InteractiveFly: GeneBrief
sidekick: Biological Overview | References
|
Gene name - sidekick
Synonyms - Cytological map position - 1C2-1C3 Function - Ig domain receptor Keywords - accumulates in specific synaptic layers of the developing motion detection circuit, necessary for normal optomotor behavior, adherens junctions, tricellular junction protein, regulates vertex dynamics to promote bicellular junction extension |
Symbol - sdk
FlyBase ID: FBgn0021764 Genetic map position - chrX:686,834-749,496 NCBI classification - Immunoglobulin domain, Fibronectin type III domain Cellular location - surface transmembrane |
| Recent literature | Finegan, T. M., Hervieux, N., Nestor-Bergmann, A., Fletcher, A. G., Blanchard, G. B. and Sanson, B. (2019). The tricellular vertex-specific adhesion molecule Sidekick facilitates polarised cell intercalation during Drosophila axis extension. PLoS Biol 17(12): e3000522. PubMed ID: 31805038
Summary: In epithelia, tricellular vertices are emerging as important sites for the regulation of epithelial integrity and function. Compared to bicellular contacts, however, much less is known. In particular, resident proteins at tricellular vertices were identified only at occluding junctions, with none known at adherens junctions (AJs). A previous study discovered that in Drosophila embryos, the adhesion molecule Sidekick (Sdk), well-known in invertebrates and vertebrates for its role in the visual system, localises at tricellular vertices at the level of AJs. This study surveyed a wide range of Drosophila epithelia and establish that Sdk is a resident protein at tricellular AJs (tAJs), the first of its kind. Clonal analysis showed that two cells, rather than three cells, contributing Sdk are sufficient for tAJ localisation. Super-resolution imaging using structured illumination reveals that Sdk proteins form string-like structures at vertices. Postulating that Sdk may have a role in epithelia where AJs are actively remodelled, the phenotype of sdk null mutant embryos during Drosophila axis extension was analyzed using quantitative methods. Apical cell shapes were found to be abnormal in sdk mutants, suggesting a defect in tissue remodelling during convergence and extension. Moreover, adhesion at apical vertices is compromised in rearranging cells, with apical tears in the cortex forming and persisting throughout axis extension, especially at the centres of rosettes. Finally, it was shown that polarised cell intercalation is decreased in sdk mutants. Mathematical modelling of the cell behaviours supports the notion that the T1 transitions of polarised cell intercalation (referring to the rearrangement of groups of four cells) are delayed in sdk mutants, in particular in rosettes. It is proposed that this delay, in combination with a change in the mechanical properties of the converging and extending tissue, causes the abnormal apical cell shapes in sdk mutant embryos. |
The assembly of functional neuronal circuits requires growth cones to extend in defined directions and recognize the correct synaptic partners. Homophilic adhesion between vertebrate Sidekick proteins promotes synapse formation between retinal neurons involved in visual motion detection. This study shows that that Drosophila Sidekick accumulates in specific synaptic layers of the developing motion detection circuit and is necessary for normal optomotor behavior. Sidekick is required in photoreceptors, but not their target lamina neurons, to promote the alignment of lamina neurons into columns and subsequent sorting of photoreceptor axons into synaptic modules based on their precise spatial orientation. Sidekick is also localized to the dendrites of the direction-selective T4 and T5 cells, and is expressed in some of their presynaptic partners. In contrast to its vertebrate homologues, Sidekick is not essential for T4 and T5 to direct their dendrites to the appropriate layers or to receive synaptic contacts. These results illustrate a conserved requirement for Sidekick proteins to establish visual motion detection circuits that is achieved through distinct cellular mechanisms in Drosophila and vertebrates (Astigarraga, 2018).
Establishing synaptic circuits requires neurons to identify and interact with the correct partners within a highly complex cellular environment. These interactions depend on the combinatorial actions of a wide variety of cell-surface adhesion molecules. Proteins of the cadherin, neurexin, leucine-rich repeat and immunoglobulin (Ig) superfamilies can engage in homophilic or heterophilic interactions, leading either to adhesion or repulsion of the expressing cells. It is not yet clear how such molecular interactions enable the assembly of complex neural circuits with high specificity (Astigarraga, 2018).
The Drosophila visual system has been used as a model system for many studies of the development and function of neural circuits. Each ommatidium in the compound eye contains eight photoreceptors: R1-R6 express rhodopsin Rh1 and detect visual motion, while R7 and R8 each express one of four different rhodopsins and mediate color vision. Motion detection is subdivided into 'ON' and 'OFF' pathways that are specialized to detect moving bright edges and moving dark edges, respectively. Lamina neurons L1 and L2 receive direct synaptic input from R1-R6; L1 provides input to the 'ON' pathway, while L2 is the major link to the 'OFF' pathway. Information from the lamina is conveyed to the medulla, where the 'ON' and 'OFF' signals are transmitted by distinct populations of neurons that converge on the dendrites of T4 ('ON') and T5 ('OFF') lobula plate neurons in specific layers of the medulla and lobula, respectively. Four subtypes of T4 and T5 neurons are each sensitive to one of the four cardinal directions of motion and provide input to lobula plate tangential neurons that are segregated in four corresponding layers (Astigarraga, 2018).
In the third larval instar, Hedgehog and Spitz signals from the R1-R6 photoreceptor axons induce the final division and neuronal differentiation of their target lamina neurons. Interactions between the Ig family members Hibris (Hbs) on postmitotic lamina neurons and Roughest (Rst) on photoreceptor axons then induce the lamina neurons to line up in columns along the retinal axon bundles. The R1-R6 axons from a single ommatidium subsequently diverge during pupal development to project to six different synaptic modules known as lamina cartridges, enabling each cartridge to collect information from six photoreceptors located in different neighboring ommatidia that view the same point in visual space (Agi, 2014). This sorting process requires the early polarization of the 'front' of each photoreceptor growth cone towards its target, while the 'heels' of these growth cones form a stable scaffold. Correct polarization and extension are thought to require a comparison of the relative levels of the cadherin Flamingo (Fmi) on neighboring growth cones, as well as adhesive interactions with other growth cones and target lamina neurons that are mediated by N-cadherin (Ncad). Photoreceptor axons later develop synaptic terminals that connect to their target lamina neurons through tetrad synapses, at which a single presynaptic active zone contacts four postsynaptic cells. Two of these are always the lamina neurons L1 and L2, while the others may be L3 in combination with amacrine or glial cells (Astigarraga, 2018).
The vertebrate retina is comparable in complexity and organization to the combined Drosophila retina, lamina and medulla, and the two visual systems share many design principles. In the chick retina, synapses between different types of bipolar, amacrine and ganglion cells form in distinct sublaminae of the inner plexiform layer that are specified by homophilic adhesion molecules of the Sidekick (Sdk), Down's syndrome cell adhesion molecule (Dscam) and Contactin families. Each of these proteins is localized to a specific layer and expressed by cells that synapse on each other in that layer. In the mouse retina, sidekick cell-adhesion molecules 1 and 2 (Sdk1 and Sdk2) label largely non-overlapping subsets of cells, but these are not clearly separated by layer. Sdk2 is required in both VG3 amacrine cells and W3B retinal ganglion cells to form a strong synaptic connection between these neurons, which sense object motion relative to the surroundings (Krishnaswamy, 2015; Astigarraga, 2018 and references therein).
