abdominal-A
The six Drosophila proteins that belong to the antennapedia-type Homeobox subfamily are
Antennapedia (ANTP), Abdominal-A (ABD-A), Deformed (DFD), Proboscipedia (PB),
Sex combs reduced (SCR) and Ultrabithorax (UBX).
The ExPASy World Wide Web (WWW) molecular biology server of the Geneva University Hospital and
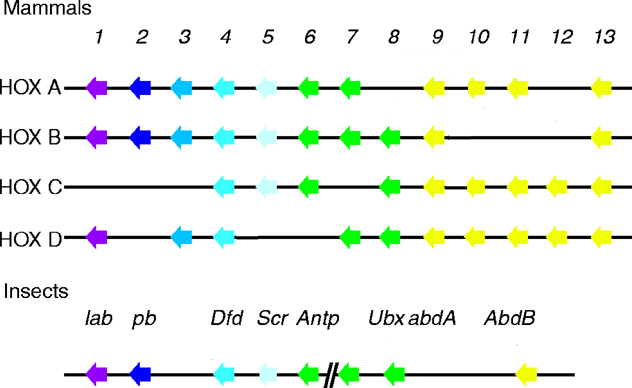
the University of Geneva provides extensive documentation for the 'Homeobox' antennapedia-type protein signature. The four paralogous Hox clusters of mammals contain eight ABD-A homologs: HoxA6, HoxA7, HoxB6, HoxB7, HoxB8, HoxC6, HoxC8 and HoxD8.
The Drosophila homeotic selector gene abdominal-A is important for determinative decisions in the
anterior abdomen. Insects vary considerably with respect to abdominal morphology, and changes in the
function of homeotic selector genes and/or downstream genes under their control presumably have
been important to the evolution of these differences. Mutations in Abdominal, the Tribolium ortholog of
abdominal-A, have been described, and have more posterior homeotic transformations than do
Drosophila variants (see Tribolium early embryonic development). The organization of the Abdominal gene and the sequences of its
predicted proteins, the first such report for a non-Drosophilid insect, is reported. Two predicted proteins share
N-terminal sequences with those proposed to be synthesized by the Drosophila ortholog. In addition,
the distribution of Abdominal transcripts during embryogenesis is described. The Tribolium expression
pattern closely resembles that of Drosophila, and does not account for the differences in mutant
phenotypes (Shippy, 1998).
Insects are easily distinguishable by the absence of legs on the adult abdomen. Studies performed on the Dipteran, Drosophila melanogaster, indicate that this is because of the repressive effects of the homeotic genes Ultrabithorax and abdominal-A on the limb promoting gene Distal-less during embryonic development. However, in many species appendage-like
structures are present on abdominal segments in embryonic and juvenile stages. By using classical genetics and double-stranded
RNA-mediated gene silencing in the red flour beetle, Tribolium castaneum, a species that develops an appendage on the first
abdominal segment, it was possible to examine the roles of Ubx and Abd-A in abdominal limb development. In Tribolium, Abd-A, but not Ubx, represses early
expression of Dll in the embryonic abdomen. Ubx appears to modify the A1 appendage. This difference in the activities of Abd-A and Ubx is critical for proper
development of this appendage. It is suggested that an ancestral role of Abd-A in insect abdominal appendage development was in the repression of Dll initiation and
that an ancestral role of Ubx was in modulation of abdominal appendage morphology (Lewis, 2000).
By examining TcDll and TcEn expression in TcUbx and
Tcabd-A mutant embryos, a better
understanding of the role of each in suppressing and modifying limb
programs in the beetle abdomen was obtained. In TcUbx mutant embryos,
TcDll expression in the abdomen remains restricted to anterior A1, whereas, in Tcabd-A, mutant embryo
TcDll is ectopically expressed in each abdominal segment,
resulting in abdominal appendage development. These results clearly
support the role for TcAbd-A as a primary TcDll
repressor (and therefore appendage repressor) in the
Tribolium abdomen. The role of TcUbx in
regulating Dll expression appears to be more complex. Although
TcDll and TcUbx are initially coexpressed during
early pleuropod development, later TcUbx is absent in the
TcDll-expressing cells, leaving open the possibility that
TcUbx represses TcDll late in development. Whether or not late expression of TcUbx represses
TcDll expression in these cells, it is evident from mutant
analysis that TcUbx is required for the proper
differentiation of these cells. In TcUbx mutants, the nuclei
of TcDll-expressing cells in the pleuropod never become
morphologically distinct as they do in the wild type. It is therefore
believed that TcUbx acts as a modifier rather than a
repressor of abdominal appendage development (Lewis 2000).
The dynamic relationship between TcUbx and TcDll
expression in the pleuropod and the effect of TcUbx
expression on the differentiation of TcDll-expressing cells
suggests that TcUbx acts to modify the way cells in the
anterior A1 compartment interpret signaling cues. In the absence of
TcUbx, cells respond to signaling cues as if they were no
longer pleuropodial. The failure of the appendage to invaginate and the
presence of the subterminal tarsal claw in TcUbx mutant
larvae support this view. In addition, the position of the subterminal
tarsal claw appears to correspond to the boundary of TcEn
expression and the cluster of TcDll-expressing cells in the
developing appendage of the embryo. This is interpreted as evidence that
these cells now respond to signaling cues as if they were leg, with the
distal-most tip, the tarsal claw in the leg, at the intersection of the
anterior-posterior boundary (Lewis 2000).
Differences in the manner in which TcUbx-expressing
cells respond to signaling cues could be because of TcUbx
acting directly on signaling pathway components or their targets.
Studies performed on Ubx control of wing vs. haltere development in
Drosophila have indeed shown that Ubx can act at multiple
levels of a genetic hierarchy. In the case of pleuropod
development, the levels of Ubx and/or the presence of Hox cofactors
are likely to be responsible for pleuropod-specific gene expression. The former explanation is favored since very high levels of TcUbx
are found in the pleuropod, compared with the levels found in other
regions of the embryo. The levels of TcUbx expression may be
important to outcompete other proteins expressed in these cells, such
as Antennapedia, which normally promotes leg patterning. In
addition, it has been shown that TcUbx levels are decreased
in TcEn-expressing cells of the thorax and abdomen in
wild-type embryos. Differences in TcUbx levels in these
compartments may also explain why, in Tcabd-A mutants, only
the cells in the anterior compartment of the abdominal segments are
able to differentiate as pleuropodial cells, whereas the
TcEn-expressing cells in the posterior compartment differentiate as leg cells. The possible effect of Ubx levels on
pleuropod patterning is consistent with data obtained in Drosophila on the effects of Ubx levels on patterning ps6 in
the embryo and bristles on the T2 leg in the adult (Lewis 2000).
