Select image to enlarge
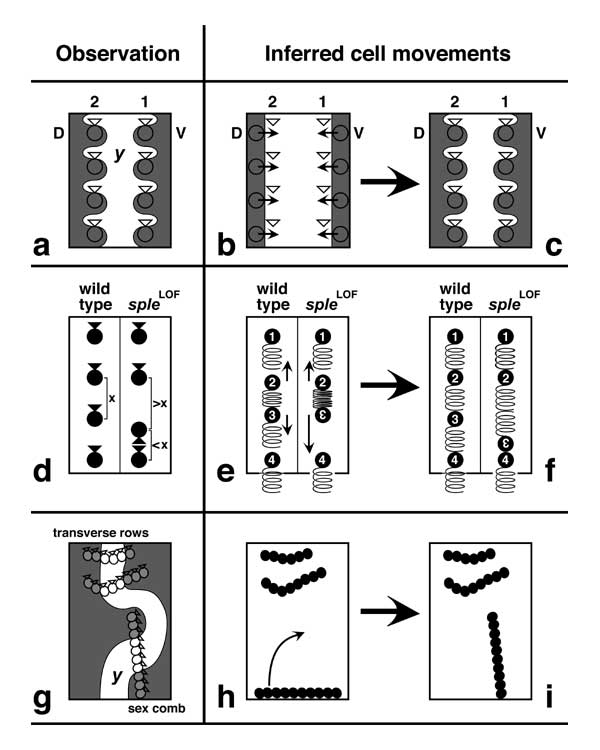
Figure 3.11
Movements of bristle cells inferred from various studies. Bristles are denoted by circles and bracts by triangles. Three types of hypothetical movements are shown -- none of which have actually been witnessed yet.
a-c. Lateral shifts that may align SOPs in longitudinal rows [1800]. a. Sections of rows 1 and 2 from a left leg (D, dorsal; V, ventral). Wavy lines indicate the kinds of boundaries seen in gynandromorphs. Male tissue is marked with yellowLOF (y). Curiously, bristles within a row tend to be more closely related to each other than to their own (physically closer) bracts. These affiliations are explicable if SOPs in each proneural stripe (Fig. 3.9) migrate in one direction (dorsally for row 1, ventrally for row 2) to a different column of cells where they induce neighbors to become bracts (b, c) [1800, 1808]. Shaded zones in b and c denote y+ tissue as in a (they are not Ac stripes), and triangles in b mark future bracts.
d-f. Proximal-distal shifts that may fine-tune SOP intervals within those rows [1803, 1810, 1813]. d. LOF mutations of spiny legs (sple) cause bristles to be misoriented along with their bracts -- usually a 180° polarity reversal (third bristle). Strangely, intervals between bristles that face one another tend to be larger (>x) than wild-type (x), while spaces between bristles that away from each other tend to be smaller (<x). These correlations are explicable if SOPs repel one another via filopodia (distally pointing springs) [1809, 1810]. If SOPs #2 and 3 happen to be unusually close, then they would be pushed apart by the cramped filopodia of SOP #2. In wild-type flies, such repulsions would make spaces more uniform (left half of e, f), but reversed-polarity SOPs (#3) should feel more force from the proximal neighbor (due to 2 intervening springs) than from the distal one (0 springs), resulting in the observed correlations (right half of e, f). Filopodia have indeed been seen on bristle SOPs [2387, 3630], and they are longer in one direction. Similar filopodia appear to mediate rearrangements of scale SOPs in moth wings [3051]. (Lepidopteran scales are thought to be homologous to dipteran bristles [1340, 2173, 2422, 4640, 4641].)
g-i. Sex comb rotation [4344]. g. Clones that pass through the centers of the transverse rows (two shown) often include the center of the sex comb, despite its perpendicular orientation. This zigzag is explicable if the sex comb arises as a transverse row and rotates 90° (h, i) [4344]. Conceivably, movements of this sort might also be guided by filopodia: apical 'cytonemes' have been seen in leg and wing discs [3507], and basal 'epidermal feet' [2175, 2585-2588] have been seen in leg, wing, and eye discs [1133, 2863, 3426] and on leg and wing disc cells in culture [3303]. N.B: The boundary in g is not actually traceable in such smooth detail since yLOF does not affect the background cuticle, and the same is true for a.
|
|
{kind=link}