|
Gene name - rutabaga Synonyms - Cytological map position - 12F5-13A1 Function - adenylate cyclase Keywords - neural, cAMP signaling, learning pathway, calcium dependent enzymes |
Symbol - rut FlyBase ID:FBgn0003301 Genetic map position - 1-[46] Classification - Type 1 - Ca++ CaM sensitive Cellular location - cytoplasmic |
| Recent literature | Dissel, S., Angadi, V., Kirszenblat, L., Suzuki, Y., Donlea, J., Klose, M., Koch, Z., English, D., Winsky-Sommerer, R., van Swinderen, B. and Shaw, P.J. (2015). Sleep restores behavioral plasticity to Drosophila mutants. Curr Biol [Epub ahead of print]. PubMed ID: 25913403
Summary: Given the role that sleep plays in modulating plasticity, this study hypothesized that increasing sleep would restore memory to canonical memory mutants without specifically rescuing the causal molecular lesion. Sleep was increased using three independent strategies: activating the dorsal fan-shaped body, increasing the expression of Fatty acid binding protein (dFabp), or by administering the GABA-A agonist 4,5,6,7-tetrahydroisoxazolo-[5,4-c]pyridine-3-ol (THIP). Short-term memory (STM) or long-term memory (LTM) was evaluated in rutabaga (rut) and dunce (dnc) mutants using aversive phototaxic suppression and courtship conditioning. Each of the three independent strategies increased sleep and restored memory to rut and dnc mutants. Importantly, inducing sleep also reversed memory defects in a Drosophila model of Alzheimer's disease. Together, these data demonstrate that sleep plays a more fundamental role in modulating behavioral plasticity than previously appreciated and suggest that increasing sleep may benefit patients with certain neurological disorders. |
Naganos, S., Ueno, K., Horiuchi, J. and Saitoe, M. (2016). Learning defects in Drosophila growth restricted chico mutants are caused by attenuated adenylyl cyclase activity. Mol Brain 9: 37. PubMed ID: 27048332
Summary: Reduced insulin/insulin-like growth factor signaling (IIS) is a major cause of symmetrical intrauterine growth retardation (IUGR), an impairment in cell proliferation during prenatal development that results in global growth defects and mental retardation. In Drosophila, chico encodes the only insulin receptor substrate. The physiological and molecular bases of learning defects caused by chico mutation are not clear. This study found that chico mutations impair memory-associated synaptic plasticity in the mushroom bodies (MBs), neural centers for olfactory learning. Mutations in chico reduce expression of the rutabaga-type adenylyl cyclase (rut), leading to decreased cAMP synthesis in the MBs. Expressing a rut + transgene in the MBs restores memory-associated plasticity and olfactory associative learning in chico mutants, without affecting growth. Thus chico mutations disrupt olfactory learning, at least in part, by reducing cAMP signaling in the MBs. These results suggest that some cognitive defects associated with reduced IIS may occur, independently of developmental defects, from acute reductions in cAMP signaling. |
Vonhoff, F. and Keshishian, H. (2016). Cyclic nucleotide signaling is required during synaptic refinement at the Drosophila neuromuscular junction. Dev Neurobiol [Epub ahead of print]. PubMed ID: 27281494
Summary: The removal of miswired synapses is a fundamental prerequisite for normal circuit development, leading to clinical problems when aberrant. However, the underlying activity-dependent molecular mechanisms involved in synaptic pruning remain incompletely resolved. This study examined the dynamic properties of intracellular calcium oscillations and tested a role for cAMP signaling during synaptic refinement in intact Drosophila embryos using optogenetic tools. In vivo evidence at the single gene level is provided that the calcium-dependent adenylyl cyclase rutabaga, the phosphodiesterase dunce, the kinase PKA, and Protein Phosphatase 1 (PP1) all operate within a functional signaling pathway to modulate Sema2a-dependent chemorepulsion. Presynaptic cAMP levels are required to be dynamically maintained at an optimal level to suppress connectivity defects. It is also proposed that PP1 may serve as a molecular link between cAMP signaling and CaMKII in the pathway underlying refinement. These results introduce an in vivo model where presynaptic cAMP levels, downstream of electrical activity and calcium influx, act via PKA and PP1 to modulate the neuron's response to chemorepulsion involved in the withdrawal of off-target synaptic contacts. |
Ueno, K., Suzuki, E., Naganos, S., Ofusa, K.,
Horiuchi, J. and Saitoe, M. (2017). Coincident
postsynaptic activity gates presynaptic dopamine release to induce
plasticity in Drosophila mushroom bodies. Elife 6.
PubMed ID: 28117664 Summary: Simultaneous stimulation of the antennal lobes (ALs) and the ascending fibers of the ventral nerve cord (AFV), two sensory inputs to the mushroom bodies (MBs), induces long-term enhancement (LTE) of subsequent AL-evoked MB responses. LTE induction requires activation of at least three signaling pathways to the MBs, mediated by nicotinic acetylcholine receptors (nAChRs), NMDA receptors (NRs), and D1 dopamine receptors (D1Rs). This study demonstrates that inputs from the AL are transmitted to the MBs through nAChRs, and inputs from the AFV are transmitted by NRs. Dopamine signaling occurs downstream of both nAChR and NR activation, and requires simultaneous stimulation of both pathways. Dopamine release requires the activity of the rutabaga adenylyl cyclase in postsynaptic MB neurons, and release is restricted to MB neurons that receive coincident stimulation. These results indicate that postsynaptic activity can gate presynaptic dopamine release to regulate plasticity. |
Chouhan, N. S., Mohan, K. and Ghose, A. (2017). cAMP signaling mediates behavioral flexibility and consolidation of social status in Drosophila aggression. J Exp Biol 220(Pt 23): 4502-4514. PubMed ID: 28993465
Summary: Social rituals, such as male-male aggression in Drosophila, are often stereotyped and the component behavioral patterns modular. The likelihood of transition from one behavioral pattern to another is malleable by experience and confers flexibility to the behavioral repertoire. Experience-dependent modification of innate aggressive behavior in flies alters fighting strategies during fights and establishes dominant-subordinate relationships. Dominance hierarchies resulting from agonistic encounters are consolidated to longer-lasting, social-status-dependent behavioral modifications, resulting in a robust loser effect. This study shows that cAMP dynamics regulated by the calcium-calmodulin-dependent adenylyl cyclase, Rut, and the cAMP phosphodiesterase, Dnc, but not the Amn gene product, in specific neuronal groups of the mushroom body and central complex, mediate behavioral plasticity necessary to establish dominant-subordinate relationships. rut and dnc mutant flies were unable to alter fighting strategies and establish dominance relationships during agonistic interactions. This real-time flexibility during a fight was independent of changes in aggression levels. Longer-term consolidation of social status in the form of a loser effect, however, required additional Amn-dependent inputs to cAMP signaling and involved a circuit-level association between the alpha/beta and gamma neurons of the mushroom body. These findings implicate cAMP signaling in mediating the plasticity of behavioral patterns in aggressive behavior and in the generation of a temporally stable memory trace that manifests as a loser effect. |
Baggett, V., Mishra, A., Kehrer, A. L., Robinson, A. O., Shaw, P. and Zars, T. (2018). Place learning overrides innate behaviors in Drosophila. Learn Mem 25(3): 122-128. PubMed ID: 29449456
Summary: Animals in a natural environment confront many sensory cues. Some of these cues bias behavioral decisions independent of experience, and action selection can reveal a stimulus-response (S-R) connection. How animals use learning to modify S-R relationships is a largely open question. Three sensory stimuli, air, light, and gravity sources were presented to individual Drosophila melanogaster in both naive and place conditioning situations. Flies were tested for a potential modification of the S-R relationships of anemotaxis, phototaxis, and negative gravitaxis by a contingency that associated place with high temperature. With two stimuli, significant S-R relationships were abandoned when the cue was in conflict with the place learning contingency. The role of the dunce (dnc) cAMP-phosphodiesterase and the rutabaga (rut) adenylyl cyclase were examined in all conditions. Both dnc1 and rut2080 mutant flies failed to display significant S-R relationships with two attractive cues, and have characteristically lower conditioning scores under most conditions. Thus, learning can have profound effects on separate native S-R relationships in multiple contexts, and mutation of the dnc and rut genes reveal complex effects on behavior. |
Sato, S., Ueno, K., Saitoe, M. and Sakai, T. (2018). Synaptic depression induced by postsynaptic cAMP production in the Drosophila mushroom body calyx. J Physiol. PubMed ID: 29659025
Summary: Synaptic plasticity has been studied to reveal the molecular and cellular mechanisms of associative and nonassociative learning. The fruit fly Drosophila melanogaster can be used to identify the molecular mechanisms of synaptic plasticity because vast genetic information or tools are available. By ex vivo Ca(2+) imaging of an isolated cultured Drosophila brain, this study examined the novel activity-dependent synaptic depression between the projection neurons of the antennal lobe (AL) and mushroom body (MB). Ex vivo Ca(2+) imaging analysis revealed that electrical stimulation of AL elicits Ca(2+) responses in the dendritic (calyx) and axonal (alpha lobe) regions of MB neurons, and the responses are reduced after repetitive AL stimulation. Since the cAMP signalling pathway plays an important role in synaptic plasticity in invertebrates and vertebrates, this study examined whether the reduction of Ca(2+) responses is also regulated by the cAMP signalling pathway. The expression of rutabaga (rut), which encodes Ca(2+) /calmodulin-dependent adenylyl cyclase, was essential for the reduction of Ca(2+) responses in the calyx and alpha lobe. Furthermore, imaging analysis using a fluorescence resonance energy transfer-based cAMP indicator revealed that the cAMP level increased in the wild-type calyx during repetitive AL stimulation, whereas it decreased in rut1 mutant flies with a loss-of-function mutation of rut. Thus, this study suggests that an increase in postsynaptic cAMP level during repetitive AL stimulation contributes to the attenuation of inputs at the AL-MB synapses. |
Sabandal, J. M., Sabandal, P. R., Kim, Y. C. and Han, K. A. (2020). Concerted Actions of Octopamine and Dopamine Receptors Drive Olfactory Learning. J Neurosci. PubMed ID: 32277043
Summary: Aminergic signaling modulates associative learning and memory. Substantial advance has been made in Drosophila on the dopamine receptors and circuits mediating olfactory learning, however knowledge on other aminergic modulation lags behind. To address this knowledge gap, this study investigated the role of octopamine in olfactory conditioning. Octopamine activity through the beta adrenergic-like receptor Octbeta1R is shown to drive aversive and appetitive learning: Octbeta1R in the mushroom body alphabeta neurons processes aversive learning whereas Octbeta1R in the projection neurons mediates appetitive learning. Genetic interaction and imaging studies pinpoint cAMP signaling as a key downstream effector for Octbeta1R function. The rutabaga-adenylyl cyclase synthesizes cAMP in a Ca(2+)/calmodulin-dependent manner, serving as a coincidence detector for associative learning and likely representing a downstream target for Octbeta1R. Supporting this notion, the double heterozygous rutabaga/+;;octbeta1r/+ flies perform poorly in both aversive and appetitive conditioning, while individual heterozygous rutabaga/+ and octbeta1r/+ behave like the wild-type control. Consistently, the mushroom body and projection neurons in the octbeta1r brain exhibit blunted responses to octopamine when cAMP levels are monitored through the cAMP sensor. Previous work demonstrated the pivotal functions of the D1 receptor dDA1 in aversive and appetitive learning, and the alpha1 adrenergic-like receptor OAMB in appetitive learning. As expected, octbeta1r genetically interacts with dumb (dDA1 mutant) in aversive and appetitive learning, but it interacts with oamb only in appetitive learning. This study uncovers the indispensable contributions of dopamine and octopamine signaling to aversive and appetitive learning. All experiments were performed on mixed sex unless otherwise noted. |
Melnattur, K., Kirszenblat, L., Morgan, E., Militchin, V., Sakran, B., English, D., Patel, R., Chan, D., van Swinderen, B. and Shaw, P. J. (2020). A conserved role for sleep in supporting Spatial Learning in Drosophila. Sleep. PubMed ID: 32959053
