Antennapedia
The homeobox-complex (HOX-C) programs much of the basic body plan and forms, in most metazoans, one continuous cluster with the order of genes paralleling their order of expression along the body axis. The HOX-C organization of several Drosophila species and of Anopheles gambiae, a representative of lower Dipterans, have been compared. A model was developed to account for the evolution of the HOX-C in the Diptera (Lewis, 2003).
In D. melanogaster, the HOX-C is split into the Antennapedia complex (ANT-C) and the bithorax complex (BX-C). In D. virilis, a split is found between Ubx and abd-A, which corresponds to a separation of approximately 3-4 Mb, based on estimates in D. melanogaster (Lewis, 2003).
Recently, a third species, D. pseudoobscura, has been sequenced and aligned to D. melanogaster. In D. pseudoobscura, the split occurs between Antp and Ubx. Thus, D. pseudoobscura is more similar to D. melanogaster than to D. virilis, which is consistent with the phylogeny of these species. In contrast, D. repleta, like D. virilis, carries the split between Ubx and abd-A, which is consistent with the two species being closely related. This breakdown of the contiguity of the HOX-C in four Drosophila species is presumably a relatively recent event, since in A. gambiae the HOX-C has remained intact (Lewis, 2003).
Sequencing of the Antp and Ubx genes and the intervening region in D. virilis, comprising 308,092 base pairs (bp) (AY333070) and including the Antp gene, confirms that there is no split between Antp and Ubx. However, between Antp and Ubx a gene, CG31217, is inserted immediately downstream of Ubx. In D. melanogaster, CG31217 is located adjacent to Ubx and immediately downstream of CG31217 is the breakpoint of the split (Lewis, 2003).
The length of stretch from the end of the last exon of Ubx to the beginning of the first exon of Antp is 275 kb in A. gambiae and 190 kb in D. virilis. The length of the homologous region in D. melanogaster is 143 kb, excluding the 9.6 Mb represented by the gap. CG31217 is a newly defined gene that was formerly considered to be two separate genes. CG10013 is either absent from A. gambiae or poorly conserved compared to the Drosophila genera (Lewis, 2003).
A. gambiae lacks any obvious protein-coding genes in the interval between Antp and Ubx and the homolog of CG31217 is located distant from the HOX-C on the same chromosome arm, in a 16.2 Mb sequence scaffold (AAAB01008987) (Lewis, 2003).
These results suggest a model for the evolution of the HOX-C of the higher Diptera. At the base of the Dipterans, an ancestor is assumed with the primordial HOX-C, from which two evolutionary branches can be inferred. One branch led to A. gambiae, and the other, after insertion of CG31217, to a postulated ' vir-mel' chromosome, which subsequently led to the chromosome organization in D. virilis and to D. melanogaster (Lewis, 2003).
A hypothetical ancestor of the Diptera contains a colinear HOX-C. The hypothetical chromosome ' vir-mel' has arisen by transposition (Tp) of the gene CG31217 between the Antp and Ubx genes. To account for additional genes present in the Drosophila lineage, ' vir-mel' is postulated to have contained the Sodh gene closely linked to the Cctg gene. The ' pre-vir' chromosome aroses by a postulated inversion, Inv(pre-vir). D. virilis differs from pre-vir in having a Tp of the gene CG10013 between Antp and Ubx. In the D. melanogaster lineage, a precursor chromosome, ' pre-mel' is assumed to have arisen by an inversion, Inv(pre-mel), with breakpoints proximal to CG31217 and between the Sodh and Cctg genes of ' vir-mel' . For the modern D. melanogaster gene arrangement, a second inversion Inv(mel) is postulated that included the BX-C and extended just beyond Cctg (Lewis, 2003).
In the D. virilis lineage, an inversion, Inv(pre-vir), gave rise to a ' pre-vir' chromosome, and insertion of a second gene, CG10013, led to the present configuration in D. virilis. In D. melanogaster, CG10013 is remote from both Antp and Ubx (Lewis, 2003).
In the lineage leading to D. melanogaster, the split between Ubx and Antp is postulated to be an inversion, Inv(pre-mel) that upon an additional inversion Inv(mel) could give rise to the standard gene order. These two events placed Ubx and CG31217 next to a chaperonin-containing gamma gene (Cctg), and Antp next to a sorbitol dehydrogenase I (Sodh) gene (AE001572) (Lewis, 2003).
The simplest assumption is that Sodh and Cctg were closely linked in ' vir-mel' and that the inversion had one breakpoint between them. However, in A. gambiae these well conserved genes are located distant from each other and from the HOX-C (Lewis, 2003).
Interestingly, Inversion(3R)Payne, which is widespread among wild D. melanogaster populations, shows a gene order similar to ' pre-mel' . Moreover, the closely related D. simulans contains the only other known chromosome that is similar to ' pre-mel' . The major cytological difference between D. simulans and D. melanogaster is a large inversion with breakpoints in sections 84B and 92C. The 84B breakpoint is adjacent to, or identical with, the one that separated Antp and Ubx, whereas the 92C breakpoint is distal to the BX-C (Lewis, 2003).
Several hypotheses have been invoked to explain the persistence of the HOX-C as a single cluster. A common view holds that the cis-regulatory regions between adjacent HOX genes are bifunctional. In this scenario, splitting can occur only if preceded or accompanied by a duplication of the enhancer (Lewis, 2003).
The persistence of an intact and colinear HOX-C over hundreds of millions of years suggests that its organization is advantageous. Thus, any rearrangement that splits the HOX-C will generally become fixed only if it confers a selective advantage. Such a rearrangement is evidently a rare event. Possibly it has occurred in Drosophila for a combination of reasons: the extreme fecundity and short life cycle of Drosophila; the fact that paracentric inversions do not reduce fertility when heterozygous (which is due to the elimination of dicentric chromatids that arise from single crossovers within the inverted regions); the rarity of crossing over in the Drosophila male, and, finally, the possibility of a high frequency of transposon-mediated inversions, as has been found for D. buzzatii (Lewis, 2003).
Two nested Antp transcription units extend over 103 kb and produce sixteen different transcripts.
The Antp genes of D. virilis, D. subobscura and D.
melanogaster were compared to determine which structural features are conserved and therefore may be important
to the gene's function. The overall gene structures are similar. There are many conserved sequence
blocks throughout the large introns at least 15 kb upstream of the first promoter, and at least 3 kb
downstream of the last polyadenylation site. Intron and exon sequence conservation around
alternative splice sites indicates that alternative protein coding forms may also be conserved.
Protein coding potential is perfectly conserved around the C-terminal homeodomain, well
conserved in the N-terminal region, and more variable in the middle. The large size of the Antp
gene may reflect a large number of control elements necessary for appropriate ANTP protein
expression. The conservation of transcript complexity suggests functional requirements for the
different protein forms (Hooper, 1992).
During the embryogenesis of Drosophila, the homeotic genes are required to
specify proper cell fates along the anterior-posterior axis
of the embryo. Partial cDNAs of homologs
of the Drosophila homeotic gene teashirt and five of the
homeotic-complex (HOM-C) genes were cloned from the thysanuran
insect, Thermobia domestica (the firebrat), and these genes were assayed for their embryonic
expression patterns. The HOM-C genes examined
were labial, Antennapedia, Ultrabithorax, abdominal-A and Abdominal-B. Since the expression pattern of these HOM-C genes is largely conserved among insects
and since Thermobia is a member of a phylogenetically basal
order of insects, the ancestral expression patterns of these genes in insects could be inferred. The expression patterns of the Thermobia HOM-C genes were compared with their
expression in Drosophila and other insects;
the potential roles these genes may have played in insect
evolution are discussed. Interestingly, the teashirt homolog shows
greater variability between Thermobia and Drosophila
than any of the HOM-C genes. In particular, teashirt is
not expressed strongly in the Thermobia abdomen, unlike
Drosophila teashirt. It is proposed that teashirt expression
has expanded posteriorly in Drosophila and contributed to a homogenization of the Drosophila larval thorax and abdomen (Peterson, 1999).
Firebrat and grasshopper Antp are expressed in a very similar embryonic pattern
and they are essentially equivalent, with the exception
that early Td-Antp (Thermobia domestica/firebrat Antp) shows 'striping', while early Schistocerca
expression is uniform throughout the thorax. It is
concluded that the ancestral pattern of Antp is similar to
the firebrat/grasshopper Antp pattern and includes the
entire thorax, with little modulation during development.
