The Interactive Fly
Genes involved in tissue and organ development
The tracheaee constitute the respiratory system of the fly. Made up of tubules that ramify throughout the body, trachea terminate in enlargements called air sacs, through which gas exchange takes place. Spiracles are the external tracheal apertures, repeated segmentally on either side of the thorax and abdomen.
The embryonic tracheal system is an epithelial tubular network established from defined sets of ectodermal precursor cells. At stage 10 of embryonic development, lateral ectodermal clusters of cells on both sides of the 10 posterior (thoracic 2 and 3 and abdominal 1 through 8) parasegments assume a tracheal placode fate [Images]. These cells, about 80 in each placode invaginate at stage 11 to form tracheal pits. The subsequent formation of the tracheal tree occurs without further cell division. First the cells migrate in a distinct pattern, then they fuse with other tracheal cells from adjacent segments to form a continuous tubular network. Terminal tracheal cells send long extentions toward the target cells. (Wilk, 1996).
Adaptation to diverse habitats has prompted the development of distinct organs in different animals to better exploit their living conditions. This is the case for the respiratory organs of arthropods, ranging from tracheae in terrestrial insects to gills in aquatic crustaceans. Although Drosophila tracheal development has been studied extensively, the origin of the tracheal system has been a long-standing mystery. Tracheal placodes and leg primordia arise from a common pool of cells in Drosophila, with differences in their fate controlled by the activation state of the wingless signalling pathway. Early events that trigger leg specification have been elucidated and it is shown that cryptic appendage primordia are associated with the tracheal placodes even in abdominal segments. The association between tracheal and appendage primordia in Drosophila is reminiscent of the association between gills and appendages in crustaceans. This similarity is strengthened by the finding that homologues of tracheal inducer genes are specifically expressed in the gills of crustaceans. It is concluded that crustacean gills and insect tracheae share a number of features that raise the possibility of an evolutionary relationship between these structures. An evolutionary scenario is proposed that accommodates the available data (Franch-Marro, 2006).
The Drosophila tracheal system has a clearly metameric origin, arising from clusters of cells, on either side of each thoracic and abdominal segment, that express the tracheal inducer genes trachealess (trh) and ventral veinless (vvl). Conversely, the leg precursors can be recognized as clusters of cells that express the Distal-less (Dll) gene, on either side of each thoracic segment; these will give rise both to the Keilin's Organs (KOs, the rudimentary legs of the larvae) and to the three pairs of imaginal discs that will give rise to the legs of the adult fly (Franch-Marro, 2006).
To investigate whether there is a direct physical association between the leg and tracheal primordia, Drosophila embryos co-stained for the expression of trh and early markers of leg primordia were examined. Although Dll is one of the most commonly used markers for the leg primordia, it is not the earliest gene required for their specification. Instead, a couple of related and apparently redundant genes, buttonhead (btd) and Sp1, act upstream of Dll in the specification of these primordia (Estella, 2003). Examining the specification of tracheal cells with respect to btd expression, tracheal cells were observed to appear in close apposition to btd-expressing cells, from the earliest stages of their appearance (by stage 9/early stage 10). Interestingly, unlike Dll, btd is initially expressed both in the thoracic and abdominal segments, and its expression is restricted to the thoracic segments later, under the influence of the BX-C. Thus, the cells of the respiratory system in Drosophila always arise in close proximity to the cells that are fated to give rise to the legs (Franch-Marro, 2006).
To fully endorse this conclusion it is necessary to show that the btd-expressing cells in the abdomen correspond to cryptic leg primordia. This may be a key point because, although many of the genes required for leg development are already known, it has not yet been possible to induce leg development in abdominal segments (except by transforming these segments into thoracic ones). In particular, although the Dll promoter contains BX-C binding sites that repress its expression in the abdominal segments, no ectopic appendage has been reported by misexpressing Dll in the abdomen. These observations have lead to some doubts as to whether a leg developmental program is at all compatible with abdominal segmental identity (Franch-Marro, 2006).
Since the initial expression of btd in the abdominal segments is downregulated by the BX-C genes, it was reasoned that sustained expression of btd might overcome the repressive effect of the BX-C genes and force the induction of leg structures in the abdomen. To test this, a btd-GAL4 driver was used to drive btd expression, expecting that the perdurance of the GAL4/UAS system would ensure a more persistent expression of btd in its endogenous expression domain. No sign was ever obtained of ectopic Dll expression or KOs in the abdominal segments, but the increased expression of btd had an effect on the KOs of the thoracic segments, which had more sensory hairs than the three normally found in wild-type KOs. Thus, on its own, btd seems unable to overcome BX-C repression of leg development (Franch-Marro, 2006).
One possibility would be that the BX-C genes could suppress appendage development in the abdomen by independently repressing both btd and Dll in this region. To assess this possibility, the same btd-GAL4 driver was used to simultaneously induce the expression of both btd and Dll. Under these circumstances, it was observed that KOs develop in otherwise normal abdominal segments; as in the previous experiment, the newly formed KOs have more than three sensory hairs. These results suggest that expression of btd and Dll in the btd-expressing abdominal primordia is sufficient to induce the development of leg structures in the abdomen, overcoming the repressive effect of the BX-C genes. Furthermore, these results demonstrate that these clusters of btd-expressing cells in the abdomen are indeed cryptic leg primordia. These results clearly show that tracheal cells are specified in close proximity to the leg primordia, in both thoracic and abdominal segments (Franch-Marro, 2006).
Previous results have shown that the leg primordia are specified straddling the segmental stripes of wingless (wg) expression in the early embryonic ectoderm, whereas tracheal cells are specified in between these stripes. To investigate whether wg might play a role in determining the fate of these primordia, what happens when the normal pattern of wg expression is disrupted was studied. In wg mutant embryos, trh and vvl from the earliest stages of their expression are no longer restricted to separate clusters of cells; instead larger patches of expression add up to a continuous band of cells running along the anteroposterior axis of the embryo, while btd expression is suppressed in this part of the embryonic ectoderm. Conversely, ubiquitous expression of wg suppresses trh expression, while causing an expansion of btd expression along the embryo. Restricted activation or inactivation of the wg pathway by the expression of a constitutive form of armadillo or a dominant-negative form of dTCF, respectively, are also able to specifically induce or repress trh and btd expression. trh/vvl and btd seem to respond independently to wg signalling and there is no sign of cross-regulation among them, since btd expression is normal in trh vvl double mutants, and trh and vvl expression is normal in mutants for a deficiency uncovering btd and Sp1 (Franch-Marro, 2006).