The Drosophila genome encodes a single Sdk protein. Null sdk alleles have previously been shown to produce small changes in the numbers of photoreceptors and non-neuronal cells in the eye (Nguyen, 1997). This study shows that Sdk is a homophilic adhesion molecule that is expressed in several cells of the visual motion detection circuit and is localized to specific synaptic layers. Consistent with its location, mutants lacking Sdk show defects in optomotor behavior. sdk was found to be required in photoreceptors, but not in their target lamina neurons, for the correct organization of lamina columns and cartridges, indicating that, unlike its vertebrate counterparts, Sdk does not mediate adhesion between pre- and post-synaptic cells. Although Sdk is present on the dendrites of the direction-selective T4 and T5 cells, it is not required for these neurites to arborize in the appropriate layer or to receive synaptic input. These results suggest that Sdk proteins in Drosophila and vertebrates have a common functional role in establishing visual motion detection circuits, but act through distinct cellular mechanisms (Astigarraga, 2018).
This study shows that Drosophila Sdk is required to establish a functional pathway for visual motion detection. Defective optomotor behavior in sdk mutant flies appears to result from defects in the organization of connections between R1-R6 photoreceptors and their target lamina neurons. Sdk acts in photoreceptors both to organize lamina neurons into columns and to enable photoreceptor axons to sort to the correct lamina cartridges. Sdk is also expressed in T4 and T5 neurons, the output cells of the 'ON' and 'OFF' motion detection circuits; however, no morphological or functional defects were observed in these cells in the absence of sdk (Astigarraga, 2018).
In the chicken retina, it was proposed that the homophilic adhesion molecules Sdk1 and Sdk2 each drive neurons to arborize and to form synapses with other neurons that express the same Sdk in a specific sublamina of the inner plexiform layer (Yamagata, 2002). Drosophila Sdk is also a homophilic adhesion molecule. However, because it is only required in photoreceptors, Sdk-Sdk binding cannot directly mediate the interaction of lamina neurons with photoreceptor axons. The simplest explanation for the presence of lamina neurons beneath the R1-R6 growth cone layer in sdk mutants or when sdk function is removed from the eye is that Sdk-mediated adhesion between photoreceptor growth cones normally forms a barrier to the movement of lamina neurons. An alternative possibility is that Sdk on photoreceptors might interact with a heterophilic binding partner on lamina neurons. The existence of a splice form of mouse Sdk1 that lacks the first two Ig domains (Kaufman, 2004), which are necessary for homophilic adhesion (Goodman, 2016; Hayashi, 2005), suggests the possibility of functions other than homophilic binding. The only cell-adhesion molecules known to promote interactions between lamina neurons and photoreceptor axons are Rst and Hbs, Ig superfamily members that are homologous to vertebrate Neph and Nephrin proteins, respectively. Loss of Sdk does not affect the ability of lamina neurons to associate with photoreceptor axons to the same extent as loss of Rst or Hbs, but it is possible that Sdk might modulate the interaction between them. Such an effect might explain why human Sdk1 upregulation contributes to the pathology of kidney diseases such as HIV-induced nephropathy and focal segmental glomerulosclerosis, as interacting Neph and Nephrin molecules form the slit diaphragm in the kidney (Astigarraga, 2018).
Homophilic adhesion is also a likely explanation for the requirement for Sdk in photoreceptor axon sorting. Sorting of the axons from a single ommatidium to six different lamina cartridges requires R1-R6 growth cones to polarize such that their mobile 'fronts' move away from their stable 'heels' at characteristic angles (Schwabe, 2013; Langen, 2015). The pattern of Sdk labeling in five puncta, two of which arise from and are adjacent to R4, is consistent with a location at the contact points between the six heels, which are arrayed in a similar crescent (Langen, 2015). Two cadherins, Fmi and Ncad, have previously been shown to affect photoreceptor growth cone extension and sorting in the lamina. Although they have partially redundant roles, Ncad functions largely autonomously to mediate interactions with target cells and with photoreceptor growth cones within and across ommatidia, whereas a comparison of Fmi levels between neighboring growth cones appears to promote normal polarization. It is suggested that Sdk mediates adhesion between growth cone heels, enabling them to form a stable scaffold that may facilitate the comparison of Fmi levels or provide mechanical resistance necessary for polarized extension (Astigarraga, 2018).
In contrast to knockdown studies in the chick retina, loss of Drosophila sdk does not alter the layer-specific arborization patterns of the Sdk-expressing neurons examined in this study. Sdk is localized to the dendrites of T4 and T5, and is present in at least one of their presynaptic partners. Nonetheless, T4 and T5 still extend their dendrites in the correct layers and form arbors of the normal size and shape in the absence of sdk. In the mouse retina, Sdk2 controls the strength of synaptic connections between Sdk2-expressing neurons that detect motion of the central visual field relative to the surround (Krishnaswamy, 2015). This study finds that the Sdk-expressing neuron Tm9 does not require sdk to form synapses with T5, as shown both by punctate localization of the active zone protein Brp at its axon terminals and by activity-dependent synaptic GRASP. It is possible that Sdk is required for synapse formation by other neurons that are presynaptic to T4 or T5, or that it recruits specific postsynaptic proteins to these synapses. Nevertheless, removing sdk function from T4 and T5 does not result in significant defects in optomotor behavior, arguing against an essential role for sdk in these cells (Astigarraga, 2018).
It has been suggested that the fly and vertebrate visual systems share common design principles reflecting deep evolutionary homology. The structural organization of the vertebrate retina resembles the Drosophila retina, lamina and medulla, and the computational mechanisms underlying motion detection also have many common features despite anatomical and molecular differences in the circuits. The current developmental and behavioral studies demonstrate that Sdk shares a physiological function in visual motion detection with its vertebrate counterparts, although their cellular mechanisms of action appear to differ. The functions of other molecules have also diverged between Drosophila and vertebrates; for example, the extensive alternative splicing of Drosophila Dscam1 allows it to mediate self-avoidance during growth and synaptogenesis, contrary to the adhesive function of chick Dscams. Self-avoidance in the mouse instead depends on clustered protocadherins, which achieve diversity through promoter duplication. It has been proposed that the formation of visual motion detection circuits in vertebrate and invertebrate visual systems is controlled by a genetic network derived from a common ancestor. The data suggest that evolutionary changes may include the repurposing of common elements of such a network to mediate distinct functions (Astigarraga, 2018).
Remodeling of cell-cell junctions drives cell intercalation that causes tissue movement during morphogenesis through the shortening and growth of bicellular junctions. The growth of new junctions is essential for continuing and then completing cellular dynamics and tissue shape sculpting; however, the mechanism underlying junction growth remains obscure. This study investigated Drosophila genitalia rotation where continuous cell intercalation occurs to show that myosin II accumulating at the vertices of a new junction is required for the junction growth. This myosin II accumulation requires the adhesive transmembrane protein Sidekick (Sdk), which localizes to the adherens junctions (AJs) of tricellular contacts (tAJs). Sdk also localizes to and blocks the accumulation of E-Cadherin at newly formed growing junctions, which maintains the growth rate. It is proposed that Sdk facilitates tAJ movement by mediating myosin II-driven contraction and altering the adhesive properties at the tAJs, leading to cell-cell junction extension during persistent junction remodeling (Uechi, 2019).