Summary: Sleep loss and aging impair hippocampus-dependent Spatial Learning in mammalian systems. This study used the fly Drosophila melanogaster to investigate the relationship between sleep and Spatial Learning in healthy and impaired flies. The Spatial Learning assay is modeled after the Morris Water Maze. The assay uses a 'thermal maze' consisting of a 5X5 grid of Peltier plates maintained at 36-37°C and a visual panorama. The first trial begins when a single tile that is associated with a specific visual cue is cooled to 25°C. For subsequent trials, the cold tile is heated, the visual panorama is rotated and the flies must find the new cold-tile by remembering its association with the visual cue. Significant learning was observed with two different wild-type strains - Cs and 2U, validating the design. Sleep deprivation prior to training impaired Spatial Learning. Learning was also impaired in the classic learning mutant rutabaga (rut); enhancing sleep restored learning to rut mutants. Further, flies exhibited dramatic age-dependent cognitive decline in Spatial Learning starting at 20-24 days of age. These impairments could be reversed by enhancing sleep. Finally, Spatial Learning was found to requires dopaminergic signaling and that enhancing dopaminergic signaling in aged flies restored learning. These results are consistent with the impairments seen in rodents and humans. These results thus demonstrate a critical conserved role for sleep in supporting Spatial Learning, and suggest potential avenues for therapeutic intervention during aging. |
Guo, L., Sun, Y. and Liu, S. (2023). Adaptive behaviors of Drosophila larvae on slippery surfaces. J Biol Phys 49(1): 121-132. PubMed ID: 36790728
Summary: Friction is ubiquitous but an essential force for insects during locomotion. Insects use dedicated bio-mechanical systems such as adhesive pads to modulate the intensity of friction, providing a stable grip with touching substrates for locomotion. However, how to uncover behavioral adaptation and regulatory neural circuits of friction modification is still largely understood. A novel behavior paradigm was devised in this study to investigate adaptive behavioral alternation of Drosophila larvae under low-friction surfaces. A tail looseness phenotype similar to slipping behavior in humans, was found to be a primary indicator to assess the degree of slipping. A gradual reduction on slipping level was found in wild-type larvae after successive larval crawling, coupled with incremental tail contraction, displacement, and speed acceleration. Meanwhile, a strong correlation was found between tail looseness index and length of contraction, suggesting that lengthening tail contraction may contribute to enlarging the contact area with the tube. Moreover, a delayed adaptation was found in rut mutant larvae, inferring that neural plasticity may participate in slipping adaptation. In conclusion, this paradigm can be easily and reliably replicated, providing a feasible pathway to uncover the behavioral principle and neural mechanism of acclimation of Drosophila larvae to low-friction conditions. |
Abubaker, M. B., Hsu, F. Y., Feng, K. L., Chu, L. A., de Belle, J. S., Chiang, A. S. (2024). Asymmetric neurons are necessary for olfactory learning in the Drosophila brain. Curr Biol, 34(5):946-957.e944 PubMed ID: 38320552
Summary: Animals have complementary parallel memory systems that process signals from various sensory modalities. In the brain of the fruit fly Drosophila melanogaster, mushroom body (MB) circuitry is the primary associative neuropil, critical for all stages of olfactory memory. These findings suggest that active signaling from specific asymmetric body (AB) neurons is also crucial for this process. These AB neurons respond to odors and electric shock separately and exhibit timing-sensitive neuronal activity in response to paired stimulation while leaving a decreased memory trace during retrieval. These experiments also show that rutabaga-encoded adenylate cyclase, which mediates coincidence detection, is required for learning and short-term memory in both AB and MB. Additive effects were observed when manipulating rutabaga co-expression in both structures. Together, these results implicate the AB in playing a critical role in associative olfactory learning and short-term memory. |
Coban, B., Poppinga, H., Rachad, E. Y., Geurten, B., Vasmer, D., Rodriguez Jimenez, F. J., Gadgil, Y., Deimel, S. H., Alyagor, I., Schuldiner, O., Grunwald Kadow, I. C., Riemensperger, T. D., Widmann, A., Fiala, A. (2024). The caloric value of food intake structurally adjusts a neuronal mushroom body circuit mediating olfactory learning in Drosophila. JLearn Mem, 31(5) PubMed ID: 38862177
Summary: Associative learning enables the adaptive adjustment of behavioral decisions based on acquired, predicted outcomes. The valence of what is learned is influenced not only by the learned stimuli and their temporal relations, but also by prior experiences and internal states. This study used the fruit fly Drosophila melanogaster to demonstrate that neuronal circuits involved in associative olfactory learning undergo restructuring during extended periods of low-caloric food intake. Specifically, a decrease was observed in the connections between specific dopaminergic neurons (DANs) and Kenyon cells at distinct compartments of the mushroom body. This structural synaptic plasticity was contingent upon the presence of allatostatin A receptors in specific DANs and could be mimicked optogenetically by expressing a light-activated adenylate cyclase in exactly these DANs. Importantly, this rearrangement in synaptic connections was found to influence aversive, punishment-induced olfactory learning but did not impact appetitive, reward-based learning. Whether induced by prolonged low-caloric conditions or optogenetic manipulation of cAMP levels, this synaptic rearrangement resulted in a reduction of aversive associative learning. Consequently, the balance between positive and negative reinforcing signals shifted, diminishing the ability to learn to avoid odor cues signaling negative outcomes. These results exemplify how a neuronal circuit required for learning and memory undergoes structural plasticity dependent on prior experiences of the nutritional value of food. |
Martinez-Cordera, M., Sakai, T., Saitoe, M., Ueno, K. (2025). Comparative experience shapes sucrose preference through memory in Drosophila. Mol Brain, 18(1):32 PubMed ID: 40211246
Summary: \Selection of appropriate food is an ability that allows animals to make optimal foraging choices. However, the neural mechanisms that control this food selection remain unclear. The purpose of this study was to investigate the connection between memory and the feeding behavior of Drosophila melanogaster when two sucrose solutions with different concentrations are available. Flies were placed into plates with 150 mM and 100 mM sucrose solutions and measured the preference for the 150 mM one. Flies preferred the 150 mM solution over the 100 mM when all 60 wells of the plate were filled with both solutions; this preference decreased when there were only 8 wells with food. Remarkably, prior exposure to a plate with all 60 wells filled with both solutions enhanced the preference for the 150 mM, even when there were only 8 wells with food. The memory-related gene rut and the dopamine D1 receptor on the mushroom body were required to enhance the preference after the prior exposure. These findings show that memory acquired through experiencing both solutions is stored in the mushroom body optimizing the food selection process (Martinez-Cordera. 2025. |
rutabaga encodes a calmodulin dependent adenylate cyclase that converts ATP to cyclic AMP. cAMP is a major signal transducer of the cell, and its creation and destruction is involved in just about every response of the cell to environmental changes. Calmodulin is a protein that binds the Ca++ ion, sensing its cellular concentration and interacting with the rutabaga encoded adenyl cyclase to activate adenyl cyclase mediated enzymatic conversion of ATP into cAMP.
The cyclic AMP system plays a critical role in olfactory learning in Drosophila, as does rutabaga, given this gene's crucial involvement in the same system. Calcium ions enter vertebrate cells through a type of glutamate receptor known as the NMDA (N-methyl-D-aspartate) receptor. Neural stimulation results in movement of calcium ions into the cell, leading to activation of adenyl cyclase and a rise in cAMP levels. cAMP, one of the central chemical messengers of the cell, then activates protein kinase A, which in turn initiates a phosphorylation cascade leading to the induction of genes involved in learning (Davis, 1995, Davis, 1996, Fagnon, 1995 and Nighorn, 1995,).
Rutabaga and Dunce are both found expressed at high levels in the mushroom bodies of adult flies. Mushroom bodies are paired groups of about 2500 neurons each, clustered in either hemisphere of the brain. They receive olfactory information from the antennal lobes via dentrites located in the calyx, a region of the brain just ventral to the mushroom bodies. The calyx is a neuropil, a region rich in synapses. It is in the calyx that integrates information from the thoracic ganglion (sensory information) and from the antennal nerve (olfactory information), passing the news on to the mushroom bodies. In classical operant conditioning experiments an electric shock (sensory information) is paired with a particular stimulus (olfactory information) to condition learning (avoidance behavior) upon presentation of the successfully conditioned stimulus, (in these cases, an odor) (Quinn, 1974, Han, 1992 and Nighorn, 1994).
A group of diffuse interneurons in mushroom bodies carry the information to the lateral protocerebrum. Interneurons from the protocerebrum make synaptic contact onto command neurons in a brain structure called the posterior slope, and these response messages are sent out of the brain to direct motor activity. In the case of the odor conditioned to a noxious stimulus like a shock, the message is the equivalent of "get out of here."
Biological systems are built in order to respond to environmental stimulus, whether in nature or in the laboratory. Learning takes this ability to extremes. The cellular basis of operant conditioning is the same, whether one considers Pavlov's dogs, salivating to the sound of a bell, or the conditioned fruit fly, avoiding a particular odor. Coincidence of information from two modalities (smell plus shock sensation) results in heightened neural activity, that is, an influx of Ca++ ions. This in turn stimulates gene activity and the building of new cell connections or the reinforcement of old ones. This is the chemical, genetically driven basis of learning and memory (Davis, 1995).
Memories are thought to be due to lasting synaptic modifications in the brain. The search for memory traces has relied predominantly on determining regions that are necessary for the process. However, a more informative approach is to define the smallest sufficient set of brain structures. In rut mutants, ectopic expression of rutabaga in a spatially restricted fashion, in a defined set of neurons, was used to examine the cellular localization of the memory trait. rutabaga, an enzyme that is ubiquitously expressed in the Drosophila brain and that mediates synaptic plasticity, has been found to be needed exclusively in the Kenyon cells of the mushroom bodies for a component of olfactory short-term memory. This demonstrates that synaptic plasticity in a small brain region can be sufficient for memory formation (Zars, 2000b).
In insects, much attention has been paid to the mushroom bodies as the site for olfactory learning. In Drosophila, they are made up of about 2500 intrinsic neurons (Kenyon cells), receive multimodal sensory input, preferentially from the antennal lobe to the calyx, and send axon projections to the anterior brain where they bifurcate to form the alpha/beta, alpha'/beta', and gamma lobes. Noninvasive intervention techniques can provide mushroom body-less flies. In most respects, these flies show remarkably normal behavior but are deficient in olfactory learning. Genes important for olfactory memory have elevated expression levels in the mushroom bodies (Zars, 2000b and references therein). Additionally, the mushroom bodies are necessary for context generalization in visual learning at the flight simulator and the control of spontaneous walking activity (Martin, 1998; Liu, 1999).
The approach taken is built on the assumption that synaptic plasticity is impaired in general in rut mutants and that it is this cellular defect that causes the various learning deficits. Restoring rut AC in a spatially restricted fashion in a defined set of neurons would furnish synaptic plasticity to only those cells. If in such flies a learning task is rescued, the corresponding memory trace is mapped to the set of neurons expressing the gene, or a subset of these (Zars, 2000b).
Olfactory short-term memory was tested with an apparatus in which flies are sequentially exposed to two odorants, one of which is paired with electric shocks. Shortly after training, ~95% of wild-type flies prefer the odorant not accompanied by punishment. Mutant rut flies show significantly lower memory scores. To test whether the olfactory learning defect of the rut mutant is rescuable, a P-element expressing a wild-type rut cDNA under the control of a GAL4-sensitive enhancer P[UASGAL4-rut+] was combined in the rut mutant with a driver transgene P[elav-GAL4] expressing the yeast GAL4 transcription factor in all neurons. This pan-neuronal expression of rut AC partially restores olfactory learning in the rut mutant. The incomplete rescue could be due to insufficient expression levels of the P[UASGAL4-rut+] transgene, a dominant negative effect of the P[elav-GAL4] element, or a negative effect of ectopically expressing this transgene (Zars, 2000b).
