Select image to enlarge
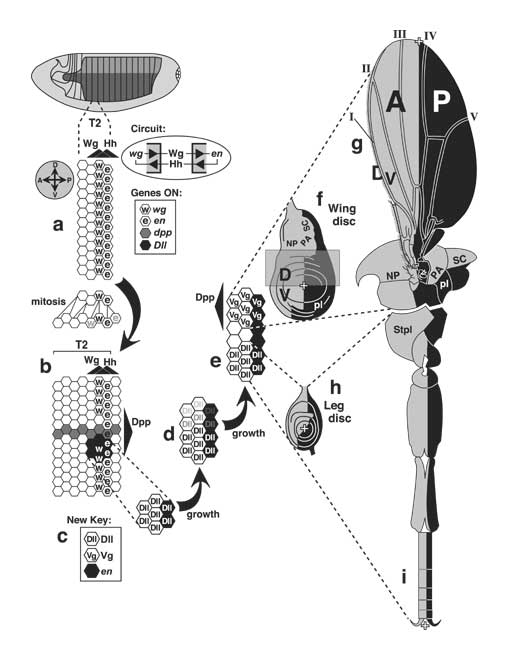
Figure 4.4
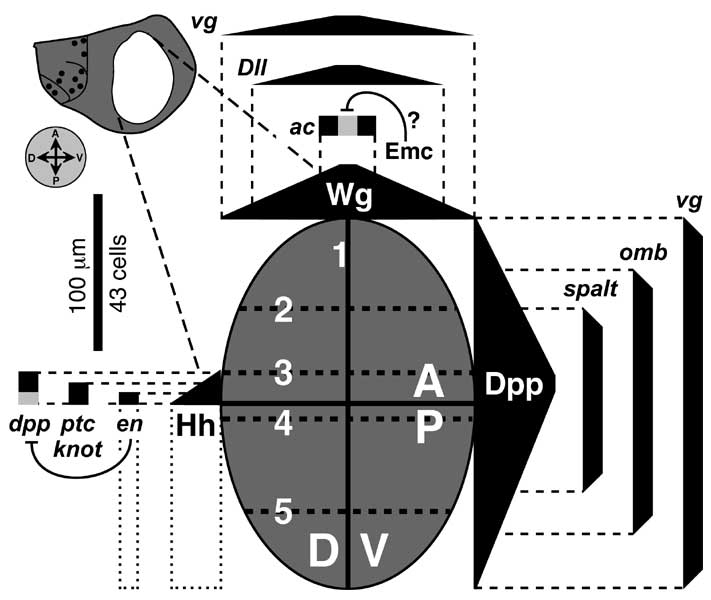
Initiation of thoracic discs at the Wingless/Engrailed interface and the later role of the A/P boundary during disc development. Gene abbreviations: dpp (decapentaplegic), Dll (Distal-less), en (engrailed), hh (hedgehog), vg (vestigial), wg (wingless).
a, b. Magnified view of the embryo's T2 (second thoracic segment, flank area only) at two stages after egg laying (AEL). Axes (A-P, anterior-posterior; D-V, dorsal-ventral) are indicated by compass at left. a. T2 at ~ 5 h AEL (extended germ band, St. 10/11). The posterior two rows of cells (hexagons) transcribe wg or en; key at right) and secrete Wg or Hh. Wg and Hh diffuse (black triangles = concentration gradients) and sustain one another's synthesis by a feedback circuit [2717] that is abridged here (inset at right; see Fig. 2.7 for key and [2816] for theory). Because the gradients are symmetric, each should create two flanking counterparts, but the response is asymmetric due to other genes (not shown) [596, 1628, 3129, 3748]. Before the stage in b, all ectoderm cells divide at least once. The pedigree (lines connect mother to daughter cells) depicts mitoses in one file of cells along the A-P axis, though divisions are not really so orderly. The important point is this: whether a cell expresses Wg or En is not heritable. To wit, as cells move away from the Wg/En interface, they stop making En [4491] and Wg [3363] (fading letters). b. T2 at ~8 h AEL (germ band shortening, St. 12). Wg and En domains do not grow in proportion to segment width (though En stripes widen more than depicted [272, 1064, 2717]) because diffusion ranges of Wg and Hh are fixed [1063]. An A-P stripe of dpp-ON cells crosses a gap in the Wg stripe. Dpp is also a morphogen (gradient at right). Around the tip of the Wg stripe remnant, cells transcribe Dll [834]. The upper limit of the Dll-ON spot is set (at ~St. 11) by Dpp (Dpp ––| Dll). The lower limit is set by Spitz (Spitz ––| Dll) -- a morphogen (not shown; cf. Fig. 6.12) from the V midline [1572, 2335, 3536].