The role of wg as a repressor of the tracheal fate is further illustrated by looking at the behaviour of transformed cells: the clusters of cells that have lost btd expression and gained trh and vvl expression in wg mutant embryos begin a process of invagination that is characteristic of tracheal cells. Furthermore, these cells also express the dof (stumps) gene, a target gene of both trh and vvl in the tracheal cells. Although further development of these cells is hard to ascertain because of gross abnormalities in wg- embryos, these results indicate that they have been specified as tracheal cells. Thus, wg appears to act as a genetic switch that decides between two mutually exclusive fates in this part of the embryonic ectoderm: the tracheal fate, which is followed in the absence of wg signalling; and the leg fate, which is followed upon activation of the wg pathway. Given that there are no cell lineage restrictions setting apart the cells of the tracheal and leg primordia, these two cell populations could be considered as a single equivalence group, with the differences in their fate controlled by the activation state of the wg signalling pathway (Franch-Marro, 2006).
A link between respiratory organs and appendages is also found in many primitively aquatic arthropods, like crustaceans, where gills typically develop as distinct dorsal branches (or lobes) of appendages called epipods. Following the current observations, which suggest a link between respiratory organs and appendages in Drosophila, whether further similarities could be found between insect tracheal cells and crustacean gills was examined. Specifically, whether homologues of the tracheal inducing genes might have a role in the development of appendage-associated gills in crustaceans was considered (Franch-Marro, 2006).
RT-PCR was used to clone fragments of the vvl and trh homologues from Artemia franciscana and from Parhyale hawaiensis, representing two major divergent groups of crustaceans (members of the branchiopod and malacostracan crustaceans, respectively). In the case of Artemia vvl, a fragment was cloned that corresponds to the APH-1 gene and an antibody was generated for immunochemical staining in developing Artemia larvae. It was observed that Artemia Vvl is initially absent from early limb buds; it becomes weakly and uniformly expressed while the limb is developing its characteristic branching morphology, and becomes strongly upregulated in one of the epipods as its cells begin to differentiate. Uniform weak expression persists in mature limbs, but expression levels in the epipod are always significantly higher. Expression of the trh homologue from Artemia appears to be restricted to the same epipod as Vvl. Similarly, homologues of vvl and trh were cloned from Parhyale hawaiensis and their expression was studied by in situ hybridization. Both genes are specifically expressed in the epipods of developing thoracic appendages. Besides epipods, the Artemia trh and vvl homologues are also expressed in the larval salt gland, an organ with osmoregulatory functions during early larval stages of Artemia development (Franch-Marro, 2006).
What is the significance of the two Drosophila tracheal inducer genes being specifically expressed in crustacean epipods/gills? One possibility is that the expression of these two genes was acquired independently in insect tracheae and in crustacean gills. Alternatively, tracheal systems and gills may have inherited these expression patterns from a common evolutionary precursor, perhaps a respiratory/osmoregulatory structure that was already present in the common ancestors of crustaceans and insects (Franch-Marro, 2006).
The latter possibility is considered unlikely by conventional views, because of the structural differences between gills and tracheae (external versus internal organs, discrete segmental organs versus fused network of tubes), and the difficulty to conceive a smooth transition between these structures. Yet, analogous transformations have occurred during arthropod evolution: tracheae can be organized as large interconnected networks or as isolated entities in each segment (as in some apterygote insects), invagination of external respiratory structures is well documented among groups that have made the transition from aquatic to terrestrial environments (terrestrial crustaceans, spiders and scorpions), and conversely evagination of respiratory surfaces is common in animals that have returned to an aquatic environment (tracheal gills or blood gills in aquatic insect larvae). A very similar (but independent) evolutionary transition is, in fact, thought to have occurred in arachnids, where gills have been internalised to give rise to book lungs, and these in turn have been modified to give rise to tracheae in some groups of spiders. Thus, a relationship between insect tracheae and crustacean gills is plausible (Franch-Marro, 2006).
A particular type of epipod/gill has also been proposed as the origin of insect wings, a hypothesis that has received support from the specific expression in a crustacean epipod of the pdm/nubbin (nub) and apterous (ap) genes - that have wing-specific functions in Drosophila. In fact, the Artemia nub and ap homologues are expressed in the same epipod as trh and vvl, raising questions as to the specific relationship of this epipod with either tracheae or wings. A resolution to this conundrum becomes apparent when one considers the different types of epipods/gills found in aquatic arthropods, and their relative positions with respect to other parts of the appendage (Franch-Marro, 2006).
The primary branches of arthropod appendages, the endopod/leg and exopod, develop straddling the anteroposterior (AP) compartment boundary, which corresponds to a widely conserved patterning landmark in all arthropods. Different types of epipods/gills, however, differ in their position with respect to this boundary. For example, in the thoracic appendages of the crayfish, some epipods develop spanning the AP boundary [visualized by engrailed (en) expression running across the epipod], whereas others develop exclusively from anterior cells (with no en expression). Given that wing primordia comprise cells from both the anterior and posterior compartments, wings probably derived from structures that were straddling the AP boundary. Conversely, given that tracheal primordia arise exclusively from cells of the anterior compartment (anterior to en and even wg-expressing cells), it seems probable that tracheal cells evolved from a population of cells that was located in the anterior compartment. In this respect, it is interesting to note that the former type of epipods express nub, whereas the latter do not (Franch-Marro, 2006).
In summary, it is suggested that the ancestors of arthropods had specific areas on the surface of their body that were specialized for osmoregulation and gas exchange. Homologues of trh and vvl were probably expressed in all of these cells and played a role in their specification, differentiation or function. Some of these structures were probably associated with appendages, in the form of epipods/gills or other types of respiratory surfaces. A particular type of gill, straddling the AP compartment boundary, is likely to have given rise to wings, whereas respiratory surfaces arising from anterior cells only may have given rise to the tracheal system of insects. Confirmation of this hypothetical scenario may ultimately come from the discovery of new fossils, capturing intermediate states in the transition of insects from an aquatic to a terrestrial lifestyle (Franch-Marro, 2006).