There are three important
distinctions between the non-drosophilid insects
(grasshoppers and firebrats) and Drosophila epidermal
Antp expression: (1) early expression in the non-drosophilid
insects is in a segmental register in all three thoracic
segments (apparently compartmental in the youngest
firebrats), while in Drosophila initial expression appears
in PS4 and 6; (2) as elongation proceeds, a
dorsal-ventral shift in Antp expression arises in the mandibulate
insects, but Drosophila Antp remains distinctly parasegmental
in both lateral and ventral domains; (3) there
is no modulation of expression levels in the mandibulate
insects, other than a uniformly higher level in the thorax
versus the abdomen. In Drosophila, however, dynamic
modulations occur within each of its four main parasegmental
domains, shifting in relative levels all through
germ band extension and into germ band shortening.
The first two of these differences reflect the homogenization
of the Drosophila thorax and abdomen. Td-Antp
expression is more similar to head homeotic genes like
Td-cap'n'collar, Td-Deformed and Td-Scr, which have
dorsal-ventral shifts, than it is to Td-Ubx, Td-abd-A and
Td-Abd-B, which have parasegmental anterior borders.
Just the opposite is true for Drosophila Antp, which shows parasegmental
registers like the abdominal genes. Likewise,
in both Drosophila and Tribolium, mutations in anterior
homeotic genes typically give segmental transformations,
while Ubx, abd-A and Abd-B (Ultrathorax, Abdominal
and extra urogomphi in Tribolium) tend to give
parasegmental transformations. Antp shows a difference: in Drosophila the homeosis appears to be parasegmental, while prothoraxless
(Tc-Antp) in Tribolium causes homeotic transformations
in a segmental register. Thus, in the non-drosophilid insects,
Antp has an expression domain typical of gnathal
homeotic genes, while that of Drosophila Antp is more like an
abdominal gene. This similarity in HOM expression register
throughout the trunk may be necessary for the consistent
patterning of denticle belts and hairs (Peterson, 1999).
The third difference (modulation of levels in each
parasegment) may reflect a greater distinction between
thoracic segments in Drosophila that is not seen in firebrats
and a reduction of the role of Antp in thoracic development.
Each ventral denticle belt and set of dorsal
hairs is different among the Drosophila larval thoracic
segments. The
presence of Scr and, in some cases, the absence of Antp
expression is important for the formation of the T1 denticles. Antp controls cell fate in T2 (PS4), and Ubx and Antp are both required to produce T3 denticles. A thorough analysis
of the cuticular structure of the firebrat thorax would be
required to determine whether the Drosophila thoracic
segments are really more distinct from each other than
they are in firebrats or whether the differences are simply
more apparent in Drosophila. However, this analysis is
consistent with the lack of Scr expression in most of T1
in firebrats, milkweed bugs and crickets and the lack of transformation of PS5 (pT2-aT3) in Tribolium Ultrathorax mutants. A
reduced role for Antp in defining the Drosophila thorax
is also consistent with the somewhat modest transformation
seen in Drosophila embryos, in comparison to the
dramatic transformations seen in Tribolium (Peterson, 1999).
Hox genes are known to control the identity of
serially repeated structures in arthropods and vertebrates.
The expression pattern of the Hox genes Deformed, Sex combs reduced, Antennapedia, and Ultrabithorax/abdominal-A (Ubx/abd-A)
from the honey bee Apis mellifera were analyzed. A
cDNA with the complete coding region of the Antennapedia
gene from Apis was cloned. Comparison with Antp proteins
from other insect species revealed several regions of homology.
The expression patterns of the isolated Hox
genes from Apis show that the original expression patterns
of Dfd, Scr, and Antp appear between late blastoderm
and early germ band stage in a temporal and spatial
sequence. Each of them shows up as a belt, spanning approximately
two segment anlagen; Dfd in the anterior
gnathal region, Scr in the posterior gnathal and anterior
thoracic region, and Antp in the thoracic region. Following
expansion of the Antp domain in the abdomen as a
gradient towards the posterior, Ubx/abd-A expression appears
laterally in the abdomen. During gastrulation and
in the germ band stage the domains of strong expression
do not overlap any more, but touch each other. After gastrulation
the borders of the expression domains partly
correlate with parasegment and partly with segment
boundaries. Laterally, gaps between the domain of each
gene may show no expression of any of the genes examined (Walldorf, 2000).
In the blastoderm stage the expression domains of all
four genes are similar in fruit fly and honey bee embryos.
Certainly these genes play very similar roles in establishing
the general character of that part of the insect
body where they have their main expression domain.
Even in Chelicerata the orthologs show comparable
patterns. However, some differences should be mentioned. In the
fly, the borders of the Dfd domain are segmental. In
the bee, the anterior expression border is initially parasegmental
in the rudiment of the intercalary segment: this changes from segmental to parasegmental during
gastrulation in the ventral part of the maxillary segment. The steady Engrailed pattern shows that this change is not a result of cell movement. The same development of that pattern is reported for Tribolium. This is most
clearly seen ventrally in the parasegmental Scr pattern in
the germ band of the bee, corresponding to segmental in
that area in the fly. While some differences in the Dfd pattern are
consistently observed between the bee and the fly, the
complicated posterior borders of the Scr and the Antp
patterns of young and old embryos of flies and bees are identical. The
same Antp pattern is also found in Thermobia domestica. However, in Manduca sexta the
anterior border of Antp is the compartment border of T1. It may be that some of these differences are only the result of different sensitivity levels of
the stainings. Probably the ventral switch from segmental
to parasegmental is more general in insects but is not
seen very clearly in some cases, such as in the anterior
part of the fruit fly head, due to the subsequent head involution.
The resulting ventrolateral corners of the expression
borders of Dfd, Scr, and Antp correlate fairly
well with the position of the appendage anlagen and may
help to pin down their position (Walldorf, 2000).
Hox genes are well-known master regulators in developmental morphogenesis along the anteroposterior axis of animals. However, the molecular mechanisms by which Hox proteins regulate their target genes and determine cell fates are not fully understood. The silk gland of Bombyx mori is a tubular tissue divided into several subparts along the anteroposterior axis, and the silk genes are expressed with specific patterns. The sericin-1 gene (ser1) is expressed in the middle silk gland (MSG) with sublocal specificity. This study shows that the Hox protein Antp is a component of the middle silk gland-specific complex, MIC (MSG-intermolt-specific complex), binds to the essential promoter element of ser1, and activates its expression. Ectopic expression of Antp in transgenic silkworms induced the expression of ser1 in the posterior silk gland (PSG), but not in the anterior part of MSG (MSG-A). Correspondingly, a MIC-like complex was formed by the addition of recombinant Antp in extracts from PSG with its cofactors Exd and Hth, but not in extracts from MSG-A. Splicing patterns of ser1 mRNA induced by the ectopic expression of Antp in PSG were almost the same as those in MSG at the fifth instar and altered depending on the induction timing of Antp. Other Hox genes were expressed with sublocal specificity in the silk gland. The Bombyx silk gland might provide a useful system for understanding how Hox proteins select and regulate their target genes (Kimoto, 2014).
Evolutional changes in homeotic gene functions have contributed to segmental diversification of arthropodan limbs, but crucial molecular changes have not been identified to date. The first leg of the crustacean Daphnia lacks a prominent ventral branch found in the second to fourth legs. This phenotype correlates with the loss of Distal-less and concomitant expression of Antennapedia in the limb primordium. Unlike its Drosophila counterpart, Daphnia Antennapedia represses Distal-less in Drosophila assays, and the protein region conferring this activity was mapped to the N terminal region of the protein. The results imply that Dapnia Antennapedia specifies leg morphology by repressing Distal-less, and this activity was acquired through a change in protein structure after separation of crustaceans and insects (Shiga, 2002).
The highly restricted expression of DapANTP in L1 of Daphnia suggests a model in which ANTP modifies the morphology of the T1 leg to a smaller one by repressing DLL. This idea would explain the observation that crustacean legs anterior to the domain of UBX/ABDA are, in general, small and resemble feeding appendages when compared with more posterior limbs specialized for locomotive functions. Given the strong limb suppressing activity of DapANTP observed in the Drosophila assays, expression of ANTP seems to be tightly regulated in Daphnia, and the post-transcriptional regulation of Antp expression observed in this study is one mechanism assuring limited expression of ANTP. Modification of T5 limbs may be due to activities of the posterior Hox gene AbdB that has been shown to repress limb development in Drosophila (Shiga, 2002).
Diversification of the ANTP protein outside the homeodomain contributes to its functional variation in modifying limb patterns. The region responsible for Daphnia-specific activity maps to the N terminal region of ANTP that is highly diverged. Two recently works on Ubx proteins have reported that functional alteration of homeotic proteins play a significant role in restricting the number of insect limbs. This work demonstrates that an evolutional change in Antennapedia protein has contributed to a
micro-evolutionary event that has produced the difference in the shape of T1 leg and T2-4 legs of Daphnia. Taken together, homeotic proteins have undergone a number of alterations in regions outside the homeodomain to change their target specificity and the way they control limb development. More importantly, Daphnia-specific ANTP activity and the pattern of its expression account for segment-specific limb morphology of Daphnia, suggesting that protein-coding regions of Hox genes serve as rich substrates for evolutional alterations that have generated segmental diversities of the crustacean limb (Shiga, 2002).
