Sex combs reduced
The six Drosophila proteins that belong to the antennapedia-type Homeobox subfamily are
Antennapedia (ANTP), Abdominal-A (ABD-A), Deformed (DFD), Proboscipedia (PB),
Sex combs reduced (SCR) and Ultrabithorax (UBX).
The ExPASy World Wide Web (WWW) molecular biology server of the Geneva University Hospital and
the University of Geneva provides extensive documentation for the 'Homeobox' antennapedia-type protein signature. See four paralogous Hox clusters of mammals for homologies of Sex combs reduced with mammalian Hox proteins.
To analyze how the silk glands of the lepidopteran Bombyx mori develop, two genes were cloned and identified that encode the homeodomain and its flanking regions identical to
the corresponding regions of Drosophila Deformed and Sex combs reduced. The Bombyx silk gland is assumed to be an evolutionally homologous organ to the Drosophila salivary gland because both structures are formed in the labial segment and share some similar functions. Bombyx Deformed is expressed in the mandibular and maxillary segments, whereas
expression of Bombyx Sex combs reduced is first limited to the labial segment and at later stages
extended to the anterior part of the prothoracic segment. The expression of Bombyx Sex combs
reduced then disappears from the invaginating placodes of silk glands where expression of Bombyx
fork head/SGF-1 follows. In the mutant embryos, which lack the 3' end region of Bombyx
Antennapedia, Bombyx Sex combs reduced is
expressed ectopically in the thoracic and abdominal regions, in addition to expression in the labial segment. Bombyx fork head/SGF-1 is also
ectopically expressed in the T1, T2, and T3 segments, resulting in the ectopic induction of the silk gland
invaginations. These results suggest that Bombyx homeobox genes such as the Bombyx Deformed and
Sex combs reduced are associated with determination of the segment identities and Bombyx Sex
combs reduced is involved in the induction of silk gland development (Kokubo, 1997).
Hox genes are known to control the identity of
serially repeated structures in arthropods and vertebrates.
The expression pattern of the Hox genes Deformed, Sex combs reduced, Antennapedia, and Ultrabithorax/abdominal-A (Ubx/abd-A)
from the honey bee Apis mellifera were analyzed. A
cDNA with the complete coding region of the Antennapedia
gene from Apis was cloned. Comparison with Antp proteins
from other insect species revealed several regions of homology.
The expression patterns of the isolated Hox
genes from Apis show that the original expression patterns
of Dfd, Scr, and Antp appear between late blastoderm
and early germ band stage in a temporal and spatial
sequence. Each of them shows up as a belt, spanning approximately
two segment anlagen; Dfd in the anterior
gnathal region, Scr in the posterior gnathal and anterior
thoracic region, and Antp in the thoracic region. Following
expansion of the Antp domain in the abdomen as a
gradient towards the posterior, Ubx/abd-A expression appears
laterally in the abdomen. During gastrulation and
in the germ band stage the domains of strong expression
do not overlap any more, but touch each other. After gastrulation
the borders of the expression domains partly
correlate with parasegment and partly with segment
boundaries. Laterally, gaps between the domain of each
gene may show no expression of any of the genes examined (Walldorf, 2000).
In the blastoderm stage the expression domains of all
four genes are similar in fruit fly and honey bee embryos.
Certainly these genes play very similar roles in establishing
the general character of that part of the insect
body where they have their main expression domain.
Even in Chelicerata the orthologs show comparable
patterns. However, some differences should be mentioned. In the
fly, the borders of the Dfd domain are segmental. In
the bee, the anterior expression border is initially parasegmental
in the rudiment of the intercalary segment: this changes from segmental to parasegmental during
gastrulation in the ventral part of the maxillary segment. The steady Engrailed pattern shows that this change is not a result of cell movement. The same development of that pattern is reported for Tribolium. This is most
clearly seen ventrally in the parasegmental Scr pattern in
the germ band of the bee, corresponding to segmental in
that area in the fly. While some differences in the Dfd pattern are
consistently observed between the bee and the fly, the
complicated posterior borders of the Scr and the Antp
patterns of young and old embryos of flies and bees are identical. The
same Antp pattern is also found in Thermobia domestica. However, in Manduca sexta the
anterior border of Antp is the compartment border of T1. It may be that some of these differences are only the result of different sensitivity levels of
the stainings. Probably the ventral switch from segmental
to parasegmental is more general in insects but is not
seen very clearly in some cases, such as in the anterior
part of the fruit fly head, due to the subsequent head involution.
The resulting ventrolateral corners of the expression
borders of Dfd, Scr, and Antp correlate fairly
well with the position of the appendage anlagen and may
help to pin down their position (Walldorf, 2000).
Expression patterns of six homeobox containing genes in a model chelicerate, the oribatid mite
Archegozetes longisetosus, were examined to establish homology of chelicerate and insect head
segments and to investigate claims that the chelicerate deutocerebral segment has been reduced or
lost. engrailed (en) expression, which has been used to demonstrate the presence of segments in
insects, fails to demonstrate a reduced deutocerebral segment. Expression patterns of the chelicerate
homologs of the Drosophila genes Antennapedia (Antp),Sex combs reduced (Scr), Deformed (Dfd),
proboscipedia (pb), and orthodenticle (otd) confirm the direct correspondence of head segments. The
chelicerate deutocerebral segment has not been reduced or lost (Telford, 1998).
Insects have evolved a large variety of specialized feeding
strategies, with a corresponding variability in mouthpart
morphology. In the Hemiptera, the mandibular and
maxillary segments give rise to two similar pairs of long, thin
stylets. The paired maxillary appendages form
channels for liquid flow and the piercing mandibles lie on
either side. These four interlocked stylets run down a groove
in the long, fused labium, which provides support. Although
in typical mandibulate insects the maxillary and labial
appendages are very similar, in the Hemiptera, it is
the mandibular and maxillary appendages that share a highly
unusual morphology very different from the labium.
These specialized mouthparts represent an important
evolutionary innovation that allows this order of insects to
feed by extracting fluids from other organisms.
There is little understanding of the
developmental mechanisms that underlie mouthpart diversity.
Until recently it was difficult to perform any analysis of
gene function outside of the genetic model insects
Drosophila melanogaster and Tribolium castaneum. In this
paper, the use of dsRNA-mediated interference
(RNAi) is described to dissect gene function in the development of the
milkweed bug Oncopeltus fasciatus, which has specialized
suctorial mouthparts. The Hox genes Deformed (Dfd),
proboscipedia (pb) and Sex combs reduced (Scr) have
previously been shown to be expressed in the gnathal
appendages of this species. Strikingly, the milkweed bug
was found to have an unusual expression pattern of pb.
Here, by analyzing single and combination RNAi
depletions, it has been found that Dfd, pb and Scr are used in the
milkweed bug to specify the identity of the mouthparts. The
exact roles of the genes, however, are different from what
is known in the two genetic model insects. The maxillary
appendages in the bug are determined by the activities of
the genes Dfd and Scr, rather than Dfd and pb as in the fly and beetle. The mandibular appendages are specified by
Dfd, but their unique morphology in Oncopeltus suggests
that Dfd's target genes are different. As in flies and beetles,
the labium is specified by the combined activities of pb and
Scr, but again, the function of pb appears to be different.
Additionally, the regulatory control of pb by the other two
genes seems to be different in the bug than in either of the
other species. These novelties in Hox function, expression
pattern and regulatory relationships may have been
important for the evolution of the unique Hemipteran head (Hughes, 2000).
Dfd is the sole gene responsible for mandibular identity. This conforms with the expression of Dfd in the mandibular segment. The single depletion of Dfd transforms the mandibular appendages to distal antennal identity, rather than the long thin stylets normally formed. Thus Dfd is necessary for proper mandibular development. In contrast, the depletion of pb leaves the mandibular appendages untouched. Thus pb is not necessary for mandibular development. The phenotype of the Scr depletion is more difficult to interpret, since the mandibular stylets generally fail to grow, and are merely short bristles in a mass of undifferentiated tissue. Since Scr is not expressed in the mandibular segment, it is suspected that this is an indirect effect of the Scr phenotype on other segments of the head. Because head development is integrated to some degree, non-local, indirect effects are often seen in Drosophila Hox mutants, particularly labial and Deformed. In the case of milkweed bug Scr, disruption of the proximal labium may be interfering with the normal development of the adjacent mandibular and maxillary appendages. The Dfd Scr double depletion corroborates this view. The mandibles in the double form distal antennae, indistinguishable from either the Dfd depletion alone or the triple. Therefore it can be inferred that, when present, Scr is not acting in the mandibular appendage to directly specify any identity over that of the default state (Hughes, 2000).
The inferred activity of Dfd, but not pb and Scr, in the mandibular appendages matches the predictions based on expression patterns. While Dfd is expressed strongly in the mandibular appendages, pb and Scr are not. The role of Dfd as the sole Hox gene regulating mandibular identity also matches the situation in Tribolium, where Dfd mutants transform Mn structures, while maxillopedia (mxp, the pb homolog) and Cephalothorax (Cx, the Scr homolog) mutants leave the Mn unaffected. The effect of Hox mutations on the reduced mandibular structures of Drosophila, however, is more difficult to determine. In the embryo, Dfd mutations disrupt the dorsolateral papillae of the terminal sense organ, which are thought to derive from the Mn segment. In the Drosophila adult, Dfd mutations disrupt parts of the head capsule. Mutations in pb and Scr do not appear to affect these presumed mandibular structures. Although both beetles and milkweed bugs use Dfd to specify the mandibular segment, the resulting appendages are very different. In contrast to the chewing mandibles of the beetle, the bug mandibles are very thin stylets nearly as long as the body. Again, these are very different from the mandibular appendages in Drosophila, which are internal structures in the embryo, and are either missing or incorporated into the head capsule of the adult. Thus it is concluded that while Dfd's basic role in the mandible may be conserved, the developmental module driven by Dfd is extremely labile (Hughes, 2000).
In the maxillary segment, Dfd depletion results in only a partial transformation of the maxilla to antennal identity. However, the Dfd;Scr double depletion results in complete transformation of the maxillae to antennae. Thus it can be concluded that Dfd acts in concert with Scr in the maxillary segment. The curled phenotype of the Dfd depletion maxillary appendage suggests that Scr may be repressing growth of the transformed limb in its posterior domain; this repression is released in the Dfd;Scr double depletion. The phenotypes in the maxillary segment are somewhat in conflict however, regarding the role of pb. Two results suggest that pb is not acting to specify maxillary identity. (1) Depletion of pb alone leaves the maxillary segment unaffected, therefore pb is not necessary for wild-type maxillary development. (2) The phenotype of the Dfd;Scr double is the same as the triple depletion (i.e., antennae), so it can be inferred that pb is not acting to confer any identity over that of the default. There is, however, a subtle difference between the phenotypes of the Dfd single and Dfd;pb double depletion. While the Dfd depletion produces short, curled antenna-like appendages, the Dfd;pb double produces straight antenna-like appendages, often with pretarsal claws. This suggests that in the absence of Dfd, pb can affect the maxillary appendages. Whether this activity derives from its small dorsal maxillary domain of accumulation, or from an expanded domain of pb expression in the Dfd depletion, is not yet clear. Nevertheless, based on the pb depletion, it is concluded that, in wild-type embryos, pb is not active in the specification of the maxillary stylet (Hughes, 2000).
