The Interactive Fly
Zygotically transcribed genes
The skeleton of the cell can be thought of as a maze of tubes and ropes. The tubes are composed of the protein tubulin, the basis of the microtubular cytoskeleton, and the ropes are actin, a component of muscle that forms the actin based cytoskeleton. If the tubulin and actin "ropes" are like the rigging of a ship, then the centrioles are analogous to the mast of the ship, providing a central organizing element for the microtubular filaments. Additional elements of the cytoskeleton are present immediately under the cell membrane, and serve a supportive function, like the ribs of a ship, to stretch the analogy.
Cytoskeleton is an important aspect of cell motility, assuring that a motile cell has a front and back as it moves along a substratum. Dynamic changes in cytoskeleton, in this case the actin based cytoskeleton, take place during differentiation processes. Dorsal closure is one example of a developmental process involving cell motility and the actin based cytoskeleton. Hemipterous is involved in a signaling process that affects cell motility in dorsal closure. The process of dorsal closure is described at the Hemipterous site.
The region immediately beneath the cell membrane is known as the cortex. Here a completely different set of cytoskeletal elements establish and maintain cell polarity, help to maintain cell shape, and serve to anchor proteins embedded in the cell surface. Thus the cortex has a major role in cell-cell communication mediated by cell surface proteins. The cortex serves as an anchor for Oskar, a key protein in establishing oocyte polarity, and also anchors Prospero and Numb, proteins important in neural cell polarity.
Microtubules (MTs), built of tubulin, are the highways on which dynein and kinesin motors travel. Microtubules are hollow, cylindrical polymers of alpha and beta tubulin heterodimers. During polymerization, the dimers assemble head-to-tail into typically 13 protofilaments arranged parallel to the long axis of the MT. The asymmetry of the individual dimers imparts an intrinsic polarity to the MT, which is displayed as kinetic differences between the two ends. The plus end, originally defined as the end of the axon's microtubule distal to the cell body, elongates 2-3 times faster than the minus end, or the end proximal to the cell body. Dependent on cell type, MTs may be arrayed in a variety of configurations, and since motors are unidirectional, the arrangements of MTs dictates the effective direction of motor movement. Kinesins are plus end directed motors, while dyneins are minus end directed motors. In mitosis, the minus end of microtubules are associated with centrosomes. Gamma Tubulin is involved in the nucleation of mitotic microtubules. Two antiparallel, overlapping MT arrays are generated with the plus ends of each interdigitating in the overlap zone (or in some cases interacting with chromosomal kinetochores) (Walker, 1993).
One of the major functions of cytoskeleton carried out by microtubules during mitosis is the equitable distribution of chromosomes to the two poles of the cell. The cytoskeleton is also responsible for structuring the cytoplasm of the cell so that proteins and nucleic acids can be carried from one site to another. The polar distribution of Bicoid mRNA and Oskar mRNA in the oocyte is accomplished through employment of the oocyte microtubular cytoskeleton.
Cytoplasmic streaming, another function of the microtubular based cytoskeleton, takes place in egg chambers during stage 12 and stage 13. cappuccino and spire are required to repress this microtubule-based ooplasmic streaming in the oocyte and to ensure the proper partitioning of molecular determinants within the oocyte. In mutants, the bundling of the microtubules at the cortex of the oocyte and the streaming of the oocyte cytoplasm occurs prematurely, by stage 8 in oogenesis. It is thought that this movement within the oocyte is necessary to mix the oocyte cytoplasm with the cytoplasm being rapidly added from the nurse cells, by an actin based cytoskeleton mechanism (Theurkauf, 1994). It is likely that subcortical nurse-cell microfilaments play a role in cytoplasmic flow into the oocyte. A nonmuscle myosin is found associated with subcortical actin but not with cytoplasmic networks. These subcortical actin filaments are very sensitve to cytochalasin treatment. Contraction of the subcortical actin could play a role in the bulk movement of nurse cells into the oocyte (Cooley, 1992 and references).
Prior to late vitellogenesis, characterized by the bulk flow of cytoplasm into the oocyte, during previtellogenesis, a different type of intercellular transport occurs. During this phase, treatment with colchicine, which inhibits microtubular transport mechanisms, does not change transport processes through the ring canals. However, when the microfilament inhibitor cytochalasin B is applied, the transfer of particles through the ring canals is completely inhibited. When an inhibitor of myosin-driven motility is applied to follicles, all movements within the cytoplasm stop. It is therefore thought that cytoplasmic myosin and the actin based microfilament network play a decisive role in particle movements during previtellogenesis (Bohrmann, 1994).
The actin based microfilament cytoskeleton plays an additional role in a process known as cytoplasmic dumping. At stage 11, the nurse cells dump their contents into the oocyte through cytoplasmic bridges termed ring canals. Microfilament bundles form in the nurse cells during this process and are apparently required to hold the nurse cell nuclei in place so that they do not obstruct the ring canals and allow rapid flow of nurse cell cytoplasm into the oocyte. Mutants in chickadee, quail and singed affect actin bundle formation. Profilin, encoded by chickadee, is presumably required for the polymerization of the actin filaments that compose the bundles (Cooley, 1992), while a villin-related protein encoded by quail and a fascin-related protein encoded by singed are thought to be required to cross-link the actin filaments to form the bundles. Two components of the actin-lined ring canals have also been identified - an adducin-like protein encoded by hu-li tai shao and a protein containing scruin repeats encoded by kelch (Manseau, 1996 and references).
It appears that there is an interaction between the actin and tubulin based components of cytoskeleton. Profilin, encoded by chickadee, a component of the actin based cytoskeleton, physically interacts with Cappuccino, involved in the microtubule based cytoskeleton. Mutants in chickadee resemble cappuccino in that they fail to localize Staufen protein and Oskar mRNA in the posterior pole of the developing oocyte. A strong allele of cappuccino has multinucleate nurse cells, similar to those described for chickadee (Manseau, 1996).
Cell migration occurs through the protrusion of the actin-enriched lamella. The effects of RNAi depletion of approximately 90 proteins implicated in actin function on lamella formation have been investigated in Drosophila S2 cells. Similar to in vitro reconstitution studies of actin-based Listeria movement, it has been found that lamellae formation requires a relatively small set of proteins that participate in actin nucleation (Arp2/3 and SCAR), barbed end capping (capping protein), filament depolymerization (cofilin and Aip1), and actin monomer binding (profilin and cyclase-associated protein). Lamellae are initiated by parallel and partially redundant signaling pathways involving Rac GTPases and the adaptor protein Nck, which stimulate SCAR, an Arp2/3 activator. RNAi of three proteins (kette, Abi, and Sra-1) known to copurify with and inhibit SCAR in vitro leads to SCAR degradation, revealing a novel function of this protein complex in SCAR stability. These results have identified an essential set of proteins involved in actin dynamics during lamella formation in Drosophila S2 cells (Rogers, 2003).
Under routine culture conditions, S2 cells display a roughly spherical morphology with a diameter of ~10 µm. These cells are not motile and exhibit no obvious morphological polarity, but time-lapse microscopy of cells expressing GFP-actin reveals that their surfaces are dynamic and continuously extend and absorb membrane ruffles. S2 cells may be induced to undergo a dramatic change in their morphology when plated on glass coverslips coated with the lectin concanavalin A (con A). Within 20 to 30 min after plating on this substrate, these cells avidly attach, flatten, and spread to adopt a discoid morphology of approximately double their normal diameter (20 µm). Spread cells resemble a 'fried egg' with a domed central region containing the nuclei and majority of organelles surrounded by a thin, organelle-free zone (Rogers, 2003).
To better understand the organization of actin in S2 cells, con A-adhered S2 cells expressing GFP-actin were fixed and stained with Texas red X-phalloidin, a probe that selectively binds to filamentous actin. When examined by fluorescence microscopy, most S2 cells (90%) exhibited a highly developed, radially symmetrical actin cytoskeleton that could be divided into three zones: a dense peripheral network at the extreme periphery of the cells (~1 µm wide), a second central zone (4-6 µm wide) of lower actin density composed of filaments, and a third circular bundle of filaments that surrounded the nucleus. Arp3, cofilin, and capping protein were enriched in this first actin-dense zone at the leading edge, especially at membrane ruffles. Enabled/VASP was further restricted to the extreme edge of the periphery (<1 µm). In contrast, immunolocalization of profilin/chickadee revealed puncta that were distributed throughout the cell and particularly abundant in the inner nuclear and organelle-rich domain. These puncta were not associated with adhesion structures; immunofluorescent staining against phosphotyrosine failed to stain the ventral surface of the cells. The distributions of these well-characterized actin-binding proteins are generally similar to those described in other cell types that form actin-rich lamellae (Rogers, 2003).
A small proportion (<10%) of cells did not exhibit such well-spread lamella but rather possessed numerous and dynamic filopodia evenly spaced around their circumference. These short (1-2 µm) projections exhibited cycles of elongation and retraction. Interconversion of the two cell morphologies has not been observed. RNAi studies were restricted to the predominant population of cells that spread and form lamella on the con A-coated surfaces (Rogers, 2003).
Also, actin dynamics were directly visualized in the lamellae of living S2 cells expressing GFP-actin after plating on con A. Membrane ruffles formed at the cell periphery, folded back toward the cell center, and ultimately fused with the dorsal surface of the cell. Such ruffling activity was more or less symmetrically distributed around the cell, and polarized morphologies or cell movement was rarely observed. At sufficiently low levels of protein induction, a speckled pattern of GFP-actin was observed, and time-lapse imaging revealed a centripetal flow of actin from the periphery toward the center of the cell at a rate of ~4.0 ± 0.44 µm/min, which is somewhat faster than described in other systems, such as migrating fibroblasts or neuronal growth cones. In summary, imaging of actin and actin-binding proteins indicates that con A-induced spreading of S2 cells constitutes an attractive model system for understanding the molecular basis of lamella formation (Rogers, 2003).
To dissect the molecular basis of lamella formation, the susceptibility of S2 cells to RNAi was exploited to identify proteins involved in this process. A candidate list of ~90 proteins implicated in aspects of actin function or in cell motility during neuronal development and dorsal closure during Drosophila embryogenesis was compiled (Rogers, 2003).
DNA microarray analysis demonstrated that only five genes in this list are not expressed above background levels in S2 cells. Since very low expressing genes nevertheless may be important for cell function, these genes were still subjected to RNAi analysis. A 7-day RNAi treatment was used to deplete proteins before assaying the cells for lamella formation on con A-treated coverslips. Filamentous actin was visualized with rhodamine-phalloidin, and DNA was stained with DAPI to screen for multiple nuclei reflecting cytokinesis defects. For every treatment, at least 500 cells were examined. The efficacy of the RNAi treatments was verified by immunoblotting extracts from dsRNA-treated cells using a panel of antibodies to 13 proteins. Immunoblotting for those tested revealed that RNAi reduced protein expression by at least 90% of endogenous levels and in many cases was not detectable. This immunoblot analysis included five proteins for which RNAi did not elicit an obvious phenotype. Greater than 90% reduction in the levels of 10 motor proteins subjected to RNAi was achieved and no case was encountered where RNAi had failed to reduce protein levels. It is, therefore, speculated that dsRNAs against proteins that could not be quantitated most likely produced a similar degree of inhibition (Rogers, 2003).
Of the ~90 genes tested, RNAi produced obvious aberrant morphologies in 19 cases. The observed defects can be categorized into seven phenotypic classes that will be described below (Rogers, 2003).
The Arp2/3 complex was inactivated by targeting its crucial p20 subunit (Arc-p20 ), which mediates protein-protein interactions within the Arp 2/3 complex and, therefore, is essential for stability and actin-nucleating activity. After p20 RNAi, >90% of S2 cells exhibited a striking morphological defect when plated on con A. Instead of the circular, symmetrical shape usually induced on this substrate, p20-depleted cells adopted a stellate, radially asymmetrical cell morphology. Phalloidin staining revealed that these cells rarely formed lamellae; instead filamentous actin was enriched in the distal tips of a variable number of tapered projections. The presence of actin filaments could be due to residual Arp2/3 or to alternative actin-nucleating activities. In addition, actin filaments were sometimes observed to run radially from the center of the cell body along the lengths of these projections. These processes were also enriched in microtubules that often extended to their distal regions. The frequency of multinucleate cells was approximately the same as control cells, indicating that inhibition of Arp2/3 does not affect cytokinesis (Rogers, 2003).
Cells contain actin nucleation-promoting factors that activate the Arp2/3 complex. Genetic analysis in Drosophila has shown that one of these factors, SCAR, is essential for numerous actin-based processes during development, while WASP, another activator, mediates a subset of Arp2/3 functions in neuronal cell fate determination. WASP RNAi did not alter cell morphology or actin organization in S2 cells. In contrast, RNAi against SCAR exactly duplicated the morphological defects observed with RNAi of the p20 subunit of Arp2/3 in >80% of the cells. Interestingly, RNAi for three proteins (Kette, Sra-1, and Abi) that were recently identified to copurify with SCAR produced a phenotype indistinguishable from SCAR or p20. Thus, it is concluded that lamella formation in S2 cells is a SCAR-Arp2/3-dependent process (Rogers, 2003).
The second category of RNAi-induced morphological defect was typified by inhibition of profilin (Chickadee), an actin monomer-binding protein. After this treatment, >85% of cells failed to spread on con A and instead retained their spherical shape. Phalloidin staining was diffuse throughout these cells, however, individual filaments could not be resolved. These cells also were defective in cytokinesis, as revealed by the high incidence of multiple nuclei (39%). A similar morphology also was generated by RNAi against cyclase-associated protein (CAP/Act up), another monomeric actin-binding protein that plays an important role in actin organization in Drosophila. When bound to monomeric actin, profilin acts to restrict actin incorporation to the barbed-end of actin filaments and mediates exchange of ADP for ATP. It is speculated that the accumulation of f-actin in profilin and CAP RNAi cells, along with the failure to form lamellae, may reflect nonproductive polymerization of actin filaments from both the barbed and pointed ends (Rogers, 2003).
The actin-binding protein cofilin/Twinstar is essential for actin-based functions in many cell types, and in vitro and in vivo studies indicate a role for cofilin in actin filament severing and turnover. Inhibition of cofilin by RNAi prevented S2 cell spreading on con A in >95% of treated cells. These cells retained their spherical morphology, and phalloidin staining revealed a dramatic cortical accumulation of filamentous actin as well as a wrinkled "raisin-like" texture to the surface of the cell. The abnormal accumulation of filamentous actin within the cells suggests that actin turnover is inhibited in S2 cells depleted of either of these two proteins. Cofilin-inhibited S2 cells exhibited a high incidence of multinucleate cells, implicating a role in cytokinesis. This morphology and actin distribution was mimicked by RNAi inhibition of Aip1, a protein that acts cooperatively with cofilin in disassembling actin in Xenopus and budding yeast. Aip1 also produced a cytokinesis defect. These results indicate that both cofilin and Aip1 are essential for actin remodeling during lamella formation and that, despite the similarities in cell morphology produced by RNAi against either of them, these two proteins have distinct roles in actin regulation (Rogers, 2003).
Slingshot is a protein phosphatase that activates the actin-severing activity of cofilin; loss-of-function experiments in Drosophila have demonstrated that tissues mutant for slingshot exhibit abnormal accumulations of f-actin. S2 cells treated with dsRNA to inhibit slingshot are able to attach and spread efficiently on con A. However, the lamellae in >50% of these cells exhibited structural abnormalities as compared with controls. The distribution of f-actin was uniformly dense from the cell periphery to the center of the cell and did not show the typical distal enrichment commonly observed in spread S2 cells. Cells exhibiting this morphology typically had prominent radial bundles of actin that spanned the entire width of the lamellae. It is speculated that this cellular morphology is produced by a partial loss of cofilin activity, leading to inefficient disassembly of the dendritic array of actin filaments at the rear of the lamellae and thus producing a lamellipod that is radially wider than normal. Cytokinesis defects were not observed in these cells (Rogers, 2003).
Capping protein is an important regulatory factor that binds to the barbed ends of actin filaments to prevent actin monomer addition. Recent studies have suggested that a functional antagonism between capping protein and enabled/VASP regulates the length and polymerization rate of actin filaments in the lamella. This balance controls the rate of lamella protrusion in motile cells. S2 cells treated with dsRNA against capping protein adher and spread normally, but ~80% had lamellae exhibiting a hyper-ruffled shape. Lamellae in S2 cells lacking capping protein also exhibited an accumulation of filamentous actin at the periphery that extended 2-3 µm inwards from the cell perimeter, as compared with ~1 µm in untreated cells. An explanation for the abnormal lamella morphology has been suggested. In the absence of capping protein, enabled/VASP-mediated actin filament elongation favors the formation of abnormally long filaments at the cell margin. These filaments push against the membrane, fueling protrusion, until compressive forces exceed the flexural rigidity of long filaments, causing them to buckle and the membrane to retract. This hypothesis explains the hyper-ruffled phenotype as well as the accumulation of f-actin at the margin of the cell. No accumulation of multinucleated cells was observed, suggesting that capping protein is dispensable for cytokinesis (Rogers, 2003).
A sixth category of morphological defect was produced by depletion of Cdc42 by RNAi. Cdc42, a member of the Rho family of small G proteins, regulates actin organization and is generally thought to mediate the formation of filopodia during cellular migration. Inhibition of Cdc42 prevented formation of a normal lamella in ~50% of the cells. Instead, actin was organized into long, thin processes that projected from the entire periphery of the cell. These processes did not resemble the filopodia that spontaneously form on some S2 cells or that form in response to overexpression of constitutively active Cdc42V12, because they were typically >10 µm in length and possessed a uniform diameter. This morphology is difficult to reconcile with what is known about Cdc42 functions, although a cellular null phenotype for Cdc42 in metazoan cells has not been reported (Rogers, 2003).
A seventh category was failure of cytokinesis without inhibition of cell spreading on con A-coated surfaces. Cells in this category (>95%) possessed multiple nuclei and were much larger in diameter than control cells. Phalloidin staining revealed that, despite their larger size, cells were able to form lamellae with normal architecture. Inhibition of Rho1 and its downstream effectors citron kinase, diaphanous, AcGAP, and myosin II typified this defect. Many of these molecules were recently identified in a similar S2-based RNAi screen for genes specifically involved in cytokinesis, but Aip1, CAP, citron kinase, and diaphanous were not tested in this study (Rogers, 2003).
In addition to producing cytokinesis defects, however, cells depleted of cytoplasmic myosin II sometimes failed to form normal lamellae, in addition to producing cytokinesis defects. These cells contained abundant filamentous actin, as judged by phalloidin staining, but the actin cytoskeleton displayed an overall lack of organization with filaments criss-crossing the width of the cell in an apparently random manner. These results reveal a role for myosin II in the organization of actin in the lamellae (Rogers, 2003).
The SCAR-associated proteins kette, Sra-1, and Abi prevent degradation of SCAR: Native SCAR exists in a trans-inhibited state in a complex with the Kette, Sra-1, and Abi proteins. Given the demonstrated role of these proteins in suppressing SCAR activity in vitro, it was surprising that RNAi-mediated depletion of Sra-1, Abi, or Kette resulted in a SCAR-like phenotype rather than in excessive actin polymerization. One hypothesis to account for these observations was that SCAR is either not localized at the membrane or degraded in the absence of members of the kette-Sra-1-Abi complex. To test these ideas, Kette, Abi, or Sra-1 RNAi-treated cells were stained with anti-SCAR antibodies and the overall staining intensities were observed to be reduced or eliminated. Quantitative immunoblotting was performed and it was found that Kette, Sra-1, and Abi RNAi treatments caused a considerable reduction of SCAR levels in S2 cells. Depletion of Abi, Kette, and Sra-1 reduced SCAR protein levels to 34.3 ± 18, 17.3 ± 9.5, and 9.6 ± 2.6%, respectively. In contrast, cells treated with dsRNA versus Diaphanous did not show reduced SCAR levels. From these observations, it is concluded that the kette-Sra-1-Abi complex is required for SCAR stability (Rogers, 2003).
The small G proteins Rac1/2 and Mtl and the adaptor protein Nck mediate cell spreading and lamella formation via two independent pathways:
Activation of SCAR proteins is generally thought to be mediated by Rac GTPases. However, RNAi of Drosophila Rac 1, Rac 2, and the Rac-like protein Mtl did not prevent cell spreading or lamella formation. Genetic evidence has demonstrated that these small G proteins are functionally redundant in many tissues in the fly. Furthermore, in vitro experiments show that the inhibitory SCAR complex can be activated either by Rac1 or the SH2-SH3 adaptor protein Nck. To test whether this is the case in S2 cells, cells were treated with dsRNA designed to simultaneously inhibit Rac1 and Rac2 (Rac1/2) and Mtl for 7 d. Unexpectedly, phalloidin staining revealed that these dsRNA-treated cells spread and formed a normal lamella when plated on con A (Rogers, 2003).
Next, the in vitro finding that either Rac or Nck is able to activate SCAR was tested by simultaneously inhibiting various combinations of Rac1/2, Mtl, and the Drosophila orthologue of Nck (Dreadlocks). This treatment produced three different cell morphologies: cells with normal lamellae, cells that spread but exhibited an abnormal serrated edge, and cells exhibiting the stellate morphology observed after RNAi of Arp2/3 and SCAR. The serrated cell shape likely represents an intermediate morphology caused by incomplete inhibition of the signaling pathway. In control RNAi-treated cells, >95% of the cells formed normal lamellae with <5% of the cells exhibiting a serrated cell margin. Stellate cells were never observed in control cultures. Inhibition of Nck alone by RNAi caused a reduction in the number of S2 cells with normal lamellae to ~65% and an increase in serrate cells to ~30% and stellate cells to 5%. Double RNAi treatments to inhibit Nck and Rac1/2 or Nck and Mtl produced moderate increases in the number of serrate cells compared with Nck alone. However, simultaneous application of dsRNAs against Nck, Rac1/2, and Mtl induced a dramatic increase in serrate and stellate cells to ~30% and ~20%, respectively. These observations suggest that the Rac-like proteins and Nck are partially redundant for lamella formation in S2 cells (Rogers, 2003).
Lamellipodia are dynamic actin-rich cellular extensions that drive advancement of the leading edge during cell migration. Lamellipodia undergo periodic extension and retraction cycles, but the molecular mechanisms underlying these dynamics and their role in cell migration have remained obscure. This study shows that glia-maturation factor (GMF), which is an Arp2/3 (see Arpc1) complex inhibitor and actin filament debranching factor, regulates lamellipodial protrusion dynamics in living cells. In cultured S2R+ cells, GMF silencing resulted in an increase in the width of lamellipodial actin filament arrays. Importantly, live-cell imaging of mutant Drosophila egg chambers revealed that the dynamics of actin-rich protrusions in migrating border cells is diminished in the absence of GMF. Consequently, velocity of border cell clusters undergoing guided migration was reduced in GMF mutant flies. Furthermore, genetic studies demonstrated that GMF cooperates with the Drosophila homolog of Aip1 (flare) in promoting disassembly of Arp2/3-nucleated actin filament networks and driving border cell migration. These data suggest that GMF functions in vivo to promote the disassembly of Arp2/3-nucleated actin filament arrays, making an important contribution to cell migration within a 3D tissue environment (Poukkula, 2014).
The transcription cofactor MAL (Myocardin-related transcription factor or Mrtf) is regulated by free actin levels and thus by actin dynamics. MAL, together with its DNA-binding partner, SRF, is required for invasive cell migration and in experimental metastasis. Although MAL/SRF has many targets, this study provides genetic evidence in both Drosophila and human cellular models that actin is the key target that must be regulated by MAL/SRF for invasive cell migration. By regulating MAL/SRF activity, actin protein feeds back on production of actin mRNA to ensure sufficient supply of actin. This constitutes a dedicated homeostatic feedback system that provides a foundation for cellular actin dynamics (Salvany, 2014).
The transcription cofactor MAL is regulated by cellular actin dynamics and confers this regulation on the activity of its DNA-binding partner, SRF. Free G-actin directly binds to MAL via RPEL motifs at the N terminus of MAL and negatively regulates its activity. This regulation is conserved from mammals to insects (Somogyi 2004). In the physiological context of the animal, the function of MAL and related proteins (MRTF-A and MRTF-B in mammals and mal-d/mrtf in Drosophila) appears conserved as well, related to active changes in the cytoskeleton. For example, initiation of invasive cell migration is essentially abolished in the absence of MAL or SRF in border cell migration in the Drosophila ovary (Somogyi 2004) or in mouse bipolar neurons exiting the subventricular zone of the brain and for cancer cells in three-dimensional (3D) invasion assays and experimental metastasis (Medjkane, 2009; Salvany, 2014).
Actin is a very abundant and exquisitely conserved protein in eukaryotic cells. Cycling of actin between G-actin and F-actin pools is controlled by a vast array of regulators, which have been the focus of considerable attention. Actin protein synthesis is also a regulated process. Actin mRNA localization and localized protein synthesis are important for cell migration and axonal growth and guidance. This study presents evidence that regulation of actin gene transcription is itself a key regulatory step in the control of invasive cell migration. Using a genome-wide approach, Actin5C was identified as a major target of the MAL/SRF transcription factor complex. Loss or reduction of MAL activity impairs invasive migration in Drosophila and human cancer cell models. It was found that restoring actin expression can be sufficient to replace the requirement for MAL to support invasive migration in these models. Thus, actin and MAL form a conserved homeostatic feedback system to ensure that actin levels are appropriate to support the actin dynamics required for complex cell behavior (Salvany, 2014).
To understand why MAL is essential for invasive migration and whether the apparent similarity of its role in different organisms reflects a conserved molecular mechanism, attempts were made to identify Drosophila MAL target genes at the genome level. A combination was used of chromatin immunoprecipitation (ChIP) and gene expression analysis and focus was placed on MAL, as SRF has functions independent of MAL/mrtf in mammals and flies. To perform analysis in the relevant tissue context, a mutant in the single Drosophila mal-d gene was used that abolishes expression in the ovary (mal-dΔ7) (Somogyi, 2004). This specifically blocked invasive migration by border cells and caused overall ovary growth defects. Ubiquitous expression of a GFP-tagged version of Mal-d completely rescued the mal-d mutant phenotypes, showing that the fusion protein provides normal Mal-d function. The MAL-GFP transgene also allowed efficient identification of MAL-GFP-bound regions in the Drosophila genome by immunoprecipitation with GFP. Key MAL-GFP-bound regions were confirmed in independent samples, and their recovery was dependent on the presence of the transgene. In parallel, genome-wide expression analysis of wild-type versus mal-d mutant ovaries identified genes whose expression was dependent on MAL. These two complementary data sets allowed genome-wide identification of MAL target genes (Salvany, 2014).
There were two key findings from this genome-wide analysis. First, only a small number of genes qualified as direct MAL targets, with MAL-GFP bound to the regulatory region and a significant decrease of mRNA expression in the mutant: the cytoplasmic Actin5C gene and five other genes, most encoding heat-shock proteins. A few additional genes encoding cytoskeletal proteins or regulators were identified as potential targets in the MAL-GFP-bound set. Second, three of the four most enriched MAL-bound regions in the whole genome were associated with Actin5C. The MAL-bound regions were conserved in other species, suggesting functional importance, and in each case, these sites bracketed gene-free upstream regions of ~10 kb. The latter is noteworthy because the Drosophila genome is dense, with most 'housekeeping genes' closely spaced, and large regulatory regions generally confined to developmental regulators. These findings focused the attention of this study on Actin5C (Salvany, 2014).
The gene expression arrays indicated a modest decrease of Actin5C mRNA levels in the mal-d mutant. Quantitative RT–PCR of carefully matched ovary samples showed a twofold decrease of mature Actin5C mRNA and a threefold to fourfold decrease of primary transcript in mal-d mutants, with no change in the closely related, but less highly expressed, Actin42A gene. In FACS-sorted migratory cells, including border cells, Actin5C levels were fourfold reduced. Analysis of the Actin5C promoter and upstream region in luciferase reporter assays showed robust promoter activity and 200-fold to 600-fold up-regulation by coexpression of SRF and activated MAL (mal-d ΔN -). Conversely, knockdown of MAL or SRF by RNAi gave 50-fold to 100-fold reduction in basal Actin5C expression. The Actin5C regulatory region also conferred responsiveness to drugs affecting actin dynamics, specifically induction by Cytochalasin D and inhibition by Latrunculin B, as observed for mammalian MAL/SRF-regulated genes. This type of reporter assay generally reveals regulatory potential at the transcriptional level. The large magnitude of regulation of the Actin5C promoter/enhancer region by MAL/SRF is consistent with the abundant binding of MAL to this region. In vivo, compensatory mechanisms may contribute to sustaining Actin5C mRNA levels upon loss of MAL activity. Thus, MAL and actin dynamics have the potential to regulate Actin5C transcription over a large dynamic range (Salvany, 2014).
The genomic data raised the possibility that Actin5C might not be just one of many cytoskeletal target genes for MAL regulation but the key target gene. If a transcription factor has one key target gene in vivo, re-expression of this gene should replace the need for the transcription factor. In genetic terms, expression of the target gene should rescue the phenotype of complete loss of function for the transcription factor in specific cells (Salvany, 2014).
To investigate this hypothesis functionally, the severe defect in invasive migration observed in mal-d mutant border cells was examined. Actin5C is the major cytoplasmic actin gene, and, as expected, mutating it perturbs border cell migration. To determine whether Actin5C was the sole required target gene of MAL, whether re-expression of Actin5C in cells that appear to be null for mal-d (mal-d S2) (Somogyi, 2004) could restore invasive migration was tested. Fly strains were used in which the Actin5C gene has a Gal4-responsive transposon, an 'EP element,' in the promoter region. Surprisingly, migration was indeed restored to normal when Actin5C was activated by Gal4 in mal-d mutant cells. Thus, as long as the Actin5C gene is induced at an adequate level, border cells do not need MAL to invade and migrate (Salvany, 2014).
In fully mutant ovaries (mal-d&Delta:7 homozygous females), restricted expression of Actin5C in terminally differentiated outer border cells using slbo-Gal4 provided significant but less efficient rescue of migration despite normal expression levels. This suggested that MAL also acts in other cells, consistent with the general oogenesis phenotype. The ORF of Actin5C in a Gal4-reponsive transgene (UAS-Actin-ORF) also provided some rescue of migration, whereas a construct with stop codons present did not. This confirmed that actin protein expression was responsible for the activity of the Actin5C locus in border cells (Salvany, 2014).
To test whether this finding extended to other tissues, another mal-d mutant phenotype, bent bristles, was examined. Bristles are 'hairs' organized by actin-rich structures and are characteristically defective in mal-d mutants (Somogyi, 2004). This effect of MAL deficiency was also restored to normal by ectopic expression of the Actin5C, accomplished by placing Actin5C cDNA under the control of a heat-shock promoter and rearing at 29°C. Thus, the requirement for MAL in these contexts reflects a specific need for MAL-driven induction of Actin5C expression. Regulation of other targets, direct or indirect, is not required. These findings indicate that the primary role of Drosophila MAL is to regulate actin levels in response to free actin fluctuations, a homeostatic feedback regulation (Salvany, 2014).
It was next asked whether this role of MAL as regulator of actin homeostasis is conserved in mammalian cells. The cytoplasmic β-actin and γ-actin genes are regulated by MAL/SRF in mammalian cells. As for Drosophila Actin5C, β-actin is essential for embryonic development and proper cell migration, whereas the related γ-actin can be compensated for. However, many other genes are more dramatically regulated by SRF and mrtfs, and some of these are important for cell migration and related functions. It is therefore assumed that MAL and SRF exert their function by regulating a battery of cytoskeletal genes. For SRF, the number of direct target genes is estimated at 200-300. For MAL and MRTFs, this is less clear, as systematic ChIP analysis is missing. Based on the findings in the fly model, the hypothesis was tested that a single target gene (β-actin) could be the key effector of mammalian MAL and that its expression could replace the need for MAL-driven gene regulation in an assay of invasive cell migration (Salvany, 2014).
The requirement for MAL is most pronounced in cases of active cell shape change and cytoskeletal challenge, such as tissue or matrix invasion (Somogyi, 2004; Medjkane, 2009; Pinheiro, 2011). Therefore, a simple cellular assay was sought that could test the ability of cells to invade into a confined environment. Migration under agarose provides mechanical resistance to movement and has been used to study migration of eukaryotic cells in a constrained space. The assay can easily be adapted to human tumor cells such as MDA-MB-231 breast cancer cells. MAL activity in these cells is provided by mrtf-a and mrtf-b, and reducing their expression severely reduces invasion in an organotypic assay and in experimental metastasis in mice (Medjkane, 2009). Simultaneous knockdown of mrtf-a and mrtf-b in MDA-MB-231 cells by siRNA reduced β-actin transcript levels. It also produced severe attenuation of migration under agarose, confirming that this assay interrogates MAL-dependent cell movement (Salvany, 2014).
To assess the importance of cytoplasmic actin downstream from MAL, stable cell lines were derived from MDA-MB-231 that allowed the ORF of human β-actin, or an N-terminally Flag-tagged version of β-actin, to be induced from the strong CMV promoter by the TET-ON system. These cell lines showed some baseline expression of the transgenes and six-fold to 10-fold induction upon Tet addition. Under agarose, migration was similar to the parental cell line and not changed by Tet addition. Knockdown of mrtf-a and mrtf-b largely attenuated migration under agarose. Remarkably, ectopic induction of β-actin or Flag-β-actin in the mrtf-a/b-depleted cells rescued migration to control levels. This indicates that human cancer cells require MAL activity to perform invasive migration for the exact same reason that Drosophila border cells do: to regulate cytoplasmic actin gene expression. Regulation of any other potential MAL target genes may not be required for this cell behavior. These experiments rely on siRNA-mediated reduction of mrtf activity; thus, it remains possible that other target genes are important but require only low levels of mrtf activity (Salvany, 2014).
Elegant experiments have shown how activity of SRF and its transcriptional cofactor, MAL, is regulated by the level of free actin and thereby by the dynamics of the actin cytoskeleton. This study has shown that the key role of MAL is to regulate the expression level of cytoplasmic actin. It is suggested that this simple feedback system allows cells to produce sufficient free actin to meet the needs of their changing cellular cytoskeletal dynamics. The extensive MAL-decorated regulatory region displayed by Actin5C may serve to tune actin gene expression sensitively and accurately. This study provides evidence that cytoplasmic actin is the sole critical target gene for invasive migration in Drosophila and possibly also in human cells. It is therefore proposed that this regulation is the ancestral function of the MAL/SRF complex in animal cells. Additional target genes for MAL have been acquired in different species and are likely to contribute to cell fitness. Examples include other cytoskeletal proteins (Medjkane, 2009) as well as genes encoding heat-shock proteins, some of which may interact with actin. While some of these genes appear more dramatically regulated by MAL when considering relative mRNA levels, actin may be the 'most regulated' gene if considering the number of transcripts induced. In any case, identification of actin as the core, essential target of MAL reveals the core of this transcription regulatory 'network' to be a simple and logical feedback system (Salvany, 2014).
Why is the MAL-driven regulation of actin particularly critical for invasively migrating cells? The stimuli inducing a robust F-actin-based cytoskeleton when initiating migration into constrained space is likely to convert free G-actin into F-actin-rich structures. Maintaining the appropriate free actin pool for further cytoskeletal buildup or for other cellular functions then requires new production of actin. Other dramatic shape changes, such as cells rounding up in the stratified epidermal layer of the skin, may induce similar sudden actin pool depletion and therefore require MAL and SRF. It has been suggested that MAL forms part of a mechanical feedback system for invasive cells (Somogyi, 2004) whereby mechanical tension induces MAL activity in order to make 'robust' cells. Related concepts of tension-driven responsiveness have recently been indicated for mammalian MAL/SRF function as well (Connelly, 2010; McGee, 2011). The mechanical feedback logic is fully compatible with the simple molecular feedback system presented here. It can be regarded as an alternate point of perturbation impinging on actin homeostasis; namely, stretching or stressing cells to provoke a biomechanical cytoskeletal response. Robust feedback systems such as this one driven by MAL are likely to be well conserved through animal evolution, when the target is a crucial one. The ability of cells to occasionally migrate and invade is a characteristic of animal systems, and the dynamic actin cytoskeleton is central to this behavior (Salvany, 2014).
Wiskott-Aldrich syndrome proteins (WASP) are nucleation promoting factors (NPF) that differentially control the Arp2/3 complex. In Drosophila, three different family members, SCAR/WAVE, WASP and WASH, have been analyzed so far. This study characterizes WHAMY, the fourth Drosophila WASP family member. whamy originated from a wasp gene duplication and underwent a sub-neofunctionalization. Unlike WASP, WHAMY specifically interacts with activated Rac1 through its two CRIB domains that are sufficient for targeting WHAMY to lamellipodial and filopodial tips. Biochemical analyses showed that WHAMY promotes exceptionally fast actin filament elongation, while it does not activate the Arp2/3 complex. Loss- and gain-of function studies revealed an important function of WHAMY in membrane protrusions and cell migration in macrophages. Genetic data further imply synergistic functions between WHAMY and WASP during morphogenesis. Double mutants are late-embryonic lethal and show severe defects in myoblast fusion. Trans-heterozygous mutant animals show strongly increased defects in sensory cell fate specification. Thus, WHAMY is a novel actin polymerase with an initial partitioning of ancestral WASP functions in development and subsequent acquisition of a new function in cell motility during evolution (Brinkmann, 2015).
The actin cytoskeleton plays a central role in a number of different cellular functions, such as cell shape changes, cell motility and membrane trafficking. Members of the Wiskott–Aldrich syndrome protein (WASP) family are conserved nucleation-promoting factors (NPF) that activate the Arp2/3 complex, a major actin nucleator in eukaryotic cells. In mammals, the WASP protein family consists of eight different members: the two Wiskott-Aldrich syndrome proteins WASP and N-WASP (also known as WAS and WASL, respectively), the related WASP family Verprolin homologous proteins WAVE1–WAVE3 (also known as SCAR1–SCAR3 and WASF1–WASF3, the Wiskott–Aldrich syndrome protein and SCAR homolog WASH (also known as WASH1), and the WHAMM and JMY proteins. WASP proteins share a conserved C-terminal Arp2/3-complex-activating WCA module. This module consists of either one or multiple actin-monomer-binding WH2 (W) domains, a central domain (C) and an acidic (A) domain, which mediate Arp2/3 binding. Apart from the catalytic WCA module, WASP proteins often share a proline-rich region and a basic region, which bind SH3-domain containing proteins and acidic phosphoinositides, respectively. WASP proteins are regulated by similar molecular principles. Under resting conditions NPFs are primarily inactive and become activated upon binding of the Rho GTPases Cdc42 and Rac1. Additionally, a variety of factors further modulate proper activation and recruitment of WASP proteins (Brinkmann, 2015).
In Drosophila, only three WASP subfamily members have been described, namely WAVE, WASP and WASH (also known as CG13176). Insects like Drosophila have subsequently lost a WHAMM/JMY gene, although the common ancestor first arose in invertebrates. Genetic studies indicate that WAVE and WASP are the central activators of the Arp2/3 complex, differentially regulating most aspects of Arp2/3 function in Drosophila. These studies highlight distinct, but also overlapping cellular requirements of WAVE and WASP during development. WAVE function is in particular essential for cell shape and morphogenetic cell movements during development. By contrast, WASP function is needed for cell fate specification of sensory organ precursors (SOPs) and spermatid Both, WASP and WAVE are required for myoblast fusion (Brinkmann, 2015).
Loss of maternal and zygotic WASP results in late-embryonic lethality due to strong defects in cell fate decisions of neuronal cell lineages and myoblast fusion defects. Remarkably, animals lacking zygotic WASP function survive until early adulthood. Thus, maternally provided WASP protein is sufficient for proper embryonic and larval development. Mutant wasp flies show no strong morphological defects except a partial loss of sensory bristles. Loss of zygotic Arp2/3 function results in a similar, albeit stronger, neurogenic phenotype suggesting an involvement of additional factors in Arp2/3-dependent SOP development (Rajan et al., 2009). The loss of sensory bristles in wasp and arp2/3 mutants phenocopies Notch loss-of-function and is caused by a pIIa-to-pIIb cell fate transformation. This results in an excess of neurons at the expense of bristle sheath, shaft and socket cells. Recent work further suggests that the WASP–Arp2/3 pathway rather plays an important role in the trafficking of Delta-positive vesicles from the basal area to the apical cortex of the signal-sending pIIb cell (Brinkmann, 2015).
Remarkably, rescue experiments have implied that established activators of WASP, such as Cdc42 or phosphatidylinositol 4,5-bisphosphate (PIP2), are not required for WASP function, neither for the myoblast fusion process nor for SOP development. The identity of an independent activator that might act cooperatively to control Arp2/3 function in these contexts is unknown. This study presents a functional analysis of WHAMY, a new WASP-like protein that regulates cell motility of Drosophila blood cells but also synergizes with WASP during embryonic muscle formation and cell fate specification of adult SOPs (Brinkmann, 2015).
The identification of all WASP family homologs in all sequenced organisms allows a detailed phylogenetic analysis of the origin of diverse subfamilies evolving differential cellular functions. WASP proteins are multi-domain proteins. They share functions that are encoded by similar domains at the C-termini, whereas different N-terminal domains mainly define their diverse cellular processes. Gene duplication and domain shuffling are two important mechanisms driving novel and increasingly complex developmental programs during evolution. It is thought that this boost in domain shuffling is responsible for the apparent disconnection between greatly increased phenotypic complexity and a relatively small difference in gene number between humans and Drosophila (Brinkmann, 2015).
The whamy gene is an excellent example for how gene duplication and subsequent domain shuffling can create new gene functions after initial gene duplication. It arose through a duplication of wasp at the base of the genus Drosophila. Although the encoded protein has evolved a new function in cell motility, it also functions synergistically with WASP in muscle formation and sensory organ development. In the latter, WHAMY can even partially substitute for WASP, indicating that it has kept functionality following the duplication. This duality is reflected in the sequence of the WHAMY CRIB domains. As there is an overlap in function with WASP, selective pressure has been reduced since the duplication, leading to the observed increase of evolutionary rate. Following the duplication of the CRIB domains within WHAMY, a similar trend can be found. Whereas one domain has kept the function of binding to Cdc42-GTP, the other has lost the ability to interact. This is reflected in domain-specific conserved substitutions. The duplication of the wasp gene and subsequent subneofunctionalization of whamy might have occurred at the same time as the loss of a true WHAMM/JMY ancestor during insect evolution (Veltman, 2010). Like Drosophila WHAMY, the common ancestor of WHAMM/JMY proteins in invertebrates also lacks the characteristic C-terminal tryptophan residue in their VCA domains that is crucial for Arp2/3 binding and activation (Veltman, 2010). This further implies a primary Arp2/3-independent function of the common ancestor of invertebrate WHAMM/JMY proteins (Brinkmann, 2015).
WHAMY shows no Arp2/3-activating nucleation promoting factor (NPF) activity in vitro. However, different from WASP, WHAMY itself is able to promote fast elongation of linear actin filaments from actin-rich clusters. With respect to its activity, WHAMY resembles the WH2-domain containing Ena/VASP polymerases that actively drive processive actin-filament elongation and promote assembly of both lamellipodial and filopodia actin networks. Notably, Ena/VASP proteins are tetramers, and their oligomerization is mandatory to allow for polymerase activity in experiments in solution, as used in this study. Since fast filament elongation was exclusively observed from WHAMY clusters in total internal reflection fluorescence (TIRF) experiments, and consistent with the size exclusion chromatography experiments, it is proposed that WHAMY requires oligomerization to acquire actin polymerase activity. Concerning previously analyzed proteins of the WASP family, the filament elongation activity of WHAMY is therefore rather unique, and when compared to other fast actin polymerases, only the Drosophila formin Diaphanous achieves comparable high elongation activity in vitro. As evidenced from the pyrene data, the activity of WHAMY can further be increased by Rac1 (Brinkmann, 2015).
Rac1 seems to act on both the activity and the localization of WHAMY at lamellipodial tips. Both of the two CRIB domains of WHAMY bind equally to activated Rac1, and only loss of both CRIB domains abolishes Rac1 binding and the localization to the leading edge. Therefore, it currently remains unclear why WHAMY contains two CRIB domains and whether they differentially mediate distinct cellular functions. They might contribute to a local clustering of WHAMY and Rac1 at the leading edge. The most prominent Rac1 effector represents the WAVE regulatory complex (WRC) that drives Arp2/3-mediated branched actin nucleation. Rac1 directly binds and activates the WRC by allosterically releasing the bound Arp2/3-activating WCA domain of WAVE. Overexpression of WHAMY leads to a strong induction of filopodia, presumably due to the filament elongation activity of WHAMY. Additionally, competition between WHAMY and the WRC for Rac1 could disturb the balance between nucleation and elongation activity, and therefore might contribute to the observed overexpression phenotype. Different from WHAMY, WRC function is essential for lamellipodia formation and cell migration in most eukaryotic cells. By contrast, loss of WHAMY function does not impair lamellipodia formation but rather regulates cell spreading and contributes to cell motility (Brinkmann, 2015).
WHAMY does not compete but rather functions together with WASP in Drosophila morphogenesis. Previous studies have revealed that the major established activators of WASP, such as Cdc42 and PIP2, are not required for the function of WASP in sensory organ development or myoblast fusion. This observation already suggests that additional components, such as WHAMY, might act together with WASP in sensory organ development and myoblast fusion. Consistent with this, further reduction of whamy function in wasp mutants was found to phenocopy loss of arp2/3 function, resulting in an excess of neurons and a near absence of bristle sheath, shaft and socket cells. Rescue data further indicate that WHAMY can partially substitute for WASP function. Thus, WHAMY cooperates with WASP rather than acting redundantly in sensory organ development. Based on TIRF microscopy data, it is suggested that WHAMY might potentially generate mother filaments in close vicinity of Arp2/3 complex facilitating Arp2/3-mediated actin assembly (Brinkmann, 2015).
How might WHAMY and WASP act on actin dynamics during sensory organ development? Recent work suggests that the WASP–Arp2/3 pathway is not involved in Notch receptor endocytosis or its processing in the signal-receiving cell (pIIa) but rather plays an important role in the trafficking of Delta-positive vesicles from the basal area to the apical cortex of the signal-sending pIIb cell. This model also implies that recycled Notch ligands such as Delta and Serrate are active at apical junctions with actin-rich structures induced by WASP and the Arp2/3 complex, which in turn activate apical Notch receptor in pIIa. In vivo, WHAMY localizes at dynamic vesicles during sensory organ precursor formation and, together with WASP, becomes strongly enriched at apical junctions shortly after SOP division. Thus, a scenario is proposed in which WASP and WHAMY might act either on the assembly of actin-rich structures or directly promote apical trafficking of Delta through Rab11-recycling endosomes (Brinkmann, 2015).
A dynamic reorganization of the actin cytoskeleton into distinct cellular structures is also necessary to ensure successful myogenesis. Filopodial protrusions are crucial for the attachment of FCMs to the founder cell and growing myotube, and for the initiation of the fusion process. The recognition and adhesion of myoblasts depends on members of the immunoglobulin superfamily (IgSF) that are expressed specifically in myoblasts in a ring-like structure. The interaction of these proteins leads to the formation of a cell communication structure, which has been termed fusion-restricted myogenic adhesive structure (FuRMAS) or podosome-like structure. The cytodomains of the IgSFs trigger the activation of WAVE in founder cells, and of WAVE and WASP in FCMs. In FCMs, WAVE- and WASP-mediated Arp2/3 activation results in the formation of a dense F-actin focus that accumulates at the interface of adhering myoblasts. Electron microscopy studies have revealed that WASP is required for the formation of fusion pores at apposing myoblasts during embryonic and indirect flight muscle development. These fusion pores expand until full cytoplasmic continuity is achieved, and WASP has implicated to be required for fusion pore expansion. It has been discussed that WASP is required for the removal of membrane residuals during membrane vesiculation. WHAMY might contribute to this process, but the detailed mechanistic contribution of WHAMY in fusion pore formation needs to be addressed in future studies by ultrastructural analyses (Brinkmann, 2015).
CYtoplasmic FMRP Interacting Protein 1 (CYFIP1) is a candidate gene for intellectual disability (ID), autism, schizophrenia and epilepsy. It is a member of a family of proteins that is very conserved during evolution, sharing high homology with dCYFIP, its Drosophila homolog. CYFIP1 interacts with the Fragile X Mental Retardation Protein (FMRP), whose absence causes the Fragile X Syndrome, and with the translation initiation factor eIF4E. It is a member of the WAVE Regulatory Complex (WRC), thus representing a link between translational regulation and actin cytoskeleton. Data is presented showing a correlation between mRNA levels of CYFIP1 and other members of the WRC. This suggests a tight regulation of the levels of the WRC members not only by post-translational mechanisms, as previously hypothesized. Moreover, the impact of loss of function of both CYFIP1 and FMRP on neuronal growth and differentiation in was studied in two animal models, fly and mouse. These two proteins antagonize each other's function not only during neuromuscular junction growth in the fly but also during new neuronal differentiation in the olfactory bulb of adult mice. Mechanistically, FMRP and CYFIP1 modulate mTor signaling in an antagonistic manner, likely via independent pathways, supporting the results obtained in mouse as well as in fly at the morphological level. Collectively, these results illustrate a new model to explain the cellular roles of FMRP and CYFIP1 and the molecular significance of their interaction (Abekhoukh, 2017).
The WAVE regulatory complex (WRC) is the major Arp2/3 activator, promoting lamellipodial protrusions in migrating cells. The WRC is basally inactive but can be activated by Rac1 and phospholipids, and phosphorylation. However, the in vivo relevance of phosphorylation of WAVE remains largely unknown. This study identified the kinase CK1&apha; as a novel regulator of WAVE controlling cell shape and cell motility in Drosophila macrophages. CK1α binds and phosphorylates WAVE in vitro. Phosphorylation of WAVE by CK1α appears not to be required for activation but rather regulates its stability. Pharmacologic inhibition of CK1α promotes ubiquitin-dependent degradation of WAVE. Consistently, loss of ck1α but not ck2 function phenocopies WAVE depletion. Phosphorylation-deficient mutations in the CK1α consensus sequences within the VCA domain of WAVE can neither rescue mutant lethality nor lamellipodia defects. By contrast, phosphomimetic mutations rescue all cellular and developmental defects. Finally, RNAi-mediated suppression of 26S proteasome or E3 ligase complexes substantially rescues lamellipodia defects in CK1α depleted macrophages. Thus, it is concluded that the basal phosphorylation of WAVE by CK1α protects it from premature ubiquitin-dependent degradation, thus promoting WAVE function in vivo (Hirschhauser, 2021).
Although poleward segregation of acentric chromosomes is well documented, the underlying mechanisms remain poorly understood. This study demonstrates that microtubules play a key role in poleward movement of acentric chromosome fragments generated in Drosophila melanogaster neuroblasts. Acentrics segregate with either telomeres leading or lagging in equal frequency and are preferentially associated with peripheral bundled microtubules. In addition, laser ablation studies demonstrate that segregating acentrics are mechanically associated with microtubules. Finally, this study shows that successful acentric segregation requires the chromokinesin Klp3a. Reduced Klp3a function results in disorganized interpolar microtubules and shortened spindles. Normally, acentric poleward segregation occurs at the periphery of the spindle in association with interpolar microtubules. In klp3a mutants, acentrics fail to localize and segregate along the peripheral interpolar microtubules and are abnormally positioned in the spindle interior. These studies demonstrate an unsuspected role for interpolar microtubules in driving acentric segregation (Karg, 2017).
Actins are structural components of the cytoskeleton and muscle, and numerous actin isoforms are found in most organisms. However, many actin isoforms are expressed in distinct patterns allowing each actin to have a specialized function. Numerous studies have demonstrated that actin isoforms both can and cannot compensate for each other under specific circumstances. This allows for an ambiguity of whether isoforms are functionally distinct. This study analyzed mutants of Drosophila Act79B, the predominant actin expressed in the adult jump muscle. Functional and structural analysis of the Act79B mutants found the flies to have normal jumping ability and sarcomere structure. Analysis of actin gene expression determined that expression of Act88F, an actin gene normally expressed in the flight muscles, was significantly up-regulated in the jump muscles of mutants. This indicated that loss of Act79B caused expansion of Act88F expression. When double mutants were created of Act79B and Act88F, this abolished the jump ability of the flies and resulted in severe defects in myofibril formation. These results indicate that Act88F can functionally substitute for Act79B in the jump muscle, and that the functional compensation in actin expression in the jump muscles only occurs through Act88F (Dohn, 2018).