Representatives of the Insecta and the Malacostraca
(higher crustaceans) have highly derived body plans
subdivided into several tagma (groups of segments united
by or fused into a common function and/or morphology). The
tagmatization of segments in the trunk, the part of the body
between head and telson, in both lineages is thought to have
evolved independently from ancestors with a distinct head
but a homonomous, undifferentiated trunk. In the
branchiopod crustacean, Artemia franciscana, the trunk
Hox genes are expressed in broad overlapping domains
suggesting a conserved ancestral state. In comparison, in
insects, the Antennapedia-class genes of the homeotic
clusters are more regionally deployed into distinct domains
where they serve to control the morphology of the different
trunk segments.
In Drosophila Antp is expressed in
and required for the specification of the three-segmented
locomotory thorax. Both Ubx and abd-A are involved in the
development of the legless abdomen. Ubx is also expressed in the posterior thorax where it is known to be involved in the development of the modified hind
wings, the halteres. Thus an originally Artemia-like pattern of
homeotic gene expression has apparently been modified in
the insect lineage associated with and perhaps facilitating
the observed pattern of tagmatization. Since insects are the
only arthropods with a derived trunk tagmosis tested to
date, the expression patterns of the Hox genes
Antp, Ubx and abd-A were examined in the malacostracan crustacean
Porcellio scaber (Oniscidae, Isopoda). Unlike
the pattern seen in Artemia, these genes are expressed in
well-defined discrete domains coinciding with tagmatic
boundaries that are distinct from those of the insects. These
observations suggest that, during the independent
tagmatization in insects and malacostracan crustaceans,
the homologous 'trunk' genes evolved to perform different
developmental functions. It is also proposed that, in each
lineage, the changes in Hox gene expression pattern may
have been important in trunk tagmatization (Abzhanov, 2000).
Contemporary molecular and morphological phylogenies of
the Crustacea indicate that this group comprises a
monophyletic assembly with some classes such as the
Remipidia and Branchiopoda at a basal position and the
Malacostraca as a crown group. Additionally, according to recent phylogenies, the
Crustacea are placed as the sister group to the Insecta in the
subphylum Mandibulata. Alternatively, some studies suggest
that crustaceans may be paraphyletic with regard to the Insecta
with the Malacostraca as the closest sister group to insects. The
Mandibulata also includes the more distantly related
Myriapoda. The Chelicerates are generally regarded as a sister
group to the Mandibulata (Abzhanov, 2000 and references therein).
In the insects, epidermal Antp expression is confined
primarily to the thorax although minor but detectable
accumulation is also observed in the abdomen. This expression domain is highly
conserved in all studied members of the Insecta. For example, in the apterygote insect Thermobia domestica, Antp is detected in the three thoracic segments and
at lower levels in the abdominal segments, except the most
terminal segment bearing cerci. It has
been postulated that the strong uniform expression of Antp in
the thorax with clear segmental boundaries and weak
expression in the abdomen represents a pattern basal common to all
insects. The anterior boundary of insect
Antp resides in the posterior labial segment ventrally and in the
anterior prothoracic segment and appendages laterally.
In the branchiopod crustacean, A. franciscana, Antp is
expressed throughout the trunk in the eleven-segmented,
uniform thorax-like tagma anterior to the genital segments. The anterior boundary extends into the gnathal region and is observed in posterior mx1. Antp expression is not uniform in the trunk
but is restricted chiefly to the legs (Abzhanov, 2000 and references therein).
Since branchiopods are believed to be phylogenetically basal with
respect to malacostracan crustaceans, it is concluded that the
discrete pereonic/thoracic expression domain of Antp reported here for
P. scaber is derived. However, it must be emphasized that the
PsAntp domain is different from that seen in insects and
Artemia in at least three ways: (1) the anterior boundary is roughly one segment
more anterior than what is observed in Artemia and, in this
respect, appears to be similar to insect Antp; (2)
the posterior boundary lies in the seventh trunk
segment rather than in the third trunk segment as in insects or
in the last, eleventh trunk segment as in Artemia; (3)
unlike in insects, PsAntp expression is highly
compartmentalized within the segments suggesting that this
gene may be playing a unique role in the specification of the
pereon in spite of a virtually complete overlap with the Ubx
domain. Importantly, however, both in insects and crustaceans,
the anterior and posterior boundaries correlate well with the
morphological tagmatic borders (Abzhanov, 2000).
In the freshwater planarian Dugesia japonica, five cDNAs for HOM/HOX homeobox genes, (Plox or planarian HOM/HOX homeobox genes) were cloned and sequenced. Together with sequence data on platyhelminthes
HOM/HOX homeobox genes that have been deposited in databases, comparison of the deduced amino acid sequences reveals that planarians have at least
seven HOM/HOX homeobox genes, Plox1 to Plox7. Plox1, Plox3 and Plox7 show high similarity to the fly genes labial (82% amino acid identity in the homeodomain), Dfd (82%) and abd-a (88%), respectively. Plox4 may correspond to Sex combs reduced (Scr) rather than Dfd (75%), because of Plox4's high similarity with nematode mab5 (67%), leech Lox5-hro (77%), and paralog group 4 in vertebrates; planarian chich corresponds to Scr in the fly. Plox2 could not be classified. Both Plox5 and Plox6 resemble Antp in the fly. Whole-mount in situ hybridization and RT-PCR reveal that Plox4 (72% homologous to Sex combs reduced)
and Plox5 (77% homologous to Scr) are increasingly expressed along a spatial gradient in the posterior region of intact animals. During regeneration, Plox5 is expressed only in the
posterior region of regenerating body pieces, suggesting that the gene is involved in the anteroposterior patterning in planarians. Plox5 is not found to be expressed
in a blastema-specific manner. X-ray irradiation experiments show that at a minimum, Plox5 is expressed in some cells other than neoblasts, but that the induction of Plox5 expression during
regeneration might require neoblasts. Chromosome walking data are needed to understand the genomic organization and relationships of planarian HOM/HOX homeobox genes to those of other animal groups (Orii, 1999).
Molecular developmental studies of fly and mouse embryos have shown that the identity of individual
body segments is controlled by a suite of homeobox-containing genes called the Hox cluster. To
examine the conservation of this patterning mechanism in other segmented phyla,
four Hox gene homologs isolated from glossiphoniid leeches of the genus Helobdella are described. Based on
sequence similarity and phylogenetic analysis, the leech genes Lox7, Lox6, Lox20, and Lox5 are
deemed to be orthologs of the Drosophila genes lab, Dfd, Scr, and Antp, respectively. Sequence
similarities between Lox5 and Antp outside the homeodomain and phylogenetic reconstructions suggest
that the Antennapedia family of Hox genes had already expanded to
include at least two discrete Antp and Ubx/abdA precursors prior to the annelid/arthropod divergence.
In situ hybridization reveals that the four Lox genes described in this study are all expressed at high
levels within the segmented portion of the central nervous system (CNS), with variable levels of
expression in the segmental mesoderm. Little or no expression is seen in peripheral ectoderm or
endoderm, or in the unsegmented head region (prostomium). Each Lox gene has a distinct anterior
expression boundary within one of the four rostral segments, and the anterior-posterior (AP) order of
these expression boundaries is identical to that reported for the orthologous Hox gene products in fly
and mouse. This finding supports the idea that the process of AP axis differentiation is conserved
among the higher metazoan phyla with respect to the regional expression of individual Hox genes along
that axis. One unusual feature of leech Hox genes is the observation that some genes are only
expressed during later development -- beginning at the time of terminal cell differentiation -- whereas
others begin expression at a much earlier stage; their RNA ceases to be detectable shortly after
the onset of expression of the 'late' Hox genes. The functional significance of this temporal disparity is
unknown, but it is noteworthy that only the two 'early' Hox genes display high levels of mesodermal expression (Kourakis, 1997).
The leech homeobox gene Lox1 contains a homeodomain homologous to those of Drosophila's Sex combs
reduced and Antennapedia, and to homeodomains of the vertebrate Hox groups 5
and 6. Lox1 expression was detected during late gangliogenesis in 15-20 pairs of central neurons
repeated in most segments, suggesting that Lox1 may play a role in the differentiation of these cells.
Two pairs of Lox1-expressing neurons have been identified as the Bipolar
cells and the L1 neurons. The L1 neurons were detected late in gangliogenesis and became stable
neurons. The Bipolar cells appeared very early in gangliogenesis, extended processes in the
longitudinal connective nerves, and then degenerated, suggesting that they play a role in establishing
the longitudinal tracts of the central nerve cord (Aisemberg, 1994).
