Select image to enlarge
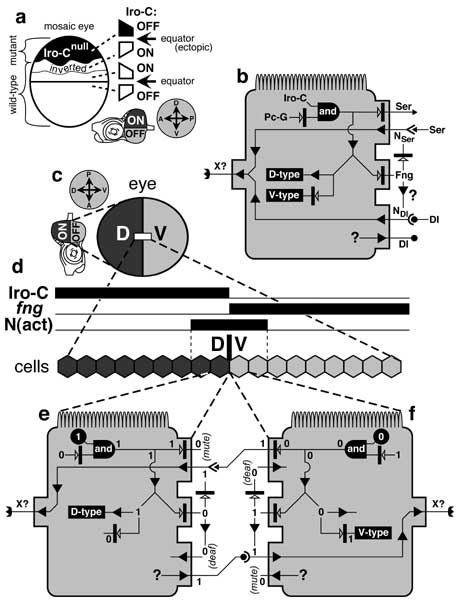
Figure 7.4
Control of ommatidial polarity by the Iroquois Complex (Iro-C).
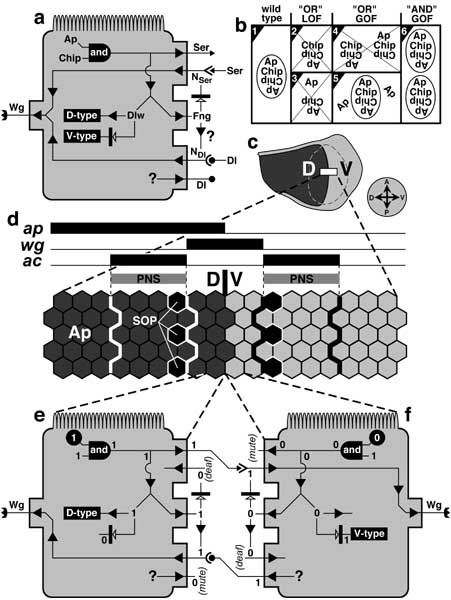
a. Iro-C genes are expressed in the D -- but not V -- half of the eye during normal development (disc schematic below). The equator (horizontal line) arises at this ON/OFF boundary, and all photoreceptor trapezoids (cf. Fig. 7.2) point away from it. When an Iro-Cnull clone (black) is induced in the D area, it causes wild-type trapezoids within ~7 rows (the eye is ~33 rows high) to invert their polarity. Hence, an ectopic equator emerges at the clone edge.
b. Generic eye cell (cf. Fig. 2.7 for icons) showing the main control circuit for emitting an unknown diffusible factor (X) that regulates polarity. Abbreviations: Dl (Delta), Fng (Fringe), N (Notch; 'act' = activated form), Pc-G (Polycomb-Group genes), Ser (Serrate). NSer and NDl are hypothetical Fng-modulated forms of Notch that are receptive to Ser or Dl [990, 1250, 1988]. The circuit works as follows. If Pc-G products are sufficiently low, then Iro-C genes can be expressed (e) [3079]. In that case, the cell adopts D-type identity and expresses Dl (by an unknown route) but not Ser or Fng. D-type cells can receive Ser signals (from across the D/V line). If Iro-C genes are OFF (f), then the cell adopts V-type identity and expresses both Ser and Fng. In that case, Fng blocks the Ser-N interaction, so V-type cells can only receive Dl signals. The circuit allows two types of dialogue: Ser-signal 'speakers' with NSer 'listeners' or Dl-signal 'speakers' with NDl 'listeners'. Both conversations should turn ON Factor X. Factor X uses Frizzled as a receptor (cf. Fig. 7.5) [869, 1194, 4366].
c. Eye portion of a mature left eye disc. Shading denotes D (dark; Iro-C = ON) vs. V (light; Iro-C = OFF) compartments. Note that this eye was rotated 90° counterclockwise relative to the eye in a (cf. compasses and disc icons) to facilitate comparison with the wing disc's circuit diagram (Fig. 6.8).
d. Enlarged part of the D/V border (box in c). In this single file, cells on the D side express Iro-C genes, while cells on the V side express fng. Interactions between cells in e and f activate Notch at the D/V boundary. The width of the N(act) stripe (here shown spanning 4 cells) is not known exactly.
e, f. Schematics of cells flanking the D/V line. The V cell is drawn as if flipped around to face the D cell. States of variables (cf. b) are recorded as '1' (present and active) or '0' (absent or inactive). Black circles indicate determining factors. 'Mute' and 'deaf' denote inability to send or receive particular signals (Ser above, Dl below). The functioning of the circuit is explained under b above.
Panel a is schematized adaptation of a photo in [696]. Circuitry in b is compiled from [696] (Iro-C), [3079] (Pc-G), and [774, 869, 1080, 1194, 3258] (Fng-N-Ser-Dl). Panel d incorporates data from the same sources.
N.B.: The D/V circuitry of the eye differs from that of the wing insofar as fng is expressed ventrally instead of dorsally (cf. Fig. 6.8). This difference is attributable to the 180° rotation that the eye disc undergoes after its inception [774, 4146] (cf. Fig. 7.1a legend). The 'Iro-C —| Ser' link is simplified from the 'Iro-C —| Fng —> Ser' route [696, 774, 3258], and the full path is probably even more complex. This model does not explain how equators arise from ectopic furrows that are triggered without altering Iro-C expression [722, 723, 4167, 4648]. Other key players in this scenario may be (1) Lobe (LobeLOF causes mirr to turn ON in the V half and hence suppresses ommatidia [758, 2560]) and (2) teashirt (tshLOF suppresses ommatidia in the D half) [3973].
|
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}