by Lewis I. Held, Jr.
purchasing information
| Quirks of Human Anatomy by Lewis I. Held, Jr. | purchasing information |
| back to Quirks index page | |
Figure Legends 1 1.1 * 1.1R * 1.2 * 1.2R * 1.3 * 1.3R figure legends 2 * 3 * 4 * 5 * 6 * 7 * A N.B.: An 'R' suffix denotes reflections (commentaries, annotations, and further references) pertaining to the numbered legend that precedes it. [Select any image to enlarge; use back button to return] Fig. 1.1 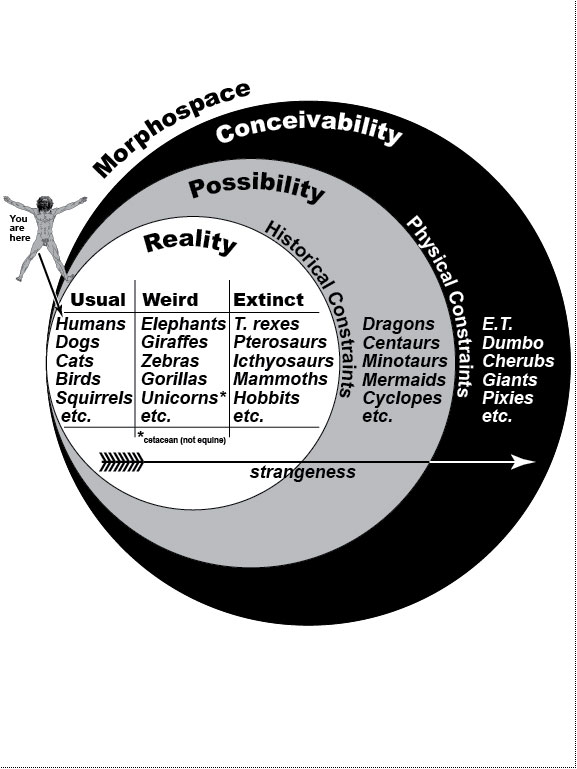 Real, possible, and conceivable subsets of vertebrate 'Morphospace' [959,2600] in Venn diagram format. Real, possible, and conceivable subsets of vertebrate 'Morphospace' [959,2600] in Venn diagram format.
The arrow points from the familiar to the fantastic. City dwellers routinely see only a tiny part of the animal world: pets, birds, and the occasional squirrel. As children, we first met exotic animals (elephants, giraffes, etc.) at the zoo or circus and extinct dinosaurs at the museum. The thrill we felt at the novelty of those beasts has faded, but we can still get a similar frisson when we see science-fiction monsters in movie theaters. Some of those fabulous creatures could have evolved if Earth’s history had unfolded differently [970], whereas others could not because they violate the laws of physics. For instance, centaurs could have evolved if the first fish to come on land had possessed three pairs of fins instead of two [823,959,2544], as some other groups of fish did at that time [2417]. Examples of conceivable, but impossible, animals include (1) Steven Spielberg’s E.T. (the Extra-Terrestrial), whose neck was too thin to support his cantilevered head [2305], and (2) Walt Disney’s Dumbo (the flying elephant), whose ears were too small (despite frantic flapping) to lift him into the air [49,1335]. The same is true for cherubs with their impotent wings. Mythical giants like Paul Bunyan could never stand because their proportionally scaled legs would not support their overly massive torso [955], nor could Disney’s pixies like Tinkerbell exist because their brains would be too tiny to afford intellect. On the other hand, hobbits (~1 m tall à la Tolkien) not only could have evolved but did [92,2611], at least on one small island [407,2212]. Unicorns also evolved, albeit in aquatic form as narwhals [298,795], and, as noted by Aristotle {PoA:3:2:663a23} [137], the Indian rhino is technically a unicorn as well, given its median nasal horn [2085]. Heavier- or lighter-gravity planets may have fostered a rich assortment of alien faunas [556,1049], which we may someday encounter. Sadly, our Moon is lifeless, and although we like to think of it as colonizable, we are ill suited to walking there [1764,2685]. Indeed, the Apollo astronauts resorted to hopping and skipping to get around [50,1812]. Fig. 1.1R Given how blasé we are today about the natural world, it is hard to imagine how amazed Europeans must have been when they first saw African wildlife in the 1700s [501] and dinosaur