Comparing the data obtained in this study on beetle abdominal
appendage development with that obtained from other holometabolous insects, it is suggested that abdominal limb repression
through direct Abd-A repression of Dll expression evolved at the latest
in the last common ancestor of the holometabola. This is the most
parsimonious interpretation, given that the repressive activity of Abd-A
is evident in species from all of the holometabolous orders examined.
However, one holometabolous insect species, the Lepidopteran
Manduca sexta, appears to be an exception. In the
developing abdominal prolegs in this species, Dll is expressed despite
the coexpression of Ubx/Abd-A. It is interesting to note that the
ability to express Dll in developing prolegs has arisen using at least
two different mechanisms within the Lepidoptera. In the butterfly
Precis coenia, activation of Dll expression in the abdomen
is correlated with regional repression of Ubx/Abd-A, whereas, in
the moth Manduca sexta, Dll expression occurs through a
different mechanism, presumably involving the escape of Dll from the
repressive effects of Abd-A. These data suggest that the release
of the repressive effect of Abd-A on abdominal limbs in higher
holometabolous insects occurred convergently through changes at
different levels of the limb regulatory hierarchy. Alternatively, it is
possible, although less likely, that the regional
repression/expression of Ubx/Abd-A has no causative effect on
proleg outgrowth, leaving open the possibility that the presence of
prolegs in these two Lepidopteran species is not convergent (Lewis 2000).
In higher holometabolous insect species, such as those found in the
orders Diptera and Lepidoptera, Ubx can act as a primary repressor of
Dll expression in the abdomen, whereas, in the more basal species such
as Tribolium, Ubx acts instead as a modifier of abdominal
limb development. Both the modifier role of Ubx in the anterior A1
compartment and the repressive role of Abd-A in the posterior
compartment are required for proper pleuropod development in
Tribolium. Because pleuropodia develop in the A1 segment of most insect orders, it is believed limb modification rather than limb
repression is a more ancient property of Ubx. Given the
conserved expression patterns of Ubx and Abd-A in the insect abdomen,
it will be of interest to examine how the functions of these genes in
regulating abdominal appendage development have changed during the
course of insect evolution (Lewis 2000).
During the embryogenesis of Drosophila, the homeotic genes are required to
specify proper cell fates along the anterior-posterior axis
of the embryo. Partial cDNAs of homologs
of the Drosophila homeotic gene teashirt and five of the
homeotic-complex (HOM-C) genes were cloned from the thysanuran
insect, Thermobia domestica (the firebrat), and these genes were assayed for their embryonic
expression patterns. The HOM-C genes examined
were labial, Antennapedia, Ultrabithorax, abdominal-A and Abdominal-B. Since the expression pattern of these HOM-C genes is largely conserved among insects
and since Thermobia is a member of a phylogenetically basal
order of insects, the ancestral expression patterns of these genes in insects could be inferred. The expression patterns of the Thermobia HOM-C genes were compared with their
expression in Drosophila and other insects;
the potential roles these genes may have played in insect
evolution are discussed. Interestingly, the teashirt homolog shows
greater variability between Thermobia and Drosophila
than any of the HOM-C genes. In particular, teashirt is
not expressed strongly in the Thermobia abdomen, unlike
Drosophila teashirt. It is proposed that teashirt expression
has expanded posteriorly in Drosophila and contributed to a homogenization of the Drosophila larval thorax and abdomen (Peterson, 1999).
Like Ubx, Drosophila abd-A promotes abdominal development
and suppresses thoracic development. It is expressed in pA1-aA7, nearly the entire
abdominal region. Similar abdominal expression is seen
in other insect species (Thermobia, Tribolium
and Schistocerca) and the pA1 anterior boundary of
the abd-A expression pattern is highly conserved, though
some variation among insects is observed in the initiation
pattern. The posterior border of abd-A shows some variability.
In both grasshoppers and firebrats, abd-A initially extends
through A10, the final abdominal segment; then it
retracts to A9. In Drosophila, which has eight abdominal
segments, the posterior border is A7 dorsally and laterally, while expression extends into
aA8 (the anterior part of A8) ventrally. Only Tribolium shows extensive variation;
Tc-Abdominal (the Tribolium abd-A ortholog) initially extends
beyond A10 at the fully elongated stage and then later
retracts to A8. The differences in the posterior boundary reflect the
differences in the morphology of the abdominal terminus,
the length of the abdomen and the expression of the
Abd-B orthologs. The posterior abdomen
has been highly modified in the evolution of higher insects
like Drosophila, Tribolium, and fleas, with strikingly
non-abdominal characteristics (Peterson, 1999).
The aA1-pA1 distinction between Ubx and abd-A expression
has been conserved among all insects and is
therefore the probable ancestral condition. In Drosophila
and Tribolium, this distinction is important for the
unique identity of A1 versus more posterior abdominal
segments. In Drosophila, A1 produces a unique abdominal denticle belt
(it is missing denticle rows 1 and 4) and in Tribolium, Abdominal (Tc-abd-A)
mutations produce ectopic pleuropodia. Since the presence of pleuropodia also serves to distinguish A1 from other abdominal segments, it is interesting
that of the two genes, only Ubx appears to be expressed
in the pleuropodia. Thus, it would appear that Ubx is required to specify a unique fate for aA1 in firebrats as it is in Drosophila and Tribolium (Peterson, 1999).
One aspect of abd-A expression that does not seem to
be conserved in all insects is its intrasegmental modulation
in Drosophila, whereby it is expressed strongly in
posterior compartments and more weakly in anterior
compartments. No modulation is apparent in firebrat or
grasshopper embryos. However, stronger expression of Tribolium
Abdominal mRNA is consistently found in the posterior
compartments. The intersegmental variation of Drosophila abd-A is thought to produce the complimentary Drosophila Ubx expression pattern by negative regulation; thus, this variation in the pattern of abd-A expression
suggests that cross regulation of Ubx by abd-A in
Drosophila, and perhaps Tribolium, may be a derived
characteristic (Peterson, 1999).
