2022).
Sleep is highly conserved across animal species. Both wake- and sleep-promoting neurons are implicated in the regulation of wake-sleep transition at dusk in Drosophila However, little is known about how they cooperate and whether they act via different mechanisms. This study demonstrated that in female Drosophila, sleep onset was specifically delayed by blocking the Shaker cognate L channels (Shal, also known as voltage-gated K(+) channel 4, Kv4) in wake-promoting cells, including large ventral lateral neurons (l-LNvs) and pars intercerebralis (PI), but not in sleep-promoting dorsal neurons (DN1s). Delayed sleep onset was also observed in males by blocking Kv4 activity in wake-promoting neurons. Electrophysiological recordings show that Kv4 channels contribute A-type currents (IA) in LNvs and PI cells, but are much less conspicuous in DN1s. Interestingly, blocking Kv4 in wake-promoting neurons preferentially increased firing rates at dusk around ZT13, when the resting membrane potentials (RMPs) and firing rates were at lower levels. Furthermore, pigment-dispersing factor (PDF) is essential for the regulation of sleep onset by Kv4 in l-LNvs, and downregulation of PDF receptor (PDFR) in PI neurons advanced sleep onset, indicating Kv4 controls sleep onset via regulating PDF/PDFR signaling in wake-promoting neurons. It is proposed that Kv4 acts as a sleep onset controller by suppressing membrane excitability in a clock-dependent manner to balance the wake-sleep transition at dusk. These results have important implications for the understanding and treatment of sleep disorders such as insomnia (Feng, 2018).
This is the first demonstration of the function of Kv4 in sleep regulation. This study has shown that Kv4 is important for night-time sleep in Drosophila, and is especially crucial to normal sleep onset. Pan-neuronal expression of DNKv4 leads to decreased night-time sleep, indicating a general sleep-promoting function of Kv4. The increased night-time sleep in Shal495-null mutant reveals a compensatory overexpression of Shaker. A transcription factor named Kruppel (Kr) was identified to be a central regulator of this process, consistent with the conclusion that the compensatory modulation occurs at the transcriptional level. However, the Kr expression is suggested to be a result from detecting Kv4 or Shaker/Kv1 conductance, since 4-Aminopyridine (4-AP) also increased Shaker RNA expression. In this study, no significant increase was seen in Shaker RNA expression in DNKv4 mutants suggesting that other transcription factor(s), rather than Kr, may be involved in regulating Shal/Kv4 and Shaker balance (Feng, 2018).
Previous studies have shown that cyclin A and GABAA receptors (or RDL) in circadian neurons also regulate sleep latency. A molecule named WAKE interacts with RDL, and the cycling manner of WAKE promotes excitability oscillation of l-LNvs. Moreover, DN1 neurons may regulate wake-sleep transition at dusk in a clock-dependent manner. The results verified that Kv4-mediated IA in l-LNvs, and function-loss of Kv4 preferentially upregulated membrane excitability at dusk, although Kv4 expression was not rhythmic. Moreover, resting membrane potential (RMP) values exhibit circadian oscillation in l-LNvs, with depolarized RMPs at dawn. Thus, less Kv4 channels would be available for opening when RMPs are depolarized. This supports data showing that blocking Kv4 did not significantly increased firing rate when circadian neurons were hyperexcitable at dawn. Although RMPs and firing rate also exhibit circadian oscillation in DN1s, blocking Kv4 activity has no effects on sleep latency. It was further demonstrated that this may be caused by the near absence of Kv4-mediated IA in DN1s. Thus, Kv4 controls wake-sleep transition at dusk in subsets wake-promoting cells, but not in DN1s. The phases of Ca2+ waves recorded with a fine temporal resolution were approximately coincident with membrane excitability and RMPs oscillations in l-LNvs, but with slight (∼1-2 h) delay in peaks. In this study, DNKv4 preferentially reduced excitability at dusk without causing daily excitability oscillation shifts, and presumably would reduce Ca2+ peaks as well. But how Kv4 exactly regulates intracellular Ca2+ waves and coordination of electrical excitability and Ca2+ waves need further study (Feng, 2018).
PI exerts its effects as downstream of clock neurons and is one of the key targets (direct or indirect) of PDF neurons, although PI cells do not express molecular clock machinery. Previous studies have suggested that EGFR/ERK signal in insulin-producing PI neurons plays important roles in regulating the consolidation and maintenance of sleep in Drosophila, indicating that PI cells act as wake-promoting neurons. These results are the first to demonstrate that the neuronal activities and PDF/PDFR signaling in subsets of PI cells (50y-GAL4 driver) participate in the regulation of sleep onset (Feng, 2018).
In this study, defects in sleep initiation were presumed to be due to the hyperactivity of specific circadian neurons caused by blocking Kv4 activity. dTrpA1 experiments further supported this notion. However, a recent study concluded that reducing the Shaker/Kv1 current would decrease, rather than increase, the action potential discharge in dFB (dorsal FB; Pimentel, 2016). Depleting Shaker from dFB neurons could shift the interspike interval distribution toward longer values, making it more difficult to generate next spike. It is supposed that the opposite effect may be due to diverse types of Kv channels and different discharge and biophysical properties of neurons. For example, Kv4 is required for maintaining excitability in cultured neurons (Ping, 2011), but not in groups of neurons from dissected brain in this study. Whether driving DNKv4 expression in other sleep-promoting or wake-promoting neurons would cause parallel or opposite effects on sleep needs to be investigated. Neurons in EB, FB, or a subset MBNs driven by 201y-GAL4 could be tested in further studies (Feng, 2018).
Previous work has provided a strong link between circadian periods with habitual sleep timing in human, and CRY1 is suggested to be associated with a familial form of delayed sleep phase disorder. However, no significant change was detected in circadian periods by blocking Kv4 in LNvs and DN1s, indicating that delayed sleep onset in the mutants was not due to changes in circadian periods. Blocking Kv4 in PI cells disrupted rhythmic activities in most flies, and a lengthened circadian period was detected in the rhythmic ones. Because PI is a circadian output region and does not express clock machinery, it is not likely that intrinsic circadian rhythm disorder contributes to the sleep onset phenotype caused by blocking Kv4 (Feng, 2018).
Studies have provided evidence that links types of potassium channels to sleep phenotypes in Drosophila and human. For example, insomnia has been delineated in patients with neurologic disorders of Voltage-gated K+ channels (Kv) autoimmunity (or named Kv antibody syndrome). Sleep onset insomnia, defined as inability to fall asleep at the desired time, is observed in patients with neurodegenerative diseases and psychiatric disorders. Abnormal sleep timing and pattern have also been observed in Drosophila disease models. This study may provide a potential alternative therapy of sleep onset insomnia by targeting Kv4 channels (Feng, 2018).
Like in mammals, Drosophila circadian clock neurons display rhythms of activity with higher action potential firing rates and more positive resting membrane potentials during the day. This rhythmic excitability has been widely observed but, critically, its regulation remains unresolved. This study characterized and modeled the changes underlying these electrical activity rhythms in the lateral ventral clock neurons (LNvs). Currents mediated by the voltage-gated potassium channels Shaw (Kv3) and Shal (Kv4) oscillate in a circadian manner. Disruption of these channels, by expression of dominant negative (DN) subunits, leads to changes in circadian locomotor activity and shortens lifespan. LNv whole-cell recordings then show that changes in Shaw and Shal currents drive changes in action potential firing rate and that these rhythms are abolished when the circadian molecular clock is stopped. A whole-cell biophysical model using Hodgkin-Huxley equations can recapitulate these changes in electrical activity. Based on this model and by using dynamic clamp to manipulate clock neurons directly, the pharmacological block of Shaw and Shal can be rescued, restoring the firing rhythm, and thus demonstrating the critical importance of Shaw and Shal. Together, these findings point to a key role for Shaw and Shal in controlling circadian firing of clock neurons and show that changes in clock neuron currents can account for this. Moreover, with dynamic clamp the LNvs between morning-like and evening-like states of electrical activity can be switched. It is concluded that changes in Shaw and Shal underlie the daily oscillation in LNv firing rate (Smith, 2019).
Imbalances in amount and timing of sleep are harmful
to physical and mental health. Therefore, the study of the underlying
mechanisms is of great biological importance. Proper timing and amount
of sleep are regulated by both the circadian clock and homeostatic
sleep drive. However, very little is known about the cellular and
molecular mechanisms by which the circadian clock regulates sleep.
This study describes a novel role for Diuretic
hormone 31 (DH31), the fly homolog of the vertebrate neuropeptide
calcitonin gene-related peptide, as a circadian wake-promoting signal
that awakens the fly in anticipation of dawn. Analysis of
loss-of-function and gain-of-function Drosophila mutants demonstrates
that DH31 suppresses sleep late at night. DH31 is expressed by a subset
of dorsal circadian clock neurons that also express the receptor for the
circadian neuropeptide pigment-dispersing factor (PDF). PDF secreted by
the ventral pacemaker subset of circadian clock neurons acts on PDF
receptors in the DH31-expressing dorsal clock neurons to increase DH31
secretion before dawn. Activation of PDF receptors in DH31-positive DN1
specifically affects sleep and has no effect on circadian rhythms, thus
constituting a dedicated locus for circadian regulation of sleep. This
study has identified a novel signaling molecule (DH31) as part of a
neuropeptide relay mechanism for circadian control of sleep. The results
indicate that outputs of the clock controlling sleep and locomotor
rhythms are mediated via distinct neuronal pathways (Kunst, 2014). Vertebrate Calcitonin gene-related peptide (CGRP) has been implicated
in controlling anxietyand stress response. While not previously
addressed experimentally, the intimate relationship between stress,
anxiety, and sleep suggests that CGRP might regulate sleep. Potentially
relevant to this possibility is the recent observation that acute
activation of CGRP signaling in zebrafish larvae increases spontaneous
locomotor activity and decreases quiescence. This analysis of
gain-of-function and loss-of-function mutant flies establishes that
DH31, the Drosophila homolog of CGRP, is a negative regulator of sleep
maintenance that awakens the animal in anticipation of dawn. This
finding motivates investigation of a potential role for CGRP in the
regulation of vertebrate sleep (Kunst, 2014). Using powerful tools for cell-specific neuronal manipulation, this
study identified a highly restricted subset of DN1 circadian clock
neurons that secrete DH31 late at night to awaken the fly in
anticipation of dawn. Neuropeptides secreted from the circadian
pacemaker in the mammalian SCN, such as prokinecticin 2 and
cardiotrophin-like cytokine, are important clock outputs, but their
cellular targets and molecular mechanisms remain unknown. In flies,
PDF-expressing sLNv pacemaker neurons are known to be upstream of DN1s,
sLNvs are most active around dawn, and PDF secretion from LNvs
suppresses sleep at night. However, how PDF signals propagate out of the
circadian clock network to regulate sleep remains unknown (Kunst, 2014).
This study shows that PDF secreted by the sLNv pacemaker neurons
activates PDFR in the DH31-expressing DN1s to increase neuronal activity
and DH31 secretion late at night, thereby awakening the fly in
anticipation of dawn. This is consistent with an earlier report
demonstrating that flies lacking PDF or PDFR sleep more late at night.
However, unlike DH31, PDF also promotes wake during the day. This
suggests that PDF regulation of daytime sleep is mediated by neurons
other than the DH31-expressing DN1s. PDFR is expressed by circadian
clock neurons in addition to DN1s, and PDF signaling to these other
clock neurons could be responsible for the wake-promoting effect of PDF
during the day (Kunst, 2014). While PDF plays a key role in circadian timekeeping, neither
constitutive activation of PDFR in the DH31-expressing DN1s nor
manipulation of their secretion of DH31 affects free-running circadian
rhythms. This establishes the PDF-to-DH31 neuropeptide relay as a novel
sleep-specific output of the circadian pacemaker network, independent
from and parallel to the outputs that drive circadian rhythms
themselves. Since the basic cellular and molecular organization of
vertebrate and insect circadian networks is conserved, this motivates
the search for similar mechanisms in the SCN. Future studies are
required to identify the neuronal targets of sleep-regulating DH31
signals and the cellular and molecular mechanisms by which DH31-R1
activation in these targets induces wake (Kunst, 2014).
The regulation of sleep and the response to sleep deprivation rely on multiple biochemical pathways. A critical connection is the link between sleep and metabolism. Metabolic changes can disrupt sleep, and conversely decreased sleep can alter the metabolic environment. There is building evidence that lipid metabolism, in particular, is a critical part of mounting the homeostatic response to sleep deprivation. This study evaluated an acyl-CoA synthetase, pudgy (pdgy), for its role in sleep and response to sleep deprivation. When pdgy transcript levels are decreased through transposable element disruption of the gene, mutant flies showed lower total sleep times and increased sleep fragmentation at night compared to genetic controls. Consistent with disrupted sleep, mutant flies had a decreased lifespan compared to controls. pdgy disrupted fatty acid handling as pdgy mutants showed increased sensitivity to starvation and exhibited lower fat stores. Moreover, the response to sleep deprivation is reduced when compared to a control flies. When the transcript levels for pdgy were decreased using RNAi, the response to sleep deprivation was decreased compared to background controls. In addition, when pdgy transcription is rescued throughout the fly, the response to sleep deprivation is restored. These data demonstrate that the regulation and function of acyl-CoA synthetase plays a critical role in regulating sleep and the response to sleep deprivation. Endocrine and metabolic signals that alter transcript levels of pdgy impact sleep regulation or interfere with the homeostatic response to sleep deprivation (Thimgan, 2018).
Sleep is a dynamic process in most animals, involving distinct stages that probably perform multiple functions for the brain. Before sleep functions can be initiated, it is likely that behavioral responsiveness to the outside world needs to be reduced, even while the animal is still awake. Recent work in Drosophila has uncovered a sleep switch in the dorsal fan-shaped body (dFB) of the fly's central brain, but it is not known whether these sleep-promoting neurons also govern the acute need to ignore salient stimuli in the environment during sleep transitions. This study found that optogenetic activation of the sleep switch suppressed behavioral responsiveness to mechanical stimuli, even in awake flies, indicating a broader role for these neurons in regulating arousal. The dFB-mediated suppression mechanism and its associated neural correlates requires innexin6 expression, suggesting that the acute need to reduce sensory perception when flies fall asleep is mediated in part by electrical synapses (Troup, 2018).
Before sleep can begin to achieve any of its multiple putative functions, it would seem that behavioral responsiveness to the outside world first needs to be reduced. This is a fundamental yet poorly understood aspect of sleep in all animals. Mechanisms that regulate behavioral responsiveness are therefore likely to be involved during transitions from wakefulness to sleep, so it seems parsimonious that overlapping neuronal systems might govern both behavioral states. In this way, sleep onset has some similarity with selective attention in its capacity to gate or suppress perception rapidly. This study shows that gap junctions that are expressed in the sleep-promoting neurons of the dFB in the central fly brain are probably mediators for gating behavioral responsiveness in both awake and sleeping flies, as a consequence of acutely increased membrane potential in these neurons. Recent work has shown that sleep pressure increases the input resistance and membrane time constants in these same neurons (Donlea, 2014; Pimentel, 2016), so this seems to be a probable mechanism whereby increased sleep pressure impairs behavioral responsiveness via electrical channels, perhaps even before promoting and maintaining sleep - which may then occur via chemical (e.g. peptidergic) signaling from these neurons (Donlea, 2018). It is therefore possible that the R23E10 neurons, which have been recently characterized as comprising part of a 'sleep homeostat' in the fly brain, have two distinct arousal-related functions: to regulate behavioral responsiveness more generally -- and thus to promote sleep acutely -- and then to maintain sleep for the duration required by homeostatic demands. As increased arousal thresholds are a key criterion for sleep, it is of course difficult to disentangle these processes from each other. However, the data suggest that dFB-mediated suppression of behavioral responsiveness could be a prequel to sleep. First, on the basis of the current definitions of sleep, it does not make sense for sleep to precede changes in responsiveness, so either these processes are simultaneous or sleep functions succeed an initial increase in arousal thresholds that may already be evident during wakefulness (Troup, 2018).
The experiments described in this study show that behavioral responsiveness can be suppressed by acutely activating the dFB even in awake flies, and that this effect has little inertia, at the levels of both behavior and electrophysiology. Sleep intensity was increased however upon prolonged dFB activation, suggesting a distinct cumulative effect. Indeed, at the deepest stage of sleep (when flies are least responsive to mechanical stimuli), removal of INX6 from the dFB had no effect on behavioral responsiveness, suggesting that other factors are involved at that stage for maintaining high arousal thresholds. When flies are closer to awakening again, or when they are engaged in lighter sleep stages, it is possible that INX6-mediated mechanisms come back into play to maintain sleep. In recent work, it has been shown that transitions in and out of sleep are associated with dFB activity, which is consistent with the view that a common system might be governing behavioral responsiveness during wakefulness as well as sleep (Troup, 2018).
It is likely that other dFB neurons outside of the R23E10 circuit are also involved, as suggested by only partial overlap with INX6 labeling. However, without INX6, activated R23E10 neurons were unable to reduce behavioral responsiveness, suggesting a cell-autonomous effect. This study has shown that dFB activation is associated with increased local field potential (LFP) activity in the central brain, which could reflect increased synchronous neural firing. Without expression of INX6 in the R23E10 neurons, these induced LFP effects also disappear, suggesting that gap junctions are important for promoting synchronous activity in this circuit. This suggests a rapid mechanism through which the dFB might suppress behavioral responsiveness upon sleep onset, by for example producing synchronous low frequency activity that could interfere with ongoing sensory processing and integration in the central brain. Although acute activation of the R23E10 neurons alone is unlikely to occur in a natural context, electrical communication to other cells via gap junctions could support subtler levels of behavioral control while the flies are awake, which might consolidate into overall increased arousal thresholds as membrane potentials across the dFB increase with increasing sleep pressure. The possibility was not excluded that a fast-acting neuropeptide or neurotransmitter secreted from R23E10 neurons regulates behavioral responsiveness, but the data favor the view that a parallel channel involving electrical synapses exists. Future studies should uncover the extent of this gap-junction-coupled network, although antibody-labeling and dye-coupling experiments suggest that cells in the pars intercerebralis (PI) are probably involved. It also remains possible that dFB neurons are electrically coupled to each other, thereby generating LFP oscillations that are employed in the regulation of behavior. However, in dye-coupling experiments, no evidence was seen of labeling of another dFB cell, whereas multiple PI cells were labeled (Troup, 2018).
It is not known whether electrical communication might be employed to promote sleep in other animal brains, although there is increasing evidence that gap junctions play an important role in the regulation of behavioral state and arousal in the mammalian brain. More generally, a role for gap junctions in sleep-promoting neurons also suggests a novel plasticity mechanism for regulating behavioral states. Whereas plasticity is typically viewed as a property of chemical synapses, the parallel electrical communication channel afforded by gap junctions could provide an alternate way of regulating sleep pressure, and thereby promoting sleep functions while ensuring that arousal thresholds are tightly linked to sleep need. Recent work has shown that transient electrical activation of the dFB neurons during fly brain development permanently impairs behavioral responsiveness in adult animals, further confirming the strong link between these neurons and the regulation of arousal in Drosophila. Different INX6 expression levels in the dFB may provide a mechanism for optimally linking behavioral responsiveness with sleep need. It will be interesting to see whether INX6 expression in the dFB correlates with the striking range of individual differences that were observed for sleep duration and behavioral responsiveness in wildtype flies, and whether INX6 expression in the dFB might be co-regulated alongside other proteins that have been shown to control neuronal excitability in this arousal circuit (Troup, 2018).
Nicotinic acetylcholine receptors (nAChRs) are a highly conserved gene family that form pentameric receptors involved in fast excitatory synaptic neurotransmission. The specific roles individual nAChR subunits perform in Drosophila melanogaster and other insects are relatively uncharacterized. Of the 10 D. melanogaster nAChR subunits, only three have described roles in behavioral pathways; Dα3 and Dα4 in sleep, and Dα7 in the escape response. Other subunits have been associated with resistance to several classes of insecticides. In particular, previous work has demonstrated that an allele of the Dα1 subunit is associated with resistance to neonicotinoid insecticides. This study used ends-out gene targeting to create a knockout of the Dα1 gene to facilitate phenotypic analysis in a controlled genetic background. This is the first report of a native function for any nAChR subunits known to be targeted by insecticides. Loss of Dα1 function was associated with changes in courtship, sleep, longevity, and insecticide resistance. While acetylcholine signaling had previously been linked with mating behavior and reproduction in D. melanogaster, no specific nAChR subunit had been directly implicated. The role of Dα1 in a number of behavioral phenotypes highlights the importance of understanding the biological roles of nAChRs and points to the fitness cost that may be associated with neonicotinoid resistance (Somers, 2017).
Nicotinic acetylcholine receptors (nAChRs) belong to the Cys-loop receptor subfamily of ligand-gated ion channels (LGICs) that mediate the transduction of a chemical signal into an electrical signal. Activation in response to acetylcholine (ACh), the major excitatory neurotransmitter of the Drosophila melanogaster central nervous system, is on a micro- to submicrosecond timescale. nAChRs are expressed in a wide range of tissues, including but not limited to the nervous system. When expressed presynaptically, nAChRs can enhance neurotransmitter release, while postsynaptic expression mediates excitation. Like all LGICs, nAChRs are pentameric and can consist of several different subunits forming multiple receptor subtypes that vary in both their sensitivity to particular ligands and their permeability to particular cations. Individual subunits consist of a pore-forming transmembrane domain coupled to a large extracellular N-terminal domain, which forms the endogenous ligand-binding site with adjacent subunits. Upon ACh binding, a conformational change occurs opening the channel pore to permit the flow of cations (Somers, 2017).
D. melanogaster has 10 nAChR subunits; however, only three of these have been associated with behavioral phenotypes, with specific roles described for Dα7 in the escape response and for both Dα3 and Dα4 in sleep behavior. Mutations in three D. melanogaster nAChR subunits confer resistance to two important classes of insecticides; Dα1 and Dβ2 to neonicotinoids, and Dα6 to spinosyns. Modification of an insecticide target protein can decrease the efficacy of an insecticide through altered affinity or pharmacological response. Modifications of this nature can provide a significant fitness boost to individuals in a population during insecticide treatment periods and can rapidly increase in frequency leading to control failures. However, allele frequencies will also be shaped by fitness costs if resistant mutations negatively impact reproductive output, either by reducing viability or the capacity to mate. This could be particularly true for alleles that directly modify the neonicotinoid-binding site as neonicotinoids occupy the same binding site as ACh. Loss-of-function mutants for the Dα1, Dα6, and Dβ2 genes are insecticide-resistant and viable. The viability phenotype suggests a level of functional redundancy among nAChR subunits. Questions remain as to whether individual subunits have other, nonredundant functions in controlling behavior or if the subtle pharmacological and physiological differences from compensating subunits manifests in altered behaviors. Impacts on mating behavior are of particular interest with respect to potential fitness costs (Somers, 2017).
While invertebrate nAChR pharmacology and biochemistry with respect to insecticide binding has been examined in detail, little research has been devoted to the endogenous functions of these receptors. D. melanogaster is commonly used as a model insect system to study many facets of biology, including insecticide resistance. A large number of well-defined behavioral paradigms exist to investigate the roles of genes in traits, including sleep and cognition through to mating and auditory faculties (Somers, 2017).
This study reports the creation of a Dα1 knockout mutant and a rescue system through transgene expression. This provided a consistent genetic background to analyze several behavioral paradigms. These reagents were used to identify roles for the D. melanogaster Dα1 subunit in mating, locomotion, and sleep, demonstrating the diverse pleiotropic influences that nAChRs can have on insect behavior. This research has relevance to the consideration of fitness costs that might be associated with resistance conferring mutations in Dα1 orthologs in pest insects, and the behavioral impact that exposure to neonicotinoids may have in beneficial species such as the western honey bee, Apis mellifera (Somers, 2017).
To generate a Dα1 null mutant, a modified ends-out targeting scheme was used. A precise 57-kb deletion of the Dα1 genomic region was created, then validated by Southern blot and sequencing. This mutant is referred to as Dα1KO. Given that previously created Dα1 alleles are resistant to neonicotinoids (Perry, 2008), Dα1KO was screened on two different neonicotinoids (imidacloprid and nitenpyram) and was found to be highly resistant to both. Calculated LC50 values and resistance ratios were consistent with those measured for Dα1 mutants studied previously (Perry, 2008). Dα1 and orthologs in pest species are well-established targets for several different neonicotinoid insecticides. Heterologous expression and affinity chromatography studies have also implicated Dα1 in directly binding imidacloprid at a site that overlaps that normally occupied by the ligand, ACh. Therefore, these resistance data matched the prediction that a mutant with a genomic deletion of Dα1 would be resistant to neonicotinoid insecticides (Somers, 2017).
The GAL4-UAS system was employed to create a phenotypic rescue in which a Dα1 cDNA clone was expressed in the Dα1KO background. Expression of the Dα1 clone using the pan-neuronal elav::GAL4 driver rescued sensitivity to both imidacloprid and nitenpyram. The reversion of the resistance phenotype indicates that the subunit expressed from the Dα1 transgene is assembling into functional nAChRs that bind these insecticides (Somers, 2017).
The loss of Dα1 results in significant levels of resistance to neonicotinoids. However, the level of resistance is not of the same magnitude observed in the Dα6 knockout mutant, which is over 1000-fold resistant to spinosad. The Dα1KO mutant is still susceptible when exposed to a high enough dose, which may be due to expression of other neonicotinoid-sensitive nAChR subtypes. Mutations in the orthologs of Dα3 and Dβ1 have been identified in neonicotinoid-resistant strains of Nilaparvata lugens and Myzus persicae, respectively. While only one imidacloprid-binding site has been reported in adult D. melanogaster and other Dipteran and Lepidopteran species, multiple binding sites have been reported in several Hemipteran species. Unlike Hemipterans, Dipterans and Lepidopterans are holometabolous insects that undergo complete metamorphosis from larva to adult. It is possible that as yet undescribed imidacloprid-sensitive nAChR subtypes are expressed in the larval life stages. Another possibility is that a novel subtype is formed as a consequence of the loss of the Dα1 subunit that alters the mutant's sensitivity to neonicotinoids (Somers, 2017).
RNAi knockdown of the Dα1 subunit resulted in defects in courtship and copulation behavior. Previous microarray studies, confirmed by RT-PCR, highlighted expression of Dα1 in both neuronal and reproductive tissues. Taken together, these lines of evidence suggested the potential for Dα1 to function in mating behavior (Somers, 2017).
Mating behavior can be influenced by genetic background, for example expression of w is important for visual cues and misexpression of this gene can trigger male-male courtship. As the Dα1KO line was generated in the w1118 background, both the mutant and the control line have functionally null copies of w. Therefore, the w+ X chromosome from another isogenic line, RAL059, was used to replace the w1118 X chromosome present in both the Dα1KO mutant line and the w1118 background line. These lines will be referred to as the mutant and wild-type lines respectively (Somers, 2017).
Courtship behavior was measured in terms of the latency of courtship initiation by males after female introduction into the mating chamber. Flies that failed to initiate courtship within the allowed 10-min period were given a maximum value. Wild-type males initiated courtship in every trial, regardless of the genotype of the female partner. Mutant males only initiated courtship 65% of the time with wild-type females and 79% of the time with mutant females. Mutant males also took significantly longer to initiate courtship than wild-type males when paired with either wild-type or mutant females (Somers, 2017).
Copulation latency was also measured for the same flies. Mutant males rarely copulated within the 10-min period, only 15% of trials when paired with a wild-type female and only 3% when paired with a mutant female. Wild-type males were more successful, initiating copulation in 90% of the trials when paired with a wild-type female and in 56% of the trials when paired with a mutant female. No significant difference was observed in copulation latency of mutant males when paired with either wild-type or mutant females. In contrast, wild-type males did initiate copulation significantly faster with wild-type females compared to mutant females. This suggests that, unlike the latency of courtship initiation that is primarily influenced by the genotype of the male, both sexes contribute to the latency of copulation initiation (Somers, 2017).
The rescue system was again employed to see if expression of a Dα1 transgene could rescue the courtship and copulation phenotypes observed for the Dα1KO mutant. Appropriate driver-only and UAS-only control flies were used for comparison to rescue flies. No discernible differences in courtship initiation were observed when wild-type males were crossed to control or rescue female flies. However, when crossed to wild-type female flies, male rescue flies were more successful in initiating courtship than male control flies. Male rescue flies also showed a significant decrease in courtship initiation when crossed to female wild-type compared to male control flies. Significant rescue of copulation initiation was also observed in both female and male rescue flies. Rescue flies were both more successful and faster at initiating copulation than the appropriate controls (Somers, 2017).
It is clear from the data that Dα1 plays a role in Drosophila mating behavior; however, the underlying mechanism remains unknown. Defects in ACh signaling have previously been associated with abnormal mating behavior. Analysis of mosaic mutants, defective for cholinergic signaling, identified a neuropile in the mushroom body calyx critical for normal male courtship behavior. Male-specific, cholinergic neurons have also been identified in the abdominal ganglion, the disruption of which significantly decreased male fertility, potentially due to their innervation of the male reproductive system. The disruption of these neurons has been hypothesized to result in the uncoordinated or altered release of sperm, seminal fluid, and accessory proteins. The study, performed by Acebes (2004) used the presynaptic choline acetyltransferase marker to identify these neurons; however, the receptors receiving this signal were not identified. The data from mating experiments and expression analysis suggests that Dα1 may be one of the specific nAChR subunits expressed in this pathway. Another possibility to consider is a higher processing role for Dα1 in mating behavior. The phenotypes observed in the Dα1KO mutant are consistent with a defect in one or more sensory modalities. While there are specialized receptors responsible for detecting sensory stimuli, cholinergic signaling has been identified in connecting sensory circuitry to processing centers in the brain. Dα1 expression has previously been observed in the mushroom body calyx and lateral protocerebrum, which includes the lateral horn. Recently, the role of the lateral horn in locusts has been proposed to serve as a site for multimodal sensory integration. This may suggest that Dα1 plays a role in integrating multimodal courtship circuitry to higher sensory processing centers. Challenging Dα1KO mutants with sensory-specific behavioral paradigms and neuron-specific rescue could test this hypothesis (Somers, 2017).
The possibility was explored that impaired locomotion may be contributing to the mating phenotype observed in the Dα1KO mutant. Over a 24-hr period, the Dα1KO mutant exhibited hyperactivity; however, it moved at a slower average speed. When average speed was binned in 3-hr intervals, the difference was significant during the middle two 3-hr bins of the day and the last three 3-hr bins of the night. Most importantly, there was no significant difference in average speed during the first 3-hr bin of the day, when courtship assays were performed, indicating that general locomotion deficits did not impact the measurement of mating behavior (Somers, 2017).
The Dα1KO mutant also has an unusual pattern of sleep. Although there were no significant differences observed in total amount of sleep, mutant flies slept significantly less during the night, experiencing less sleep episodes of significantly shorter duration than wild-type controls. The total time of day sleep experienced by the mutant was significantly longer with a higher number of sleep episodes; however, there was no observable difference in episode length. Expression of the Dα1 transgene in the mutant background increased amount of sleep, both during the day and night. This increase was a result of longer sleep episodes, which may explain why both mutant and rescue flies both show less night sleep episodes. Rescue flies have less night sleep episodes due to the length of these episodes, whereas mutant flies may have trouble with sleep initiation and maintenance. The exceptional increase in episode length observed in rescue flies is likely due to nonnative expression of the transgene; however, it is clear Dα1 influences sleep (Somers, 2017).
While activation of all cholinergic neurons inhibits sleep in flies, different neuronal groups can be wake-promoting or sleep-promoting. Individual nAChR subunits also appear to have different roles in sleep regulation. From this study, Dα1 seems to have net sleep-promoting effects, especially with sleep maintenance. The increase in daytime sleep observed in mutant flies may be a compensation mechanism to cope with the reduced amount of nighttime sleep. Further experiments are needed to determine whether the Dα1KO mutant has intact sleep homeostatic regulation, and whether loss of sleep affects sleep quality. Sleep deprivation has been linked to various detrimental effects, namely reduced life span and learning deficits (Somers, 2017).
Longevity of Dα1KO mutants was measured, revealing a much shorter life span in the mutant compared to the wild-type. While longevity is not a direct measure of fitness, the mutants' reduced life span highlights the importance of the Dα1 subunit in D. melanogaster physiology. It is not clear if this effect is due to a single physiological role of Dα1, such as the subunits involvement in sleep, or cumulative effects of several impaired physiological roles that impact the mutant's longevity. In either case, it suggests that a resistance allele in the Dα1 subunit is likely to impact the fitness levels of the insect. It also supports the notion that sublethal exposures to insecticides that target receptor subunits orthologous to Dα1 encountered by beneficial insects, such as honey bees, are likely to affect their behavior in ways that may also impact their fitness (Somers, 2017).
The Dα1KO mutant generated in this study provides a useful tool to study the role of Dα1 in insecticide resistance but, more significantly, to explore the endogenous roles of this gene (Somers, 2017).
Based on prior evidence, it was expected that the Dα1KO mutant would be resistant to neonicotinoid insecticides (Perry, 2008). However, the data presented in this study allow a fresh evaluation of the value of Dα1 and orthologs in other species as insecticide targets. The ability of the Dα1 knockout flies to survive at all presents a potential issue for resistance evolution to compounds that target this receptor. Given that a wide range of mutations would lead to a total loss-of-function phenotype, such mutations will arise frequently, conferring insecticide resistance. The data presented in this study indicate that, looking beyond viability, there is a significant fitness cost associated with the total loss of Dα1 function, most obvious in terms of severe mating behavior defects, but possibly contributed to by the sleep and longevity phenotypes. Under optimal environmental conditions in the laboratory, large phenotypic differences between Dα1KO and control flies were observed. It is possible that the fitness cost associated with a null allele of this gene would be even greater under less ideal conditions experienced by natural populations. Therefore, while a nonsense mutation resulting in a resistance allele may be viable, it would likely be associated with fitness costs that would prevent it from persisting in the field. In contrast, mutations that may be null alleles have been found in Dα6 orthologs in spinosad-resistant insects in a number of species. Therefore, it is possible that the spectrum of mutations in Dα1 orthologs that can increase in frequency to confer insecticide resistance in a pest species are constrained by fitness costs (Somers, 2017).
The potential impact of neonicotinoid insecticides on the behavior of beneficial insects, such as A. mellifera, has been intensively researched. Eleven nAChR subunits have been identified in A. mellifera, including a 1:1 ortholog of Dα1. While it cannot be assumed that the functional roles of the honey bee ortholog are identical to those of Dα1, it is likely to influence a range of behaviors that may be perturbed upon exposure to sufficient concentrations of neonicotinoid insecticides (Somers, 2017).
The genetic resources that can be developed to study gene function in D. melanogaster, such as those described here, are extremely powerful. Research on nonmodel insects, both beneficial and pest species, is more challenging. Some functional analysis of Dα6 orthologs from pest species has been possible following the appropriate expression of these genes in a D. melanogaster Dα6 null mutant. A similar approach may be useful in the functional characterization of the orthologs of Dα1 (Somers, 2017).
Insecticides can be powerful probes in neuroscience. To exert toxic effects, these chemicals bind to receptors that have crucial roles in neurotransmission. Research using pyrethroids and cyclodiene insecticides has been productive in elucidating the function of sodium channels and ligand-gated chloride channels, respectively. Similarly, the use of insecticides that target nAChRs has stimulated research on their function. The data demonstrates that such research will make a vital contribution in providing a detailed knowledge of the role neurotransmission in a wide range of behaviors. The variety of behavioral traits impacted by Dα1 loss-of-function indicates a high level of involvement of Dα1 in several behavioral neural circuits, which will require further investigation. Further analysis of Dα1 and the remaining members of the nAChR gene family with a range of paradigms is likely to reveal function in a wide range of insect behaviors (Somers, 2017).
A relationship between sleep and seizures is
well-described in both humans and rodent animal models; however, the
mechanism underlying this relationship is unknown. Using Drosophila
melanogaster mutants with seizure phenotypes, this study demonstrated
that seizure activity can be modified by sleep deprivation. Seizure
activity was evaluated in an adult bang-sensitive seizure mutant, stress
sensitive B (sesB9ed4), and in an adult
temperature sensitive seizure mutant seizure
(seits1) under baseline and following 12 h of
sleep deprivation. The long-term effect of sleep deprivation on young,
immature sesB9ed4 flies was also assessed. Sleep
deprivation increased seizure susceptibility in adult sesB9ed4
and seits1 mutants. Sleep deprivation also increased
seizure susceptibility when sesB was disrupted using RNAi. The
effect of sleep deprivation on seizure activity was reduced when sesB9ed4/+
flies were given the anti-seizure drug, valproic acid. In contrast to
adult flies, sleep deprivation during early fly development resulted in
chronic seizure susceptibility when sesB9ed4/+ became
adults. These findings show that Drosophila is a model organism for
investigating the relationship between sleep and seizure activity
(Lucey, 2014). With advancing age, humans demonstrate a number of sleep disruptions,
which contribute to poor health consequences and accelerated senescence.
The mechanisms that lead to these negative health outcomes, however,
remain unclear. This study shows that both baseline and recovery sleep
change with age and these alterations can be modulated by the chemical
chaperone PBA. PBA is a low molecular weight fatty acid that has been
shown to act as a chemical chaperone for misfolded or mislocalized
proteins. It has also been shown to suppress ER-stress induced cell
death without the aid of endogenous ER chaperones. It was also
demonstrated in this study that directly inducing ER stress in young
flies fragments baseline sleep and alters recovery sleep, phenocopying
aged flies. Sleep fragmentation may contribute to accelerated aging and
it has been suggested that an intervention that consolidates sleep may
delay the aging process and increase life span (Brown, 2014). Aging also impairs quality control systems that are necessary for
protein homeostasis. Earlier work has demonstrated that aging decreases
BiP levels and increases pro-apoptotic factors such as
CCAAT/enhancer-binding protein-homologous protein (CHOP) in the cerebral
cortex of mice. Young flies experience less ER stress and their adaptive
UPR is fully functional. Aged flies, on the other hand, are under
chronic ER stress conditions with a diminished adaptive UPR. Therefore,
basal ER stresses for the two groups are vastly different and likely
contributes to the disparate responses to sleep deprivation. The young
flies are readily able to handle acute sleep deprivation, whereas aged
flies, already under stress, are overwhelmed by the secondary stressor.
Addition of PBA has little effect on recovery sleep in young flies;
however, PBA treatment ameliorates existing ER stress and suppresses
further UPR induction in aged flies. As a result, treated aged flies
display more efficient recovery sleep or are able to discharge sleep
debt more efficiently than untreated aged flies (Brown, 2014). Although no significant differences were found in BiP expression in
flies treated with PBA, XBP1 was significantly affected by PBA
treatment. PBA not only reduces the amount of spliced XBP1, it also
reduces the expression of the unspliced variant as well. PBA likely
suppresses UPR induction by chaperoning unfolded proteins, so that BiP
stays bound to IRE1 and PERK and delays or prevents its activation, even
under the condition of sleep deprivation. Phospho-IF2α levels are
significantly reduced in the PBA treated aged flies subjected to sleep
deprivation. Changes in p-eIF2α can be directly linked to the behavioral
changes in the aged flies given that increased p-eIF2α facilitates
baseline and recovery sleep. The reduced eIF2α phosphorylation in PBA
treated animals may account for the improved homeostatic response in the
aged SD flies (Brown, 2014). This study also chemically induced ER stress in young flies and then
subjected them to sleep deprivation. This acute tunicamycin treatment
was found to decrease and fragment baseline sleep and phenocopy the
recovery sleep behavior that is seen in the aged flies. There are two
possible explanations for the tunicamycin results. First, tunicamycin
activity leads to inhibition of glycosylation and general misfolding of
proteins, which will ultimately result in ER stress. Secondly,
tunicamycin could potentially prevent maturation of glycoproteins
required for sleep maintenance. This data demonstrates that ER stress
induces sleep fragmentation. Together with previous findings that sleep
deprivation caused ER stress and activated the UPR, these results
suggests that both processes are intimately linked and feedback on one
another (Brown, 2014). PBA has been shown to significantly increase life span in Drosophila,
purportedly through inhibition of HDAC activity. The moderate effects of
PBA on sleep consolidation in young sssp1 mutant flies suggest
that protein stabilization by PBA is only partially ameliorating defects
in these animals. The decrease in sleep in the ctrl sss
background strain treated with PBA supports the increase and
consolidation of wake in the wild-type lines. It has been previously
shown that induction of ER stress impairs waking in mice. The study
speculates that application of PBA also promotes wake-active neuronal
function and consolidates wake in the flies by reducing ER stress. The
issue is not the accumulation of sleep debt, but the discharge of sleep
debt that is impaired in the aged flies. By ameliorating ER stress and
suppressing activation of the UPR, PBA allows for more efficient
recovery sleep in the aged flies. Since PBA ameliorates ER stress by
directly reducing aberrant protein load, treating Drosophila
with PBA supplements chaperone activity and decreases the burden of
misfolded proteins that occurs as a consequence of an external stressor,
sleep deprivation, as well as normal aging (Brown, 2014). Circadian coordination of metabolism, physiology, and
neural functions contributes to healthy aging and disease prevention.
Clock genes govern the daily rhythmic expression of target genes whose
activities underlie such broad physiological parameters as maintenance
of redox homeostasis. Previously, it was reported that glutathione
(GSH) biosynthesis is controlled by the circadian system via effects
of the clock genes on expression of the catalytic (Gclc)
and modulatory (Gclm)
subunits comprising the glutamate cysteine ligase (GCL) holoenzyme. The
objective of this study was to determine whether and how aging, which
leads to weakened circadian oscillations, affects the daily profiles of
redox-active biomolecules. Fly aging was found to be associated with
altered profiles of Gclc and Gclm expression at both
the mRNA and protein levels. Analysis of free aminothiols and GCL
activity revealed that aging abolishes daily oscillations in GSH levels
and alters the activity of glutathione biosynthetic pathways. Unlike
GSH, its precursors and products of catabolism, methionine, cysteine and
cysteinyl-glycine, were not rhythmic in young or old flies, while
rhythms of the glutathione oxidation product, GSSG, were detectable. The
study concludes that the temporal regulation of GSH biosynthesis is
altered in the aging organism and that age-related loss of circadian
modulation of pathways involved in glutathione production is likely to
impair temporal redox homeostasis (Klichko, 2015). This study investigated a role for the circadian system in regulating
redox in the context of organismal aging. Cellular redox homeostasis
largely relies on the redox-active compound, glutathione, which is
present at concentrations many fold higher compared to concentrations of
other redox-active molecules. Previous studies have shown that the
circadian clocks modulate the de novo synthesis of GSH via
transcriptional control of GCLc and GCLm, the subunits that comprise the
GCL holoenzyme. This study broadened the investigation of the
relationship between clock and redox to determine the levels of other
redox-active molecules, involved in glutathione synthesis and
metabolism. Around the clock expression profiles of both cysteine and
its precursor methionine were investigated (Klichko, 2015). As neither cysteine nor methionine exhibited evidence for diurnal
rhythms in either young or old flies, it appears that the contribution
of the trans-sulfuration pathway to glutathione homeostasis is not
regulated by circadian clocks. Consistent with these findings,
methionine was found to be arrhythmic in the study of the human
metabolome of blood plasma and saliva although the analyses were
performed with healthy but older (57-61 years) males, where
age-dependent effects might have influenced the oscillations. In
contrast, mouse hepatic metabolome and transcriptome studies revealed
rhythmicity in metabolic sub-pathways, where oscillations in glutathione
were ascertained by oscillations in its precursors, cysteine and
methionine, albeit with a lower amplitude for the latter (Klichko,
2015). Analysis of the Drosophila heads revealed no cycling in the
concentrations of Cys-Gly, consistent with the arrhythmic behavior of
cysteine and methionine. Given that Cys-Gly also serves as a signature
of glutathione degradation, interpretation of these results are somewhat
tentative. Nevertheless, these results revealed no rhythmicity in
cysteine, methionine and Cys-Gly, and suggest that, at least in flies,
the pathways responsible for the supply of sulfur-containing precursors
for glutathione synthesis are not regulated by the circadian clocks. It
should be noted that the mammalian liver is a homogenous tissue with a
strong food-entrained clock mechanism, while fly heads are enriched in
nervous tissues with clocks entrained by light-dark cycles. Moreover
recent analysis of the circadian transcriptome shows that liver
possesses the highest number of rhythmic genes, while brain has the
lowest (Klichko, 2015). Another important finding of this study is that the diurnal
fluctuations in GSH levels were not followed by similar changes in the
products of its degradation (Cys-Gly) and oxidation (GSSG). While
Cys-Gly was completely arrhythmic, changes in GSSG profile did not
mirror those observed for GSH. Even though both shared the same slow
drop-off from ZT0 to ZT8 as well as the ZT8 trough, their peaks were
quite distinct (ZT12 for GSSG and ZT20 for GSH). Also in old flies, a
certain degree of rhythmicity is maintained for GSSG in contrast to the
absence of any diurnal GSH patterns (Klichko, 2015). Another important finding of this study is that the rhythms in
glutathione levels observed in young flies were lost in old flies,
presumably due to the loss of diurnal fluctuations of GCLc, GCLm as well
as GCL activity, in response to the weakening of the circadian clocks.
In contrast, GSSG rhythms were largely preserved in older flies,
suggesting that the daily changes in glutathione disulfide levels are
supported by enzymatic reactions that are not under clock control
(Klichko, 2015). Despite loss of circadian regulation, average daily levels of GSH
remained unchanged during aging, while the levels of GSSG were slightly
higher, mainly due to lesser drop in the early morning. In Drosophila,
it has been established that whole body GSH levels were either
relatively constant or slightly decreased during aging while GSSG rose
2-3 fold. Similar age-related changes were documented in different
mammalian tissues with the most significant reduction in GSH and
accumulation of GSSG in the brain, indicative of a more pro-oxidative
cellular environment. As such changes in GSH and GSSG were frequently
associated with increases in enzyme activities related to GSH usage, the
relatively steady glutathione concentrations observed in the heads of
old flies could point to less efficient GSH utilization (Klichko, 2015).
The rather unexpected finding of this study is that the expression of
Gclc at both mRNA and protein levels significantly increased in the
heads of old flies, and this increase was associated with about 25%
higher average daily GCL activity. Despite this increase, the average
daily levels of GSH remained unchanged suggesting a loss in GCL
catalytic efficiency or an age-related increase in GSH utilization. One
possible scenario is that the efficiency of GSH synthesis can be induced
by oxidative stress, in part through the well-documented increase in
H2O2 signaling that accompanies aging. For instance, post-translational
control of γ-glutamylcysteine (γ-Glu-Cys) synthesis is influenced by
oxidative stress, which can dramatically affect formation of GCL
holoenzyme and its stabilization (Klichko, 2015). Consistent with induction of GCL by stress, it was previously reported
that per-null mutants with disrupted clock displayed
arrhythmic as well as elevated GCL activity, which was also reflected in
arrhythmic and elevated ROS levels relative to the control. It should be
noted that previous studies comparing GCL activity and GSH levels in
young and old rats showed a decrease of both parameters in liver, while
in aging brain and heart GSH decreased but GCL activity remained
unchanged, pointing again to catalytic deficiency of the enzyme
(Klichko, 2015). Other aminothiols that did not show cycling in young flies also
remained arrhythmic in old flies, but displayed changes in their steady
state levels. Consistent with previous reports, the amounts of Cys-Gly
were ~50% higher in older flies. Cys-Gly, derived from the breakdown of
glutathione, is required for GSH synthesis as a precursor of cysteine,
but at the same time it is also a prooxidant generated during the
catabolism of glutathione. The requirement of Cys-Gly for GSH synthesis
justifies its increase with age, as the tissues require an increased
supply of precursors for GSH biosynthesis in older flies. However, an
increase in cysteine levels during aging was not observed. A more
plausible explanation is that the increase in Cys-Gly is indicative of
an increase in oxidative stress and GSH degradation. In agreement with
this view, the average daily levels of methionine were about 35% lower
in old flies suggesting the likelihood of an increase in oxidation of
methionine to methionine sulfoxide by ROS rather than an increase in
methionine consumption for cysteine biosynthesis. Together, these
changes indicate a shift in redox homeostasis in the heads of older
flies, consistent with the earlier reports in whole flies. Similar
alterations in the redox components were also indicative of heightened
oxidative stress in pathologies like systemic lupus erythematosus
(Klichko, 2015).
Gene expression is much altered in aging. This study observed age-dependent decline of core clock genes' expression in the whole body of the fruit fly. It was hypothesized that inducible overexpression of clock genes (cry, per, tim, cyc and Clk) in the nervous system can improve healthspan of D. melanogaster. The lifespan of transgenic Drosophila was studied; life extension was shown for cry, per, cyc and tim genes. There were significant positive changes in the stress-resistance of flies overexpressing core clock genes in conditions of hyperthermia, hyperoxia, starvation and persistent lighting. The overexpression of per and cry restore circadian rhythms of locomotor activity. The results presented support the hypotheses that the compensation of circadian oscillator genes expression can improve the healthspan in Drosophila melanogaster (Solovev, 2019a).
Diet restriction is one of the most accurately confirmed interventions which extend lifespan. Genes coding circadian core clock elements are known to be the key controllers of cell metabolism especially in aging aspect. The molecular mechanisms standing behind the phenomenon of diet-restriction-mediated life extension are connected to circadian clock either. This study investigated the effects of protein-rich and low-protein diets on lifespan observed in fruit flies overexpressing core clock genes (cry, per, Clk, cyc and tim). The majority of core clock genes being upregulated in peripheral tissues (muscles and fat body) on protein-rich diet significantly decrease the lifespan of male fruit flies from 5 to 61%. Nevertheless, positive increments of median lifespan were observed in both sexes, males overexpressing cry in fat body lived 20% longer on poor diet. Overexpression of per also on poor medium resulted in life extension in female fruit flies. Diet restriction reduces mortality caused by overexpression of core clock genes. Cox-regression model revealed that diet restriction seriously decreases mortality risks of flies which overexpress core clock genes. The hazard ratios are lower for flies overexpressing clock genes in fat body relatively to muscle-specific overexpression. The present work suggests a phenomenological view of how two peripheral circadian oscillators modify effects of rich and poor diets on lifespan and hazard ratios (Solovev, 2019).
Early studies from psychology suggest that sleep facilitates memory
retention by stopping ongoing retroactive interference caused by
mental activity or external sensory stimuli. Neuroscience research
with animal models, on the other hand, suggests that sleep facilitates
retention by enhancing memory consolidation. Recently, in Drosophila,
the ongoing activity of specific dopamine neurons was shown to
regulate the forgetting of olfactory memories. This study shows that
this ongoing dopaminergic activity is modulated with behavioral state,
increasing robustly with locomotor activity and decreasing with rest.
Increasing sleep-drive, with either the sleep-promoting agent
Gaboxadol or by genetic stimulation of the neural circuit for sleep,
decreases ongoing dopaminergic activity, while enhancing memory
retention. Conversely, increasing arousal stimulates ongoing
dopaminergic activity and accelerates dopaminergic-based forgetting.
Therefore, forgetting is regulated by the behavioral state modulation
of dopaminergic-based plasticity. These findings integrate
psychological and neuroscience research on sleep and forgetting (Berry
2015). While some memories are long-lasting, most others fade away and are
forgotten. Why we forget, has been an intriguing and central question
in psychology and neuroscience for more than a century. Even though
forgetting is often thought of as a failure or limitation of the
brain, recent studies support the view that forgetting is a
biologically regulated function of the brain allowing optimal
adaptability to an ever-changing environment. In the fruit fly
Drosophila, it was recently shown that the very same set of dopamine
neurons (DANs) that signal through one receptor to form aversive
olfactory memories, also signal through a separate receptor after
learning to forget these memories (Berry, 2012). However, it remains
unclear whether this dopaminergic forgetting signal is constant and
autonomous, or dynamic and regulated (Berry 2015). From fruit flies to humans, animals routinely alternate between
highly active behavioral states and long states of immobility and
quiescence called sleep. Despite the obvious disadvantages an
inanimate state conveys to survival, sleep has been proposed to have
critically important functions, including in memory and cognition.
Since the earliest experimental studies of human memory, sleep shortly
after learning has been shown to consistently lead to an increase in
retention and thus less forgetting of many forms of memory including
declarative and emotional memory in mammals and long-term courtship
memory in Drosophila. However, there exists controversy as to how
sleep benefits memory retention. Many studies in mammals support the
idea that sleep benefits memory retention because it is accompanied by
specific mechanisms, such as slow wave sleep, rapid eye movement (REM)
sleep, and sharp-wave ripple-based memory replay, that increase memory
retention by actively consolidating newly formed memories.
Alternatively, it was proposed nearly a century ago and recently
revisited, that sleep, or long periods of quiet wakefulness, benefit
memory retention by muting experience-driven plasticity and new memory
formation, thus reducing retroactive interference-based forgetting. In
addition, this state of reduced neuronal activity might then allow
consolidation to occur more efficiently, referred to as the
"opportunistic consolidation" model. Thus, the essence of how exactly
sleep benefits memory retention remains debated (Berry 2015). Previous studies have observed that, after promoting the acquisition
of olfactory memories, a small set of DANs that innervate the mushroom
body (MB) memory center, intriguingly, display synchronized and
ongoing Ca2+-based activity after learning that causes the forgetting
of early aversive olfactory memories in Drosophila(Berry, 2012). While
this activity occurs as reoccurring bursts, it was noticed that the
pattern of activity appeared temporally regulated, occurring in bouts.
In order to understand how the DAN-based forgetting signal might be
regulated, an in vivo imaging assay was developed allowing
simultaneous monitoring of a fly's DAN Ca2+ activity, via GCaMP3.0
expression using TH-gal4 and behavior while walking on a ball
supported by air. Focus was placed on two regions of the DAN processes
that form synaptic connections to the MBs, referred to as neuropils,
one that displays ongoing activity and belongs to the MV1 neuron and
an adjacent control region belonging to the V1 neuron, which is
relatively inactive. Remarkably, a 1-hr simultaneous recording of
locomotion and DAN activity revealed that the MV1 neuropil displayed
activity resembling the coarse temporal pattern of locomotor behavior.