To generate tissue shapes, cell collectives show various dynamics, such as cell division and cell deformation. Among them, cell intercalation is a multicellular behavior in which cells change their position through the remodeling of cell-cell contacts, leading to the directional elongation and expansion of tissues across species. Especially in epithelia, this cell-cell junction remodeling involves the shortening and loss of bicellular junctions and the subsequent growth of bicellular junctions in a new direction. Junction shortening initiates tissue dynamics and is driven in a conserved manner by contractile forces generated by actomyosin (actin and non-muscle myosin II complex) associating with the cadherin-catenin core complex, including E-Cadherin, β-Catenin, and other related adherens junction (AJ) components, at the AJs of shortening junctions. Junction growth is also essential for continuing and then completing cellular dynamics and tissue shape sculpting. Several studies using flies have suggested that myosin II has a role in junction growth during developmental events. In the germ band, medial pulses of myosin II in the cells surrounding junctions and toward the posterior ectoderm regulate junction growth during cell intercalation-driven convergent extension [germ band extension (GBE)]. A similar contribution of myosin II pulses in the surrounding cells to junction extension is also observed in the apical cell oscillation of amnioserosa cells during dorsal closure. In developing wing epithelia, a decrease in myosin II levels at newly formed junctions facilitates junction growth to organize the epithelial cellular pattern. However, despite its importance, the mechanisms underlying junction growth remain unclear, in contrast to junction shortening (Uechi, 2019).
Previous studies demonstrated that cell intercalation also contributes to the tissue rotational movement observed for Drosophila male genitalia. The fly genitalia are located at the animal's posterior end, and the male genitalia are surrounded by epithelia known as the A8 segment at the anterior side. At 24 h after puparium formation (APF), the genitalia and the A8 epithelia begin dextral rotation that terminates at around 36 h APF. The rotation consists of an initial 180° movement of the posterior compartment of A8 (A8p) along with the genitalia and a subsequent 180° movement of the anterior component of A8 (A8a), the latter of which starts at around 26 h APF. From 26 h APF in the A8a cells, myosin II accumulates to a greater extent at AJs, forming a right oblique angle with the anterior-posterior (AP) axis than at junctions forming a left oblique angle. This polarized myosin II distribution gives rise to right-biased junction shortening in relation to the AP axis and leads to left-right asymmetric cell intercalation, which is persistently observed during the movement. By combining numerical simulations, it was demonstrated that this repeated junction remodeling in the confined space generates the A8a movement. In this movement, newly formed junctions are sufficiently elongated within a certain time frame to execute the next round of cell intercalation. Incomplete genitalia rotation leads to male sterility (Uechi, 2019).
This study performed time-lapse imaging, developed an optogenetic tool, and analyzed the adhesive protein Sidekick (Sdk), which is known to regulate retinal development in flies and mice and showed that myosin II accumulating at the tricellular contacts (tAJs) of growing junctions is required for bicellular junction growth in A8a cells. Also, Sdk regulates the myosin II and E-Cadherin distributions at the tAJs, thereby maintaining the junction growth rate. These findings suggest that the tAJ is a specialized point promoting cell-cell junction extension (Uechi, 2019).
The process of junction shortening is well characterized and is organized by the contractile forces of actomyosin, which is transmitted to cell-cell contacts via AJ components, such as E-Cadherin. These proteins have important roles in the dynamics of multicellular deformation. Since junction formation and growth are important for the continuation and completion of multicellular dynamics and tissue architecture shaping, it is likely that active mechanisms underlie the extension of cell-cell junctions. Indeed, recent reports suggest that actin and myosin II at the bicellular junctions are involved in the junction extension in cell rearrangement and in cell-shape formation during Drosophila wing and eye development. Polarized medial pulses of myosin II in the cells surrounding junctions regulate junction extension in the Drosophila germ band and amnioserosa. This study used an optogenetic tool that allows for the spatiotemporal inactivation of endogenous myosin II and revealed that myosin II accumulating at the tAJs of newly formed junctions is required for junction growth in the A8a epithelia. This study also demonstrated that the myosin II accumulation and junction growth require the tAJ-localizing protein Sdk. Thus, this report that tAJs are an additional point promoting the extension of bicellular junctions (Uechi, 2019).
Sdk transiently localizes to newly formed junctions as well as tAJs, causing a downregulation of E-Cadherin and a slight increase in intercellular spaces at the AJs of growing junctions, indicative of less tight cell-cell contacts. Recent studies in zebrafish showed that the presence of extracellular spaces and the disassembly of cell-cell contacts contribute to fluidize tissues. During body axis elongation, the extracellular spaces render mesodermal cells fluidized and uncaged and associated with large fluctuations in the lengths of cell-cell contact. Decreases in cell-cell contacts through the destabilization of junctional E-Cadherin, accompanied by an increase in extracellular spaces, induces the fluidization of blastoderm cells and consequently allows blastoderm spreading at the onset of morphogenesis. Analogous to these properties, it is possible that the presence of Sdk at growing junctions confers flexible dynamics to the cell-cell contacts at the level of the AJs of the growing junctions. This study proposes mechanisms of junction growth in which Sdk has dual roles. First, Sdk mediates a driving force of junction growth by anchoring myosin II at tAJs; the contractility of the actomyosin then retracts the membrane of the surrounding cells at tAJs. Second, Sdk assists in the myosin II-driven junction growth by localizing to and decreasing the accumulation of E-Cadherin at the growing junctions and their tAJs; this composition of E-Cadherin and Sdk causes contacts between the vertices of the surrounding cells and the cells forming the growing junction to be less tight. Such adhesion can render the tAJs of growing junctions more sensitive to contractile forces at the vertices of the surrounding cells, supporting the retraction of the membrane of the surrounding cells at tAJs. The latter mechanism is indeed likely to contribute to junction growth since inducing sdk RNAi only in the cell forming the growing junction was sufficient to reduce the junction growth rate, even when the surrounding cells consisted of WT cells (Uechi, 2019).
The precise mechanism by which Sdk blocks the accumulation of E-Cadherin at newly formed junctions is still unclear. While Sdk was already present at growing junctions from the step of four-way vertex resolution, E-Cadherin would be newly recruited to the growing junctions since E-Cadherin is removed from remodeling junctions by endocytosis during junction shortening. A recent study using fluorescence recovery after photobleaching (FRAP) revealed two ways that E-Cadherin is re-distributed to cell-cell junctions, lateral diffusion within the plasma membrane and delivery from the cytoplasm by vesicular trafficking. Since (1) E-Cadherin and Sdk did not interact despite their localization to AJs, (2) showed complementary distributions at newly formed junctions and even at cellular edges in S2 cells where they were ectopically expressed, and (3) changed their distributions when the other protein was depleted, it is possible that there are repelling forces between E-Cadherin and Sdk molecules, which cause them to exclude each other and may delay the diffusion of E-Cadherin from neighboring junctions into newly formed junctions, where Sdk is already enriched. However, this study does not exclude another possibility that Sdk inhibits machineries that deliver E-Cadherin from the cytoplasm, such as blocking their access to growing junctions or biochemically inactivating them (Uechi, 2019).
This study observed the accumulation of myosin II and decreased E-Cadherin levels at the tAJ of growing junctions. These distributions resemble those occurring during new cell-cell contact formation between daughter cells in epithelia. After cytokinesis, myosin II accumulates at the edges of new cell-cell junctions in the neighboring cells of the daughter cells, in response to the local decrease in E-Cadherin levels at these edges, which participates in new cell-cell junction formation. These reports and the current observations suggest a possible common mechanism underlying new cell-cell contact formation among epithelial multicellular behaviors. Although the dynamics and roles of Sdk in cell division are still unclear, an intriguing possibility is that Sdk regulates the dynamics of new cell-cell junctions in concert with myosin II and E-Cadherin not only in the context of cell intercalation but also global epithelial dynamics including cytokinesis (Uechi, 2019).