Changes in the expression of the Hox genes Ultrabithorax and Abdominal in different crustaceans correlate well with the modification of their anterior thoracic limbs into feeding appendages (maxillipeds). In branchiopod crustaceans (such as Artemia), which do not have maxillipeds, Ubx and abdA are expressed throughout the thoracic region. In peracarids, the first, and sometimes second, of the eight thoracic segments bear limbs that have acquired several characteristics of feeding appendages. The modification of these segments correlates with the repression of Ubx and abdA in these segments. Uniform early expression becomes modulated within individual metameres during later development. Decapods are generally described as having three pairs of maxillipeds and five pairs of walking limbs in their thorax. In Periclimenes Ubx and abdA expression is excluded from the first three thoracic parasegments and limbs, is weaker in T4, and stronger in more posterior segments. In Homarus, only the T1 and T2 limbs appear to be distinctly reduced at hatching. Ubx and abdA staining is absent from the first two thoracic parasegments and strong in T3 and more posterior segments. Thus, the anterior boundary of embryonic expression of Ubx and abdA in Homarus appears to be shifted backwards by two metameric units corresponding to the morphological transition in thoracic limbs seen at hatching. It is suggested that spatially modulated distribution of Ubx and abdA expression and temporal changes in the expression of Hox genes are responsible for different decisions on regional identity. In some limbs identity could be determined as a mosaic, with some parts of a segment retaining a thoracic identity and others becoming homeotically transformed to a gnathal fate (Averof, 1997).
Representatives of the Insecta and the Malacostraca
(higher crustaceans) have highly derived body plans
subdivided into several tagma (groups of segments united
by or fused into a common function and/or morphology). The
tagmatization of segments in the trunk, the part of the body
between head and telson, in both lineages is thought to have
evolved independently from ancestors with a distinct head
but a homonomous, undifferentiated trunk. In the
branchiopod crustacean, Artemia franciscana, the trunk
Hox genes are expressed in broad overlapping domains
suggesting a conserved ancestral state. In comparison, in
insects, the Antennapedia-class genes of the homeotic
clusters are more regionally deployed into distinct domains
where they serve to control the morphology of the different
trunk segments.
In Drosophila Antp is expressed in
and required for the specification of the three-segmented
locomotory thorax. Both Ubx and abd-A are involved in the
development of the legless abdomen. Ubx is also expressed in the posterior thorax where it is known to be involved in the development of the modified hind
wings, the halteres. Thus an originally Artemia-like pattern of
homeotic gene expression has apparently been modified in
the insect lineage associated with and perhaps facilitating
the observed pattern of tagmatization. Since insects are the
only arthropods with a derived trunk tagmosis tested to
date, the expression patterns of the Hox genes
Antp, Ubx and abd-A were examined in the malacostracan crustacean
Porcellio scaber (Oniscidae, Isopoda). Unlike
the pattern seen in Artemia, these genes are expressed in
well-defined discrete domains coinciding with tagmatic
boundaries that are distinct from those of the insects. These
observations suggest that, during the independent
tagmatization in insects and malacostracan crustaceans,
the homologous 'trunk' genes evolved to perform different
developmental functions. It is also proposed that, in each
lineage, the changes in Hox gene expression pattern may
have been important in trunk tagmatization (Abzhanov, 2000).
Contemporary molecular and morphological phylogenies of
the Crustacea indicate that this group comprises a
monophyletic assembly with some classes such as the
Remipidia and Branchiopoda at a basal position and the
Malacostraca as a crown group. Additionally, according to recent phylogenies, the
Crustacea are placed as the sister group to the Insecta in the
subphylum Mandibulata. Alternatively, some studies suggest
that crustaceans may be paraphyletic with regard to the Insecta
with the Malacostraca as the closest sister group to insects. The
Mandibulata also includes the more distantly related
Myriapoda. The Chelicerates are generally regarded as a sister
group to the Mandibulata (Abzhanov, 2000 and references therein).
The developmental role of abd-A in insects is similar to Ubx. The products of both genes are expressed
in similar, largely overlapping domains and are used to specify
abdominal identity.
The Drosophila abd-A expression domain covering A1-A7 (out
of 10 segments) is similar in Thermobia, Schistocerca,
Tribolium and Manduca. The anterior
boundary in the posterior A1 segment is conserved amongst all
insects studied. The more basal insects, however, have a
posterior boundary that extends to the end of the abdomen into
the A10 segment, suggesting retraction of this boundary during
insect evolution, perhaps via changes in expression and
function of the Abd-B gene. In Artemia,
unlike insects, abd-A is expressed in the trunk region anterior
to the genital and postgenital segments. The anterior boundary lies in the second trunk segment
and expression is strongest in the neuromeres. The resulting
overlap with the other trunk genes, Antp and Ubx, suggests
redundant, fractional and/or mosaic control over trunk identity. Recent
studies on chelicerates have revealed broadly overlapping
expression domains of these trunk Hox genes in the
opisthosoma, the posterior chelicerate tagma.
These observations from a recognized non-mandibulate
outgroup confirm the proposed ancestral condition concluded
from studies on Artemia. Thus it
would appear that the Psabd-A pleonically restricted domain
of expression is a derived condition. The
resemblance to the insect abd-A domain in the abdomen, which
is analogous to the pleon, is intriguing and implies the
deployment of abd-A to the posterior tagmata has occurred
separately in the insect and crustacean lineages. The salient
difference being that sequestration of abd-A was accompanied
by Ubx in the insects but was accomplished singly in the case
of higher crustaceans (Abzhanov, 2000).
In summary, the expression domains of the trunk genes in P. scaber are distinct from both insects and branchiopod
crustaceans. They are better defined than the broadly
overlapping domains in A. franciscana and despite a superficial
resemblance to the discrete domains of their insect
homologs, the anterior and posterior expression domain
boundaries are quite different from those in insects. These
observations suggest that the trunk genes were co-expressed
and performed redundant roles in the homonomous trunk in the
last ancestor of insects and higher crustaceans, and that the
trunk of the ancestor has independently differentiated into the
thorax/abdomen of insects and pereon/pleon of
malacostracans via specialization in the deployment and
function of the Hox genes. This being the case, it is likely that
the homologous Hox trunk genes have evolved to acquire
different developmental functions in the closely related classes
Insecta and Malacostraca (Abzhanov, 2000 and references therein).
