|
Gene name - Abdominal-B Synonyms - Cytological map position - 89E2-5 Function - transcription factor Keywords - bithorax-complex, homeotic protein |
Symbol - Abd-B FlyBase ID: FBgn0000015 Genetic map position - 3-58.8 Classification - homeodomain Cellular location - nuclear |
| Recent literature | Kubo, A., Matsuka, M., Minami, R., Kimura, F., Sakata-Niitsu, R., Kokuryo, A., Taniguchi, K., Adachi-Yamada, T. and Nakagoshi, H. (2018). Nutrient conditions sensed by the reproductive organ during development optimize male fecundity in Drosophila. Genes Cells. PubMed ID: 29846027
Summary: Nutrient conditions affect the reproductive potential and lifespan of many organisms through the insulin signaling pathway. Although this is well characterized in female oogenesis, nutrient-dependent regulation of fertility/fecundity in males is not known. Seminal fluid components synthesized in the accessory gland are required for high fecundity in Drosophila males. The accessory gland is composed of two types of binucleated cells: a main cell and a secondary cell (SC). The transcription factors Defective proventriculus (Dve) and Abdominal-B (Abd-B) are strongly expressed in adult SCs, whose functions are essential for male fecundity. Gene expression of both Dve and Abd-B was down-regulated under nutrient-poor conditions. In addition, nutrient conditions during the pupal stage affected the size and number of SCs. These morphological changes clearly correlated with fecundity, suggesting that SCs act as nutrient sensors. This study provides evidence that Dve associates nutrient conditions with optimal reproductive potential in a target of rapamycin signaling-dependent manner. |
Ghosh, N., Bakshi, A., Khandelwal, R., Rajan, S. G. and Joshi, R. (2019). Hox gene Abdominal-B uses Doublesex(F) as a cofactor to promote neuroblast apoptosis in Drosophila central nervous system. Development. PubMed ID: 31371379
Summary: Highly conserved DM domain containing transcription factors (Doublesex/MAB-3/DMRT1) are responsible for generating sexually dimorphic features. In Drosophila CNS a set of Doublesex (Dsx) expressing neuroblasts undergo apoptosis in females while their male counterparts proliferate and give rise to serotonergic neurons crucial for adult mating behaviour. This study study demonstrates that female specific isoform of Doublesex collaborates with Hox gene Abdominal-B (AbdB) to bring about this apoptosis. Biochemical results suggest AbdB and Dsx interact through their highly conserved Homeodomain and DM domains respectively. This interaction is translated into a cooperative binding of the two proteins (AbdB and Dsx) on the apoptotic enhancer in case of females but not in case of males, resulting in female specific activation of apoptotic genes. The capacity of AbdB to utilize sex specific isoform of Dsx as a cofactor underlines the possibility that two classes of proteins are capable of cooperating in selection and regulation of target genes in tissue and sex specific manner. It is proposed that this interaction could be a common theme in generating sexual dimorphism in different tissues across different species. |
Liu, Y., Ramos-Womack, M., Han, C., Reilly, P., Brackett, K. L., Rogers, W., Williams, T. M., Andolfatto, P., Stern, D. L. and Rebeiz, M. (2019). Changes throughout a genetic network mask the contribution of Hox gene evolution. Curr Biol 29(13): 2157-2166. PubMed ID: 31257142
Summary: Hox genes pattern the anterior-posterior axis of animals and are posited to drive animal body plan evolution, yet their precise role in evolution has been difficult to determine. This study identified evolutionary modifications in the Hox gene Abd-B that dramatically altered its expression along the body plan of Drosophila santomea. Abd-B is required for pigmentation in Drosophila yakuba, the sister species of D. santomea, and changes to Abd-B expression would be predicted to make large contributions to the loss of body pigmentation in D. santomea. However, manipulating Abd-B expression in current-day D. santomea does not affect pigmentation. This is attributed to epistatic interaction to four other genes within the D. santomea pigmentation network, three of which have evolved expression patterns that do not respond to Abd-B. These results demonstrate how body plans may evolve through small evolutionary steps distributed throughout Hox-regulated networks. Polygenicity and epistasis may hinder efforts to identify genes and mechanisms underlying macroevolutionary traits. |
Romero-Pozuelo, J., Foronda, D., Martin, P., Hudry, B., Merabet, S., Graba, Y. and Sanchez-Herrero, E. (2019). Cooperation of axial and sex specific information controls Drosophila female genitalia growth by regulating the Decapentaplegic pathway. Dev Biol. PubMed ID: 31251896
Summary: The specification and morphogenesis of an organ requires the coordinate deployment and integration of regulatory information, including sex specific information when the organ is sex specific. Only a few gene networks controlling size and pattern development have been deciphered, which limits the emergence of principles, general or not, underlying the organ-specifying gene networks. This study elucidates the genetic and molecular network determining the control of size in the Drosophila abdominal A9 primordium, contributing to the female genitalia. This network requires axial regulatory information provided by the Hox protein Abdominal-BR (Abd-BR), the Hox cofactors Extradenticle (Exd) and Homothorax (Hth), and the sex specific transcription factor Doublesex Female (DsxF). These factors synergize to control size in the female A9 by the coordinate regulation of the Decapentaplegic (Dpp) growth pathway. Molecular dissection of the dpp regulatory region and in vivo protein interaction experiments suggest that Abd-BR, Exd, Hth and DsxF coordinately regulate a short dpp enhancer to repress dpp expression and restrict female A9 size. The same regulators can also suppress dpp expression in the A8, but this requires the absence of the Abd-BM isoform, which specifies A8. These results delineate the network controlling female A9 growth in Drosophila. |
Kyrchanova, O., Wolle, D., Sabirov, M., Kurbidaeva, A., Aoki, T., Maksimenko, O., Kyrchanova, M., Georgiev, P. and Schedl, P. (2019). Distinct elements confer the blocking and bypass functions of the Bithorax Fab-8 boundary. Genetics. PubMed ID: 31551239
Summary: Boundaries in the Drosophila bithorax complex (BX-C) enable the regulatory domains that drive parasegment specific expression of the three Hox genes to function autonomously. The four regulatory domains (iab-5, iab-6, iab-7 and iab-8) that control the expression of the Abdominal-B (Abd-B) gene are located downstream of the transcription unit and are delimited by the Mcp, Fab-6, Fab-7 and Fab-8 boundaries. These boundaries function to block crosstalk between neighboring regulatory domains. In addition, three of the boundaries (Fab-6, Fab-7 and Fab-8) must also have bypass activity so that regulatory domains distal to the boundaries can contact the Abd-B promoter. In these studies a functional dissection was undertaken of the Fab-8 boundary using a boundary replacement strategy. The studies indicate that the Fab-8 boundary has two separable sub-elements. The distal sub-element blocks crosstalk, but can not support bypass. The proximal sub-element has only minimal blocking activity but is able to mediate bypass. A large multiprotein complex, the LBC, binds to sequences in the proximal sub-element and contributes to its bypass activity. The same LBC complex has been implicated in the bypass activity of the Fab-7 boundary. |
Bakshi, A., Sipani, R., Ghosh, N. and Joshi, R. (2020). Sequential activation of Notch and Grainyhead gives apoptotic competence to Abdominal-B expressing larval neuroblasts in Drosophila Central nervous system. PLoS Genet 16(8): e1008976. PubMed ID: 32866141
Summary: Neural circuitry for mating and reproduction resides within the terminal segments of central nervous system (CNS) which express Hox paralogous group 9-13 (in vertebrates) or Abdominal-B (Abd-B) in Drosophila. Terminal neuroblasts (NBs) in A8-A10 segments of Drosophila larval CNS are subdivided into two groups based on expression of transcription factor Doublesex (Dsx). While the sex specific fate of Dsx-positive NBs is well investigated, the fate of Dsx-negative NBs is not known so far. Studies with Dsx-negative NBs suggests that these cells, like their abdominal counterparts (in A3-A7 segments) use Hox, Grainyhead (Grh) and Notch to undergo cell death during larval development. This cell death also happens by transcriptionally activating RHG family of apoptotic genes through a common apoptotic enhancer in early to mid L3 stages. However, unlike abdominal NBs (in A3-A7 segments) which use increasing levels of resident Hox factor Abdominal-A (Abd-A) as an apoptosis trigger, Dsx-negative NBs (in A8-A10 segments) keep the levels of resident Hox factor Abd-B constant. These cells instead utilize increasing levels of the temporal transcription factor Grh and a rise in Notch activity to gain apoptotic competence. Biochemical and in vivo analysis suggest that Abdominal-A and Grh binding motifs in the common apoptotic enhancer also function as Abdominal-B and Grh binding motifs and maintains the enhancer activity in A8-A10 NBs. Finally, the deletion of this enhancer by the CRISPR-Cas9 method blocks the apoptosis of Dsx-negative NBs. These results highlight the fact that Hox dependent NB apoptosis in abdominal and terminal regions utilizes common molecular players (Hox, Grh and Notch), but seems to have evolved different molecular strategies to pattern CNS. |
Postika, N., Schedl, P., Georgiev, P. and Kyrchanova, O. (2021). Redundant enhancers in the iab-5 domain cooperatively activate Abd-B in the A5 and A6 abdominal segments of Drosophila. Development. PubMed ID: 34473267
Summary: The Abdominal-B (Abd-B) gene belongs to Bithorax complex and its expression is controlled by four regulatory domains, iab-5, iab-6, iab-7 and iab-8, each of which is thought to be responsible for directing the expression of Abd-B in one of the abdominal segments from A5 to A8. A variety of experiments have supported the idea that BX-C regulatory domains are functionally autonomous and that each domain is both necessary and sufficient to orchestrate the development of the segment they specify. Unexpectedly, this study discovered that this model does not always hold. Instead, it was found that tissue-specific enhancers located in the iab-5 domain are required for the proper activation of Abd-B not only in A5 but also in A6. These findings indicate that the functioning of the iab-5 and iab-6 domains in development of the adult cuticle A5 and A6 in males fit better with an additive model much like that first envisioned by Ed Lewis. |
Pu, J., Wang, Z., Cong, H., Chin, J. S. R., Justen, J., Finet, C., Yew, J. Y. and Chung, H. (2021). Repression precedes independent evolutionary gains of a highly specific gene expression pattern. Cell Rep 37(4): 109896. PubMed ID: 34706247 Summary: Highly specific expression patterns can be caused by the overlapping activities of activator and repressor sequences in enhancers. However, few studies illuminate how these sequences evolve in the origin of new enhancers. This study shows that expression of the bond gene in the semicircular wall epithelium (swe) of the Drosophila melanogaster male ejaculatory bulb (EB) is controlled by an enhancer consisting of an activator region that requires Abdominal-B driving expression in the entire EB and a repressor region that restricts this expression to the EB swe. Although this expression pattern is independently gained in the distantly related Scaptodrosophila lebanonensis and does not require Abdominal-B, this study shows that functionally similar repressor sequences are present in Scaptodrosophila and also in species that do not express bond in the EB. It is suggested that during enhancer evolution, repressor sequences can precede the evolution of activator sequences and may lead to similar but independently evolved expression patterns. |
Hafer, T. L., Patra, S., Tagami, D. and Kohwi, M. (2022). Enhancer of trithorax/polycomb, Corto, regulates timing of hunchback gene relocation and competence in Drosophila neuroblasts. Neural Dev 17(1): 3. PubMed ID: 35177098