Tricellular adherens junctions are points of high tension that are central to the rearrangement of epithelial cells. However, the molecular composition of these junctions is unknown, making it difficult to assess their role in morphogenesis. This study shows that Sidekick, an immunoglobulin family cell adhesion protein, is highly enriched at tricellular adherens junctions in Drosophila. This localization is modulated by tension, and Sidekick is itself necessary to maintain normal levels of cell bond tension. Loss of Sidekick causes defects in cell and junctional rearrangements in actively remodeling epithelial tissues like the retina and tracheal system. The adaptor proteins Polychaetoid and Canoe are enriched at tricellular adherens junctions in a Sidekick-dependent manner; Sidekick functionally interacts with both proteins and directly binds to Polychaetoid. It is suggested that Polychaetoid and Canoe link Sidekick to the actin cytoskeleton to enable tricellular adherens junctions to maintain or transmit cell bond tension during epithelial cell rearrangements (Letizia, 2019).
Epithelial cells are linked together by multiple types of junctions that must be dynamically rearranged as the cells change their relative positions during tissue remodeling. Cadherin-based adherens junctions (AJs) are essential for cells to sense and respond to the mechanical forces that drive such rearrangements, while tight junctions (TJs) in vertebrates and septate junctions (SJs) in invertebrates seal the epithelium to form a permeability barrier. A specialized class of junctions is found at points where three or more cell membranes meet. These tricellular tight junctions (tTJs) or tricellular septate junctions (tSJs) maintain the integrity of the barrier. The molecular components that are specifically localized or enriched at these junctions include transmembrane proteins with sealing extracellular domains, such as angulins and tricellulins in vertebrates, and Gliotactin (Gli), Anakonda (Aka), and M6 in Drosophila (Letizia, 2019).
Tricellular adherens junctions (tAJs) are thought to be points of high tension, at which the ends of actin filaments must be anchored to the cell surface. They are much less characterized than tTJs and tSJs, and no molecular components specific to tAJs have yet been identified. A few intracellular proteins are known to be enriched at tAJs, although they also localize continuously along bicellular adherens junctions (bAJs). One of them is the adaptor protein Afadin/Canoe (Cno), which links actin filaments to the junctional proteins E-cadherin (Ecad) and Echinoid (Ed). In the early Drosophila embryo, Cno enrichment at tAJs requires Rap1 activation by the guanine nucleotide exchange factor (GEF) Dizzy (Dzy), and in cultured Madin-Darby Canine Kidney (MDCK) cells tAJ localization of Afadin is enhanced by knocking down Zonula occludens 1 (ZO-1) family proteins. ZO-1 and Afadin physically interact and the single Drosophila ZO-1 homologue Polychaetoid (Pyd) has embryonic functions very similar to those of Cno, suggesting that the two proteins act together. As Ed is not necessary for the enrichment of Cno at tAJs, the protein that organizes tAJs by physically linking Cno to the cell surface at these positions remains unknown (Letizia, 2019).
Sidekick (Sdk) proteins are members of the immunoglobulin (Ig) superfamily that can mediate homophilic adhesion through their four N-terminal Ig domains. Their 13 Fibronectin type III (FnIII) domains interact with lipid membranes, allowing Sdks to facilitate tight cell-cell adhesion (Tang, 2018). Sdks are best known for their role in promoting interactions between specific neuronal processes in the retina. However, the identification of the original Drosophila sdk gene was based on defects in eye development in sdk mutants, including abnormalities in the pigment cell lattice that results from epithelial cell rearrangements and shape changes at the pupal stage. A Sdk protein trap accumulates at cell vertices in the epithelium of early embryos. Sdks are also expressed during branching morphogenesis in the developing mouse and human kidney, and their upregulation in glomeruli induces podocyte dysfunction. Sdk-1 controls actin cytoskeleton organization in prostate cancer cells and is required for their migration. The cytoplasmic domain of all Sdks ends in a conserved PDZ-binding motif that can interact with multiple PDZ domain proteins, including members of the MAGI family that control Sdk localization and function in the retina and kidney (Letizia, 2019).
This study shows that Sdk accumulates at tAJs through a mechanism that is influenced by mechanical tension. Sdk is in turn necessary to maintain normal levels of cell bond tension, and in its absence cell and junctional rearrangements are perturbed. Sdk assembles a tAJ complex by directly interacting with Pyd and enriching both Pyd and Cno at tAJs. These results suggest that Sdk links Cno and Pyd to the plasma membrane at tAJs, where they anchor the ends of actin filaments to maintain cellular tension and allow vertex remodeling during tissue rearrangements (Letizia, 2019).
Sdk is the only protein so far shown to be specifically localized to tAJs and almost excluded from bAJs in most epithelia. Although several proteins were known to be enriched at tricellular contacts at the level of AJs, most of them are also present along the whole bicellular junction. This study found find that Sdk is required for the specific enrichment of Cno, Pyd, and actin at tAJs, but not their localization at bAJs. Consistent with this, cno and pyd mutants have stronger phenotypes than sdk mutants, indicating that these proteins have functions independent of Sdk. In contrast, Sdk localizes to tAJs even in the absence of Cno and Pyd. It is proposed that Sdk is the hub that organizes a protein complex specifically at tAJs to modulate the actin cytoskeleton (Letizia, 2019).
This study haa shown that the C-terminal predicted PDZ-binding motif is required for Sdk function, and directly interacts with Pyd. The requirement for Sdk to recruit Cno to tAJs and the ability of misexpressed Cno to expand Sdk suggest that Cno is also in a complex with Sdk. As the vertebrate homologues of Cno and Pyd can bind to each other, all three proteins may be present in the same complex. However, knocking down ZO-1 proteins in MDCK cells increases the recruitment of Afadin to tAJs, so it is also possible that Cno and Pyd compete for the same binding site on Sdk. Although it is not possible to detect a direct interaction between Sdk and Cno, the experiments did not exclude this possibility. Alternatively, Sdk might recruit Cno indirectly, for example by binding to a GEF that increases Rap activity. Cno is known to interact with Ed, which is necessary for Ecad recruitment to bAJs; it is not yet clear whether Ed is in the same complex as Sdk or whether Cno interacts with the two proteins in a mutually exclusive manner. In addition to anchoring the ends of actin filaments, Cno might recruit other regulators of tension such as the LIM domain protein Smallish. pyd mutant embryos show defects in tracheal intercalation similar to sdk mutants, and pyd and cno genetically interact with sdk in this context, supporting a general role for these factors downstream of Sdk (Letizia, 2019).
Although Cno and Pyd are likely to be important mediators of the effects of Sdk on tension and junctional stability, other partners may also contribute to Sdk function. The cytoplasmic domain of Sdk has several regions of strong evolutionary conservation, which could serve as interaction domains for non-PDZ proteins. Elucidating the nature of the complex that has Sdk as its hub will provide important clues to the structure and function of tAJs. The expression of mouse Sdks in tissues other than the nervous system, such as the ureteric bud during branching morphogenesis of the kidney, suggests that the role of Sdk at tAJs may be conserved, although it is not yet known whether Sdks accumulate at tAJs in epithelia in other organisms (Letizia, 2019).
While Sdk can mediate homophilic interactions at bicellular contacts in cultured cells, in epithelial tissues it localizes almost exclusively to vertices. Several mechanisms might contribute to this localization pattern. It is possible that the topology of membranes at vertices imposes a particular structural conformation on Sdk that promotes its clustering there. The FnIII domains of Sdk molecules can interact with lipids and are thought to lie flat along the plasma membrane at bicellular contacts between parallel membranes, while the first four Ig domains mediate homophilic adhesion; the fourth and fifth Ig domains meet at a relatively rigid angle of 126°. This might make Sdk-mediated adhesion at tricellular interfaces (the point at which cells meet at an average angle of 120°) more energetically favorable. Alternatively or additionally, this angle might favor cis interactions between Sdk molecules through their FnIII domains over FnIII-membrane interactions, contributing to Sdk clustering. Such a geometric model could explain why Sdk is still enriched at tAJs when it is present on only two of the three cells that are in contact. However, this cannot be the only mechanism for Sdk localization, as tricellular vertices can form at a range of angles (Letizia, 2019).
