Select image to enlarge
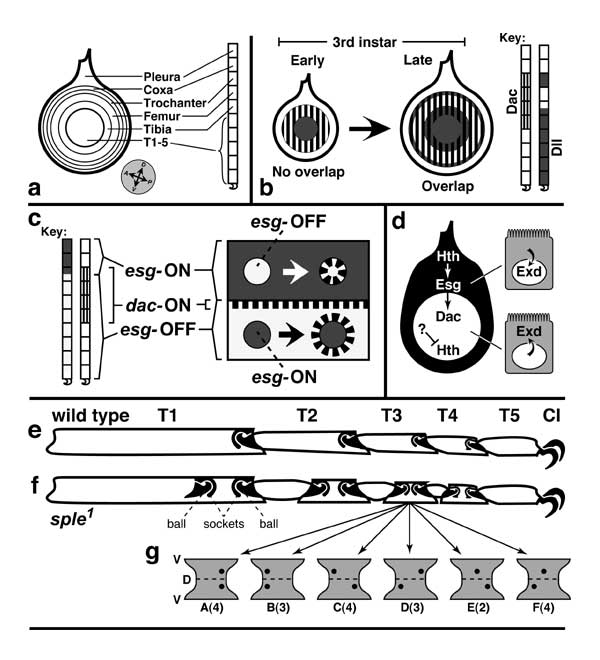
Figure 5.12
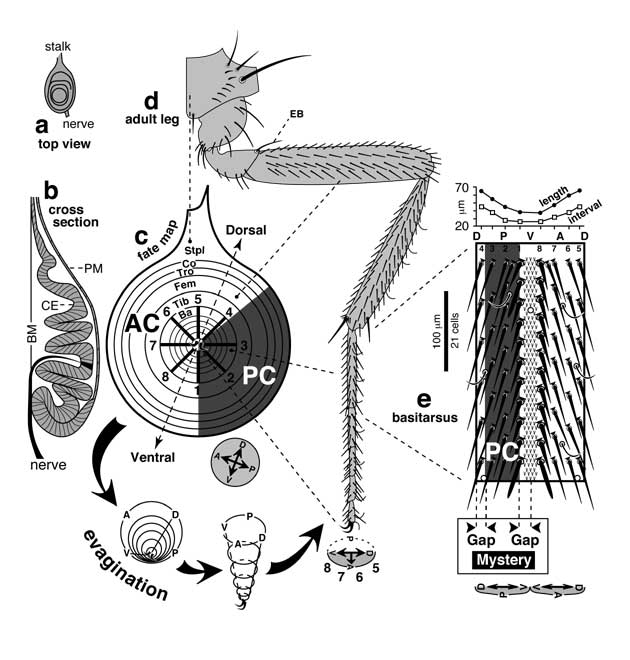
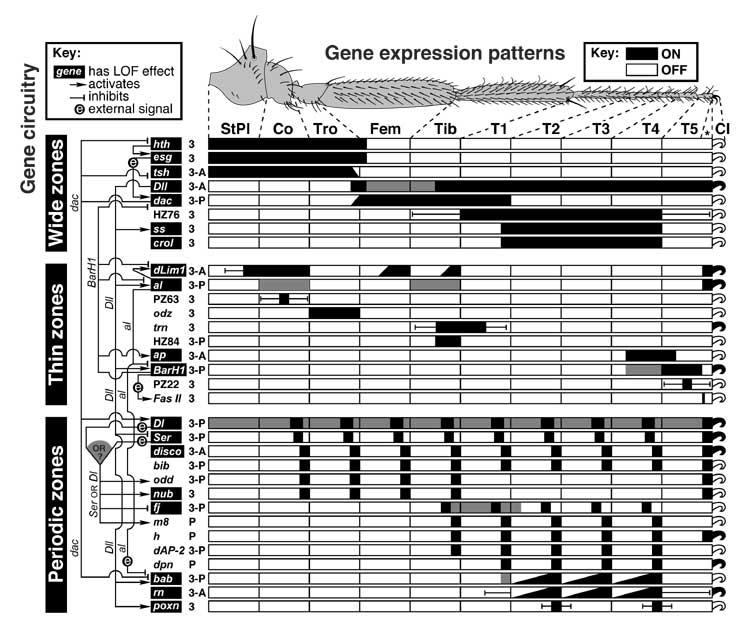
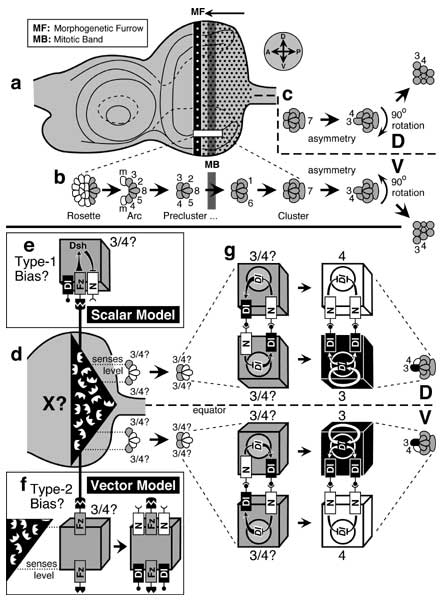
Proximal-distal axis of the leg and how it is subdivided into discrete domains.