Summary: Neural progenitors produce diverse cells in a stereotyped birth order, but can specify each cell type for only a limited duration. In the Drosophila embryo, neuroblasts (neural progenitors) specify multiple, distinct neurons by sequentially expressing a series of temporal identity transcription factors with each division. Hunchback (Hb), the first of the series, specifies early-born neuronal identity. Neuroblast competence to generate early-born neurons is terminated when the hb gene relocates to the neuroblast nuclear lamina, rendering it refractory to activation in descendent neurons. Mechanisms and trans-acting factors underlying this process are poorly understood. This study identified Corto, an enhancer of Trithorax/Polycomb (ETP) protein, as a new regulator of neuroblast competence. The GAL4/UAS system was used to drive persistent misexpression of Hb in neuroblast 7-1 (NB7-1), a model lineage for which the early competence window has been well characterized, to examine the role of Corto in neuroblast competence. immuno-DNA Fluorescence in situ hybridization (DNA FISH) was used in whole embryos to track the position of the hb gene locus specifically in neuroblasts across developmental time, comparing corto mutants to control embryos. Finally, immunostaining was used in whole embryos to examine Corto's role in repression of Hb and a known target gene, Abdominal B (Abd-B). In corto mutants, the hb gene relocation to the neuroblast nuclear lamina was found to be delayed and the early competence window is extended. The delay in gene relocation occurs after hb transcription is already terminated in the neuroblast and is not due to prolonged transcriptional activity. Further, it was found that Corto genetically interacts with Posterior Sex Combs (Psc), a core subunit of polycomb group complex 1 (PRC1), to terminate early competence. Loss of Corto does not result in derepression of Hb or its Hox target, Abd-B, specifically in neuroblasts. These results show that in neuroblasts, Corto genetically interacts with PRC1 to regulate timing of nuclear architecture reorganization and support the model that distinct mechanisms of silencing are implemented in a step-wise fashion during development to regulate cell fate gene expression in neuronal progeny. |
Clarembaux-Badell, L., Baladron-de-Juan, P., Gabilondo, H., Rubio-Ferrera, I., Millan, I., Estella, C., Valverde-Ortega, F. S., Cobeta, I. M., Thor, S. and Benito-Sipos, J. (2022). Dachshund acts with Abdominal-B to trigger programmed cell death in the Drosophila central nervous system at the frontiers of Abd-B expression. Dev Neurobiol 82(6): 495-504. PubMed ID: 35796156
Summary: A striking feature of the nervous system pertains to the appearance of different neural cell subtypes at different axial levels. Studies in the Drosophila central nervous system reveal that one mechanism underlying such segmental differences pertains to the segment-specific removal of cells by programmed cell death (PCD). One group of genes involved in segment-specific PCD is the Hox homeotic genes. However, while segment-specific PCD is highly precise, Hox gene expression is evident in gradients, raising the issue of how the Hox gene function is precisely gated to trigger PCD in specific segments at the outer limits of Hox expression. The Drosophila Va neurons are initially generated in all nerve cord segments but removed by PCD in posterior segments. Va PCD is triggered by the posteriorly expressed Hox gene Abdominal-B (Abd-B). However, Va PCD is highly reproducible despite exceedingly weak Abd-B expression in the anterior frontiers of its expression. This study found that the transcriptional cofactor Dachshund supports Abd-B-mediated PCD in its anterior domain. In vivo bimolecular fluorescence complementation analysis lends support to the idea that the Dachshund/Abd-B interplay may involve physical interactions. These findings provide an example of how combinatorial codes of transcription factors ensure precision in Hox-mediated PCD in specific segments at the outer limits of Hox expression. |
Bu, S., Lau, S. S. Y., Yong, W. L., Zhang, H., Thiagarajan, S., Bashirullah, A. and Yu, F. (2023). Polycomb group genes are required for neuronal pruning in Drosophila. BMC Biol 21(1): 33. PubMed ID: 36793038
Summary: Pruning that selectively eliminates unnecessary or incorrect neurites is required for proper wiring of the mature nervous system. During Drosophila metamorphosis, dendritic arbourization sensory neurons (ddaCs) and mushroom body (MB) γ neurons can selectively prune their larval dendrites and/or axons in response to the steroid hormone ecdysone. An ecdysone-induced transcriptional cascade plays a key role in initiating neuronal pruning. However, how downstream components of ecdysone signalling are induced remains not entirely understood. This study identified that Scm, a component of Polycomb group (PcG) complexes, is required for dendrite pruning of ddaC neurons. Two PcG complexes, PRC1 and PRC2, are important for dendrite pruning. Interestingly, depletion of PRC1 strongly enhances ectopic expression of Abdominal B (Abd-B) and Sex combs reduced, whereas loss of PRC2 causes mild upregulation of Ultrabithorax and Abdominal A in ddaC neurons. Among these Hox genes, overexpression of Abd-B causes the most severe pruning defects, suggesting its dominant effect. Knockdown of the core PRC1 component Polyhomeotic (Ph) or Abd-B overexpression selectively downregulates Mical expression, thereby inhibiting ecdysone signalling. Finally, Ph is also required for axon pruning and Abd-B silencing in MB γ neurons, indicating a conserved function of PRC1 in two types of pruning. This study demonstrates important roles of PcG and Hox genes in regulating ecdysone signalling and neuronal pruning in Drosophila. Moreover, our findings suggest a non-canonical and PRC2-independent role of PRC1 in Hox gene silencing during neuronal pruning. |
Schroeder, A. M., Nielsen, T., Lynott, M., Vogler, G., Colas, A. R. and Bodmer, R. (2022). Nascent polypeptide-Associated Complex and Signal Recognition Particle have cardiac-specific roles in heart development and remodeling. PLoS Genet 18(10): e1010448. PubMed ID: 36240221
Summary: Establishing a catalog of Congenital Heart Disease (CHD) genes and identifying functional networks would improve understanding of its oligogenic underpinnings. The current studies identified protein biogenesis cofactors Nascent polypeptide-Associated Complex (NAC) and Signal-Recognition-Particle (SRP) as disease candidates and novel regulators of cardiac differentiation and morphogenesis. Knockdown (KD) of the α-subunit (Nacα) or beta-subunit (bicaudal, bic) of NAC in the developing Drosophila heart disrupted cardiac developmental remodeling resulting in a fly with no heart. Heart loss was rescued by combined KD of Nacα with the posterior patterning Hox gene Abd-B. Consistent with a central role for this interaction in cardiogenesis, KD of Nacα in cardiac progenitors derived from human iPSCs impaired cardiac differentiation while co-KD with human HOXC12 and HOXD12 rescued this phenotype. These data suggest that Nacα KD preprograms cardioblasts in the embryo for abortive remodeling later during metamorphosis, as Nacα KD during translation-intensive larval growth or pupal remodeling only causes moderate heart defects. KD of SRP subunits in the developing fly heart produced phenotypes that targeted specific segments and cell types, again suggesting cardiac-specific and spatially regulated activities. Together, this study demonstrated directed function for NAC and SRP in heart development, and that regulation of NAC function depends on Hox genes. |
Moniot-Perron, L., Moindrot, B., Manceau, L., Edouard, J., Jaszczyszyn, Y., Gilardi-Hebenstreit, P., Hernandez, C., Bloyer, S. and Noordermeer, D. (2023). The Drosophila Fab-7 boundary modulates Abd-B gene activity by guiding an inversion of collinear chromatin organization and alternate promoter use. Cell Rep 42(1): 111967. PubMed ID: 36640345
Summary: Hox genes encode transcription factors that specify segmental identities along the anteroposterior body axis. These genes are organized in clusters, where their order corresponds to their activity along the body axis, a feature known as collinearity. In Drosophila, the BX-C cluster contains the three most posterior Hox genes, where their collinear activation incorporates progressive changes in histone modifications, chromatin architecture, and use of boundary elements and cis-regulatory regions. To dissect functional hierarchies, this study compareed chromatin organization in cell lines and larvae, with a focus on the Abd-B gene. This work establishes the importance of the Fab-7 boundary for insulation between 3D domains carrying different histone modifications. Interestingly, a non-canonical inversion of collinear chromatin dynamics was detected at Abd-B, with the domain of active histone modifications progressively decreasing in size. This dynamic chromatin organization differentially activates the alternative promoters of the Abd-B gene, thereby expanding the possibilities for fine-tuning of transcriptional output. |
Hajirnis, N., Pandey, S. and Mishra, R. K. (2023). CRISPR/Cas9 and FLP-FRT mediated regulatory dissection of the BX-C of Drosophila melanogaster. Chromosome Res 31(1): 7. PubMed ID: 36719476
Summary: The homeotic genes or Hox define the anterior-posterior (AP) body axis formation in bilaterians and are often present on the chromosome in an order collinear to their function across the AP axis. However, there are many cases wherein the Hox are not collinear, but their expression pattern is conserved across the AP axis. The expression pattern of Hox is attributed to the cis-regulatory modules (CRMs) consisting of enhancers, initiators, or repressor elements that regulate the genes in a segment-specific manner. In the Drosophila melanogaster Hox complex, the bithorax complex (BX-C) and even the CRMs are organized in an order that is collinear to their function in the thoracic and abdominal segments. In the present study, the regulatorily inert regions were targeted using CRISPR/Cas9 to generate a series of transgenic lines with the insertion of FRT sequences. These FRT lines are repurposed to shuffle the CRMs associated with Abd-B to generate modular deletion, duplication, or inversion of multiple CRMs. The rearrangements yielded entirely novel phenotypes in the fly suggesting the requirement of such complex manipulations to address the significance of higher order arrangement of the CRMs. The functional map and the transgenic flies generated in this study are important resources to decipher the collective ability of multiple regulatory elements in the eukaryotic genome to function as complex modules. |
Kyrchanova, O., Ibragimov, A., Postika, N., Georgiev, P. and Schedl, P. (2023). Boundary bypass activity in the abdominal-B region of the Drosophila bithorax complex is position dependent and regulated. Open Biol 13(8): 230035. PubMed ID: 37582404
Summary: Expression of Abdominal-B (Abd-B) in abdominal segments A5-A8 is controlled by four regulatory domains, iab-5-iab-8. Each domain has an initiator element (which sets the activity state), elements that maintain this state and tissue-specific enhancers. To ensure their functional autonomy, each domain is bracketed by boundary elements (Mcp, Fab-7, Fab-7 and Fab-8). In addition to blocking crosstalk between adjacent regulatory domains, the Fab boundaries must also have bypass activity so the relevant regulatory domains can 'jump over' intervening boundaries and activate the Abd-B promoter. This study investigated the parameters governing bypass activity. The bypass elements in the Fab-7 and Fab-8 boundaries must be located in the regulatory domain that is responsible for driving Abd-B expression. It is suggested that bypass activity may also be subject to regulation. |
Hodkinson, L. J., Smith, C., Comstra, H. S., ...., Zeng, H., Schmidt, C. A., Rieder, L. E. (2023). A bioinformatics screen reveals hox and chromatin remodeling factors at the Drosophila histone locus. BMC genomic data, 24(1):54 PubMed ID: 37735352