In addition, this study found that tension enhances Sdk recruitment to tAJs. Although the Pyd homologue ZO-1 is stretched by tension, which controls its interactions with some of its binding partners, neither Pyd nor Cno is required for the initial localization of Sdk to tAJs. This argues that Sdk can detect tension by a mechanism independent of and prior to its own role in anchoring actin filaments through Cno and Pyd. Sdk might itself be mechanosensitive, as FnIII domains can stretch in response to force. Force exerted on the plasma membrane to pull it away from Sdk-Sdk adhesions could extend the FnIII domains to open up more interaction sites. Alternatively, as Ecad is necessary for Sdk localization, Sdk could detect tension through the cadherin-catenin complex, perhaps by interacting with α-catenin or another protein that is deformed by mechanical force (Letizia, 2019).
Mechanical forces can regulate tissue morphogenesis by promoting cell intercalation, oriented cell division or cell extrusion. AJs are known to transduce mechanical force, with one prominent mechanism being the force-dependent unfolding of α-catenin, which exposes binding sites for vinculin and other proteins. Vinculin is enriched at tAJs, indicating that tension is highest at these positions and suggesting that tAJs might sense and regulate tension. At bAJs, the cadherin-catenin complex resists tension by linking cell adhesions to the actin cytoskeleton through catch bonds. tAJs may require additional mechanisms to resist contractility because they anchor the ends of actin filaments, generating tension in an actomyosin network both along the cell cortex and in radially directed filaments. The requirement for Sdk to maintain normal levels of tension along cell bonds makes it a good candidate to mediate this tension-modulating function of tAJs. The homophilic adhesive properties of the extracellular domain of Sdk and its interaction with the actin cytoskeleton could allow it to transmit tension between cells or organize supracellular contractile networks that facilitate mechanical coordination across a tissue. Sdks mediate strong and compact adhesions with tightly packed molecules (Tang, 2018), potentially allowing them to resist force. However, the Ecad complex is sufficient to maintain adhesion at tAJs in the absence of Sdk (Letizia, 2019).
Cell shape changes and cell rearrangements often involve oscillations in apical area driven by pulses of actomyosin contraction. Both reduced tension on cell bonds in the embryo and defects in cell intercalation in the retina in sdk mutants could result from failure to connect the oscillatory network to tAJs. A lack of mechanical coupling of tensile actomyosin networks to tAJs would disrupt cell intercalation by reducing the efficiency of T1 transitions mediated by changes in junction length. In the trachea, Sdk accumulates at autocellular-intercellular vertices and is required for the replacement of intercellular junctions by autocellular ones. Tracheal cell intercalation does not require myosin contractility, suggesting that other actin regulators provide local forces to resolve cell rearrangements. Sdk may mediate the replacement of intercellular junctions by homophilic adhesion and/or by modulating the actin cytoskeleton to provide polarized tension or polarized membrane growth that allows vertex displacement during the zipping process. In general, vertex displacements in complex cell rearrangements may rely on the Sdk complex to transmit tension (Letizia, 2019).
As tTJs can also contribute to actin organization and tension, it will be important to investigate the interactions between the two types of tricellular junctions. Such studies may shed light on the role of tricellular junctions in regulating cell proliferation, stem cell homeostasis, and tissue integrity (Letizia, 2019).
In epithelia, tricellular vertices are emerging as important sites for the regulation of epithelial integrity and function. Compared to bicellular contacts, however, much less knowledge is available. In particular, resident proteins at tricellular vertices were identified only at occluding junctions, with none known at adherens junctions. In a previous study, it was discovered that in Drosophila embryos, the adhesion molecule Sidekick (Sdk), well known in invertebrates and vertebrates for its role in the visual system, localises at tricellular vertices at the level of adherens junctions. This study surveyed a wide range of Drosophila epithelia and establish that Sdk is a resident protein at tricellular adherens junctions, the first of its kind. Clonal analysis suggests that pair-wise homophilic adhesion is necessary and sufficient for Sdk tricellular vertex localisation. Super-resolution imaging using structured illumination reveals that Sdk proteins form string-like structures at vertices. Postulating that Sdk may have a role in epithelia where adherens junctions are actively remodelled, the phenotype of sdk null mutant embryos during Drosophila axis extension was determined using quantitative methods. Apical cell shapes were found to be strikingly abnormal in sdk mutants. Moreover, adhesion at apical vertices is compromised in rearranging cells, with holes forming and persisting throughout axis extension. Finally, it was shown that polarized cell intercalation is decreased and abnormal in sdk mutants. Mathematical modeling of the cell behaviours supports the conclusion that the T1 transitions of polarized cell intercalation are delayed in sdk mutants. It is proposed that this delay, in combination with a change in the mechanical properties of the converging and extending tissue, causes the striking cell shape phenotype of sdk mutant embryos (Finegan, 2019).
In the Drosophila eye imaginal disc the photoreceptor cells (R cells) differentiate according to a precise spatial and temporal order. The sidekick (sdk) gene is necessary to prevent extra R cells from differentiating during eye disc development. The extra cell appears between R3 and R4 early in R cell clusters and is most likely the result of the mystery cell inappropriately differentiating as an R cell. Mosaic analysis shows that sdk is required neither in the R cells nor in the extra cell, suggesting that sdk is necessary in the surrounding undifferentiated cells. The sdk gene codes for a protein that is a member of the immunoglobulin superfamily, having six immunoglobulin domains, thirteen fibronectin repeats and a transmembrane domain. The protein structure is consistent with its participation in cell-cell interaction during eye development (Nguyen, 1997).
Cell-cell adhesion is important for cell growth, tissue development, and neural network formation. Structures of cell adhesion molecules have been widely studied by crystallography, revealing the molecular details of adhesion interfaces. However, due to technical limitations, the overall structure and organization of adhesion molecules at cell adhesion interfaces has not been fully investigated. This study combines electron microscopy and other biophysical methods to characterize the structure of cell-cell adhesion mediated by the cell adhesion molecule Sidekick (Sidekick-1 and Sidekick-2) and obtained 3D views of the Sidekick-mediated adhesion interfaces as well as the organization of Sidekick molecules between cell membranes by electron tomography. The results suggest that the Ig-like domains and the fibronectin III (FnIII) domains of Sidekicks play different roles in cell adhesion. The Ig-like domains mediate the homophilic transinteractions bridging adjacent cells, while the FnIII domains interact with membranes, resulting in a tight adhesion interface between cells that may contribute to the specificity and plasticity of cell-cell contacts during cell growth and neural development (Tang, 2018).
Cell adhesion molecules such as Ig superfamily (IgSF) molecules usually have a long extracellular portion containing multiple domains. The N-terminal domains usually form homophilic pairs mediating the trans interaction between cell membranes. For the IgSF adhesion molecules, the N-terminal Ig-like domains could adopt either linear or the horseshoe-like conformation for trans interactions. A potential advantage of the horseshoe-shaped conformation is that it may provide relatively larger binding interfaces with higher specificity and selectivity. In the case of Dscam, its N-terminal Ig-like domains form a unique S-shaped conformation (dual horseshoe) with a larger dimerization interface comparing to the horseshoe-shaped conformation, hence providing the structural bases for the dimerization of a large number of isoforms. Therefore, the number and the conformation of the N-terminal Ig-like domains may correlate with the selectivity and specificity of the IgSF adhesion molecules (Tang, 2018).
