Select image to enlarge
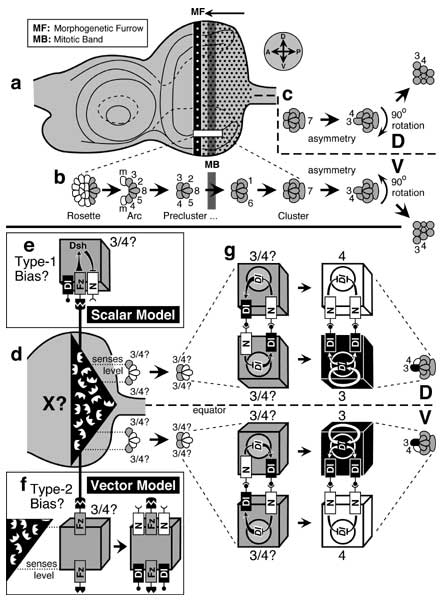
Figure 7.5
'Symmetry breaking' in nascent ommatidia. Precursor cells are denoted by a 'p' suffix and equivalent pairs by a slash mark (e.g., 'R3/4p'). Abbreviations: Dl (Delta), Fz (Frizzled), N (Notch). a-c. Histological appearance of photoreceptor clusters during development. d-g. Control of D/V chirality by Fz and N signaling. Cubes are individual cells, and inscribed circles are nuclei. Naïve R3/4p ('3/4?') cells are shaded (left). Committed R3p and R4p cells (far right) are black or white respectively. See also App. 7.
a. Mature left eye disc. Dots are ommatidial sites. Vertical stripes are the morphogenetic furrow (MF) and mitotic band (MB; see [185, 4291, 4293, 4355] for details). Ahead of the MF are scattered mitoses (not shown). Compass (at right) gives axes in the fate map (cf. Fig. 7.1). 'Rows' and 'columns' run along A-P and D-V axes respectively.
b. Stages in development of a cluster. Because the MF moves as a wave from P to A, this series is manifest in each row (magnified from box in a). Shading indicates future photoreceptors. Numbers denote the prospective fate of precursors ('3' = R3, etc.), not their current state of determination (which may be indefinite). Mystery (m) cells leave the arc as it collapses into a precluster [186, 4364, 4715]. Rotation is more gradual than implied here: it starts after the precluster stage (~ column 3-6) and ends by the 4-cone stage (~ column 15) [4361, 4364] (T. Wolff, pers. comm.).
c. Clusters become asymmetric at the 2-cone cell stage [4355, 4873] (≈ column 11 [4364]) when R4p severs its connection to R8p and undergoes a slight shift (D or V) jointly with R3p. At about this time, the clusters rotate clockwise or counterclockwise [777, 1639, 2794, 4873] on either side of the equator (thick dashed line). Trapezoid shapes arise later. (Only rhabdomeres really adopt this shape -- not whole cells as shown here.)
d. Posterior (eye) part of the disc. The polarizing signal (X = E-shaped ligands) is assumed to be under N control (cf. Fig. 7.4) and to form 2 gradients by diffusing from the equator. Fz is the receptor for X. At the Arc Stage the naïve R3/4p cell nearer the equator should sense a higher level of X (dotted line) and hence activate more of its Fz receptors [4366, 4873].
e. According to the Scalar Model [869], this higher Fz activity represses N (via Dsh) more strongly than in the rival cell. When the cells touch (Precluster Stage, d), this bias causes the cell nearer the equator to have less active N, so it wins the 'shouting' contest (i.e., whoever has the most Dl) to become R3.
f. The Vector Model [4366] invokes a different kind of bias (Type 2 vs. 1). The side of the cell that faces the equator should have more of its Fz receptors occupied than the other side. If X-bound Fz receptors recruit N while vacant Fz receptors recruit Dl, then each R3/4p cell will have more N on its equatorial side and more Dl on its polar side. When the two such R3/4p cells meet (d), the one nearer the equator will have more Dl at the interface and so will always win and go on to become R3. (Other variations on this scheme are possible.)
g. Enlargement of R3/4p pairs after these na�ve (gray) cells abut (Precluster Stage). The outcome of the contest (black '3' or white '4') is preordained by the bias introduced earlier (e or f), though no bias is depicted here (g). The positive feedback loop between the cells constitutes a 'Delta-Notch Flip-flop' device, whose instability eventually forces the cells to adopt binary (alternative) R3 vs. R4 fates (Cluster Stage at right) by amplifying the small initial bias [2883].
Relative locations of MF and MB in a are based on [604, 3539]. Panels b and c are adapted from [293, 604, 3539, 3563, 4355, 4715, 4873], though not all of the depicted stages (b) may occupy successive columns [605, 2887], and the mitotic band may actually span several columns [185, 228]. Models and schematics in d-g are modified from [359, 447, 869, 4173, 4366]. For details of rosette geometries see [184, 4712]. For critiques of earlier models see [3562, 3563]. See also App. 7.
|
|
{kind=link}
{kind=link}