Mitotic cells assume a spherical shape by increasing their surface tension and osmotic pressure by extensively reorganizing their interphase actin cytoskeleton into a cortical meshwork and their microtubules into the mitotic spindle. Mitotic entry is known to interfere with tissue morphogenetic events that require cell-shape changes controlled by the interphase cytoskeleton, such as apical constriction. However, this study shows that mitosis plays an active role in the epithelial invagination of the Drosophila tracheal placode. Invagination begins with a slow phase under the control of epidermal growth factor receptor (EGFR) signalling; in this process, the central apically constricted cells, which are surrounded by intercalating cells, form a shallow pit. This slow phase is followed by a fast phase, in which the pit is rapidly depressed, accompanied by mitotic entry, which leads to the internalization of all the cells in the placode. It was found that mitotic cell rounding, but not cell division, of the central cells in the placode is required to accelerate invagination, in conjunction with EGFR-induced myosin II contractility in the surrounding cells. It is proposed that mitotic cell rounding causes the epithelium to buckle under pressure and acts as a switch for morphogenetic transition at the appropriate time (Kondo, 2013).
The invagination of epithelial placodes converts flat sheets into the three-dimensional structures that form complex organs, and it is a key morphogenetic process in animal development. A major mechanism of invagination is apical constriction, which is driven by actomyosin contraction. However, not all constricted cells invaginate, and some cell internalization occurs without apical constriction, suggesting that additional mechanisms of inward cell movement contribute to invagination (Kondo, 2013).
To obtain three-dimensional information about cell behaviour during invagination, live imaging was performed of the Drosophila tracheal placode. Ten pairs of tracheal placodes, each of which is composed of about 40 cells, are formed in the ectoderm at mid-embryogenesis, and each placode initiates invagination simultaneously. Using an adherens junction marker, DE-cadherin-green fluorescent protein (E-cad-GFP), it was found that the adherens junctions of the central placode cells slowly created a depression by apical constriction, which became the tracheal pit. After 30 to 60 min of slow movement (slow phase), the tracheal pit was suddenly enlarged, and the tracheal cells were rapidly internalized (fast phase) and eventually formed L-shaped tube structures (Kondo, 2013).
After the fast transition, all the tracheal cells and surrounding epidermal cells entered mitosis 16, the final round of embryonic mitosis. It was noticed that the fast invagination was always associated with the mitotic entry of central cells that were frequently the first to enter mitosis 16. Intriguingly, mitotic rounding of the central constricted cells occurred simultaneously with the rapid depression of their apices, followed by chromosome condensation 10 min later. In this study, this atypical mitotic rounding associated with apical depression in an internalized cell is called 'rounding', to distinguish it from canonical surface mitosis (surface cell rounding) (Kondo, 2013).
To determine whether cell rounding is required for invagination, zygotic mutants were examined of the cell-cycle gene Cyclin A (CycA), which fail to enter mitosis 16, and double parkeda3 (dupa3), which show a prolonged S phase 16 and delayed entry into mitosis 16. Tracheal invagination was initiated normally in the CycA and dupa3 mutants, but proceeded more slowly than in controls, indicating that entry into mitosis 16 is required for proper timing of the fast phase (Kondo, 2013).
Although delayed, the accelerated invagination in the CycA or dupa3 mutants eventually occurred, allowing the formation of tube structures and suggesting that additional mechanisms are involved. After invagination, fibroblast growth factor (FGF) signalling is activated in the tracheal cells to induce branching morphogenesis through chemotaxis. To examine the contribution of FGF signalling to invagination, mutants of the FGF ligand branchless (bnl) or the FGF receptor breathless (btl) were analyzed. These mutants invaginated normally, indicating that chemoattraction to FGF is dispensable for invagination (Kondo, 2013).
Next, to assess FGF's role in the mitosis-defective condition, double mutants were analyzed for CycA and bnl or CycA and btl, and it was found that they showed slower invagination than CycA single mutants. Furthermore, the invagination in these double mutants was incomplete, in that the cells failed to form L-shaped tubular structures. Therefore, FGF signalling is critical for invagination when mitosis is blocked, serving a back-up role. Tracheal-specific CycA expression rescued the defects in invagination speed and tube structure in the CycA btl mutants. In addition, mitosis of cells outside the pit was occasionally observed that occurred before the mitosis of the central apically constricted cells and was not correlated with the fast invagination phase. Thus, mitosis of the surrounding epidermal cells is dispensable for tracheal invagination. Taken together, it is concluded that mitotic entry of central cells is a major mechanism for accelerating tracheal invagination (Kondo, 2013).
To distinguish the role of cell rounding from that of cell division in the fast phase, the microtubule inhibitor colchicine was used to arrest the cell cycle after cell rounding. Colchicine treatment after mitosis 15 induced M-phase arrest at mitosis 16, but the fast invagination movement accompanied by cell rounding was not affected. This result indicates that cell rounding, but not cell division, is responsible for the acceleration phase of the tracheal invagination (Kondo, 2013).
Mitosis of cells in the columnar epithelium normally occurs at the apical surface after surface rounding. It was next asked how the apical surface of the central cells becomes depressed during internalized cell rounding. One possible model explains internalized cell rounding as cell-autonomously controlled by the association of the cells with the basement membrane or underlying mesodermal cells. However, genetic removal of basement-membrane adhesion by the maternal and zygotic mutation of βPS-integrin (also known as mys) did not compromise the speed of invagination, and snail-twist double-mutant embryos, which lack mesodermal cells, still showed tracheal invagination with internalized cell rounding. These results suggest that anchoring to the basal side is probably not required (Kondo, 2013).
A second model proposes that the apical depression of the rounding cells is driven by local planar interactions among the tracheal cells. Before and during tracheal invagination, myosin II is enriched at the cell boundaries tangential to the centre of the placode and regulates cell intercalation. It was noted that the myosin II level in the central cells was lower than in the surrounding, intercalating cells. Nevertheless, the apices of the central cells were constricted during the slow phase, strongly suggesting that the surrounding cells exerted centripetal pressure on the central cells through myosin II cables. Myosin II cables fail to form in EGFR signalling mutants (such as rho, the rhomboid endopeptidase required for EGF ligand maturation, and Egfr), and apical constriction is impaired in these mutants. The first few cells undergoing mitosis 16 in the tracheal placode of rho or Egfr mutants showed surface cell rounding with expanded apices, indicating that EGFR signalling is required to couple the mitotic cell rounding with fast apical depression. It is speculated that the columnar shape of the central cells resists centripetal movements, resulting in the accumulation of inward pressure during the slow phase. The existence of such resistance was supported by the results of a physical perturbation experiment using a pulsed ultraviolet lase. The cell rounding associated with mitotic entry would release the stored inward pressure by means of cytoskeletal remodelling that causes rapid depression of apical surface together with the active shortening of cell height, leading to rapid buckling of the apical surface and the fast phase of invagination (Kondo, 2013).