Proper muscle contraction requires the assembly and maintenance of sarcomeres and myofibrils. Although the protein components of myofibrils are generally known, less is known about the mechanisms by which they individually function and together synergize for myofibril assembly and maintenance. For example, it is unclear how the disruption of actin filament (F-actin) regulatory proteins leads to the muscle weakness observed in myopathies. This study shows that knockdown of Drosophila Tropomodulin (Tmod), results in several myopathy-related phenotypes, including reduction of muscle cell (myofiber) size, increased sarcomere length, disorganization and misorientation of myofibrils, ectopic F-actin accumulation, loss of tension-mediating proteins at the myotendinous junction, and misshaped and internalized nuclei. These findings support and extend the tension-driven self-organizing myofibrillogenesis model. Like its mammalian counterpart, Drosophila Tmod was shown to cap F-actin pointed-ends, and it is proposed that this activity is crucial for cellular processes in different locations within the myofiber that directly and indirectly contribute to the maintenance of muscle function. These findings provide significant insights to the role of Tmod in muscle development, maintenance and disease (Zapater. 2023).
Heterochromatin mainly comprises repeated DNA sequences that are prone to ectopic recombination. In Drosophila cells, 'safe' repair of heterochromatic double-strand breaks by homologous recombination relies on the relocalization of repair sites to the nuclear periphery before strand invasion. The mechanisms responsible for this movement were unknown. This study shows that relocalization occurs by directed motion along nuclear actin filaments assembled at repair sites by the Arp2/3 complex. Relocalization requires nuclear myosins associated with the heterochromatin repair complex Smc5/6 and the myosin activator Unc45, which is recruited to repair sites by Smc5/6. ARP2/3, actin nucleation and myosins also relocalize heterochromatic double-strand breaks in mouse cells. Defects in this pathway result in impaired heterochromatin repair and chromosome rearrangements. These findings identify de novo nuclear actin filaments and myosins as effectors of chromatin dynamics for heterochromatin repair and stability in multicellular eukaryotes (Caridi, 2018).
The establishment of left-right (LR) asymmetry is fundamental to animal development, but the identification of a unifying mechanism establishing laterality across different phyla has remained elusive. A cilia-driven, directional fluid flow is important for symmetry breaking in numerous vertebrates, including zebrafish. Alternatively, LR asymmetry can be established independently of cilia, notably through the intrinsic chirality of the acto-myosin cytoskeleton. This study shows that Myosin1D (Myo1D), a previously identified regulator of Drosophila LR asymmetry, is essential for the formation and function of the zebrafish LR organizer (LRO), Kupffer's vesicle (KV). Myo1D controls the orientation of LRO cilia and interacts functionally with the planar cell polarity (PCP) pathway component VanGogh-like2 (Vangl2; see Drosophila Van Gogh), to shape a productive LRO flow. These findings identify Myo1D as an evolutionarily conserved regulator of animal LR asymmetry, and show that functional interactions between Myo1D and PCP are central to the establishment of animal LR asymmetry (Juan, 2018).
Myosin-1D (myo1D) is important for Drosophila left-right asymmetry, and its effects are modulated by myosin-1C (myo1C). De novo expression of these myosins in nonchiral Drosophila tissues promotes cell and tissue chirality, with handedness depending on the paralog expressed. Remarkably, the identity of the motor domain determines the direction of organ chirality, rather than the regulatory or tail domains. Myo1D, but not myo1C, propels actin filaments in leftward circles in in vitro experiments, but it is not known if this property contributes to establishing cell and organ chirality. To further explore if there are differences in the mechanochemistry of these motors, the ATPase mechanisms of myo1C and myo1D were determined. Myo1D was found to have a 12.5-fold higher actin-activated steady-state ATPase rate, and transient kinetic experiments revealed myo1D has an 8-fold higher MgADP release rate compared to myo1C. Actin-activated phosphate release is rate limiting for myo1C, whereas MgADP release is the rate-limiting step for myo1D. Notably, both myosins have among the tightest MgADP affinities measured for any myosin. Consistent with ATPase kinetics, myo1D propels actin filaments at higher speeds compared to myo1C in in vitro gliding assays. Finally, the ability of both paralogs to transport 50 nm unilamellar vesicles along immobilized actin filaments was tested, and robust transport was found by myo1D and actin binding but no transport by myo1C. These findings support a model where myo1C is a slow transporter with long-lived actin attachments, whereas myo1D has kinetic properties associated with a transport motor (Baez-Cruz, 2023).
Protein histidine methylation is a rare post-translational modification of unknown biochemical importance. In vertebrates, only a few methylhistidine-containing proteins have been reported, including beta-actin as an essential example. The evolutionary conserved methylation of beta-actin H73 is catalyzed by an as yet unknown histidine N-methyltransferase. This study reports that the protein SETD3 is the actin-specific histidine N-methyltransferase. In vitro, recombinant rat and human SETD3 methylated beta-actin at H73. Knocking-out SETD3 in both human HAP1 cells and in Drosophila melanogaster resulted in the absence of methylation at beta-actin H73 in vivo, whereas beta-actin from wildtype cells or flies was > 90% methylated. As a consequence, it was shown that Setd3-deficient HAP1 cells have less cellular F-actin and an increased glycolytic phenotype. In conclusion, by identifying SETD3 as the actin-specific histidine N-methyltransferase, this work pioneers new research into the possible role of this modification in health and disease and questions the substrate specificity of SET-domain-containing enzymes (Kwiatkowski. 2018).
Epithelial folding transforms simple sheets of cells into complex three-dimensional tissues and organs during animal development. Epithelial folding has mainly been attributed to mechanical forces generated by an apically localized actomyosin network, however, contributions of forces generated at basal and lateral cell surfaces remain largely unknown. This study shows that a local decrease of basal tension and an increased lateral tension, but not apical constriction, drive the formation of two neighboring folds in developing Drosophila wing imaginal discs. Spatially defined reduction of extracellular matrix density results in local decrease of basal tension in the first fold; fluctuations in F-actin lead to increased lateral tension in the second fold. Simulations using a 3D vertex model show that the two distinct mechanisms can drive epithelial folding. This combination of lateral and basal tension measurements with a mechanical tissue model reveals how simple modulations of surface and edge tension drive complex three-dimensional morphological changes (Sui, 2018).
This work has uncovered two new mechanisms of epithelial fold formation. First, a locally defined basal decrease of surface and edge tension, associated with local reduction of ECM density, leads to basal cell expansion and folding. Second, a lateral increase of surface tension at the future fold location, associated with F-actin flows and pulsatile contractions, leads to a local reduction of tissue height and fold formation. It is conceivable that both mechanisms may also operate in combination during epithelial folding (Sui, 2018).
A simplified picture resulting from mechanical analysis of how basal tension reduction can induce fold formation is as follows. Higher basal tension in the cells outside the fold compared to cells inside the fold stretches the basal surface areas of fold cells. Consequently, fold cells widen basally and reduce cell height to maintain cell volume. The new force balance state is characterized by apical indentation and wedge-shaped, shortened cells. How is ECM depletion linked to a decrease in basal cell edge and surface tension? In one scenario, following ECM depletion, the actomyosin network lacks stabilization via binding to integrins, reducing the active tension it can generate with myosin molecular motors. Alternatively, the ECM and cortical actomyosin network, linked together via integrins and other molecules, can be seen as a single composite material under tension. Elastic straining of the ECM, e.g. during tissue growth, could give rise to a passive mechanical tension within the ECM. As the ECM is depleted, the composite material is reorganized and passive ECM stress due to ECM straining could be lost, also contributing to the overall decrease in basal tension in the fold (Sui, 2018).
Lateral tension increase can also induce fold formation. This can be outlined in a simplified picture (see Differential lateral and basal tension drive folding of Drosophila wing discs through two distinct mechanisms). Increased lateral tension leads to a reduction in cell height. Since basal tension is high, the shortened cells deform the apical surface inwards, while the basal surface resists deformation. As the cells resist volume changes, they widen. Conceivably, increased apical tension in the fold cells favors further basal expansion of the fold cells (Sui, 2018).
Folding requires the transition of cells from a columnar to a wedge-shape where the apical surface is smaller than the basal surface. Previous work has stressed the role of mechanical stresses generated by apical actomyosin networks driving apical constriction during folding. This work shows that for the epithelial folds, in the case of the wing, apical constriction is not important. Instead, they rely either on the basal widening of cells due to the decrease of basal tension or alternatively on increased lateral tension. Interestingly, two fundamentally different mechanisms generate similar morphologies of neighboring folds. This implies that the mechanical processes shaping a tissue cannot be deduced from the tissue morphology alone. Cell shortening and an active role for the ECM is also required for the folding of the zebrafish embryonic brain. Basal decrease of tension and lateral increase of tension may therefore represent two important mechanisms driving the folding of epithelia in different organisms (Sui, 2018).
Spectrins are membrane cytoskeletal proteins generally thought to function as heterotetramers comprising two α-spectrins and two β-spectrins. They influence cell shape and Hippo signaling, but the mechanism by which they influence Hippo signaling has remained unclear. This study has investigated the role and regulation of the Drosophila β-heavy spectrin (β(H)-spectrin, encoded by the karst gene) in wing imaginal discs. The results establish that β(H)-spectrin regulates Hippo signaling through the Jub biomechanical pathway due to its influence on cytoskeletal tension. While this study found that α-spectrin also regulates Hippo signaling through Jub, unexpectedly, it was found that β(H)-spectrin localizes and functions independently of α-spectrin. Instead, β(H)-spectrin co-localizes with and reciprocally regulates and is regulated by myosin. In vivo and in vitro experiments support a model in which β(H)-spectrin and myosin directly compete for binding to apical F-actin. This competition can explain the influence of β(H)-spectrin on cytoskeletal tension and myosin accumulation. It also provides new insight into how β(H)-spectrin participates in ratcheting mechanisms associated with cell shape change (Ibar, 2023).
Multiple models for how spectrins regulate Hippo signaling have been proposed. This study focused on investigating claims that spectrins could alter cytoskeletal tension in wing discs by regulating pMLC levels without affecting localization of myosin or Jub. In contrast to prior studies, this study observed that when βH-Spec or α-Spec levels are decreased by RNAi, levels of junctional myosin are increased. This increase in junctional myosin levels is associated with increased junctional tension and with increased recruitment of Jub to AJ. It is uncertain why these effects were missed in prior studies, but it is noted that these observations are consistent with studies linking recruitment of both myosin and Jub to AJ under tension. Moreover, the results are further supported by the observation that jub is genetically required for the influence of spectrin knockdown on Yki activity and wing growth. Taken together, these observations imply that βH-Spec and α-Spec regulate Hippo signaling in wing discs through the Jub biomechanical pathway, rather than through hypothesized alternate mechanisms (Ibar, 2023).
Spectrin has been suggested to form two distinct complexes in Drosophila epithelial cells, (αβ)2 and (αβH)2 heterotetramers, which localize to the lateral and apical sides of cells, respectively. However, the current observations indicate that βH-Spec functions independently of α-Spec in wing imaginal discs. α-Spec and βH-Spec do not exhibit significant co-localization. Moreover, α-Spec and βH-Spec are not required for each other's localization to apical cell junctions. This contrasts with the requirement for β-Spec for recruitment of α-Spec to lateral membranes in wing discs. The requirement is not entirely reciprocal, as α-Spec knockdown only partially reduces β-Spec recruitment, but this likely reflects mechanisms that recruit spectrins to cell membranes: β-Spec subunits, but not α-Spec subunits, have a pleckstrin homology domain that can mediate membrane association, as well as possessing the CH domains that mediate F-actin association. Thus β-Spec can associate with lateral membranes without α-Spec, but α-Spec does not have a way to associate with lateral membranes without β-Spec (Ibar, 2023).
An independent role for βH-Spec has also been suggested in mammalian photoreceptors, where it was reported that that mammalian βV-Spec does not co-localize with αII-spectrin, and that βV-Spec could form homodimers, potentially allowing it to cross-link actin by itself, enabling α-Spec-independent functions. The observation that a mutation in Drosophila α-Spec that disrupts binding to β-Spec in vitro has only mild phenotypes also suggests that spectrin functions do not depend entirely on αβ interactions. Collectively, these results together with these earlier studies emphasize that the dogma that "spectrin comprises α- and β-subunits that interact in an antiparallel manner to form an αβ dimer" should be revised. Nonetheless, the current results do not exclude the possibility that βH-Spec and α-Spec might act together in a physical complex in other tissues, or under distinct physiological conditions (Ibar, 2023).
The conclusion that βH-Spec and α-Spec act independently in wing discs implies that they influence tension at apical junctions through distinct mechanisms. It is suggested that α-Spec could influence AJ tension in wing discs through the mechanism proposed to explain the influence of α- and β-Spec in pupal eyes. It was inferred that α- and β-Spec maintain cell rigidity by linking F-actin to membranes. In the absence of α- or β-Spec, it was proposed that dissociation between F-actin and the membrane leads to an expansion of the apical regions. This expansion increases cytoskeletal tension at AJs, which bind F-actin independently of spectrins. Consistent with this suggested mechanism, a decrease in cell height was observed in α-Spec knockdown cells in wing discs, in conjunction with increased tension at apical junctions. The alteration in cell shape was not observed in βH-Spec knockdown cells, further supporting the conclusion that βH-Spec and α-Spec act in different ways to regulate junctional tension (Ibar, 2023).
Instead, experiments analyzing the relationship between βH-Spec and myosin revealed an entirely different mechanism by which βH-Spec influences tension at AJ. A mutual antagonism was observed between βH-Spec and myosin in vivo for localization to apical F-actin: decreasing βH-Spec increases junctional myosin, while increasing βH-Spec decreases junctional myosin. Reciprocally, increasing myosin activity decreases βH-Spec localization to apical F-actin, while decreasing myosin activity increases βH-Spec localization to apical F-actin. In vitro studies with purified protein domains revealed that myosin can compete with βH-Spec for binding to F-actin. Finally, computational modeling of protein structures revealed that myosin and βH-Spec would interfere with each other's binding to F-actin. Together, these observations indicate that βH-Spec and myosin can directly compete with each other for localization to F-actin. It is therefore suggested that the influence of βH-Spec on junctional tension is likely to be a direct consequence of its competition with myosin for overlapping binding sites on F-actin (Ibar, 2023).
Despite βH-Spec and myosin sharing overlapping binding sites on F-actin, and competing reciprocally in vivo, in co-sedimentation experiments only a partial ability of myosin to compete for βH-Spec binding was detecte, and an ability of βH-Spec to compete for myosin binding could not be detected. Several factors are likely to contribute to these observations. First, higher protein concentrations of myosin or βH-Spec could not be detected due to the need to keep the salt concentration constant and close to physiological levels, and to prevent protein precipitation. Second, it has been suggested for β-Spec that the CH2 domain regulates the actin binding function of the CH1 domain through steric hindrance when the two domains are associated. A specific mutation in β-Spec CH2 (L253P) has been shown to lower the energetic barrier between closed and open structural states, increasing the affinity of β-Spec for F-actin around 1000-fold, but it is unknown how β-Spec or βH-Spec conformational changes are normally regulated in vivo and whether both proteins share this regulatory feature. Additionally, phosphorylation of myosin regulatory light chain shifts myosin from a compact, autoinhibited conformation to a filamentous, active conformation . The autoinhibited conformation binds F-actin very weakly (KD>100 μM) compared to the active conformation, suggesting that spectrin could outcompete autoinhibited myosin more effectively than active myosin for binding to F-actin. In vitro experiments used an active form of myosin. In addition, other factors including other actin-binding proteins and cytoskeletal tension are likely to influence the dynamic localization and actin-binding properties of both proteins (Ibar, 2023).
The competition between βH-Spec and myosin also provides key insights into how βH-Spec contributes to ratcheting of apical constriction. The apical constriction of cells in the ventral furrow that initiates Drosophila mesoderm invagination occurs through fast constriction pulses interrupted by pauses during which cells must stabilize their constricted state before reinitiating constriction. This ratcheting-like behavior is thought to be a consequence of the finite length of actin filaments. Myosin contracts the cytoskeleton by driving filaments past each other, and extensive contractions require release and reassociation with new pairs of filaments. βH-Spec participates in ratcheting of apical constriction. When βH-Spec is knocked down, cells can undergo cycles of unratcheted apical constriction during which they alternately constrict and then expand. Consequently, most βH-Spec-depleted embryos fail to complete normal mesoderm invagination. It was proposed that the actin cross-linking function of βH-Spec could hold F-actin in place for the next cycle of myosin-mediated contraction, but this raises the question of how myosin and βH-Spec association with F-actin are coordinated so that βH-Spec prevents relaxation without interfering with constriction. The results suggest a simple solution: since they compete for the same binding site, the release of myosin from F-actin at the end of a cycle of contraction would naturally be coupled to the accessibility of F-actin for binding by βH-Spec. Thus, the competition between myosin and βH-Spec for binding to F-actin enables myosin-mediated cell contraction to effectively alternate with βH-Spec-mediated stabilization (Ibar, 2023).
Different memory components are forgotten through distinct molecular mechanisms. In Drosophila, the activation of 2 Rho GTPases (Rac1 and Cdc42), respectively, underlies the forgetting of an early labile memory (anesthesia-sensitive memory, ASM) and a form of consolidated memory (anesthesia-resistant memory, ARM). This study dissected the molecular mechanisms that tie Rac1 and Cdc42 to the different types of memory forgetting. Two WASP family proteins, SCAR/WAVE and WASp, act downstream of Rac1 and Cdc42 separately to regulate ASM and ARM forgetting in mushroom body neurons. Arp2/3 complex, which organizes branched actin polymerization, is a canonical downstream effector of WASP family proteins. However, this study found that Arp2/3 complex is required in Cdc42/WASp-mediated ARM forgetting but not in Rac1/SCAR-mediated ASM forgetting. Instead, Rac1/SCAR may function with formin Diaphanous (Dia), a nucleator that facilitates linear actin polymerization, in ASM forgetting. The present study, complementing the previously identified Rac1/cofilin pathway that regulates actin depolymerization, suggests that Rho GTPases regulate forgetting by recruiting both actin polymerization and depolymerization pathways. Moreover, Rac1 and Cdc42 may regulate different types of memory forgetting by tapping into different actin polymerization mechanisms (Gao, 2019).
There are 3 major findings. First, 2 WASP family proteins, SCAR/WAVE and WASp, act as downstream effectors of Rac1-mediated ASM forgetting and Cdc42-mediated ARM forgetting, respectively. Second, although the Arp2/3 complex is a well-established effector that links activation of WASP family proteins to actin polymerization, it is only required in Cdc42/WASp-mediated ARM forgetting. Instead, formin Dia functions together with Rac1/SCAR in ASM forgetting. Third, feeding inhibitors of the Arp2/3 complex and Dia to fruit flies led to rather specific effects on ASM and ARM forgetting, raising the possibility of developing drugs on these molecular targets to treat memory-related diseases (Gao, 2019).
The effect of Rac1 on ASM forgetting has been tied to the activation of an actin depolymerization regulator cofilin presumably through a PAK/LIMK phosphorylation cascade. However, actin dynamics is a balanced play that requires continuous turnover between polymerization and depolymerization. It is not known whether signaling pathways regulating actin polymerization also play a role. There are 3 families of proteins that nucleate and promote actin polymerization, Arp2/3 complex, WH2-domain proteins, and formin. The finding that Arp2/3 complex and formin Dia function in ARM and ASM forgetting suggests that both actin polymerization and depolymerization pathways contribute to forgetting. How Arp2/3 complex and Dia separately contribute to ARM and ASM forgetting remains an open question. It is yet to be determined whether these proteins have different expression or subcellular locations in the MB neurons. However, it is interesting that Arp2/3 complex and formins are specialized in different types of actin polymerization (Gao, 2019).
In a working model, Cdc42 activates Arp2/3 complex via a canonical pathway (Cdc42/WASp/Arp2/3 complex), while Rac1-mediated ASM forgetting depends on SCAR/WAVE complex. This complex, in addition to SCAR/WAVE, includes at least 4 other members: Sra-1, Abi, HSPC300, and Kette. These additional members are thought to hold SCAR/WAVE in the complex in an inactive state, until GTP-bound Rac1 binds to Sra-1 and relieves the inhibition. On the other hand, the intact complex is essential for the stability of the SCAR/WAVE protein as well (i.e., failure to keep the intact complex can lead to SCAR degradation). This latter effect may explain the observation that RNAi knockdown of SCAR complex members has the same effect on inhibiting forgetting as the knockdown of SCAR. As a WASP family protein, SCAR/WAVE is able to associate with and activate Arp2/3 complex through its C-terminal region. However, RNAi knockdown of Arp2 and Arp3 and pharmacological inhibition of Arp2/3 complex specifically affects ARM forgetting, while no effects on ASM retention were observed. It is therefore proposed that Rac1/SCAR may function through Arp2/3 complex-independent mechanisms. SCAR/WAVE complex is reported to physically associates with Dia through one of its members, Abi, to regulate actin dynamics. Behavioral characterization of Dia knockdown and overexpression, as well as the genetic epistasis experiment, support the idea that Dia could be downstream of Rac1/SCAR in ASM forgetting. Details about the functional coordination between SCAR/WAVE and Dia therefore await further clarification (Gao, 2019).
Cell migration is hypothesized to involve a cycle of behaviours beginning with leading edge extension. However, recent evidence suggests that the leading edge may be dispensable for migration, raising the question of what actually controls cell directionality. This study exploits the embryonic migration of Drosophila macrophages to bridge the different temporal scales of the behaviours controlling motility. This approach reveals that edge fluctuations during random motility are not persistent and are weakly correlated with motion. In contrast, flow of the actin network behind the leading edge is highly persistent. Quantification of actin flow structure during migration reveals a stable organization and asymmetry in the cell-wide flowfield that strongly correlates with cell directionality. This organization is regulated by a gradient of actin network compression and destruction, which is controlled by myosin contraction and cofilin-mediated disassembly. It is this stable actin-flow polarity, which integrates rapid fluctuations of the leading edge, that controls inherent cellular persistence (Yolland, 2019).
The actin cytoskeleton is regulated by many proteins including capping proteins that stabilize actin filaments (F-actin) by inhibiting actin polymerization and depolymerization. This study report two pediatric probands who carry damaging heterozygous de novo mutations in CAPZA2 (HGNC: 1490) and exhibit neurological symptoms with shared phenotypes including global motor development delay, speech delay, intellectual disability, hypotonia and a history of seizures. CAPZA2 encodes a subunit of an F-actin-capping protein complex (CapZ). CapZ is an obligate heterodimer consisting of alpha and beta heterodimer conserved from yeast to human. Vertebrate genomes contain three alpha subunits encoded by three different genes and CAPZA2 encodes the alpha2 subunit. The single orthologue of CAPZA genes in Drosophila is cpa. Loss of cpa leads to lethality in early development and expression of the human reference CAPZA2 rescues this lethality. However, the two CAPZA2 variants identified in the probands rescue this lethality at lower efficiency than the reference. Moreover, expression of the CAPZA2 variants affects bristle morphogenesis, a process that requires extensive actin polymerization and bundling during development. Taken together, these findings suggest that variants in CAPZA2 lead to a non-syndromic neurodevelopmental disorder in children (Huang, 2020).
The actin cytoskeleton drives cell motility and is essential for neuronal development and function. LIM and SH3 Protein 1 (LASP1) is a unique actin-binding protein that is expressed in a wide range of cells including neurons, but its roles in cellular motility and neuronal development are not well understood. LASP1 is expressed in rat hippocampus early in development, and this expression is maintained through adulthood. High-resolution imaging reveals that LASP1 is selectively concentrated at the leading edge of lamellipodia in migrating cells and axonal growth cones. This local enrichment of LASP1 is dynamically associated with the protrusive activity of lamellipodia, depends on the barbed ends of actin filaments, and requires both the LIM domain and nebulin repeats of LASP1. Knockdown of LASP1 in cultured rat hippocampal neurons results in a substantial reduction in axonal outgrowth and arborization. Finally, loss of the Drosophila homolog Lasp from a subset of commissural neurons in the developing ventral nerve cord produces defasciculated axon bundles that do not reach their targets. Together, these data support a novel role for LASP1 in actin-based lamellipodial protrusion and establish LASP1 as a positive regulator of both in vitro and in vivo axon development (Pollitt, 2020).
During morphogenesis, cells exhibit various behaviours, such as migration and constriction, which need to be coordinated. How this is achieved remains elusive. During morphogenesis of the Drosophila adult abdominal epidermis, larval epithelial cells (LECs) migrate directedly before constricting apically and undergoing apoptosis. This study investigates the mechanisms underlying the transition from migration to constriction. LECs possess a pulsatile apical actomyosin network, and a change in network polarity correlates with behavioural change. Exploring the properties of the contractile network, it was found that cell contractility, as determined by myosin activity, has an impact on the behaviour of the network, as well as on cytoskeletal architecture and cell behaviour. Pulsed contractions occur only in cells with intermediate levels of contractility. Furthermore, increasing levels of the small Rho GTPase Rho1 disrupts pulsing, leading to cells that cycle between two states, characterised by a junctional cortical and an apicomedial actin network. These results highlight that behavioural change relies on tightly controlled cellular contractility. Moreover, constriction can occur without pulsing, raising questions why constricting cells pulse in some contexts but not in others (Companys, 2020).
During development, morphogenetic processes ultimately shape the organism. Such processes are driven by various cell behaviours, e.g. intercalation, division, migration and shape change, all of which need to be coordinated. Little is known about how this coordination is achieved, how cells switch behaviour and how different behaviours occur simultaneously (Companys, 2020).
Cell behaviour is often directional. Planar cell polarity (PCP) directs cell behaviour in the plane of the tissue, coordinating, for example, junctional remodelling, division orientation and migration. Migrating cells have a protruding front and a contracting back, further indicating cytoskeletal polarity. In contrast, apically constricting cells show radial cell polarity (RCP) (Companys, 2020).
Ultimately, cell behaviour depends on the actin cytoskeleton. Cell migration relies on protrusive activity, such as lamellipodia formation. Cell shape changes, including apical constriction, which reduces apical cell area, depend on actomyosin contractility. Increasing evidence suggests that the actin cytoskeleton shows rhythmical activity, such as actin flows during migration and pulsed contractions, e.g. during junctional remodelling, neuroblast ingression, apical constriction, basal constriction and vertebrate neural tube closure. Pulsed contractions are driven by periodic actomyosin contractions, which are thought to lead to cycles of cell area fluctuation, followed by stabilisation of the resulting smaller area ('ratchet model') (Companys, 2020).
Pulsed contractions are regulated by phosphorylation of Myosin II regulatory light chain [MRLC; Spaghetti squash (Sqh) in Drosophila] by Rho kinase (Rok), and its dephosphorylation by Myosin phosphatase. In addition, Rok inhibits the Myosin-binding subunit of the Myosin phosphatase complex (Mbs). Upstream of Rok, the small GTPase Rho1 is involved in regulating actomyosin contractility in many contexts, from rear retraction in migrating cells to pulsed contractions. The activity of Rho1 is regulated by activating guanine nucleotide exchange factors (GEFs) and inhibitory GTPase-activating proteins (GAPs) (Companys, 2020).
To gain insights into the mechanisms underlying the coordination of cell behaviour, larval epithelial cells (LECs) were studied during formation of the adult abdominal epidermis of Drosophila. During this process, the LECs are replaced by the adult histoblasts. Previous work has shown that LECs undergo directed migration followed by a transition to apical constriction, which eventually leads to delamination and apoptosis (Bischoff, 2012). During migration, LECs form crescent-shaped lamellipodia and migrate posteriorly (Bischoff, 2012). LECs can only move if neighbours provide space by either migrating, reducing apical area or undergoing apoptosis (Companys, 2020).
Studying the LEC cytoskeleton during abdominal morphogenesis, this study showed that LECs possess an apicomedial actomyosin network that undergoes pulsed contractions. The network is planar polarised during migration, undergoing pulsed contractions in the cell back, while the front protrudes. Contractions then re-localise to the cell centre, displaying radial polarity during constriction. Thus, behavioural change correlates with a change in the polarity of the contractile cytoskeletal network. To explore how manipulating actomyosin contractility affects the contractile network, Rho1, Rok and Myosin phosphatase were interfered with. Cellular contractility levels have an impact on the behaviour of the contractile network, with pulsed contractions only occurring in cells with intermediate contractility levels. 'Contractility' is referred to as the ability of a cell to contract its actomyosin network (or increase stress in this network, if cell deformation is resisted), which is ultimately determined by its amount of active myosin II. Interestingly, increasing Rho1 levels interferes with pulsed contractions and leads instead to a cycling of cells between two states characterised by a junctional cortical and an apicomedial actin network. Thus, increasing contractility is sufficient to have an impact on cytoskeletal architecture and, consequently, cell behaviour. Additionally, apical constriction can take place without pulsing, raising questions why constricting cells pulse in some contexts but not in others (Companys, 2020).
A cell's level of contractility determines the behaviour of its actin cytoskeleton, with pulsed contractions depending on intermediate levels of contractility, as well as cytoskeletal architecture and cell behaviour. Moderately increasing contractility causes LECs to cycle between two states, characterised by a junctional cortical and an apicomedial actin network, respectively. Moreover, the data suggest that apical constriction can occur without pulsed contractions, raising questions why constricting cells pulse in some contexts but not in others (Companys, 2020).
During migration, LECs are planar polarised. They protrude at the front and undergo pulsed contractions in the back, where they also create d-v oriented actin bundles. In their junctional cortical network, Sqh::GFP localises preferentially to the a-p junctions, similar to embryonic cells during germband extension. As LECs constrict, their apicomedial network becomes radially polarised, with a central actin focus and radial actin bundles that connect the network to the junctions, resembling a spider's web. In addition, Sqh::GFP is distributed evenly across all junctions. Thus, LEC behavioural change is accompanied by a change in cell polarity from PCP to RCP that underlies a change in cytoskeletal activity as well as an overall reorganisation of cytoskeletal architecture (Companys, 2020).
The polarity of LEC migration depends on PCP signalling, but little is known about the link between PCP and cytoskeletal asymmetry. Cytoskeletal asymmetry could be due to localised activity of small Rho GTPases, with Rho1 controlling pulsed contractions in the cell back and Rac1 promoting lamellipodia formation at the front. Such mutually exclusive localisation has been shown in various cell types. How RCP is established in constricting LECs is still unknown (Companys, 2020).
LECs create very large actin foci when contracting, but their foci period is comparable to other systems that create smaller foci. A possible explanation for the short period of the large foci is that foci in LECs are initiated within a large area in the periphery of the cell and not in a small region from which the contractile event then expands (which would potentially slow down foci formation). Focus formation involves both the recruitment of novel actin and Sqh as well as advection, where already recruited actin and Sqh 'flow' towards the centre of the focus (Companys, 2020).
Interestingly, actin dynamics in LECs involves the formation of distinct regions of contractile activity, depending on LEC behaviour and polarity. Constricting LECs create a single focus in their centre, while migrating LECs generate two alternating foci in their back. This difference cannot easily be explained by a change in cellular polarity. Instead, it could be due to the different shapes of migrating and constricting cells. In migrating cells, which are elongated along the d-v axis, one contractile event might not suffice to constrict the whole apicomedial network. Constricting cells, however, are rounder, which might allow radial actin recruitment over the whole apical area. Alternatively, the two foci could be a consequence of the cytoskeletal architecture of a migrating cell, as seen in keratinocytes, where myosin localises to two areas in the back of the cell, flanking the nucleus (Companys, 2020).
LECs that create actin foci migrate faster than earlier stage LECs, which do not undergo pulsed contractions. However, pulsed contractions are not part of the migratory machinery, as early migrating LECs do not show foci and Rho-CA LECs migrate without pulsatile activity. Thus, occurrence of pulsed contractions in migratory LECs appears to be associated with constriction rather than migration (Companys, 2020).
In contrast to other systems that show pulsed contractions, LECs have a very large apical area, up to 70 μm in diameter. This large area might determine the architecture of the LEC apicomedial network, which consists of persistent actin bundles that connect to the cell cortex, as well as dynamic flows and foci. In large cells, a robust, persistent apicomedial network might be crucial to maintain apical area and transduce contractile forces. In line with these observations, a persistent and a dynamic pool of actin have recently been described in ectodermal cells during germband extension and in amnioserosa cells during dorsal closure in Drosophila, but only the larger amnioserosa cells showed visible actin bundles (Companys, 2020).
Altering levels of contractility in LECs showed that pulsed contractions depend on Rok, Myosin phosphatase and Rho1 activity. In this respect, LECs resemble other cells that undergo pulsed contractions. As shown for Drosophila germband cells and amnioserosa cells, the current results indicate that cytoskeletal network dynamics depends on the level of cell contractility. Only with intermediate, wild-type levels of contractility did LECs show pulsed contractions. Reducing contractility (Rok-RNAi or MbsN300) interfered with pulsed contractions, as did increasing contractility (Rho1-CA or Rok-CAT). In addition to the amount of activated myosin II, contractility is also determined by the availability of G-actin as well as F-actin nucleators and crosslinkers (Companys, 2020).
As well as interfering with pulsed contractions, increasing contractility in LECs had a more far-reaching impact on cytoskeletal architecture and cell behaviour. (1) Over-expression of Rho1-CA resulted in F-actin disappearing apicomedially and localising mostly to the junctional cortex. A similar phenotype has been observed during Drosophila gastrulation, suggesting that Rho1 is involved in determining the ratio of apicomedial versus cortical contractility. (2) While over-expression of Rho1-CA can only activate endogenous Rok, over-expression of Rok-CAT can supply large amounts of activated Rok and thus lead to a stronger and more specific activation of Myosin II. In LECs, this not only resulted in F-actin disappearing apicomedially and localising mostly to the junctional cortex (and thus a loss of pulsed contractions), but also in the formation of cortical actin bundles. In cell culture, activation of myosin has also been shown to create actin bundles during stress fibre formation. (3) Increased junctional cortical contractility in both Rho1-CA and Rok-CAT LECs furthermore induced cell blebbing. Blebbing is driven by strong cortical myosin activation. During Drosophila dorsal closure, activation of Myosin light chain kinase and Mbs led to cell blebbing. (4) In 'strong' phenotypes of both Rok-CAT and Rho-CA pupae, LECs do not generate lamellipodia-like protrusions and do not migrate. Thus, high levels of activated myosin appear to over-ride protrusive activity and force cells to constrict. A comparable hyper-contractile phenotype has been described in CHO.K1 cells, where over-expressing a phosphomimetic form of myosin light chain leads to a loss of cell spreading and migration (Companys, 2020).
Increasing wild-type Rho1 levels (UAS.rho1) proved particularly interesting. Cells appear to cycle between a state in which actin accumulates at the junctional cortex and cells show blebbing, and a state in which the apicomedial network is present but non-pulsatile. This cycling phenotype could be explained by the rhythmical activation and de-activation of Rho1 via the endogenous machinery in a context of increased overall contractility of the network. With increasing levels of activated Rho1 (and thus increased levels of activated myosin), LECs appear to shift their ratio of apicomedial versus junctional cortical actin towards cortical. This is reversed when levels of activated Rho1 (and thus levels of activated myosin) decrease. The current results support this hypothesis, as not only the disappearance of apicomedial actin was observed, but also the beginning of junctional cortical blebbing, which suggests an increase in contractility in the junctional cortical network. These observations highlight the importance of specifically coordinating the junctional cortical and the apicomedial actomyosin networks (Companys, 2020).
Although pulsed contractions are affected by the manipulation of contractility, most LECs in Rho-RNAi, MbsN300, UAS.Rho1, Rho1-CA or Rok-CAT pupae constricted successfully. In all these cases, pulsed contractility of the apicomedial actin network was impaired, but cells still had actin at their junctional cortex. This indicates that contractility at the junctional cortex alone can drive apical constriction. Similar observations have been made in amnioserosa cells and during neural tube formation. The observation that boundary LECs constrict without pulsed contractions and with insignificant cell area fluctuation further supports the notion that the contractile force needed for constriction can be created by the junctional cortical network, acting like a contractile ring during cell extrusion, as suggested for boundary LEC extrusion. Another possibility is that the apicomedial network creates non-pulsatile tension that helps drive constriction. For instance, in boundary LECs, diffuse apical Sqh activity can be observed, which might contribute to apical area reduction (Companys, 2020).
Interestingly, Rok-RNAi LECs labelled with Sqh::GFP showed a phenotype comparable to that of constricting early boundary LECs in controls. This raises the possibility that the change from early boundary cell behaviour to the behaviour of cells undergoing pulsed contractions during later stages of development might be due to an increase of contractility in these cells over time (Companys, 2020).
That LECs can constrict apically without showing pulsed contractions raises important questions about the role of pulsed contractions in constriction, as also raised by others. LECs begin pulsed contractions while still migrating, at a time when LEC shape changes, and thus tissue remodelling intensifies. Also, for most of morphogenesis, LECs undergo pulsed contractions without apically constricting, merely fluctuating their cell area rhythmically and changing shape . This suggests that pulsed contractions do not drive apical constriction per se. Instead, they might play other roles, such as helping to maintain cell shape and to withstand pushing and pulling forces created by neighbouring cells during morphogenesis. Ultimately, this could help to maintain tissue integrity during morphogenesis. Alternatively, pulsed contractions could cooperate with junctional cortical contractility to create sufficient forces to drive apical constriction more effectively; LEC constriction that is accompanied by pulsed contractions is faster than constriction of boundary LECs without actin foci. To gain further insights into the role of pulsed contractions in LECs, future studies need to consider the interactions between neighbouring LECs to assess the impact of external forces on the behaviour of the apicomedial actomyosin network (Companys, 2020).
This study provides insights into the complexity of the cytoskeleton of a cell during morphogenesis, as well as the importance of the level of cell contractility for the regulation of pulsed contractions, cellular architecture and, consequently, cell behaviour. It also highlights the complex interplay between the junctional cortical and the apicomedial network in changing cell shape and cell area. For LEC migration and the subsequent behavioural transition to constriction, a polarised cytoskeletal network and intermediate contractility levels seem to be crucial. What regulates the transition from migration to constriction is unknown. As LECs transit from PCP to RCP while they change behaviour, studying signals that regulate this change in polarity will be crucial in future studies (Companys, 2020).
Flightless-I is a unique member of the gelsolin superfamily alloying six gelsolin homology domains and leucine-rich repeats. Flightless-I is an established regulator of the actin cytoskeleton, however, its biochemical activities in actin dynamics are still largely elusive. To better understand the biological functioning of Flightless-I, the actin activities of Drosophila Flightless-I were studied by in vitro bulk fluorescence spectroscopy and single filament fluorescence microscopy, as well as in vivo genetic approaches. Flightless-I was found to interact with actin and affects actin dynamics in a calcium-independent fashion in vitro. This work identifies the first three gelsolin homology domains (1-3) of Flightless-I as the main actin-binding site; neither the other three gelsolin homology domains (4-6) nor the leucine-rich repeats bind actin. Flightless-I inhibits polymerization by high-affinity (~nM) filament barbed end capping, moderately facilitates nucleation by low-affinity (~µM) monomer binding, and does not sever actin filaments. This work reveals that in the presence of profilin Flightless-I is only able to cap actin filament barbed ends but fails to promote actin assembly. In line with the in vitro data, while gelsolin homology domains 4-6 have no effect on in vivo actin polymerization, overexpression of gelsolin homology domains 1-3 prevents the formation of various types of actin cables in the developing Drosophila egg chambers. This study also shows that the gelsolin homology domains 4-6 of Flightless-I interact with the C-terminus of Drosophila Disheveled-associated activator of morphogenesis formin and negatively regulates its actin assembly activity (Pinter, 2020).
Epithelial sheets undergo highly reproducible remodeling to shape organs. This stereotyped morphogenesis depends on a well-defined sequence of events leading to the regionalized expression of developmental patterning genes that finally triggers downstream mechanical forces to drive tissue remodeling at a pre-defined position. However, how tissue mechanics controls morphogenetic robustness when challenged by intrinsic perturbations in close proximity has never been addressed. Using Drosophila developing leg, this study shows that a bias in force propagation ensures stereotyped morphogenesis despite the presence of mechanical noise in the environment. Knockdown of the Arp2/3 complex member Arpc5 specifically affects fold directionality while altering neither the developmental nor the force generation patterns. By combining in silico modeling, biophysical tools, and ad hoc genetic tools, these data reveal that junctional myosin II planar polarity favors long-range force channeling and ensures folding robustness, avoiding force scattering and thus isolating the fold domain from surrounding mechanical perturbations (Martin, 2021).
Cadherins are transmembrane adhesion proteins required for the formation of cohesive tissues. Intracellular interactions of E-cadherin with the Catenin family proteins, α- and β-catenin, facilitate connections with the cortical actomyosin network. This is necessary for maintaining the integrity of cell-cell adhesion in epithelial tissues. The supra-molecular architecture of E-cadherin is an important feature of its adhesion function; cis and trans interactions of E-cadherin are deployed to form clusters, both in cis and trans. Studies in Drosophila embryo have also shown that Drosophila E-cadherin (dE-cad) is organized as finite-sized dynamic clusters that localize with actin patches at cell-cell junctions, in continuous exchange with the extra-junctional pool of dE-cad surrounding the clusters. This study used the ectopic expression of dE-cad in larval hemocytes, which lack endogenous dE-cad to recapitulate functional cell-cell junctions in a convenient model system. While dE-cad at cell-cell junctions in hemocytes exhibits a clustered trans-paired organization similar to that reported previously in embryonic epithelial tissue, extra-junctional dE-cad is also organized as relatively immobile nanoclusters as well as more loosely packed diffusive oligomers. Oligomers are promoted by cis interactions of the ectodomain, and their growth is counteracted by the activity of cortical actomyosin. Oligomers in turn promote assembly of dense nanoclusters that require cortical actomyosin activity. Thus, cortical actin activity remodels oligomers and generates nanoclusters. The requirement for dynamic actin in the organization of dE-cad at the nanoscale may provide a mechanism to dynamically tune junctional strength (Chandran, 2021).
Vesicular trafficking defects, particularly those in the autophagolysosomal system, have been strongly implicated in the pathogenesis of Parkinson's disease and related α-synucleinopathies. However, mechanisms mediating dysfunction of membrane trafficking remain incompletely understood. Using a Drosophila model of α-synuclein neurotoxicity with widespread and robust pathology, this study found that human α-synuclein expression impairs autophagic flux in aging adult neurons. Genetic destabilization of the actin cytoskeleton rescues F-actin accumulation, promotes autophagosome clearance, normalizes the autophagolysosomal system, and rescues neurotoxicity in α-synuclein transgenic animals through an Arp2/3 dependent mechanism. Similarly, mitophagosomes accumulate in human α-synuclein-expressing neurons, and reversal of excessive actin stabilization promotes both clearance of these abnormal mitochondria-containing organelles and rescue of mitochondrial dysfunction. These results suggest that Arp2/3 dependent actin cytoskeleton stabilization mediates autophagic and mitophagic dysfunction and implicate failure of autophagosome maturation as a pathological mechanism in Parkinson's disease and related α-synucleinopathies (Sarkar, 2021).
Atxn7, a subunit of SAGA chromatin remodeling complex, is subject to polyglutamine expansion at the amino terminus, causing spinocerebellar ataxia type 7 (SCA7), a progressive retinal and neurodegenerative disease. Within SAGA, the Atxn7 amino terminus anchors Non-stop, a deubiquitinase, to the complex. To understand the scope of Atxn7-dependent regulation of Non-stop, substrates of the deubiquitinase were sought. This revealed Non-stop, dissociated from Atxn7, interacts with Arp2/3 and WAVE regulatory complexes (WRC), which control actin cytoskeleton assembly. There, Non-stop countered polyubiquitination and proteasomal degradation of WRC subunit SCAR. Dependent on conserved WRC interacting receptor sequences (WIRS), Non-stop augmentation increased protein levels, and directed subcellular localization, of SCAR, decreasing cell area and number of protrusions. In vivo, heterozygous mutation of SCAR did not significantly rescue knockdown of Atxn7, but heterozygous mutation of Atxn7 rescued haploinsufficiency of SCAR (Cloud, 2019).
Purification of Atxn7-containing complexes indicated that Atxn7 functions predominantly as a member of SAGA. In yeast, the Atxn7 orthologue, Sgf73, can be separated from SAGA along with the deubiquitinase module by the proteasome regulatory particle. Without Sgf73, the yeast deubiquitinase module is inactive. In higher eukaryotes, Atxn7 increases, but is not necessary for Non-stop/USP22 enzymatic activity in vitro. In Drosophila, loss of Atxn7 leads to a Non-stop over activity phenotype, with reduced levels of ubiquitinated H2B observed (Mohan, 2014). In this study, purification of Non-stop revealed the active SAGA DUBm associates with multi-protein complexes including WRC and Arp2/3 complexes separate from SAGA. SCAR was previously described to be regulated by a constant ubiquitination/deubiquitination mechanism. SCAR protein levels increased upon knockdown of Atxn7 and decreased upon knockdown of non-stop. Decreases in SCAR protein levels in the absence of non-stop required a functional proteasome (Cloud, 2019).
Conversely, overexpression of Non-stop in cells led to increased SCAR protein levels and this increased SCAR protein colocalized to subcellular compartments where Non-stop was found. Nuclear Arp2/3 and WRC have been linked to nuclear reprogramming during early development, immune system function, and general regulation of gene expression. Distortions of nuclear shape alter chromatin domain location within the nucleus, resulting in changes in gene expression. Nuclear pore stability is compromised in SAGA DUBm mutants, resulting in deficient mRNA export. Similarly, mutants of F-actin regulatory proteins, such as Wash, show nuclear pore loss. Non-stop may contribute to nuclear pore stability and mRNA export through multiple mechanisms (Cloud, 2019).
When the basis for this unexpected regulatory mechanism was examined, a series was uncovered of WIRS motifs conserved in number and distribution between flies and mammals. These sequences functionally modulate Non-stop ability to increase SCAR protein levels. Point mutants of each WIRS resulted in less SCAR protein per increase in Non-stop protein. WIRS mutant Non-stop retained the ability to incorporate into SAGA, indicating these are separation of function mutants (Cloud, 2019).
Overall, these findings suggest that the cell maintains a pool of Non-stop that can be made available to act distally from the larger SAGA complex to modulate SCAR protein levels (see Non-stop regulates SCAR protein levels and location). In yeast, the proteasome regulatory particle removes the DUBm from SAGA. In higher eukaryotes, caspase-7 cleaves Atxn7 at residues which would be expected to release the DUBm, although this remains to be shown explicitly. The mechanisms orchestrating entry and exit of the DUBm from SAGA remain to be explored (Cloud, 2019).
Rho family GTPases regulate both linear and branched actin dynamics by activating downstream effectors to facilitate the assembly and function of complex cellular structures such as lamellipodia and contractile actomyosin rings. Wiskott-Aldrich Syndrome (WAS) family proteins are downstream effectors of Rho family GTPases that usually function in a one-to-one correspondence to regulate branched actin nucleation. In particular, the WAS protein Scar/WAVE has been shown to exhibit one-to-one correspondence with Rac GTPase. This study shows that Rac and SCAR are recruited to cell wounds in the Drosophila repair model and are required for the proper formation and maintenance of the dynamic actomyosin ring formed at the wound periphery. Interestingly, it was found that SCAR is recruited to wounds earlier than Rac and is still recruited to the wound periphery in the presence of a potent Rac inhibitor. It was also shown that while Rac is important for actin recruitment to the actomyosin ring, SCAR serves to organize the actomyosin ring and facilitate its anchoring to the overlying plasma membrane. These differing spatiotemporal recruitment patterns and wound repair phenotypes highlight the Rac-independent functions of SCAR and provide an exciting new context in which to investigate these newly uncovered SCAR functions (Nakamura, 2023).
Filamins are highly conserved actin-crosslinking proteins that regulate organization of the actin cytoskeleton. As key components of versatile signaling scaffolds, filamins are implicated in developmental anomalies and cancer. Multiple isoforms of filamins exist, raising the possibility of distinct functions for each isoform during development and in disease. This study provides an initial characterization of jitterbug (jbug), which encodes one of the two filamin-type proteins in Drosophila. Jbug antiserum was generated that recognizes all of the spliced forms and reveals differential expression of different Jbug isoforms during development, and a significant maternal contribution of Jbug protein. To reveal the function of Jbug isoforms, new genetic tools were developed, including a null allele that deletes all isoforms, hypomorphic alleles that affect only a subset, and UAS lines for Gal4-driven expression of the major isoforms. Using these tools, it was demonstrated that Jbug is required for viability and that specific isoforms are required in the formation of actin-rich protrusions including thoracic bristles in adults and ventral denticles in the embryo. Specific isoforms of Jbug show differential localization within epithelia, and maternal and zygotic loss of jbug disrupts Crumbs (Crb) localization in several epithelial cell types (Chung, 2021).
Structures and machines require smoothening of raw materials. Self-organized smoothening guides cell and tissue morphogenesis and is relevant to advanced manufacturing. Across the syncytial Drosophila embryo surface, smooth interfaces form between expanding Arp2/3-based actin caps and surrounding actomyosin networks, demarcating the circumferences of nascent dome-like compartments used for pseudocleavage. This study found that forming a smooth and circular boundary of the surrounding actomyosin domain requires Arp2/3 in vivo. To dissect the physical basis of this requirement, the interacting networks were reconstituted using node-based models. In simulations of actomyosin networks with local clearances in place of Arp2/3 domains, rough boundaries persisted when myosin contractility was low. With addition of expanding Arp2/3 network domains, myosin domain boundaries failed to smoothen, but accumulated myosin nodes and tension. After incorporating actomyosin mechanosensitivity, Arp2/3 network growth locally induced a surrounding contractile actomyosin ring that smoothened the interface between the cytoskeletal domains, an effect also evident in vivo. In this way, a smooth structure can emerge from the lateral interaction of irregular active materials (Sharma, 2021).
Most actin-related proteins (Arps) are highly conserved and carry out well-defined cellular functions in eukaryotes. However, many lineages like Drosophila and mammals encode divergent non-canonical Arps whose roles remain unknown. To elucidate the function of non-canonical Arps, this study focused on Arp53D, which is highly expressed in testes and retained throughout Drosophila evolution. Arp53D localizes to fusomes and actin cones, two germline-specific actin structures critical for sperm maturation, via a unique N-terminal tail. Surprisingly, it was found that male fertility is not impaired upon Arp53D loss, yet population cage experiments reveal that Arp53D is required for optimal fitness in Drosophila melanogaster. To reconcile these findings, focus was placed on Arp53D function in ovaries and embryos where it is only weakly expressed. Under heat stress Arp53D-knockout (KO) females lay embryos with reduced nuclear integrity and lower viability; these defects are further exacerbated in Arp53D-KO embryos. Thus, despite its relatively recent evolution and primarily testis-specific expression, non-canonical Arp53D is required for optimal embryonic development in Drosophila (Schroeder, 2021).
Polarization of the actin cytoskeleton is vital for the collective migration of cells in vivo. During invasive border cell migration in Drosophila, actin polarization is directly controlled by the Hippo signaling complex, which resides at contacts between border cells in the cluster. This study identified, in a genetic screen for deubiquitinating enzymes involved in border cell migration, an essential role for nonstop/USP22 in the expression of Hippo pathway components expanded and merlin. Loss of nonstop (not) function consequently leads to a redistribution of F-actin and the polarity determinant Crumbs, loss of polarized actin protrusions, and tumbling of the border cell cluster. Nonstop is a component of the Spt-Ada-Gcn5-acetyltransferase (SAGA) transcriptional coactivator complex, but SAGA's histone acetyltransferase module, which does not bind to Expanded or Merlin, is dispensable for migration. Taken together, these results uncover novel roles for SAGA-independent nonstop/USP22 in collective cell migration, that may help guide studies in other systems where USP22 is necessary for cell motility and invasion (Badmos, 2021).