Lox2 in the leech is
expressed in an iterated set of neurons in the posterior two-thirds of the CNS. On the basis of cell
body position and relative size, two pairs of Lox2-expressing cells were identified as the RPE-like
neurons and the CV motor neurons. Other Lox genes are also expressed in segmentally repeated
subpopulations of neurons. These neuronal subpopulations appear to be different from one another
but partially overlapping. Different combinations of Lox genes that may be expressed in individual
cells could, in theory, generate enough variability to specify all central neurons in a leech ganglion (Aisemberg, 1993).
The nematode C. elegans stands out as having a greatly reduced Hox gene complement. To address this, orthologs of C. elegans Hox genes were identified in six species from across the Nematoda, and they show that rapid homeodomain sequence evolution is a general feature of nematode Hox genes. Some nematodes express additional Hox genes belonging to orthology groups that are absent from C. elegans but present in other bilaterian animals. A central antennapaedia-like gene (ant-1) was found in three species: B. malayi, A. suum, and T. spiralis. These genes were not recovered by using identical procedures from P. pacificus. Hox cluster reduction in nematodes is therefore not due to a single event, but has been occurring through the evolution of the phylum. Loss of hox-3 and ant-1 can be inferred to have taken place since C. elegans last shared a common ancestor with nematodes in Clades IV and III, respectively. The remaining Hox gene losses either occurred before the radiation of the nematode crown group, or they occurred within the nematode lineage. Analysis of the genomic environment of a newly identified Brugia malayi Hox6-8 ortholog (Bm-ant-1) reveals that it lay downstream of the Bm-egl-5 Hox gene and that their homeodomain exons are alternately cis spliced to the same 5' exon. This organization may represent an intermediate state in Hox gene loss via redundancy. The two Brugia malayi genes have overlapping but distinct temporal expression patterns, with that of Bm-ant-1 contained entirely within that of Bm-egl-5. The Hox clusters of nematodes are the product of a dynamic mix of gene loss and rapid sequence evolution, with the most derived state observed in the model C. elegans (Aboobaker, 2003).
In C. elegans, the
Antennapedia-like HOM-C gene mab-5 not only specifies postembryonic fates of cells in a posterior
body region, but also influences the migration of mesodermal and neural cells that move through this
region. As one neuroblast migrates into this posterior region, it switches on mab-5
gene expression; mab-5 then acts as a developmental switch to control the migratory behaviour of
the neuroblast descendants. HOM-C genes can therefore not only direct region-specific patterns of
cell division and differentiation, but can also act within migrating cells to program region-specific
migratory behaviour (Salser, 1992).
Two homeotic cluster (HOM-C) genes of C. elegans have antagonistic roles in the ability of vulval precursor cells (VPCs) to respond to the inductive signal from the anchor cell of the somatic gonad. The vulva develops from a subset of ectodermal vulval precursor cells distributed along the anteroposterior axis. Vulval patterning depends on both a localized inductive signal by the epidermal growth factor homolog LIN-3 (see Drosophila Vein for more information) and lateral signaling between inducing VPCs. One HOM-C gene, the Antennapedia homolog mab-5, is expressed in the posterior two VPCs. By examining the response of single VPCs to controlled doses of inductive signal in wild-type and in mab-5 mutant animals, it can be seen that mab-5 mutation increases the responsiveness of these two cells. Moreover, a gain-of-function allele of mab-5 that causes ectopic expression of MAB-5 in all VPCs reduces the sensitivity of all VPCs to inductive signal. Additional experiments suggest that another HOM-C gene, the Sex combs reduced homolog lin-39, is required for VPCs in wild-type animals to respond to activation of inductive signal. Genetic epistasis tests are consistent with models in which lin-39 acts downstream of the RAS pathway to regulate response to inductive signal. It is proposed that the spatial pattern of HOM-C gene expression may enhance the precision of vulval fate patterning (Clandinin, 1997).
ceh-7 is a small Caenorhabditis elegans homeobox gene consisting of 84 amino acids that at present is not known to be closely related to any others. Examination of the
expression pattern of ceh-7 using reporter constructs reveals that is expressed in a few cells of the male tail, which form a
ring around the rectum. The most posterior member of the C. elegans Hox cluster, egl-5, an Abd-B homolog, is required for the proper development of several blast cells in the male tail. The expression
of ceh-7 has been examined in mutant backgrounds of egl-5 and also mab-5, an Antp/Ubx/Abd-A homolog. Although ceh-7 is not
expressed in egl-5 mutants, it is still expressed in mab-5 mutants. The late expression argues that ceh-7 is not involved in sex determination itself; rather, its role appears to be in the generation or differentiation of cell types specific to the male tail. Given the conserved nature of the Hox cluster, it appears possible that in vertebrates and flies, homeobox genes, perhaps even a ceh-7 homolog, might exist which control the development of sex-specific appendages (Kagoshima, 1999).
The specification of body pattern along the anteroposterior (A/P) body axis is achieved largely by the actions of
conserved clusters of Hox genes. Precise control of the hox gene mab-5 expression is crucial for achieving wild-type development of at least two cell types, the V cells and the migratory Q neuroblasts. In the V5 lineage, mab-5 is switched on and off multiple times: each change in expression regulates a different type of cell-fate decision. The two Q cells are neuroblasts born in identical A/P postions but on opposite sides of the animal: QL on the left and QR on the right. In wild-type animals, after a short poserior migration, QL switches on mab-5, which, in turn, causes descendants of QL to migrate to the posterior. In contrast, mab-5 remains off in QR and its descendants, and as a result the descendants of QR continue to migrate toward the anterior. Limiting expression of Hox genes to localized regional domains and controlling the
precise patterns of expression within those domains is critically important for normal patterning.
egl-20, a C. elegans gene required to activate expression of the Hox gene mab-5 in the migratory neuroblast QL,
encodes a member of the Wnt family of secreted glycoproteins. A second Wnt pathway gene,
bar-1, which encodes a beta-catenin/Armadillo-like protein, is also required for activation of mab-5 expression in
QL. In addition, the gene pry-1, phenotypically resembling Drosophila Polycomb-group mutants, is required to limit expression of the Hox genes lin-39, mab-5 and
egl-5 to their correct local domains. egl-20, pry-1 and bar-1 all function in a linear genetic pathway with
conserved Wnt signaling components, suggesting that a conserved Wnt pathway activates expression of mab-5 in the
migratory neuroblast QL. Members of this Wnt signaling system are found to play a major role in both the
general and fine-scale control of Hox gene expression in other cell types along the A/P axis. Although a similar global role for Wingless in regulating Hox gene expression in Drosophila has not been found, in at least one instance, the expression of Ultrabithorax in the midgut, a Drosophila Hox gene acts downstream of a Wnt gene (Maloof, 1999).
Hox genes pattern the fates of the ventral ectodermal Pn.p
cells that lie along the anteroposterior (A/P) body axis of C.
elegans. In these cells, the Hox genes are expressed in
sequential overlapping domains where they control the
ability of each Pn.p cell to fuse with the surrounding
syncytial epidermis. The activities of Hox proteins are sex-specific
in this tissue, resulting in sex-specific patterns of
cell fusion: in hermaphrodites, the mid-body cells remain
unfused, whereas in males, alternating domains of syncytial
and unfused cells develop. The wild-type Pn.p cell fusion pattern
arises through the regulation of both Hox gene expression
and Hox protein activity. In both wild-type males and
hermaphrodites, lin-39 (Scr homolog) is expressed in P(3-8).p in the mid-body
and mab-5 (Antp homolog) is expressed in an overlapping domain that spans P(7-11).p in the posterior. At the beginning of the second larval stage,
only P(3-8).p in the mid-body remains unfused in wild-type
hermaphrodites. The
posterior Pn.p cells adopt the syncytial fate despite mab-5
expression because the activity of MAB-5 protein is inhibited in
hermaphrodite Pn.p cells (Chng, 1999 and references).
The gene egl-27,
which encodes a C. elegans homolog of a chromatin
regulatory factor, specifies these patterns by regulating
both Hox gene expression and Hox protein function.
Mutations in the C. elegans gene egl-27 were first isolated in
a screen for egg-laying defective mutants.
More recently, egl-27 has been found to encode nuclear protein
isoforms similar to a protein encoded by mta1, whose expression is elevated in
metastatic tumor cell lines and functions as a component of
NURD, a human chromatin regulatory complex. egl-27 appears to be widely expressed
and is involved in the control of cell polarity, cell migration
and embryonic morphogenesis (Chng, 1999 and references).