Summary: Cells orchestrate histone biogenesis with strict temporal and quantitative control. To efficiently regulate histone biogenesis, the repetitive Drosophila melanogaster replication-dependent histone genes are arrayed and clustered at a single locus. Regulatory factors concentrate in a nuclear body known as the histone locus body (HLB), which forms around the locus. Historically, HLB factors are largely discovered by chance, and few are known to interact directly with DNA. It is therefore unclear how the histone genes are specifically targeted for unique and coordinated regulation. RESULTS: To expand the list of known HLB factors, we performed a candidate-based screen by mapping 30 publicly available ChIP datasets of 27 unique factors to the Drosophila histone gene array. We identified novel transcription factor candidates, including the Drosophila Hox proteins Ultrabithorax (Ubx), Abdominal-A (Abd-A), and Abdominal-B (Abd-B), suggesting a new pathway for these factors in influencing body plan morphogenesis. Additionally, six other factors were identified that target the histone gene array: JIL-1, hormone-like receptor 78 (Hr78), the long isoform of female sterile homeotic (1) (fs(1)h) as well as the general transcription factors TBP associated factor 1 (TAF-1), Transcription Factor IIB (TFIIB), and Transcription Factor IIF (TFIIF). This foundational screen provides several candidates for future studies into factors that may influence histone biogenesis. Further, this study emphasizes the powerful reservoir of publicly available datasets, which can be mined as a primary screening technique. |
Matsuka, M., Otsune, S., Sugimori, S., Tsugita, Y., Ueda, H., Nakagoshi, H. (2024). Fecundity is optimized by levels of nutrient signal-dependent expression of Dve and EcR in Drosophila male accessory gland. Dev Biol, 508:8-23 PubMed ID: 38199580
Summary: Steroid hormones play various physiological roles including metabolism and reproduction. Steroid hormones in insects are ecdysteroids, and the major form in Drosophila melanogaster is ecdysone. In Drosophila males, the accessory gland is responsive to nutrient-dependent regulation of fertility/fecundity. The accessory gland is composed of two types of binucleated epithelial cells: a main cell and a secondary cell (SC). The transcription factors Defective proventriculus (Dve), Abdominal-B, and Ecdysone receptors (EcRs) are strongly expressed in adult SCs. This EcR expression is regulated by parallel pathways of nutrient signaling and the Dve activity. Induction of Dve expression is also dependent on nutrient signaling, and it becomes nutrient signal-independent during a restricted period of development. Forced dve expression during the restricted period significantly increased the number of SCs. This study provides evidence that the level of nutrient signal-dependent Dve expression during the restricted period determines the number of SCs, and that ecdysone signaling is also crucial to optimize male fecundity through nutrient signal-dependent survival and maturation of SCs. |
Hemba-Waduge, R. U., Liu, M., Li, X., Sun, J. L., Budslick, E. A., Bondos, S. E., Ji, J. Y. (2024). Metabolic control by the Bithorax Complex-Wnt signaling crosstalk in Drosophila. bioRxiv, PubMed ID: 38853890
Summary: Adipocytes distributed throughout the body play crucial roles in lipid metabolism and energy homeostasis. Regional differences among adipocytes influence normal function and disease susceptibility, but the mechanisms driving this regional heterogeneity remain poorly understood. This study reports a genetic crosstalk between the Bithorax Complex ( BX-C ) genes and Wnt/Wingless signaling that orchestrates regional differences among adipocytes in Drosophila larvae. Abdominal adipocytes, characterized by the exclusive expression of abdominal A ( abd-A )and Abdominal B ( Abd-B ), exhibit distinct features compared to thoracic adipocytes, with Wnt signaling further amplifying these disparities. Depletion of BX-C genes in adipocytes reduces fat accumulation, delays larval-pupal transition, and eventually leads to pupal lethality. Depleting Abd-A or Abd-B reduces Wnt target gene expression, thereby attenuating Wnt signaling-induced lipid mobilization. Conversely, Wnt signaling stimulated abd-A transcription, suggesting a feedforward loop that amplifies the interplay between Wnt signaling and BX-C in adipocytes. These findings elucidate how the crosstalk between cell-autonomous BX-C gene expression and Wnt signaling define unique metabolic behaviors in adipocytes in different anatomical regions of fat body, delineating larval adipose tissue domains. |
Kyrchanova, O., Kudryashova, K., Ibragimov, A., Dubrovskaya, V., Schedl, P., Georgiev, P. (2025). Embryonic enhancers help transmit positional information to the initiator cores that control Drosophila Abd-B regulatory domains. Development, 152(19) PubMed ID: 41020347
Summary: Drosophila homeotic gene Abdominal-B (Abd-B) is controlled throughout development by four infraabdominal (iab) regulatory domains, the active or repressed state of which is determined by initiators that have parasegment-specific enhancer activity at an early stage of embryonic development. For this reason, it has long been assumed that the enhancer activity and initiation function of these elements are synonymous. This study examined two initiators that regulate the activity of the iab-5 and iab-6 domains responsible for Abd-B expression in embryonic parasegment PS10 (adult segment A5) and PS11(A6), respectively. In both initiators, core regions were identified that do not stimulate reporter gene transcription, but retain the ability to establish the appropriate activity state of the corresponding iab domains. Other initiator sequences are responsible for parasegment-specific reporter activation in early embryos and enhance the activity of the core initiators. Taken together, these results indicate that initiators represent a new type of regulatory element that function as on/off switches for regulatory domains controlling segment-specific Abd-B expression during Drosophila development. |
Li, J., Taniguchi, K., Ye, W., Kondo, S., Kobayashi, T., Matsuyama, M., Saito, K., Ohsawa, S., Igaki, T. (2025). Epithelial cell-fate switch triggering ectopic ligand-receptor-mediated JAK-STAT signaling promotes tumorigenesis in Drosophila. iScience, 28(4):112191 PubMed ID: 40230533
Summary: Disruption of epithelial architecture is a hallmark of human malignant cancers, yet whether and how epithelial deformation influences tumor progression has been elusive. Through a genetic screen in Drosophila eye disc, this study explored mutations that potently promoted Ras-activated (Ras(V12)) tumor growth and identified eyes absent (eya), an eye determination gene, whose mutation compromised tissue growth but synergized with Ras(V12) to cause massive overgrowth. Furthermore, induction of cell-fate switch by mis-expression of Abd-B in the eye disc also induced massive Ras(V12) overgrowth. Mechanistically, cell-fate switch caused epithelial invagination accompanied by partial mislocalization of the transmembrane receptor Domeless (Dome) from the apical to the basal membrane of the eye epithelium, where its ligand Unpaired3 (Upd3) is present. This led to JAK-STAT activation that cooperates with Ras(V12) to drive tumor progression. These data provide a mechanistic explanation for how cell-fate switch and subsequent epithelial deformation creates a cancer-prone environment in the epithelium. |
Pair-rule and segment polarity genes are responsible for determining the uniformity of different segments, in contrast to homeodomain proteins that are responsible for establishing the diversity between segments. Abdominal-B acts in three germ cell layers to fulfill this latter function.
Abdominal-B is the last in linkage order and the most posterior acting of the linked homeodomain proteins of the bithorax. Abdominal-B is unique among the homeotics in that it is transcribed in two forms; a regulatory (r) protein and a morphogenic (m) protein. Regulatory transcripts of Abdominal-B act as repressors, suppressing embryonic ventral epidermal structures in the 8th and 9th segments of the abdomen. Thus ABD-B r and m proteins are critically involved in establishing cell fate in the tail segments of the fly.
Expression is driven by two promoters. The m form is transcribed in parasegments 10-13, corresponding to adult abdominal segments 5-8, while the regulatory protein is transcribed in parasegment 14, corresponding to adult abdominal segment 9. This division of labor does not appear until stage 13, relatively late in development. Earlier, in stage 10, both forms are transcribed in epidermis. In stage 11, both forms are found in epidermis and mesoderm. By stage 12 central nervous system (ventral cord) expression is evident for both forms. In stage 13 m expression becomes restricted to segments 11-13 for all tissues, while r expression becomes restricted to segment 14 for all tissues (De Lorenzi, 1990b). The r protein's designation as regulatory stems from its unique role in segment 14. There it suppresses myogenesis. It is believed that it also represses transcription of the m form.
The distinction between r and m functions was based on the discovery of three classes of regulatory mutations affecting Abdominal-B (Casanova, 1986). One class affects expression in five parasegments (10-14), a second affects expression in only four (parasegments 10-13) and a third class affects expression in just parasegment 14. The regulatory transcript of ABD-B is thought to suppress the proximal morphogenetic (m) function (Casanova, 1986). The smaller r protein differs from its m counterpart in its lack of an M repeat region. This is a particular amino acid segment that lies upstream of the homeobox. The M repeat is rich in glutamines, a classical transcription activation motif (De Lorenzi, 1988).
P>
Genetic evidence shows that lines, a Drosophila segment polarity gene that has yet to be cloned, is required for the function of the Abdominal-B protein. In lines mutant embryos Abdominal-B protein expression is normal but is incapable of promoting its normal function: formation of the posterior spiracles and specification of an eighth abdominal denticle belt. The tail and A8 segment of lin embryos are highly abnormal. The A8 denticle belt is replaced by naked cuticle that occasionally forms a few denticles less pigmented than the normal ventral denticles. This abnormal A8 cuticle does not resemble the cuticle of any region of the wild-type or of the lin mutant embryo. The absence of anal pads and the abnormal hindgut suggests abnormal development of abdominal segment 11, however, other aspects of the tail development are normal, such as the formation of an anal tuft. In lin embryos the sensory organs are formed at roughly correct positions but have an abnormal shape (Castelli-Gair, 1998). <>The Abd-B gene directs the formation of the posterior spiracles by controlling downstream target genes. The defects associated with lines mutation arise because in lines mutant embryos the Abdominal-B protein cannot activate its direct target empty spiracles (ems) or other downstream genes, such as cut(ct) and spalt(sal), while it can still function as a repressor of Ultrabithorax and abdominal-A. empty spiracles is one gene required for the formation of posterior spiracles. ems expression in the posterior spiracles is regulated by Abd-B. In lin embryos the transcription of ems is not activated in the posterior spiracles, showing that lin is required for Abd-B to activate its direct downstream target. The other putative Abd-B downstream targets cut and spalt are also required for the normal development of the posterior spiracles. The activation of ct and sal in the anlage of the posterior spiracles requires Abd-B function but their activation remains independent of one another and of ems, suggesting that all three genes are independently controlled by Abd-B. In lin mutants neither ct nor sal are activated in the anlage of the posterior spiracles. These results show that in lin mutant embryos, Abd-B is incapable of activating some of its targets. The requirement of lines for Abd-B function is not a specific property of the A8 segment. In wild-type embryos, ectopic Abd-B expression using the GAL4 targeting system results in the formation of ectopic posterior spiracles in segments anterior to A8. In contrast, ectopic Abd-B expression in lin mutants does not form ectopic posterior spiracles showing that no matter where the Abd-B protein is expressed in the embryo it requires lines to be fully functional (Castelli-Gair, 1998).