Classical embryological experiments suggest that a posterior signal is required for
patterning the developing anteroposterior axis. In this paper, a potential role in Xenopus is investigated
for FGF signaling during this process. During normal development, embryonic fibroblast growth
factor (eFGF) (See Drosophila Branchless) is expressed in the dorsal mesoderm, specifically, in the notochord and in the posterior mesoderm around the closing blastopore. Overexpression of eFGF from the start of gastrulation results in a posteriorised
phenotype of reduced head and enlarged proctodeum. The overexpression of eFGF causes the up-regulation of a number of posteriorly expressed genes, and prominent among these are Xcad3, a caudal homolog, and the Hox genes, in particular HoxA7 (a homolog of Drosophila abd-A). There is both an increase of expression
within the normal domains and an extension of expression towards the anterior. Application
of eFGF-loaded beads to specific regions of gastrulae reveals that anterior truncations arise
from an effect on the developing dorsal axis. Similar anterior truncations are caused by the
dorsal overexpression of Xcad3 or HoxA7. This suggests that this aspect of the eFGF
overexpression phenotype is caused by the ectopic activation of posterior genes in anterior
regions. Further results using the dominant negative FGF receptor show that the normal expression
of posterior Hox genes is dependent on FGF signaling and that this regulation is likely
mediated by the activation of Xcad3. It has been demonstrated that the eFGF regulates the transcription of Xbra (Drosophila homolog: T-related gene) and that Xbra can in turn activate eFGF expression. Xbra does not directly activate Hox gene expression. However, at the very least, Xbra clearly plays an indirect role in anteroposterior specification through its regulation of eFGF expression in the notochord and the posterior of the embryo. The biological activity of eFGF, together with its
expression in the posterior of the embryo, make it a good candidate to fulfil the role of the
'transforming' activity proposed by Nieuwkoop in his 'activation and transformation' model
for neural patterning (Pownall, 1996).
The biological activities of the Xenopus caudal (Cdx) family member Xcad3 have been examined. A series of
domain-swapping experiments demonstrate that the N-terminus of Xcad3 is necessary for it to activate Hox gene expression
and that this function can be replaced by the activation domain from the viral protein VP16. Injection of 50 pg or more of
Xcad3 mRNA leads to activation of HoxC6 and HoxA7, which are normally expressed in both
the mesoderm and neuroectoderm, and HoxB7 and HoxB9, which are expressed predominantly in
the neuroectoderm. Xcad3 does not upregulate the expression of the general mesodermal
marker Xbra, indicating that it does not induce the formation of ectopic mesoderm. Experiments using an
Xcad3 repressor mutant (XcadEn-R), which potently blocks the activity of wild-type Xcad3, are reported. Overexpression of
XcadEn-R in embryos inhibits the activation of the same subset of Hox genes that are activated by wild-type Xcad3 and leads
to a dramatic disruption of posterior development. Xcad3 is shown to be an immediate early target of the FGF signaling
pathway: Xcad3 and FGF both posteriorize anterior neural tissue in similar ways. Xcad3 is also required for the
activation of Hox genes by FGFs. These data provide strong evidence that Xcad3 is required for normal posterior development
and that it regulates the expression of the Hox genes downstream of FGF signaling (Isaacs,1998).
Is there any evidence that the role for caudal-related genes in regulating Hox genes is conserved
outside the vertebrates? In Drosophila, homologs of the vertebrate Hox genes (Hom-C complex)
are considered to be largely epistatic to caudal, but there is now evidence suggesting that some aspects
of expression from the HOM-C complex member Abdominal-B are regulated by caudal. Interestingly, ectopic anterior expression of caudal results in a disruption of head
development, part of which appears to be due to the suppression of expression by the deformed
gene, which is also a member of the Drosophila Hom-C complex.
Certain parallels can be seen with the posterior-promoting/anterior-suppressing activity of the
Xenopus Xcad proteins. With regard to the role of caudal-related genes in other invertebrates, it
has been suggested that pal-1 is involved in regulation of the C.elegans Abd-B homolog mab-5 (Isaacs, 1998).
Transposition of anatomical structures along the anteroposterior axis has been a commonly used mechanism for changing body
proportions during the course of evolutionary time. Transposition in
mesodermal derivatives (vertebrae) could be attributed to transposition in the expression of Hox genes along the axial series of
somites. Transposition in the segmental arrangement of the spinal nerves can also be correlated with shifts in
the expression domains of Hox genes. Specifically, the expression domains of Hoxa-7, a-9 and a-10 in spinal
ganglia correspond in both mouse and chick to the positions of the brachial and lumbosacral plexuses, and this is
true even though the brachial plexus of chick is shifted posteriorly, relative to mouse, by seven segmental units. In spite of
these marked species differences in the boundaries of Hoxa-7 expression, cis regulatory elements located up to 5 kb upstream
of the chick Hoxa-7 gene show much functional and structural conservation with those described in the mouse. Chick Hoxa-7 and a-10 expression domains spread forward into regions of somites that are
initially negative for the expression of these genes. This is discussed as evidence that Hox expression in paraxial mesoderm
spreads forward, as earlier found for neurectoderm and lateral plate mesoderm, in a process that occurs independent of cell
movement (Gaunt, 1999).
Studies of pattern formation in the vertebrate central nervous system indicate that anteroposterior positional information is
generated in the embryo by signaling gradients of an as yet unknown nature. Transcription factors were sought that
transduce this information to the Hox genes. Based on the assumption that the activity levels of such factors might vary with
position along the anteroposterior axis, an in vivo assay was devised to detect responsiveness of cis-acting sequences to such
differentially active factors. This assay was used to analyze a Hoxb8 regulatory element, and the most pronounced
response was detected in a short stretch of DNA containing a cluster of potential CDX binding sites. Differentially expressed
DNA binding proteins are present in gastrulating embryos that bind to these sites in vitro (included among these proteins are cdx gene products). Binding site mutations that abolish binding of these proteins completely destroy the ability of the regulatory
element to drive regionally restricted expression in the embryo. Ectopic expression of cdx gene products
anteriorizes expression of reporter transgenes driven by the CDX binding regulatory element, as well as that of the endogenous Hoxb8 gene,
in a manner that is consistent with CDX genes being essential transducers of positional information. These data suggest that, in
contrast to Drosophila Caudal, vertebrate cdx gene products transduce positional information directly to the Hox genes, acting
through CDX binding sites in their enhancers. This may represent the ancestral mode of action for caudal homologs, which
are involved in anteroposterior patterning in organisms with widely divergent body plans and modes of development (Charite, 1998).