Several GAL4 enhancer trap lines were selected for local rescue because of their expression patterns. Mutant rut flies with the enhancer trap GAL4 elements 247, c772, 30y, 238y, and H24 in combination with the P[UASGAL4-rut+] transgene show memory scores statistically indistinguishable from wild-type flies. The GAL4 line 201y partially rescues the rut learning defect. Finally, rut mutant flies with three other GAL4 enhancer trap elements (c232, 189y, and 17d) and the P[UASGAL4-rut+] effector gene have rut mutant-like short-term memory scores (Zars, 2000b).
Four of the nine enhancer trap lines were previously used to study olfactory learning after locally expressing a constitutively activated G-protein alpha subunit (Galphas*). In the present experiments, the magnitude of rescue was similar to the suppressive effect of the Galphas* protein in the respective lines. In c232, Galphas* has no effect; in 201y, suppression is about 50%, whereas in c772 [the same expression pattern as c747] and 238y, suppression is nearly complete (Zars, 2000b).
Neither the rescuing GAL4 enhancer trap lines without P[UASGAL4-rut+] nor the P[UASGAL4-rut+] line without driver has a dominant rescue effect. Nor do the nonrescuing GAL4 enhancer trap inserts have a negative effect on wild-type flies. Thus, it is the specific interaction of the GAL4 enhancer trap element with the P[UASGAL4-rut+] effector that can rescue the memory defect in rut mutant flies (Zars, 2000b).
Control experiments were conducted with naive wild-type, rut mutant, and potentially rescued rut mutant flies to assure that none of the memory scores were due to changes in shock reactivity or perception of the odorants. All genotypes avoid electric shocks at similar levels. Although 189y and 17d have somewhat reduced shock reactivity, their responses here are similar to that of c772, which shows a wild-type-like learning score. Thus, these shock reactivity scores cannot be responsible for the low memory scores. In addition, all genotypes tested avoid the aversive odorants used in the training protocol (Zars, 2000b).
To determine what brain structures are minimally sufficient for olfactory short-term memory, the expression patterns of the rescue and nonrescue GAL4 enhancer trap lines were examined. Serial sections show that the common structure labeled in all rescuing GAL4 lines is specifically the mushroom bodies. Indeed, comparing expression patterns of rescuing and nonrescuing lines indicates that the gamma lobes may be especially important. In contrast to the GAL4 lines used, the rescuing line 247 lacks expression in the median bundle. The latter, therefore, is not part of the set of minimally sufficient neurons (Zars, 2000b).
On the basis of the current model of how type I ACs function in synaptic plasticity and on the connectivity of the mushroom bodies, the short-term memory trace of odors is localized to a single level in the olfactory pathway: the presynaptic sites in the Kenyon cells contacting extrinsic output neurons and possibly other Kenyon cells in the peduncle and lobes. Modulating neurons carrying the reinforcer must project to the peduncle or lobes and contact presynaptic endings of Kenyon cells there. No rut-dependent synaptic plasticity is required in the antennal lobe or calyx for olfactory learning (Zars, 2000b).
Different brain structures are involved in different learning tasks. In the heat box paradigm, the median bundle, antennal lobes, and ventral ganglion are sufficient for rescue of rut-dependent short-term memory. To find olfactory and heat box memory at different locations was not unexpected, since mushroom body-less flies do well in heat box learning. The task-specific rescue in different GAL4 lines strongly supports the claim that it is the spatial distribution of the rut AC that matters (Zars, 2000b).
Several open questions remain. (1)The conclusions refer only to rut-dependent synaptic plasticity. Although unlikely, the 60% short-term memory remaining in rut mutant flies may reside outside the mushroom bodies. (2) The current understanding of the role of type I ACs in synaptic plasticity and learning is not complete. (3) The P[UASGAL4-TAU] reporter was used to visualize GAL4 expression patterns, and coincidence with P[UASGAL4-rut+] is inferred. (4) Temporal control of transgene expression in these lines is not yet possible, leaving the faint possibility that the behavioral rescue in some cases might be due to developmental expression. (5) Whether all memory traces of odors reside in the mushroom bodies and how memory traces of odors are organized within the mushroom bodies await further investigation (Zars, 2000b).
The technique of restoring synaptic plasticity in minimally sufficient
brain regions has, in two cases, revealed simple, locally confined
memory traces. This result is probably due to the simplicity of the
learning tasks, requiring the animal to store a single sensory modality
for a binary orientation response. It will be of considerable interest
to map memory traces of more complex learning paradigms (Zars, 2000b).
Some memories last longer than others, with some lasting a lifetime. Using several approaches memory phases have been identified. How are these different phases encoded, and do these different phases have similar temporal properties across learning situations? Place memory in Drosophila using the heat-box provides an excellent opportunity to examine the commonalities of genetically-defined memory phases across learning contexts. This study determines optimal conditions to test place memories that last up to three hours. An aversive temperature of 41°C was identified as critical for establishing a long-lasting place memory. Interestingly, adding an intermittent-training protocol only slightly increased place memory when intermediate aversive temperatures were used, and slightly extended the stability of a memory. Genetic analysis of this memory identified four genes as critical for place memory within minutes of training. The role of the rutabaga type I adenylyl cyclase was confirmed, and the latheo Orc3 origin of recognition complex component, the novel gene encoded by pastrel, and the small GTPase rac were all identified as essential for normal place memory. Examination of the dopamine and ecdysone receptor (DopEcR) did not reveal a function for this gene in place memory. When compared to the role of these genes in other memory types, these results suggest that there are genes that have both common and specific roles in memory formation across learning contexts. Importantly, contrasting the timing for the function of these four genes, plus a previously described role of the radish gene, in place memory with the temporal requirement of these genes in classical olfactory conditioning reveals variability in the timing of genetically-defined memory phases depending on the type of learning (Ostrowski, 2014).
Temperature as an aversive reinforcer interacts with training conditions to induce place memories of different stabilities. Previous work showed that intermittent training for Drosophila in space and place memory increases memory performance up to two hours after training. Shown in this study is that temperatures at or above 41°C are needed for induction of this longer lasting memory. That is, 37°C and below can act as an aversive reinforcer and condition flies to avoid a part of the training chamber, but continued avoidance decays within minutes of training. It is only with a temperature of 41°C that an hours-long memory is induced with massed and intermittent training. This abrupt difference in the length of the memory after training with the higher temperature may reflect a threshold of some sort, the steepness of which is currently unknown. This could arise from a differential input to the reinforcing circuit from separate sensory systems, like the Trp family of receptors, or from altered output from one of these sensory systems. Future studies on different temperature responsive proteins may differentiate between these possibilities (Ostrowski, 2014).
Genetic analysis challenges the use of time as a critical factor in determining a memory phase. Memory phases in the fly were initially examined after classical olfactory conditioning where an odorant is typically paired with an aversive electric shock or a rewarding sugar. Four different memory phases have been classified based roughly on time after training and genetic/pharmacological manipulations. Short-term memory after olfactory learning is measured within minutes of training; long-term memory and anesthesia resistant memory start to be active within hours and are increasingly important for memories at the 24 h range and longer. An intermediate memory is thought to be important in the interval between short-term and long-term memories. That time alone is a critical factor in determining these phases loses support when comparing flies with different mutations in aversive and rewarded olfactory memory. For example, the long-known mutant radish was originally shown to be important in the hours-long range after aversive olfactory training and genetically classified the anesthesia-resistant memory. Interestingly, this gene is important within minutes of training in rewarded olfactory memory (Ostrowski, 2014).
Several genes that are important for early to late phases of classical olfactory conditioning are critical on a finer time scale in place memory. Mutation of both the rut and lat genes leads to reduced aversive olfactory memory tested immediately after training, as well as longer time points. Although it is currently unclear when during the life-cycle these genes are important for place memory, mutation of rut and lat reduces memory directly after training. Furthermore, both the rut and lat products have been implicated in synaptic plasticity at the neuromuscular junction (NMJ), which suggests a role for these genes in early stages of learning and memory. It is pretty straight-forward that the rut-encoded type I adenylyl cyclase is also acting early on in associative processes in place learning. The lat gene encoding a subunit of the origin of replication (orc3) is also localized to the pre-synaptic specializations at the NMJs). The lat-orc3 also acts early-on in associative processes for place learning. How the lat-orc3 product is related to regulation of cAMP levels is, however, not as clear. The rut and lat results add to our understanding of an apparently common set of short-term changes in memory between olfactory and place memory, which include a common function of the S6 kinase II, an atypical tribbles kinase, and the arouser EPS8L3. And, the recently identified role of the foxp transcription factor specifically in operant learning, as tested in a flight simulator, suggests another set of genes that could be important for operant place memory in the minutes range (Ostrowski, 2014).
Late memory phases in classical olfactory conditioning depend on a set of genes that are important for place memory within minutes. The first challenge to a common timing of a memory phase came from the radish gene. In contrast to a role in the hours range after olfactory learning, radish mutant flies have a deficit in operant place memory within minutes of training. Furthermore, the pst gene (CG8588), encoding a novel product, has been previously shown to have a specific defect in aversive olfactory memory 24 h after spaced training. That is, the pst mutant flies have a normal short-term olfactory memory but a defective memory 1 day later. Interestingly, in the heat-box pst mutant flies already show a significant decrement in place memory immediately after training. This place memory defect seems to get worse within the first hour after training, reduced to ~50% of normal after 60 min. Thus, this 'long-term memory gene' is also involved in a memory within minutes of training in a second learning situation (Ostrowski, 2014).
Using the classical aversive olfactory learning paradigm the rac small GTPase has been identified as a key regulator in memory retention. Inhibition of Rac activity slows early olfactory memory decay, leading to elevated memory levels one hour after training, but becoming increasingly important 2 h after training. There does not appear to be an effect of Rac inhibition in olfactory memory in the minutes range after training. Transgenic flies with inhibited Rac function also have an increase in memory retention after place memory training. However, the first evidence of an increase in memory performance is within 10 min. Impressively, significant place memory was still evident up to 5 h after training, far beyond the range that can be typically measured in wild-type flies. Thus, while rac has a more general role in stabilizing memories, the timing of this function depends again on the type of memory trace that is formed (Ostrowski, 2014).
Not all memory genes first identified in other contexts, however, play a significant role in place memory. The DopEcR gene has been implicated in several behaviors, including a 30 min memory after courtship conditioning. This G-protein linked receptor is responsive to both dopamine and the steroid hormone ecdysone. Remarkably, DopEcR has been shown to interact with the cAMP cascade through double mutant and pharmacological tests. Using conditions that induce a robust and lasting place memory, the DopEcR mutant flies do not show a defect in memory directly after training or at 1 h post-training. This is despite the fact that the rut and cAMP-phosphodiesterase genes (dunce) are critical for place memory. It may be that DopEcR is not required for this type of learning and would be consistent with the independence of place memory from dopamine signaling. Alternatively, other redundant pathways may compensate for the reduction in DopEcR function caused by the DopEcRPB1 allele. One might further speculate that other types of behavioral plasticity, such as reversal learning or memory enhancement after unpredicted high temperature exposures in the heat-box might be more sensitive to DopEcR changes. Future experiments will determine if this is the case (Ostrowski, 2014).
