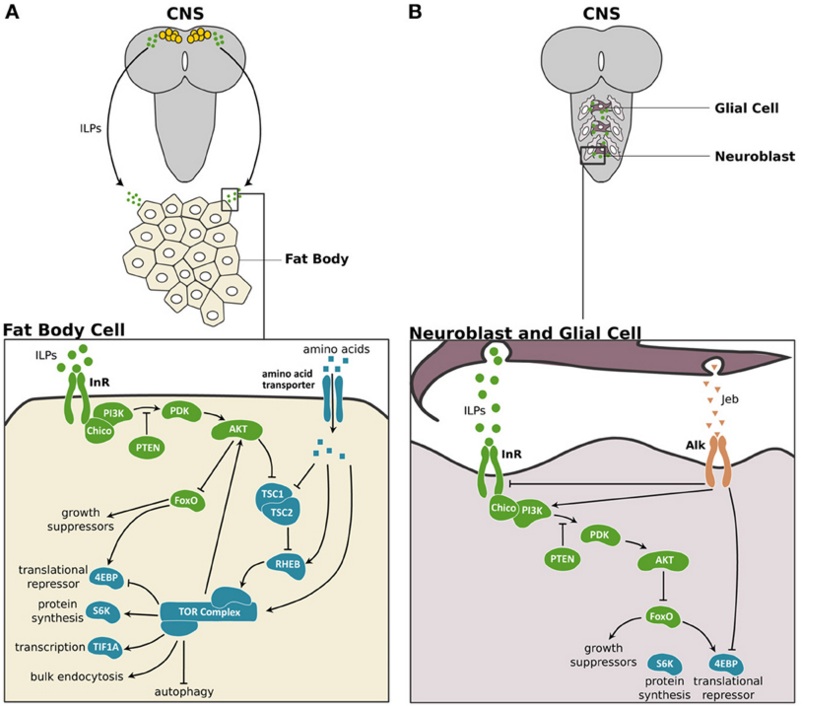
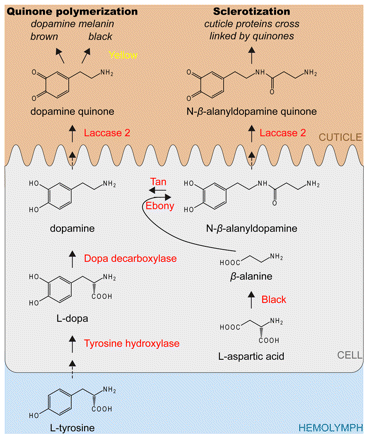
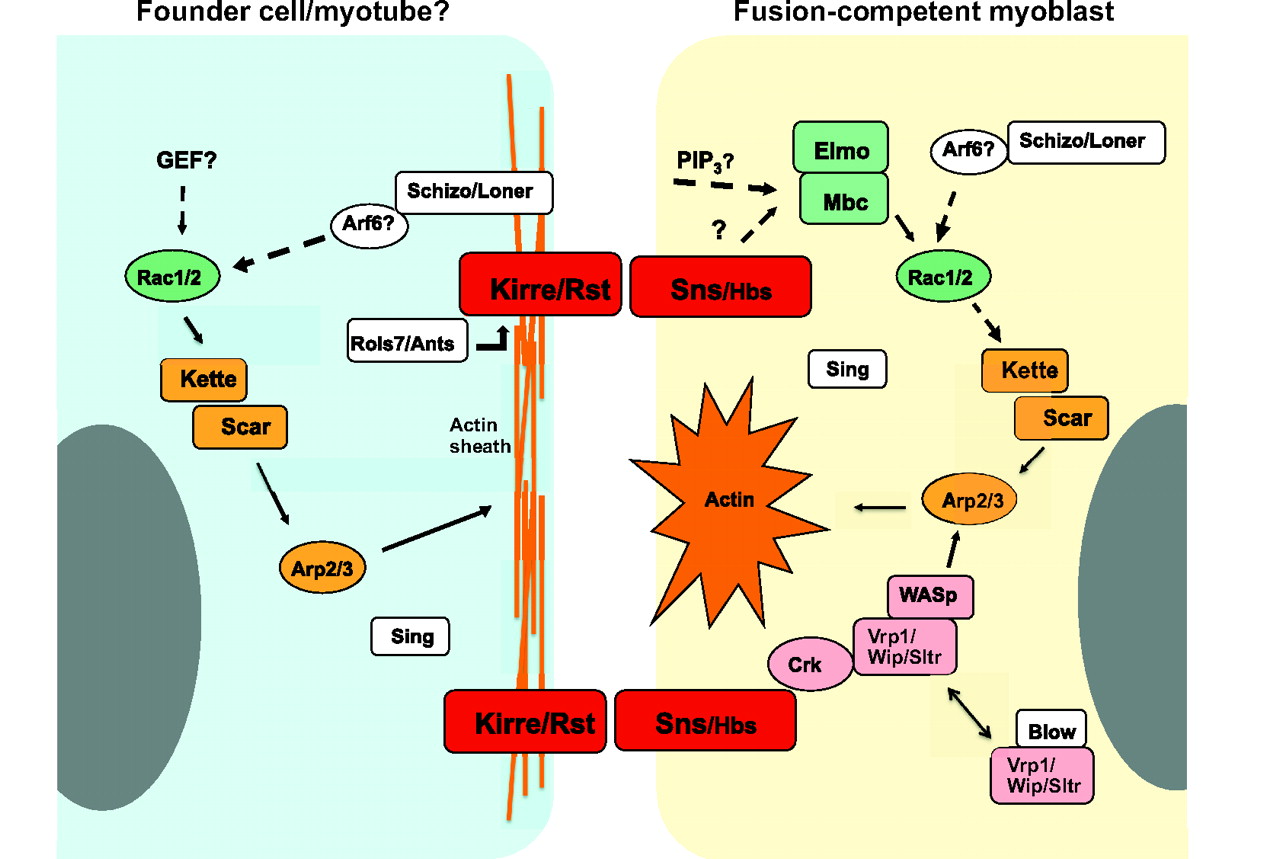
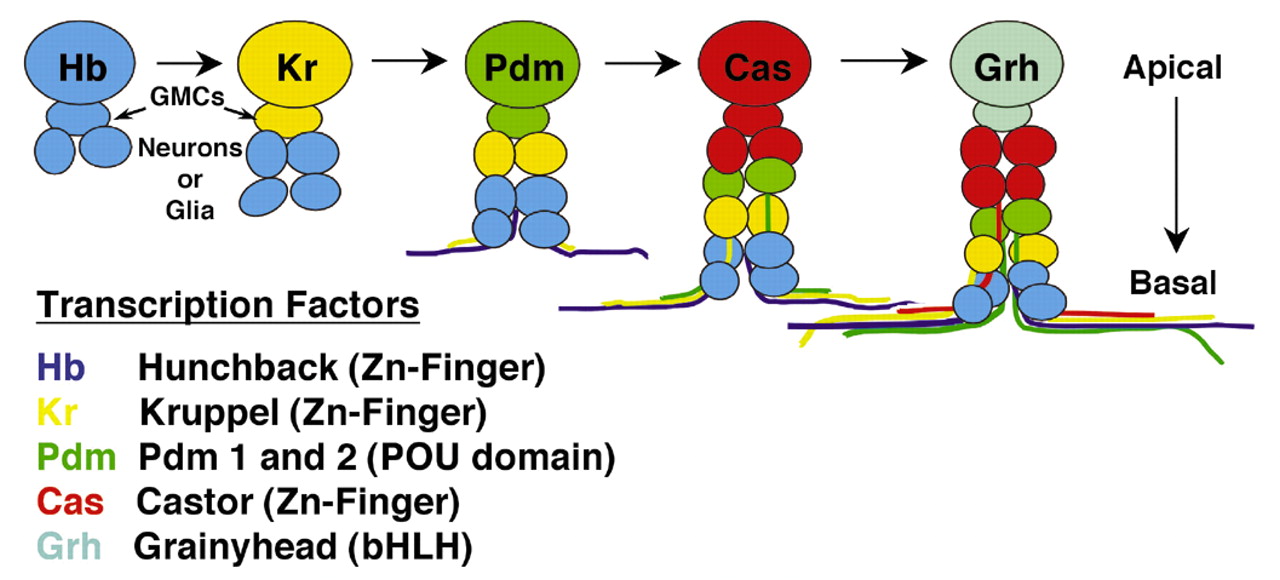
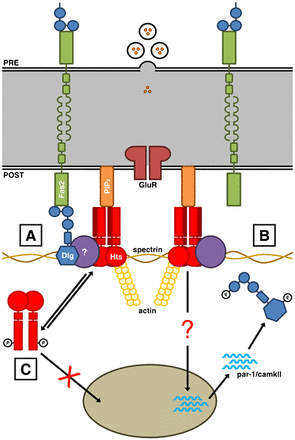
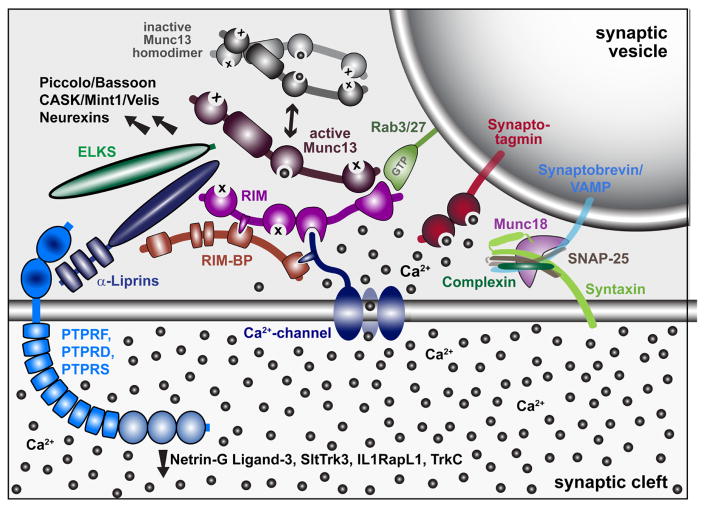
Postsynapse of the larval neuromuscular junction
(A) At the postsynaptic membrane, shown at the bottom, Discs large (Dlg) localizes to spectrin-actin complexes. Homophilic adhesion between Fasciclin 2 (Fas2) transmembrane proteins links the presynaptic and postsynaptic sides, with the intracellular C-terminal domains anchored to the first and second PDZ domains of Dlg. The adducin Hu-li tai shao (Hts) is in a complex with Dlg at the postsynaptic membrane, though the