fossils in the 1800s [1472]. We take too much for granted in our anatomy (and behavior). One aim of this book is to rekindle our sense of wonder about life in general [594] and ourselves in particular. "The way we walk, for example, teetering on long, paired stilts of articulated bone, is unique among mammals, and as preposterous in its way as elephant trunks and platypus feet. We also communicate by tossing oddly intricate noises at one another, which somehow carry complex packages of feeling, thought, and information. We share and understand these sounds as if they were scents drifting on the wind, and our minds ... sniff the fragrance of their meaning" [2737]. During the Middle Ages, narwhal tusks were marketed as "unicorn horns" and sold for 10 times their weight in gold [1876]! Now, of course, we know better: they’re just teeth after all, so they sell nowadays for only $125/foot. But wait! If you think that narwhal tusks are any less mysterious, majestic, or magical just because we "know" what they really are, then you haven’t thought about (1) what it takes for them to develop or (2) what it took for them to evolve (cf. Fig. 2.4). You should read Richard Dawkins’s Unweaving the Rainbow [594] ... or just read on. This book was written as a kind of amusement park. Its thematic "pretend game" is to inspect each body part through the eyes of an alien visitor who asks, "Why is it this way and not that?" (cf. [2037]; his p. 523). Why, for instance, is there no Earthling that makes its skeleton out of metal, considering the ubiquity of metal ions and their use in other roles [2699]? In the face of such questions, the neophyte is on an equal footing with the expert. No one knows! Therein lies the fun of interrogating Nature. Asking the right questions is an art form unto itself, and some people are naturally gifted. Darwin was one of them. For an even wilder ride through Fantasyland, see Dougal Dixon’s Man After Man: An Anthropology of the Future (1990, St. Martin’s Press, New York). Fig. 1.2 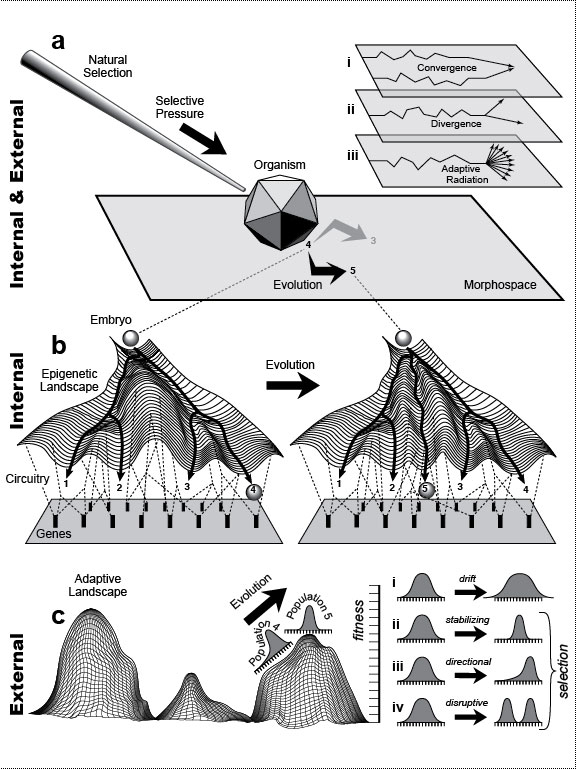 Metaphors for evo-devo that illustrate internal factors, external factors, or both. Modified from ref. [1135]. Metaphors for evo-devo that illustrate internal factors, external factors, or both. Modified from ref. [1135].a. Galton’s polyhedron analogy [960]. Francis Galton thought that species go from one stable anatomy to another when exposed to selective pressure [855]. He likened the stability to a polyhedron that stays poised on one facet unless pushed hard enough to topple it onto an adjacent facet. The organism would thus zigzag through its available "Morphospace" [2129]—the set of all possible shapes (cf. Fig. 1.1) [493,1702]. Insets (upper right) depict some trends in this context, where the y axis represents a one-dimensional Morphospace and the x axis is time. a.i. Convergence. Similar anatomies often evolve in separate lineages [1989,2821] because of physical constraints that allow few solutions for any given problem [1105,1176]. Similar behavioral traits can also evolve independently—for example, intelligence in humans and dolphins [1651]. a.ii. Divergence. Splitting of lineages is thought to occur mainly through geographic separation (i.e., allopatric speciation), where different niches demand different adaptations [1154,2007,2580]. Initially, the splitting may only entail subspecific morphological specializations (e.g., dog breeds), but the greater the differences become, the more likely they will lead to reproductive isolation (i.e., separate species) [531]. a.iii. Adaptive radiation [870,2209]. Species can ramify rapidly into multiple offshoots [1523,2265] when they colonize ecosystems full of vacant niches [2108,2304,2446]. Mounting evidence suggests that the stress of habitat colonization itself may reveal hitherto-untapped potential for wild variability, on which natural selection can then act [1180,2525,2790]. c. Wright’s adaptive landscape [869,2857]. In this imaginary example, height (scale at right) denotes fitness of genotypes (points in the mesh) [613]. If the environment changes (not shown), then contours would change accordingly. The three hills represent different ways to elevate fitness [593] (cf. [1304]). One interbreeding population is plotted as a bell curve, where the height of each point denotes the number of individuals having the genotype beneath it on the hill. The population is plotted before (Pop. 4) and after (Pop. 5) selection has exerted an effect, where 4 and 5 correspond to anatomies from the pathways shown in b. Panels at the right show how selection varies with topography [815]. c.i. On terrain that is relatively flat, there is negligible selection, so the span of genotypes can broaden via random mutation and recombination [118,738,1840]. In small populations, this "genetic drift" [1480,1606] can lead to fixation of neutral (or even deleterious) mutations [891,1224,2581,2632]. A case in point was the bottleneck ~5 MYA that led to fixation of our chromosome #2 from a harmless (but useless?) fusion of two ape chromosomes [1317]. c.ii. At an adaptive peak (hilltop) any deviations from the mean will be disfavored, thus narrowing ("stabilizing") the bell curve. c.iii. While a population is in the process of climbing a hill, individuals at the leading edge will leave more offspring, thus skewing the distribution ("directional" selection) [714]. Climbing can only occur by (1) mutational creation of new alleles or (2) recombination of old ones, both of which are random events. c.iv. If a population straddles a valley, then cohorts will be pulled apart ("disruptive" selection) [1702], and this divergence can lead to speciation [1407]. Fig. 1.2R Such metaphors give us a feeling for how genes control anatomy. For more on how "Genospace" maps onto "Phenospace," see articles by Lieberman and Hall [1545] and Weiss [2778]. Ultimately, it was our ricocheting through "Ecospace" that caused us to land at the point we now occupy in Morphospace [482,493]. a. In Morphospace, a