Memory stability across learning contexts in Drosophila has some common genetic mechanisms, but the timing for gene action depends on the type of learning. That this study has added several genes here, including lat, pst, and rac as regulators of memory stability in operant place memory suggests that there are at least some common molecular processes in memory stability across different learning types. However, the timing of these genetically-defined phases depends on what is learnt. It is speculated that an ideal system to regulate memory stability would be one that activates its own decline. That is, a given memory type should activate the process of decreasing memory expression. This might work with the recruitment of a reinforcing pathway, like the dopaminergic signal that is important for both the acquisition of an associative olfactory memory and the active process of forgetting that association. In this case an odor associated with shock gives rise to a memory trace in mushroom body neurons that depends on a set of dopamine neurons that is important for both memory acquisition and decline. Whether this type of aminergic-based system applies to other forms of memory is not yet known. However, if an aminergic-based signal is common in memory decline, as appears to be the case with the Rac-based mechanism, differences in the types of aminergic neurons or innervation targets could give rise to the altered stabilities of behaviorally expressed memories (Ostrowski, 2014).
This study defines a role of the cAMP intermediate EPAC in
Drosophila aversive odor learning
by means of null epac mutants. Complementation analysis revealed
that EPAC acts downstream from the rutabaga adenylyl cyclase and in
parallel to protein kinase A. By
means of targeted knockdown and genetic rescue, mushroom body Kenyon cells (KCs) were
identified as a necessary and sufficient site of EPAC action.
Mechanistic insights were provided by analyzing acquisition dynamics and
using the 'performance increment' as a means to access the trial-based
sequential organization of odor learning. Thereby it was shown that
versatile cAMP-dependent mechanisms are engaged within a sequential
order that correlate to individual trials of the training session (Richlitzki, 2017).
The cAMP signaling pathway is central to the regulation of plasticity
and can mediate cellular responses via different intermediaries, i.e.,
PKA (protein kinase A), EPACs (exchange proteins directly activated by
cAMP), and CNGs (cyclic nucleotide gated channels). While
the numerous contributions of PKA to the regulation of plasticity have
been described in great detail, the role of EPAC was not
recognized until 1998.
Since then, its operation as a noncanonical cAMP sensor has been proven
in numerous studies, aided by the development of selective cAMP analogs
and/or genetic models that allow discrimination between PKA and EPAC
functions. Epac has been shown to
enhance neurotransmitter release, activate neuronal excitability via Ca2+-dependent
K+-channels, and enhance hippocampal long-term
potentiation and memory consolidation. This study investigated a potential role of Epac
in the Drosophila aversive odor-learning paradigm (Richlitzki, 2017).
This study has distinguish epac-dependent from epac-independent learning by
means of a Drosophila null-epac mutant and used the performance increment
as a means to address the functional disparity of individual training
trials. Disparity impacts on the learning rate suggesting an
evolutionary benefit of alternative cAMP mediators as this provides a
mechanism for transforming rut-derived cAMP signals into behavioral
output following different strategies (Richlitzki, 2017).
Rutabaga adenylyl cyclase (rut-AC1) is supposed to act as
coincidence detector between US- and CS-derived impulses that converge
at the level of KC synapses and consequently induce cAMP-dependent
plasticity within an odor specific matrix of KCs. This KC synapse matrix
is widely accepted as a neuronal representation of learning and thought
of as an engram of an odor memory. Dopaminergic
neurons (DANs) provide the major share of KCs' dopamine-induced cAMP
signals, however KC synapses are not uniform but exhibit different
sensitivities for cAMP. Likewise,
learning is not an instantaneous process but develops over the multiple
trials of a training session. As a consequence, cAMP gain over the time
course of training would not be uniform but effectively determined by
the sensitivity of a particular KC synapse within the odor specific
matrix. This disparity would proceed to the next levels, i.e.,
activation gain of alternative downstream cAMP intermediaries that, in
turn, are unequally sensitive as reflected by characteristic
half-maximal cAMP concentrations (IC50), i.e., IC50 values for PKA ~80
nm; for HCN channels ~ 100 nm; and for EPAC ~ 800 nm. By this design, KCs diversify into high and low
cAMP gain fractions that activate mediators responsive to either high
and/or low cAMP amplitudes at quite different rates. Thereby, a
particular odor-specific KC matrix can interconnect to multiple outputs
via different mechanisms of cAMP-dependent plasticity as its synaptic
elements exhibit disparate hysteresis, i.e., different time-dependent
changes in the cellular cAMP levels (Richlitzki, 2017).
The surprising result that insensitive EPAC recruits early, i.e., during the
second trial of the training session, suggests existence of a fast,
high-amplitude cAMP signal that meets EPAC's thresholds early during
training. Similarly, one has to assume that the efficacy of later
training trials is mediated by a perpetuating low-amplitude signal that
develops with low gain over multiple trials. Given the fact that PKA
acts isogenic to EPAC and null epac mutants show regular learning during
late training trials-that is from the third trial onward-neither of
these intermediates is likely to account for late trial learning. One
plausible mechanism would be HCN channels, i.e., subthreshold,
voltage-gated ion channels that reduce membrane resistance and promote
neuronal firing probability. Within sparsely firing KCs
such channels might stabilize an odor-specific synaptic matrix, i.e.,
the CS-representation, over the 1-min time course of the training cycle
and promote its transfer to mushroom body output neurons (MBONs), the
recognized convergence site for KCs. In contrast, epac has been shown to
enhance neurotransmitter release and/or activated neuronal excitability via different mechanisms (Richlitzki, 2017).
While epac-dependent and independent learning mechanisms clearly
dissociate at the molecular level, their learning rates appear
counter-intuitive as a high-amplitude signal precedes a low-amplitude
one, but both originate from rut-AC1. If one considers the animal's need
to trade off certainty of a prediction against its computation time,
this seemingly counterintuitive design appears deliberate and beneficial
as it holds the possibility of combining both contrarian needs (DasGupta, 2014): First, epac-dependent learning requires a short
computation time, i.e., after two trials, compared with the slow and
cumbersome integration over multiple trials needed for epac-independent
learning. Second, epac-dependent learning is restricted to salient
conditions, i.e., high voltages that represent a serious threat to the
animal's health, while this part of the animal's memory is spared with
the 15 V DC US. In fact, wild-type strains trained with 15
V exhibited similar learning to epac-null mutants trained with 120 V
suggesting that epac amplifies conditioned avoidance under trusted
environmental circumstances (Richlitzki, 2017).
Do training trials clock recurrent computation within the learning
network? How individual training trials are represented within the fly
brain is unclear. However, functional studies have identified DANs as
critical substrates of the US that tightly innervate KCs at the level of
the MBs. In general, DANs and KCs work together with MBONs, the
recognized readout routes of aversive odor memory. Moreover, their anatomy suggests that MBONs serve as critical inter-loops that reiterate the MBs' computational output to DANs, i.e., its major modulatory input: This recurrent connectivity exhibits a
remarkable zonal architecture as dendrites of MBONs tile the length of
KC axons in a nonoverlapping manner, where they meet with dopaminergic
neurons (DANs), the other main innervation of the MB lobes. The DANs
tile the MB lobes in a corresponding manner so that the dendrites of a
particular MBON meet axonal projections of cognate DANs. Moreover,
dendrites of DANs overlap with MBON axons within the MBON projection
zones outside the MBs suggesting that MBONs modulate the activity of
DANs and thereby generate recurrent loops. By this
design MBON activity is dually modulated by DANs, first via a direct
connection, and second via a KC detour that undergoes cAMP-dependent
plasticity. However, further research will be required to understand the
rules by which repetitive trials clock the computation within the
DAN/KC/MBON network (Richlitzki, 2017).
Learning and memory rely on dopamine and downstream cAMP-dependent plasticity across diverse organisms. Despite the central role of cAMP signaling, it is not known how cAMP-dependent plasticity drives coherent changes in neuronal physiology that encode the memory trace, or engram. In Drosophila, the mushroom body (MB) is critically involved in olfactory classical conditioning, and cAMP signaling molecules are necessary and sufficient for normal memory in intrinsic MB neurons. To evaluate the role of cAMP-dependent plasticity in learning, this study examined how cAMP manipulations and olfactory classical conditioning modulate olfactory responses in the MB with in vivo imaging. Elevating cAMP pharmacologically or optogenetically produced plasticity in MB neurons, altering their responses to odorants. Odor-evoked Ca(2+) responses showed net facilitation across anatomical regions. At the single-cell level, neurons exhibited heterogeneous responses to cAMP elevation, suggesting that cAMP drives plasticity to discrete subsets of MB neurons. Olfactory appetitive conditioning enhanced MB odor responses, mimicking the cAMP-dependent plasticity in directionality and magnitude. Elevating cAMP to equivalent levels as appetitive conditioning also produced plasticity, suggesting that the cAMP generated during conditioning affects odor-evoked responses in the MB. Finally, this plasticity was found to be dependent on the Rutabaga type I adenylyl cyclase, linking cAMP-dependent plasticity to behavioral modification. Overall, these data demonstrate that learning produces robust cAMP-dependent plasticity in intrinsic MB neurons, which is biased toward naturalistic reward learning. This suggests that cAMP signaling may serve to modulate intrinsic MB responses toward salient stimuli (Louis, 2018).
Learning generates plasticity in neuronal responses to input stimuli, which is distributed across multiple cells and synapses in the brain. Molecularly, dopamine and downstream cAMP signaling are involved in multiple forms of memory, including olfactory learning. For instance, dopamine is required in the amygdala for olfactory classical conditioning in mammals. Similarly, dopamine and downstream cAMP signaling molecules play a central role in olfactory classical conditioning in Drosophila. This pathway is particularly critical in the mushroom body (MB), a brain region that receives olfactory information and is required for olfactory learning. Dopaminergic neurons are postulated to convey a reinforcement signal to the MB-stimulating certain subsets of MB-innervating dopaminergic neurons drives aversive or appetitive reinforcement in lieu of a physical reinforcer. The dopamine released from these neurons acts directly on intrinsic MB neurons, and possibly other neurons in the area as well. The D1-like receptor DopR, type I adenylyl cyclase Rutabaga (Rut), catalytic domain of protein kinase A, and Dunce phosphodiesterase (Dnc) are all required for olfactory classical conditioning. Importantly, rescuing the expression of DopR or Rut -- specifically in intrinsic MB neurons of otherwise mutant animals -- restores normal olfactory learning and memory. Further downstream, both Epac and PKA, as well as phosphorylation targets such as synapsin, have been shown to regulate learning and memory via effects in MB neurons. Thus, dopamine and cAMP are critical in intrinsic MB neurons for normal memory. Furthermore, broadly elevating cAMP generates plasticity in MB neurons, demonstrating that this pathway influences the responsivity of MB neurons. However, the role of this pathway in driving coherent patterns of plasticity that encode memory is unknown (Louis, 2018).
Recent advances have opened up the possibility of understanding how olfactory memory is encoded in exquisite detail. Recent studies of memory encoding in the Drosophila MB have suggested that mushroom body neurons are highly plastic, exhibiting learning-related changes in odor responses. This is supported by observations of memory traces using in vivo Ca2+ imaging of neurons innervating the MB. However, the neuronal changes associated with cAMP-dependent, short-term memory are unclear. Conditioning generates plasticity in α'/β'-neurons within a few minutes of training, a time point at which the animals exhibit robust short-term memory. However, the Rut cyclase is not required in α'/β'-neurons for learning, leaving the functional role of cAMP-dependent plasticity in the MB unclear. MB γ-neurons exhibit depression in response to an aversive conditioned odor that is sensitive to manipulations of G αo-signaling, though it is not clear how this relates to dopaminergic modulation via G αs. Finally, blocking the synaptic output of MB neurons during conditioning does not impair aversive learning, suggesting that a significant proportion of the engram resides in the MB neurons and/or upstream connections (Louis, 2018).