Using a xenograft model of fetal intestinal anlagen implanted under the
skin of nude mice, the expression of five homeobox genes
(HoxA-4, HoxA-9, HoxC-8, Cdx-1 and Cdx-2) was examined. In homotypic associations of fetal endoderm and mesenchyme that
recapitulate normal development, the overall pattern of homeobox gene expression is
maintained: HoxA-9 (homologous to Drosophila Abd-B) and HoxC-8 (homologous to Drosophila abd-A) were the highest in the colon and ileum, respectively, and
HoxA-4 (homologous to Drosophila Deformed) is expressed all along the intestine. Cdx-1 and Cdx-2 (Both homologs of Drosophila caudal) exhibit an increasing
gradient of expression from small intestine to colon. Grafting per se causes a faint
upregulation of HoxA-9 and HoxC-8 in small intestinal regions where these genes are not
normally expressed, while the endoderm-mesenchyme dissociation-association step
provokes a decay of Cdx-1 in the colon. In heterotopic associations of colonic endoderm
with small intestinal mesenchyme, the colonic epithelium exhibits heterodifferentiation into a
small intestinal-like phenotype. In this case, a decay of HoxA-9 expression and
an upregulation of HoxC-8 is observed. Heterodifferentiation of the colonic epithelium is
accompanied by a downregulation of Cdx-1 and Cdx-2 to a level similar to that found in the
normal small intestine. To demonstrate that mesenchyme-derived cells can influence Cdx-1
and Cdx-2 expression in the bowel epithelium, fetal jejunal endoderm was associated with
intestinal fibroblastic cell lines that either support small intestinal-like or colonic-like
morphogenesis. A lower expression of both homeobox genes occurs in grafts presenting
the small intestinal phenotype than in those showing glandular colonic-like differentiation.
Taken together, these results suggest that homeobox genes participate in the control of the
positional information and/or cell differentiation in the intestinal epithelium. They also
indicate that the level of Cdx-1 and Cdx-2 homeobox gene expression is influenced by
epithelial-mesenchymal cell interactions in the intestinal mucosa (Duluc, 1997).
The expression patterns of two distantly clustered Hox genes were studied:cHoxc-8, a median
paralog, and cHoxd-13, located at the 5' extremity of the HoxD cluster. These could, respectively, be involved
in specification of dorsal feather- and foot scale-forming skin in the chick embryo. The cHoxc-8 transcripts are
present at embryonic day 3.5 (E3.5) in the somitic cells, which give rise to the dorsal dermis by E5, and
at E6.5-8.5 in the dorsal dermal and epidermal cells during the first stages of feather morphogenesis.
The cHoxd-13 transcripts are present at E4.5-9.5 in the autopodial mesenchyme and at E10.5-12.5 in
the plantar dermis during the initiation of reticulate scale morphogenesis. Both the cHoxc-8 and
cHoxd-13 transcripts are no longer detectable after the anlagen stage of cutaneous appendage
morphogenesis. Heterotopic dermal-epidermal recombinations of dorsal, plantar, and
apteric tissues reveal that the epidermal ability or inability to form feathers is already established by
the time of skin formation. Retinoic acid (RA) treatment at E11 induces after 12 hr an inhibition of
cHoxd-13 expression in the plantar dermis, followed by the formation of feather filaments on the
reticulate scales. When E7.5 dorsal explants are treated with RA for 6 days, they form scale-like
structures where the Hox transcripts are no longer detectable. Protein analysis reveals that the plantar
filaments, made up of feather beta-keratins, correspond to a homeotic transformation, whereas the
scale-like structures, composed also of feather beta-keratins, are teratoid. These results strengthen
the hypothesis that different homeobox genes play a significant role in specifying the regional identity of
the different epidermal territories (Kanzler, 1997).
In Drosophila and mouse, Polycomb group genes are involved in the maintenance of homeotic gene
expression patterns throughout development. Skeletal phenotypes are found in mouse compound mutants
for two Polycomb group genes bmi1 (Drosophila homolog Sex combs reduced) and M33 (Drosophila homolog: Polycomb). Mice deficient for both bmi1 and M33 present
stronger homeotic transformations of the axial skeleton as compared to each single Polycomb group mutant,
indicating strong dosage interactions between those two genes. These skeletal transformations are
accompanied with an enhanced shift of the anterior limit of expression of several Hox genes in the somitic
mesoderm. These results demonstrate that in mice the Polycomb group genes act in synergy to control the
nested expression pattern of some Hox genes in somitic mesodermal tissues during development (Bel, 1998).
When Pc-G mutant mice are compared, loss of each Pc-G
gene shows a unique subset of affected Hox genes. For example, in M33 mutant mice, only anterior shifts for Hoxa3 can be detected and, in some cases, for Hoxc8. mel18 -/-, bmi1 -/- and rae28 -/- mice
present a more extensive overlap in affected Hox genes,
encompassing one prevertebrae anterior shifts of Hoxa5 and Hoxc8.
However, Hoxc6 and Hoxc5 are uniquely affected in bmi1 -/-
mice, while Hoxa7 and Hoxd4 are only affected in mel18 -/-
mice and Hoxb5 is unaffected in all those mutant mice. In M33 bmi1 double mutant mice, the
anterior limit of expression of at least two Hox genes, Hoxc9
and Hoxc8, is significantly more severely shifted as compared
to both single mutants, demonstrating the additive effect of
Pc-G products in maintaining the boundaries of selected Hox
genes. One striking observation is that deleting three gene doses
of these Pc-G genes does not increase the derepressive effect
of Hoxc8 or Hoxc9 expression; full deficiency of M33 and
bmi1 is required to induce extensive ectopic expression of
these two genes in mesodermal tissues. Nevertheless,
according to the differential expression of Hoxc8 and Hoxc9
in both single mutants, it seems that Hoxc8 is more sensitive
to M33 regulation since a one-segment anterior
shift is observed in M33 -/- bmi1 +/- (prevertebrae 10), whereas that shift is not visible
in bmi1 -/- M33 +/- (prevertebrae 11); reciprocally, Hoxc9 is more
sensitive to bmi1 regulation.