Ball rotation data was used to cluster time points into either
behaviorally active or rest states, and MV1 neuropil activity was
found to be robustly elevated during active states, whereas the V1
neuropil activity remained low in both states, but had a slight
decrease during active states. Furthermore, the MV1 neuropil Ca2+
signal was strongly correlated with ball rotation, particularly in the
lower frequency domains (frequency < 0.002 Hz, or ~1 cycle every 8
min or more, timescales consistent with that of locomotor bout
structure). Finally, DAN activity was examined during stable
transitions into and out of behaviorally active states ( by aligning
transition segments of recordings across all animals. Interestingly,
MV1 DAN activity robustly increased upon transition into active
states, while, conversely, dropped during rest states. V1 activity
remained low and was not significantly regulated with behavioral
transitions. Together, these data, along with observations of
synchronized activity between MV1 and another DAN, MP1, indicate that
the ongoing activity from specific sets of DAN involved in forgetting,
including MV1, is regulated with the behavioral state of the animal
(Berry 2015). Given the strong correlation between DAN activity and locomotor
activity, tests were performed to see whether DAN activation might
promote locomotor activity, that is, whether DAN activity is upstream
of locomotor circuits. Two prior studies found no role for these MB
innervating DANs in regulating locomotor activity. When the synaptic
output from these DANs was blocked with restricted expression in MV1,
MP1, and V1 DANs to drive temperature-sensitiveUAS-shits1function,
no decreased locomotor activity was seen between temperatures,
although the experimental genotype exhibited less activity at high
temperature compared to one but not both control genotypes. It was
also noted from imaging experiments that locomotor activity
occasionally occurs while the MV1 neuron is not active, thus further
supporting that locomotor behavior does not require c150-gal4>DAN
output. Furthermore, stimulation of these neurons, using UAS-trpA1,
did not produce genotype specific and robust increases in locomotor
activity. But similar to the blocking experiments, high temperature
increased the locomotor activity of the two control genotypes
(UAS-trpA1 and c150-gal4 alone). These data indicate that c150-gal4
DAN output is neither necessary nor sufficient to acutely drive
locomotor activity. It is therefore concluded that the ongoing signal
in MV1 is either downstream of locomotor behavior itself, or is
regulated in parallel, by other brain areas that promote arousal and
locomotor activity (Berry 2015). Given that the ongoing activity in MV1 was highest during
behaviorally active states, the hypothesis was tested that reducing
behavioral activity with increased sleep drive would reduce this
ongoing activity. The GABAA agonist, Gaboxadol (or THIP), has been
shown to specifically promote deep non-REM sleep in humans, while
leaving REM sleep intact; sleep characteristics similar to those
occurring during normal homeostatic regulation of sleep. Recently, it
was shown that Gaboxadol also induces sleep in Drosophila. To confirm
this, attempts were made to induce sleep in Drosophila by feeding
flies various doses of Gaboxadol. Shortly after Gaboxadol feeding,
long periods of quiescence, (>5 min) conventionally defined as
'sleep' in Drosophila, significantly increased during both day and
night in a dose-dependent manner. Next, flies were fed Gaboxadol (0.1
mg/ml) for 1 day and then removed the drug to test whether these
effects were reversible. Once again, Gaboxadol treatment increased
sleep, occurring as bouts with increased duration, but remarkably,
total sleep and bout duration actually decreased after drug removal
compared to control fed flies. These data indicate that less sleep is
needed in flies given Gaboxadol the prior day, suggestive of a
homeostatic response. Finally, to test the arousability of flies fed
Gaboxadol, a single mechanical stimulus was delivered every hour for 1
day followed by a day of drug treatment. Interestingly, the
average-evoked activity, post-stimulus was significantly reduced with
increasing Gaboxadol dosage. These data suggest that having Gaboxadol
onboard increases arousal thresholds. Altogether, these data indicate
that Gaboxadol, similar to effects on mammals, induces bona fide sleep
in Drosophila, with hallmark characteristics that include reversible
quiescence, homeostatic regulation, and increased arousal thresholds
(Berry 2015). In order to observe the effects of Gaboxadol on DAN activity, varying
concentrations of this sleep agent were perfused across the brain
while performing in vivo imaging of MV1 activity and fly body movement
was monitored in a recording chamber. Like walking on the ball,
ongoing MV1 activity was also regulated with behavioral state in this
assay, increasing during bouts of body movement. Remarkably, Gaboxadol
perfusion rapidly and robustly attenuated both fly movement and MV1
activity at 0.01 and 0.1 mg/ml. Furthermore, it was found that the
quiescent behavioral state and reduced MV1 activity was fully
reversible with wash out, thus eliminating pharmacological-induced
damage as a cause of decreased physical and DAN activity (Berry 2015).
To rule out non-specific effects of Gaboxadol and extend these
results, sleep drive was increased sharply by thermogenetic
stimulation of the sleep circuit. Recent studies identified a dorsal
fan shaped body (dfsb) circuit in the central brain, specifically
represented in the R23E10-gal4 line, which acts as the effector arm of
the sleep homeostat (Donlea; 2011 and Donlea; 2014b). Consistent with
these studies, TrpA1-based stimulation of R23E10-gal4-expressing
neurons caused a rapid and robust increase in daytime sleep followed
by a negative sleep rebound the day after stimulation, confirming the
dfsb circuit's role in homeostatic sleep regulation. In order to
measure DAN activity in vivo while using the gal4-uas system to
modulate the sleep circuit, a TH-lexA line was developed to express
GCaMP3.0 in the MV1 and V1 DAN neurons, their associated MB
neuropil regions, as well as DAN innervation of the anterior inferior
medial protocerebrum (PR), a region also exhibiting ongoing activity
like MV1 (Berry, 2012). While simultaneously measuring movement and
DAN activity, before ('Pre'), during ('Treat'), and after ('Post')
stimulation of dfsb neurons was recorded. As predicted, stimulation of
the sleep circuit rapidly decreased fly behavioral activity and was
accompanied with a robust decrease in MV1 and PR DAN activity, with no
change in the control V1 region. Fly behavioral activity was partially
restored and ongoing activity in MV1 and PR completely restored to
pre-stimulation levels after stimulation of the sleep circuit was
ceased. These results, along with those from Gaboxadol administration,
indicate that increased sleep drive dramatically reduces the ongoing
activity of DANs involved in forgetting (Berry 2015). Since ongoing MV1 activity is decreased with increasing sleep drive,
it was hypothesized that acutely and reversibly increasing sleep drive
specifically after learning would reduce DAN-mediated forgetting. To
test this, sleep was modulated with Gaboxadol after aversive olfactory
conditioning, where populations of flies learn to associate an odor
with electric shock. Memory to this association is then tested in a
T-maze, giving flies the choice between the trained odor and an
unconditioned odor. Since memory from this kind of training decays
quickly after training, attempts were made to increase the rate of
Gaboxadol consumption and thus the rate of sleep onset, by increasing
the hunger of flies via starvation prior to feeding. As was observed
previously, flies fed Gaboxadol experienced more sleep than controls,
and a 16-hr starvation period increased this effect. Furthermore, it
was found that flies removed from Gaboxadol food 1 hr after learning
partially returned to control sleep and activity levels by the sixth
hour and completely by the eighth hour after learning, indicating that
these time points were appropriate for testing memory retrieval. This
Gaboxadol feeding protocol led to increased sleep during the period of
memory retention. Remarkably, it was found that flies forced to sleep
with Gaboxadol treatment after learning exhibited enhanced memory
retention at both 6 and 8 hr. Similarly, sleep circuit stimulation
after conditioning also rapidly and reversibly induced sleep and
enhanced both 3- and 6-hr memory retention. Importantly, simultaneous
stimulation of the dfsb sleep circuit and c150-gal4 DANs also led to
strong sleep induction. Memory retention, in contrast, was markedly
decreased, similar to that observed with stimulation of DANs alone.
Therefore, sleep, after learning, loses its protective qualities when
DAN signaling is artificially potentiated. This circuit level
epistasis experiment indicates that DAN-mediated forgetting is
downstream of sleep networks. Together, the data indicate that
increased sleep and reduced arousal after learning reduces
DAN-mediated forgetting of aversive olfactory memories (Berry 2015). If the DANs innervating the MB memory center are downstream of
arousal circuitry, then they should respond to arousing stimuli. In
fact, these neurons have already been shown to respond to many salient
stimuli, including odors and electric shock and temperature changes.
Since mechanical stimuli have been extensively used to arouse flies
for sleep deprivation, airpuffs were delivered to the fly using a
protocol shown to induce arousal in flies, while simultaneously
recording fly movement and DAN activity. It was found that the DAN
processes in all three areas (MV1, PR, V1) of the mushroom body
neuropil exhibited robust responses to each airpuff. However, MV1
responsiveness was maintained across stimuli while the other regions
showed attenuated responsiveness across stimuli. Importantly, both fly
movement and ongoing MV1 activity continued at an elevated level just
after stimulation, indicating a stimulus-induced elevation in arousal
and MV1 DAN activity (Berry 2015). Next, it was reasoned that increasing the arousal after learning
would accelerate DAN-mediated forgetting. To test this, a population
arousal device was developed that allowed delivery of a mechanical
stimuli (2-s stimulus every 1 min over 80 min) to flies in population
vials after aversive olfactory learning. It was found that mechanical
stimuli delivered for the first 80 min after learning significantly
aroused populations of flies, leading to an overall increase in
activity between each stimulus, with activity levels dropping back to
control levels after treatment. Importantly, mechanical stimulation
after learning caused a robust decrease in 3-hr memory for wild-type
Canton-S flies. However, acquisition and immediate memory retrieval
were not disrupted by prior mechanical stimulation, indicating that
the stimuli must be delivered after learning to observe its disruptive
effects. Remarkably, it was found that blocking neuronal output of
c150-gal4DANs specifically during mechanical stimulation blocked the
forgetting induced by this treatment. Therefore, these data indicate
that increasing arousal after learning accelerates DAN-mediated
forgetting (Berry 2015). The following conclusions are made from the data. First, after
learning, the ongoing DA forgetting signal is not constant but instead
is regulated with behavioral state. Thus, the forgetting signal does
not chronically remove memories at a constant rate. Second, the
ongoing forgetting signal is coupled directly to the arousal level of
the animal, being suppressed with low levels of arousal such as with
the state of sleep and being enhanced by activation of sensory
pathways. As a result, forgetting decreases when flies rest or sleep
and increases when flies are aroused by external stimuli (Berry 2015).
DA is known to regulate various types of plasticity in mammals. In
flies, DA has been shown to elicit presynaptic plasticity within the
Kenyon cells of the MB memory center proposed to underlie learning.
Additionally, it was previously found that DA after learning regulates
forgetting (Berry, 2012), thus implicating a DA-based plasticity
mechanism that weakens memories. Synthesizing these previous
observations with the current data, it is proposed that the behavioral
state-coupled DA signal, discovered in this study, regulates the
plasticity of the memory system, making it malleable for memory
updating so that memories of current events can be formed and old,
unused memories can be forgotten. While it was found previously that
different DA receptors underlie learning and forgetting, more work
remains to distinguish the molecular cascades involved and the
cellular events that underlie these forms of behavioral plasticity
(Berry 2015). These findings add compelling mechanistic evidence to support the
model that sleep, which begins with and is accompanied by inactivity
or rest, benefits newly formed labile memories by reducing the level
of plasticity induced by behavioral activity. Furthermore, as sleep
progresses and arousal thresholds increase, DANs become less reactive
to stimuli. Thus, the molecular/cellular model is congruent with early
psychological models of sleep benefitting memory by muting the
retroactive interference that causes forgetting. Nevertheless, the
data do not eliminate the possibility that sleep-specific mechanisms
exist that enhance memory consolidation, as often proposed from
studies with mammalian systems. Mechanistically, the effects of sleep
on memory consolidation and forgetting may operate in parallel and
independently of one another or more intriguingly; they may operate in
serial in a dependent fashion, with reduced forgetting being a
prerequisite for sleep-facilitated consolidation, similar to the
'opportunistic consolidation' model proposed by Mednick, 2011 (Berry
2015). This study has observed that multiple DANs
produce the ongoing DA signal, synchronized across the MB memory
center and protocerebrum, that leads to forgetting of olfactory
memories. It remains to be determined if this network activity is but
one segment of a larger and more diffuse DA network that operates on
memory types other than olfactory; whether there exist multiple,
independent forgetting networks regulated by arousal levels; and
whether forgetting of non-olfactory memories occurs through DA-based
mechanisms or involves other neuromodulatory transmitters (Berry
2015). Alzheimers
disease (AD) is a neurodegenerative disease characterized by
severe cognitive deterioration. While causes of AD pathology are
debated, a large body of evidence suggests that increased cleavage of
Amyloid Precursor Protein (APP) producing the neurotoxic Amyloid-β (Aβ)
peptide plays a fundamental role in AD pathogenesis. One of the
detrimental behavioral symptoms commonly associated with AD is the
fragmentation of sleep-activity cycles with increased nighttime activity
and daytime naps in humans. Sleep-activity cycles, as well as
physiological and cellular rhythms, which may be important for neuronal
homeostasis, are generated by a molecular system known as the circadian
clock. Links between AD and the circadian system are increasingly
evident but not well understood. This study examined whether genetic
manipulations of APP-like (APPL) protein cleavage in Drosophila
melanogaster affect rest-activity rhythms and core circadian
clock function in this model organism. It was shown that the increased
β-cleavage of endogenous APPL by the β-secretase (dBACE)
severely disrupts circadian behavior and leads to reduced expression of
clock protein PER
in central clock neurons of aging flies. The study's data suggest that
behavioral rhythm disruption is not a product of APPL-derived Aβ
production but rather may be caused by a mechanism common to both α and
β-cleavage pathways. Specifically, it was shown that increased
production of the endogenous Drosophila Amyloid Intracellular
Domain (dAICD) causes disruption of circadian rest-activity rhythms,
while flies overexpressing endogenous APPL maintain stronger circadian
rhythms during aging. In summary, this study offers a novel entry point
toward understanding the mechanism of circadian rhythm disruption in
Alzheimer's disease (Blake, 2015). Loss of rest-activity rhythms is a well-established early symptom of AD
in humans. Because disruption of circadian rhythms is detrimental to
neuronal homeostasis, it is important to understand relationships
between AD and circadian rhythms at the cellular and molecular levels.
To address this question, this study examined how manipulations of the
fly ortholog of APP and its cleaving enzymes affect endogenous
rest-activity rhythms and clock mechanism in Drosophila.
Over-expression of dBACE was found to disrupt behavioral rest-activity
rhythms, and this effect is most severe in aged flies suggesting an
age-dependent mechanism. Furthermore, dBACE expression resulted in
dampened oscillation of the core clock protein PER in central pacemaker
neurons, which are master regulators of rest activity rhythms.
Significantly reduced PER levels are observed in the sLNv and lLNv
neurons of age 50d flies expressing dBACE in all clock cells (including
glia), all neurons, or only in PDF-positive sLNv and lLNv neurons. These
data suggest that manipulation of APP-cleavage by dBACE over-expression
directly affects the oscillation of PER protein in central pacemaker
neurons in a cell-autonomous manner. Since a functional clock mechanism
in sLNv is necessary and sufficient to maintain free running activity
rhythms, reduced oscillations of PER in these neurons could be
responsible for the loss of activity rhythms in age 50d flies.
Importantly, the decline in PER levels occurrs only in flies with
manipulated dBACE, not in old control flies. This is in agreement with
earlier findings that aging does not dampen PER oscillations in
pacemaker neurons of wild type flies, while it reduces clock
oscillations in peripheral clocks (Blake, 2015). While this study reports that the loss of behavioral rhythms after
manipulation of dBACE is associated with reduced expression of clock
genes in the central pacemaker, other recent work shows that expression
of human Aβ peptides leads to disruption of rest activity rhythms
without interfering with PER oscillations in the central pacemaker. Even
strongly neurotoxic Aβ peptides, such as Aβ42 arctic, do not cause
rhythm disruption when expressed in central pacemaker neurons; rather,
pan-neuronal expression is required. The fact that even the most
neurotoxic Aβ peptides are not capable of dampening PER oscillation in
pacemaker neurons suggests that Aβ production does not affect clock
oscillations and that it is not Aβ production that causes the phenotype
observed in this study upon over-expression of dBACE. This was confirmed
by expression of KUZ,
whose activity does not increase dAβ production; however, it also leads
to disruption of rest-activity rhythms. Similar rhythm disruption by
dBACE and KUZ suggests that an excess cleavage product of both pathways
might be responsible for the disruption. Like in the mammalian APP
cleavage pathway, in Drosophila cleavage of APPL by KUZ or
dBACE results in a C-terminal fragment (CTF) that is subsequently
cleaved by the ϒ-secretase resulting in the production of dAICD. Indeed,
it was shown that expression of dAICD results in an age-dependent
decline in rhythmic locomotor activity. As with dBACE and KUZ
expression, dAICD expression causes weakening or complete loss of
behavioral rhythms while age-matched control flies remain highly
rhythmic. In this context, it is worth noting that α-secretase
activators are considered for clinical trials to reduce Aβ production in
AD patients. However, according to results in this study, this could
lead to disruptions of circadian rhythms and sleep patterns thus
negatively impacting the lives of patients and their caretakers (Blake,
2015). This study's data suggest that increased dAICD may be the proximal
cause of decay in rest-activity rhythms. The role of AICD in AD is
increasingly evident but poorly understood. AICD is able to enter the
nucleus and has been implicated in transcriptional regulation that may
affect cell death, neurite outgrowth and neuronal excitability.
Interestingly, transgenic mice expressing AICD have increased activity
of GSK-3, which in flies affects the circadian clock. Over-expression of
GSK-3
in Drosophila leads to altered circadian behavior by
hyper-phosphorylation of
TIMELESS (TIM), a key circadian protein which forms dimers with
PER that enter the nucleus and regulate the clock mechanism. Of further
interest, increased GSK-3 activity has been implicated in AD, and in Drosophila,
increased GSK-3 activity mediates the toxicity of Aβ peptides (Blake,
2015). Cleavage of APPL likely results in a significant decline in intact
APPL, and this could be detrimental as APPL has neuroprotective effects.
It was also recently shown that loss of full-length APPL induces
cognitive deficits in memory. This study reports that flies
over-expressing full-length APPL in central pacemaker neurons maintain
stronger behavioral rest-activity rhythms during aging than control
flies; however this effect is not observed when APPL is expressed
pan-neuronally. This could be caused by negative effects of APPL when
expressed in other unspecified neurons, or could be related to driver
strength. Overall, the study suggests that the loss of full-length APPL
might negatively affect circadian behavior by way of the central
pacemaker neurons (Blake, 2015). Over-expression of dAICD induces a severe phenotype, disrupting
rest-activity rhythms as early as age 5d when expressed in central
pacemaker neurons and by age 35d with pan-neuronal expression. Taken
together these results suggest that while loss of full-length APPL by
over-expression of its secretases might negatively impact circadian
behavior, the cleavage product dAICD induces the most severe behavioral
rest-activity disruption. Interestingly, the observed effect is not
likely a product of neurodegeneration as it was previously shown that
dAICD has no effect on neurodegeneration, and this study shows that the
pacemaker cells appear intact in pdf > dAICD flies. In
addition, it was shown that dAICD, like the vertebrate AICD, can be
found in the nucleus. Therefore, this study suggests that dAICD may
directly or indirectly affect the expression of clock genes. This offers
a novel entry point toward understanding the mechanism of circadian
rhythm disruption in Alzheimer's disease (Blake, 2015). Sleep is an essential and conserved behavior whose
regulation at the molecular and anatomical level remains to be
elucidated. This study identifies Taranis
(Tara), a Drosophila homolog of the Trip-Br (SERTAD) family of
transcriptional coregulators, as a molecule that is required for normal
sleep patterns. Through a forward-genetic screen, tara was
isolated as a novel sleep gene associated with a marked reduction in
sleep amount. Targeted knockdown of tara suggests that it
functions in cholinergic neurons to promote sleep. tara
encodes a conserved cell-cycle protein that contains a Cyclin
A (CycA)-binding homology domain. Tara regulates CycA protein levels and
genetically and physically interacts with CycA to promote sleep.
Furthermore, decreased levels of Cyclin-dependent
kinase 1 (Cdk1), a kinase partner of CycA, rescue the
short-sleeping phenotype of tara and CycA mutants,
while increased Cdk1 activity mimics the tara and CycA
phenotypes, suggesting that Cdk1 mediates the role of Tare and CycA in
sleep regulation. Finally, a novel wake-promoting role was described for
a cluster of ∼14 CycA-expressing neurons in the pars
lateralis (PL), previously proposed to be analogous to the
mammalian hypothalamus. The study proposes that Taranis controls sleep
amount by regulating CycA protein levels and inhibiting Cdk1 activity in
a novel arousal center. Most animals sleep, and evidence for the essential nature of this
behavior is accumulating. However, how sleep is controlled at a
molecular and neural level is far from understood. The fruit fly,
Drosophila, has emerged as a powerful model system for understanding
complex behaviors such as sleep. Mutations in several Drosophila genes
have been identified that cause significant alterations in sleep. Some
of these genes were selected as candidates because they were implicated
in mammalian sleep. However, others (such as Shaker and CREB)
whose role in sleep was first discovered in Drosophila have later been
shown to be involved in mammalian sleep, validating the use of
Drosophila as a model system for sleep research. Since the strength of
the Drosophila model system is the relative efficiency of large-scale
screens, unbiased forward-genetic screens have been conducted to
identify novel genes involved in sleep regulation. Previous genetic
screens for short-sleeping fly mutants have identified genes that affect
neuronal excitability, protein degradation, and cell-cycle progression.
However, major gaps remain in understanding of the molecular and
anatomical basis of sleep regulation by these and other genes (Afonso,
2015). Identifying the underlying neural circuits would facilitate the
investigation of sleep regulation. The relative simplicity of the
Drosophila brain provides an opportunity to dissect these sleep circuits
at a level of resolution that would be difficult to achieve in the more
complex mammalian brain. Several brain regions, including the mushroom
bodies, pars intercerebralis, dorsal fan-shaped body, clock neurons, and
subsets of octopaminergic and dopaminergic neurons, have been shown to
regulate sleep. However, the recent discovery that Cyclin A (CycA) has a
sleep-promoting role and is expressed in a small number of neurons
distinct from brain regions suggests the existence of additional neural
clusters involved in sleep regulation (Afonso, 2015). From an unbiased forward-genetic screen, this study discovered taranis
(tara), a mutant that exhibits markedly reduced sleep amount. tara
encodes a Drosophila homolog of the Trip-Br (SERTAD) family of mammalian
transcriptional coregulators that are known primarily for their role in
cell-cycle progression. TARA and Trip-Br proteins contain a conserved
domain found in several CycA-binding proteins. This research shows that
tara regulates CycA levels and genetically interacts with CycA
and its kinase partner Cyclin-dependent kinase 1 (Cdk1) to
regulate sleep. Furthermore, a cluster of CycA-expressing neurons in the
dorsal brain was shown to lie in the pars lateralis (PL), a
neurosecretory cluster previously proposed to be analogous to the
mammalian hypothalamus, a major sleep center. Knockdown of tara
and increased Cdk1 activity in CycA-expressing PL neurons, as well as
activation of these cells, reduces sleep. Collectively, these data
suggest that TARA promotes sleep through its interaction with CycA and
Cdk1 in a novel arousal center (Afonso, 2015). From an unbiased forward genetic screen, this study has identified a
novel sleep regulatory gene, tara. The data demonstrate that
TARA interacts with CycA to regulate its levels and promote sleep. Cdk1
was also identified as a wake-promoting molecule that interacts
antagonistically with TARA. Given the fact that TARA regulates CycA
levels, the interaction between TARA and Cdk1 may be mediated by CycA.
The finding that Cdk1 and CycA also exhibit an
antagonistic interaction supports this view. The previous discovery that
CycE sequesters its binding partner Cdk5 to repress its kinase activity
in the adult mouse brain points to a potential mechanism, namely that
TARA regulates CycA levels, which in turn sequesters and inhibits Cdk1
activity. TARA and its mammalian homologs (the Trip-Br family of
proteins) are known for their role in cell-cycle progression. However,
recent data have shown that Trip-Br2 is involved in lipid and oxidative
metabolism in adult mice, demonstrating a role beyond cell-cycle
control. Other cell-cycle proteins have also been implicated in
processes unrelated to the cell cycle. For example, CycE functions in
the adult mouse brain to regulate learning and memory. Based on the
finding that CycA and its regulator Rca1 control sleep, it was
hypothesized that a network of cell-cycle genes was appropriated for
sleep regulation. The current data showing that two additional
cell-cycle proteins, TARA and Cdk1, control sleep and wakefulness
provide support for that hypothesis. Moreover, the fact that TARA and
CycA, factors identified in two independent unbiased genetic screens,
interact with each other highlights the importance of a network of
cell-cycle genes in sleep regulation (Afonso, 2015). There are two main regulatory mechanisms for sleep: the circadian
mechanism that controls the timing of sleep and the homeostatic
mechanism that controls the sleep amount. This study has shown that TARA
has a profound effect on total sleep time. TARA also affects rhythmic
locomotor behavior. Since TARA is expressed in clock cells, whereas CycA
is not, it is possible that TARA plays a non-CycA dependent role in
clock cells to control rhythm strength. The finding that tara
mutants exhibit severely reduced sleep in constant light suggests that
the effect of TARA on sleep amount is not linked to its effect on
rhythmicity. Instead, TARA may have a role in the sleep homeostatic
machinery, which will be examined in an ongoing investigation (Afonso,
2015). To fully elucidate how sleep is regulated, it is important to identify
the underlying neural circuits. This study has shown that activation of
the CycA-expressing neurons in the PL suppresses sleep while blocking
their activity increases sleep, which establishes them as a novel
wake-promoting center. Importantly, knockdown of tara and
increased Cdk1 activity specifically in the PL neurons leads to
decreased sleep. A simple hypothesis, consistent with the finding that
both activation of PL neurons and increased Cdk1 activity in these
neurons suppress sleep is that Cdk1 affects neuronal excitability and
synaptic transmission. Interestingly, large-scale screens for
short-sleeping mutants in fruit flies and zebrafish have identified
several channel proteins such as SHAKER, REDEYE, and ETHER-A-GO-GO and
channel modulators such as SLEEPLESS and WIDE AWAKE. Thus, it is
plausible that Cdk1 regulates sleep by phosphorylating substrates that
modulate the function of synaptic ion channels or proteins involved in
synaptic vesicle fusion, as has previously been demonstrated for Cdk5 at
mammalian synapses (Afonso, 2015). Whereas the data mapped some of TARA-s role in sleep regulation to a
small neuronal cluster, the fact that pan-neuronal tara knockdown
results in a stronger effect on sleep than specific knockdown in PL
neurons suggests that TARA may act in multiple neuronal clusters.
PL-specific restoration of TARA expression did not rescue the tara sleep
phenotype (data not shown), further implying that the PL cluster may not
be the sole anatomical locus for TARA function. Given that CycA is
expressed in a few additional clusters, TARA may act in all
CycA-expressing neurons including those not covered by PL-Gal4. TARA may
also act in non-CycA-expressing neurons. The data demonstrate that tara
knockdown using Cha-Gal4 produces as strong an effect on sleep as
pan-neuronal knockdown. This finding suggests that TARA acts in
cholinergic neurons, although the possibility cannot be ruled out that
the Cha-Gal4 expression pattern includes some non-cholinergic cells.
Taken together, these data suggest that TARA acts in PL neurons as well
as unidentified clusters of cholinergic neurons to regulate sleep
(Afonso, 2015). Based on genetic interaction studies, tara has been classified
as a member of the trithorax group genes, which typically act as
transcriptional coactivators. However, TARA and Trip-Br1 have been shown
to up- or downregulate the activity of E2F1 transcription factor
depending on the cellular context, raising the possibility that they
also function as transcriptional corepressors. Interestingly, TARA
physically interacts with CycA and affects CycA protein levels but not
its mRNA expression. These findings suggest a novel non-transcriptional
role for TARA, although an indirect transcriptional mechanism cannot be
ruled out. The hypothesis that TARA plays a non-transcriptional role in
regulating CycA levels and Cdk1 activity at the synapse may provide an
exciting new avenue for future research (Afonso, 2015). Despite an established link between epilepsy and sleep
behavior, it remains unclear how specific epileptogenic mutations
affect sleep and subsequently influence seizure susceptibility.
Recently, a fly knock-in model of human generalized epilepsy has been
established that exhibits febrile seizures plus (GEFS+), a
wide-spectrum disorder characterized by fever-associated seizing in
childhood and lifelong affliction. GEFS+ flies carry a disease-causing
mutation in their voltage-gated
sodium channel (VGSC) gene and display semidominant heat-induced
seizing, likely due to reduced GABAergic inhibitory activity at high
temperature. This study shows that at room temperature the GEFS+
mutation dominantly modifies sleep, with mutants exhibiting rapid sleep
onset at dusk and increased nighttime sleep as compared to controls.
GEFS+ mutant sleep phenotypes were more resistant to pharmacologic
reduction of GABA transmission by carbamazepine (CBZ) than controls, and
were mitigated by reducing GABAA receptor expression specifically in
wake-promoting pigment
dispersing factor (PDF) neurons. These findings are consistent
with increased GABAergic transmission to PDF neurons being mainly
responsible for the enhanced nighttime sleep of GEFS+ mutants. This
study reveals the sleep architecture of a Drosophila VGSC
mutant that harbors a human GEFS+ mutation, and provides unique insight
into the relationship between sleep and epilepsy (Petruccelli, 2015). Sleep is under the control of homeostatic and circadian
processes, which interact to determine sleep timing and duration, but
the mechanisms through which the circadian system modulates sleep are
largely unknown. This study used adult-specific, temporally controlled
neuronal activation and inhibition to identify an interaction between
the circadian clock and a novel population of sleep-promoting neurons
in Drosophila. In this study, transgenic flies expressed either dTRPA1,
a neuronal activator, or Shibirets1,
an inhibitor of synaptic release, in small subsets of neurons. Sleep, as
determined by activity monitoring and video tracking, was assessed
before and after temperature-induced activation or inhibition using
these effector molecules. This study compared the effect of these
manipulations in control flies and in mutant flies that lacked
components of the molecular circadian clock. Adult-specific activation
or inhibition of a population of neurons that projects to the
sleep-promoting dorsal Fan-Shaped Body resulted in bidirectional control
over sleep. Interestingly, the magnitude of the sleep changes were
time-of-day dependent. Activation of sleep-promoting neurons was
maximally effective during the middle of the day and night, and was
relatively ineffective during the day-to-night and night-to-day
transitions. These time-of-day specific effects were absent in flies
that lacked functional circadian clocks. In conclusion, the circadian
system functions to gate sleep through active inhibition at specific
times of day. These data identify a mechanism through which the
circadian system prevents premature sleep onset in the late evening,
when homeostatic sleep drive is high (Cavanaugh, 2015). Sleep is conserved across phyla and can be measured
through electrophysiological or behavioral characteristics. The fruit
fly, Drosophila melanogaster, provides an excellent model
for investigating the genetic and neural mechanisms that regulate sleep.
Multiple systems exist for measuring fly activity, including video
analysis and single-beam (SB) or multi-beam (MB) infrared (IR)-based
monitoring. This study compared multiple sleep parameters of individual
flies using a custom-built video-based acquisition system, and
commercially available SB- or MB-IR acquisition systems. It was found
that all three monitoring systems appear sufficiently sensitive to
detect changes in sleep duration associated with diet, age, and mating
status. It was also demonstrated that MB-IR detection appears more
sensitive than the SB-IR for detecting baseline nuances in sleep
architecture, while architectural changes associated with varying
life-history and environment are generally detected across all
acquisition types. Finally, video recording of flies in an arena allows
measurement of the effect of ambient environment on sleep. These
experiments demonstrate a robust effect of arena shape and size as well
as light levels on sleep duration and architecture, and highlighting the
versatility of tracking-based sleep acquisition. These findings provide
insight into the context-specific basis for choosing between Drosophila
sleep acquisition systems, describe a novel cost-effective system for
video tracking, and characterize sleep analysis using the MB-IR sleep
analysis. Further, the study also describes a modified dark-place
preference sleep assay using video tracking, confirming that flies
prefer to sleep in dark locations (Garbe, 2015). Though evidence is mounting that a major function
of sleep is to maintain brain plasticity and consolidate memory,
little is known about the molecular pathways by which learning and
sleep processes intercept. Anaplastic
lymphoma kinase (Alk), the gene encoding a tyrosine
receptor kinase whose inadvertent activation is the cause of many
cancers, is implicated in synapse formation and cognitive functions. In
particular, Alk genetically interacts with Neurofibromatosis
1 (Nf1) to regulate growth and associative learning in
flies. This study shows that Alk mutants have increased sleep.
Using a targeted RNAi screen the negative effects of Alk on
sleep was localized to the mushroom body, a structure important for both
sleep and memory. Mutations in Nf1 produce a sexually
dimorphic short sleep phenotype, and suppress the long sleep phenotype
of Alk. Thus Alk and Nf1 interact in both
learning and sleep regulation, highlighting a common pathway in these
two processes (Bai, 2015). Though a few studies implicate Alk orthologs in regulating
behaviors such as decision-making, cognition, associative learning and
addiction, most functional studies demonstrate various developmental
roles for Alk. This study acutely induce a long-sleep
phenotype by taking advantage of a temperature-sensitive allele, Alkts,
revealing that Alk regulates sleep directly rather than
through developmental processes. Mutations in Nf1, a gene
encoding a GAP that regulates the Ras/ERK pathway activated by ALK, also
cause a sexually dimorphic short-sleep phenotype. Thus this study
establishes a novel in vivo function for both Alk
and Nf1 and shows they interact with each other to regulate
sleep (Bai, 2015). Many downstream signaling pathways have been proposed for ALK, among
them Ras/ERK, JAK/STAT, PI3K and PLCγ signaling. ERK activation through
another tyrosine receptor kinase Epidermal growth factor receptor (EGFR)
has been linked to increased sleep, while this study shows that Alk,
a positive regulator of ERK, inhibits sleep. It is noted that ERK is a
common signaling pathway targeted by many factors, and may have circuit-
specific effects, with different effects on sleep in different brain
regions. Indeed, neural populations that mediate effects of ERK on sleep
have not been identified. The dose of ALK required for ERK activation
might also differ in different circuits. Region-specific effects of Alk
are supported by a GAL4 screen, in which down-regulation of Alk
in some brain regions even decreased sleep. The overall effect, however,
is to increase sleep, evident from the pan-neuronal knockdown. It was
found that the mushroom body, a site previously implicated in sleep
regulation and learning, requires Alk to inhibit sleep.
Interestingly, the expression patterns of Alk and Nf1
overlap extensively in the mushroom body, suggesting that they may
interact here to regulate both sleep and learning. However, it was
previously shown that Alk activation in the mushroom body has
no effect on learning. The mushroom body expression in that study was
defined with MB247 and c772, both of which also had no effects on sleep
when driving Alk RNAi. The spatial requirement for Nf1
in the context of learning has been disputed in previous studies with
results both for and against a function in the mushroom body. The
discrepancies between these studies could result from: 1) varied
expression of different drivers within lobes of the mushroom body, with
some not even specific to the mushroom body; 2) variability in the
effectiveness and specificity of MB-Gal80 in combination with different
GAL4s. It was confirmed that the MB-Gal80 manipulation eliminated all
mushroom body expression and preserved most if not all other cells with
30Y, 386Y and c309. Future work will further define the cell populations
in which Alk and Nf1 interact to affect sleep (Bai,
2015). A substantial sleep decrease was observed in Nf1 male flies
compared to control flies. However, sleep phenotypes in Nf1
female flies are inconsistent. It is unlikely that unknown mutations on
the X chromosome cause the short-sleeping phenotype because 7 generation
outcrosses into the control iso31 background started with
swapping X chromosomes in Nf1P1 and Nf1P2
male flies with those of iso31 flies. In support of a function
in sleep regulation, restoring Nf1 expression in neurons of Nf1
mutants reverses the short sleep phenotype to long sleep in both males
and females. This does not result from ectopic expression of the
transgene as expressing the same UAS-Nf1 transgene in wild-type
flies has no effect. It was hypothesized that Nf1 promotes
sleep in some brain regions and inhibits it in others, and sub-threshold
levels of Nf1, driven by the transgene in the mutant
background, tilt the balance towards more sleep. As reported in this
study, Alk also has differential effects on sleep in different
brain regions, as does protein kinase A, thus such effects are not
unprecedented. Severe sleep fragmentation was also observed in Nf1
mutants, which suggests that they have trouble maintaining sleep (Bai,
2015). The sex-specific phenotypes of Nf1 mutants may reflect
sexually dimorphic regulation of sleep. A recently published genome-wide
association study of sleep in Drosophila reported that an
overwhelming majority of single nucleotide polymorphisms (SNPs) exhibit
some degree of sexual dimorphism: the effects of ~80% SNPs on sleep are
not equal in the two sexes. Interestingly, sex was found to be a major
determinant of neuronal dysfunction in human NF1 patients and Nf1
knock-out mice, resulting in differential vision loss and learning
deficits. The sex-dimorphic sleep phenotype in Nf1 flies
provides another model to study sex-dimorphic circuits involving Nf1.
Interestingly, a prevalence of sleep disturbances have recently been
reported in NF1 patient, suggesting that NF1 possibly play a conserved
function in sleep regulation (Bai, 2015). An attractive hypothesis for a function of sleep is that plastic
processes during wake lead to a net increase in synaptic strength and
sleep is necessary for synaptic renormalization. There is structural
evidence in Drosophila to support this synaptic homeostasis
hypothesis (SHY): synapse size and number increase during wake and after
sleep deprivation, and decrease after sleep. However, little is known
about the molecular mechanisms by which waking experience induces
changes in plasticity and sleep. FMRP, the protein encoded by the Drosophila
homolog of human fragile X mental retardation gene FMR1, mediates some
of the effects of sleep/wake on synapses. Loss of Fmr1 is
associated with synaptic overgrowth and strengthened neurotransmission
and long sleep. Overexpressing Fmr1 results in dendritic and
axonal underbranching and short sleep. More importantly, overexpression
of Fmr1 in specific circuits eliminates the wake-induced increases in
synapse number and branching in these circuits. Thus, up-regulation of
FMR accomplishes a function normally associated with sleep (Bai, 2015).
It is hypothesized that Alk and Nf1 similarly play
roles in synaptic homeostasis. They are attractive candidates for
bridging sleep and plastic processes, because: 1) Alk is
expressed extensively in the developing and adult CNS synapses. In
particular, both Alk and Nf1 are strongly expressed
in the mushroom body, a major site of plasticity in the fly brain. 2)
Functionally, postsynaptic hyperactivation of Alk negatively
regulates NMJ size and elaboration. In contrast, Nf1 is
required presynaptically at the NMJ to suppress synapse branching. 3) Alk
and Nf1 affect learning in adults and they functionally
interact with each other in this process. It is tempting to speculate
that in Alk mutants, sleep is increased to prune the excess
synaptic growth predicted to occur in these mutants. Such a role for
sleep is consistent with the SHY hypothesis. The SHY model would predict
that Alk flies have higher sleep need, which is expected to
enhance rebound after sleep deprivation. While the data show equivalent
quantity of rebound in Alk mutants, this study found that they
fall asleep faster than control flies the morning after sleep
deprivation, suggesting that they have higher sleep drive. Increased
sleep need following deprivation could also be reflected in greater
cognitive decline, but this has not yet been tested for Alk
mutants. It is noted that Nf1 mutants have reduced sleep
although their NMJ phenotypes also consist of overbranched synapses. It
is postulated that their sleep need is not met and thus results in
learning deficits. Clearly, more work is needed to test these hypotheses
concerning the roles of Alk and Nf1 in sleep,
learning, and memory circuits (Bai, 2015). The fruit fly Drosophila melanogaster is a
diurnal insect active during the day with consolidated sleep at night.
Social interactions between pairs of flies have been shown to affect
locomotor activity patterns, but effects on locomotion and sleep
patterns have not been assessed for larger populations. This study
used a commercially available locomotor activity monitor (LAM25H)
system to record and analyze sleep behavior. Surprisingly, it was
found that same-sex populations of flies synchronize their sleep/wake
activity, resulting in a population sleep pattern, which is similar
but not identical to that of isolated individuals. Like individual
flies, groups of flies show circadian and homeostatic regulation of
sleep, as well as sexual dimorphism in sleep pattern and sensitivity
to starvation and a known sleep-disrupting mutation (amnesiac).
Populations of flies, however, exhibit distinct sleep characteristics
from individuals. Differences in sleep appear to be due to
olfaction-dependent social interactions and change with population size
and sex ratio. These data support the idea that it is possible to
investigate neural mechanisms underlying the effects of population
behaviors on sleep by directly looking at a large number of animals in
laboratory conditions (Liu, 2015). Shared neurophysiologic features between sleep
and anesthetic-induced hypnosis indicate a potential overlap in neuronal
circuitry underlying both states. The authors explored the hypothesis
that propofol anesthesia also dispels sleep pressure in the fruit fly.
Propofol was administered by transferring flies onto food containing
various doses of propofol. High-performance liquid chromatography was
used to measure the tissue concentrations of ingested propofol. To
determine whether propofol anesthesia substitutes for natural sleep, the
flies were subjected to 10-h sleep deprivation (SD), followed by 6-h
propofol exposure, and monitored for subsequent sleep. Oral propofol
treatment was shown to cause anesthesia in flies as indicated by a
dose-dependent reduction in locomotor activity and increased arousal
threshold. Recovery sleep in flies fed propofol after SD was delayed
until after flies had emerged from anesthesia. SD was also associated
with a significant increase in mortality in propofol-fed flies.
Together, these data indicate that fruit flies are effectively
anesthetized by ingestion of propofol and suggest that homologous
molecular and neuronal targets of propofol are conserved in Drosophila.
However, behavioral measurements indicate that propofol anesthesia does
not satisfy the homeostatic need for sleep and may compromise the
restorative properties of sleep (Gardner, 2015). Sleep is thought to be controlled by two main processes:
a circadian clock that primarily regulates sleep timing and a
homeostatic mechanism that detects and responds to sleep need. Whereas
abundant experimental evidence suggests that sleep need increases with
time spent awake, the contributions of different brain arousal systems
have not been assessed independently of each other to determine
whether certain neural circuits, rather than waking per se,
selectively contribute to sleep homeostasis. This study found that
flies sustained thermogenetic activation of three independent
neurotransmitter systems promoted nighttime wakefulness. However, only
sleep deprivation resulting from activation of cholinergic neurons was
sufficient to elicit subsequent homeostatic recovery sleep, as
assessed by multiple behavioral criteria. In contrast, sleep
deprivation resulting from activation of octopaminergic
neurons suppressed homeostatic recovery sleep, indicating that
wakefulness can be dissociated from accrual of sleep need. Neurons that
promote sleep homeostasis were found to innervate the central brain and
motor control regions of the thoracic ganglion. Blocking activity of
these neurons suppressed recovery sleep but did not alter baseline
sleep, further differentiating between neural control of sleep
homeostasis and daily fluctuations in the sleep/wake cycle. Importantly,
selective activation of wake-promoting neurons without engaging the
sleep homeostat impaired subsequent short-term memory, thus providing
evidence that neural circuits that regulate sleep homeostasis are
important for behavioral plasticity. Together, these data suggest a
neural circuit model involving distinct populations of wake-promoting
neurons, some of which are involved in homeostatic control of sleep and
cognition (Seidner, 2015). Drosophila melanogaster's large lateral
ventral neurons (lLNvs) are part of both the circadian
and sleep-arousal neuronal circuits. In the past,
electrophysiological analysis revealed that lLNvs fire action potentials
(APs) in bursting or tonic modes and that the proportion of neurons
firing in those specific patterns varies circadianly. This study
provides evidence that lLNvs fire in bursts both during the day and at
night and that the frequency of bursting is what is modulated in a
circadian fashion. Moreover, lLNvs AP firing is not only under cell
autonomous control, but is also modulated by the network, and in the
process a novel preparation was developed to assess this. lLNv bursting
mode was shown to rely on a cholinergic input because application of
nicotinic acetylcholine receptor antagonists impairs this firing
pattern. Finally, bursting of lLNvs depends was found to depend on an
input from visual circuits that includes the cholinergic L2 monopolar
neurons from the lamina.
This work sheds light on the physiological properties of lLNvs and on a
neuronal circuit that may provide visual information to these important
arousal neurons (Muraro, 2015). Given the relationship between sleep and plasticity, this study
examined the role of Extracellular signal-regulated kinase (ERK,
Rolled in Drosophila) in regulating baseline sleep, and modulating the
response to waking experience. Both sleep deprivation and social
enrichment increase ERK phosphorylation in wild-type flies. The
effects of both sleep deprivation and social enrichment on structural
plasticity in the within the Pigment Dispersing Factor
(PDF)-expressing ventral lateral neurons (LNvs) can be recapitulated
by expressing an active version of ERK (UAS-ERKSEM)
pan-neuronally in the adult fly using GeneSwitch (Gsw) Gsw-elav-GAL4.
Conversely, disrupting ERK reduces sleep and prevents both the
behavioral and structural plasticity normally induced by social
enrichment. Finally, using transgenic flies carrying a cAMP response
Element (CRE)-luciferase reporter it was shown that activating ERK
enhances CRE-Luc activity while disrupting ERK reduces it. These data
suggest that ERK phosphorylation is an important mediator in
transducing waking experience into sleep (Vanderheyden, 2013). ERK plays a key role in regulating not only cell differentiation and
proliferation during development, but is also critical for regulating
long-term potentiation and plasticity related events in the fully
developed adult. Recent studies have highlighted the important
relationship between plasticity induced by waking-experience and sleep
need. With that in mind, it was hypothesized that ERK may provide a
molecular link between plasticity and sleep. Since, ERK
phosphorylation has been previously correlated with sleep time
following rhomboid mediated activation of EGFR, this study
over-expressed an active version of ERK pan-neuronally in the adult
fly and found a significant increase in sleep. ERK activation
increased sleep during the day, was rapidly reversible, and was
associated with increased activity during waking. In contrast,
disrupting ERK signaling by feeding adults the MEK inhibitor SL327
decreased daytime sleep and lowered waking activity. Together these
data indicate ERK activation plays a role in sleep regulation
(Vanderheyden, 2013). As mentioned, inducing EGFR signaling resulted in an increase in
sleep which seemed to correlate nicely with the increase in ppERK.
Although these data strongly suggested that the increase in sleep was
due to ppERK activation, ppERK was not directly manipulated. Thus
these studies confirm and extend data demonstrating that directly
activating ppERK can increase sleep. While the largest effects of
sleep were obtained using a pan neuronal activation of EGFR signaling,
it has also been reported that the EGFR induced increase in sleep
could be mapped to the PI. Surprisingly, this study did not see any
changes in sleep when ppERK was expressed in the PI using the same
GAL4 drivers as a previous study. These data suggest that in the PI,
EGFR activation may recruit additional factors along with ppERK to
alter sleep. Such regulation may be particularly important for
allowing ppERK to carry out multiple functions in various circuits as
needed (Vanderheyden, 2013). In flies, sleep homeostasis is primarily observed during
the subjective day. Similarly, social enrichment also produces
increases in daytime sleep. Thus, the data indicate that modulating
ERK activity in the adult produces a change in sleep during the
portion of the circadian day during which sleep deprivation and social
enrichment modify sleep time. Interestingly, ERK activation not only
increases daytime sleep, but it also results in the proliferation of
terminals in the wake-promoting LNvs. The ability of ERK activation to
increase synaptic terminals is reminiscent of the change in synaptic
markers and structural morphology that are independently observed
following sleep deprivation and social enrichment. Interestingly, arouser
mutants show both enhanced ethanol sensitivity and an increase in
terminals from the LNvs through its activation by EGFR/ERK. Given that a
well characterized function of ERK is to regulate synaptic morphology,
the current results suggest that ERK activation may be a common
mechanism linking waking experience, plasticity and sleep (Vanderheyden,
2013). As mentioned, ERK activation has been correlated with sleep time
following rhomboid mediated activation of EGFR. Interestingly,
in that study, ppERK was not detectable in cell bodies following rho
mediated increases in sleep suggesting that, during rho
activation, ppERK might be modifying sleep at the level of translation
initiation. The current data extend these observations and provide
genetic evidence that ERK activation may also play a role in regulating
sleep and plasticity by activating gene transcription. That is,
pan-neuronal expression of RSK, which retains ERK in the cytoplasm and
prevents its nuclear translocation, results in a decrease in daytime
sleep similar to that observed in wild type flies fed SL327. Although
previous studies have established a link between CREB and sleep this
study evaluated CRE-Luc solely as a reporter of transcriptional
activation. The data indicates that the expression of UAS-ERKSEM
increased CRE-Luc activity. In contrast, transgenic CRE-Luc reporter
flies show reduced bioluminescence when crossed into a rl10a
mutant background. Finally, flies fed SL327 also showed a reduction of
CRE-Luc activity. These data are consistent with a recent report
demonstrating that activating MEK increases bioluminescence in flies
carrying a CRE-Luc reporter. Together these data suggest that ERK
activation may alter plasticity and sleep, in part, by activating gene
transcription (Vanderheyden, 2013). Interestingly, expressing UAS-ERKSEM in
PDF neurons does not change the number of PDF-terminals and does not
alter sleep time. This is in contrast to the effects of expressing UAS-ERKSEM
pan-neuronally which increases both the number of PDF-terminals and
increases sleep. These data suggest that ERK activation can either
influence PDF neurons in a non-cell autonomous fashion or that ERK
activation is required in multiple circuits to modulate plasticity.
Indeed, it has been recently shown that increasing sleep by activating
the dorsal Fan Shaped Body significantly reduces the number of
PDF-terminals. Thus, PDF terminal number provides an accessible read-out
of brain plasticity that can be used to elucidate molecular mechanisms
linking sleep and plasticity at the circuit level (Vanderheyden, 2013).
It is important to note that in flies there is a critical window of
adult development that can influence sleep and learning. For example,
0-3 day old rut2080 mutants are able to respond to
social enrichment with an increase in sleep but their older siblings
(>3days) cannot. In other words, rut2080 mutants
can exhibit higher or lower amounts of sleep as adults depending upon
environmental context, not levels of rutabaga per se. Indeed,
rutabaga mutants have been reported to have significant
variations in sleep (both longer and shorter) compared to controls.
Given that the environment can stably modify sleep during adult
development, even in the absence of memory related genes, care must be
taken when classifying a mutant as either long or short sleepers. It
should be emphasize that the current experiments were designed to avoid
making manipulations during this critical time window to avoid such
confounds. However, it remains possible that ERK may modify sleep by
activating additional downstream targets and/or by regulating
translation initiation at the synapse (Vanderheyden, 2013). Recent studies have shown that waking experience, including both
prolonged wakefulness and exposure to enriched environments,
independently produce dramatic increases in both synaptic markers and
structural morphology throughout the fly brain and that these changes
are reversed during sleep. To date, most studies have evaluated
mutations that disrupt synaptic plasticity to identify the molecular
mechanisms linking sleep with plasticity. Given that ERK is a key
molecule for the regulation of synaptic plasticity and long-term
potentiation, this study evaluated its ability to alter both sleep and
structural plasticity. The data indicate that both sleep deprivation and
social enrichment independently increase ERK phosphorylation in
wild-type flies. It is also reported that expressing an active version
of ERK (UAS-ERKSEM) in the adult fly
results in an increase in sleep and an increase in structural plasticity
in the LNvs. These data suggest that ERK phosphorylation is an important
mediator in transducing waking experience into sleep (Vanderheyden,
2013). Sleep fragmentation, particularly reduced and interrupted night
sleep, impairs the quality of life of older people. Strikingly similar
declines in sleep quality are seen during ageing in laboratory
animals, including the fruit fly Drosophila. This study investigated
whether reduced activity of the nutrient- and stress-sensing
insulin/insulin-like growth factor (IIS)/TOR signalling network, which
ameliorates ageing in diverse organisms, could rescue the sleep
fragmentation of ageing Drosophila. Lowered IIS/TOR network activity
improved sleep quality, with increased night sleep and day activity
and reduced sleep fragmentation. Reduced TOR activity, even when
started for the first time late in life, improved sleep quality. The
effects of reduced IIS/TOR network activity on day and night
phenotypes were mediated through distinct mechanisms: Day activity was
induced by adipokinetic hormone, dFOXO, and enhanced octopaminergic
signalling. In contrast, night sleep duration and consolidation were
dependent on reduced S6K and dopaminergic signalling. These findings
highlight the importance of different IIS/TOR components as potential
therapeutic targets for pharmacological treatment of age-related sleep
fragmentation in humans (Metaxakis, 2014). Sleep syndromes are highly prevalent in elderly humans and, with a
continuing increase in life expectancy and a greater proportion of
elderly people worldwide, effective treatments with fewer side effects
are becoming increasingly needed. Sleep in flies shares striking
similarities with sleep in humans, including an age-related reduction
in sleep quality. This study used Drosophila to examine
age-related sleep pathologies and to suppress these pathologies
through genetic and pharmacological perturbation of insulin/IGF and
TOR signaling (Metaxakis, 2014). This study has shown that the highly conserved IIS pathway, with
roles in growth and development, metabolism, fecundity, stress
resistance, and lifespan, also affects sleep patterns in Drosophila.
Reduced IIS increases and consolidates night sleep, while decreasing
day sleep and inducing day activity. Interestingly, dilp2-3
double mutant flies as well as flies with neuron or fat-body-specific
down-regulation of IIS showed no obvious or only mild sleep phenotypes
in a previous study, suggesting that a strong and/or systemic
reduction in IIS activity may be necessary to induce the activity and
sleep phenotypes. Consistently, dilp2-3 double mutant flies
have very mild growth, lifespan, and metabolic phenotypes compared to
the dilp2-3,5 triple mutant flies used in this study.
Reduced IIS activity resulted in increased sleep consolidation in
young flies. Importantly, reduced IIS ameliorated the age-related
decline in sleep consolidation seen in wild-type flies, thus showing
that it is malleable. Contrary to the increased sleep consolidation
with reduced IIS, high calorie diets have been reported to accelerate
sleep fragmentation. Furthermore, dietary sugar affects sleep pattern
in flies. Taken together, these findings reveal a role of nutrition
and metabolism in sleep regulation and age-related sleep decline in
flies (Metaxakis, 2014). In humans, several studies suggest a link between nutrition and
sleep. The amino acid tryptophan can promote sleep, possibly by
affecting synthesis of the sleep regulators serotonin and melatonin.