In contrast, other studies have described a major role for plasticity in downstream MB output neurons (MBONs), which may arise via pre- and/or postsynaptic plasticity. Robust, dopamine-dependent plasticity has been observed in MBONs, but not at the cellular level in MB neurons. This emphasizes the role of the MB in encoding sparse, relatively invariant olfactory representations. Learning-induced plasticity is then layered in at the MB-MBON synapses, possibly via synaptic depression. This leaves the requirement of cAMP signaling molecules in the MB, and the dispensability of MB output during memory acquisition, unresolved. Thus, there is a paradoxical dissociation of anatomical loci between where cAMP signaling is required and where robust, short-term, learning-induced plasticity has been reported. This study has examined the role of cAMP-dependent plasticity in the MB using in vivo imaging, combined with pharmacological and optogenetic manipulation of cAMP levels. Results suggest that cAMP-dependent plasticity localizes to intrinsic MB neurons and mirrors the plasticity induced during olfactory classical conditioning, with a bias toward appetitive conditioning (Louis, 2018).
The present data support several major conclusions about the role of cAMP-dependent plasticity in the memory-encoding MB: (1) Intrinsic MB neurons exhibit robust cAMP-dependent plasticity; (2) cAMP-dependent plasticity is heterogeneous, both across and within anatomical classes of MB neurons; (3) the directionality and magnitude of plasticity parallel Rut-dependent associative changes in MB responsivity following appetitive classical conditioning; and (4) appetitive conditioning produces changes in cAMP of a magnitude that generates plasticity in odor-evoked responses. Thus, cAMP-dependent plasticity plays a major role in modulating intrinsic MB neurons, directly linking the physiology of MB neurons to the behavioral roles for cAMP signaling molecules in learning and memory. One caveat to the interpretation of imaging studies is that the preparations require tethering the animal under a microscope. Future developments enabling recording of brain activity in freely behaving animals will be necessary to test how responses in the MB neurons facilitate behavioral output in real time (Louis, 2018).
In the context of olfactory learning, the MB encodes a sparse representation of olfactory space, which is computationally advantageous for learning and potentially modulated by learned valence. If neurons responded homogeneously to input stimuli, coincidence detection would result in uniform plasticity across the sparse set of neurons that encode the odor and receive a reinforcement signal. However, the heterogeneity observed in cAMP-dependent plasticity in this study suggests that olfactory memory traces may be driven to specific subsets of 'eligible' neurons in the MB. This could play an analogous role to memory allocation, which drives memory traces to subsets of eligible neurons in the mammalian amygdala during fear conditioning. Molecularly, heterogeneity may be driven by differential expression of genes that function downstream of cAMP/Epac/PKA to regulate neuronal excitability or presynaptic function. Such differences in expression could be set up via developmental or epigenetic mechanisms (Louis, 2018).
It is proposed that the cAMP-dependent plasticity in the MB plays two roles during olfactory learning: filtering MB responses based on salience and encoding valence. A role for MB plasticity in salience filtering is suggested by the observation that appetitive conditioning produced enhancement of MB responses across spatial compartments. These compartments have been suggested to route olfactory signals to valence-encoding output neurons, driving learned approach or avoidance via heterosynaptic plasticity. Therefore, the broad pattern of plasticity observed in this study would affect multiple downstream output pathways of opposing valence, suggesting that it does not encode valence per se. Rather, it may function to heighten relative MB sensitivity to salient stimuli. Across multiple sensory systems, ascending information is filtered according to salience, typically enhancing responses to stimuli that are biologically important. Alternatively, appetitive conditioning may modulate MB neurons in a fundamentally different way from aversive conditioning. While these opposing forms of memory require many overlapping MB-associated neurons, there are some differences in the circuits recruited during these forms of learning, and plasticity across MB neurons may be one difference. Regardless of the interpretation, the data reveal cAMP-dependent plasticity at the cellular level in intrinsic MB neurons. This may be layered on top of synaptic plasticity at MB output synapses, which have been proposed to encode valence by altering how olfactory signals flow through the neuronal networks that mediate behavioral approach or avoidance (Louis, 2018).
Several additional lines of evidence support the idea that cAMP-dependent plasticity serves as an overall gain control, regulating MB responses based on stimulus salience. First, the MB and MB-innervating dopaminergic neurons modulate salience-based decision making in a visual flight simulator paradigm. Second, dopaminergic neurons innervating the MB respond broadly to sensory stimuli that do not have an acquired valence, and exhibit activity that is correlated with locomotion. Activation of these neurons elevates cAMP in the downstream MB neurons in a compartmentalized manner, which in turn modulates their sensitivity and neurotransmission at the MB-MBON synapses. Thus, the MB neurons receive dynamically regulated dopaminergic inputs that alter the function of both the MB and downstream network components as a function of behavioral state. This may facilitate learning in situations in which the animal is likely to experience biologically important events (e.g., during foraging). Similar modulatory mechanisms modulate plasticity and memory in other animals as well. For instance, in honeybees, appetitive conditioning prolongs odor responses in MB neurons. Likewise, in the mammalian amygdala, coactivation of neuromodulatory and Hebbian plasticity is necessary for plasticity and memory (Louis, 2018).
Aversive conditioning produced no significant plasticity in the current study, consistent with results from some optogenetic reinforcement substitution imaging experiments. However, since Rut is required in MB neurons for normal aversive memory, cAMP-dependent plasticity is likely present in some form. Indeed, a previous study detected plasticity in the γ-lobe following aversive conditioning, which could be tightly localized to specific output synapses or neuronal subsets. Pairing odor with stimulation of tyrosine hydroxylase Gal4-labeled dopaminergic neurons produces aversive memory and detectable plasticity in MB γ-neurons. In this study, robust plasticity differentially following appetitive conditioning. This may be due to a bias toward learning about stimuli that guide motivationally relevant behaviors, such as approaching food-associated odors. Consistent with such an interpretation, appetitive conditioning produces memory that is more stable over time than aversive memory. A single trial of appetitive conditioning leads to the formation of long-term memory, while aversive conditioning requires multiple-spaced trials. The cAMP-dependent plasticity during appetitive conditioning could trigger downstream molecular pathways necessary to engage long-term memory formation. This presumably interacts with Ca2+ levels in neurons to regulate short- and long-term memory. In honeybees, elevating intracellular Ca2+ during a single-trial conditioning, which normally only triggers short-term memory, can induce long-term memory, whereas decreasing intracellular Ca2+ during multiple-spaced training impaired long-term memory formation. In addition, appetitive memory retrieval is motivationally gated by hunger state, suggesting a tie-in with motivational state. Integrating these observations, this suggests that motivationally relevant stimuli may enhance the sensitivity of MB neurons via cAMP-dependent plasticity, modulating the overall gain of the system in a salience-dependent manner (Louis, 2018).
The MB is involved in multiple distinct yet potentially interrelated behaviors, including several forms of learning and memory, regulating sleep and activity, context generalization, habituation, temperature preference, context dependence of olfactory behaviors, and salience-based decision making. The common thread among these behaviors is that they revolve around selection of an appropriate action based on context. Thus, a primary function of the MB and its modulatory input may to be alter the probability of action based on integrating environmental cues and internal state. In such a scenario, modulating the overall gain of the circuit could function in concert with fine-scale synapse-specific plasticity to alter the flow of information to downstream motor areas. Thus, these data support a model in which dopaminergic neurons and downstream cAMP-dependent plasticity modulate MB responses to stimuli based on their salience, priming the animal to engage in appropriate goal-oriented behaviors (Louis, 2018).
Habituation is the process that enables salience filtering, precipitating perceptual changes that alter the value of environmental stimuli. To discern the neuronal circuits underlying habituation to brief inconsequential stimuli, a novel olfactory habituation paradigm was developed, identifying two distinct phases of the response that engage distinct neuronal circuits. Responsiveness to the continuous odor stimulus is maintained initially, a phase termed habituation latency; it requires Rutabaga Adenylyl-Cyclase-depended neurotransmission from GABAergic Antennal Lobe Interneurons and activation of excitatory Projection Neurons (PNs) and the Mushroom Bodies. In contrast, habituation depends on the inhibitory PNs of the middle Antenno-Cerebral Track, requires inner Antenno-Cerebral Track PN activation and defines a temporally distinct phase. Collectively, these data support the involvement of Lateral Horn excitatory and inhibitory stimulation in habituation. These results provide essential cellular substrates for future analyses of the molecular mechanisms that govern the duration and transition between these distinct temporal habituation phases (Semelidou, 2018).
Drosophila is a premier system for molecular approaches to understand habituation because of its advanced molecular and classical genetics. In fact, it is a well-established model for habituation of various sensory modalities such as taste, vision, mechanosensory and escape responses, reflecting that habituation is apparent in most, if not all, circuits and modalities of the nervous system. However, in most of these paradigms, the circuits engaged to process the stimulus and establish the experimentally measured attenuated behavioral response are unclear. Importantly, the advanced understanding of the Drosophila olfactory circuitry and stimulus processing facilitates exploration of the mechanisms mediating decreased stimulus responsiveness and habituation to inconsequential odors. Such a recently described paradigm of olfactory habituation in Drosophila required 30 min of odor exposure and was mediated entirely by antennal lobe neurons. In contrast, habituation to repetitive 30 s odor pulses required functional Mushroom Bodies, neurons on the central brain also implicated in associative learning and memory in flies (Semelidou, 2018).
To resolve this paradox, this study focused on the early behavioral dynamics of habituation upon continuous odor stimulation. To that end, a novel habituation paradigm was developed and characterized to rather brief continuous odors. The behavioral responses define two distinct phases, an initial phase termed habituation latency, when stimulus responsiveness is maintained, which is followed by a significant response decrement reflecting habituation. Analogous response dynamics have been reported for footshock habituation. In addition, whether these phases engage and are mediated by distinct neuronal circuits was. The results highlight the stimulus duration-dependent activation of specific neuronal subsets and their distinct roles in securing timely habituation latency and habituation induction (Semelidou, 2018).
This study describes a novel olfactory habituation paradigm to brief odor stimuli and operationally defines two distinct phases in the response dynamics. The initial period of ~120 s is termed habituation latency and is characterized by maintenance of responsiveness to the odor. This is followed by manifestation of the habituated response, characterized behaviorally by attenuated osmotaxis. Focusing on the behavioral dynamics early in the process complements previous work olfactory habituation to continuous odor stimulation in Drosophila. A number of criteria differentiate these two paradigms from other types of habituation to olfactory stimuli as discussed below (Semelidou, 2018).
Drosophila habituate equally well to continuous or pulsed olfactory stimuli. This likely reflects the nature of olfactory stimuli, which typically are continuous rather than pulsed. On the other hand, habituation of the startle response to ethanol vapor may specifically require short (30 s) pulses due to its sedative properties and this may also be reflected by the rather long 6 min ITIs compared to the 15 s to 2.5-min intervals used herein for OCT. Short odor pulses are also required for the odor-mediated jump and flight response habituation, suggesting that pulsing may be necessary to evoke the startle response per se (Semelidou, 2018).
An important property shared with all habituation paradigms in Drosophila and other systems is spontaneous recovery of the response. This is another differentiating parameter among habituation paradigms in Drosophila. For the olfactory habituation paradigms, whereas 6 min suffice for spontaneous recovery after 4 and 30 min continuous odor exposure, 15-30 to surprisingly 60 min
are required for recovery in the olfactory startle paradigms. Habituation to mechanosensory stimuli typically also requires shorter spontaneous recovery times, with habituation of the giant fiber-mediated jump-and-flight response requiring a mere 2 min and electric footshock habituation 6 min. Interestingly, other non-mechanosensory habituation paradigms require long spontaneous recovery periods with 30 min for habituation of the proboscis extension reflex (PER), and surprisingly, 2 hr for habituation of odor-induced leg response. It is posited that these differences reflect the engagement of distinct neuronal circuits mediating habituation to these diverse stimuli and the properties and connections of the neuronal types that comprise them (Semelidou, 2018).