This study reports that Drosophila USP22, encoded by not, is necessary for F-actin polarity and collective cell migration of invasive BCs. Collective BC migration requires actomyosin polymerization and contraction at the cortex around the cluster as it moves over the nurse cell substrate; F-actin is effectively excluded from the center of the cluster where polarity determinants acting via the Hippo complex block the activity of the F-actin regulator Enabled. Not has been reported to regulate the actin cytoskeleton directly by promoting the stability of Scar/WAVE. However, this study did not observe a reduction in Scar levels in not mutant clones, and scar loss of function did not disrupt F-actin polarity. Furthermore, no significant change was observed in the number of actin protrusions following not loss of function. This might be expected if Scar were a target in BCs. Interestingly, scar RNAi weakly suppressed not loss of function, suggesting that accumulation of branched actin, mediated by Scar at BC-BC junctions, may contribute to disrupted cell polarity and impaired migration. The data suggest that not regulates inside-out F-actin polarity by regulating the expression of Hippo signaling components ex and mer, that are direct Not targets, in a yki-independent manner. Reanalysis of ChIP-Seq data from embryos indicates that Not and Ada2b bind other core Hippo pathway components, so expression of multiple components may be affected by loss of SAGA components. However, ex and mer are targets for Not, but not Ada2b, which is largely dispensable for migration. Notably, this study found that overexpression of ex suppressed not1-induced F-actin accumulation at inner BC junctions, consistent with partial restoration of Hippo function and inhibition of Enabled function. It was also observed that cpb overexpression rescued loss of not, again consistent with disruption of Enabled function due to competitive binding of Cpb to F-actin barbed ends and the inhibition of F-actin polymerization at inner BC junctions. Incomplete rescue of not1 with overexpressed ex or cpb means that other parallel downstream targets that contribute to not function may exist. Interestingly, the data suggest that not is dispensable in polar cells for BC migration. It will be interesting to examine whether the requirement for not in Hippo pathway function is limited to situations where the Hippo complex acts in a yki-independent fashion. The nature of putative noncell autonomous signaling mediated by not controlling polar cell number remains to be elucidated, but altered signaling may be an indirect consequence of changes in polarity or via direct changes in the expression of affected signaling molecules (Badmos, 2021).
A striking effect of not loss of function in BCs is the redistribution of Crb from inner to outer BC junctions. When possible effects of this on other polarity determinants were examined, it was found that localization of aPKC to the inside apical junction between BCs was disrupted, consistent with studies showing that Crb, acting together with the Par complex and endocytic recycling machinery, is necessary for ensuring its correct distribution. Mislocalized aPKC generates protrusions at the side and back of BCs, just as were seen in not1 clusters. Why is Crb mislocalized to the cortex of the BC complex? Complementation experiments suggest that this is partially accounted for by loss of expression of the FERM domain proteins Ex and Mer, which in follicle cells act together with Moe to recruit Crb to the apical surface. Moe stabilizes Crb at the apical membrane of epithelia by linking Crb to cortical actin. Although the physical interaction between Moe and Crb may be weak, Moe is an important regulator of dynamic Crb localization because it acts to antagonize interactions between Crb and aPKC at the marginal zone of the apical membrane domain while stabilizing interactions between Crb and the apical surface. Importantly, in BCs, Moe is cortically localized where it organizes a supercellular actin cytoskeleton network and promotes cortical stiffness. An attractive hypothesis, therefore, is that Moe, along with other proteins, is a sink for Crb at the cortex of the BC cluster following loss of Ex and Mer at inner BC junctions in not mutants. When ex was overexpressed, the normal pattern of Crb localization was partially restored in support of there being competitive binding. Interestingly, weak rescue of Crb localization was also observed following Cpb overexpression. This might be because Moe, or other proteins that tether Crb on the outer membrane, is only accessible in the absence of a strong supercellular F-actin cortex and that restoration of cortical F-actin in not1 cpb+ cells displaces Crb. In WT BCs, Crb needs to be constantly moved from the outside membrane in a dynamin- and Rab5-dependent manner. Another possibility therefore, which is not mutually exclusive from the first, is that polarization of the F-actin cytoskeleton is important for correct trafficking of Crb in BCs as it is in follicle cells (Badmos, 2021).
The growth, specification, and migration of cells during tissue development requires precisely regulated patterns of gene expression that depend on numerous cues for temporal and spatial gene activation involving crosstalk with multiple signaling pathways. Strikingly, it has emerged that factors once considered to be ubiquitous regulators of transcription, including the SAGA chromatin-modifying complex, can have specific roles in discrete developmental processes. Although it has been suggested that SAGA is required for all transcribed genes in some contexts, numerous studies have shown that loss of SAGA components affects the expression of only a subset of genes and that different components modulate distinct and overlapping subsets. These differences in expression are likely to explain their different physiological roles; for instance, during female germline development in Drosophila, ada2B affects the expression of many genes and is required for oogenesis, whereas not affects relatively few and is dispensable. Genome-wide ChIP studies indicate that even though both DUB and HAT modules bind the same genes, many of the targets do not require the DUB module for expression, explaining the observed dependencies. These experiments also revealed nonoverlapping sites of chromatin occupancy for the DUB and HAT modules of SAGA in Drosophila, but the significance of differences in transcriptional targeting for cell function had not been established. Notably, in this respect, this study found that the requirement for not in BC migration is not matched by a requirement for HAT components, including ada2b or gcn5. Furthermore, Ada2b has not been found to bind the ex and mer promoters, providing a molecular explanation for not's SAGA-independent role. Importantly, these findings challenge the perceived view that transcriptional roles for not/USP22 are mediated solely by SAGA. This may have broader relevance to situations where USP22, but not other members of SAGA, is associated with human disease states, particularly where cell polarity is frequently disrupted, such as cancer. Current efforts are directed at identifying SAGA-independent factors that facilitate Not's chromatin binding and function (Badmos, 2021).
Assemblies of actin and its regulators underlie the dynamic morphology of all eukaryotic cells. To understand how actin regulatory proteins work together to generate actin-rich structures such as filopodia, the localization was analyzed of diverse actin regulators within filopodia in Drosophila embryos and in a complementary in vitro system of filopodia-like structures (FLSs). The composition of the regulatory protein complex where actin is incorporated (the filopodial tip complex) is remarkably heterogeneous both in vivo and in vitro. The data reveal that different pairs of proteins correlate with each other and with actin bundle length, suggesting the presence of functional subcomplexes. This is consistent with a theoretical framework where three or more redundant subcomplexes join the tip complex stochastically, with any two being sufficient to drive filopodia formation. An explanation is provided for the observed heterogeneity and it is suggested that a mechanism based on multiple components allows stereotypical filopodial dynamics to arise from diverse upstream signaling pathways (Dobramysl, 2021).
The regulation of actin polymerization is crucial for numerous cell functions, including cell migration, adhesion, and epithelial closure and is often disrupted in disease, such as cancer metastasis and intracellular infection by pathogens. Micron-scale actin superstructures and their associated regulators form transient membrane-bound complexes that orchestrate large-scale cytoskeletal remodeling and provide the mechanical infrastructure for the cell. One of the best examples is filopodia, with their characteristic membrane-associated 'tip complex' where new actin monomers are incorporated, leading to rapid extension of the filopodia from the cell surface. The tip complex contains many components, including formins such as diaphanous-related formin 3 (Diaph3), barbed-end polymerases Enabled (Ena), vasodilator-stimulated phosphoprotein (VASP), actin bundling proteins including Fascin, and the molecular motor myosin X. There are currently three main models for filopodia formation, each identifying specific tip complex proteins as the key players: (1) formins mediating de novo actin nucleation; (2) a preexisting actin network generated by the Arp2/3 complex becoming bundled by Fascin; and (3) membrane-bound adaptor proteins recruiting Ena/VASP, which could coexist with either formin or Arp2/3 complex-based mechanisms. One way to reconcile these models is to postulate the existence of subtypes of filopodia on the basis of their mechanism of formation. What is not yet clear is whether the subtypes reflect differences between cell types or coexist in the same cell and whether they impart particular properties to the growing filopodia. This question was recently examined by measuring whether the amount of Ena and VASP at the tip complex correlated with the protrusion velocity of filopodia, using cultured Xenopus retinal ganglion cells. A correlation was observed in only a subset of filopodia, suggesting that the accumulation of Ena/VASP proteins is not essential and there are diverse molecular mechanisms that lead to filopodial elongation (Dobramysl, 2021).
This study comprehensively analyzed the role of heterogeneity in the filopodial tip complex. By measuring endogenously tagged actin regulators in Drosophila, similar heterogeneity to exogenous expression in Xenopus retinal ganglion cells was confirmed. This study found that a cell-free system of filopodia-like structures (FLSs) is characterized by similar heterogeneities, and it allowed making of large-scale combinatorial measurements of the correlations of actin regulators with each other and with the morphology of the actin bundle. The emergence of FLSs and their resulting lengths are remarkably insensitive to the presence or absence of any individual tip complex protein. By measuring the momentary rates of growth and shrinkage of the actin bundle and incorporating theoretical modeling, a simple theory was identified that suggests a mechanistic role for tip complex heterogeneity, and its predictions were tested in vitro and in vivo. This work explains how diverse combinations of tip complex proteins give rise to filopodia (Dobramysl, 2021).
This study found that actin regulatory proteins form heterogeneous semidynamic assemblies on membranes composed of at least three or four different subcomplexes where actin bundles nucleate and grow. The resulting actin bundles grow and shrink with velocities that fall on a Laplace distribution, which results in exponentially distributed filopodial lengths. Using the mathematics governing probability distributions, it was possible to link the observed velocity distributions to pairs of fluctuating actin regulators. The subcomplexes are reminiscent of proteins and interactions that were previously thought to be important in filopodium formation. Cdc42-GTP was most highly correlated with VASP and N-WASP; Ena and VASP correlated with each other, and Diaph3, previously implicated in de novo filopodia nucleation, correlated with membrane-adaptor protein TOCA-1, although not with Cdc42-GTP. However, with the complex composition of the extracts and multiple interaction partners for all the proteins involved, it is not yet concluded that no correlation means no relevant interaction (Dobramysl, 2021).
Previous theoretical work considered actin filament length distributions resulting from monomer addition-removal processes together with fragmentation driven by gelsolin and how length control can emerge from other properties of cytoskeletal regulation (such as limited monomer availability, active transport of monomers, capping protein, and formin inhibitors). In contrast, long-tailed exponential length distributions were observed both in vitro and in vivo, suggesting that stochastic processes are governing filopodial dynamics. The primary result shows that FLS and filopodial growth velocities follow a Laplace distribution. These observations are not compatible with simple monomer addition/removal processes, yet still point to a simple emergent dynamic arising from molecular complexity. The fluctuations of components on which the theory depends may originate from many different biochemical possibilities. For example, ubiquitination cycles of VASP have been observed to alter its dynamics within the tip complex, together with filopodial properties, downstream of netrin-1 signaling. Other possible molecular candidates include phosphorylation cycles, GTP/GDP exchanges, or specific protein-protein interactions (Dobramysl, 2021).
The heterogeneity reported resembles observations made for clathrin-mediated endocytosis in mammalian cells and components of the adhesome present in filopodia, suggesting that a similar mechanism based on a heterogeneity of multiple players is a more general feature of cell regulation. The redundancy in molecular composition allows a robustness and may also allow a variety of upstream and downstream components to intersect with the control of filopodia and co-opt them in diverse biological contexts. A multicomponent system could also ensure that signals regulating filopodia must be multiple and coincident, as only rarely will a single input be sufficient to cause an effect, and it takes an overexpression scenario to subvert the normal homeostatic mechanisms, such as Fascin in cancer. In FLSs, the membrane interactions together with SH3 domain and proline-rich regions in Ena, N-WASP, VASP, and Diaph3 are similar to observations with N-WASP and Nck in purified systems that have phase separation properties. It may be that a Laplace-distributed output and the harnessing of fluctuations is the reason for such organization. This study shows that in spite of a dynamic and heterogeneous tip complex, a constraint emerges in the resulting activity, which may be what allows actin machinery to be co-opted in a stereotypical manner, accommodating different tissue regulatory programs without any alteration to its underlying functional properties (Dobramysl, 2021).
The infiltration of immune cells into tissues underlies the establishment of tissue-resident macrophages and responses to infections and tumors. Yet the mechanisms immune cells utilize to negotiate tissue barriers in living organisms are not well understood, and a role for cortical actin has not been examined. This study found that the tissue invasion of Drosophila macrophages, also known as plasmatocytes or hemocytes, utilizes enhanced cortical F-actin levels stimulated by the Drosophila member of the fos proto oncogene transcription factor family (Dfos, Kayak). RNA sequencing analysis and live imaging show that Dfos enhances F-actin levels around the entire macrophage surface by increasing mRNA levels of the membrane spanning molecular scaffold tetraspanin TM4SF, and the actin cross-linking filamin Cheerio, which are themselves required for invasion. Both the filamin and the tetraspanin enhance the cortical activity of Rho1 and the formin Diaphanous and thus the assembly of cortical actin, which is a critical function since expressing a dominant active form of Diaphanous can rescue the Dfos macrophage invasion defect. In vivo imaging shows that Dfos enhances the efficiency of the initial phases of macrophage tissue entry. Genetic evidence argues that this Dfos-induced program in macrophages counteracts the constraint produced by the tension of surrounding tissues and buffers the properties of the macrophage nucleus from affecting tissue entry. This study thus identifies strengthening the cortical actin cytoskeleton through Dfos as a key process allowing efficient forward movement of an immune cell into surrounding tissues (Belyaeva, 2022).
Cells extend membrane protrusions like lamellipodia and filopodia from the leading edge to sense, to move and to form new contacts. The Arp2/3 complex sustains lamellipodia formation, and in conjunction with the actomyosin contractile system, provides mechanical strength to the cell. Drosophila p53-related protein kinase (Prpk), a Tsc5p ortholog, has been described as essential for cell growth and proliferation. In addition, Prpk interacts with proteins associated to actin filament dynamics such as α-spectrin and the Arp2/3 complex subunit Arpc4. This study investigated the role of Prpk in cell shape changes, specifically regarding actin filament dynamics and membrane protrusion formation. Reductions in Prpk alter cell shape and the structure of lamellipodia, mimicking the phenotypes evoked by Arp2/3 complex deficiencies. Prpk co-localize and co-immunoprecipitates with the Arp2/3 complex subunit Arpc1 and with the small GTPase Rab35. Importantly, expression of Rab35, known by its ability to recruit upstream regulators of the Arp2/3 complex, could rescue the Prpk knockdown phenotypes. Finally, the requirement of Prpk was evaluated in different developmental contexts, where it was shown to be essential for correct Arp2/3 complex distribution and actin dynamics required for hemocytes migration, recruitment, and phagocytosis during immune response (Molina, 2022).
In the early syncytial Drosophila embryo, rapid changes in filamentous actin networks and membrane trafficking pathways drive the formation and remodeling of cortical and furrow morphologies. Interestingly, genomic integrity and the completion of mitoses during cell cycles 10-13 depends on the formation of transient membrane furrows that serve to separate and anchor individual spindles during division. While substantial work has led to a better understanding of the core network components that are responsible for the formation of these furrows, less is known about the regulation that controls cytoskeletal and trafficking function. The DOCK protein Sponge was one of the first proteins identified as being required for syncytial furrow formation, and disruption of Sponge deeply compromises F-actin populations in the early embryo, but how this occurs is less clear. Quantitative analysis was performed of the effects of Sponge disruption on cortical cap (actin structures at the apical surface) growth, furrow formation, membrane trafficking, and cytoskeletal network regulation through live-imaging of the syncytial embryo. Membrane trafficking was found to be relatively unaffected by the defects in branched actin networks that occur after Sponge disruption, but Sponge was found to act as a master regulator of a diverse cohort of Arp2/3 regulatory proteins. As DOCK family proteins have been implicated in regulating GTP exchange on small GTPases, it is also suggested that Rac GTPase activity bridges Sponge regulation to the regulators of Arp2/3 function. Finally, the phasic requirements were demonstrated for branched F-actin and linear F-actin networks in potentiating furrow ingression. In total, these results provide quantitative insights into how a large DOCK scaffolding protein coordinates the activity of a variety of different actin regulatory proteins to direct the remodeling of the apical cortex into cytokinetic-like furrows (Henry, 2022).
This work demonstrates the F-actin networks that support the phased ingression of syncytial furrows, while also revealing that Sponge is critically required for the recruitment of several Arp2/3 regulators to the apical and furrow-supporting cortex in the early Drosophila blastoderm. Sponge knockdown causes the mislocalization as well as altered cortical levels of Arp2/3 subunits and Arp regulators such as Scar, Coronin, Pod1, and Cortactin (see Proposed model of Sponge activity in syncytial embryos). Sponge regulation of F-actin is essential for the transition of these regulators from caps onto apical regions of the growing furrows, as Cortactin, DPod1, Coronin, and Scar are absent on ingressing furrows in Sponge knockdown embryos. This leads to inadequate branched actin network function resulting in short, broad furrows that extend no longer than 2.1 μm in length through cycle 13, and small residual cap-like structures, as small as 33% the wild-type cap area. Based on these data, a mechanism is proposed in which Sponge regulates and recruits Scar, Coronin, and DPod1 to the apical caps, while antagonizing Cortactin localization. These proteins in turn modulate filamentous actin and activate Arp2/3 activity and/or stability. As the new apical actin cap forms and expands, Arp2/3 and its regulators remain associated with the branched Actin network and are present in the ring-like structure at the transition point from cap to furrow. This appears necessary for proper linear F-actin nucleation and polymerization by Diaphanous which enables the building of sufficiently extended furrows in each syncytial cycle (Henry, 2022).
Successful cell division relies on an ingressing plasma membrane furrow physically separating neighboring nuclei. In the early Drosophila blastoderm syncytium, as successive cell cycles progress and nuclei become more densely packed, the risk of chromosomal missegregation or mitotic collapse rises if furrows do not adequately segregate neighboring nuclei or provide appropriate anchoring points for spindles. Previous work has shown that furrow length is negatively correlated with mitotic defects. In Sponge embryos, where furrows do not ingress past ~2 μm, severe missegregation in the syncytial stages causes embryos to fail to survive past cellularization (cycle 14). In control embryos, segregation defects are avoided by building furrows in two phases, Ingression I and Ingression II, with a Stabilization period juxtaposed in between. Sponge knockdown does not affect the biphasic nature of furrow ingression, as each cycle maintains two separate ingression periods as well as a measurable stabilization phase that is not significantly different in duration than in control embryos. However, the rate of ingression that Sponge furrows reach in any given ingression phase is slower than in control embryos. This reveals that Sponge does not affect select portions of furrow ingression, but is acting on all phases that promote invagination of the plasma membrane. It also remains a possibility that defects in actin and/or early ingression events may indirectly affect the degree to which later phases such as Ingression II proceeds, though Sponge protein, and the actin and/or nucleator regulatory proteins (ANRP) proteins it regulates, are present at caps and furrows throughout the syncytial cycle. Whether direct or indirect, further evidence of the requirement for Sponge during both early and late furrow phases is observed in measurements of the width of syncytial furrows. As furrows transition from Ingression I to Ingression II, furrows change in morphology from a broad and diffuse appearance to very sharply delineated furrows. In Sponge embryos, furrows possess a broader morphology throughout cycles 10-13, but still partially sharpen as later cycles transition into Ingression II phasic behaviors. Together, these data show that furrow ingression after Sponge disruption occurs in a predictable phasic pattern but that the formation, efficiency, and organization of the furrow is compromised, suggesting that Sponge acts as a master regulator of the furrow ingression process. As the results suggest that Sponge primarily impacts cortical F-actin cap components, this further indicates the importance of the actin cortex in directing proper furrow ingression dynamics (Henry, 2022).
F-actin levels are dramatically reduced both on apical caps and on furrows in Sponge embryos. As these regions have different contributions from branched and unbranched actin populations (branched is more predominant in cap regions while linear is more strongly present at the furrow) this raises the possibility that Sponge could regulate both pathways of actin polymerization. Reducing linear F-actin populations through dia knockdown results in a shortened furrow phenotype reminiscent of what is seen in Sponge. However, several characteristics of dia furrows indicate they may be controlled by a separate mechanism than those in Sponge disrupted embryos. First, furrows are able to reach a significantly greater maximum depth of 3.5 μm, which also reduces the occurrence of mitotic defects. To achieve this greater length, dia furrows ingress at maximal rates closer to those in control during the early phases of furrow formation. In the Ingression I phases of cycles 12 and 13, Sponge furrows show reduced ingression rates, while dia maximal rates are slightly higher than or equal to control furrows, showing no significant difference. It is only in Ingression II phases that dia maximum ingression rates fall behind control. Similarly, dia furrow morphologies are thinner and sharper than those in Sponge throughout cycles 10-13, and are not significantly different from control furrows during Ingression I phases. It is only in Ingression II phases that dia furrows are significantly wider than control, although they do still transition to a sharper morphology than in respective Ingression I phases. In contrast to Sponge, which is needed for both Ingression I and Ingression II, Dia appears more important for Ingression II phases that are responsible for the bulk of a given cycles maximum furrow length. However, it should be noted that both shRNA-driven disruptions and a Dia genetic allele that was used previously, although similar in phenotype, may only be partial disruptions of function, and thus additional defects could be observed with amorphic loss-of-function approaches (Henry, 2022).
When branched F-actin networks are reduced through Many factors are involved in activating, stabilizing, and otherwise regulating Arp2/3 activity during the syncytial stage of embryogenesis, including Scar, Coronin, DPod1, and Cortactin. Each of these proteins can be found on the apical caps where Arp2/3 mediated branched F-actin is prominent (Xie, 2021). These regulators are all disrupted to varying degrees on caps when Sponge is knocked down. Scar, Coronin, and DPod1 are each reduced after Sponge disruption, with remaining protein mislocalized as random puncta throughout the cytoplasm. Within these cap-like structures, Scar is the most severely diminished; from cycle 10-13, Scar intensity is on average 70% reduced from control embryos. Coronin is the next most severely affected factor with Coronin levels at residual caps being 67% lower in intensity than on control caps. The Coronin-family member DPod1 also shows a 49% reduction over cycles 10-13. DPod1 has been shown to have the strongest impact on overall F-actin and Arp2/3 intensities at the cap, and thus its reduction is likely a major contributing factor to the observed loss of F-actin intensities in Sponge embryos, with changes in Coronin and Scar function also contributing to changes in cap growth rates and sizes. One regulator that does not appear to require Sponge activity for localization to the apical cap, however, is Cortactin. While Cortactin is only present on the small residual structures in Sponge embryos, it is present on these structures in significantly increased intensity levels, suggesting that Sponge and Cortactin may possess an antagonistic relationship. Prior work has demonstrated an inhibitory function of Coronin on Cortactin function, so it may be that Sponge regulation of Coronin in turn affects Cortactin levels at the cortical cap in an opposing fashion. Alternatively, it may be that Sponge affects Cortactin levels through its regulation of small GTPase activity or through its other scaffolding domains (Henry, 2022).
Drosophila trachea is a classical model for analyzing epithelial, especially tubular epithelial biology. This study identified lateral E-cadherin mediated junctions that encircle the cells just basal to the zonula adherens in the larval trachea. The lateral junction is associated with downstream adapters, including catenins, and has a distinct junctional actin cortex. The lateral cortex contributes to the development of a supracellular actomyosin mesh in the late larvae. Establishment of this cytoskeletal structure depends on lateral junction associated Rho1 and Cdc42 GTPases, and Arp and WASP pathways. The supracellular network takes the character of stress fibers along the AP axis in the early hours of pupation. It contributes to the shortening of the epithelial tube albeit in a manner redundant to ECM-mediated compression mechanism. In conclusion, this study shows the in vivo existence of functional lateral adherens junction and suggest a role for it in mediating dynamic cytoskeletal events during tissue scale morphogenesis (Pradhan, 2023).
The actin cytoskeleton is a key determinant of cell and tissue homeostasis. However, tissue-specific roles for actin dynamics in aging, notably brain aging, are not understood. This study shows that there is an age-related increase in filamentous actin (F-actin) in Drosophila brains, which is counteracted by prolongevity interventions. Critically, modulating F-actin levels in aging neurons prevents age-onset cognitive decline and extends organismal healthspan. Mechanistically, autophagy, a recycling process required for neuronal homeostasis, was shown to be disabled upon actin dysregulation in the aged brain. Remarkably, disrupting actin polymerization in aged animals with cytoskeletal drugs restores brain autophagy to youthful levels and reverses cellular hallmarks of brain aging. Finally, reducing F-actin levels in aging neurons slows brain aging and promotes healthspan in an autophagy-dependent manner. These data identify excess actin polymerization as a hallmark of brain aging, which can be targeted to reverse brain aging phenotypes and prolong healthspan (Walker, 2023).
Collective cell movements contribute to tissue development and repair and spread metastatic disease. In epithelia, cohesive cell movements require reorganization of adherens junctions and the actomyosin cytoskeleton. However, the mechanisms that coordinate cell-cell adhesion and cytoskeletal remodeling during collective cell migration in vivo are unclear. This study investigated the mechanisms of collective cell migration during epidermal wound healing in Drosophila embryos. Upon wounding, the cells adjacent to the wound internalize cell-cell adhesion molecules and polarize actin and the motor protein non-muscle myosin II to form a supracellular cable around the wound that coordinates cell movements. The cable anchors at former tricellular junctions (TCJs) along the wound edge, and TCJs are reinforced during wound closure. The small GTPase Rap1 was necessary and sufficient for rapid wound repair. Rap1 promoted myosin polarization to the wound edge and E-cadherin accumulation at TCJs. Using embryos expressing a mutant form of the Rap1 effector Canoe/Afadin that cannot bind Rap1, it was found that Rap1 signals through Canoe for adherens junction remodeling, but not for actomyosin cable assembly. Instead, Rap1 was necessary and sufficient for RhoA/Rho1 activation at the wound edge. The RhoGEF Ephexin localized to the wound edge in a Rap1-dependent manner, and Ephexin was necessary for myosin polarization and rapid wound repair, but not for E-cadherin redistribution. Together, these data show that Rap1 coordinates the molecular rearrangements that drive embryonic wound healing, promoting actomyosin cable assembly through Ephexin-Rho1, and E-cadherin redistribution through Canoe, thus enabling rapid collective cell migration in vivo (Rothenberg, 2023).
Stress-induced cell death, mainly apoptosis, and its subsequent tissue repair is interlinked although knowledge of this connection is still very limited. An intriguing finding is apoptosis-induced proliferation (AiP), an evolutionary conserved mechanism employed by apoptotic cells to trigger compensatory proliferation of their neighboring cells. Studies using Drosophila as a model organism have revealed that apoptotic caspases and c-Jun N-terminal kinase (JNK) signaling play critical roles to activate AiP. For example, the initiator caspase Dronc, the caspase-9 ortholog in Drosophila, promotes activation of JNK leading to release of mitogenic signals and AiP. Recent studies further revealed that Dronc relocates to the cell cortex via Myo1D, an unconventional myosin, and stimulates production of reactive oxygen species (ROS) to trigger AiP. During this process, ROS can attract hemocytes, the Drosophila macrophages, which further amplify JNK signaling cell non-autonomously. However, the intrinsic components connecting Dronc, ROS and JNK within the stressed signal-producing cells remain elusive. This study identified LIM domain kinase 1 (LIMK1), a kinase promoting cellular F-actin polymerization, as a novel regulator of AiP. F-actin accumulates in a Dronc-dependent manner in response to apoptotic stress. Suppression of F-actin polymerization in stressed cells by knocking down LIMK1 or expressing Cofilin, an inhibitor of F-actin elongation, blocks ROS production and JNK activation, hence AiP. Furthermore, Dronc and LIMK1 genetically interact. Co-expression of Dronc and LIMK1 drives F-actin accumulation, ROS production and JNK activation. Interestingly, these synergistic effects between Dronc and LIMK1 depend on Myo1D. Therefore, F-actin remodeling plays an important role mediating caspase-driven ROS production and JNK activation in the process of AiP (Farrell, 2022).
Kinesins constitute a superfamily of microtubule (MT)-based motor proteins, which can perform diverse biological functions in cells such as transporting vesicle, regulating MT dynamics, and segregating chromosome. Some motors such as kinesin-1, kinesin-2, and kinesin-3 do the activity mainly on the MT lattice, while others such as kinesin-7 and kinesin-8 do the activity mainly at the MT plus end. To perform the different functions, it is required that the former motors can reside on the MT lattice for longer times than at the end, while the latter motors can reside at the MT plus end for long times. This study presents a simple but general theory of the MT-end residence time of the kinesin motor is presented, with which the factors dictating the residence time are determined. The theory is further used to study specifically the MT-end residence times of Drosophila kinesin-1, kinesin-2/KIF3AB, kinesin-3/Unc104, kinesin-5/Eg5, kinesin-7/CENP-E, and kinesin-8/Kip3 motors, with the theoretical results being in agreement with the available experimental data (Xie, 2023).
Actin filament polymerization can be branched or linear, which depends on the associated regulatory proteins. Competition for actin monomers occurs between proteins that induce branched or linear actin polymerization. Cell specialization requires the regulation of actin filaments to allow the formation of cell type-specific structures, like cuticular hairs in Drosophila, formed by linear actin filaments. This study reports the functional analysis of CG34401/pelado, a gene encoding a SWIM domain-containing protein, conserved throughout the animal kingdom, called ZSWIM8 in mammals. Mutant pelado epithelial cells display actin hair elongation defects. This phenotype is reversed by increasing actin monomer levels or by either pushing linear actin polymerization or reducing branched actin polymerization. Similarly, in hemocytes, Pelado is essential to induce filopodia, a linear actin-based structure. This study further showed that this function of Pelado/ZSWIM8 is conserved in human cells, where Pelado inhibits branched actin polymerization in a cell migration context. In summary, these data indicate that the function of Pelado/ZSWIM8 in regulating actin cytoskeletal dynamics is conserved, favoring linear actin polymerization at the expense of branched filaments (Molina-Pelayo, 2022).
The evolution of the ancestral eukaryotic flagellum is an example of a cellular organelle that became dispensable in some modern eukaryotes while remaining an essential motile and sensory apparatus in others. To help define the repertoire of specialized proteins needed for the formation and function of cilia (see Kimball's Cilia and Flagella site or UTMB Cell Biology Topics), comparative genomics was used to analyze the genomes of organisms with prototypical cilia, modified cilia, or no cilia and 200 genes were identified that are absent in the genomes of nonciliated eukaryotes but are conserved in ciliated organisms. Importantly, over 80% of the known ancestral proteins involved in cilia function are included in this small collection. Using Drosophila as a model system, a novel family of proteins (OSEGs: outer segment) was then characterized that is essential for ciliogenesis. Osegs encode components of a specialized transport pathway unique to the cilia compartment and are related to prototypical intracellular transport proteins (Avidor-Reiss, 2004).
Cilia are microtubule-rich, hair-like cellular extensions that perform essential motile and sensory functions. In sperm and in unicellular eukaryotes, a motile form of cilia called flagellum propels cells to their destination, while in epithelial cells, multiple motile cilia beat synchronously to stir extracellular fluid. In vertebrate photoreceptor cells and invertebrate mechano- and chemoreceptor neurons, the entire sensory transduction machinery is housed in a specialized cellular compartment derived from the cilium. This domain, known as the outer segment, is a hallmark of these sensory neurons and an outstanding example of subcellular compartmentalization as a strategy to optimize function. The ancestral nature of the eukaryotic cilia is evident by its presence in organisms from both lineages: D. melanogaster (Dm), H. sapiens (Hs), T. brucei (Tb), and C. reinhardtii (Cr). In three independent events (indicated by red stars), cilia were lost in lineages leading to A. thaliana (At), D. discoideum (Dd), and S. cerevisiae (Sc). Similarly, compartmentalized cilia were lost in P. falciparum (Pf), while motile cilia were lost in C. elegans (Ce) (Avidor-Reiss, 2004).
Given the wide range of cells and tissues that contain cilia, and the extraordinary diversity of roles performed by cilia, a basic question in cell biology is how ciliogenesis is orchestrated and to what extent common mechanisms underlie this process. Cilia formation begins when the basal body, a centriole-related structure, serves as a template for the assembly of the axoneme. This process can proceed through two different mechanisms. In most motile and sensory cilia, the basal body docks to the plasma membrane, and a bud-like structure containing the axoneme and the ciliary membrane projects out from the cell body; since the ciliary membrane and the axoneme are assembled concurrently as a compartment separated from the cell body, this process is referred to as compartmentalized ciliogenesis. In a few cases, however, such as in the sperm cells of Drosophila and the flagella of the parasite Plasmodium, the entire axoneme is first assembled inside the cytosol and only later is either extruded or matures into a flagellum (i.e., cytosolic biogenesis) (Avidor-Reiss, 2004).
Unlike cytosolic biogenesis, the process of compartmentalized ciliogenesis requires that cilia, flagella, and outer segments transport their building blocks -- proteins and metabolites -- from the cell soma. Genetic and biochemical studies in the biflagellated green alga Chlamydomonas have singled out kinesin II, dynein 1b, and 17 additional proteins named intraflagellar transport (IFT) particle proteins as candidate proteins involved in flagella biogenesis. IFT particle proteins are proposed to function as macromolecular rafts traveling up and down the flagellum, via kinesin and dynein, transporting axonemal precursor proteins to their growing tips. Consistent with this postulate, mutations in the Chlamydomonas IFT particle proteins IFT88 and IFT52 produce very short flagella. Similar results are seen in C. elegans mutants defective in the IFT orthologs OSM-5 and OSM-6 (Avidor-Reiss, 2004 and references therein).
This study reports the development of a novel bioinformatics approach to identify genes involved in ciliogenesis. The strategy is based on the hypothesis that the ancestral eukaryote was a ciliated unicellular organism, and that cilia and flagella were independently lost throughout evolution from several eukaryotic groups. By comparing the genomes of ciliated and nonciliated organisms, a collection of candidate genes important for cilia formation and function was identified. In addition, by phylogenetically examining orthologs in organisms with 'compartmentalized' versus 'cytosolic' axonemes, a large subgroup selectively expressed in Drosophila sensory outer segments, but not in sperm, a novel family of proteins (OSEGs: outer segment) essential for compartmentalized ciliogenesis was isolated and characterized. Together, these studies establish a compelling bioinformatics strategy to help decode gene function and lay the foundation for a comprehensive dissection of eukaryotic ciliogenesis and outer segment development (Avidor-Reiss, 2004).
In order to identify specialized genes essential for cilia biogenesis and function, a phylogenetic screen was undertaken that identified genes conserved in the genomes of ciliated organisms but absent in nonciliated eukaryotes. It was reasoned that gene loss can be used as a powerful tactic to map gene function, particularly if the biological process in question (e.g., cilia biogenesis in this case) is conserved in distantly related species and if it was lost more than once during evolution. Eight species were chosen representing the two major lineages of eukaryotic evolution, and which included nodes where cilia were lost or modified during the evolution of eukaryotes (Avidor-Reiss, 2004).
Because Drosophila contains experimentally tractable motile and sensory cilia and has an extensively annotated genome, it was selected as the anchor for these studies. BLAST searches were performed against the proteome of five ciliated (H. sapiens [Hs], C. elegans [Ce], P. falciparum [Pf], C. reinhardtii [Cr], and T. brucei [Tb]) and three nonciliated organisms (A. thaliana [At], S. cerevisiae [Sc], D. discoideum [Dd]) and orthologs were sought of the 14,000 Drosophila genes in each of these species using a 'reciprocal best hit' algorithm. Because the T. brucei and C. reinhardtii genomes are incomplete, a ciliary protein was considered as conserved in Bikonts (ancestorally biciliate eukaryotes) if it was present in either of those two species (Cr/Tb). Similarly, a partial draft of the Dictyostelium discoideum (Dd) proteome is now available; this organism displays exquisite motility, yet it lacks ciliated structures, thus providing a robust bioinformatics counterscreen (Avidor-Reiss, 2004).
Because all ciliated organisms have an axoneme but may differ in their mode of ciliogenesis, or whether they have motile or nonmotile cilia, it is suspected that distinct sets of proteins might be required during biogenesis of the various forms of cilia. Therefore, the screening strategy was applied to four different search routines: (1) to identify genes involved in processes common to all cilia, like axoneme formation, all ciliated versus all nonciliated eukaryotes were compared (i.e., genes conserved in Hs, Dm, Pf, Cr/Tb, and Ce but not in At, Sc, or Dd); (2) to identify genes involved in cilia motility (either of compartmentalized or cytosolic origin), organisms with motile cilia versus those with nonmotile or no-cilia were compared (i.e., genes conserved in Hs, Dm, Pf, and Cr/Tb but not in Ce, At, Sc, or Dd); (3) to identify genes involved in cilia compartmentalization, organisms with compartmentalized cilia biogenesis versus cytosolic biogenesis were compared (e.g., genes conserved in Hs, Dm, Ce, and Cr/Tb but not in Pf, At, Sc, or Dd) and (4) to identify genes that may be unique to organisms that have both motile and compartmentalized cilia, genes were sought that are shared between Drosophila and organisms with prototypical cilia (i.e., Hs and Cr/Tb but not in Ce, Pf, At, Sc, or Dd) (Avidor-Reiss, 2004).
From a total of 121,243 predicted transcriptional units and 141,000 ESTs (ESTs were used in Chlamydomonas due to the lack of an assembled partial proteome), a total of 187 ancestral genes were identified: (1) 16 conserved in all ciliated organisms, but absent in nonciliated (all-cilia subset); (2) 18 present only in organisms with motile cilia (motility subset); (3) 103 common only to organisms with compartmentalized cilia biogenesis (compartment subset), and (4) 50 shared only between organisms with prototypical cilia (both motile and compartmentalized; prototypical-cilia subset) (Avidor-Reiss, 2004).
To evaluate the performance of the screen, it was asked whether known genes implicated in ciliogenesis are indeed enriched in this collection. A search of the literature revealed that there are 36 genes that have been implicated in ciliogenesis in either flies or in other organisms and were part of the likely ancestral repertoire of genes in the primitive eukaryotic cell (e.g., conserved in organisms from both ancestral eukaryotic lineages). This set includes specialized genes whose primary role is in cilia biogenesis and function (e.g., dynein arms, IFTs), as well as genes that may also participate in other cellular processes (e.g., dynein light chains: see Dynein and intracellular transport). Remarkably, 30 out of the 36 known genes (>80%) are included in the 187 ancestral gene collection obtained in the bioinformatics screen; of the remaining six, five also function outside the cilia and were filtered out because they are present in nonciliated organisms (four dynein subunits and myosin VIIA), and one (left/right-dynein) was eliminated because it did not have an ortholog in Tb or Cr (Avidor-Reiss, 2004).
The selectivity of the screen is also illustrated by examining the genes in the motility subset: all six known ciliary genes recovered in this collection, in fact, encode proteins involved in motility (four axonemal dynein subunits, a radial spoke protein, and Mbo2, a protein important for flagella waveform). In addition, of the remaining 12 candidate motility genes in this subgroup, five are specifically expressed in testis, a tissue highly enriched in motile cells. Taken together, these results substantiate the logic of the approach and the search criteria that were used (Avidor-Reiss, 2004).
Of particular interest was the formation of sensory outer segments, therefore focus was placed on the genes in the cilia-compartment subset both as a platform for gene discovery and for dissecting mechanisms of outer segment biogenesis. Curation of the 103 candidates in this group suggested that several may not have a direct role in ciliogenesis, yet they cosegregated with the selection criteria. These included ion channels, signal-transduction components, transcription factors, and metabolic enzymes. In order to extract 'ciliary' genes from this subset, it was demanded that candidates meet two additional search criteria. (1) Many genes involved in sensory cilia formation share an upstream regulatory sequence known as the X box, often at 150 to 50 nucleotides upstream from the translation start site. A general search of the D. melanogaster and C. elegans genomes for the presence of the 14 nucleotides consensus X box motif has demonstrated that this sequence is much too abundant to be used as a primary screen (for instance, 2449 of Dm and 1897 of Ce genes contain such a motif); however, as a secondary screen, it selected 41 candidates from the cilia-compartment subset. Notably, over 90% of the known ciliary genes in the compartment subset (14/15) are included in these 41 genes. (2) Compartmentalized cilia in Drosophila are found only in chemo- and mechano-sensory neurons. Because these neurons are scattered all over the fly body and comprise a minute fraction of the fly cells, available EST databases contain none, or very few, representatives ESTs. Based on this premise, the compartment subset was searched for genes that contained 0-4 ESTs and 48 candidates were identified. Importantly, these 48 candidates contain nearly all of the known ciliary genes in the original collection (13/15). Together, these two secondary screens identified a total of 30 genes that overlapped both the X box and EST filters: these were chosen for biological validation (Avidor-Reiss, 2004).
Genes involved in compartmentalized ciliogenesis should satisfy two important requirements: (1) the genes should be expressed in ciliated sensory cells; (2) the proteins must be essential for outer segment formation or function.
The genes selected in the compartment subset encode members of several protein groups, including IFT proteins, Bardet-Biedl syndrome (BBS)-related polypeptides, C2 domain-containing proteins, small G proteins, a group of 'coil-coil' proteins, and a family of six WD-domain proteins (OSEGs: outer segment). Below a short summary of these families is provided (Avidor-Reiss, 2004).
BBS is a heterogeneous genetic disorder that is characterized by retinal dystrophy, renal malformation, learning disabilities, and obesity. Six BBS genes have been cloned, and several were recently implicated in ciliogenesis. Drosophila has three BBS orthologs, and all three were selected in this screen (BBS1 and BBS8 as part of the compartment subset, and BBS4 as part of the prototypical-cilia subset). Interestingly, this collection also includes two additional proteins (CG5142 and CG4525) sharing a similar domain organization; it is suggested these proteins encode new BBS members (Avidor-Reiss, 2004).
The C2 domain is a 120 amino acid sequence that functions as a Ca2+-dependent membrane-targeting module in proteins involved in signal transduction (e.g., protein kinase C, cytosolic phospholipase A2) or transport processes (e.g., synaptotagmin I, rabphilin). The analysis identified three novel C2 domain-containing proteins (CG18631, CG9227, and CG14870). Given the central role of calcium in regulating cilia function, as well as processes as diverse as membrane fusion, protein transport, and protein breakdown, these are worthy candidates for sensors of the calcium signals. Small G proteins are known to function as universal molecular switches in a wide range of intracellular processes. Recently, Leishmania ARL3 (LdARL-3A) was implicated in flagellum biogenesis. Notably, the screen identified ARL3 and ARL6, two Arf-like proteins, as components of the compartment group. The cilia-compartment subset also contains orthologs of all seven known IFT particle proteins. In addition, this group also contains two novel WD domain-containing proteins (OSEGs) and three novel coiled-coiled candidate IFT members. OSEGs are a family of six related polypeptides sharing the same predicted topology and signature sequences: an N terminus with seven tandem WD repeats (300 residues), a β sheet rich interdomain (300 residues), and multiple TPR-like repeats (tetratricopeptide repeats; 300 residues). WD repeats are 44-60 residue sequence motifs that fold as parts of two adjacent blades of a typically seven blade propeller structure. TPR-like repeats comprise a TPR-related sequence motif that folds into two antiparallel α helices; these in turn assemble into large right-handed helices. WD- and TPR-like-repeats are often found in large macromolecular assemblies and are thought to function as structural platforms for reversible protein-protein interactions (Avidor-Reiss, 2004).
To identify the cells that express the candidate ciliary compartment genes, 15 genes representing the various gene families were selected, plus a control each from the all-cilia (Tctex2) and prototypical-cilia subsets (BBS4), and transgenic flies were generated expressing Gal4 promoter fusions. Individual lines were crossed to flies containing UAS reporters and examined for GFP expression in larvae and adult animals (Avidor-Reiss, 2004).
In Drosophila, there are three types of ciliated
cells: sperm, mechanosensory, and chemosensory neurons. Mechanosensory and chemosensory cilia are assembled through compartmentalized ciliogenesis, while the sperm tail is assembled via cytoplasmic ciliogenesis. Refined specificity is demonstrated in the anatomical sites of expression of all 17 genes: each transgene is restricted to ciliated cells, with BBS4 and the 15 candidate compartment genes expressed exclusively in neurons of mechanosensory and chemosensory organs. The remaining one, Tctex2/LC2 (a dynein light chain subunit from dynein arms and cytosolic dyneins), was also expressed in sperm cells. No other sites of expression were observed for any of the transgenes. Taken together, these results strongly authenticate the bioinformatics strategy, provide a new perspective into the evolution of cilia, and set the foundation for a comprehensive use of this approach in other biological processes (Avidor-Reiss, 2004).
To gain insights into the biology of outer segment biogenesis, mutants defective in candidate cilia-compartment genes were sought. Drosophila mutants with outer segment defects are expected to be mechanosensory defective. Mutagenized F3 lines were screened for the presence of mechanoinsensitive flies and mechanoreceptor currents (MRC) and transepithelial potentials (TEP) were recorded from candidate lines. Mutations that affect the cilia are predicted to show defective MRC. In contrast, mutations that affect the function or development of the support and accessory cells should abolish both the MRC and the TEP. MRCs and TEPs were recorded from multiple bristles in various uncoordinated lines and complementation groups were selected with normal TEP but defective MRC and they were tested in chemo-sensory and sperm motility assays. Two complementation groups with abnormal mechano- and chemosensory responses but normal sperm motility mapped near the location of oseg1 and oseg2, respectively. It is expected that mutant alleles would carry missense or nonsense mutations, and that introduction of the wild-type gene into mutant animals should rescue their behavioral and physiological phenotype. Indeed, oseg1179 and oseg110 alleles had stop codons in oseg1, and the oseg2 allele contained a nonconservative substitution in the oseg2 gene. More importantly, introduction of the wild-type oseg1 and oseg2 genes by germline transformation rescued the uncoordinated and MRC defects of oseg1 and oseg2 mutants (Avidor-Reiss, 2004).
To analyze the phenotype of oseg1 and oseg2 mutants in detail, the ultra-structure of the sensory cilium was examined by EM serial section analyses. Wild-type mechano- and chemosensory dendrites contain a striated rootlet, two basal bodies, a connecting cilium, and the outer segment. oseg1 and oseg2 mutants have normal inner segments and an intact rootlet, basal bodies, and connecting cilium. However, both mutants display dramatic defects in outer segment morphology: In mechanoreceptor neurons, oseg1 has a striking reduction of the distal-most end of the outer segment (the dendritic tip and tubular body), while oseg2 has a total loss of the tubular body. In chemoreceptors, oseg1 and oseg2 both show severely shortened outer segments. Together, these results firmly implicate oseg genes in ciliogenesis, and outer segment formation (Avidor-Reiss, 2004).
HMM analyses and secondary structure predictions indicate that OSEGs are related to α- and β'-coatomer, two proteins involved in intracellular trafficking. Significantly, clathrin heavy chain (Chc) also displays prominent domain similarity to OSEG family members. Because outer segments (and cilia) are separated from the rest of the cell by a connecting cilium, they need to import their proteins from the cell soma and therefore might be expected to require specialized machinery to assemble a functional compartment (Avidor-Reiss, 2004).
If the OSEG proteins were essential for the transport of selective macromolecules into ciliary compartments, they would be expected to meet several criteria. (1) In contrast to structural or signal transduction components of the outer segment, OSEGs should travel in and out of the outer segment, while concentrating primarily at the base of the cilia. This region of the cell is considered the cilium's 'hub', a strategic place between the cell soma and the outer segment, and is hypothesized to function as the site where molecules targeted to the cilium are loaded and transported via the microtubule-based motors. (2) Ciliary cargo should be transported normally from the cell soma to the cilia base of oseg mutants, but it should be unable to enter the cilia and therefore may accumulate near the cilia base (Avidor-Reiss, 2004).
To examine the subcellular localization of OSEG proteins, translational fusions between all six OSEG family members and GFP were engineered. Each fusion protein was then targeted to ciliated sensory cells using a pan-neuronal promoter. In order to mark the position of the cilium, the cells were co-labeled with mab21A6, a monoclonal antibody that labels the base of the cilium at the inner/outer segment boundary. As predicted, all GFP-tagged OSEG proteins localize primarily at the base of the cilium and can be found inside the sensory cilia (Avidor-Reiss, 2004).
To examine transport into outer segments, it was necessary to identify a candidate cargo protein, ideally one that requires either of the available mutants (oseg1 or oseg2). Mechanosensory outer segments contain at their distal-most end a unique microtubule-rich structure known as the tubular body; this is the proposed site of channel anchoring and force generation in mechanosensory bristles. The α-tubulin isoform in the tubular body is encoded by the α1tub84B gene in Drosophila. Therefore, it was hypothesized that α1tub84B would be an ideal OSEG cargo. Overexpression of a plain GFP reporter, or even a membrane tagged-GFP, does not label the outer segment of ciliated neurons. However, if GFP is linked to α1tub84B (i.e., a GFP-α1tub84B fusion protein), it is now co-targeted with tubulin and functions as a robust reporter of α1tub84B transport into the outer segment (Avidor-Reiss, 2004).
Next, the GFP-α1tubulin 84B reporter was introduced into oseg mutant backgrounds and its localization was examined. The GFP-α1tub84B cargo completely
fails to enter the outer segment of oseg2 mutants, but is efficiently transported to the outer segments of controls and oseg1 mutants. Furthermore, EM examination of oseg2 mutant cells revealed a dramatic accumulation of microtubules at the base of the cilium. These results prove that oseg2, but not oseg1, is essential for tubulin transport into the cilium and illustrate an important aspect of OSEGs function: OSEGs may play distinct roles, and different cargo are likely to be matched to specific OSEG members. Notably, the N-terminal WD domains of α-coatomers and clathrin have been implicated in cargo recognition and sorting. The identification of six OSEG members with distinct N-terminal WD domains may provide the structural basis for selective cargo recognition within this family (Avidor-Reiss, 2004).
The bioinformatics approach also identified two kinesin II subunits as cilia-compartment genes. <Kinesin II has been shown to be required for cilia assembly in a variety of organisms and was proposed to function as the anterograde motor carrying cargo from the base of the cilia to its distal tip. If OSEGs mediate the kinesin-based intraciliary transport, and if this transport were specifically required for outer segment formation, it was reasoned that mutations in
Kinesin-like protein at 64D
(klp64D), the central component of Drosophila kinesin II, should generate in vivo phenotypes that resemble oseg defects. Thus, flies were generated defective in klp64D function and mechano- and chemo-sensory physiology and the transport and accumulation of α1tub84B into sensory cilia were examined. klp64D mutant animals share all of the hallmarks of oseg2 mutants: (1) severe chemoinsensitivity, (2) a total loss of mechanoreceptor currents, (3) GFP-α1tub84B completely failing to enter the outer segments, and (4) microtubules dramatically accumulating at the base of the cilia. Furthermore, klp64D animals, just like oseg2 mutants, have an almost complete loss of the tubular body, but have normal basal bodies and connecting cilia; thus, kinesin II is also not essential for the assembly of the proximal ciliary structures, including axoneme components. Together, these results substantiate kinesin II as a critical player in OSEG function and validate the fundamental importance of intraciliary transport in outer segment (compartmentalized cilia) biogenesis (Avidor-Reiss, 2004).
In this study, a novel bioinformatics screen was used, relying on evolutionary gene conservation and gene loss as a paradigm to discover loci selectively involved in cilia formation and function. This strategy efficiently identified a wide spectrum of known ciliary proteins and dramatically enriched the repertoire of candidate ciliary genes. Because of the focus on identifying ciliary genes of the ancestral eukaryotic cell (e.g., by selecting ciliary genes found in both Bikonts and Unikonts lineages), the recovery of genes unique to specific lineages was not expected. However, by using selective combinations of genomes in the search algorithm, it was possible to define and distinguish between genes involved in cilia motility versus cilia compartmentalization: as additional genomes are completed, it should be possible to target new categories (Avidor-Reiss, 2004).
Approximately 200 genes were selected in the four searches described in this paper. The cilia-compartment subset was analyzed in detail and 27 genes were identified as strong ciliary compartment candidates. Fifteen were selected for detailed in vivo expression studies and it was demonstrated that all were specifically expressed in compartmentalized cilia. Many of the genes in the motility and prototypical-cilia subsets were also examined using a spectrum of curation strategies. This analysis identified an additional collection of novel candidate ciliary genes. It will be of great interest to determine whether mutations in the human orthologs of these genes underlie cilia-based sensory, developmental, or reproductive disorders (Avidor-Reiss, 2004).
Ciliary genes that serve multiple cellular functions were not selected in this screen, mainly because they are still present in organisms that have lost ciliated structures. For example, dyneins are critical components of the ciliary motility apparatus, yet many were filtered out in these screens because they are also involved in intracellular transport in nonciliated organisms. Indeed, it is suggested that the reason so few candidate genes were recovered in the 'all ciliated organisms' subgroup is because proteins common to all cilia, like those involved in axoneme assembly, are also required in basic cellular processes and therefore conserved in nonciliated organisms (e.g., α-tubulin, β-tubulin, γ-tubulin, centrin, pericentrin, etc.) (Avidor-Reiss, 2004).