Sdks, Dscams, and contactins are the IgSF adhesion molecules playing important roles in establishing sublaminar specificity in the retina and have been proposed as cell surface recognition codes for forming precise synaptic connectivity in the nervous system. Crystal structures of these molecules have revealed the molecular details of the N-terminal Ig-like domains as well as some Fn domains. Although these N-terminal domains adopt a conserved horseshoe-shaped conformation, their dimerization patterns are different. The dimers of Sdks are formed through the interactions between Ig1 and Ig2 of the monomers, while the dimerization of Dscam is formed by the interactions from Ig2, Ig3, and Ig7. Different dimerization patterns are also found for other horseshoe-shaped heads, suggesting that the dimerization modes may also be relevant to the functional activities of IgSF adhesion molecules (Tang, 2018).
A common feature shared among many IgSF adhesion molecules is that they usually contain different types of domains. The Ig-like domains usually locate at the N terminus and are involved in homophilic or heterophilic interactions by forming trans interactions between cell membranes. The FnII or FnIII domains are also frequently found in the IgSF adhesion molecules, and the number of the Fn domains varies in different cases. However, the roles of the Fn domains in cell adhesion have not been clearly defined, although it has been suggested that Fn domains may be involved in the cis interactions and facilitate the cluster formation in adhesion interfaces. Similar to other IgSF adhesion molecules, the N-terminal Ig-like domains of Sdks mediate the trans-homophilic adhesion. A notable feature of Sdk molecules is that they have 13 FnIII domains, which may be one of the largest FnIII fragments among IgSF adhesion molecules and makes two-thirds of the Sdk ectodomains. The extended Sdk ectodomain is roughly 70-80 nm in length, and the EM images show that the isolated Sdk ectodomains exhibit highly flexible conformations, it would be difficult to imagine how these long, flexible molecules could mediate stable interactions between cell membranes. However, the EM results show that the adhesion interfaces mediated by Sdks are rather narrow with a constant intermembrane distance that is much shorter than the length of Sdk ectodomains, suggesting that a large part of Sdk ectodomain may not contribute to the intermembrane spacing directly. Indeed, the current data also show that the FnIII domains are associated with membranes and probably lying down on the membrane surface, acting as anchors for Sdks on cell membranes. In the meantime, the FnIII domains may also mediate cis interactions among Sdk molecules and lead to a network formation on cell surface, which may stabilize the adhesion between membranes . Moreover, attempts are being made to identify potential 2D patterns of Sdk in the interfaces, but the current data do not show any obvious 2D pattern in the interfaces, although some small local patterns may exist. It is unclear whether any regular 2D patterns are required for forming functional adhesion interfaces or the packing density of Sdk molecules in the interfaces could be important for adhesion (Tang, 2018).
Since a large number of IgSF adhesion molecules have been identified, especially in neural systems, the major questions about cell adhesion are how these molecules are organized in the interfaces and how they regulate the specificity and plasticity of adhesion. These results for Sdks may provide a generic model to address these questions. Although adhesion molecules may have multiple domains and flexible conformations, they could lead to compact adhesion interfaces with constant intermembrane spacing, and different types of domains are playing different roles in establishing adhesion interfaces. The Ig-like domains might behave as 'hooks' to mediate the trans-homophilic pair formation, while the Fn domains act as 'anchors' associating with membrane and also introduce cis interactions among each other, resulting in a tightly packed stable interface. The number and the conformation of the Ig-like domains may reflect the specificity and selectivity of adhesion interactions, while the number of the Fn domains may be relevant to the plasticity or stability of adhesion interfaces. Therefore, the combination of different types of domains might be evolved as structural codes of IgSF adhesion molecules for establishing precise cell-cell contacts with specificity and plasticity (Tang, 2018).
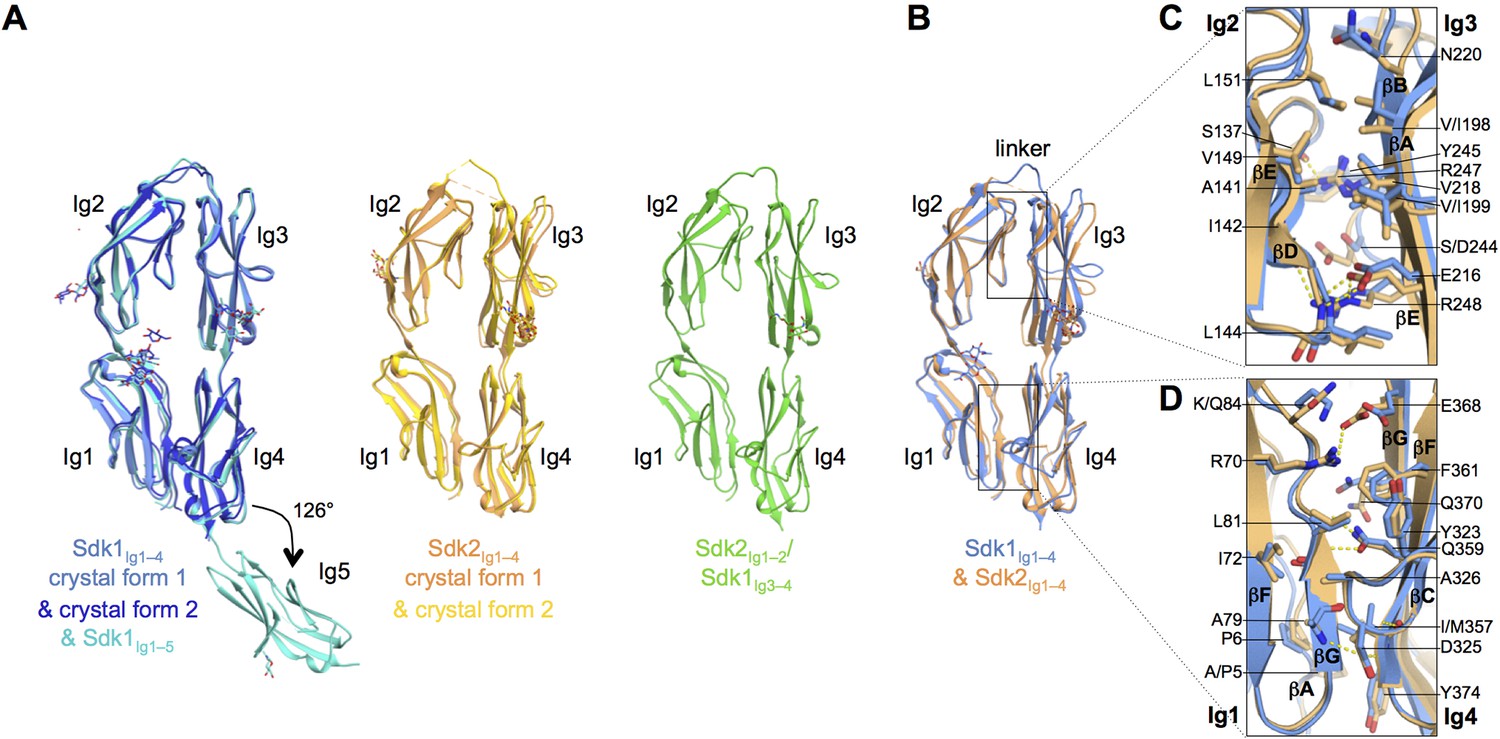
Sidekick (Sdk) 1 and 2 are related immunoglobulin superfamily cell adhesion proteins required for appropriate synaptic connections between specific subtypes of retinal neurons. Sdks mediate cell-cell adhesion with homophilic specificity that underlies their neuronal targeting function. This study reports crystal structures of Sdk1 and Sdk2 ectodomain regions, revealing similar homodimers mediated by the four N-terminal immunoglobulin domains (Ig1-4), arranged in a horseshoe conformation (see The N-terminal four Ig domains of Sdk1 and Sdk2 adopt a stable horseshoe conformation). These Ig1-4 horseshoes interact in a novel back-to-back orientation in both homodimers through Ig1:Ig2, Ig1:Ig1 and Ig3:Ig4 interactions. Structure-guided mutagenesis results show that this canonical dimer is required for both Sdk-mediated cell aggregation (via trans interactions) and Sdk clustering in isolated cells (via cis interactions). Sdk1/Sdk2 recognition specificity is encoded across Ig1-4, with Ig1-2 conferring the majority of binding affinity and differential specificity. It is suggested that competition between cis and trans interactions provides a novel mechanism to sharpen the specificity of cell-cell interactions (Goodman, 2016).