Overall, these data suggest that latency and habituation to brief odor exposure involve modulation of lateral horn (LH) output, a neuropil innately encoding response valence to odor stimuli. It is propose dthat habituation latency involves processes that are not permissive to, or actively prevent stimulus devaluation. Latency duration depends on stimulus strength and is consistent with the notion that it is adaptive not to devalue strong, hence potentially important stimuli, expediently. In fact, it is posited that habituation latency serves to facilitate associations with concurrent stimuli, a requirement for associative learning. Shortened latency leading to premature habituation is predicted to compromise associative learning (Semelidou, 2018).
Importantly, maintaining responsiveness early upon odorant exposure requires activity of GABAergic inhibitory neurons], which are essential for lateral inhibition of antennal lobe glomeruli. LN activation appears to prevent saturation by strong continuous odors and hence reduce PN activity. Therefore, shortening habituation latency by blocking GABAergic neurotransmission in the antennal lobe may effectively reduce stimulus intensity, expediting habituation as suggested by the dilute odor experiments. This interpretation is further supported by the decreased habituation latency upon silencing the iACT PNs conveying olfactory signals to the MBs and the LH, but not by the mACT neurons innervating only the LH. Since iACT PNs are mainly excitatory, it appears that response maintenance requires excitatory signaling to the LH and the MBs (Semelidou, 2018).
All MB neuronal types except the γ, are essential for habituation latency. This suggests that at least part of the excitatory signal conveyed by the iACT PNs impinges upon the αβ and α' β' MB neurons, which is consistent with their role in associative learning and the proposal that habituation latency facilitates it. Neurotransmission from the MBs to LH neurons mediating aversive responses likely engages MB output neurons (MBONs), to maintain the valence and intensity of the odor and sustain aversion. Distinct MBONs are known to drive both attraction and aversion to odors and their potentially differential involvement in habituation is currently under investigation (Semelidou, 2018).
Dishabituation results in stimulus value recovery and apparently resets habituation latency. Clearly it requires neurotransmission via the GH146-marked neurons and MBs because silencing these neurons disables dishabituation, consistent with their role in response maintenance. These results lead to the hypothesis that dishabituating stimuli might converge on the MBs and/or iACT, possibly stimulating excitatory neurotransmission to the LH, to reinstate stimulus aversion. This hypothesis is currently under investigation as well (Semelidou, 2018).
In contrast, habituation requires prolonged or repeated exposure to the odorant and functional iACT and mACT PNs converging on the LH. Interestingly, the mainly GABAergic mACT PNs receive input both from the olfactory sensory neurons and the excitatory iACT PNs. Their depolarization also activates the excitatory iACT neurons via direct chemical synapses. This apparent feedback loop may be required for mACT activation after prolonged exposure to aversive odors, since these neurons were reported to respond mainly to attractive stimuli. It is proposed that prolonged aversive odor exposure enhances iACT activation, which in turn leads to habituation, while shorter exposure does not activate the iACT neurons, reflected by their dispensability for habituation latency. Importantly, the mACT innervates the LH downstream of the iACT PNs, providing feedforward inhibition. These characteristics likely underlie the necessary and sufficient role of mACT PNs in habituation upon 4-min odor stimulation. Collectively, these results are consistent with the proposal that mACT activation inhibits the innate LH-mediated avoidance response to the aversive odorant, establishing habituation (see A model of the neuronal subsets underlying (A) Habituation Latency and (B) Habituation, after exposure to aversive stimuli.). However, full mACT activation appears to also require iACT neurotransmission, which if abrogated eliminates habituation but is insufficient to establish it on its own (Semelidou, 2018).
Because the MZ699 Gal4 driver also marks ventrolateral protocerebrum (vlpr) neurons it is possible that they also play a role in habituation. In fact, vlpr neurons function in aversive odor responses, are activated by excitatory iACT PNs, inhibited by the inhibitory PNs, and are afferent to the LH. Thus, they could act in parallel or synergistically to mACT PNs to establish the habituated response. As no specific vlpr driver is available, it is impossible at the moment to address this possibility directly. Briefly then, the current collective results strongly suggest a novel role for the inhibitory PNs innervating the LH, and possibly vlpr neurons, in inhibition of the innate response and habituation. The kinetics of inhibitory projection neuron activation and their output on downstream neurons could serve as a measure of the duration of odor exposure. Upon prolonged exposure, these neurons mediate inhibition of odor avoidance, thus devaluing the stimulus (Semelidou, 2018).
Analysis of the neuronal subsets underlying habituation has focused on aversive odors. However, considering the neuronal clusters involved in the process, it would be relatively safe to assume that the results extend to attractive odor habituation as well. It is possible that the neuronal circuitry comprised of PNs, the LH and MBs may be mediating habituation independently of odor valence. However, specific neuronal clusters may differ in odor valance-dependent activation or inhibition of other circuit components with opposing effects on the behavioral readout. For example, inhibitory PNs (iPNs) mediate attraction by releasing GABA in the LH to inhibit avoidance. If inhibited themselves, the resultant attenuated attraction will likely drive a behavioral output of habituation to an attractive odor (Semelidou, 2018).
In accord with this notion, attractive and aversive odors are represented in different AL glomerular clustersand this valence-dependent organization is preserved into higher brain centers. In fact, the posterior-dorsal LH responds to attractive and its ventral complement to aversive odors, while third order neurons convey information from ventral LH to the vlpr and from the dorsal LH to the superior medial protocerebrum. This organization potentially reflects differential recruitment of these neuronal clusters in habituation to aversive and attractive odors. The circuits involved in habituation to attractive odors and their specific contribution to the process will be the focus of future work (Semelidou, 2018).
Although behaviorally there is significant osmotactic attenuation after both 4 and 30 min aversive odor exposure, the experiments suggest that these represent distinct types of olfactory habituation. Habituation after 4 min of odor exposure does not require the MBs, but rather the projection neurons innervating the LH. Habituation after 30 min of exposure is also independent of MB function, but appears to be entirely mediated by iLNs and reside within the AL. This clear difference suggests that the specific potentiation of inhibitory synapses shown to underlie habituation after 30 min of exposure is not necessary for habituation to the brief 4-min exposure. Additionally, while Rut is required within the iLNs during the latency period upon brief odor exposure, it is surprisingly required within the same neurons for habituation to long odor exposure. Therefore, Rut-driven activity within the iLNs yields opposing time-dependent behavioral outputs in accord with the abovementioned notion that the same circuit components may drive opposing outputs (Semelidou, 2018).
Furthermore, the fact that mechanosensory stimuli are not effective dishabituators after 30 min of odor exposure as they are after 4 min, augments the conclusion these are different types of olfactory habituation and suggests that distinct dishabituators likely recruit different neuronal subsets to modulate the habituated response. Such neuronal circuits and the effect of different dishabituators in response recovery are currently under investigatio (Semelidou, 2018).
Altogether, the results indicate different mechanisms for 4 min and 30 min habituation to aversive odors with the former mediated by the interaction between iPNs, ePNs and their targets in the LH, while the latter is based on the inhibition of ePNs by iLNs at the AL level. However, it is possible that the potentiated PN inhibition would decrease their output to the LH to drive reduced avoidance. This argues that the LH could be involved in the behavioral output indicating habituation after 30 min of OCT exposure as well. An AL-mediated reduction in the perceived intensity or valence of a chronically present odor probably serves an adaptive evolutionary role distinct from short exposure to the same stimulus. In fact, filtering away the chronic odor at the antenna, the first olfactory synaptic station, might facilitate evaluation of additional odors at higher order neurons such as the MBs or the LH (Semelidou, 2018).
Significantly, this interpretation is congruent with timescale habituation in mice, where short-timescale odor habituation is mGluR-dependent and mediated by the anterior piriform cortex while long-timescale habituation requires NMDAR and is mediated by the olfactory bulb. In addition, studies in mice, rats and primates have shown that habituation of the higher order neurons is faster and more prominent than in olfactory bulb neurons. Therefore, temporal and spatial principles for olfactory habituation appear broadly conserved between insects and mammals, despite their evolutionary distance (Semelidou, 2018).
Odor-based learning and innate odor-driven behavior have been hypothesized to require separate neuronal circuitry. Contrary to this notion, innate behavior and olfactory learning were recently shown to share circuitry that includes the Drosophila mushroom body (MB). But how a single circuit drives two discrete behaviors remains unknown. This study defines an MB circuit responsible for both olfactory learning and innate odor avoidance and the distinct dDA1 dopamine receptor-dependent signaling pathways that mediate these behaviors. Associative learning and learning-induced MB plasticity require rutabaga-encoded adenylyl cyclase activity in the MB. In contrast, innate odor preferences driven by naive MB neurotransmission are rutabaga independent, requiring the adenylyl cyclase ACXD. Both learning and innate odor preferences converge on PKA and the downstream MBON-γ2α'1. Importantly, the utilization of this shared circuitry for innate behavior only becomes apparent with hunger, indicating that hardwired innate behavior becomes more flexible during states of stress (Noyes, 2023).
These data reveal the shared use of a discrete circuit for both state-dependent odor-driven behavior and experience-dependent odor learning. The shared components include the upstream DA neurons, the MBn-expressed DA receptor dDA1, and the downstream MBON. Odor response processing for state-dependent behavior and odor learning diverge at the level of the dDA1 receptor-activated adenylyl cyclase, with ACXD employed for innate state-dependent odor driven behavior and rut employed for olfactory learning. The unique activation of rut for olfactory learning is explained by the fact that this adenylyl cyclase functions as a coincidence detector, synergistically responding to both DA receptor activation from the unconditioned stimulus and Ca2+ increases due to the conditioned stimulus (Noyes, 2023).
ACXD is a transmembrane AC that is expressed in a number of tissues including the brain (flyatlas.org) and is orthologous to the mammalian AC2. Mammalian AC2 activity is Ca2+ independent.
If ACXD is also Ca2+ independent, it would provide a mechanism for the engagement of distinct cAMP pathways by dDA1 for state-dependent versus experience-dependent olfactory behavior. Thus, common neural circuitry is employed for both state-dependent and conditioned behaviors with the unique changes of MBn output influenced by the intracellular signaling pathways that are mobilized.
Dopaminergic input to the dDA1 DA receptor expressed in the γ2 compartment of MBn activates an intracellular signaling pathway that includes the ACXD adenylyl cyclase, PKA activity, and the release of ACh. The downstream MBON-γ2α'1 responds to the MB ACh release through the α2 nACh receptor, with the activity of the MBON-γ2α'1 ultimately dictating the balance in state-dependent odor approach/avoidance. The simplest model to account for the state-dependent MBON activity would have the internal state modulating DA input into to the MBn to increase or decrease ACh release onto the MBON. However, the current data failed to detect a significant change in ACh release between the fed and starved conditions. Nevertheless, the activity of the PPL1-γ2α'1 that influences the MB γ2 compartment is required for state-dependent behavioral responses to odor. A proposed model for reconciling these observations envisions that the basal activity of this circuit is required for behavioral-state odor choice but that starvation mobilizes a qualitative or quantitative signal independent of the magnitude ACh release by the MBn to increase MBON activity. An unidentified signal representing hunger could directly enhance MBON excitability. For example, octopamine has been proposed as a feeding signal that acts directly on MBONs (Noyes, 2023).
Similarly, a hunger signal could act on neurons elsewhere in the brain that ultimately connect to MBONs through intermediary neurons. The hunger-responsive neuropeptide leucokinin acts on DAns that connect to MBONs (Noyes, 2023).
Alternatively, there may be a co-neurotransmitter released by the MBn due to starvation that works to increase MBON activity. Finally, the possibility that starvation does modulate MBn ACh output is left open, since the reporters employed lack the sensitivity to detect this change. Future investigations into state-dependent changes in MBON-γ2α'1 physiology will need to grapple with numerous competing hypotheses (Noyes, 2023).