The effect of lin on Abd-B can be explained at the molecular level if lin is required for protein posttranscriptional modification or as a transcriptional cofactor of Abd-B. There is some evidence that the Abd-B protein is posttranslationally modified. If Lin were mediating this process, it would imply that such posttranscriptional modification is functional in vivo. Alternatively if Lines is a transcriptional cofactor of Abd-B, Lines would be interacting with Abd-B in a similar way to that proposed for Extradenticle with Ubx and Abd-A, or Ftz-F1 with Ftz. It is interesting that Exd does not have any effect on Abd-B protein binding or function, and that lin is specific for Abd-B but not for the other Hox genes tested. This suggests that different HOX proteins use different cofactors that contribute to the DNA binding specificity of the HOX proteins (Castelli-Gair, 1998).
Abdominal-B is required to specify the posterior abdomen and the genitalia. Homologs of Abdominal-B in other species are also needed to determine the posterior part of the body. The function of Abdominal-B in the formation of Drosophila genitalia has been studied, and the absence of Abdominal-B in the genital disc of Drosophila has been shown to transform male and female genitalia into leg or, less frequently, into antenna. These transformations are accompanied by the ectopic expression of genes such as Distal-less or dachshund, which are normally required in these appendages. The extent of wild-type and ectopic Distal-less expression depends on the antagonistic activities of the Abdominal-B gene (as a repressor), and of the decapentaplegic and wingless genes (as activators). Absence of Abdominal-B also changes the expression of Homothorax, a Hox gene co-factor. These results suggest that Abdominal-B forms genitalia by modifying an underlying positional information and repressing appendage development. It is proposed that the genital primordia should be subdivided into two regions, one of them competent to be transformed into an appendage in the absence of Abdominal-B (Estrada, 2001).
Abd-B clones were induced, and they transform posterior abdominal segments into more anterior ones but are normal in the analia. Rare clones transform to distal antennae (second and/or third antennal segment and arista). Transformations to legs or antennae are cell autonomous. The formation of legs requires the activity of genes such as homothorax (hth), dac and Dll, which specify the proximal, medial and distal parts of the leg, respectively. Dll expression in wild-type discs is regulated by the combined activities of wingless and dpp in the genital primordia, and is confined to two groups of cells in male and female discs, the female domains being smaller and expressing lower levels of Dll protein. Since Abd-B is transcribed in the entire genital primordia of the two sexes, some cells co-express Abd-B and Dll. In the male disc, hth is not expressed in the Dll-expressing cells and is also excluded from a large group of cells surrounding them. Levels of antibody signal vary within the disc, and are higher in the female repressed primordium. In females, the hth domain of expression occupies the whole primordium. Lower levels of Hth are detected in a region encompassing the Dll-expressing cells, whereas higher levels are observed in the male repressed primordium. In both sexes, hth expression is absent from the anal primordium. dac is expressed differently in male and female genital primordia: in male discs, Dac protein is detected in two broad lateral bands, while in female discs it is found in the central region, almost coincident with the wg-expressing region. Therefore, the expression patterns of hth, dac and Dll differ substantially from those observed in legs (Estrada, 2001).
It is known that expression of Dll is not required to make male genitalia and that it has only a minor role in the formation of the female one. To ascertain the role of hth in the genitalia, hth minus clones were induced during the third larval period and they were examined in the adult structures. In the female genitalia, hth minus clones cause extra growths with additional vaginal teeth. In males, these clones show occasionally some abnormalities in the clasper teeth. hth clones in the analia are wild type. Possible interactions between Dll and hth in the genital disc were sought. In these experiments, unless stated, the results apply both to male and female genital primordia. Dll minus clones in the Dll domain of the male disc have no hth expression. Similarly, in hth minus clones Dll is not ectopically expressed. Dll was also expressed ectopically and the effect on hth expression was examined. Dll-expressing cells close to the wild-type Dll domain repress hth expression, although not all the cells do so. By contrast, clones far from the Dll domain do not affect hth expression (Estrada, 2001).
To characterize the transformation of genitalia into leg or antennal tissues, Abd-B minus clones were examined. Abd-B minus clones in the genital primordia tend to segregate from the rest of the tissue, round up and have smooth borders, suggesting they have acquired different affinities. By contrast, clones in the analia have indented borders and do not segregate. Abd-B minus clones in the genital primordium close to the normal Dll domain show ectopic, cell-autonomous Dll expression, whereas those far apart do not show such expression. dac is also activated cell autonomously in many Abd-B minus clones. As expected, Dll target genes, such as Bar, also become activated in these clones (Estrada, 2001).
Abd-B minus clones exhibit differential effects on hth, depending on their position: those close to the Dll domain show no hth expression, whereas those located away from the Dll domain show a slight increase in hth signal. Clones in intermediate positions do not significantly change hth levels. This distribution, however, is clearer in females, since in males there is a wide region with no hth expression. The repression of hth observed in some Abd-B minus clones may be mediated by the ectopic Dll (Estrada, 2001).
In the genital disc, the transcription of Dll depends, as in the leg disc, on dpp and wg signals. Abd-B represses Dll expression. Moreover, increasing Abd-B levels in the Dll domain suppresses Dll transcription. The antagonistic activities of dpp/wg and Abd-B in determining the Dll distribution was analyzed. Mutations in PKA ectopically activate wg and dpp expression. PKA minus clones in the genital primordia activate Dll, although only in some places. This activation is not mediated by changes in Abd-B levels. Similarly, although Dll is derepressed in many late Abd-B minus clones, derepression of either dpp or wg was not observed. It is cncuded that there is an antagonism between the activation of Dll by dpp/wg signaling and its repression by Abd-B. This is not mediated by changes in the expression of either dpp, wg or Abd-B (Estrada, 2001).
To characterize this antagonism further, Abd-B minus clones that were made were also unable to transduce the dpp signal. This signal requires the presence of the type II receptor encoded by the gene punt. In put;Abd-B double mutant clones, Dll is not activated, indicating that, in the absence of Abd-B, Dpp (and possibly Wg) are still required to activate Dll. Abd-B minus clones far from the wild-type Dll domain fail to activate Dll ectopically, suggesting that activation of Dll in the absence of Abd-B depends on the range of diffusion of Dpp and Wg, as in the leg disc and in the anal primordium (Estrada, 2001).
Dll is required for the development of legs and antennae, and induces these structures when expressed ectopically in the wing or eye-antennal discs. However, although Dll is also expressed in the genital primordia this expression does not lead to the formation of any of these appendages. To test if Abd-B prevents Dll function Abd-B was eliminated in Dll-expressing cells; these cells ome leg tissue. However, it is possible that the high levels of Dll observed in these mutant cells account for the leg transformation. To test this, use was made of the GAL4/UAS system to increase Dll expression in the genital disc (dpp-GAL4/ UAS-Dll flies). Male and female genitalia of this genotype are abnormal, but not transformed into leg. To extend these observations, the ability of Dll to promote Bar transcription, a gene expressed in the leg disc and activated by Dll, was examined. Bar is not expressed in the female genital primordium and only in a few cells within the Dll domain in the male genital primordium; however, Abd-B minus clones show Bar expression in both sexes. When Dll is ectopically expressed in the genital disc, Bar expression is activated in some of the cells that express Dll. These results suggest that, in females, Dll levels are insufficient to activate Bar when Abd-B is present, but that increasing Dll expression or removing Abd-B activates Bar transcription. Abd-B, therefore, prevents some Dll activity in females. In males, although there is Bar transcription, leg tissue is not formed, probably because Abd-B modifies or prevents the activation of other Dll target genes. A similar case has been reported in the wing disc: ectopic Dll activates bric a brac, a gene downstream of Dll, both in the wing pouch and the body wall region of the wing disc; however, legs appear in the wing, but not in the notum (Estrada, 2001).
The Hox gene Antennapedia is involved in leg development. Therefore, an examination was performed to see whether Antp is derepressed in Abd-B minus clones. Antp is not transcribed in the wild-type genital disc, but some Abd-B coesshow Antp signal. The presence of the Antp product, however, is not required to transform the genitalia into a leg, since Antp:Abd-B double mutant clones still form ectopic legs. This result is consistent with the view that the role of Antp in leg specification is simply to repress hth expression. It seems that Dll alone is able to direct leg development, provided that Hox and hth genes are not transcribed. Under these conditions, leg tissue can be formed in several appendages: leg, wing, antennal and genital primordia (Estrada, 2001).
Ubx and abdominal A expression were examined
in Abd-B minus clones. Ubx was not derepressed in these clones,
whereas some clones presented weak ectopic abd-A
expression, but only in some cells (Estrada, 2001).
The genitalia of Drosophila derive from the genital disc and
require the activity of the Abdominal-B (Abd-B) Hox gene.
This gene encodes two different proteins, Abd-B M and Abd-B R. The embryonic genital disc, like the larval genital disc, is formed by
cells from the eighth (A8), ninth (A9) and tenth (A10) abdominal segments,
which most likely express the Abd-B M, Abd-B R and Caudal products,
respectively. Abd-B m is needed for the development of A8 derivatives
such as the external and internal female genitalia, the latter also requiring
abdominal-A (abd-A), whereas Abd-B r shapes male
genitalia (A9 in males). Although Abd-B r represses Abd-B m
in the embryo, in at least part of the male A9 such regulation does not occur.
In the male A9, some Abd-B m–r– or
Abd-B r– clones activate Distal-less and
transform part of the genitalia into leg or antenna. In the female A8, many
Abd-B m–r– mutant clones produce
similar effects, and also downregulate or eliminate abdominal-A
expression. By contrast, although Abd-B m is the main or only
Abd-B transcript present in the female A8, Abd-B
m– clones induced in this primordium do not alter
Distal-less or abd-A expression, and transform the A8
segment into the A4. The relationship between Abd-B and
abd-A in the female genital disc is opposite that of the embryonic
epidermis, and contravenes the rule that posteriorly expressed Hox genes
downregulate more anterior ones (Foronda, 2006).
Abd-B is a complex gene: the use of four different promoters and the existence of specific exons give rise to several transcripts that encode two different proteins. The A (m) transcript encodes the Abd-B M (or Abd-B I) protein, and the B, C (r) and gamma RNAs encode the
Abd-B R (or Abd-B II) protein. The Abd-B M protein has 221 amino acids more than the Abd-B R product does in its N-terminal domain but both proteins share a common
C-terminal region, which includes the homeodomain.
In the embryonic epidermis, the Abd-B M transcript and protein are expressed
in parasegments (PS) 10-13 (A5-A8 segments), whereas the Abd-B R transcript
and protein are present in PS14-PS15 (A9-A10) initially, and in PS14 (A9) at
late stages. The gamma RNA is transcribed in just a few
cells of PS14 or PS15 (Foronda, 2006 and references therein).