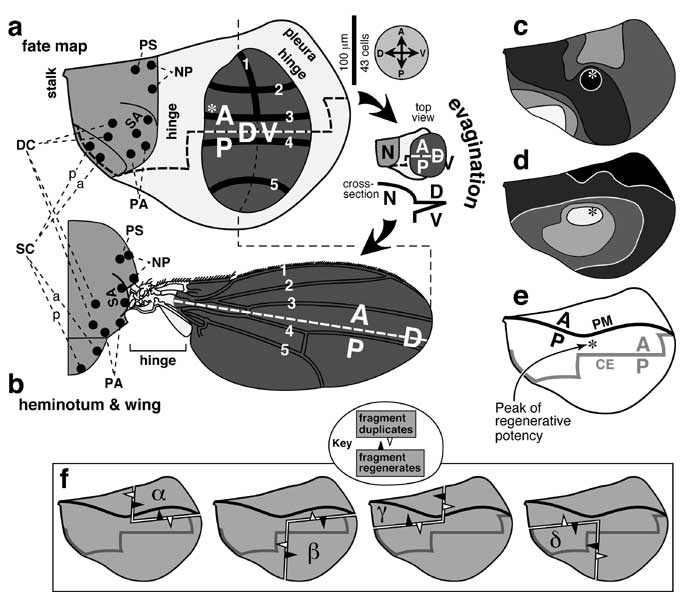
c-e. Subsequent changes in the T2 Dll cluster. Reshaping is apparently due to mitosis (vs. active cell migration) [827, 1571], but cells also rearrange to some extent [1572, 2335]. c. Note the new symbol key. d. As cells move dorsally with the Dpp stripe (see e), they stop making Dll [1571] (fading letters). e. Eventually a dorsal cohort starts transcribing vg -- a wing (and haltere) marker. Thus, wing and leg anlagen arise from a common pool of cells [827]. The pool is partitioned (St. 15) based on how much Dpp (D signal) the cells perceive relative to Spitz (V signal; not shown) [2335]:
High Dpp/Spi ratio --> wing state (vg-ON/Dll-OFF)
Low Dpp/Spi ratio --> leg state (vg-OFF/Dll-ON)
f-i. Mature (3rd-instar) discs and their adult derivatives, showing regions that do (black) or do not (gray) make En (oriented as per compass in a). Within both discs, the en ON/OFF boundary becomes a reference line for specifying cell positions along the A-P axis, using either Wg or Dpp as a morphogen (not shown). In the leg disc, a remnant of wg-ON cells is retained ventrally, and a stripe of dpp-ON cells is induced dorsally (cf. Fig. 5.4). In the wing disc a stripe of dpp-ON cells is induced along the entire boundary (cf. Fig. 6.3). Plusses mark the appendage tips, which would come out of the page during eversion and then flop down. f. Left wing disc. Tissue in the translucent rectangle (D wing surface) is not visible in g because it folds behind (cf. Fig. 6.1). Eversion brings the notum (NP, PA, and SC = notopleural, postalar, and scutellar areas) into contact with the pleura (pl). g. Adult wing (V surface only) and heminotum in side view. Bristles are omitted except at wing margin. A and P denote lineage compartments. The A edge of the en-ON region per se is actually closer to vein III due to a shift during late 3rd instar (cf. Fig. 6.7d) [350]. h. Left 2nd-leg disc. i. Adult leg, drawn as if fileted along the ventral A/P boundary and spread flat so that its whole surface is visible. Thoracic structures also come from the leg disc, including StPl (sternopleural sclerite). Holes in joints are cartographic artifacts due to odd shapes of proximal leg segments.
Panel a is based on [1063]; b-e are adapted from [834, 884, 1571]; f is traced from [2467]; g is modified from [1376, 4076]; h is sketched from stained discs pictured in [495, 2754]; and i is amalgamated from [1800, 2449, 4076].
N.B.: Cells are not usually packed so neatly [1064], nor are they so constant in shape or size [4743]. Indeed, they may not even stay in a monolayer [1572]. In b, dpp transcription is actually greater posteriorly along its stripe (possibly due to upstream control by segment-polarity genes) [2003]. In c-e a ring of escargot-ON cells (not shown; future pleura and coxa) surrounds the Dll-ON core (all other leg segments) [1542, 1572, 1573]. Unlike the leg disc whose axes persist from the blastoderm, the wing disc appears to rotate ~90° during development (not shown) [4652]. As shown in h, the A/P (en-ON/OFF) line of the leg disc is diagonal near the center but zigzags toward the stalk above [495, 3747, 4254]. This irregularity has led to confusion since some authors schematize the A/P line vertically (D pole at 12 o'clock) like the outer part, while others slant it (D pole at 1 or 2 o'clock) like the central part (cf. Fig. 5.1).
|
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}