Also, the carbohydrate/fat content of the diet seemingly affects sleep
parameters. However, most of these studies are based on correlational
methods and small sample size, and it is not yet clear how diet
affects sleep. Interestingly, sleep duration can affect metabolism,
risk for obesity and diabetes, and even food preference. These
findings associate sleep and metabolism; thus, manipulation of
nutrient-sensing pathways, such as IIS and TOR signalling, may affect
activity and sleep in humans (Metaxakis, 2014). The transcription factor FoxO is an important downstream mediator of
IIS. In C. elegans all aspects of IIS are dependent on daf-16,
the worm ortholog of foxO. In contrast, in Drosophila
IIS-mediated lifespan extension is dependent on dfoxo,
whereas several phenotypes of reduced IIS are dfoxo-independent.
Activity and sleep were unaffected by the loss of dfoxo in
wild-type flies. In contrast, under low IIS conditions loss of dfoxo
specifically affected daytime behaviour, with night time behaviours
unaffected. Reduced IIS therefore affects day and night sleep and
activity through distinct mechanisms. It also uncouples the effects of
IIS on lifespan and on night sleep consolidation, since dfoxo
is essential for extended longevity of flies with reduced IIS. dFOXO
has been previously shown to increase neuronal excitability, possibly
via transcription of ion channel subunits or other (Metaxakis, 2014).
It is suggested that a possible such regulator could be
octopaminergic signalling, known to promote arousal in Drosophila.
Octopamine, the arthropod equivalent of noradrenaline, regulates
several behavioural/physiological processes, including glycogenolysis
and fat metabolism, as well as synaptic and behavioural plasticity.
Moreover, octopamine can affect sleep by acting on insulin-producing
cells in the fly brain, thus linking IIS and sleep/activity. Indeed,
this study found that IIS mutants have increased octopamine levels
and, importantly, pharmacological inhibition of octopaminergic
signalling reverted the increased day activity of IIS mutants.
Noteworthy, mRNA expression of octopamine biosynthetic enzymes was not
changed, but tyramine levels were significantly reduced, suggesting
that increased translation, reduced degradation, or increased activity
of the tyramine-β hydroxylase regulates octopamine levels in IIS
mutants. In contrast to day activity, increased lifespan of IIS
mutants was not affected by pharmacological inhibition of
octopaminergic signalling, thus separating longevity from the day
activity phenotype (Metaxakis, 2014). The effect of reduced IIS on day sleep/activity was mediated through
AKH, the equivalent of human glucagon, an antagonist of. In flies, AKH
coordinates the response to hunger through mobilizing energy stores
and increasing food intake, as well as inducing a starvation-like
hyperactivity. Loss of AKH receptor (AkhR) abrogated the
increased activity of IIS mutants without affecting night sleep. These
results demonstrate that day and night phenotypes of IIS mutants can
be uncoupled, suggesting that the increased night sleep of IIS mutants
is not just a compensatory consequence of increased day activity
(Metaxakis, 2014). dilp2-3,5 mutants have increased octopamine levels, and loss
of AkhR in the dilp2-3,5 mutant background reduced
their octopamine level back to wild-type levels, suggesting that AkhR-mediated
regulation of octopamine controls day hyperactivity in IIS mutants. In
support of these findings, octopaminergic cells mediate the increased
activity effect of AKH in other insects. Flies lacking dFOXO did not
respond to chemically induced AKH release, suggesting that AKH affects
activity through dFOXO. Therefore, it is suggested that dFOXO and AkhR
act through overlapping mechanisms to enhance octopaminergic
signalling and induce activity (Metaxakis, 2014). In flies, AkhR is highly expressed in fat body and its loss
alters lipid and carbohydrate store levels. Therefore, AkhR
might indirectly enhance octopaminergic signalling through alterations
in lipid and carbohydrate metabolism. In support of this idea, lipid
metabolism affects sleep homeostasis in flies. Additionally, AkhR
expression in octopaminergic cells could regulate octopamine synthesis
and release in flies. Interestingly, expression of AkhR is
altered in dfoxo mutants, thus implicating dFOXO in AkhR
regulation. Both are highly expressed in fat body, an important organ
for metabolism in flies, and fat-body-specific insulin receptor may
regulate AkhR function through dFOXO activation (Metaxakis,
2014). In larval motor neurons, dFOXO increases neuronal excitability and
octopamine increases glutamate release, suggesting there is at least a
spatial functional link between the two. Thus, together with a
possible role in AkhR synthesis, dFOXO could act downstream
of octopamine to increase activity (Metaxakis, 2014). To determine the mechanism underlying the IIS-dependent amelioration
of age-related sleep decline, downstream components and genetic
interactors of IIS were investigated. One such interactor that affects
health and ageing is TORC1. TORC1 is a major regulator of translation,
through S6K, 4E-BP, and of autophagy, through ATG1. Inhibiting TOR
signalling, and thus translation, by rapamycin treatment in wild-type
flies recapitulated the sleep features of IIS mutants, even in old
flies. This rescue of sleep quality was blocked by ubiquitous
expression of activated S6K, suggesting that reduced S6K activity is
required for the rescue. These findings, together with previous
results showing S6K to regulate hunger-driven behaviours, highlight
the importance of S6K as a regulator of behaviour in flies. Thus,
manipulating TOR signalling can improve sleep quality through S6K
(Metaxakis, 2014). In mammals, rapamycin treatment has beneficial effects on behaviour
throughout lifespan. Although complete block of TOR activity is
detrimental for long-term memory, a moderate decrease through
rapamycin treatment can improve cognitive function, abrogate
age-related cognitive deterioration, and reduce anxiety and
depression. Moreover, increased TOR activity throughout development is
detrimental for neuronal plasticity and memory. In flies, rapamycin
prevents dopaminergic neuron loss in mutants with parkinsonism.
Although the role of TOR in brain function has not been well studied
in flies, the advantageous effect of rapamycin in both mammalian brain
function and sleep in flies may be mediated through common
neurophysiological mechanisms (Metaxakis, 2014). Gene expression studies have suggested that protein synthesis is
up-regulated during sleep, which may be an essential stage in
macromolecular biosynthesis. Consistent with this, inhibiting protein
synthesis in specific brain domains prolongs sleep duration in
mammals, suggesting that sleep is maintained until specific levels of
biosynthesis occur and aids in explaining the ubiquitously conserved
need for sleep. Brief cycloheximide treatment has been shown to
prolong night sleep and increased consolidation in flies, indicating
an evolutionarily conserved role for protein synthesis inhibition on
sleep regulation. Contrary to reduced IIS, cycloheximide reduced day
activity, possibly due to the global effect of cycloheximide on
protein synthesis or due to toxic defects in flies' physiology.
Decreased protein synthesis rates may enhance the necessity for
increased sleep duration, to allow sufficient synthesis of proteins
and other macromolecules during sleep, allowing organisms to be
healthy and functional during the day (Metaxakis, 2014). Alternatively, the effect of protein synthesis inhibition on night
sleep could be the result of reduced expression of specific sleep
regulators. This study found that DopR1 and dilp2-3,5
mutants share night phenotypes and that rapamycin did not affect sleep
of DopR1 mutants, suggesting that TOR acts on dopaminergic
signalling to affect night sleep. Reduced IIS elevated expression of DopR1,
independently of dFOXO, in accordance with data from mammals. This
effect may be feedback caused by down-regulation of dopaminergic
signalling in IIS mutants, although not through direct regulation of
DopR. Under normal physiological conditions, dopamine signalling is
determined by the level of extracellular dopamine and the rate of
DAT-mediated dopamine clearance from the synaptic cleft. The rate of
dopamine clearance is dependent on the turnover rate of DAT and the
number of functional transporters at the plasma membrane. This study
found that reduced IIS and rapamycin treatment induced increased
expression of DAT, suggesting an increased rate of dopamine clearance
from the synaptic cleft, and thus a reduction in the amplitude of
dopamine signalling, without changes in total dopamine levels. DAT
function and IIS have recently been linked in mammals. DAT function
increases upon insulin stimulation and is diminished on insulin
depletion, through alterations in DAT membrane localization. However,
IIS-dependent regulation of DAT subcellular localization in Drosophila
has not yet been demonstrated. The current data suggest
down-regulating dopaminergic signalling, either by loss of DopR1 or
increasing DAT levels, is beneficial for sleep quality. In agreement
with this it was shown that artificially increasing dopaminergic
signalling, through short-term methamphetamine treatment, increases
both day and night activity and reduces night sleep, and reverts the
beneficial effect of reduced IIS on night behaviours. In mammals,
cocaine administration, which enhances dopaminergic signalling,
increases TOR activity. Also, rapamycin blocks cocaine-induced
locomotor sensitization. Interestingly, cocaine stimulates S6K
phosphorylation in rat brains, and this effect is blocked by
rapamycin. Taken together, these results show that in flies and
mammals dopaminergic and IIS/TOR signalling may interact in similar
ways (Metaxakis, 2014). In conclusion, reduced IIS extends lifespan in
diverse organisms. This study has have shown that it can also
ameliorate age-related sleep fragmentation, but that the mechanisms by
which it does so are distinct from those by which it extends lifespan.
Reduced IIS affected day activity and sleep phenotypes through
increased octopaminergic signalling, but enhanced octopaminergic
signalling did not increase lifespan. Similarly, in Drosophila
increased lifespan from reduced IIS requires dfoxo, but the
night sleep phenotypes of IIS mutants were independent of this
transcription factor. Reduced IIS thus acts through multiple pathways
to ameliorate different aspects of loss of function during ageing. IIS
links metabolism and behaviour through its components, such as S6K and
dFOXO, which act through different neuronal circuits and neurons to
affect sleep. The strong evolutionary conservation of these circuits
and their functions suggests that pharmacological manipulation of IIS
effectors could be beneficial in treatments of sleep syndromes in
humans (Metaxakis, 2014). Mammalian T-type Ca(2+) channels are encoded by three
separate genes (Cav3.1, 3.2, 3.3). These channels are reported to be
sleep stabilizers important in the generation of the delta rhythms of
deep sleep, but controversy remains. The identification of precise
physiological functions for the T-type channels has been hindered, at
least in part, by the potential for compensation between the products
of these three genes and a lack of specific pharmacological
inhibitors. Invertebrates have only one T-type channel gene, but its
functions are even less well-studied.
Ca2+-channel protein α1 subunit T (Ca-alpha1T),
the only Cav3 channel gene in Drosophila melanogaster, was cloned and
expressed in Xenopus oocytes and HEK-293 cells and was confirmed to pass
typical T-type currents. Voltage-clamp analysis revealed the biophysical
properties of Ca-alpha1T show mixed similarity, sometimes falling closer
to Cav3.1, sometimes to Cav3.2, and sometimes to Cav3.3. Ca-alpha1T was
found to be broadly expressed across the adult fly brain in a pattern
vaguely reminiscent of mammalian T-type channels. In addition, flies
lacking Ca-alpha1T show an abnormal increase in sleep duration most
pronounced during subjective day under continuous dark conditions
despite normal oscillations of the circadian clock. Thus, this study
suggests invertebrate T-type Ca(2+) channels promote wakefulness rather
than stabilizing sleep (Jeong, 2015). T-type Ca2+ channels are a subfamily of voltage-dependent Ca2+
channels (VDCCs) that produce low-voltage-activated (LVA) Ca2+
currents implicated in NREM sleep in mammals. Three different genes
encode the pore-forming alpha subunits of mammalian T-type channels, Cav3.1,
3.2, and 3.3. Of these, Cav3.1 and 3.3 are highly expressed
in the thalamus, where the oscillations required for NREM sleep are
generated. Mice lacking Cav3.1 show reduced delta-wave
activity and reduced sleep stability, suggesting that mammalian T-type
currents have a sleep-promoting or stabilizing function (Jeong, 2015). Unlike mammals, Drosophila melanogaster has only one T-type
Ca2+ channel, Ca-α1T, which is also known as DmαG. A
recent study found that motor neurons in flies lacking Ca-α1T show
reduced LVA but also reduced high-voltage-activated (HVA) Ca2+ currents,
suggesting that although Ca-α1T seems to be a genuine T-type channel, it
may have interesting biophysical properties. Therefore a single isoform
of Ca-α1T was cloned, it was expressed in Xenopus oocytes or HEK-293
cells, and its biophysical properties were compared with those of the
rat T-type channel Cav3.1. Several Ca-α1T mutant alleles were generated,
and a defect was generated in their sleep/wake cycles. Contrary to
results in mammals, the fly T-type Ca2+ channel destabilizes sleep. It
is anticipated that these findings will help clarify species-dependent
differences in the in vivo functions of T-type Ca2+ channels,
particularly their role in sleep physiology (Jeong, 2015). Ca-α1T is the largest T-type channel cloned to date, measuring 3205
amino acids. Electrophysiological characterization of Ca-α1T in Xenopus
oocytes showed that Ca-α1T has the hallmark properties of a T-type
channel: low-threshold activation at around -60 mV, a maximal current
output at -20 mV, transient current kinetics elicited by a step-pulse
protocol producing a criss-crossing pattern, and slow deactivation of
tail currents. These biophysical properties are also consistent with
previous studies that implicated Ca-α1T in
low-voltage-activated (LVA) currents in both the central and peripheral
nervous systems of the fly (Jeong, 2015 and references therein). Mammalian genomes contain three T-type Ca2+ channel genes
(i.e., Cav3.1-3.3), while the fly genome contains only one.
Therefore Ca-α1T was measured for some of the characteristics
that distinguish the three mammalian channels. In terms of current
kinetics, Ca-α1T is more similar to mammalian Cav3.1
and Cav3.2 than Cav3.3, which exhibits
considerably slower kinetics. In terms of both its relative permeability
to Ba2+ over Ca2+ and its sensitivity to nickel
inhibition, Ca-α1T is most similar to Cav3.2
(Jeong, 2015). The three mammalian T-type Ca2+ channels, each with their
own distinct biophysical properties, are expressed in largely
complementary patterns of neurons throughout the brain, conferring
considerable functional diversity. Areas of particularly strong
expression include those important for the gating and processing of
sensory inputs, motor control, learning and memory, as well as reward
circuits (Talley, 1999). Using a GFP-tagged knock-in allele, this study
reports that Ca-α1T is expressed broadly across the adult fly
brain in structures reminiscent of the mammalian T-type Ca2+
channels. These include sensory neuropils (i.e., the optic and antennal
lobes, the antennal mechanosensory and motor centers, the anterior
ventrolateral protocerebrum, and the subesophageal zone),
motor-associated neuropils (i.e., the central complex), and those
associated with learning, memory, and reward (i.e., the mushroom
bodies). It is still unclear, however, whether the different isoforms
predicted to originate from the Ca-α1T locus will have
different biophysical properties or different distributions around the
brain (Jeong, 2015). Considering their broad expression, T-type knockout mice appear healthy
and subtle mutant phenotypes emerge only upon close inspection. Sleep,
in particular, has become a focal point in the search for a
physiological function for the T-type channels. Mammalian T-type Ca2+
channels may act as sleep stabilizers and may help generate the burst
firing necessary for the sleep oscillations of deep NREM sleep.
Unfortunately, the three separate mammalian T-type genes all undergo
alternative splicing to produce various channel isoforms that each have
specific biophysical properties, neuroanatomical and subcellular
localizations, and varying abilities to interact with other ion
channels. All these variables and more combine to make it difficult if
not impossible to define a precise physiological role in sleep for
T-type channels as a group. Although Cav3.1 knockout mice
lack the delta oscillations characteristic of deep sleep and show
reduced total sleep, when the knockout is limited to the rostral midline
thalamus, sleep is still reduced, but delta waves are mildly increased.
Another more recent study showed that treatment with the T-type-specific
channel blocker TTA-A2 enhances sleep and delta rhythms in wild type
mice but not Cav3.1/Cav3.3 double knockout mice.
In other words, manipulation of T-type channels can both enhance and
reduce total sleep and deep delta-wave sleep depending on the
experimental context (Jeong, 2015). Although perhaps an underestimate of the actual complexity of the
situation, the subtlety of the phenotypes of the homozygous viable Cav3
mutant mice are often ascribed to functional compensation among the
various Cav3.1-3 isoforms. It was therefore expected that a
behavioral investigation of the sole fly T-type channel, Ca-α1T,
would uncover less subtle sleep phenotypes. It was thus surprising to
find, that despite its broad and relatively strong expression across
adult fly brains, Ca-α1T-null mutants, like the Cav3.1-null
mice, are homozygous viable and lack any overt phenotypes. Upon closer
examination, however, it was observed that Ca-α1T-null mutants
sleep more than controls, especially in constant darkness (Jeong, 2015).
The reason for this relative specificity in the sleep phenotype caused
by Ca-α1T loss-of-function to constant darkness is still
unclear. Flies exhibit a burst of activity upon exposure to the early
morning light but then sleep through most of the rest of the day. Since
control flies show less sleep during subjective daytime under continuous
darkness than under the light phase of light-dark conditions, it is
clear that light exposure can also have sleep-promoting effects. Through
a series of imaging experiments, Although dopamine (DA) is potently
wake-promoting, light exposure can suppress this action of DA at least
partly by causing the up-regulation of the inhibitory DA receptor D2R in
PDF neurons, which are themselves wake-promoting. This modulation of the
wake-promoting PDF neurons by light may help explain why the Ca-α1T
loss-of-function phenotype is biased toward continuous dark conditions
if Ca-α1T functions downstream of the PDF neurons. It would
mean the responsible Ca-α1T-positive neurons are also modulated
by light (Jeong, 2015 and references therein). It was possibe to replicate the increased sleep phenotype of Ca-α1T-null
mutants via pan-neuronal knock-down of Ca-α1T, but it was not
possible to further narrow the cause of this phenotype to a more
specific neuronal subpopulation. This was in spite of numerous attempts
with neuronal Gal4 driver lines ranging from broadly expressed enhancer
traps and neurotransmitter Gal4 drivers to much more narrowly expressed
neuropeptide drivers. This difficulty suggests Ca-α1T may
function in novel sleep circuits (Jeong, 2015). In addition to their sleep phenotype, Ca-α1T-null mutants also
have a circadian phenotype: an elongated circadian period and a
reduction in rhythmic power. It is difficult to say, however, whether
these altered circadian parameters are independent of or secondary to
the sleep phenotype. Rhythmic power is proportional to the magnitude of
the changes in activity level and the regularity with which they occur.
Since the increased sleep observed in the Ca-α1T-null mutants
does reduce the change in overall activity level between subjective day
and subjective night, the increased sleep must also cause a reduction in
rhythmic power (Jeong, 2015). The length of time animals spend sleeping is controlled by both the
circadian clock and by a homeostatic drive to sleep that is proportional
to time spent awake. Thus, most 'sleep mutants' described so far have
had defects in one or the otherthey are either circadian sleep mutants
or homeostatic sleep mutants. After 24 hours of mechanically-induced
sleep deprivation, it was observed that Ca-α1T-null mutants
regain slightly more of their lost sleep than control flies, although
the increase was not statistically significant. This suggests that, in
addition to their circadian phenotype, Ca-α1T-null mutants may
also have a slightly stronger homeostatic drive to sleep than controls.
Although neither the circadian phenotype nor the homeostatic phenotype
are particularly strong, together they produce a robust increase in
sleep (Jeong, 2015). The 'three channel' compensation hypothesis in mice may yet turn out to
be correct, but the current results in flies suggest that other factors
-- isoform-specific differences, differences related to protein-protein
interactions, or even something completely unforeseen -- may allow mice
and flies lacking these broadly expressed and highly conserved ion
channels to still function remarkably well. It will be interesting to
see whether future studies focused on the technically demanding study of
isoform-specific expression patterns and isoform-specific rescues in
both mice and flies will clarify how T-type channels can at various
times and in various contexts both enhance and reduce sleep (Jeong,
2015). Oh, Y., Jang, D., Sonn, J. Y. and Choe, J. (2013). Histamine-HisCl1
receptor axis regulates wake-promoting signals in Drosophila
melanogaster. PLoS One 8: e68269. PubMed ID: 23844178
Histamine and its two receptors, Histamine-gated
chloride channel subunit 1 (HisCl1) and Ora transientless (Ort),
are known to control photoreception and temperature sensing in
Drosophila. However, histamine signaling in the context of neural
circuitry for sleep-wake behaviors has not yet been examined in detail.
Mutant flies were obtained with compromised or enhanced histamine
signaling, and their baseline sleep was tested. Hypomorphic mutations in
histidine
decarboxylase (HDC), an enzyme catalyzing the conversion from
histidine to histamine, caused an increase in sleep duration.
Interestingly, hisCl1 mutants but not ort mutants showed
long-sleep phenotypes similar to those in hdc mutants. Increased
sleep duration in hisCl1 mutants was rescued by overexpressing hisCl1
in circadian pacemaker neurons expressing a neuropeptide pigment
dispersing factor (PDF). Consistently, RNA interference (RNAi)-mediated
depletion of hisCl1 in PDF neurons was sufficient to mimic hisCl1
mutant phenotypes, suggesting that PDF neurons are crucial for sleep
regulation by the histamine-HisCl1 signaling. Finally, either hisCl1
mutation or genetic ablation of PDF neurons dampened wake-promoting
effects of elevated histamine signaling via direct histamine
administration. Taken together, these data clearly demonstrate that the
histamine-HisCl1 receptor axis can activate and maintain the wake state
in Drosophila and that wake-activating signals may travel via the PDF
neurons (Oh, 2013). Using genetic and pharmacological methods to manipulate histamine
signaling, this study shows that the HisCl1 receptor and its downstream
signaling cascade regulate wake-evoking behavior in Drosophila,
while Ort receptor does not show any sleep/wake regulatory function.
Histamine promotes activity via the HisCl1 receptor. Reduced histamine
in HDC mutants or loss of the HisCl1 receptor both show reduced activity
and enhanced sleep. Additionally, the relevant signaling pathway
downstream of the HisCl1 receptor may function in the PDF neurons.
Finally, it was demonstrated that the histamine-HisCl1 receptor axis can
activate and maintain wakefulness in PDF neurons (Oh, 2013). These data show the complete functional segregation of the two
histamine receptors for the first time. Ort receptor is expressed in
large monopolar cells (LMC), postsynaptic to photoreceptors in the
lamina and is a major target of photoreceptor synaptic transmission in Drosophila.
In contrast to Ort, HisCl1 receptor is not expressed in postsynaptic
neurons of photoreceptors. It is expressed in lamina glia and shapes the
LMC postsynaptic response of Ort signaling. Both Ort and HisCl1 receptor
are involved in temperature-preference behaviors, but the major
independent function of HisCl1 receptor remains elusive. This study
showed that sleep regulation is a novel and independent function of
HisCl1 receptor. Additionally, this finding is an important clue in
understanding the functional evolution of the two histamine receptors in
Drosophila (Oh, 2013). It is proposed that wake-activation by histamine signaling in Drosophila
is similar to that found in mammals. hdc mutant flies have
increased sleep durations compared to controls and a previous study
showed that HDC-knockout mice have increased paradoxical sleep compared
to controls. This suggests that the HDC enzyme has a common
wake-promoting function in mammals and flies. However, the structures of
histamine receptors differ between flies and mammals; the histamine
receptors of Drosophila are histamine-gated chloride channels,
whereas the mammalian histamine receptors belong to the rhodopsin-like
G-protein-coupled receptor family. Currently, researchers are working to
identify a metabotropic histamine receptor in Drosophila.
Despite the structural differences of the mammalian and Drosophila
receptors, they share a wake-activating function. This functional
homology may be the result of evolution and provides a hint to find out
the metabotropic histamine receptors in Drosophila (Oh, 2013).
Surprisingly, functional conservations between flies and mammals are
also found among the histamine receptor subtypes. The HisCl1 receptor
has a wake-activating role, whereas the Ort receptor does not. This
result parallels differences in the wake-activation roles of the H1
and H2 receptors in mammals: the H1 receptor can
activate wakefulness, but the H2 receptor cannot. Thus, the
data provide a more detailed understanding of the potential functional
relationship between the HisCl1 and H1 receptors. A
functional connection between the Ort receptor and the H2
receptor is also possible, since the two have little effect on
sleep/wake regulation in their corresponding model systems. No
auto-receptor of histamine has yet been found in Drosophila,
suggesting that there may not be a Drosophila homolog for the
mammalian H3 receptor. Further research should shed greater
light on the evolutionary relationship between the histamine receptors
of flies and mammals (Oh, 2013). Histamine signaling modulates the maintenance of wakefulness and
controls light sensing, and it is speculated that a number of
interactions are possible between these two different pathways. Previous
studies on light-perception mechanisms showed that histamine mutants
exhibit light-sensing defects. However, this study found that the sleep
duration was increased in histamine signaling mutants compared to
wild-type flies in constant darkness. Thus, the perception of light in
the context of evoking wakefulness is independent of vision-related
light perception in Drosophila. Further research will be
required to definitively establish the relationship between light
perception and sleep regulation (Oh, 2013). Previous studies revealed that the PDF neurons promote wakefulness in Drosophila.
The current findings show that histamine signaling acts as a
wake-promoting pathway in PDF neurons. The HisCl1 receptor is a chloride
channel, which would be expected to inhibit the function of the neurons.
However, since previous studies showed that chloride channels can
activate the function of the neurons, hence the HisCl1 receptor might be
an activator of the PDF neurons. The downstream signaling of
histamine-HisCl1 receptor in PDF neurons should be further studied using
genetic manipulation and electro-physiological methods (Oh, 2013). Orexin is a neuropeptide that acts as an important wake-activating
neurotransmitter in mammals, as shown by the demonstration that defects
in orexin synthesis can cause narcoleptic symptoms in human and animals.
Orexin neurons activate wakefulness in the lateral hypothalamic area and
the feedback loop between orexin neurons and monoaminergic neurons such
as histaminergic and serotonergic neurons (tuberomammillary nucleus,
TMN, and dorsal raphe nucleus, DR) controls wakefulness in the
hypothalamus and the brain stem. Histamine receptors are essential for
wake-activation by orexin treatment, indicating that orexin and
histamine signaling constitute an interactive wake-activating system in
mammals. However, orexin has not been found in Drosophila. A
previous study suggested that the PDF neuropeptide functions similar to
those of orexin in Drosophila (Parisky, 2008), potentially
explaining many aspects of the wake-activation cascade in Drosophila.
Histamine and orexin have similar wake-activating function, but
mammalian histamine mutants do not show narcoleptic symptoms. This study
has shown that histamine and one of its receptors, HisCl1, constitute an
important wake-evoking axis in Drosophila. Moreover, it was
demonstrated that histamine-signaling mutants cannot maintain
wakefulness during the daytime, which is similar to the phenotype of
orexin mutants in mammals. Hence, it is proposed that, in Drosophila,
histamine may have a function similar to that of the mammalian orexin.
Further research is required to establish the functional relationship
between wake activation of histamine signaling in Drosophila
and wake-promoting function of orexin and histaminergic system in
mammals (Oh, 2013).
Fruit flies exposed to highly enriched social
environment are found to show increased synaptic connections and a
corresponding increase in sleep. This study investigated if social
environment comprising a pair of same-sex individuals could enhance
sleep in the participating individuals. Same-sex pairs were maintained
for a period of 1 to 4 days, and after separation, sleep of the
previously socialized and solitary individuals was monitored under
similar conditions. Males maintained in pairs for 3 or more days were
found to sleep significantly more during daytime and showed a tendency
to fall asleep sooner as compared to solitary controls (both measures
together are henceforth referred to as "sleep-enhancement").
Sleep-enhancement was found to occur without any significant increase
in wakefulness. Furthermore, while sleep-enhancement due to group-wise
social interaction requires Pigment
Dispersing Factor (PDF) positive neurons; PDF positive and Cryptochrome
(CRY) positive circadian clock neurons and the core circadian clock
genes are not required for sleep-enhancement to occur when males
interact in pairs. Pair-wise social interaction mediated
sleep-enhancement requires dopamine and olfactory signaling, while
visual and gustatory signaling systems seem to be dispensable. These
results suggest that socialization alone (without any change in
wakefulness) is sufficient to cause sleep-enhancement in fruit fly D.
melanogaster males, and that its neuronal control is
context-specific (Lone, 2015). The Drosophila mushroom body (MB) is a key associative memory center that has also been implicated in the control of sleep. However, the identity of MB neurons underlying homeostatic sleep regulation, as well as the types of sleep signals generated by specific classes of MB neurons, has remained poorly understood. We recently identified two MB output neuron (MBON) classes whose axons convey sleep control signals from the MB to converge in the same downstream target region: a cholinergic sleep-promoting MBON class and a glutamatergic wake-promoting MBON class. Here, we deploy a combination of neurogenetic, behavioral, and physiological approaches to identify and mechanistically dissect sleep-controlling circuits of the MB. Our studies reveal the existence of two segregated excitatory synaptic microcircuits that propagate homeostatic sleep information from different populations of intrinsic MB "Kenyon cells" (KCs) to specific sleep-regulating MBONs: sleep-promoting KCs increase sleep by preferentially activating the cholinergic MBONs, while wake-promoting KCs decrease sleep by preferentially activating the glutamatergic MBONs. Importantly, activity of the sleep-promoting MB microcircuit is increased by sleep deprivation and is necessary for homeostatic rebound sleep (i.e., the increased sleep that occurs after, and in compensation for, sleep lost during deprivation). These studies reveal for the first time specific functional connections between subsets of KCs and particular MBONs and establish the identity of synaptic microcircuits underlying transmission of homeostatic sleep signals in the MB (Sitarraman, 2015).
In Drosophila, a crucial component of the machinery for sleep homeostasis is a cluster of neurons innervating the dorsal fan-shaped body (dFB) of the central complex. dFB neurons in sleep-deprived flies tend to be electrically active, with high input resistances and long membrane time constants, while neurons in rested flies tend to be electrically silent. This study demonstrates state switching by dFB neurons, identifies dopamine as a neuromodulator that operates the switch, and delineates the switching mechanism. Arousing dopamine causes transient hyperpolarization of dFB neurons within tens of milliseconds and lasting excitability suppression within minutes. Both effects are transduced by Dop1R2 receptors and mediated by potassium conductances. The switch to electrical silence involves the downregulation of voltage-gated A-type currents carried by Shaker and Shab, and the upregulation of voltage-independent leak currents through a two-pore-domain potassium channel that was termed Sandman. Sandman is encoded by the CG8713 gene and translocates to the plasma membrane in response to dopamine. dFB-restricted interference with the expression of Shaker or Sandman decreases or increases sleep, respectively, by slowing the repetitive discharge of dFB neurons in the ON state or blocking their entry into the OFF state. Biophysical changes in a small population of neurons are thus linked to the control of sleep-wake state (Pimentel, 2016).
Recordings were made from dFB neurons (which were marked by R23E10-GAL4 or R23E10-lexA-driven green fluorescent protein (GFP) expression) while head-fixed flies walked or rested on a spherical treadmill. Because inactivity is a necessary correlate but insufficient proof of sleep, the analysis was restricted to awakening, which is defined as a locomotor bout after >5 min of rest, during which the recorded dFB neuron had been persistently spiking. To deliver wake-promoting signals, the optogenetic actuator CsChrimson was expressed under TH-GAL4 control in the majority of dopaminergic neurons, including the PPL1 and PPM3 clusters, whose fan-shaped body (FB)-projecting members have been implicated in sleep control. Illumination at 630 nm, sustained for 1.5 s to release a bolus of dopamine, effectively stimulated locomotion. dFB neurons paused in successful (but not in unsuccessful) trials, and their membrane potentials dipped by 2-13 mV below the baseline during tonic activity. When flies bearing an undriven CsChrimson transgene were photostimulated, neither physiological nor behavioural changes were apparent. The tight correlation between the suppression of dFB neuron spiking and the initiation of movement might, however, merely mirror a causal dopamine effect elsewhere, as TH-GAL4 labels dopaminergic neurons throughout the brain. Because localized dopamine applications to dFB neuron dendrites similarly caused awakening, this possibility is considered remote (Pimentel, 2016).
Flies with enhanced dopaminergic transmission exhibit a short-sleeping phenotype that requires the presence of a D1-like receptor in dFB neurons, suggesting that dopamine acts directly on these cells. dFB-restricted RNA interference (RNAi) confirmed this notion and pinpointed Dop1R2 as the responsible receptor, a conclusion reinforced by analysis of the mutant Dop1R2MI08664 allele. Previous evidence that Dop1R1, a receptor not involved in regulating baseline sleep, confers responsiveness to dopamine when expressed in the dFB indicates that either D1-like receptor can fulfill the role normally played by Dop1R2. Loss of Dop1R2 increased sleep during the day and the late hours of the night, by prolonging sleep bouts without affecting their frequency. This sleep pattern is consistent with reduced sensitivity to a dopaminergic arousal signal (Pimentel, 2016).
To confirm the identity of the effective transmitter, avoid dopamine release outside the dFB, and reduce the transgene load for subsequent experiments, optogenetic manipulations of the dopaminergic system were replaced with pressure ejections of dopamine onto dFB neuron dendrites. Like optogenetically stimulated secretion, focal application of dopamine hyperpolarized the cells and suppressed their spiking. The inhibitory responses could be blocked at several nodes of an intracellular signalling pathway that connects the activation of dopamine receptors to the opening of potassium conductances: by RNAi-mediated knockdown of Dop1R2; by the inclusion in the patch pipette of pertussis toxin (PTX), which inactivates heterotrimeric G proteins of the Gi/o family; and by replacing intracellular potassium with caesium, which obstructs the pores of G-protein-coupled inward-rectifier channels. Elevating the chloride reversal potential above resting potential left the polarity of the responses unchanged, corroborating that potassium conductances mediate the bulk of dopaminergic inhibition (Pimentel, 2016).
Coupling of Dop1R2 to Gi/o, although documented in a heterologous system, represents a sufficiently unusual transduction mechanism for a predicted D1-like receptor to prompt verification of its behavioural relevance. Like the loss of Dop1R2, temperature-inducible expression of PTX in dFB neurons increased overall sleep time by extending sleep bout length (Pimentel, 2016).
While a single pulse of dopamine transiently hyperpolarized dFB neurons and inhibited their spiking, prolonged dopamine applications (50 ms pulses at 10 Hz, or 20 Hz optogenetic stimulation, both sustained for 2-10 min) switched the cells from electrical excitability (ON) to quiescence (OFF). The switching process required dopamine as well as Dop1R2, but once the switch had been actuated the cells remained in the OFF state-and flies, awake-without a steady supply of transmitter. Input resistances and membrane time constants dropped to 53.3 ± 1.8 and 24.0 ± 1.3% of their initial values (means ± s.e.m.), and depolarizing currents no longer elicited action potentials (15 out of 15 cells). The biophysical properties of single dFB neurons, recorded in the same individual before and after operating the dopamine switch, varied as widely as those in sleep-deprived and rested flies (Pimentel, 2016).
Dopamine-induced changes in input resistance and membrane time constant occurred from similar baselines in all genotypes and followed single-exponential kinetics with time constants of 1.07-1.10 min. The speed of conversion points to post-translational modification and/or translocation of ion channels between intracellular pools and the plasma membrane as the underlying mechanism(s). In 7 out of 15 cases, recordings were held long enough to observe the spontaneous recommencement of spiking, which was accompanied by a rise to baseline of input resistance and membrane time constant, after 7-60 min of quiescence (mean ± s.e.m. = 25.86 ± 7.61 min). The temporary suspension of electrical output is thus part of the normal activity cycle of dFB neurons and not a dead end brought on by the experimental conditions (Pimentel, 2016).
dFB neurons in the ON state expressed two types of potassium current: voltage-dependent A-type (rapidly inactivating) and voltage-independent non-A-type currents. The current-voltage (I-V) relation of iA resembled that of Shaker, the prototypical A-type channel: no current flowed below -50 mV, the approximate voltage threshold of Shaker; above -40 mV, peak currents increased steeply with voltage and inactivated with a time constant of 7.5 ± 2.1 ms (mean ± s.e.m.). Non-A-type currents showed weak outward rectification with a reversal potential of -80 mV, consistent with potassium as the permeant ion, and no inactivation (Pimentel, 2016).
Switching the neurons OFF changed both types of potassium current. iA diminished by one-third, whereas inon-A nearly quadrupled when quantified between resting potential and spike threshold. The weak rectification of inon-A in the ON state vanished in the OFF state, giving way to the linear I-V relationship of an ideal leak conductance. dFB neurons thus upregulate iA in the sleep-promoting ON state. When dopamine switches the cells OFF, voltage-dependent currents are attenuated and leak currents augmented. This seesaw form of regulation should be sensitive to perturbations of the neurons' ion channel inventory: depletion of voltage-gated A-type (KV) channels (which predominate in the ON state) should tip the cells towards the OFF state; conversely, loss of leak channels (which predominate in the OFF state) should favour the ON state. To test these predictions, sleep was examined in flies carrying R23E10-GAL4-driven RNAi transgenes for dFB-restricted interference with individual potassium channel transcripts (Pimentel, 2016).
RNAi-mediated knockdown of two of the five KV channel types of Drosophila (Shaker and Shab) reduced sleep relative to parental controls, while knockdown of the remaining three types had no effect. Biasing the potassium channel repertoire of dFB neurons against A-type conductances thus tilts the neurons' excitable state towards quiescence, causing insomnia, but leaves transient and sustained dopamine responses unaffected. The seemingly counterintuitive conclusion that reducing a potassium current would decrease, not increase, action potential discharge is explained by a requirement for A-type channels in generating repetitive activity of the kind displayed by dFB neurons during sleep. Depleting Shaker from dFB neurons shifted the interspike interval distribution towards longer values, as would be expected if KV channels with slow inactivation kinetics replaced rapidly inactivating Shaker as the principal force opposing the generation of the next spike. These findings identify a potential mechanism for the short-sleeping phenotypes caused by mutations in Shaker, its β subunit Hyperkinetic, or its regulator sleepless (Pimentel, 2016).
Leak conductances are typically formed by two-pore-domain potassium (K2P) channels. dFB-restricted RNAi of one member of the 11-strong family of Drosophila K2P channels, encoded by the CG8713 gene, increased sleep relative to parental controls; interference with the remaining 10 K2P channels had no effect. Recordings from dFB neurons after knockdown of the CG8713 gene product, which this study termed Sandman, revealed undiminished non-A-type currents in the ON state and intact responses to a single pulse of dopamine but a defective OFF switch: during prolonged dopamine applications, inon-A failed to rise, input resistances and membrane time constants remained at their elevated levels, and the neurons continued to fire action potentials (7 out of 7 cells). Blocking vesicle exocytosis in the recorded cell with botulinum neurotoxin C (BoNT/C) similarly disabled the OFF switch. This, combined with the absence of detectable Sandman currents in the ON state, suggests that Sandman is internalized in electrically active cells and recycled to the plasma membrane when dopamine switches the neurons OFF (Pimentel, 2016).
Because dFB neurons lacking Sandman spike persistently even after prolonged dopamine exposure, voltage-gated sodium channels remain functional in the OFF state. The difficulty of driving control cells to action potential threshold in this state must therefore be due to a lengthening of electrotonic distance between sites of current injection and spike generation. This lengthening is an expected consequence of a current leak, which may uncouple the axonal spike generator from somatodendritic synaptic inputs or pacemaker currents when sleep need is low (Pimentel, 2016).
The two kinetically and mechanistically distinct actions of dopamine on dFB neurons-instant, but transient, hyperpolarization and a delayed, but lasting, switch in excitable state-ensure that transitions to vigilance can be both immediate and sustained, providing speedy alarm responses and stable homeostatic control. The key to stability lies in the switching behaviour of dFB neurons, which is driven by dopaminergic input accumulated over time. Unlike bistable neurons, in which two activity regimes coexist for the same set of conductances, dFB neurons switch regimes only when their membrane current densities change. This analysis of how dopamine effects such a change, from activity to silence, has uncovered elements familiar from other modulated systems: simultaneous, antagonistic regulation of multiple conductances; reduction of iA; and modulation of leak currents. Currently little is known about the reverse transition, from silence to activity, except that mutating the Rho-GTPase-activating protein Crossveinless-c locks dFB neurons in the OFF state, resulting in severe insomnia and an inability to correct sleep deficits. Discovering the signals and processes that switch sleep-promoting neurons back ON will hold important clues to the vital function of sleep (Pimentel, 2016).
Food consumption is thought to induce sleepiness. However, little is known
about how postprandial sleep is regulated. This study simultaneously
measured sleep and food intake of
individual flies and found a transient rise in sleep following meals.
Depending on the amount consumed, the effect ranges from slightly arousing
to strongly sleep inducing. Postprandial sleep was positively correlated
with ingested volume, protein, and salt-but not sucrose-revealing meal
property-specific regulation. Silencing of leucokinin receptor (Lkr)
neurons specifically reduces sleep induced by protein consumption.
Thermogenetic stimulation of leucokinin (Lk) neurons decreases whereas Lk
downregulation by RNAi increases postprandial sleep, suggestive of an
inhibitory connection in the Lk-Lkr circuit. A subset of
non-leucokininergic cells proximal to Lkr neurons were identified to
rhythmically increase postprandial sleep when silenced, suggesting that
these cells are cyclically gated inhibitory inputs to Lkr neurons.
Together, these findings reveal the dynamic nature of postprandial sleep (Murphy, 2016). Little is known about the ability of Drosophila circadian
neurons to promote sleep.
This study, using optogenetic manipulation and video recording, shows that
a subset of dorsal clock neurons (DN1s) are potent sleep-promoting cells
that release glutamate to directly inhibit key pacemaker neurons. The
pacemakers promote morning arousal by activating these DN1s, implying that
a late-day feedback circuit drives midday siesta and night-time sleep. To
investigate more plastic aspects of the sleep program, the study used a
calcium assay to monitor and compare the real-time activity of DN1 neurons
in freely behaving males and females. It was found that DN1 neurons are
more active in males than in females, consistent with the finding that
male flies sleep more during the day. DN1 activity is also enhanced by
elevated temperature, consistent with the ability of higher temperatures
to increase sleep. These new approaches indicate that DN1s have a major
effect on the fly sleep-wake profile and integrate environmental
information with the circadian molecular program (Guo, 2016). Lack of quality sleep increases central nervous system oxidative stress and impairs removal of neurotoxic soluble metabolites from brain parenchyma. During aging poor sleep quality, caused by sleep fragmentation, increases central nervous system cellular stress. Currently, it is not known how organisms offset age-related cytotoxic metabolite increases in order to safeguard neuronal survival. Furthermore, it is not understood how age and sleep fragmentation interact to affect oxidative stress protection pathways. This study demonstrates that sleep fragmentation increases systems that protect against oxidative damage and neuroprotective endoplasmic reticulum molecular chaperones, as well as neuronal insulin and dopaminergic expression in middle-aged Drosophila males. Interestingly, even after sleep recovery the expression of these genes was still upregulated in middle-aged flies. Finally, sleep fragmentation generates higher levels of reactive oxygen species (ROS) in middle-aged flies and after sleep recovery these levels remain significantly higher than in young flies. The fact that neuroprotective pathways remain upregulated in middle-aged flies beyond sleep fragmentation suggests it might represent a strong stressor for the brain during later life (Williams, 2016).
In mammals, there is evidence that glutamate has a role as a wake-active neurotransmitter. So using video-based analysis of Drosophila behavior, a study was undertaken to examine if glutamate, which has been previously shown to have an excitatory role in neuromuscular junctions in Drosophila, may have a conserved wake-active role in the adult brain. Using 6- to 9-day-old female flies, the effect was examined of perturbations of the glutamatergic signaling on total wakefulness and wake bout architecture. Neuronal activity of glutamatergic neurons in the brains of adult flies was increased or decreased using Upstream Activating Sequence (UAS) NaChBac (a voltage-gated bacterial Na+ channel) and UAS EKO ('electrical knockout channel'), respectively. Neurotransmission was blocked from glutamatergic neurons in adult flies using the UAS-driven temperature-sensitive dynamin mutation shibirets. The behavior of flies was examined with loss of function mutations of individual subunits of brain-specific ionotropic glutamate receptors. Increasing the activity of glutamatergic neurons in the adult brain led to a significant increase in wakefulness compared to the control groups both in the daytime and nighttime and decreasing the activity of these same neurons reduced wakefulness in the nighttime. Blocking neurotransmitter release in glutamatergic neurons significantly reduced wake in the nighttime. The ionotropic receptor mutants had significantly less wake in the nighttime than their respective genetic background controls. The results show the following: glutamate is indeed a wake-active neurotransmitter in Drosophila; there is a major time of day effect associated with loss of glutamatergic neurotransmission; and it is a major wake-active neurotransmitter in the nighttime (Zimmerman, 2017).
This study developed a system to simultaneously measure sleep and metabolic rate in individual Drosophila, allowing for interrogation of neural systems governing interactions between sleep and metabolic rate. Like mammals, metabolic rate in flies is reduced during sleep and increased during sleep deprivation suggesting sleep-dependent regulation of metabolic rate is conserved across phyla. The reduction of metabolic rate during sleep is not simply a consequence of inactivity because metabolic rate is reduced ~30 minutes following the onset of sleep, raising the possibility that CO2 production provides a metric to distinguish different sleep states in the fruit fly. It was determined that basal and sleep-dependent changes in metabolic rate are reduced in starved flies, suggesting that starvation inhibits normal sleep-associated effects on metabolic rate. Further, translin mutant flies that fail to suppress sleep during starvation demonstrate a lower basal metabolic rate, but this rate was further reduced in response to starvation, revealing that regulation of starvation-induced changes in metabolic rate and sleep duration are genetically distinct. Therefore, this system provides the unique ability to simultaneously measure sleep and oxidative metabolism, providing novel insight into the physiological changes associated with sleep and wakefulness in the fruit fly (Stahl, 2017).
Complex interactions of environmental cues and transcriptional clocks drive rhythmicity in organismal physiology. Light directly affects the circadian clock; however, little is known about its relative role in controlling metabolic variations in vivo. This study used high time-resolution sampling in Drosophila at every 2 h to measure metabolite outputs using a liquid-chromatography tandem mass spectrometry (LC-MS/MS) approach. Over 14% of detected metabolites oscillated with circadian periodicity under light-dark (LD) cycles. Many metabolites peaked shortly after lights-on, suggesting responsiveness to feeding and/or activity rather than the preactivity anticipation, as observed in previous transcriptomics analyses. Roughly 9% of measured metabolites uniquely oscillated under constant darkness (DD), suggesting that metabolite rhythms are associated with the transcriptional clock machinery. Strikingly, metabolome differences between LD and constant darkness were observed only during the light phase, highlighting the importance of photic input. Clock mutant flies exhibited strong 12-h ultradian rhythms, including 4 carbohydrate species with circadian periods in wild-type flies, but lacked 24-h circadian metabolic oscillations. A meta-analysis of these results with previous circadian metabolomics experiments uncovered the possibility of conserved rhythms in amino acids, keto-acids, and sugars across flies, mice, and humans and provides a basis for exploring the chrono-metabolic connection with powerful genetic tools in Drosophila (Rhoades, 2018).
In all animals, sleep pressure is under continuous tight regulation. It is universally accepted that this regulation arises from a two-process model, integrating both a circadian and a homeostatic controller. This study has explored the role of environmental social signals as a third, parallel controller of sleep homeostasis and sleep pressure. It was shown that, in Drosophila melanogaster males, sleep pressure after sleep deprivation can be counteracted by raising their sexual arousal, either by engaging the flies with prolonged courtship activity or merely by exposing them to female pheromones (Beckwith, 2017).
This study provides new evidence that the neuropeptide SIFa and its G protein-coupled receptor SIFR are novel mediators for promoting sleep in Drosophila. Either genetic ablation of SIFa-expressing neurons or depletion of SIFa expression shortened baseline sleep and caused sleep fragmentation by decreasing sleep-bout length. Consistent with these observations, a recent study independently revealed a possible sleep promoting role of SIFa-expressing neurons in DD conditions. Using neuron-specific SIFR depletion, this study further mapped the sleep-promoting SIFR function to Dilp2-negative, SIFR-positive PI neurons (Park 2014).
The PI in adult fly brain is homologous to the mammalian hypothalamus, the control center for neurotransmitter regulation. Several therapeutic targets for human sleep disorders are concentrated in the hypothalamus. For instance, dopaminergic neurons blocked by amphetamine-like drugs induce wake-promoting signals to cure narcolepsy. Benzodiazepine compounds increase gamma-aminobutryic acid (GABA)ergic neuronal transmission to enhance sleep-promoting signals to treat insomnia. Octopamine, which is similar to mammalian norepinephrine, has been identified as a wake-promoting molecule in Drosophila. When octopamine biosynthesis is compromised, flies exhibit enhanced sleep. On the other hand, octopamine promotes wakefulness in flies, particularly at night. Moreover, octopamine and OAMB, an octopamine receptor, act in Dilp2-expressing PI neurons to promote wakefulness through the cyclic AMP (cAMP) pathway. This is in contrast with the current finding that Dilp2-negative PI neurons are important for SIFR-dependent sleep promotion. Therefore, this study has defined a novel PI circuit that promotes sleep via the SIFa-SIFR signaling pathway (Park 2014).
Additional genes have been identified as sleep regulators in the PI region of adult fly brain, including members of the rhomboid family, which are integral membrane proteases and star, a transmembrane cargo receptor. They process epidermal growth factor receptor (EGFR)- activating ligands, such as spitz, gurken, and keren, so that extracellular signal-regulated kinase (ERK) is activated by phosphorylation. When EGFR is activated, flies exhibit excessive sleep. Interestingly, depletion of rhomboid, one of the processors for the ligand of the EGF receptor in c767-Gal4 expressing PI neurons shortened sleep. Given that SIFR and rhomboid promote sleep in the same PI neurons, it might be possible that SIFR and rhomboid function together to regulate sleep through the EGFR-ERK signaling pathway. Not much is known about the SIFR in terms of its downstream effectors and how it exerts its physiological effects. In general, GPCR activates the protein kinase A (PKA)-cAMP pathway via Gs or Ca2+ through a Gq regulator. It was recently shown that lethality in flies with SIFR knock-down is rescued by the overexpression of dSTIM, one of the key regulators of store-operated Ca2+ entry. Furthermore, the nuclear factor of activated T cells (NFAT), a Ca2+-activated transcription factor, is regulated by SIFR in a Schneider 2 (S2) cell-based dsRNA screening, suggesting that sleep regulation by SIFR might involve Ca2+ signaling. Future studies will address which signaling pathways SIFR affects to regulate neuronal activity and sleep behavior (Park, 2014).
Individuals frequently find themselves confronted
with a variety of challenges that threaten their wellbeing. While some
individuals face these challenges efficiently and thrive (resilient)
others are unable to cope and may suffer persistent consequences
(vulnerable). Resilience/vulnerability to sleep disruption may
contribute to the vulnerability of individuals exposed to challenging
conditions. With that in mind this study exploited individual
differences in a fly's ability to form short-term memory (STM)
following 3 different types of sleep disruption to identify the
underlying genes. The analysis showed that in each category of flies
examined, there are individuals that form STM in the face of sleep
loss (resilient) while other individuals show dramatic declines in
cognitive behavior (vulnerable). Molecular genetic studies revealed
that Antimicrobial Peptides, factors important for innate
immunity, were candidates for conferring resilience/vulnerability
to sleep deprivation. Specifically, Metchnikowin
(Mtk), drosocin
(dro) and Attacin
(Att) transcript levels seemed to be differentially increased by sleep
deprivation in glia
(Mtk), neurons (dro) or primarily in the head fat
body (Att). Follow-up genetic studies confirmed that expressing Mtk
in glia but not neurons, and expressing dro in neurons but not
glia, disrupted memory while modulating sleep in opposite directions.
These data indicate that various factors within glia or neurons can
contribute to individual differences in resilience/vulnerability to
sleep deprivation (Dissel, 2015).
Juvenile hormone (JH) is one of the most important hormones in insects since it is essential for insect development. The mechanism by which JH affects the central nervous system still remains a mystery. This study demonstrates that one of the JH receptors, Methoprene-tolerant (Met), is important for the control of neurite development and sleep behavior in Drosophila. With the identification of Met-expressing glial cells, the mechanism that Met negatively controls the mushroom body (MB) β lobes fusion and positively maintains pigment-dispersing factor sLNvs projection pruning has been established. Furthermore, despite the developmental effects, Met can also maintain nighttime sleep in a development-independent manner through the α/β lobe of MB. Combining analyses of neuronal morphology and entomological behavior, this study advances understanding of how the JH receptor regulates the nervous system (Wu, 2021).
Phase response curves (PRCs) for light or temperature stimuli have been shown to be most valuable in understanding how circadian clocks are entrained to daily environmental cycles. Nowadays, PRC experiments in which clock neurons are manipulated in a temporally restricted manner by thermogenetic or optogenetic tools are also useful to comprehend clock network properties. In this study, specific clock neurons of Drosophila melanogaster were temporally depolarized by activating temperature-sensitive dTrpA1 channels to unravel their role in phase shifting the flies' activity rhythm. The depolarization of all clock neurons causes a PRC resembling the flies' light PRC, with strong phase delays in the first half of the subjective night and modest phase advances in its second half. However, the activation of the flies' pigment-dispersing factor (PDF)-positive morning (M) neurons (s-LNvs) only induces phase advances, and these reach into the subjective day, where the light PRC has its dead zone. This indicates that the M neurons are very potent in accelerating the clock, which is in line with previous observations. In contrast, the evening (E) neurons together with the PDF-positive l-LNvs appear to mediate phase delays. Most interestingly, the molecular clock (Period protein cycling) of the depolarized clock neurons is shifted in parallel to the behavior, and this shift is already visible within the first cycle after the temperature pulse. The cAMP response element binding protein B (CREB) was identified as a putative link between membrane depolarization and the molecular clock (Eck, 2016).
Biological clocks allow organisms to anticipate daily environmental changes such as temperature fluctuations, abundance of daylight, and nutrient availability. Many circadian-controlled physiological states are coordinated by the release of systemically acting hormones, including steroids and insulin. Thus, hormones relay circadian outputs to target tissues, and disrupting these endocrine rhythms impairs human health by affecting sleep patterns, energy homeostasis, and immune functions. It is largely unclear, however, whether circadian circuits control hormone levels indirectly via central timekeeping neurons or whether peripheral endocrine clocks can modulate hormone synthesis directly. This study shows that perturbing the circadian clock, specifically in the major steroid hormone-producing gland of Drosophila, the prothoracic gland (PG), unexpectedly blocks larval development due to an inability to produce sufficient steroids. This is surprising, because classic circadian null mutants are viable and result in arrhythmic adults. Timeless and Period, both core components of the insect clock, are required for transcriptional upregulation of steroid hormone-producing enzymes. Timeless couples the circadian machinery directly to the two canonical pathways that regulate steroid synthesis in insects, insulin and PTTH signaling, respectively. Activating insulin signaling directly modulates Timeless function, suggesting that the local clock in the PG is normally synced with systemic insulin cues. Because both PTTH and systemic insulin signaling are themselves under circadian control, it is concluded that de-synchronization of a local endocrine clock with external circadian cues is the primary cause for steroid production to fail (Di Cara, 2016).