This lack of pb function in determining the maxillary stylet matches the prediction made based on the expression pattern of pb in the bug. Thus far this is the only insect known not to use pb to determine maxillary identity. In Drosophila and Tribolium, pb/mxp mutations cause transformation of the maxillary palps, which become reduced in Drosophila, and transform to legs in Tribolium. In the bug, it would appear that Scr assumes much of this role. This is in contrast to Drosophila or Tribolium, where the Scr/Cx mutation does not affect the maxillary appendage. While Dfd and Scr work together to specify maxillary identity in the bug, it seems that Dfd has the principal role. Perhaps by activating a similar set of target genes as in the mandible, Dfd may induce a similar stylet identity in the maxillary appendage. This contrasts with mandibulate insects, in which the maxillary morphology is most similar to the labium, probably due to the activity of pb in both of those appendages (Hughes, 2000).
By analyzing the depletion phenotypes, it can be inferred that pb and Scr cooperate to pattern the labium. Dfd depletion leaves the labium unaffected, so it can be concluded that Dfd is not necessary for wild-type labial development. Depletion of pb results in a labium that is fused and wild-type basally, but splits distally into a pair of normal T1 legs. From this, it can be inferred that pb is necessary for distal, but not proximal, labial development. Scr depletion results in transformation of the entire appendage to a mixed identity between leg and antenna. From this it can be inferred that Scr is necessary for development of both proximal and distal labium. The pb;Scr double depletion, as well as the triple, transforms the labium to a pair of full-length antennae, which is very different from either single depletion alone. Therefore in the bug as well as the fly and beetle, pb and Scr specify the labium. The Scr depletion phenotype in the labium supports the hypothesis that Scr normally functions to fuse the two labial appendages. This conclusion is derived from the observation that, in wild-type and the pb depletion animals, the transformed labial appendage is basally fused, while the Scr depletion limbs are often well-separated (Hughes, 2000).
By comparing the pb depletion to the pb;Scr double, it can be inferred that the function of Scr in the absence of pb is to induce T1 legs. In light of the Drosophila model, this may seem reasonable, since Scr is also expressed in T1 but, in the case of the milkweed bug, it is actually a bit surprising. Based on its ectodermal pattern of accumulation in a single discrete spot on the T1 leg, Scr would not appear to be capable of conferring overall leg identity to this appendage. Rather, in ventral T1 Scr appears to function solely in the specification of the T1 leg comb. The role of pb in the gnathal appendages is provided by a comparison of the Scr single depletion animals to the pb;Scr doubles. The Scr depletion labium is transformed to a pair of appendages with features of both antenna and leg. This suggests that the activity of pb alone is conferring some leg identity over the default antennal state. This contrasts strongly with the function of pb in flies. In two situations in Drosophila, the presence of pb alone appears to induce maxillary identity: first, in Scr hypomorphic mutant adults, the labium is partially transformed to maxillae; second, when pb+ is ectopically expressed in the antennae, they are transformed to maxillary palps. Since in the bug pb is inducing partial leg identity rather than maxillary identity, this suggests that pb may have a very different function than in the fly. Since both pb and Scr, when acting alone, each seem to be specifying some leg identity, this suggests that pb and Scr have overlapping functions. It is known, however, that the two genes are not entirely redundant, because their depletion phenotypes are different and because when both are present the result is labial identity. Thus the activity of wild-type pb or Scr alone induces leg identity, but the combined synergistic activity of the two together creates a new overriding labium identity. Whether the combinatorial effect of pb and Scr is at the level of transcription of their target genes, or in the activity of their combined target gene products, is not yet clear (Hughes, 2000).
One aspect of the Scr depletion phenotype deserves special mention. Although expression studies have shown that Scr is not expressed in the milkweed bug labium distally, the Scr depletion results in a change of identity for the distal labium. Instead of the sensory structures normally formed there, pretarsal claws and pulvilli appear. This suggests that the presence of Scr elsewhere in the appendage normally acts non-cell autonomously to induce labial identity. Non-cell-autonomous functions for Scr and other Hox genes have been suggested for other tissues in Drosophila (Hughes, 2000).
Unlike the broad band of Scr expression across the first thoracic segment in fly embryos, in milkweed bugs and other insects, the pattern of Scr is restricted to three thoracic domains: a T1 leg patch, a dorsal T1 patch and the mesoderm of all legs. A hypothesis about the function of Scr in one domain can be addressed, but the functions of the other thoracic domains in the milkweed bug cannot yet be described. Scr is expressed in the T1 limb in an anterior patch midway down the leg. This accumulation correlates with the position of a comb on the distal tibia of this leg and it has been proposed that Scr expression is necessary for the development of that structure. The Scr depletions reported here support this hypothesis. Although comb transformation is not invariably associated with labium transformation, of the animals that also had a transformed labium, 80% of the legs had a transformed comb, with bristles resembling those found in a comparable position of the more posterior legs. This suggests that, in wild-type, the patch of Scr expression is promoting the formation of the T1 comb from a row of bristles on the distal tibia, as compared to the serially homologous structures found on the second and third legs (Hughes, 2000).
The second domain of Scr expression is a patch on dorsal T1. Tribolium and Drosophila Cx/Scr mutants develop ectopic wings on dorsal T1; thus it has been proposed that the dorsal patch of Scr functions to repress wing development in that segment in the other insects as well. Unfortunately, this could not be tested with the Scr depletions reported here since, in the bug, the wings do not develop until several instars after hatching. Because depletions of Scr also disrupt the mouthparts, affected animals could not be raised to determine if wing rudiments developed on the first thoracic segment. The third thoracic domain of Scr is in the mesoderm of all three segments. This expression of Scr has been proposed to non-cell autonomously direct tarsus development in the fly. In the Scr depletions, no consistent effect on the tarsi was observed. It would be difficult, however, to ensure that the depletion had completely removed Scr function from all cells of the mesoderm; activity in just a few cells might be sufficient to create enough downstream diffusible signal.
Thus determining the generality of this proposed function for Scr requires further experimentation (Hughes, 2000).
Since the triple depletion animals should lack all Hox activity in the gnathal segments, the default state of these appendages in the absence of Hox function can be inferred. Antennae have been thought to represent the default appendage state in the absence of Hox activity, since a large deletion of much of the Hom-C in the beetle was found to produce a larva bearing a series of antennae instead of mouthparts and legs. Likewise, in the adult fly, the pb and Scr double mutant combination in the adult labium gives rise to antennae. Triple depletion indicates that, in the milkweed bug as well, antennae represent the no-Hox state for the gnathal appendages. In the triple depletion, the maxillary and labial segments form full-length antennae. Since wild-type antennae are the only paired appendages that lack Hox expression, it is reasonable that this morphology is produced in the absence of Hox function. Furthermore, this result indicates that the genetic program that specifies the presence of an appendage is independent of Hox expression, and that the function of the Hox gene products is to confer a specific morphology onto a generic appendage program. In the absence of Hox activity, this generic appendage turns on the antennal differentiation pathway (Hughes, 2000).
In the absence of Hox activity, the mandibular appendages, however, each form only a distal antenna on a short base. This suggests that a portion of the generic appendage is missing. Previous work has suggested that insect mandibles, which lack Distal-less (Dll) expression, are gnathobasic (lack distal portions of the appendage). In Drosophila and Tribolium, Dll is repressed by Dfd in the mandibular segment. In the milkweed bug, therefore, depletion of Dfd is likely associated with Dll derepression. Consistent with this hypothesis, the morphology of the transformed mandibular appendage is primarily to distal antenna. Interestingly, as noted, the basal portions of the appendage are distorted or missing, compared to the maxillary or labial default appendages. This suggests that some aspects of the reduced structure of the insect mandible are independent of the Dfd/Dll pathway (Hughes, 2000).
In this study, evidence has been found of remodeling in the role of Hox genes in head appendage development. This change in the role of the Hox genes is due to three mechanisms: (1) change in expression patterns; (2) change in regulatory relationships, and (3) change in function. Previous work had shown that pb possesses a unique expression pattern in the milkweed bug. The RNAi analysis confirms that the unusual expression pattern does in fact reflect an unusual domain of activity. Unlike Drosophila, Tribolium and probably most other insects, pb is not necessary for development of the maxillary appendages in the milkweed bug. The pb gene is also regulated differently in the bug, where it appears to be independent of the activity of Dfd and Scr. This contrasts strongly with the regulatory interactions described for the fly and the beetle. Studies of more distant insects are needed, however, before the ancestral state and the direction of change for these regulatory interactions can be confidently inferred. Lastly, evidence was found of novel functions in the bug that probably reflects differences in the suite of target genes activated by the Hox genes. For example, the function of pb is different in the bug, where it induces a mixed leg-like identity rather than a maxillary palp. Even in the mandibular segment, where Dfd is the conserved controller of identity, the resulting mandibular appendage is strikingly different from the mandibles of other insects, indicating a very different developmental module at work downstream of the Hox signal. It is not known, however, whether the repertoire of target genes is different, whether the orthologous target genes themselves have different functions or if both possibilities are correct (Hughes, 2000).
These differences in Hox function are provocative, but a likely scenario for the evolution of the Hemipteran mouthparts cannot yet be described. The exclusion of pb from the maxillary segment may have caused the loss of the maxillary palp -- or the expression of pb may be the result of loss of the palp, which would remove selection to maintain pb in an inactive tissue. Moreover, the evolution of Hemipteran stylate-haustellate mouthparts was clearly a multistep process within the Hemipteroid group: the Psocoptera (booklice) and some Phthiraptera (chewing lice) have pronounced lacinia; the Thysanoptera (thrips), some Phthiraptera (sucking lice) and the Hemiptera have long stylets; and finally, the Hemipterans have lost the maxillary palps entirely. Understanding the changes that accompanied the evolution of these mouthparts will require additional study of the embryology, homologies and gene expression of these various insect orders (Hughes, 2000).
The role of Hox genes in the development of insect gnathal appendages has been examined in three insects: the fruitfly, Drosophila melanogaster; the milkweed bug, Oncopeltus fasciatus; and the red
flour beetle, Tribolium castaneum. In each of these organisms, the identity of the labium depends on the homeotic genes Sex combs reduced (Scr) and proboscipedia (pb). Loss of pb function in each of
the three insects results in homeotic transformation of the labial appendages to legs. In contrast, loss of Scr function yields a different transformation in each species. Interestingly, mutations in Cephalothorax (Cx), the Tribolium ortholog of Scr, transform the labial appendages to antennae, a result seen in the other insects only when both pb and Scr are removed. The Tribolium labial appendages also develop as antennae in double mutants. Further, expression of the Tribolium proboscipedia ortholog maxillopedia (mxp) is greatly reduced or absent in the labium of Cx mutant larvae. Thus, in the wild-type labial segment, Cx function is required (directly or indirectly) for mxp transcription. A similar interaction between Scr and pb during Drosophila embryogenesis has been described recently. Thus, this regulatory paradigm appears to be conserved at least within the Holometabola (DeCamillis, 2001).