In egl-27
mutants, the expression domains of Hox genes in these
cells are shifted posteriorly, suggesting that egl-27
influences A/P positional information. In addition, egl-27
controls Hox protein function in the Pn.p cells in two ways:
in hermaphrodites, it inhibits MAB-5 activity, whereas in
males it permits a combinatorial interaction between LIN-39
and MAB-5. Thus, by selectively modifying the activities
of Hox proteins, egl-27 elaborates a simple Hox expression
pattern into complex patterns of cell fates. Taken together,
these results implicate egl-27 in the diversification of cell
fates along the A/P axis and suggest that chromatin
reorganization is necessary for controlling Hox gene
expression and Hox protein function (Chng, 1999).
egl-27 is required for both of the
post-translational forms of Hox regulation that occur in the
Pn.p cells. egl-27 is required for the inhibition of MAB-5
activity in hermaphrodite Pn.p cells. In C. elegans
hermaphrodites, the sex-determination pathway culminates in
the activation of tra-1, which acts cell-autonomously to specify
hermaphrodite-specific cell fates. Since tra-1 encodes zinc-finger
transcription factor isoforms, it has been postulated
that tra-1 might directly interfere with MAB-5s ability to bind
a promoter or activate transcription. Since there is no logical
necessity for additional component(s) outside the sex-determination
pathway for MAB-5 inhibition, it is
surprising that this process also requires egl-27. egl-27 is not
specifically involved in sex-determination because most of its
other phenotypes occur in both sexes. Since MAB-5 inhibition
involves both the sex-determination pathway and egl-27, tra-1
(or another sex-specific factor) might interact directly or
indirectly with egl-27 to inhibit MAB-5.
egl-27 is also required for the combinatorial interaction
between LIN-39 and MAB-5 in male Pn.p cells.
When multiple Hox genes function to specify a single body
part, it is difficult to distinguish whether they do so by acting
in distinct cells within that structure or by a combinatorial
interaction within the same cells. In C. elegans, it was possible to bypass this complication by examining the fate of single
Pn.p cells that are controlled cell-autonomously by mab-5 and
lin-39 (Chng, 1999).
How do LIN-39 and MAB-5 neutralize each other in this
combinatorial interaction? This post-translational process is not
regulated by nuclear access or protein degradation because it
occurs when both LIN-39 and MAB-5 proteins are present in
the nucleus of the same Pn.p cell. Moreover, this neutralization has been
shown to be insensitive to the relative levels of LIN-39 or MAB-5,
arguing against the formation of inactive LIN-39/MAB-5
heterodimers or competition between LIN-39 and MAB-5 for
target promoter(s) or co-factor(s). Instead,
this suggests that the two proteins might act together on another
target that is limiting, such as a promoter that could integrate
their activities.
EGL-27 (or other components of the NURD complex)
might interact directly or indirectly with MAB-5 and/or LIN-39
on a promoter of a cell fusion target gene in a manner
dependent on the activity of tra-1 (or another sex-specific
factor). Recruiting EGL-27 and the NURD complex to this
promoter could direct its chromatin reorganization activities to
this promoter and thus alter the transcriptional state of the target
gene, resulting in different Pn.p fates (Chng, 1999).
Members of the Hox family of homeoproteins and their
cofactors play a central role in pattern formation of all
germ layers. During postembryonic development of C.
elegans, non-gonadal mesoderm arises from a single
mesoblast cell M. Starting in the first larval stage, M
divides to produce 14 striated muscles, 16 non-striated
muscles, and two non-muscle cells (coelomocytes). The role of the C. elegans Hox cluster and of the exd ortholog ceh-20 in patterning of the postembryonic
mesoderm has been investigated. By examining the M lineage and its
differentiation products in different Hox mutant
combinations, an essential but overlapping role was found
for two of the Hox cluster genes, lin-39 (Scr homolog) and mab-5 (Antp homolog), in
diversification of the postembryonic mesoderm. This role
of the two Hox gene products requires the CEH-20
cofactor. One target of these two Hox genes is the C. elegans
twist ortholog hlh-8. Using both in vitro and in vivo assays,
it has been demonstrated that twist is a direct target of Hox
activation. Evidence from mutant phenotypes is presented
that twist is not the only target for Hox genes in the M
lineage: in particular lin-39 mab-5 double
mutants exhibit a more severe M lineage defect than the
hlh-8 null mutant (Liu, 2000).
The M lineage defects of mutants of Hox genes lin-39 and mab-5 combined with ceh-20 mutants are intriguing. These defects do not appear to
be a result of homeotic transformation of the fate of M or its
descendants. Instead, the mutants exhibit either (1) a loss of all
differentiated M-lineage descendants or (2) the precocious
production of abnormal mesodermal fates with certain
properties of later M lineage products. The precocious
appearance of large cells that exhibit SM- and sex muscle-like
characteristics suggests that this program might be a default
state of M in the absence of Hox function (Liu, 2000).
The shared role of MAB-5 and LIN-39 in the M lineage
appearsto be specific to these two Hox factors for the following reasons: (1) forced
expression of either lin-39 or mab-5, but not of the neighboring
Hox genes ceh-13 and egl-5, is sufficient to activate ectopic
expression of M lineage reporters; (2) egl-5 mutants
(which are viable either alone or in combination with lin-39
and mab-5 mutants) have no M lineage defects on their own and
show no synergistic effects with lin-39 and mab-5.
Mesodermal roles of Hox and exd genes have also been
shown in Drosophila. In the visceral mesoderm, Ubx and abd-A
are involved in morphogenesis of the midgut. In this case, a
few targets for Hox genes have been described: Ubx in the
visceral mesoderm is directly required with an Exd cofactor
for activating expression of the signaling molecule dpp. In the somatic
mesoderm, Ubx and abd-A can each promote the formation
of specific sets of muscle precursors. None of the Drosophila Hox mutants or
combinations that have been analyzed show as drastic an
effect on postembryonic mesoderm as was seen with the
lin-39 mab-5 double mutant in C. elegans. This apparent
discrepancy may reflect a fundamental difference between
the biological systems; alternatively, a more drastic
postembryonic requirement for Hox factors in the Drosophila
mesoderm might have been missed due to the embryonic
lethality of multiple-Hox mutants (Liu, 2000 and references therein).
The rescue experiments suggest partially overlapping roles
for mab-5 and lin-39 in the M lineage. The modest M-lineage
defects seen in mab-5 single mutants, compared with the lack
of any M-lineage defects in lin-39 single mutants suggest that
under normal circumstances the contribution of mab-5 may be
somewhat more substantial at early time points. One
conceivable explanation for the 'either/or' requirement would
involve cross-regulation between Hox genes. In particular, the possibility that lin-39 expression in the M lineage only occurs in the absence of functional mab-5 was tested. This is apparently not the case, since mab-5 mutants show an
apparently normal pattern of M lineage staining with antibodies to LIN-39 (Liu, 2000).
Several types of interactions between lin-39 and mab-5
activities in determining cell fate have been reported. In a
subset of Pn.aap cells that normally express both lin-39 and
mab-5, the lin-39 activity is dominant, preventing mab-5 from
functioning in these cells. A distinct interaction is seen in male Pn.p cells, where
lin-39 and mab-5 are both expressed and act combinatorially
to specify a fate that is different from that specified by either
alone. A third situation is seen in hermaphrodite vulval precursor cells, for which the loss of Hox (lin-39) activity after specification results in a failure to
differentiate; in this lineage, lin-39 and mab-5 activities have
the capability to promote distinct and non-overlapping
consequences in terms of cell fate. The functional and
simultaneous requirement in the M lineage for either mab-5 or
lin-39 function represents a further degree of freedom in using
these genes to build an organism (Liu, 2000).
The highly conserved structure of Hox factors is consistent
with a view that these genes have evolved by duplication of
a single precursor gene. Under these circumstances, it is not surprising that certain roles for Hox factors would still be maintained as shared (or redundant)
between several genes in the cluster. While the individual
genes might have acquired position-specific roles based on
their acquisition of intricate patterns of expression, it is
certainly conceivable that the entire family (or a large subset)
will have maintained a shared role equivalent to that of the
ancestral (and unique) Hox factor. While the role of that factor
will remain a mystery, the appearance of Hox factors in the
developing embryo just prior to the start of differentiation
suggests that the ancestral factor could have played a role in
developmental timing, perhaps modulating the start of
differentiation in a subset of cells (Liu, 2000).
The C. elegans twist ortholog hlh-8 is a direct and
critical target of Hox genes and ceh-20 in the
postembryonic M lineage. A critical site has been identified in the hlh-8
promoter that is a binding site for the LIN-39/CEH-20 protein
complex. The similarity between core binding sequences for
Drosophila Antp and Dfd proteins in vitro, and the functional equivalence of mab-5 and lin-39 in activating hlh-8 expression in the M lineage, strongly suggest
that this site is also a binding site for MAB-5/CEH-20.
Although hlh-8 is a target for Hox/CEH-20 function in the
M lineage, it is not the only such target. Several indirect
observations demonstrate the existence of additional targets.