In the mammalian retina, processes of approximately 70 types of interneurons form specific synapses on roughly 30 types of retinal ganglion cells (RGCs) in a neuropil called the inner plexiform layer. Each RGC type extracts salient features from visual input, which are sent deeper into the brain for further processing. The specificity and stereotypy of synapses formed in the inner plexiform layer account for the feature-detecting ability of RGCs. This study analysed the development and function of synapses on one mouse RGC type, called the W3B-RGC. These cells have the remarkable property of responding when the timing of the movement of a small object differs from that of the background, but not when they coincide. Such cells, known as local edge detectors or object motion sensors, can distinguish moving objects from a visual scene that is also moving. This study shows that W3B-RGCs receive strong and selective input from an unusual excitatory amacrine cell type known as VG3-AC (vesicular glutamate transporter 3). Both W3B-RGCs and VG3-ACs express the immunoglobulin superfamily recognition molecule sidekick 2 (Sdk2), and both loss- and gain-of-function studies indicate that Sdk2-dependent homophilic interactions are necessary for the selectivity of the connection. The Sdk2-specified synapse is essential for visual responses of W3B-RGCs: whereas bipolar cells relay visual input directly to most RGCs, the W3B-RGCs receive much of their input indirectly, via the VG3-ACs. This non-canonical circuit introduces a delay into the pathway from photoreceptors in the centre of the receptive field to W3B-RGCs, which could improve their ability to judge the synchrony of local and global motion (Krishnaswamy, 2015).
Focal segmental glomerulosclerosis (FSGS) is a leading cause of nephrotic syndrome and end-stage renal disease worldwide. Although the mechanisms underlying this important disease are poorly understood, the glomerular podocyte clearly plays a central role in disease pathogenesis. The current work demonstrates that the homophilic adhesion molecule sidekick-1 (sdk-1) is up-regulated in podocytes in FSGS both in rodent models and in human kidney biopsy samples. Transgenic mice that have podocyte-specific overexpression of sdk-1 develop gradually progressive heavy proteinuria and severe FSGS. It was also shown that sdk-1 associates with the slit diaphragm linker protein MAGI-1, which is already known to interact with several critical podocyte proteins including synaptopodin, alpha-actinin-4, nephrin, JAM4, and beta-catenin. This interaction is mediated through a direct interaction between the carboxyl terminus of sdk-1 and specific PDZ domains of MAGI-1. In vitro expression of sdk-1 enables a dramatic recruitment of MAGI-1 to the cell membrane. Furthermore, a truncated version of sdk-1 that is unable to bind to MAGI-1 does not induce podocyte dysfunction when overexpressed. It is concluded that the up-regulation of sdk-1 in podocytes is an important pathogenic factor in FSGS and that the mechanism involves disruption of the actin cytoskeleton possibly via alterations in MAGI-1 function (Kaufman, 2010).
Four transmembrane adhesion molecules-Sidekick-1, Sidekick-2, Down's syndrome cell adhesion molecule (Dscam), and Dscam-like-are determinants of lamina-specific synapse formation in the vertebrate retina. Their C termini are predicted to bind postsynaptic density (PSD)-95/Discs Large/ZO-1 (PDZ) domains, which are present in many synaptic scaffolding proteins. This study identified members of the membrane-associated guanylate kinase with inverted orientation (MAGI) and PSD-95 subfamilies of multi-PDZ domain proteins as binding partners for Sidekicks and Dscams. Specific MAGI and PSD-95 family members are present in distinct subsets of retinal synapses, as are Sidekicks and Dscams. Using Sidekick-2 as an exemplar, this study shows that its PDZ-binding C terminus is required for both its synaptic localization in photoreceptors and its ability to promote lamina-specific arborization of presynaptic and postsynaptic processes in the inner plexiform layer. In photoreceptor synapses that contain both MAGI-1 and PSD-95, Sidekick-2 preferentially associates with MAGI-1. Depletion of MAGI-1 from photoreceptors by RNA interference blocks synaptic localization of Sidekick-2 in photoreceptors without affecting localization of PSD-95. Likewise, depletion of MAGI-2 from retinal ganglion cells and interneurons interferes with Sidekick-2-dependent laminar targeting of processes. These results demonstrate that localization and function of Sidekick-2 require its incorporation into a MAGI-containing synaptic scaffold (Yamagata, 2010).
Synaptic circuits in the retina transform visual input gathered by photoreceptors into messages that retinal ganglion cells (RGCs) send to the brain. Processes of retinal interneurons (amacrine and bipolar cells) form synapses on dendrites of RGCs in the inner plexiform layer (IPL). The IPL is divided into at least 10 parallel sublaminae; subsets of interneurons and RGCs arborize and form synapses in just one or a few of them. These lamina-specific circuits determine the visual features to which RGC subtypes respond. This study shows that four closely related immunoglobulin superfamily (IgSF) adhesion molecules--Dscam (Down's syndrome cell adhesion molecule), DscamL (refs 6-9), Sidekick-1 and Sidekick-2--are expressed in chick by non-overlapping subsets of interneurons and RGCs that form synapses in distinct IPL sublaminae. Moreover, each protein is concentrated within the appropriate sublaminae and each mediates homophilic adhesion. Loss- and gain-of-function studies in vivo indicate that these IgSF members participate in determining the IPL sublaminae in which synaptic partners arborize and connect. Thus, vertebrate Dscams, like Drosophila Dscams, play roles in neural connectivity. Together, these results on Dscams and Sidekicks suggest the existence of an IgSF code for laminar specificity in retina and, by implication, in other parts of the central nervous system (Yamagata, 2008).
Sidekick-1, a cell adhesion molecule of the immunoglobulin superfamily, is up-regulated in glomerular podocytes in the collapsing glomerulopathy of HIV-associated nephropathy (HIVAN). Sidekick-1 and its ortholog sidekick-2 have also been shown to function as neuronal targeting molecules, guiding developing neurons to specific synapses. The current work overexpresses mouse sidekick-1 and -2 in HEK 293 T cells in order to characterize their binding specificities. Cells transiently transfected with either sidekick-1 or -2 cDNA formed separate aggregates when mixed together, demonstrating that sidekicks are homophilic adhesion molecules. The transfection of the short splice variant (lacking the first two Ig domains) or a construct encoding sidekick-1 with the second Ig domain deleted both resulted in nearly abolished adhesion. A beta-sheet strand peptide containing the sequence QLVILA corresponding to an amino acid sequence in the second Ig domain of sidekick-1 showed specific interaction with the recombinant first Ig domain-His protein of sidekick-1. Cells expressing a mutant sidekick-1 where the binding sequence QLVILA is deleted failed to mediate significant adhesion. Furthermore, cells transfected with a chimeric sidekick, where the first two Ig domains of sidekick-2 are replaced with the corresponding two Ig domains of sidekick-1, form aggregates with sidekick-1-transfected cells. The reverse chimera, where the first two Ig domains of sidekick-2 are substituted onto sidekick-1, was similarly able to form aggregates with sidekick-2-transfected cells. These results establish that the first and second Ig domains of sidekick-1 and -2 are necessary and sufficient to mediate and target homophilic adhesion, and the QLVILA sequence is critical to the interaction. Understanding these functional domains has widespread implications in normal development and HIVAN pathogenesis (Hayashi, 2005).