<>herole of Abd-B M and Abd-B R products in genital development remains
unclear. Abd-B m mutations transform the A5-A8 segments into the A4
segment, both in males and females; the female genitalia are lost whereas male
genitalia remain intact. Significantly, the transformations obtained in either Abd-B m or Abd-B r mutants clearly differ from those observed when all Abd-B functions are eliminated: in some of the clones mutant for
Abd-B (m and r), part of the male or female
genitalia are transformed into leg or antenna. Therefore, the precise role of abd-A, Abd-B m and Abd-B r in genitalia development is not well defined (Foronda, 2006).
PTisstudy has analyzed homeotic expression and requirement in terminalia
development. It is proposed that in the embryonic genital disc, as in the larval
discs, Abd-B m, Abd-B r and cad are expressed in the A8, A9
and A10, respectively. It is also reported that abd-A, Abd-B m and
Abd-B r are needed for development of the internal female genitalia,
Abd-B m for the development of female external genitalia and
Abd-B r for the development of male genitalia. Strikingly,
abd-A and Abd-B bear unexpected relationships in mature
genital discs. In the A8 of the female genital disc, Abd-B M maintains
abd-A expression. In Abd-B m mutant clones, however, another
Abd-B protein maintains abd-A expression but does not prevent
abd-A function, since these clones transform the A8 segment into the A4.
In the male A9, Abd-B r function does not repress the Abd-B
m transcript, at least in part of the primordium, and some Abd-B
r mutant clones transform male genitalia into leg or antenna. These
relationships between Hox genes are different from those reported in the
embryonic epidermis and contravene the rule that posteriorly expressed Hox
genes repress those expressed more anteriorly (Foronda, 2006). <>nth third instar genital disc of Drosophila, Abd-B is expressed
in the A8 and A9 segments, and cad in the A10. To
study whether these expression domains are established early in development,
Abd-B and cad transcription were examined in the
embryonic genital disc. This disc is identified by the expression of genes
like snail, escargot or headcase (hdc), and the hdc-lacZ B5 line, which reproduces the pattern of hdc RNA expression, was selected to mark the genital disc. At about stage 15, hdc is expressed in three clusters of cells, two anterior ones placed bilaterally, and a third one
located in a more posterior and central position. The three clusters
fuse later in development to form the genital disc. At stage 15,
xt evn cells were counted at each of the two anterior groups, and two to three cells in the posterior one, making up a total of 14-17 cells. Double
staining with anti-Abd-B and anti-ß-galactosidase antibodies (in
hdc-lacZ embryos), or with GFP and anti-ß-galactosidase antibody
(in cad-Gal4/UAS-GFP; hdc-lacZ/+ embryos), shows that
Abd-B is expressed in the two anterior clusters and cad in
the posterior one (Foronda, 2006).
To ascertain whether the two Abd-B products (Abd-B M and Abd-B R) are
present in the genital disc primordium, the expression driven by
an Abd-B m-Gal4 line was compared with the signal
detected with an antibody that recognizes both Abd-B M and Abd-B R proteins. In
UAS-myc-EGFPF/+;
Abd-B-Gal4LDN/hdc-lacZ embryos, a GFP
signal was seen in about two cells located laterally in each of the two anterior
clusters; these cells most likely express Abd-B m, and, therefore,
are also labelled with the anti-Abd-B antibody. There are also
8-10 Abd-B-expressing cells not labelled with GFP, and these,
probably, correspond to those expressing the Abd-B R protein. Taken together,
these results suggest that the embryonic genital primordium includes three
groups of cells that probably express Abd-B m, Abd-B r and
cad, respectively (Foronda, 2006).
Study of mutant phenotypes reveals that as in the
embryonic cuticle, abd-A and Abd-B m are needed in the A8
whereas Abd-B r is required in the A9. The relationship between these
homeotic products in the mature genital discs, however, clearly differs from
what is observed in the embryonic epidermis. The embryonic genital disc has three distinct cell populations at stages 15/16: some anterior-lateral cells transcribe Abd-B m, anterior-central and middle cells express Abd-B r and
posterior cells transcribe cad, although the expression of these
products may overlap. Because the genital disc is formed by the fusion of
cells coming from the A8, A9 and A10 segments,
and by analogy to the expression of these genes in the mature genital discs, it is concluded that Abd-B m, Abd-B r and cad are
probably expressed in the A8, A9 and A10 segments, respectively, of the
embryonic genital disc (Foronda, 2006).
Abd-B is not only expressed, but also required in the embryonic
genital primordium. In the absence of Abd-B m, the number of
hdc-expressing cells in the disc is reduced, most likely because
these cells adopt now a more anterior fate, as occurs in the cuticle. When Abd-B r is absent, the genital primordium lacks some cells and is disorganized, and when both Abd-B products are absent, the primordium is reduced to a few, dispersed cells, some of which express Dll ectopically, suggesting a transformation into a leg primordium (Foronda, 2006).
The A8, A9 and A10 primordia of the mature genital discs bear anterior and
posterior compartments, with expression of en and wg in each
of these three primordia. Curiously, although three primordia in the
embryonic disc can be defined, based on the expression of Abd-B m, Abd-B r and cad, neither en nor wg is expressed in the three
separate domains at this stage. This may suggest, as was also recently proposed, that new bands of en and wg expression may be formed later in
development, in precise concordance with the three primordia defined
by the Abd-B m, Abd-B r and cad genes. It is noted
that late en expression is also characteristic of the antennal
primordium of the eye-antennal disc (Foronda, 2006).
abd-A is expressed in the whole internal female
genitalia except for the parovaria, and this is consistent with experiments
indicating that parovaria derive from the female A9 segment.
abd-A has been shown to be required for gonad development, and in the
abd-Aiab-3/Df mutant, combinations ovaries are also absent.
However, the defects observed in the female internal genitalia are not
simply due to an indirect effect of the lack of gonads, since iab-4
mutations prevent the formation of the ovaries but do not alter internal
genitalia formation (Foronda, 2006).
The results indicate that Abd-B m is required for the development
of female external and internal genitalia, both derived from the female A8.
The internal genitalia of Abd-B-Gal4LDN/UAS-lacZ
females (driving expression only where Abd-B m levels are high)
were stained with X-gal except in two structures, the oviducts and
parovaria. The absence of oviduct staining in Abd-B-Gal
4LDN/UAS-lacZ females is probably due to the
particular expression driven by this reporter, and does not imply an absence
of Abd-B m transcription in these organs, for two reasons: (1)
Abd-B m transcripts are present in the whole A8 segment of the female
genital disc, and (2) oviduct development is affected in Abd-B m
mutant females. Parovaria, by contrast, are not stained in Abd-B-Gal
4LDN/UAS-lacZ or abd-A-lacZ females, and
this agrees with their A9 provenance. This is supported by the observation that in some Abd-B m mutant females parovaria are the only structures that remain in the internal female genitalia (Foronda, 2006).
Abd-B M seems to be the main or only Abd-B product present in the
female A8, so it was expected that elimination in this segment of just Abd-B M
or of all Abd-B proteins would give similar results. This is not so. Some
Abd-B– clones transform part of the female genitalia
into leg or antenna, whereas Abd-B m mutant clones convert the eighth tergite, and probably the female genitalia, into an anterior abdominal segment. The differences between Abd-B m– and AbdB– clones in the A8 of the female genital disc reveal the existence of unsuspected regulatory interactions between the abd-A and Abd-B genes: whereas Abd-B m– clones do not affect abd-A, in AbdB– clones abd-A expression is
eliminated. This is a surprising result, because it is contrary to what is
observed in the embryo, where Abd-B represses abd-A (Foronda, 2006).
Abd-B m– clones induced in the female A8 do not
alter abd-A expression but do not change Abd-B expression
levels either. This is observed with mutations that do not make Abd-B M
protein, so the Abd-B protein detected is not the Abd-B M product.
Surprisingly, although some Abd-B r expression is detected in the
female A8, uniform Abd-B r expression is not seen throughout this
primordium and Abd-B r transcripts seem not to be derepressed in
Abd-BM5 mutant clones. No explanation is available for this
conundrum. Perhaps the probe used, although it includes sequences complementary to all of the Abd-B r cDNA sequences that have been published, does not efficiently detect all of the non-Abd-B m transcripts (Foronda, 2006).
The differences in regulatory and functional interactions among gene
products in the embryo and the genital discs are not limited to those of
Abd-B and abd-A that have been discussed above. Three other possibilities should be considered. (1) There
may be changes in phenotypic suppression: the transformation of the eighth
tergite to the fourth one in Abd-B m– clones is due
to abd-A. Because in these clones Abd-B protein is still present, this suggests that abd-A may phenotypically suppress Abd-B, differently from what is generally observed in the embryo. (2) Abd-B r represses Abd-B m in the embryo, but some Abd-B r– clones do not activate Abd-B m in the male disc. (3) abd-A represses Dll in the embryo, but not in the female genital disc, and ectopic Dll can repress abd-A instead. abd-A does not repress Dll in the leg discs either, and this resembles Ubx function, which represses Dll only early in development. By contrast, Abd-B represses Dll in the embryo, in the larval genital disc, and in the leg disc when ectopically expressed (Foronda, 2006).
Abd-B r expression is restricted to the A9 segment in male genital
discs, but shows expression in the A9 and in some cells of the A8 in female
genital discs. In spite of this, Abd-B r– clones in
the external female genitalia (A8) are phenotypically wild type. In the male
A9, some Abd-B r mutant clones eliminate Abd-B, activate
Dll and transform part of the genitalia into distal leg or antenna.
This is similar to the result obtained in some
Abd-B– clones, and it implies that Abd-B m
is not derepressed in these mutant clones. However, Abd-B m is
perhaps derepressed in those Abd-B r mutant clones where
Abd-B signal remains (Foronda, 2006).
Although Abd-B r– clones affect, almost
exclusively, male genitalia development, Abd-B r hemizygous or
trans-heterozygous flies lack genitalia and analia in both sexes. This
probably reflects the absence of proper interactions between the different
primordia needed for the growth of the genital disc. In
Abd-B r mutant females, the internal genitalia are abnormal, and in
some of these females, an absence of parovaria and the presence of
three or four spermathecae is observed. This phenotype is consistent with a
segment-autonomous transformation of A9 derivatives (parovaria) into A8
structures (spermathecae), similar to the embryonic cuticular transformation
of A9 into A8 observed in Abd-B r mutations. A transformation of parovaria into spermathecae has been described in Polycomblike mutants, and may also indicate a transformation of A9 to A8 (Foronda, 2006).
These results illustrate that there are quite different Hox cross-regulatory
interactions in the embryo and in the genital disc. The effects in the
genital discs contradict the general rule that genes transcribed more
posteriorly suppress or downregulate the expression of more anterior ones. This
rule has, nevertheless, some exceptions in genes of the Antennapedia complex.
Further, differences in Hox cross-regulation between the embryo and imaginal
discs are not unprecedented: the proboscipedia (pb) Hox gene
is positively regulated by Sex combs reduced in the embryo, but
pb activates Sex combs reduced in the labial imaginal disc (Foronda, 2006).