In contrast, some other Hox genes like
Hoxb1, Hoxd4 and Hoxd11 are not affected, either in single
or in double mutant mice. This suggests that several murine
multimeric Pc-G protein complexes of different composition
might exist that differ in their affinities for specific Hox
genes. Alternatively, since in mammals Pc-G genes exist as
highly related gene pairs (such as mel18/bmi1, Enx1/Enx2,
M33/MPc2, hPc1/hPc2 and Hph1/rae28/Hph2), a potential redundancy likely exists. This
suggests that the homologous gene complements part of the
function and thus maintains the boundaries of expression for
a subset of Hox genes. Analysis of double mutant mice for a
related pair such as mel18 and bmi1 should clarify the degree
of redundancy. Differential effects of Pc-G mutations on Hox
gene expression in different tissues like the somitic
mesoderm and the neural tube, also suggest that different Pc-G
complexes may regulate a specific subset of Hox genes in
a tissue-dependent manner (Bel, 1998).
Hoxb8 mutant mice were generated by inserting the lacZ coding sequence in frame with the first exon of Hoxb8. These mice express a
fusion protein with a functional beta-galactosidase activity instead of Hoxb8. Mutant embryos were analyzed for anatomical changes. The
results indicate that Hoxb8 is not an indispensable regulator of A-P patterning in the forelimb, unlike models suggested by Hoxb8 gain of function
experiments. The null mutant phenotypic traits include degeneration
of the second spinal ganglion (C2), an abnormality opposite to the alteration in the gain of function transgenic mice. Partial or complete degeneration of the most rostral definitive dorsal root ganglion, spinal ganglion C2, is
observed with full penetrance in Hoxb8 mutants, with an increase in severity
from heterozygous to homozygous mutants. Subtle changes in the
thoracic part of the vertebral column are observed as well. Adult homozygous mutants exhibit an abnormal clasping reflex of the limbs (van den Akker, 1999).
A detailed study is presented of the genetic basis of mesodermal axial patterning
by paralogous group 8 Hox genes in the mouse. The phenotype of Hoxd8
loss-of-function mutants is presented, and compared with that of Hoxb8- and
Hoxc8-null mice. This analysis of single mutants reveals common features for the Hoxc8 and Hoxd8 genes in patterning lower thoracic and lumbar vertebrae. In the Hoxb8 mutant, more anterior axial regions are affected. The three paralogous Hox genes are expressed up to similar rostral boundaries in the mesoderm, but at levels that strongly vary with the axial position. The axial region
affected in each of the single mutants mostly corresponds to the area with the
highest level of gene expression. However, analysis of double and triple mutants
reveals that lower expression of the other two paralogous genes also plays a
patterning role when the gene that is most highly expressed is defective. It is therefore
concluded that paralogous group 8 Hox genes are involved in patterning quite an
extensive anteroposterior (AP) axial region. Phenotypes of double and triple
mutants reveal that Hoxb8, Hoxc8 and Hoxd8 have redundant functions at upper thoracic and sacral levels, including positioning of the hindlimbs.
Interestingly, loss of functional Hoxb8 alleles partially rescues the phenotype of Hoxc8- and Hoxc8/Hoxd8-null mutants at lower thoracic and lumbar levels. This suggests that Hoxb8 affects patterning at these axial positions differently from the other paralogous gene products. It is concluded that paralogous Hox genes can have a unique role in patterning specific axial regions in addition to their redundant function at other AP levels (van Den Akker, 2001).
The regulatory interactions are described that cause anterior extension of the mouse 5' Hoxb expression domains from spinal cord levels to their definitive boundaries in the posterior hindbrain between embryonic day E10 and E11.5. This anterior expansion is retinoid dependent since it does not occur in mouse embryos deficient for the retinoic acid-synthesizing enzyme retinaldehyde dehydrogenase 2. A retinoic acid response element (RARE) was identified downstream of Hoxb5 and shown to be essential for expression of Hoxb5 and Hoxb8 reporter transgenes in the anterior neural tube. The spatio-temporal activity of this element overlaps with rostral extension of the expression domain of endogenous Hoxb5, Hoxb6 and Hoxb8 into the posterior hindbrain. The RARE and surrounding sequences are found at homologous positions in the human, mouse and zebrafish genome, which supports an evolutionarily conserved regulatory function (Oosterveen, 2003).
Fibroblast growth factor (Fgf) and retinoic acid (RA) signals control the
formation and anteroposterior patterning of posterior hindbrain. They are also
involved in development processes in other regions of the embryo. Therefore,
responsiveness to Fgf and RA signals must be controlled in a context-dependent
manner. Inhibiting the caudal-related genes cdx1a and
cdx4 in zebrafish embryos caused ectopic expression of genes that are
normally expressed in the posterior hindbrain and anterior spinal cord, and
ectopic formation of the hindbrain motor and commissure neurons in the
posteriormost neural tissue. Combinational marker analyses suggest
mirror-image duplication in the Cdx1a/4-defective embryos, and cell
transplantation analysis further revealed that Cdx1a and Cdx4 repress a
posterior hindbrain-specific gene expression cell-autonomously in the
posterior neural tissue. Expression of fgfs and retinaldehyde
dehydrogenase 2 suggested that in the Cdx1a/4-defective embryos, the Fgf
and RA signaling activities overlap in the posterior body and display opposing
gradients, compared with those in the hindbrain region. Fgf and
RA signals were required for ectopic expression. Expression of the posterior
hox genes hoxb7a, hoxa9a or hoxb9a, which function
downstream of Cdx1a/4, or activator fusion genes of hoxa9a or
hoxb9a (VP16-hoxa9a, VP16-hoxb9a) suppressed this
loss-of-function phenotype. These data suggest that Cdx suppresses the
posterior hindbrain fate through regulation of the posterior hox
genes; the posterior Hox proteins function as transcriptional activators and
indirectly repress the ectopic expression of the posterior hindbrain genes in
the posterior neural tissue. These results indicate that the Cdx-Hox code
modifies tissue competence to respond to Fgfs and RA in neural tissue (Shumizu, 2006).