The Tribolium Cx single mutant phenotype (transformation of larval labial appendages to antennae) differs from the phenotypes of Drosophila Scr mutant larvae and adults. Understanding the underlying regulatory pathways helps resolve this discrepancy. In both Drosophila and Tribolium embryos, Scr/Cx positively regulates pb/mxp. However, pb lacks an embryonic function in Drosophila, and some other gene is presumably responsible for the residual gnathal identity in the labial segment of Scr mutant larvae. In adult flies, pb functions to specify the maxillary palps. Scr hypomorphs and mitotic clones lacking Scr cause transformation of the adult labial appendages to maxillary palp. However, when clones lack both Scr and pb, cells assume antennal identity. These results suggest that, in contrast to the embryonic paradigm, pb is present and functional during Drosophila adult development even in the absence of Scr. Indeed, expression of pb in the labial imaginal disc does not require Scr function. In fact, to some extent, the reverse appears to be true. Scr expression is greatly reduced in the distal portions of pb null labial imaginal discs (DeCamillis, 2001 and references therein).
Orthologs of the Hox genes Sex combs reduced and proboscipedia are active in the developing labial appendages of all insect species tested. The remarkable variation among insect gnathal structures, particularly in the distal podomeres, suggests two Hox genes may enhance the adaptive potential of gnathal appendage morphology. Functional studies in Drosophila melanogaster, the flour beetle Tribolium castaneum and the milkweed bug Oncopeltus fasciatus show that cooperation between Scr and pb has been generally conserved, but specific mechanisms have been altered during evolution. Cross-regulation of pb by Scr is evident in Drosophila and Tribolium, the more closely related of the three species, but not in Oncopeltus. In all three species, pb function is restricted to the distal podomeres, but details are only known for Drosophila and Oncopeltus, two species exhibiting specialized stylate-haustellate mouthparts. Drosophila pb is required for distal Scr expression, and to repress the appendage patterning genes dachshund and Distal-less. Oncopeltus pb has the novel capacity to specify leg fates. Little is known about distal functions of Tribolium pb. Hypomorphic mutations of the Tribolium pb ortholog maxillopedia can be arranged in a graded phenotypic series of palp to leg transformations along both the proximodistal and dorsoventral axes. Mid-embryonic expression profiles of Tribolium pb, Scr, wingless and Dll genes were examined in maxillopedia hypomorphic and null mutant backgrounds. Levels of pb and Scr are significantly reduced in the distal appendage field. Tribolium pb therefore positively regulates distal Scr expression, a role that it has in common with Drosophila pb. Tribolium wg is normally down-regulated in the distal domain of the embryonic gnathal appendage buds. It becomes activated distally in maxillopedia hypomorphs. Repression of wg by pb has not been reported in the labial imaginal discs of Drosophila. Alterations of Tribolium Scr and wg expression occur in Dll-expressing cells, however, unlike in Drosophila labial imaginal discs, Dll expression appears unaffected in pb hypomorphic backgrounds. It is concluded that the Hox genes Sex combs reduced and proboscipedia control an appendage organizer and cell autonomous fate determination during embryonic labial palp development in Tribolium (DeCamillis, 2003).
During Drosophila segmentation, gap genes function as short-range
gradients that determine the boundaries of pair-rule stripes. A classical
example is Drosophila Krüppel (Dm'Kr) which is
expressed in the middle of the syncytial blastoderm embryo. Patterning defects
in Dm'Kr mutants are centred symmetrically around its bell-shaped
expression profile. The role of Krüppel was examined in the
short-germ beetle Tribolium castaneum where the pair-rule stripes
corresponding to the 10 abdominal segments arise during growth stages
subsequent to the blastoderm. The previously described mutation
jaws is an amorphic Tc'Kr allele. Pair-rule gene expression
in the blastoderm is affected neither in the amorphic mutant nor in
Tc'Kr RNAi embryos. Only during subsequent growth of the germ band
does pair-rule patterning become disrupted. However, only segments arising
posterior to the Tc'Kr expression domain are affected, i.e., the
deletion profile is asymmetric relative to the expression domain. Moreover,
stripe formation does not recover in posterior abdominal segments, i.e., the
Tc'Krjaws phenotype does not constitute a gap in segment
formation but results from a breakdown of segmentation past the 5th
eve stripe. Alteration of pair-rule gene expression in
Tc'Krjaws mutants does not suggest a direct role of
Tc'Kr in defining specific stripe boundaries as in
Drosophila. Together, these findings show that the segmentation
function of Krüppel in this short-germ insect is fundamentally
different from its role in the long-germ embryo of Drosophila. The
role of Tc'Kr in Hox gene regulation, however, is in better
accordance to the Drosophila paradigm (Cerny, 2005).
The most obvious differences between the phenotypes of Krüppel
in Tribolium and Drosophila are the homeotic transformations
in Tc'Krjaws and Tc'Kr RNAi larvae that are not
evident in Dm'Kr mutants. Such transformations are not entirely
unexpected given that in Drosophila the expression boundaries of Hox
genes are also set by gap genes, including Dm'Kr. However, in
Drosophila gap mutants all segments that would be transformed because
of misregulation of homeotic genes usually also suffer segmentation defects
and fail to develop. By contrast, Tribolium segment primordia
anterior of, and within, the Krüppel expression domain do
differentiate, such that homeotic transformations can manifest themselves in
the differentiated larva (Cerny, 2005).
The expression of homeotic genes in Tc'Krjaws embryos
is consistent with the morphological transformations observed. The results with
Tc'Dfd, Tc'Scr, Tc'Antp and Tc'Ubx confirm and extend
earlier findings for Tc'pb and Tc'UBX/Tc'ABD-A expression.
Notably, the complementary double-segmental expression of Dfd and
Scr in Tc'Krjaws embryos explains the phenotype
of alternating maxillary and labial segments. These expression
patterns indicate that the posterior limit of Tc'Dfd and
Tc'Scr domains is set through inhibition by Tc'Kr. In this
respect, Tc'Kr fulfils a function similar to Drosophila gap
genes (Cerny, 2005).
The homeotic phenotype of Tc'gt RNAi embryos
could suggest a similar function in Hox regulation for Tc'gt. Indeed
Tc'Antp anteriorly expands and gnathal Hox genes
(Tc'Scr) repress in Tc'gt RNAi embryos, consistent with
the expansion of thoracic fates found in differentiated Tc'gt RNAi
larvae. These transformations are just opposite to those of
Tc'Krjaws larvae. Interestingly, in embryos that lack
Tc'Kr and at the same time have reduced Tc'gt activity, the
homeotic effect of Tc'Krjaws clearly is epistatic. This shows that the
ectopic Tc'gt stripes in the Tc'Kr mutant do not contribute
to the Tc'Kr phenotype. However, this experiment suggests that the
homeotic transformation of gnathal segments into thorax in Tc'gt RNAi
embryos is indeed an indirect effect and comes about through misregulation of
Tc'Kr in these embryos. This interpretation is supported by the finding that the Tc'Kr expression domain expands anteriorly in
Tc'gt RNAi embryos. Evidently, it is
expansion of Tc'Kr that results in repression of gnathal Hox genes in
maxilla and labium of Tc'gt RNAi embryos, not loss of gnathal Hox
gene activation. Similarly, expansion of Tc'Antp in Tc'gt
RNAi larvae could be due to activation by anteriorly expanded Tc'Kr.
However, as Antp is not significantly reduced in
Tc'Krjaws, it seems more likely that Tc'gt acts
directly to define the anterior boundary of the Tc'Antp domain (Cerny, 2005).
In addition to gap gene input, Drosophila Hox genes also receive
input from pair-rule genes. The near-pair-rule pattern of Tc'Dfd and
Tc'Scr in Tc'Krjaws embryos reveals an important
role of pair-rule genes also in defining Tribolium Hox domain
boundaries. It seems likely that regulation of Tc'Dfd and
Tc'Scr by pair-rule genes is responsible for the precision of their
expression boundaries in wild-type Tribolium embryos, while input
from gap genes defines the broad region where a particular Hox gene can become
active (Cerny, 2005).
In Drosophila, Krüppel is expressed in a bell-shaped profile
centered over the primordia of segments T2 to A3. In
the Tribolium blastoderm, only one such gradient is present; the
Tc'Kr domain covers the posterior pole. When
the germ rudiment has formed, the Tc'Kr domain retracts from the
posterior end and forms a distinct domain overlapping the three thoracic
segment primordia. At
this stage, therefore, the Tc'Kr domain covers more anterior segment
primordia (and more anterior pair-rule stripes) than does its
Drosophila counterpart (Cerny, 2005).
The results for Tc'Kr deviate from those obtained for
Krüppel in Oncopeltus fasciatus. In
this short-germ insect, knock-down of Kr also results in
mis-expression of Hox genes, although the effects are more limited as only one
ectopic Of'Dfd domain is detected. Interestingly, expression of
Of'en in such embryos seems to indicate a clear gap phenotype, i.e.
perfect segmental stripes reappear posterior to a region of segmental
disruption. Incomplete inactivation of Of'Kr could be responsible for
this difference; it is noted, however, that weak Tc'Kr RNAi situations do
not result in obvious gap phenotypes. Rather, in such embryos the segmentation process simply breaks down somewhat later than in Tc'Krjaws, i.e., the additional segments present in weak Tc'Kr RNAi embryos appear to represent anterior abdominal rather than posterior (post-gap) abdominal segments.
Oncopeltus is sometimes denoted an intermediate-germ insect, because
a few more segments are formed already in the blastoderm than, for example, in
Tribolium. It will be interesting to see if the 'next posterior' gap
gene in Oncopeltus will also display a 'gap' phenotype, and to find
out whether pair-rule gene expression in Of'Kr RNAi embryos indicates
a role in the regulation of specific stripes boundaries (Cerny, 2005).
In insects, the boundary between the embryonic head and thorax is formed by the dorsal ridge, a fused structure composed of portions of the maxillary and labial segments. However, the mechanisms that promote development of this unusual structure remain a mystery. In Drosophila, mutations in the Hox genes Sex combs reduced and Deformed have been reported to cause abnormal dorsal ridge formation, but the significance of these abnormalities is not clear. Three mutant allele classes of Cephalothorax, the Tribolium castaneum (red flour beetle) ortholog of Sex combs reduced were detected, each of which has a different effect on dorsal ridge development. By using Engrailed expression to monitor dorsal ridge development in these mutants, it has been demonstrated that Cephalothorax promotes the fusion and subsequent dorsolateral extension of the maxillary and labial Engrailed stripes (posterior compartments) during dorsal ridge formation. Molecular and genetic analysis of these alleles indicates that the N terminus of Cephalothorax is important for the fusion step, but is dispensable for Engrailed stripe extension. Thus, specific regions of Cephalothorax are required for discrete steps in dorsal ridge formation (Shippy, 2006; full text of article).
The first evidence of dorsal ridge formation in wild-type embryos is the connection of the maxillary and labial En stripes by the appearance of En expression along the lateral edges of the anterior compartment of the labial segment. Since this 'fusion' does not occur in Cx null embryos, it is conclude that Cx is responsible for the unique behavior of these En stripes. The maxillary and labial En stripes also fail to fuse in Cx20 and CxE homozygotes. Thus, this event apparently requires the N terminus of Cx, as well as expression of Cx in at least one of the domains affected in CxE mutants. At the time the maxillary and labial En stripes begin to fuse, Cx is expressed in the posterior compartment of the maxillary segment and the anterior compartment of the labial segment. CxE homozygotes have little or no Cx expression in the lateral regions of the posterior maxillary segment, but apparently normal expression in the anterior compartment of the labial segment. This could mean that maxillary Cx expression is required for fusion of the En stripes. Taken together, the phenotypes produced by each of these allele classes indicate that fusion of the maxillary and labial En stripes is an essential step in normal head development. One unresolved issue is the mechanism by which these En stripes fuse. Do En-expressing posterior compartment cells move along the edges of the labial segment? If so, do the migrating cells originate from the maxillary or labial segment, or both? Or does En expression appear de novo in anterior compartment cells instead? There have been previous reports of en expression in the anterior compartment of Drosophila wing discs, but in that case hh expression remains limited to the posterior compartment. Likewise, En is expressed in the anterior compartment of the eighth abdominal segment in Drosophila, but hh is not. Tc hh is coexpressed with En along the edges of the labial segment, suggesting that these cells derive from the posterior compartment. However, the simultaneous appearance of En expression along the entire anterior compartment might suggest that 'de novo' expression is more likely (Shippy, 2006).