It has been proposed that the primordia of female and male genitalia could
be subdivided into an 'appendage-like' and a 'trunk-like' region). These two regions of the female A8 can now be defined more
precisely. The 'appendage-like' region would be that expressing abd-A
and low levels of Abd-B, and corresponds approximately to the
presumptive internal female genitalia. This domain is
roughly coincident with the region of expression of a reporter insertion in
buttonhead, the gene that defines ventral appendage development, and
this is also, approximately, the domain where Abd-B–
clones may activate Dll. If this subdivision is correct,
the 'appendage' specification defined by buttonhead would be
repressed in the wild type by Abd-B, which both limits Dll
expression to a few cells of the A8 primordium and prevents Dll
function. Abd-B– clones in this region eliminate abd-A expression and promote leg or antenna development. This subdivision may also apply to the male disc, the penis apparatus presumptive region being the main 'appendage' domain. Similar to what is described in this study, the labial disc possesses a large 'appendage' region that is revealed by Dll derepression in pb mutations. This characteristic, and the changes in Hox gene cross-regulation between the embryo and the imaginal disc, are two features shared by pb/labial disc and Abd-B/genital disc (Foronda, 2006).
A cis-regulatory region of nearly 300 kb controls the expression of the three bithorax complex (BX-C) homeotic genes: Ubx, abd-A and Abd-B. Interspersed between the numerous enhancers and silencers within the complex are elements called domain boundaries. Many pieces of evidence have suggested that boundaries function to create autonomous domains by interacting among themselves and forming chromatin loops. In order to test this hypothesis, Dam identification was used to probe for interactions between the Fab-7 boundary and other regions in the BX-C. It was surprising to find that the targeting of Dam methyltransferase (Dam) to the Fab-7 boundary results in a strong methylation signal at the Abd-Bm promoter, ~35 kb away. Moreover, this methylation pattern is found primarily in the tissues where Abd-B is not expressed and requires an intact Fab-7 boundary. Overall, this work provides the first documented example of a dynamic, long-distance physical interaction between distal regulatory elements within a living, multicellular organism (Cleard, 2006).
For decades, the Drosophila BX-C has served as a model system for studying complex gene regulation. Through the years, results from numerous experiments have indicated that the large cis-regulatory region of the BX-C is divided into parasegment-specific chromatin domains in which each domain controls the expression of a single homeotic gene in a particular parasegment. Notably, these domains are aligned along the chromosome in an order corresponding to the parasegment in which they function. To keep the enhancers and silencers of each domain autonomous, elements known as domain boundaries have been found to separate each domain. The Fab-7 boundary, for example, normally separates the iab-6 from the iab-7 cis-regulatory domain. Both iab-6 and iab-7 control the expression of the Abd-B gene, in different parasegments and at different levels (in parasegment 11 (PS11) and PS12 respectively. Removal of the Fab-7 element causes a fusion between these two domains and results in the inappropriate use of iab-7 enhancers in PS11, where only iab-6 enhancers are usually functional. The resulting hyperactivation of Abd-B in PS11 causes a homeotic transformation of PS11 into PS12 (Cleard, 2006).
In transgenic contexts, the Fab-7 boundary (like other boundaries isolated from the BX-C) has been shown to behave as an insulator, blocking enhancer-promoter interactions. Although the mechanism for this activity is still unknown, this insulator activity is almost paradoxical within the BX-C because Fab-7 is situated between the Abd-B promoter and the iab-5 and iab-6 enhancers within the BX-C. These distal enhancers must therefore bypass the Fab-7 boundary to interact with the Abd-B promoter. To resolve this apparent paradox, two models have been proposed. The first of these models relies on the ability of elements called promoter targeting sequences (PTS) to aid distal enhancers in bypassing intervening insulators. The second of these models suggests that boundaries interact with one another to allow distal enhancers to bypass them. Each of these models proposes chromatin loops as the mode of action (Cleard, 2006).
In order to directly test for boundary-mediated long-distance chromatin loop formation, the Dam identification method (DamID) was employed. Previous studies in Saccharomyces cerevisiae have shown that DamID is a viable method for probing long-distance chromatin interactions. And, given the in vivo nature of the procedure, DamID provides an opportunity to monitor long-distance chromatin interactions occurring in a living organism without the generation of artifacts due to fixation and DNA isolation (Cleard, 2006).
In order to target Dam to a specific region of the BX-C, 14 Gal4-UAS sites were engineered into the native BX-C by gene conversion, just distal to the Fab-7 boundary (relative to the Abd-B promoter). Using these sites, it was possible to target a Dam-Gal4DBD fusion protein to the Fab-7 region and monitor distant regions of the BX-C for methylation occurring in trans. Two loxP recombination sites were added, flanking the Fab-7 boundary element (leaving the iab7PRE and the PTS-6 element), to determine if any discovered interactions were Fab-7 dependent. Flies homozygous for the converted chromosome or the converted chromosome deleted for Fab-7 are viable, and Fab-7 deleted lines display the expected Fab-7 mutant phenotype (Cleard, 2006).
Nine primer pairs within the BX-C spanning the region from Fab-7 to the Abd-Bm promoter were chosen to probe for transmethylation upon targeting of Dam to the Fab-7 region. Each primer pair amplifies a region centered on a Dam target site (DpnII site). For the first experiment, many primers were chosen that cluster near possible interacting elements like the Fab-8 boundary and the Abd-Bm promoter. To control for nontargeted methylation, primers centered on a GATC-containing region of the TBP gene were also monitored. In lines expressing the Dam-Gal4DBD fusion protein alone, the background methylation seems to follow roughly the patterns previously established for DNAseI hypersensitivity. In order to control for this low level of background, the level of methylation in the untargeted line was subtracted from each corresponding point in the targeted lines (Cleard, 2006).
As expected, the targeting of Dam methyltransferase to the Fab-7 region results in a strong enrichment of methylation near the site of targeting that decreases to lower levels in moving farther away. An additional peak of DNA methylation was observed at a place that corresponds to the Abd-Bm promoter, ~35 kb away. Although a small increase in methylation was found around the Fab-8 boundary element, thus far statistical analysis is inconclusive regarding its significance. Previously, Dam methylation activity had been found to spread only to a distance of ~2.5 kb on each side of the targeting site. Therefore, it is believed that the methylation found at the Abd-Bm promoter, ~35 kb away, indicates that Fab-7 and the Abd-Bm promoter are in very close proximity to each other within the nucleus (Cleard, 2006).
Upon the removal of the Fab-7 boundary, there was a significant drop in the methylation levels at the Abd-Bm promoter. This drop occurs even though the targeting site is now 1 kb closer to the promoter in these flies than in flies carrying an intact Fab-7 element. The finding that the methylation pattern changes between lines carrying Fab-7 and lines lacking Fab-7 strongly supports the idea that Fab-7 itself participates in the long-distance interaction (Cleard, 2006).
It was next asked if the interaction between Fab-7 and the Abd-Bm promoter occurs in areas where Abd-B is expressed or silenced. To do this, the experiments were repeated using two different tissues: one in which Abd-B is expressed and one in which Abd-B is silenced. For simplicity, whole adult abdomens were chosen as representative of tissues where Abd-B is expressed and whole adult heads as representative of tissues where Abd-B is silenced. Four new primer pairs were added to these experiments to obtain better overall coverage of the region (Cleard, 2006).
The results highlight two differences between the methylation patterns found in head tissues versus abdominal tissues. First, it was found that there is a significantly higher level of methylation in abdominal tissues relative to head tissues. This difference probably represents an overall change in the chromatin structure in the regions where Abd-B is expressed. However, a change in the general pattern of methylation was also observed at the Abd-Bm promoter region. In order to eliminate statistical artifacts arising from the generally higher level of methylation in abdominal tissues, the two samples were normalized to the level of methylation found at the targeting site to examine the change in pattern. Upon doing this, it was found that in abdominal tissues, where many cells have Abd-B turned on, strong methylation was seen only at the targeting site. Meanwhile, in the head tissues, where Abd-B is turned off, strong methylation was found at the Abd-B promoter as well as at the targeting site. Although the deletion of the Fab-7 boundary had no effect on the methylation pattern in abdominal tissues, a clear decrease was seen in the level of transmethylation at the Abd-Bm promoter upon deletion of the Fab-7 element in head tissues (Cleard, 2006).
Overall, this work demonstrates two important points: first, that the Fab-7 boundary region mediates a long-distance interaction between itself and a region near the Abd-Bm promoter, and second, that this interaction takes place primarily in areas where Abd-B is not expressed. Although these results were quite unexpected, previous data had hinted at the possibility of a tethering element in the region around the Abd-Bm promoter. It has been shown previously that transvection at the Abd-B locus could be enhanced by a deletion of the region upstream of the Abd-Bm promoter. To explain this observation, it was proposed that the Abd-Bm promoter region might be tethering the Abd-B cis-regulatory elements. The current data indicate that this hypothesis is probably true and that this process is mediated by the BX-C boundary elements (Cleard, 2006).
At first glance, the finding that this interaction occurs only in the inactive domain seems to point to a role for Fab-7 and other boundary elements in the silencing of the Abd-B gene. Indeed, Fab-7 is located adjacent to the iab-7PRE silencer. However, although a role for Fab-7 in silencing cannot be strictly ruled out, this model is disfavored based on the phenotype of Fab-7 mutations. Two main facets of the Fab-7 mutant phenotype cause doubt with regard to a role for Fab-7 in silencing. First, Fab-7 deletions result in a net increase of Abd-B expression only in A6, where Abd-B is normally expressed, and do not affect segments where Abd-B is silenced. And second, in Fab-7 mutants, some of the cells in A6 actually show ectopic silencing of Abd-B. This silencing can even be enhanced by crossing in a trithorax-group mutation. Therefore, Fab-7 mutants are not deficient in silencing capability but instead simply make inappropriate decisions regarding silencing. It was based on these observations and others that Fab-7 has been characterized as a boundary element and not a silencer (Cleard, 2006).
In recent years, a number of experiments have suggested that the ability of insulators to block distal enhancer activity represents the formation of chromatin domain loops. This study presents the first direct, in vivo evidence for this, at the Fab-7 locus. However, unlike with other insulators, which are thought to interact with each other, Fab-7 was found to interact with the Abd-B promoter. Based on previous data that suggest that tethering might be required to bring enhancers to the Abd-B promoter, and based on the phenotype of flies with Fab-7 mutations, which affect only the Abd-B expressing region, it is believed that this interaction is probably important for targeting specific enhancer regions to the Abd-B promoter. However, regardless of whether Fab-7 targets enhancers or silencers to the Abd-B promoter, it now seems clear that the role of Fab-7 in the BX-C is to bring elements of the cis-regulatory regions into close proximity to the Abd-B promoter. Because other BX-C boundary elements share such similar characteristics, it is believed the other BX-C boundaries will share a similar purpose (Cleard, 2006).