Infection of podocytes by HIV-1 induces unique changes in phenotype, which contribute to the pathogenesis of glomerular disease in HIV-associated nephropathy (HIVAN). The host genetic pathways altered by HIV-1 infection that are responsible for these phenotypic changes are largely unknown. For identifying such pathways, representational difference analysis was performed comparing cDNA from HIV-1 transgenic podocytes with nontransgenic controls. In this way, a gene named sidekick-1 (sdk-1) was cloned, a transmembrane protein of the Ig superfamily that is highly upregulated in HIV-1 transgenic podocytes. Sdk-1 and its ortholog, sidekick-2 (sdk-2), were recently shown to guide axonal terminals to specific synapses in developing neurons. Their presence and role in other organs, including the kidney, has not been described. The current study demonstrates developmental expression of both sdk-1 and sdk-2 and a tight spatial and temporal regulation of these genes in kidney. During nephrogenesis, sidekick expression was observed first in ureteric bud and ureteric bud-derived tissues in a pattern similar to other genes known to play important roles in branching morphogenesis. In adult murine renal tissue, sidekick proteins were seen in glomeruli at low levels, and expression of sdk-1 was greatly upregulated in diseased HIV-1 transgenic mouse kidneys. In a human HIVAN kidney biopsy, sidekick expression was increased in glomeruli in a pattern consistent with the mouse model. It is proposed that the dysregulation of sdk-1 protein may play an important role in HIVAN pathogenesis (Kaufman, 2004).
A major determinant of specific connectivity in the central nervous system is that synapses made by distinct afferent populations are restricted to particular laminae in their target area. This study identified Sidekick (Sdk)-1 and -2, homologous transmembrane immunoglobulin superfamily molecules that mediate homophilic adhesion in vitro and direct laminar targeting of neurites in vivo. sdk-1 and -2 are expressed by nonoverlapping subsets of retinal neurons; each sdk is expressed by presynaptic (amacrine and bipolar) and postsynaptic (ganglion) cells that project to common inner plexiform (synaptic) sublaminae. Sdk proteins are concentrated at synaptic sites, and Sdk-positive synapses are restricted to the 2 (of > or =10) sublaminae to which sdk-expressing cells project. Ectopic expression of Sdk in Sdk-negative cells redirects their processes to a Sdk-positive sublamina. These results implicate Sdks as determinants of lamina-specific synaptic connectivity (Yamagata, 2002).
Search PubMed for articles about Drosophila Sidekick
Agi, E., Langen, M., Altschuler, S. J., Wu, L. F., Zimmermann, T. and Hiesinger, P. R. (2014). The evolution and development of neural superposition. J Neurogenet 28(3-4): 216-232. PubMed ID: 24912630
Astigarraga, S., Douthit, J., Tarnogorska, D., Creamer, M. S., Mano, O., Clark, D. A., Meinertzhagen, I. A. and Treisman, J. E. (2018). Drosophila Sidekick is required in developing photoreceptors to enable visual motion detection. Development. PubMed ID: 29361567
Finegan, T. N., Hervieux, N, Nestor-Bergmann, A., Fletcher, A. G., Blanchard, G. B., Sanson, B. (2019). The tricellular vertex-specific adhesion molecule Sidekick facilitates polarised cell intercalation during Drosophila axis extension. BioRxiv
Goodman, K. M., Yamagata, M., Jin, X., Mannepalli, S., Katsamba, P. S., Ahlsen, G., Sergeeva, A. P., Honig, B., Sanes, J. R. and Shapiro, L. (2016). Molecular basis of sidekick-mediated cell-cell adhesion and specificity. Elife 5. PubMed ID: 27644106
Hayashi, K., Kaufman, L., Ross, M. D. and Klotman, P. E. (2005). Definition of the critical domains required for homophilic targeting of mouse sidekick molecules. FASEB J 19(6): 614-616. PubMed ID: 15703275
Kaufman, L., Hayashi, K., Ross, M. J., Ross, M. D. and Klotman, P. E. (2004). Sidekick-1 is upregulated in glomeruli in HIV-associated nephropathy. J Am Soc Nephrol 15(7): 1721-1730. PubMed ID: 15213259
Kaufman, L., Potla, U., Coleman, S., Dikiy, S., Hata, Y., Kurihara, H., He, J. C., D'Agati, V. D. and Klotman, P. E. (2010). Up-regulation of the homophilic adhesion molecule sidekick-1 in podocytes contributes to glomerulosclerosis. J Biol Chem 285(33): 25677-25685. PubMed ID: 20562105
Krishnaswamy, A., Yamagata, M., Duan, X., Hong, Y. K. and Sanes, J. R. (2015). Sidekick 2 directs formation of a retinal circuit that detects differential motion. Nature 524(7566): 466-470. PubMed ID: 26287463
Langen, M., Agi, E., Altschuler, D. J., Wu, L. F., Altschuler, S. J. and Hiesinger, P. R. (2015). The developmental rules of neural superposition in Drosophila. Cell 162(1): 120-133. PubMed ID: 26119341
Letizia, A., He, D., Astigarraga, S., Colombelli, J., Hatini, V., Llimargas, M. and Treisman, J. E. (2019). Sidekick is a key component of tricellular adherens junctions that acts to resolve cell rearrangements. Dev Cell 50(3):313-326. PubMed ID: 31353315
Nguyen, D. N., Liu, Y., Litsky, M. L. and Reinke, R. (1997). The sidekick gene, a member of the immunoglobulin superfamily, is required for pattern formation in the Drosophila eye. Development 124(17): 3303-3312. PubMed ID: 9310325
Schwabe, T., Neuert, H. and Clandinin, T. R. (2013). A network of cadherin-mediated interactions polarizes growth cones to determine targeting specificity. Cell 154(2): 351-364. PubMed ID: 23870124
Tang, H., Chang, H., Dong, Y., Guo, L., Shi, X., Wu, Y., Huang, Y. and He, Y. (2018). Architecture of cell-cell adhesion mediated by sidekicks. Proc Natl Acad Sci U S A 115(37): 9246-9251. PubMed ID: 30150416
Uechi, H. and Kuranaga, E. (2019). The tricellular junction protein Sidekick regulates vertex dynamics to promote bicellular junction extension. Dev Cell 50(3):327-338. PubMed ID: 31353316
Yamagata, M., Weiner, J. A. and Sanes, J. R. (2002). Sidekicks: synaptic adhesion molecules that promote lamina-specific connectivity in the retina. Cell 110(5): 649-660. PubMed ID: 12230981
Yamagata, M. and Sanes, J. R. (2008). Dscam and Sidekick proteins direct lamina-specific synaptic connections in vertebrate retina. Nature 451(7177): 465-469. PubMed ID: 18216854
Yamagata, M. and Sanes, J. R. (2010). Synaptic localization and function of Sidekick recognition molecules require MAGI scaffolding proteins. J Neurosci 30(10): 3579-3588. PubMed ID: 20219992
date revised: 25 November 2019
Home page: The Interactive Fly © 2011 Thomas Brody, Ph.D.
{kind=link}