What do cilia-compartment genes do? At a basic level, these genes should encode components of the intraciliary transport system and the cilia pore, a supramolecular structure that forms the gate into the cilia (see Scholey Intraflagellar transport motors in Caenorhabditis elegans neurons). Indeed, the current screen identified all of the known IFT homologs found in Drosophila, including novel OSEG members. By extension, it is suggested that the compartment group also contains the molecular components of the cilia pore complex (Avidor-Reiss, 2004).
This study shows that oseg1 and oseg2 have distinct roles in ciliogenesis, but neither oseg1 nor oseg2, or even kinesin II, are required for formation of the connecting cilium. These results demonstrate that the assembly of outer segment is orchestrated independently of the connecting cilia (and its axoneme). It will be of great interest to determine which cilia-compartment genes have a role in the biogenesis of this structure (Avidor-Reiss, 2004).
OSEGs are characterized by the presence of two major protein-protein interaction domains, WD and TPR repeats, implicated in the assembly of multiprotein complexes. Significantly, the most closely related proteins outside of the family are α- and β'-coatomer, two cargo-carrying proteins intimately involved in intracellular trafficking. Furthermore, clathrin heavy chains display striking domain similarity to the OSEG family: an N terminus, consisting of 7 WD repeats and a C terminus consisting of 35 TPR-like repeats known as CHCR motifs. Interestingly, coatomers and clathrin-mediated transport systems use small G proteins of the Arf subfamilies as regulators of the transport process. Notably, the screen also identified ARL3 and ARL6, two Arf-like proteins, as components of the ciliary compartment group, with ARL6 expression restricted to mechano- and chemo-sensory neurons (Avidor-Reiss, 2004 and references therein).
What do OSEGS do? The Drosophila oseg2 gene shares significant similarity with a 20 amino acid tryptic peptide from Chlamydomonas IFT172. IFTs were originally identified as a group of proteins enriched in the flagella of Chlamydomonas dynein-1b mutants and absent in the flagella of kinesin II mutants. Because anterograde transport is blocked in kinesin mutants, and retrograde transport is abolished in dynein mutants, IFT particle proteins were proposed to function as molecular rafts transporting cargo up and down the axoneme. Multiple lines of evidence strongly support the proposal that OSEGs function as ciliary transport proteins: (1) OSEGs are specifically expressed in ciliated cells, and the proteins are selectively localized to the cilia and cilia base; (2) OSEGs share structural similarity to prototypical intracellular transport proteins (e.g., clathrin, COP1); (3) oseg2 mutants have specific defects in intraciliary transport; (4) Drosophila OSEGS are required for compartmentalized ciliogenesis (sensory cilia) but not for cytosolic ciliogenesis (sperm tail), and (5) flies defective in oseg2 have nearly the same phenotype as mutants defective in klp64D, the ciliary motor. While there is very limited available data on oseg orthologs in Chlamydomonas, several of the oseg orthologs in C. elegans genes map at, or near, the location of worm mutations leading to sensory cilia defects and implicated in cilia formation and maintenance. For example, OSEG2 and OSEG5 are orthologs of OSM-1 and CHE-2, and OSEG1 and OSEG3 are probably orthologs of DAF-10 and CHE-3, respectively. Surprisingly, the integration of these proteins into a group of genes related to the main families of intracellular transport proteins had escaped notice. These results illustrate a common foundation in the organization of intracellular transport systems, whether mediating internalization of surface proteins, transferring cargo between organelles, or delivering components from the cell body to distal ciliary compartments (Avidor-Reiss, 2004 and references therein).
Nuclear shape is different in stem cells and differentiated cells and reflects important changes in the mechanics of the nuclear envelope (NE). The current framework emphasizes the key role of the nuclear lamina in nuclear mechanics and its alterations in disease. Whether active stress controls nuclear deformations and how this stress interplays with properties of the NE to control NE dynamics is unclear. This study addresses this problem in the early Drosophila embryo, in which profound changes in NE shape parallel the transcriptional activation of the zygotic genome. Microtubule (MT) polymerization events are shown to produce the elementary forces necessary for NE dynamics. Moreover, large-scale NE deformations associated with groove formation require concentration of MT polymerization in bundles organized by Dynein. However, MT bundles cannot produce grooves when the farnesylated inner nuclear membrane protein Kugelkern (Kuk) is absent. Although it increases stiffness of the NE, Kuk also stabilizes NE deformations emerging from the collective effect of MT polymerization forces concentrated in bundles. Finally, it is reported that MT-induced NE deformations control the dynamics of chromatin and its organization at steady state. Thus, the NE is a dynamic organelle, fluctuations of which increase chromatin dynamics. It is proposed that such mechanical regulation of chromatin dynamics by MTs might be important for gene regulation (Hampoelz, 2011).
In summary, these results indicate that nuclear shape in Drosophila embryos is not simply determined by nuclear factors that control deformability but instead requires the interplay between active stresses exerted by polymerization of MTs organized in bundles and properties of the NE. Surprisingly, MTs do not shape the NE like a static scaffold that constrains inherent dynamics of the NE. Rather, it was found that MT dynamics is essential. Polymerization of MTs produces small high-frequency fluctuations of the NE but is not capable of large-scale deformations into grooves. Groove formation requires MT polymerization within bundles, a property which depends on Dynein. It is thus proposed that pushing forces emanating from MT polymerization events are the fundamental active process underlying nuclear deformations. However, their organization in bundles is essential for lobulation. Bundling of growing MTs along a stationary core probably increases their ability to produce force. In vitro experiments and simulations showed that MTs in a bundle reach pushing forces much higher than the stall force of individual MTs. Moreover, relaxation of grooves might be facilitated within bundles, as pushing of bundles towards obstacles can facilitate collective catastrophe. Bundle integrity is ensured by cytoplasmic Dynein. Although no enrichment of Dynein was observed at the NE, Dynein could be localized at the NE, where it would allow bundle cohesion and attachment. In Drosophila photoreceptors and other systems, Dynein is recruited to the NE by the LINC (linker of nucleoskeleton and cytoskeleton) complex. The LINC complex consists of a KASH domain protein and a SUN domain protein, and spans the NE. In contrast to Dynein inhibition, reduced activity of the Drosophila LINC complex component CG3287/Klaroid did not affect nuclear morphology. Instead, this enhanced the MT-induced fluctuations of the NE, indicating that MT polymerization forces are less efficiently buffered. By bridging the NE, LINC proteins are believed to dampen cytoskeletal forces at the lamina or heterochromatin. The observations made in this study are consistent with this interpretation (Hampoelz, 2011).
The LINC complex also transmits cytoskeletal forces across the NE to direct chromosome movement during meiosis. In fission yeast, LINC proteins mediate MT- and Dynein-dependant oscillating movements of whole nuclei. This results in nucleoplasmic agitation, which promotes pairing of homologous regions. By analogy, MT-induced fluctuations of the NE could serve as a means to generally enhance chromatin mobility at the onset of zygotic transcription. This would increase the probability of interactions with other loci or the NE and could tune cis-regulatory interactions. The recently established concept of transcription factories where active loci are pulled into pre-assembled sites of mRNA production demands a mobile chromatin. Interestingly, transcription is affected in kuk mutants, including up- and downregulation of predominantly early zygotic genes. Although a specific role in directly regulating these target genes cannot be excluded, a more likely scenario is a global contribution of Kuk to transcription at this stage owing to its effects on NE mechanics and dynamics as well as chromatin mobility. It will be important to study this quantitatively through direct visualization of dynamics at specific loci and transcription with fluorescence in situ hybridization (Hampoelz, 2011).
Although necessary, MT polymerization forces are not sufficient to produce grooves in the NE. These deformations require specific material or structural properties of the NE. This work sheds new light on this process. Comparison of human embryonic stem cells and differentiated cells indicates that deformability is usually increased when stiffness is reduced (and vice versa), for instance owing to absence or knockdown of A-type lamins. A Drosophila A-type lamin is not expressed during cellularization, and nuclear deformability is instead controlled by the farnesylated, INM-protein Kuk. Kuk increases the stiffness of the NE, and it is required for large deformations probably because stiffness is required for the pre-stressed NE to buckle. However, Kuk is likely to control other properties of the NE as its depletion cannot be rescued by elevated levels of the B-type lamin Dm0, which also increase NE stiffness. Likewise, overexpressed Dm0 does not enhance lobulation. Kuk could stabilize transient and small deformations imposed by MTs. Stabilization of NE curvature would work as a ratchet and allows the temporal integration of small polymerization forces contributed by individual MTs in bundles. The relative amounts of MT polymerization forces and NE stiffness would define the threshold above which buckling is possible (Hampoelz, 2011).
Kinesins and dyneins play important roles during cell division. Using RNA interference (RNAi) to deplete individual (or combinations of) motors followed by immunofluorescence and time-lapse microscopy, the mitotic functions were examined of cytoplasmic dynein and all 25 kinesins in Drosophila S2 cells. Four kinesins are involved in bipolar spindle assembly, four kinesins are involved in metaphase chromosome alignment, Dynein plays a role in the metaphase-to-anaphase transition, and one kinesin is needed for cytokinesis. Functional redundancy and alternative pathways for completing mitosis were observed for many single RNAi knockdowns, and failure to complete mitosis was observed for only three kinesins. As an example, inhibition of two microtubule-depolymerizing kinesins initially produced monopolar spindles with abnormally long microtubules, but cells eventually formed bipolar spindles by an acentrosomal pole-focusing mechanism. From this phenotypic data, a model is constructed for the distinct roles of molecular motors during mitosis in a single metazoan cell type (Goshima, 2003).
Before beginning functional analysis, the sequences of all Drosophila kinesin superfamily proteins were identified and analyzed. A BLAST search was performed on the fly database using the conserved motor domain of fly conventional kinesin (1-340 aa). 25 genes emerged as exhibiting significant (E-value < 1e-15) sequence homology. Sequence alignments of the motor and non-motor domains with kinesins from other organisms were used to assign the Drosophila kinesins to different subfamilies. This analysis identified clear subfamilies and mammalian homologues for 21 of the 25 genes. The remaining four are divergent kinesins that have no homology in their tail domains to kinesins in other organisms. Five kinesins may not be present or are expressed at very low levels in S2 cells. Nevertheless, RNAi was performed for all 25 kinesins so as not to miss a potential mitotic involvement of a low copy number kinesin (Goshima, 2003).
RNAi of eight kinesins and cytoplasmic DHC caused a variety of mitotic defects,
including monopolar spindle formation, chromosome misalignment, anaphase delay, and cytokinesis failure. The involvement of other motors in mitosis cannot be ruled out due to several experimental caveats. (1) Due to unavailability of antibodies, the reduction of 16 motors by RNAi could not be confirmed. However, for the 10 motors that were examined, drastic (>90%) protein reductions by RNAi were confirmed. Moreover, similar reductions of 24 other cytoskeleton-related proteins was confirmed by immunoblotting and/or immunostaining after dsRNA treatment, regardless of the appearance of phenotypes, no case has been encountered where protein levels were not dramatically reduced. Based on such results, it is highly likely that all motors were reduced to very low levels by RNAi. (2) Even the small amount of residual protein that remains after RNAi treatment may be sufficient for cellular function. (3) Functional redundancy may obscure a mitotic phenotype for some motors. This could be potentially examined by exhaustive double or triple RNAi for many combinations of motor proteins (Goshima, 2003).
The results provide several general insights into the roles of molecular motors in mitosis. First, cells appear to have redundant or alternate mechanisms for completing mitosis, despite the absence of a kinesin motor. These experiments, for example, uncovered a novel rescue mechanism for converting monopolar to bipolar spindles by an acentrosomal pole-focusing mechanism. By performing multiple RNAi, it could also be show that several kinesin act in a partially redundant manner to ensure congression of chromosomes to the metaphase plate. Only three kinesins appear to be absolutely essential for completing mitosis; Klp61F [BimC/Eg5] and Klp67A [Kip3] RNAi cells could not proceed into anaphase, and Pavarotti [MKLP1]-depleted cells could not execute cytokinesis. The phenotypes from comprehensive RNAi analysis enable a model to be derived for the roles of microtubule-based motors in the sequential steps of mitosis in S2 cells (Goshima, 2003).
The establishment of spindle bipolarity, the first step of mitosis, requires Klp61F [BimC/Eg5], a bipolar homotetrameric kinesin. Time-lapse imaging results are consistent with proposals that Klp61F [BimC/Eg5] cross-links and slides apart antiparallel spindle microtubules (Goshima, 2003).
Ncd [Kin C] depletion causes defects in spindle pole integrity. MTOC fusion is inhibited, and multiple poles, frequently acentrosomal, are produced. Time-lapse observation of GFP-tubulin in Ncd [Kin C] RNAi cells suggests that microtubules are frequently released from the existing pole in prometaphase, and these released microtubules can build additional acentrosomal spindles. Ncd [Kin C] may move toward the microtubule minus ends and physically tether the microtubules to the poles. The S2 Ncd [Kin C] RNAi phenotype is quite different from the Drosophila Ncd [Kin C] null mutant, in which meiotic spindle defects are observed, but spindle formation defects in mitotic cells are either not severe or not observed. Variations in phenotype after inhibition of Kin C kinesins are also evident in comparing results from different experimental systems. For example, antibody inhibition of XCTK2 induced accumulation of asters and half spindle in Xenopus egg extract. It is speculated that Ncd [Kin C] may serve as a primary motor for spindle pole coalescence in S2 cells, whereas in most of the fly tissues, Ncd [Kin C]'s function may be secondary and/or compensated by cytoplasmic dynein (Goshima, 2003).
The regulation of microtubule dynamics by motor proteins is essential for efficient spindle assembly. RNAi of Klp10A [Kin I], a member of the microtubule-destabilizing Kin I kinesins, resulted in excessively long microtubules emanating radially from the MTOC, suggesting that Klp10A [Kin I] destabilizes most, if not all, spindle microtubules. Monopolar spindles were frequently formed, as found in fixed images after protein inhibition of the vertebrate homologue KCM1/MCAK. Live-cell imaging clearly shows that separated MTOCs fuse together after NEB, despite the presence of Klp61F [BimC/Eg5]. Longer than normal microtubules may be physically difficult for Klp61F [BimC/Eg5] to cross-link into antiparallel bundles. Alternatively, pushing forces against the cell cortex exerted by long astral microtubules might overcome the forces generated by Klp61F [BimC/Eg5]. However, Klp10A [Kin I] RNAi cells eventually succeed in forming monastral bipolar spindles, which segregate sister chromatids in anaphase, strongly suggesting that stable kinetochore-spindle interaction can be achieved in the absence of Klp10A [Kin I]. A mitotic function was not detected for the two other Kin I motor proteins (Klp59C and Klp59D), although another analysis involving antibody microinjection found anaphase A defects for Klp59C in fly embryos (Goshima, 2003).
Klp67A [Kip3] was originally localized at mitochondria. However, a mitochondria transport defect after Klp67A [Kip3] RNAi was not detected in S2 cells Instead, RNAi of Klp67A [Kip3] produces long mitotic microtubules, indicating that it destabilizes microtubules. This result is consistent with sequence analysis showing that Klp67A [Kip3] is related to S. cerevisiae Kip3 and Schizosaccharomyces pombe Klp5/Klp6, null mutants of which have longer than normal spindle microtubules. Excessively long microtubules may give rise to monopolar spindle formation in Klp67A [Kip3] RNAi cells by the mechanism described above for Klp10A [Kin I]. However, unlike Klp10A [Kin I], which destabilizes all spindle microtubules, Klp67A [Kip3] appears to act selectively on microtubules between the poles and chromosomes; astral microtubules have normal length in Klp67A [Kip3] RNAi cells. Thus, Klp67A [Kip3] and Klp10A [Kin I] have distinct roles as microtubule-destabilizing proteins in the spindle (Goshima, 2003).
Three and possibly four chromosomal kinesins are important for chromosome movement. Previous reports have shown that the kinetochore-localized CENP-meta [CENP-E] is essential for chromosome congression at early developmental stages. The genetic phenotypes of the two chromosome-associated kinesins, No distributive disjunction [Nod] and Klp3A [chromokinesin], for mitotic chromosome alignment are less evident. Nod [Kid] is important for chromosome positioning in meiosis, but is nonessential for fly viability. Klp3A [chromokinesin] is also nonessential for fly viability, and null mutation causes cytokinesis failure in meiosis. However, the data suggest that these two chromokinesins act redundantly in mitotic cells for prometaphase chromosome movement; double RNAi of Klp3A [chromokinesin] and Nod [Kid] causes more severe chromosome misalignment phenotypes than single RNAi treatments. The survival of adult fly without either Nod [Kid] or Klp3A [chromokinesin] might be due to this redundancy. However, phenotypic analyses show that the actions of these two chromokinesins are distinct. The chromosome arms were abnormally extended along the direction of spindle axis during prometaphase and metaphase after Nod [Kid] RNAi, suggesting that Nod [Kid] functions to transport chromosomal arms away from the pole. In contrast, Klp3A [chromokinesin] may be needed for both kinetochore and arm-directed chromosome motility. The redundant function of chromatin- and kinetochore-localized kinesins for chromosome congression may be a general feature of eukaryotes (Goshima, 2003).
The Klp67A [Kip3] RNAi phenotype also suggests an additional role for this motor in proper chromosome-spindle interaction, most probably at the kinetochore. After depletion of Klp67A [Kip3], chromosomes are scattered in the spindle (in contrast to Klp10A [Kin I] RNAi), possibly reflecting unbalanced spindle tension on chromosomes. Additionally, very few cells enter anaphase, presumably because of activation of the spindle checkpoint that monitors spindle-kinetochore interaction. In S. pombe, two Klp67A [Kip3]-like proteins, Klp5 and Klp6, are localized at mitotic kinetochores, and have been proposed to generate stable kinetochore-spindle interactions and tension at kinetochores. Because RNAi phenotypes of Klp67A [Kip3] are similar in many aspects to Klp5/Klp6 mutants, Klp67A [Kip3] also might act at kinetochores (Goshima, 2003).
In addition to Klp67A [Kip3], cytoplasmic dynein plays a role in the metaphase-to-anaphase transition. Dhc64C
(cytoplasmic dynein) may control the timing of anaphase onset, possibly by transporting Rod or other checkpoint proteins away from kinetochores as proposed for the fly embryo. A similar checkpoint inactivation model was proposed for mammalian dynein/dynactin based on inhibition analyses (Goshima, 2003).
The roles of motors in anaphase A (chromosome to pole motion) and anaphase B (spindle elongation) are less clear from this paper, since no defects were detected by fixed cell images after RNAi. This may be due to the fact that RNAi depletion of a single motor protein may cause kinetic defects in anaphase, which may be difficult to detect in fixed population images. For example, recent live-cell imaging of anaphase in living Drosophila embryos shows that antibodies against Kin I motors slow down rather than completely block anaphase. It is also possible that Klp61F [BimC/Eg5] or Klp67A [Kip3] play roles in anaphase; however, RNAi of these motors arrests cells earlier in mitosis (Goshima, 2003).
Finally, central spindle formation and cytokinesis in S2 cells require Pavarotti [MKLP1]. However, it has not been clear whether Pav is needed for the formation or maintenance of the central spindle. Live-cell observations of Pav [MKLP1] RNAi cells shows that some cells formed central spindles, but could not maintain them, whereas others showed a complete failure in formation (Goshima, 2003).
An interesting phenomenon is the conversion of a monopolar spindle to a monastral bipolar spindle, in which one of the poles lacks a centrosome. Anastral bipolar spindles, in which both poles lack centrosomes, are commonly observed in higher plant mitosis and animal female meiosis, but the formation and relevance of acentrosomal poles in animal mitotic cells have been less clear. It has been found that bipolar spindles containing one centrosomal and one acentrosomal pole can be formed if one centrosome is destroyed by laser ablation. In addition, there has been a previous report of asymmetrical bipolar spindles in nonmeiotic cells) (Goshima, 2003).
The live-cell analysis provides a mechanism for monastral bipolar spindle formation. Initially, microtubules emerge from the chromosomal region in the monopolar spindle. Similar observations of chromosome-directed microtubule formation have been made for acentrosomal spindle formation in meiosis. Simultaneous RNAi experiments show that Klp61F [BimC/Eg5], but not cytoplasmic dynein, is required for monastral bipolar spindle formation. Bipolar kinesins may be necessary for bundling the chromosome-generated microtubules. The role of Ncd [Kin C] during monopolar to bipolar conversion is ambiguous by double the RNAi method, since Ncd [Kin C] depletion dominantly inhibits monopolar spindle formation. However, considering Ncd [Kin C]'s role in MTOC fusion in early prometaphase and pole maintenance, it also may be needed for acentrosomal pole-focusing in the final step of the conversion. Klp67A [Kip3] may also play a minor role in this process because monopolar spindles generated after Klp67A [Kip3] RNAi do not always form an acentrosomal pole, perhaps due to problems in chromosome-microtubule interaction as discussed above (Goshima, 2003).
Monopolar to monastral bipolar spindle conversion also appears to occur in untreated cells as well, because both monopolar (<5% of mitotic cells) and monastral bipolar spindles (10% of bipolar spindles) were observed in untreated S2. Moreover, it has been reported that 0.9% of mitotic spindles of the wild-type Drosophila larval neuroblasts display monastral bipolar spindles. In the monastral bipolar spindle of Klp10A [Kin I] RNAi and untreated cells, anaphase chromosome movement can occur. Thus, the mitotic checkpoint is activated during the monopolar spindle phase, but is properly down-regulated once metaphase is achieved in monastral bipolar spindle. Thus, in the presence of a functional spindle checkpoint system, monastral bipolar spindle formation enables completion of mitosis in rare cases when the spindle gets trapped in a monopolar state after nuclear envelope breakdown. Although originally believed to be a property of plant and meiotic animal cell, it is proposed that acentrosomal pole formation constitutes a general backup mechanism for mitosis in somatic animal cells (Goshima, 2003).
Cytoskeletal organization is central to establishing cell polarity in various cellular contexts, including during messenger ribonucleic acid sorting in Drosophila melanogaster oocytes by microtubule (MT)-dependent molecular motors. However, MT organization and dynamics remain controversial in the oocyte. This study used rapid multichannel live-cell imaging with novel image analysis, tracking, and visualization tools to characterize MT polarity and dynamics while imaging posterior cargo transport. All MTs in the oocyte were highly dynamic and were organized with a biased random polarity that increased toward the posterior. This organization originated through MT nucleation at the oocyte nucleus and cortex, except at the posterior end of the oocyte, where PAR-1 suppressed nucleation. These findings explain the biased random posterior cargo movements in the oocyte that establish the germline and posterior (Parton, 2011).
Despite the importance of MTs in the oocyte, how they are organized and to what extent they are dynamic have remained highly controversial. Moreover, the prevailing models for MT organization have mostly relied on a static view of MT distribution and on indirect measures of polarity, such as the steady-state distribution of motor fusions and cargoes. By using live-cell imaging and developing novel image analysis and global visualization tools, the dynamics and polarity of MTs were characterized directly in living oocytes. MTs were found to form a dynamic cortical network extending into the posterior with a bias in net orientation that increases toward the posterior. It was established that posterior-directed cargo is actively transported on these dynamic MTs, with no evidence for preferential transport by a subpopulation of more stable, posttranslationally modified MTs. Significantly, the magnitude and distribution of the observed bias in cargo movements parallels closely the polarity of the MT network. These findings explain the previously reported subtle biased random transport of posterior cargoes and lead to the proposal of the following model for posterior cargo localization: posterior cargo is transported on the entire dynamic MT network and the overall net bias in MT orientation directs the net movement of cargo to the posterior cap, where it becomes anchored (Parton, 2011).
The results reveal that the establishment of the biased MT network is dependent on a specific distribution of MT nucleation sites around the oocyte cortex, with a critical, PAR-1-dependent exclusion of MT nucleation from the posterior cortex. This extends upon previous observations that PAR-1 affects MT organization, leading to an increased density of MTs at the posterior. By using highly sensitive imaging techniques in live oocytes, it was demonstrated that, in contrast to previous work, in stage 9 oocytes, MT nucleation is also distributed along the anterior and lateral cortexes. Initiation of MTs is predominantly from the anterior of the oocyte with a sharp decrease in nucleation along the posterior two thirds of the oocyte cortex. Those MTs nucleating along the anterior are constrained to grow in a more posterior orientation, whereas nucleation along the lateral cortex is more random in orientation. The combination of these two contributions to the network of MTs present in the oocyte results in a slight excess of plus ends extending in a posterior direction, which increases in magnitude closer to the posterior. Despite the fact that, at the extreme posterior, there are no MT nucleation sites, it was found that, even at the extreme posterior, a percentage of MTs appear to orient toward the anterior. This is caused by MTs bending around as they extend into the posterior. Importantly, this detailed analysis of cargo movements reveals a bias in cargo movement directionality at the posterior that matches precisely the bias in MT orientation (Parton, 2011).
It is interesting to consider how the PAR-1 kinase might prevent the nucleation of MTs at the posterior cortex. The PAR genes are conserved polarity determinants with common functions in a variety of organisms. PAR-1 is known to function in association with other PAR proteins, so it is possible that the other PAR proteins also function together with PAR-1 to inhibit MT nucleation in the oocyte. However, several other factors may also be involved. PAR-1 could affect MTs through its association with Tau, which has been shown in mammalian cells and proposed in the Drosophila oocyte, but this remains contentious, as the presence of Tau is not absolutely required for PAR-1 function. Another possibility is that PAR-1 could act through the components of the γ-TuRC complex or some other MT nucleation components, rather than through a direct affect on MTs. Whatever the molecular mechanism of PAR-1 inhibition of MT nucleation, it is most likely to involve the phosphorylation of a downstream target of the PAR-1 kinase (Parton, 2011).
The live-imaging results highlight the role of a dynamic MT network in establishing cell polarity in the oocyte, in which no stable, posttranslationally modified MTs were detected. This raises the question of why this should be the case when, in some other cells, subsets of either stably bundled or completely stable, posttranslationally modified MTs have been observed and proposed to have functional roles in directing cell polarity. Moreover, in many polarized cell types, including the blastoderm embryo and secretory columnar epithelial cells of egg chambers, MTs are organized with a very strong apical-basal polarity and include stable MTs. This makes functional sense in both cases, as cargoes have to be transported very rapidly either apically or basally. In contrast, in the oocyte, MTs perform three key functions that are not necessarily all compatible with having a very strict apical-basal polarity. First, they provide a means of randomly distributing generic components, such as mitochondria and lipid droplets, throughout the cytoplasm. Second, they provide a network to gather cargoes and redistribute them to distinct intracellular destinations, initiating and maintaining cell polarity. Third, they provide a scaffold that maintains structural integrity. It is proposed that the dynamic, subtly biased network of MTs in the oocyte provides an efficient compromise for dealing with these multiple conflicting biological requirements. During mid-oogenesis, the oocyte undergoes a huge expansion in size, when many cellular components are transported from the nurse cells or secreted from the overlying follicle cells. Although generic cellular components, such as Golgi, mitochondria, and lipid droplets, must be kept distributed throughout the ooplasm, the nucleus and specific mRNAs and proteins must be transported to different poles to establish the embryonic axes. A biased random network of MTs enables the mixing of generic components by continuous transport using molecular motors with opposing polarities, while, at the same time, allowing specific components to be transported by motors with single polarities to the anterior or posterior poles for anchoring. Furthermore, a network of highly dynamic MTs would allow efficient capturing of cargo by the motors throughout the entire ooplasm in a rapidly growing and developing oocyte. The fact that the MT network is highly dynamic also makes considerable functional sense for such a rapidly developing system. The MT cytoskeleton is reorganized extensively during Drosophila oogenesis but most dramatically during stage 7. This fits well with observations in other cell types showing that MTs are highly dynamic in nature and are often reorganized to direct cellular polarization (Parton, 2011).
MTs certainly play critical roles in driving cell polarization and extension in many kinds of eukaryotic cells, for example, during guidance of extending neuronal growth cones, in migratory cells, in dorsal closure, and in fields of bristles with planar polarity in fly wings. In all these cases, the polarity and dynamics of MTs have tended to be studied quite crudely because of an inability to follow the subtleties of global MT polarity and dynamics. Therefore, it is highly likely that MTs in such cells are more complex and subtle than previously thought. Interestingly, at least in Xenopus laevis oocytes in which hook decoration and EM were used as the previous gold standard for determining the polarity of individual MTs, a network of MTs is nucleated at the cortex, leading to a bias of polarity rather than an absolute polarity. The methods used in this study are significantly easier to apply technically than hook decoration methods and are, therefore, more generally applicable to study the orientation and dynamics of MTs in most cells. For example, it has been possible to apply these analysis tools to examine subtleties of MT organization in migratory border cells (Parton, 2011).
It is proposed that during cellular reorganization and repolarization, as in the oocyte, the establishment of a dynamic, subtly biased MT network is a widespread phenomenon and provides a general mechanism by which strong cell polarity can be initiated and maintained while efficiently handling the transport requirements of cargoes distributed throughout the cytoplasm. The tools developed in this study to quantitate global or local bias in a complex field of MTs can now be applied widely to other oocytes and other cell types to test the generality of the proposed biased random model for MT organization (Parton, 2011).
Epithelial tissues are lined with a sheet-like basement membrane (BM) extracellular matrix at their basal surfaces that plays essential roles in adhesion and signaling. BMs also provide mechanical support to guide morphogenesis. Despite their importance, little is known about how epithelial cells secrete and assemble BMs during development. BM proteins are sorted into a basolateral secretory pathway distinct from other basolateral proteins. Because BM proteins self-assemble into networks, and the BM lines only a small portion of the basolateral domain, it was hypothesized that the site of BM protein secretion might be tightly controlled. Using the Drosophila follicular epithelium, this study shows that kinesin-3 and kinesin-1 motors work together to define this secretion site. Similar to all epithelia, the follicle cells have polarized microtubules (MTs) along their apical-basal axes. These cells collectively migrate, and they also have polarized MTs along the migratory axis at their basal surfaces. This study found follicle cell MTs form one interconnected network, which allows kinesins to transport Rab10+ BM secretory vesicles both basally and to the trailing edge of each cell. This positions them near the basal surface and the basal-most region of the lateral domain for exocytosis. When kinesin transport is disrupted, the site of BM protein secretion is expanded, and ectopic BM networks form between cells that impede migration and disrupt tissue architecture. These results show how epithelial cells can define a subdomain on their basolateral surface through MT-based transport and highlight the importance of controlling the exocytic site of network-forming proteins (Zajac, 2022).
In epithelial tissues, polarisation of microtubules and actin microvilli occurs along the apical-basal axis of each cell, yet how these cytoskeletal polarisation events are coordinated remains unclear. This study examines the hierarchy of events during cytoskeletal polarisation in Drosophila and human epithelia. Core apical-basal polarity determinants polarise the Spectrin cytoskeleton to recruit the microtubule-binding proteins Patronin (CAMSAP1/2/3 in humans) and Shortstop (Shot; MACF1/BPAG1 in humans) to the apical membrane domain. Patronin and Shot then act to polarise microtubules along the apical-basal axis to enable apical transport of Rab11 endosomes by the Nuf-Dynein microtubule motor complex. Finally, Rab11 endosomes are transferred to the MyoV actin motor to deliver the key microvillar determinant Cadherin99C to the apical membrane to organise the biogenesis of actin microvilli (Khanal, 2016).
These results reveal a mechanism linking determinants of cell polarity with stepwise polarisation of the spectrin cytoskeleton, microtubule cytoskeleton and biogenesis of actin microvilli through apical trafficking of Cad99C. The results suggest that polarisation of the apical spectrin βH-Spectrin is dependent on polarity determinants, likely through interactions with the FERM domain proteins and the apical polarity determinant Crb. The spectraplakin Shot is highly similar to βH-Spectrin, and is able to bind to and colocalise with it at the apical domain of epithelial cells, suggesting that the two proteins might have a similar function. βH-Spectrin is linked to microtubules through Patronin, whereas Shot can directly bind microtubules. Consequently, redundancy is anticipated between βH-Spectrin and Shot, or between Patronin and Shot. Accordingly, this study found that mutation of βH-spectrin only had a mild phenotype, whereas mutation of α-spectrin simultaneously disrupted both pairs of proteins in parallel and caused a drastic phenotype, completely disrupting the apical trafficking of Cad99C and microvillar biogenesis. More importantly, double mutants for shot and βH-spectrin had a more severe effect on microtubule and Cad99C localisation than either alone, therefore demonstrating that the two proteins act in a redundant fashion (Khanal, 2016).
Downstream of the spectrin cytoskeleton, Patronin and Shot are required in parallel to drive apical-basal polarisation of microtubules, which is then responsible for orienting the apical transport of Cad99C, within Rab11 endosomes, by the Dynein motor protein. Eliminating microtubules from cells by overexpressing Katanin60 results in loss of Nuf-Dynein-based apical Rab11 endosome transport and failure to efficiently deliver Cad99C to the apical membrane. The effect on Cad99C polarisation is not an indirect effect of loss of polarity due to impaired Rab11 and Dynein function in localising the apical polarity determinant Crumbs to the apical membrane because, firstly, polarity is maintained in cells expressing Rab11 or Dynein RNAi, as indicated by the normal localisation of aPKC and, secondly, loss of Crb does not strongly affect cell polarity in the follicle cell epithelium owing to redundancy with Bazooka. The results indicate that even under conditions with severe depletion of microtubules, the overall shape of the follicle cell epithelium is relatively normal, indicating that polarised microtubules are required to influence formation of apical microvilli, rather than for other functions of the actin cytoskeleton in epithelial cells. Similarly, no strong effects are seen on cell shape upon loss of either Patronin or Shot (or both), raising questions over the claimed requirement for Patronin homologs and microtubules in formation or maintenance of adherens junctions epithelial cells in culture (Khanal, 2016).
The final step in delivery of Cad99C to the apical membrane also requires actin-based transport through the action of Rip11-MyoV complex. Compromising normal MyoV function in Drosophila follicle cells by expressing a dominant-negative version of the protein, results in loss of Rab11 polarisation from the apical membrane and its abnormal accumulation in the sub-apical region. This phenotype in Drosophila shows similarities with the human microvillus inclusion disease, where mutations in the Myo5b gene also cause loss of Rab11 endosomes from the apical membrane (Khanal, 2016).
In summary, these results reveal how the spectrin cytoskeleton acts to polarise microtubules in epithelial cells, and how polarised microtubules then direct trafficking of Rab11 endosomes carrying Cad99C to the apical membrane. This process relies on a hierarchy of events, and disruption at any stage can lead to failure in delivering Cad99C to the apical membrane, resulting in defective biogenesis of microvilli. These findings are directly relevant to human diseases such as Usher's Syndrome Type 1 and microvillus inclusion disease, helping to outline the molecular and cellular basis for these conditions (Khanal, 2016).
Mitotic spindles are primarily composed of microtubules (MTs), generated by polymerization of α- and β-Tubulin hetero-dimers. Defects in Tubulin polymerization dramatically affect spindle formation and disrupt chromosome segregation. Recently studies have described a role for the product of the conserved misato (mst) gene in regulating mitotic MT generation in flies, but the molecular function of Mst remains unknown. This study used affinity purification mass spectrometry (AP-MS) to identify interacting partners of Mst in the Drosophila embryo. Mst was shown to associate stoichiometrically with the hetero-octameric Tubulin Chaperone Protein-1 (TCP-1) complex, with the hetero-hexameric Tubulin Prefoldin complex, and with proteins having conserved roles in generating MT-competent Tubulin. RNAi-mediated in vivo depletion of any TCP-1 subunit phenocopies the effects of mutations in mst or the Prefoldin-encoding gene merry-go-round (mgr), leading to monopolar and disorganized mitotic spindles containing few MTs. Crucially, it was demonstrated that Mst, but not Mgr, is required for TCP-1 complex stability and that both the efficiency of Tubulin polymerization and Tubulin stability are drastically compromised in mst mutants. Moreover, structural bioinformatic analyses indicate that Mst resembles the three-dimensional structure of Tubulin monomers and might therefore occupy the TCP-1 complex central cavity. Collectively, these results suggest that Mst acts as a co-factor of the TCP-1 complex, playing an essential role in the Tubulin-folding processes required for proper assembly of spindle MTs (Palumbo, 2015).
Spatially organized macromolecular complexes are essential for cell and tissue function, but the mechanisms that organize micron-scale structures within cells are not well understood. Microtubule-based structures such as mitotic spindles scale with cell size, but less is known about the scaling of actin structures within cells. Actin-rich denticle precursors cover the ventral surface of the Drosophila embryo and larva and provide templates for cuticular structures involved in larval locomotion. Using quantitative imaging and statistical modeling, denticle number and spacing were demonstrated to scale with cell size over a wide range of cell lengths in embryos and larvae. Denticle number and spacing are reduced under space-limited conditions, and both features robustly scale over a ten-fold increase in cell length during larval growth. The relationship between cell length and denticle spacing can be recapitulated by specific mathematical equations in embryos and larvae, and accurate denticle spacing requires an intact microtubule network and the microtubule minus-end-binding protein, Patronin. These results identify a novel mechanism of microtubule-dependent actin scaling that maintains precise patterns of actin organization during tissue growth (Spencer, 2017).
INT6/eIF3e is a highly conserved component of the translation initiation complex that interacts with both the 26S proteasome and the COP9 signalosome, two complexes implicated in ubiquitin-mediated protein degradation. The INT6 gene was originally identified as the insertion site of the mouse mammary tumor virus (MMTV), and later shown to be involved in human tumorigenesis. Here we show that depletion of the Drosophila orthologue of INT6 (Int6) results in short mitotic spindles and deformed centromeres and kinetochores with low intra-kinetochore distance. Poleward flux of microtubule subunits during metaphase is reduced, although fluorescence recovery after photobleaching (FRAP) demonstrates that microtubules remain dynamic both near the kinetochores and at spindle poles. Mitotic progression is delayed during metaphase due to the activity of the spindle assembly checkpoint (SAC). Interestingly, a deubiquitinated form of the kinesin Klp67A (a putative orthologue of human Kif18A) accumulates near the kinetochores in Int6-depleted cells. Consistent with this finding, Klp67A overexpression mimics the Int6 RNAi phenotype. Furthermore, simultaneous depletion of Int6 and Klp67A results in a phenotype identical to RNAi of just Klp67A, which indicates that Klp67A deficiency is epistatic over Int6 deficiency. It is proposed that Int6-mediated ubiquitination is required to control the activity of Klp67A. In the absence of this control, excess of Klp67A at the kinetochore suppresses microtubule plus-end polymerization, which in turn results in reduced microtubule flux, spindle shortening, and centromere/kinetochore deformation (Renda, 2017).
Kinetochores connect centromeric chromatin to spindle microtubules during mitosis. Neurons are post-mitotic, so it was surprising to identify transcripts of structural kinetochore (KT) proteins and regulatory chromosome passenger complex (CPC) and spindle assembly checkpoint (SAC) proteins in Drosophila neurons after dendrite injury. To test whether these proteins function during dendrite regeneration, post-mitotic RNAi was performed and dendrites or axons were removed using laser microsurgery. Reduction of KT, CPC and SAC proteins decreased dendrite regeneration without affecting axon regeneration. To understand whether neuronal functions of these proteins rely on microtubules, microtubule behavior was analyzed in uninjured neurons. The number of growing plus, but not minus, ends increased in dendrites with reduced KT, CPC and SAC proteins, while axonal microtubules were unaffected. Increased dendritic microtubule dynamics was independent of DLK-mediated stress, but was rescued by concurrent reduction of γTubulin, the core microtubule nucleation protein. Reduction of γTubulin also rescued dendrite regeneration in backgrounds containing kinetochore RNAi transgenes. It is concluded that kinetochore proteins function post-mitotically in neurons to suppress dendritic microtubule dynamics by inhibiting nucleation (Hertzler, 2020).
The kinetochore is a quintessential mitosisand meiosis-specific structure that attaches chromosomes to the mitotic spindle for segregation of genetic material to daughter cells. In many species, including Drosophila, it is built on centromeric DNA that is recognized by the histone Cid/Cenp-A that recruits inner kinetochore protein Cenp-C to serve as a binding site for the KMN network (Knl1, Mis12, and Ndc80 complexes). Of these, the Mis12 complex is most central, while the remainder make up more distal parts of the structure that is involved in connecting to microtubules. Many animals have other inner kinetochore proteins, including the constitutive centromere-associated network (CCAN), but these are absent in Drosophila (Drinnenberg, 2016). In cycling mammalian cells, inner centromere proteins including Cenp-A, Cenp-C, and CCAN are found on the centromere in interphase as well as mitosis. The Mis12 complex has some nuclear staining in interphase, but it is not as clearly punctate as CENP-A. In Drosophila, Cenp-A/Cid, Cenp-C, and Mis12 localize to centromeres in interphase and mitosis. In mammals and Drosophila, the other kinetochore components including the Knl1 and Ndc80 complexes are recruited to the kinetochore only in mitosis. Once the kinetochore is fully assembled, Nuf2 and Ndc80 in the outer kinetochore capture microtubule plus ends growing into the central spindle to allow chromosome segregation (Hertzler, 2020).
In addition to the kinetochore proteins that bridge chromosomes to microtubules, several regulatory complexes are present at the kinetochore before final bioriented microtubule attachment. The chromosome passenger complex (CPC) targets the kinase Aurora B to the centromere, where it helps correct errors in chromosome attachment to microtubules by phosphorylation of substrates including Ndc80. The targeting subunits of the CPC are borealin (or in flies borealin-related, borr), survivin (or in flies Deterin, Det), and Incenp, which are tightly associated with one another through a three-helix bundle. Aurora B is also critical for recruiting spindle assembly checkpoint (SAC) proteins to unattached kinetochores. The major effector of the SAC is cdc20/fzy, which is a subunit of the anaphase-promoting complex or cyclosome (APC/C). When bound by SAC subunits at the kinetochore, cdc20/fzy is inactive. A third complex, Rod1-Zw10-Zwilch (RZZ) complex, interacts with both SAC and KMN proteins. Together these regulatory complexes ensure that anaphase does not begin until all sister chromatids are correctly attached to opposite spindle poles. Attachment causes a shift in kinase and phosphatase balance, such that Aurora B starts to lose, and 'stripping' of the SAC by dynein connected to the RZZ complex is initiated. As a result, cdc20/Fzy is freed to activate APC/C, the ubiquitin ligase that triggers degradation of Cyclin B and Securin to initiate sister chromatid separation. KMN network proteins remain at the kinetochore to mediate attachment of separating sister chromatids to spindle microtubules (Hertzler, 2020).
Most of the proteins that make up the kinetochore and its associated regulatory complexes have not been linked to functions in interphase or postmitotic cells. However, two recent studies demonstrated neuronal defects after postmitotic reduction of kinetochore proteins, suggesting that they do function in noncycling cells (Cheerambathur, 2019; Zhao, 2019). In Caenorhabditis elegans, KMN network components localize to ciliated dendrites of amphid neurons and play a role in their extension (Cheerambathur, 2019). Although the microtubule-binding domains of Ndc80 are required to support normal amphid development, specific defects in microtubules were not detected (Cheerambathur, 2019). In Drosophila, mis12 mutants were identified in a genetic screen to identify modulators of synaptic bouton structure (Zhao, 2019). Reduction of other KMN network proteins had similar effects on neuromuscular junction structure, and intriguingly, so did targeting the centromeric histone cid/CENP-A (Zhao, 2019). Knockdown of Mis12 in primary rodent hippocampal neurons increased the number of protrusions from dendrites (Zhao, 2019), indicating that kinetochore proteins likely function in neurons broadly across evolution. Specific alterations in the neuronal cytoskeleton were not reported in this study either. It therefore remains to be determined how kinetochore proteins impact neuronal structure (Hertzler, 2020).
One of the most intense structural challenges neurons face is rebuilding axons or dendrites after injury. Key regulators of axon regeneration have been identified, including sensors like the dual leucine zipper kinase (DLK), epigenetic regulators like HDAC5, transcription factors including fos, jun, and cebp-1, and a myriad of downstream regeneration-associated genes (RAGS). However, it was only recently shown that neurons survive removal of all dendrites and can regrow a new arbor. Only a handful of genes including Akt and Ror have been linked to dendrite regeneration. To identify dendritic RAGs, RNA was isolated from Drosophila neurons after in vivo dendrite removal. Surprisingly, many kinetochore gene transcripts were coordinately up-regulated 6 h after dendrite injury. These included not only transcripts encoding structural kinetochore proteins in the inner kinetochore and KMN network, but also transcripts encoding regulatory proteins in the CPC, SAC, and RZZ complexes. Postmitotic RNA interference (RNAi) was used to confirm a role of some of these proteins in dendrite regeneration. In the same genetic backgrounds, no effect was seen on axon regeneration. In uninjured neurons, kinetochore protein reduction specifically increased microtubule plus-end number (also referred to as microtubule dynamics) in dendrites without altering other metrics of microtubule behavior such as polarity, speed, or minus-end density. Unexpectedly, axonal microtubule dynamics was unaffected. Therefore, kinetochore proteins act in a compartment-specific manner to suppress microtubule dynamics in dendrites. The dendritic microtubule phenotype and dendrite regeneration defect were rescued by concurrent reduction of γ-tubulin, suggesting that kinetochore proteins normally function to temper microtubule nucleation in dendrites. Together the data suggest that the KMN network and CPC, SAC, and RZZ complexes function together to promote dendrite regeneration and modulate dendritic microtubule dynamics through control of nucleation in postmitotic neurons (Hertzler, 2020).
On the basis of the surprising appearance of transcripts of structural and regulatory kinetochore proteins in RNASeq data from injured neurons, their role in dendrite regeneration was investigated. Regeneration after dendrite, but not axon, injury was impaired when they were reduced using postmitotic RNAi. Because kinetochore proteins function to attach, and monitor attachment, to microtubules in mitotic spindles, microtubule behavior was surveyed in neurons in which they were reduced. RNAi knockdowns of many kinetochore proteins caused an up-regulation of dendritic microtubule dynamics, an effect not seen in axons. Other parameters of microtubule behavior, including number of growing minus ends, were not affected. The increase in plus-end, but not minus-end, number suggested that nucleation rather than severing was responsible. The involvement of nucleation was confirmed by rescue of the plus end and regeneration phenotypes in neurons with reduced γ-tubulin. The increase in microtubule dynamics when kinetochore proteins were reduced suggests that they normally function to dampen nucleation of dendritic microtubules in postmitotic neurons (Hertzler, 2020).
Two recent studies indicate that kinetochore proteins are likely to function broadly in different neuron types. In one study, the authors identified Mis12 mutants in a Drosophila forward genetic screen. Other components of the KMN network complexes (Ndc80, Knl1, and Nnf1a), as well as centromeric protein Cenp-A (Cid), had similar neuromuscular junction phenotypes and reduced neuropil in the CNS (Zhao, 2019). The authors went on to show that mammalian neurons with reduced Mis12 also had structural defects, this time in dendrites (Zhao, 2019). The group reported some localization of KMN network proteins to spots in peripheral nerves and neuropil in the CNS (Zhao, 2019), but the pattern did not help to suggest how the proteins might function in neurons (Hertzler, 2020).
The other study demonstrated that Cenp-C, Ndc80, and Nuf2 localize postmitotically to the ciliated dendrites of amphid neuron bundles in C. elegans. When these proteins were knocked down with an elegant GFP-degrader system, amphid dendrite extension was impaired (Cheerambathur, 2019). Effects on egg laying and fertility were also seen. Notably, a deletion of only the microtubule-binding domains of Ndc80 was able to phenocopy the knockdowns, indicating that an interaction with microtubules likely mediates the phenotypes. Degradation of GIP2, an essential γTuRC subunit, showed the same neuron extension deficits, supporting the hypothesis that microtubules in these neurons are affected by KMN network protein knockdown, although the authors were unable to pinpoint any specific changes in the microtubule cytoskeleton (Hertzler, 2020).
The two previous studies on neuronal roles for kinetochore proteins focused on the structural components of the kinetochore, including inner centromere components Cenp-A and C and KMN network proteins (Cheerambathur, 2019; Zhao, 2019). The three regulatory complexes-CPC, RZZ, and SAC- are added to the list of kinetochore proteins with neuronal function. Moreover, it was found that players in all structural and regulatory complexes have similar roles in controlling microtubule dynamics. This suggests that, as in mitosis, these complexes function together in a single pathway in neurons. It is therefore likely that they are not only involved in attaching microtubules to something, but are also acting as sensors of microtubule behavior. If they have a similar role in neurons as in mitosis, they may recognize plus-end arrival at a specific cellular location. Again, in analogy with mitosis, plus-end arrival could trigger release of regulatory complexes and free them to send signals to other cellular locations about the status of the microtubule cytoskeleton. These signals could involve ubiquitination mediated by association of Cdc20/Fzy with an E3 ligase complex, or phosphorylation by Aurora B. One target of either output signal could be suppression of γTuRC activity (Hertzler, 2020).
One intriguing aspect of these findings is that changes were only observed in the microtubule cytoskeleton in dendrites, and dynamics in the axon was unaffected. While phenotypes in C. elegans and mammalian neurons occurred in dendrites, changes in motor axon terminals were seen in Drosophila. One possible way to reconcile the axonal and dendritic phenotypes would be if kinetochore proteins are important throughout dendrites but function specifically at synaptic regions of axons. There are several hints that microtubule nucleation might be particularly important near presynaptic sites. In Drosophila motor neurons, concentrations of γ-tubulin were seen in large synaptic boutons at the neuromuscular junction. In cultured mammalian neurons, γ-tubulin also concentrates at presynaptic sites, and increases in neuronal activity lead to increases in EB comet formation at these sites, suggesting that regulation of nucleation is important in axons. It is therefore possible that kinetochore proteins regulate presynaptic nucleation in addition to dendritic nucleation (Hertzler, 2020).
In mitotic cells, kinetochore proteins localize to tight spots around centromeric DNA, and their localization has provided invaluable clues to their function. So far, localization patterns in neurons have not helped pin down specific sites of action. In C. elegans, expression levels in amphid neurons were high enough to detect tagged proteins expressed at endogenous levels, and they were seen throughout the linear ciliated dendrite region (Cheerambathur, 2019). In contrast, expression seems low in Drosophila neurons, and beyond being present in neuropil, a pattern has not been discernible (Zhao, 2019). Attempts of this study to visualize meaningful localization in sensory neurons also did not provide any insights. Use of amplification systems like SunTag may be required to acquire meaningful information about where structural and regulatory kinetochore proteins function in neurons (Hertzler, 2020).
One possible model for neuronal kinetochore protein function is that, in analogy with mitosis, the structural and regulatory complexes colocalize at a specific cellular site, perhaps in the cell body, when microtubule plus ends are absent or in low abundance. Upon plus-end arrival, the RZZ complex in conjunction with dynein could transport regulatory proteins toward microtubule minus ends. Dendrites contain minus-end-out microtubules, so this would mean that regulatory proteins could be transported outward into dendrites to suppress microtubule dynamics distally after plus-end arrival. This type of mechanism could allow for global homeostatic control of microtubule dynamics in dendrites (Hertzler, 2020).
Dendritic morphogenesis requires dynamic microtubules (MTs) to form a coordinated cytoskeletal network during development. Dynamic MTs are characterized by their number, polarity and speed of polymerization. Previous studies described a correlation between anterograde MT growth and terminal branch extension in Drosophila dendritic arborization (da) neurons, suggesting a model that anterograde MT polymerization provides a driving force for dendritic branching. This study recently found that the Ste20-like kinase Tao specifically regulates dendritic branching by controlling the number of dynamic MTs in a kinase activity-dependent fashion, without affecting MT polarity or speed. This finding raises the interesting question of how MT dynamics affects dendritic morphogenesis, and if Tao kinase activity is developmentally regulated to coordinate MT dynamics and dendritic morphogenesis. This study explored the possible correlation between MT dynamics and dendritic morphogenesis together with the activity changes of Tao kinase in C1da and C4da neurons during larval development. The data show that spatiotemporal changes in the number of dynamic MTs, but not polarity or polymerization speed, correlate with dendritic branching and Tao kinase activity. These findings suggest that Tao kinase limits dendritic branching by controlling the abundance of dynamic MTs and a novel model is proposed on how regulation of MT dynamics might influence dendritic morphogenesis (Hu, 2022).