Drosophila bithorax complex (BX-C) is one of the best model systems for studying the role of boundaries (insulators) in gene regulation. Expression of three homeotic genes, Ubx, abd-A, and Abd-B, is orchestrated by nine parasegment-specific regulatory domains. These domains are flanked by boundary elements, which function to block crosstalk between adjacent domains, ensuring that they can act autonomously. Paradoxically, seven of the BX-C regulatory domains are separated from their gene target by at least one boundary, and must 'jump over' the intervening boundaries. To understand the jumping mechanism, the Mcp boundary was replaced with Fab-7 and Fab-8. Mcp is located between the iab-4 and iab-5 domains, and defines the border between the set of regulatory domains controlling abd-A and Abd-B. When Mcp is replaced by Fab-7 or Fab-8, they direct the iab-4 domain (which regulates abd-A) to inappropriately activate Abd-B in abdominal segment A4. For the Fab-8 replacement, ectopic induction was only observed when it was inserted in the same orientation as the endogenous Fab-8 boundary. A similar orientation dependence for bypass activity was observed when Fab-7 was replaced by Fab-8. Thus, boundaries perform two opposite functions in the context of BX-C-they block crosstalk between neighboring regulatory domains, but at the same time actively facilitate long distance communication between the regulatory domains and their respective target genes (Postaka, 2018).
The three homeotic (HOX) genes in the Drosophila Bithorax complex (BX-C), Ultrabithorax (Ubx), abdominal-A (abd-A) and Abdominal-B (Abd-B), are responsible for specifying cell identity in parasegments (PS) 5-14, which form the posterior half of the thorax and all of the abdominal segments of the adult fly. Parasegment identity is determined by the precise expression pattern of the relevant HOX gene and this depends upon a large cis-regulatory region that spans 300 kb and is subdivided into nine PS domains that are aligned in the same order as the body segments in which they operate. Ubx expression in PS5 and PS6 is directed by abx/bx and bxd/pbx, while abd-A expression in PS7, PS8, and PS9 is controlled by iab-2, iab-3, and iab-4. Abd-B is regulated by four domains, iab-5, iab-6, iab-7 and iab-8, which control expression in PS10, PS11, PS12 and PS13 respectively (Postaka, 2018).
Each regulatory domain contains an initiator element, a set of tissue-specific enhancers and Polycomb Response Elements (PREs) and is flanked by boundary/insulator elements. BX-C regulation is divided into two phases, initiation and maintenance. During the initiation phase, a combination of gap and pair-rule proteins interact with initiation elements in each regulatory domain, setting the domain in the on or off state. In PS10, for example, the iab-5 domain, which regulates Abd-B, is activated by its initiator element, while the more distal Abd-B domains, iab-6 to iab-8 are set in the off state. In PS11, the iab-6 initiator activates the domain, while the adjacent iab-7 and iab-8 domains are set in the off state. Once the gap and pair-rule gene proteins disappear during gastrulation, the on and off states of the regulatory domains are maintained by Trithorax (Trx) and Polycomb (PcG) group proteins, respectively (Postaka, 2018).
In order to select and then maintain their activity states independent of outside influence, adjacent regulatory domains are separated from each other by boundary elements or insulators. Mutations that impair boundary function permit crosstalk between positive and negative regulatory elements in adjacent domains and this leads to the misspecification of parasegment identity. This has been observed for deletions that remove five of the BX-C boundaries (Front-ultraabdominal (Fub), Miscadestral pigmentation (Mcp), Frontadominal-6 (Fab-6), Frontadominal-7 (Fab-7), and Frontadominal-8 (Fab-8)) (Postaka, 2018).
While these findings indicate that boundaries are needed to ensure the functional autonomy of the regulatory domains, their presence also poses a paradox. Seven of the nine BX-C regulatory domains are separated from their target HOX gene by at least one intervening boundary element. For example, the iab-6 regulatory domain must 'jump over' or 'bypass' Fab-7 and Fab-8 in order to interact with the Abd-B promoter. That the blocking function of boundaries could pose a significant problem has been demonstrated by experiments in which Fab-7 is replaced by heterologous elements such as scs, gypsy or multimerized binding sites for the architectural proteins dCTCF, Pita or Su(Hw). In these replacements, the heterologous boundary blocked crosstalk between iab-6 and iab-7 just like the endogenous boundary, Fab-7. However, the boundaries were not permissive for bypass, preventing iab-6 from regulating Abd-B (Postaka, 2018).
A number of models have been proposed to account for this paradox. One is that BX-C boundaries must have unique properties that distinguish them from generic fly boundaries. Since they function to block crosstalk between enhancers and silencers in adjacent domains, an appealing idea is that they would be permissive for enhancer/silencer interactions with promoters. However, several findings argue against this notion. For one, BX-C boundaries resemble those elsewhere in the genome in that they contain binding sites for architectural proteins such as Pita, dCTCF, and Su(Hw). Consistent with their utilization of these generic architectural proteins, when placed between enhancers (or silencers) and a reporter gene, BX-C boundaries block regulatory interactions just like boundaries from elsewhere in the genome. Similarly, there is no indication in these transgene assays that the blocking activity of BX-C boundaries are subject to parasegmental regulation. Also arguing against the idea that BX-C boundaries have unique properties, the Mcp boundary, which is located between iab-4 and iab-5, is unable to replace Fab-7. Like the heterologous boundaries, it blocks crosstalk, but it is not permissive for bypass. A second model is that there are special sequences, called promoter targeting sequence (PTS), located in each regulatory domain that actively mediate bypass. While the PTS sequences that have been identified in iab-6 and iab-7 enable enhancers to 'jump over' an intervening boundary in transgene assays, they do not have a required function in the context of BX-C and are completely dispensable for Abd-B regulation (Postaka, 2018).
A third model is suggested by transgene 'insulator bypass' assays. In one version of this assay, two boundaries instead of one are placed in between an enhancer and the reporter. When the two boundaries pair with each other, the enhancer is brought in close proximity to the reporter, thereby activating rather than blocking expression. Consistent with a possible role in BX-C bypass, these pairing interactions can occur over large distances and even skip over many intervening boundaries. The transgene assays point to two important features of boundary pairing interactions that are likely to be relevant in BX-C. First, pairing interactions are specific. For this reason boundaries must be properly matched with their neighborhood in order to function appropriately. A requirement for matching is illustrated in transgene bypass experiments in which multimerized binding sites for specific architectural proteins are paired with themselves or with each other. Bypass was observed when multimerized dCTCF, Zw5 or Su(Hw) binding sites were paired with themselves; however, heterologous combinations (e.g., dCTCF sites with Su(Hw) sites) did not support bypass. A second feature is that pairing interactions between boundaries are typically orientation dependent For example, scs pairs with itself head-to-head, not head-to-tail (Postaka, 2018).
If both blocking and bypass activities are intrinsic properties of fly boundaries then the BX-C boundaries themselves may facilitate contacts between the regulatory domains and their target genes. Moreover, the fact that both blocking and bypass activity are non-autonomous (in that they depend on partner pairing) could potentially explain why heterologous Fab-7 replacements like gypsy and Mcp behave anomalously while Fab-8 functions appropriately. Several observations fit with this idea. There is an extensive region upstream of the Abd-B promoter that has been implicated in tethering the Abd-B regulatory domains to the promoter and this region could play an important role in mediating bypass by boundaries associated with the distal Abd-B regulatory domains (iab-5, iab-6, iab-7). Included in this region is a promoter tethering element (PTE) that facilitates interactions between iab enhancers and the Abd-B promoter in transgene assays. Just beyond the PTE is a boundary-like element, AB-I. In transgene assays AB-I mediates bypass when combined with either Fab-7 or Fab-8. In contrast, a combination between AB-I and Mcp fails to support bypass. The ability of both Fab-7 and Fab-8 to pair with AB-I is recapitulated in Fab-7 replacement experiments. Unlike Mcp, Fab-8 has both blocking and bypass activity when inserted in place of Fab-7. Moreover, its bypass but not blocking activity is orientation-dependent. When inserted in the same orientation as the endogenous Fab-8 boundary, it mediates blocking and bypass, while it does not support bypass when inserted in the opposite orientation (Postaka, 2018).
Boundaries flanking the Abd-B regulatory domains must block crosstalk between adjacent regulatory domains but at the same time allow more distal domains to jump over one or more intervening boundaries and activate Abd-B expression. While several models have been advanced to account for these two paradoxical activities, replacement experiments argued that both must be intrinsic properties of the Abd-B boundaries. Thus Fab-7 and Fab-8 have blocking and bypass activities in Fab-7 replacement experiments, while heterologous boundaries including multimerized dCTCF sites and Mcp from BX-C do not. One idea is that Fab-7 and Fab-8 are simply 'permissive' for bypass. They allow bypass to occur, while boundaries like multimerized dCTCF or Mcp are not permissive in the context of Fab-7. Another is that they actively facilitate bypass by directing the distal Abd-B regulatory domains to the Abd-B promoter. Potentially consistent with an 'active' mechanism that involves boundary pairing interactions, the bypass activity of Fab-8 and to a lesser extent Fab-7 is orientation dependent (Postaka, 2018).
This study has tested these two models further. For this purpose the Mcp boundary was used for in situ replacement experiments. Mcp defines the border between the regulatory domains that control expression of abd-A and Abd-B. In this location, it is required to block crosstalk between the flanking domains iab-4 and iab-5, but it does not need to mediate bypass. In this respect, it differs from the boundaries that are located within the set of regulatory domains that control either abd-A or Abd-B, as these boundaries must have both activities. If bypass were simply passive, insertion of a 'permissive' Fab-7 or Fab-8 boundary in either orientation in place of Mcp would be no different from insertion of a generic 'non-permissive' boundary such as multimerized dCTCF sites. Assuming that Fab-7 and Fab-8 can block crosstalk out of context, they should fully substitute for Mcp. In contrast, if bypass in the normal context involves an active mechanism in which more distal regulatory domains are brought to the Abd-B promoter, then Fab-7 and Fab-8 replacements might also be able to bring iab-4 to the Abd-B promoter in a configuration that activates transcription. If they do so, then this process would be expected to show the same orientation dependence as is observed for bypass of the Abd-B regulatory domains in Fab-7 replacements (Postaka, 2018).
Consistent with the idea that a boundary located at the border between the domains that regulate abd-A and Abd-B need not have bypass activity, this study found that multimerized binding sites for the dCTCF protein fully substitute for Mcp. Like the multimerized dCTCF sites, Fab-7 and Fab-8 are also able to block crosstalk between iab-4 and iab-5. Blocking activity of Fab-7 is incomplete and there are small clones of cells in which the mini-y reporter is activated in A4. In contrast, the blocking activity of Fab-8 is comparable to the multimerized dCTCF sites and the mini-y reporter is off throughout A4. One plausible reason for this difference is that Mcp and the boundaries flanking Mcp (Fab-4 and Fab-6) utilize dCTCF as does Fab-8, while this architectural protein does not bind to Fab-7(Postaka, 2018).