During dorsal closure in wild-type embryos, En stripes in the thorax and abdomen extend to the dorsolateral edges of the embryo. Likewise, the fused dorsal ridge En stripe (apparently composed of the maxillary and labial En stripes) extends to the dorsolateral edge of the embryo. The maxillary and labial En stripes fail to fuse in all three Cx mutant classes, but the subsequent fate of these stripes differs in each class. In Cx null mutants, neither En stripe extends dorsally. This indicates that Cx is required for dorsolateral extension of En in the maxillary/labial region. The ability to promote dorsolateral En expression seems to be a function shared by many Tribolium Hox genes, since in Df(HOMC) homozygotes (which lack most of the Hox genes) all of the En stripes fail to extend (Shippy, 2006).
In CxE homozygotes, the labial En stripe extends, but the maxillary En stripe does not. This difference seems to correlate with the more severe reduction in Cx expression in the posterior maxillary segment in CxE mutants. In Cx20 homozygotes, the labial En stripe extends dorsolaterally to the edges of the embryo, thus behaving like the thoracic and abdominal En stripes. In addition, a second En stripe appears anterior to the labial En stripe and extends dorsolaterally. This suggests that the Cx protein missing its N terminus cannot promote fusion of the maxillary and labial En stripes, but is sufficient for En stripe extension (Shippy, 2006).
The simplest interpretation of the data is that the N-terminal region of Cx includes a domain that controls maxillary and labial En stripe fusion. Interestingly, the region missing from the Cx20 protein contains the octapeptide motif that is present in many Hox genes. Deletion of the N terminus of Hox-a5 (a mouse Scr homolog) has been shown to decrease its transactivation ability in in vitro assays and its ability to produce homeotic transformations when ectopically expressed in Drosophila. Furthermore, Ultrabithorax lacking its N terminus can repress target genes when expressed in Drosophila, but its ability to activate target genes is greatly reduced. Likewise, deletion of the Scr N terminus reduces its ability to activate target genes. Extrapolating from these observations, the current model is that the N-terminally truncated protein produced from the Cx20 allele is capable of repression but not activation of target genes. This model is consistent with the loss of particular functions seen in Cx20 mutants and predicts that fusion of the maxillary and labial En stripe during dorsal ridge formation requires activation of target genes, while dorsolateral extension of En stripes requires repression of target genes (perhaps genes that negatively regulate en) (Shippy, 2006).
This study has shown that function of the homeotic gene Cx is crucial for formation of the dorsal ridge in Tribolium. Given that the events of dorsal ridge development are conserved in a wide variety of insects, it will be interesting to determine whether the upstream factors governing this process are also conserved. Closer analysis of dorsal ridge formation in Drosophila Scr mutants will be a first step, but perturbation of Scr ortholog function(s) in other insects (perhaps by RNAi) will be important as well (Shippy, 2006).
Specialized insect mouthparts, such as those of Drosophila, are derived from an ancestral mandibulate state, but little is known about the developmental genetics of mandibulate mouthparts. The metamorphic patterning of mandibulate mouthparts of the beetle Tribolium castaneum was studied RNA interference to deplete the expression of 13 genes involved in mouthpart patterning. These data were used to test three hypotheses related to mouthpart development and evolution. First, the prediction was tested that maxillary and labial palps are patterned using conserved components of the leg-patterning network. This hypothesis was strongly supported: depletion of Distal-less and dachshund led to distal and intermediate deletions of these structures while depletion of homothorax led to homeotic transformation of the proximal maxilla and labium, joint formation required the action of Notch signaling components and odd-skipped paralogs, and distal growth and patterning required epidermal growth factor (EGF) signaling. Additionally, depletion of abrupt or pdm/nubbin caused fusions of palp segments. Second, the hypotheses was tested for how adult endites, the inner branches of the maxillary and labial appendages, are formed at metamorphosis. The data reveal that Distal-less, Notch signaling components, and odd-skipped paralogs, but not dachshund, are required for metamorphosis of the maxillary endites. Endite development thus requires components of the limb proximal-distal axis patterning and joint segmentation networks. Finally, adult mandible development is considered in light of the gnathobasic hypothesis. Interestingly, while EGF activity is required for distal, but not proximal, patterning of other appendages, it is required for normal metamorphic growth of the mandibles (Angelini, 2012).
In D. melanogaster, Dll mutants lack maxillary structures and portions of the proboscis (i.e., labium), although Dll expression in the maxillary anlagen is weaker than in the leg or antennal discs. Paralleling the results for T. castaneum, in the horned beetle Onthophagus taurus distal regions of the adult mouthparts were deleted with larval Dll RNAi (Simonnet 2011). The embryonic and metamorphic functions of Dll in T. castaneum are also similar: the gene is required for the development of distal structures at both stages, and during embryogenesis Dll is expressed throughout the developing palps. Interestingly, removal of T. castaneum Dll expression earlier during larval life led to delayed metamorphosis, as well as changes in appendage morphology (Suzuki, 2009). Many insects delay molting after appendage loss to allow time for regeneration, and this dual role of Dll suggests a mechanism linking these processes (Angelini, 2012).
The data from T. castaneum provide evidence for a conserved gap gene role of dac during patterning of mouthparts and legs of this species. dachshund is not expressed in or required for development of the labial and maxillary anlagen of D. melanogaster. In T. castaneum embryos dac is expressed strongly in the proximal maxilla and part of the developing endite. Embryonic dac expression is weaker in the distal maxillary palp and the labium. The current data show a clear metamorphic requirement for dac in the intermediate regions of the maxillary and labial palps, as does a recent study of O. taurus (Simonnet, 2011). A function for dac in the development of an intermediate portion of the maxillary and labial appendages has so far only been observed in these two beetles, while data from two species with specialized mouthparts (the milkweed bug O. fasciatus and D. melanogaster) found that dac is not required for PD patterning of the mouthparts. Thus, comparative data from other species do not support the hypothesis that this mouthpart patterning role is ancestral. However, if mandibulate mouthparts evolved from leg-like structures similarities in the expression and function of genes patterning both legs and mouthparts are expected to be plesiomorphic. This hypothesis can be further tested by examining the role of dac in mouthpart development in additional insect orders, particularly those that retain mandibulate mouthparts, and in other arthropods (Angelini, 2012).
The effects of hth depletion are distinct in different species, but typically involve some degree of homeotic transformation. In D. melanogaster, hth is expressed in the labial discs, but without nuclear expression of its cofactor Extradenticle. Maxillary palps are retained in hth loss-of-function flies, but they may possess bristles typical of legs, indicating a partial proboscis-to-leg transformation. In the cricket Gryllus bimaculatus, which has mandibulate mouthparts, hth depletion causes transformation of proximal mouthpart structures towards antennal identity, with a loss of endites, while distal structures are transformed towards leg identity (Ronco, 2008). hth RNAi in T. castaneum transformed intermediate regions of the maxilla and labium towards distal mouthpart identity. Proximal regions also appeared transformed, but their identity could not be established, while distal regions appeared wild type. In the beetle O. taurus, proximal regions of the labium are transformed towards maxillary endite identity, but distal regions of the labium and the entire maxilla remain relatively unaffected (Angelini, 2012).
These results highlight the similarity between patterning of the maxilla, labium and legs in T. castaneum. Functional data from two species with highly derived mouthpart morphologies, D. melanogaster and the milkweed bug Oncopeltus fasciatus, suggest only limited similarity between mouthpart and leg patterning. One explanation for this low degree of conservation is that evolution of the ancestral patterning mechanism has occurred in concert with the functional and morphological diversification of these mouthparts. A correlation between generative mechanisms and structural morphology has been used as a common null hypothesis, although exceptions in which similar morphologies result from different developmental pathways are documented. Nevertheless, this hypothesis predicts that developmental patterning should be more highly conserved across appendage types in species that retain the ancestral mandibulate mouthpart morphology (Angelini, 2012).
The maxillary and labial palps are an interesting case of serial homology. Despite a difference in overall size, their shape and arrangement of sensillae are similar. The intermediate segments of each palp type are also similar, but differ in number, which suggests that segment number is regulated independently from other morphological traits. The RNAi depletion of pdm in T. castaneum caused the reduction and deletion of the third maxillary palp segment, producing a phenotype closely resembling the wildtype morphology of the labial palps. While a role for pdm in the labium cannot be excluded, the absence of observed labial phenotypes was significant compared to maxillary results. Therefore, it is hypothesized that the difference in the number of palp segments results from specific activation of pdm in the maxillary palp. Loss of function in the Hox gene Deformed during T. castaneum embryogenesis causes a transformation of the larval maxillae towards labial identity. Since Hox genes are the primary determinants of body segment identity, it is proposed that pdm is activated by Deformed, and repressed by the labial Hox gene Sex combs reduced. RNAi targeting pdm in another mandibulate insect, the cricket Acheta domesticus, generated defects in the antenna and legs, but no defects in the mouthparts, despite similar pdm expression in these appendages (Turchyn; 2011; Angelini, 2012).
Endites are a primitive component of arthropod appendages, and they are retained in insect mouthparts, as well as in the mouthparts and thoracic appendages of many crustaceans (Boxshall 2004). At least three hypotheses have been put forward for how endites are patterned, and these hypotheses are not mutually exclusive. The first hypothesis states that multiple PD axes result from redeployment of a PD axis patterning mechanism shared by palps and endites. A second hypothesis posits that endites and appendage segments form by the same mechanism, Notch-mediated in-folding of the cuticle. A third hypothesis states that dac expression initiates endite branching from the main appendage axis. The axis redeployment hypothesis predicts that depletion of genes involved in PD axis patterning will have similar effects on the development of palps and endites. Some support for this hypothesis comes from studies of endite morphogenesis and the expression and function of leg gap genes in the embryos of T. castaneum and the orthopteran Schistocerca americana, but not all data are consistent with it. The segmentation hypothesis predicts that endites will fail to differentiate if genes required for joints are depleted. This hypothesis was posed based on a comparative developmental study of segmented and phyllopodous crustacean limbs. Finally, the dac-mediated hypothesis predicts that depletion of dac will lead to reduced endites. This hypothesis emerged from the observation that dac expression is reiterated along the medial edges of larval endites in the crustacean Triops longicaudatus. Comparative expression data from the isopod Porcellio scaber are also consistent with the dac-mediated hypothesis (Angelini, 2012).