The relocalisation of some genes to positions outside chromosome
territories, and the visible decondensation or unfolding of interphase
chromatin, are two striking facets of nuclear reorganisation linked to gene
activation that have been assumed to be related to each other. Here, in a
study of nuclear reorganisation around the Hoxd cluster, it is suggested
that this may not be the case. Despite its very different genomic environment
from Hoxb, Hoxd also loops out from its chromosome territory, and
unfolds, upon activation in differentiating embryonic stem (ES) cells and in
the tailbud of the embryo. However, looping out and decondensation are not
simply two different manifestations of the same underlying change in chromatin
structure. In the limb bud of the embryonic day 9.5 embryo,
where Hoxd is also activated, there is visible decondensation of
chromatin but no detectable movement of the region out from the chromosome
territory. During ES cell differentiation, decondensed alleles can also be
found inside of chromosome territories, and loci that have looped out of the
territories can appear to still be condensed. It is concluded that evolutionarily
conserved chromosome remodelling mechanisms, predating the duplication of
mammalian Hox loci, underlie Hox regulation along the rostrocaudal embryonic
axis. However, it is suggested that separate modes of regulation can modify
Hoxd chromatin in different ways in different developmental contexts (Morey, 2007).
To dissect the events of nuclear reorganisation, an ES cell differentiation system was used. Gene expression and nuclear reorganisation could be induced at Hoxb by
triggering the differentiation of murine ES cells with retinoic acid (RA). To determine whether similar activation occurs at
Hoxd RT-PCR was used to analyse the expression of Hoxd genes in undifferentiated OS25 ES cells, and during 18 days after the withdrawal of LIF and the addition of RA. As for Hoxb, there was no detectable expression of
Hoxd genes in undifferentiated cells. The extinction of Oct4
expression upon differentiation is accompanied by the rapid induction (by day
2) of Hoxd1 expression, but not of the more 5' genes
Hoxd3 through to Hoxd12. Expression of
Hoxd3 and Hoxd4 were detected by day 6, Hoxd8 by
day 8, Hoxd9 and Hoxd10 by day 10 and Hoxd12 expression was not detected until day 18. Hoxd1 expression declined at later stages of differentiation, but not as rapidly as seen for Hoxb1 (Morey, 2007).
Hoxd is flanked by structurally and functionally unrelated genes.
Expression of Mtx2, located 3' of Hoxd1, is induced by day 2, suggesting that this gene might also be subject to temporal colinearity. However, at the 5'
end of Hoxd, the early detection of Hoxd13 expression (by
day 2) suggests a break in the temporal colinearity at this end of the cluster
in this system. A large conserved noncoding region 5' of Hoxd, termed a global
control region (GCR), contains digit enhancers that act on Hoxd13,
Lnp and Evx2. Neural enhancers in the GCR also act on
Evx2 and Lnp. As Evx2 is also activated early in the
timecourse of differentiation, this suggests that the GCR may have some
activity in ES cells. Lnp expression in ES cells is constitutive. This
analysis shows that the colinear activation of the Hoxd cluster is
mostly recapitulated upon RA-induced ES cell differentiation (Morey, 2007).
The generation of distinct classes of motor neurons is an early step in the control of vertebrate motor behavior. To study the interactions that control the generation of motor neuron subclasses in the developing avian spinal cord in vivo grafting studies were performed in which either the neural tube or flanking mesoderm were displaced between thoracic and brachial levels. The positional identity of neural tube cells and motor neuron (MN) subtype identity was assessed by Hox and LIM homeodomain protein expression. Brachial (B) levels of the median motor column (MMC) are organized into three columns: neurons of the medial MMC (MMCM) co-express Isl1, Isl2 and Lim3, neurons of the medial lateral motor column (LMCM) co-express Isl1 and Isl2, and motoneurons of the lateral LMC (LMCL) coexpress Isl2 and Lim1. At thoracic (T) levels motoneurons are also organized into three columns: MMCM neurons; lateral MMC neurons that coexpress Isl1 and Isl2 but not Lim3, and dorsomedially positioned Column of Terni (CT) neurons that express only Isl1. Grafts of 13-15 segment quail T neural tube were placed rostrally at the B level of 12-15 segment chick hosts. Marker and morphological analysis reveals that grafted neural cells divert their normal T fates and their neuronal progeny acquire the molecular properties of B MNs. These changes in the neural tube are restricted to a limited time frame. The rostrocaudal identity of neural cells is plastic at the time of neural tube closure and is sensitive to positionally restricted signals from the paraxial mesoderm. Such paraxial mesodermal signals appear to control the rostrocaudal identity of neural tube cells and the columnar subtype identity of motor neurons. Analysis of neural Hoxc8 expression provides evidence that the change in cell identity after neural tube displacement is not restricted to the MNs; the change occurs in a graded manner along the rostrocaudal axos of the spinal cord, and is associated with both a rostral and caudal respecification in cell fate. In contrast, neural tube grafts between B and T levels do not change the pattern of Hoxc8 expression in the flanking paraxial mesodem.
These results suggest that the generation of motor neuron subtypes in the developing spinal cord involves the integration of distinct rostrocaudal and dorsoventral patterning signals that derive, respectively, from paraxial and axial mesodermal cell groups (Ensini, 1998).
Retinoic acid (RA) activity plays sequential roles during the development of the ventral spinal cord. The functions of local RA synthesis in the process of motoneuron specification and early differentiation have been investigated using a conditional knockout strategy that ablates the function of the retinaldehyde dehydrogenase 2 (Raldh2) synthesizing enzyme essentially in brachial motoneurons, and later in mesenchymal cells at the base of the forelimb. Mutant (Raldh2L–/–) embryos display an early embryonic loss of a subset of Lim1+ brachial motoneurons, a mispositioning of Islet1+ neurons and inappropriate axonal projections of one of the nerves innervating extensor limb muscles, which lead to an adult forepaw neuromuscular defect. The molecular basis of the Raldh2L–/– phenotype relies in part on the deregulation of Hoxc8, which in turn regulates the RA receptor RARß. Hoxc8 mutant mice, which exhibit a similar congenital forepaw defect, display at embryonic stages molecular defects that phenocopy the Raldh2L–/– motoneuron abnormalities. Thus, interdependent RA signaling and Hox gene functions are required for the specification of brachial motoneurons in the mouse (Vermot, 2005).