The evolutionarily conserved Glycogen Synthase Kinase 3β (GSK3β), a negative regulator of microtubules, is crucial for neuronal polarization, growth and migration during animal development. However, it remains unknown whether GSK3β regulates neuronal pruning, which is a regressive process. This study reports that the Drosophila GSK3β homologue Shaggy (Sgg) is cell-autonomously required for dendrite pruning of ddaC sensory neurons during metamorphosis. Sgg is necessary and sufficient to promote microtubule depolymerization, turnover and disassembly in the dendrites. Although Sgg is not required for the minus-end-out microtubule orientation in dendrites, hyperactivated Sgg can disturb the dendritic microtubule orientation. Moreover, pharmacological and genetic data suggest that Sgg is required to promote dendrite pruning at least partly via microtubule disassembly. This study showed that Sgg and Par-1 kinases act synergistically to promote microtubule disassembly and dendrite pruning. Thus, Sgg and Par-1 might converge on and phosphorylate a common downstream microtubule-associated protein(s) to disassemble microtubules and thereby facilitate dendrite pruning (Dzaki, 2022).
The evolutionarily conserved CLASPs (cytoplasmic linker-associated proteins) are microtubule-associated proteins that inhibit microtubule catastrophe and promote rescue. CLASPs can regulate axonal elongation and dendrite branching in growing neurons. However, their roles in microtubule orientation and neurite pruning in remodeling neurons remain unknown. This study identified the Drosophila CLASP homolog Orbit/MAST, which is required for dendrite pruning in ddaC sensory neurons during metamorphosis. Orbit is important for maintenance of the minus-end-out microtubule orientation in ddaC dendrites. Structural analysis reveals that the microtubule lattice-binding TOG2 domain is required for Orbit to regulate dendritic microtubule orientation and dendrite pruning. In a genetic modifier screen, the conserved Par-1 kinase was further identified as a suppressor of Orbit in dendritic microtubule orientation. Moreover, elevated Par-1 function impairs dendritic microtubule orientation and dendrite pruning, phenocopying orbit mutants. Overall, this study demonstrates that Drosophila CLASP governs dendritic microtubule orientation and dendrite pruning at least partly via suppressing Par-1 kinase (Bu, 2022).
The mechanisms regulating synapse numbers during development and aging are essential for normal brain function and closely linked to brain disorders including dementias. Using Drosophila, this study demonstrates roles of the microtubule-associated protein Tau in regulating synapse numbers, thus unravelling an important cellular requirement of normal Tau. In this context, it was found that Tau displays a strong functional overlap with microtubule-binding spectraplakins, establishing new links between two different neurodegenerative factors. Tau and the spectraplakin Short Stop act upstream of a three-step regulatory cascade ensuring adequate delivery of synaptic proteins. This cascade involves microtubule stability as the initial trigger, JNK signalling as the central mediator, and kinesin-3 (see Drosophila Unc-103) mediated axonal transport as the key effector. This cascade acts during development (synapse formation) and aging (synapse maintenance) alike. Therefore, these findings suggest novel explanations for intellectual disability in Tau deficient individuals, as well as early synapse loss in dementias including Alzheimer's disease (Voelzmann, 2016).
The correct formation and subsequent maintenance of synapses is a key prerequisite for brain development, function and longevity. Precocious loss of synapses is observed in late onset neurodegenerative diseases including Alzheimer's disease (AD) and Frontotemporal Dementia (FTD), likely contributing to the cognitive decline and neuronal decay observed in patients. Therefore, the characterisation of mechanisms maintaining synapses during ageing would have major implications for understanding of dementias (Voelzmann, 2016).
The development of synapses and their maintenance during ageing is dependent on sustained transport of synaptic proteins from the distant soma, driven by motor proteins which trail along the bundles of microtubules in axons and dendrites. Microtubules are regulated by microtubule binding proteins which are therefore in a key position to regulate synapse formation and maintenance (Voelzmann, 2016).
Tau is a microtubule associated protein (MAP) discovered in the mid-seventies. Reduction in Tau levels has been linked to intellectual disability and a class of brain disorders termed 'dementias which lack distinctive histopathology' (DLDH). Tau detachment from MTs is linked to prominent neurodegenerative diseases such as Alzheimer's disease, Frontotemporal Dementia and some forms of Parkinson's disease. In vitro, Tau has the ability to regulate microtubule properties including stability, cross-linkage and polymerisation. Through such functions, Tau would be expected to regulate multiple aspects of neuronal cell biology, but its physiological roles are still not understood and highly debated. This might partly be due to experimental challenges posed by functional redundancy, where other MAPs are proposed to mask physiological roles of Tau (Voelzmann, 2016).
A good model in which to deal with functional redundancy is the fruit fly Drosophila melanogaster. As is ideal for studies of Tau, Drosophila neurons provide access to powerful genetics, they are readily established for research on the neuronal cytoskeleton, on neuronal transport and on synapses. Importantly, concepts and mechanisms gained from work in flies are often well conserved in higher organisms (Voelzmann, 2016).
Work in Drosophila suggested that the spectraplakin Short Stop (Shot), a large actin-MT linker molecules and potent regulators of microtubules, could display potential functional overlap with Tau during microtubule stabilisation. This hypothesis is attractive because the well-conserved mammalian spectraplakin Dystonin is already linked to a neurodegenerative disease (type VI hereditary sensory autonomic neuropathy; OMIM #614653), and its paralogue ACF7/MACF1 plays important roles during brain development). Since ACF7 continues to be expressed in the brain, it is tempting to speculate that it might be required for neuronal maintenance (Voelzmann, 2016).
This study used Drosophila neurons, in culture and in vivo alike, to demonstrate novel roles of Tau in regulating the formation and maintenance of synapses during ageing, by coordinating the intracellular trafficking of synaptic proteins. Thus, this study shows that the role of Tau in synapse regulation occurs in functional overlap with Shot. The robust shot-tau double-mutant phenotypes enabled study of the mechanistic cascade composed of three steps: microtubule stability as the trigger, the JNK signalling pathway as the mediator and kinesin-3 mediated axonal transport of synaptic proteins as the key effector. It is propose that a new mechanism based on the loss of Tau function which could explain intellectual disability in MAPT (the human tau gene) mutant individuals and precocious synapse loss in tau-related neurodegeneration (Voelzmann, 2016).
The aim of these studies was to understand the role of endogenous Tau in neurons with particular attention to synapses. This effort was essentially aided by the finding that Tau and Shot are functionally redundant, and the subsequent incorporation of Shot into these studies. The robust phenotypes of shot-tau double-mutant neurons enabled this study to demonstrate roles of Shot-Tau during the formation and maintenance of pre-synaptic sites in axons, and unravel the underlying mechanistic cascade which involves three major steps. Firstly, the absence of Shot-Tau causes microtubule destabilisation. Secondly, this cytoskeletal stress causes aberrant JNK activity patterns with upregulation in somata and downregulation at axon tips. Thirdly, aberrant JNK activation leads to a somatic roadblock for kinesin-3 mediated transport, thus inhibiting the delivery of synaptic proteins and eventually causing synapse loss. Depending on whether the functions of Tau and/or Shot are removed during development or ageing, either the formation or the maintenance of synapses are affected, respectively (Voelzmann, 2016).
The model explaining the function of Tau and Shot in synapse establishment and maintenance by regulating intracellular transport, is supported by loss- and gain-of-function experiments, genetic interactions and cross-rescue experiments. The initial finding that shot-tau mutant neurons had reduced branch numbers, could have suggested that defects on synapse numbers is indirect. However, experiments with double knock-down in culture and in the adult brain clearly showed strong synapse reduction whilst maintaining normal branch patterns, and Unc-104 rescued synapse reduction in shot-tau mutant neurons without major increases of the branch pattern in these neurons. These results clearly demonstrate that changes in neuronal morphology are not the cause of changes in synapse number (Voelzmann, 2016).
Notably, the synaptic function of Tau described in this study for Drosophila might be conserved in higher animals or humans, since also aged Tau knock-out mice develop a reduction of synaptic proteins from the hippocampus (Voelzmann, 2016 and references therein).
These findings provide potential new mechanistic explanations for various tau related brain disorders. For example, microdeletions in the region of MAPT (the human tau gene) cause intellectual disability, and Tau's synapse-promoting roles may well contribute to this pathology. Furthermore, various tauopathies are characterised by precocious pathological loss of synapses. The currnet data suggest that loss of tau could lead to defective synapse maintenance and eventually synapse loss. For example, a prominent group of dementias which lacks distinctive histopathology (DLDH) are characterised by the loss of Tau. Further tauopathies including Alzheimer disease, typically involve hyper-phosphorylation and aggregate formation of Tau. In this scenario, there are two parallel, non-exclusive modalities through which Tau can cause pathology. Firstly, detached hyper-phosphorylated tau attains gain-of-function roles in the cytoplasm damaging neurons through a number of mechanisms. Secondly, hyper-phosphorylation of tau causes a loss-of-function condition by depleting Tau from microtubules. However, since Tau knock-out mouse models mostly failed to show significant phenotypes and the neuronal functions of endogenous tau remain little understood, the pathological importance of Tau loss from microtubules has been marginalised. The current results now re-emphasise the notion that loss of Tau from microtubules could contribute to neurodegenerative pathology and deliver mechanistic explanations (Voelzmann, 2016).
To unravel pathomechanisms caused by the loss of Tau, a combined depletion of Shot and Tau gave strong phenotypes, ideal for short-term experimental approaches. However, similar, yet milder phenotypes were found if only Tau was depleted, suggesting that the mechanisms described in this study could well contribute to slow disease progression in tauopathies. The discovery that spectraplakins are MAPs which functionally overlap with Tau, opens up new experimental avenues for Tau studies. So far, spectraplakins have been linked to the degeneration of sensory and autonomous neurons, and it remains to be elucidated whether they may have similar roles also in the brain. These results clearly hint at this possibility (Voelzmann, 2016).
The loss of Tau and/or Shot inhibits kinesin-3 mediated transport leading to accumulation of synaptic proteins in the soma of neurons. A road-block mechanism is proposed suppressing the initiation of axonal transport in somata of Shot-Tau depleted neurons, which is caused indirectly through microtubule stress and mediated by JNK (Voelzmann, 2016).
The involvement of microtubules in causing a transport block is supported by experiments using microtubule stabilising and de-stabilising drugs which rescued or mimicked the shot-tau mutant phenotypes, respectively. Similarly, axonal transport defects and cognitive deficits of PS19Tg mice (expressing the P301S mutant form of human tau) and various other mouse and fly tauopathy models were shown to be rescued by microtubule-stabilising drugs, suggesting that the mechanisms described may be conserved and relevant to disease (Voelzmann, 2016).
The somatic road-block is a novel mechanism through which the loss of Tau can interfere with the transport of synaptic proteins and provides potential explanations also for somatic accumulations of postsynaptic proteins such as PSD-95, AMPA and NMDA receptors observed in mouse tauopathy models. A likely mechanism causing a roadblock in intracellular transport could be the direct inactivation of Unc-104 or its associated adaptor proteins, for example through JNK or other kinases within its pathway. This mode of regulation has a clear precedent in kinesin-1 and its adaptor Jip which are directly phosphorylated by JNK leading to transport inhibition. Unfortunately, extensive attempts to co-immunoprecipitate JNK and Kinesin-3 were unsuccessful, leaving open for now the exact molecular mechanism (Voelzmann, 2016).
It is proposed that aberrant JNK activation downstream of microtubule destabilisation or stress is the ultimate cause for the defective delivery of synaptic proteins in Tau and/or Shot loss of function. Also in mouse, microtubule stress leads to somatic activation of the JNK pathway, suggesting this mechanism is likely to be conserved with vertebrates (Voelzmann, 2016).
The JNK pathway is emerging as a central player in neurodegenerative diseases. Its activation is prompted by various neurodegeneration risk factors including oxidative stress, inflammation, and ageing. Furthermore, JNK is activated in AD patients and in several AD models where it triggers progression of the pathology. The new link between Tau/spectraplakins, JNK and synapses proposed in this study, is therefore likely to provide mechanistic explanations for synaptic pathology observed in AD and other tauopathies (Voelzmann, 2016).
This study has delivered an important conceptual advance by revealing a new mechanistic cascade which can explain synaptic decay as the consequence of Tau loss from microtubules. Furthermore, a previously unknown functional redundancy with spectraplakins was identified as a promising new avenue for research on Tau. These findings emphasize that Tau detachment from microtubules can be an important aspect contributing to the pathology of tauopathies in parallel to roles of hyper-phosphorylated Tau in the cytoplasm. Synaptic decay, axonal transport and alterations in the JNK pathway are emerging as central players in a wider range of adult-onset neurodegenerative diseases, and here this study has aligned these factors into a concrete mechanistic cascade (Voelzmann, 2016).
Coordinated rearrangements of cytoskeletal structures are the principal source of forces that govern cell and tissue morphogenesis. However, unlike for actin-based mechanical forces, knowledge about the contribution of forces originating from other cytoskeletal components remains scarce. This study has establish microtubules as central components of cell mechanics during tissue morphogenesis. Individual cells were found to be mechanically autonomous during early Drosophila wing epithelium development. Each cell contains a polarized apical non-centrosomal microtubule cytoskeleton that bears compressive forces, whereby acute elimination of microtubule-based forces leads to cell shortening. It was further established that the Fat planar cell polarity (Ft-PCP) signalling pathway couples microtubules at adherens junctions (AJs) and patterns microtubule-based forces across a tissue via polarized transcellular stability, thus revealing a molecular mechanism bridging single cell and tissue mechanics. Together, these results provide a physical basis to explain how global patterning of microtubules controls cell mechanics to coordinate collective cell behaviour during tissue remodelling. These results also offer alternative paradigms towards the interplay of contractile and protrusive cytoskeletal forces at the single cell and tissue levels (Singh, 2018).
During development individual cells assemble into complex tissues and organs with specialized forms and functions. Tissue morphogenesis is driven by mechanical forces that are generated by the cytoskeleton within cells and transmitted in a coordinated manner through adhesion molecules across neighbouring cells. The best-studied cytoskeletal component is actin, which, together with other proteins, forms protrusive and contractile arrays, a fundamental constituent of rearrangements on the single cell and tissue levels. Recent work has suggested that microtubules, similar to actin, can also generate forces in cells. However, understanding of the contribution of microtubules to cell mechanics, cell shape changes and force coordination during morphogenesis remains poor. This is mainly due to the fact that many current models describing the mechanical state of tissues during shape changes focus on actomyosin dynamics and/or rely on continuum mechanics. These studies, which are based on coarse-grain observations of cell movements or cell shape changes, reveal only part of the physical mechanisms that drive morphogenesis and do not directly investigate the physicomechanical context of tissue remodelling. To understand the relationship between cell mechanics, force patterning and molecular structure, this study investigated the mechanical properties of microtubules at high spatiotemporal resolution using wing development in Drosophila melanogaster as a paradigm (Singh, 2018).
During pupal wing development, non-centrosomal microtubules form an apical array of parallel microtubule bundles that are globally aligned along the proximal-distal (P-D) axis. Patterning of the microtubule cytoskeleton depends on the Ft-PCP signalling pathway and occurs during the early phase of wing reshaping (that is, between 14 and 18 h after puparium formation, or APF). This patterning is associated with extensive changes in cell shape, cell divisions and cell-cell rearrangements. In the Drosophila wing, the Ft-PCP pathway further orients cell elongation and cell divisions along the P-D axis to induce wing tissue elongation. Intriguingly, rescue of the Hippo pathway in Ft-PCP mutant animals, in which microtubule alignment is impaired, aberrant development results in perturbed cell elongation and an abnormal rounded wing shape, suggesting that there is an interdependence between these events. Therefore this study explored the possibility that microtubule-based cell mechanics control cell and tissue shape during early wing development between 16 and 18 h APF (Singh, 2018).
Tissue remodelling is driven by intrinsic and extrinsic mechanisms, and it has previously been shown that extrinsic mechanical forces act during the late phase of wing reshaping (starting 18 h APF). These forces are generated by hinge contraction of the wing that is attached to the cuticle on the distal side. This study evaluated the mechanical autonomy of individual cells before hinge contraction at an earlier developmental stage (that is, between 16 and 18 h APF). This was done by isolating a single cell (or a small patch of cells) using a single-pulse multipoint procedure to cut AJs, thus mechanically uncoupling individual cells from their surrounding. Strikingly, the shape of individual isolated cells did not change significantly upon laser ablation at 17-18 h APF, when cells in the wing are already elongated. The same result was obtained when patches of cells were isolated. Additional analyses of the Feret's diameter before and after ablation showed a small isotropic decrease in cell size, providing evidence that at this early stage, individual cells are not influenced by the neighbouring cells or by tissue-scale forces in a polarized fashion. Consistently, analysis of animals expressing a mutant form of dumpy protein, an extracellular matrix protein associated with tissue remodelling at later developmental stages, showed no substantial differences in wing shape compared to wild-type wings at 18 h APF. Together, these experiments argue that unlike later stages, cell autonomous forces are the major drivers of initial cell shape changes between 16 and 18 h APF (Singh, 2018).
To identify the molecular mechanism underlying cell autonomous shape formation, the distribution and dynamics of two cytoskeletal force-generators were investigated: microtubules and non-muscle myosin II (MyoII) as a component of the actomyosin cytoskeleton. MyoII was detected at the apical cell cortex at the level of AJs. A subsequent analysis of the signal distribution within single cells revealed a planar polarized distribution of MyoII along the P-D axis, which correlated with increased tension along the same junctions. As MyoII provides contractile forces, this should result in P-D junctional shortening upon laser ablation. However, this is inconsistent with the current ablation experiments, suggesting that there is an opposing force present. Interestingly, staining of microtubules showed planar polarized apical microtubules along the P-D axis at the level of AJs. Microtubules are the stiffest cytoskeletal filaments, with a persistence length on the order of millimetres. Microtubules are therefore well suited to balance the tension generated by actomyosin contraction. Consistently, the distribution of microtubules and MyoII in mechanically isolated cells remain polarized. In addition, microtubule and MyoII polarity was preserved in dumpy mutant wings at 18 h APF, indicating that they are polarized in a cell autonomous fashion. The possible role of the atypical myosin Dachs, a downstream component of the Fat signalling pathway, was also analyzed. Dachs mutant wings showed no change in cell elongation or microtubule polarity, which is consistent with recent work reporting that recombinant Dachs does not have ATPase activity and can therefore not function as a molecular motor. Together, these observations argue that planar polarized microtubules may balance actomyosin tensional forces that pull on P-D junctions and stabilize cell shape (Singh, 2018).
To validate this hypothesis, and to elucidate the dynamic and functional role of microtubules in cell mechanics, their properties were investigated during wing development. Live cell imaging of EOS-α-tubulin (EOS-Tub) showed that microtubules were not static but engaged in complex and dynamic rearrangements. An analysis of microtubule straightness showed that in wing cells, virtually all microtubules along the P-D axis were bent, consistently undergoing short wavelength buckling (~3 μm) near the cell cortex. It was further observed that growing microtubules remain straight and only start to buckle after they reach the cell cortex, exhibiting local short wavelength buckling near these sites. This result indicates that microtubule polymerization can generate considerable compressive forces to induce microtubule buckling (Singh, 2018).
Next, whether buckling of microtubules in Drosophila wing epithelium is indeed a result of forces acting on microtubules was also investigated, as suggested by the current experiments and in vitro studies, or whether the cellular environment yields more flexible microtubules. This is important, as buckling of flexible microtubules would rule out a role in balancing actomyosin contractility. To probe the forces of single microtubule filaments in vivo, individual microtubules were cut by laser ablation and the subsequent relaxation was monitored using live imaging. Previously curved microtubules rapidly straighten out, thus verifying that microtubules are indeed loaded with compressive forces. Finally, it was also observed that local ablation of microtubules triggers a rapid translocation of the adjacent junction. This finding supports the idea that non-centrosomal microtubules continuously generate pushing forces via polymerization that may then be stored as compressive forces in a polarized fashion to balance contractile forces generated by junctional actomyosin (Singh, 2018).
How are microtubules polarized along the P-D axis? While the molecular mechanism has remained elusive, previous work has established that the Ft-PCP signalling pathway aligns the apical microtubule network along the P-D axis by regulating association sites of microtubules with AJs. Considering the observed stability of aligned microtubules, whether directional differences in microtubule dynamics could serve as a mechanism for the planar polarization of microtubules was tested. Monitoring of EB1 tagged with green fluorescent protein (EB1-GFP) revealed two populations of microtubule-plus ends: fast growing microtubules with a growth velocity of 24.43 ± 0.43 7mi;m min-1 (mean ± s.e.m.), and slow growing microtubules with a velocity of 17.06 ± 0.26 7mi;m min-1. A further analysis showed that the microtubule growth rates depended on relative localization within cells as well as the growth angle relative to the P-D axis. Microtubule growth rates in the cell interior were higher compared to the cell cortex. Similarly, microtubules along the P-D axis grew faster than microtubules growing perpendicular to the P-D axis along the A-P axis, establishing a spatial gradient in microtubule growth velocity. The lower growth rate along the A-P axis close to the cell periphery suggests that there is more frequent pausing and switching between polymerization and depolymerization of microtubules, thus indicating a decreased stability of A-P oriented microtubules (Singh, 2018).
It was reasoned that over time, such differences in dynamics and stability may result in predominantly P-D aligned microtubules. To test this hypothesis, the cortical residence time was analyzed of microtubules as a function of their angle with respect to the P-D axis. Intriguingly, it found that microtubules that interact with the P-D cell cortex have a longer lifetime than microtubules interacting with the A-P cortex. Upon closer inspection, dynamic cycles of short-lived interactions of microtubules with A-P junctions were noted followed by depolymerization. Importantly, A-P oriented microtubules do not show buckling behaviour, which is in contrast to P-D oriented microtubules, but rather undergo catastrophe soon after interaction with A-P oriented cell junctions. This result suggests that microtubule-plus ends are less stable at these sites and thus cannot sustain long-lasting interactions with the cell cortex, which are required to generate compressive forces. Building on these observations, in silico probing was performed to see whether the angular difference in lifetime may indeed be sufficient for microtubule polarization. Assuming a random orientation for de novo formed microtubules, the lifetime of each microtubule was defined as a function of the angle with a maximal lifetime along the P-D axis. Upon expiration, individual microtubules were re-introduced into the system at random angles, therefore keeping the total number of microtubules constant. Consistent with the in vivo observations, the simulation reached a steady-state at which a constant fraction of microtubules polarized along the P-D axis. Taken together, these observations point to a mechanism whereby microtubule stability regulates the planar alignment of the microtubule cytoskeleton along the P-D axis, which in turn directs cell mechanics along this axis. These data place directional microtubule stability upstream of proposed mechanisms of how cell shape influences microtubule alignment. Furthermore, these results are consistent with previous findings that microtubule association with P-D oriented AJs during the initial stage of wing development depends on Ft-PCP signalling (Singh, 2018).
Having established that planar polarized microtubule-based forces shape single cells, their mechanical coupling and integration into tissue-level mechanics were investigated. In a first round of experiments, transcellular coupling of microtubules were investigated on the ultrastructural level using transmission electron microscopy (TEM). In agreement with previous work, AJs were juxtaposed in neighbouring cells associated with microtubule filaments that span across cells in wild-type wings, forming supracellular cables analogous to myosin cables. Notably, no such association was observed in ftl(2)fdd1 / ftl(2)fd dGC13 (ft d) and ftl2 fd/ftGRV;ActP-Gal4/UAS-FtΔECDΔN-1 (N1) mutant wings, in which microtubules are randomly oriented in wing cells, therefore providing structural support for the Ft-PCP-dependent stabilization of microtubule-based forces at P-D oriented AJs. Consistently, ft mutant clones showed a fragmented microtubule cytoskeleton, arguing that there is Ft-PCP-dependent stabilization of microtubules via coupling at AJs (Singh, 2018).
To further validate the role of polarized transcellular microtubule stability in tissue mechanics and organization, tissue shape changes were observed upon acute perturbation of microtubule-based forces. To control microtubule dynamics in a precise spatial and temporal manner, he recently developed photostatin (PST1)35, a photo-switchable analogue of combretastatin A-4 (CA4)36 was used. The drug was applied to directly test the requirement of microtubules for cell shape maintenance. Notably, it was found that the exposed wing area contracted along the P-D axis upon microtubule inhibition. Quantitative cell shape analysis showed a small but significant reduction in the elongation index (EI) in selective regions where the drug was activated, arguing that polarized tissue stabilization is via microtubule-based forces. Finally, overexpression of the microtubule-severing protein Spastin increased cell shape heterogeneity. These results are consistent with the hypothesis that an intact polarized microtubule cytoskeleton is not only required for the maintenance of anisotropic cell shape but also critically involved in shaping the whole tissue during morphogenesis via polarized transcellular force stability (Singh, 2018).
Understanding the role of microtubules during animal development has so far been limited, especially because of a shortage of methods suitable to demonstrate causality in vivo. Taking advantage of complementary genetic, chemical, numerical and microscopy approaches, these experiments unveil polarized microtubule-based compressive forces as an alternative principle for stabilizing and maintaining cell and tissue shape during morphogenesis. Alignment of microtubules along the P-D axis was found to be based on increased longevity and polymerization of microtubules interacting with P-D oriented AJs compared to non-polarized microtubules. The result of this microtubule patterning along the P-D axis is an asymmetric distribution of protruding forces, which are stored in a polarized fashion via compressive loads on microtubules. Considering that actomyosin and microtubules are both planar polarized, it is plausible to envision that the observed compressive load on microtubules plays an active role in balancing actin-based contractile forces, resulting in the cell mechanical autonomy observed in the laser ablation experiments. Intriguingly, it was recently shown that acetylation of microtubules increases their mechanical resistance and that microtubules undergo self-repair upon damage. These important features support the role of the microtubule cytoskeleton as a site of long-term compressive force storage. Finally, evidence is provided that planar polarized microtubules are coupled at AJs across individual cells, bridging forces on the tissue level via polarized transcellular stability. Although the molecular identity remains elusive, the data suggests an involvement of AJ-associated proteins organized by the Ft-PCP pathway in this process (Singh, 2018).
Collectively, this work provides evidence that PCP-based planar patterning of the microtubule cytoskeleton not only results in polarized cell-autonomous forces but also coordinates global force patterning during tissue morphogenesis. The proposed mechanism establishes the Ft-PCP pathway at the onset of cell and wing elongation, before shape changes, due to extrinsic mechanical forces. Consistently, in a Ft-PCP mutant, in which initial elongation fails, consecutive remodelling by extrinsic tensile forces cannot rescue these length defects, therefore leading to shorter and rounder adult wings. Considering that the Ft-PCP signalling pathway controls a variety of dynamic cell population in vertebrates, the microtubule-based mechanism described in this study is likely to be physiologically relevant beyond wing development (Singh, 2018).
Axons of neurons are contractile, i.e., they actively maintain a rest tension. However, the spatial origin of this contractility along the axon and the role of the cytoskeleton in generating tension and sustaining rigidity are unknown. Using a microfluidic platform, this study exposed a small segment of the axons of embryonic Drosophila motor neurons to specific cytoskeletal disruption drugs. A local actomyosin disruption led to a total loss in axonal tension, with the stiffness of the axon remaining unchanged. A local disruption of microtubules led to a local reduction in bending stiffness, while tension remained unchanged. These observations demonstrated that contractile forces are generated and transferred along the entire length of the axon in a serial fashion. Thus, a local force disruption results in a collapse of tension of the entire axon. This mechanism potentially provides a pathway for rapid tension regulation to facilitate physiological processes that are influenced by axonal tension (Fan, 2019).
In many eukaryotic cells, direcsted molecular transport occurs along microtubules. Within neuronal axons, transport over vast distances particularly relies on uniformly oriented microtubules, whose plus-ends point towards the distal axon tip (anterogradely polymerizing, or plus-end-out). However, axonal microtubules initially have mixed orientations, and how they orient during development is not yet fully understood. Using live imaging of primary Drosophila melanogaster neurons, this study found that, in the distal part of the axon, catastrophe rates of plus-end-out microtubules were significantly reduced compared to those of minus-end-out microtubules. Physical modelling revealed that plus-end-out microtubules should therefore exhibit persistent long-term growth, while growth of minus-end-out microtubules should be limited, leading to a bias in overall axonal microtubule orientation. Using chemical and physical perturbations of microtubule growth and genetic perturbations of the anti-catastrophe factor p150, which was enriched in the distal axon tip, it was confirmed that the enhanced growth of plus-end-out microtubules is critical for achieving uniform microtubule orientation. Computer simulations of axon development integrating the enhanced plus-end-out microtubule growth identified in this studya with previously suggested mechanisms, that is, dynein-based microtubule sliding and augmin-mediated templating, correctly predicted the long-term evolution of axonal microtubule orientation as found in the experiments. This study thus leads to a holistic explanation of how axonal microtubules orient uniformly, a prerequisite for efficient long-range transport essential for neuronal functioning (Jakobs, 2022).
Microtubules are essential to neuron shape and function. Acetylation of tubulin has the potential to directly tune the behavior and function of microtubules in cells. Although proteomic studies have identified several acetylation sites in α-tubulin, the effects of acetylation at these sites remains largely unknown. This includes the highly conserved residue lysine 394 (K394), which is located at the α-tubulin dimer interface. Using a fly model, this study showed that α-tubulin K394 is acetylated in the nervous system and is an essential residue. Acetylation-blocking mutation in endogenous α-tubulin, K394R, was found to perturb the synaptic morphogenesis of motoneurons and reduces microtubule stability. Intriguingly, the K394R mutation has opposite effects on the growth of two functionally and morphologically distinct motoneurons, revealing neuron-type-specific responses when microtubule stability is altered. Eliminating the deacetylase HDAC6 increases K394 acetylation, and the over-expression of HDAC6 reduces microtubule stability similar to the K394R mutant. Thus, these findings implicate α-tubulin K394 and its acetylation in the regulation of microtubule stability and suggest that HDAC6 regulates K394 acetylation during synaptic morphogenesis (Saunders, 2022).
In response to nutritional stress, microtubules in cells of the Drosophila female germline are depleted from the cytoplasm and accumulate cortically. This triggers aggregation of mRNPs into large processing bodies (P-bodies) and oogenesis arrest. This study shows that hyperacetylation of α-tubulin at lysine 40 (K40) alters microtubule dynamics and P-body formation. Depletion of histone deacetylase 1 (HDAC1) by RNAi phenocopies the nutritional stress. response, causing α-tubulin hyperacetylation and accumulation of maternally deposited mRNPs in P-bodies. Through in vitro and in vivo studies, this study identified HDAC1 as a direct regulator of α-tubulin K40 acetylation status. In well-fed flies, HDAC1 maintains low levels of α-tubulin acetylation, enabling the microtubule dynamics required for mRNP transport. Using quantitative phosphoproteomics, nutritional stress-induced changes were identified in protein phosphorylation that act upstream of α-tubulin acetylation, including phosphorylation of HDAC1 at S391, which reduces its ability to deacetylate α-tubulin. These results reveal that Drosophila HDAC1 senses and relays the nutritional status, which regulates germline development through modulation of cytoskeleton dynamics (Wippich, 2023)
Microtubule acetylation is found in populations of stable, long-lived microtubules, occurring on the conserved lysine 40 (K40) residue of α-tubulin by α-tubulin acetyltransferases (αTATs). α-tubulin K40 acetylation has been shown to stabilize microtubules via enhancing microtubule resilience against mechanical stress. This study shows that a previously uncharacterized αTAT, Drosophila CG17003/leaky (lky), is required for α-tubulin K40 acetylation in early germ cells in Drosophila ovary. Loss of lky resulted in a progressive egg chamber fusion phenotype accompanied with mislocalization of germline-specific Vasa protein in somatic follicle cells. The same phenotype was observed upon replacement of endogenous α-tubulin84B with non-acetylatable α-tubulin84BK40A, suggesting α-tubulin K40 acetylation is responsible for the phenotype. Chemical disturbance of microtubules by Colcemid treatment resulted in a mislocalization of Vasa in follicle cells within a short period of time (~30 min), suggesting that the observed mislocalization is likely caused by direct leakage of cellular contents between germline and follicle cells. Taken together, this study provides a new function of α-tubulin acetylation in maintaining the cellular identity possibly by preventing the leakage of tissue-specific gene products between juxtaposing distinct cell types (Antel, 2022).
The formation and maintenance of microtubules requires their polymerisation, but little is known about how this polymerisation is regulated in cells. Focussing on the essential microtubule bundles in axons of Drosophila and Xenopus neurons, this study showed that the plus-end scaffold Eb1, the polymerase XMAP215/Msps and the lattice-binder Tau co-operate interdependently to promote microtubule polymerisation and bundle organisation during axon development and maintenance. Eb1 and XMAP215/Msps promote each other's localisation at polymerising microtubule plus-ends. Tau outcompetes Eb1-binding along microtubule lattices, thus preventing depletion of Eb1 tip pools. The three factors genetically interact and show shared mutant phenotypes: reductions in axon growth, comet sizes, comet numbers and comet velocities, as well as prominent deterioration of parallel microtubule bundles into disorganised curled conformations. This microtubule curling is caused by Eb1 plus-end depletion which impairs spectraplakin-mediated guidance of extending microtubules into parallel bundles. This demonstration that Eb1, XMAP215/Msps and Tau co-operate during the regulation of microtubule polymerisation and bundle organisation, offers new conceptual explanations for developmental and degenerative axon pathologies (Hahn, 2021).
Centrosome-mediated microtubule (MT) nucleation has been well characterized; however, numerous noncentrosomal MT nucleation mechanisms exist. The branching MT nucleation pathway envisages that the gamma-tubulin ring complex (gamma-TuRC) is recruited to MTs by the augmin complex to initiate nucleation of new MTs. While the pathway is well conserved at a molecular and functional level, branching MT nucleation by core constituents has never been directly observed in animal cells. In this study, multicolor TIRF microscopy was applied to visualize and quantitatively define the entire process of branching MT nucleation in dividing Drosophila cells during anaphase. The steps of a stereotypical branching nucleation event entailed augmin binding to a mother MT and recruitment of gamma-TuRC after 15 s, followed by nucleation 16 s later of a daughter MT at a 36 degrees branch angle. Daughters typically remained attached throughout their approximately 40-s lifetime unless the mother depolymerized past the branch point. Assembly of branched MT arrays, which did not require Drosophila TPX2, enhanced localized RhoA activation during cytokinesis (Verma, 2019).
Abl family kinases (see Drosophila Abl) are evolutionarily conserved regulators of cell migration and morphogenesis. Genetic experiments in Drosophila suggest that Abl family kinases interact functionally with microtubules to regulate axon guidance and neuronal morphogenesis. Vertebrate Abl2 binds to microtubules and promotes their plus-end elongation, both in vitro and in cells, but the molecular mechanisms by which Abl2 regulates microtubule (MT) dynamics are unclear. This study reports that Abl2 regulates MT assembly via condensation and direct interactions with both the MT lattice and tubulin dimers. Abl2 promotes MT nucleation, which is further facilitated by the ability of the Abl2 C-terminal half to undergo liquid-liquid phase separation (LLPS) and form co-condensates with tubulin. Abl2 binds to regions adjacent to MT damage, facilitates MT repair via fresh tubulin recruitment, and increases MT rescue frequency and lifetime. Cryo-EM analyses strongly support a model in which Abl2 engages tubulin C-terminal tails along an extended MT lattice conformation at damage sites to facilitate repair via fresh tubulin recruitment. Abl2Δ688-790, which closely mimics a naturally occurring splice isoform, retains binding to the MT lattice but does not bind tubulin, promote MT nucleation, or increase rescue frequency. In COS-7 cells, MT reassembly after nocodazole treatment is greatly slowed in Abl2 knockout COS-7 cells compared with wild-type cells, and these defects are rescued by re-expression of Abl2, but not Abl2Δ-790. It is proposed that Abl2 locally concentrates tubulin to promote MT nucleation and recruits it to defects in the MT lattice to enable repair and rescue.
Chromosome congression, the process of positioning chromosomes in the midspindle, promotes the stable transmission of the genome to daughter cells during cell division.
Congression is typically facilitated by DNA-associated, microtubule (MT)
plus end-directed motors called chromokinesins. The Drosophila melanogaster chromokinesin No distributive disjunction (NOD) contributes to congression, but the means by which it does so are unknown in large part because NOD has been classified as a nonmotile, orphan kinesin. It has been postulated that NOD promotes congression, not by conventional plus
end-directed motility, but by harnessing polymerization forces by
end-tracking on growing MT plus ends via a mechanism that is also
uncertain. This study demonstrates that NOD possesses MT plus
end-directed motility. Furthermore, NOD directly binds EB1 through unconventional
EB1-interaction motifs that are similar to a newly characterized MT tip
localization sequence. It is proposed that NOD produces congression
forces by MT plus end-directed motility and tip-tracking on polymerizing
MT plus ends via association with EB1 (Ye, 2018).
Equal distribution of duplicated DNA is required to maintain genomic stability through cell division. The microtubule (MT) cytoskeleton is reorganized to form a bipolar spindle as cells enter mitosis or meiosis. Chromosomes are positioned at the spindle equator during a process known as congression. Chromosome movements within the spindle are predominantly mediated by motor proteins that walk directionally on spindle MTs. An important class of congression motors is the chromokinesins, which include the kinesin families, kinesin-4, kinesin-10, and kinesin-12. Chromokinesins work cooperatively to promote chromosome alignment during cell division. Kinesin-10 and kinesin-4 are chromosome-associated, plus end-directed motors, although KIF4A suppresses plus end MT dynamics and may dampen polar ejection forces (PEFs) that push chromosome arms away from spindle poles, whereas kinesin-10's role is more intuitive and likely the predominant PEF-producing motor. The function of vertebrate kinesin-10 (Kid) was first described in Xenopus laevis egg extracts in which Xkid was required to establish and maintain chromosome arm congression. Although data from human cells have consistently shown that hKid contributes to congression, the alignment defects observed in tissue culture cells have not been as severe as in egg extracts. Vertebrate kinesin-10s have been shown to possess plus end-directed motility and to generate force when bound to chromatin (Ye, 2018).
The Drosophila melanogaster chromokinesin NOD shares sequence homology in its N-terminal motor with both kinesin-10 and kinesin-4 motor domains and in its C terminus with the kinesin-10 DNA-binding motif (helix-hairpin-helix), but it has been designated an orphan kinesin as a result of significant divergence in its structural elements and organization relative to conventional kinesins. NOD was initially identified and characterized genetically as the mutant no distributive disjunction (nod), which exhibited high frequencies of nondisjunction and chromosome loss in female meiosis. The nod gene encodes a kinesin-like protein (NOD) with an N-terminal motor domain. Achiasmate (nonexchange) chromosomes frequently failed to associate with spindles or were mispositioned near spindle poles in oocytes lacking functional NOD. The characterization of DNA binding activities in the C terminus of NOD, its N-terminal motor domain, and the misalignment phenotype of nod mutants led to the hypothesis that NOD is the PEF motor in fly oocytes, and later work in Drosophila tissue culture cells revealed a role for NOD in mitotic chromosome congression. Although NOD possesses a conserved N-terminal motor domain and MT-stimulated ATPase activity, efforts to reconstitute directional motility in vitro have failed leading to NOD being classified as a nonmotile kinesin. How could a nonmotile kinesin generate force? An alternative theory posits that, as a result of the unique mechanochemical properties of its motor domain, NOD moves chromosomes by associating with the plus ends of polymerizing MTs, although direct evidence for this mechanism is lacking. High resolution imaging of NOD-coated chromatin stretching events in living cells were suggestive of both plus end-directed motility and end-tracking coincident with EB1 comets (Ye, 2018).
NOD possesses an N-terminal motor domain and two distinct C-terminal DNA binding regions comprised of high mobility group (HMG) repeats and a helix-hairpin-helix motif that mediate chromatin-association of full length (FL) NOD-mCherry throughout the cell cycle in Drosophila S2 cells. Between the motor and DNA-binding domains, NOD is predicted to contain intrinsically disordered regions (~50%), as well as four α-helices, one of which has a low probability of forming a parallel two-stranded coiled coil (CC). To dissect NOD motor function in vivo, truncations of NOD tagged at their C termini with mCherry were expressed in GFP-α-tubulin expressing Drosophila S2 cells. The motor domain (1-324) did not bind MTs in mitosis or interphase and was diffuse in the cytosol. Thus, unlike many well-characterized motor proteins, the NOD324 motor domain exhibits weak or no MT binding activity in cells. Given the high physiological concentration of ATP, this observation is consistent with in vitro studies in which NOD motor domain exhibited significantly lower MT binding affinities in the presence of excess ATP than conventional kinesin motors (Ye, 2018).
Most kinesins with N-terminal motor domains possess a neck linker and a well-defined neck CC downstream of the motor; however, some kinesins, including NOD, have noncanonical neck regions that contribute to their functions in cells. The localization was examined of a NOD truncation (1-485) that encompasses the motor domain and the noncanonical neck extension region. Addition of the neck extension conferred MT-binding activity in cells as NOD485 uniformly coated MTs throughout the cell cycle. Inclusion of the nonconventional extension could promote MT binding through numerous nonexclusive mechanisms, including the introduction of a second MT binding site, which has been shown for hKID, posttranslational modifications, association with regulatory factors, or dimerization through the low probability CC. The MT localization of NOD485 was significantly reduced upon fixation conditions suggestive of the interaction, while stronger than NOD324, being relatively low affinity. Nonetheless, the localization of NOD485 led to the idea that the oligomeric state of NOD warranted further investigation. Several structural and functional aspects of NOD are reminiscent of the monomeric plus end-directed kinesin Unc104/KIF1A. Kinesin-10 family members have been shown to be monomeric in vitro. Furthermore, NOD and Unc104/KIF1A possess low-probability/weak CC regions adjacent to their N-terminal motors, whereas their C termini contain domains that cluster the motors on the surface of cargos: chromosomes and synaptic vesicles, respectively. Prior work on Unc104 demonstrated that constitutive dimerization of the Unc104 motor domain and its adjacent 'weak' CC converted the monomer into a processive, plus end-directed motor with physiological velocities. A comparable approach as that applied to Unc104, specifically fusion to the kinesin-1 'stalk' CC, was next used to examine how dimerization of the NOD motor domain affected its behavior (Ye, 2018).
The motor domain alone (1-324) was first dimerized. NOD324CC evenly coated interphase MTs and robustly associated with spindle MTs during mitosis, exhibiting a slight enrichment toward the plus ends of kinetochore fibers in the vicinity of aligned chromosomes with some evident MT bundling. NOD485CC exhibited a striking localization pattern in which it was highly enriched near MT plus ends in mitosis and interphase. Significant MT bundling was also observed throughout the cell cycle especially in cells with high NOD485CC levels, which was not observed in the NOD485-expressing cells. The localization patterns and behavior of the dimerized NOD truncations were not attributable to the kinesin-1 CC because CC-mCherry only weakly associated with a subset of MTs in some cells and exhibited no obvious or consistent localization pattern. Furthermore, the neck extension region, while necessary, was not sufficient for MT binding, as the localization patterns of NOD325-485 and NOD325-485CC were identical to mCherry and CC-mCherry, respectively (Ye, 2018).
The localization pattern of NOD485CC suggested dimerized NOD could possess plus end-directed motility in cells. This possibility was further examined by imaging cells using total internal reflection fluorescence (TIRF) microscopy to visualize NOD on the MTs closest to the cell cortex. Puncta of both NOD485 and NOD324CC were visible on MTs, but motility was not obvious. The behavior of NOD485CC was markedly different from the other truncations as NOD puncta were clearly observed moving processively on MTs toward the cell periphery. The distribution of fluorescence intensities of motile NOD485CC-mCherry puncta was indistinguishable from that of motile kinesin-1-mCherry dimers, demonstrating that NOD485CC was assembling into motile dimers. Measurements of motile NOD485CC dimers in cells yielded a mean velocity of 8.70 ± 3.61 micrometers/min. These data demonstrate that dimerization of the NOD motor domain alone (NOD324CC) promotes MT binding but not motility, most likely because it lacks a functional neck linker, which plays an important role in coordinating motor head functions, whereas dimerization of the motor domain and nonconventional neck extension region converts NOD into a directional motor (Ye, 2018).
To further characterize NOD motility in vitro, TIRF-based imaging was applied to cell extracts in which NOD485CC was the only fluorescently tagged component. Using this technique, NOD motility could be studied in near physiological conditions, but in a chamber in which the state of the MTs, which are attached to the cover-glass, as well as the buffering conditions can be controlled. In cell lysates prepared from NOD485CC-expressing cells, NOD walked unidirectionally on taxol stabilized MTs in an ATP-dependent manner. The mean velocity of 8.62 2.32 micrometers/min measured for motile NOD485CC dimers in cell lysates was indistinguishable from live-cell measurements (Ye, 2018).
NOD motility has never been reconstituted in vitro and, like others,this study could not purify active NOD485CC from bacteria. Furthermore, NOD485CC activity in cell lysates was labile and sensitive to buffer conditions. Thus, NOD485CC-mCherry was purified from Drosophila S2 cells using a C-terminal Strep tag that allows for affinity purification on a streptavidin-based matrix and gentle elution using biotin-containing buffers to better preserve protein activity. Silver staining of the purification revealed a unique band, when compared with a mock purification from wild-type cell extracts, at the predicted size of NOD485CC-mCherry (~110 kD), which was confirmed by Western blot using an mCherry antibody. Although it is probable that NOD-associated proteins were copurified in the preparation, the majority of background bands were likely biotinylated proteins or nonspecific because a nearly identical banding pattern was observed in mock purifications from wild-type cell extracts. In TIRF assays, the affinity purified NOD485CC-mCherry exhibited ATP-dependent, unidirectional motility, although the mean velocity of 5.79 ± 1.56 micrometers/min was slower than velocities measured in cells and lysates (Ye, 2018).
NOD485CC dimers often moved toward the cell periphery where the MT plus ends are typically oriented during interphase. To define the directionality of NOD485CC motility, purified GFP-labeled human kinesin-1 (Kif5B) motor, a plus end-directed motor, was added to lysates from NOD485CC-mCherry expressing cells. NOD485CC and kinesin-1 walked in the same direction, establishing that NOD is a plus end-directed motor. Collectively, the data in cells and in vitro demonstrate that dimerized NOD is a plus end-directed motor with velocities similar to those measured for Xkid and hKID, which in combination with the presence of a C-terminal helix-hairpin-helix DNA-binding motif warrants consideration of NOD as the Drosophila kinesin-10 orthologue (Ye, 2018).
Next, it was tested if chemically induced dimerization was sufficient to support NOD motility by building cell lines coexpressing dark (no fluorescent tag) NOD485-FRB and NOD485-FKBP-EGFP. Like NOD485, NOD485-FKBP-EGFP localized uniformly to MTs in the absence of rapamycin. Upon addition of 100 nM rapamycin, motile puncta of NOD485-FKBP-EGFP moved on MTs toward the cell periphery. Extracts were next prepared from NOD485-FKBP-EGFP, NOD485-FRB-expressing cells to visualize NOD on MTs by TIRF microscopy in vitro. Without rapamycin, nonmotile puncta of NOD485-FKBP-EGFP associated with MTs. Unidirectional motility of NOD485-FKBP-EGFP puncta was achieved within minutes of adding 100 nM rapamycin to the same cell extracts that did not exhibit motility in the absence of rapamycin. Cell lines coexpressing NOD485-FKBP-EGFP and NOD485-FRB-mCherry were next visualized. In the absence of rapamycin, the mCherry and EGFP-tagged NOD uniformly coated the MTs similar to NOD485, but NOD localization changed upon addition of 100 nM rapamycin as EGFP- and mCherry-tagged, chemically dimerized NOD485 accumulated toward MT plus ends at the cell periphery similar to NOD485CC. Altogether, the kinesin-1 CC and chemically induced dimerization data support the conclusion that multiple modes of NOD485 dimerization can support directional motility (Ye, 2018).
Although earlier work visualizing MT-associated stretching of NOD-coated chromatin was indicative of plus end-directed motility (Cane, 2013), it is technically challenging to assess the activity of nonartificially dimerized NOD in cells as a result of its constitutive chromatin localization. Fortuitously, NOD contains an endogenous nuclear export signal (NES) computationally predicted and borne out by the localization of various NOD truncations. Although NOD possesses an NES, it is not typically observed in the cytosol because it is tightly associated with chromatin throughout the cell cycle. It was reasoned that the affinity of NOD for chromatin could be reduced by deleting one of its two chromosome-associating domains (HMG14/17 repeats), a truncation called NODΔHMG. In ~10% of cells expressing NODΔHMG, possibly as a result of reduced affinity for the chromatin and the presence of an NES, cytosolic NOD puncta were visible. Therefore, it was possible to visualize nearly FL cytosolic NOD that was not artificially dimerized with CC or FKBP-FRB domains. Importantly, the cytosolic NODΔHMG exhibited plus end-directed motility toward the cell periphery along interphase MTs. Compared with the kinesin-1-mCherry standard, motile NODΔHMG puncta spanned a range of fluorescence intensities corresponding to ~5-200 NOD molecules with a mean of 52 motors per puncta, suggesting that NOD may function as clusters of dimers or more complex oligomers. Nevertheless, the velocity of NODΔHMG clusters was comparable to the velocities measured for NOD485CC dimers in cells and cell lysates in vitro (7.6 micrometers/min for NODΔHMG in cells vs. 8.6-8.7 micrometers/min for NOD485CC in cells and lysates) (Ye, 2018).
Prior work characterized NOD-coated chromatin stretch events that colocalized with EB1 comets, as well as occasional cytoplasmic NOD fragments that tracked polymerizing and depolymerizing MT plus ends (Cane, 2013), and NODΔHMG puncta also tracked growing plus ends. Spinning disc confocal imaging of cells coexpressing NOD485CC-EGFP and EB1-TagRFP-T revealed that a subset of NOD485CC puncta colocalized with EB1 tracks on MT plus ends. It is worth noting that NOD often remained associated with depolymerizing MTs through a mechanism that is presently unclear, but that could be similar to the bidirectional tracking activity of CENP-E because NOD, like hKID, may possess low-affinity MT binding regions outside of the motor domain. Plus end-directed NOD485CC dimers were evident behind tip-tracking NOD puncta in kymographs from NOD485CC-, EB1-expressing cells. Given the utility of cell lysates in visualizing NOD motility, extracts from NOD485CC-EGFP-expressing cells were added to dynamic rather than taxol-stabilized MTs in an effort to better visualize NOD end tracking by TIRF microscopy in vitro. The cell-based observations were fully validated in vitro using dynamic MTs as NOD485CC clearly walked toward the faster growing plus ends while a subpopulation of NOD485CC tip tracked on growing MT plus ends (Ye, 2018).
NOD's end-tracking activity combined with the observed colocalization of NOD with EB1 comets led to an investigation of whether NOD and EB1 directly interact in pulldown assays using purified components. GST-EB1-TagRFP-T pulled down with MBP-NOD485CC-EGFP, but not the MBP-EGFP control protein. To further characterize the NOD-EB1 interaction, attention turned to amino acids 325-485 because NOD485CC exhibited MT plus end-tracking activity, and NOD324CC did not. NOD contains multiple regions that are predicted to be disordered, centered around proline (P) and threonine (T) residues between residues 430-480. These motifs are similar to unconventional EB1-binding motifs recently identified in fungal species in the Saccharomyces cerevisiae Kar9 motor and Schizosaccharomyces pombe Dis1/Tog MAP. To identify EB1-interacting regions in NOD, overlapping peptide SPOT arrays encompassing what were deemed the 'PT' motifs were synthesized onto a cellulose membrane, and their interactions with purified EB1 were probed. The SPOT arrays comprised 15 amino acid peptides spanning NOD residues 430-480 with an offset of two residues and included a 'perfect' SxIP aptamer as a positive control peptide. The NOD peptides exhibiting the strongest association with purified Drosophila EB1-TagRFP-T centered on the PT motifs (Ye, 2018).
The amino acids surrounding SxIP motifs impact their affinities for EB1. To examine how PT-flanking amino acids contributed to EB1 binding, the peptides that exhibited the strongest association with EB1 on the array were subjected to alanine scanning. In each peptide, mutation of a surrounding arginine (R) residue approximately three to six amino acids upstream or downstream of the PT sequence eliminated EB1 binding. The results suggest that, like the SxIP motif, neighboring basic residues contribute to the affinity of NOD PT motifs for EB1. During the preparation of this manuscript, the unconventional Ka9-EB1 interaction was further characterized, leading to the designation of LxxPTPh as a novel tip-localization motif. Although the PT motifs in NOD resemble the LxxPTPh motif, the compositional differences may indicate that the Drosophila EB1 PT motif has diverged somewhat and/or that variability in the motif still supports EB1 interactions (Ye, 2018).