In animals, networks of clock neurons containing molecular clocks
orchestrate daily rhythms in physiology and behavior. However, how various
types of clock neurons communicate and coordinate with one another to
produce coherent circadian rhythms is not well understood. This study
investigated clock neuron
coupling in the brain of Drosophila
and demonstrates that the fly's various groups of clock neurons display
unique and complex coupling relationships to core pacemaker neurons.
Furthermore, coordinated free-running rhythms require molecular clock
synchrony not only within the well-characterized lateral clock neuron
classes but also between lateral clock neurons and dorsal clock neurons.
These results uncover unexpected patterns of coupling in the clock neuron
network and reveal that robust free-running behavioral rhythms require a
coherence of molecular oscillations across most of the fly's clock neuron
network (Yao, 2016). There are no general methods for reliably assessing the firing properties or even calcium profiles of specific neurons in freely moving flies. To this end, this study adapted a GFP-based calcium reporter to luciferase that was expressed in small subsets of circadian neurons. This Tric-LUC reporter allowed a direct comparison of luciferase activity with locomotor activity, which was assayed in the same flies with video recording. The LUC profile from activity-promoting E cells paralleled evening locomotor activity, and the LUC profile from sleep-promoting glutamatergic DN1s (gDN1s) paralleled daytime sleep. Similar profiles were generated by novel reporters recently identified based on transcription factor activation. As E cell and gDN1 activity is necessary and sufficient for normal evening locomotor activity and daytime sleep profiles, respectively, it is suggest that their luciferase profiles reflect their neuronal calcium and in some cases firing profiles in wake-behaving flies (Guo, 2017).
Ventral Lateral Neurons (LNvs), which are essential in the control of rest-activity cycles in Drosophila, undergo circadian remodeling of their axonal projections. This structural plasticity gives rise to changes in the degree of connectivity, which could provide a means of transmitting time of day information. Several neuronal types undergoing circadian remodeling hint to a differential effect of clock genes; while per mutants exhibited poorly developed axonal terminals giving rise to low complexity arbors, tim mutants displayed a characteristic hyper branching phenotype, suggesting these genes could be playing additional roles to those ascribed to core clock function. To shed light onto this possibility clock gene levels were altered through RNAi- mediated downregulation and expression of a dominant negative form exclusively in the adult LNvs. These experiments confirmed that the LNv clock is necessary to drive the remodeling process. The contribution of glia to the structural plasticity of the small LNvs was explored through acute disruption of their internal clock. Interestingly, impaired glial clocks also abolished circadian structural remodeling, without affecting other clock-controlled outputs. Taken together these data shows that both neuronal and glial clocks are recruited to define the architecture of the LNv projections along the day, thus enabling a precise reconfiguration of the circadian network (Herrero, 2017).
Sleep is a dynamic process comprising multiple stages, each associated with distinct electrophysiological properties and potentially serving different functions. While these phenomena are well described in vertebrates, it is unclear if invertebrates have distinct sleep stages. Local field potential (LFP) recordings were performed on flies spontaneously sleeping, and their brain activity was compared to flies induced to sleep using either genetic activation of sleep-promoting circuitry or the GABAA agonist Gaboxadol. A transitional sleep stage was found associated with a 7-10 Hz oscillation in the central brain during spontaneous sleep. Oscillatory activity is also evident when sleep-promoting neurons were acutely activated in the dorsal fan-shaped body (dFB) of Drosophila. In contrast, sleep following Gaboxadol exposure is characterized by low-amplitude LFPs, during which dFB-induced effects are suppressed. Sleep in flies thus appears to involve at least two distinct stages: increased oscillatory activity, particularly during sleep induction, followed by desynchronized or decreased brain activity (Yap, 2017).
Circadian clocks ensure that behavioral and physiological processes occur at optimal times of day and in the correct temporal order. It is becoming increasingly clear that chronic circadian misalignment (CCM), such as occurs in shift workers, has profound metabolic and cognitive consequences, but the proximate mechanisms connecting CCM with reduced organismal health are unknown. This study investigated the consequences of CCM in the powerful model system of the fruit fly, Drosophila melanogaster. Flies were subjected to daily 4-h phase delays in the light-dark schedule, and the Drosophila Activity Monitoring (DAM) system was used to continuously track locomotor activity and sleep while simultaneously monitoring fly lifespan. Consistent with previous results, exposing flies to CCM leads to a ~ 15% reduction in median lifespan in both male and female flies. Importantly, it was demonstrated that the reduced longevity occurs independent of changes in overall sleep or activity. To uncover potential molecular mechanisms of CCM-induced reduction in lifespan, whole body RNA-sequencing was conducted to assess differences in gene transcription between control and misaligned flies. CCM caused progressive, large-scale changes in gene expression characterized by upregulation of genes involved in response to toxic substances, aging and oxidative stress, and downregulation of genes involved in regulation of development and differentiation, gene expression and biosynthesis. Many of these gene expression changes mimic those that occur during natural aging, consistent with the idea that CCM results in premature organismal decline, however, genes involved in lipid metabolism are overrepresented among those that are differentially regulated by CCM and aging. This category of genes is also among the earliest to exhibit CCM-induced changes in expression, thus highlighting altered lipid metabolism as a potentially important mediator of the negative health consequences of CCM (Boomgarden, 2019).
Sleep appears to be a universally conserved phenomenon among the animal kingdom, but whether this notable evolutionary conservation underlies a basic vital function is still an open question. Using a machine learning-based video-tracking technology, a detailed high-throughput analysis of sleep was conducted in the fruit fly Drosophila melanogaster, coupled with a lifelong chronic and specific sleep restriction. These results show that some wild-type flies are virtually sleepless in baseline conditions and that complete, forced sleep restriction is not necessarily a lethal treatment in wild-type D. melanogaster. It was also shown that circadian drive, and not homeostatic regulation, is the main contributor to sleep pressure in flies. These results offer a new perspective on the biological role of sleep in Drosophila and, potentially, in other species (Geissmann, 2019).
Insomnia is the most common sleep disorder among adults, especially affecting individuals of advanced age or with neurodegenerative disease. Insomnia is also a common comorbidity across psychiatric disorders. Cognitive behavioral therapy for insomnia (CBT-I) is the first-line treatment for insomnia; a key component of this intervention is restriction of sleep opportunity, which optimizes matching of sleep ability and opportunity, leading to enhanced sleep drive. Despite the well-documented efficacy of CBT-I, little is known regarding how CBT-I works at a cellular and molecular level to improve sleep, due in large part to an absence of experimentally-tractable animals models of this intervention. Guided by human behavioral sleep therapies, this study developed a Drosophila model for sleep restriction therapy (SRT) of insomnia. It was demonstrated that restriction of sleep opportunity through manipulation of environmental cues improves sleep efficiency in multiple short-sleeping Drosophila mutants. The response to sleep opportunity restriction requires ongoing environmental inputs, but is independent of the molecular circadian clock. This sleep opportunity restriction paradigm was applied to aging and Alzheimer's disease fly models; sleep impairments in these models are reversible with sleep restriction, with associated improvement in reproductive fitness and extended lifespan. This work establishes a model to investigate the neurobiological basis of CBT-I, and provides a platform that can be exploited toward novel treatment targets for insomnia (Belfer, 2019).
One output arm of the sleep homeostat in Drosophila appears to be a group of neurons with projections to the dorsal fan-shaped body (dFB neurons) of the central complex in the brain. However, neurons that regulate the sleep homeostat remain poorly understood. Using neurogenetic approaches combined with Ca(2+) imaging, synaptic connections were characterized between dFB neurons and distinct sets of upstream sleep-regulatory neurons. One group of the sleep-promoting upstream neurons is a set of circadian pacemaker neurons that activates dFB neurons via direct glutaminergic excitatory synaptic connections. Opposing this population, a group of arousal-promoting neurons downregulates dFB axonal output with dopamine. Co-activating these two inputs leads to frequent shifts between sleep and wake states. dFB neurons were shown to release the neurotransmitter GABA and inhibit octopaminergic arousal neurons. It is proposed that dFB neurons integrate synaptic inputs from distinct sets of upstream sleep-promoting circadian clock neurons, and arousal neurons (Ni, 2019).
Slow-wave rhythms characteristic of deep sleep oscillate in the delta band (0.5-Hz) and can be found across various brain regions in vertebrates. Across phyla, however, an understanding of the mechanisms underlying oscillations and how these link to behavior remains limited. This study discovered compound delta oscillations in the sleep-regulating R network of Drosophila. The power of these slow-wave oscillations increases with sleep need and is subject to diurnal variation. Optical multi-unit voltage recordings reveal that single R neurons get synchronized by activating circadian input pathways. This synchronization depends on NMDA receptor (NMDAR) coincidence detector function, and an interplay of cholinergic and glutamatergic inputs regulates oscillatory frequency. Genetically targeting the coincidence detector function of NMDARs in R5, and thus the uncovered mechanism underlying synchronization, abolished network-specific compound slow-wave oscillations. It also disrupted sleep and facilitated light-induced wakening, establishing a role for slow-wave oscillations in regulating sleep and sensory gating. It is therefore proposed that the synchronization-based increase in oscillatory power likely represents an evolutionarily conserved, potentially 'optimal,' strategy for constructing sleep-regulating sensory gates (Raccuglia, 2019).
In vertebrates, oscillatory electrical compound patterns are associated with fundamental brain functions and specific behaviors. Characteristic of vertebrate deep sleep are compound slow-wave oscillations in the delta band (0.5- Hz), which are thought to derive from the synchronization of neuronal activity. However, how specific neural networks contribute to generating compound oscillations and whether these oscillations represent a functional unit for sleep regulation largely remains unclear. This is partially due to methodological constraints, as readouts either focus on single-cell/unit or local compound potential recordings (electroencephalograms and local field potentials) that, confined by poor spatial resolution, do not permit for dissecting interactions of genetically identified multi-units (Raccuglia, 2019).
Like vertebrates, invertebrates' sleep and behavior selection is sensitive to an animal's sleep need. However, in invertebrates, it is unknown whether electrical oscillations can gate specific behaviors and whether an electrophysiological sleep correlate, such as slow-wave oscillations, exists or is involved in sleep regulation. Local field potential (LFP) measurements in the Drosophila brain indicate that the frequency of large-scale compound neuronal activity is reduced during sleep, opening up the possibility that, comparable to vertebrates, slow oscillatory activity could be involved in mediating sleep. Yet such oscillations have not been identified, and it remains unknown which and how neural networks would generate slow-wave oscillations that could be crucial for sleep regulation (Raccuglia, 2019).
This study made use of recent technological advancements in Drosophila melanogaster and combined targeted expression of a genetically encoded voltage indicator (GEVI) with that of optogenetic actuators. This all-optical electrophysiological approach bypasses common methodological constraints, allowing monitoring of multi-unit electrical patterns within a specific network and gaining of mechanistic insight into how sleep-relevant neural activity might be generated (Raccuglia, 2019).
This study has discovered sleep-regulating, network-specific delta oscillations within the R network of the Drosophila ellipsoid body, which is situated at a crossroad involved in sleep regulation and sensory processing. Compound delta oscillations are not detectable in the morning (Zeitgeber time [ZT] 0-3) but become apparent, and increase in power, over the day. Activating circadian input pathways leads to multi-unit synchronization that depends on NMDA receptor (NMDAR)-mediated coincidence detection. Disrupting this synchronization and thus the emergence of compound delta oscillations affects the flies' sleep patterns and alters sensory gating during sleep. This study thus identified delta oscillations as an electrophysiological correlate of sleep pressure regulation in invertebrates and ties these oscillatory patterns to behavioral readouts. This work suggests that slow-wave oscillations and sleep could be fundamentally interconnected across phyla. Slow-wave oscillations may therefore potentially represent an evolutionarily conserved strategy for network mechanisms regulating internal states and sleep (Raccuglia, 2019).
The data suggest that compound delta oscillations specific to the sleep-regulating R network are generated by circadian drive transduced via TuBu neurons. Optogenetic activation of TuBu neurons increases single-unit power and synchronizes R neurons, which should result in an increase of compound delta power and thus internal sleep drive. This is consistent with thermogenetic activation of TuBu neurons increasing the total amount of sleep in flies. High levels of TuBu neuron output could be generated by altering activity in sleep-modulating DN circadian clock neurons, which form direct connections with the TuBu neurons. Of note, following R synchronization mediated via activation of TuBu neurons, a prolonged hyperpolarization becomes apparent that typically can be observed following highly synchronized states (Raccuglia, 2019).
The R network also receives excitatory input from Helicon cells, a potential source of cholinergic input. This study provides evidence that nAChRs act as prime candidates to provide concurrent depolarization required for NMDAR coincidence detection in R neurons. Analogous to the relationship between AMPARs and NMDARs in mammals, the interplay of nAChRs and NMDARs could provide a substrate for coincidence detection in Drosophila. By reducing the temporal variability of mixed synaptic inputs, NMDARs could promote the temporal summation of excitatory postsynaptic potentials, thus increasing the synchrony between R neurons. Indeed, rhythmic application of acetylcholine to R dendrites is sufficient to induce oscillations but only in the presence of functional NMDARs. Helicon cells also receive visual information and are part of a recurrent circuit mediating homeostatic sleep pressure regulation. Thus, R oscillatory activity is likely regulated via a complex interplay of sensory input, circadian rhythms, and homeostatic sleep pressure regulation (Raccuglia, 2019).
These experiments further indicate that additional inhibitory input via glutamate-gated chloride channels could antagonize excitatory drives to retain a frequency within the delta range, potentially defining the time window for recurrent input. It is therefore proposed that the interplay between glutamatergic and cholinergic signals sets a frequency within the delta range (0.5-1. Hz), which permits temporal synchronization of single units to define the power and frequency of compound oscillatory activity and thus the strength of sleep drive (Raccuglia, 2019).
Expression levels of NMDARs in R neurons have previously been associated with the regulation of sleep drive. However, the physiological contribution of NMDARs remained open. The current data suggest that NMDAR coincidence detection gates neuronal synchronization of delta-wave activity within the R network to increase the power of sleep-relevant compound oscillations. Indeed, at the single-cell level, expressing Mg2+ block-deficient NMDARs in R neurons led to irregular activity patterns, which could be at the basis of impaired synchronization and disrupted compound oscillations. It is therefore hypothesized that NMDAR-coincidence detection can lead to a state-dependent transition of activity patterns permitting synchronization. Disrupting synchronization by expressing Mg2+-block-deficient NMDARs interfered with multi-unit synchronization, directly altering the animals' sleep drive, sleep quality, and stimulus-induced wakening. This tightly links the mechanisms underlying network-specific delta-band oscillations to shaping animal behavior (Raccuglia, 2019).
This study has identified slow-wave oscillations as an electrophysiological correlate of sleep pressure regulation involved in sensory gating in Drosophila. Interestingly, oscillatory activity in Drosophila R neurons is reminiscent of up- and down-states occurring at the level of mammalian cortical networks during deep sleep. It is thus hypothesized that the oscillations observed in this study are comparable to sleep-regulating thalamocortical oscillations as well as network-specific oscillations observed during sleep deprivation in vertebrates (local sleep). Thus, the R network could be functionally analogous to the thalamus, as network-specific synchronization of slow-wave activity within the thalamus plays a crucial role in maintaining sleep and sensory gating. Comparable to the potential function of network-specific oscillations during 'local sleep' in vertebrates, slow-wave oscillations within the flies' R network may well be involved in the homeostatic regulation of synaptic strength. It is thus suggested that oscillatory network synchronization may represent an evolutionarily selected optimal strategy for signaling sleep pressure as well as for the internal representation of sleepiness. One prediction from this analogy would be that slow-wave oscillations would also be detected in other Drosophila brain regions. Indeed, extracellular local field potential recordings suggest that overall activity in the fly brain does change during sleep. Future work should address whether network oscillations in the delta band can also be detected in other brain regions, for instance, the sleep-inducing fan-shaped bodies or the mushroom bodies. Additionally, whether cell-autonomous conductances contribute to sustained rhythmic activities of single R neuron remains an open question. This framework should pave the road for identifying evolutionarily conserved fundamental principles that link slow-wave oscillations as electrophysiological hallmarks of sleep to the neuronal processes underlying memory consolidation (Raccuglia, 2019).
Consumption of foods that are high in fat contribute to obesity and metabolism-related disorders. Dietary lipids are comprised of triglycerides and fatty acids, and the highly palatable taste of dietary fatty acids promotes food consumption, activates reward centers in mammals, and underlies hedonic feeding. Despite the central role of dietary fats in the regulation of food intake and the etiology of metabolic diseases, little is known about how fat consumption regulates sleep. The fruit fly, Drosophila melanogaster, provides a powerful model system for the study of sleep and metabolic traits, and flies potently regulate sleep in accordance with food availability. To investigate the effects of dietary fats on sleep regulation, fatty acids were supplemented into the diet of Drosophila and their effects on sleep and activity were measured. Flies fed a diet of hexanoic acid, a medium-chain fatty acid that is a by-product of yeast fermentation, slept more than flies starved on an agar diet. To assess whether dietary fatty acids regulate sleep through the taste system, sleep was assessed in flies with a mutation in the hexanoic acid receptor Ionotropic receptor 56D, which is required for fatty acid taste perception. These flies also sleep more than agar-fed flies when fed a hexanoic acid diet, suggesting the sleep promoting effect of hexanoic acid is not dependent on sensory perception. Taken together, these findings provide a platform to investigate the molecular and neural basis for fatty acid-dependent modulation of sleep (Sah Pamboro, 2019).
Sleep is essential but incompatible with other behaviors, and thus sleep drive competes with other motivations. Previous work has shown Drosophila males balance sleep and courtship via octopaminergic neurons that act upstream of courtship-regulating P1 neurons. This study shows that nutrition modulates the sleep-courtship balance and identify sleep-regulatory neurons downstream of P1 neurons. Yeast-deprived males exhibited attenuated female-induced nighttime sleep loss yet normal daytime courtship, which suggests male flies consider nutritional status in deciding whether the potential benefit of pursuing female partners outweighs the cost of losing sleep. Trans-synaptic tracing and calcium imaging identified dopaminergic neurons projecting to the protocerebral bridge (DA-PB) as postsynaptic partners of P1 neurons. Activation of DA-PB neurons led to reduced sleep in normally fed but not yeast-deprived males. Additional PB-projecting neurons regulated male sleep, suggesting several groups of PB-projecting neurons act downstream of P1 neurons to mediate nutritional modulation of the sleep-courtship balance (Duhart, 2020b).
Regulation of circadian behavior and physiology by the Drosophila brain clock requires communication from central clock neurons to downstream output regions, but the mechanism by which clock cells regulate downstream targets is not known. This study shows that the pars intercerebralis (PI), previously identified as a target of the morning cells in the clock network, also receives input from evening cells. It was determined that morning and evening clock neurons have time-of-day-dependent connectivity to the PI, which is regulated by specific peptides as well as by fast neurotransmitters. Interestingly, PI cells that secrete the peptide DH44, and control rest:activity rhythms, are inhibited by clock inputs while insulin-producing cells (IPCs) are activated, indicating that the same clock cells can use different mechanisms to drive cycling in output neurons. Inputs of morning cells to IPCs are relevant for the circadian rhythm of feeding, reinforcing the role of the PI as a circadian relay that controls multiple behavioral outputs. These findings provide mechanisms by which clock neurons signal to nonclock cells to drive rhythms of behavior (Barber, 2021).
Many physiological and behavioral processes across organisms exhibit daily rhythms controlled by an internal circadian system. The temporal organization conferred by circadian clocks allows anticipation of environmental changes and the coordination of biochemical and physiological processes within and across cells and tissues. Work in Drosophila and other organisms has elucidated the molecular basis of circadian pacemaking in the brain, but a molecular understanding of how time-of-day information is relayed from brain clock circuitry to downstream output regions that regulate circadian physiology and behavior is still lacking (Barber, 2021).
The Drosophila clock network consists of ~150 neurons that express the core clock transcription-translation feedback loop genes (1). Of these, the ventrolateral neurons (LNvs) and dorsal lateral (LNd) clock neurons are important pacemakers for driving circadian rhythms in constant darkness, and coordinating the activity of other neurons in the clock network. Indeed, robust rhythms are an emergent property of the clock network as a whole. The rhythmic pattern of locomotor activity in light:dark (LD) cycles, which consists of morning and evening peaks that anticipate dawn and dusk, respectively, is also attributed to specific clock cells. The LNvs act through the neuropeptide pigment-dispersing factor (PDF) to drive the morning bout of locomotor activity while LNd clock neurons are termed evening neurons due to their role in regulating evening anticipatory activity. LNd neurons are a molecularly heterogeneous population with subpopulations expressing different signaling molecules including the classical transmitter acetylcholine and the neuropeptides Ion transport peptide (ITP), NPF, and sNPF DN1 dorsal neurons, which receive input from LNvs, also promote morning activity and exhibit highest activity in the predawn and morning hours. DN1 neurons express glutamate and the neuropeptide DH31, both of which have previously described roles in regulating circadian rhythms. The diverse array of signaling molecules expressed by neurons in the clock network offers many possibilities for signaling to clock output brain regions that lack their own clocks but are important for the generation of behavioral rhythm (Barber, 2021).
A major clock output region in Drosophila is the pars intercerebralis (PI), a protohypothalamic region implicated in regulating circadian locomotor rhythms and peripheral cycling. Like the hypothalamus, the PI controls aspects of sleep and feeding and comprises a population of neurosecretory cells with heterogeneous peptide expression. PI neurons that express the neuropeptide diuretic hormone 44 (DH44), the fly ortholog of corticotropin-releasing factor, modulate rest:activity rhythms. On the other hand, PI neurons producing Drosophila insulin-like peptides are implicated in circadian gene expression in the fat body, but have not been linked to behavioral rhythms (Barber, 2021).
As PI cells do not express the molecular clock machinery, time-of-day information must be relayed directly or indirectly from clock neurons to the PI. Previous work showed that the DN1 clock neurons project to two groups of PI neurons, the diuretic hormone 44 positive (DH44+) cells and the insulin-producing cells (IPCs). However, while hyperactivation of DH44+ neurons or RNAi knockdown of DH44 peptide significantly dampens rest:activity rhythms in Drosophila, silencing of DN1 neurons has a considerably weaker effect. Thus, there must be additional upstream inputs to PI neurons that maintain rest:activity rhythms in the absence of DN1 signals. It was hypothesized that robust circadian control of the PI would require inputs from both morning- and evening-active clock neurons, including perhaps long-distance signals directly from LNvs. This study sought to determine if evening cells project to the PI and also investigated clock-PI signaling at different times of day. It was demonstrated that both DH44+ neurons and IPCs receive time-of-day-dependent inputs from both CRYPTOCHROME-negative LNds and DN1s. Surprisingly, clock neurons inhibit DH44+ neurons while activating IPCs, indicating that the same clock cells use multiple mechanisms simultaneously to drive cycling in output neurons. It was also shown that morning clock cells and PI-secreted insulin-like peptides are required for rhythms of feeding (Barber, 2021).
The Drosophila PI was previously described as a protohypothalamic region that serves as a circadian output hub; while inputs from DN1 clock neurons were shown, relevance of other clock neurons or the signaling molecules involved in transducing time-of-day cues from the clock to the PI were not described. This study demonstrates that time-of-day information arrives at the PI from multiple clock neuron populations through cholinergic and glutamatergic signaling. This study further identified an output role for the IPCs of the PI in regulating circadian feeding, which depends on inputs from morning-active, but not evening-active, clock neurons (Barber, 2021).
The response of PI neurons to clock neuron stimulation is time-of-day dependent. DH44+ neurons are inhibited-i.e., they exhibit a reduction in intracellular calcium-by both DN1 and LNd neuron stimulation in the morning, with little to no effect of stimulation of clock neurons in the evening. Given that LNds drive the evening peak of locomotor activity, it is surprising that their stimulation only inhibits DH44+ neurons in the morning; however, inhibition in the morning is consistent with the normal pattern of activity of DH44+ neurons. DH44+ neurons are evening active in both LD and DD conditions, and while they regulate both the morning and evening peaks of locomotor activity in LD, their contribution to the evening peak is larger. It is likely that DH44+ neurons, which are implicated in several processes, receive nonclock inputs that drive locomotor activity, and they require silencing by the clock to inhibit locomotion outside the morning and evening activity peaks. Thus, silencing by DN1s may serve to delineate the morning peak of activity; on the other hand, early day silencing of DH44+ neurons by LNds may be one mechanism by which LNds drive evening activity (Barber, 2021).
The response of IPCs to DN1 stimulation is time-of-day dependent. The response to LNd stimulation also shows an apparent difference between morning and evening, but, in fact, some cells respond at both times. The IPC population is larger than the DH44+ population and is known to be heterogeneous in terms of basal activity and signaling molecule expression. The functional relevance of LNd signaling to IPCs is unclear, but could be linked to the role of these cells in promoting arousal. However, the DN1 inputs likely contribute to the circadian rhythm of feeding. As noted above, this is consistent with the higher firing of IPCs in the morning and the morning peak of the feeding rhythm in w1118 flies, as well as with the finding that DILPs are required for rhythmic feeding. In further support of this idea, this study reports that feeding rhythms depend upon the activity of morning-active LNvs, which signal through DN1s, but not evening-active LNds. These finding of loss of feeding rhythms on yeast/sucrose diet with genetic ablation of two insulin-like peptides differs from a previous study that observed maintenance of rhythmic feeding on sucrose-only food upon adult silencing of IPCs. It is speculated that developmental changes caused by genetic ablation of insulin(s) result in disorganized feeding patterns and/or that rhythmic protein, but not sugar, feeding is regulated by brain insulins (Barber, 2021).
The finding that nAChR and mGluR signaling contribute to inhibition in one case (input from DN1 and LNds on to DH44 neurons) and activation in another (input from DN1 and LNds on to IPCs) raises the question of what determines the nature of the effect. The most straightforward explanation would be a difference in the receptor subtypes expressed in the two cell groups. Alternatively, other signaling components specific to each cell type could account for the differential response. Regardless, it is clear that the actions of known neurotransmitters can be regulated by the clock in different ways to effect rhythmic output. As noted above, acetylcholine is secreted by the LNds and likely also DN1s, but to date, the only known source of glutamate in the clock network are the DN1s (16, 20). This suggests that glutamatergic input from the DN1s can modulate the effect of LNds on PI neurons (Barber, 2021).
Despite the effect of clock cell-secreted neuropeptides on PI cells and the role of fast neurotransmitters in signaling from clock cells to the PI, knockdown of neuropeptide/neurotransmitter receptors in the PI has little effect on behavioral rhythms. There are many possible explanations for lack of a phenotype-low mRNA levels remaining after knockdown are sufficient for function; receptor subtypes act redundantly in the regulation of rhythms; knockdown in the PI alone is insufficient to yield a phenotype; or these molecules affect physiological rhythms other than those of rest:activity. While DH31 and NPF have been implicated in the control of rest:activity rhythms, loss of acetylcholine and glutamate have not been linked to behavioral rhythms. Although knockdown of receptors for these molecules in the PI did not yield a phenotype, reduced evening activity was seen with DH44-driven knockdown of PDFR. This is in contrast to the effect of loss-of-function pdf mutants in which evening activity remains high, but is shifted ~1 h early. It is possible that DH44+ cell-mediated reduced evening activity is compensated in pdfr mutants. PDF is secreted by LNvs, which are not known to synapse onto the PI, but do project to the dorsal brain, suggesting diffusion of PDF across a small region. Limited diffusion of PDF is supported by a previous study showing that overexpression of PDF in cells that project to the dorsal brain produces behavioral phenotypes (Barber, 2021).
These findings elucidate some of the complexity underlying circadian control in neural circuits. Through the use of neuropeptides and fast neurotransmitters coupled with time-of-day-specific actions on downstream neurons, clock cells are able to drive multiple outputs at varying timescales. Behavioral outputs require that rhythmic patterns of activity be propagated through many parts of the brain, and this is likely the case. Previous work has shown that hugin+ neurons, which are downstream of the PI, show rhythmic peptide release, indicating that cycling is propagated to second order output neurons. In addition, clock neurons are linked to rhythmic neural activity in locomotor centers of the fly brain. While a rhythm in spiking activity may be sufficient to transmit some outputs, it is known that other variations in electrical activity (e.g., temporal coding from DN1 neurons) may also contribute to circadian behaviors. As in mammals, different output rhythms also map to different output circuits in flies. Thus, Drosophila take advantage not only of diverse neural output mechanisms to regulate aspects of circadian behavior, but also use distinct neuropeptidergic output cells to regulate locomotor and feeding rhythms. DH44+ neurons are important for locomotor, but not feeding, rhythms, while the opposite is true for IPCs. At the same time, proximity of output neurons controlling locomotor activity with those that control feeding likely allows for integration of multiple signals to coordinate behavior, thus contributing to organismal fitness (Barber, 2021).
Drosophila melanogaster is a long-standing model organism in the circadian clock research. A major advantage is the relative small number of about 150 neurons, which built the circadian clock in Drosophila. A recent work focused on the neuroanatomical properties of the lateral neurons of the clock network. By applying the multicolor-labeling technique Flybow it was possible to identify the anatomical similarity of the previously described E2 subunit of the evening oscillator of the clock, which is built by the 5th small ventrolateral neuron (5th s-LNv ) and one ITP positive dorsolateral neuron (LNd ). These two clock neurons share the same spatial and functional properties. Both neurons were found innervating the same brain areas with similar pre- and postsynaptic sites in the brain. The anatomical findings support their shared function as a main evening oscillator in the clock network like also found in previous studies. A second quite surprising finding addresses the large lateral ventral PDF-neurons (l-LNv s). It was shown that the four hardly distinguishable l-LNv s consist of two subgroups with different innervation patterns. While three of the neurons reflect the well-known branching pattern reproduced by PDF immunohistochemistry, one neuron per brain hemisphere has a distinguished innervation profile and is restricted only to the proximal part of the medulla-surface. This neuron was named "extra" l-LNv (l-LNv x). The anatomical findings reflect different functional properties of the two l-LNv subgroups (Schubert, 2018).
The brain clock that drives circadian rhythms of locomotor activity relies on a multi-oscillator neuronal network. In addition to synchronizing the clock with day-night cycles, light also reformats the clock-driven daily activity pattern. How changes in lighting conditions modify the contribution of the different oscillators to remodel the daily activity pattern remains largely unknown. Data in Drosophila indicate that light readjusts the interactions between oscillators through two different modes. This study shows that a morning s-LNv > DN1p circuit works in series, whereas two parallel evening circuits are contributed by LNds and other DN1ps. Based on the photic context, the master pacemaker in the s-LNv neurons swaps its enslaved partner-oscillator-LNd in the presence of light or DN1p in the absence of light-to always link up with the most influential phase-determining oscillator. When exposure to light further increases, the light-activated LNd pacemaker becomes independent by decoupling from the s-LNvs. The calibration of coupling by light is layered on a clock-independent network interaction wherein light upregulates the expression of the PDF neuropeptide in the s-LNvs, which inhibits the behavioral output of the DN1p evening oscillator. Thus, light modifies inter-oscillator coupling and clock-independent output-gating to achieve flexibility in the network. It is likely that the light-induced changes in the Drosophila brain circadian network could reveal general principles of adapting to varying environmental cues in any neuronal multi-oscillator system (Chatterjee, 2018).
Circadian rhythms are orchestrated by a master clock that emerges from a network of circadian pacemaker neurons. The master clock is synchronized to external light/dark cycles through photoentrainment, but the circuit mechanisms underlying visual photoentrainment remain largely unknown. This study reports that Drosophila has eye-mediated photoentrainment via a parallel pacemaker neuron organization. Patch-clamp recordings of central circadian pacemaker neurons reveal that light excites most of them independently of one another. Light-responding pacemaker neurons were shown to send their dendrites to a neuropil called accessary medulla (aMe), where they make monosynaptic connections with Hofbauer-Buchner eyelet photoreceptors and interneurons that transmit compound-eye signals. Laser ablation of aMe and eye removal both abolish light responses of circadian pacemaker neurons, revealing aMe as a hub to channel eye inputs to central circadian clock. Taken together, this study demonstrates that the central clock receives eye inputs via hub-organized parallel circuits in Drosophila (Li, 2018).
Animals choose among sleep, courtship, and feeding behaviors based on the integration of both external sensory cues and internal states; such choices are essential for survival and reproduction. These competing behaviors are closely related and controlled by distinct neural circuits, but whether they are also regulated by shared neural nodes is unclear. This study investigated how a set of male-specific P1 neurons controls sleep, courtship, and feeding behaviors in Drosophila males. Mild activation of P1 neurons was sufficient to affect sleep, but not courtship or feeding, while stronger activation of P1 neurons labeled by four out of five independent drivers induced courtship, but only the driver that targeted the largest number of P1 neurons affected feeding. These results reveal a common neural node that affects sleep, courtship, and feeding in a threshold-dependent manner, and provide insights into how competing behaviors can be regulated by a shared neural node (Zhang, 2018).
Circadian clocks play conserved roles in gating sleep and wake states throughout the day-night cycle. In the fruit fly Drosophila melanogaster, DN1p clock neurons have been reported to play both wake- and sleep-promoting roles, suggesting a complex coupling of DN1p neurons to downstream sleep and arousal centers. However, the circuit logic by which DN1p neurons modulate sleep remains poorly understood. This study shows that DN1p neurons can be divided into two morphologically distinct subsets. Projections from one subset surround the pars intercerebralis, a previously defined circadian output region. In contrast, the second subset also sends presynaptic termini to a visual processing center, the anterior optic tubercle (AOTU). Within the AOTU, DN1p neurons inhibit a class of tubercular-bulbar (TuBu) neurons that act to promote consolidated sleep. These TuBu neurons in turn form synaptic connections with R neurons of the ellipsoid body, a region linked to visual feature detection, locomotion, spatial memory, and sleep homeostasis. These results define a second output arm from DN1p neurons and suggest a role for TuBu neurons as regulators of sleep drive (Lamaze, 2018).
The Drosophila core circadian circuit contains distinct groups of interacting neurons that give rise to diurnal sleep-wake patterns. Previous work showed that a subset of dorsal neurons 1 (DN1s) are sleep-promoting through their inhibition of activity-promoting circadian pacemakers. This study shows that these anterior-projecting DNs (APDNs) also "exit" the circadian circuitry and communicate with the homeostatic sleep center in higher brain regions to regulate sleep and sleep-wake arousal. These APDNs connect to a small, discrete subset of tubercular-bulbar neurons, which are connected in turn to specific sleep-centric ellipsoid body (EB)-ring neurons of the central complex. Remarkably, activation of the APDNs produces sleep-like oscillations in the EB and affects arousal. The data indicate that this APDN-TuBusup-EB circuit temporally regulates sleep-wake arousal in addition to the previously defined role of the TuBu-EB circuit in vision, navigation, and attention (Guo, 2018).
Animals consolidate some, but not all, learning experiences into long-term memory. Across the animal kingdom, sleep has been found to have a beneficial effect on the consolidation of recently formed memories into long-term storage. However, the underlying mechanisms of sleep dependent memory consolidation are poorly understood. This study shows that consolidation of courtship long-term memory in Drosophila is mediated by reactivation during sleep of dopaminergic neurons that were earlier involved in memory acquisition. Specific fan-shaped body neurons were identified that induce sleep after the learning experience and activate dopaminergic neurons for memory consolidation. Thus, this study provide a direct link between sleep, neuronal reactivation of dopaminergic neurons, and memory consolidation (Dag, 2019).
The activity of DAN-aSP13s, which is essential for courtship memory acquisition, is also necessary during a discrete post-learning time window for LTM consolidation. Because neuronal reactivation occurs during sleep in rodents, it was hypothesized that post-learning activation of DAN-aSP13s involves a sleep-dependent mechanism. Using behavioral analysis and neuronal activity monitoring and perturbation approaches, thus study shows that DAN-aSP13s display an increased activity in freely behaving animals during sleep after a prolonged learning experience. This sleep is necessary for LTM consolidation, and it can be mediated by a specific class of sleep promoting neurons in the ventral layer of the fan shaped body (vFB). These vFB neurons consolidate courtship LTM in a discrete time window and provide an excitatory input to DAN-aSP13s. Thus, these data provide a causal link between sleep promoting neurons in the vFB, post-learning activation of dopaminergic neurons, and LTM consolidation (Dag, 2019).
Based on these data, the following model is proposed for sleep-dependent consolidation of courtship LTM in Drosophila (see Post-learning activation of DAN-aSP13 neurons mediates LTM consolidation). During a prolonged learning experience, γKCs and DAN-aSP13s are repeatedly activated by olfactory and behavioral cues presented by an unreceptive female, respectively. Whereas prolonged wakefulness leads to an increase in homeostatic sleep drive in ellipsoid body neurons that in turn is conveyed to dFB, it is hypothesized that an extended learning experience generates a learning-dependent sleep drive that is transmitted to vFB. In turn, the vFB neurons enhance sleep after learning and provide an excitatory input back on DAN-aSP13s. It is thought that one potential site of a learning-dependent sleep drive are the MB neurons since they have been implicated in both memory formation and sleep regulation. Dopamine released upon DAN-aSP13 reactivation stimulates molecular processes in γKCs that are different from those engaged during STM acquisition and involve protein synthesis that is essential for LTM formation and persistence (Dag, 2019).
Dopaminergic pathways are thought to convey information about whether an experience is rewarding or punishing and thus, worth remembering. Post-learning neuronal activity of the dopaminergic hippocampal inputs from the Ventral Tagmental Area (VTA) has been implicated in the consolidation of fear memory in rodents. Interestingly, this activity is critical during a discrete time window after learning. Post-learning activity of the VTA dopamine neurons has been also implicated in the reactivation during sleep of the hippocampal cells involved earlier in encoding of the spatial experience. This study shows that post-learning activation of DAN-aSP13s mediates the consolidation of courtship LTM in Drosophila. It is proposed that reactivation during sleep of the dopamine neurons that were previously active during memory acquisition ensures that spurious experiences are not admitted into LTM storage and thus only experiences that are either sufficiently salient or persistent become long-lasting memories. Specifically, reactivation of DAN-aSP13s during post-learning sleep enhances reactivation of the γKCs and cognate MBON-M6, which together with DAN-aSP13s form a recurrent circuit necessary for courtship memory acquisition. This study considers two hypotheses to account for this selectivity. During sleep, vFB neurons might selectively reactivate only the relevant DANs, or alternatively, they might activate all DANs but only the relevant subset is able to consolidate LTM. The selective-reactivation model would require some marker to distinguish which DANs were activated, whereas the selective-consolidation model would require a marker in the synapses of the γKCs that were earlier active during memory acquisition, for example translational regulator Orb2, which regulates translation upon neuronal activity during LTM consolidation (Dag, 2019).
Activity of dopaminergic neurons regulates sleep-wake states in animals, including flies. Artificial activation of DAN-aSP13s has been shown to increase wakefulness. In contrast, the data presented in this study imply that activation of vFB neurons, although they activate DAN-aSP13s, do not promote wakefulness. These results suggest that activation of DAN-aSP13s by vFB neurons during sleep is qualitatively different from a direct optogenetic or thermogenetic activation used in previous studies. One potential explanation is that post-learning sleep involves activation of the vFB circuit, which provides both excitatory stimulus to DAN-aSP13s and inhibitory input to motor neurons. Another possibility is that post-learning sleep activates a subset od DAN-aSP13s that do not affect wakefulness (Dag, 2019).
Thia study has identified a class of sleep-promoting neurons in the ventral layer of FB that are distinct from the well-studied sleep-promoting neurons in the dorsal layer of FB, which regulate sleep homeostasis. Given that vFB neurons enhance sleep and activate DAN-aSP13s for LTM consolidation, whereas dFB neurons are neither necessary nor sufficient for LTM consolidation, it is hypothesized that dFB and vFB neurons promote distinct components of sleep that have different functions. Homeostatic sleep is thought to facilitate memory encoding by downscaling synaptic weights and clearing metabolites from the brain accumulated during wakefulness. In contrast, the function of learning-dependent sleep might be to facilitate memory consolidation by strengthening synaptic connections that were engaged earlier during memory acquisition. Thus, the co-operation of homeostatic- and experience-dependent sleep would facilitate optimal conditions for learning new information and, if appropriate, incorporating it into long-term storage (Dag, 2019).
Recent studies have implied that sleep in flies, as in humans and rodents, exhibits sleep stages characterized by distinct electrophysiological signatures. Interestingly, the signature of sleep that is induced by activation of the dFB neurons seems to have a simpler oscillatory pattern, recorded by local field potentials, than sleep that is induced by activation of the FB neurons comprising both dFB and vFB neurons. Thus, these data support the hypothesis that dFB and vFB neurons promote sleep with different properties and likely different functions (Dag, 2019).
It is thought that sleep evolved in animals that are capable of complex learning which requires selective attention. Courtship learning is a multisensory form of learning that requires selective attention of a male to associate multiple learning cues presented by the mated female with the outcome of his own behavior. Accordingly, studies in bees have shown that sleep affects a complex form of learning such as spatial memory but has no role in the simple learning paradigm of proboscis extension. Hence, it would be interesting to investigate whether post-learning sleep is involved in the consolidation of other types of memory in Drosophila, such as the well-studied Pavlovian olfactory associative learning whereby animals associate an individual learning cue with a behavioral contingency (Dag, 2019).
This work has established a functional link between a novel class of sleep-promoting neurons in the FB, post-learning reactivation of dopaminergic neurons and consolidation of courtship LTM. Moreover, the data suggest that sleep promoting vFB neurons mediate a learning-dependent regulation of sleep that is distinct from the homeostatic control which is facilitated by dFB neurons. Thus, this study uncovered a causal link between sleep-mediated neuronal reactivation and LTM consolidation in Drosophila. In addition, courtship LTM was establish in Drosophila as a tractable model to investigate the mechanisms that link learning-dependent sleep, neuronal reactivation and LTM consolidation (Dag, 2019).
Lei, Z., Henderson, K. and Keleman, K. (2022). A neural circuit linking learning and sleep in Drosophila long-term memory. Nat Commun 13(1): 609. PubMed ID: 35105888
Animals retain some but not all experiences in long-term memory (LTM). Sleep supports LTM retention across animal species. It is well established that learning experiences enhance post-learning sleep. However, the underlying mechanisms of how learning mediates sleep for memory retention are not clear. Drosophila males display increased amounts of sleep after courtship learning. Courtship learning depends on Mushroom Body (MB) neurons, and post-learning sleep is mediated by the sleep-promoting ventral Fan-Shaped Body neurons (vFBs). This study shows that post-learning sleep is regulated by two opposing output neurons (MBONs) from the MB, which encode a measure of learning. Excitatory MBONs-γ2α'1 becomes increasingly active upon increasing time of learning, whereas inhibitory MBONs-β'2mp is activated only by a short learning experience. These MB outputs are integrated by SFS neurons, which excite vFBs to promote sleep after prolonged but not short training. This circuit may ensure that only longer or more intense learning experiences induce sleep and are thereby consolidated into LTM (Lei, 2022).
This study has identified a neural circuit that regulates learning-induced sleep for LTM consolidation. This circuit links neurons essential for learning and memory in Drosophila, the MB neurons, with those critical for post-learning sleep, the vFBs8. It is proposed that only a longer learning experience is sufficient to induce sleep, and thereby be consolidated into LTM. Given that the increasing duration of a learning experience correlates with the total amount of time males spend on futile courtship towards mated females during training, selective activation of vFBs likely depends on the amount or intensity of a learning experience, rather than just its duration. Post-learning sleep induction requires integration of two MB outputs, previously implicated in courtship memory in SFSs. Post-learning activity of the excitatory MBONs-γ2α'1 increases linearly with the duration of the prolonged learning experience. In contrast, activity of the inhibitory MBONs-β'2mp peaks after a short experience sufficient to induce STM. As a result, only when the males court mated females sufficiently long or intensely, the activity of MBONs-γ2α'1 reaches the threshold required to activate SFSs. This in turn leads to activation of vFBs to promote post-learning sleep and the reactivation of those dopaminergic neurons (DANs) that were involved in memory encoding. Consequently, biochemical processes essential for LTM consolidation become engaged (Lei, 2022).
How might MBONs-γ2α'1 and MBONs-β'2mp measure the learning experience to control post-learning sleep? In homeostatic sleep regulation, the potentiation of R2 neurons reflects a measure of sleep loss that is sensed by dFBs, likely in response to the accumulation of byproducts of oxidative stress during sleep loss. In the case of learning-induced sleep, it is envisioned that learning results in lasting changes in the molecular pathways essential for memory formation in the MB. For example, the cAMP pathway along with the dopamine receptor are activated during sleep in a discrete 3-h time window after learning in rodents and Drosophila males lacking a dopamine receptor, and hence unable to learn, do not display increased post-learning sleep. Thus, the accumulation of changes in the cAMP signaling pathway upon increasing learning experience with mated females might lead to the increasing potentiation of MBONs-γ2α'1 and MBONs-β'2mp after learning. Interestingly, MBONs-γ2α'1 and MBONs-β'2mp display distinct temporal activity patterns upon learning which likely reflects their distinct neuronal properties (Lei, 2022).
This study reveals a circuit mechanism that ensures that only persistent, and thus likely significant, learning experiences generate post-learning sleep to consolidate LTM. Recent findings suggest that dFBs, involved in sleep homeostasis, might mediate a paradoxical type of sleep, in humans also called Rapid Eye Movement (REM) sleep. This in conjunction with the current data, provide an opportunity to investigate whether the post-learning sleep, mediated by vFBs, might represent another type of sleep implicated in mammals in memory consolidation (Lei, 2022).
Sleep plays an important role in regulating plasticity. In Drosophila, the relationship between sleep and learning and memory has primarily focused on mushroom body dependent operant-learning assays such as aversive phototaxic suppression and courtship conditioning. In this study, sleep was increased in the classic mutant rutabaga (rut2080) and dunce (dnc1) by feeding them the GABA-A agonist gaboxadol (Gab). Performance was evaluated in each mutant in response to social enrichment and place learning, tasks that do not require the mushroom body. Gab-induced sleep did not restore behavioral plasticity to either rut2080 or dnc1 mutants following social enrichment. However, increased sleep restored place learning to rut2080 mutants. These data extend the positive effects of enhanced sleep to place learning and highlight the utility of Gab for elucidating the beneficial effects of sleep on brain functioning (Dissel, 2020).
Sleep loss and aging impair hippocampus-dependent Spatial Learning in mammalian systems. This study used the fly Drosophila melanogaster to investigate the relationship between sleep and Spatial Learning in healthy and impaired flies. The Spatial Learning assay is modeled after the Morris Water Maze. The assay uses a 'thermal maze' consisting of a 5X5 grid of Peltier plates maintained at 36-37°C and a visual panorama. The first trial begins when a single tile that is associated with a specific visual cue is cooled to 25°C. For subsequent trials, the cold tile is heated, the visual panorama is rotated and the flies must find the new cold-tile by remembering its association with the visual cue. Significant learning was observed with two different wild-type strains - Cs and 2U, validating the design. Sleep deprivation prior to training impaired Spatial Learning. Learning was also impaired in the classic learning mutant rutabaga (rut); enhancing sleep restored learning to rut mutants. Further, flies exhibited dramatic age-dependent cognitive decline in Spatial Learning starting at 20-24 days of age. These impairments could be reversed by enhancing sleep. Finally, Spatial Learning was found to requires dopaminergic signaling and that enhancing dopaminergic signaling in aged flies restored learning. These results are consistent with the impairments seen in rodents and humans. These results thus demonstrate a critical conserved role for sleep in supporting Spatial Learning, and suggest potential avenues for therapeutic intervention during aging (Melnattur, 2020).
Memory forms when a previously neutral stimulus (CS+) becomes competent to predict a biologically potent stimulus (US). However, if the CS+ is repeatedly presented without the US after the memory formation, this memory will be suppressed by newly formed extinction memory. The striking feature of extinction learning is that it requires repeated trials to robustly form extinction. Extended repetition only yields memories that remain transient in nature, thus imposing challenges in understanding the underlying mechanisms of extinction learning. This study took advantage of the versatile genetic tools and the well-characterized circadian system of Drosophila to link these unique features to clock neurons. Inhibiting the activity of clock neurons blocks the formation of extinction memory. Further investigation attributes this role to a subset of cryptochrome-positive dorsal neurons 1 (DN1s) and their downstream SIFamide neurons. The requirement of clock neurons from a gating mechanism of extinction for a single extinction learning trial robustly causes typical extinction when coupled with acute activation of DN1s, as marked by the initially enhanced but eventually diminished memory suppression. Accordingly, specific neural responses were detected to extinction training in a few DN1s via calcium imaging fulfilled by the TRIC tool, but not in dorsal neuron 2 or dorsolateral neurons. Based on these findings, it is proposed that in extinction of appetitive long-term memory, multiple trials of extinction learning robustly activate DN1 clock neurons to open the gate of extinction, which may contribute to the transient nature of extinction memory (Zhang, 2021).
The current study focused on the extinction of 24-h appetitive long-term memory (apLTM) because only 24-h apLTM shows spontaneous recovery. Examination of extinction with a transient nature revealed that clock neurons are a key component in the mechanism by which multiple trials of extinction learning are required to robustly form the transient extinction memory. The data presented in this study strongly support the proposed model, as multiple trials of extinction learning are required to robustly activate four pairs of CRY-positive, Spl-DN1-Gal4-labeled DN1s to permit extinction. Therefore, this study shows that multi-trial extinction learning is functionally heterogeneous, with multiple trials needed in the robust activation of a subset of clock neurons and a single trial for acquiring the CS-no US association. The coupling of these two factors results in the extinction of the 24-h apLTM, i.e., clock neuron gate extinction (Zhang, 2021).
Four lines of evidence support these conclusions. First, multiple, but not single, trials of extinction learning or the presentation of CS+ for multiple times robustly activates DN1s, but not DN2s or LNds, as detected by accumulating signals for TRIC (a tool for imaging intracellular calcium), an intracellular Ca2+ indicator. Second, suppression of the activity of only CRY-positive, but not CRY-negative, clock neurons, particularly four pairs of CRY-positive, Spl-DN1-Gal4-labeled DN1s, blocked extinction. Third, the artificial activation of Spl-DN1-Gal4-labeled DN1s coupled with a single extinction trial potentiated the transient extinction of 24-h apLTM. Fourth, suppression of the neuronal activity of downstream target SIFa neurons also blocked natural extinction (Zhang, 2021).
The results suggest the role of clock neurons in memory regulation by extinction. In previous studies, the circadian system has been reported to regulate multiple physiological processes. In general, these regulatory effects have been attributed to oscillation of the circadian system. However, this study showed that extinction learning itself can evoke neural responses in DN1s to open the gate for extinction memory, instead of its passive modulation driven by circadian rhythm. Thus, it would be of considerable interest to determine whether such active regulation of clock neurons also occurs in other physiological processes (Zhang, 2021).
Recent studies on the fly's extinction have identified the PPL-1 dopaminergic neurons and γ mushroom body neurons as the key components in mediating the extinction process. In addition, the downstream SIFa neurons are known to send neurites throughout the adult brain. It would be of interest to determine how DN1 neurons, as well as their downstream targets, are related to functions of previously identified neural networks mediating extinction (Zhang, 2021).
Many animals exhibit morning and evening peaks of locomotor behavior. In Drosophila, two corresponding circadian neural oscillators-M (morning) cells and E (evening) cells-exhibit a corresponding morning or evening neural activity peak. Yet little is known of the neural circuitry by which distinct circadian oscillators produce specific outputs to precisely control behavioral episodes. This study shows that ring neurons of the ellipsoid body (EB-RNs) display spontaneous morning and evening neural activity peaks in vivo: these peaks coincide with the bouts of locomotor activity and result from independent activation by M and E pacemakers. Further, M and E cells regulate EB-RNs via identified PPM3 dopaminergic neurons, which project to the EB and are normally co-active with EB-RNs. These in vivo findings establish the fundamental elements of a circadian neuronal output pathway: distinct circadian oscillators independently drive a common pre-motor center through the agency of specific dopaminergic interneurons (Liang, 2019).
The circadian clock is a timekeeper but also helps adapt physiology to the outside world. This is because an essential feature of clocks is their ability to adjust (entrain) to the environment, with light being the most important signal. Whereas cryptochrome-mediated entrainment is well understood in Drosophila, integration of light information via the visual system lacks a neuronal or molecular mechanism. This study shows that a single photoreceptor subtype is essential for long-day adaptation. These cells activate key circadian neurons, namely the large ventral-lateral neurons (lLNvs), which release the neuropeptide pigment-dispersing factor (PDF). RNAi and rescue experiments show that PDF from these cells is necessary and sufficient for delaying the timing of the evening (E) activity in long-day conditions. This contrasts to PDF that derives from the small ventral-lateral neurons (sLNvs), which are essential for constant darkness (DD) rhythmicity. Using a cell-specific CRISPR/Cas9 assay, this study shows that lLNv-derived PDF directly interacts with neurons important for E activity timing. Interestingly, this pathway is specific for long-day adaptation and appears to be dispensable in equinox or DD conditions. The results therefore indicate that external cues cause a rearrangement of neuronal hierarchy, which contributes to behavioral plasticity (Schlichting, 2019a).
Circadian clocks evolved as an adaptation to the continuous change of day and night and are believed to provide organisms a fitness advantage. The underlying molecular machinery includes a transcriptional-translational feedback loop, which generates oscillations of clock gene expression with an endogenous period close to 24 h. This period is approximately 24.2 h in humans, whereas a Drosophila period was reported to be 23.8 h. A key feature of circadian clocks is the ability to entrain to the 24 h environment. This means that the human clock has to be accelerated by about 0.2 h each day, whereas this Drosophila clock has to be slowed down to the same extent. To do so, clocks must integrate external cues, so-called zeitgebers, which are used to synchronize the molecular and physiological properties of the organism (Schlichting, 2019a).
