Select image to enlarge
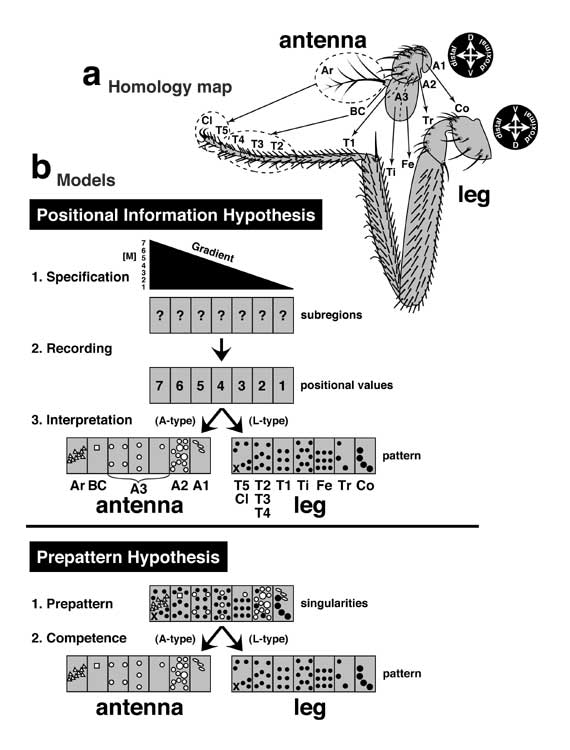
Figure 4.3
Demise of the Prepattern Hypothesis in 1969 and ascendancy of the Positional Information Hypothesis, due in part to their clumsy or elegant explanations for homeotic autonomy.
a. Correspondence (arrows) between antennal and leg parts, as deduced from AntennapediaR (AntpR) flies, whose antennae are patchily transformed into 2nd legs [3445]. Abbreviations: A1-3 (antennal segments 1-3), BC (basal cylinder), Ar (arista), Co (coxa), Tr (trochanter), Fe (femur), Ti (tibia), T1-5 (tarsal segments 1-5), Cl (claws). Compasses denote adult polarities (D = dorsal, V = ventral; the inversion is explained below). Antennae of AntpR flies (not shown) have patches of leg tissue. Whenever leg tissue arises at a particular proximal-distal level, it makes leg structures that are appropriate for that level. The implication is that these leg cells can somehow figure out their location along this axis of the antenna. Wolpert and Stern agreed that the antenna and leg probably share some kind of ground plan that both types of cells can read. They disagreed over the kind of ground plan. In Wolpert's scheme it is a gradient; in Stern's, it is a prepattern.
b. The two models. Each of the 7 gray rectangles represents a segmental region (eventually containing 102-103 cells) of the antennal or leg rudiment along the proximal-distal axis.
Lewis Wolpert's Positional Information (PI) Hypothesis explained antenna-to-leg homeosis in terms of a shared coordinate system [4724]. Antennal and leg cells are supposed to use the same positional signal -- a diffusible molecule, M. This morphogen is produced at one end of the organ (left) and diffuses to form a concentration gradient (triangle). The gradient could be linear (as shown) but is more likely exponential. In Step 1, cells assess their distance from the source by measuring the amount of M: the more M that a cell 'tastes' at its site, the nearer it 'thinks' that it must be to the source. In Step 2, cells record these levels (1-7) as positional values (ON or OFF states of memory genes?) that persist after M vanishes. In Step 3, cells translate these values into structures that suit the disc to which they belong. For example, antennal cells interpret the number 7 as arista, while leg cells interpret it as T5 and claws. The interpretation mode that a cell uses (A vs. L) is predetermined by the turning ON or OFF of a particular homeotic gene in one of the cell's ancestors. Mutations in that gene cause homeosis: mutant cells think that they are leg cells, but they can still sense M properly, so they make leg structures that are appropriate for their proximal-distal level in the antenna. Finer-grain patterning within subregions may rely on a second echelon of (segmental) gradients (not shown) [386]. The elegance of Wolpert's model has been its universality: the same gradients can theoretically pattern all of the discs [20, 523, 3701].
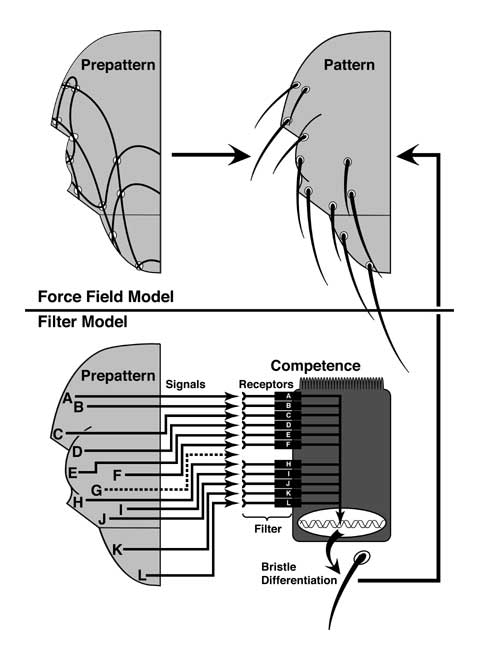
Curt Stern's Prepattern Hypothesis had a more difficult time explaining homeosis. Its basic assumption was that every pattern element is preceded by a discrete signal (singularity) [4095]. Unlike the morphogen, these signals (circles, ovals, square, triangles, and x) differ qualititively: they are not graded. In Step 1, the disc lays out an array of these signals (the prepattern). The autonomy of the AntpR mosaicism implied that antenna and leg must express one another's signals. (How else could nouveau-leg cells that find themselves stranded in the antenna figure out what structures to make?) In other words, both prepatterns must coexist in each disc [4346]. Antennal cells would normally read one subset of signals (white symbols), while leg cells read the other (black symbols). The filtering of these subsets (cf. Fig. 3.2) would depend on a cell's competence, which in turn would be determined early in development when one of the cell's ancestors turned a particular homeotic gene ON or OFF. Mutations in such genes would lead to inter-disc transformations (cf. Ch. 8). This argument was tolerable for a single type of transformation (e.g., antenna-to-leg), but the large number of inter-disc homeoses [3214] implied that every disc must contain the prepattern for virtually every other disc. While not impossible, this notion seemed implausibly clumsy -- especially when contrasted with how easily the PI Hypothesis handled this same phenomenon.
Panel a is redrawn from [3447] which depicts a right antenna and left leg (see [1587] for A3 subregions, [1516, 1561, 2287] for molecular homology, and [4522] for a different map). Panels b and c are based on the ideas of Stern (Fig. 58 in [4100]) and Wolpert [4734], as contrasted inter se by Bryant [526] and Tokunaga [4346]. Black or white symbols in the antennal and leg schematics are merely abstract representations of the actual patterns. See Figure 8.3 for recent data on how antennal vs. leg identities are controlled. See [2956, 3744] for the origin of the gradient concept. See also App. 7.
|
|
{kind=link}
{kind=link}