a. Blank templates for charting zones of gene expression: disc (left) and derivatives (right) consisting of pleura (body wall) and the leg proper (9 segments; cf. Figs. 5.1 and 5.11). Abbreviations: Dac (Dachshund), Dll (Distal-less), Esg (Escargot), Exd (Extradenticle), Hth (Homothorax).
b. Areas where Dac and Dll are expressed during 3rd instar. The Dll circle initially abuts the Dac ring but later expands to overlap it. (Note also the emergence of an outer ring.) An opposite situation occurs more distally (not shown) where an Aristaless circle and a Bar ring first overlap and later abut [2287].
c. Areas where Dac and Esg are expressed (left) and intercalation of a dac-ON zone wherever esg-ON and esg-OFF cells abut (right). Dac and Esg zones normally overlap slightly (omitted at right). Two clones (circles) are cartooned. When an esgnull clone (above) is induced in esg-ON territory (where Esg and Dac overlap [1573]), its peripheral cells make Dac. When an esgGOF clone (below) arises in esg-OFF territory, Dac is expressed by cells around the clone. Evidently, esg-ON cells emit a signal that causes nearby esg-OFF cells to turn dac ON (Esg —> ? —> dac). An analogous induction (Bar —> contact? —> Fasciclin II) occurs more distally (not shown) [2287].
d. Proximal vs. distal domains of the leg disc. In the proximal area (black; dorsal region exaggerated), hth and esg are expressed (Hth —> esg), and Hth enables Exd to enter the nucleus (Hth —> Exdnuc) [9, 130, 677, 2673, 3589]. In the distal area (white), hth is inhibited jointly by Dpp and Wg so that Exd cannot enter the nucleus [8]. The inhibition involves high doses of Dpp and Wg but apparently is not mediated by Dac or Dll [4760], so its route has been unclear. The likely agent until early 3rd instar is Antennapedia (Antp —| hth), but Antp fades from the endknob by mid-3rd [677].
e, f. Extra tarsal joints caused by mutations in 'planar polarity' genes. T1-T5 are tarsal segments, and 'Cl' denotes claws (dorsal is at top, ventral at bottom). e. Tarsus of wild-type 2nd leg (bristles omitted) showing normal ball-and-socket joints (solid black). Within each joint the ball belongs to the distal segment, while the socket belongs to the proximal one. Pivoting of balls inside sockets lets the tarsus curl ventrally when an apodeme (not shown) is pulled by tibial muscles (like a marionette) [236, 2857, 3421]. The joints arise dorsally [1812] and constrain movement to the D-V plane (like an elbow), though the details of their morphogenesis are unclear [4009]. Note unequal lengths of T1-T5 and flared tip of T4/T5 socket. f. 2nd-leg tarsus from a fly homozygous for the spiny legs1 (sple1) mutation (a.k.a. pksple1) at the prickle locus [1641, 1795, 1810, 2884]. In each segment (except T5 which is immune) there is an extra joint near the midpoint (except T1 where it lies distally and only makes a partial intersegmental membrane). All extra joints are upside down (balls above sockets; cf. the embryo's segment polarity phenotypes [3151]). The extra joint in T4 has a flared socket like the T4/T5 joint, so joints are probably copies of their segment's distal end. Mutations in other planar-polarity genes -- e.g., fz or dsh -- cause milder phenotypes [1810, 4716]. One clue to the etiology is that ommatidia -- like joints -- are ensembles controlled by the same polarity circuit. In that case, the whole proneural group is oriented by a few key cells (cf. Fig. 7.5) [4366], so maybe the same is true for joints.
g. 'Fickle Sensilla Mystery' associated with the double-joint syndrome (cf. Fickle Bristle Mystery; Fig. 7.8). Hourglass shapes are full-surface views of the distal half of T3 from legs of various sple1 individuals. Dorsal (D) and ventral (V) midlines are marked. Black dots are sensilla campaniformia (stretch receptors [4342]). Types of sensillar patterns in a sample of sple1 legs are shown, with numbers of legs per class (A-F) in parentheses (total = 20). In wild-type flies two sensilla always arise at the distal end of T3 (A) and T1 (not shown) [3705]. In the mutants, sensilla must only arise near the normal joints (not near the ectopic ones) since otherwise there would be more than two sensilla. However, they must then be able to move to the extra joint. Normal and extra joints seem to attract sensilla equally (cases of A ≈ B, and C ≈ D), with occasional cases (E or F) where a sensillum cannot seem to decide and hence has been frozen in midsegment.
Panel b is adapted from [2456, 4497, 4575], c depicts data from [1573], d is based on [8], e and f are traced from [1810] (the sple1 tarsus is actually ~10% shorter than the wild-type one), and g depicts data from [1810]. See [883, 1341] for an intercalation scheme.
|
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}