The most important zeitgeber is light. In mammals, a combination of the traditional photoreception pathway (rods and cones) and the circadian photoreceptor melanopsin in retinal ganglion cells allows for fine-tuning of clock synchronization. Similarly, Drosophila uses the visual system and possibly Rhodopsin 7 (Rh7) within the clock neurons as well as the circadian photoreceptor cryptochrome (CRY) for light synchronization. CRY-mediated entrainment is well understood in Drosophila, whereas less is known about the mechanism of entrainment via the visual system. It consists of seven eye structures: three ocelli, two Hofbauer-Buchner eyelets, and two compound eyes (Schlichting, 2019a).
The compound eye consists of approximately 800 ommatidia, each harboring 8 photoreceptor cells (Rs): R1-6 are located in the periphery and span the whole depth of the ommatidium. These cells were previously shown to be important for motion vision and express Rhodopsin 1 (Rh1). In the center, R7 is located above R8. These cells have a complex expression pattern of Rh4 and/or Rh3 in R7 and Rh5 or Rh6 in R8. Although the mechanism of light transduction from the visual system to the central clock is still not completely understood, recent work indicates a special role for R8. These cells specifically target the sLNvs in standard conditions of 12 h light and 12 h darkness (LD 12:12). R8 photoreceptors additionally express and react to HisCl1 and can therefore not only act as photoreceptors but also as interneurons (Alejevski, 2109; Schlichting, 2019a and references therein).
Recent electrophysiological results further suggest that the visual system is able to activate an array of circadian clock neurons, e.g., it can activate the small ventral-lateral neurons (sLNvs), an important center for morning (M) activity. Furthermore, the visual system increases neuronal firing in the large LNvs (lLNvs), the arousal center within the circadian network. The 5th sLNv and the NPF+ dorsal-lateral neurons (LNds), previously implicated as necessary for evening (E) activity, also increase their firing rates in response to visual system stimulation. In addition, the visual system activates several dorsal neurons (DNs), which were recently implicated in connecting the circadian clock to central brain sleep centers. These data suggest that visual input is integrated into the clock network in a parallel fashion, which contradicts a master-oscillator point of view. The latter posits that these are the pigment-dispersing factor (PDF)-expressing neurons (sLNvs and lLNvs), which receive light input and release PDF upon illumination, thereby adjusting their downstream target neurons to the LD cycle (Schlichting, 2019a).
To investigate the impact of the visual input pathway at the behavioral and neuronal level, this study investigated fly behavior under long-day conditions. Long days cause plastic changes in fly behavior: in standard light-dark cycles (LD 12:12), flies show a bimodal activity pattern with a M anticipation peak around lights on and an E anticipation peak around lights off; this results in a phase relationship of approximately 12 h between the two peaks appropriately adjust their peak timings, i.e., their E peak is phase advanced compared to wild-type flies. Moreover, visual input appears to modulate PDF release from the lateral neurons, which in turn modulates cells important for E activity. In summary, complex interactions between CRY- and PDF-expressing neurons appear to be essential for the behavior under long days: expressing these proteins in different parts of the fly brain alters the behavior of flies and mimics the behavior of high-latitude species using Drosophila melanogaster as a model. It is still unknown, however, which receptors and which neuronal pathways are involved in this adjustment (Schlichting, 2019a).
This study shows that R8 of the compound eyes is essential for long-day adaptation. These photoreceptor cells connect to the PDF-containing lLNvs and trigger the release of this neuropeptide. Using a cell-specific CRISPR/Cas9 strategy, it was demonstrated that light-mediated PDF directly signals to the PDF receptor (PDFR) on E cells and hence delays E activity. The data implicate a mammal-like structure of clock entrainment, with the visual system activating PDF-expressing clock neurons. These data further support a shift of PDF targets between LD and constant darkness (DD) conditions as well as a more quantitative reorganization of neuronal dominance within the clock network by changes in photoperiod (Schlichting, 2019a).
The circadian clock is able to entrain to the changes of day and night, with light being the most important zeitgeber. The adaptation to summer-like days is especially important for insects, as they are prone to predator visibility and even more importantly desiccation. Therefore, the circadian clock has to be plastic and be able to adjust behavior in response to changing environments. For example, flies show an additional afternoon peak during summer days under semi-natural conditions, which is thought to be an escape response of the fly from heat. This study shows that Drosophila adjusts its behavior to extremely long photoperiods (LD 20:4) by delaying its E peak as reported previously. Even though this photoperiod can only be found in very northern countries, Drosophila melanogaster is still able to adjust to this extreme light condition, which exemplifies its ability to adapt to various environmental conditions. This delay allows the animal to reduce its activity during the unfavorable midday, when temperatures are highest. Most interestingly, this phenotype is easily visible even without temperature changes, underscoring the importance of light as a major entrainment cue (Schlichting, 2019a).
A central finding is that flies lacking the compound eyes show an entrainment deficit, i.e., they have an advanced E peak under long-day conditions. Similarly, they show a reduction in M peak amplitude; this is likely due to a failure to properly activate PDF-expressing neurons. Using rhodopsin mutants and by manipulating specific photoreceptors using the GAL4/UAS system, only R8 of the compound eyes appears essential for this summer day response; R8 was previously implicated in the adaptation to nature-like light conditions (Schlichting, 2019a).
Notably, even flies lacking all compound eyes significantly delay their E peak timing under long photoperiod conditions but to a much smaller extent. This indicates that other photoreceptors also contribute to the E peak delays under these conditions. One likely candidate is the HB-eyelet. Recent work has shown that this photoreceptor contributes to delaying the E peak under high-light-intensity conditions [48]. Similar mechanisms might apply under long days, which is supported by the strongly advanced E peak when rh5-GAL4 and rh6-GAL4-positive neurons, which includes the HB eyelets, were silenced or ablated (Schlichting, 2019a).
lLNv arbors in the optic lobe are in close proximity to R8 termini, where they most likely interact via cholinergic interneurons in addition to the accessory medulla, which was recently shown to be important for light-evoked responses of the lLNvs. This interaction results in a change of neuronal bursting behavior and hence neuropeptide release. Indeed, this study shows that release of PDF from the lLNvs is necessary and sufficient for proper long-day adaptation (Schlichting, 2019a).
These results are surprising given a recently published study on how the visual system is connected to the clock neuron network vv. It shows that the visual system can activate a broad spectrum of lateral and dorsal neurons; they include sLNvs, lLNvs, ITP+ LNds, and DN2s, among others. Ablation of PDF neurons left the other neurons responsive to visual input, suggesting a parallel model for clock synchronization, i.e., information from the visual system can be directly transferred to independent classes of clock neurons rather than only via PDF. This new pathway might be involved in the residual delay of E activity in pdf01 flies under long photoperiods, suggesting a potential PDF-independent contribution to long-day adaptation (Schlichting, 2019a).
PDF stimulates different adenylate-cyclases and increases cAMP, which leads to the stabilization of PER and TIM and consequently a longer period or phase delay. Therefore, one view is that removing the compound eyes decreases PDF release from the lLNvs and phase advances the molecular clock in downstream target neurons like the LNds. This newly discovered 'visual system to LNd pathway' might also enhance CRY-mediated photoentrainment: CRY was shown to activate neurons upon stimulation, similar to the newly identified light activation of clock neuron pathway. Additional activation of the E cells could therefore contribute to the kinetics of TIM degradation, which was recently shown to be important for the phase advance of E activity under long-day conditions (Schlichting, 2019a).
An intriguing inference of this work is that the principal targets of PDF must change with the environmental conditions. Previous work established the sLNvs as essential for DD rhythmicity, and recent work shows that these neurons are tightly coupled to the dorsal clock neurons in DD: speeding up the PDF neurons forced the DN1s to follow the short period of the sLNvs. In LD, however, this connection is much weaker, and the cell-type-specific CRISPR/Cas9 knockout strategy shows that it is the PDF-expressing lLNvs that communicate with the LNd neurons. The data show that the lLNv to LNd connection is important in LD conditions but does not affect DD behavior (Schlichting, 2019a).
Importantly, the data not only indicate a qualitative shift of PDF targets between DD and LD but also suggest a quantitative shift of dominance, depending on photoperiod or the time of light exposure. In DD, the sLNvs are necessary for rhythmic behavior and show robust cycling in PER oscillations, whereas the lLNvs lose PER rhythms as early as the second day of DD. In equinox conditions, both groups may be relevant: PDF from either the sLNvs or lLNvs is sufficient for WT behavior, and only knockdown in both sets of neurons is able to reproduce the pdf01 mutant phenotype. In long photoperiods, however, PDF from the lLNvs is necessary and sufficient for proper entrainment, whereas the sLNvs do not contribute to E peak timing. The data therefore point to a profound circuit switch in response to photoperiod, analogous to the neurotransmitter switching that occurs in the mammalian paraventricular nucleus in response to long photoperiods (Schlichting, 2019a).
A similar circuit reorganization might also occur in the principal mammalian brain clock neuron location, the suprachiasmatic nucleus (SCN). Light information from the visual system is transferred to cells in the ventral part of the SCN, which expresses VIP. VIP functions similarly to Drosophila PDF and is not only important for communication between different parts of the SCN but also essential for seasonal encoding. This is because VIP knockout mice show no change in peak width as measured by in vivo electrophysiological recordings in response to entrainment to different photoperiods]. This suggests that VIP is not only involved in relaying light information beyond the ventral SCN but also in the response to light duration as shown in this study for PDF in Drosophila. It will be interesting to see whether different VIP-expressing SCN neurons are involved in this response (Schlichting, 2019a).
In Drosophila, the clock that controls rest-activity rhythms synchronizes with light-dark cycles through either the blue-light sensitive Cryptochrome (Cry) located in most clock neurons, or rhodopsin-expressing histaminergic photoreceptors. This study shows that, in the absence of Cry, each of the two histamine receptors Ort and HisCl1 contribute to entrain the clock whereas no entrainment occurs in the absence of the two receptors. In contrast to Ort, HisCl1 does not restore entrainment when expressed in the optic lobe interneurons. Indeed, HisCl1 is expressed in wild-type photoreceptors and entrainment is strongly impaired in flies with photoreceptors mutant for HisCl1. Rescuing HisCl1 expression in the Rh6-expressing photoreceptors restores entrainment but it does not in other photoreceptors, which send histaminergic inputs to Rh6-expressing photoreceptors. The results thus show that Rh6-expressing neurons contribute to circadian entrainment as both photoreceptors and interneurons, recalling the dual function of melanopsin-expressing ganglion cells in the mammalian retina (Schlichting, 2019a).
Circadian rhythms in behavior and physiology are produced by central brain clock neurons that can be divided into subpopulations based on molecular and functional characteristics. It has become clear that coherent behavioral rhythms result from the coordinated action of these clock neuron populations, but many questions remain regarding the organizational logic of the clock network. This study used targeted genetic tools in Drosophila to eliminate either molecular clock function or neuronal activity in discrete clock neuron subsets. Neuronal firing was found to be necessary across multiple clock cell populations to produce free-running rhythms of rest and activity. In contrast, such rhythms are much more subtly affected by molecular clock suppression in the same cells. These findings demonstrate that network connectivity can compensate for a lack of molecular oscillations within subsets of clock cells. It was further shown that small ventrolateral (sLNv) clock neurons, which have been characterized as master pacemakers under free-running conditions, cannot drive rhythms independent of communication between other cells of the clock network. In particular, an essential contribution of the dorsolateral (LNd) clock neurons was pinpointed and manipulations that affect LNd function were shown to reduce circadian rhythm strength without affecting molecular cycling in sLNv cells. These results suggest a hierarchical organization in which circadian information is first consolidated among one or more clock cell populations before accessing output pathways that control locomotor activity (Bulthuis, 2019).
Sleep is a behavior that exists broadly across animal phyla, from flies to humans, and is necessary for normal brain function. Recent studies in both vertebrates and invertebrates have suggested a role for glial cells in sleep regulatory processes. Changes in neural-glial interactions have been shown to be critical for synaptic plasticity and circuit function. This study tested the hypothesis that changes in sleep pressure alters neural-glial interactions. In the fruit fly, Drosophila melanogaster, sleep is known to be regulated by mushroom body (MB) circuits. The technique GFP Reconstitution Across Synaptic Partners (GRASP) was used to test whether changes in sleep pressure affect neural-glial interactions between MB neurons and astrocytes, a specialized glial cell type known to regulate sleep in flies and mammals. The MB-astrocyte GRASP signal was reduced after 24 h of sleep deprivation, whereas the signal returned to baseline levels following 72 h of recovery. Social enrichment, which increases sleep drive, similarly reduced the MB-astrocyte GRASP signal. No changes were observed in the MB-astrocyte GRASP signal over time-of-day, or following paraquat exposure or starvation. These data suggest that changes in sleep pressure are linked to dynamic changes in neural-glial interactions between astrocytes and neuronal sleep circuits, which are not caused by normal rest-activity cycles or stressors (Vanderheyden, 2019).
Circadian rhythms allow animals to coordinate behavioral and physiological processes with respect to one another and to synchronize these processes to external environmental cycles. In most animals, circadian rhythms are produced by core clock neurons in the brain that generate and transmit time-of-day signals to downstream tissues, driving overt rhythms. The neuronal pathways controlling clock outputs, however, are not well understood. Furthermore, it is unclear how the central clock modulates multiple distinct circadian outputs. Identifying the cellular components and neuronal circuitry underlying circadian regulation is increasingly recognized as a critical step in the effort to address health pathologies linked to circadian disruption, including heart disease and metabolic disorders. Building on the conserved components of circadian and metabolic systems in mammals and Drosophila melanogaster, this study used a recently developed feeding monitor to characterize the contribution to circadian feeding rhythms of two key neuronal populations in the Drosophila pars intercerebralis (PI; the central neuroendocrine system), which is functionally homologous to the mammalian hypothalamus. Thermogenetic manipulations of PI neurons expressing the neuropeptide SIFamide (SIFa) as well as mutations of the SIFa gene degrade feeding:fasting rhythms. In contrast, manipulations of a nearby population of PI neurons that express the Drosophila insulin-like peptides (DILPs) affect total food consumption but leave feeding rhythms intact. The distinct contribution of these two PI cell populations to feeding is accompanied by vastly different neuronal connectivity as determined by trans-Tango synaptic mapping. These results for the first time identify a non-clock cell neuronal population in Drosophila that regulates feeding rhythms and furthermore demonstrate dissociable control of circadian and homeostatic aspects of feeding regulation by molecularly-defined neurons in a putative circadian output hub (Dreyer, 2019).
At its core, the circadian system is made up of central clock neurons in the brain that keep time through the presence of cell-autonomous molecular clocks. To enact behavioral rhythms, these clock cells must be connected through output pathways to downstream neuronal populations that directly control behavioral outputs; therefore, a complete understanding of circadian regulation of behavior depends on the delineation of output circuitry. This study identified a population of SIFa+ neurons in the pars intercerebralis that comprises part of the output pathway controlling feeding:fasting rhythms in flies. Constitutive activation of these cells strongly compromises normal patterns of feeding behavior, including producing a substantial percentage of flies that feed arrhythmically. This study also pinpointed a specific contribution of SIFa peptide to feeding rhythms, as SIFa mutant and RNAi knockdown lines show similar reductions of feeding rhythm strength (Dreyer, 2019).
The identification of a neuronal population and associated signaling molecule for the control of feeding:fasting rhythms should facilitate future studies aimed at further dissecting feeding output circuits, with the ultimate aim of tracing the pathway to motor neurons that directly control feeding. To that end, the trans-Tango analysis demonstrated that many neurons throughout the brain are postsynaptic to SIFa+ PI cells, including in areas such as the AL, which is involved in olfactory processing, and the SEZ, which is involved in gustatory processing and also contains feeding-related motor neurons. It will be of interest to more definitively determine the functional and neurochemical identity of postsynaptic neurons and to assess whether manipulations of SIFa receptor expression in these putative downstream output cells can recapitulate the feeding phenotypes observed following SIFa+ cell manipulations (Dreyer, 2019).
A role for SIFa in feeding regulation is supported by a recent study that demonstrated that SIFa modulates olfactory processing under conditions of starvation. Flies normally show sensitized AL projection neuron responses to food odors following starvation, however, this sensitization is absent in flies in which SIFa expression has been reduced through RNAi mechanisms. Martelli (2017) also showed that SIFa+ cells exhibit increased activity in response to starvation, and that thermogenetic activation of SIFa+ cells increases food consumption in satiated flies. Their experiments suggest that SIFa tunes sensory responsiveness to food cues according to the energy status of the fly, which subsequently increases feeding propensity in energy-depleted states. The described effects in this study identify an additional function of SIFa in dictating temporal patterns of feeding (Dreyer, 2019).
Interestingly, although the current findings of increased food consumption following SIFa+ cell activation are in line with those of Martelli (2017), this study found that feeding amount was also elevated in SIFa mutant flies, which is not predicted by a model in which SIFa peptide solely serves to increase appetitive and feeding behavior. This suggests that the exact nature of the regulation of feeding by SIFa is complex and may vary depending on environmental conditions and internal state. It is unclear why food consumption would be similarly affected by manipulations that eliminate SIFa peptide and those that hyperactivate SIFa+ cells, which should result in heightened SIFa signaling. One possibility is that constitutive SIFa+ cell activity could ultimately deplete SIFa stores, thus mimicking the SIFa mutant phenotype. Alternatively, feeding phenotypes may be affected differentially by SIFa mutations, which are present throughout development, compared to adult-specific thermogenetic activation. Regardless of whether acute SIFa signaling stimulates or inhibits food consumption, the fact that SIFa+ cell activation and reduction of SIFa signaling via mutations, cell ablation, or RNAi knockdown consistently degrade feeding:fasting rhythms provides strong evidence for a central contribution to the determination of the timing of feeding (Dreyer, 2019).
Flies in which SIFa+ cells are constitutively activated or that lack SIFa peptide due to cellular ablation or mutation also exhibit significantly reduced rest:activity rhythms, which is consistent with previous results demonstrating weakened locomotor rhythms following ablation of these cells. This effect was most pronounced in SIFa>reaper flies, indicating a potential for additional neurotransmitters emanating from SIFa+ cells to contribute to the regulation of locomotor activity. The overlap of feeding:fasting and rest:activity disruption raises the question of whether SIFa cells independently regulate feeding and locomotor rhythms, or whether one of these is indirectly affected secondary to changes in the other. Feeding and locomotor activity are interconnected behaviors that usually coincide, as animals primarily feed during their active phase. Nevertheless, feeding and locomotor rhythms can be dissociated in both flies and mammals. For example, adipocyte-specific knockout of the mammalian clock gene Arntl attenuates feeding rhythms in mice while leaving rest:activity rhythms intact, and mutations in mammalian per1 and per2 genes have differential effects on the phasing of locomotor and feeding rhythms. A similar phenotype has been noted in flies, as cell-specific abrogation of the molecular clock in the Drosophila fat body, a peripheral metabolic tissue, selectively alters the phase and magnitude of feeding rhythms without changing cycles of rest and activity (Xu, 2008). More recently, it was shown that manipulations that downregulate DH44 signaling or silence neurons expressing the hugin peptide significantly degrade rest:activity rhythm strength in DD conditions but leave the strength of DD feeding:fasting intact. Taken together, these results confirm that feeding:fasting rhythms are under de facto circadian control and do not simply occur secondary to rest:activity rhythms. Because locomotor rhythm disruption can occur independent of changes in feeding behavior, it is concluded that the effects of the SIFa manipulations likely reflect direct feeding:fasting rhythm regulation (Dreyer, 2019).
In addition to affecting rest:activity and feeding:fasting rhythms, adult-specific SIFa+ cell manipulations also resulted in high lethality, particularly in the case of adult-specific neuronal silencing. This suggests that SIFa+ cells perform some necessary function in the adult animal, though it seems that SIFa peptide itself is dispensable for survival, as mutants eclose at expected Mendelian ratios. Intriguingly, it was found that a substantial number of flies eclosed from genetic crosses that result in SIFa+ cell ablation during developmental stages due to expression of the apoptotic gene reaper. The lack of a lethality phenotype in SIFa+ ablated flies is perhaps due to compensatory changes in these flies that are not present following adult-specific manipulations. It is unclear whether the lethality phenotype is related to alterations in feeding behavior following SIFa+ cell manipulations, or whether other, yet unidentified contributions of SIFa+ cells are responsible, but as there is little evidence for SIFa expression in cells outside of the PI, it is likely that the phenotype stems from dysregulation of these cells (Dreyer, 2019).
Together with previous findings, the current results add to a growing understanding of the PI in the control of circadian outputs. The PI is situated in a region of the Drosophila brain that is near the axon terminals of multiple groups of core clock cells, and previous work has shown anatomical and functional connections between clock cells and multiple PI populations, including those expressing DH44, SIFa and DILPs. These clock cell inputs could allow PI cells, which lack molecular clocks, to transmit circadian information to downstream output regions. Interestingly, the PI cell populations appear to differentially contribute to circadian outputs. As detailed above, DH44+ cells selectively regulate rest:activity rhythms while SIFa+ cells contribute to both rest:activity and feeding:fasting rhythms. DILP+ PI cells contribute to neither behavioral rhythm but instead have been shown to modulate circadian gene expression in the fat body. These results support the hypothesis that the PI is a circadian output hub that channels core clock input into anatomically distinct output pathways to coordinately regulate different circadian outputs (Dreyer, 2019).
Though no effect of DILP+ cell manipulations on feeding:fasting rhythms were observed, changes in overall food intake following IPC activation were observed in two independent assays, which is consistent with a homeostatic role for these cells. The IPCs receive feedback from a range of circulating peptides and are also indirectly targeted by satiety signals secreted from the fat body integrating information regarding the nutritional status of a fly as one component of the intricate regulation of energy homeostasis. DILP+ neurons have also recently been shown to play a role in nutrient sensing in female flies, contributing to the modulation of reproductive dormancy by affecting overall feeding and maintaining females in a metabolically active state. Generally, IPC neuronal activity is regulated by feeding status, as the cells are more active in the fed versus starved state, which likely results in increased DILP secretion in fed flies. In turn, insulin/IGF signaling (IIS) is an integral regulator of growth and development and affects a range of physiological attributes including metabolism, reproduction, stress response, and aging (Dreyer, 2019).
DILPs have also been directly implicated in regulating feeding behavior, with several studies demonstrating anorexigenic effects of increased DILP signaling, as well as of drosulfakinin peptides, which are an additional output of the IPCs and act as a satiety signal. These effects are in line with evidence demonstrating increased activation of IPCs and release of DILPs in the fed state. In contrast, it has also been shown that DILP+ cell silencing can result in hypophagia, as indicated by reduced fecal output, and that thermogenetic DILP+ cell activation can either stimulate or inhibit feeding depending on metabolic status. Thus, the role of the DILP+ PI cells in determining overall food consumption, similar to SIFa peptides, is likely complex. Given this, the finding of increased feeding following IPC stimulation, though counterintuitive, is not without precedent, and may occur as a result of an interaction between diet type, insulin signaling, and the metabolic condition of the flies, especially as they are exposed to a carbohydrate-only diet in the FLIC and CAFE assays. Alternatively, increased feeding could occur if DILPs are depleted by extended IPC activation. This possibility could be directly tested using recently-developed DILP2 reporter flies, which allow for sensitive measurements of circulating DILP2 levels (Dreyer, 2019).
The contrasting effects of DILP+ and SIFa+ PI cell activation demonstrate dissociable control over homeostatic and circadian regulation of feeding by these two populations of PI cells. The results of the trans-Tango analyses provide a potential anatomical basis for this and suggest that DILP+ and SIFa+ PI cells rely on different signaling paradigms. Given their limited connectivity to other brain regions, the IPCs likely release DILPs systemically to act on target tissues, including the brain, via long-distance diffusion through the hemolymph. In contrast, SIFamidergic cells appear to act via direct synaptic connections to impact widespread brain areas. The differences in kinetics between these two signaling mechanisms could underlie the functional differences of these cell populations with respect to feeding regulation, with IPC activity reflecting overall energy status and therefore controlling homeostatic aspects of feeding, and SIFa cells regulating moment-to-moment feeding decisions and therefore controlling circadian patterns of feeding. In addition, the downstream connections of SIFa+ cells to central clock neurons, including l-LNvs and s-LNvs as well as LNds, implicates SIFa+ cells in feedback control of the central clock. Previous work found no alterations in central clock timing following SIFa+ cell ablation; however, as SIFa+ cells appear to lie at a crossroads of energetic signaling, it follows that they would have the capacity to relay that information back to the core clock and affect behavioral changes that are attuned to the circadian patterns of activity as necessary, perhaps under conditions in which food access is limited (Dreyer, 2019).
Research into metabolic and feeding control continues to uncover a dense web of interconnected regulators, indicative of how integral proper nutrient signaling is to overall organismal health. The partial reduction of feeding rhythm strength ascribed to SIFa here leaves room for the discovery of additional signals affecting circadian feeding rhythms. Two promising neuropeptides that have been shown to affect feeding behaviors are short neuropeptide F and allatostatin A, both of which also have been associated with sleep regulation in flies. Characterizing the complete output circuit of circadian feeding behavior in flies will help identify the most important contributors to synchronized feeding patterns and increase understanding of the profound metabolic consequences of circadian disruption (Dreyer, 2019).
Animal circadian rhythms persist in constant darkness and are driven by intracellular transcription-translation feedback loops. Although these cellular oscillators communicate, isolated mammalian cellular clocks continue to tick away in darkness without intercellular communication. To investigate these issues in Drosophila, behavior as well as molecular rhythms were assayed within individual brain clock neurons while blocking communication within the ca. 150 neuron clock network. CRISPR-mediated neuron-specific circadian clock knockouts were also generated. The results point to two key clock neuron groups: loss of the clock within both regions but neither one alone has a strong behavioral phenotype in darkness; communication between these regions also contributes to circadian period determination. Under these dark conditions, the clock within one region persists without network communication. The clock within the famous PDF-expressing s-LNv neurons however was strongly dependent on network communication, likely because clock gene expression within these vulnerable sLNvs depends on neuronal firing or light (Schlichting, 2019b).
Neuronal networks make myriad contributions to behavior and physiology. By definition, individual neurons within a network interact, and different networks also interact to coordinate specialized functions. For example, the visual cortex and motor output centers must coordinate to react properly to environmental changes. In a less immediate fashion, sleep centers and circadian clocks are intertwined to properly orchestrate animal physiology. The brain clock is of special interest: it not only times and coordinates physiology within neuronal tissues but also sends signals to the body to keep the entire organism in sync with the cycling external environment (Schlichting, 2019b).
The small, circumscribed Drosophila clock network is ideal to address circadian communication issues. The comparable region in mammals, the suprachiasmatic nucleus, is composed of thousands of cells depending on the species. There are in contrast only 75 clock neurons per hemisphere in Drosophila. These different clock neurons can be divided into several subgroups according to their location within the fly brain. There are 4 lateral and three dorsal neuron clusters, which have different functions in controlling fly physiology (Schlichting, 2019b).
The four small ventro-lateral neurons (sLNvs) are arguably the most important of the 75 clock neurons. This is because ablating or silencing these neurons abolishes rhythms in constant darkness (DD). They reside in the accessory medulla region of the fly brain, an important pacemaker center in many insects, and express the neuropeptide PDF. In addition, they are essential for predicting dawn. A very recent study suggests that the sLNvs are also able to modulate the timing of the evening (E) peak of behavior via PDF. The other ventral-lateral group, the four large-ventro-lateral neurons (lLNvs), also express PDF and send projections to the medulla, the visual center of the fly brain; they are important arousal neurons. Consistent with the ablation experiments mentioned above, the absence of pdf function or reducing PDF levels via RNAi causes substantial arrhythmic behavior in DD (Schlichting, 2019b).
Other important clock neurons include the dorso-lateral neurons (LNds), which are essential for the timing of the E peak and adjustment to long photoperiods. Two other clock neuron groups, the lateral-posterior neurons (LPN and a subset of the dorsal neurons (DN1s), were recently shown to connect the clock network to sleep centers in the fly central complex. The DN2 neurons are essential for temperature preference rhythms, whereas no function has so far been assigned to the DN3s (Schlichting, 2019b).
Despite these distinct functions, individual clock neuron groups are well-connected to each other. At the anatomical level, all lateral neuron clusters and even DN1 dorsal neurons send some of their projections into the accessory medulla, where they can interact. A second area of common interaction is the dorsal brain; only the lLNvs do not project there (Schlichting, 2019b).
Several studies have investigated interactions between different clock neurons. Artificially expressing kinases within specific clock neurons causes their clocks to run fast or slow and also changes the overall free-running period of the fly, indicating that network signaling adjusts behavior. Similarly, speeding up or slowing down individual neurons is able to differentially affect behavioral timing in standard light-dark (LD) cycles. A high level of neuronal plasticity within the network also exists: axons of individual cells undergo daily oscillations in their morphology, and neurons change their targets depending on the environmental condition (Schlichting, 2019b).
How neuronal communication influences the fly core feedback loop is not well understood. The latter consists of several interlocked transcriptional-translational feedback loops, which probably underlie rhythms in behavior and physiology. A simplified version of the core feedback loop consists of the transcriptional activators Clock (CLK) and Cycle (CYC) and the transcriptional repressors Period (PER) and Timeless (TIM). CLK and CYC bind to E-boxes within the period (per) and timeless (tim) genes (among other clock-controlled genes) and activate their transcription. After PER and TIM synthesis in the cytoplasm, they form a heterodimer and enter the nucleus toward the end of the night. There they interact with CLK and CYC, release them from their E-box targets and thereby inhibit their own transcription. All 75 pairs of clock neurons contain this canonical circadian machinery, which undergoes daily oscillations in level. Indeed, the immunohistochemical cycling of PER and TIM within these neurons is a classic assay to visualize these molecular oscillations (Schlichting, 2019b).
Silencing PDF neurons stops their PER cycling, indicating an important role of neuronal firing in maintaining circadian oscillations. However, only two time points were measured, and the results were possibly confounded by developmental effects. PDF neuron silencing also phase advances PER cycling in downstream neurons, suggesting that PDF normally serves to delay cycling in target neurons. This is consistent with experiments showing that PDF signaling stabilizes PER. In addition, neuronal activation is able to mimic a light pulse and phase shift the clock due to firing-mediated TIM degradation (Schlichting, 2019b).
To investigate more general features of clock neuron interactions on the circadian machinery, the majority of the fly brain clock neurons were silenced, and behavior and clock protein cycling within the circadian network was examined in a standard light-dark cycle (LD) as well as in constant darkness (DD). Silencing abolished rhythmic behavior but had no effect on clock protein cycling in LD, indicating that the silencing affects circadian output but not oscillator function in a cycling light environment. Silencing similarly abolished rhythmic behavior in DD but with very different effects on clock protein cycling. Although protein cycling in the LNds was not affected by neuronal silencing in DD, the sLNvs dampened almost immediately. Interestingly, this differential effect is under transcriptional control, suggesting that some Drosophila clock neurons experience activity-regulated clock gene transcription. Cell-specific CRISPR/Cas9 knockouts of the core clock protein Per further suggests that network properties are critical to maintain wild-type activity-rest rhythms. The data taken together show that clock neuron communication and firing-mediated clock gene transcription are essential for high amplitude and synchronized molecular rhythms as well as rhythmic physiology (Schlichting, 2019b).
The central clock of animals is essential for dictating the myriad diurnal changes in physiology and behavior. Knocking out core clock components such as period or Clock severely disrupts circadian behavior as well as molecular clock properties in flies and mammals. This study show that similar behavioral effects occur when the central clock neurons are silenced and thereby abolish communication within this network and with downstream targets, that is fly behavior becomes arrhythmic in LD as well as DD conditions and resembles the phenotypes of core clock mutant strains (Schlichting, 2019b).
Despite the loss of all rhythmic behavior, silencing did not impact the molecular machinery in LD conditions: PER and PDP1 protein cycling was normal. These findings suggest that (1) rhythmic behavior requires clock neuron output, which is uncoupled from the circadian molecular machinery by network silencing, and (2) synchronized molecular rhythms of clock neurons do not require neuronal activity. These findings are in agreement with previous work showing that silencing the PDF neurons had no effect on Per cycling within these neurons. The results presumably reflect the strong effect of the external light-dark cycle on these oscillators (Schlichting, 2019b).
In DD however, the individual neurons change dramatically: the different neurons desynchronize, and their protein cycling damps to different extents. Interestingly, sLNv cycling relies most strongly on neuronal communication: these neurons cycle robustly in controls but apparently not at all in the silenced state. sLNvs were previously shown to be essential for DD rhythms. Unfortunately, the sensitivity of immunohistochemistry precludes determining whether the molecular clock has actually stopped or whether silencing has only (dramatically) reduced cycling amplitude. However, a simple interpretation of the adult-specific silencing experiment favors a stopped clock: decreasing the temperature to 18 degrees after a week at high temperature failed to rescue rhythmic behavior. A similar experiment in mammals gave rise to the opposite result, suggesting an effect of firing on circadian amplitude in that case. However, different explanations cannot be excluded, for example chronic effects of neuronal silencing or a too large phase difference between the different neuronal subgroups to reverse after a week without communication (Schlichting, 2019b).
In either case, a stopped clock or an effect on clock protein oscillation amplitude, these results make another link to the mammalian literature: modeling of the clock network suggests that different neurons resynchronize more easily if the most highly-connected cells are intrinsically weak oscillators. The sLNvs are essential for DD rhythms, known to communicate with other clock neurons and are situated in the accessory medulla; this is an area of extensive neuronal interactions in many insects. These considerations rationalize weak sLNv oscillators (Schlichting, 2019b).
An important role of interneuron communication in DD is in agreement with previous work showing that altering the speed of individual neuron groups can change the phase of downstream target neurons. An important signaling molecule is the neuropeptide PDF: its absence changes the phase of downstream target neurons, and silencing PDF neurons causes an essentially identical phenotype to the lack of PDF. However, the effects reported in this study are much stronger and show different levels of autonomy than PDF ablation, suggesting that other signaling molecules and/or the neuronal activity of additional clock neurons are essential to maintain proper rhythmic clock protein expression (Schlichting, 2019b).
To address these possibilities, this study took two approaches. First, clock gene RNA levels were investigated after silencing. The goal was to assess whether the damping of silenced neurons is under gene expression control, likely transcriptional control. Indeed, tim mRNA profiles nicely reproduced the protein cycling profiles: robust cycling of all (assayed) clock neurons was maintained in LD even with silencing, but tim-mRNA levels in the sLNvs stopped cycling in DD; in contrast, robust cycling was maintained in the LNds. This suggests that the changes in protein cycling amplitude and also possibly phase are under transcriptional control. Importantly, the tim signal in the sLNvs disappeared upon silencing, suggesting that neuronal activity promotes clock gene transcription at least in this subset of neurons. This recapitulates for the first time in Drosophila the robust positive relationship between neuronal firing and clock gene transcription in mammals. To date, Drosophila neuronal firing had only been connected to post-transcriptional clock protein regulation, namely TIM degradation. Conceivably, these two effects are connected: TIM degradation might be required to relieve transcriptional repression and maintain cycling (Schlichting, 2019b).
The second approach was a cell-specific knockout strategy, applied to the clock neuron network. Three guides were generated targeting the CDS of per and also CAS9 was expressed in a cell-specific manner. The guides caused double strand breaks in the per gene, which in turn led to cell-specific per mutations. This adult brain knockout strategy worked reliably and specifically, in glial cells as well as neurons, with high efficiency and with no apparent background effects. This strategy was successfully used to knock out most if not all Drosophila GPCRs and it is believed to be superior to RNAi for most purposes. Importantly, expression of the guides with the clk856-GAL4 driver phenocopied per01 behavior. To focus on individual clock neurons, cell-specific knockouts were generated in different clock neurons. PERKO in the PDF cells did not increase the level of arrhythmicity, only a PERKO in most lateral neurons, E cells as well as PDF cells, generated high levels of arrhythmic behavior (Schlichting, 2019b).
Both the structure and the amount of sleep are important for brain function. Entry into deep, restorative stages of sleep is time dependent; short sleep bouts selectively eliminate these states. Fragmentation-induced cognitive dysfunction is a feature of many common human sleep pathologies. Whether sleep structure is normally regulated independent of the amount of sleep is unknown. This study shows that in Drosophila melanogaster, activation of a subset of serotonergic neurons fragments sleep without major changes in the total amount of sleep, dramatically reducing long episodes that may correspond to deep sleep states. Disruption of sleep structure results in learning deficits that can be rescued by pharmacologically or genetically consolidating sleep. Two reciprocally connected sets of ellipsoid body neurons were identified that form the heart of a serotonin-modulated circuit that controls sleep architecture. Taken together, these findings define a circuit essential for controlling the structure of sleep independent of its amount (Liu, 2019).
This study describes a circuit that regulates of sleep structure without affecting the total amount of sleep. 5HT acts to enhance the response of 5HT7-GAL4+ neurons to basally active excitatory inputs. 5HT-dependent calcium signals are blocked by TTX, while its ability to increase cAMP is not, supporting the existence of these active excitatory inputs to 5HT7-GAL4+ cells. In contrast, VT-GAL4+ cells do not have basally active excitatory inputs. 5HT modulation of the circuit likely occurs primarily via inputs to 5HT7-GAL4+ neurons since the response of VT038828-GAL4 (VT-GAL4)+ neurons is weaker and lower affinity. Whether there are other, perhaps situationally active, inputs to this circuit is currently unknown (Liu, 2019).
Within the ellipsoid body (EB) the circuit is complex. VT-GAL4+ neurons are functionally connected with the 5HT7-GAL4+ group. VT-GAL4+ neurons provide feedback inhibition to a subset of 5HT7-GAL4+ neurons, which enhances fragmentation, likely via output to non-central complex regions. How inhibition of a subset of the 5HT7-GAL4+ cells acts to modulate the behavioral output of the rest of the population is not yet clear, but it is noted that many of the 5HT7-GAL4+ cells are GABAergic. While all the details of the circuit's complex dynamics remain to be discovered, it is clear that this circuit has a profound and selective effect on sleep architecture (Liu, 2019).
The circuit described in this study is modulated by 5HT, a neurochemical known to be important for regulation of behavioral states in many species. While 5HT in mammals is important in a wide variety of contexts, it was controversial for nearly half a century whether it promoted sleep or wakefulness. In Drosophila, 5HT has only been thought to promote sleep. The current data show that upregulation of serotonergic signaling can also induce sleep fragmentation, suggesting that 5HT's role in sleep in flies exhibits a complexity similar to that of its roles in mammals. The genesis of this apparently conserved complexity may be the extensive involvement of 5HT in non-sleep processes. For an animal in the wild, sleep has inherent risks: predation and loss of opportunities for mating or feeding are just a few. Sleep/wake systems in the brain must control arousal state in collaboration with systems that assess competing needs. 5HT, because it is central to so many critical behavioral circuits, is ideally poised to be an integration point for sleep and the general state of the animal. The diverse, circuit-specific, roles in sleep that 5HT exhibits across phyla may be a result of its ubiquity (Liu, 2019).
The role this study has uncovered for 5HT as a regulator of sleep architecture aligns well with this idea. The daily neuronal activity profile reported by Tric-LUC, a calcium sensor that drives luciferase expression in response to neuronal activity, in sleep fragmentation-generating neurons maps to dawn and dusk, when crepuscular organisms such as fruit flies are most active. Fragmentation of sleep at these times would presumably be beneficial since flies would not enter into deep sleep states at times when they should be feeding and mating. Interestingly, the circuit described in this study accomplishes this feat by increasing P(doze), the probability of falling asleep from a wake state, leaving the scaling of P(wake), a parameter associated with dopamine and arousal, free to be modulated by other factors (e.g., danger from predation, appearance of potential mates). The fact that long sleep bouts can be prevented without putting the animal into a hyperaroused state is advantageous, allowing flexible responsiveness to changing conditions. The involvement of P(Doze), a parameter associated with sleep drive, is also congruent with the sleep-promoting role of 5HT in other brain regions (Liu, 2019).
While controlled sleep fragmentation appears to assist in active period behavior, there is also a need for consolidated sleep. In both mammals and Drosophila, sleep has electrophysiologically distinct substrates with progressively higher arousal thresholds that appear in an ordered fashion during a sleep episode. The deeper sleep stages in mammals, REM and slow wave sleep, are strongly associated with maintenance of cognitive function. Fragmentation of sleep, because it truncates sleep episodes before deeper stages are reached, can result in a selective deprivation of deep sleep stages even when total sleep is not changed. In this study, it was demonstrated that decreasing sleep consolidation, without changing the amount of sleep, can disrupt associative learning. These results suggest that in Drosophila, like in mammals, there are time-dependent changes in the depth of sleep that are important for its beneficial effects. This idea is also supported by modeling and analysis of the structure of fly sleep, which indicate that there are time-dependent changes in the probability of sleep-wake transitions consistent with the existence of deep sleep stages that are only accessed in long sleep episodes (Liu, 2019).
Fragmentation of sleep induced by activation of 5HT inputs to the EB also produced an increase in sleep after the activation was terminated. Excess sleep in the recovery day after a perturbation is a hallmark of a homeostatic process. Homeostatic regulation of total sleep time has been previously demonstrated in Drosophila, but the data suggest that there is also homeostatic regulation of sleep quality. In mammals, individual sleep substates have been demonstrated to be homeostatically regulated- selective deprivation of REM or slow wave sleep, in the absence of loss of total sleep time, drive rebound increases of the deprived stage and mechanical sleep fragmentation has been shown to lead to an increase in total sleep. The ability of the EB circuit in Drosophila to selectively modulate sleep structure, without changing the total amount of sleep, has allowed for the first time the selective probing of the cognitive importance of long sleep bouts and deep sleep stages in the fly. The fact that fragmentation triggers rebound sleep implies that these long sleep bouts may also be important for the general health benefits of sleep (Liu, 2019).
The Cambridge Protein Trap resource (CPTI) was used to screen for flies whose locomotor rhythms are rhythmic in constant light (LL) as a means of identifying circadian photoreception genes. From the screen of approximately 150 CPTI lines, seven hits were obtained, two of which targeted the glutamate pathway, Got1 (Glutamate oxaloacetate transaminase 1) and Gs2 (Glutamine synthetase 2). The genetic background was important with a strong interaction observed with the common and naturally occurring timeless (tim) polymorphisms, ls-tim and s-tim. The less circadian photosensitive ls-tim allele generated high levels of LL rhythmicity in combination with Got1 or Gs2, even though ls-tim and s-tim alleles do not, by themselves, generate the LL phenotype. The use of dsRNAi for both genes as well as for Gad (Glutamic acid decarboxylase) and the metabotropic glutamate receptor DmGluRA driven by clock gene promoters also revealed high levels of LL rhythmicity compared to controls. It is clear that the glutamate pathway is heavily implicated in circadian photoreception. TIM levels in Got1 and Gs2 mutants cycled and were more abundant than in controls under LL. Got1 but not Gs2 mutants showed diminished phase shifts to 10 min light pulses. Neurogenetic dissection of the LL rhythmic phenotype using the gal4/gal80 UAS bipartite system suggested that the more dorsal CRY-negative clock neurons, DNs and LNds were responsible for the LL phenotype. Immunocytochemistry using the CPTI YFP tagged insertions for the two genes revealed that the DN1s but not the DN2 and DN3s expressed Got1 and Gs2, but expression was also observed in the lateral neurons, the LNds and s-LNvs. These results suggest a model whereby the DN1s and possibly CRY-negative LNds use glutamate signaling to suppress the pacemaker s-LNvs in LL (de Azevedo, 2020).
Networks of circadian timekeeping in the brain display marked daily changes in neuronal morphology. In Drosophila melanogaster, the striking daily structural remodeling of the dorsal medial termini of the small ventral lateral neurons has long been hypothesized to mediate endogenous circadian timekeeping. To test this model, this study has specifically abrogated these sites of daily neuronal remodeling through the reprogramming of neural development and assessed the effects on circadian timekeeping and clock outputs. Remarkably, the loss of these sites has no measurable effects on endogenous circadian timekeeping or on any of the major output functions of the small ventral lateral neurons. Rather, their loss reduces sites of glutamatergic sensory neurotransmission that normally encodes naturalistic time cues from the environment. These results support an alternative model: structural plasticity in critical clock neurons is the basis for proper integration of light and temperature and gates sensory inputs into circadian clock neuron networks (Fernandez, 2020).
The daily changes of light and dark exemplify a prominent cue for the synchronization of circadian clocks with the environment. The match between external and internal time is crucial for the fitness of organisms, and desynchronization has been linked to numerous physical and mental health problems. Organisms therefore developed complex and not fully understood mechanisms to synchronize their circadian clock to light. In mammals and in Drosophila, both the visual system and non-image-forming photoreceptors contribute to circadian clock resetting. In Drosophila, light-dependent degradation of the clock protein TIMELESS by the blue light photoreceptor Cryptochrome is considered the main mechanism for clock synchronization, although the visual system also contributes. To better understand the visual system contribution, a genetic variant exhibiting extremely slow phototransduction kinetics, yet normal sensitivity, was generated. In this variant, the visual system is able to contribute its full share to circadian clock entrainment, both with regard to behavioral and molecular light synchronization. This function depends on an alternative phospholipase C-β enzyme, encoded by PLC21C, presumably playing a dedicated role in clock resetting. This pathway requires the ubiquitin ligase CULLIN-3, possibly mediating CRY-independent degradation of TIMELESS during light:dark cycles. These results suggest that the PLC21C-mediated contribution to circadian clock entrainment operates on a drastically slower timescale compared with fast, norpA-dependent visual phototransduction. These findings are therefore consistent with the general idea that the visual system samples light over prolonged periods of time (h) in order to reliably synchronize their internal clocks with the external time (Ogueta, 2020).
Light is one of the most important factors regulating rhythmical behavior of Drosophila melanogaster. It is received by different photoreceptors and entrains the circadian clock, which controls sleep. The retina is known to be essential for light perception, as it is composed of specialized light-sensitive cells which transmit signal to deeper parts of the brain. This study examined the role of specific photoreceptor types and peripheral oscillators located in these cells in the regulation of sleep pattern. Sleep was shown to be controlled by the visual system in a very complex way. Photoreceptors expressing Rh1, Rh3 are involved in night-time sleep regulation, while cells expressing Rh5 and Rh6 affect sleep both during the day and night. Moreover, Hofbauer-Buchner (HB) eyelets which can directly contact with s-LN (v) s and l-LN (v) s play a wake-promoting function during the day. In addition, this study showed that L2 interneurons, which receive signal from R1-6, form direct synaptic contacts with l-LN (v) s, which provides new light input to the clock network (Damulewicz, 2020).
Sleep rebound - the increase in sleep that follows sleep deprivation
(SD) - is a hallmark of homeostatic sleep regulation. This, this study
developed an efficient thermogenetic method of inducing SD in
Drosophila that produces a strong rebound. To develop a thermogenetic
method of SD suitable for screening, different populations of
wake-promoting neurons labeled by Gal4 drivers were thermogenetically
stimulated. Thermogenetic activation of neurons marked by the
c584-Gal4 driver produces both strong sleep loss and a substantial
rebound that is more subject to genetic regulation and more consistent
within genotypes than rebound following mechanical or caffeine-induced
SD. This driver was then used to induce SD in a screen of 1,741
mutagenized lines generated by the Drosophila Gene Disruption Project.
Flies were subjected to 9 h of SD during the dark period and released
from SD 3 h before lights-on. Recovery was measured over the 15 h
following SD. Following identification of lines with reduced sleep
rebound, baseline sleep and sleep depth was characterized before and
after SD. Two lines were identified that consistently exhibit a
blunted increase in the duration and depth of sleep after
thermogenetic SD. Neither of the two genotypes has reduced total
baseline sleep. Statistical analysis across all screened lines shows
that genotype is a strong predictor of recovery sleep, independent
from effects of genotype on baseline sleep. These data show that
rebound sleep following thermogenetic SD can be genetically separated
from sleep at baseline. This suggests that genetically controlled
mechanisms of sleep regulation not manifest under undisturbed
conditions contribute to sleep rebound following thermogenetic SD
(Dubowy, 2016). Apterous (Ap), the best studied LIM-homeodomain transcription factor in Drosophila, cooperates with the cofactor Chip (Chi) to regulate transcription of specific target genes. Although Ap regulates various developmental processes, its function in the adult brain remains unclear. This study reports that Ap and Chi in the neurons expressing PDF, a neuropeptide, play important roles in proper sleep/wake regulation in adult flies. PDF-expressing neurons consist of two neuronal clusters: small ventral-lateral neurons (s-LNvs) acting as the circadian pacemaker and large ventral-lateral neurons (l-LNvs) regulating light-driven arousal. Ap localizes to the nuclei of s-LNvs and l-LNvs. In light-dark (LD) cycles, RNAi knockdown or the targeted expression of dominant-negative forms of Ap or Chi in PDF-expressing neurons or l-LNvs promoted arousal. In contrast, in constant darkness, knockdown of Ap in PDF-expressing neurons did not promote arousal, indicating that a reduced Ap function in PDF-expressing neurons promotes light-driven arousal. Furthermore, Ap expression in l-LNvs showed daily rhythms (peaking at midnight), which are generated by a direct light-dependent mechanism rather than by the endogenous clock. These results raise the possibility that the daily oscillation of Ap expression in l-LNvs may contribute to the buffering of light-driven arousal in wild-type flies (Shimada, 2016).
Many biological and behavioural processes of animals are governed by an endogenous circadian clock, which is dependent on transcriptional regulation. This study addresses post-transcriptional regulation and the role of miRNAs in Drosophila circadian rhythms. At least six miRNAs show cycling expression levels within the pigment dispersing factor (PDF) Like many other insects in temperate regions, Drosophila melanogaster exploits the photoperiod shortening that occurs during the autumn as an important cue to trigger a seasonal response. Flies survive the winter by entering a state of reproductive arrest (diapause), which drives the relocation of resources from reproduction to survival. This study profiled the expression of microRNA (miRNA) in long and short photoperiods and identified seven differentially expressed miRNAs (dme-mir-2b, dme-mir-11, dme-mir-34, dme-mir-274, dme-mir-184, dme-mir-184*, and dme-mir-285). Misexpression of dme-mir-2b, dme-mir-184, and dme-mir-274 in pigment-dispersing, factor-expressing neurons largely disrupted the normal photoperiodic response, suggesting that these miRNAs play functional roles in photoperiodic timing. This study also analyzed the targets of photoperiodic miRNA by both computational predication and by Argonaute-1-mediated immunoprecipitation of long- and short-day RNA samples. Together with global transcriptome profiling, these results expand existing data on other Drosophila species, identifying genes and pathways that are differentially regulated in different photoperiods and reproductive status. These data suggest that post-transcriptional regulation by miRNA is an important facet of photoperiodic timing (Pegoraro, 2022).
This study describes a genome-wide microRNA (miRNA)-based screen to identify brain glial cell functions required for circadian behavior. To identify glial miRNAs that regulate circadian rhythmicity, a collection of 'miR-sponges' was employed to inhibit miRNA function in a glia-specific manner. The initial screen identified 20 glial miRNAs that regulate circadian behavior. Two miRNAs, miR-263b and miR-274, were studied in detail; both function in adult astrocytes to regulate behavior. Astrocyte-specific inhibition of miR-263b or miR-274 in adults acutely impairs circadian locomotor activity rhythms with no effect on glial or clock neuronal cell viability. To identify potential RNA targets of miR-263b and miR-274, 35 predicted miRNA targets were screenexd, employing RNA interference-based approaches. Glial knockdown of two putative miR-274 targets, CG4328 and MESK2, resulted in significantly decreased rhythmicity. Homology of the miR-274 targets to mammalian counterparts suggests mechanisms that might be relevant for the glial regulation of rhythmicity (You, 2018).
Animals partition their daily activity rhythms through their internal circadian clocks, which are synchronized by oscillating day-night cycles of light. The fruitfly Drosophila melanogaster senses day-night cycles in part through rhodopsin-dependent light reception in the compound eye and photoreceptor cells in the Hofbauer-Buchner eyelet. A more noteworthy light entrainment pathway is mediated by central pacemaker neurons in the brain. The Drosophila circadian clock is extremely sensitive to light. However, the only known light sensor in pacemaker neurons, the flavoprotein Cryptochrome (Cry), responds only to high levels of light in vitro. These observations indicate that there is an additional light-sensing pathway in fly pacemaker neurons. This study describes a previously uncharacterized rhodopsin, Rh7, which contributes to circadian light entrainment by circadian pacemaker neurons in the brain. The pacemaker neurons respond to violet light, and this response depends on Rh7. Loss of either cry or rh7 caused minor defects in photoentrainment, whereas loss of both caused profound impairment. The circadian photoresponse to constant light was impaired in rh7 mutant flies, especially under dim light. The demonstration that Rh7 functions in circadian pacemaker neurons represents the first role for an opsin in the central brain (Ni, 2017).
The sodium leak channel Narrow abdomen (NA)/ NALCN is an important component of circadian pacemaker neuronal output. In Drosophila, rhythmic expression of the NA channel regulator Nlf-1 in a subset of adult pacemaker neurons has been proposed to contribute to circadian regulation of channel localization or activity. This study restricted expression of Drosophila NA channel subunits or the Nlf-1 regulator to either development or adulthood using the temperature-inducible tubulin-GAL80ts system. Surprisingly, it was found that developmental expression of endogenous channel subunits and Nlf-1 is sufficient to promote robust rhythmic behavior in adults. Moreover, channel complex proteins produced during development persist in the Drosophila head with little decay for at least 5-7 days in adults. In contrast, restricting either endogenous or transgenic gene expression to adult stages produces only limited amounts of the functional channel complex. These data indicate that much of the NA channel complex that functions in adult circadian neurons is normally produced during development, and that the channel complex is very stable in most neurons in the Drosophila brain. Based on these findings, it is proposed that circadian regulation of NA channel function in adult pacemaker neurons is mediated primarily by post-translational mechanisms that are independent of Nlf-1 (Moose, 2017).