species would actually occupy an area, not a point, and the size of that area would reflect its span of variation [653]. Indeed, as mentioned in the text, a better metaphor for a species (≈bell curves in c) might be an amoeba that moves over the terrain by extending and retracting pseudopodia. Species can retain their anatomy for eons [833,2750] (i.e., stay put in Morphospace), although structural stasis need not imply genetic stasis [1124]. Darwin called such species "living fossils" and inferred that they must have "inhabited a confined area and ... thus [were] exposed to less severe competition" [559] (p. 107). a.i. One clear example of convergence involves hooves. Any animal that runs en pointe (like a ballerina) risks injuring its toes, and the same protective devices evolved separately in even- and odd-toed mammals (e.g., pigs vs. horses)—namely, fused toes and thicker toenails ("hooves") [2085]. Convergence also arises when lineages fill similar niches [482,2007,2692]. Thus, marsupials evolved species that look eerily like the placental wolf, cat, mole, squirrel, and anteater [631,2153,2446,2861]. a.ii. A classic example of divergence is Darwin’s finches [1691,2770]. The molecular basis for finch beak divergence has finally been revealed by some recent evo-devo analyses [8]. a.iii. Explosive speciation can be sparked not only by access to uninhabited areas [811,2304] but also by the appearance of novel structures [237,1548,1890,2873]. For example, the debut of the neural crest created jaws that allowed a predatory (vs. filter-feeding) lifestyle [1624,1900] and hence spawned clades of carnivorous fish [472,796]. Radiation is aided by founder effects in small populations [1755] (e.g., colonizing of the Galapagos by Darwin’s finches [997,998]). Surprisingly, recent data refute the old cliché that mammals only radiated after dinosaurs disappeared [200,1597]. b. The uniformity of the depicted pegs may leave the impression that all genes are equal [2778]. Far from it! A tiny subset (the "toolkit") is most critical for building anatomy [392], and evolution results mainly from mutations therein [390]. Fig. 1.3 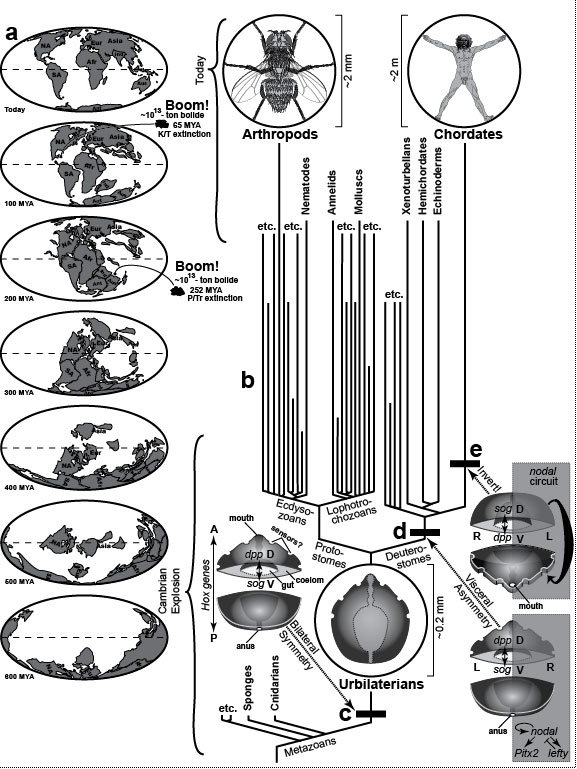 Overview of bilaterian evolution. The intent of this diagram is to convey the vast amount of time that has been available for evolution to reshape the anatomies of metazoan phyla. In particular, the Hox genes that form the scaffolding for all bilaterian bodies (c) are >20 times older than the Himalayan Alps [604,2023]. Humans and flies shared a multicellular ancestor who lived more than half a billion years ago. Overview of bilaterian evolution. The intent of this diagram is to convey the vast amount of time that has been available for evolution to reshape the anatomies of metazoan phyla. In particular, the Hox genes that form the scaffolding for all bilaterian bodies (c) are >20 times older than the Himalayan Alps [604,2023]. Humans and flies shared a multicellular ancestor who lived more than half a billion years ago.