Microscale thermophoresis (MST) was used to measure the binding affinity of the NOD PT motif-1 for EB1. The N terminus of either the PT motif-1 or a bona fide 'perfect' SxIP peptide (positive control) was labeled with fluorescein (FITC) dye, and the movement of each fluorescently labeled peptide (fixed at 50 nM) in a temperature gradient was measured by MST while varying concentrations of unlabeled Drosophila GST-EB1. The binding affinity measured by MST for the SxIP motif was 807 nM, which is relatively close to the Kd (~570 nM) measured by isothermal titration calorimetry using the 'perfect' SxIP peptide and Drosophila EB1. The affinity of the PT motif-1 for Drosophila EB1 was measured to be 725 nM, slightly higher than the Kd of the positive control SxIP aptamer. Thus, the NOD PT motif-1 possesses an affinity for Drosophila EB1 that is comparable to other bona fide SxIP motifs and higher than affinities measured for EB1-CAP-Gly interactions (Ye, 2018).
How the Drosophila kinesin-10 NOD generates PEFs has long been a mystery. Prior work led to a hypothesis that NOD possesses two force producing activities: (1) MT plus end-directed motility, which had never been directly demonstrated and (2) end tracking on polymerizing MTs, the mechanism of which was speculative. This study demonstrates, for the first time, that NOD exhibits plus end-directed motility in cells and in vitro and that NOD directly interacts with the tip-tracking protein EB1 via a new type of conserved MT tip localization sequence. It is noteworthy that both MT polymerization and motor activity were originally proposed as sources of the PEF when the phenomenon was first described, more than 30 years later the current findings reveal NOD as a molecular nexus of both force-producing mechanisms (Ye, 2018).
Cortical collapse factors affect microtubule (MT) dynamics at the plasma membrane. They play important roles in neurons. How cortical collapse factors influence axon growth is little understood. This study focussed on the function of Drosophila Efa6 in experimentally and genetically amenable fly neurons. First, it was shown that Drosophila Efa6 can inhibit MTs directly without interacting molecules via an N-terminal 18 amino acid motif (MT elimination domain/MTED) that binds tubulin and inhibits microtubule growth in vitro and cells. If N-terminal MTED-containing fragments are in the cytoplasm they abolish entire microtubule networks of mouse fibroblasts and whole axons of fly neurons. Full-length Efa6 is membrane-attached, hence primarily blocks MTs in the periphery of fibroblasts, and explorative MTs that have left axonal bundles in neurons. Accordingly, loss of Efa6 causes an increase of explorative MTs: in growth cones they enhance axon growth, in axon shafts they cause excessive branching, as well as atrophy through perturbations of MT bundles. Efa6 over-expression causes the opposite phenotypes. Taken together, this work conceptually links molecular and sub-cellular functions of cortical collapse factors to axon growth regulation and reveals new roles in axon branching and in the prevention of axonal atrophy. Furthermore, the MTED delivers a promising tool that can be used to inhibit MTs in a compartmentalised fashion when fusing it to specifically localising protein domains (Qu, 2019).
Axons are the cable-like neuronal extensions that wire the nervous system. They are only 0.1–15 μm in diameter, but can be up to a meter long in humans. It is a fascinating challenge to understand how axons can extend over these enormous distances and branch in orderly manners, but also how these delicate structures can be maintained for a lifetime, that is many decades in humans. It is not surprising that humans gradually lose about 40% of their axons towards old age, and that axon decay is a prominent neurodegenerative phenomenon (Qu, 2019).
Essential for axon biology are the parallel bundles of microtubules (MTs) running all along the axon shaft; these bundles provide (1) structural support, (2) highways for life-sustaining cargo transport, and (3) a source of MTs that can leave these bundles to drive morphogenetic changes. Through being organised in this way, MTs essentially drive processes of axon growth, branching and maintenance. The dynamics of MTs are orchestrated through MT-binding and -regulating proteins, for most of what is known about the molecular mechanisms of function. However, such knowledge alone is usually not sufficient to explain their cellular roles (Qu, 2019).
For example, cortical collapse factors are cell surface-associated proteins which specifically inhibit MTs that approach the cell periphery. Previous reports suggested important roles for cortical collapse factors in regulating axon growth: the ARF activator Efa6 (exchange factor for ARF6) in C. elegans negatively impacts on developmental and regenerative axon growth (Chen, 2015; Chen, 2011; O'Rourk., 2010); the mammalian type four kinesin KIF21A also affects axon growth and links to the neurodevelopmental eye movement disorder 'congenital fibrosis of extraocular muscles' (OMIM reference #135700). However, it can currently only be hypothesized how the molecular functions of these two collapse factors link to axon growth, most likely by acting in growth cones (GCs) (Qu, 2019).
GCs are the amoeboid tip structures through which axons extend to wire the nervous system during development or regeneration. The axonal MT bundles terminate in the centre of GCs; from here, single MTs splay into the actin-rich periphery of GCs. These explorative MTs can trigger extension of the entire MT bundle into their direction, thus elongating the axon; by inhibiting such explorative MTs, cortical collapse factors could negatively impact on axon growth (Qu, 2019).
In line with this argument, and depending on where cortical collapse factors are present and functionally active, further functional predictions could be made: for example, collateral branching of axons along their shafts has been described to depend on explorative MTs that leave the parallel axonal bundles and polymerise towards the periphery. Cortical collapse factors might therefore be negative regulators of axon branching (Qu, 2019).
Other roles might concern axon maintenance: the model of 'local axon homeostasis' states that the force-enriched environment in axons biases MTs to buckle or project out of the bundle to seed pathological areas of MT disorganisation. By inhibiting off-track MTs in the axon shaft, cortical collapse factors might prevent such processes, acting in parallel to other bundle-maintaining factors. For example, spectraplakins serve as spacers that keep polymerising MTs away from the cortex by linking the tips of extending MTs to the axonal surface and guiding them into parallel bundles. Their deficiency in any organism causes severe MT disorganisation, potentially explaining human dystonin-linked HSAN6 ('type six hereditary sensory and autonomic neuropathy'; OMIM ID: 614653). If the hypothesis is correct, loss of cortical collapse factors in axon shafts would also cause MT disorganisation, but through a very different mechanistic route (Qu, 2019).
This study makes use of Drosophila neurons as a well-established, powerful model for studying roles of MT regulators. Using in vitro and cellular assays, this study showed that Drosophila Efa6 is a cortical collapse factor acting through its N-terminal MT elimination domain (MTED). The MTED binds tubulin and blocks MT polymerisation in vitro which indicates that the effect of the peptide is due to a direct interaction between the peptide and tubulin. By localising to neuronal membranes, it only abolishes explorative MTs. This subcellular role translates into negative regulation of axon growth and branching and the prevention of pathological MT disorganisation, both in cultured neurons and in vivo. It is proposed that Efa6 functions as a quality control or axonal maintenance factor that keeps explorative MTs in check, thus playing a complementary role to spectraplakins that prevent MTs from leaving axonal bundles (Qu, 2019).
Axons are the structures that wire the brain and body and are therefore fundamental to nervous system function. To understand how axons are formed during development, can be maintained in a plastic state thereafter, and why they deteriorate in pathological conditions, it is necessary to increase knowledge of axonal cell biology. The MT bundles that form the core of axons are an essential aspect of this cell biology, and understanding how these bundles are regulated and contribute to axon morphogenesis will provide essential insights into axon development and maintenance. This study has addressed fundamental contributions made by cortical collapse factors. This work started from reports that two such factors from distinct protein families both negatively impact on axon growth in species as diverse as C. elegans (CeEfa6) and mouse (Kif21A) (Qu, 2019).
This study found that DmEfa6 likewise acts as a negative regulator of axon growth. Efa6 is a cortical collapse factor, inhibiting MTs primarily via the 18aa long MTED. Since the MTED is the only shared motif with CeEfa6 in an otherwise entirely divergent N-terminus, this clearly demonstrates that the MTED is functionally conserved between both species (Qu, 2019).
Capitalising on Drosophila neurons as a conceptually well-established model for studies of axonal MT regulation, two novel roles were demonstrated for Efa6: as a negative regulator of axon branching and a quality control factor maintaining MT bundle organisation. To perform these functions, Efa6 does not affect the dynamics of MTs contained within the central axonal bundles, but it inhibits mainly those MTs that leave these bundles (see A model for axonal roles of Efa6). By inhibiting explorative MTs in GCs, it negatively impacts on a key event underlying axon growth. By inhibiting off-track MTs in the axon shaft, it tones down the machinery that seeds new interstitial branches, but also prevents these MTs from going astray and causing MT disorganisation (Qu, 2019).
Therefore, this work provides conceptual understanding of cortical collapse factors, which can explain how their molecular functions and subcellular roles in MT regulation link to their reported axonal growth phenotypes during development and regeneration, and to their additional functions in axon branching and maintenance. Apart from existing links of cortical collapse factors to neurodevelopmental disorders, it is therefore predicted that future links will be made to neurodegeneration (Qu, 2019).
During axon growth, MTs constantly polymerise towards the periphery of GCs; the advance of many of these MTs is inhibited at the leading edge, and this work shows that cortical collapse factors are key mediators to this end. Only a fraction of MTs enters filopodia, potentially helped by active guidance mechanisms such as MT-actin cross-linkage (e.g. through spectraplakins, tau, drebrin-EB3). The widely accepted protrusion-engorgement-consolidation model of axon growth proposes that stabilised MTs in filopodia can seed axon elongation events. This model is consistent with the current findings for Efa6. Thus loss of Efa6 can contribute to enhanced axon growth in two ways: firstly, through allowing more MTs to enter filopodia; secondly, by allowing them to dwell in filopodia for longer, thus enhancing the likelihood of their stabilisation. This scenario can explain why loss of Efa6 in C. elegans improves axon re-growth after injury and growth overshoot during development, and why the higher levels of Kif21A levels in GCs causes stalled axon growth (Qu, 2019).
In C. elegans it was shown that axonal injury leads to a re-localisation of CeEfa6 to MT minus ends in the axon core. None of the conditions used in the current study reproduced such behaviour with fly Efa6. Furthermore, it was shown that such central pools of CeEfa6 require their MTED to recruit two kinases: TAC-1 (homologue of TACC/transforming-acidic-coiled-coil) and ZYG-8 (homologue of DCLK/Doublecortin-Like-Kinase). However, in contrast to Efa6, both of these kinases perform growth-enhancing functions and play a secondary, delayed role downstream of Efa6. They are therefore unsuited to explain the direct MT-inhibiting roles of the MTED. In contrast, virtually all structure-function analyses performed with CeEfa6 in developing and regenerating axons perfectly match the data and can be explained through the proposed model. Based on these findings, one might argue that CeEfa6 detachment from the membrane could be the consequence of injury-induced physiological changes that would then pose a threat to axonal MT bundles; localisation to MT minus ends could therefore represent a protective sequestration mechanism. Another C. elegans study reported that loss of Efa6 has no impact on MT length in developing axons, which appears consistent with the current data. They also found an increase in MT numbers, but there is currently no mechanism to explain this in non-injury conditions where CeEfa6 stays at the membrane (Qu, 2019 and references therein).
Interestingly, mammalian Efa6 also plays a role in axon regeneration. However, this mechanism is entirely different, in that it requires the C-terminus to activate Arf6 which, in turn, regulates integrin trafficking at the axon initial segment (Qu, 2019 and references therein).
Axon branching can occur via GC split, in that diverging MTs get stabilised in parallel in the same GC. Alternatively, it can occur through interstitial branching which involves the active generation (e.g. through MT severing) and then stabilisation of off-track MTs. Both models agree with observations in Efa6-deficient/over-expressing neurons: greater/lower numbers of MTs were found in GC and shaft filopodia at 6 hours in vitro, which then correlate with enhanced/reduced axonal branch numbers in mature neurons (Qu, 2019).
If interstitial branch formation is negatively regulated by Efa6, this poses the question as to whether Efa6 has to be actively down-regulated in healthy neurons for branching to occur. Efa6 could either be physically removed from future branch points or its MT inhibition function could be switched off. However, no such regulation appears to be required because Efa6 seems to be in a well-balanced equilibrium. Enough Efa6 appears to be present to inhibit occasional, likely accidental off-track MTs; this capacity is surpassed when the number of off-track MTs is actively increased, for example through MT severing proteins during axonal branch formation. Such a saturation model is supported by experiments with shot: filopodial MT numbers are elevated in shot mutant neurons, although Efa6 is present and functional (as demonstrated by the further increase in filopodial MT numbers in shot Efa6 double-mutant neurons). This is consistent with a model where Efa6 function occurs at a level that is easily saturated when increasing the number of explorative MTs. Such a view would also explain why loss of CeEfa6 promotes axon regeneration in C. elegans, in that the constant base-line of MT inhibition present in the wild-type, is removed in the mutant condition, thus favouring growth-mediating explorative MTs (Qu, 2019).
Axonal MT disorganisation in Efa6-deficient neurons occurs gradually and can even be induced by knock-down of Efa6 at mature stages. Therefore, Efa6 appears to prevent MT disorganisation during axon development and maintenance, as is consistent with its continued expression in the nervous system. Such a continued role makes sense in a scenario where MT bundles remain highly dynamic throughout a neuron's lifetime, constantly undergoing polymerisation to drive renewal processes that prevent senescence (Qu, 2019).
Based on these findings, it is proposed that Efa6 acts as a quality control or maintenance factor within a model of 'local axon homeostasis'. This model states that MTs in the force-enriched environment of axons have a tendency to go off-track and curl up, thus potentially seeding MT disorganisation. Different classes of MT-binding regulators, amongst them spectraplakins, prevent this by actively promoting the bundled conformation. It is proposed that cortical collapse factors act in a complementary way to spectraplakins in that they play no role in maintaining MTs in bundles, but they inhibit those MTs that have escaped the bundling mechanisms (Qu, 2019).
In this scenario, MTs are protected from cortical collapse as long as they are actively maintained in axonal bundles; this can explain the long known conundrum of how axonal MTs extend hundreds of micrometres in relative proximity to the cell cortex in axons, whereas in non-neuronal cells cortical proximity of MTs tends to trigger either their inhibition or tethered stabilisation (Qu, 2019).
This study found that the MTED motif correlates well with MT inhibiting functions of Efa6 family members, whereas the rest of the N-terminus bears no obvious further similarity. Experiments with N-terminal protein and synthetic MTED peptide, both reveal association with MTs/tubulin. The MTED strongly interferes with MT polymerisation. Future co-crystallisation experiments are required to reveal how the MTED works. Given its small size it is hypothesised that it simply blocks assembly, rather than acting via more complex mechanisms such as active promotion of depolymerisation (e.g. kinesin-8 and −13, XMap215) or severing (e.g. spastin, katanin, fidgetin (Qu, 2019).
In any case, the small size of MTEDs might come in handy as experimental tools to inhibit MTs, potentially displaying complementary properties to existing genetic tools such as the kinesin-13 Kif2C, stathmin or spastin. Importantly, the experiments with the CAAX domain have shown that Efa6's MT inhibiting function can be targeted to specific subcellular compartments to clear them of MTs, thus opening up a wide range of future applications (Qu, 2019).
Interestingly, the MT-inhibiting role of Efa6 seems not to be conserved in chordates when taking the MTED as indicator for this function. However, roles of cortical collapse factors in neurons seem to have been taken over by other proteins such as the kinesin-4 family member Kif21A. The CFEOM1-linked Kif21AR954W mutation causes the protein to relocate from the axon shaft to the growth cone of cultured hippocampal neurons. In consequence, increased Kif21A levels in GCs cause reduced axon growth - and this study observed the same with Efa6 over-expression. The decreased levels of Kif21A in proximal axons correlate with a local increase in side branches - and the same is observed with Efa6 loss of function (Qu, 2019).
Finally, this study found that the C-terminal domains of Efa6 might display some degree of functional conservation. So far, work on mammalian PSDs has revealed functions for C-terminal domains in regulating ARF6, ARF1 or ARL14 during actin cytoskeletal reorganisation and membrane ruffling, tumour formation, axon regeneration and immune regulation. The finding that PSD1 and C-terminal Efa6 constructs cause similar membrane ruffling phenotypes in fibroblasts, suggests that some conserved functions reside in this region and might further contribute, together with N-terminally mediated MT inhibition, to the neuronal or non-neuronal defects that cause semi-lethality displayed by Efa6 mutant flies (Qu, 2019).
It is proposed that Efa6 acts as a cortical collapse factor which is important for the regulation of axonal MTs and relevant for axon growth, maintenance and branching. Although this function of Efa6 is evolutionarily not widely conserved, these findings provide a helpful paradigm for studies of other classes of cortical collapse factors also in mammalian neurons. Promising research avenues will be to refine the mechanistic understanding of how Efa6 blocks MT polymerisation, not only to better understand how it can be regulated in axons, but also to better exploit MTEDs as molecular tools in cell biological research (Qu, 2019).
Drosophila Ringmaker (Ringer) is homologous to the human Tubulin Polymerization Promoting Proteins (TPPPs) that are implicated in the stabilization and bundling of microtubules (MTs) that are particularly important for neurons and are also implicated in synaptic organization and plasticity. No in vivo functional data exist that have addressed the role of TPPP in synapse organization in any system. This study presents the phenotypic and functional characterization of ringer mutants during Drosophila larval neuromuscular junction (NMJ) synaptic development. ringer mutants show reduced synaptic growth and transmission and display phenotypic similarities and genetic interactions with the Drosophila homolog of vertebrate Microtubule Associated Protein (MAP)1B, futsch. Immunohistochemical and biochemical analyses show that individual and combined loss of Ringer and Futsch cause a significant reduction in MT loops at the NMJs and reduced acetylated-tubulin levels. Presynaptic over-expression of Ringer and Futsch causes elevated levels of acetylated-tubulin and significant increase in NMJ MT loops. These results indicate that Ringer and Futsch regulate synaptic MT organization in addition to synaptic growth. Together these findings may inform studies on the close mammalian homolog, TPPP, and provide insights into the role of MTs and associated proteins in synapse growth and organization (Shi, 2019).
While regulation of synaptic MTs and the range of proteins that affect synaptic MT organization and function are not well-characterized, synaptic MTs have been implicated in regulating synaptic bouton growth. Thus, understanding the regulation of MT assembly, organization and dynamics in synaptic terminals is crucial for understanding synapse development and function. The current findings demonstrate that loss of Ringer affects synaptic bouton growth at the NMJ. The growth of NMJ synapses in Drosophila has been postulated to occur either through a process called intercalation where existing synaptic boutons space apart with new boutons inserted between them, or by end addition where new boutons are added at the ends of existing string of boutons. Synapse growth is also thought to occur from budding of existing boutons. While future studies will determine which of these processes may be compromised in ringer mutants leading to a reduction in the number of NMJ synaptic boutons, Ringer can be added to the increasing repertoire of proteins involved in the modulation of synaptic growth. It is likely that the regulation of the cytoskeleton by Ringer may have a profound impact on the balance between synaptic growth and stability. Since synaptic growth can be under both positive and negative regulations, one issue of interest would be to determine what genes are upstream and downstream of Ringer and define a signaling cascade that modulate synaptic growth (Shi, 2019).
The observations that the apposition of the presynaptic AZ protein, BRP, with the GluR receptor fields were not severely disrupted in ringer mutants suggest that Ringer might not be crucial for proper placement of pre- and post-synaptic specializations at the synaptic boutons. Interestingly, number of BRP-positive puncta/bouton area was significantly increased in ringer mutants than wild type. The synaptic ultrastructure of ringer mutants also revealed an increase in AZ number as well as disrupted AZ morphology. Thus, one possibility is that Ringer may directly play a role in AZ organization by interacting with BRP or indirectly through other proteins. It is also possible that disorganized MTs due to loss of Ringer may simply impact the proper assembly of AZs in the synaptic boutons. While elucidating the role of Ringer in AZ organization is an interesting topic of future research, it is important to note that this role of Ringer may or may not be dependent on Futsch. Recent findings report that futsch mutants, contrary to ringer mutants, have a decrease in AZ number and density at the larval NMJs but normal AZ ultrastructural morphology) further underscoring the fact that these proteins may coordinate unique axonal cytoskeletal functions during synapse organization (Shi, 2019).
ringer mutants showed a decrease in bouton numbers but an increase in AZs/bouton area as revealed both by Brp immunostaining and EM analyses. This phenotype could result in unchanged spontaneous firing of the minis as is reflected from no significant changes in mEJP frequency. At the same time the evoked EJP amplitude was decreased in ringer mutants. The increase in AZ numbers did not translate directly into increased miniature frequency, as loss of Ringer may also affect synapse ultrastructure that could still be abnormal at the more molecular level. Analysis of the synaptic vesicles (clustered and docked) at the AZs in the presynaptic terminals of ringer mutants also did not reveal any significant differences compared to controls. It is quite likely that significant decrease in EJP amplitude and quantal content might reflect a lower release probability and possibly defects in the synaptic release machinery. Altogether, Ringer loss reflects a presynaptic defect in neurotransmission machinery (Shi, 2019).
Both present and previously published studies on Ringer (Mino, 2016) reveal an interesting spatio-temporal pattern and differential levels of Ringer localization in the Drosophila nervous system during development. Ringer displayed a temporally dynamic expression in neurons during early embryonic stages followed by an expression at the midline glia during later stages of embryonic ventral nerve cord development (Mino, 2016). Interestingly, in vertebrates, TPPP is predominantly expressed in the CNS oligodendrocytes and plays a critical role in myelin maturation (Skjoerringe, 2006; Lehotzky, 2010; Ota, 2014). Given Ringer's localization in both neuronal and glial cell types in the Drosophila embryonic CNS, it is possible that mammalian TPPP may also be expressed at lower/undetectable levels in neurons in physiological conditions. Under pathological conditions though, TPPPs are reported to be enriched and colocalize with α-Synuclein in neuronal and oligodendroglial inclusions that are characteristic of Synucleinopathies. Ringer also has differential levels of wild type localization in third instar larvae as it is expressed at higher levels in larval axons (Mino, 2016) but at much lower levels at the presynaptic NMJ terminals. The NMJ localization is mostly cytoplasmic but also seems to associate with Futsch, which localizes at higher levels to the core MT cytoskeleton (Shi, 2019).
MT assembly and dynamics are regulated by several factors and mechanisms, such as MT-assembly promoting factors, MT stabilizing/destabilizing factors, MT severing proteins and MT post-translational modifications that affect MT stability. As cells respond to physiological needs, they constantly adapt their MT arrays by modulating the balance between dynamic and stable MT subpopulations. This is also achieved through acetylation which occurs primarily on MTs and can be abundant on long lived stable MTs. These studies revealed that Ringer together with Futsch regulates levels of Ac-Tub at the NMJ with single and double mutants displaying significantly decreased levels of acetylation. These in vivo findings are in line with previously reported cell culture data showing down regulation of TPPP by specific si-RNA resulted in decrease of Ac-Tub levels. The control of acetylation level of MT network is an important factor for the regulation of MT architecture and maintenance of its integrity. The current data suggest that one of the aspects of Ringer functions would thus be to regulate the MT architecture possibly by regulating levels of MT acetylation (Shi, 2019).
The stabilization of MTs during neuronal maturation also underlies axonal specification and growth. Data from Drosophila have shown that the conversion of a motile growth cone into a presynaptic terminal is associated with the appearance of a hairpin MT loop in the growth cone. Homozygous mutations in both ringer and futsch alter MT loop formation, a process that has been implicated as a phenomenon reflective of MT stability and budding of new boutons. While individual and combined loss of ringer and futsch resulted in reduced levels of synaptic Ac-Tub and reduction in NMJ MT loops, overexpression of Ringer and Futsch showed the opposite. These findings are consistent with the in vitro cell culture experiments and biochemical Tubulin assays that showed that Ringer affects MT polymerization; with Ringer-expressing cells forming a circular ring instead of regularly distributed MTs (Mino, 2016). Vertebrate MAP1B may also be involved in MT loop formation as revealed by in vitro overexpression of MAP1B (Shi, 2019).
There is also a group of MT-severing proteins that regulate synaptic MT stability and growth at the NMJ. These are Spastin and Katanin 60. Spastin is enriched in axons and is highly abundant in presynaptic terminals. Knockdown of Spastin causes a severe reduction in synaptic arbor and an increase in stable and looped MTs at synaptic terminals. Similarly, loss of Katanin 60 also resulted in increased MT loops and levels of Ac-Tub suggesting that these protein functions are contrasting to that of Ringer and Futsch. Vertebrate Spastin is critically required for axonal outgrowth during zebrafish embryonic development. Also, axon branch loss at the developing mouse NMJ is mediated by branch-specific MT severing by Spastin, which results in local disassembly of the MT cytoskeleton with subsequent dismantling of branches. Mutations in Spastin have also been associated with increased stabilization of MT network. Recently, it has also been shown that in HeLa cells, the two isoforms of Spastin harboring a missense mutation increases the levels of Ac-Tub. Thus, the broader implications from all of these findings could be that a fine balance of acetylation/de-acetylation kinetics may underlie proper MT organization and synaptogenesis (Shi, 2019).
The primary intracellular target of TPPP is tubulin/MT under both in vitro and in vivo conditions and displays extensive MT bundling activity (Hlavanda, 2002; Mino, 2016). One of the crucial factors affecting the function of MT network is its acetylation by the action of acetyltransferase complex as well as histone deacetylase 6 (HDAC6) and Sirtuin-2 (SIRT2). In vitro studies suggest that mammalian TPPP modulates MT acetylation by binding to HDAC6 and inhibits its activity, resulting in a reciprocal increase in MT acetylation (Tokesi, 2010). HDAC6 is commonly considered to be a tubulin-deacetylase because chemical inhibition of this enzyme significantly increases MT acetylation in neurons. Similar to HDAC6, a more recent study showed the tubulin deacetylase (SIRT2) to play a role with TPPP in regulating MT dynamics and stability. Thus, TPPP-directed deacetylase inhibition can be speculated as one of the mechanisms for the fine control of the dynamics and stability of the MT network. It will be interesting to further investigate whether Drosophila Ringer and/or Futsch may form a larger molecular complex that involves aspects of HDAC6 and SIRT2 in regulating MT dynamics and potentially synaptic growth at the NMJs. In vitro studies have also demonstrated that TPPP influences MT dynamics by decreasing the growth velocity of MT plus ends. While future studies will investigate how Drosophila Ringer modulates the dynamics and stability of the MT network, one can speculate based on the findings from the vertebrate TPPP, that these mechanisms could involve its MT assembly promoting, cross-linking and/or acetylation enhancing activities (Shi, 2019).
The biochemical analyses of Ringer reported in this study provide important insights into its role in regulating the MT cytoskeleton. It is interesting that the overall levels of Ringer did not change in futsch mutants compared to the control. This finding was consistent whether the total Ringer levels were assayed from larval tissues or adult head lysates. However, while the total Ringer levels were unchanged, the synaptic Ringer localization displayed a significant alteration compared to control raising the possibilities that, in the absence of Futsch, either Ringer levels significantly decreases in the presynaptic terminals or Ringer just fails to localize in its proper place and instead gets diffuse. However, total Tub levels and that of Ac-Tub were consistent with what was observed at the synapses. Irrespective of tissue type, these findings reveal a remarkable consistency in demonstrating that Ringer and Futsch regulate synaptic and overall MT stability: (1) Ac-Tub levels in synapses, (2) synaptic MT loops and (3) total Ac-Tub levels, each of these parameters were found to be affected similarly with a reduction in individual and combined loss of ringer and futsch and an elevation in their respective overexpression (Shi, 2019).
Although not in the context of intercellular protein-protein interactions in the synapses, there are reports of some TPPP interacting proteins. Consistent with published reports, Ringer being a Tub-binding protein was further reiterated by their presence in the IP complex. As expected, the MAP1B/Futsch also existed in a complex with Tub. Interestingly, while endogenous Ringer and Futsch could not be detected in the same IP complex, m-Cherry tagged Ringer was detected from an overexpression experimental paradigm. These datasets are reflective of an inability of the endogenous proteins to be detected either due to: (1) a huge difference in their molecular weights (Ringer being ~25 kDa and Futsch over 550 kDa); (2) the relative abundance of the endogenous proteins; and (3) the binding affinity or the stoichiometry of the complex. However, the GST pull-down assays further established Futsch as an interacting partner of Ringer. Having established Ringer and Futsch as a complex, it will be interesting to investigate what other known as well as yet to be identified proteins will likely be recruited to this complex. Moreover, the large size and multiple domains of Futsch alone may allow it to complex with several others in the presynaptic terminals. An issue of interest, then, will be to determine how these complexes are assembled together with the variety of interactions with the post-synaptic targets. Also interesting will be to see if these protein-protein interactions are conserved across species, particularly in vertebrates and what role they will play in regulating MT dynamics. Together these results reveal that changes in MT organization are an essential aspect of synapse development and function and Ringer, a member of the unique and highly conserved TPPP family of proteins, plays a role in regulating MT stability and synaptic organization (Shi, 2019).
Promoting axon regeneration in the central and peripheral nervous system is of clinical importance in neural injury and neurodegenerative diseases. Both pro- and anti-regeneration factors are being identified. Previous work has shown that the Rtca mediated RNA repair/splicing pathway restricts axon regeneration by inhibiting the nonconventional splicing of Xbp1 mRNA under cellular stress. However, the downstream effectors remain unknown. Through transcriptome profiling this study has shown that the tubulin polymerization-promoting protein (TPPP) ringmaker/ringer In recent years, several strategies have shown efficacy augmenting nerve regeneration in various experimental models. Unfortunately, therapeutic interventions to promote nerve regeneration and functional recovery still do not exist. Previous work has also helped shape the approach researchers have taken toward better understanding regeneration and drawing connections between successful paradigms. This study reports a link between two cellular mechanisms that are essential for regeneration: RNA processing and microtubule dynamics (Monahan Vargas, 2020).
In Drosophila, sensory dendritic arborization (da) neurons show differential regenerative potentials between the periphery and the central nervous system (CNS), resembling that of mammalian neurons. Moreover, distinct subclasses of da neurons also regenerate differently. A previous study developed a two-photon-based axon injury model that assays class III (C3da) and class IV (C4da) da neurons to identify and analyze targets that enhance regeneration. Using this model, Rtca (RNA 3'-terminal phosphate cyclase), an RNA-binding protein (RBP), was identified as an inhibitor of axon regeneration. Rtca is involved in stress induced Xbp1 mRNA splicing, and its knockout or neuronal knockdown promotes axon regeneration both in the peripheral nervous system (PNS) and CNS. However, its downstream effectors and signaling mechanisms remain unexplored. RBPs are increasingly shown to regulate complex cellular processes associated with neurodegenerative diseases and regeneration. This study reports the results from transcriptome profiling revealing that a microtubule associated protein, Ringer (also known as Ringmaker, which is the fly homolog of the mammalian tubulin polymerization-promoting proteins [TPPPs]), is strongly increased following Rtca removal (Monahan Vargas, 2020).
Microtubules and the cytoskeletal network are essential for neuronal function and are paramount to an axon's ability to respond to guidance cues, transport proteins and organelles, grow, survive, and regenerate. Microtubule-binding small molecules and microtubule-associated proteins (MAPs) that regulate microtubule dynamics are attractive therapeutic targets to augment axon regeneration. Ringer belongs to the brain-specific protein, p25α, also known as the TPPP protein family. TPPPs regulate tubulin polymerization and are implicated in neurodegenerative disorders such as α-synucleinopathies and Multiple System Atrophy. Drosophila has only one TPPP ortholog, Ringer, and it directly binds tubulin, promotes microtubule bundling and polymerization in vitro, and is critical for microtubule stabilization and developmental axon growth. This study shows that transcription of ringer is negatively regulated by Rtca via Xbp1. ringer was found to function as a neuronal intrinsic promoter of axon regeneration, working in concert with other MAPs, specifically Futsch/MAP1B and HDAC6, which have been previously shown to be integral for axonal health and integrity. The results reveal MAPs as important arbiters of axon regeneration, and ringer (TPPP homologs) is proposed as an attractive therapeutic target for promoting axon regeneration (Monahan Vargas, 2020).
RBPs have been shown to be crucial in regulating complex cellular processes such as mRNA editing, transport and local translation. Aberrant processing of RNA is present in neuronal diseases and injury. How these processes are affected after nervous system trauma and their regulation during neural repair are poorly understood. Previous work has identified Rtca, an RNA-binding protein regulating RNA repair and splicing, as a potential damage sensor that inhibits axon regeneration. Rtca LOF enhances axon regeneration in both fly and mammalian neurons. To better understand its underlying mechanism, RNA-seq was performed to assess the transcriptome of Rtca mutant neurons; ringer transcripts were found to be highly expressed. Ringer is a MAP homologous to the mammalian tubulin polymerization-promoting proteins (TPPPs), in particular TPPP3 or TPPP1, which has been shown to be a regulator of axonal microtubule organization by promoting microtubule polymerization, assembly, and stability both in vitro and in vivo. This study has revealed a connection between the injury-evoked RNA repair/splicing system and the MAP ringer; it is proposed that Rtca suppresses Xbp1 via nonconventional mRNA splicing, which in turn reduces ringer expression to inhibit axon regeneration. Furthermore, evidence is provided for an association between Futsch and HDAC6, additional MAPs capable of regulating microtubule stability and posttranslational modifications. Ringer is also inhibited by HDAC6, and it cooperates with Futsch to relay a cellular stress signal to the microtubule network. In addition, these data suggest that Rtca and Xbp1 likely have additional downstream effectors independent of ringer, and that Futsch likely receives additional inputs, in parallel to Ringer, to support axonal regeneration. Future studies to directly monitor microtubule dynamics in Rtca LOF mutants will help further validate this model and offer clues to the identity of additional players in this pathway (Monahan Vargas, 2020).
The capacity of an axon to regenerate depends on both the external environment and cell-intrinsic mechanisms, which ultimately converge onto axonal microtubules. MAPs have become popular targets for augmenting nerve regeneration given the importance of microtubule stability and polymerization in both the nascent axon and the regenerating axon's growth cone. As an axon elongates, microtubules engorge the growth cone to fill it with microtubule mass. As the growth cone advances, microtubules bundle and consolidate within the nascent axon to provide structure and support. Ringer has been shown to be essential for proper microtubule bundling. Microtubules are inherently polarized because newly added tubulin dimers only assemble and disassemble at the 'plus' end of the lattice, whereas the minus end of a microtubule is highly stabilized with special tubulin variants, abundant post translational modifications (e.g., acetylation of α-tubulin), and minus-end associating proteins. Therefore, a single microtubule can be thought of as having two general domains; a plus-end that is labile (i.e., where dynamic instability occurs) and a minus end that is stable and resists depolymerization. Microtubule stabilization prevents depolymerization and favors microtubule growth, which is beneficial for the axon's growth cone to advance. Inducing microtubule stabilization using extremely low doses of the drugs paclitaxel or epithilones has resulted in significant augmentation of nerve regeneration in vivo. The results of this study demonstrated a loss of microtubule acetylation in whole-cell lysate and specifically within the proximal axon of injured neurons in ringer mutants. This is in line with the function of Ringer, which has been associated with microtubule polymerization and stability. Future experiments to dynamically track Ringer proteins in accordance with microtubule polymerization during axon regeneration, and an extensive investigation of microtubule posttranslational modifications following axotomy are warranted (Monahan Vargas, 2020).
Futsch, a MAP1B homolog, was recently shown to associate with ringer. Together, Ringer and Futsch were found to regulate synapse formation at neuromuscular junctions via a microtubule-based mechanism. It can be inferred that Ringer and Futsch may help promote the formation of a growth cone rather than a retracting dystrophic end within injured axons, similar to its maintenance of synaptic integrity. Ringer mutation led to a decrease in futsch mRNAs and immunolabeling, suggesting a role in regulating futsch transcription, localization, and protein levels. Both ringer and futsch mutations impaired axon regeneration, albeit futsch had a more dramatic effect, suggesting that futsch may contribute to additional signaling independent of ringer. While heterozygous mutants for futsch and ringer did not have a reduction in regeneration, transheterozygotes of ringer and futsch mutations exhibited a similar reduction in regeneration as ringer mutants alone. Coimmunoprecipitation experiments showed that ringer, futsch, and tubulin physically interact and form a molecular complex, and that Ringer facilitates Futsch binding to tubulin. Epistasis analysis further demonstrated that overexpression of Futsch failed to rescue the reduced axon regeneration in ringer mutants, while overexpression of futsch is sufficient to promote axon regeneration despite the absence of futsch. Importantly, this study found that microtubule turnover is faster in injured versus uninjured axons, and that futsch LOF dysregulates microtubule dynamics, accelerating its turnover after injury. Taken together, sthe data suggest that Ringer and Futsch cooperate in the same complex with tubulin, to maintain microtubule dynamics/stability, and that both are critical to the ability of sensory neurons to regenerate. Futsch is phosphorylated by GSK3 and sustained GSK3 activity promotes axon regeneration and increases the pool of dynamic microtubule mass, which further leads to a speculation that futsch might be regulated by additional signaling pathways (Monahan Vargas, 2020).
Elucidating how microtubule stability properties are altered following an injury and the MAPs responsible for mediating those changes may identify novel therapeutic targets. This study found that acetylation properties were altered by ringer mutations and, therefore, attempts were made to explore the role HDAC6, the primary tubulin deacetylase, may play in instructing regeneration. HDAC6 knockout and pharmacological inhibition increased regeneration in C3da neurons, a subtype of sensory neurons incapable of regeneration in WT flies. Previous studies have shown that HDAC6 inhibition and deletion leads to the hyperacetylation of microtubules. Early studies found that HDAC6 was neuroprotective after a CNS injury and associated these findings with HDAC6's role in transcriptional regulation. However, more recent studies found that HDAC6 is neuroprotective in a manner that was associated with its deacetylation of microtubules. Other studies have shown that HDAC6 is essential for healthy axonal transport and influences MAP-microtubule interactions. This study showd that HDAC6 LOF leads to increased protein levels of ringer and futsch, likely through posttranscriptional mechanisms. It may also be possible that HDAC6 knockout affects microtubule-binding kinetics and the protein localization of Ringer and Futsch (i.e., concentrated versus diffuse). Augmented regeneration following HDAC6 knockout was lost with a ringer mutation. These results, along with the changes observed in Ac-Tub levels, suggest an interaction between HDAC6 and Ringer, where Ringer may function to either directly or indirectly restrict HDAC6 deacetylase activity with respect to α tubulin acetylation. This is likely, given that Ringer has been shown to regulate microtubule bundling and stability, which are associated with highly acetylated domains of microtubules. Ringer may be essential to protecting highly acetylated and stable microtubule domains from HDAC6 deacetylation by occluding its interaction with α tubulin or directly blocking deacetylase activity. This would be consistent with in vitro studies suggesting that mammalian TPPP modulates microtubule acetylation by binding to HDAC6 and inhibiting its activity. Alternatively, HDAC6 could inhibit TPPP nucleation by binding to TPPP and preventing its association to tubulin. Furthermore, HDAC6 can also physically modify kinases shown to negatively interrupt TPPP function such as ERK2. This network hypothesis could help explain an underlying positive feedback loop regulating microtubule stability: Increase of TPPP would inhibit HDAC6 leading to an enhancement of acetylated, potentially stable microtubule; in contrast, modification of kinases by HDAC6 could lead to kinase activation and downstream phosphorylation of TPPP, limiting its microtubule binding activity. It is believed that HDAC6 and ringer are involved in a pathway that ultimately affects the stability and dynamics of microtubules. Future studies will explore whether Ringer and HDAC6 expression, along with posttranslational modifications of tubulin, can predict the regenerative potential of da sensory neurons. C4da neurons show only ~75% regeneration and it is proposed that the other 25% will show differences in the expression of MAPs and microtubule posttranslational modifications, specifically acetylation of α-tubulin (Monahan Vargas, 2020).
The future treatments for nerve regeneration will most likely be combinatorial, with a need to address the extrinsic and intrinsic barriers to regeneration. This study has identified a link between RNA repair/splicing and microtubule organization via a damage-evoked mechanism involving Rtca and Ringer. Further evidence is presented that therapeutic targets capable of augmenting nerve regeneration ultimately converge on microtubules. Microtubules are a bottleneck to regeneration and identifying intrinsic signaling cascades that regulate microtubule dynamics using fly genetics will reveal pathways critical to microtubule-mediated nerve regeneration. Given the complexity of MAPs and the increasing number of candidate proteins, utilizing the fly injury model system allows screening for promising targets that warrant an investigation into their mammalian homologs with in vitro and in vivo mammalian nerve injury models. Excitingly, the zebrafish homolog of TPPP3 was recently shown to promote axon regeneration in Mauthner cells and is regulated at the transcript level by microRNA 133b. This corroborates the current findings, leading to the proposal that ringer/TPPP is tightly regulated and may function as a relay station at multiple levels. Moreover, HDAC6 was also recently shown to be inhibitory in a regeneration screen performed in C. elegans. In summary, this study has identified a RNA repair/splicing pathway that up-regulates the MAP Ringer, which interacts with other MAPs associated with microtubule stability/dynamics and tubulin posttranslational modifications, processes that are evolutionarily conserved and promising targets for regenerative therapies (Monahan Vargas, 2020).
Dendrite microtubules are polarized with minus-end-out orientation in Drosophila neurons. Nucleation sites concentrate at dendrite branch points, but how they localize is not known. Using Drosophila, this study found that canonical Wnt signaling proteins regulate localization of the core nucleation protein gammaTubulin (gammaTub). Reduction of frizzleds (fz), arrow (low-density lipoprotein receptor-related protein [LRP] 5/6), dishevelled (dsh), casein kinase Igamma, G proteins, and Axin reduced gammaTub-green fluorescent protein (GFP) at branch points, and two functional readouts of dendritic nucleation confirmed a role for Wnt signaling proteins. Both dsh and Axin localized to branch points, with dsh upstream of Axin. Moreover, tethering Axin to mitochondria was sufficient to recruit ectopic gammaTub-GFP and increase microtubule dynamics in dendrites. At dendrite branch points, Axin and dsh colocalized with early endosomal marker Rab5, and new microtubule growth initiated at puncta marked with Fz, Dsh, Axin, and Rab5. It is proposed that in dendrites, canonical Wnt signaling proteins are housed on early endosomes and recruit nucleation sites to branch points (Weiner, 2020).
Branching MT nucleation is dependent upon Augmin and γ-TuRC and generates the bulk of MTs required for both meiotic and mitotic spindle formation and has been visualised in vivo in Drosophila, Xenopus, plants, and humans. However, understanding, and in vitro reconstitution of, this phenomenon has been hampered by methodological constraints relating to purification of functional protein complexes); Augmin is composed of 8 subunits, while the γ-TuRC is a ~ 2 MD protein complex containing multiple copies of at least six proteins, including 14 molecules of γ-Tubulin. In vitro studies generally use proteins that have been individually- or co-expressed and purified in heterologous systems, where folding and post translational modifications crucial to function may not occur. Although purification of protein complexes from autogenous cells can be achieved using affinity-based methods, non-specific binding of contaminating proteins and difficulties in releasing purified proteins from affinity matrices are major problems (Tariq, 2020).
This study therefore developed an approach to allow the isolation of intact, functional Augmin and γ-TuRC, to test the hypothesis that these two complexes are necessary and sufficient for branched MT nucleation. The approach is based on biotinylated, amine-reactive thiol- or photo-cleavable linkers, Sulfo-NHS-SS-Biotin and PC-Biotin-NHS (see Isolation of functional γ-TuRC and Augmin using cleavable affinity purification). Stepwise incubation of the ~12 kD camelid anti-GFP nanobody, GFP-binding protein, with either of these linkers resulted in covalent linkage, while subsequent incubation with a Streptavidin Agarose matrix led to stable tri-partite reagents - GFP-TRAP-Sulfo beads and GFP-TRAP-PC beads; where GFP-binding protein is immobilised, but cleavable through the addition of DTT or exposure to UV light, respectively (Tariq, 2020).
This work confirms a long-standing hypothesis, first articulated to explain the loss of γ-Tubulin on the mitotic spindle when the expression of Augmin subunits is reduced. It demonstrates conclusively that Augmin directly recruits γ-TuRC to MTs and that these two protein complexes are sufficient for robust and efficient branching MT nucleation. Although calculations suggest that 65% of the mitotic γ-TuRCs are competent to bind MT minus ends and Augmin, and that they are able to nucleate at a rate of >1 MT/min, it is expected that, in vivo, other proteins further enhance these γ-TuRC activities. For example, a clear role has been reported for the MT associated protein, TPX2, in stimulating Augmin-dependent branched MT nucleation in Xenopus meiotic extracts. However, in support of these conclusions, Drosophila TPX2 has recently been shown to be dispensible for the phenomenon in vivo. These experiments also highlight the intriguing possibility that, in some cells, Augmin might recruit pre-existing γ-TuRC-containing MTs, nucleated elsewhere in the cell, anchoring them to specific sites and increasing local MT density (Tariq, 2020).
The generation of stable MT-Augmin-γ-TuRC-MT junctions using the methodologies pioneered in this study also provide a route to finally defining the molecular detail of MT branching at the ultrastructural level. More broadly, cleavable affinity purification provides the basis to generate more complex, but molecularly defined, mixes of purified proteins, complete with in vivo post-translational modifications, in order to reconstitute higher-order aspects of spindle formation. Indeed, by isolating and combining purified, active proteins and protein complexes from any biological system of interest, 'cl-AP TRAP' overcomes the limitations of traditional 'bottom-up' approaches, allowing exploration of the level of biological organisation between individual protein and biological process - the level at which emergence of cellular phenomena often occurs (Tariq, 2020).
Muscle myofibrils and sarcomeres present exceptional examples of highly ordered cytoskeletal filament arrays, whose distinct spatial organization is an essential aspect of muscle cell functionality. This study utilized ultra-structural analysis to investigate the assembly of myofibrils and sarcomeres within developing myotubes of the indirect flight musculature of Drosophila. A temporal sequence composed of three major processes was identified: subdivision of the unorganized cytoplasm of nascent, multi-nucleated myotubes into distinct organelle-rich and filament-rich domains; initial organization of the filament-rich domains into myofibrils harboring nascent sarcomeric units; and finally, maturation of the highly-ordered pattern of sarcomeric thick (myosin-based) and thin (microfilament-based) filament arrays in parallel to myofibril radial growth. Significantly, organized microtubule arrays were present throughout these stages and exhibited dynamic changes in their spatial patterns consistent with instructive roles. Genetic manipulations confirm these notions, and imply specific and critical guidance activities of the microtubule-based cytoskeleton, as well as structural interdependence between the myosin- and actin-based filament arrays. These observations highlight a surprisingly significant, behind-the-scenes role for microtubules in establishment of myofibril and sarcomere spatial patterns and size, and provide a detailed account of the interplay between major cytoskeletal elements in generating these essential contractile myogenic units (Dhanyasi, 2020).
Mitotic divisions depend on the timely assembly and proper orientation of the mitotic spindle. Malfunctioning of these processes can considerably delay mitosis, thereby compromising tissue growth and homeostasis, and leading to chromosomal instability. Loss of functional Mms19 drastically affects the growth and development of mitotic tissues in Drosophila larvae, and this study demonstrates that Mms19 is an important factor that promotes spindle and astral microtubule (MT) growth, and MT stability and bundling. Mms19 function is needed for the coordination of mitotic events and for the rapid progression through mitosis that is characteristic of neural stem cells. Surprisingly, Mms19 performs its mitotic activities through two different pathways. By stimulating the mitotic kinase cascade, it triggers the localization of the MT regulatory complex TACC/Msps (Transforming Acidic Coiled Coil/Minispindles, the homolog of human ch-TOG) to the centrosome. This activity of Mms19 can be rescued by stimulating the mitotic kinase cascade. However, other aspects of the Mms19 phenotypes cannot be rescued in this way, pointing to an additional mechanism of Mms19 action. This study provides evidence that Mms19 binds directly to MTs and that this stimulates MT stability and bundling (Chippalkatti, 2020).
Proper assembly of mitotic spindles requires microtubule nucleation not only at the centrosomes but also around chromatin. This study found that the Drosophila tubulin-specific chaperone dTBCE is required for the enrichment of tubulin in the nuclear space after nuclear envelope breakdown and for subsequent promotion of spindle microtubule nucleation. These events depend on the CAP-Gly motif found in dTBCE and are regulated by Ran and lamin proteins. These data suggest that during early mitosis, dTBCE and nuclear pore proteins become enriched in the nucleus, where they interact with the Ran GTPase to promote dynamic tubulin enrichment. It is proposed that this novel mechanism enhances microtubule nucleation around chromatin, thereby facilitating mitotic spindle assembly (Metivier, 2020).
Membrane organelle function, localization, and proper partitioning upon cell division depend on interactions with the cytoskeleton. Whether membrane organelles also impact the function of cytoskeletal elements remains less clear. This study shows that acute disruption of the ER around spindle poles affects mitotic spindle size and function in Drosophila syncytial embryos. Acute ER disruption was achieved through the inhibition of ER membrane fusion by the dominant-negative cytoplasmic domain of atlastin. When centrosome-proximal ER membranes are disrupted, specifically at metaphase, mitotic spindles become smaller, despite no significant changes in microtubule dynamics. These smaller spindles are still able to mediate sister chromatid separation, yet with decreased velocity. Furthermore, by inducing mitotic exit, this study found that nuclear separation and distribution are affected by ER disruption. These results suggest that ER integrity around spindle poles is crucial for the maintenance of mitotic spindle shape and pulling forces. In addition, ER integrity also ensures nuclear spacing during syncytial divisions (Araujo, 2023).
Asymmetric cell division gives rise to two daughter cells that inherit different determinants, thereby acquiring different fates. Polarized trafficking of endosomes containing fate determinants recently emerged as an evolutionarily conserved feature of asymmetric cell division to enhance the robustness of asymmetric cell fate determination in flies, fish and mammals. In particular, polarized sorting of signalling endosomes by an asymmetric central spindle contributes to asymmetric cell division in Drosophila melanogaster. However, how central spindle asymmetry arises remains elusive. This study identified a moonlighting function of the Elongator complex-an established protein acetylase and tRNA methylase involved in the fidelity of protein translation-as a key factor for central spindle asymmetry. Elongator controls spindle asymmetry by stabilizing microtubules differentially on the anterior side of the central spindle. Accordingly, lowering the activity of Elongator on the anterior side using nanobodies mistargets endosomes to the wrong cell. Molecularly, Elongator regulates microtubule dynamics independently of its acetylation and methylation enzymatic activities. Instead, Elongator directly binds to microtubules and increases their polymerization speed while decreasing their catastrophe frequency. These data establish a non-canonical role of Elongator at the core of cytoskeleton polarity and asymmetric signalling (Planelles-Herrero, 2022).
Sperm elongation and nuclear shaping in Drosophila largely depends on the microtubule cytoskeleton that in early spermatids has centrosomal and non-centrosomal origins. This study reports an additional γ-tubulin focus localized on the anterior pole of the nucleus in correspondence of the apical end of the perinuclear microtubules that run within the dense complex. The perinuclear microtubules are nucleated by the pericentriolar material, or centriole adjunct, that surrounds the basal body and are retained to play a major role in nuclear shaping. However, it was found that both the perinuclear microtubules and the dense complex are present in spermatids lacking centrioles. Therefore, the basal body or the centriole adjunct seem to be dispensable for the organization and assembly of these structures. These observations shed light on a novel localization of γ-tubulin and open a new scenario on the distribution of the microtubules and the organization of the dense complex during early Drosophila spermiogenesis (Riparbelli, 2020).
Microtubule nucleation in eukaryotes is primarily promoted by γ-tubulin and the evolutionary conserved protein complex, γ-Tubulin Ring Complex (γ-TuRC). γ-TuRC is part of the centrosome and basal body, which are the best-known microtubule-organizing centers. Centrosomes undergo intensive and dynamic changes during spermatogenesis, as they turn into basal bodies, a prerequisite for axoneme formation during spermatogenesis. This study describes the existence of a novel, tissue-specific γ-TuRC in Drosophila. Three genes (CG7716 (t-Grip84), CG18109 (t-Grip91), and CG32232 (t-Grip128) were characterized encoding testis-specific components of γ-TuRC (t-γ-TuRC) testis-specific Grip paralogs. Presence of t-γ-TuRC is essential to male fertility. The diverse subcellular distribution was shown of the t-γ-TuRC proteins during post-meiotic development, at first at the centriole adjunct and then also on the anterior tip of the nucleus, and finally, they appear in the tail region, close to the mitochondria. The physical interactions between the t-γ-TuRC members, γ-tubulin and Mozart1 were also proven. These results further indicate heterogeneity in γ-TuRC composition during spermatogenesis and suggest that the different post-meiotic microtubule organizing centers are orchestrated by testis-specific gene products, including t-γ-TuRC (Alzyoud, 2021).