Changes in odor responses in MBONs have suggested that learning induces a change in connectivity in the MBn-MBON synapse. In addition, compartment specific plasticity in MB ACh release was discovered that fits with the idea that plasticity observed in MBONs occurs from the input of MB compartments (Noyes, 2023).
However, there has been a lack of data connecting the MBn ACh release plasticity with MBON plasticity and particularly to the central role for the rut adenylyl cyclase. The current data offer this important connection. It shows that the MB γ2 compartment undergoes a rapid depression in response to odor/shock pairings during aversive learning and that rut is required for the acquisition of this depression. Downstream of the MB γ2 compartment, MBON-γ2α'1 drives approach and also undergoes a learning-induced depression (Noyes, 2023).
These results put prior speculation about how the genetic regulation of cAMP signaling through the rut adenylyl cyclase drives Drosophila memory on concrete ground. The work does not conclusively delineate a role for dDA1 in the MB plasticity. Loss of MB dDA1 dramatically reduced naive odor responses in MB γ2. This precluded attempts to measure dDA1 effects on MB γ2 depression because the naive responses were already low. Interestingly, both learning- and starvation-dependent odor avoidance require PKA. A likely explanation is that rut and ACXD are spatially segregated, creating distinct cAMP microdomains or signaling platforms. Thus, PKA activity would result in the phosphorylation of unique substrates within those microdomains (Noyes, 2023).
The characterization of the MB as a brain region for learned, but not innate, olfactory behavior was motivated by experiments eliminating MBn or blocking MB output. Disrupting the MB eliminates odor-associated memory but has no effect on innate avoidance of those same odors. Recent work has overturned this simple categorization demonstrating some DANs, MBONs, and MBns do contribute to innate olfactory behavior in certain circumstances. Interestingly, the majority of reports define a role for MBns in innate behavioral responses to food-related odors (Noyes, 2023).
The results, using more general, non-food odors, puts the hypothesis that MBn regulates innate behavior on more solid ground. Importantly, it was found that MB dDA1 is required for state-dependent behavior to general odors. This is in contrast to a report finding that dDA1 is not involved but that DAMB is required for state-dependent behavior to food odors (Noyes, 2023).
This difference will be a key element to understand state-dependent behavior moving forward.
The Drosophila and mammalian olfactory systems are remarkably similar in terms of anatomical organization and function. In both, odorant molecules activate olfactory sensory neurons (OSNs), with each OSN only expressing one type of odorant receptor (OR). Each OSN expressing the same type of OR project to the same glomeruli (Noyes, 2023).
Within the glomeruli, the OSNs synapse onto projection neuron (PN) dendrites, and PN activity is modified in the glomeruli by local inhibitory interneurons before being sent on to multiple higher-order brain regions. PN neurons connect to downstream neurons in the mammalian piriform cortex
and in the Drosophila MB in a seemingly random manner. Like the Drosophila MB, the piriform cortex is critical for olfactory memory. It is not clear how the piriform cortex is involved in state-dependent olfactory behavior. However, in humans, odor coding changes in the piriform cortex with hunger and sleep deprivation, and piriform cortex neuron activity levels are inversely correlated with sexual satiety in rats (Noyes, 2023).
It is concluded from these results demonstrating dDA1-dependent MBn Ach release and a dDA1-dependent MBON-γ2α'1 Ca2+ in response to odor that dDA1 directly modulates the MBn/MBON connection. However, due to a limitation in the sensitivity of the ACh sensor employed, it was not possible to directly record ACh input to MBON-γ2α'1. Based on the established direct cholinergic connection between these MBn and MBONs and the lack of any known non-MB cholinergic innervation to this brain region, it is believed that the conclusions are merited. However, a formal possibility that other intermediary neurons mediate this relationship must be left open (Noyes, 2023).
Mate choice constitutes a major fitness-affecting decision often involving social learning leading to copying the preference of other individuals (i.e., mate copying). While mate copying exists in many taxa, its underlying neurobiological mechanisms remain virtually unknown. This study shows in Drosophila melanogaster that the rutabaga gene is necessary to support mate copying. Rutabaga encodes an adenylyl cyclase (AC-Rut(+)) acting as a coincidence detector in associative learning. Since the brain localization requirements for AC-Rut(+) expression differ in classical and operant learning, this study determine the functional localization of AC-Rut(+) for mate copying by artificially rescuing the expression of AC-Rut(+) in neural subsets of a rutabaga mutant. It was found that AC-Rut(+) has to be expressed in the mushroom bodies' Kenyon cells (KCs), specifically in the γ-KCs subset. Thus, this form of discriminative social learning requires the same KCs as non-social Pavlovian learning, suggesting that pathways of social and asocial learning overlap significantly (Nobel, 2023).
Previous studies showed that young fruit flies use social information to choose a mate and develop a preference for male phenotypes that they previously saw being chosen by demonstrator females, i.e., perform mate copying. In other words, they copy the mate choice of conspecifics. Mate copying occurs when, after observing another females' mate choice, an observer female tends to preferentially mate with the same male ("individual based" mate copying) or with males of the same phenotype ('trait-based' mate copying) as the one chosen during the demonstration.
In fruit flies, mate-copying experiments involve a demonstration during which a virgin, naive observer female can watch another female copulating with a male of a given phenotype while a male of a contrasting phenotype stands by, followed by a mate-choice test in which the observer female can mate with one of the two male phenotypes. Mate copying in Drosophila is quite sophisticated and has the potential to lead to long-lasting traditions of preferring a certain male phenotype. Although the behavioral patterns are well described, the underlying neurobiological mechanisms are unknown (Nobel, 2023).
This study showed that the adenylyl cyclase AC-Rut+ protein is involved in mate copying. Expressing AC-Rut+ in the Central Complex did not rescue mate copying, suggesting that the AC-rut+ in the CC is not necessary for that behavioral pattern. This is in contrast to previous studies showing that the CC plays a role in operant visual learning. Contrastingly, this study found that the γ-KCs of the MBs are necessary and sufficient for mate copying, since re-establishing the expression of AC-Rut+ in the γ-KCs fully rescues mate copying in rut observer females in the different contexts in which it was tested. This suggests that mate copying shares some mechanisms with classical associative non-social learning. Furthermore, the fact that expressing AC-Rut+ only at the adult stage rescues the full behavioral pattern rules out any developmental issue putatively due to the rut mutation.
Interestingly, AC-Rut+ appears as a key protein required in several associative non-social learning paradigm such as classical learning or operant learning. Imaging technique showed previously that AC-Rut+ acts as a coincidence detector in non-social contexts, as MBs AC-Rut+ is activated more strongly when two neurotransmitters conveying information of unconditional and conditional stimuli are both applied simultaneously to a preparation of fly than when the two neurotransmitters are applied independently (Nobel, 2023).
The fact that AC-Rut+ is required also in this form of social learning strongly suggests the existence of tight links between social and non-social associative learning. In mate copying, the male color can be considered as the CS and the copulation of the demonstrators as the US.
This, thus, closely recalls the classical conditioning in olfactory learning in which the odor is the CS, and the electric shocks or the sugar the US, and in which the expression of AC-Rut+ is needed in the same γ-KCs of the MB. Furthermore, γ-KCs output are required also in non-social associative visual learning. Their similar roles in olfactory and visual learning,
as well as in mate copying (this study), and reacting to courtship conditioning show that both, visual and olfactory cues of social or non-social origin elicit the functionality of MB γ-neurons. Thus, these neurons appear to constitute a hub in the neuronal pathways of a large series of types of Drosophila associative learning (Nobel, 2023).
Inhibiting the expression of a gene like rutabaga and restoring its expression in a few neurons in a mutant context is a powerful way to show its involvement in any function, especially as the nature of Rutabaga (usually considered as a coincidence detector) strongly supports these findings. Altogether, these three independent experiments show that γ-KCs are necessary for mate copying and that this pathway involves the Rutabaga protein. Although the first statistical test only reveals a trend for γ-KCs, the fact that highly significant results were found supporting that trend in two independent experiments replicating the same kind of test in different contexts (photo demos and temperature-dependent expression) leads to the conclusion that the lack of significance in the first test was probably due to a lack of power because groups that did not copy were slightly, but non-significantly, above 0.5. Remarkably, binomial tests of the individual treatments all support the current interpretation. In sum, since this study found in three independent experiments (real demos, photo demos, and temperature-dependent expression) evidence that the γ-KCs are required for mate copying, the conclusions can be trusted. Finally, the fact that photos are efficient in triggering social learning involving the same mechanistic pathways opens the way to further studies, like calcium imaging, to further decipher the neurobiology of mate copying. This study opens a new avenue of research to unravel the full pathways of social learning, either upstream or downstream of the γ-KCs (Nobel, 2023).
At the Drosophila NMJ model glutamatergic synapse, presynaptic Vav-Rac1 signaling triggers actin polymerization, driving BMP-induced presynaptic macropinocytosis and activity-induced RP vesicle mobilization, two spatiotemporally distinct membrane trafficking mechanisms regulating synaptic growth and functional potentiation, respectively. The present study provides genetic evidence that the small GTPase Rap1 operates upstream of Vav-Rac1 signaling in both of these two actin-dependent processes. Our findings indicate that Rap1 is a key regulatory node for the presynaptic actin dynamics that independently controls Gbb-induced macropinocytosis and tetanus-induced RP mobilization. This study also found that BMP-induced activation of Abelson (Abl) kinase is coupled to the induction of presynaptic macropinocytosis via the Gef26-Rap1 pathway during synaptic growth. This study also found that Rut-dependent cAMP signaling plays a direct obligatory role in tetanus-induced RP mobilization and PTP, and that this action of cAMP signaling is mediated via the Epac-Rap1 pathway. Thus, this work demonstrates two distinct signaling mechanisms that independently control Rap1/Vav/Rac1-dependent actin cytoskeleton remodeling regulating structural and functional presynaptic plasticity.
Macropinocytosis is initiated by Rac1-SCAR/WAVE signaling membrane ruffles that subsequently develop into macropinocytic cups and intracellular macropinosomeset. Abl kinase and Vav play central roles in this actin-dependent process. In mammalian cells, Abl phosphorylation of SCAR2/3 promotes membrane ruffling. At the Drosophila NMJ, Abl phosphorylation of SCAR complex Abi drives presynaptic macropinocytosis induced by BMP ligand Gbb, and the Rho GEF Vav acts upstream of Rac1 in this process. In this study, multiple lines of evidence is provided that Vav is a major Abl target during Gbb-induced, Rac1-mediated presynaptic macropinocytosis. First, induced Abl kinase activity from a constitutively active variant or DPH application alone is sufficient to induce presynaptic macropinocytosis even in the absence of Gbb signaling. Second, full activation of Abl kinase activity almost completely occludes the action of Gbb, further supporting a role for Abl as the key mediator of Gbb-induced macropinocytosis. Third, loss of Gef26 or its downstream effector Rap1 results in impairments of both Gbb-induced and DPH-induced macropinocytosis. Finally, consistent with a critical role for presynaptic macropinocytosis in restraining BMP signaling and presynaptic growth, genetic interaction tests suggest that the Gef26-Rap1 pathway connects Abl kinase activation with Vav-Rac1 signaling in the restraint of presynaptic growth. Together, these results indicate that Abl activates Vav-Rac1-SCAR signaling via the Gef26-Rap1 pathway to drive Gbb-induced presynaptic macropinocytosis, thereby restraining synaptic growth. However, the molecular mechanism underlying the Abl activation of Gef26 has not yet been determined (Kim, 2024).