The current data are consistent with predictions of the axis redeployment and segmentation hypotheses but do not support a role for dac in endite metamorphosis. Adult endites were disrupted by depletion of Dll, Krn, the odd-related genes, and Notch signaling, and to a lesser degree hth. In the maxilla depletion of most of these genes led to the failure of the single larval endite to divide into two distinct branches, while in the labium, their depletion caused reduction of the ligula. Their requirement in the endites is consistent with the hypothesis that these structures are generated by redeploying appendage PD axis determinants. Depletion of Notch signaling components and the odd paralogs produced reductions and fusions between palp segments, between the palps and endites, and between the lacinia and galea. Thus, these data are compatible with both the hypothesis that a reiterated PD axis is used to pattern the endites and the hypothesis that endite formation is linked to joint formation. Normal endite development in dac-depleted specimens is inconsistent with the dac-mediated hypothesis (Angelini, 2012).
It is noteworthy that endite specification and the division of the single larval endite into the adult galea and lacinia appear to be separable functions. For example, Ser RNAi resulted in a single endite lobe with lacinia identity medially and galea identity laterally. In contrast, severe Dll RNAi individuals had a single endite that lacked also obvious lacinia identity (Angelini, 2012).
The mandibulate structure of Tribolium mouthparts is the pleisomorphic state for insects and is shared by a majority of insect orders. These mouthparts are characterized by robust mandibles, lacking segmentation. A classic debate in arthropod morphology concerns whether the mandibles of insects and myriapods are derived from a whole appendage or only from proximal appendage regions; the latter are called gnathobasic mandibles. Palps are retained on the mandibles of many crustaceans, making it clear that the biting regions of their mandibles are gnathobasic. Phylogenetic support for the gnathobasic hypothesis comes from phlyogenetic studies that place insects nested within crustaceans (Regier, 2010). The first developmental genetic support for the gnathobasic hypothesis came from the discovery that insect mandibles lack Dll expression. Furthermore, neither mutations in Dll nor its depletion through RNAi have been observed to alter mandible development in insects, including T. castaneum. This evidence has led to widespread acceptance of the gnathobasic hypothesis. Of the 13 genes depleted in this study, two (Krn and hth) produced results that would not be predicted by the most straightforward form of the gnathobasic hypothesis for mandible origins (Angelini, 2012).
Loss of EGF function in insects leads to distal appendage defects, including pretarsal or tarsal deletions. The role of EGF signaling in distal appendage regions is conserved in T. castaneum metamorphosis, since depletion of the EGF ligand Krn leads to reduction of the antennal flagellum, and maxillary and labial palps, as well as to deletion of the pretarsus and malformation of the tarsus. In light of the restriction of Krns role to distal appendage regions and regulation of distal EGF ligand expression by Dll in D. melanogaster, the gnathobasic hypothesis predicts that Krn should not be required for normal development of the mandible in T. castaneum. In contrast to this prediction, depletion of Krn produced a significant reduction in mandible length (Angelini, 2012).
The hypothesis of a gnathobasic mandible also predicts that hth depletion should produce effects in the mandible similar to those in the proximal regions of other appendage types. In T. castaneum, hth RNAi during metamorphosis caused homeotic transformation of proximal regions of the maxilla, labium and legs. However, the mandibles were not affected by hth depletion. In the beetle O. taurus, hth depletion slightly altered mandible shape, but also without apparent homeosis. In contrast, hth RNAi in embryos of the cricket G. bimaculatus transformed the mandible towards a leg-like structure distally and an antenna-like structure proximally, paralleling the transformation observed in other appendages. Because these results come from only two lineages and from different life stages, additional data are needed to determine whether a homeotic role for hth was present ancestrally in insect mandibles (Angelini, 2012).
These data must be weighed alongside other evidence bearing on the gnathobasic hypothesis. In T. castaneum, the lack of phenotypic effects on mandible metamorphosis of other genes in this study is consistent with the gnathobasic hypothesis. In particular, it was observed that mandible metamorphosis was normal following depletion of genes involved in distal growth and patterning or joint formation. Moreover, homology at one biological level, such as anatomy, does not preclude divergence at other levels, such as development. Nevertheless, since developmental genetic studies of Dll and other appendage-patterning genes have been used as strong support for the gnathobasic homology of the insect mandible, the findings of Krn function highlight the difficulties in establishing serial homology based solely on developmental data (Angelini, 2012).
This study provides a genetic model of adult mouthpart development in Tribolium castaneum based on 13 genes. While previous studies have examined patterning in species with derived mouthpart morphologies, T. castaneum retains the pleisomorphic, mandibulate state of insect mouthparts. These results demonstrate the conservation of many gene functions in the maxilla and labium, relative to the legs, thus supporting the interpretation of novel gene functions in groups with derived mouthpart morphology as indicative of their specialized morphogenetic roles in those species. Mandibulate mouthparts such as those of T. castaneum include medial maxillary and labial endites, and the current data are consistent with hypotheses of reiteration in the PD axis and specification by Notch signaling, but rule out a direct role for dac in branch generation or patterning at metamorphosis. Additionally these results demonstrate that a regulator of distal leg development, Krn, which encodes an EGF ligand, is required for normal mandible elongation. This finding underscores the complex relationship between homology at the levels of anatomy and developmental patterning (Angelini, 2012).
The establishment of the anterior-posterior segmentation in insects requires the concerted action of a hierarchical gene network. The orthologue of Kruppel gap gene was studied in the hemipteran Rhodnius prolixus (Rp-Kr). Its structure, expression pattern and function were characterized. The genomic sequence upstream of the Rp-Kr transcriptional unit shows a putative regulatory region conserved in the orthologue genes from Drosophila melanogaster and Tribolium castaneum. Rp-Kr expression is zygotic and it is expressed in the anterior half of the embryo (the posterior half of the egg) during the blastoderm stage and germ band formation; later, during germ band extension, it is expressed in a central domain, from T2 to A3. The Rp-Kr loss of function phenotypes shows disrupted thoracic and abdominal segmentation. Embryos with weak segmentation phenotypes show homeotic transformations, in which an ectopic tibial comb, typical of T1 leg, appears in T2, which correlates with the ectopic expression of Rp-sex-comb reduced in this leg. It is concluded that Kruppel of Rhodnius prolixus is required for segmentation and for repression of the homeotic gene sex comb-reduced (Lavore, 2014).
Hox genes control the choice of cell fates along the anteroposterior (AP) body
axis of many organisms. In C. elegans, two Hox genes, lin-39 and mab-5, control
the cell fusion decision of the 12 ventrally located Pn.p cells. Specific Pn.p
cells fuse with an epidermal syncytium, hyp7, in a sexually dimorphic pattern.
In hermaphrodites, Pn.p cells in the mid-body region remain unfused whereas in
males, Pn.p cells adopt an alternating pattern of syncytial and unfused fates.
The complexity of these fusion patterns arises because the activities of these
two Hox proteins are regulated in a sex-specific manner. MAB-5 activity is
inhibited in hermaphrodite Pn.p cells and thus MAB-5 normally only affects the
male Pn.p fusion pattern. A gene has been identified, ref-1, that regulates the hermaphrodite Pn.p cell fusion pattern largely by regulating MAB-5 activity in
these cells. Mutation of ref-1 also affects the fate of other epidermal cells in distinct AP body regions. ref-1 encodes a protein with two basic
helix-loop-helix domains distantly related to those of the hairy/Enhancer of
split family. ref-1, and another hairy homolog, lin-22, regulate similar cell fate decisions in different body regions along the C. elegans AP body axis (Alper, 2001).
Much of the C. elegans epidermal layer, the hypodermis, is composed
of several multinucleate cells (syncytia) that are formed by
the fusion of mononucleate cells throughout embryonic and
postembryonic development. One such syncytium, hyp7, extends over most of the length of
the worm and contains 133 nuclei, close to 15% of all somatic
nuclei in the worm. How is the fusion of all these cells
coordinately regulated to allow formation of hyp7?
To understand how the hyp7 syncytium is generated, the regulation of the fusion decision of one
group of cells called the Pn.p cells that line the ventral surface
of the worm during the first larval stage (L1) has been studied. Pn.p
cell fusion is regulated by two genes of the C. elegans Hox
gene cluster. The Hox cluster consists of six genes: ceh-13, lin-39 and mab-5, homologs of Drosophila labial, Sex combs reduced and Antennapedia, respectively, and egl-5, php-3 and
nob-1, three Abdominal-B homologs. In C. elegans, as in other organisms, the
Hox genes regulate the choice of cell fates along the AP body
axis. However, the simple Hox gene
expression pattern in C. elegans is insufficient to explain the
complex Pn.p cell fusion pattern. This is due to the sex-specific,
post-translational regulation of two Hox genes, lin-39
and mab-5. In hermaphrodites, MAB-5 is
inactive and only LIN-39 influences Pn.p cell fusion fate. In
males, both LIN-39 and MAB-5 are active, but the two proteins
interact in an unusual way to control cell fusion. It is quite
likely that in most species, Hox proteins interact with each
other and with other factors to generate more complexity than
their expression patterns alone would allow (Alper, 2001).
Understanding how these interactions modulate Hox protein
activity is therefore necessary to understand fully how an
animal body plan is laid out.
At the end of the first larval stage, some of the 12 Pn.p cells
fuse with the hyp7 syncytium in a sex-specific pattern. In
hermaphrodites, anterior (P1.p and P2.p) and posterior P(9-11).p cells fuse with the hyp7 syncytium while the six central
cells P(3-8).p remain unfused. These six unfused cells, the vulval precursor cells,
remain competent to develop further, and some of these cells
generate the hermaphrodite vulva later in development. The
Pn.p cell fusion pattern is different in males, with P1.p, P2.p,
P7.p and P8.p fusing with hyp7 and P(3-6).p and P(9-11).p
remaining unfused. The posterior unfused
cells generate male-specific copulatory structures later in
development (Alper, 2001).
Two Hox genes, lin-39 and mab-5, are known to influence
Pn.p cell fusion. lin-39 is expressed in P(3-8).p in both
hermaphrodites and males. In hermaphrodites, lin-39 prevents fusion
of those Pn.p cells in which it is expressed and therefore P(3-8).p remain unfused. Thus, in a lin-39 mutant, all hermaphrodite Pn.p cells
fuse with the hyp7 syncytium and are unable to generate a
vulva. The regulation of Pn.p cell fusion in males is more
complex because both lin-39 and mab-5 can affect the fusion
decision. mab-5 is expressed in P(7-11).p in both sexes, but
only functions in males. Acting alone, either Hox gene is able to prevent fusion of those cells within which it is expressed: P(3-6).p for lin-39 and P(9-
11).p for mab-5. However, when cells express both
Hox genes (P7.p and P8.p), those cells fuse with hyp7, much
like cells that contain neither Hox gene (P1.p and P2.p). The ability of these two Hox genes to negate each other's effects in males occurs post-translationally; that is, LIN-39 and MAB-5 proteins can somehow inhibit each
other's activity when both proteins are present in the same cell.
Moreover, the relative levels of the two proteins do not matter
because the two proteins are still capable of inhibiting each
other when one of the Hox genes is strongly overexpressed. This result argues against a model in
which the two Hox proteins simply sequester each other and, as
a consequence, titrate each other's activity. Instead, something
else appears to be limiting in this cell fate decision. One
possibility is that both proteins bind to regulatory sites in the
same target gene, which in turn encodes a protein that directly
affects cell fusion. In this model, the binding of either protein
alone influences the activity of the fusion gene, whereas the
binding of both Hox proteins together does not (Alper, 2001).