Even with the loss of both EGFR and FGF signalling, the tracheal placodes form moderately invaginated structures, compared to the flat tracheal placode observed in the rho-bnl-CycA triple mutant at the same stage, indicating that cells needed to undergo mitosis 16 to induce invagination, independent of EGFR and FGF signalling. In rho bnl double mutants, although the cells undergoing the earliest mitoses showed surface cell rounding, some of the subsequent mitotic events were coupled to apical depression and internalized cell rounding. Unlike the earlier mitotic events on the surface, the internalized rounding cells in the rho bnl embryos showed constricted apices and were surrounded by apically rounded cells before mitosis. Internalized rounding with a constricted apical surface were shared properties of cells in mitoses leading to invagination, in both control and rho bnl embryos. It is suggested that the first few cells undergoing surface cell rounding compress the adjacent interphase cells and restrict their apical area, so that they are forced to move internally after rounding, causing the epithelial layer to buckle and invaginate (Kondo, 2013).
Although invagination was largely blocked in the rho-bnl-CycA triple mutants, any double mutant combination permitted invagination to some degree, indicating that three qualitatively distinct mechanisms, mitotic cell rounding, myosin II contractility (EGFR) and active cell motility (FGFR), can independently trigger invagination. In the normal context of wild-type development the combination of cell rounding and EGFR signalling may optimize the timing and speed of invagination, and then invaginated tracheal sacs subsequently respond to FGF emanating from several target tissues guiding branching morphogenesis (Kondo, 2013).
These observations demonstrates a new role for mitosis in tissue morphogenesis to generate mechanical force through cell rounding, independent of cell division. This is distinct from previously described invagination mechanisms involving cell-autonomous constriction by the apical activation of actomyosin contractility, which is incompatible with mitosis. Mitosis 16 outside the tracheal placode occurs in clusters on the ectoderm surface, but does not lead to invagination, suggesting that the tracheal placode is sensitized to invaginate upon mitosis, independent of EGFR and FGFR signalling. Future research to uncover the properties of the tracheal placode that enables it to respond to clustered mitosis will explain not only this new mode of morphogenesis, but also the homeostasis mechanisms of epithelial architecture (Kondo, 2013).
The Drosophila brain is tracheated by the cerebral trachea, a branch of the first segmental trachea of the embryo. During larval stages the cerebral trachea splits into several main (primary) branches that grow around the neuropile, forming a perineuropilar tracheal plexus (PNP) at the neuropile surface. Five primary tracheal branches whose spatial relationship to brain compartments is relatively invariant can be distinguished, although the exact trajectories and branching pattern of the brain tracheae is surprisingly variable. Immuno-histochemical and electron microscopic demonstrate that all brain tracheae grow in direct contact with the glial cell processes that surround the neuropile. To investigate the effect of glia on tracheal development, embryos and larvae lacking glial cells as a result of a genetic mutation or a directed ablation, using the glial cells missing gene and a UAS-hid;rpr construct expressed in glial cells by the nrv2-Gal4 driver. In these animals, the tracheal branching pattern was highly abnormal. In particular, the number of secondary branches entering the central neuropile was increased. Wild type larvae possess only two central tracheae, typically associated with the mushroom body and the antenno-cerebral tract. In larvae lacking glial cells, six to ten tracheal branches penetrate the neuropile in a variable pattern. This finding indicates that glia-derived signals constrained tracheal growth in the Drosophila brain and restrict the number of branches entering the neuropile (Pereanu, 2006).
Outgrowth and branching morphogenesis of the tracheal system has been investigated in great detail for the tracheal network underlying the epidermis. Three phases, all of them dependent on the FGF signaling pathway, have been distinguished. In the early embryo, shortly after gastrulation, the epidermal ectoderm of segments T2-A8 gives rise to metameric pairs of tracheal placodes. Shortly after invagination, placodes resemble simple, round cups. Soon primary tracheal branches grow out at stereotypic positions from each cup. These branches grow in length and form secondary and tertiary branches. Growth of the primary (and some of the secondary) tracheal branches is induced by the FGF homolog Bnl which is expressed in strategically positioned clusters of epidermal cells. The pattern of the Bnl expressing epidermal cells foreshadows the later pattern of primary branches; both patterns are highly invariant (Pereanu, 2006).
The second phase of tracheal branching morphogenesis includes the formation of secondary branches which occur mainly at the growing tips of primary branches. High levels of the Bnl signal induce the expression of intrinsic activators of branching behavior, such as the ETS transcription factor Pointed, as well as inhibitory factors (e.g., sprouty) that restrict the formation of secondary branches to positions that are somewhat remote from the primary branch tip. The resulting pattern of secondary and higher order branches is considerably more variable, given that no 'hard-wired' prepattern exists. The third and terminal phase of tracheal branching describes the formation of a filigrane network of tracheoles that sprout from the tips of secondary tracheal branches. This process occurs largely after hatching of the embryo and depends on extrinsic factors, among them local oxygen levels. It is therefore a plastic, demand-based process. Again, the FGF signaling cascade plays a central role in terminal branching (Pereanu, 2006).
The development of the ganglionic tracheal branches (GBs) that supply oxygen to the ventral nerve cord is initiated as a typical 'phase 1' process where a ventrally located cluster of epidermal cells guides the formation of a secondary branch towards the edge of the neural primordium. Subsequently, the tip of the GB follows the roots of the peripheral nerve towards the neuropile. It curves around the ventral edge of the neuropile and then turns dorsally. The formation of short tertiary tracheal branches that interconnect the corresponding ganglionic branches of the left and right side, as well as neighboring ganglionic branches of the same side, constitutes a 'phase 2' event and, accordingly, leads to a relatively variable plexus of tertiary tracheae. Aside from FGF signaling, the Slit/Robo pathway forms part of the molecular network that controls the phase 2 branching morphogenesis of the GB. Slit is expressed by midline glial cells, a subset of neuropile glia derived from the mesectoderm. GB cells express the Slit receptors Robo and Robo 2. The latter is required to attract the GB towards midline; the former inhibits crossing (Pereanu, 2006).