interaction may not be direct. Hts also binds to the lipid Phosphatidylinositol 4,5-bisphosphate (PIP2) via the MARCKS-homology domain. (B) Hts promotes the accumulation of par-1 and camkII transcripts in the muscle cytoplasm through an as of yet identified mechanism. PAR-1 and CaMKII phosphorylate Dlg. Phosphorylation disrupts Dlg postsynaptic targeting. (C) Phosphorylation translocates Hts away from the postsynaptic membrane and hinders Hts' ability to regulate Dlg localization, presumably through the control of PAR-1 and CaMKII at the transcriptional level. Phosphorylation of the MARCKS-homology domain also inhibits Hts' ability to bind to PIP2 (Wang, 2014).
Neto engages the iGluR complexes extrajunctionally and together they traffic and cluster at the synapses, opposite from the active zones marked by T-bars. Neto and the essential iGluR subunits are limiting for formation of functional iGluR complexes at the NMJ and for growth of synaptic structures (Kim, 2014).
Kim, Y. J. and Serpe, M. (2013). Building a synapse: a complex matter. Fly (Austin) 7: 146-152. PubMed ID: 23680998
Wang, S. J., Tsai, A., Wang, M., Yoo, S., Kim, H. Y., Yoo, B., Chui, V., Kisiel, M., Stewart, B., Parkhouse, W., Harden, N. and Krieger, C. (2014). Phospho-regulated Drosophila adducin is a determinant of synaptic plasticity in a complex with Dlg and PIP2 at the larval neuromuscular junction. Biol Open 3: 1196-1206. PubMed ID: 25416060