Importantly, in spite of their normal (or near normal) ability to block crosstalk, both boundaries still perturb Abd-B regulation. In the case of Fab-8, the misregulation of Abd-B is orientation dependent just like the bypass activity of this boundary when it is used to replace Fab-7. When inserted in the reverse orientation, Fab-8 behaves like multimerized dCTCF sites and it fully rescues the Mcp deletion. In contrast, when inserted in the forward orientation, Fab-8 induces the expression of Abd-B in A4 (PS9), and the misspecification of this parasegment. Unlike classical Mcp deletions or the McpPRE replacement described here, expression of the Abd-B gene in PS9 is driven by iab-4, not iab-5. This conclusion is supported by two lines of evidence. First, the mini-y reporter inserted in iab-5 is off in PS9 cells indicating that iab-5 is silenced by PcG factors as it should be in this parasegment. Second, the ectopic expression of Abd-B is eliminated when the iab-4 regulatory domain is inactivated (Postaka, 2018).
These results, taken together with previous studies, support a model in which the chromatin loops formed by Fab-8 inserted at Mcp in the forward orientation brings the enhancers in the iab-4 regulatory domain in close proximity to the Abd-B promoter, leading to the activation of Abd-B in A4 (PS9). In contrast, when inserted in the opposite orientation, the topology of the chromatin loops formed by the ectopic Fab-8 boundary are not compatible with productive interactions between iab-4 and the Abd-B promoter. Moreover, it would appear that boundary bypass for the regulatory domains that control Abd-B expression is not a passive process in which the boundaries are simply permissive for interactions between the regulatory domains and the Abd-B promoter. Instead, it seems to be an active process in which the boundaries are responsible for bringing the regulatory domains into contact with the Abd-B gene. It also seems likely that bypass activity of Fab-8 (and also Fab-7) may have a predisposed preference, namely it is targeted for interactions with the Abd-B gene. This idea would fit with transgene bypass experiments, which showed that both Fab-7 and Fab-8 interacted with an insulator like element upstream of the Abd-B promoter, AB-I, while the Mcp boundary didn't (Postaka, 2018).
Similar conclusions can be drawn from the induction of Abd-B expression in A4 (PS9) when Fab-7 is inserted in place of Mcp. Like Fab-8, this boundary inappropriately targets the iab-4 regulatory domain to Abd-B. Unlike Fab-8, Abd-B is ectopically activated when Fab-7 is inserted in both the forward and reverse orientations. While the effects are milder in the reverse orientation, the lack of pronounced orientation dependence is consistent with experiments in which Fab-7 was inserted at its endogenous location in the reverse orientation. Unlike Fab-8 only very minor iab-6 bypass defects were observed. In addition to the activation of Abd-B in A4 (PS9) the Fab-7 Mcp replacements also alter the pattern of Abd-B regulation in more posterior segments. In the forward orientation, A4 and A5 are transformed towards an A6 identity, while A6 is also misspecified. Similar though somewhat less severe effects are observed in these segments when Fab-7 is inserted in the reverse orientation. At this point the mechanisms responsible for these novel phenotypic effects are uncertain. One possibility is that pairing interactions between the Fab-7 insert and the endogenous Fab-7 boundary disrupt the normal topological organization of the regulatory domains in a manner similar to that seen in boundary competition transgene assays. An alternative possibility is that Fab-7 targets iab-4 to the Abd-B promoter not only in A4 (PS9) but also in cells in A5 (PS10) and A6 (PS11). In this model, Abd-B would be regulated not only by the domain that normally specifies the identity of the parasegment (e.g., iab-5 in PS10), but also by interactions with iab-4. This dual regulation would increase the levels of Abd-B, giving the weak GOF phenotypes. Potentially consistent with this second model, inactivating iab-4 in the McpF8 replacement not only rescues the A4 (PS9) GOF phenotypes but also suppresses the loss of anterior trichomes in the A6 tergite (Postaka, 2018).
The evolutionary origins of complex morphological structures such as the vertebrate eye or insect wing remain one of the greatest mysteries of biology. Recent comparative studies of gene expression imply that new structures are not built from scratch, but rather form by co-opting preexisting gene networks. A key prediction of this model is that upstream factors within the network will activate their preexisting targets (i.e., enhancers) to form novel anatomies. This study shows how a recently derived morphological novelty present in the genitalia of D. melanogaster employs an ancestral Hox-regulated network deployed in the embryo to generate the larval posterior spiracle. This study demonstratea how transcriptional enhancers and constituent transcription factor binding sites are used in both ancestral and novel contexts. These results illustrate network co-option at the level of individual connections between regulatory genes and highlight how morphological novelty may originate through the co-option of networks controlling seemingly unrelated structures (Glassford, 2015).
This study has shown how a gene regulatory network (GRN) underlying a novel structure, the posterior lobe, is composed of components that are active in the embryonic posterior spiracle, an ancestral Hox-regulated structure that was present at the inception of this novelty (see Model Depicting the Co-option of Genes, Enhancers, and Transcription Factor Binding Sites during the Origination of the Novel Posterior Lobe). These findings confirm previous speculation that network co-option proceeds through the re-use of individual transcription factor binding sites within enhancer sequences. Further, the data help calibrate expectations concerning the degree of physical similarity between novel and ancestral structures during co-option events. Below, it is discussed how the architecture of the posterior spiracle network may have predisposed it for co-option in the genitalia, and the general implications of these findings are explored with regard to the origins of morphological novelty (Glassford, 2015).
While these results illustrate the downstream consequences of co-option, the upstream causative events await characterization. It is suspected that some number of high-level regulators of the posterior spiracle network recently evolved novel genital expression patterns through alterations within their regulatory regions. Currently, Unpaired represents the best candidate upstream factor, as it is positioned near the top of the spiracle network, differs in expression greatly between lobed and non-lobed species (unlike Spalt and Abd-B), and is the only high-level factor in the spiracle network for which a shared lobe and spiracle enhancer has yet to be identified. Indeed, a reporter screen of the 30 kb of regulatory DNA immediately surrounding the upd gene identified a posterior spiracle enhancer that is not deployed in the posterior lobe, marking an important point of divergence separating the posterior spiracle and posterior lobe networks. However, the identification of enhancers controlling upd expression in the posterior lobe will be required to resolve its role in this structure’s origination (Glassford, 2015).
The architecture of the posterior spiracle network may have shaped the possible developmental contexts in which it could be co-opted. The Hox factor Abd-B has a deeply conserved role in the insect abdomen and genitalia. The top-level factors of the posterior spiracle network depend upon Abd-B for activation in the embryo. This regulation by Abd-B extends to lower tiers of the network, such as Poxn. The tight integration of Abd-B with multiple tiers of the posterior spiracle network may have limited this network’s re-deployment to posterior body segments that express Abd-B. Indeed, several components of this network (Poxn, ems, upd) are activated early during genital development in the presumptive cleavage furrow separating the lateral plate from the clasper. This may represent the aftereffect of multiple waves of re-deployment in Abd-B expressing tissues. Examination of additional examples of network co-option at the level of constituent regulatory sequences could reveal general rules that govern and bias network redeployment (Glassford, 2015).
Historically, the identification of co-option events has relied upon comparative analyses of gene co-expression. The first examples of co-option were diagnosed by finding novel gene expression patterns near zones of ancestral function, such as the deployment of the posterior wing patterning circuit within novel butterfly eyespots. Subsequently, many examples of co-option have involved educated guesses of the types of networks that contribute to the novelty, such as the role of the appendage specification network within beetle horns or the sharing of the biomineralization network between adult and larval skeletons of sea urchins. The data suggest that tracing the evolutionary origins of individual enhancers provides a less biased path for connecting novelties to their ancestral beginnings, as any of the seven enhancers that this study has characterized in the posterior lobe would have led to the spiracle network. Further, this approach is likely to illuminate the underlying cellular mechanisms by which the co-option of a network is translated into a novel developmental outcome (Glassford, 2015).
Rather than generating a serial homolog of the posterior spiracle, the co-option event forming the posterior lobe resulted in an epithelial outgrowth, likely owing to the deployment of only a portion of the spiracle network in the genitalia. This is reflected by the absence of the Cut transcription factor and downstream genes that control the spiracular chamber’s development. Of the 10 genes this study has identified in both networks, 9 are active in the stigmatophore, the outer sheath of the posterior spiracle that protrudes from the body through a process that involves convergent extension. Collectively, these findings imply that similar morphogenic processes are activated by this shared network in the novel setting of the posterior lobe. It is proposed that the inspection of enhancers underlying other novel three-dimensional structures may reveal similar networks that have been used over and over again to generate "unique styles of architecture" within developing tissues (Glassford, 2015).
Four classes of overlapping transcripts are generated from the Abd-B gene The transcription initiation sites for the class A (4.6 kb) and class B (3.4kb) transcripts show that they are generated from separate promoters. Both of these transcripts are present throughout the period during which the ABD-B subfunctions are required. A mutation that inactivates the morphogenetic function is associated with a 411bp deletion of the initiation site for the 4.6 kb RNA. Regulatory function mutations disrupt the transcription unit for the 3.4 kb RNA, but not the 4.6 kb RNA. A morphogenetic (m) function is assigned to the 4.6 kb RNA and a regulatory (r) function to the 3.4 kb RNA.
A 7.8 kb RNA expressed during embryogenesis may also contribute to the regulatory function. Sequence analysis of cDNAs indicates that the 4.6 kb RNA encodes the 55-kD morphogenetic protein, whereas the 3.4 kb RNA encodes a 30-kD regulatory protein. The m and r proteins share a carboxy-terminal sequence that includes the homeodomain, but the r protein lacks a glutamine-rich amino-terminal domain found in the m protein (Celniker,1989 and Zavortink, 1989).
Exons - four for class A transcript and five for class B transcript
Bases in 3' UTR - 1604
There are two variants to the ABD-B protein. One, a morphogenetic unction can be assigned to the 55 KD protein and a second, regulatory function can be assigned to a 30 KD protein (Boulet, 1991).
The class r 4.6 kb transcript encodes for a protein of 493 amino acids. The class m 3.4 kb transcript encodes for a protein of 270 amino acids (Zavortink, 1989).
See Four paralogous Hox clusters of mammals for homologies of Abd-B with mammalian Hox cluster genes.
date revised: 29 January 2026> RF="../aimain/1aahome.htm">Home page: The Interactive Fly © 2026 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
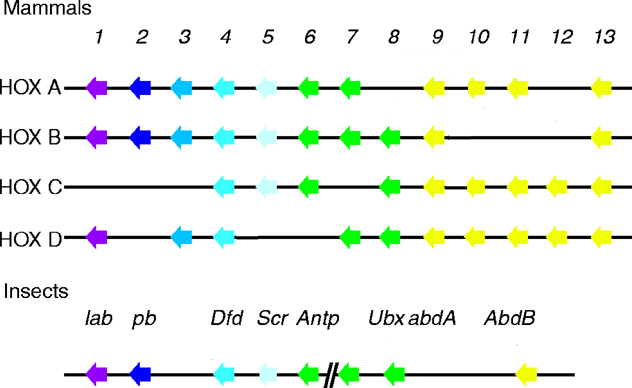{kind=link}