To address the molecular mechanisms underlying Polycomb group (PcG)-mediated repression of Hox gene expression, this study focused on the binding patterns of PcG gene products to the flanking regions of the Hoxb8 gene in expressing and non-expressing tissues. In parallel, the distribution of histone marks of transcriptionally active H3 acetylated on lysine 9 (H3-K9) and methylated on lysine 4 (H3-K4) was followed, and of transcriptionally inactive chromatin trimethylated on lysine 27 (H3-K27). Chromatin immunoprecipitation revealed that the association of PcG proteins, and H3-K9 acetylation and H3-K27 trimethylation around Hoxb8 were distinct in tissues expressing and not expressing the gene. Developmental changes of these epigenetic marks temporally coincide with the misexpression of Hox genes in PcG mutants. Functional analyses, using mutant alleles impairing the PcG class 2 component Rnf2 (Homolog of Drosophila Ring) or the Suz12 mutation decreasing H3-K27 trimethylation, revealed that interactions between class 1 and class 2 PcG complexes, mediated by trimethylated H3-K27, play decisive roles in the maintenance of Hox gene repression outside their expression domain. Within the expression domains, class 2 PcG complexes appeared to maintain the transcriptionally active status via profound regulation of H3-K9 acetylation. The present study indicates distinct roles for class 2 PcG complexes in transcriptionally repressed and active domains of Hoxb8 gene (Fujimura, 2006; full text of article).
The main outcome of this study was to show that binding of a specific,
Rnf2-containing form of the class 2 PcG complex, as well as H3-K27
trimethylation marking inactive chromatin, correlates with the maintenance of
transcriptional silencing of a Hox gene in developing embryos. Moreover, the
results demonstrated that genetic impairment of both PcG binding, and H3-K27
trimethylation leads to Hox gene derepression, and that H3-K27 trimethylation
is required for PcG binding. In addition, the establishment of
differential PcG binding and histone marks in expressing and non-expressing
embryonic tissues occur in the same developmental time window as when Hox
genes are deregulated in PcG mutants (Fujimura, 2006).
Rnf2 association to known regulatory elements of the Hoxb8 gene is
seen predominantly in transcriptionally silent anterior embryonic tissues,
whereas the binding of other PcG class 2 members, Phc1 and Cbx2, is observed
at all AP levels, irrespective of transcriptional status. This implies that
different forms of class 2 PcG complexes bind to the Hoxb genomic region in
embryonic domains where the gene is transcriptionally active and repressed.
This is reminiscent of previous findings in the Engrailed/Inv/GeneVI
complex in Drosophila SL-2 cells, where the Pc protein is exclusively
associated with transcriptionally silent genes, while Ph and Psc are present
irrespective of the transcriptional status.
Therefore the complete, 'perfect' form of the class 2 PcG core complex may
mediate transcriptional repression more efficiently than form(s) lacking the
Rnf2 component. If this is the case, incorporation of the Rnf2 component into
the complex might be a limiting process to mediate transcriptional repression
and regulate its stability. It is also possible that the role of Rnf2 is mediated
through its E3 ubiquitin ligase activity directed to histone H2A (Fujimura, 2006).
Transcriptional repression of Hox genes in the developing embryo has been
shown to correlate with the association of Rnf2-containing class 2 PcG
complexes and H3-K27 trimethylation. De-repression of Hox genes in
Rnf2 and Suz12 mutant cells reveal the requirement of both
Rnf2 association and H3-K27 trimethylation in the mediation of this
transcriptional repression. Since Rnf2 association to Hox genes is reduced in
Suz12 mutant ES cells and Rnf2 mutation alters Hox
expression without changing local levels of H3-K27 trimethylation, H3-K27
trimethylation mediated by class 1 PcG complexes at Hox genes may facilitate
subsequent binding of Rnf2-containing PcG complexes. Recruitment of
Rnf2-containing PcG complexes may in turn prevent the access of nucleosome
remodeling factors, such as SWI/SNF complex, leading to the formation of a
repressed chromatin status. Therefore, molecular circuitry underlying PcG silencing of
Hox genes seems to have been evolutionarily conserved between
Drosophila and mammals. It is also notable that Cbx2, a homologue of
Drosophila Pc, binds to Hoxb8 in transcriptionally active
embryonic tissues, despite the lack of histone H3 trimethylated at K27. This
is consistent with biochemical data that have shown the association
of purified or reconstituted PcG complexes with the nucleosomal templates
lacking histone tails. The implication of these findings is that there are at
least two different means by which class 2 PcG complexes bind to the
chromatin, and that the association, which involves trimethylated H3-K27,
mediates the repression at the Hox genes in vivo (Fujimura, 2006).
The maintenance of regionally restricted expression of Hox genes is likely
to involve H3-K9 acetylation and H3-K4 methylation. This study has shown that these modifications of the histone tail increases
craniocaudally along the axis. Although the transcriptionally active posterior
tissues of 9.5 dpc and older embryos are more heavily acetylated at H3-K9 than
the anterior, non-Hox expressing tissues, some acetylation of H3-K9 at
Hoxb8 is seen in anterior regions where Hoxb8 expression is
repressed at early and later developmental stages. De-repression of
Hoxb8 expression upon depletion of Rnf2 in MEFs derived from the
cranial part of 9.5 dpc embryos suggests the involvement of Rnf2-containing
class 2 PcG complexes to mediate this transcriptional repression. Therefore,
these data suggest that the associations of Rnf2-containing PcG complexes and
acetylated H3-K9 may counteract each other and cooperate to maintain the
anterior boundaries of Hoxb8 expression at mid-gestational stages and
later. This is consistent with the antagonistic properties of Mll and
Bmi1 mutations. Moreover, the establishment of the differential binding of
the Rnf2 and H3-K9 acetylation at Hoxb8 during embryogenesis
temporally coincides with de-repression of that Hox gene in
Bmi1/Rnf110 and Phc1/Phc2 double homozygotes, and loss of its transcription in Mll homozygotes. Intriguingly, class 2 PcG complexes, which lack the Rnf2 component, are also involved in the maintenance of H3-K9 acetylation in embryonic tissues where Hox genes are expressed. This is consistent with predominant subnuclear localization of several PcG proteins in the perichromatin compartment where
most pre-mRNA synthesis takes place. The molecular
mechanisms underlying this positive action remain unaddressed (Fujimura, 2006).
In conclusion, class 2 PcG gene products play distinct roles in embryonic
territories, which are silent or active for Hoxb8 transcription, by
forming complexes of different composition. Interaction between class 1 and class 2 PcG complexes mediated by trimethylated H3-K27 play decisive roles in Hox gene repression outside their expression domains, as seen in Drosophila. In addition, within the Hox expression domain, class 2 PcG complexes are involved in maintaining a transcriptionally active status, independent of H3-K27 trimethylation (Fujimura, 2006).
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
abdominal-A:
Biological Overview
| Transcriptional Regulation
| Targets of activity
| Protein Interactions
| Developmental Biology
| Effects of Mutation
| References
Society for Developmental Biology's Web server.
{kind=link}