Tracheation of the brain appears to be fundamentally similar to that of the epidermis and ventral nerve cord, and it is reasonable to apply to brain tracheation the three-step model. Complicating this analysis is the fact that the entire brain, including the gnathal part of the ventral nerve cord that later becomes the subesophageal ganglion, is tracheated by a single trachea, the cerebral trachea, which splits into five relatively invariant main branches (called 'primary' branches in this paper) that form the perineuropilar plexus around the brain neuropile. Strictly speaking, the cerebral trachea represents a primary branch of the first tracheal primordium, and the main tracheae [basomedial cerebral trachea (BMT), basolateral cerebral trachea (BLT), basocentral cerebral trachea (BCT), centromedial cerebral trachea (CMT), and centroposterior cerebral trachea (CPT)] would therefore constitute secondary branches. In line with such interpretation, the trajectories and branching patterns of the brain tracheae is much more variable than that of typical primary branches in the trunk. In contrast, with respect to their diameter and expansion, brain tracheae resemble primary tracheal branches of the trunk. More analysis is required to evaluate brain tracheation in comparison to the segmentally organized tracheation of the ventral nerve cord. Of particular importance will be comparative studies that reconstruct tracheation patterns in more primitive insects. Thus, it is likely that the lack of segmental tracheal primordia in the gnathal segments and, possibly, even in the preoral head segments of Drosophila, is derived from a primitive condition in which such segmental primordia were present. The main ('primary') brain tracheae which in Drosophila arise as secondary branches of a single tracheal primordium could in the primitive condition have been formed as primary branches of one or more segmental tracheal primordia (Pereanu, 2006).
From the existing descriptions of tracheation of the ventral nerve cord it seems likely that throughout development, the GBs and their branches are in contact with glial cell precursors that surround the peripheral nerve roots and the neuropile. A direct contact between tracheae and glia does certainly occur for all tracheae of the brain, as shown in this study. Furthermore, experimental ablation of glia reveals inhibitory interactions between glial and tracheal cells. The molecular nature of these interactions remains to be established. It is tempting to invoke FGF signaling as part of the mechanism, given its pervasive involvement in other aspects of tracheal development, and the fact that the FGF receptor heartless (htl) is expressed and required for glial development. Loss of Htl prevents the formation of glial processes enwrapping the neuropile, and application of FGF soaked beads in grasshopper embryos provokes the outgrowth of such processes. Thus a scenario exists where two different FGF receptors, Htl and Btl, are expressed by adjacent tissues, namely glia and trachea, respectively. Competition for a common source of the FGF signal (from brain neurons?) or other, more complex interactions could guide the formation of glia and tracheae, and form the basis of the inhibitory action of the former on the latter (Pereanu, 2006).
The comparison of vascular patterning in the vertebrate brain and tracheal patterning in Drosophila reveals a number of similarities. In both systems, epithelial vessels penetrate the neural primordium from the outside, follow radially organized glial elements, and branch out to form a vascular/tracheal plexus, the subependymal plexus in vertebrates and the perineuropilar plexus in Drosophila. Furthermore, vascular growth in vertebrates and tracheal growth in flies is guided by specialized tip cells. Similar to the tips of extending axons, vascular/tracheal tip cells have a growth cone whose filopodia explore cues of the microenvironment, which primarily consists of glia and the extracellular matrix produced by these cells (Pereanu, 2006).
Numerous signaling mechanisms guiding vascular/tracheal tip cells are shared between vertebrates and Drosophila, among them FGF signaling which plays a preeminent role. In addition to FGF signaling, other receptor tyrosine kinases and their ligands were identified as mediators of vascular patterning in the vertebrate brain. During their radial growth into the neural primordium, vertebrate capillary tip cells, expressing VEGF and PDGF receptors, follow a gradient of VEGF. A corresponding role of the Drosophila VEGFR/PDGFR homolog, PVR, cannot be ascertained, at least during the embryonic and larval phase. Drosophila PVR is expressed and required for hemocyte differentiation and migration, but has not been found in the tracheal system (Pereanu, 2006).
Other signal-receptor systems, among them members of the semaphorin and neuropilin families of proteins, as well as adhesion molecules such as N-cadherin and integrin, modify the response of developing capillaries to FGF and VEGF. Many of these signaling mechanisms require the close interaction between capillaries and radial glia/astrocytes. Astrocyte-derived cues are also responsible for the structural changes in capillary endothelia that underlie the formation of the blood-brain barrier (BBB), which consists of specialized tight junctions, as well as a number of enzymatic transport systems which regulate molecular movements across the endothelial cell membrane. In addition, factors derived from endothelial cells, including leukemia inhibitory factor (LIF), modulate astrocytic differentiation, demonstrating that endothelium and astrocytes are engaged in a two-way inductive process. In Drosophila, cadherins (e.g. E-cadherin) are essential for tracheal morphogenesis. A more specific role of these and other adhesion systems in the tracheation of the nervous system awaits to be studied (Pereanu, 2006).
In conclusion, as in so many other instances where comparisons between organogenetic processes in Drosophila and vertebrates were conducted, one is confronted with a surprising degree of similarity, both in structural morphogenetic mechanisms and their molecular control. Given that there is little support for true homologies [tracheae are highly derived structures that only appear in insects, and even glia as a tissue might not have been present in the bilaterian ancestor. it is reasonable to assume that the constraints that acted during the independent evolution of a tracheated insect nervous system and a vascularized vertebrate nervous system were similar. In both scenarios, epithelial tubes, branching in a dichotomous way, penetrate the neural primordium at an early stage. In branching out and permeating the volume of the growing nervous system, tracheae/blood vessels had to adopt to the special microenvironment presented by the neural primordium, a microenvironment that is composed of conserved elements like axonal growth cones, a specific extracellular matrix, signaling systems controlling midline crossing, and others. Some of these elements are most likely homologous, i.e., existed in the bilaterian ancestor; one only need to look at the conserved role of the Slit/Robo system, which regulates midline crossing of axons in animals ranging from flies to humans. Novel elements that, later in evolution and possibly multiple times, were added to this conserved microenvironment presented by the neural primordium, made use of the elements already present for their own development, and thereby convergently evolved a large number of similar characteristics (Pereanu, 2006).