One line of evidence comes from the observation that forced
expression of hlh-8 in lin-39 mab-5 mutants
fails to rescue the M lineage defects. An independent line of
evidence comes from a comparison of mutant phenotypes: lin-39 mab-5 mutants show a more severe patterning defect in the M lineage than null hlh-8
mutants: (1) while lin-39 mab-5 animals lack both M-derived coelomocytes, the
majority of hlh-8 mutants contain normal
numbers of M-derived coelomocytes; (2) while lin-39 mab-5 mutants lack all M-derived bodywall muscle, hlh-8 mutants produce variable number of these cells;
(3) sex muscles can be produced in hlh-8 mutants,
although they are not fully differentiated.
The identity of other Hox targets in the M lineage is not known (Liu, 2000).
Polycomb group (PcG) chromatin proteins regulate homeotic genes in both animals and plants. In Drosophila and vertebrates, PcG proteins form complexes and maintain early patterns of Hox gene repression, ensuring fidelity of developmental patterning. PcG proteins in C. elegans form a complex and mediate transcriptional silencing in the germline, but no role for the C. elegans PcG homologs in somatic Hox gene regulation has been demonstrated. Surprisingly, it is found that the PcG homologs MES-2 [E(Z)] and MES-6 (ESC), along with MES-3, a protein without known homologs, do repress Hox expression in C. elegans. mes mutations cause anteroposterior transformations and disrupt Hox-dependent neuroblast migration. Thus, as in Drosophila, vertebrates, and plants, C. elegans PcG proteins regulate key developmental patterning genes to establish positional identity (Ross, 2003).
The three mes genes act upstream of the Hox genes mab-5 and egl-5 during V ray differentiation, and loss of mes activity can restore normal ray development and mating ability to males mutant in the mab-5 activator pal-1. Males lacking mes activity display anterior expansions of tail structures and ectopic expression of the Hox reporter egl-5::gfp and the Hox target lin-32::gfp. This regulation is not restricted to the male tail: mes-2, -3, and -6 also repress lin-39::lacZ expression in the midbody and head and mab-5 activity in a migrating neuroblast. Consistent with a general somatic regulay function, MES protein expression is widespread in larvae, particularly males. These findings suggest that the regulatory relationship between PcG chromatin proteins and thtore Hox genes has been conserved in nematodes (Ross, 2003).
Polycomb group (PcG)-mediated repression of C. elegans Hox genes has not been demonstrated, and genes homologous to components of one of the PcG complexes (PRC1) have not been identified in the C. elegans genome. A mechanism of general Hox gene repression exists in C. elegans, carried out in part by SOP-2, a protein related to, but not orthologous with, any PcG protein. sop-2 mutations lead to widespread ectopic expression of Hox genes and homeotic transformations. SOP-2 contains a SAM domain, a self-associating protein domain found in other repressors, including a core component of PRC1 and ETS transcription factors. Phylogenetic analysis indicates that this domain is more closely related to those of the ETS family than to those of PcG proteins. The results suggest that global repression of Hox genes has been taken over by a different branch of the SAM domain family during the evolution of nematodes (Zhang, 2003).
As seen in fly or mouse PcG mutants, in sop-2 mutants, Hox genes are not expressed in every cell, and each Hox gene is ectopically expressed at a different level. For instance, mab-5 is not expressed in the tail region, and the ectopic expression domains of mab-5 and egl-5 appear to be much broader than those of ceh-13 and lin-39. The simplest explanation for these gene-specific patterns is that, for each gene, transcription factors that will activate expression in the absence of sop-2 gene function are present or active in only a subset of cells. Alternatively, sop-2 repression may be redundant with other repressive mechanisms in some tissues. Crossregulation between Hox genes may also contribute to the Hox gene expression patterns observed in sop-2 mutants, as the data suggest for lin-39 and egl-5 (Zhang, 2003).
sop-2 also appears to play a role in regulating the expression of nonhomeotic genes, since sop-2 mutants have pleiotropic effects not known to be associated with Hox gene misexpression, including abnormalities in body size, sex determination, and vulva development. PcG mutants in other organisms also cause defects that may be caused by inappropriate expression of nonhomeotic genes. For instance, M33 (Pc homolog) mutant mice have slow gonad growth that leads to male to female sex reversal, and loss of function of mel-18, mph-1/rae28, bmi-1 (Pc, ph, and Psc homologs, respectively), and M33 result in cell proliferation defects. Thus, regulation of these distinct pathways may be inherent properties of some PcG genes (Zhang, 2003).
During Caenorhabditis elegans development, the HSN neurons and the right Q
neuroblast and its descendants undergo long-range anteriorly directed
migrations. Both of these migrations require EGL-20, a C. elegans Wnt
homolog. Through a canonical Wnt signaling pathway, EGL-20/Wnt
transcriptionally activates the Hox gene mab-5 in the left Q neuroblast and
its descendants, causing the cells to migrate posteriorly. CAM-1,
a Ror receptor tyrosine kinase (RTK) family member,
inhibits EGL-20 signaling. Excess EGL-20, like loss of cam-1, causes the
HSNs to migrate too far anteriorly. Excess CAM-1, like loss of egl-20,
shifts the final positions of the HSNs posteriorly and causes the left Q
neuroblast descendants to migrate anteriorly. The reversal in the migration
of the left Q neuroblast and its descendants results from a failure to
express mab-5, an egl-20 mutant phenotype. These data suggest
that CAM-1 negatively regulates EGL-20 (Forrester, 2004).
In Caenorhabditis elegans males, different subsets of ventral epidermal precursor (Pn.p) cells adopt distinct fates in a position-specific manner: three posterior cells, P(9-11).p, comprise the hook sensillum competence group (HCG) with three potential fates (1°, 2°, or 3°), while eight anterior cells, P(1-8).p, fuse with the hyp7 epidermal syncytium. This study shows that activation of the canonical BAR-1 β-catenin pathway of Wnt signaling alters the competence of P(3-8).p and specifies ectopic HCG-like fates. This fate transformation requires the Hox gene mab-5. In addition, misexpression of mab-5 in P(1-8).p is sufficient to establish HCG competence among these cells, as well as to generate ectopic HCG fates in combination with LIN-12 or EGF signaling. While increased Wnt signaling induces predominantly 1° HCG fates, increased LIN-12 or EGF signaling in combination with MAB-5 overexpression promotes 2° HCG fates in anterior Pn.p cells, suggesting distinctive functions of Wnt, LIN-12, and EGF signaling in specification of HCG fates. Lastly, wild-type mab-5 function is necessary for normal P(9-11).p fate specification, indicating that regulation of ectopic HCG fate formation revealed in anterior Pn.p cells reflect mechanisms of pattern formation during normal hook development (Yu, 2010).
Overall, vulval precursor cell (VPC) and HCG patterning are quite similar: the precise cell fate is generated by progressive restriction through competence, induction, and lateral inhibition mediated by multiple signal integration at different steps, representing a general scenario of complex pattern formation (Yu, 2010).
Specifically, both VPC and HCG competence are established by Wnt signaling and one of the two Hox genes, lin-39 and mab-5, respectively. Expression patterns of both Hox genes are the same in both hermaphrodite and male, with lin-39 expression in P(3-8).p and mab-5 expression in P(7-11).p. However, sex-specific utilization of these two Hox genes, lin-39 and mab-5, determines whether a hermaphrodite vulva or a male hook, respectively, is formed. In hermaphrodites, lin-39 function is favored in the central Pn.p cells, and the ability of mab-5 to prevent P(9-11).p fusion with hyp7 is somehow blocked. As a transcription factor, mab-5 regulates target gene expression. One possibility is that a negative regulator in hermaphrodites sequesters mab-5 from its targets. Alternatively, mab-5 may act with a co-regulator that is missing in hermaphrodites. The Hox genes appear to play a permissive role in VPC and HCG induction because neither multi-vulvae nor multi-hooks are observed when lin-39 or mab-5, respectively, are overexpressed (Yu, 2010).
A major difference between VPC and HCG development is the major inductive signal used to specify the 1° fate: the EGF pathway induces the 1° VPC fate while Wnt signaling promotes the 1° HCG fate. However, both EGF and Wnt act to induce HCG as well as VPC fates, and it has been observed that excessive Wnt signaling can at least partially substitute for EGF signaling in VPC induction and vice versa in HCG specification. The local abundance of the signal could explain why different inductive signals are utilized in VPC and HCG patterning. The availability of the Wnt and EGF inductive signals differ spatially in hermaphrodites and males, contributing further to the sex-specific bias of Hox gene expression. Although Wnts are present in the central region of the body and the EGF ligand is produced in the tail, the EGF signal emanates from a concentrated source, the gonadal anchor cell, only in the hermaphrodite, while Wnt signaling is more abundant in the tail region as elucidated by extensive tail defects caused by deficient Wnt signaling. As such, only the required Hox gene is promoted in each region in a sex-specific manner -- for example, lin-39 activity in males is not reinforced due to lack of a strong extrinsic signal in the central region. Therefore, different signaling pathways may not be the direct cause of sexually dimorphic organogenesis. The specificity of signaling relies on Hox genes to direct sex-specific pattern formation among competent precursor cells (Yu, 2010).