The daily light-dark cycles represent a key signal for synchronizing circadian clocks. Both insects and mammals possess dedicated "circadian" photoreceptors but also utilize the visual system for clock resetting. In Drosophila, circadian clock resetting is achieved by the blue-light photoreceptor cryptochrome (CRY), which is expressed within subsets of the brain clock neurons. In addition, rhodopsin-expressing photoreceptor cells contribute to light synchronization. Light resets the molecular clock by CRY-dependent degradation of the clock protein Timeless (TIM), although in specific subsets of key circadian pacemaker neurons, including the small ventral lateral neurons (s-LNvs), TIM and Period (PER) oscillations can be synchronized by light independent of CRY and canonical visual Rhodopsin phototransduction. This study shows that at least three of the seven Drosophila rhodopsins can utilize an alternative transduction mechanism involving the same alpha-subunit of the heterotrimeric G protein operating in canonical visual phototransduction (Gq). Surprisingly, in mutants lacking the canonical phospholipase C-beta (PLC-beta) encoded by the no receptor potential A (norpA) gene, a novel transduction pathway using a different PLC-beta encoded by the Plc21C gene. This novel pathway is important for behavioral clock resetting to semi-natural light-dark cycles and mediates light-dependent molecular synchronization within the s-LNv clock neurons. The same pathway appears to be responsible for norpA-independent light responses in the compound eye. Rhodopsin 5 (Rh5) and Rh6, present in the R8 subset of retinal photoreceptor cells, drive both the long-term circadian and rapid light responses in the eye (Ogueta, 2018).
Circadian clocks impose daily periodicities to animal behavior and physiology. At their core, circadian rhythms are produced by intracellular transcriptional/translational feedback loops (TTFL). TTFLs may be altered by extracellular signals whose actions are mediated intracellularly by calcium and cAMP. In mammals these messengers act directly on TTFLs via the calcium/cAMP-dependent transcription factor, CREB. In the fruit fly, Drosophila melanogaster, calcium and cAMP also regulate the periodicity of circadian locomotor activity rhythmicity, but whether this is due to direct actions on the TTFLs themselves or are a consequence of changes induced to the complex interrelationship between different classes of central pacemaker neurons is unclear. This question was investigated by focusing on the peripheral clock housed in the non-neuronal prothoracic gland (PG), which, together with the central pacemaker in the brain, controls the timing of adult emergence. Genetic manipulations that increased and decreased the levels of calcium and cAMP in the PG caused, respectively, a shortening and a lengthening of the periodicity of emergence. Importantly, knockdown of CREB in the PG caused an arrhythmic pattern of eclosion. Interestingly, the same manipulations directed at central pacemaker neurons caused arrhythmicity of eclosion and of adult locomotor activity, suggesting a common mechanism. These results reveal that the calcium and cAMP pathways can alter the functioning of the clock itself. In the PG, these messengers, acting as outputs of the clock or as second messengers for stimuli external to the PG, could also contribute to the circadian gating of adult emergence (Palacios-Munoz, 2018).
Circadian pacemaker neurons in the Drosophila brain display daily rhythms in the levels of intracellular calcium. These calcium rhythms are driven by molecular clocks and are required for normal circadian behavior. To study their biological basis, this study employed genetic manipulations in conjunction with improved methods of in vivo light-sheet microscopy to measure calcium dynamics in individual pacemaker neurons over complete 24-h durations at sampling frequencies as high as 5 Hz. This technological advance unexpectedly revealed cophasic daily rhythms in basal calcium levels and in high-frequency calcium fluctuations. Further, the rhythms of basal calcium levels and of fast calcium fluctuations were found to reflect the activities of two proteins that mediate distinct forms of calcium fluxes. One is the inositol trisphosphate receptor (ITPR), a channel that mediates calcium fluxes from internal endoplasmic reticulum calcium stores, and the other is a T-type voltage-gated calcium channel, which mediates extracellular calcium influx. These results suggest that Drosophila molecular clocks regulate ITPR and T-type channels to generate two distinct but coupled rhythms in basal calcium and in fast calcium fluctuations. It is proposed that both internal and external calcium fluxes are essential for circadian pacemaker neurons to provide rhythmic outputs and thereby, regulate the activities of downstream brain centers (Liang, 2022).
Circadian rhythms in multiple aspects of cellular physiology help organisms across taxa, from unicellular cyanobacteria to multicellular animals, adapt to environmental day-night changes. In mammals, neurons in the hypothalamic suprachiasmatic nucleus (SCN) show circadian rhythms in gene expression, intracellular calcium, neural activity, and other cellular properties. Circadian rhythms in SCN neuronal outputs coordinate circadian rhythms in other cells throughout the body and generate behavioral rhythms. The rhythms of SCN neuronal outputs can be generated cell intrinsically by the negative transcription/translation feedback loop of core clock genes as a molecular clock, which then generates 24-h oscillations in a series of genes. These gene oscillations then regulate different aspects of membrane physiology, such as the expression levels of channels for potassium, sodium, and calcium. The mechanisms by which the molecular clockworks coordinate complex membrane physiology to generate neural activity rhythms within individual circadian pacemakers remain to be defined (Liang, 2022).
Calcium signaling regulates many cellular processes, such as neural excitability, neurotransmitter release, and gene expression. Cytoplasmic calcium can be regulated from extracellular calcium influx as well as from intracellular calcium stored in the endoplasmic reticulum (ER) and mitochondria. Studies on SCN neurons in vitro and recently, in vivo measured circadian calcium rhythms (CCRs) in SCN neurons. Some studies suggested that calcium rhythms were driven by neuronal firing and voltage-gated calcium channels, while others suggested they were driven by intracellular stores via the ER channel ryanodine receptor (RyR). These alternative hypotheses may derive from the technical differences in the various studies, including the details of in vitro preparations, but also, due to a lack of single-cell resolution in the calcium measurements (Liang, 2022).
In Drosophila, circadian pacemaker neurons also show clock-driven CCRs. The dynamics can be resolved across all five major pacemaker groups (the small ventral lateral neurons [s-LNv], the large ventral lateral neurons [l-LNv], the dorsal lateral neurons [LNd], the group #1 dorsal neurons [DN1], and the group #3 Dorsal Neurons [DN3]), and each group exhibits distinct and sequential daily peak phases. Within such groups, the rhythms can be measured in single identified cells. The multihour phase diversity exhibited by this network requires a series of delays effected by environmental light and by noncell-autonomous modulation mediated by different neuropeptides. Precisely how neuropeptide signaling regulates calcium activity in pacemaker neurons over long (many-hour) durations is unknown. To begin to understand these critical mechanisms of pacemaker modulation, this study began by addressing the cellular and molecular basis of pacemaker calcium rhythms with physiological, genetic, and behavioral measures (Liang, 2022).
In this study, in vivo calcium imaging began at single-cell resolution, using a high-speed light-sheet microscope termed OCPI-2; the acronym OCPI stands for objective-coupled planar illumination. OCPI-2 represents a fundamental technical advance because it permits sampling frequencies to capture stacks of large tissue volumes, without compromising photon efficiency or spatial resolution. Whereas OCPI-1 methods permitted sampling a whole-brain volume once every 10 min across the 24-h day, OCPI-2 methods permit sampling volumetrically at rates as high as 5 Hz. Thus, both basal calcium levels and fast calcium fluctuations were simultaneously measured at single-cell resolution over entire 24-h durations. Circadian rhythmicity was found in both measures. The fast fluctuations are considered to represent events closely coupled to neuronal firing, as have previous studies conducted in much more restricted temporal durations (i.e., not circadian). In all the Drosophila pacemaker neurons studied, these two layers of calcium rhythms shared the same daily temporal pattern (i.e., they were cophasic). To gain insights into the mechanism of these patterns, the fact was exploited that in Drosophila, many calcium channels are encoded by single genes, and genetics was used to study the roles of individual channels in generating daily pacemaker calcium rhythms. This study presents results of experiments in which RNAs encoding different calcium channels were knocked down selectively in all or a subset of pacemakers. The impact of individual channels in setting both slow daily changes in basal calcium levels and in fast fluctuations was evaluated. Finally, PERIOD (PER) protein staining levels and behavior were measure to determine which channels provide feedback to the molecular clock and which are required for normal circadian output from the pacemaker network (Liang, 2022).
This study used in vivo 24-h high-frequency calcium imaging and genetic screening to study the cellular biology of daily calcium rhythms in circadian pacemaker neurons of Drosophila. It was found that the calcium rhythm is in fact a composite; it reflects daily fluctuations in both a slow component (basal levels) and a fast one (high-frequency fluctuations). Fast calcium fluctuations were interpreted as representations of calcium dynamics that occur as neurons fire single action potentials or bursts of them. While it is not sufficient to resolve single action potentials, GCaMP6-induced fluorescence is a good index of neuronal electrical activity. For individual identified pacemakers, these two calcium rhythms share the same daily pattern, yet distinct calcium sources appear to contribute differentially to these two rhythms. An extracellular calcium influx, through plasma membrane calcium channels that include the α1T subunit, is critical for the fast calcium fluctuations. In contrast, calcium fluxes from the ER via the channel ITPR are required for both the slow rhythms in the basal calcium levels and the fast ones. Importantly, both channels are essential for normal circadian behavior. Thus, the molecular clocks may drive circadian rhythms in pacemaker neuron output by regulating different calcium sources to generate coordinate but distinct rhythms in its calcium activities (Liang, 2022).
CCRs are widespread across taxa. Calcium rhythms are required for circadian pacemaker functions in both rodents and Drosophila. Studies on mammalian circadian pacemakers in the SCN are controversial regarding the temporal relationship between the CCR and rhythms in electrical activity, such as in spontaneous firing rate (SFR) and in resting membrane potential (RMP). Recordings from SCN slice cultures showed that the phases of CCR in individual pacemakers are diverse and could be different from the populational phase of SFR rhythms (16, 33). However, the populational SFR phase is composed of many diverse phases of SFR rhythms on the individual cell level; it is unclear whether SFR phases align with the phases of CCR. Imaging with both voltage sensor and calcium sensors in SCN slices, another study concluded that RMP rhythms and CCR were in phase, yet even a third study concluded that RMP rhythms and CCR were in phase in the ventral SCN but that in dorsal SCN, the CCR phase led the RMP rhythms by ~2 h. Because the voltage sensor signal measured from dorsal SCN may derive from the neural processes of ventral SCN neurons, the cellular interpretation of these results is unclear. Another source for the inconsistency might be the culture conditions; in fact, when SCN neurons were recorded in vivo by photometry, the rhythms in fast calcium activity were in phase with slow calcium rhythms. In general, comparisons of population rhythms and rhythms in single cells are not easily reconciled. Recordings that tracked pacemaker neurons from different identified groups in vivo for 24 h showed that at the single cell level, slow calcium rhythms (CCRs) were unambiguously in phase with rhythms in fast calcium fluctuations; the latter likely reflects rhythms in SFR (Liang, 2022).
To measure the fast calcium fluctuations, a high-frequency light-sheet scanning microscope termed OCPI-2 was used. OCPI-2 resolved limitations present in previous microscope designs that created a bottleneck in terms of the rate at which tissue volumes can be repeatedly scanned. Importantly, it does so without compromising photon efficiency or spatial resolution. Its use was instrumental in allowing sampling of the whole-brain volume at frequencies as high as 5 Hz periodically throughout the day. cry01 flies were used to avoid the direct light responses of pacemaker neurons, and it was found that all pacemaker groups displayed slow calcium rhythms, with patterns that were comparable with those previously reported, except for that of l-LNv. That group displayed an additional calcium peak in the early evening and another just before lights on. Because l-LNv innervates the optic lobes and receives large-scale visual inputs, it is speculated that the repeated optical scanning might anomalously activate l-LNv in the evening via visual systems. In the later experiments, when the illumination duration per hour was reduced from 31.5 s to 2.4 s, l-LNv did not show the additional evening peaks; this was true even when measured in flies wild type for cry. Shorter durations of light exposure permit all pacemaker groups to display slow calcium rhythms with phases similar to those obtained with the slow-frequency (every 10-min) recording sessions. Therefore, the concern to avoid technical artifacts when imaging from light-sensitive pacemaker appears especially acute in the case of l-LNv. Nevertheless, by carefully tuning the illumination intensity for calcium imaging, it is proposed that it is possible to monitor normal slow calcium rhythms and fast calcium fluctuations from the same individual pacemaker neurons (Liang, 2022).
The causal relationships between clock gene rhythms, calcium rhythms, and electrical activity rhythms in the SCN remain generally unresolved. Treating SCN slices with Tetrodotoxin (TTX, to block Na-dependent action potentials) diminished SFR rhythms, partially affected RMP rhythms and CCRs, and slowly affected clock gene rhythms over several days. Dispersed SCN cells in vitro showed a TTX-resistant CCR, suggesting that CCR is driven by clock gene rhythms. Thus, the variation in CCR sensitivity to TTX treatment might be caused by the degree to which clock gene rhythms in vitro become progressively dysfunctional. In Drosophila, the findings suggested that clock gene rhythms drive two components of the CCR-both basal calcium levels and fast calcium fluctuations-via circadian regulation of the ER channel ITPR and membrane voltage-gated calcium channel α1T. Both channels might then contribute to SFR and RMP rhythms. Similarly, in SCN pacemakers, pharmacologically blocking another ER channel RyR affected both CCR and SFR rhythms, suggesting that rhythms in basal calcium levels are regulated by calcium from ER and are required for fast electric activity rhythms. In addition, SCN pacemakers also showed a circadian rhythm in fast calcium activity mediated by L-type voltage-gated calcium channels. Pharmacologically blocking these membrane channels affected SFR rhythms and in some case, affected CCR. In these studies, manipulating a membrane voltage-gated calcium channel in all or a subset of pacemakers selectively affected rhythms in fast calcium fluctuations, which likely reflected SFR rhythms and thus, impaired circadian outputs; however, it did not significantly affect the slow rhythms in basal calcium levels. Manipulating the ER calcium channel IP3R in all pacemakers affected rhythms in both slow and fast calcium rhythms. Therefore, in parallel to mammalian SCN neurons, Drosophila circadian pacemakers generate calcium rhythms by regulating both ER and extracellular calcium sources. Since these results suggest little or no role for the RyR channel, the daily rhythmic regulation in fly pacemakers likely acts on a different set of ER and cytoplasmic membrane channels from those in mammalian pacemakers (Liang, 2022).
This study also presented RNAi evidence implicating the Itpr and Ca-&alpha1T genes in pacemaker cell calcium fluxes. However, for each, only a single RNAi proved effective in reducing rhythmic power and increasing percentage of arrhythmicity. Corroboration for these findings is provided by a prior report, which found that the same RNAi line used decreases Itpr RNA levels. Likewise, corroboration was found in a prior report, that described a specific Ca-alpha1T insertion line as a protein null mutation that exhibited reduced percentage of rhythmicity and power in DD but without a change in circadian period or in the daily per rhythm. They concluded that Ca-alpha1T expression does not affect the central clock mechanism and surmised instead that it likely affects the output of the circadian system, specifically via a requirement for the channel to permit normal physiological activation of pacemaker neurons. That work provides independent genetic confirmation of the RNAi results, and its conclusions align precisely with the current work (Liang, 2022).
The RNAi knockdown experiments indicate a role for the ER calcium channel SERCA in supporting slow calcium rhythms in Drosophila pacemakers and behavioral rhythms. However, SERCA knockdown also produced high rates of lethality and among survivors, strong effects on the PER molecular oscillation. It is concluded that SERCA, which maintains the ER cytoplasmic calcium gradient, is essential for the normal physiology of the cells but that these additional phenotypes precluded an assessment of its precise role in circadian rhythmicity. In contrast to the situation with SERCA, the results support a hypothesis that ITPR is a crucial actuator of the molecular clock. Previous transcriptomic analysis also supports that possibility; in circadian neurons, Itpr displays rhythmic expression, while SERCA does not. Knocking down Itpr in all circadian neurons (with tim-GAL4) generally caused stronger deficits in both behavioral rhythms and calcium rhythms than in just the PDF-positive neurons (with pdf-GAL4). One possibility might be that the RNAi expression level is lower when driven by pdf-GAL4 than when driven by tim-GAL4. However, in SI knocking down Itpr in all circadian neurons caused stronger deficits in the amplitude of calcium rhythms of non-PDF-positive neurons (LNd, DN1, and DN3) than in those of PDF-positive neurons (s-LNv and l-LNv). Hence, a difference in the vulnerability to IP3R disruption between PDF-negative and PDF-positive neurons might provide an additional or alternative explanation for why the pdf-GAL4-driven knockdown of Itpr affected neither calcium rhythms nor behavior. It is also noted that the literature provides examples, wherein a single RNAi line produces stronger behavioral effects when coupled with pdf-Gal4 than with tim-GAL4. Such findings suggest that the final result in such experiments will represent a mixture of GAL4 driver line strength and the differential contributions of the UAS:Responder gene product across different positions within an affected neural circuit (Liang, 2022).
Finally, the RNAi knockdown of the plasma membrane calcium channel α1T indicated a role for voltage-gated T-type calcium channels in the final rhythmic output of the pacemakers. Consistent with a role in the presumed output pathway, impairing the rhythm of fast calcium activity strongly affected circadian behavior but did not affect the molecular clock or the slow calcium rhythms. T-type channels play a crucial role in other pacemakers, such as the Sino-Atrial node of the mammalian heart. Their conduction in the hyperpolarized state and closure at more depolarized potentials are central to their role in generating bursting dynamics with periods much longer than the membrane time constant. Given the power spectrum of the fluctuations observed, it seems possible that α1T channels play a similar role in the fast (~0.1-Hz) fluctuations of Drosophila circadian neurons. Remarkably, the expression of α1T also displays a circadian rhythm, with distinct phases in different groups of pacemaker neurons. Collectively, these results suggest that Itpr and α1T channel activity are together critical to produce clock regulation of rhythms in both slow and fast calcium activities of key pacemaker neurons (Liang, 2022).
Although alternative pre-mRNA splicing (AS) significantly diversifies the neuronal proteome, the extent of AS is still unknown due in part to the large number of diverse cell types in the brain. To address this complexity issue, this study used an annotation-free computational method to analyze and compare the AS profiles between small specific groups of Drosophila circadian neurons. The method, the Junction Usage Model (JUM), allows the comprehensive profiling of both known and novel AS events from specific RNA-seq libraries. The results show that many diverse and novel pre-mRNA isoforms are preferentially expressed in one class of clock neuron and also absent from the more standard Drosophila head RNA preparation. These AS events are enriched in potassium channels important for neuronal firing, and there are also cycling isoforms with no detectable underlying transcriptional oscillations. The results suggest massive AS regulation in the brain that is also likely important for circadian regulation (Wang, 2018).
Tissues of the nervous and germline systems, such as brain, testes and ovaries, have more complex transcriptomes than other cell types due to extensive alternative pre-mRNA splicing or AS. The nervous system especially exhibits vast numbers of AS isoforms, many of which are novel and are only beginning to be comprehensively identified. This increase in transcript isoform complexity likely contributes to the specification and functional diversity of cell types within the nervous system (Wang, 2018).
This study applied a novel computational algorithm called JUM to characterize the transcript isoform diversity generated by alternative splicing in three circadian neuronal subtypes (LNv, LNd and DN1), as well as a non-circadian dopaminergic neuron population (TH neurons) of the Drosophila central nervous system. JUM can comprehensively analyze, quantitate and compare tissue- or cell-type-specific AS patterns without requiring a priori annotations of known transcripts or transcriptomes. The analysis revealed a previously unappreciated diversity and complexity of alternatively spliced transcript isoform patterns in these four neuronal subtypes, suggesting that they contribute to neuronal identity, connectivity, activity and circadian functions. This is because many of these novel, previously undetected and unannotated isoforms were unique to a given neuronal population and occurred in transcripts from genes implicated in neuronal activity or circadian rhythms (Wang, 2018).
For example, the kinase Shaggy and the blue light photoreceptor Cryptochrome play central roles in circadian clock regulation and have novel AS patterns in discrete subsets of the circadian neurons. In addition, nine different transcripts involved in potassium transport undergo differential AS in circadian neurons compared to non-circadian neurons. These transcripts encode six different potassium channels. Many of these genes have a complex organization known to encode populations of functionally distinct proteins isoforms, which change the activation kinetics as well as calcium sensitivity of the channels. Neuronal firing is known to play a key role in the circadian circuit with recent studies illustrating that different subgroups of circadian neurons have characteristic time-of-day neuronal firing patterns. Although it is not yet fully understood which potassium channels play a critical role in each circadian neuron subgroup, several channel pre-mRNAs that undergo differential splicing in circadian neurons impact circadian behavior and sleep, such as slowpoke (slo), Shaker (Sh) and Hyperkinetic (Hk). It is therefore likely that AS adds diversity and distinct physiological properties to these protein isoforms, which then impacts neuron-specific firing patterns. From a more general perspective, AS augments transcriptional regulation in giving different circadian neurons individual identities and distinct functions (Wang, 2018).
Approximately 5% of the AS events identified in circadian neurons also undergo time-of-day dependent changes in alternative splicing (cycling splicing). It is important to note that all experiments carried out in this study were conducted under 12 hr of light and 12 hr of dark conditions, making it impossible to distinguish between light and clock control. Nonetheless, these data indicate that splicing adds a dramatic layer of gene regulation to diurnal changes in gene expression. Moreover, many of the cycling AS transcripts show constant overall mRNA levels, which suggests the existence of neuron-specific splicing factors that are expressed or activated only at specific times of the day. Indeed, this study has identified several candidate cycling neuron-enriched transcripts that encode RBPs that may help to drive cycling AS patterns (Wang, 2018).
A recent trend in biological research is to generate transcriptome profiles from single cells. For example, this strategy is part of the 'human cell atlas' project aimed at personalized genomic medicine or the 'brain initiative' project to generate profiles of all neurons in the mouse brain. One recent study was able to obtain about 20M sequence reads per isolated human iPS cell but only managed to analyze splicing patterns for the most highly expressed genes. The current study in contrast used ~100 isolated Drosophila neurons for each of the four neuron subtypes along with judicious use of both oligo-dT and random hexamer priming of the cDNA libraries. This strategy obtained about 10-30M sequence reads for each sample, including substantial information from the 5' ends of transcripts, and JUM was able to detect and classify a large number of previously unannotated pre-mRNA isoforms. Many of them are missing from the fly head RNA-seq data assayed and analyzed in parallel, indicating that these new isoforms are cell-type specific. Not surprisingly, the novel isoforms from the three circadian neuron groups fall into many gene ontology (GO) categories associated with specific circadian clock activity and function (Wang, 2018).
Taken together, the work presented in this study indicates that the number of alternative splicing events that take place in neuronal tissues is grossly underestimated, even though publically-funded genome projects, such as the NIH modENCODE projects deeply sequenced transcriptomes from a variety of Drosophila tissues and developmental stages. This is despite the appreciation of how much AS occurs in the nervous system, for example recent comprehensive analysis of splicing patterns through deep sequencing of ~50 mouse and human tissues revealed about 2500 neuronally-regulated alternative splicing events. It is therefore suggested that these events will need to be comprehensively evaluated by much deeper sequencing than is currently afforded by most contemporary single cell RNA-seq studies and by AS analysis software like JUM that is not constrained by a priori knowledge of known splicing events (Wang, 2018).
The circadian clock is an evolutionarily conserved mechanism that generates the rhythmic expression of downstream genes. This study explored the role of Achilles in regulating the rhythmic transcriptome in the fly head. Achilles is a clock-controlled gene in Drosophila that encodes a putative RNA-binding protein. Achilles expression is found in neurons throughout the fly brain, and legacy data suggest it is not expressed in core clock neurons. To assess its impact on circadian mRNA rhythms, RNA sequencing (RNAseq) was performed to compare the rhythmic transcriptomes of control flies and those with diminished Achilles expression in all neurons. Dramatic upregulation of immune response genes was observed upon knock-down of Achilles. Furthermore, many circadian mRNAs lose their rhythmicity in Achilles knock-down flies, suggesting that a subset of the rhythmic transcriptome is regulated either directly or indirectly by Achilles. These Achilles-mediated rhythms are observed in genes involved in immune function and in neuronal signaling, including Prosap, Nemy and Jhl-21. A comparison of RNAseq data from control flies reveals that only 42.7% of clock-controlled genes in the fly brain are rhythmic in both males and females. As mRNA rhythms of core clock genes are largely invariant between the sexes, this observation suggests that sex-specific mechanisms are an important, and heretofore under-appreciated, regulator of the rhythmic transcriptome (Li, 2019).
Most organisms on the earth exhibit circadian rhythms in behavior and physiology, which are driven by endogenous clocks. Phosphorylation plays a central role in timing the clock, but how this contributes to overt rhythms is unclear. Phosphoproteomics in conjunction with transcriptomic and proteomic profiling was conducted using fly heads. By developing a pipeline for integrating multi-omics data, 789 (~17%) phosphorylation sites with circadian oscillations were identified. It is predicted that 27 potential circadian kinases participate in phosphorylating these sites, including 7 previously known to function in the clock. The remaining 20 kinases were screened for effects on circadian rhythms, and an additional 3 were found to be involved in regulating locomotor rhythm. A signal web was re-constructed that includes the 10 circadian kinases, and GASKET was identified as a potentially important regulator. Taken together, this study uncovered a circadian kinome that potentially shapes the temporal pattern of the entire circadian molecular landscapes (Wang, 2020).
The impact of global warming on the life of the earth is increasingly concerned. Previous studies indicated that temperature changes have a serious impact on insect sleep. Sleep is critical for animals as it has many important physiological functions. It is of great significance to study the regulation mechanism of temperature-induced sleep changes for understanding the impact of global warming on insects. More importantly, understanding how these pressures regulate sleep can provide insights into improving sleep. This study found that extra sex combs (ESC) is a regulatory factor in this process. The data showed that ESC was an upstream negative regulatory factor of Heat shock proteins (Hsps), and it could regulate sleep in mushroom and ellipsoid bodies of Drosophila. ESC mutation exaggerates the sleep change caused by temperature, while buffering the shortening of life caused by sleep deprivation. These phenotypes can be rescued by Hsps mutants. Therefore, it is concluded that the ESC buffers sleep-related stresses through regulating Hsps (Zhao, 2020).
Circadian rhythms have been extensively studied in Drosophila, however, still little is known about how the electrical properties of clock neurons are specified. A behavioral genetic screen was performed in this study through the downregulation of candidate ion channels in the lateral ventral neurons (LNvs); it was shown that the hyperpolarization-activated cation current I(h) is important for the behaviors that the LNvs influence: temporal organization of locomotor activity, analyzed in males, and sleep, analyzed in females. Using whole-cell patch clamp electrophysiology this study demonstrated that small LNvs are bursting neurons, and that I(h) is necessary to achieve the high frequency bursting firing pattern characteristic of both types of LNvs in females. Since firing in bursts has been associated to neuropeptide release, it is hypothesized that I(h) would be important for LNvs communication. Indeed, this study demonstrates that I(h) is fundamental for the recruitment of PDF filled dense core vesicles to the terminals at the dorsal protocerebrum and for their timed release, and hence for the temporal coordination of circadian behaviors (Fernandez-Chiappe, 2020).
The integration of circadian and metabolic signals is essential for maintaining robust circadian rhythms and ensuring efficient metabolism and energy use. Using Drosophila as an animal model, this study shows that cellular protein O-GlcNAcylation exhibits robust 24-hour rhythm and represents a key post-translational mechanism that regulates circadian physiology. Strong correlation was observed between protein O-GlcNAcylation rhythms and clock-controlled feeding-fasting cycles, suggesting that O-GlcNAcylation rhythms are primarily driven by nutrient input. Interestingly, daily O-GlcNAcylation rhythms are severely dampened when flies are subjected to time-restricted feeding at unnatural feeding time. This suggests the presence of clock-regulated buffering mechanisms that prevent excessive O-GlcNAcylation at non-optimal times of the day-night cycle. This buffering mechanism is mediated by the expression and activity of GFAT, OGT, and OGA, which are regulated through integration of circadian and metabolic signals. Finally, a mathematical model was generated to describe the key factors that regulate daily O-GlcNAcylation rhythm (Liu, 2021).
The Drosophila circadian clock is driven by a transcriptional feedback loop in which CLOCK-CYCLE (CLK-CYC) binds E-boxes to transcribe genes encoding the PERIOD-TIMELESS (PER-TIM) repressor, which releases CLK-CYC from E-boxes to inhibit transcription. CLOCKWORK ORANGE (CWO) reinforces PER-TIM repression by binding E-boxes to maintain PER-TIM bound CLK-CYC off DNA, but also promotes CLK-CYC transcription through an unknown mechanism. To determine how CWO activates CLK-CYC transcription, CWO target genes were identified that are upregulated in the absence of CWO repression, conserved in mammals, and preferentially expressed in brain pacemaker neurons. Among the genes identified was a putative ortholog of mouse Clock Interacting Protein Circadian (Cipc), which represses CLOCK-BMAL1 transcription. Reducing or eliminating Drosophila Cipc expression shortens period, while overexpressing Cipc lengthens period. Cipc represses CLK-CYC transcription in vivo, but not uniformly, as per is strongly repressed, tim less so, and vri hardly at all. Long period rhythms in cwo mutant flies are largely rescued when Cipc expression is reduced or eliminated, indicating that increased Cipc expression mediates the period lengthening of cwo mutants. These results suggest a mechanism for CWO-dependent CLK-CYC activation: CWO inhibition of CIPC repression promotes CLK-CYC transcription. This mechanism may be conserved since cwo and Cipc perform analogous roles in the mammalian circadian clock (Rivas, 2021).
The Pallidin protein is a central subunit of a multimeric complex called biogenesis of lysosome-related organelles complex 1 (BLOC1) that regulates specific endosomal functions and has been linked to schizophrenia. Downregulation of Pallidin and other members of BLOC1 in the surface glia, the Drosophila equivalent of the blood-brain barrier, reduces and delays nighttime sleep in a circadian-clock-dependent manner. In agreement with BLOC1 involvement in amino acid transport, downregulation of the large neutral amino acid transporter 1 (LAT1)-like transporters JhI-21 and mnd, as well as of TOR (target of rapamycin) amino acid signaling, phenocopy Pallidin knockdown. Furthermore, supplementing food with leucine normalizes the sleep/wake phenotypes of Pallidin downregulation, and this study identified a role for Pallidin in the subcellular trafficking of JhI-21. Finally, evidence is provided that Pallidin in surface glia is required for GABAergic neuronal activity. These data identify a BLOC1 function linking essential amino acid availability and GABAergic sleep/wake regulation (Li, 2023).
Biogenesis of lysosome-related organelles complex 1 (BLOC1) is an octameric complex linked to endosomal compartments and the cytoskeleton (Cheli, 2010). The genes coding for the 8 subunits in mice (Pallidin, dysbindin, BLOS1, BLOS2, BLOS3, cappuccino, muted, and snapin) are broadly expressed within the brain and in peripheral tissues. The complex regulates the trafficking of various receptors and transporters and appears to play a prominent role in the biogenesis of recycling endosomes. Mice bearing severe or complete loss-of-function mutations in BLOC1 genes are viable and fertile. They display common phenotypes, originating from defects in highly specialized lysosome-related organelles, such as reduced pigmentation due to impaired retinal and epidermal melanosomes or extended bleeding times resulting from the lack of dense granules in platelets. In humans, mutations in BLOC1 genes and other functionally related genes are found in the Hermansky-Pudlak syndrome (Li, 2023).
In addition, genetic studies have identified variants of the dysbindin gene and other BLOC1 genes as risk factors for developing schizophrenia. Although the latter results are debated, several postmortem studies have reported reduced levels of dysbindin mRNA and protein in the brain of schizophrenics. Furthermore, genome-wide association studies also reported a link between dysbindin genetic variants and cognitive abilities (Li, 2023).
These findings have led to investigations aiming at deciphering the role of BLOC1 in neuronal function using not only mouse models defective for individual BLOC1 gene function but also the Drosophila model, in which the complex is well conserved. These studies confirmed the involvement of BLOC1 in behavior and memory. They identified potential cellular and molecular mechanisms such as abnormal glutamatergic, GABAergic, and dopaminergic transmission. However, the vast majority of these studies have been carried out in animals with spontaneous or artificial mutations. This approach makes the interpretation of the resulting phenotypes challenging, given the broad expression of BLOC1 in many cell types. Furthermore, the complex is expressed during development, when it can be involved in neurodevelopmental diseases such as autism spectrum disorders (Li, 2023).
Apart from one recent report (Lee, 2018) the involvement of BLOC1 in sleep/wake regulation has not been investigated. Interestingly, it has been found that Pallidin is upregulated in a somnolent mouse model with defective histaminergic transmission (Seugnet, 2023).
Thus, further investigation is required given the critical implication of sleep in brain function, and in particular in schizophrenia, in which sleep disruption could be in part a consequence of the pathology, or an aggravating factor as seen in human
and rodent models. This study investigated the potential role of BLOC1 in sleep/wake regulation in Drosophila. Neurotransmission systems, ion channels, and glial functions in Drosophila and mammals are globally conserved (Li, 2023).
There are differences regarding sleep/wake in insects compared with mammals, such as the absence of slow wave and paradoxical sleep. Nevertheless, the evidence for conserved regulatory principles and functions is compelling.
The flexibility and short generation time of the Drosophila model is an asset in molecular genetic studies, providing hypotheses that can be tested in rodents and ultimately used for therapy. A conditional knockdown strategy was used to target Pallidin, a major component of BLOC1. The pool of Pallidin in the cell is associated almost entirely with the complex, and this protein plays a central role through its interactions with dysbindin, BLOS1, and cappuccino (Cheli, 2010), with its loss leading to their degradation (Li, 2023).
This study focused on glia, as a previous results suggest a function for Pallidin in non-neuronal cells. Glial cells are increasingly proven to play very significant roles in the control of sleep/wake and circadian rhythms both in Drosophila and in rodent models. Glia-dependent neurotransmitter reuptake, neurotransmitter metabolism, and glial calcium transients critically influence neuronal networks that control sleep timing and sleep homeostasis. Neuroglia signaling pathways have been shown to modulate sleep-deprivation-induced learning impairments. Recent reports have demonstrated the prominent role of glial cells at the interface between the brain and the circulating fluids during the sleep/wake cycle (Li, 2023 and references therein).
This study provides evidence in Drosophila that Pallidin and other BLOC1 components regulate the initiation of sleep in the early part of the night, at the level of surface glial (SG) cells. In Drosophila, SG cells express tight junctions and molecular components in common with the mammalian BBB. As in the BBB this organization prevents the entry of macromolecules and tightly regulates the exchanges of solutes between the circulatory system (the hemolymph in Drosophila) and the brain, thus maintaining the particular interstitial fluid composition necessary for neuronal activity. This function relies on intense transporter activity. The data support a mechanism whereby BLOC1 regulates LAT1-like transporters subcellular trafficking in both perineurial and subperineurial cells, leading to adequate essential amino acid supply and GABAergic neuronal activity promoting sleep, in a circadian-clock-dependent manner. This study observed that downregulating Pallidin using a pan-neuronal driver also resulted in reduced night sleep, suggesting that SG is not the only cell type in the brain where Pallidin is required to regulate sleep (Li, 2023).
Blocking the cycling of the molecular clock by placing flies in a per0 mutant background completely abolishes the Pallidin knockdown sleep/wake phenotype, suggesting that the circadian clock is implicated. Accordingly, it was found that hyperpolarization of PDF-expressing clock neurons is sufficient to normalize the main Pallidin knockdown phenotype: increased sleep latency at night. This raises the possibility that Pallidin affects clock-dependent sleep/wake regulatory networks and in particular, those under the control of the large PDF-expressing neurons (lLNv). In lLNv neurons, GABAergic input is modulated in a circadian manner, resulting in higher inhibition in the early night, promoting sleep, while a disruption of this input results in delayed and fragmented sleep (Li, 2023).
The GABAergic neurons responsible for lLNv inhibition are so far unidentified. The delayed and reduced night sleep observed upon Pallidin knockdown in SG is also similar to the phenotype previously reported with pan-GABAergic neuron inhibition.
Conversely, feeding flies the GABA agonist THIP65
or increasing GABAergic neuronal firing by expressing TrpA1 can potently induce sleep. The latter manipulation cannot promote sleep when Pallidin is downregulated in SG, suggesting that the gene is required to allow sustained GABAergic neuronal activity. Alternatively, Pallidin function could be regulated directly by the clock present in perineurial cells, which modulates the efflux properties of subperineurial cells in a circadian manner (Li, 2023).
Downregulation of several members of BLOC1 in SG phenocopies the knockdown of Pallidin, strongly suggesting that the whole complex is involved in this Pallidin-dependent sleep/wake regulation. This is consistent with the observation that the pool of Pallidin and dysbindin present in the cell is almost entirely associated with the complex and that the lack of one subunit leads to a destabilization of the others (Li, 2023).
The lack of sleep alteration with snapin manipulation may result from lower efficiency of the genetic tools used for downregulation. Alternatively, it may also reflect different gene dosage sensitivity and functionality for the different members of the complex, as previously reported, or the existence of multiple subunits within the complex2
or different complexes containing a subset of the subunits (Li, 2023).
In Drosophila larval neuromuscular junction, Snapin has, for example, been shown to regulate synaptic homeostasis (Dickman, 2012),
while Pallidin does not seem to affect baseline neurotransmission and is required during sustained neuronal activity to replenish the pool of releasable synaptic vesicles (Chen, 2017). BLOC1 has been shown to be required for the biogenesis of recycling endosomes through its interactions with sorting endosomes and the cytoskeleton (Delevoye, 2016).
These BLOC1 functions and the regulation of sleep by BLOC1 members reported in this study are in agreement with a recent report showing bidirectional interactions between sleep and endocytosis in SG (Lee, 2018).
Intriguingly, the endocytosis in SG was reported to be the most intense in the early part of the night,
when sleep is the deepest and the phenotype of Pallidin downregulation is the most pronounced. In addition, the activity of the recycling endosome associated small GTPase Rab11 appears to play a prominent role in this context.
Consistent with these findings, extensive protein-protein interaction analyses identified rab11 as the best interacting partner for BLOC1 in both Drosophila and humans.5
Thus, BLOC1 may facilitate the high endocytic activity of SG during the early part of the night, notably the biogenesis of recycling endosomes, while having a less prominent role at other times during the day. In line with the results, Pallidin mutant mice display reduced sleep amount during the light phase of the day, the primary sleep period in rodents, and shorter average sleep-bout duration (Li, 2023).
However, BBB-specific and conditional knockout models in mice would be necessary to determine whether the model outlined in this study applies to mammals. In humans, the incidence of sleep/wake disorders in the rare Hermansky-Pudlak syndrome, bearing deficits in BLOC1 or other functionally related complexes, is unknown but deserves scrutiny. In contrast, sleep disruption is common among patients suffering from schizophrenia, another pathology associated with BLOC1 deficits. Interestingly, one of the prominent sleep abnormalities in schizophrenia patients is a reduction in sleep spindle density, which strongly relies on GABAergic activity, and the latter is also affected in this pathology. Accordingly, previous studies in dysbindin mutant mice have shown that BLOC1 can disrupt GABAergic activity (Li, 2023).
Thus, a potential role for amino acid transport at the BBB in those contexts would deserve investigation given these results. Early studies of Pallidin function pointed to a role in amino acid import into the brain transport, as suggested by lower sensitivity to intraperitoneal injections of the LAT1 substrates l-DOPA and tryptophan. The LAT1 transporter plays a major role in the import of these amino acids and in the import of large neutral essential amino acids such as leucine, isoleucine, and histidine (Li, 2023).
A conditional knockout of LAT1 in brain endothelial cells confirmed the crucial role of LAT1 in the regulation essential amino acid abundance within the brain, its impact on GABAergic transmission, and its potential implication in autism spectrum disorders.
Intriguingly, the relative abundance of amino acids in the brain of Pallidin mutant mice resembles those found in mice with a BBB specific knockout LAT1 (Li, 2023).
This study provides evidence that Pallidin function in SG is linked to essential amino acid supply and to LAT1-like transporter activity. First, downregulation of the LAT1-like transporters and of TOR signaling phenocopy knock down, to a large extent or completely, the BLOC1 genes. Second, supplementing the food with leucine can normalize the JhI-21, Pallidin, and Blos2 phenotypes. Interestingly, the phenotype of the JhI-21 transporter knockdown could be rescued by valine and tryptophan supplementation, while Pallidin knockdown could be rescued only by leucine. This suggests that Pallidin does not solely modulate LAT1-like transporter function and has complex effect on multiple transport systems, as previously suggested (Li, 2023).
JhI-21 and minidisc are the Drosophila closest homologs of LAT1 light-chain.
JhI-21 is expressed broadly in the Drosophila brain, with most of the signal in perinuclear punctae that colocalizes partially with the lysosomal marker ATG8. The subcellular trafficking of JhI-21 in SG is abnormal following Pallidin downregulation, with a substantial fraction of the transporter localized in the nucleus. This abnormal trafficking is likely to reduce the functionality of the transporter, explaining the similar sleep/wake phenotypes of JhI-21 and Pallidin downregulation in SG and their normalization by essential amino acid supplementation. A nuclear localization has already been reported in glioma cell lines for the solute carrier (SLC) Eaat1 and Eaat2 transporters, and for the K+ inwardly rectifying channel. In both cases, this unusual subcellular localization was associated with an overall reduced functionality of the protein in the cell. Aside transporters, several full-length transmembrane receptors have been repeatedly reported to be localized in the nucleus where some of them may act as a transcription factors (Li, 2023).
The mechanisms underlying this phenomenon are mostly unknown. In this study, it was not possible to detect obvious changes in autophagy, suggesting that Pallidin downregulation does not induce major changes in lysosomal activity and rather affects more specifically particular cargos, as previously suggested. Interestingly, in addition to their cytoplasmic localization, a nuclear localization has previously been reported for two BLOC1 subunits: BLOS2102
and, in particular, dysbindin. ATG8 is also found both in lysosomes and in the nucleus where it can regulate gene expression in association with transcription factors. Such lysosomal-nuclear connections open the possibility of trapping a diversity of unexpected proteins in the nucleus in normal as well as in abnormal conditions (Li, 2023).
In Drosophila, protein intake, threonine intake, and D-serine levels play a role in sleep/wake regulation. Although the changes in amino acid intake presumably change global free amino acid levels within the brain, the effect on sleep/wake regulation seems to originate from specific sleep/wake regulatory networks or neurotransmission systems. In humans, amino-acid-supplemented diets have been designed to improve sleep and wakefulness, on the basis of the principle that this will increase the synthesis of monoamines. The results point to a model independent from these previously identified mechanisms: higher recycling endosome activity in the early part of the night, mediated by BLOC1, would lead to high LAT1-like activity and essential amino acid import in the brain. The TOR signaling appears also to be required and may facilitate LAT1-like transporter function. This assessment suggests that Pallidin-dependent sleep regulation does not involve a modulation of monoamine levels, in contrast with previous reports. However, in these reports the conclusions were reached using complete knock out and not by downregulation in specific cell types, making comparison with the present results difficult. For example, Drosophila Dysbindin has been suggested to regulate global brain dopamine level by modulating its recycling through glial cells (Li, 2023).
In contrast, the current data suggest that essential amino acid import to the brain facilitated by Pallidin would enhance GABAergic transmission required for sleep. This study showed that a large number of GABAergic neurons in the adult fly brain are in direct cellular contacts with SG. Although the significance and the function of these contacts remain to be determined, these observations fit the idea that a significant subset of these neurons have particular metabolic needs (Li, 2023).
How could amino acid transport regulate GABAergic transmission? Amino acids are at the interplay among many processes, being involved in protein synthesis, energy metabolism, neurotransmitter synthesis, and degradation. Amino acid transporters may further intertwine these processes: as a prime example, LAT1 is an antiporter that can associate the import of leucine to the export of glutamine, two amino acids that are crucial in the glutamate/GABA/glutamine cycle in the brain (Li, 2023).
Pharmacological experiments have indeed suggested that LAT1 in the BBB regulates GABA homeostasis in the interstitial fluid. Branched-chain amino acids
(BCAA) and leucine in particular are thought to be critical providers of nitrogen for glutamate synthesis through transamination,
affecting glutamate/GABA/glutamine cycling. Enzymes for BCAA metabolism are conserved in Drosophila, suggesting similar metabolic regulation in this model organism. An interesting example is Drosophila mutants for GABA transaminase (GABAT), which sleep 2-3 h longer than their genetic controls because of impaired GABA degradation. For survival, these mutants require that food include glutamate or BCAAs (leucine and valine), whose transamination can provide glutamate in the cells. These data indicate that GABA metabolism and BCAAs as a source of glutamate play a substantial role in sleep/wake regulation and brain energy metabolism. Interestingly, this study found that the GABA/glutamate ratio tends to be decreased in Pallidin knockdown flies, while the glutamate/aspartate ratio is increased compared with controls. These results suggest that BLOC1 function in SG modulates neurotransmitter and brain amino acid metabolism on a global level and may control the inhibitory/excitatory balance (Li, 2023).
The data presented in this study emphasize the implication of circulating amino acids, in particular BCAA, in sleep/wake regulation, corroborating several recent metabolomics studies in mammalian models and in humans, in which branched-chain amino acid (BCAA) levels have been repeatedly shown to be modulated by the circadian clock and/or the sleep homeostat. For example, a recent study identified in insomniac patients prominent changes in the levels of several circulating BCAA, including increased levels of leucine during the night.
Food supplemented with BCAA can correct sleep disorders in a mouse model with chronic sleep disruption. It is worth noting that genes involved in amino acid transport are among those commonly disrupted in autism and schizophrenia. In conclusion, this study provides potential mechanisms at the blood-brain interface that may be relevant to both sleep disruption and psychosis and emphasizes the possibility of diet-based therapies (Li, 2023).
One limitation of this study has been the inability to monitor Pallidin protein expression using immunofluorescence. A previously published antibody
produced a non-specific signal in the adult brain, and attempts to generate a new antibody were unsuccessful. This precluded determining the subcellular localization of the protein and assessing the local efficiency of the knockdown constructs. The lack of appropriate drivers prevented more precise manipulation of the neuronal targets affected by Pallidin function: the GABAergic neurons presynaptic to lLNv neurons are unknown, and therefore there is no identified driver making it possible to monitor or control their activity, and in addition, there are no available LexA construct specifically expressed in lLNv neurons. For future investigations, it will be important to develop an efficient conditional system, independent of temperature, to limit the knockdown of Pallidin to the adult stage. Finally, further work is required to elucidate the involvement of the TOR signaling pathway in this context and the Pallidin-JhI-21-dependent interactions between SG and GABAergic neurons (Li, 2023).
While endogenous lipids are known to exhibit rhythmic oscillations, less is known about how specific lipids modulate circadian behavior. Through a series of loss-of-function and gain-of-function experiments on ceramide phosphoethanolamine (CPE) synthase of Drosophila, it was demonstrated that pan-glial-specific deficiency in membrane CPE, the structural analog of mammalian sphingomyelin (SM), leads to arrhythmic locomotor behavior and shortens lifespan, while the reverse is true for increasing CPE. Comparative proteomics uncovered dysregulated synaptic glutamate utilization and transport in CPE-deficient flies. An extensive genetic screen was conducted to verify the role of differentially expressed proteins in circadian regulation. Arrhythmic locomotion under cpes(1) mutant background was rescued only by restoring endogenous CPE or SM through expressing their respective synthases. These results underscore the essential role of CPE in maintaining synaptic glutamate homeostasis and modulating circadian behavior in Drosophila. The findings suggest that region-specific elevations of functional membrane lipids can benefit circadian regulation (Chen, 2022)
In the fruit fly Drosophila melanogaster, as in
mammals, acute exposure to a high dose of ethanol leads to
stereotypical behavioral changes beginning with increased activity,
followed by incoordination, loss of postural control, and eventually,
sedation. The mechanism(s) by which ethanol impacts the CNS leading to
ethanol-induced sedation and the genes required for normal sedation
sensitivity remain largely unknown. This study identified the gene apontic
(apt), an Myb/SANT-containing transcription factor that is
required in the nervous system for normal sensitivity to ethanol
sedation. Using genetic and behavioral analyses, it was shown that apt
mediates sensitivity to ethanol sedation by acting in a small set of
neurons that express This work identifies apt and crz that mediate the
fly's sensitivity to ethanol-induced sedation. Flies with reduced
expression of either gene display dramatically decreased ethanol
sedation sensitivity; thus, both apt and crz
normally promote ethanol sedation. Normal sensitivity to ethanol
sedation requires apt expression in neurons during two
distinct life stages, metamorphosis and adulthood. Apt function in a
subset of crz-expressing neurons (approximately 6 of the 12-16
crz-expressing cells) is necessary and sufficient for normal
sensitivity to ethanol sedation. Acute manipulations of the activity of
crz neurons led to altered ethanol sedation sensitivity,
demonstrating that these neurons play an active role in regulating the
behavioral response to ethanol-induced sedation. The neuropeptide Crz is
also involved in ethanol sedation, as flies with reduced crz
expression, specifically during adulthood, show dramatically decreased
ethanol sedation sensitivity. Finally, in response to acute ethanol
exposure the expression of both apt and crz are
rapidly upregulated during ethanol exposure. It is hypothesized that the
Apt-Crz system, functioning in a very small group of neurosecretory
cells, may be an early target of ethanol in the fly brain whose function
is crucial for normal sensitivity to ethanol (McClure, 2013). How does Apt regulate ethanol-induced sedation? Although Apt's role
could be to regulate the expression of crz, no such function
was observed. It is thus postulated that Apt functions to regulate the
activity of crz neurons and/or neuropeptide release. For
example, Apt, acting as a transcription factor, could regulate the
transcription of proteins required for synthesis, packaging, and/or
release of Crz and possibly other neuropeptides. Alternatively, Apt
could regulate the expression of proteins required for synapse
formation. In support of these two possibilities, apt mutant
embryos were observed to have defective synaptic transmission at the
neuromuscular junction, as well as fewer numbers of active zones within
motoneurons, indicating a presynaptic defect (McClure, 2013). Another possible function for Apt in regulating ethanol sedation
behavior may be found in its neuronal requirement during metamorphosis,
a time of intense remodeling to construct the adult CNS. During
embryogenesis, Apt functions in multiple morphogenetic processes,
including tracheal, head, CNS, and heart morphogenesis, as well as
border cell migration. It is therefore possible that during
metamorphosis Apt establishes proper development and neuronal
connectivity of the adult CNS, and in particular the Crz neurons.
However, this possibility seems somewhat unlikely given that the adult
CNS in apt13-66 flies appeared normal, as was the
number and morphology of Crz-expressing neurons. Additionally, it was
found that adult-specific expression of apt in neurons was
necessary for normal sedation sensitivity. However, there may be subtle
defects in the adult CNS of apt mutant flies, which were not
detected, that could contribute to their altered sedation sensitivity
(McClure, 2013). Apt shows highest sequence conservation with the human FSBP, a negative
regulator of transcription of the gamma chain of fibrinogen
(see Starz-Gaiano, 2008). Sequence conservation between apt
and FSBP is observed within the DNA-binding domain.
Interestingly, moderate alcohol consumption in humans has been known to
exert a cardioprotective effect, in part by lowering levels of
circulating Fibrinogen. The mechanism for how alcohol consumption
regulates Fibrinogen is currently unknown, but in light of the current
findings it is speculated that it may occur at the level of
transcription. Ethanol exposure in flies was found to acutely upregulate
apt expression. A similar situation may occur in humans,
whereby alcohol consumption could upregulate the transcription of FSBP,
ultimately leading to negative regulation of the gamma chain of fibrinogen
and lowered levels of circulating Fibrinogen, which in turn would
provide cardioprotection (McClure, 2013). The observations implicate crz-expressing neurons in the
regulation of ethanol sedation behavior, a function not previously
attributed to these neurons. This study demonstrated that adult-specific
silencing of crz neurons significantly reduced ethanol
sedation sensitivity, while increasing their activity resulted in the
opposite phenotype, an increase in ethanol sedation sensitivity. Based
on the observation that inhibiting crz expression also reduced
ethanol sedation sensitivity, it is believed that the phenotypes
associated with crz neuronal manipulations reflect changes in
the release of the neuropeptide Crz and activation of its signaling
pathway. However, a few Crz neurons also express the short Neuropeptide
F (sNPF). sNPF is considered to be a multifunctional neuropeptide due to
its broad expression in diverse neuronal types, and its possible role in
crz-expressing neurons and in ethanol sedation sensitivity has
not been excluded. However, the observation that Apt function is
required in a subset of crz neurons to promote ethanol
sedation behavior, firmly establishes the importance of these neurons in
mediating the behavioral response to ethanol. The data also suggest that
the function of both genes, crz and apt, overlaps in
a small set of neurons likely located in the pars lateralis (PL) to
mediate the behavioral response to ethanol-induced sedation (McClure,
2013). It has been hypothesized that Crz is released in response to various
types of stress in insects (Veenstra, 2009; Boerjan, 2010), and that
this could explain its pleiotropic effects. This hypothesis was
bolstered by a recent study showing that flies deficient in Crz are
resistant to metabolic, osmotic, and oxidative stress, as measured by
survival (Zhao, 2010). In addition, Crz plays a role in stress
physiology through its association with well characterized stress
hormones. For instance, crz-expressing neurons in the PL also
express receptors for two diuretic hormones, DH44 and DH31.
By virtue of receptor similarity, DH44 and DH31
are related to corticotrophin-releasing factor (CRF) and calcitonin-gene
related peptide (CGRP), respectively, both of which mediate the
mammalian physiological and behavioral responses to stress.
Interestingly, both CRF and CGRP act to inhibit secretion of GnRH in the
mammalian hypothalamus. This is significant because Crz is thought to be
the homolog of mammalian GnRH, and suggests that analogous regulation
occurs in Drosophila (Cazzamali, 2002). It is thus possible
that in flies a stress signal or the animal's stress status may be
relayed to Crz neurons and alters their function. Thus, based on its
functional and molecular associations with stress physiology, it is
tempting to speculate that the role of Crz signaling in ethanol sedation
sensitivity is related to a stress response. A previous study has shown
that stress, in the form of heat shock, induces tolerance to a
subsequent ethanol exposure, and that ethanol tolerance relies on the
gene hangover, a large nuclear zinc-finger protein, that
mediates various other stress responses (Scholz, 2005). In addition,
several genes related to stress responses have been shown to be
upregulated by ethanol exposure in transcriptional profiling studies,
including nearly half of all Drosophila heat shock protein
genes, as well as genes involved in the regulation of oxidative stress
and aging. Importantly, a maladaptive response to stress has been shown
in humans to be a major and common element contributing to drug
addiction. Finally, an increase in ethanol self-administration has been
observed in animal models with physical, social, and emotional stress.