The development of air-filled respiratory organs is crucial for survival at birth. A combination of live imaging and genetic analysis was used to dissect respiratory organ maturation in the embryonic Drosophila trachea. Tracheal tube maturation was found to entail three precise epithelial transitions. Initially, a secretion burst deposits proteins into the lumen. Solid luminal material is then rapidly cleared from the tubes, and shortly thereafter liquid is removed. To elucidate the cellular mechanisms behind these transitions, gas-filling-deficient mutants were identified showing narrow or protein-clogged tubes. These mutations either disrupt endoplasmatic reticulum-to-Golgi vesicle transport or endocytosis. First, Sar1 was shown to be required for protein secretion, luminal matrix assembly, and diametric tube expansion. sar1 encodes a small GTPase that regulates COPII vesicle budding from the endoplasmic reticulum (ER) to the Golgi apparatus. Subsequently, a sharp pulse of Rab5-dependent endocytic activity rapidly internalizes and clears luminal contents. The coordination of luminal matrix secretion and endocytosis may be a general mechanism in tubular organ morphogenesis and maturation (Tsarouhas, 2007).
Branched tubular organs are essential for oxygen and nutrient transport. Such organs include the blood circulatory system, the lung and kidney in mammals, and the tracheal respiratory system in insects. The optimal flow of transported fluids depends on the uniform length and diameter of the constituting tubes in the network. Alterations in the distinct tube shapes and sizes cause pronounced defects in animal physiology and lead to serious pathological conditions. For example, tube overgrowth and cyst formation in the collecting duct are intimately linked to the pathology of Autosomal Dominant Polycystic Kidney Disease. Conversely, stenoses, the abnormal narrowing of blood vessels and other tubular organs, are associated with ischemias and organ obstructions (Tsarouhas, 2007 and references therein).
While the early steps of differentiation, lumen formation, and branch patterning begin to be elucidated in several tubular organs, only scarce glimpses into the cellular events of lumen expansion and tubular organ maturation are available. De novo lumen formation can be induced in three-dimensional cultures of MDCK cells. Recent studies in this system revealed that PTEN activation, apical cell membrane polarization, and Cdc42 activation are key events in lumen formation in vitro (Martin-Belmonte, 2007). In zebrafish embryos and cultured human endothelial cells, capillary vessels form through the coalescence and growth of intracellular pinocytic vesicles (Kamei, 2006). These tubular vacuoles then fuse with the plasma membranes to form a continuous extracellular lumen. Salivary gland extension in Drosophila requires the transcriptional upregulation of the apical membrane determinant Crumbs (Crb), but the cellular mechanism leading to gland expansion remains unclear (Tsarouhas, 2007 and references therein).
The epithelial cells of the Drosophila tracheal network form tubes of different sizes and cellular architecture, and they provide a genetically amenable system for the investigation of branched tubular organ morphogenesis. Tracheal development begins during the second half of embryogenesis when 20 metameric placodes invaginate from the epidermis. Through a series of stereotypic branching and fusion events, the tracheal epithelial cells generate a tubular network extending branches to all embryonic tissues. In contrast to the wealth of knowledge about tracheal patterning and branching, the later events of morphogenesis and tube maturation into functional airways have yet to be elucidated. As the nascent, liquid-filled tracheal network develops, the epithelial cells deposit an apical chitinous matrix into the lumen. The assembly of this intraluminal polysaccharide cable coordinates uniform tube growth. Two luminal, putative chitin deacetylases, Vermiform (Verm) and Serpentine (Serp), are selectively required for termination of branch elongation. The analysis of verm and serp mutants indicates that modifications in the rigidity of the matrix are sensed by the surrounding epithelium to restrict tube length. What drives the diametric expansion of the emerging narrow branches to their final size? How are the matrix- and liquid-filled tracheal tubes cleared at the end of embryogenesis (Tsarouhas, 2007)?
This study used live imaging of secreted GFP-tagged proteins to identify the cellular mechanisms transforming the tracheal tubes into a functional respiratory organ. The precise sequence and cellular dynamics were characterized of a secretory and an endocytic pulse that precede the rapid liquid clearance and gas filling of the network. Analysis of mutants with defects in gas filling reveals three distinct but functionally connected steps of airway maturation. Sar1-mediated luminal deposition of secreted proteins is tightly coupled with the expansion of the intraluminal matrix and tube diameter. Subsequently, a Rab5-dependent endocytotic wave frees the lumen of solid material within 30 min. The precise coordination of secretory and endocytotic activities first direct tube diameter growth and then ensure lumen clearance to generate functional airways (Tsarouhas, 2007).
Two strong, hypomorphic sar1 alleles were identified in screens for mutants with tracheal tube defects. In wild-type embryos, the bulk of luminal markers 2A12, Verm, and Gasp has been deposited inside the DT lumen by stage 15. However, in zygotic sar1P1 mutants (hereafter referred to as sar1), luminal secretion of 2A12, Verm, and Gasp was incomplete. The tracheal cells outlined by GFP-CAAX partially retained those markers in the cytoplasm. sar1 zygotic mutant embryos normally deposited the early luminal marker Piopio by st/ge 13. Luminal chitin was also detected in sar1 mutants at stage 15. However, the luminal cable was narrow, more dense, and distorted compared to wild-type. To test if the sar1 secretory phenotype in the trachea is cell autonomous, Sar1 was reexpressed specifically in the trachea of sar1 mutants by using btl-GAL4. Such embryos showed largely restored secretion of 2A12, Verm, and Gasp. Thus, it is concluded that tracheal sar1 is required for the efficient secretion of luminal markers, which are predicted to associate with the growing intraluminal chitin matrix (Tsarouhas, 2007).
sar1 mRNA has been reported to be abundantly maternally contributed. At later stages, zygotic expression of sar1 mRNA is initiated in multiple epithelial tissues. To monitor Sar1 zygotic expression in the trachea, a Sar1-GFP protein trap line was used. Embryos carrying only paternally derived Sar1-GFP show a strong zygotic expression of GFP in the trachea. An anti-Sar1 antibody was used to analyze Sar1 expression in the trachea of wild-type, zygotic sar1P1, and sar1EP3575Δ28 null mutant embryos were generated. Both zygotic mutants showed a clear reduction, but not complete elimination, of Sar1 expression in the trachea. To test the effects of a more complete inactivation of Sar1, transgenic flies were generated expressing a dominant-negative sar1T38N form in the trachea. In btl > sar1T38N-expressing embryos, early defects were observed in tracheal branching and epithelial integrity as well as a complete block in Verm secretion. In contrast to btl > sar1T38N-expressing embryos, zygotic sar1P1 mutant embryos show normal early tracheogenesis with no defects in branching morphogenesis and epithelial integrity (Tsarouhas, 2007).
In summary, tracheal expression of Sar1 is markedly reduced in zygotic sar1 mutant embryos. While maternally supplied Sar1 is sufficient to support early tracheal development, zygotic Sar1 is required for efficient luminal secretion (Tsarouhas, 2007).