Many animals are indirect developers with distinct larval and adult body plans. This study compared the larval and adult body plans of an indirect developing hemichordate, Schizocardium californicum. The expression is described of 27 transcription factors with conserved roles in deuterostome ectodermal anteroposterior (AP) patterning in developing embryos, tornaria larvae, and post-metamorphic juveniles and show that the tornaria larva of S. californicum is transcriptionally similar to a truncated version of the adult. The larval ectoderm has an anterior molecular signature, while most of the trunk, defined by the expression of hox1-7, is absent. Posterior ectodermal activation of Hox is initiated in the late larva prior to metamorphosis, in preparation for the transition to the adult form, in which the AP axis converges on a molecular architecture similar to that of the direct developing hemichordate Saccoglossus kowalevskii. These results identify a molecular correlate of a major difference in body plan between hemichordate larval and adult forms and confirm the hypothesis that deuterostome larvae are "swimming heads". This will allow future comparative studies with hemichordates to take into account molecular differences caused by early life history evolution within the phylum. Additionally, comparisons with other phyla suggest that a delay in trunk development is a feature of indirect development shared across distantly related phyla (Gonzalez, 2016).
The normal Pbx1 homeodomain protein (Homolog of Drosophila Extradenticle), as well as its oncogenic
derivative, E2A-Pbx1, binds the DNA sequence ATCAATCAA cooperatively with the murine Hox-A5 (homolog of Drosophila Scr) and
Hox-B7, Hox-B8, and Hox-C8 (Antennapedia class: see four paralogous Hox clusters of mammals) homeodomain proteins, which are themselves known oncoproteins, as well
as with the Hox-D4 (homolog of Deformed) homeodomain protein. Cooperative binding to ATCAATCAA required the
homeodomain-dependent DNA-binding activities of both Pbx1 and the Hox partner. In cotransfection
assays, Hox-B8 suppresses transactivation by E2A-Pbx1. These results suggest that (1) Pbx1 may
participate in the normal regulation of Hox target gene transcription in vivo and thereby contribute to
aspects of anterior-posterior patterning and structural development in vertebrates; (2) that E2A-Pbx1
could abrogate normal differentiation by altering the transcriptional regulation of Hox target genes in
conjunction with Hox proteins, and (3) that the oncogenic mechanism of certain Hox proteins may
require their physical interaction with Pbx1 as a cooperating, DNA-binding partner (Lu, 1995).
Specific residues located within the Pbx
homeodomain are essential for cooperative DNA binding with Hox and
Engrailed gene products. Within the N-terminal region of the Pbx
homeodomain, a residue has been identified that is required for cooperative
DNA binding with three Hox Antennapedia class gene products (Hoxb-7, Hoxb-8 and Hoxc-6) but not for cooperativity with
Engrailed-2 (En-2) (See Drosophila Engrailed). There are similarities between
heterodimeric interactions involving the yeast mating type homeodomain proteins MATa1
and MATalpha2, and those that allow the formation of Pbx/Hox and Pbx/En-2
heterodimers. Specifically, residues located in the a1 homeodomain that form a hydrophobic pocket allowing the alpha2
C-terminal tail to bind, are also required for Pbx/Hox and Pbx/En-2
cooperativity (Peltenburg, 1997).
Three residues located at another site, in the turn
between helix 1 and helix 2 are characteristic of many atypical homeodomain
proteins. These residues, present in Pbx type homeodomains, are required for cooperative DNA binding involving both Hox and
En-2. Replacement of the three residues located in the turn between helix 1
and helix 2 of the Pbx homeodomain with those of the atypical homeo-domain
proteins controlling cell fate in the basidiomycete Ustilago maydis, bE5 and
bE6, allows cooperative DNA binding with three Hox members but abolishes
interactions with En-2. The data suggest that the molecular mechanism of
homeodomain protein interactions that control cell fate in Saccharomyces
cerevisiae and in the basidiomycetes may well be conserved in part in
multicellular organisms. While a number of structural determinates, such as the hydrophobic pocket, are required for cooperativity involving both Hox and Engrailed, others, such as the three amino acid insert, are clearly more specific (Peltenburg, 1997).
A binding site selection strategy was used to determine the optimal binding sites
for Pbx proteins by themselves and as heterodimeric partners with various Hox gene
products. Among the Pbx proteins by themselves, only Pbx3 binds with high affinity,
as a monomer or as a homodimer, to an optimal binding site: TGATTGATTTGAT. An
inhibitory domain located N terminal to the Pbx1 homeodomain prevents intrinsic Pbx1
binding to this sequence. When complexed with Hoxc-6, each of the Pbx gene
products binds the same consensus sequence, TGATTTAT, which differs from the
site bound by Pbx3 alone. Three members of the Antennapedia family, Hoxc-6,
Hoxb-7, and Hoxb-8, select the same binding site in conjunction with Pbx1. These
proteins show similar affinities as heterodimeric partners with Pbx1 for the selected
optimal binding site. However, the binding specificity of Hox proteins for
optimal binding sites is increased, compared to nonspecific DNA, in the presence of
Pbx proteins. Thus, while cooperative DNA binding involving heterodimers of Pbx and
Hox gene products derived from members within the Antennapedia family does not
increase binding site selectivity, DNA binding specificity of the Hox gene products is
significantly enhanced in the presence of Pbx (Neuteboom, 1997).
The distribution of Hoxb-8 transcripts through the chick flank and early forelimb
mirrors the distribution of polarizing activity in the flank at these early stages.
Polarizing activity displayed by Hoxb-8-expressing tissue is only realized when placed
adjacent to the apical ectodermal ridge (AER) and appears to be mediated through Sonic hedgehog (see Drosophila Hedgehog) induction, suggesting that Hoxb-8 may lie genetically upstream of Shh. Accordingly, Hoxb-8 expression is rapidly induced by retinoic acid (RA) treatment in the anterior of the forelimb in a
spatial and temporal manner, consistent with the induction of Shh and formation
of the ZPA. Inhibition of RA synthesis in the flank downregulates the
expression of endogenous Hoxb-8 and results in the loss of Shh expression. However,
once the ZPA has become established the posterior limb mesoderm displays
resistance to the induction of Hoxb-8 expression. Grafting of ZPA cells to the anterior
of a host limb renders the host anterior tissue resistant to RA-induced Hoxb-8
expression. These results indicate that Hoxb-8 expression may be regulated by the
established ZPA through a negative feedback loop. The anterior AER also secretes an
inhibitory factor, preventing RA-induced or already established Hoxb-8 expression in
the cells immediately underneath the AER. Consistent with a role for Hoxb-8 in
positioning of the forelimb ZPA, Hoxb-8 expression is not seen in RA-induced
duplications at the anterior of the hindlimb. However, grafting of Hoxb-8-expressing
tissue to the hindlimb can lead to Shh expression and similar duplications, suggesting
that factors mediating ZPA formation are very similar in both wing and leg (Stratford, 1997).
Mice deficient for the homeotic gene Hoxc-8 suffer from a congenital prehension deficiency
of the forepaw. During embryogenesis, Hoxc-8 is highly expressed in motoneurons within
spinal cord segments C7 to T1. These motoneurons innervate forelimb distal muscles that
move the forepaw. In Hoxc-8 mutant embryos, formation of these muscles is normal, but
their innervation is perturbed. From E13.5 onward, distal muscles normally supplied by
C7-8 MNs also receive ectopic projections from C5-6 and T1 motoneurons. Coordinates of
motor pools are altered along both the rostrocaudal and mediolateral axes. Following this
aberrant connectivity pattern and during the time of naturally occuring cell death, apoptosis
is specifically enhanced in C7-T1 motoneurons. Loss of Hox-encoded regional specifications
subsequently leads to a numerical deficit of motoneurons and an irreversible disorganization
of motor pools. In Hoxc-8 null mutants, C7-8 motoneurons lose their selective advantage in
growth cone pathfinding behavior and/or target recognition, two essential steps in the
establishment and maintenance of a functional nervous system (Tiret, 1998).