In summary, Hox protein activity is regulated in two key
ways to control the Pn.p cell fusion decision. (1) MAB-5 is
present in the same cells in both sexes but only functions in
male Pn.p cells. Thus, something keeps MAB-5 inactive in the
hermaphrodite Pn.p cells. (2) Both Hox proteins can
interact to inhibit each other when present in the same Pn.p cell
in males. To identify genes that affect Pn.p cell fusion by regulating
Hox protein activity, mutations were isolated that alter the Pn.p
cell fusion pattern. One such mutation, ref-1(mu220)
(REgulator of Fusion-1) prevents fusion of posterior Pn.p cells
in hermaphrodites, largely, but not completely, by affecting the
sex-specific activity of MAB-5. ref-1 mutants also exhibit a
defect in the specification of the fate of a hypodermal cell
located on the lateral surface of the worm in this same posterior
body region as well as other defects in the anterior part of the
worm. ref-1 has been cloned and it encodes a
transcription factor with two basic helix-loop-helix (bHLH)
domains, both of which are distantly related to the hairy/Enhancer of split [E(spl)] subfamily of such proteins (Alper, 2001).
Hox transcription factors have been implicated in playing a central role in the evolution of animal morphology. Many studies indicate the
evolutionary importance of regulatory changes in Hox genes, but little is known about the role of functional changes in Hox proteins. In
the nematodes Pristionchus pacificus and Caenorhabditis elegans, developmental processes can be compared at the cellular, genetic,
and molecular levels and differences in gene function can be identified. The Hox gene lin-39 is involved in the regulation of nematode
vulva development. Comparison of known lin-39 mutations in P. pacificus and C. elegans reveals both conservation and changes of
gene function. Evolutionary changes of lin-39 function were studied using hybrid transgenes and site-directed mutagenesis in an in vivo assay using C. elegans lin-39 mutants. Despite the functional differences of LIN-39 between the two species, Ppa-LIN-39, when driven by Cel-lin-39 regulatory
elements, can functionally replace Cel-lin-39. Furthermore, the MAPK docking and phosphorylation motifs unique for Cel-LIN-39 are dispensable for
Cel-lin-39 function. Therefore, the evolution of lin-39 function is driven by changes in regulatory elements rather than changes in the protein itself (Grandien, 2001).
Development of the vulva in C. elegans is mediated by the
combinatorial action of several convergent regulatory inputs, three of which
(the Ras, Wnt and Rb-related pathways) act by regulating expression of the
lin-39 Hox gene. LIN-39 specifies cell fates and regulates cell
fusion in the mid-body region, leading to formation of the vulva. In the
lateral seam epidermis, differentiation and cell fusion have been shown to be
regulated by two GATA-type transcription factors, ELT-5 and -6.
ELT-5 is encoded by the egl-18 gene, which
promotes formation of a functional vulva. Furthermore, EGL-18
(ELT-5), and its paralogue ELT-6, are redundantly required to regulate cell
fates and fusion in the vulval primordium and are essential to form a vulva.
Elimination of egl-18 and elt-6 activity results in arrest
by the first larval stage; however, in animals rescued for this larval
lethality by expression of ELT-6 in non-vulval cells, the post-embryonic cells
(P3.p-P8.p) that normally become vulval precursor cells often fuse with the
surrounding epidermal syncytium or undergo fewer than normal cell divisions,
reminiscent of lin-39 mutants. Moreover, egl-18/elt-6
reporter gene expression in the developing vulva is attenuated in
lin-39(rf) mutants, and overexpression of egl-18 can
partially rescue the vulval defects caused by reduced lin-39
activity. LIN-39/CEH-20 heterodimers bind two consensus HOX/PBC sites in a
vulval enhancer region of egl-18/elt-6, one of which is essential for
vulval expression of egl-18/elt-6 reporter constructs. These findings
demonstrate that the EGL-18 and ELT-6 GATA factors are essential, genetically
redundant regulators of cell fates and fusion in the developing vulva and are
apparent direct transcriptional targets of the LIN-39 Hox protein (Kohl, 2002).
Hox proteins appear to require co-factors to achieve DNA-binding
specificity. The most extensively studied of the Hox co-factor genes are
the Drosophila extradenticle (exd) and mammalian pre-B
cell homeobox 1 genes, collectively referred to as PBC genes. Hox and PBC
proteins form heterodimers that bind DNA in vitro. C. elegans
contains one known Exd homolog, CEH-20, which appears to act as a Hox
co-factor. Consistent with the possibility that egl-18 and
elt-6 are direct targets of LIN-39 Hox, several consensus
Hox/PBC-binding sites (TGATNNAT) were found in the egl-18 and elt-6 genomic region. Two of these [site 1 (TGATATAT) and site 2 (TGATTGAT)] are present in intron 2 of egl-18, which is included in the ~800 bp promoter element that directs GFP expression in the VPC lineages and VC neurons. Several lines of evidence indicate that site 1, but not site 2, is important for vulval-specific expression of egl-18/elt-6: (1) alteration of 6 bp in site 1 eliminates expression in the VPC lineages and VC neurons, whereas a similar mutation that alters 4 bp of site 2 has no obvious effect on reporter expression; (2) a reporter in which 544 base pairs surrounding only site 1 is present showed expression in the vulva and VC neurons, albeit at an attenuated level compared with the reporter containing both sites; (3) mutation of site 1 from this construct eliminated vulval and VC expression, and (4) comparison of the egl-18 sequence of C. elegans and C. briggsae revealed a highly conserved 27 bp element surrounding Site 1 but no conservation of site 2. Thus, the site 1 Hox/PBC site is apparently necessary and sufficient for vulva-specific expression of
egl-18/elt-6::GFP (Kohl, 2002).
Electrophoretic mobility shift assays were performed to test the hypothesis
that egl-18 and elt-6 are direct targets of LIN-39/CEH-20
heterodimers in the vulva. Indeed, LIN-39 and CEH-20
heterodimers bind in vitro to 30 bp oligonucleotides centered on either the
Hox/PBC site 1 or site 2. Whereas binding of LIN-39/CEH-20 to site 1 oligos could be competed away with excess unlabeled site 1 or 2 oligos, unlabeled site 1 oligos could not compete with site 2 oligos, implying that site 2
has a higher in vitro affinity for LIN-39/CEH-20 than does site 1. These results indicate that LIN-39/CEH-20 heterodimers can bind cooperatively to site 1, which is essential for expression of the egl-18/elt-6 reporter in the vulva. Based on these results and the phenotypes of egl-18/elt-6
mutants, it seems likely that LIN-39 regulates vulval development by directly
activating EGL-18 and ELT-6, which in turn repress epidermal fusion and
activate vulval differentiation (Kohl, 2002).
The tumor suppressor Rb and the NuRD (nucleosome remodeling and histone deacetylation) complex have been implicated in transcriptional repression during cell cycle progression and cell fate specification. The Rb/E2F complex physically interacts with and thus recruits the NuRD complex to actively repress transcription. C. elegans counterparts of Rb, E2F/DP, and some NuRD complex components appear to function in a common class B synthetic Multivulva (synMuv) pathway to antagonize RTK/Ras signaling during vulval fate specification. Therefore, it has been suggested that they function together in a single complex to repress vulva-specific gene transcription. However, little is known about the in vivo interactions between these class B synMuv genes and their relationships with other pathways in specific cellular processes during vulval development. It has been shown that C. elegans Rb/E2F and NuRD complexes antagonize Ras activity by controlling a lin-39 Hox-mediated cell fusion event that regulates the competence of vulval cells. Interestingly, Rb/E2F and NuRD complexes exhibit very different genetic properties. While the NuRD complex negatively regulates lin-39 Hox activity, likely by downregulating its expression, RB/E2F appears to play dual roles in regulating lin-39: a negative role in controlling its activity and a previously uncharacterized positive role in regulating its expression (Chen, 2001).
The Ras signaling pathway specifies a variety of cell fates in many organisms. However, little
is known about the genes that function downstream of the conserved signaling cassette, or
what imparts the specificity necessary to cause Ras activation to trigger different responses
in different tissues. In C. elegans, activation of the Ras pathway induces cells in the central
body region to generate the vulva. Vulval induction takes place in the domain of the Hox
gene lin-39, a homolog of Drosophila Sexcombs reduced. lin-39 is absolutely required for Ras signaling to induce
vulval development. During vulval induction, the Ras pathway, together with basal lin-39
activity, up-regulates lin-39 expression in vulval precursor cells. If lin-39
function is absent at this time, no vulval cell divisions occur. If lin-39 is
replaced with the posterior Hox gene mab-5, then posterior structures are induced instead
of a vulva (Maloof, 1998).
Animal lacking lin-1, which encodes an ETS-like transcription factor, have a multivulva phenotype: in lin-1 mutants, all the vulval precursor cells generate vulval lineages in an anchor-cell independent fashion. Because lin-39 increases in lin-1 mutants, it seems likely that Lin-39 protein acts downstream of lin-1, and thus is required for the multivulva phenotype of lin-1.
Lin39 alone cannot trigger vulval development. Thus, the Ras pathway must have other functions in vulval development in addition to inducing lin-39 expression These findings suggest that in addition to permitting vulval cell divisions to occur, lin-39 is also required to specify the outcome of Ras signaling by selectively activating vulva-specific genes (Maloof, 1998).
Hox genes pattern the fates of the ventral ectodermal Pn.p
cells that lie along the anteroposterior (A/P) body axis of C.
elegans. In these cells, the Hox genes are expressed in
sequential overlapping domains where they control the
ability of each Pn.p cell to fuse with the surrounding
syncytial epidermis. The activities of Hox proteins are sex-specific
in this tissue, resulting in sex-specific patterns of
cell fusion: in hermaphrodites, the mid-body cells remain
unfused, whereas in males, alternating domains of syncytial
and unfused cells develop. The wild-type Pn.p cell fusion pattern
arises through the regulation of both Hox gene expression
and Hox protein activity. In both wild-type males and
hermaphrodites, lin-39 (Scr homolog) is expressed in P(3-8).p in the mid-body
and mab-5 (Antp homolog) is expressed in an overlapping domain that spans P(7-11).p in the posterior. At the beginning of the second larval stage,
only P(3-8).p in the mid-body remains unfused in wild-type
hermaphrodites. The
posterior Pn.p cells adopt the syncytial fate despite mab-5
expression because the activity of MAB-5 protein is inhibited in
hermaphrodite Pn.p cells (Chng, 1999 and references).
The gene egl-27,
which encodes a C. elegans homolog of a chromatin
regulatory factor, specifies these patterns by regulating
both Hox gene expression and Hox protein function.
Mutations in the C. elegans gene egl-27 were first isolated in
a screen for egg-laying defective mutants.
More recently, egl-27 has been found to encode nuclear protein
isoforms similar to a protein encoded by mta1, whose expression is elevated in
metastatic tumor cell lines and functions as a component of
NURD, a human chromatin regulatory complex. egl-27 appears to be widely expressed
and is involved in the control of cell polarity, cell migration
and embryonic morphogenesis (Chng, 1999 and references).