Given the conserved role of Sar1 in vesicle budding from the ER, its subcellular localization in the trachea was determined by using anti-Sar1 antibodies. Sar1 localizes predominantly to the ER (marked by the PDI-GFP trap). Continuous COPII-mediated transport from the ER is required to maintain the Golgi apparatus and ER structure. To test if zygotic loss of Sar1 compromises the integrity of the ER and Golgi in tracheal cells, sar1 mutant embryos were stained with antibodies against KDEL (marking the ER lumen) and gp120 (highlighting Golgi structures). In sar1 mutant embryos, a strongly disrupted ER structure and loss of Golgi staining was observed in dorsal trunk (DT) cells at stage 14. Additionally, TEM of stage-16 wild-type and sar1 mutant embryos showed a grossly bloated rough ER structure in DT tracheal cells. Consistent with its functions in yeast and vertebrates, Drosophila Sar1 localizes to the ER and is not only required for efficient luminal protein secretion, but also for the integrity of the early secretory apparatus (Tsarouhas, 2007).
To analyze tracheal maturation defects in sar1 mutant embryos, sar1 strains were generated and imaged that carry either btl > ANF-GFP btl-mRFP-moe or btl > Gasp-GFP. In sar1 mutants, luminal deposition of both ANF-GFP and Gasp-GFP is reduced. Like endogenous Gasp in the mutants, Gasp-GFP was clearly retained in the cytoplasm of sar1 embryos. ANF-GFP was also retained in the tracheal cells of sar1 mutants, but to a lesser extent. Strikingly, sar1 mutants failed to fully expand the luminal diameter of the DT outlined by the apical RFP-moe localization. This defect was quantified by measuring diametric growth rates in metamere 6 for wild-type and sar1 mutant embryos. While early lumen expansion commences in parallel in both genotypes, the later diametric growth of sar1 mutants falls significantly behind compared to wild-type embryos. The DT lumen in sar1 mutants reaches only an average of 70% of the wild-type diameter at early stage 16. Identical diametric growth defects were detected in fixed sar1 mutant embryos expressing btl > GFP-CAAX by analysis of confocal yz sections or TEM. Reexpression of sar1 in the trachea of sar1 mutant embryos not only rescued secretion, but also the lumen diameter phenotype at stage 16. In contrast to the diametric growth defects, DT tube elongation in sar1 embryos was indistinguishable from that in wild-type. This demonstrates distinct genetic requirements for tube diameter and length growth. It also reveals that the sar1 DT luminal volume reaches less than half of the wild-type volume. Prolonged live imaging showed that sar1 mutants are also completely deficient in luminal protein and liquid clearance. Up to 80% of the rescued embryos also completed luminal liquid clearance, suggesting that efficient tracheal secretion and the integrity of the secretory apparatus are prerequisites for later tube maturation steps. Taken together, the above-described results show that tracheal Sar1 is selectively required for tube diameter expansion. Additionally, subsequent luminal protein and liquid clearance fail to occur in sar1 mutants (Tsarouhas, 2007).
Do the tracheal defects of sar1 reflect a general requirement for the COPII complex in luminal secretion and diameter expansion? To test this, lethal P element insertion alleles were examined disrupting two additional COPII coat subunits, sec13 and sec23. Mutant sec13 and sec23 embryos were stained for luminal Gasp and for Crb and α-Spectrin to highlight tracheal cells. At stage 15, embryos of both mutants show a clear cellular retention of Gasp. Furthermore, stage-16 sec13 and sec23 embryos show significantly narrower DT tubes when compared to wild-type. The average diameter of the DT branches in metamere 6 was 4.8 μm and 4.4 μm in fixed sec13 and sec23 embryos, respectively, compared to 6.3 μm in wild-type. Therefore, sec13 and sec23 mutants phenocopy sar1. The phenotypic analysis of three independent mutations disrupting ER-to-Golgi transport thus provides a strong correlation between deficits in luminal protein secretion and tube diameter expansion (Tsarouhas, 2007).
The live-imaging approach defines the developmental dynamics of functional tracheal maturation. At the organ level, three sequential and rapid developmental transitions were identified: (1) the secretion burst, followed by massive luminal protein deposition and tube diameter expansion, (2) the clearance of solid luminal material, and (3) the replacement of luminal liquid by gas. Live imaging of each event additionally revealed insights into the startlingly dynamic activities of the tracheal cells. ANF-GFP-containing structures and apical GFP-FYVE-positive endosomes rapidly traffic in tracheal cells during the secretion burst and protein clearance. The direct live comparison between wild-type and mutant embryos further highlights the dynamic nature of epithelial activity during each pulse (Tsarouhas, 2007).
This study identified several mutations that selectively disrupt distinct cellular functions and concurrently interrupt the maturation process at specific steps. This clearly demonstrates the significance of phenotypic transitions in epithelial organ maturation and establishes that secretion is required for luminal diameter expansion and endocytosis for solid luminal material clearance (Tsarouhas, 2007).
The sudden initiation of an apical secretory burst tightly precedes diametric tube expansion. The completion of both events depends on components of the COPII complex, further suggesting that the massive luminal secretion is functionally linked to diametric growth. How does apical secretion provide a driving force in tube diameter expansion? In mammalian lung development, the distending internal pressure of the luminal liquid on the epithelium expands the lung volume and stimulates growth. Cl− channels in the epithelium actively transport Cl− ions into luminal liquid. The resulting osmotic differential then forces water to enter the lung lumen, driving its expansion (Olver, 2004). By analogy, the tracheal apical exocytic burst may insert protein regulators such as ion channels into the apical cell membrane or add additional membrane to the growing luminal surface. Since the ER is a crucial cellular compartment for intracellular traffic and lipogenesis, its disruption in sar1 mutants may disrupt the efficient transport of so far unknown specific regulators or essential apical membrane addition required for diametric expansion. Alternatively, secreted chitin-binding proteins (ChB) may direct an increase of intraluminal pressure and tube dilation. Overexpression of the chitin-binding proteins Serp-GFP or Gasp-GFP was insufficient to alter the diametric growth rate of the tubes, suggesting that lumen diameter expansion is insensitive to increased amounts of any of the known luminal proteins. In sar1 mutants, the secretion of at least two chitin-binding proteins, Gasp and Verm, is reduced. Chitin, however, is deposited in seemingly normal quantities, but assembles into an aberrantly narrow and dense chitinous cable. This phenotype suggests that the correct ratio between chitin and multiple interacting proteins may be required for the correct assembly of the luminal cable. Interestingly, sar1, sec13, and sec23 mutant embryos form a severely defective and weak epidermal cuticle (Abrams, 2005). The luminal deposition of ChB proteins during the tracheal secretory burst may orchestrate the construction and swelling of a functional matrix, which, in turn, induces lumen diameter dilation. While this later hypothesis is favored, it cannot be excluded that other mechanisms, either separately or in combination with the dilating luminal cable, drive luminal expansion (Tsarouhas, 2007).