Despite synaptic overgrowth, the basal EJP and spontaneous mEJP amplitudes in a physiological condition (1.5 mM Ca2+) are not significantly altered in Rap1 mutants compared with controls. However, the frequency of mEJPs is significantly increased in Rap1 mutants. Interestingly, a selective increase in mEJP frequency, but not EJP and mEJP amplitudes, is also observed in Abl and Vav mutants, as well as in animals depleted of presynaptic CtBP and Rabankyrin (unpublished data), two key regulators of macropinocytosis. Thus, it is highly plausible to propose that genetic conditions impairing presynaptic BMPR macropinocytosis commonly cause synaptic overgrowth, thereby leading to parallel increases of functional release sites and mEJP frequency (Kim, 2024).
In many model synapses, the presynaptic elevation of cAMP has been found to facilitate neurotransmitter release for sustained periods, leading to short- or long-term increases in neurotransmission strength. Studies of mammalian cerebral and cerebellar synapses have suggested that PKA is the major target of this cAMP action. More recent studies, however, have shown that the effects of cAMP on transmitter release are independent of PKA. For example, Epac2 has been found to partly mediate cAMP-induced potentiation at the mossy fiber-CA3 synapse of the mouse hippocampus. In addition, Ih channels and Epac, but not PKA, have been found to contribute to cAMP-induced potentiation at the crayfish NMJ (Beaumont and Zucker, 2000; Zhong and Zucker, 2005), and Epac activation is shown to fully mediate cAMP-induced potentiation at the calyx of Held in the mammalian brain stem (Kaneko and Takahashi, 2004). Thus, the role of cAMP in the regulation of presynaptic transmitter release appear largely independent of PKA. This study provides evidence that Epac is the major target of cAMP-induced potentiation of synaptic transmission at the Drosophila NMJ. This study also shows that the potentiation of EJP amplitudes by the adenylyl cyclase activator FSK is completely blocked by the loss of Epac but not by PKA and Ih channel activity. This study also showed that this modulation is fully mimicked by an Epac-specific cAMP analog (Kim, 2024).
Our findings further suggest that this FSK/cAMP-induced potentiation and PTP appear to share a common mechanism. Both potentiation processes display similar decay time constants and are associated with increases in the synaptic vesicle Pr, readily releasable pool (RRP) size, and replenishment rate. Moreover, as previously demonstrated for PTP, this study found that FSK-induced potentiation is completely blocked by the MLCK inhibitor ML-7, which interferes with the mobilization of RP SVs. Importantly, the prior application of FSK completely occludes the manifestation of PTP. These findings, together with results showing that genetic ablation or pharmacological inhibition of the Rut adenylate cyclase as well as Epac disrupts PTP, indicate that activation of Rut-cAMP-Epac signaling plays a direct, obligatory role in this functional potentiation at the Drosophila NMJ. In comparison with these findings, cAMP-PKA signaling has been implicated in a presynaptic tetanic stimulation-induced long-term potentiation (LTP) at mammalian hippocampal synapses. These functional time course comparisons across species are compelling. However, the mechanisms by which cAMP signaling may play differential roles in short-term versus long-term synaptic potentiation at different synapses remain unclear (Kim, 2024).
Vav acts via the actin-regulatory Rac1-SCAR pathway to mediate tetanus-induced RP mobilization and PTP. Consistent with previous results showing that Epac is a direct activator of Rap1 , our new findings indicate that the Rut-cAMP-Epac pathway signals through Rap1 to mediate Vav/Rac1-dependent RP mobilization and PTP. The loss of Rap1 was found to impairs tetanus-induced RP mobilization and PTP, mimicking the rut, Epac, Vav, and Rac1 loss-of-function phenotypes. Moreover, dosage-sensitive transheterozygous interactions between rut and Epac, Rap1, or Vav during PTP expression, strongly indicate that Epac, Rap1, and Vav work together in a common, Rut-dependent cAMP pathway. Furthermore, Nuronal overexpression of constitutively active Rap1 and Vav fully mimics the effects of FSK/8-pCPT on neurotransmission, with FSK/8-pCPT-induced potentiation being almost completely abolished in Rap1 and Vav mutants. These results strongly indicate that Rap1-Vav signaling acts downstream of Epac to facilitate transmitter release. Finally, as previously demonstrated for both Vav and Rac1 mutants the RP mobilization and PTP defects in Epac and Rap1 mutants are completely rescued by stabilizing filamentous actin (F-actin) with jasplakinolide (Jasp). Taken together, this work indicates that Rut-cAMP-Epac signaling acts through the actin-regulatory Rap1-Vav pathway to mediate tetanus-induced RP mobilization and PTP (Kim, 2024).
Our data indicate that Abl-Gef26 signaling does not contribute to functional presynaptic plasticity. Both tetanus-induced RP mobilization and PTP are not affected by acute pharmacological blockade of Abl kinase activity and in Gef26 mutantsThis study also found that loss of Abl or Gef26 function does not impair FSK/8-pCPT-induced synaptic potentiation. Moreover, in contrast to FSK/8-pCPT, the Abl activator DPH fails to potentiate synaptic transmission. On the other hand, our data indicate that Rut-cAMP-Epac signaling plays no role in regulating structural presynaptic plasticity. x Gbb-induced presynaptic macropinocytosis is not impaired in rut1 mutants.This study also found that, in contrast to DPH, FSK fails to induce presynaptic macropinocytosis in the absence of Gbb. Finally, Epac mutants show normal levels of Gbb-induced presynaptic macropinocytosis and do not interact with the Abl-Gef26-Rap1 pathway during synaptic growth. Thus, this work demonstrates the differential roles of the Abl/Gef26- and Rut/cAMP/Epac-mediated Rap1 pathways at the Drosophila NMJ. Consistent with this conclusion, enrichment was found of Rap1 expression at the periactive zone, where endocytosis occurs, as well as in CSP-associated SV compartments. Given that mammalian Rap1 signals long-term depression via lysosomal p38MAPK, it is of paramount importance to define the detailed mechanisms by which Rap1 achieves signal diversity and specificity to mediate different forms of synaptic plasticity (Kim, 2024).
How might Rut-cAMP-Epac signaling mediate PTP at the cellular level? The strong correlation between tetanus-induced RP mobilization and PTP, coupled with the essential roles of Vav-Rac1 signaling and F-actin cytoskeleton networks in RP mobilization, implies that Epac-dependent cAMP signaling acts by promoting Rap1/Vav/Rac1-dependent actin polymerization and thus SV mobilization via actin-based molecular motors. This scenario is in general agreement with previous reports showing that RP mobilization and PTP depend on the activity of MLCK, a major activator of myosin motor activity. It remains unclear, however, whether RP mobilization per se is sufficient to increase neurotransmitter release in the absence of tetanic stimulation or whether RP mobilization simply confers competence on synapses to manifest PTP. In the latter case, Epac and Rap1/Vav/Rac1 signaling may directly act on the components of the presynaptic release machinery to increase RRP size and/or vesicular Pr, two presynaptic parameters that ultimately determine the amount of transmitter release. In mammals, the Epac regulatory role in exocytosis is best characterized in studies of insulin secretion by pancreatic β-cells. Epac mediates the cAMP-induced potentiation of insulin secretion via interaction with Rab3-interacting molecule 2 (RIM2), a protein family also involved in regulating synaptic vesicle exocytosis. Thus, it will be interesting to determine whether, in addition to mobilizing RP vesicles, Epac/Rap1 signaling acts on the presynaptic release machinery to mediate PTP. In conclusion, this study demonstrates that Abl-Gef26 and Rut-cAMP-Epac pathways converge on actin-regulatory Rap1-Vav signaling to mediate presynaptic plasticity (Kim, 2024).
Exons - 16
Four putative adenylate cyclase genes from Drosophila melanogaster have been identified by virtue of their extensive sequence homology with mammalian cyclases. One is coded for by the learning and memory gene rutabaga and is most similar to the mammalian brain Ca2+/calmodulin (CaM)-responsive cyclase. The hydrophobicity profile of the N-terminal half is similar to that of the mammalian Type I enzyme and suggests a similar arrangement of twelve membrane-spanning segments. There are two potential N-linked glycosylation sites in the predicted extracellular loop between membrane spans 9 and 10 (Levin, 1992).
date revised: 25 March 2026
Home page: The Interactive Fly © 2026 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
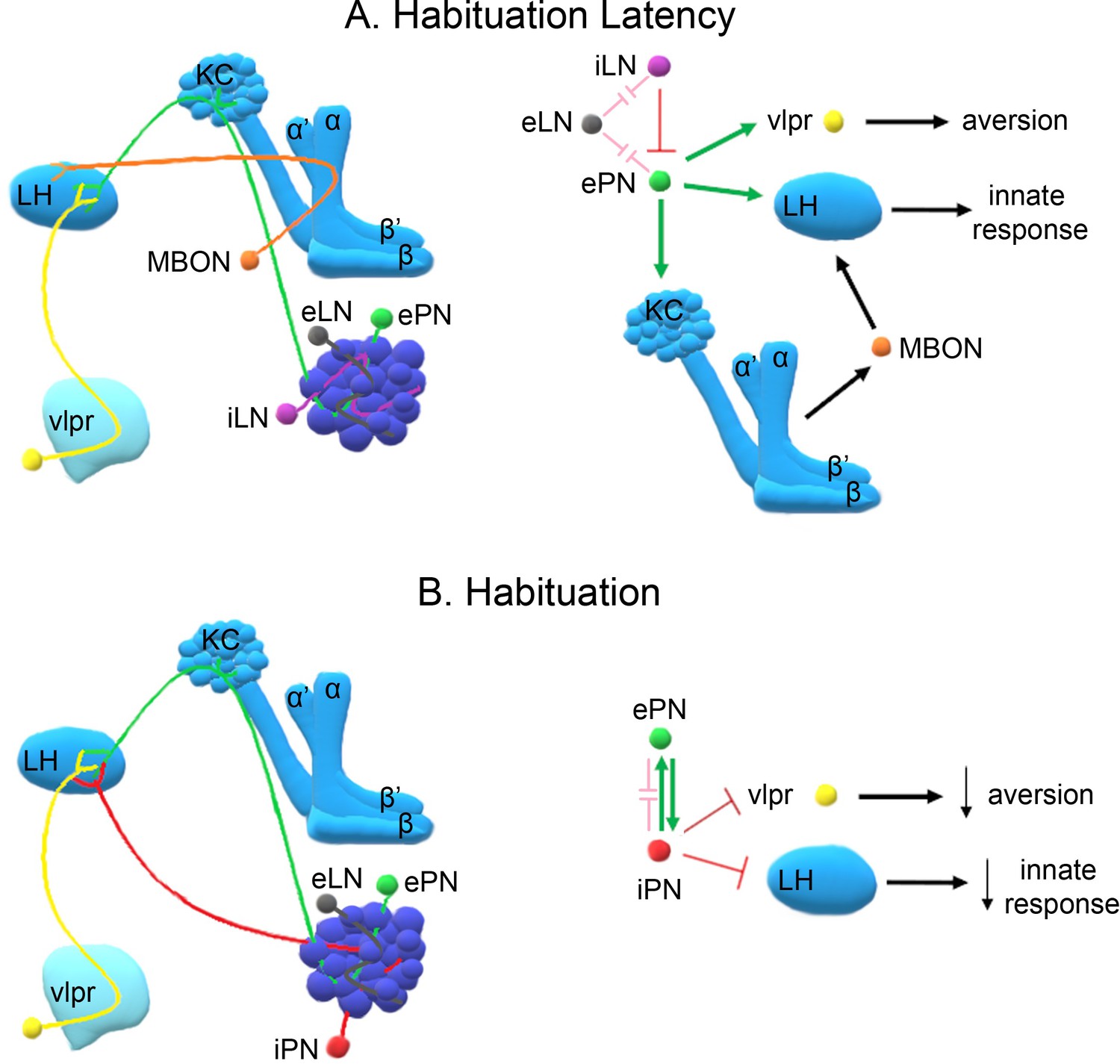{kind=link}