In light of these findings, it will be interesting to further explore
the role of Crz and its function in stress physiology and the regulation
of ethanol-related behaviors (McClure, 2013). Neuropeptides are diverse signaling molecules that mediate a broad
spectrum of physiological and behavioral processes. Several studies have
linked neuropeptides to behavioral responses to ethanol. For instance,
one of the first ethanol sensitivity mutants described in Drosophila, amnesiac,
encodes a neuropeptide homologous to the vertebrate pituitary adenylate
cyclase-activating peptide. In addition, mice lacking either
neuropeptide Y (NPY), a neuromodulator abundantly expressed in many
regions of the CNS, or its Y1 receptor subtype, display increased
ethanol consumption and resistance to ethanol sedation, whereas animals
overexpressing NPY show the opposite behavioral phenotypes. Neuropeptide
F (NPF), the sole member of the NPY family in Drosophila, and
its receptor NPFR1, has similarly been shown to mediate the fly's
sensitivity to ethanol-induced sedation. Finally, flies with neuronal
perturbations in the insulin signaling pathway displayed increased
ethanol sedation sensitivity. These studies, implicating the
neuropeptide Crz in sensitivity to ethanol sedation, suggest that
neuropeptides are important regulators of the behavioral response to
ethanol, and it would therefore be interesting to survey all known Drosophila
neuropeptides and their downstream signaling components for possible
role(s) in ethanol-related behaviors (McClure, 2013). Circadian rhythms in transcription are generated by rhythmic abundances and DNA binding activities of transcription factors. Propagation of rhythms to transcriptional initiation involves the core promoter, its chromatin state, and the basal transcription machinery. This study characterize core promoters and chromatin states of genes transcribed in a circadian manner in mouse liver and in Drosophila. The core promoter was shown to be a critical determinant of circadian mRNA expression in both species. A distinct core promoter class, strong circadian promoters (SCPs), is identified in mouse liver but not Drosophila. There was no apparent SCP-like subpopulation of promoters in Drosophila with high amounts of paused Pol II and low nucleosome occupancy, at the same time also driving transcriptional rhythms with both high amplitudes and mean levels. SCPs are defined by specific core promoter features, and are shown to drive circadian transcriptional activities with both high averages and high amplitudes. Data analysis and mathematical modeling further provided evidence for rhythmic regulation of both polymerase II recruitment and pause release at SCPs. The analysis provides a comprehensive and systematic view of core promoters and their link to circadian mRNA expression in mouse and Drosophila, and thus reveals a crucial role for the core promoter in regulated, dynamic transcription (Westermark, 2016).
The circadian clock is a transcriptional/translational feedback loop that
drives the rhythmic expression of downstream mRNAs. Termed
"clock-controlled genes," these molecular outputs of the circadian
clock orchestrate cellular, metabolic, and behavioral rhythms. This
study identified a novel clock-controlled gene in Drosophila
melanogaster, Achilles
(Achl), which is rhythmic at the mRNA level in the brain and
which represses expression of anti-microbial peptides in the immune
system. Achilles knock-down in neurons dramatically
elevates expression of crucial immune response genes, including IM1
(Immune induced molecule 1), Mtk
(Metchnikowin), and Drs
(Drosomysin). As a result, flies with knocked-down Achilles
expression are resistant to bacterial challenges. Meanwhile, no
significant change in core clock gene expression and locomotor activity is
observed, suggesting that Achilles influences rhythmic mRNA
outputs rather than directly regulating the core timekeeping mechanism.
Notably, Achilles knock-down in the absence of immune challenge
significantly diminishes the fly's overall lifespan, indicating a
behavioral or metabolic cost of constitutively activating this pathway.
Together, these data demonstrate that (1) Achilles is a novel
clock-controlled gene that (2) regulates the immune system, and (3)
participates in signaling from neurons to immunological tissues (Li, 2016). Activity-regulated genes (ARGs) are important for neuronal functions like long-term memory and are well-characterized in mammals but poorly studied in other model organisms like Drosophila. This study stimulated fly neurons with different paradigms and identified ARGs using high-throughput sequencing from brains as well as from sorted neurons: they included a narrow set of circadian neurons as well as dopaminergic neurons. Surprisingly, many ARGs are specific to the stimulation paradigm and very specific to neuron type. In addition and unlike mammalian immediate early genes (IEGs), fly ARGs do not have short gene lengths and are less enriched for transcription factor function. Chromatin assays using ATAC-sequencing show that the transcription start sites (TSS) of ARGs do not change with neural firing but are already accessible prior to stimulation. Lastly based on binding site enrichment in ARGs, transcription factor mediators of firing were identified and neuronal activity reporters were created. Several of the induced transcription factors encode proteins with high sequence identity to well-known mammalian IEGs: HR38 shows 67% identity to nuclear receptor subfamily four group A member 2 (NR4A2); Stripe has 82% identity to early growth response protein 3 (EGR3); Cabut is 61% identical to Krueppel-like factor 11 (KLF11) (Chen, 2016)
Locomotor activity rhythms are controlled by a network of ~150 circadian neurons within the adult Drosophila brain. They are subdivided based on their anatomical locations and properties. Transcripts "around the clock" were profiled from three key groups of circadian neurons with different functions. A non-circadian outgroup, dopaminergic (tyrosine hydroxylase; TH) neurons, was also profiled. They have cycling transcripts but fewer than clock neurons as well as low expression and poor cycling of clock gene transcripts. This suggests that TH neurons do not have a canonical circadian clock and that their gene expression cycling is driven by brain systemic cues. The three circadian groups are surprisingly diverse in their cycling transcripts and overall gene expression patterns, which include known and putative novel neuropeptides. Even the overall phase distributions of cycling transcripts are distinct, indicating that different regulatory principles govern transcript oscillations. This surprising cell-type diversity parallels the functional heterogeneity of the different neurons (Abruzzi, 2017).
Kohlschutter-Tonz syndrome (KTS) is a rare genetic disorder with neurological dysfunctions including
seizure and intellectual impairment. Mutations at the Rogdi locus have
been linked to development of KTS, yet the underlying mechanisms remain
elusive. This study demonstrates that a Drosophila homolog of Rogdi acts as a
novel sleep-promoting factor by supporting a specific subset of gamma-aminobutyric acid (GABA)
transmission. Rogdi mutant flies displayed insomnia-like behaviors
accompanied by sleep fragmentation and delay in sleep initiation. The
sleep suppression phenotypes were rescued by sustaining GABAergic
transmission primarily via metabotropic GABA receptors or by blocking
wake-promoting dopaminergic pathways. Transgenic rescue further mapped
GABAergic neurons as a cell-autonomous locus important for
Rogdi-dependent sleep, implying metabotropic GABA transmission upstream
of the dopaminergic inhibition of sleep. Consistently, an agonist
specific to metabotropic but not ionotropic GABA receptors titrated the
wake-promoting effects of dopaminergic neuron excitation. Taken
together, these data provide the first genetic evidence that implicates
Rogdi in sleep regulation via GABAergic control of dopaminergic
signaling. Given the strong relevance of GABA to epilepsy, it is proposed that similar mechanisms might underlie the neural pathogenesis of Rogdi-associated KTS (Kim, 2017).
Neurological disorders caused by single-gene mutations are important genetic models to understand how individual genes execute their roles to support the development and function of the brain, as in the case of Kohlschutter-Tönz syndrome (KTS). KTS patients display developmental delays and psychomotor regression. The most prominent symptoms include amelogenesis imperfecta, early-onset seizures, and intellectual disabilities. Linkage analyses followed by genomic sequencing have revealed that most, if not all, KTS patients have homozygous nonsense, frameshift deletion, or splicing site mutations at the Rogdi locus. ROGDI protein expression is not detectable in affected individuals, indicating that the loss of Rogdi function is responsible for the pathogenesis of KTS. In wild-type human tissues, Rogdi transcripts are ubiquitously expressed while the highest enrichment is evident in adult brain and spinal cord. This observation is consistent with the neurological phenotypes observed in KTS patients (Kim, 2017).
Given that Rogdi homologs are relatively well conserved in higher eukaryotes, animal models may facilitate understanding of Rogdi-dependent neural processes. In fact, Rogdi was initially identified in a Drosophila genetic screen as a memory-relevant gene and was thus named after one of Pavlov’s dogs. Sequence analyses of Rogdi homologs revealed a putative leucine zipper (ZIP) motif, which could mediate the dimerization of DNA-binding basic ZIP (bZIP) transcription factors. Interestingly, ROGDI proteins localize to the nuclear envelope in cultured human cells, although they lack basic amino acid residues that are typically located at the N-terminus of the ZIP domain and are required for the DNA-binding and nuclear localization of the bZIP transcription factors. Nonetheless, few or no studies have demonstrated the biological activity of Rogdi and genetic models for Rogdi homologs have not been reported yet. Therefore, how Rogdi exerts its physiological roles particularly in the central nervous system and how its mutation leads to the development of KTS are largely unknown (Kim, 2017).
In the course of genetic studies to elucidate genes and regulatory pathways involved in sleep behaviors, novel sleep mutant alleles were identified in the Drosophila Rogdi gene. This study employed the sleep-promoting effects of Rogdi as a readout of its neural function and demonstrated that Rogdi acts cell-autonomously in GABAergic neurons to enhance metabotropic GABA transmission and thereby sustain sleep. In addition, dopaminergic rescue of Rogdi mutant sleep revealed a novel sleep-regulatory mechanism that functionally links a specific subset of sleep-promoting GABAergic neurons to a wake-promoting dopaminergic pathway. Since epilepsy, a well penetrated phenotype in KTS patients, implicates GABAergic transmission, the current findings provide an important genetic clue to understanding the molecular and neural pathogenesis of KTS (Kim, 2017).
Modeling of neurological diseases and disease-relevant genes has greatly advanced understanding of the fundamental principles that underlie disease pathogenesis as well as brain function. This study has established the first genetic model of the KTS-associated disease gene Rogdi to demonstrate that Rogdi functions as a novel sleep-promoting factor in GABAergic neurons by promoting GABA transmission. While GABA-dependent sleep regulation via ionotropic GABA receptors have been well documented in Drosophila, the data suggest that GABAergic transmission via metabotropic GABA receptors might be primarily compromised by Rogdi mutation. Furthermore, the wake-promoting DA pathway was identified as a neural locus downstream of Rogdi-dependent GABA signaling given that Rogdi mutant sleep could be rescued by pharmacological or genetic manipulation of dopaminergic transmission. This sleep-regulatory pathway was further supported by the observation that wake-promoting effects of TH-expressing dopaminergic neurons could be selectively titrated by an agonist of metabotropic GABA receptors. Rogdi thus defines a novel pathway coupling these two neurotransmitters to promote baseline sleep in Drosophila as exemplified in other behavioral paradigms across species. On the other hand, Rogdi-dependent GABA transmission might have inhibitory effects on a sleep-promoting neural pathway for sleep homeostasis to suppress recovery sleep after sleep loss (Kim, 2017).
What is the molecular basis by which Rogdi supports GABAergic transmission and promotes sleep? A possible role of ROGDI as a transcription factor has been suggested by the nuclear localization of human ROGDI protein, particularly in the nuclear envelope of blood mononuclear cells and dermal fibroblasts, and by the conservation of a putatively dimerizing leucine zipper (ZIP) motif among ROGDI homologs. Several lines of evidence, however, argue against this possibility. bZIP transcription factors possess basic residues followed by their ZIP domains whereas ROGDI protein lacks the canonical motif (i.e., basic residues) for DNA-binding activity and nuclear localization. Drosophila ROGDI actually displays its subcellular distribution in both nucleus and cytoplasm of cultured cells or adult fly neurons although the exclusive nuclear localization might not be a prerequisite for transcriptional activities. The crystal structure of human ROGDI protein showed that, unlike other bZIP transcription factors, human ROGDI protein exists as a monomer containing two structurally distinguishable domains (designated as α and β domains, respectively). The α domain exhibits an α-helical bundle that consists of H1, H2, H3, and H6 helices. In fact, the ZIP-like motif in the α domain appears to mediate their intramolecular interactions, contributing to the overall structure and stability of a monomeric ROGDI protein. Based on sequence homology between Drosophila and human ROGDI proteins, it is predicted that Rogdi[del] allele removes the majority of the first helix including the repeated leucine residues in the α domain and the first three strands in the β domain, explaining the instability of ROGDIdel proteins. A smaller but comparable deletion of the ZIP-like motif has been reported in a KTS patient with a splicing mutation in human Rogdi gene. In addition, a functional study in cervical cancer cell lines demonstrated Rogdi effects on cell cycle progression and radio-sensitivity. However, further investigations will be required to understand how these cellular phenotypes could be linked to the molecular and neural function of ROGDI protein (Kim, 2017).
What will be the relevance of these findings to KTS pathogenesis? Genetic heterogeneity has been reported among KTS patients, indicating that Rogdi-independent genetic mutations could contribute to KTS pathogenesis. A recent study indeed showed that familial mutations in a sodium-citrate transporter gene SLC13A5 are the second genetic cause of KTS. The pathogenic phenotypes commonly found in Rogdi- and SLC13A5-associated KTS gives rise to the intriguing possibility that these two genes might work together to control the intracellular levels of citrate. This idea is further supported by the relevance of citrate metabolism to neurological phenotypes in KTS patients. Neurons are energetically dependent on astrocytes because neurons lack pyruvate carboxylase, an enzyme that converts pyruvate to oxaloacetate in the citric acid cycle. SLC13A5 plays an important role in the transport of glial citrate into neurons to supplement the neuronal citric acid cycle and thereby supply cellular energy. Furthermore, citrate, an intermediate in the citric acid cycle, acts as a precursor of α-ketoglutarate, which can be metabolized to glutamate and GABA, implicating SLC13A5 in the biogenesis of GABA. Consistently, anti-epileptic drugs that elevate GABAergic transmission rescued the seizure phenotypes in SLC13A-associated KTS patients. In addition, this study showed that the pharmacological enhancement of GABAergic transmission by oral administration of GABA-T or GAT inhibitors was sufficient to rescue the short sleep phenotypes in Rogdi mutant flies (Kim, 2017).
These genetic studies strongly implicate Rogdi function in GABAergic transmission, providing the first clue to understanding the neurological phenotypes observed in KTS patients. Molecular and neural deficits selectively caused by Rogdi mutation might explain why seizures in Rogdi-associated KTS are often resistant to anti-epileptic drugs. Future studies should thus address if Rogdi mutant flies display seizure-like behaviors similarly as in KTS patients and if Rogdi-dependent neural relay of GABAergic transmission controls seizure susceptibility in parallel with baseline sleep. In addition, it will be important to determine whether sleep deficiencies are also observed in KTS patients and whether reduced GABAergic transmission in Rogdi- and, possibly, SLC13A5-associated KTS patients is responsible for their neural dysfunctions, including early-onset seizures. Taken together, this genetic model would constitute an important platform for elucidating the molecular and neural pathogenesis underlying KTS and hint towards a precise development of a therapeutic strategy for KTS in the future (Kim, 2017).
Starvation reduces sleep in various animal species including humans and fruit flies. Immediate hunger and the following insufficient nutritional status resulting from starvation may affect sleep and arousal differently. In order to clarify the mechanism underlying the relationship between diet and sleep, the sleep behaviour of Drosophila melanogaster was analyzed that were either starved or fed with different types of sugars. Starved flies showed longer activity bouts, short sleep bouts and a decreased arousal threshold. Non-nutritive sweeteners such as sucralose and arabinose, which are sweet but not nutritive, induced sleep in starved flies, but sleep bout length and the arousal threshold was short and decreased, respectively. On the other hand, sorbitol, which is not sweet but nutritive, did not induce sleep, but slightly increased the lowered arousal threshold. Activation of sweetness receptor expressing neurons induced sleep in starved flies. These results suggest that sweetness alone is sufficient to induce sleep in starved flies and that the nutritional status affects sleep homeostasis by decreasing the arousal threshold, which resulted in short sleep bouts in Drosophila.
Although sleep appears to be broadly conserved in animals, the physiological functions of sleep remain unclear. This study sought to identify a physiological defect common to a diverse group of short-sleeping Drosophila mutants, which might provide insight into the function and regulation of sleep. These short-sleeping mutants share a common phenotype of sensitivity to acute oxidative stress, exhibiting shorter survival times than controls. It was further shown that increasing sleep in wild-type flies using genetic or pharmacological approaches increases survival after oxidative challenge. Moreover, reducing oxidative stress in the neurons of wild-type flies by overexpression of antioxidant genes reduces the amount of sleep. Together, these results support the hypothesis that a key function of sleep is to defend against oxidative stress and also point to a reciprocal role for reactive oxygen species (ROS) in neurons in the regulation of sleep (Hill, 2018).
Circadian clocks coordinate behaviour, physiology and metabolism with Earth's diurnal cycle. These clocks entrain to both light and temperature cycles, and daily environmental temperature oscillations probably contribute to human sleep patterns. However, the neural mechanisms through which circadian clocks monitor environmental temperature and modulate behaviour remain poorly understood. This study has elucidate how the circadian clock neuron network of Drosophila melanogaster processes changes in environmental temperature. In vivo calcium-imaging techniques demonstrate that the posterior dorsal neurons 1 (DN1ps), which are a discrete subset of sleep-promoting clock neurons, constantly monitor modest changes in environmental temperature. These neurons are acutely inhibited by heating and excited by cooling; this is an unexpected result when considering the strong correlation between temperature and light, and the fact that light excites clock neurons. The DN1ps rely on peripheral thermoreceptors located in the chordotonal organs and the aristae. The DN1ps and their thermosensory inputs are required for the normal timing of sleep in the presence of naturalistic temperature cycles. These results identify the DN1ps as a major gateway for temperature sensation into the circadian neural network, which continuously integrates temperature changes to coordinate the timing of sleep and activity (Yadlapalli, 2018).
Circadian clocks display a remarkable combination of imperturbability and sensitivity with regard to temperature. Circadian clocks maintain free-running periods that are strongly temperature-compensated, and brief temperature pulses do not reset the phase of free-running circadian rhythms. Nevertheless, circadian rhythms entrain to low-amplitude step-function temperature cycles. The neural mechanisms by which temperature modulates circadian rhythms are not well understood. This study used Drosophila melanogaster, an organism with a highly conserved yet relatively simple circadian clock neuron network, to investigate how the clock neuron network processes changes in environmental temperature.
To determine whether and how the various clock neuron classes respond to changes in temperature, experiments were performed using CaMPARI15, a fluorescent protein in which photoconversion from green to red is proportional to Ca2+ levels. Individual flies were affixed to a Peltier module and exposed to heating, cooling or constant temperature in the presence of 405-nm light. Upon heating, the DN1ps and DN2s displayed reduced photoconversion compared to controls, whereas the remaining classes showed no significant changes. Cooling resulted in increased photoconversion within the DN1p, DN2 and DN3 classes, whereas the remaining classes showed no changes. These results suggest-in contrast to longstanding expectations-that nodes within the circadian network are unexpectedly sensitive to brief changes in temperature, and reveal that DN1ps are acutely inhibited by heating and excited by cooling. Levels of environmental light and temperature rise and fall concurrently, with temperature lagging slightly behind light. The results show that light and temperature are processed in distinct ways in the clock neuron network: light is excitatory and heating is inhibitory. Cooling had no effect on clock neurons in isolated brains, suggesting a requirement for peripheral thermosensors (Yadlapalli, 2018).
Next, the kinetics of temperature response were examined by monitoring intracellular Ca2+ concentrations ([Ca2+]i) using the Ca2+ sensor GCaMP6m. In agreement with the results from the CaMPARI experiments, the DN1ps showed increases in [Ca2+]i during cooling and decreases in [Ca2+]i during heating, whereas the LNd neurons displayed no significant responses. The DN1ps exhibited significant responses even to modest (4 °C) temperature changes and responded repeatedly to rapid cycles of cooling and heating. To understand whether the DN1ps were tracking the environmental temperature, or were simply excited when the temperature decreased below a threshold, the responses of the DN1ps were measured to a double cooling step, and they were found to respond robustly to both steps. DN1p cooling responses in flies that were entrained to light:dark cycles were higher in the morning than in the evening in a manner dependent on the circadian clock, consistent with previous findings that DN1ps are more excitable in the morning. These results establish that the DN1ps constantly monitor environmental temperature, are excited by cooling and inhibited by heating. At first, these results may appear to contradict recent work describing increases in Ca2+ levels within clock cells in response to heating. However, these studies did not measure the acute physiological responses of clock neurons to brief temperature changes; rather they examined the effects of prolonged increased temperature. The findings reveal that the circadian network transduces brief and transient temperature changes and prolonged increases in temperature in distinct ways (Yadlapalli, 2018).
In Drosophila, thermoreceptors reside in structures in the antennae, called the aristae and the sacculi, and within chordotonal organs in the body. Each arista contains both cold-sensitive and heat-sensitive cells, whereas a sacculus contains only cold-sensitive cells. Each chordotonal organ contains heat-activated cells, which are required for behavioural synchronization to low-amplitude, step-function temperature cycles. In addition, a group of Transient receptor potential cation channel A1 (TrpA1)-expressing anterior cells in the brain act as internal heat sensors. GCaMP imaging experiments were performed on flies from which the aristae or the body had been removed. The responses to cooling and heating were attenuated when the aristae or the body were removed, with only the ipsilateral arista required for normal-amplitude cooling responses. In isolated brains, which lack both the aristae and the body, DN1p cooling responses were completely abolished, whereas slight heat-induced decreases in GCaMP fluorescence persisted. These persistent heating responses appear to be an artefact, as they persist in the absence of neuronal firing, synaptic transmission and anterior-cell neuron function (Yadlapalli, 2018).
Next, using GCaMP imaging, the responses of DN1ps in nocte1 mutants, which have defects in both chordotonal organ structure and temperature entrainment. The responses of DN1ps to temperature changes are reduced in nocte1 mutants, and almost completely abolished in nocte1 mutants from which the aristae were removed. Finally, RNAi-mediated knockdown of nocte expression in the chordotonal organs caused a nocte1-like reduction in the responses of DN1ps to temperature. The chordotonal organs therefore appear to be the primary source of temperature input from the body to the DN1ps. It is concluded that peripheral thermoreceptors in both the aristae and the chordotonal organs contribute to the thermal sensitivity of DN1ps (Yadlapalli, 2018).
The role of the DN1ps in the timing of sleep and activity under constantly cycling temperature was examined, as such cycles are more similar to those found in natural environments than the step-function temperature cycles typically used in the laboratory. Drosophila locomotor activity readily entrains to gradually ramping temperature cycles between 18°C and 25°C that mimic rising temperatures during the morning and falling temperatures during the evening. Under these conditions, flies display locomotor activity that slowly increases throughout the heating phase followed by a precipitous increase in sleep that anticipates the onset of cooling. This sharp transition to sleep, which persists under free-running conditions, is notable considering that the maximal rate of temperature change was only around 0.6°C h-1. Flies entrained to ramping temperature cycles respond to unexpected bouts of heating and cooling with increased activity and sleep, respectively. In the absence of clock function, flies are sensitive to the heating and cooling transitions of ramping temperature cycles: per01 mutants, for example, precipitously increase their activity at the onset of heating and decrease their activity at the onset of cooling, failing to gradually increase activity throughout the heating phase or to anticipate cooling. Outside the laboratory, flies display a mid-afternoon activity peak, with a relationship to the environmental temperature cycle that is markedly similar to that seen for ramping temperature cycles (Yadlapalli, 2018).
The nocte1 mutants and flies that lacked aristae displayed reduced anticipation to cooling and a slow transition to sleep, and nocte1 mutant flies from which aristae were removed showed clearly abnormal behaviour under ramping temperature cycles. Although the nocte1 flies that lacked aristae were able to entrain to light:dark cycles, a majority of them were not able to entrain to temperature cycles and were arrhythmic under constant conditions after the temperature-entrainment period. Similar defects were observed in flies that both lacked aristae and in which nocte had been knocked down in the chordotonal organs. Therefore, the thermoreceptors that are required for the temperature sensitivity of DN1ps are also required for the normal entrainment to ramping temperature cycles, and flies that lack both aristae and chordotonal function are 'circadian blind' to such cycles (Yadlapalli, 2018).
To determine the extent to which the DN1ps are required for the normal pattern of sleep and activity in the presence of ramping temperature cycles, tetanus-toxin (TNT) was exoressed to block synaptic transmission in approximately half of the DN1ps using the Clk4.1m-GAL4 driver. The effects were markedly reminiscent of the effects of the per01 mutation. Consistent with previous reports that identified DN1ps as sleep-promoting neurons,Clk4.1M/UAS-TNT flies were found to sleep less than control flies during the heating phase, but surprisingly, they displayed normal levels of sleep during the cooling phase. glass60j mutants, which lack all DN1ps in addition to all external photoreceptors were tested, and they neither gradually increase their activity during heating nor display cooling-associated increases in sleep, despite the presence of aristae and chordotonal organs. The expression of the toxin within DN2s did not affect the rhythmic behaviour of flies under ramping temperature cycles. Taken together, the results from behavioural experiments are consistent with those of previous work, which established a role for DN1ps in entrainment to step-function temperature cycles, and further revealed that DN1ps have a critical role in the timing of sleep and activity under constantly changing temperature cycles. It is noted, however, that these neurons are not necessary for the response to unexpected bouts of heating and cooling. Finally, the activity peak under ramping temperature cycles is controlled by the DN1p clocks: it is advanced or delayed when the circadian clock is sped up or slowed down specifically in these neurons, respectively (Yadlapalli, 2018).
These results reveal a central role for DN1ps in temperature entrainment and the generation of sleep and activity rhythms in the presence of constantly changing environmental temperature cycles, through the integration of temperature inputs from two distinct peripheral thermosensors. The finding that DN1ps are inhibited by heating and excited by cooling provides important insights into how the clock-neuron network of the fly is likely to coordinate behaviour under constantly changing environmental temperatures. It is proposed that the slow and gradual increase in activity observed during the heating phase of the temperature cycle is promoted through increasing inhibition of the DN1ps, neurons recently shown to promote sleep, whereas the precipitous increase in sleep preceding the onset of cooling is mediated by the excitation of DN1ps. This work reveals how a circadian clock modulates sleep and activity by constantly integrating temperature signals into its neural network. The duration and timing of sleep are closely correlated to both environmental temperature cycles and clock-controlled body temperature rhythms throughout the animal kingdom. It is therefore expected that the mechanisms reported in this study will be broadly relevant to sleep, including sleep in humans (Yadlapalli, 2018).
The fruit fly Drosophila melanogaster survives thermally stressful conditions in a state of reproductive dormancy (diapause), manifested by reduced metabolic activity and arrested ovarian development in females. Unlike insects that rely primarily on photoperiodic stimuli to initiate the diapause program, in this species dormancy is regulated by low temperature and enhanced by shorter photoperiods. Overwintering phenotypes are usually studied under simple laboratory conditions, where animals are exposed to rectangular light-dark (LD) cycles at a constant temperature. This study sought to adopt more realistic diapause protocols by generating LD profiles that better mimic outdoor conditions. Experimental flies were subjected to semi-natural late autumn and summer days, while control females received the same amounts of light but in rectangular LD cycles (LD 8:16 and LD 15:9, respectively). Semi-natural autumnal days were found to induce a higher proportion of females to enter dormancy, while females in semi-natural summer days showed reduced diapause compared with their corresponding rectangular controls, generating an impressive photoperiodic response. In contrast, under rectangular light regimes, the diapause of Drosophila field lines exhibited minimal photoperiodicity. This semi-natural method reveals that D. melanogaster diapause is considerably more photoperiodic than previously believed and suggests that this seasonal response is best studied under simulated natural lighting conditions (Nagi, 2018).
Adel, M., Chen, N., Zhang, Y., Reed, M. L., Quasney, C. and Griffith, L. C. (2022). Pairing-Dependent Plasticity in a Dissected Fly Brain Is Input-Specific and Requires Synaptic CaMKII Enrichment and Nighttime Sleep. J Neurosci 42(21): 4297-4310. PubMed ID: 35474278
Abhilash, L., Ghosh, A. and Sheeba, V. (2019). Selection for timing of eclosion results in co-evolution of temperature responsiveness in Drosophila melanogaster. J Biol Rhythms: 748730419877315. PubMed ID: 31608742
Abhilash, L. and Sharma, V. K. (2020). Mechanisms of photic entrainment of activity/rest rhythms in populations of Drosophila selected for divergent timing of eclosion. Chronobiol Int: 1-16. PubMed ID: 32079418
Abhilash, L., Kalliyil, A. and Sheeba, V. (2020). Responses of activity rhythms to temperature cues evolve in Drosophila populations selected for divergent timing of eclosion. J Exp Biol. PubMed ID: 32291322
Abhilash, L., Shafer, O. T. (2023). Parametric effects of light acting via multiple photoreceptors contribute to circadian entrainment in Drosophila melanogaster. Proceedings Biological sciences, 290(2006):20230149 PubMed ID: 37700655
Aboudhiaf, S., Alves, G., Parrot, S., Amri, M., Simonnet, M. M., Grosjean, Y., Maniere, G. and Seugnet, L. (2018). LAT1-like transporters regulate dopaminergic transmission and sleep in Drosophila. Sleep. PubMed ID: 30016498
Abruzzi, K. C., et al. (2011). Drosophila CLOCK target gene
characterization: implications for circadian tissue-specific gene
expression. Genes Dev. 25(22): 2374-86. PubMed Citation: 22085964
Abruzzi, K. C., Zadina, A., Luo, W., Wiyanto, E., Rahman, R., Guo, F., Shafer, O. and Rosbash, M. (2017). RNA-seq analysis of Drosophila clock and non-clock neurons reveals neuron-specific cycling and novel candidate neuropeptides. PLoS Genet 13(2): e1006613. PubMed ID: 28182648
Adams, G. J. and O'Brien, P. A. (2023). The unified theory of sleep: Eukaryotes endosymbiotic relationship with mitochondria and REM the push-back response for awakening. Neurobiol Sleep Circadian Rhythms 15: 100100. PubMed ID: 37484687
Adewoye, A.B., Kyriacou, C.P. and Tauber, E. (2015). Identification
and functional analysis of early gene expression induced by circadian
light-resetting in Drosophila. BMC Genomics 16: 570. PubMed
ID: 26231660
Adewoye, A. B., Nuzhdin, S. V. and Tauber, E. (2017). Mapping quantitative trait loci underlying circadian light sensitivity in Drosophila. J Biol Rhythms 32(5): 394-405. PubMed ID: 28990443
Agosto, J., Choi, J. C., Parisky, K. M., Stilwell, G., Rosbash, M.
and Griffith, L. C. (2008). Modulation of GABAA receptor desensitization uncouples sleep onset and maintenance in Drosophila. Nat Neurosci 11: 354-359. PubMed ID: 18223647
Afonso, D.J., Liu, D., Machado, D.R., Pan, H., Jepson, J.E., Rogulja,
D. and Koh, K. (2015). TARANIS functions with Cyclin A and Cdk1 in a
novel arousal center to control sleep in Drosophila. Curr
Biol 25(13):1717-26. PubMed ID: 26096977
Alejevski, F., Saint-Charles, A., Michard-Vanhee, C., Martin, B., Galant, S., Vasiliauskas, D. and Rouyer, F. (2019). The HisCl1 histamine receptor acts in photoreceptors to synchronize Drosophila behavioral rhythms with light-dark cycles. Nat Commun 10(1): 252. PubMed ID: 30651542
Alpert, M. H., Gil, H., Para, A. and Gallio, M. (2022). A thermometer circuit for hot temperature adjusts Drosophila behavior to persistent heat. Curr Biol. PubMed ID: 35981537
Altarejos, J. Y. and Montminy, M. (2011). CREB and the CRTC co-activators: sensors for hormonal and metabolic signals. Nat Rev Mol Cell Biol 12: 141-151. PubMed ID: 21346730
Amatobi, K. M., Ozbek-Unal, A. G., Schabler, S., Deppisch, P., Helfrich-Forster, C., Mueller, M. J., Wegener, C. and Fekete, A. (2023). The circadian clock is required for rhythmic lipid transport in Drosophila in interaction with diet and photic condition. J Lipid Res: 100417. PubMed ID: 37481037
Andreani, T., Rosensweig, C., Sisobhan, S., Ogunlana, E., Kath, W. and Allada, R. (2022). Circadian programming of the ellipsoid body sleep homeostat in Drosophila. Elife 11. PubMed ID: 35735904
Andreatta, G., Montagnese, S. and Costa, R. (2022). Natural alleles of the clock gene timeless differentially affect life-history traits in Drosophila. Front Physiol 13: 1092951. PubMed ID: 36703932
Anthoney, N., Tainton-Heap, L. A. L., Luong, H., Notaras, E., Zhao, Q., Perry, T., Batterham, P., Shaw, P. J. and van Swinderen, B. (2023). Experimentally induced active and quiet sleep engage non-overlapping transcriptomes in Drosophila. bioRxiv. PubMed ID: 37066182
Aschoff, J. (1979). Circadian rhythms: influences of internal and external factors on the period measured in constant conditions. Z. Tierpsychol. 49: 225-249. PubMed citation: 386643
Alsehli, A. M., Liao, S., Al-Sabri, M. H., Vasionis, L., Purohit, A., Behare, N., Clemensson, L. E., Williams, M. J. and Schioth, H. B. (2022). The Statin Target HMG-Coenzyme a Reductase (Hmgcr) Regulates Sleep Homeostasis in Drosophila. Pharmaceuticals (Basel) 15(1). PubMed ID: 35056136
Asirim, E. Z., Humberg, T. H., Maier, G. L. and Sprecher, S. G. (2020). Circadian and Genetic Modulation of Visually-Guided Navigation in Drosophila Larvae. Sci Rep 10(1): 2752. PubMed ID: 32066794
Aso, Y., Ray, R. P., Long, X., Bushey, D., Cichewicz, K., Ngo, T. T., Sharp, B., Christoforou, C., Hu, A., Lemire, A. L., Tillberg, P., Hirsh, J., Litwin-Kumar, A. and Rubin, G. M. (2019). Nitric oxide acts as a cotransmitter in a subset of dopaminergic neurons to diversify memory dynamics. Elife 8. PubMed ID: 31724947
Au, D. D., Foden, A. J., Park, S. J., Nguyen, T. H., Liu, J. C., Tran, M. D., Jaime, O. G., Yu, Z. and Holmes, T. C. (2022). Mosquito cryptochromes expressed in Drosophila confer species-specific behavioral light responses. Curr Biol 32(17): 3731-3744. PubMed ID: 35914532
Au, D. D., Liu, J. C., Park, S. J., Nguyen, T. H., Dimalanta, M., Foden, A. J. and Holmes, T. C. (2023). Drosophila photoreceptor systems converge in arousal neurons and confer light responsive robustness. Front Neurosci 17: 1160353. PubMed ID: 37274190
Axelrod, S., Li, X., Sun, Y., Lincoln, S., Terceros, A., O'Neil, J., Wang, Z., Nguyen, A., Vora, A., Spicer, C., Shapiro, B., Young, M. W. (2023). The Drosophila blood-brain barrier regulates sleep via Moody G protein-coupled receptor signaling. Proc Natl Acad Sci U S A, 120(42):e2309331120 PubMed ID: 37831742
Axelrod, S., Li, X., Sun, Y., Lincoln, S., Terceros, A., O'Neil, J., Wang, Z., Nguyen, A., Vora, A., Spicer, C., Shapiro, B., Young, M. W. (2023). The Drosophila blood-brain barrier regulates sleep via Moody G protein-coupled receptor signaling. Proc Natl Acad Sci U S A, 120(42):e2309331120 PubMed ID: 37831742
Bachleitner, W., et al. (2007). Moonlight shifts the endogenous clock
of Drosophila melanogaster. Proc. Natl. Acad. Sci. 104(9): 3538-43. PubMed citation Bai, L. and Sehgal, A. (2015). Anaplastic lymphoma kinase acts in the Drosophila mushroom body to negatively regulate sleep. PLoS Genet 11: e1005611. PubMed ID: 26536237
Baik, L. S., Recinos, Y., Chevez, J. A. and Holmes, T. C. (2018). Circadian modulation of light-evoked avoidance/attraction behavior in Drosophila. PLoS One 13(8): e0201927. PubMed ID: 30106957
Barber, A. F., Erion, R., Holmes, T. C. and Sehgal, A. (2016). Circadian and feeding cues integrate to drive rhythms of physiology in Drosophila insulin-producing cells. Genes Dev 30(23): 2596-2606. PubMed ID: 27979876
Barber, A. F., Fong, S. Y., Kolesnik, A., Fetchko, M. and Sehgal, A. (2021). Drosophila clock cells use multiple mechanisms to transmit time-of-day signals in the brain. Proc Natl Acad Sci U S A 118(10). PubMed ID: 33658368
Bartok, O., Teesalu, M., Ashwall-Fluss, R., Pandey, V., Hanan, M., Rovenko, B.M., Poukkula, M., Havula, E., Moussaieff, A., Vodala, S., Nahmias, Y., Kadener, S. and Hietakangas, V. (2015). The transcription factor Cabut coordinates energy metabolism and the circadian clock in response to sugar sensing. EMBO J 34(11):1538-53. PubMed ID: 25916830
Beckwith, E. J., Geissmann, Q., French, A. S. and Gilestro, G. F. (2017). Regulation of sleep homeostasis by sexual arousal. Elife 6. PubMed ID: 28893376
Beckwith, E. J., Hernando, C. E., Polcownuk, S., Bertolin, A. P., Mancini, E., Ceriani, M. F. and Yanovsky, M. J. (2017). Rhythmic behavior is controlled by the SRm160 splicing factor in Drosophila melanogaster. Genetics [Epub ahead of print]. PubMed ID: 28801530
Belfer, S. J., Bashaw, A. G., Perlis, M. L. and Kayser, M. S. (2019). A Drosophila model of sleep restriction therapy for insomnia. Mol Psychiatry. PubMed ID: 30824866
Benoit, J. B., Bose, J., Ajayi, O. M., Webster, A., Grieshop, K., Lewis, D., Talbott, H., Polak, M. (2025). Shifted levels of sleep and activity during the night as mechanisms underlying ectoparasite resistance. NPJ Biol Timing Sleep, 2(1):15 PubMed ID: 40191448
Berry, J. A., Cervantes-Sandoval, I., Nicholas, E. P. and Davis, R. L. (2012). Dopamine is required for learning and forgetting in Drosophila. Neuron 74: 530-542. PubMed ID: 22578504
Berry, J.A., Cervantes-Sandoval, I., Chakraborty, M. and Davis, R.L. (2015). Sleep facilitates memory by blocking dopamine neuron-mediated forgetting. Cell 161(7):1656-67. PubMed ID: 26073942
Bertolini, E., Schubert, F. K., Zanini, D., Sehadova, H., Helfrich-Forster, C. and Menegazzi, P. (2019). Life at high latitudes does not require circadian behavioral rhythmicity under constant darkness. Curr Biol 29(22): 3928-3936. PubMed ID: 31679928
Bhattacharya, D., Gorska-Andrzejak, J., Abaquita, T. A. L. and Pyza, E. (2023). Effects of adenosine receptor overexpression and silencing in neurons and glial cells on lifespan, fitness, and sleep of Drosophila melanogaster. Exp Brain Res 241(7): 1887-1904. PubMed ID: 37335362
Blake, M.R., Holbrook, S.D., Kotwica-Rolinska, J., Chow, E.S., Kretzschmar, D. and Giebultowicz, J.M. (2015). Manipulations of amyloid precursor protein cleavage disrupt the circadian clock in aging Drosophila. Neurobiol Dis 77: 117-126. PubMed ID: 25766673
Blum, I. D., Keles, M. F., Baz, E. S., Han, E., Park, K., Luu, S., Issa, H., Brown, M., Ho, M. C. W., Tabuchi, M., Liu, S. and Wu, M. N. (2020). Astroglial calcium signaling encodes sleep need in Drosophila. Curr Biol. PubMed ID: 33186550
Boomgarden, A. C., Sagewalker, G. D., Shah, A. C., Haider, S. D., Patel, P., Wheeler, H. E., Dubowy, C. M. and Cavanaugh, D. J. (2019). Chronic circadian misalignment results in reduced longevity and large-scale changes in gene expression in Drosophila. BMC Genomics 20(1): 14. PubMed ID: 30616504
Boto, T., Louis, T., Jindachomthong, K., Jalink, K. and Tomchik, S.
M. (2014). Dopaminergic modulation of cAMP drives nonlinear plasticity
across the Drosophila mushroom body lobes. Curr Biol 24: 822-831.
PubMed ID: 24684937
Brisevac, D., Prakash, C. and Kaiser, T. S. (2023). Genetic analysis of a phenotypic loss in the mechanosensory entrainment of a circalunar clock. PLoS Genet 19(6): e1010763. PubMed ID: 37347791
Brown, M.K., Chan, M.T., Zimmerman, J.E., Pack, A.I., Jackson, N.E.
and Naidoo, N. (2014). Aging induced endoplasmic reticulum stress
alters sleep and sleep homeostasis. Neurobiol Aging 35: 1431-1441.
PubMed ID: 24444805
Buchert, S. N., Murakami, P., Kalavadia, A. H., Reyes, M. T. and Sitaraman, D. (2021).
Sleep correlates with behavioral decision making critical for reproductive output in Drosophila melanogaster. Comp Biochem Physiol A Mol Integr Physiol: 111114. PubMed ID: 34785379
Buhl, E., Bradlaugh, A., Ogueta, M., Chen, K. F., Stanewsky, R. and Hodge, J. J. (2016). Quasimodo mediates daily and acute light effects on Drosophila clock neuron excitability. Proc Natl Acad Sci U S A [Epub ahead of print]. PubMed ID: 27821737
Bulthuis, N., Spontak, K. R., Kleeman, B. and Cavanaugh, D. J. (2019). Neuronal activity in non-LNv clock cells is required to produce free-running rest:activity rhythms in Drosophila. J Biol Rhythms 34(3):249-271. PubMed ID: 30994046
Bushey, D., Tononi, G. and Cirelli, C. (2011). Sleep and synaptic homeostasis: structural evidence in Drosophila. Science 332: 1576-1581. PubMed ID: 21700878
Busza, A., Emery-Le, M., Rosbash, M. and Emery, P. (2004). Roles of the two Drosophila CRYPTOCHROME structural domains in circadian photoreception. Science 304: 1503-1506. PubMed citation: 15178801
Cascallares, G., Riva, S., Franco, D. L., Risau-Gusman, S. and Gleiser, P. M. (2018). Role of the circadian clock in the statistics of locomotor activity in Drosophila. PLoS One 13(8): e0202505. PubMed ID: 30138403
Cavanaugh, D. J., Geratowski, J. D., Wooltorton, J. R., Spaethling,
J. M., Hector, C. E., Zheng, X., Johnson, E. C., Eberwine, J. H. and
Sehgal, A. (2014). Identification of a circadian output circuit for
rest:activity rhythms in Drosophila. Cell 157: 689-701. PubMed ID: 24766812
Cavanaugh, D. J., Vigderman, A. S., Dean, T., Garbe, D. S. and Sehgal, A. (2015). The circadian clock gates sleep through time-of-day dependent modulation of sleep-promoting neurons. Sleep [Epub ahead of print]. PubMed ID: 26350473
Cavey, M., Collins, B., Bertet, C. and Blau, J. (2016). Circadian
rhythms in neuronal activity propagate through output circuits. Nat
Neurosci [Epub ahead of print]. PubMed ID: 26928065
Chatterjee, A., Lamaze, A., De, J., Mena, W., Chelot, E., Martin, B., Hardin, P., Kadener, S., Emery, P. and Rouyer, F. (2018). Reconfiguration of a multi-oscillator network by light in the Drosophila circadian clock. Curr Biol. PubMed ID: 29910074
Chaturvedi, R., Stork, T., Yuan, C., Freeman, M. R. and Emery, P. (2022). Astrocytic GABA transporter controls sleep by modulating GABAergic signaling in Drosophila circadian neurons. Curr Biol. PubMed ID: 35303417
Cheli, V. T., Daniels, R. W., Godoy, R., Hoyle, D. J., Kandachar, V., Starcevic, M., Martinez-Agosto, J. A., Poole, S., DiAntonio, A., Lloyd, V. K., Chang, H. C., Krantz, D. E., Dell'Angelica, E. C. (2010). Genetic modifiers of abnormal organelle biogenesis in a Drosophila model of BLOC-1 deficiency. Hum Mol Genet, 19(5):861-878 PubMed ID: 20015953
Chen, C., Buhl, E., Xu, M., Croset, V., Rees, J. S., Lilley, K. S., Benton, R., Hodge, J. J. and Stanewsky, R. (2015). Drosophila Ionotropic Receptor 25a mediates circadian clock resetting by temperature. Nature 527: 516-520. PubMed ID: 26580016
Chen, C., Xu, M., Anantaprakorn, Y., Rosing, M. and Stanewsky, R. (2018). nocte is required for integrating light and temperature inputs in circadian clock neurons of Drosophila. Curr Biol 28(10): 1595-1605. PubMed ID: 29754901
Chen, K. F., Lowe, S., Lamaze, A., Kratschmer, P. and Jepson, J. (2019). Neurocalcin regulates nighttime sleep and arousal in Drosophila. Elife 8. PubMed ID: 30865587
Chen, N., Zhang, Y., Rivera-Rodriguez, E. J., Yu, A. D., Hobin, M., Rosbash, M. and Griffith, L. C. (2023). Widespread posttranscriptional regulation of cotransmission. Sci Adv 9(22): eadg9836. PubMed ID: 37267358
Chen, S. C., Tang, X., Goda, T., Umezaki, Y., Riley, A. C., Sekiguchi, M., Yoshii, T. and Hamada, F. N. (2022). Dorsal clock networks drive temperature preference rhythms in Drosophila. Cell Rep 39(2): 110668. PubMed ID: 35417715
Chen, W., Xue, Y., Scarfe, L., Wang, D. and Zhang, Y. (2019). Loss of Prune in circadian cells decreases the amplitude of the circadian locomotor rhythm in Drosophila. Front Cell Neurosci 13: 76. PubMed ID: 30881291
Chen, W. F., Maguire, S., Sowcik, M., Luo, W., Koh, K. and Sehgal, A. (2015). A neuron-glia interaction involving GABA transaminase contributes to sleep loss in sleepless mutants. Mol Psychiatry 20(2): 240-251. PubMed ID: 24637426
Chen, X., Rahman, R., Guo, F. and Rosbash, M. (2016). Genome-wide identification of neuronal activity-regulated genes in Drosophila. Elife 5. PubMed ID: 27936378
Chen, X. and Rosbash, M. (2017). MicroRNA-92a is a circadian modulator of neuronal excitability in Drosophila. Nat Commun 8: 14707. PubMed ID: 28276426
Chen, X., Ma, W., Zhang, S., Paluch, J., Guo, W., Dickman, D. K. (2017). The BLOC-1 Subunit Pallidin Facilitates Activity-Dependent Synaptic Vesicle Recycling. eNeuro, 4(1) PubMed ID: 28317021
Chen, X., Li, J., Gao, Z., Yang, Y., Kuang, W., Dong, Y., Chua, G. H., Huang, X., Jiang, B., Tian, H., Wang, Y., Huang, X., Li, Y., Lam, S. M. and Shui, G. (2022). Endogenous ceramide phosphoethanolamine modulates circadian rhythm via neural-glial coupling in Drosophila. Natl Sci Rev 9(12): nwac148. PubMed ID: 36713590
Cho, E., Oh, J. H., Lee, E., Do, Y. R. and Kim, E. Y. (2016). Cycles of circadian illuminance are sufficient to entrain and maintain circadian locomotor rhythms in Drosophila. Sci Rep 6: 37784. PubMed ID: 27883065
Cho, E., Lee, E. and Kim, E. Y. (2016). Diversification of molecular clockwork for tissue specific function: insight from novel Drosophila Clock mutant homologous to mouse Clock allele. BMB Rep 49(11):587-589. PubMed ID: 27756446
Cho, E., Kwon, M., Jung, J., Hyun Kang, D., Jin, S., Choi, S. E., Kang, Y. and Kim, E. Y. (2019). AMP-activated protein kinase regulates circadian rhythm by affecting CLOCK in Drosophila. J Neurosci. PubMed ID: 30819799
Chouhan, N. S., Wolf, R. and Heisenberg, M. (2017). Starvation promotes odor/feeding-time associations in flies. Learn Mem 24(7): 318-321. PubMed ID: 28620079
Chouhan, N. S., Griffith, L. C., Haynes, P. and Sehgal, A. (2021). Availability of food determines the need for sleep in memory consolidation. Nature 589(7843): 582-585. PubMed ID: 33268891
Chouhan, N. S. and Sehgal, A. (2022). Consolidation of Sleep-Dependent Appetitive Memory Is Mediated by a Sweet-Sensing Circuit. J Neurosci 42(18): 3856-3867. PubMed ID: 35361706
Chow, E. S., Long, D. M. and Giebultowicz, J. M. (2016). Circadian rhythm in mRNA expression of the glutathione synthesis gene Gclc is controlled by peripheral glial clocks in Drosophila melanogaster. C. Physiol Entomol 41(4): 369-377. PubMed ID: 28503020
Chung, B. Y, et al. (2009). The GABAA receptor RDL acts in
peptidergic PDF neurons to promote sleep in Drosophila. Curr. Biol.
19: 386-390. PubMed Citation: 19230663
Churgin, M. A., Szuperak, M., Davis, K. C., Raizen, D. M., Fang-Yen, C. and Kayser, M. S. (2019). Quantitative imaging of sleep behavior in Caenorhabditis elegans and larval Drosophila melanogaster. Nat Protoc. PubMed ID: 30953041
Chyb, S., et al. (2009). Modulation of the light response by cAMP in
Drosophila photoreceptors. J. Neurosci. 19: 8799-8807. PubMed
Citation: 10516299
Claridge-Chang, A., et al. (2001). Circadian regulation of gene
expression systems in the Drosophila head. Neuron 32: 657-671. 11719206
Colizzi, F. S., Veenstra, J. A., Rezende, G. L., Helfrich-Forster, C. and Martinez-Torres, D. (2023). Pigment-dispersing factor is present in circadian clock neurons of pea aphids and may mediate photoperiodic signalling to insulin-producing cells. Open Biol 13(6): 230090. PubMed ID: 37369351
Collins, B., Mazzoni, E. O., Stanewsky, R. and Blau, J. (2006). Drosophila CRYPTOCHROME is a circadian transcriptional repressor. Curr. Biol. 16: 441-449. PubMed citation: 16527739
Collins, B., Kane, E. A., Reeves, D. C., Akabas, M. H. and Blau, J.
(2012). Balance of activity between LNvs and glutamatergic dorsal clock neurons promotes robust circadian rhythms in Drosophila. Neuron 74(4): 706-18. PubMed Citation: 22632728
Crocker, A., Shahidullah, M., Levitan, I. B. and Sehgal, A. (2010).
Identification of a neural circuit that underlies the effects of
octopamine on sleep:wake behavior. Neuron 65: 670-681. PubMed ID: 20223202
Cusumano, P., Biscontin, A., Sandrelli, F., Mazzotta, G. M., Tregnago, C., De Pitta, C. and Costa, R. (2018). Modulation of miR-210 alters phasing of circadian locomotor activity and impairs projections of PDF clock neurons in Drosophila melanogaster. PLoS Genet 14(7): e1007500. PubMed ID: 30011269
Dag, U., Lei, Z., Le, J. Q., Wong, A., Bushey, D. and Keleman, K. (2019). Neuronal reactivation during post-learning sleep consolidates long-term memory in Drosophila. Elife 8 pii: e42786. PubMed ID: 30801246
Dahdal, D., Reeves, D. C., Ruben, M., Akabas, M. H. and Blau, J.
(2010). Drosophila pacemaker neurons require g protein signaling and
GABAergic inputs to generate twenty-four hour behavioral rhythms.