In egl-27
mutants, the expression domains of Hox genes in these
cells are shifted posteriorly, suggesting that egl-27
influences A/P positional information. In addition, egl-27
controls Hox protein function in the Pn.p cells in two ways:
in hermaphrodites, it inhibits MAB-5 activity, whereas in
males it permits a combinatorial interaction between LIN-39
and MAB-5. Thus, by selectively modifying the activities
of Hox proteins, egl-27 elaborates a simple Hox expression
pattern into complex patterns of cell fates. Taken together,
these results implicate egl-27 in the diversification of cell
fates along the A/P axis and suggest that chromatin
reorganization is necessary for controlling Hox gene
expression and Hox protein function (Chng, 1999).
egl-27 is required for both of the
post-translational forms of Hox regulation that occur in the
Pn.p cells. egl-27 is required for the inhibition of MAB-5
activity in hermaphrodite Pn.p cells. In C. elegans
hermaphrodites, the sex-determination pathway culminates in
the activation of tra-1, which acts cell-autonomously to specify
hermaphrodite-specific cell fates. Since tra-1 encodes zinc-finger
transcription factor isoforms, it has been postulated
that tra-1 might directly interfere with MAB-5s ability to bind
a promoter or activate transcription. Since there is no logical
necessity for additional component(s) outside the sex-determination
pathway for MAB-5 inhibition, it is
surprising that this process also requires egl-27. egl-27 is not
specifically involved in sex-determination because most of its
other phenotypes occur in both sexes. Since MAB-5 inhibition
involves both the sex-determination pathway and egl-27, tra-1
(or another sex-specific factor) might interact directly or
indirectly with egl-27 to inhibit MAB-5.
egl-27 is also required for the combinatorial interaction
between LIN-39 and MAB-5 in male Pn.p cells.
When multiple Hox genes function to specify a single body
part, it is difficult to distinguish whether they do so by acting
in distinct cells within that structure or by a combinatorial
interaction within the same cells. In C. elegans, it was possible to bypass this complication by examining the fate of single
Pn.p cells that are controlled cell-autonomously by mab-5 and
lin-39 (Chng, 1999).
How do LIN-39 and MAB-5 neutralize each other in this
combinatorial interaction? This post-translational process is not
regulated by nuclear access or protein degradation because it
occurs when both LIN-39 and MAB-5 proteins are present in
the nucleus of the same Pn.p cell. Moreover, this neutralization has been
shown to be insensitive to the relative levels of LIN-39 or MAB-5,
arguing against the formation of inactive LIN-39/MAB-5
heterodimers or competition between LIN-39 and MAB-5 for
target promoter(s) or co-factor(s). Instead,
this suggests that the two proteins might act together on another
target that is limiting, such as a promoter that could integrate
their activities.
EGL-27 (or other components of the NURD complex)
might interact directly or indirectly with MAB-5 and/or LIN-39
on a promoter of a cell fusion target gene in a manner
dependent on the activity of tra-1 (or another sex-specific
factor). Recruiting EGL-27 and the NURD complex to this
promoter could direct its chromatin reorganization activities to
this promoter and thus alter the transcriptional state of the target
gene, resulting in different Pn.p fates (Chng, 1999).
Members of the Hox family of homeoproteins and their
cofactors play a central role in pattern formation of all
germ layers. During postembryonic development of C.
elegans, non-gonadal mesoderm arises from a single
mesoblast cell M. Starting in the first larval stage, M
divides to produce 14 striated muscles, 16 non-striated
muscles, and two non-muscle cells (coelomocytes). The role of the C. elegans Hox cluster and of the exd ortholog ceh-20 in patterning of the postembryonic
mesoderm has been investigated. By examining the M lineage and its
differentiation products in different Hox mutant
combinations, an essential but overlapping role was found
for two of the Hox cluster genes, lin-39 (Scr homolog) and mab-5 (Antp homolog), in
diversification of the postembryonic mesoderm. This role
of the two Hox gene products requires the CEH-20
cofactor. One target of these two Hox genes is the C. elegans
twist ortholog hlh-8. Using both in vitro and in vivo assays,
it has been demonstrated that twist is a direct target of Hox
activation. Evidence from mutant phenotypes is presented
that twist is not the only target for Hox genes in the M
lineage: in particular lin-39 mab-5 double
mutants exhibit a more severe M lineage defect than the
hlh-8 null mutant (Liu, 2000).
The M lineage defects of mutants of Hox genes lin-39 and mab-5 combined with ceh-20 mutants are intriguing. These defects do not appear to
be a result of homeotic transformation of the fate of M or its
descendants. Instead, the mutants exhibit either (1) a loss of all
differentiated M-lineage descendants or (2) the precocious
production of abnormal mesodermal fates with certain
properties of later M lineage products. The precocious
appearance of large cells that exhibit SM- and sex muscle-like
characteristics suggests that this program might be a default
state of M in the absence of Hox function (Liu, 2000).
The shared role of MAB-5 and LIN-39 in the M lineage
appearsto be specific to these two Hox factors for the following reasons: (1) forced
expression of either lin-39 or mab-5, but not of the neighboring
Hox genes ceh-13 and egl-5, is sufficient to activate ectopic
expression of M lineage reporters; (2) egl-5 mutants
(which are viable either alone or in combination with lin-39
and mab-5 mutants) have no M lineage defects on their own and
show no synergistic effects with lin-39 and mab-5.
Mesodermal roles of Hox and exd genes have also been
shown in Drosophila. In the visceral mesoderm, Ubx and abd-A
are involved in morphogenesis of the midgut. In this case, a
few targets for Hox genes have been described: Ubx in the
visceral mesoderm is directly required with an Exd cofactor
for activating expression of the signaling molecule dpp. In the somatic
mesoderm, Ubx and abd-A can each promote the formation
of specific sets of muscle precursors. None of the Drosophila Hox mutants or
combinations that have been analyzed show as drastic an
effect on postembryonic mesoderm as was seen with the
lin-39 mab-5 double mutant in C. elegans. This apparent
discrepancy may reflect a fundamental difference between
the biological systems; alternatively, a more drastic
postembryonic requirement for Hox factors in the Drosophila
mesoderm might have been missed due to the embryonic
lethality of multiple-Hox mutants (Liu, 2000 and references therein).
The rescue experiments suggest partially overlapping roles
for mab-5 and lin-39 in the M lineage. The modest M-lineage
defects seen in mab-5 single mutants, compared with the lack
of any M-lineage defects in lin-39 single mutants suggest that
under normal circumstances the contribution of mab-5 may be
somewhat more substantial at early time points. One
conceivable explanation for the 'either/or' requirement would
involve cross-regulation between Hox genes. In particular, the possibility that lin-39 expression in the M lineage only occurs in the absence of functional mab-5 was tested. This is apparently not the case, since mab-5 mutants show an
apparently normal pattern of M lineage staining with antibodies to LIN-39 (Liu, 2000).
Several types of interactions between lin-39 and mab-5
activities in determining cell fate have been reported. In a
subset of Pn.aap cells that normally express both lin-39 and
mab-5, the lin-39 activity is dominant, preventing mab-5 from
functioning in these cells. A distinct interaction is seen in male Pn.p cells, where
lin-39 and mab-5 are both expressed and act combinatorially
to specify a fate that is different from that specified by either
alone. A third situation is seen in hermaphrodite vulval precursor cells, for which the loss of Hox (lin-39) activity after specification results in a failure to
differentiate; in this lineage, lin-39 and mab-5 activities have
the capability to promote distinct and non-overlapping
consequences in terms of cell fate. The functional and
simultaneous requirement in the M lineage for either mab-5 or
lin-39 function represents a further degree of freedom in using
these genes to build an organism (Liu, 2000).
The highly conserved structure of Hox factors is consistent
with a view that these genes have evolved by duplication of
a single precursor gene. Under these circumstances, it is not surprising that certain roles for Hox factors would still be maintained as shared (or redundant)
between several genes in the cluster. While the individual
genes might have acquired position-specific roles based on
their acquisition of intricate patterns of expression, it is
certainly conceivable that the entire family (or a large subset)
will have maintained a shared role equivalent to that of the
ancestral (and unique) Hox factor. While the role of that factor
will remain a mystery, the appearance of Hox factors in the
developing embryo just prior to the start of differentiation
suggests that the ancestral factor could have played a role in
developmental timing, perhaps modulating the start of
differentiation in a subset of cells (Liu, 2000).
The C. elegans twist ortholog hlh-8 is a direct and
critical target of Hox genes and ceh-20 in the
postembryonic M lineage. A critical site has been identified in the hlh-8
promoter that is a binding site for the LIN-39/CEH-20 protein
complex. The similarity between core binding sequences for
Drosophila Antp and Dfd proteins in vitro, and the functional equivalence of mab-5 and lin-39 in activating hlh-8 expression in the M lineage, strongly suggest
that this site is also a binding site for MAB-5/CEH-20.
Although hlh-8 is a target for Hox/CEH-20 function in the
M lineage, it is not the only such target. Several indirect
observations demonstrate the existence of additional targets.
One line of evidence comes from the observation that forced
expression of hlh-8 in lin-39 mab-5 mutants
fails to rescue the M lineage defects. An independent line of
evidence comes from a comparison of mutant phenotypes: lin-39 mab-5 mutants show a more severe patterning defect in the M lineage than null hlh-8
mutants: (1) while lin-39 mab-5 animals lack both M-derived coelomocytes, the
majority of hlh-8 mutants contain normal
numbers of M-derived coelomocytes; (2) while lin-39 mab-5 mutants lack all M-derived bodywall muscle, hlh-8 mutants produce variable number of these cells;
(3) sex muscles can be produced in hlh-8 mutants,
although they are not fully differentiated.
The identity of other Hox targets in the M lineage is not known (Liu, 2000).
During Caenorhabditis elegans vulval development, activation
of receptor tyrosine kinase/Ras and Notch signaling pathways causes
three vulval precursor cells (VPCs) to adopt induced cell fates. A Wnt
signaling pathway is also activated in cell fate specification within the VPCs, via
regulation of the Hox gene lin-39. Either
mutation of pry-1 or expression of an activated BAR-1 ß-catenin protein causes an Overinduced phenotype, in which greater than three VPCs adopt induced cell fates. This indicates that pry-1, which encodes a C. elegans axin homolog, acts as
a negative regulator of Wnt signaling in the VPCs. Loss of activity of
the APC homolog apr-1 increases the penetrance of this
Overinduced phenotype, suggesting that APR-1 may play a negative role
in Wnt signaling in this process in C. elegans, similar to APC
proteins in other systems. The Overinduced phenotype is suppressed by
reduction of function of the genes pop-1 TCF and lin-39
Hox. Surprisingly, the Overinduced phenotype caused by
hyperactivated Wnt signaling is not dependent on signaling through the Ras pathway. These data suggest that hyperactivation of Wnt signaling is sufficient to cause VPCs to adopt induced fates and that a canonical Wnt pathway may play an important role during C. elegans vulval induction (Gleason, 2002).