Retinoid signaling is required for the establishment of a zone of polarizing activity (ZPA) and for the expression of
Hoxb-8, a mediator of ZPA formation in the chick. Retinoid receptor antagonists applied to the presumptive wing region block
the formation of a ZPA. Although Hoxd-13 is still expressed after treatment with anti-retinoids, both sonic hedgehog and bmp-2 expression are absent in limb bud mesenchyme of wing regions treated with anti-retinoid activity. This suggests a direct relationship
between retinoid signaling and the establishment of the ZPA. The
Hox gene Hoxb-8 is a direct target of retinoid signaling since exogenously applied RA
rapidly induces this gene in the absence of protein synthesis; moreover, retinoid
receptor antagonists down-regulate Hoxb-8 expression. In addition in the
lateral plate mesoderm, the domains of Hoxb-8 expression and of polarizing activity are
coextensive. Taken together, these findings support the hypothesis that retinoids are
required for the establishment of a ZPA, and that retinoids act, at least in part, through
Hoxb-8, a gene associated with ZPA formation. Apparently there is only a brief temporal overlap between Hoxb-8 and shh expression. Furthermore, only a subset of Hoxb-8 expressing cells eventually express shh. It is thought that the RA-induced ectodermal factor is required for the formation of the ZPA. It is thought that shh is expressed in those Hoxb-8 positive cells that are in proximity to the AER and thus receive the FGF signal (Lu, H.-C., 1997).
A short sequence element (L7ATE) within the proximal
promoter of a Purkinje cell-specific gene, pcp-2(L7), is required for the normal pattern of expression of
this gene in the cerebellum of transgenic mice. The presence of a series of TAAT sequence motifs in
this element suggests its interaction with homeodomain proteins. To extend these observations,
degenerate oligonucleotides were used to clone by reverse-transcriptase polymerase chain reaction
members of the mouse Hox gene family expressed in neonatal cerebellum but not forebrain. Two of
these, HoxB7 and HoxA5, are continuously expressed from the neonatal period into adult stages in
cerebellar Purkinje cells. These Hox proteins are shown to synergistically activate the L7 promoter by
cotransfection assay in vitro. In contrast, another homeodomain protein that is normally expressed in
Purkinje cells only during the embryonic period, En-2, has a negative effect on L7 gene expression.
These data suggest a biphasic, combinatorial control mechanism for the Purkinje cell-specific
expression of the pcp-2(L7) gene (Sanliouglu, 1998).
The proper expression of Hox genes is necessary for the accurate patterning of the body plan. The elucidation of the developmental genetic basis of transcriptional regulation of Hox genes by the study of their cis-regulatory elements provides crucial information regarding the establishment of axial specification. The role of the early enhancer (EE) of the murine Hoxc8 gene has been investigated to better understand its role in pattern formation. Knockouts of the endogenous Hoxc8 coding region result in a combination of neural, behavioral and skeletal phenotypes. This report is limited to a consideration of the skeletal abnormalities. A 200 bp non-coding element 3 kb upstream of the Hoxc8 promoter is a crucial enhancer that regulates the transcription of Hoxc8. This regulatory region has been deleted from the endogenous genome using embryonic stem cell technology. Deletion of the EE results in a significant delay in the temporal expression of Hoxc8. Deletion of the EE does not eliminate the expression of the Hoxc8 protein, but delays the attainment of control levels of expression and anterior and posterior boundaries of expression on the AP axis. The temporal delay in Hoxc8 expression is sufficient to produce phenocopies of many of the axial skeletal defects associated with the complete absence of Hoxc8 gene product as previously reported for the Hoxc8-null mutation. These results are consistent with emerging evidence that the precise temporal expression of Hox genes is crucial for the establishment of regional identities. The fact that the EE deletion does not eliminate Hoxc8 expression indicates the existence of a Hoxc8 transcriptional regulatory apparatus independent to some degree of the Hoxc8 EE. In a comparison of these results with those reported previously by others investigating temporal control of Hox gene expression, a structural similarity was found between the Hoxc8 EE reported here and a transcriptional control element located in the Hoxd11 region. It is speculated that a distributed system of expression timing control may exist that is similar to the one proposed for Hoxc8. These data are consistent with the idea that disparate regulatory pathways are responsible for the expression of Hoxc8 in the organogenesis of somites, neural tube and limb bud (Juan, 2003).
To analyze requirements for Notch signalling in patterning the
paraxial mesoderm, transgenic mice were generated that express in the paraxial mesoderm a dominant-negative version of Delta1. Transgenic mice with reduced Notch activity in the presomitic mesoderm as indicated by loss of Hes5 expression were viable and displayed defects in somites and vertebrae consistent with known roles of Notch signalling in somite compartmentalization. In addition, these mice showed with variable
expressivity and penetrance alterations of vertebral identities resembling
homeotic transformations, and subtle changes of Hox gene expression in day
12.5 embryos. Mice that carried only one functional copy of the endogenous
Delta1 gene also showed changes of vertebral identities in the lower cervical region, suggesting a previously unnoticed haploinsufficiency for Delta1. Likewise, in mice carrying a null allele of the oscillating Lfng gene, or in transgenic mice expressing Lfng constitutively in the presomitic mesoderm, vertebral identities were changed and numbers of segments in the cervical and thoracic regions were reduced, suggesting anterior shifts of axial identity. Together, these results provide genetic evidence that precisely regulated levels of Notch activity as well as cyclic Lfng activity are critical for positional specification of the
anteroposterior body axis in the paraxial mesoderm (Cordes, 2004).
Transformations of vertebral identities, anterior shifts of Hoxb6 expression, and of the position of both fore and hind limb buds were detected in mice lacking Lfng function or expressing Lfng constitutively. An apparent anterior shift of Hoxb6 expression in Lfng mutant embryos could also be expected if fewer segments were generated in the prospective cervical region, whereas the absolute position of the anterior Hoxb6 expression border along the anteroposterior body axis was maintained. Recent models of somite segmentation suggest that the interaction of graded FGF or WNT signals with the segmentation clock generates the periodic somite pattern. Conceptually, increasing the steepness of the gradient or slowing the periodicity of the clock would lead to fewer segments, which in either case would be larger. Thus, if the loss of Lfng would affect the clock (output) and fewer segments would be formed in the prospective cervical region, they should be larger than normal. However, the five cervical segments in Lfng mutant embryos occupied essentially the same space as the anterior five segments in WT embryos, strongly supporting the idea that the rostral Hoxb6 expression border is indeed shifted anteriorly. The positions of the fore and hind limb buds are invariant in WT embryos and correspond to the transition between the cervical and thoracic, and lumbar and sacral regions, respectively. Their anterior shift suggests homeotic transformations throughout the trunk region along the anterior posterior body axis that led to an overall reduction of the number of segments in the trunk (Cordes, 2004).
Retinoic acid (RA) signaling plays an important role in determining the anterior boundary of Hox gene expression in the neural tube during embryogenesis. In particular, RA signaling is implicated in a rostral expansion of the neural expression domain of 5' Hoxb genes (Hoxb9-Hoxb5) in mice. However, underlying mechanisms for this gene regulation have remained elusive due to the lack of RA responsive element (RARE) in the 5' half of the HoxB cluster. To identify cis-regulatory elements required for the rostral expansion, a recombineering technology was developed to serially label multiple genes with different reporters in a single bacterial artificial chromosome (BAC) vector containing the mouse HoxB cluster. This allowed simultaneous monitoring of the expression of multiple genes. In contrast to plasmid-based reporters, transgenic BAC reporters faithfully recapitulated endogenous gene expression patterns of the Hoxb genes including the rostral expansion. Combined inactivation of two RAREs, DE-RARE and ENE-RARE, in the BAC completely abolished the rostral expansion of the 5' Hoxb genes. Knock-out of endogenous DE-RARE lead to significantly reduced expression of multiple Hoxb genes and attenuated Hox gene response to exogenous RA treatment in utero. Regulatory potential of DE-RARE was further demonstrated by its ability to anteriorize 5' Hoxa gene expression in the neural tube when inserted into a HoxA BAC reporter. These data demonstrate that multiple RAREs cooperate to remotely regulate 5' Hoxb genes during CNS development, providing a new insight into the mechanisms for gene regulation within the Hox clusters (Ahn, 2014).
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Antennapedia:
Biological Overview
| Regulation
| Targets of activity
| Protein Interactions
| Developmental Biology
| Effects of Mutation
| References
Society for Developmental Biology's Web server.
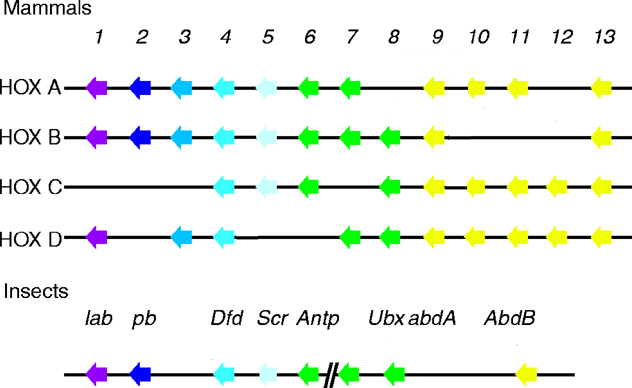{kind=link}