This study investigated planar cell polarity (PCP) in the Drosophila larval epidermis. The intricate pattern of denticles depends on only one system of PCP, the Dachsous/Fat system. Dachsous molecules in one cell bind to Fat molecules in a neighbour cell to make intercellular bridges. The disposition and orientation of these Dachsous-Fat bridges allows each cell to compare two neighbours and point its denticles towards the neighbour with the most Dachsous. Measurements of the amount of Dachsous reveal a peak at the back of the anterior compartment of each segment. Localization of Dachs and orientation of ectopic denticles help reveal the polarity of every cell. Whether these findings support the gradient model of Dachsous activity is discussed. Several groups have proposed that Dachsous and Fat fix the direction of PCP via oriented microtubules that transport PCP proteins to one side of the cell. This proposition was tested in the larval cells; most microtubules grow perpendicularly to the axis of PCP. No meaningful bias was found in the polarity of microtubules aligned close to that axis. Published data from the pupal abdomen was reexamined, and no evidence was found supporting the hypothesis that microtubular orientation draws the arrow of PCP (Pietra, 2020).
Many neurons in bilaterian animals are polarized with functionally distinct axons and dendrites. Microtubule polarity, microtubule stability, and the axon initial segment (AIS) have all been shown to influence polarized transport in neurons. Each of these cytoskeletal cues could act independently to control axon and dendrite identity, or there could be a hierarchy in which one acts upstream of the others. This study tested the hypothesis that microtubule polarity acts as a master regulator of neuronal polarity by using a Drosophila genetic background in which some dendrites have normal minus-end-out microtubule polarity and others have the axonal plus-end-out polarity. In these mosaic dendrite arbors, this study found that ribosomes, which are more abundant in dendrites than axons, were reduced in plus-end-out dendrites, while an axonal cargo was increased. In addition, it was determined that microtubule stability was different in plus-end-out and minus-end-out dendrites, with plus-end-out ones having more stable microtubules like axons. Similarly, it was found that ectopic diffusion barriers, like those at the AIS, formed at the base of dendrites with plus-end-out regions. Thus, changes in microtubule polarity were sufficient to rearrange other cytoskeletal features associated with neuronal polarization. However, overall neuron shape was maintained with only subtle changes in branching in mosaic arbors. It is concluded that microtubule polarity can act upstream of many aspects of intracellular neuronal polarization, but shape is relatively resilient to changes in microtubule polarity in vivo (Thyagarajan, 2022).
Robustness of biological systems is crucial for their survival, however, for many systems its origin is an open question. This study analyzed one subcellular level system, the microtubule cytoskeleton. Microtubules self-organize into a network, along which cellular components are delivered to their biologically relevant locations. While the dynamics of individual microtubules is sensitive to the organism's environment and genetics, a similar sensitivity of the overall network would result in pathologies. Large-scale stochastic simulations show that the self-organization of microtubule networks is robust in a wide parameter range in individual cells. This robustness was confirmed in vivo on the tissue-scale using genetic manipulations of Drosophila epithelial cells. Finally, a minimal mathematical model shows that the origin of robustness is the separation of time-scales in microtubule dynamics rates. Altogether, this study demonstrates that the tissue-scale self-organization of a microtubule network depends only on cell geometry and the distribution of the microtubule minus-ends (Plochocka, 2021).
Regulation of microtubule stability is crucial for the maintenance of cell structure and function. This study identified an N-terminal acetyltransferase, Mnat9, that regulates cell signaling and microtubule stability in Drosophila. Loss of Mnat9 causes severe developmental defects in multiple tissues. In the wing imaginal disc, Mnat9 RNAi leads to the ectopic activation of c-Jun N-terminal kinase (JNK) signaling and apoptotic cell death. These defects are suppressed by reducing the level of JNK signaling. Overexpression of Mnat9 can also inhibit JNK signaling. Mnat9 colocalizes with mitotic spindles, and its loss results in various spindle defects during mitosis in the syncytial embryo. Furthermore, overexpression of Mnat9 enhances microtubule stability. Mnat9 is physically associated with microtubules and shows a catalytic activity in acetylating N-terminal peptides of α- and β-tubulin in vitro. Cell death and tissue loss in Mnat9-depleted wing discs are restored by reducing the severing protein Spastin, suggesting that Mnat9 protects microtubules from its severing activity. Remarkably, Mnat9 mutated in the acetyl-CoA binding site is as functional as its wild-type form. This study also found that human NAT9 can rescue Mnat9 RNAi phenotypes in flies, indicating their functional conservation. Taken together, it is proposed that Mnat9 is required for microtubule stability and regulation of JNK signaling to promote cell survival in developing Drosophila organs (Mok, 2021).
Repair after traumatic injury often starts with mitotic activation around the lesion edges. Early midline cells in the Drosophila embryonic CNS can enter into division following the traumatic disruption of microtubules. Microtubule disruption activates non-canonical TNF signaling by phosphorylation of TGF-β activated kinase 1 (Tak1) and its target IkappaB kinase (Ik2), culminating in Dorsal/NfkappaB nuclear translocation and Jra/Jun expression. Tak1 and Ik2 are necessary for the damaged-induced divisions. Microtubule disruption caused by Tau accumulation is also reported in Alzheimer's disease (AD). Human Tau expression in Drosophila midline cells is sufficient to induce Tak1 phosphorylation, Dorsal and Jra/Jun expression, and entry into mitosis. Interestingly, activation of Tak1 and Tank binding kinase 1 (Tbk1), the human Ik2 ortholog, and NfkappaB upregulation are observed in AD brains (Barros, 2021).
Non-centrosomal microtubule arrays serve crucial functions in cells, yet the mechanisms of their generation are poorly understood. During budding of the epithelial tubes of the salivary glands in the Drosophila embryo, it has been demonstrated that the activity of pulsatile apical-medial actomyosin depends on a longitudinal non-centrosomal microtubule array. This study uncovered that the exit from the last embryonic division cycle of the epidermal cells of the salivary gland placode leads to one centrosome in the cells losing all microtubule-nucleation capacity. This restriction of nucleation activity to the second, Centrobin-enriched, centrosome is key for proper morphogenesis. Furthermore, the microtubule-severing protein Katanin and the minus-end-binding protein Patronin accumulate in an apical-medial position only in placodal cells. Loss of either in the placode prevents formation of the longitudinal microtubule array and leads to loss of apical-medial actomyosin and impaired apical constriction. A mechanism is proposed whereby Katanin-severing at the single active centrosome releases microtubule minus-ends that are then anchored by apical-medial Patronin to promote formation of the longitudinal microtubule array crucial for apical constriction and tube formation (Gillard, 2021).
Dendrite
shape impacts functional connectivity and is mediated by organization
and dynamics of cytoskeletal fibers. Identifying the molecular factors
that regulate dendritic cytoskeletal architecture is therefore important
in understanding the mechanistic links between cytoskeletal
organization and neuronal function. This study identified Formin 3 (Form3) as an essential regulator of cytoskeletal architecture in nociceptive sensory neurons
in Drosophila larvae. Time course analyses reveal that Form3 is
cell-autonomously required to promote dendritic arbor complexity. form3 is required for the maintenance of a population of stable dendritic microtubules (MTs),
and mutants exhibit defects in the localization of dendritic
mitochondria, satellite Golgi, and the TRPA channel Painless. Form3
directly interacts with MTs via FH1-FH2 domains. Mutations in human
inverted formin 2 (INF2; ortholog of form3) have been causally
linked to Charcot-Marie-Tooth (CMT) disease. CMT sensory neuropathies
lead to impaired peripheral sensitivity. Defects in form3
function in nociceptive neurons result in severe impairment of noxious
heat-evoked behaviors. Expression of the INF2 FH1-FH2 domains partially
recovers form3 defects in MTs and nocifensive behavior,
suggesting conserved functions, thereby providing putative mechanistic
insights into potential etiologies of CMT sensory neuropathies (Das, 2021).
The ability of stem cells to switch between quiescence and proliferation
is crucial for tissue homeostasis and regeneration. Drosophila
quiescent neural stem cells (NSCs) extend a primary cellular protrusion
from the cell body prior to their reactivation. However, the structure
and function of this protrusion are not well established. This study
shows that in the protrusion of quiescent NSCs, microtubules are
predominantly acentrosomal and oriented plus-end-out toward the tip of
the primary protrusion. This study has identified Mini Spindles (Msps)/XMAP215
as a key microtubule regulator in quiescent NSCs that governs NSC
reactivation via regulating acentrosomal microtubule growth and
orientation. Quiescent NSCs form membrane contact with the neuropil and
E-cadherin, a cell adhesion molecule, localizes to these NSC-neuropil
junctions. Msps and a plus-end directed motor protein Kinesin-2 promote NSC cell cycle re-entry and target E-cadherin
to NSC-neuropil contact during NSC reactivation. Together, this work
establishes acentrosomal microtubule organization in the primary
protrusion of quiescent NSCs and the Msps-Kinesin-2 pathway that governs
NSC reactivation, in part, by targeting E-cad to NSC-neuropil contact
sites (Deng, 2021).
Axons and dendrites are distinguished by microtubule polarity. In Drosophila, dendrites are dominated by minus-end-out microtubules while axons contain plus-end-out microtubules. Local nucleation in dendrites generates microtubules in both orientations. To understand why dendritic nucleation does not disrupt polarity, this study used live imaging to analyze the fate of microtubules generated at branch points. It was found that they had different rates of success exiting the branch based on orientation: correctly oriented minus-end-out microtubules succeeded in leaving about twice as often as incorrectly oriented microtubules. Increased success relied on other microtubules in a parallel orientation. From a candidate screen, Trim9 and kinesin-5 (Klp61F) were identified as machinery that promoted growth of new microtubules. In S2 cells, EB1 recruited Trim9 to microtubules. Klp61F promoted microtubule growth in vitro and in vivo, and could recruit Trim9 in S2 cells. In summary, the data argue that Trim9 and kinesin-5 act together at microtubule plus ends to help polymerizing microtubules parallel to pre-existing ones resist catastrophe (Feng, 2021).
The almost uniform minus-end-out microtubule polarity in Drosophila dendrites has proven a useful system for the identification of mechanisms that control microtubule organization and polarity. One surprise is that multiple mechanisms operate in parallel even in this very confined space with simple microtubule layout. Two basic types of polarity control mechanisms have been identified: those that can establish microtubule polarity independently of existing microtubules, and those that act as positive feedback loops to reinforce the predominant polarity (Feng, 2021).
Two mechanisms have been identified that can autonomously contribute to dendritic minus-end-out microtubule polarity: local nucleation and minus-end growth. Very early in the development of dendritic arborization neurons in the Drosophila embryo, new dendrites are populated by plus-end-out microtubules that grow in from the cell body (Feng, 2019). The next step is for slower-growing minus ends to enter dendrites from the cell body adding minus-end-out microtubules. Dendrites then remain with mixed polarity for the rest of embryogenesis, and eventually this resolves to minus-end-out in larvae. Minus-end-out microtubules can also be generated locally in dendrites by nucleation. Like microtubule growth from the cell body, nucleation can contribute plus-end-out and minus-end-out microtubules. In mature dendritic arborization neurons, nucleation at branch points is biased towards generating minus-end-out microtubules. Several mechanisms to bias nucleation have been shown to operate in neurons, but they have not been shown to act in mature dendritic arborization neurons. In developing C. elegans sensory dendrites, nucleation sites cluster close to the tip, resulting in a short region of plus-end-out microtubules beyond the cluster and the proximal dendrite dominated by minus-end-out microtubules (Liang, 2020). However, in ddaE dendrites, nucleation sites are found not just at the dendrite tip but throughout the arbor at branch points. In ddaE dendrites, nucleation has been proposed to be biased by recruitment to only one side of Golgi outposts. However, nucleation sites have recently been shown to be recruited to endosomes rather than Golgi in ddaE dendrites (Weiner, 2020), making the earlier findings difficult to interpret. In summary, both growth from microtubule ends and nucleation of new microtubules create minus-end-out, as well as plus-end-out, microtubules in dendrites (Feng, 2021).
In addition to the de novo mechanisms that add minus-end-out microtubules to dendrites described above, feedback mechanisms reinforce the dominant polarity. One of these controls the direction of microtubule growth at branch points. As microtubules grow from distal regions of the dendrite into branch points, they encounter a choice to grow towards the cell body or away from the cell body. Kinesin-2, together with Eb1, Apc and Apc2, allows the growing plus end to track existing microtubules, reinforcing polarity. If this mechanism is eliminated, polarity in dendrites remains mixed. The final percentage of minus-end-out microtubules seems to depend on the angle of branches such that in ddaE dendrites, in which branch angles are close to 90° and microtubules can easily turn either direction at branch points, ~50% of microtubules are minus-end-out. In dendrites with more acute branch angles that help direct growing plus ends towards the cell body, 70% of microtubules remain minus-end-out, even when steering is eliminated. The mechanism described in this study, that selectively promotes growth of new microtubules out of branch points along parallel microtubules, seems to act as an additional feedback loop that helps align newly nucleated microtubules with pre-existing ones. Impairing this mechanism seems to have little impact on overall polarity in dendrites, unlike eliminating steering. Quality control of newly nucleated microtubules may have a small effect on overall polarity because nucleation itself is somewhat biased and nucleation may contribute at relatively low levels to the overall microtubule population at steady state in mature neurons. Quality control of new microtubules may exist as a backup mechanism to help maintain microtubule organization under stressful conditions like axon injury, which can upregulate nucleation (Feng, 2021).
The data suggest that Trim9 and Klp61F are both needed for new microtubules to grow in parallel bundles with pre-existing microtubules. The fact that reduction of either results in a phenotype suggests that they act together, rather than in parallel, to control plus-end behavior. A vertebrate Trim9 family member, Trim46, organizes parallel microtubules at the axon initial segment and has autonomous parallel bundling activity in vitro. A role for the single Drosophila Trim9 family member in parallel orientation of microtubules suggests that this may be an ancestral function of this family, rather than a new function that evolved with the expansion of the family in vertebrates. If this is the case, then the other vertebrate family members in the C-1 subfamily containing a microtubule-binding COS box may all engage parallel microtubules, perhaps in different cell types or subcellular regions. In vitro, Trim46 has a strong preference for interacting with parallel bundles of microtubules over individual microtubules, and so tends to accumulate some distance behind dynamic plus ends. When a depolymerizing plus end encounters a bundled region decorated with Trim46, catastrophe is strongly inhibited. Although the ability of Trim46 to rescue catastrophes would be expected to promote growth like Drosophila Trim9, the site of action seems to be somewhat different. Trim46 acts at bundled regions of microtubules behind the dynamic plus end, and Drosophila Trim9 prevents catastrophes from happening, likely at the plus end, with Eb1 and Klp61F (Feng, 2021).
The in vivo role of Klp61F in promoting continued growth of plus ends out of the branch point is consistent with previous in vitro studies showing that Eg5 dimers can accumulate at the plus end and promote polymerization. Klp61F tetramers form rods ~95 nm in length that crosslink microtubules with spacing of at least 60 nm. The spacing of microtubules bundled with Trim46 is ~37 nm. Based on these general size considerations, as well as in vitro activities of Klp61F and Trim46, the following model is proposed. Klp61F could be constantly traveling along stable bundles of microtubules. If a new microtubule approaches within 60 nm, then it could be captured. As Klp61F interacts with the growing plus end it could promote polymerization. At the same time, Trim9 could be recruited to the growing plus end by Eb1, perhaps through [S/T]-x-[I/L]-P motifs, which interact with Eb1. Both Drosophila Trim9 proteins contain two potential motifs at amino acids: the first is at 42-SALP-46 in both, and the second is 479-TILP-483 in the RB isoform and 468-TILP-472 in the RA isoform. After Klp61F grabs the growing microtubule with Eb1 and Trim9 at its tip, Trim9 could reinforce the parallel interaction. Although this model provides an initial framework for the function of Klp61F and Trim9 in dendrites, it is quite speculative and raises many additional questions. For example, if Klp61F is in constant flux towards the plus end of microtubules, how is it transported into dendrites? And do these proteins really act sequentially? If so, perhaps family members collaborate in a similar way in other cellular scenarios; for example, at the axon initial segment (Feng, 2021).
It has long been thought that microtubule disassembly, one of the earliest cellular events, contributes to neuronal pruning and neurodegeneration in development and disease. However, how microtubule disassembly drives neuronal pruning remains poorly understood. This study conducted a systematic investigation of various microtubule-destabilizing factors and identified exchange factor for Arf6 (Efa6) and Stathmin (Stai) as new regulators of dendrite pruning in ddaC sensory neurons during Drosophila metamorphosis. Efa6 is both necessary and sufficient to regulate dendrite pruning. Interestingly, Efa6 and Stai facilitate microtubule turnover and disassembly prior to dendrite pruning without compromising the minus-end-out microtubule orientation in dendrites. Moreover, pharmacological and genetic manipulations strongly support a key role of microtubule disassembly in promoting dendrite pruning. Thus, this systematic study highlights the importance of two selective microtubule destabilizers in dendrite pruning and substantiates a causal link between microtubule disassembly and neuronal pruning (Bu, 2021).
Microtubule disassembly, one of the earliest cellular alterations in pruning dendrites or axons, has been thought to be a prerequisite for the execution of neuronal pruning. This systematic study reveals that dendrite pruning selectively requires two microtubule destabilizers, Efa6 and Stai. Moreover, pharmacological and genetic manipulations strongly support a causative role of microtubule disassembly in promoting dendrite pruning of ddaC sensory neurons (Bu, 2021).
The Efa6 protein family is conserved from yeast to mammals and was originally identified in mammals to regulate endosomal membrane recycling and actin cytoskeletal rearrangement via its C-terminal Sec7 GEF domain. In the mammalian nervous systems, Efa6, via the GEF domain, regulates dendritic spine formation and axon regeneration in an Arf6-dependent manner. However, in worms and flies, Efa6 orthologs act independently of Arf6 to inhibit microtubule polymerization via their respective N-terminal MTED domains. The MTED domain is conserved in worms and flies, but not in mammals. Through their MTEDs, Efa6s function as negative regulators of developmental axon growth/branching and axonal regeneration after injury. This study reports an important role of Efa6 in promoting dendrite pruning of sensory neurons via its MTED domain. Multiple genetic manipulations with loss or gain of Efa6 function demonstrate that Efa6 is both necessary and sufficient to promote dendrite pruning. Moreover, the structure-function analysis reveals that the MTED domain is essential for Efa6 to promote dendrite pruning, whereas the C-terminal Sec7 GEF domain is dispensable. Overexpression of the MTED-deleted Efa6 variant was unable to induce precocious dendrite pruning phenotype. Unlike other microtubule regulators, such as Patronin and Msps/TACC, Efa6 does not modulate the minus-end-out microtubule orientation in ddaC dendrites, but rather inhibits microtubule growth and promotes microtubule disassembly at the proximal regions of the dendrites (Bu, 2021).
How does Efa6 disassemble microtubules in the dendrites during pruning? This study found that loss of Efa6 function resulted in increased number of EB1-GFP-labelled plus ends of microtubules and reduced microtubule depolymerization/turnover at the proximal dendrites, whereas gain of Efa6 function led to enhanced microtubule depolymerization and disassembly. The model that Efa6 may directly disassemble microtubules in vivo at the proximal dendrites is preferred, leading to initial severing of dendrites. In line with this model, fly Efa6 directly interacts with tubulins and inhibits microtubule polymerization per se. In the case of axon injury and regeneration in nematodes, Efa6 was reported to indirectly inhibit microtubule dynamics by suppressing its binding partners, TAC-1 (TACC in Drosophila) and ZYG-8 (Doublecortin-like kinase, CG17528 in Drosophila;). However, fly TACC and Efa6 may function independently during dendrite pruning. A previous study reported that TACC regulates the minus-end-out microtubule orientation to promote dendrite pruning in ddaC dendrites. By contrast, Efa6 does not affect dendritic microtubule orientation in these neurons. Moreover, mutants of the fly zyg8/CG17528 were generated that did not show any dendrite pruning defect, and double mutant of efa6 and zyg8/CG17528 displayed no genetic interaction. Efa6 has a cortical localization pattern in worm neurons/embryos. Interestingly, the cytoplasmic distribution of fly Efa6 in ddaC neurons was found despite its membrane association in epidermal cells. This finding raises the possibility that Efa6 might be released from the plasma membrane to the cytoplasm where it might more potently eliminate microtubules during dendrite pruning. In support of this notion, worm Efa6 can relocalize from the plasma membrane to microtubule minus ends and inhibit microtubule dynamics in axons upon injury (Bu, 2021).
This study has systematically interrogated various microtubule-destabilizing factors for their possible roles in dendrite pruning using both RNAi and clonal approaches. Dendrite pruning was demonstrated to require two microtubule destabilizers, Efa6 and Stai. Stai and its mammalian homologues negatively regulate microtubule dynamics and disassemble microtubules via a mechanism distinct from Efa6. Stai can either sequester α/β-tubulin dimers and prevent their incorporation into growing microtubules or directly interact with microtubules to promote their disassembly. Stai has been involved in diverse models of neurodegeneration, neuronal migration axonal transport, neuronal polarization and regeneration as well as plasticity. Drosophila Stai regulates axonal microtubule integrity, synapse stability and neuronal functions. This study identified an additive function of Stai and Efa6 in dendrite pruning of ddaC neurons. Like Efa6, Stai promotes microtubule turnover/disassembly but does not regulate microtubule orientation in the dendrites of ddaC sensory neurons. Loss of Stai function resulted in an increase in polymerized microtubules; conversely, overexpression of Stai caused strong reductions in microtubule growth and mass. Thus, Stai is essential for inhibiting microtubule polymerization and promoting the disassembly during dendrite pruning. However, in contrast to this finding, a previous study has also reported that loss of Stai function results in decreased microtubule levels in the axons of Drosophila CNS neurons. It is possible that differential roles of Stai in PNS and CNS neurons might be due to its different phosphorylation. In Xenopus, the microtubule-destabilizing activity of Stai is dependent on its phosphorylation state, which is regulated by the serine/threonine type-2A phosphatase (Bu, 2021).
The microtubule-severing enzymes (katanin, fidgetin and spastin), which hydrolyse ATP to sever microtubules into small pieces in vitro, are apparent candidates that disassemble dendritic microtubules to promote neuronal pruning. These enzymes were also reported to sever microtubules and regulate the neuromuscular junction development or dendrite arborization in Drosophila and mammals . Moreover, spastin was reported to promote axon pruning at the neuromuscular junctions in postnatal mice by locally destabilizing microtubules. However, RNAi knockdown of these severing enzymes did not cause any prominent dendrite pruning defects in Drosophila ddaC neurons in the previous studies. To exclude the possibility that RNAi knockdown was inefficient for these genes, several mutant alleles were generated for these genes, and their clonal analysis was conducted in this study. The results further substantiate that these individual enzymes are indeed dispensable for dendrite pruning. Kat-60L1, an AAA ATPase analogous to Kat-60, is involved in ddaC dendrite pruning. However, Kat-60L1 had no apparent microtubule-disassembly function in ddaC neurons. Unlike Kat-60, overexpression of the Kat-60L1 short isoform neither impaired microtubule turnover nor affected the microtubule levels in the dendrites of ddaC neuron. The effect of the long isoform of Kat-60L1 in ddaC neurons was also analyzed, as the long isoform, but not the short isoform, was reported to possess microtubule-disassembly function in adult mechanosensory neurons. Unexpectedly, similar to the short isoform, reduced Futsch-positive microtubules were not observed in the dendrites when the long isoform of Kat-60L1 was overexpressed in ddaC neurons. Moreover, no significant alteration in the levels of polymerized microtubules was observed in dendrites and soma of kat-60L1F1 mutant ddaC neurons. Thus, none of the data suggest that Kat-60L1 acts as a microtubule-severing enzyme in ddaC neurons, which contrasts with a recent finding showing that purified Kat-60L1 protein possesses the microtubule-severing activity in vitro. However, the data cannot rule out the possibility that Kat-60L1 promotes dendrite pruning via severing dendritic microtubules. Further studies will be necessary to determine its potential microtubule-severing function during ddaC dendrite pruning (Bu, 2021).
Microtubule disassembly precedes membrane scission during neuronal pruning. This study has solated two microtubule-destabilizing factors, Efa6 and Stai, whose mutants exhibited excessive microtubule polymerization without affecting microtubule orientation in dendrites. These findings further provided the opportunity to investigate a relationship between microtubule disassembly and dendrite pruning. First, efa6 RNAi larvae were treated with colchicine, a microtubule-destabilizing drug. A low concentration of colchicine not only reduced microtubule polymerization but also fully rescued the dendrite pruning defects in efa6 loss-of-function neurons. Second, the treatment with the microtubule-stabilizing drug taxol caused excessive microtubule polymerization in dendrites and inhibited dendrite pruning in wild-type neurons. Third, taxol treatment significantly enhanced dendrite pruning defects in the efa6 RNAi background. Finally, reduced γ-tubulin in efa6 mutant neurons, which likely downregulates microtubule nucleation and in turn polymerization, almost fully rescued the pruning defects. Taken together, multiple lines of genetic and pharmacological evidence support that microtubule disassembly drives dendrite pruning in ddaC sensory neurons (Bu, 2021).
In summary, this systematic study highlights important roles of two negative regulators of microtubule polymerization, Efa6 and Stai, in facilitating dendrite pruning via microtubule disassembly. Moreover, this study supports a causal relationship between microtubule disassembly and dendrite pruning during early metamorphosis (Bu, 2021).
Dendritic morphology underlies the source and processing of neuronal signal inputs. Previous work demonstrated that microtubules and actin filaments are associated with arbor elongation and branching, fully constraining dendrogram topology. This study relate the local distribution of these two primary cytoskeletal components with dendritic spatial embedding. First, 167 sensory neurons from the Drosophila larva encompassing multiple cell classes and genotypes were reconstructed and analyzed. It was observed that branches with higher microtubule concentration are overall straighter and tend to deviate less from the direction of their parent branch. F-actin displays a similar effect on the angular deviation from the parent branch direction, but its influence on branch tortuosity varies by class and genotype. A computational model was created of dendritic morphology purely constrained by the cytoskeletal composition imaged from real neurons. The model quantitatively captures both spatial embedding and dendrogram topology across all tested neuron groups. These results suggest a common developmental mechanism regulating diverse morphologies, where the local cytoskeletal distribution can fully specify the overall emergent geometry of dendritic arbors (Nanda, 2023).
Regulation of microtubule stability is crucial for diverse biological processes, including cell division, morphogenesis, and signaling. Various in vitro assays for microtubule stability have been developed to identify and characterize proteins involved in controlling microtubule stability. This study introduced a simple ex-vivo assay for identifying potential microtubule regulators in the wing imaginal disc of Drosophila melanogaster. This assay utilizes silicon rhodamine-tubulin (SiR-Tub) as a cell-permeable fluorogenic dye for labeling microtubules. In an attempt to increase the sensitivity of the screen, an assay was designed using a sensitized microtubule condition. Wing discs are treated with SiR-Tub followed by demecolcine, a microtubule inhibitor, to partially label impaired microtubules. Under this sensitized condition, it was possible to test whether overexpression or downregulation of a gene can enhance or suppress the weakened SiR-Tub labeling. This assay allows highly sensitive detection of microtubules in developing larval tissues. Hence, it provides a useful tool for identifying new microtubule regulators in both unfixed and fixed imaginal discs in Drosophila. This strategy may also be applied to characterize microtubule regulators in tissues from other model organisms (Mok, 2021).
Mutations in the gene encoding vesicle-associated membrane protein B (VAPB) cause a familial form of amyotrophic lateral sclerosis (ALS). Expression of an ALS-related variant of vapb (vapbP58S) ) in Drosophila motor neurons results in morphologic changes at the larval neuromuscular junction (NMJ) characterized by the appearance of fewer, but larger, presynaptic boutons. Although diminished microtubule stability is known to underlie these morphologic changes, a mechanism for the loss of presynaptic microtubules has been lacking. By studying flies of both sexes, this study demonstrate the suppression of vapbP58S) -induced changes in NMJ morphology by either a loss of endoplasmic reticulum (ER) Ca(2+) release channels or the inhibition Ca(2+)/calmodulin (CaM)-activated kinase II (CaMKII). These data suggest that decreased stability of presynaptic microtubules at vapbP58S NMJs results from hyperactivation of CaMKII because of elevated cytosolic [Ca(2+)]. The Ca(2+) dyshomeostasis is attributed to delayed extrusion of cytosolic Ca(2+) Suggesting that this defect in Ca(2+) extrusion arose from an insufficient response to the bioenergetic demand of neural activity, depolarization-induced mitochondrial ATP production was diminished in vapbP58S neurons. These findings point to bioenergetic dysfunction as a potential cause for the synaptic defects in vapbP58S -expressing motor neurons (Karagas, 2022).
The expulsion of dying epithelial cells requires well-orchestrated remodelling steps to maintain tissue sealing. This process, named cell extrusion, has been mostly analysed through the study of actomyosin regulation. Yet, the mechanistic relationship between caspase activation and cell extrusion is still poorly understood. Using the Drosophila pupal notum, a single layer epithelium where extrusions are caspase-dependent, this study showed that the initiation of cell extrusion and apical constriction are surprisingly not associated with the modulation of actomyosin concentration and dynamics. Instead, cell apical constriction is initiated by the disassembly of a medio-apical mesh of microtubules which is driven by effector caspases. Importantly, the depletion of microtubules is sufficient to bypass the requirement of caspases for cell extrusion, while microtubule stabilisation strongly impairs cell extrusion. This study shows that microtubules disassembly by caspases is a key rate-limiting step of extrusion, and outlines a more general function of microtubules in epithelial cell shape stabilisation (Villars, 2022).
Global regulation of spindle-associated proteins is crucial in oocytes due to the absence of centrosomes and their very large cytoplasmic volume, but little is known about how this is achieved beyond involvement of the Ran-importin pathway. Previous work has uncovered a novel regulatory mechanism in Drosophila oocytes, in which the phospho-docking protein 14-3-3 suppresses microtubule binding of Kinesin-14/Ncd away from chromosomes. This paper reports systematic identification of microtubule-associated proteins regulated by 14-3-3 from Drosophila oocytes. Proteins from ovary extract were co-sedimented with microtubules in the presence or absence of a 14-3-3 inhibitor. Through quantitative mass-spectrometry, proteins or complexes were identified whose ability to bind microtubules is suppressed by 14-3-3, including the chromosomal passenger complex (CPC), the centralspindlin complex and Kinesin-14/Ncd. 14-3-3 binds to the disordered region of Borealin, and this binding is regulated differentially by two phosphorylations on Borealin. Mutations at these two phospho-sites compromised normal Borealin localisation and centromere bi-orientation in oocytes, showing that phospho-regulation of 14-3-3 binding is important for Borealin localisation and function (Repton, 2022).
Diversity in cytoskeleton organization and function may be achieved through alternative tubulin isotypes and by a variety of post-translational modifications. The Drosophila genome contains five different β-tubulin paralogs, which may play an isotype tissue-specific function in vivo. One of these genes, the β-tubulin60D gene, which is expressed in a tissue-specific manner, was found to be essential for fly viability and fertility. To further understand the role of the β-tubulin60D gene, new β-tubulin60D null alleles (β-tubulin60D M) using the CRISPR/Cas9 system and found that the homozygous flies were viable and fertile. Moreover, using a combination of genetic complementation tests, rescue experiments, and cell biology analyses, Pin 1, an unknown dominant mutant with bristle developmental defects, was identified as a dominant-negative allele of β-tubulin60D. A missense mutation in the Pin1 mutant that results in an amino acid replacement from the highly conserved glutamate at position 75 to lysine (E75K). Analyzing the β-tubulin structure suggests that this E75K alteration destabilizes the alpha-helix structure and may also alter the GTP-Mg(2+) complex binding capabilities. These results revisited the credence that β-tubulin60D is required for fly viability and revealed for the first time in Drosophila, a novel dominant-negative function of missense β-tubulin60D mutation in bristle morphogenesis (Krishnan, 2021).
The spectraplakin family of proteins includes ACF7/MACF1 and BPAG1/dystonin in mammals, VAB-10 in Caenorhabditis elegans, Magellan in zebrafish, and Short stop (Shot), the sole Drosophila member. Spectraplakins are giant cytoskeletal proteins that cross-link actin, microtubules, and intermediate filaments, coordinating the activity of the entire cytoskeleton. This study examined the role of Shot during cell migration using two systems: the in vitro migration of Drosophila tissue culture cells and in vivo through border cell migration. RNA interference (RNAi) depletion of Shot increases the rate of random cell migration in Drosophila tissue culture cells as well as the rate of wound closure during scratch-wound assays. This increase in cell migration prompted an analysis of focal adhesion dynamics. The rates of focal adhesion assembly and disassembly were faster in Shot-depleted cells, leading to faster adhesion turnover that could underlie the increased migration speeds. This regulation of focal adhesion dynamics may be dependent on Shot being in an open confirmation. Using Drosophila border cells as an in vivo model for cell migration, this study found that RNAi depletion led to precocious border cell migration. Collectively, these results suggest that spectraplakins not only function to cross-link the cytoskeleton but may regulate cell-matrix adhesion (Zhao, 2022).
Tubulin s-palmitoylation involves the thioesterification of a cysteine residue in tubulin with palmitate. The palmitate moiety is produced by the fatty acid synthesis pathway, which is rate-limited by acetyl-CoA carboxylase (ACC). While it is known that ACC is phosphorylated at serine 79 (pSer(79)) by AMPK and accumulates at the spindle pole (SP) during mitosis, a functional role for tubulin palmitoylation during mitosis has not been identified. This study found that modulating pSer(79)-ACC level at the SP using AMPK agonist and inhibitor induced spindle defects. Loss of ACC function induced spindle abnormalities in cell lines and in germ cells of the Drosophila germarium, and palmitic acid (PA) rescued the spindle defects in the cell line treated transiently with the ACC inhibitor, TOFA. Furthermore, inhibition of protein palmitoylating or depalmitoylating enzymes also induced spindle defects. Together, these data suggested that precisely regulated cellular palmitate level and protein palmitoylation may be required for accurate spindle assembly. It was then shown that tubulin was largely palmitoylated in interphase cells but less palmitoylated in mitotic cells. TOFA treatment diminished tubulin palmitoylation at doses that disrupt microtubule (MT) instability and cause spindle defects. Moreover, spindle MTs comprised of α-tubulins mutated at the reported palmitoylation site exhibited disrupted dynamic instability. It was also found that TOFA enhanced the MT-targeting drug-induced spindle abnormalities and cytotoxicity. Thus, this study reveals that precise regulation of ACC during mitosis impacts tubulin palmitoylation to delicately control MT dynamic instability and spindle assembly, thereby safeguarding nuclear and cell division (Fang, 2023).
In epithelial cells, planar polarisation of subapical microtubule networks is thought to be important for both breaking cellular symmetry and maintaining the resulting cellular polarity. Studies in the Drosophila pupal wing and other tissues have suggested two alternative mechanisms for specifying network polarity. On one hand mechanical strain and/or cell shape have been implicated as key determinants, on the other the Fat-Dachsous planar polarity pathway has been suggested to be the primary polarising cue. Using quantitative image analysis in the pupal wing, this study reassessed these models. It was found that cell shape was a strong predictor of microtubule organisation in the developing wing epithelium. Conversely Fat-Dachsous polarity cues do not play any direct role in the organisation of the subapical microtubule network, despite being able to weakly recruit the microtubule minus-end capping protein Patronin to cell boundaries. It is concluded that any effect of Fat-Dachsous on microtubule polarity is likely to be indirect, via their known ability to regulate cell shape (Moreno, 2023).
Microtubules (MTs) are built from α-/β-tubulin dimers and used as tracks by kinesin and dynein motors to transport a variety of cargos, such as mRNAs, proteins, and organelles, within the cell. Tubulins are subjected to several post-translational modifications (PTMs). Glutamylation is one of them, and it is responsible for adding one or more glutamic acid residues as branched peptide chains to the C-terminal tails of both α- and β-tubulin. However, very little is known about the specific modifications found on the different tubulin isotypes in vivo and the role of these PTMs in MT transport and other cellular processes in vivo. This study found that in Drosophila ovaries, glutamylation of α-tubulin isotypes occurred clearly on the C-terminal ends of αTub84B and αTub84D (αTub84B/D). In contrast, the ovarian α-tubulin, αTub67C, is not glutamylated. The C-terminal ends of αTub84B/D are glutamylated at several glutamyl sidechains in various combinations. Drosophila TTLL5 is required for the mono- and poly-glutamylation of ovarian αTub84B/D and with this for the proper localization of glutamylated microtubules. Similarly, the normal distribution of kinesin-1 in the germline relies on TTLL5. Next, two kinesin-1-dependent processes, the precise localization of Staufen and the fast, bidirectional ooplasmic streaming, depend on TTLL5, too, suggesting a causative pathway. In the nervous system, a mutation of TTLL5 that inactivates its enzymatic activity decreases the pausing of anterograde axonal transport of mitochondria. These results demonstrate in vivo roles of TTLL5 in differential glutamylation of α-tubulins and point to the in vivo importance of α-tubulin glutamylation for cellular functions involving microtubule transport (Bao, 2023).
Microtubules and their associated motors are important players in nucleus positioning. Although nuclear migration in Drosophila oocytes is controlled by microtubules, a precise role for microtubule-associated molecular motors in nuclear migration has yet to be reported. This study characterized novel landmarks that allow a precise description of the pre-migratory stages. Using these newly defined stages, it is reported that, before migration, the nucleus moves from the oocyte anterior side toward the center and concomitantly the centrosomes cluster at the posterior of the nucleus. In the absence of Kinesin-1, centrosome clustering is impaired and the nucleus fails to position and migrate properly. The maintenance of a high level of Polo-kinase at centrosomes prevents centrosome clustering and impairs nuclear positioning. In the absence of Kinesin-1, SPD-2, an essential component of the pericentriolar material, is increased at the centrosomes, suggesting that Kinesin-1-associated defects result from a failure to reduce centrosome activity. Consistently, depleting centrosomes rescues the nuclear migration defects induced by Kinesin-1 inactivation. Our results suggest that Kinesin-1 controls nuclear migration in the oocyte by modulating centrosome activity (Loh, 2023).
Dynein and kinesin motors mediate long-range intracellular transport, translocating towards microtubule minus and plus ends, respectively. Cargoes often undergo bidirectional transport by binding to both motors simultaneously. However, it is not known how motor activities are coordinated in such circumstances. In the Drosophila female germline, sequential activities of the dynein-dynactin-BicD-Egalitarian (DDBE) complex and of kinesin-1 deliver oskar messenger RNA from nurse cells to the oocyte, and within the oocyte to the posterior pole. This study shows through in vitro reconstitution that Tm1-I/C, a tropomyosin-1 isoform, links kinesin-1 in a strongly inhibited state to DDBE-associated oskar mRNA. Nuclear magnetic resonance spectroscopy, small-angle X-ray scattering and structural modeling indicate that Tm1-I/C suppresses kinesin-1 activity by stabilizing its autoinhibited conformation, thus preventing competition with dynein until kinesin-1 is activated in the oocyte. Thus work reveals a new strategy for ensuring sequential activity of microtubule motors (Heber, 2024).
oskar (osk) mRNA localization in the Drosophila egg chamber is an attractive system for studying dual motor transport. Delivery of osk to the posterior pole of the developing oocyte, which drives abdominal patterning and germline formation in the embryo, is driven by the successive activities of dynein and kinesin-1. In early oogenesis, osk mRNA that is synthesized in the nurse cells is transported into the interconnected oocyte by dynein in complex with dynactin and the activating adaptor Bicaudal D (BicD), which is linked to double-stranded mRNA localization signals by the RNA-binding protein Egalitarian (Egl). Association of Egl with BicD and consequent dynein activation are enhanced by binding of Egl to RNA, indicating a role for the cargo in promoting dynein activity. In early oogenesis, microtubule minus ends are nucleated in the oocyte, consistent with the dynein-based delivery of mRNAs into this cell. During mid-oogenesis, the polarity of the microtubule network shifts dramatically, with plus ends pointing towards the oocyte posterior. At this stage, Khc translocates osk to the posterior pole. This process is independent of Klc, raising the question of how Khc is linked to osk and how its motor activity is regulated (Heber, 2024).
Transport of osk RNA by Khc requires the unique I/C isoform of tropomyosin-1, Tm1-I/C (hereafter Tm1). Tm1 binds to a noncanonical but conserved cargo-binding region in the Khc tail and stabilizes interaction of the motor with RNA, suggesting a function as an adaptor. Both Khc and Tm1 are loaded onto osk ribonucleoprotein particles (RNPs) shortly after their export from the nurse cell nuclei, although the motor only appears to become active in the mid-oogenesis oocyte. Similarly, dynein remains associated with osk RNPs during Khc-mediated transport within the oocyte, but is inactivated by displacement of Egl by Staufen. How the two motors are linked simultaneously to osk RNPs, and how Khc is inhibited during dynein-mediated transport into the oocyte, is not known (Heber, 2024).
This study shows that Tm1 inhibits Khc by stabilizing its autoinhibited conformation through a new mechanism involving the motor’s regulatory tail domain and stalk. Tm1 also links Khc to the dynein-transported osk RNP, thereby allowing cotransport of inactive Khc on osk RNA by dynein. In vivo, such a mechanism would avoid competition between the two motors during delivery of osk RNPs to the oocyte by dynein, while ensuring that Khc is available on these structures to mediate their delivery to the oocyte posterior in mid-oogenesis. With its cargo-binding and motor regulatory functions, it is proposed that Tm1 is a noncanonical light chain for kinesin-1 (Heber, 2024).
Tm1 was recently implicated as an RNA adaptor for kinesin-1. The current study reveals a previously unknown role of Tm1 in osk transport. Tm1 is shown to negatively regulates Khc activity, which is proposed to occurs a conformational change in the Khc stalk that stabilizes the Khc motor-tail interaction and thereby enhances autoinhibition (Heber, 2024).
With its functions in cargo binding and motor regulation, it is speculated that Tm1 is an alternative Klc in the Drosophila female germline. Because precise osk RNA localization during oogenesis is critical for development, positive regulators of Khc function such as PAT1 and negative regulators such as Tm1 may have replaced Klc to provide nuanced control of Khc activity. If this hypothesis is correct, Klc should also be dispensable for Tm1-dependent RNA localization in somatic tissues (Heber, 2024).
Tm1 also stimulates association of Khc in a strongly inhibited state with dynein-associated osk RNPs. This mechanism would allow dynein-mediated transport of osk RNPs from the nurse cells to the oocyte to proceed without competition with Khc, while positioning the plus-end directed motor on the RNPs for their posteriorward transport within the ooplasm in mid-oogenesis (Heber, 2024).
Kinesin-1 autoinhibition is not fully understood, in part because of a paucity of structural information for the full-length molecule. It has been proposed that interaction of the tail’s IAK motif with the motor domain plays a key role in kinesin-1 autoinhibition. Consistent with this notion, strongly enhanced motility was obserced of Drosophila Khc when the IAK motif was deleted. However, it was recently shown that Klc inhibits Khc activity independently of the IAK-motor interaction. It was also found that the IAK motif is not needed for inhibition of Khc by Tm1. Instead, it was found that Tm1-mediated inhibition occurs via the Khc AMB domain, a region adjacent to the IAK motif that is essential for osk localization (Heber, 2024).
Structural analyses suggest that Tm1 stabilizes the autoinhibited folded conformation of Khc by inducing rearrangement of the Khc coiled-coil stalk. Therefore, a model is proposed in which both the motor-IAK interaction and interactions within the Tm1-bound stalk that include the AMB domain act synergistically to achieve the stable inhibited conformation of Khc (Heber, 2024).
Two recent studies have proposed compact structural arrangements for mammalian kinesin-1. Those studies used chemical crosslinking and cryo-EM, which are likely to enrich for a homogeneous population of compact conformations of Khc and Khc–Klc tetramers, to provide detailed static structural information. By contrast, in the current study of Drosophila Khc and Khc–Tm1 complexes, NMR and SAXS were employed providing insight into the different conformational states, and thus flexibility, of kinesin-1 by enabling analysis of structures in solution. A previous study observed the compact, inhibited conformation in the isolated Khc, showing that Klc-binding does not induce a new fold of Khc but rather stabilizes the inhibited conformation. This is in agreement with the current model, in which Tm1, a putative alternative Klc, shifts the structural equilibrium of Khc towards the autoinhibited state by stabilizing its compact conformation (Heber, 2024).
Although it is known that many cargo types are transported by the concerted action of dynein and kinesins, the underlying regulatory mechanisms have been elusive. This study has identified one of very few examples of a factor that not only links dynein and kinesin-mediated transport, but also modulates transport through differential motor regulation. Linkage of dynein and kinesin-3 by the dynein-activating adaptor Hook3 has been demonstrated, but the cellular events for which this is relevant are still emerging. Recently, reconstituted coupling of dynein and kinesin-1 by TRAK1 and TRAK2 has provided insight into how the motors are recruited and regulated for mitochondrial transport. However, unlike this study's integration of DDBE and Khc in reconstituted osk RNPs, these systems lacked intact native cargoes and thereby excluded potential cargo-directed positioning and modulation of motor complexes (Heber, 2024).
Several studies have reported that active dynein and kinesin motors engage in a tug-of-war when artificially coupled. This study also observe motor opposition in reconstituted RNPs containing DDBE and Khc, presumably because of the stochastic engagement of autoinhibited Khc with microtubules. However, the observation that Tm1 supports efficient dynein-mediated RNA transport through robust inhibition of Khc highlights the importance of regulatory factors in addition to mechanical coupling in native transport complexes. Supporting the in vivo relevance of negative regulation of Khc during bidirectional transport, kinesin-1-activating IAK mutations were recently shown to impair dynein-mediated transport processes in Aspergillus nidulans Collectively, these observations point to complex interplay between opposite-polarity motors that are bound simultaneously to cargoes. Further reconstitutions of dual motor systems on native cargoes should reveal generalities of dynein-kinesin crosstalk, as well as any cargo-specific regulatory mechanisms (Heber, 2024).
This study provides mechanistic insight into two critical aspects of osk mRNA transport—assembly of the dual motor complex and how Khc activity is suppressed during dynein-mediated delivery of the transcript from the nurse cells to the oocyte. However, it is not understood how Khc takes over from dynein after osk RNPs arrive in the oocyte. Although recent work has shown that inactivation of dynein by the RNA-binding protein Stau is part of this process, how Tm1- and IAK-mediated inhibition of Khc is alleviated to allow delivery of the mRNA to the oocyte posterior is an open question. One candidate to fulfill this role is Ensconsin, which is required for posterior osk localization and is enriched in the oocyte relative to the nurse cells. Strikingly, the human counterpart of Ensconsin (MAP7) was recently shown to stimulate activity of mammalian kinesin-1 in vitro. Other candidate Khc activators include the exon junction complex, which, together with the SOLE RNA structure, is essential for transport of osk to the oocyte posterior. Because Tm1 needs to remain bound to the osk RNP throughout its posterior translocation, it is likely that the activating factor(s) induces a conformational change in the Khc–Tm1 complex rather than dissociation of Tm1. Future investigations of these regulatory mechanisms are likely to elucidate how kinesin-1 activity is orchestrated in other systems (Heber, 2024).
Mechanoreceptor cells develop a specialized cytoskeleton that plays structural and sensory roles at the site of mechanotransduction. However, little is known about how the cytoskeleton is organized and formed. Using electron tomography and live-cell imaging, this study resolve the 3D structure and dynamics of the microtubule-based cytoskeleton in fly campaniform mechanosensory cilia. Investigating the formation of the cytoskeleton, serves two functions. First, it amplifies the mass of microtubules to form the dense microtubule arrays inside the sensory cilia. Second, it generates short microtubules that are required to build the nanoscopic cytoskeleton at the mechanotransduction site. Additional analyses further reveal the functional roles of Patronin and other potential factors in the local regulatory network. In all, these results characterize the specialized cytoskeleton in fly external mechanosensory cilia at near-molecular resolution and provide mechanistic insights into how it is formed (Sun, 2021).
The structural reconstruction reveals that microtubules in the outer segment are highly organized and stable. They form two structurally separate arrays (see Molecular organization of microtubules in the outer segment). In particular, the array in the mechanoreceptive organelle (MO) contains mostly short microtubules, which facilitate the structural optimization of the MO and provide anchoring sites to a large number of mechanosensory molecules (e.g., NompC), thereby serving key sensory functions. Furthermore, the microtubule arrays in the tubular body (TB) and MO are differentially regulated, suggesting dedicated patterning and regulatory mechanisms in the outer segment of fly external mechanoreceptors (Sun, 2021).
The present study provides two implications in understanding how the microtubule arrays in fly sensory cilia are formed. First, a large quantity of microtubules is required to form dense arrays in the outer segment. Kat-60L1 and Patronin together amplify the mass of microtubules. In addition, the presence of a small number of microtubules in one piggyBAC insertion mutant/null mutant (c01236/BE6) implies that there is a kat-60L1–independent mechanism (denoted as X1) to assemble microtubules. These microtubules may serve as the initial substrates of kat-60L1 to create more templates. Second, the nanoscopic cytoskeleton in the MO determines the number and spatial arrangement of NompC force-sensitive channels that can be accommodated in the MO. Kat-60L1 coordinates the number and length of microtubules to build the cytoskeleton in the MO, which likely facilitates the structural–mechanical design of the MO, for example, providing adequate anchoring sites for NompC or withstanding resting tensions for optimal mechanosensitivity. An ensuing issue is how short microtubules in the MO can be stabilized. The lower susceptibility of the MO microtubules to the reduction in the level of Patronin suggests the presence of additional stabilizing mechanisms (Sun, 2021).
The results suggest that kat-60L1 has two functions in fly ES cilia. First, Kat-60L1 increases the mass of microtubules in the outer segment. This agrees with the proposed function of microtubule-severing enzymes in amplifying the local mass of microtubules. Second, kat-60L1 is required to generate short microtubules. It has been shown that microtubule-severing enzymes sculpt cellular microtubule networks; for example, the noncentrosomal microtubule arrays in plant cells and the meiotic spindle. Previous studies have also implicated the potential functions of microtubule-severing enzymes in neurons. However, due to the limited optical resolution, visualizing dense microtubule networks (e.g., in neurons) where the severing activity is potentially important remains challenging. ET could be useful in such cases. In the present study, structural reconstruction provides direct evidence to demonstrate the severing activity of Kat-60L1 in fly ciliated sensory neurons, thereby providing new insights into the neuronal and ciliary functions of microtubule-severing enzymes (Sun, 2021).
A potential caveat to the above conclusion is that the loss of kat-60L1 also changes the homeostatic microtubule dynamics in the inner segment, so it is formally possible that the structural and functional defects in the outer segment might be a secondary effect. However, this is not thought to be the case based on three reasons. First, in kat-60L1i, no significant microtubule accumulation was observed in the inner segment, but the cytoskeleton in the outer segment was also disrupted, suggesting that microtubules in the outer segment are more susceptible to the level of kat-60L1, and that the defects in the outer segment are a primary consequence of losing kat-60L1. Second, more microtubule polymerizations, suggested by the observation of more frequent EB1 comets, are observed in the inner segment in c01236/BE6, suggesting that the level of free tubulin dimers is fairly normal. Because tubulin is transported or diffuses into cilium as dimers, it is argued that tubulin influx into the cilia, which matters for microtubule dynamics in the outer segment, is not down-regulated in c01236/BE6. Third, no major defects in the morphology of dendritic inner segments were observed in c01236/BE6, suggesting that the functional defects can be mostly attributed to the defects in the outer segment (Sun, 2021).
Previous in vitro studies proposed that the cellular effects of microtubule-severing enzymes depend on the ratio between the amount of severing enzymes and the local microtubule dynamics. This study shows that in the outer segment, where microtubules are stable and the signal of Kat-60L1 is weak, Kat-60-L1 acts as a constructor, while in the inner segment, where microtubules are more dynamic and Kat-60L1 shows an enriched signal, Kat-60-L1 tends to act as a destructor. These results provide an in vivo example that supports the predictions from the previously proposed model (Sun, 2021).
The function of katanin family members has been implicated in ciliogenesis, especially in the assembly of central pair microtubules in motile cilia. Fly mechanosensory cilia contain no central pair microtubules but numerous noncentrosomal microtubules. The disruption of the cytoskeleton in the absence of Kat-60L1 in fly mechanosensory cilia resembles the loss of central pair microtubules in the katanin mutants of other species. This implies a common component in the assembly mechanism of noncentrosomal microtubules inside cilia, which, however, remains a hypothesis waiting for future studies (Sun, 2021).