How does overactivation of the Wnt pathway lead to extra vulval induction, even when activation of the Ras pathway has been compromised? It is believed that
overexpression of one or more Wnt target genes causes these extra inductions. Currently, the only known target of the Wnt pathway in the VPCs is lin-39. It is also known that LIN-39 protein levels increase in P6.p in a Ras pathway-dependent manner. This suggests that LIN-39 may function in the adoption of induced vulval fates when expressed at higher levels. Therefore, it is proposed that overactivation of the Wnt
pathway may cause levels of LIN-39 to exceed some threshold in P3.p, P4.p, and P8.p, causing those cells to sometimes adopt induced cell fates. In this
model, the Wnt pathway normally plays a permissive role in the maintenance of LIN-39 expression in all VPCs to prevent these cells from adopting the Fused fate,
but overactivation of the Wnt pathway can phenocopy a cell in which both the Wnt and Ras pathways are active. This would be consistent with the dependence of
the Overinduced phenotype on lin-39 activity in these experiments, and the independence of that phenotype with regard to Ras signaling. Preliminary experiments have suggested that expression of high amounts of LIN-39 alone in L2/L3 larvae is not sufficient to phenocopy the Overinduced phenotype described here. This suggests that there may be additional targets of the Wnt pathway that contribute to the adoption of induced fates when Wnt signaling is overactivated. Future experiments will attempt to identify target genes regulated by overactivation of the Wnt pathway, as well as to determine the functional relationship between factors acting downstream of Wnt signaling, such as LIN-39, and transcription factors known to act downstream of Ras signaling during vulval induction, such as the winged helix protein LIN-31 and the Ets domain protein LIN-1 (Gleason, 2002).
The Hox gene lin-39 plays a crucial role in the establishment of
the nematode vulva equivalence group. Mutations in lin-39 in
Caenorhabditis elegans and Pristionchus pacificus result in
a vulvaless phenotype because presumptive vulva precursor cells adopt
non-vulval fates. Interestingly, the non-vulval fate of anterior and posterior
epidermal cells differs between Caenorhabditis and
Pristionchus; in C. elegans, non-vulval cells fuse with the
hypodermis, whereas, in P. pacificus, they die as a result of
programmed cell death. C. elegans lin-39 (Cel-lin-39)
indirectly controls the cell fusion gene eff-1 by regulating the GATA
transcription factors egl-18 and elt-6. In P.
pacificus, the genetic context of its lin-39 (Ppa-lin-39) function was unknown. This study describes the isolation and characterization of gev-2, a second generation-vulvaless mutant in P. pacificus. gev-2 is the Ppa-pax-3 gene, and it has distinct functions in the cell fate specification of epidermal cells. Whereas Ppa-pax-3 regulates cell survival of the presumptive vulval precursor cells, it controls cell death of posterior epidermal cells. Molecular studies indicate that Ppa-pax-3 is a direct target of Ppa-LIN-39. Thus, this study describes the first specific developmental defect of a nematode pax-3 gene and the data reveal different regulatory networks for the specification of the vulva equivalence group (Yi, 2007).
Hox genes encode master regulators of regional fate specification during early metazoan development. Much is known about the initiation and regulation of Hox gene expression in Drosophila and vertebrates, but less is known in the non-arthropod invertebrate model system, C. elegans. The C. elegans Hox gene lin-39 (homolog of Drosophila Sex-combs reduced) is required for correct fate specification in the midbody region, including the Vulval Precursor Cells (VPCs). To better understand lin-39 regulation and function, transcription factors necessary for lin-39 expression in the VPCs, factors were sought that initiate lin-39 expression in the embryo. The yeast one-hybrid (Y1H) method was used to screen for factors that bound to 13 fragments from the lin-39 region: twelve fragments contained sequences conserved between C. elegans and two other nematode species, while one fragment was known to drive reporter gene expression in the early embryo in cells that generate the VPCs. Sixteen transcription factors that bind to eight lin-39 genomic fragments were identified in yeast, and several factors were characterized by verifying their physical interactions in vitro, and showing that reduction of their function leads to alterations in lin-39 levels and lin-39::GFP reporter expression in vivo. Three factors, the orphan nuclear hormone receptor NHR-43, the hypodermal fate regulator LIN-26, and the GATA factor ELT-6 positively regulate lin-39 expression in the embryonic precursors to the VPCs. In particular, ELT-6 interacts with an enhancer that drives GFP expression in the early embryo, and the ELT-6 site that was identified is necessary for proper embryonic expression. These three factors, along with the factors ZTF-17, BED-3 and TBX-9, also positively regulate lin-39 expression in the larval VPCs. These results significantly expand the number of factors known to directly bind and regulate lin-39 expression, identify the first factors required for lin-39 expression in the embryo, and hint at a positive feedback mechanism involving GATA factors that maintains lin-39 expression in the vulval lineage. This work indicates that, as in other organisms, the regulation of Hox gene expression in C. elegans is complicated, redundant and robust (Liu, 2014).
In the freshwater planarian Dugesia japonica, five cDNAs for HOM/HOX homeobox genes, (Plox or planarian HOM/HOX homeobox genes) were cloned and sequenced. Together with sequence data on platyhelminthes
HOM/HOX homeobox genes that have been deposited in databases, comparison of the deduced amino acid sequences reveals that planarians have at least
seven HOM/HOX homeobox genes, Plox1 to Plox7. Plox1, Plox3 and Plox7 show high similarity to the fly genes labial (82% amino acid identity in the homeodomain), Dfd (82%) and abd-a (88%), respectively. Plox4 may correspond to Sex combs reduced (Scr) rather than Dfd (75%), because of Plox4's high similarity with nematode mab5 (67%), leech Lox5-hro (77%), and paralog group 4 in vertebrates; planarian chich corresponds to Scr in the fly. Plox2 could not be classified. Both Plox5 and Plox6 resemble Antp in the fly. Whole-mount in situ hybridization and RT-PCR reveal that Plox4 (72% homologous to Sex combs reduced)
and Plox5 (77% homologous to Scr) are increasingly expressed along a spatial gradient in the posterior region of intact animals. During regeneration, Plox5 is expressed only in the
posterior region of regenerating body pieces, suggesting that the gene is involved in the anteroposterior patterning in planarians. Plox5 is not found to be expressed
in a blastema-specific manner. X-ray irradiation experiments show that at a minimum, Plox5 is expressed in some cells other than neoblasts, but that the induction of Plox5 expression during
regeneration might require neoblasts. Chromosome walking data are needed to understand the genomic organization and relationships of planarian HOM/HOX homeobox genes to those of other animal groups (Orii, 1999).
Molecular developmental studies of fly and mouse embryos have shown that the identity of individual
body segments is controlled by a suite of homeobox-containing genes called the Hox cluster. To
examine the conservation of this patterning mechanism in other segmented phyla,
four Hox gene homologs isolated from glossiphoniid leeches of the genus Helobdella are described. Based on
sequence similarity and phylogenetic analysis, the leech genes Lox7, Lox6, Lox20, and Lox5 are
deemed to be orthologs of the Drosophila genes lab, Dfd, Scr, and Antp, respectively. Sequence
similarities between Lox5 and Antp outside the homeodomain and phylogenetic reconstructions suggest
that the Antennapedia family of Hox genes had already expanded to
include at least two discrete Antp and Ubx/abdA precursors prior to the annelid/arthropod divergence.
In situ hybridization reveals that the four Lox genes described in this study are all expressed at high
levels within the segmented portion of the central nervous system (CNS), with variable levels of
expression in the segmental mesoderm. Little or no expression is seen in peripheral ectoderm or
endoderm, or in the unsegmented head region (prostomium). Each Lox gene has a distinct anterior
expression boundary within one of the four rostral segments, and the anterior-posterior (AP) order of
these expression boundaries is identical to that reported for the orthologous Hox gene products in fly
and mouse. This finding supports the idea that the process of AP axis differentiation is conserved
among the higher metazoan phyla with respect to the regional expression of individual Hox genes along
that axis. One unusual feature of leech Hox genes is the observation that some genes are only
expressed during later development -- beginning at the time of terminal cell differentiation -- whereas
others begin expression at a much earlier stage; their RNA ceases to be detectable shortly after
the onset of expression of the 'late' Hox genes. The functional significance of this temporal disparity is
unknown, but it is noteworthy that only the two 'early' Hox genes display high levels of mesodermal
expression (Kourakis, 1997).
Homeotic genes are known to be involved in patterning
morphological structures along the antero-posterior axis of
insects and vertebrates. Because of their important roles in
development, changes in the function and expression
patterns of homeotic genes may have played a major role
in the evolution of different body plans. For example, it has
been proposed that during the evolution of several
crustacean lineages, changes in the expression patterns of
the homeotic genes Ultrabithorax and abdominal-A have
played a role in transformation of the anterior thoracic
appendages into mouthparts termed maxillipeds.
In particular, the
absence of Ubx and Abd-A from anterior segments of certain
derived malacostracan crustaceans correlates with the
appearance of maxillipeds on those segments in adults
This
homeotic-like transformation is recapitulated at the late
stages of the direct embryonic development of the
crustacean Porcellio scaber [the common woodlouse (Oniscidea, Isopoda)].
The
development of Porcellio scaber is direct (epimorphic) and
results in the hatching of a larva that possesses
all the morphological features of the adult
isopod except that it lacks a seventh pair of
thoracic (pereonic) appendages. The archetypal adult
malacostracan has three tagma: the mandibulate
head of six segments; the 'thorax' (pereon) with
eight pairs of uniramous legs, and the 'abdomen'
(pleon) with five pairs of appendages used for
locomotion and/or breathing and one pair of
terminal appendages called uropods. In the lineage leading to the isopods, the
first pair of thoracic limbs have apparently been
transformed into maxillipeds.
In adult isopods these appendages often have
flat proximal articles (basipodites) that
posteriorly and ventrally enclose the mandibles
and the two pairs of maxillae and serve a
function analogous to that of the insect labium (Abzhanov, 1999).
This morphological change of thoracic limbs into maxillipeds is associated with
apparent novelties both in the transcriptional and post-transcriptional regulation of the Porcellio scaber ortholog
of the Drosophila homeotic gene, Sex combs reduced (Scr), considered a 'head' homeotic gene in insects.
Specifically, SCR mRNA is present in the second
maxillary segment and the first pair of thoracic legs (T1)
in early embryos, whereas protein accumulates only in the
second maxillae. However, in later stages, high levels of
SCR appear in the T1 legs, which correlates temporally
with the transformation of these appendages into
maxillipeds. It is possible that T1/Mxp expression of
Scr is related to the retraction of Ubx and abd-A from that segment and that Scr transcription is normally negatively
regulated by Ubx and/or abd-A in the rest of the thorax. This transformation is reminiscent of homeotic
mutations in Drosophila associated with the ectopic expression
of the anterior genes in segments posterior to their normal
expression domains. If correct, this
recruitment of thoracic appendages to mouthparts provides the
first example of a true homeotic-like morphological
transformation of this kind occurring during the evolution of
animals (Abzhanov, 1999).
Continued: Sex combs reduced Evolutionary homologs part 2/2
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Sex combs reduced:
Biological Overview
| Regulation
| Targets of Activity, Homeotic Effects, Post-Transcriptional Regulation and Protein Interactions
| Developmental Biology
| Effects of Mutation
| References
Society for Developmental Biology's Web server.
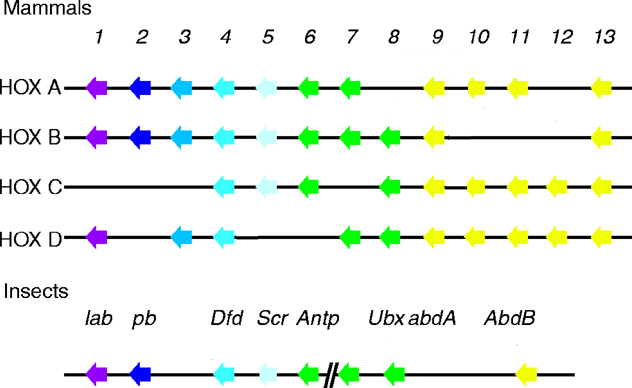{kind=link}