a. Timeline spanning one seventh of Earth’s 4.55-billion-year history [620]. Modern continents or their pieces are traced back millions of years ago (MYA). The "Cambrian Explosion" [506] was an abrupt radiation of body plans ~543 MYA [1297,2023,2210,2663]. Back then, the continents looked nothing like they do today, and days were shorter (21 hours) because Earth spun faster (425 days per year) [2122]. b. Pedigree of metazoans, simplified from refs. [25,1048,1189]. Arthropods and chordates are singled out but are only 2 of ~37 crown phyla [2663]. (There are just 4 extant deuterostome phyla [252,2536].) Lines that end before today denote extinctions but are only meant to give a general impression. Black bars mark key events. c. Urbilaterians are the ancestors of all phyla that have a medial plane of symmetry, a two-ended gut, and a coelom [195,886,2663]. Cross-section shows the A-P (anterior-posterior) axis [1189,1193] where Hox genes specify an "x" coordinate [1519,2316], and the D-V (dorsal-ventral) axis where the genes decapentaplegic (dpp; a.k.a. BMP4) and short gastrulation (sog; a.k.a. chordin) specify a "y" coordinate [39,1246,1779,2890]. Urbilaterians are depicted with the D-V polarity of protostomes for reasons explained in e. The tick-shaped animal (circled) belongs to the oldest known bilaterian species, Vernanimalcula guizhouena (580-600 MYA) [431], although it may not be our direct ancestor [2493,2675], which was probably more wormlike [119,1462]. Note its tiny size versus the fly and man. It has 3 pairs of concave pits (sense organs? [1190,1267,2301]), but no sign of segments [443,1226] or appendages [1761,2092,2096], which apparently evolved later in several descendant clades [195,604,1048]. The gut (dotted outline) is straight without even a hint of the left-right asymmetry seen in larger metazoans (cf. Fig. 2.1). The ventral mouth leads to a thin pharynx, a wide stomach, a narrow rectum, and a posterior anus. d. All deuterostomes thus far examined share asymmetric expression of the gene nodal (see Ch. 2), which marks left versus right through the circuit shown (or parts thereof) [243,681], although the trigger upstream of nodal varies among classes of vertebrates [108,1528]. Arrows indicate activation; T-bars inhibition. Basal deuterostomes are drawn with this circuit on the right for reasons explained in e. How asymmetry arose in protostomes is unclear [528,1156,1216,2255,2429], nor is it known how often they recruited different signaling pathways for different asymmetric organs [13,1625,2430,2571]. e. An inversion of the bilaterian body must have occurred here because (1) chordates are upside-down versions of annelids and arthropods with regard to the dpp-sog axis [86,606,1457,1778], (2) the D-V polarity of hemichordates resembles arthropods rather than chordates [886,1583], and (3) the nodal circuit is on the right side in sea urchins but the left side in chordates [680,681,2367]. Our protochordate progenitor must have flipped over like a swimmer switching from breaststroke to backstroke at the end of the pool [629,2579]. What had been its belly became its back and what had been its left became its right [778,977]. The cross-section shows the sog-dpp axis in chordate orientation and the nodal circuit (shaded rectangle) on what has become our left side. Thus, the animals diagrammed in d and e are facing nose-to-nose. Fig. 1.3R a. Global extinctions at the K/T (Cretaceous/Tertiary) [62,248,2322] and P/Tr (Permian/Triassic) boundaries [162,730] likely occurred as a consequence of bolide impacts (meteors or comets) [620], although other theories exist [1485]. Not shown here is the really bad day (~4500 MYA) when a Mars-sized object hit Earth so hard that we belched a molten bolus that became our moon [363,2619]. The moon’s pockmarked face records the subsequent shower of smaller bolides that finally abated ~3800 MYA [1075]. Only then could life gain a foothold. For a pithy and poetic survey of evolution—both geological and biological—see Nigel Calder’s Timescale [341]. Continents are redrawn from Scotese [2326] except that his artifactual gap along the Rocky Mountains is omitted [1834]. Dates, weights, and impact sites of bolides (asteroids, comets, or meteors) are from ref. [620]. c. Hox control of the anterior-posterior axis may actually predate cnidarians [646,764,2247]. d. Echinoderms somehow evolved radial, 5-fold symmetry as adults [2415], but they retain bilateral symmetry as larvae [2536]. Strangest of all, they metamorphose by an asymmetric process as bizarre as having a wart on your left cheek that gradually expands to become an umbrella [681,1766]. e. One abiding mystery of the dorsal-ventral axis inversion is how the chordate mouth later migrated down the head to a ventral site [456,1463,1464]. If we can figure out this riddle, the answer might shed light on why our pituitary splits apart and rejoins [1348,2192] and why it evolved with our oral region in the first place (cf. Fig. 3.1) [356,1191,1637]. top of page
Lewis I. Held, Jr. is Associate Professor in the Department of Biology at Texas Tech University.
|
 © 2009 Thomas B. Brody, Ph.D. |