Cytoplasmic microtubules are tubular polymers that can harbor small proteins or filaments inside their lumen. The identities of these objects and mechanisms for their accumulation have not been conclusively established. This study used cryogenic electron tomography of Drosophila S2 cell protrusions and found filaments inside the microtubule lumen, which resemble those reported recently in human HAP1 cells. The frequency of these filaments increased upon inhibition of the sarco/endoplasmic reticulum Ca(2+) ATPase with the small molecule drug thapsigargin. Subtomogram averaging showed that the luminal filaments adopt a helical structure reminiscent of cofilin-bound actin (cofilactin). Consistent with this, cofilin dephosphorylation, an activating modification, was observed in cells under the same conditions that increased luminal filament occurrence. Furthermore, RNA interference knock-down of cofilin reduced the frequency of luminal filaments with cofilactin morphology. These results suggest that cofilin activation stimulates its accumulation on actin filaments inside the microtubule lumen (Ventura Santos, 2023)
Mechanoreceptor cells develop specialized mechanosensory organelles (MOs), where force-sensitive channels and supporting structures are organized in an orderly manner to detect forces. It is intriguing how MOs are formed. This issue was addressed by studying the MOs of fly ciliated mechanoreceptors. The main structure of the MOs is shown to be a compound cytoskeleton formed of short microtubules and electron-dense materials (EDMs). In a knock-out mutant of microtubule associated protein DCX-EMAP, this cytoskeleton is nearly absent, suggesting that DCX-EMAP is required for the formation of the MOs and in turn fly mechanotransduction. Further analysis reveals that DCX-EMAP expresses in fly ciliated mechanoreceptors and localizes to the MOs. Moreover, it plays dual roles by promoting the assembly/stabilization of the microtubules and the accumulation of the EDMs in the MOs. Therefore, DCX-EMAP serves as a core ultrastructural organizer of the MOs, and this finding provides novel molecular insights as to how fly MOs are formed (Song, 2023).
Mechanoreceptor cells start the neural pathway of mechanosensation by converting physical stimuli (e.g., force or deformation) into cellular signals. To accomplish this task, they develop specialized mechanosensory organelles (MOs), which are structural–mechanical entities that consist of force-sensitive channels and supporting components, such as the cytoskeleton and extracellular matrix. While force-sensitive ion channels are key signal transducers, supporting components were thought to refine the sensory features of mechanoreceptor cells (e.g., sensitivity and dynamic range). For example, vertebrate inner ear hair cells grow stereocilia that contain intracellular actin bundles and extracellular tip-links, which serve to match the mechanical impedance when stereocilia deflection is converted into a conformational change of the mechanotransduction channels. The touch receptors of C. elegans form a specialized sensory complex containing the extracellular matrix (e.g., MEC-5), membrane channels (MEC-4 and MEC-10), and 15-protofilament microtubules (MEC-7 and MEC-12) to sense tactile signals. Recent studies suggest that Piezo, a force-sensitive channel that contributes to the perception of various mechanical stimuli, may be tethered to and regulated by F-actin in cells (Wang, 2022). Therefore, MOs are structurally specialized to match the sensory modality of the mechanoreceptors. This raises the question of how the MOs are formed (Song, 2023).
The MOs of Drosophila ciliated mechanoreceptors (i.e., type I mechanoreceptors) have been extensively studied to understand the structural basis of mechanotransduction. Early ultrastructural analysis showed that the main intracellular structure of the MOs is a compound cytoskeletal structure consisting of arrayed microtubules and electron-dense materials (EDMs). Later, it was shown that NompC force-sensitive channels are structurally linked to the microtubules and also formed into arrays in the MO membrane. These observations suggest that the entire MO acts as an integrated mechanosensor with a nanoscopic size. In recent work, it was revealed that short microtubules are required for the formation of the MOs and that the 'kat-60L1/Patronin' module is required to generate the short microtubules (Sun, 2021). An ensuing question is how these short microtubules are stabilized and organized in the MOs (Song, 2023).
Previous work showed that knocking down the expression level of Patronin, a microtubule minus-end–stabilizing protein, reduces the amount of the MO microtubules (Sun, 2021), suggesting that Patronin stabilizes the MO microtubules. However, this study also found that in addition to Patronin, there is a MO-specific microtubule-stabilizing mechanism (Sun, 2021). A previous study showed that DCX-EMAP, a doublecortin (DCX) domain-containing protein, is highly enriched in fly haltere tissue based on the DNA array analysis and that a piggyBac insertion mutant of DCX-EMAP, which may express a truncated protein (Liang, 2014), contains a disrupted microtubule array in the outer segment of campaniform mechanoreceptors. This implies that DCX-EMAP may be key for the assembly or stability of microtubules inside the mechanosensory cilia. However, due to the lack of further genetic and cell biological analysis, the cellular functions and biochemical mechanism of DCX-EMAP remain elusive, especially in the context of the recently resolved structure of the mechanosensory cilia (Song, 2023).
DCX-EMAP belongs to the EMAP (echinoderm-microtubule-associated proteins) family, the members of which are regulators for microtubule dynamics. All EMAP proteins share a conserved TAPE (tandem atypical propeller in EMLs) domain at the carboxyl-terminal, including the HELP (hydrophobic echinoderm-microtubule-associated-like protein) domain and multiple WD40 repeats. The amino-terminal part of EMAP proteins often contains a coiled-coil domain that binds to microtubules through trimerization. Among all EMAP family members, DCX-EMAP is unique because the coiled-coil domain is replaced by a tandem of two DCX domains (Bechstedt, 2010), which has a microtubule-binding/stabilizing activity. It was found that the DCX-domain-containing (DCDC) proteins express in a wide range of cells and show diverse cellular functions. Therefore, it is intriguing to understand how DCX-EMAP couples the functions of the DCX and EMAP families in a specific cellular process, such as the formation of a modified cilium (Song, 2023).
This work studied the formation of fly MOs by analyzing the cellular functions of DCX-EMAP, an essential molecule for fly mechanosensation. The results show that DCX-EMAP specifically expresses in fly mechanosensory cilia and acts as a core organizer for the ultrastructure of the MOs, thereby having a direct contribution to fly mechanotransduction. These findings help take an important step forward in understanding how fly MOs are formed. Additionally, this study provides implications to understand the cellular roles of the DCDC and EMAP family members in ciliary assembly and maintenance (Song, 2023).
This study resolved the 3D ultrastructural organization of the compound cytoskeleton in fly MOs. By studying the function and working mechanism of DCX-EMAP, novel insights were provided into understanding how fly MOs are formed. The key finding is that DCX-EMAP, an essential molecule for fly mechanotransduction, serves as the core ultrastructural organizer of the MOs by locally stabilizing and organizing the microtubule-EDM complex (Song, 2023).
The results demonstrate that the DCX tandem and the TAPE domain (i.e., the HELP + WD40 domains) are all required for the in vivo function of DCX-EMAP. First, in vitro analysis shows that the DCX tandem of DCX-EMAP has a microtubule-binding/stabilizing activity, in which both DCX domains and the structural linkage in between are required. In vivo experiments show that the mutant DCX-EMAP with no or only one DCX domain cannot rescue the cellular and functional phenotypes of DCX-EMAPKO, suggesting that the intact microtubule-binding/stabilizing activity is required for the formation of the MOs. Second, the HELP domain is conserved among all EMAP family members. The HELP domain of DCX-EMAP was shown to be key for the MO-specific localization. However, it is not yet clear how the HELP works at the molecular level. It might interact with other components of the MOs (such as the EDMs) or ciliary motors that are responsible for the directional transport to the MOs. Third, the WD40 domain, a known molecular platform to mediate protein–protein interactions, is also conserved in the EMAP family. It is noted that ΔWD40 could still localize to the MO but the 3D porous structure of the EDMs is absent, suggesting that the WD40 domain of DCX-EMAP is key for the local organization of the EDMs. To refine the in vivo working mechanism of the HELP and WD40 domains in this model, it would be essential to identify more components of the EDMs in future studies (Song, 2023).
Based on these findings, it is proposed that DCX-EMAP serves as a component in the structural link between the microtubules and EDMs in the MOs. More specifically, the DCX tandem promotes the assembly and stabilization of short microtubules in the MOs. The HELP and WD40 domains mediate the localization signal and organize the EDMs. In this model, DCX-EMAP promotes the assembly and stability of the MO microtubules, which would facilitate local accumulation of the EDMs and DCX-EMAP by providing more landing or binding sites. Then, an increase in the amount of DCX-EMAP would in turn promote the assembly or stability of more MO microtubules, thereby mediating a positive feedback loop (Song, 2023).
This study showed that short microtubules, generated by the 'kat-60L1-patronin' module, provide constructional flexibility in the formation of the compound cytoskeleton within the nanoscopic space of the MOs. As a concurrent mechanism, the positive feedback loop mediated by DCX-EMAP would facilitate the full assembly of all other components into a compound cytoskeleton in the MOs. The sensillar structures, in particular the MO membrane and extracellular sheath, could serve as a physical boundary to constrain this positive feedback and control the overall shape/size of the MOs. This point is supported by the observation that in the nompA mutants, where the extracellular contact of the MOs is lost, the morphology of the MOs is largely altered. Based on these considerations, it is concluded that DCX-EMAP acts as a core ultrastructural organizer for the MOs of fly ciliated mechanoreceptors (Song, 2023).
The functions of other DCDCs have also been implicated in cilia assembly and ciliopathy. For example, a missense mutation in dcdc2, which encodes DCDC2, causes human recessive deafness, likely by interfering with the structures of sensory hair cells and the supporting cells. Similar to DCX-EMAP, DCDC2 has a tandem pair of DCX domains at the amino terminus and an unstructured tail of over 200 residues at the carboxyl terminus, where the pathogenetic mutation is. Moreover, it also tends to localize to the distal end of cilia, similar to DCX-EMAP. Although the cellular functions and working mechanism of DCDC2 still await further studies, the expression of the deaf mutant of DCDC2 leads to disrupted ciliary structure, such as cilium branching and dysregulation of ciliary length, suggesting that DCDC2 has an essential role in organizing ciliary structures. This is to some extent similar to the function of DCX-EMAP in fly mechanosensory cilia. The resemblance in the cell biological features of DCDC2 and DCX-EMAP may suggest a common way of how DCDCs work in regulating ciliary structure, e.g., the ciliary tip compartment (Song, 2023).
Biological systems are highly complex, yet notably ordered structures can emerge. During syncytial stage development of the Drosophila melanogaster embryo, nuclei synchronously divide for nine cycles within a single cell, after which most of the nuclei reach the cell cortex. The arrival of nuclei at the cortex occurs with remarkable positional order, which is important for subsequent cellularisation and morphological transformations. Yet, the mechanical principles underlying this lattice-like positional order of nuclei remain untested. Using quantification of nuclei position and division orientation together with embryo explants, this study shows that short-ranged repulsive interactions between microtubule asters ensure the regular distribution and maintenance of nuclear positions in the embryo. Such ordered nuclear positioning still occurs with the loss of actin caps and even the loss of the nuclei themselves; the asters can self-organise with similar distribution to nuclei in the wild-type embryo. The explant assay enabled deduction of the nature of the mechanical interaction between pairs of nuclei. This was used to predict how the nuclear division axis orientation changes upon nucleus removal from the embryo cortex, which was confirmed in vivo with laser ablation. Overall, this study shows that short-ranged microtubule-mediated repulsive interactions between asters are important for ordering in the early Drosophila embryo and minimising positional irregularity (de-Carvalho, 2022).
The coiled-coil domains of intermediate filament (IF) proteins are flanked by regions of low sequence complexity. Whereas IF coiled-coil domains assume dimeric and tetrameric conformations on their own, maturation of eight tetramers into cylindrical IFs is dependent on either "head" or "tail" domains of low sequence complexity. This study confirms that the tail domain required for assembly of Drosophila Tropomyosin 1 (Tm1-I/C) IFs functions by forming labile cross-β interactions. These interactions are seen in polymers made from the tail domain alone, as well as in assembled IFs formed by the intact Tm1-I/C protein. The ability to visualize such interactions in situ within the context of a discrete cellular assembly lends support to the concept that equivalent interactions may be used in organizing other dynamic aspects of cell morphology (Sysoev, 2020).
Uzquiano, A., Cifuentes-Diaz, C., Jabali, A., Romero, D. M., Houllier, A., Dingli, F., Maillard, C., Boland, A., Deleuze, J. F., Loew, D., Mancini, G. M. S., Bahi-Buisson, N., Ladewig, J., Francis, F. (2019). Mutations in the Heterotopia Gene Eml1/EML1 Severely Disrupt the Formation of Primary Cilia. Cell Rep, 28(6):1596-1611 e1510 PubMed ID: 31390572
Apical radial glia (aRGs) are predominant progenitors during corticogenesis. Perturbing their function leads to cortical malformations, including subcortical heterotopia (SH), characterized by the presence of neurons below the cortex. EML1/Eml1 mutations lead to SH in patients, as well as to heterotopic cortex (HeCo) mutant mice. In HeCo mice, some aRGs are abnormally positioned away from the ventricular zone (VZ). Thus, unraveling EML1/Eml1 function will clarify mechanisms maintaining aRGs in the VZ. This study pinpoints an unknown EML1/Eml1 function in primary cilium formation. In HeCo aRGs, cilia are shorter, less numerous, and often found aberrantly oriented within vesicles. Patient fibroblasts and human cortical progenitors show similar defects. EML1 interacts with RPGRIP1L, a ciliary protein, and RPGRIP1L mutations were revealed in a heterotopia patient. This study also identified Golgi apparatus abnormalities in EML1/Eml1 mutant cells, potentially upstream of the cilia phenotype. It was thus reveal primary cilia mechanisms impacting aRG dynamics in physiological and pathological conditions (Uzquiano, 2029).
Abekhoukh, S., Sahin, H. B., Grossi, M., Zongaro, S., Maurin, T., Madrigal, I., Kazue-Sugioka, D., Raas-Rothschild, A., Doulazmi, M., Carrera, P., Stachon, A., Scherer, S., Nascimento, M. R., Trembleau, A., Arroyo, I., Peter, S., Smith, I. M., Mila, M., Smith, A. C., Giangrande, A., Caille, I. and Bardoni, B. (2017). New insights into the regulatory function of CYFIP1 in the context of WAVE- and FMRP-containing complexes. Dis Model Mech [Epub ahead of print]. PubMed ID: 28183735
Alzyoud, E., Vedelek, V., Rethi-Nagy, Z., Lipinszki, Z. and Sinka, R. (2021).
Microtubule Organizing Centers Contain Testis-Specific γ-TuRC Proteins in Spermatids of Drosophila. Front Cell Dev Biol 9: 727264. PubMed ID: 34660584
Antel, M., Simao, T., Bener, M. B. and Inaba, M. (2022). Drosophila CG17003/leaky (lky) is required for microtubule acetylation in early germ cells in Drosophila ovary. PLoS One 17(11): e0276704. PubMed ID: 36342916
Araujo, M., Tavares, A., Vieira, D. V., Telley, I. A. and Oliveira, R. A. (2023). Endoplasmic reticulum membranes are continuously required to maintain mitotic spindle size and forces. Life Sci Alliance 6(1). PubMed ID: 36379670
Avidor-Reiss, T., Maer, A. M., Koundakjian, E., Polyanovsky, A., Keil, T., Subramaniam, S. and Zuker, C. S. (2004). Decoding cilia function: defining specialized genes required for compartmentalized cilia biogenesis. Cell 117(4): 527-39. 15137945
Badmos, H., Cobbe, N., Campbell, A., Jackson, R. and Bennett, D. (2021). Drosophila USP22/nonstop polarizes the actin cytoskeleton during collective border cell migration. J Cell Biol 220(7). PubMed ID: 33988679
Baez-Cruz, F. A. and Ostap, E. M. (2023). Drosophila class-I myosins that can impact left-right asymmetry have distinct ATPase kinetics. J Biol Chem 299(8): 104961. PubMed ID: 37380077
Bao, M., Dorig, R. E., Vazquez-Pianzola, P. M., Beuchle, D. and Suter, B. (2023). Differential modification of the C-terminal tails of different α-tubulins and their importance for microtubule function in vivo. Elife 12. PubMed ID: 37345829
Barros, C. S. and Bossing, T. (2021). Microtubule disruption upon CNS damage triggers mitotic entry via TNF signaling activation. Cell Rep 36(1): 109325. PubMed ID: 34233183
Belyaeva, V., Wachner, S., Gyoergy, A., Emtenani, S., Gridchyn, I., Akhmanova, M., Linder, M., Roblek, M., Sibilia, M. and Siekhaus, D. (2022). Fos regulates macrophage infiltration against surrounding tissue resistance by a cortical actin-based mechanism in Drosophila. PLoS Biol 20(1): e3001494. PubMed ID: 34990456
Bischoff, M. (2012). Lamellipodia-based migrations of larval epithelial cells are required for normal closure of the adult epidermis of Drosophila. Dev Biol 363(1): 179-190. PubMed ID: 22230614
Bohrmann, J. and Biber, K. (1994). Cytoskeleton-dependent transport of cytoplasmic particles in previtellogenic to mid-vitellogenic ovarian follicles of Drosophila. time-lapse analysis using video-enhanced contrast microscopy. J. Cell Sci. 107: 849-858. 8056841
Brinkmann, K., Winterhoff, M., Onel, S. F., Schultz, J., Faix, J. and Bogdan, S. (2015). WHAMY is a novel actin polymerase promoting myoblast fusion, macrophage cell motility and sensory organ development. J Cell Sci 129(3):604-20. PubMed ID: 26675239
Bu, S., Yong, W. L., Lim, B. J. W., Kondo, S. and Yu, F. (2021). A systematic analysis of microtubule-destabilizing factors during dendrite pruning in Drosophila. EMBO Rep: e52679. PubMed ID: 34338441
Bu, S., Tang, Q., Wang, Y., Lau, S. S. Y., Yong, W. L. and Yu, F. (2022). Drosophila CLASP regulates microtubule orientation and dendrite pruning by suppressing Par-1 kinase. Cell Rep 39(9): 110887. PubMed ID: 35649352
Caridi, C. P., D'Agostino, C., Ryu, T., Zapotoczny, G., Delabaere, L., Li, X., Khodaverdian, V. Y., Amaral, N., Lin, E., Rau, A. R. and Chiolo, I. (2018). Nuclear F-actin and myosins drive relocalization of heterochromatic breaks. Nature. PubMed ID: 29925946
Chandran, R., Kale, G., Philippe, J. M., Lecuit, T. and Mayor, S. (2021). Distinct actin-dependent nanoscale assemblies underlie the dynamic and hierarchical organization of E-cadherin. Curr Biol. PubMed ID: 33607036
Cheerambathur, D. K., Prevo, B., Chow, T. L., Hattersley, N., Wang, S., Zhao, Z., Kim, T., Gerson-Gurwitz, A., Oegema, K., Green, R. and Desai, A. (2019). The kinetochore-microtubule coupling machinery is repurposed in sensory nervous system morphogenesis. Dev Cell 48(6): 864-872 e867. PubMed ID: 30827898
Chippalkatti, R., Egger, B. and Suter, B. (2020). Mms19 promotes spindle microtubule assembly in Drosophila neural stem cells. PLoS Genet 16(11): e1008913. PubMed ID: 33211700
Chung, S., Le, T. P., Vishwakarma, V., Cheng, Y. L. and Andrew, D. J. (2021). Isoform-specific roles of the Drosophila filamin-type protein Jitterbug (Jbug) during development. Genetics. PubMed ID: 34173831
Cloud, V., Thapa, A., Morales-Sosa, P., Miller, T. M., Miller, S. A., Holsapple, D., Gerhart, P. M., Momtahan, E., Jack, J. L., Leiva, E., Rapp, S. R., Shelton, L. G., Pierce, R. A., Martin-Brown, S., Florens, L., Washburn, M. P. and Mohan, R. D. (2019). Ataxin-7 and Non-stop coordinate SCAR protein levels, subcellular localization, and actin cytoskeleton organization. Elife 8. PubMed ID: 31348003
Companys, P. P, Norris, A. and Bischoff, M. (2020). Coordination of cytoskeletal dynamics and cell behaviour during Drosophila abdominal morphogenesis. J Cell Sci 133(6). PubMed ID: 32229579
Connelly, J. T., Gautrot, J. E., Trappmann, B., Tan, D. W., Donati, G., Huck, W. T. and Watt, F. M. (2010). Actin and serum response factor transduce physical cues from the microenvironment to regulate epidermal stem cell fate decisions. Nat Cell Biol 12: 711-718. PubMed ID: 20581838
Cooley, L., Verheyen, E. and Ayers, K. (1992).
chickadee encodes a profilin required for intercellular cytoplasm
transport during Drosophila oogenesis. Cell 69: 173-84. 1339308
Das, R., Bhattacharjee, S., Letcher, J. M., Harris, J. M., Nanda, S., Foldi, I.,
Lottes, E. N., Bobo, H. M., Grantier, B. D., Mihaly, J., Ascoli, G. A.
and Cox, D. N. (2021). Formin 3 directs dendritic architecture via microtubule regulation and is required for somatosensory nociceptive behavior. Development 148(16). PubMed ID: 34322714
de-Carvalho, J., Tlili, S., Hufnagel, L., Saunders, T. E. and Telley, I. A. (2022). Aster repulsion drives short-ranged ordering in the Drosophila syncytial blastoderm. Development 149(2). PubMed ID: 35001104
Deng, Q., Tan, Y. S., Chew, L. Y. and Wang, H. (2021). Msps governs acentrosomal microtubule assembly and reactivation of quiescent neural stem cells. Embo j: e104549. PubMed ID: 34368973
Dhanyasi, N., VijayRaghavan, K., Shilo, B. Z. and Schejter, E. D. (2020). Microtubules provide guidance cues for myofibril and sarcomere assembly and growth. Dev Dyn. PubMed ID: 32725855
Dobramysl, U., Jarsch, I. K., Inoue, Y., Shimo, H., Richier, B., Gadsby, J. R., Mason, J., Szalapak, A., Ioannou, P. S., Correia, G. P., Walrant, A., Butler, R., Hannezo, E., Simons, B. D. and Gallop, J. L. (2021). Stochastic combinations of actin regulatory proteins are sufficient to drive filopodia formation. J Cell Biol 220(4). PubMed ID: 33740033
Dohn, T. E. and Cripps, R. M. (2018). Absence of the Drosophila jump muscle actin Act79B is compensated by up-regulation of Act88F. Dev Dyn [Epub ahead of print]. PubMed ID: 29318731
Duan, D., Lyu, W., Chai, P., Ma, S., Wu, K., Wu, C., Xiong, Y., Sestan, N., Zhang, K., Koleske, A. J. (2023). Abl2 repairs microtubules and phase separates with tubulin to promote microtubule nucleation. Curr Biol, 33(21):4582-4598.e4510 PubMed ID: 37858340
Dzaki, N., Bu, S., Lau, S. S. Y., Yong, W. L. and Yu, F. (2022). Drosophila GSK3β promotes microtubule disassembly and dendrite pruning in sensory neurons. Development 149(22). PubMed ID: 36264221
Fan, A., Joy, M. S. H. and Saif, T. (2019). A connected cytoskeleton network generates axonal tension in embryonic Drosophila. Lab Chip 19(18): 3133-3139. PubMed ID: 31435630
Fang, C. T., Kuo, H. H., Amartuvshin, O., Hsu, H. J., Liu, S. L., Yao, J. S. and Yih, L. H. (2023). Inhibition of acetyl-CoA carboxylase impaired tubulin palmitoylation and induced spindle abnormalities. Cell Death Discov 9(1): 4. PubMed ID: 36617578
Farrell, L., Puig-Barbe, A., Haque, M. I., Amcheslavsky, A., Yu, M., Bergmann, A. and Fan, Y. (2022). Actin remodeling mediates ROS production and JNK activation to drive apoptosis-induced proliferation. PLoS Genet 18(12): e1010533. PubMed ID: 36469525
Feng, C., Thyagarajan, P., Shorey, M., Seebold, D. Y., Weiner, A. T., Albertson, R. M., Rao, K. S., Sagasti, A., Goetschius, D. J. and Rolls, M. M. (2019). Patronin-mediated minus end growth is required for dendritic microtubule polarity. J Cell Biol 218(7): 2309-2328. PubMed ID: 31076454
Feng, C., Cleary, J. M., Kothe, G. O., Stone, M. C., Weiner, A. T., Hertzler, J. I., Hancock, W. O. and Rolls, M. M. (2021). Trim9 and Klp61F promote polymerization of new dendritic microtubules along parallel microtubules. J Cell Sci. PubMed ID: 33988240
Gao, Y., Shuai, Y., Zhang, X., Peng, Y., Wang, L., He, J., Zhong, Y. and Li, Q. (2019). Genetic dissection of active forgetting in labile and consolidated memories in Drosophila. Proc Natl Acad Sci U S A. 116(42):21191-21197. PubMed ID: 31488722
Gillard, G., Girdler, G. and Roper, K. (2021). A release-and-capture mechanism generates an essential non-centrosomal microtubule array during tube budding. Nat Commun 12(1): 4096. PubMed ID: 34215746
Goshima, G. and Vale, R. D. (2003). The roles of microtubule-based motor proteins in mitosis : comprehensive RNAi analysis in the Drosophila S2 cell line. J. Cell Bio. 162: 1003-1016. 12975346
Hahn, I., Voelzmann, A., Parkin, J., Fulle, J. B., Slater, P. G., Lowery, L. A., Sanchez-Soriano, N. and Prokop, A. (2021). Tau, XMAP215/Msps and Eb1 co-operate interdependently to regulate microtubule polymerisation and bundle formation in axons. PLoS Genet 17(7): e1009647. PubMed ID: 34228717
Hampoelz, B., et al. (2011). Microtubule-induced nuclear envelope fluctuations control chromatin dynamics in Drosophila embryos. Development 138(16): 3377-86. PubMed Citation: 21752932
Heber, S., McClintock, M. A., Simon, B., Mehtab, E., Lapouge, K., Hennig, J., Bullock, S. L., Ephrussi, A. (2024). Tropomyosin 1-I/C coordinates kinesin-1 and dynein motors during oskar mRNA transport. Nat Struct Mol Biol, 31(3):476-488 PubMed ID: 38297086
Henry, S. M., Xie, Y., Rollins, K. R. and Blankenship, J. T. (2022). Sponge/DOCK-dependent regulation of F-actin networks directing cortical cap behaviors and syncytial furrow ingression. Dev Biol 491: 82-93. PubMed ID: 36067836
Hertzler, J. I., Simonovitch, S. I., Albertson, R. M., Weiner, A. T., Nye, D. M. R. and Rolls, M. M. (2020). Kinetochore proteins suppress neuronal microtubule dynamics and promote dendrite regeneration. Mol Biol Cell: mbcE20040237T. PubMed ID: 32673176
Hirschhauser, A., van Cann, M. and Bogdan, S. (2021). CK1alpha protects WAVE from degradation to regulate cell shape and motility in immune response. J Cell Sci. PubMed ID: 34730182
Hu, C., Feng, P., Chen, M., Tang, Y. and Soba, P. (2022).
Spatiotemporal changes in microtubule dynamics during dendritic morphogenesis.
Fly (Austin) 16(1): 13-23. PubMed ID: 34609266
Huang, Y., Mao, X., van Jaarsveld, R. H., Shu, L., Terhal, P. A., Jia, Z., Xi, H., Peng, Y., Yan, H., Yuan, S., Li, Q., Wang, H. and Bellen, H. J. (2020). Variants in CAPZA2, a member of a F-actin capping complex, cause intellectual disability and developmental delay. Hum Mol Genet. PubMed ID: 32338762
Ibar, C., Chinthalapudi, K., Heiβler, S. M. and Irvine, K. D. (2023). Competition between myosin II and β(H)-spectrin regulates cytoskeletal tension. Elife 12. PubMed ID: 37367948
Jakobs, M. A. H., Zemel, A. and Franze, K. (2022). Unrestrained growth of correctly oriented microtubules instructs axonal microtubule orientation. Elife 11. PubMed ID: 36214669
Juan, T., Geminard, C., Coutelis, J. B., Cerezo, D., Poles, S., Noselli, S. and Furthauer, M. (2018). Myosin1D is an evolutionarily conserved regulator of animal left-right asymmetry. Nat Commun 9(1): 1942. PubMed ID: 29769531
Karg, T., Elting, M. W., Vicars, H., Dumont, S. and Sullivan, W. (2017). The chromokinesin Klp3a and microtubules facilitate acentric chromosome segregation. J Cell Biol. PubMed ID: 28500183
Karagas, N. E., Gupta, R., Rastegari, E., Tan, K. L., Leung, H. H., Bellen, H. J., Venkatachalam, K. and Wong, C. O. (2022). Loss of Activity-Induced Mitochondrial ATP Production Underlies the Synaptic Defects in a Drosophila Model of ALS. J Neurosci 42(42): 8019-8037. PubMed ID: 36261266
Khanal, I., Elbediwy, A., Diaz de la Loza, M.D., Fletcher, G.C. and Thompson, B.J. (2016). Shot and Patronin polarise microtubules to direct membrane traffic and biogenesis of microvilli in epithelia. J Cell Sci 129(13):2651-9. PubMed ID: 27231092
Krishnan, R. K., Halachmi, N., Baskar, R., Bakhrat, A., Zarivach, R., Salzberg, A. and Abdu, U. (2021). Revisiting the Role of beta-Tubulin in Drosophila Development: β-tubulin60D is not an Essential Gene, and its Novel Pin (1) Allele has a Tissue-Specific Dominant-Negative Impact. Front Cell Dev Biol 9: 787976. PubMed ID: 35111755
Kwiatkowski, S., Seliga, A. K., Vertommen, D., Terreri, M., Ishikawa, T., Grabowska, I., Tiebe, M., Teleman, A. A., Jagielski, A. K., Veiga-da-Cunha, M. and Drozak, J. (2018). SETD3 protein is the actin-specific histidine N-methyltransferase. Elife 7. PubMed ID: 30526847
Li, P., Messina, G. and Lehner, C. F. (2023). Nuclear elongation during spermiogenesis depends on physical linkage of nuclear pore complexes to bundled microtubules by Drosophila Mst27D. PLoS Genet 19(7): e1010837. PubMed ID: 37428798
Liang, X., Kokes, M., Fetter, R. D., Sallee, M. D., Moore, A. W., Feldman, J. L. and Shen, K. (2020). Growth cone-localized microtubule organizing center establishes microtubule orientation in dendrites. Elife 9. PubMed ID: 32657271
Loh, M., Bernard, F. and Guichet, A. (2023). Kinesin-1 promotes centrosome clustering and nuclear migration in the Drosophila oocyte. Development 150(13). PubMed ID: 37334771
Manseau, L., Calley, J. and Phan, H. (1996). Profilin is required for posterior patterning of the Drosophila oocyte. Development 122: 2109-16. 8681792
Martin, E., Theis, S., Gay, G., Monier, B., Rouviere, C. and Suzanne, M. (2021). Arp2/3-dependent mechanical control of morphogenetic robustness in an inherently challenging environment. Dev Cell 56(5): 687-701.e687. PubMed ID: 33535069
McGee, K. M., Vartiainen, M. K., Khaw, P. T., Treisman, R. and Bailly, M. (2011). Nuclear transport of the serum response factor coactivator MRTF-A is downregulated at tensional homeostasis. EMBO Rep 12: 963-970. PubMed ID: 21799516
Medjkane, S., Perez-Sanchez, C., Gaggioli, C., Sahai, E. and Treisman, R. (2009). Myocardin-related transcription factors and SRF are required for cytoskeletal dynamics and experimental metastasis. Nat Cell Biol 11: 257-268. PubMed ID: 19198601
Metivier, M., Gallaud, E., Thomas, A., Pascal, A., Gagne, J. P., Poirier, G. G., Chretien, D., Gibeaux, R., Richard-Parpaillon, L., Benaud, C. and Giet, R. (2020). Drosophila Tubulin-Specific Chaperone E Recruits Tubulin around Chromatin to Promote Mitotic Spindle Assembly. Curr Biol. PubMed ID: 33259793
Mohan, R. D., Dialynas, G., Weake, V. M., Liu, J., Martin-Brown, S., Florens, L., Washburn, M. P., Workman, J. L. and Abmayr, S. M. (2014). Loss of Drosophila Ataxin-7, a SAGA subunit, reduces H2B ubiquitination and leads to neural and retinal degeneration. Genes Dev 28(3): 259-272. PubMed ID: 24493646
Mok, J. W. and Choi, K. W. (2021a). Novel function of N-acetyltransferase for microtubule stability and JNK signaling in Drosophila organ development. Proc Natl Acad Sci U S A 118(4). PubMed ID: 33479178
Mok, J. W. and Choi, K. W. (2021b). Ex-vivo Microtubule Stability Assay Using Drosophila Wing Disc. Bio Protoc 11(23): e4241. PubMed ID: 35005086
Molina, E., Cataldo, V. F., Eggers, C., Munoz-Madrid, V. and Glavic, A. (2022). p53 Related Protein Kinase is Required for Arp2/3-Dependent Actin Dynamics of Hemocytes in Drosophila melanogaster. Front Cell Dev Biol 10: 859105. PubMed ID: 35721516
Molina-Pelayo, C., Olguin, P., Mlodzik, M. and Glavic, A. (2022). The conserved Pelado/ZSWIM8 protein regulates actin dynamics by promoting linear actin filament polymerization. Life Sci Alliance 5(12). PubMed ID: 35940847
Monahan Vargas, E. J., Matamoros, A. J., Qiu, J., Jan, C. H., Wang, Q., Gorczyca, D., Han, T. W., Weissman, J. S., Jan, Y. N., Banerjee, S. and Song, Y. (2020). The microtubule regulator ringer functions downstream from the RNA repair/splicing pathway to promote axon regeneration. Genes Dev 34(3-4): 194-208. PubMed ID: 31919191
Moreno, M. R., Hunton, R., Strutt, D. and Bulgakova, N. A. (2023). Deciphering the roles of cell shape and Fat and Dachsous planar polarity in arranging the Drosophila apical microtubule network through quantitative image analysis. Mol Biol Cell: mbcE22090442. PubMed ID: 36735484
Nakamura, M., Hui, J., Stjepic, V. and Parkhurst, S. M. (2023). Scar/WAVE has Rac GTPase-independent functions during cell wound repair. Sci Rep 13(1): 4763. PubMed ID: 36959278
Nanda, S., Bhattacharjee, S., Cox, D. N. and Ascoli, G. A. (2023). Local Microtubule and F-Actin Distributions Fully Constrain the Spatial Geometry of Drosophila Sensory Dendritic Arbors. Int J Mol Sci 24(7). PubMed ID: 37047715
Ryniawec, J. M., Hannaford, M. R., Zibrat, M. E., Fagerstrom, C. J., Galletta, B. J., Aguirre, S. E., Guice, B. A., Dean, S. M., Rusan, N. M., Rogers, G. C. (2023). Cep104 is a component of the centriole distal tip complex that regulates centriole growth and contributes to Drosophila spermiogenesis. Curr Biol, 33(19):4202-4216 PubMed ID: 37729913
Song, X., Cui, L., Wu, M., Wang, S., Song, Y., Liu, Z., Xue, Z., Chen, W., Zhang, Y., Li, H., Sun, L., Liang, X. (2023). DCX-EMAP is a core organizer for the ultrastructure of Drosophila mechanosensory organelles. J Cell Biol, 222(10). PubMed ID: 37651176
Palumbo, V., et al. (2015). Misato controls mitotic microtubule generation by stabilizing the tubulin chaperone protein-1 complex. Curr Biol 25: 1777-1783. PubMed ID: 26096973
Parton, R. M., Hamilton, R. S., Ball, G., Yang, L., Cullen, C. F., Lu, W., Ohkura, H. and Davis, I. (2011). A PAR-1-dependent orientation gradient of dynamic microtubules directs posterior cargo transport in the Drosophila oocyte. J Cell Biol 194(1): 121-135. PubMed ID: 21746854
Pietra, S., Ng, K., Lawrence, P. A. and Casal, J. (2020). Planar cell polarity in the larval epidermis of Drosophila and the role of microtubules. Open Biol 10(12): 200290. PubMed ID: 33295841
Pinheiro, E. M., Xie, Z., Norovich, A. L., Vidaki, M., Tsai, L. H. and Gertler, F. B. (2011). Lpd depletion reveals that SRF specifies radial versus tangential migration of pyramidal neurons. Nat Cell Biol 13: 989-995. PubMed ID: 21785421
Pinter, R., Huber, T., Bukovics, P., Gaszler, P., Vig, A. T., Toth, M., Gazso-Gerhat, G., Farkas, D., Migh, E., Mihaly, J. and Bugyi, B. (2020). The activities of the gelsolin homology domains of flightless-I in actin dynamics. Front Mol Biosci 7: 575077. PubMed ID: 33033719
Planelles-Herrero, V. J., Bittleston, A., Seum, C., Gaitan, M. G. and Derivery, E. (2022). Elongator stabilizes microtubules to control central spindle asymmetry and polarized trafficking of cell fate determinants. Nat Cell Biol 24(11): 1606-1616. PubMed ID: 36302967
Plochocka, A. Z., Ramirez Moreno, M., Davie, A. M., Bulgakova, N. A. and Chumakova, L. (2021). Robustness of the microtubule network self-organization in epithelia. Elife 10. PubMed ID: 33522481
Pollitt, S. L., Myers, K. R., Yoo, J. and Zheng, J. Q. (2020). LIM and SH3 Protein 1 Localizes to the Leading Edge of Protruding Lamellipodia and Regulates Axon Development. Mol Biol Cell: mbcE20060366. PubMed ID: 32997597
Poukkula, M., Hakala, M., Pentinmikko, N., Sweeney, M. O., Jansen, S., Mattila, J., Hietakangas, V., Goode, B. L. and Lappalainen, P. (2014). GMF promotes leading-edge dynamics and collective cell migration in vivo. Curr Biol 24(21):2533-40. PubMed ID: 25308079
Pradhan, R., Kumar, S. and Mathew, R. (2023). Lateral adherens junctions mediate a supracellular actomyosin cortex in Drosophila trachea. iScience 26(4): 106380. PubMed ID: 37009223
Qu, Y., Hahn, I., Lees, M., Parkin, J., Voelzmann, A., Dorey, K., Rathbone, A., Friel, C. T., Allan, V. J., Okenve-Ramos, P., Sanchez-Soriano, N. and Prokop, A. (2019). Efa6 protects axons and regulates their growth and branching by inhibiting microtubule polymerisation at the cortex. Elife 8. PubMed ID: 31718774
Renda, F., Pellacani, C., Strunov, A., Bucciarelli, E., Naim, V., Bosso, G., Kiseleva, E., Bonaccorsi, S., Sharp, D. J., Khodjakov, A., Gatti, M. and Somma, M. P. (2017). The Drosophila orthologue of the INT6 onco-protein regulates mitotic microtubule growth and kinetochore structure. PLoS Genet 13(5): e1006784. PubMed ID: 28505193
Repton, C., Cullen, C. F., Costa, M. F. A., Spanos, C., Rappsilber, J. and Ohkura, H. (2022). The phospho-docking protein 14-3-3 regulates microtubule-associated proteins in oocytes including the chromosomal passenger Borealin. PLoS Genet 18(6): e1009995. PubMed ID: 35666772
Riparbelli, M. G., Persico, V. and Callaini, G. (2020). The Microtubule Cytoskeleton during the Early Drosophila Spermiogenesis. Cells 9(12). PubMed ID: 33327573
Rogers, S. L., Wiedemann, U., Stuurman, N. and Vale, R. D. (2003). Molecular requirements for actin-based lamella formation in Drosophila S2 cells. J. Cell Biol. 162: 1079-1088. 12975351
Rothenberg, K. E., Chen, Y., McDonald, J. A. and Fernandez-Gonzalez, R. (2023). Rap1 coordinates cell-cell adhesion and cytoskeletal reorganization to drive collective cell migration in vivo. Curr Biol 33(13): 2587-2601. PubMed ID: 37244252
Salvany, L., Muller, J., Guccione, E. and Rorth, P. (2014). The core and conserved role of MAL is homeostatic regulation of actin levels. Genes Dev 28: 1048-1053. PubMed ID: 24831700
Saunders, H. A. J., Johnson-Schlitz, D. M., Jenkins, B. V., Volkert, P. J., Yang, S. Z. and Wildonger, J. (2022). Acetylated alpha-tubulin K394 regulates microtubule stability to shape the growth of axon terminals. Curr Biol. PubMed ID: 35081332
Sarkar, S., Olsen, A. L., Sygnecka, K., Lohr, K. M. and Feany, M. B. (2021). alpha-synuclein impairs autophagosome maturation through abnormal actin stabilization. PLoS Genet 17(2): e1009359. PubMed ID: 33556113
Schroeder, C. M., Tomlin, S. A., Mejia Natividad, I., Valenzuela, J. R., Young, J. M. and Malik, H. S. (2021). An actin-related protein that is most highly expressed in Drosophila testes is critical for embryonic development. Elife 10. PubMed ID: 34282725
Sengupta, S., Rath, U., Yao, C., Zavortink, M., Wang, C., Girton, J., Johansen, K.M. and Johansen, J. (2016). Digitor/dASCIZ has multiple roles in Drosophila development. PLoS One 11: e0166829. PubMed ID: 27861562
Sharma, M., Jiang, T., Jiang, Z. C., Moguel-Lehmer, C. E. and Harris, T. J. (2021). Emergence of a smooth interface from growth of a dendritic network against a mechanosensitive contractile material. Elife 10. PubMed ID: 34423780
Shi, Q., Lin, Y. Q., Saliba, A., Xie, J., Neely, G. G. and Banerjee, S. (2019). Tubulin polymerization promoting protein, Ringmaker, and MAP1B homolog Futsch coordinate microtubule organization and synaptic growth. Front Cell Neurosci 13: 192. PubMed ID: 31156389
Singh, A., Saha, T., Begemann, I., Ricker, A., Nusse, H., Thorn-Seshold, O., Klingauf, J., Galic, M. and Matis, M. (2018). Polarized microtubule dynamics directs cell mechanics and coordinates forces during epithelial morphogenesis. Nat Cell Biol 20(10): 1126-1133. PubMed ID: 30202051
Somogyi, K. and Rorth, P. (2004). Evidence for tension-based regulation of Drosophila MAL and SRF during invasive cell migration. Dev Cell 7: 85-93. PubMed ID: 15239956
Spencer, A. K., Schaumberg, A. J. and Zallen, J. A. (2017). Scaling of cytoskeletal organization with cell size in Drosophila. Mol Biol Cell 28(11):1519-1529. PubMed ID: 28404752
Sui, L., Alt, S., Weigert, M., Dye, N., Eaton, S., Jug, F., Myers, E. W., Julicher, F., Salbreux, G. and Dahmann, C. (2018). Differential lateral and basal tension drive folding of Drosophila wing discs through two distinct mechanisms. Nat Commun 9(1): 4620. PubMed ID: 30397306
Sun, L., Cui, L., Liu, Z., Wang, Q., Xue, Z., Wu, M., Sun, T., Mao, D., Ni, J., Pastor-Pareja, J. C., Liang, X. (2021). Katanin p60-like 1 sculpts the cytoskeleton in mechanosensory cilia. J Cell Biol, 220(1) PubMed ID: 33263729
Sysoev, V. O., Kato, M., Sutherland, L., Hu, R., McKnight, S. L. and Murray, D. T. (2020). Dynamic structural order of a low-complexity domain facilitates assembly of intermediate filaments. Proc Natl Acad Sci U S A 117(38): 23510-23518. PubMed ID: 32907935
Tariq, A., Green, L., Jeynes, J. C. G., Soeller, C. and Wakefield, J. G. (2020). In vitro reconstitution of branching microtubule nucleation. Elife 9. PubMed ID: 31933481
Theurkauf, W. E. (1994). Premature microtubule-dependent cytoplasmic streaming in cappuccino and spire mutant oocytes. Science 265: 2093-96. 8091233
Thyagarajan, P., Feng, C., Lee, D., Shorey, M. and Rolls, M. M. (2022). Microtubule polarity is instructive for many aspects of neuronal polarity. Dev Biol 486: 56-70. PubMed ID: 35341730
Tokamov, S. A., Nouri, N., Rich, A., Buiter, S., Glotzer, M., Fehon, R. G. (2023). Apical polarity and actomyosin dynamics control Kibra subcellular localization and function in Drosophila Hippo signaling. Dev Cell, 58(19): 1864-1879 PubMed ID: 37729921>
Veltman, D. M. and Insall, R. H. (2010). WASP family proteins: their evolution and its physiological implications. Mol Biol Cell 21: 2880-2893. PubMed ID: 20573979
Ventura Santos, C., Rogers, S. L., Carter, A. P. (2023). CryoET shows cofilactin filaments inside the microtubule lumen. EMBO reports, 24(11):e57264 PubMed ID: 37702953
Verma, V. and Maresca, T. J. (2019). Direct observation of branching MT nucleation in living animal cells. J Cell Biol. PubMed ID: 31340987
Villars, A., Matamoro-Vidal, A., Levillayer, F. and Levayer, R. (2022). Microtubule disassembly by caspases is an important rate-limiting step of cell extrusion. Nat Commun 13(1): 3632. PubMed ID: 35752632
Voelzmann, A., Okenve-Ramos, P., Qu, Y., Chojnowska-Monga, M., Del Caño-Espinel, M., Prokop, A. and Sanchez-Soriano, N. (2016). Tau and spectraplakins promote synapse formation and maintenance through Jun kinase and neuronal trafficking. Elife 5. PubMed ID: 27501441
Walker, D., Schmid, E., Schinaman, J. and Williams, K. (2023). Accumulation of F-actin drives brain aging and limits healthspan in Drosophila. Res Sq. PubMed ID: 37577708
Walker, R. A. and Sheetz, M. P. (1993). Cytoplasmic microtubule-associated motors. Annu. Rev. Biochem. 62: 429-451
Weiner, A. T., Seebold, D. Y., Torres-Gutierrez, P., Folker, C., Swope, R. D., Kothe, G. O., Stoltz, J. G., Zalenski, M. K., Kozlowski, C., Barbera, D. J., Patel, M. A., Thyagarajan, P., Shorey, M., Nye, D. M. R., Keegan, M., Behari, K., Song, S., Axelrod, J. D. and Rolls, M. M. (2020). Endosomal Wnt signaling proteins control microtubule nucleation in dendrites. PLoS Biol 18(3): e3000647. PubMed ID: 32163403
Wippich, F., Vaishali, Hennrich, M. L. and Ephrussi, A. (2023). Nutritional stress-induced regulation of microtubule organization and mRNP transport by HDAC1 controlled α-tubulin acetylation. Commun Biol 6(1): 776. PubMed ID: 37491525
Xie, P. (2023). Determinant factors for residence time of kinesin motors at microtubule ends. J Biol Phys. PubMed ID: 36645568
Ye, A. A., Verma, V. and Maresca, T. J. (2018). NOD is a plus end-directed motor
that binds EB1 via a new microtubule tip localization sequence. J
Cell Biol. PubMed ID: 29899040
Yolland, L., Burki, M., Marcotti, S., Luchici, A., Kenny, F. N., Davis, J. R., Serna-Morales, E., Muller, J., Sixt, M., Davidson, A., Wood, W., Schumacher, L. J., Endres, R. G., Miodownik, M. and Stramer, B. M. (2019). Persistent and polarized global actin flow is essential for directionality during cell migration. Nat Cell Biol 21(11): 1370-1381. PubMed ID: 31685997
Zajac, A. L. and Horne-Badovinac, S. (2022). Kinesin-directed secretion of basement membrane proteins to a subdomain of the basolateral surface in Drosophila epithelial cells. Curr Biol. PubMed ID: 35021047
Zapater, I. M. C., Carman, P. J., Soffar, D. B., Windner, S. E., Dominguez, R. and Baylies, M. K. (2023). Drosophila Tropomodulin is required for multiple actin-dependent processes within developing myofibers. Development 150(6). PubMed ID: 36806912
Zhao, A. J., Montes-Laing, J., Perry, W. M. G., Shiratori, M., Merfeld, E., Rogers, S. L. and Applewhite, D. A. (2022). The Drosophila spectraplakin Short stop regulates focal adhesion dynamics by cross-linking microtubules and actin. Mol Biol Cell 33(5): ar19. PubMed ID: 35235367
Zhao, G., Oztan, A., Ye, Y. and Schwarz, T. L. (2019). Kinetochore proteins have a post-mitotic function in neurodevelopment. Dev Cell. PubMed ID: 30827899
list of genes involved in cytoskeleton Home page: The Interactive Fly © 2025 Thomas B. Brody, Ph.D.
The Interactive Fly resides on theActin cytoskeleton and associated proteins
Microtubular cytoskeleton and associated proteins
Spectrin Cytoskeleton
Genes affecting the cytoskeleton
Society for Developmental Biology's Web server.
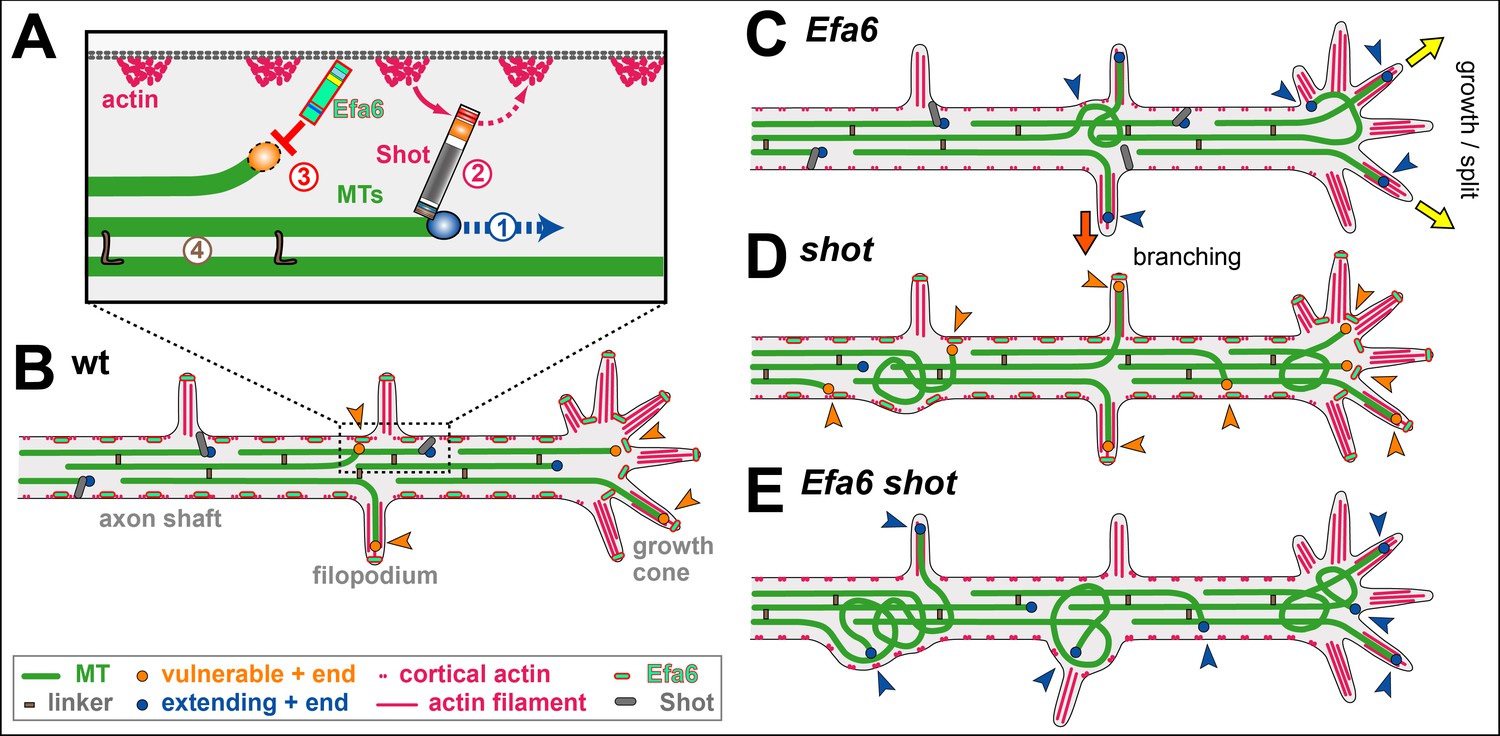{kind=link}