Neuron 68: 964-977. PubMed ID: 21145008
Damulewicz, M. and Pyza, E. (2011). The clock input to the first
optic neuropil of Drosophila melanogaster expressing neuronal
circadian plasticity. PLoS One 6(6): e21258. PubMed Citation: 21760878
Damulewicz, M., Loboda, A., Jozkowicz, A., Dulak, J. and Pyza, E. (2017). Haeme oxygenase protects against UV light DNA damages in the retina in clock-dependent manner. Sci Rep 7(1): 5197. PubMed ID: 28701782
Damulewicz, M., Swiatek, M., Loboda, A., Dulak, J., Bilska, B., Przewlocki, R. and Pyza, E. (2018). Daily regulation of phototransduction, circadian clock, DNA repair, and immune gene expression by Heme Oxygenase in the retina of Drosophila. Genes (Basel) 10(1). PubMed ID: 30583479
Damulewicz, M., Ispizua, J. I., Ceriani, M. F. and Pyza, E. M. (2020). Communication Among Photoreceptors and the Central Clock Affects Sleep Profile. Front Physiol 11: 993. PubMed ID: 32848895
Damulewicz, M., Woznicka, O., Jasinska, M. and Pyza, E. (2020). CRY-dependent plasticity of tetrad presynaptic sites in the visual system of Drosophila at the morning peak of activity and sleep. Sci Rep 10(1): 18161. PubMed ID: 33097794
Dapergola, E., Menegazzi, P., Raabe, T. and Hovhanyan, A. (2021). Light Stimuli and Circadian Clock Affect Neural Development in Drosophila melanogaster. Front Cell Dev Biol 9: 595754. PubMed ID: 33763414
De, J., Wu, M., Lambatan, V., Hua, Y. and Joiner, W. J. (2023). Re-examining the role of the dorsal fan-shaped body in promoting sleep in Drosophila. Curr Biol. PubMed ID: 37552985
de Azevedo, R. V. M., Hansen, C., Chen, K. F., Rosato, E. and Kyriacou, C. P. (2020). Disrupted Glutamate Signaling in Drosophila Generates Locomotor Rhythms in Constant Light. Front Physiol 11: 145. PubMed ID: 32210832
Delbare, S. Y. N., Venkatraman, S., Scuderi, K., Wells, M. T., Wolfner, M. F., Basu, S. and Clark, A. G. (2023). Time series transcriptome analysis implicates the circadian clock in the Drosophila melanogaster female's response to sex peptide. Proc Natl Acad Sci U S A 120(5): e2214883120. PubMed ID: 36706221
Delventhal, R., O'Connor, R. M., Pantalia, M. M., Ulgherait, M., Kim, H. X., Basturk, M. K., Canman, J. C. and Shirasu-Hiza, M. (2019). Dissection of central clock function in Drosophila through cell-specific CRISPR-mediated clock gene disruption. Elife 8. PubMed ID: 31613218
Depetris-Chauvin, A., et al. (2011). Adult-specific electrical silencing of pacemaker neurons uncouples molecular clock from circadian outputs. Curr. Biol. 21(21): 1783-93. PubMed Citation: 22018542
Deppisch, P., Prutscher, J. M., Pegoraro, M., Tauber, E., Wegener, C. and Helfrich-Forster, C. (Deppisch2022). Adaptation of Drosophila melanogaster to Long Photoperiods of High-Latitude Summers Is Facilitated by the ls-Timeless Allele. J Biol Rhythms 37(2): 185-201. PubMed ID: 35301885
Diaz, M. M., Schlichting, M., Abruzzi, K. C., Long, X. and Rosbash, M. (2019). Allatostatin-C/AstC-R2 is a novel pathway to modulate the circadian activity pattern in Drosophila. Curr Biol 29(1): 13-22.e13. PubMed ID: 30554904
Di Cara, F. and King-Jones, K. (2016). The Circadian clock is a key driver of steroid hormone production in Drosophila. Curr Biol [Epub ahead of print]. PubMed ID: 27546572
Delevoye, C., Heiligenstein, X., Ripoll, L., Gilles-Marsens, F., Dennis, M. K., Linares, R. A., Derman, L., Gokhale, A., Morel, E., Faundez, V., Marks, M. S., Raposo, G. (2016). BLOC-1 Brings Together the Actin and Microtubule Cytoskeletons to Generate Recycling Endosomes. Curr Biol, 26(1):1-13 PubMed ID: 26725201
Dickman, D. K., Tong, A. and Davis, G. W. (2012). Snapin is critical for presynaptic homeostatic plasticity. J Neurosci 32: 8716-8724. PubMed ID: 22723711
Dissel, S., et al. (2004). A constitutively active cryptochrome in
Drosophila melanogaster. Nat. Neurosci. 7: 834-840. PubMed citation: 15258584
Dissel, S., Seugnet, L., Thimgan, M. S., Silverman, N., Angadi, V., Thacher, P. V., Burnham, M. M. and Shaw, P. J. (2015). Differential activation of immune factors in neurons and glia contribute to individual differences in resilience/vulnerability to sleep
disruption. Brain Behav Immun 47: 75-85. PubMed ID: 25451614
Dissel, S., Morgan, E., Duong, V., Chan, D., van Swinderen, B., Shaw, P. and Zars, T. (2020). Sleep restores place learning to the adenylyl cyclase mutant rutabaga. J Neurogenet: 1-9. PubMed ID: 31997683
Dollish, H. K., Kaladchibachi, S., Negelspach, D. C. and Fernandez, F. X. (2022). The Drosophila circadian phase response curve to light: Conservation across seasonally relevant photoperiods and anchorage to sunset. Physiol Behav 245: 113691. PubMed ID: 34958825
Donlea, J. M., Ramanan, N. and Shaw, P. J. (2009). Use-dependent plasticity in clock neurons regulates sleep need in Drosophila. Science 324: 105-108. PubMed ID: 19342592
Donlea, J. M., Thimgan, M. S., Suzuki, Y., Gottschalk, L. and Shaw,
P. J. (2011). Inducing sleep by remote control facilitates memory
consolidation in Drosophila. Science 332: 1571-1576. PubMed ID: 21700877
Donlea, J. M., Ramanan, N., Silverman, N. and Shaw, P. J. (2014a). Genetic rescue of functional senescence in synaptic and behavioral plasticity. Sleep 37(9): 1427-37. PubMed ID: 25142573
Donlea, J. M., Pimentel, D. and Miesenbock, G. (2014b). Neuronal
machinery of sleep homeostasis in Drosophila. Neuron 81: 860-872.
PubMed ID: 24559676
Donlea, J. M., Pimentel, D., Talbot, C. B., Kempf, A., Omoto, J. J., Hartenstein, V. and Miesenbock, G. (2018). Recurrent Circuitry for Balancing Sleep Need and Sleep. Neuron 97(2): 378-389 e374. PubMed ID: 29307711
Dreyer, A. P., Martin, M. M., Fulgham, C. V., Jabr, D. A., Bai, L., Beshel, J. and Cavanaugh, D. J. (2019). A circadian output center controlling feeding:fasting rhythms in Drosophila. PLoS Genet 15(11): e1008478. PubMed ID: 31693685
Driscoll, M., Buchert, S. N., Coleman, V., McLaughlin, M., Nguyen, A. and Sitaraman, D. (2021). Compartment specific regulation of sleep by mushroom body requires GABA and dopaminergic signaling. Sci Rep 11(1): 20067. PubMed ID: 34625611
Du, J., Zhang, Y., Xue, Y., Zhao, X., Zhao, X., Wei, Y., Li, Z., Zhang, Y. and Zhao, Z. (2018). Diurnal protein oscillation profiles in Drosophila head. FEBS Lett. PubMed ID: 30311939
Dubowy, C., Moravcevic, K., Yue, Z., Wan, J. Y., Van Dongen, H. P. and Sehgal, A. (2016). Genetic dissociation of daily sleep and sleep following thermogenetic sleep deprivation in Drosophila. Sleep [Epub ahead of print]. PubMed ID: 26951392
Duhart, J. M., Herrero, A., de la Cruz, G., Ispizua, J. I., Pirez, N. and Ceriani, M. F. (2020a). Circadian structural plasticity drives remodeling of E cell output. Curr Biol. PubMed ID: 33065014
Duhart, J. M., Baccini, V., Zhang, Y., Machado, D. R. and Koh, K. (2020b). Modulation of sleep-courtship balance by nutritional status in Drosophila. Elife 9. PubMed ID: 33084567
Duhart, J. M., Buchler, J. R., Inami, S., Kennedy, K. J., Jenny, B. P., Afonso, D. J. S. and Koh, K. (2023). Modulation and neural correlates of postmating sleep plasticity in Drosophila females. Curr Biol 33(13): 2702-2716. PubMed ID: 37352854
Durkin, J., Poe, A. R., Belfer, S. J., Rodriguez, A., Tang, S. H., Walker, J. A. and Kayser, M. S. (2023). Neurofibromin 1 regulates early developmental sleep in Drosophila. Neurobiol Sleep Circadian Rhythms 15: 100101. PubMed ID: 37593040
Dusik, V., Senthilan, P. R., Mentzel, B., Hartlieb, H., Wulbeck, C., Yoshii, T., Raabe, T. and Helfrich-Forster, C. (2014). The MAP kinase p38 is part of Drosophila melanogaster's circadian clock. PLoS Genet 10: e1004565. PubMed ID: 25144774 Duvall, L. B. and Taghert, P. H. (2012). The circadian neuropeptide PDF signals preferentially through a specific adenylate cyclase isoform AC3 in M pacemakers of Drosophila. PLoS Biol 10(6): e1001337. PubMed ID: 22679392
Duvall, L. B. and Taghert, P. H. (2013). E and M circadian pacemaker neurons use different PDF receptor signalosome components in drosophila. J Biol Rhythms 28(4): 239-248. PubMed ID: 23929551
Duvall, L. B. and Taghert, P. H. (2013). E and M circadian pacemaker neurons use different PDF receptor signalosome components in drosophila. J Biol Rhythms 28(4): 239-248. PubMed ID: 23929551
Eck, S., Helfrich-Förster, C. and Rieger, D. (2016). The timed depolarization of morning and evening oscillators phase shifts the circadian clock of Drosophila. J Biol Rhythms [Epub ahead of print]. PubMed ID: 27269519
Eick, A. K., Ogueta, M., Buhl, E., Hodge, J. J. L. and Stanewsky, R. (2022). The opposing chloride cotransporters KCC and NKCC control locomotor activity in constant light and during long days. Curr Biol 32(6): 1420-1428.e1424. PubMed ID: 35303416
El Alaoui, A. A., Buhl, E., Galizia, S., Hodge, J. J. L., de Vivo, L. and Bellesi, M. (2023). Increased interaction between endoplasmic reticulum and mitochondria following sleep deprivation. BMC Biol 21(1): 1. PubMed ID: 36600217
Enell, L., Hamasaka, Y., Kolodziejczyk, A. and Nassel, D. R. (2007). gamma-Aminobutyric acid (GABA) signaling components in Drosophila: immunocytochemical localization of GABA(B) receptors in relation to the GABA(A) receptor subunit RDL and a vesicular GABA transporter. J Comp Neurol 505: 18-31. PubMed ID: 17729251
Ertekin, D., Kirszenblat, L., Faville, R. and van Swinderen, B. (2020). Down-regulation of a cytokine secreted from peripheral fat bodies improves visual attention while reducing sleep in Drosophila. PLoS Biol 18(8): e3000548. PubMed ID: 32745077
Estrella, M. A., Du, J., Chen, L., Rath, S., Prangley, E., Chitrakar, A., Aoki, T., Schedl, P., Rabinowitz, J. and Korennykh, A. (2019). The metabolites NADP(+) and NADPH are the targets of the circadian protein Nocturnin (Curled). Nat Commun 10(1): 2367. PubMed ID: 31147539
Feng, G., Zhang, J., Li, M., Shao, L., Yang, L., Song, Q. and Ping, Y. (2018). Control of sleep onset by Shal/Kv4 channels in Drosophila circadian neurons. J Neurosci. PubMed ID: 30185460
Ferguson, L., Petty, A., Rohrscheib, C., Troup, M., Kirszenblat, L., Eyles, D. W. and van Swinderen, B. (2017). Transient dysregulation of dopamine signaling in a developing Drosophila arousal circuit permanently impairs behavioral responsiveness in adults. Front Psychiatry 8: 22. PubMed ID: 28243212
Fernandez, M. P., Pettibone, H. L., Bogart, J. T., Roell, C. J., Davey, C. E., Pranevicius, A., Huynh, K. V., Lennox, S. M., Kostadinov, B. S. and Shafer, O. T. (2020). Sites of Circadian Clock Neuron Plasticity Mediate Sensory Integration and Entrainment. Curr Biol. PubMed ID: 32386535
Fernandez-Chiappe, F., Frenkel, L., Colque, C. C., Ricciuti, A., Hahm, B., Cerredo, K., Muraro, N. I. and Ceriani, M. F. (2020). High frequency neuronal bursting is essential for circadian and sleep behaviors in Drosophila. J Neurosci. PubMed ID: 33262246
Flores, C. C., Loschky, S. S., Marshall, W., Spano, G. M., Cenere, M. M., Tononi, G. and Cirelli, C. (2021). Identification of Ultrastructural Signatures of Sleep and Wake in the Fly Brain. Sleep. PubMed ID: 34536282
Flores-Valle, A., Goncalves, P. J. and Seelig, J. D. (2021). Integration of sleep homeostasis and navigation in Drosophila. PLoS Comput Biol 17(7): e1009088. PubMed ID: 34252086
Flourakis, M., Kula-Eversole, E., Hutchison, A. L., Han, T. H., Aranda, K., Moose, D. L., White, K. P., Dinner, A. R., Lear, B. C., Ren, D., Diekman, C. O., Raman, I. M. and Allada, R. (2015). A conserved bicycle model for circadian clock control of membrane excitability. Cell 162: 836-848. PubMed ID: 26276633
Fogg, P. C., O'Neill, J. S., Dobrzycki, T., Calvert, S., Lord, E. C., McIntosh, R. L., Elliott, C. J., Sweeney, S. T., Hastings, M. H. and Chawla, S. (2014). Class IIa histone deacetylases are conserved regulators of circadian function. J Biol Chem [Epub ahead of print]. PubMed ID: 25271152
Fogle, K. J., Baik, L. S., Houl, J. H., Tran, T. T., Roberts, L., Dahm, N. A., Cao, Y., Zhou, M. and Holmes, T. C. (2015). CRYPTOCHROME-mediated phototransduction by modulation of the potassium ion channel β-subunit redox sensor. Proc Natl Acad Sci U S A 112(7): 2245-50. PubMed ID: 25646452
Foley, L., Ling, J., Joshi, R., Evantal, N., Kadener, S. and Emery, P. (2019). Drosophila PSI controls circadian period and the phase of circadian behavior under temperature cycle via tim splicing. Elife 8. PubMed ID: 31702555
Frenkel, L., Muraro, N. I., Beltran Gonzalez, A. N., Marcora, M. S., Bernabo, G., Hermann-Luibl, C., Romero, J. I., Helfrich-Forster, C., Castano, E. M., Marino-Busjle, C., Calvo, D. J. and Ceriani, M. F. (2017). Organization of circadian behavior relies on glycinergic transmission. Cell Rep 19(1): 72-85. PubMed ID: 28380364
Fujii, S., Emery, P. and Amrein, H. (2017). SIK3-HDAC4 signaling regulates Drosophila circadian male sex drive rhythm via modulating the DN1 clock neurons. Proc Natl Acad Sci U S A 114(32): E6669-e6677. PubMed ID: 28743754
Fulgham, C. V., Dreyer, A. P., Nasseri, A., Miller, A. N., Love, J., Martin, M. M., Jabr, D. A., Saurabh, S. and Cavanaugh, D. J. (2021).
Central and Peripheral Clock Control of Circadian Feeding Rhythms.
J Biol Rhythms: 7487304211045835. PubMed ID: 34547954
Galvez, T., Duthey, B., Kniazeff, J., Blahos, J., Rovelli, G., Bettler, B., Prezeau, L. and Pin, J. P. (2001). Allosteric interactions between GB1 and GB2 subunits are required for optimal GABA(B) receptor function. EMBO J 20: 2152-2159. PubMed ID: 11331581
Garbe, D.S., Bollinger, W.L., Vigderman, A., Masek, P., Gertowski, J., Sehgal, A. and Keene, A.C. (2015). Context-specific comparison of sleep acquisition systems in Drosophila. Biol Open [Epub ahead of print]. PubMed ID: 26519516
Gorostiza, E. A., Depetris-Chauvin, A., Frenkel, L., Pirez, N. and Ceriani, M. F. (2014). Circadian pacemaker neurons change synaptic contacts across the day. Curr Biol 24(18): 2161-2167. PubMed ID: 25155512
Guo, F., Yu, J., Jung, H. J., Abruzzi, K. C., Luo, W., Griffith, L. C. and Rosbash, M. (2016). Circadian neuron feedback controls the Drosophila sleep--activity profile. Nature 536(7616): 292-297. PubMed ID: 27479324
Gardner, B., Strus, E., Meng, Q. C., Coradetti, T., Naidoo, N. N., Kelz, M. B. and Williams, J. A. (2015). Sleep homeostasis and general anesthesia: Are fruit flies well rested after emergence from Propofol?. Anesthesiology [Epub ahead of print]. PubMed ID: 26556728
Geissmann, Q., Beckwith, E. J. and Gilestro, G. F. (2019). Most sleep does not serve a vital function: Evidence from Drosophila melanogaster. Sci Adv 5(2): eaau9253. PubMed ID: 30801012
Giesecke, A., Johnstone, P. S., Lamaze, A., Landskron, J., Atay, E., Chen, K. F., Wolf, E., Top, D. and Stanewsky, R. (2023). A novel period mutation implicating nuclear export in temperature compensation of the Drosophila circadian clock. Curr Biol 33(2): 336-350.e335. PubMed ID: 36584676
Gilestro, G. F., Tononi, G. and Cirelli, C. (2009). Widespread changes in synaptic markers as a function of sleep and wakefulness in Drosophila. Science 324: 109-112. PubMed ID: 19342593
Glaser, F. T. and Stanewsky, R. (2005). Temperature synchronization of the Drosophila circadian clock. Curr. Biol. 15: 1352-1363. PubMed Citation: 16085487
Gmeiner, F., Kolodziejczyk, A., Yoshii, T., Rieger, D., Nassel, D. R., Helfrich-Forster, C. (2013). GABAB receptors play an essential role in maintaining sleep during the second half of the night in Drosophila melanogaster. J Exp Biol 216: 3837-3843. PubMed ID: 24068350
Goda, T., Tang, X., Umezaki, Y., Chu, M. L. and Hamada, F. N. (2016). Drosophila DH31 neuropeptide and PDF receptor regulate night-onset temperature preference. J Neurosci 36: 11739-11754. PubMed ID: 27852781
Goda, T. and Hamada, F. N. (2019). Drosophila temperature preference rhythms: An innovative model to understand body temperature rhythms. Int J Mol Sci 20(8). PubMed ID: 31018551
Goh, G. H., Blache, D., Mark, P. J., Kennington, W. J. and Maloney, S. K. (2021). Daily temperature cycles prolong lifespan and have sex-specific effects on peripheral clock gene expression in Drosophila melanogaster. J Exp Biol. PubMed ID: 33758022
Goldsmith, S. L. and Newfeld, S. J. (2023). dSmad2 differentially regulates dILP2 and dILP5 in insulin producing and circadian pacemaker cells in unmated adult females. PLoS One 18(1): e0280529. PubMed ID: 36689407
Gorostiza, E. A., Depetris-Chauvin, A., Frenkel, L., Pirez, N., Ceriani, M. F. (2014) Circadian pacemaker neurons change synaptic contacts across the day. Curr Biol. PubMed ID: 25155512
Grima, B., Chelot, E., Xia. R. and Rouyer, F. (2004). Morning and evening peaks of activity rely on different clock neurons of the Drosophila brain. Nature 431: 869-873. PubMed citation: 15483616
Guo, F., Yu, J., Jung, H.J., Abruzzi, K.C., Luo, W., Griffith, L.C. and Rosbash, M. (2016). Circadian neuron feedback controls the Drosophila sleep-activity profile. Nature [Epub ahead of print]. PubMed ID: 27479324
Guo, F., Chen, X. and Rosbash, M. (2017). Temporal calcium profiling of specific circadian neurons in freely moving flies. Proc Natl Acad Sci U S A 114(41): E8780-e8787. PubMed ID: 28973886
Guo, F., Holla, M., Diaz, M. M. and Rosbash, M. (2018). A circadian output circuit controls sleep-wake arousal in Drosophila. Neuron. PubMed ID: 30269992
Guo, X., Yu, Z. and Yin, D. (2023). Sex-dependent obesogenic effect of tetracycline on Drosophila melanogaster deteriorated by dysrhythmia. J Environ Sci (China) 124: 472-480. PubMed ID: 36182155
Gupta, T., Morgan, H. R., Bailey, J. A. and Certel, S. J. (2016). Functional conservation of MBD proteins: MeCP2 and Drosophila MBD proteins alter sleep. Genes Brain Behav [Epub ahead of print]. PubMed ID: 27489246
Hamasaka, Y., Wegener, C. and Nassel, D. R. (2005). GABA modulates Drosophila circadian clock neurons via GABAB receptors and decreases in calcium. J Neurobiol 65: 225-240. PubMed ID: 16118795
Hao, H., Song, L. and Zhang, L. (2023). Wolfram syndrome 1 regulates sleep in dopamine receptor neurons by modulating calcium homeostasis. PLoS Genet 19(7): e1010827. PubMed ID: 37399203
Hao, Y., Shao, L., Hou, J., Zhang, Y., Ma, Y., Liu, J., Xu, C., Chen, F., Cao, L. H. and Ping, Y. (2023). Resveratrol and Sir2 Reverse Sleep and Memory Defects Induced by Amyloid Precursor Protein. Neurosci Bull. PubMed ID: 37041405
Harbison, S. T., Serrano Negron, Y. L., Hansen, N. F. and Lobell, A. S. (2017). Selection for long and short sleep duration in Drosophila melanogaster reveals the complex genetic network underlying natural variation in sleep. PLoS Genet 13(12): e1007098. PubMed ID: 29240764
Harrisingh, M. C., Wu, Y., Lnenicka, G. A. and Nitabach, M. N. (2007). Intracellular Ca2+ regulates free-running circadian clock oscillation in vivo. J Neurosci 27(46): 12489-12499. PubMed ID: 18003827
Hasegawa, T., Tomita, J., Hashimoto, R., Ueno, T., Kume, S. and Kume, K. (2017). Sweetness induces sleep through gustatory signalling independent of nutritional value in a starved fruit fly. Sci Rep 7(1): 14355. PubMed ID: 29084998
He, L., Wu, B., Shi, J., Du, J. and Zhao, Z. (2023). Regulation of feeding and energy homeostasis by clock-mediated Gart in Drosophila. Cell Rep 42(8): 112912. PubMed ID: 37531254
He, Q., Du, J., Wei, L. and Zhao, Z. (2020). AKH-FOXO pathway regulates starvation-induced sleep loss through remodeling of the small ventral lateral neuron dorsal projections. PLoS Genet 16(10): e1009181. PubMed ID: 33104699
Helfrich-Forster, C., Yoshii, T., Wulbeck, C., Grieshaber, E., Rieger, D., Bachleitner, W., Cusamano, P. and Rouyer, F. (2007). The lateral and dorsal neurons of Drosophila melanogaster: new insights about their morphology and function. Cold Spring Harb Symp Quant Biol 72: 517-525. PubMed ID: 18419311
Herrero, A., Duhart, J. M. and Ceriani, M. F. (2017). Neuronal and glial clocks underlying structural remodeling of pacemaker neurons in Drosophila. Front Physiol 8: 918. PubMed ID: 29184510
Hidalgo, S., Anguiano, M., Tabuloc, C. A. and Chiu, J. C. (2023). Seasonal cues act through the circadian clock and pigment-dispersing factor to control EYES ABSENT and downstream physiological changes. Curr Biol. PubMed ID: 36708710
Hill, V. M., O'Connor, R. M., Sissoko, G. B., Irobunda, I. S., Leong, S., Canman, J. C., Stavropoulos, N. and Shirasu-Hiza, M. (2018). A bidirectional relationship between sleep and oxidative stress in Drosophila. PLoS Biol 16(7): e2005206. PubMed ID: 30001323
Hirsh, J., et al. (2010). Roles of dopamine in circadian rhythmicity and extreme light sensitivity of circadian entrainment. Curr. Biol. 20: 209-214. PubMed Citation: 20096587
Hobin, M., Dorfman, K., Adel, M., Rivera-Rodriguez, E. J., Kuklin, E. A., Ma, D. and Griffith, L. C. (2022). The Drosophila microRNA bantam regulates excitability in adult mushroom body output neurons to promote early night sleep. iScience 25(9): 104874. PubMed ID: 36034229
Hodge, B. A., Meyerhof, G. T., Katewa, S. D., Lian, T., Lau, C., Bar, S., Leung, N. Y., Li, M., Li-Kroeger, D., Melov, S., Schilling, B., Montell, C. and Kapahi, P. (2022). Dietary restriction and the transcription factor clock delay eye aging to extend lifespan in Drosophila Melanogaster. Nat Commun 13(1): 3156. PubMed ID: 35672419
Hou, X., Hayashi, R., Itoh, M. and Tonoki, A. (2023). Small-molecule screening in aged Drosophila identifies mGluR as a regulator of age-related sleep impairment. Sleep. PubMed ID: 36721967
Hsieh, A. L., Zheng, X., Yue, Z., Stine, Z. E., Mancuso, A., Rhoades, S. D., Brooks, R., Weljie, A. M., Eisenman, R. N., Sehgal, A. and Dang, C. V. (2019). Misregulation of Drosophila Myc disrupts circadian behavior and metabolism. Cell Rep 29(7): 1778-1788.e1774. PubMed ID: 31722196
Hsu, C. T., Choi, J. T. Y. and Sehgal, A. (2020). Manipulations of the olfactory circuit highlight the role of sensory stimulation in regulating sleep amount. Sleep. PubMed ID: 33313876
Huang, H., Possidente, D. R. and Vecsey, C. G. (2021). Optogenetic activation of SIFamide (SIFa) neurons induces a complex sleep-promoting effect in the fruit fly Drosophila melanogaster. Physiol Behav 239: 113507. PubMed ID: 34175361
Huang, S., Piao, C., Beuschel, C. B., Zhao, Z. and Sigrist, S. J. (2022). A brain-wide form of presynaptic active zone plasticity orchestrates resilience to brain aging in Drosophila. PLoS Biol 20(12): e3001730. PubMed ID: 36469518
Hwangbo, D. S., Kwon, Y. J., Iwanaszko, M., Jiang, P., Abbasi, L., Wright, N., Alli, S., Hutchison, A. L., Dinner, A. R., Braun, R. I. and Allada, R. (2023). Dietary Restriction Impacts Peripheral Circadian Clock Output Important for Longevity in Drosophila. bioRxiv. PubMed ID: 36711760
Inami, S., Sato, T., Kurata, Y., Suzuki, Y., Kitamoto, T. and Sakai, T. (2021). Consolidation and maintenance of long-term memory involve dual functions of the developmental regulator Apterous in clock neurons and mushroom bodies in the Drosophila brain. PLoS Biol 19(12): e3001459. PubMed ID: 34860826
Iyer, A. R., Scholz-Carlson, E., Bell, E., Biondi, G., Richhariya, S., Fernandez, M. P. (2025). Circadian rhythms are more resilient to pacemaker neuron disruption in female Drosophila. PLoS Biol, 23(5):e3003146 PubMed ID: 40327674
Jagannath, A., Butler, R., Godinho, S. I., Couch, Y., Brown, L. A., Vasudevan, S. R., Flanagan, K. C., Anthony, D., Churchill, G. C., Wood, M. J., Steiner, G., Ebeling, M., Hossbach, M., Wettstein, J. G., Duffield, G. E., Gatti, S., Hankins, M. W., Foster, R. G. and Peirson, S. N. (2013). The CRTC1-SIK1 pathway regulates entrainment of the circadian clock. Cell 154: 1100-1111. PubMed ID: 23993098
Jarabo, P., Barredo, C. G., de Pablo, C., Casas-Tinto, S. and Martin, F. A. (2022). Alignment between glioblastoma internal clock and environmental cues ameliorates survival in Drosophila. Commun Biol 5(1): 644. PubMed ID: 35773327
Jauregui-Lozano, J., Hall, H., Stanhope, S. C., Bakhle, K., Marlin, M. M. and Weake, V. M. (2022). The Clock:Cycle complex is a major transcriptional regulator of Drosophila photoreceptors that protects the eye from retinal degeneration and oxidative stress. PLoS Genet 18(1): e1010021. PubMed ID: 35100266
Jeong, K., Lee, S., Seo, H., Oh, Y., Jang, D., Choe, J., Kim, D., Lee, J. H. and Jones, W. D. (2015). Ca-alpha1T, a fly T-type Ca(2+) channel, negatively modulates sleep. Sci Rep 5: 17893. PubMed ID: 26647714
Jin, X., Tian, Y., Zhang, Z. C., Gu, P., Liu, C. and Han, J. (2021). A subset of DN1p neurons integrates thermosensory inputs to promote wakefulness via CNMa signaling. Curr Biol. PubMed ID: 33740429
Jeong, E. M., Kwon, M., Cho, E., Lee, S. H., Kim, H., Kim, E. Y. and Kim, J. K. (2022). Systematic modeling-driven experiments identify distinct molecular clockworks underlying hierarchically organized pacemaker neurons. Proc Natl Acad Sci U S A 119(8). PubMed ID: 35193959
Johnstone, P. S., Ogueta, M., Akay, O., Top, I., Syed, S., Stanewsky, R. and Top, D. (2022). Real time, in vivo measurement of neuronal and peripheral clocks in Drosophila melanogaster. Elife 11. PubMed ID: 36190119
Jones, J. D., Holder, B. L., Eiken, K. R., Vogt, A., Velarde, A. I., Elder, A. J., McEllin, J. A. and Dissel, S. (2023). Regulation of sleep by cholinergic neurons located outside the central brain in Drosophila. PLoS Biol 21(3): e3002012. PubMed ID: 36862736
Joshi, R., Cai, Y. D., Xia, Y., Chiu, J. C. and Emery, P. (2022). PERIOD Phosphoclusters Control Temperature Compensation of the Drosophila Circadian Clock. Front Physiol 13: 888262. PubMed ID: 35721569
Joyce, M., Falconio, F. A., Blackhurst, L., Prieto-Godino, L., French, A. S., Gilestro, G. F. (2024). Divergent evolution of sleep in Drosophila species. Nat Commun, 15(1):5091 PubMed ID: 38876988
Sitarraman, D., Aso, Y., Jin, X., Chen, N., Felix, M., Rubin, G.M. and Nitabach, M.N. (2015). Propagation of homeostatic sleep signals by segregated synaptic microcircuits of the Drosophila mushroom body. Curr Biol [Epub ahead of print]. PubMed ID: 26455303
Smith, B. R. and Macdonald, S. J. (2020). Dissecting the Genetic Basis of Variation in Drosophila Sleep Using a Multiparental QTL Mapping Resource. Genes (Basel) 11(3). PubMed ID: 32168738
Smith, P., Buhl, E., Tsaneva-Atanasova, K. and Hodge, J. J. L. (2019). Shaw and Shal voltage-gated potassium channels mediate circadian changes in Drosophila clock neuron excitability. J Physiol. PubMed ID: 31612994
Solovev, I., Dobrovolskaya, E., Shaposhnikov, M., Sheptyakov, M. and Moskalev, A. (2019a). Neuron-specific overexpression of core clock genes improves stress-resistance and extends lifespan of Drosophila melanogaster. Exp Gerontol 117: 61-71. PubMed ID: 30415070
Solovev, I., Shegoleva, E., Fedintsev, A., Shaposhnikov, M. and Moskalev, A. (2019b). Circadian clock genes' overexpression in Drosophila alters diet impact on lifespan. Biogerontology 20(2): 159-170. PubMed ID: 30470951
Somers, J., Luong, H. N., Mitchell, J., Batterham, P. and Perry, T. (2017). Pleiotropic effects of loss of the Dα1 subunit in Drosophila melanogaster: Implications for insecticide resistance. Genetics 205(1): 263-271. PubMed ID: 28049707
Song, B. J., Sharp, S. J. and Rogulja, D. (2021). Daily rewiring of a neural circuit generates a predictive model of environmental light. Sci Adv 7(13). PubMed ID: 33762336
Sonn, J. Y., Lee, J., Sung, M. K., Ri, H., Choi, J. K., Lim, C. and Choe, J. (2018). Serine metabolism in the brain regulates starvation-induced sleep suppression in Drosophila melanogaster. Proc Natl Acad Sci U S A. PubMed ID: 29915051
Sorkac, A., Savva, Y. A., Savaş, D., Talay, M. and Barnea, G. (2022). Circuit analysis reveals a neural pathway for light avoidance in Drosophila larvae. Nat Commun 13(1): 5274. PubMed ID: 36071059
Souto-Maior, C., Serrano Negron, Y. L. and Harbison, S. T. (2023). Nonlinear expression patterns and multiple shifts in gene network interactions underlie robust phenotypic change in Drosophila melanogaster selected for night sleep duration. PLoS Comput Biol 19(8): e1011389. PubMed ID: 37561813
Srivastava, M., Varma, V., Abhilash, L., Sharma, V. K. and Sheeba, V. (2019). Circadian clock properties and their relationships as a function of free-running period in Drosophila melanogaster. J Biol Rhythms: 748730419837767. PubMed ID: 30939971
Stahl, B. A., Slocumb, M. E., Chaitin, H., Diangelo, J. R. and Keene, A. C. (2017). Sleep-dependent modulation of metabolic rate in Drosophila. Sleep [Epub ahead of print]. PubMed ID: 28541527
Stoleru, D., Peng, Y. Agosto, J. and Rosbash, M. (2004) Coupled oscillators control morning and evening locomotor behaviour of Drosophila. Nature 431: 862-868. PubMed citation: 15483615
Stoleru, D., Peng, Y., Nawathean, P. and Rosbash, M. (2005). A resetting signal between Drosophila pacemakers synchronizes morning and evening activity. Nature 438: 238-242. PubMed citation: 16281038
Stoleru, D., et al. (2007). The Drosophila circadian network is a seasonal timer. Cell 129(1): 207-19. PubMed citation: 17418796
Sun, Y., Liu, R., Bi, Y., Feng, X., Wang, J., Li, T., Wu, H., Zhang, C. and Sun, Y. (2022). Characterization of sleep-related neurochemicals in the different developmental stages and insomnia models of Drosophila melanogaster. Biomed Chromatogr: e5341. PubMed ID: 35045589
Sun, Y., Shui, K., Li, Q., Liu, C., Jin, W., Ni, J. Q., Lu, J., Zhang, L. (2025). Upstream open reading frames dynamically modulate CLOCK protein translation to regulate circadian rhythms and sleep. PLoS Biol, 23(5):e3003173 PubMed ID: 40354412
Suzuki, Y., Kurata, Y. and Sakai, T. (2022). Dorsal-lateral clock neurons modulate consolidation and maintenance of long-term memory in Drosophila. Genes Cells 27(4): 266-279. PubMed ID: 35094465
Szuperak, M., Churgin, M. A., Borja, A. J., Raizen, D. M., Fang-Yen, C. and Kayser, M. S. (2018). A sleep state in Drosophila larvae required for neural stem cell proliferation. Elife 7. PubMed ID: 29424688
Tabuchi, M., Monaco, J. D., Duan, G., Bell, B., Liu, S., Liu, Q., Zhang, K. and Wu, M. N. (2018). Clock-generated temporal codes determine synaptic plasticity to control sleep. Cell 175(5): 1213-1227. PubMed ID: 30318147
Tabuloc, C. A., Cai, Y. D., Kwok, R. S., Chan, E. C., Hidalgo, S. and Chiu, J. C. (2023). CLOCK and TIMELESS regulate rhythmic occupancy of the BRAHMA chromatin-remodeling protein at clock gene promoters. PLoS Genet 19(2): e1010649. PubMed ID: 36809369
Tainton-Heap, L. A. L., Kirszenblat, L. C., Notaras, E. T., Grabowska, M. J., Jeans, R., Feng, K., Shaw, P. J. and van Swinderen, B. (2020). A Paradoxical Kind of Sleep in Drosophila melanogaster. Curr Biol. PubMed ID: 33238155
Tanizawa, F. and Takemoto, H. (2021). Sleep contributes to preference for novel food odours in Drosophila melanogaster. Sci Rep 11(1): 9395. PubMed ID: 33931708
Tang, M., Cao, L. H., Yang, T., Ma, S. X., Jing, B. Y., Xiao, N., Xu, S., Leng, K. R., Yang, D., Li, M. T. and Luo, D. G. (2022). An extra-clock ultradian brain oscillator sustains circadian timekeeping. Sci Adv 8(35): eabo5506. PubMed ID: 36054358
Tasman, K., Hidalgo, S., Zhu, B., Rands, S. A. and Hodge, J. J. L. (2021). Neonicotinoids disrupt memory, circadian behaviour and sleep. Sci Rep 11(1): 2061. PubMed ID: 33479461
Teles-de-Freitas, R., Rivas, G. B. S., Peixoto, A. A. and Bruno, R. V. (2020). The Summer Is Coming: nocte and timeless Genes Are Influenced by Temperature Cycles and May Affect Aedes aegypti Locomotor Activity. Front Physiol 11: 614722. PubMed ID: 33424639
Thakkar, N., Giesecke, A., Bazalova, O., Martinek, J., Smykal, V., Stanewsky, R. and Dolezel, D. (2022). Evolution of casein kinase 1 and functional analysis of new doubletime mutants in Drosophila. Front Physiol 13: 1062632. PubMed ID: 36589447
Thompson, J. B., Su, O. O., Yang, N. and Bauer, J. H. (2020). Sleep-length differences are associated with altered longevity in the fruit fly Drosophila melanogaster. Biol Open 9(9). PubMed ID: 32938639
Thimgan, M. S., Kress, N., Lisse, J., Fiebelman, C. and Hilderbrand, T. (2018). The acyl-CoA synthetase, pudgy, promotes sleep and is required for the homeostatic response to sleep deprivation. Front Endocrinol (Lausanne) 9: 464. PubMed ID: 30186232
Toda, H., Williams, J. A., Gulledge, M. and Sehgal, A. (2019). A sleep-inducing gene, nemuri, links sleep and immune function in Drosophila. Science 363(6426): 509-515. PubMed ID: 30705188
Tomita, J., Ban, G., Kato, Y. S. and Kume, K. (2021). Protocerebral Bridge Neurons That Regulate Sleep in Drosophila melanogaster. Front Neurosci 15: 647117. PubMed ID: 34720844
Troup, M., Yap, M. H., Rohrscheib, C., Grabowska, M. J., Ertekin, D., Randeniya, R., Kottler, B., Larkin, A., Munro, K., Shaw, P. J. and van Swinderen, B. (2018). Acute control of the sleep switch in Drosophila reveals a role for gap junctions in regulating behavioral responsiveness. Elife 7. PubMed ID: 30109983
Uchizono, S., Tabuki, Y., Kawaguchi, N., Tanimura, T. and Itoh, T. Q. (2017). Mated Drosophila melanogaster females consume more amino acids during the dark phase. PLoS One 12(2): e0172886. PubMed ID: 28241073
Ueno, T., Tomita, J., Tanimoto, H., Endo, K., Ito, van Alphen, B., Semenza, E. R., Yap, M., van Swinderen, B. and Allada, R. (2021). A deep sleep stage in Drosophila with a functional role in waste clearance. Sci Adv 7(4). PubMed ID: 33523916
Ulgherait, M., Midoun, A. M., Park, S. J., Gatto, J. A., Tener, S. J., Siewert, J., Klickstein, N., Canman, J. C., Ja, W. W. and Shirasu-Hiza, M. (2021).
Circadian autophagy drives iTRF-mediated longevity. Nature 598(7880): 353-358. PubMed ID: 34588695
Vaccaro, A., Issa, A. R., Seugnet, L., Birman, S. and Klarsfeld, A. (2017). Drosophila Clock is required in brain pacemaker neurons to prevent premature locomotor aging independently of its circadian function. PLoS Genet 13(1): e1006507. PubMed ID: 28072817
Vanderheyden, W. M., Gerstner, J. R., Tanenhaus, A., Yin, J. C. and Shaw, P. J. (2013). ERK phosphorylation regulates sleep and plasticity in Drosophila. PLoS One 8: e81554. PubMed ID: 24244744
Vanderheyden, W. M., Van Dongen, H. P. A., Frank, M. G. and Gerstner, J. R. (2019). Sleep pressure regulates mushroom body neural-glial interactions in Drosophila. Matters Sel 2019. PubMed ID: 31938713
Vanin, S., et al. (2012). Unexpected features of Drosophila circadian behavioural rhythms under natural conditions. Nature 484(7394): 371-5. PubMed Citation: 22495312
Vaughen, J. P., Theisen, E., Rivas-Serna, I. M., Berger, A. B., Kalakuntla, P., Anreiter, I., Mazurak, V. C., Rodriguez, T. P., Mast, J. D., Hartl, T., Perlstein, E. O., Reimer, R. J., Clandinin, M. T. and Clandinin, T. R. (2022). Glial control of sphingolipid levels sculpts diurnal remodeling in a circadian circuit. Neuron. PubMed ID: 35961319
Vaze, K. M. and Helfrich-Forster, C. (2021). The Neuropeptide PDF Is Crucial for Delaying the Phase of Drosophila's Evening Neurons Under Long Zeitgeber Periods. J Biol Rhythms: 7487304211032336. PubMed ID: 34428956
Versteven, M., Ernst, K. M. and Stanewsky, R. (2020). A Robust and Self-Sustained Peripheral Circadian Oscillator Reveals Differences in Temperature Compensation Properties with Central Brain Clocks. iScience 23(8): 101388. PubMed ID: 32798967
Walsh, E. N., Shetty, M. S., Diba, K. and Abel, T. (2023). Chemogenetic Enhancement of cAMP Signaling Renders Hippocampal Synaptic Plasticity Resilient to the Impact of Acute Sleep Deprivation. eNeuro 10(1). PubMed ID: 36635248
Wang, C., Shui, K., Ma, S., Lin, S., Zhang, Y., Wen, B., Deng, W., Xu, H., Hu, H., Guo, A., Xue, Y. and Zhang, L. (2020). Integrated omics in Drosophila uncover a circadian kinome. Nat Commun 11(1): 2710. PubMed ID: 32483184
Wang, J., Fan, J. Y., Zhao, Z., Dissel, S. and Price, J. (2022). DBT affects sleep in both circadian and non-circadian neurons. PLoS Genet 18(2): e1010035. PubMed ID: 35139068
Wang, Q., Abruzzi, K. C., Rosbash, M. and Rio, D. C. (2018). Striking circadian neuron diversity and cycling of Drosophila alternative splicing. Elife 7. PubMed ID: 29863472
Wang, Y. Y., Ma, W. W. and Peng, I. F. (2020). Screening of sleep assisting drug candidates with a Drosophila model. PLoS One 15(7): e0236318. PubMed ID: 32726319
Wang, Y., Zhang, H., Zhang, Z., Sun, B., Tang, C., Zhang, L., Jiang, Z., Ding, B., Liao, Y. and Cai, P. (2021). Simulated mobile communication frequencies (3.5 GHz) emitted by a signal generator affects the sleep of Drosophila melanogaster. Environ Pollut 283: 117087. PubMed ID: 33894629
Wei, Y., Du, J. and Zhao, Z. (2021). Integrative Role of 14-3-3epsilon in Sleep Regulation.
Int J Mol Sci 22(18). PubMed ID: 34575915
Weiss, J. T. and Donlea, J. M. (2021). Sleep deprivation results in diverse patterns of synaptic scaling across the Drosophila mushroom bodies. Curr Biol. PubMed ID: 34107302
Wen, D., Xie, J., Yuan, Y., Shen, L., Yang, Y., Chen, W. (2024). The endogenous antioxidant ability of royal jelly in Drosophila is independent of Keap1/Nrf2 by activating oxidoreductase activity. Insect Sci. PubMed ID: 37632209
Westermark, P. O. (2016). Linking core promoter classes to circadian transcription. PLoS Genet 12: e1006231. PubMed ID: 27504829
Wijnen, H., Naef, F., Boothroyd, C., Claridge-Chang, A. and Young, M. W. (2006). Control of daily transcript oscillations in Drosophila by light and the circadian clock. PLoS Genet. 2(3): e39. 16565745
Williams, M. J., Perland, E., Eriksson, M. M., Carlsson, J., Erlandsson, D., Laan, L., Mahebali, T., Potter, E., Frediksson, R., Benedict, C. and Schioth, H. B. (2016). Recurrent sleep fragmentation induces insulin and neuroprotective mechanisms in middle-aged flies. Front Aging Neurosci 8: 180. PubMed ID: 27531979
Wilson, R. I. and Laurent, G. (2005). Role of GABAergic inhibition in shaping odor-evoked spatiotemporal patterns in the Drosophila antennal lobe. J Neurosci 25: 9069-9079. PubMed ID: 16207866
Wu, B., He, L., Xiao, Y., Du, J., Wang, X. and Zhao, Z. (2021). Juvenile hormone receptor MET regulates sleep and neuronal morphology via glial-neuronal crosstalk. J Genet Genomics. PubMed ID: 34376377
Wu, X., Wang, S., Zhao, X., Wen, J., Li, Y., Zhao, Z. and Du, J. (2022). Analysis of sleep in individual Drosophila melanogaster reveals a self-regulatory role for cuticular hydrocarbons pheromones. Insect Sci. PubMed ID: 35199930
Xia, X., Fu, X., Du, J., Wu, B., Zhao, X., Zhu, J. and Zhao, Z. (2020). Regulation of circadian rhythm and sleep by miR-375-timeless interaction in Drosophila. Faseb J. PubMed ID: 33078445
Xiao, Y., Yuan, Y., Jimenez, M., Soni, N. and Yadlapalli, S. (2021). Clock proteins regulate spatiotemporal organization of clock genes to control circadian rhythms. Proc Natl Acad Sci U S A 118(28). PubMed ID: 34234015
Xie, J., Wang, D., Ling, S., Yang, G., Yang, Y. and Chen, W. (2019). High-salt diet causes sleep fragmentation in young Drosophila through circadian rhythm and dopaminergic Systems. Front Neurosci 13: 1271. PubMed ID: 31849585
Xu, K., Zheng, X. and Sehgal A. (2008). Regulation of feeding and metabolism by neuronal and peripheral clocks in Drosophila. Cell Metab. 8(4):289-300. PubMed ID: 18840359
Xu, X., Yang, W., Tian, B., Sui, X., Chi, W., Rao, Y. and Tang, C. (2021). Quantitative investigation reveals distinct phases in Drosophila sleep. Commun Biol 4(1): 364. PubMed ID: 33742082
Yadlapalli, S., Jiang, C., Bahle, A., Reddy, P., Meyhofer, E. and Shafer, O. T. (2018). Circadian clock neurons constantly monitor environmental temperature to set sleep timing. Nature 555(7694): 98-102. PubMed ID: 29466329
Yamaguchi, S. T., Tomita, J. and Kume, K. (2022). Insulin signaling in clock neurons regulates sleep in Drosophila. Biochem Biophys Res Commun 591: 44-49. PubMed ID: 34998032
Yan, W., Lin, H., Yu, J., Wiggin, T. D., Wu, L., Meng, Z., Liu, C. and Griffith, L. C. (2023). Subtype-Specific Roles of Ellipsoid Body Ring Neurons in Sleep Regulation in Drosophila. J Neurosci 43(5): 764-786. PubMed ID: 36535771
Yao, Z., Bennett, A.J., Clem, J.L. and Shafer, O.T. (2016). The Drosophila clock neuron network features diverse coupling modes and requires network-wide coherence for robust circadian rhythms. Cell Rep 17: 2873-2881. PubMed ID: 27974202
Yap, M. H. W., Grabowska, M. J., Rohrscheib, C., Jeans, R., Troup, M., Paulk, A. C., van Alphen, B., Shaw, P. J. and van Swinderen, B. (2017). Oscillatory brain activity in spontaneous and induced sleep stages in flies. Nat Commun 8(1): 1815. PubMed ID: 29180766
Van De Poll, M. N., van Swinderen, B. (2025). Long-term multichannel recordings in Drosophila flies reveal altered predictive processing during sleep compared with wake. J Exp Biol, 228(11) PubMed ID: 40296777
Yin, J. C. P., Cui, E., Hardin, P. E. and Zhou, H. (2023). Circadian disruption of memory consolidation in Drosophila. Front Syst Neurosci 17: 1129152. PubMed ID: 37034015
Yin, L., Wang, J., Klein, P. S. and Lazar, M. A. (2006). Nuclear receptor Rev-erbalpha is a critical lithium-sensitive component of the circadian clock. Science 311: 1002-1005. PubMed citation: 16484495
You, S., Fulga, T. A., Van Vactor, D. and Jackson, F. R. (2018). Regulation of circadian behavior by astroglial microRNAs in Drosophila. Genetics 208(3): 1195-1207. PubMed ID: 29487148
Yuan, Q., Xiang, Y., Yan, Z., Han, C., Jan, L. Y. and Jan, Y. N. (2011). Light-induced structural and functional plasticity in Drosophila larval visual system. Science 333: 1458-1462. Pubmed: 21903815
Yuan, Y., Chen, Q., Brovkina, M., Clowney, E. J., Yadlapalli, S. (2023). Clock-dependent chromatin accessibility rhythms regulate circadian transcription. bioRxiv, PubMed ID: 37645872
Yurgel, M. E., Shah, K. D., Brown, E. B., Burns, C., Bennick, R. A., DiAngelo, J. R. and Keene, A. C. (2018). Ade2 functions in the Drosophila fat body to promote sleep. G3 (Bethesda). PubMed ID: 30249751
Zadeh-Haghighi, H. and Simon, C. (2022). Radical pairs can explain magnetic field and lithium effects on the circadian clock. Sci Rep 12(1): 269. PubMed ID: 34997158
Zakharenko, L. P., Petrovskii, D. V. and Putilov, A. A. (2018). Larks, owls, swifts, and woodcocks among fruit flies: differential responses of four heritable chronotypes to long and hot summer days. Nat Sci Sleep 10: 181-191. PubMed ID: 29950910
Zhang, C., Daubnerova, I., Jang, Y. H., Kondo, S., Zitnan, D. and Kim, Y. J. (2021). The neuropeptide allatostatin C from clock-associated DN1p neurons generates the circadian rhythm for oogenesis. Proc Natl Acad Sci U S A 118(4). PubMed ID: 33479181
Zhang, M. Y., Lear, B. C. and Allada, R. (2021). The microtubule associated protein tau suppresses the axonal distribution of PDF neuropeptide and mitochondria in circadian clock neurons. Hum Mol Genet. PubMed ID: 34750631
Zhang, R., Du, J., Zhao, X., Wei, L. and Zhao, Z. (2020). Regulation of circadian behavioural output via clock-responsive miR-276b. Insect Mol Biol. PubMed ID: 33131172
Zhang, W., Guo, C., Chen, D., Peng, Q. and Pan, Y. (2018). Hierarchical control of Drosophila sleep, courtship, and feeding behaviors by male-specific P1 neurons. Neurosci Bull. PubMed ID: 30182322
Zhang, Y., Zhou, Y., Zhang, X., Wang, L. and Zhong, Y. (2021). Clock neurons gate memory extinction in Drosophila. Curr Biol. PubMed ID: 33545046
Zhao, X., Li, Y., Zhao, Z. and Du, J. (2020). Extra sex combs buffers sleep-related stresses through regulating Heat shock proteins. Faseb j. PubMed ID: 33220007
Zhao, Z., Zhao, X., He, T., Wu, X., Lv, P., Zhu, A. J. and Du, J. (2021). Epigenetic regulator Stuxnet modulates octopamine effect on sleep through a Stuxnet-Polycomb-Octbeta2R cascade. EMBO Rep: e47910. PubMed ID: 33410264
Zheng, Y., Xue, Y., Ren, X., Liu, M., Li, X., Jia, Y., Niu, Y., Ni, J. Q., Zhang, Y. and Ji, J. Y. (2018). The lysine demethylase dKDM2 is non-essential for viability, but regulates circadian rhythms in Drosophila. Front Genet 9: 354. PubMed ID: 30233643
Zhu, B. and Liu, S. (2023). Preservation of ~12-hour ultradian rhythms of gene expression of mRNA and protein metabolism in the absence of canonical circadian clock. bioRxiv PubMed ID: 37205336
Zimmerman, J.E., Chan, M.T., Lenz, O.T., Keenan, B.T., Maislin, G. and Pack, A.I. (2016). Glutamate is a wake active neurotransmitter in Drosophila melanogaster. Sleep [Epub ahead of print]. PubMed ID: 27855736
Zurl, M., Poehn, B., Rieger, D., Krishnan, S., Rokvic, D., Veedin Rajan, V. B., Gerrard, E., Schlichting, M., Orel, L., Coric, A., Lucas, R. J., Wolf, E., Helfrich-Forster, C., Raible, F. and Tessmar-Raible, K. (2022). Two light sensors decode moonlight versus sunlight to adjust a plastic circadian/circalunidian clock to moon phase. Proc Natl Acad Sci U S A 119(22): e2115725119. PubMed ID: 35622889
Aged flies exhibit less total baseline sleep and an altered homeostatic
response in that they recover sleep over an extended 12 h period,
whereas young flies recoup lost sleep within a much shorter window.
Similar sleep disturbances are seen in humans and mammals, where aged
individuals lose the ability to initiate and maintain nighttime sleep
and recovery sleep after prolonged wakefulness is altered. Similar to
this study, a significant age by time interaction was demonstrated,
where young flies recover sleep in the first 45 h following sleep
deprivation. Sleep deprived aged flies however, recover sleep over much
a longer period. Variations in behavioral results compared with those
existing in the literature are possibly due to use of video
analysis, which has been shown to be more accurate than DAMS in
measurements of fly sleep, in particular daytime sleep and architecture
(Brown, 2014).
During the course of this study, an increase in locomotor activity in
both young and aged flies treated with PBA was observed. A previous
study found that PBA maintains locomotor vigor (activity) during aging
in Drosophila. These results suggest that PBA is in fact
increasing sleep and not merely reducing movement in the aged flies.
Although total wake did not change in the young flies treated with PBA,
wake became more consolidated. Not observing changes in total wake is
possibly due to a ceiling effect in the young healthy flies. These
results indicate that there is an age by drug interaction in the
modification of sleep by PBA that remains to be elucidated (Brown,
2014).
{kind=link}
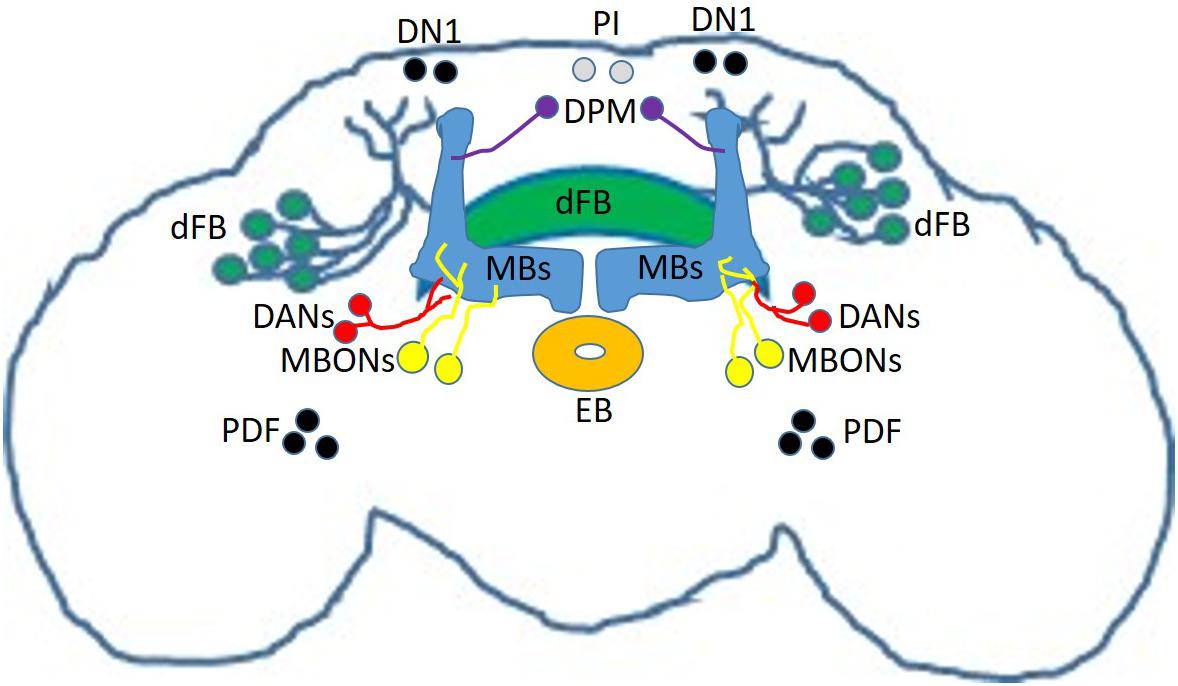{kind=link}