Select image to enlarge
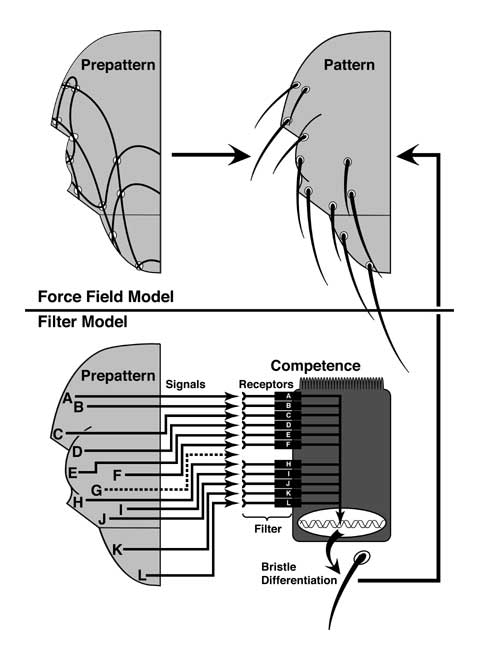
Figure 3.2
Curt Stern's Prepattern Hypothesis in its original (above) and final (below) forms, both of which were proposed in 1954 [4095, 4096].
Force Field Model. Initially, Stern envisioned patterns as the steady-state outcomes of dynamic physical or chemical forces. In this illustration, uneven growth deforms the epithelium so that each half mesonotum acquires 7 stress lines. Each of the 11 sites ('singularities') where the lines intersect then induces a macrochaete (MC). Because mutations can only change the pattern by altering the force vectors, patches of mutant tissue in a wild-type background should distort the entire web of forces. Contrary to expectation, virtually every pattern-altering mutation Stern studied (including achaete1, Fig. 3.3c) affected differentiation autonomously in mosaics.
Filter Model. To explain why cells exhibit such insularity, Stern postulated that (1) each MC must use a different signal (A-L, though signals could differ quantitatively instead) and (2) a MC will only form at a site if the cell there is 'competent' to respond to that specific signal. Inserted between the prepattern and the pattern is an extra step wherein each cell 'filters out' signals that it cannot 'hear'. In this cartoon, the competence of each cell (depicted with apical microvilli) depends on surface receptors. In reality, positional signals are 'filtered' through enhancer elements within the achaete-scute Complex (Fig. 3.4) [2891]. Wild-type cells (like this one) cannot respond to Signal G, a 'cryptic singularity' that Stern discovered [4097]. Similarly, ac1 cells would be 'deaf' to Signal I (not shown).
N.B.: Stern described these variations of his basic theme in general terms, but never formalized or named them as specific models. Here they are made explicit to highlight their differences. The idea that tensions affect gene expression (top model) seemed farfetched at the time, but various examples are now known [301, 761, 762, 1942, 2580].
|
|
{kind=link}
{kind=link}