During tube expansion, massive amounts of luminal material, including the chitinous cable, fill the tracheal tubes. This study found that Dynamin, Clathrin, and the tracheal function of Rab5 are required to rapidly remove luminal contents, indicating that endocytosis is required for this process. Several lines of evidence argue that the tracheal epithelium activates Rab5-dependent endocytosis to directly internalize luminal material. First, the tracheal cells of rab5 mutants show defects in multiple endocytic compartments. These phenotypes of rab5 mutants become apparent during the developmental period matching the interval of luminal material clearance in wild-type embryos. Second, tracheal cells internalize two luminal markers, the endogenously encoded Gasp and the dextran reporter, exactly prior and during luminal protein clearance. The number of intracellular dextran puncta reaches its peak during the clearance process and ceases shortly thereafter. Lastly, intracellular puncta of both Gasp and dextran colocalize with defined endocytic markers inside tracheal cells. The colocalization of Gasp and dextran with GFP-Rab7 and of Gasp with GFP-LAMP1 suggests that the luminal material may be degraded inside tracheal cells. Taken together, these data show that the tracheal epithelium activates a massive wave of endocytosis to clear the tubes (Tsarouhas, 2007).
Endocytic routes are defined by the nature of the internalized cargoes and the engaged endocytic compartments. What may be the features of the endocytic mechanisms mediating the clearance of luminal material? The phenotype of chc mutants and the presence of intracellular Gasp in CCVs indicate that luminal clearance at least partly relies on Clathrin-mediated endocytosis (CME). In addition to CME, Dynamin and Rab5 have also been implicated in other routes of endocytosis, suggesting that multiple endocytic mechanisms may be operational in tracheal maturation. The nature of the endocytosed luminal material provides an additional perspective. While cognate uptake receptors may exist for specific cargos such as Gasp, Verm, and Serp, the heterologous ANF-GFP, degraded chitin, and the fluid-phase marker dextran may be cleared by either fluid-phase internalization or multifunctional scavenger receptors. Interestingly, Rab5 can regulate fluid-phase internalization in cultured cells by stimulating macro-pinocytosis and the activation of Rabankyrin-5. The defective tracheal internalization of dextran in rab5 mutants provides further loss-of-function evidence for Rab5 function in fluid-phase endocytosis in vivo. The above-described arguments lead to the speculation that additional Rab5-regulated endocytic mechanisms most likely cooperate with CME in the clearance of solid luminal material (Tsarouhas, 2007).
How is liquid cleared from the lumen? While very little is known about this fascinating process, some developmental and mechanistic arguments suggest that this last maturation step is mechanistically distinct. First, the interval of luminal liquid clearance is clearly distinct from the period of endocytic clearance of solids. Second, the dynamic internalization of dextran and the abundance of GFP-marked endocytic structures decline before liquid clearance. Finally, assessment of liquid clearance further suggests that it requires a distinct cellular mechanism (Tsarouhas, 2007).
Viewing the entire process of airway maturation in conjunction, some general conclusions may be drawn. First, the three epithelial pulses are highly defined by their sequence and exact timing, suggesting that they may be triggered by intrinsic or external cues. Second, the analysis of mutants that selectively reduce the amplitude of the secretory or endocyic pulses demonstrates the requirement for each epithelial transition in the completion of the entire maturation process. These pulses are induced in the background of basal secretory and endocytic activities that operate throughout development. Third, specific cellular activities exactly precede each morphological transition. Finally, the separate transitions are interdependent in a sequential manner. Efficient secretion is a prerequisite for the endocytic wave. Similarly, protein endocytosis is a condition for luminal liquid clearance. This suggests a hierarchical coupling of the initiation of each pulse to completion of the previous one in a strict developmental sequence (Tsarouhas, 2007).
This study provides a striking example of how pulses of epithelial activity drive distinct developmental events and mold the nascent tracheal lumen into an air delivery tube. These findings are likely to be relevant beyond the scope of tracheal development. The uniform growth of salivary gland tubes in flies and the excretory canal and amphid channel lumen in worms also require the assembly of a luminal matrix for uniform tube growth (Abrams, 2006; Perens, 2005). Luminal material is also transiently present during early developmental stages in the distal nephric ducts of lamprey. Thus, the coordinated, timely deposition and removal of transient luminal matrices may represent a general mechanism in tubulogenesis (Tsarouhas, 2007).
Apical extracellular matrix filling the lumen controls the morphology and geometry of epithelial tubes during development, yet the regulation of luminal protein composition and its role in tube morphogenesis are not well understood. This study shows that an endosomal-retrieval machinery consisting of Rab9, retromer and actin nucleator WASH (Wiskott-Aldrich Syndrome Protein and SCAR Homolog) regulates selective recycling of the luminal protein Serpentine in the Drosophila trachea. Secreted Serpentine is endocytosed and sorted into the late endosome. Vps35, WASH and actin filaments differentially localize at the Rab9-enriched subdomains of the endosomal membrane, where Serpentine containing vesicles bud off. In Rab9, Vps35 and WASH mutants, Serpentine was secreted normally into the tracheal lumen, but the luminal quantities were depleted at later stages, resulting in excessively elongated tubes. In contrast, secretion of many luminal proteins was unaffected, suggesting that retrograde trafficking of a specific class of luminal proteins is a pivotal rate-limiting mechanism for continuous tube length regulation (Dong, 2013).
Maturation of the tracheal tube involves diameter expansion triggered by a cell-intrinsic programme of luminal material secretion and apical membrane growth, and the tube’s longitudinal growth is negatively regulated by the presumed conversion of chitin to chitosan through deacetylation by chitin deacetylase. Each step involves temporally regulated apical secretion, but the regulatory mechanism underlying this selective cargo secretion has not been understood. This study identified Serp as a novel cargo for the Rab9-mediated retrograde recycling pathway. Using time-lapse imaging, it was demonstrated that Serp in the endocytic compartment is sorted out by the budding of actin- and WASH-enriched portions of the LEs. The similarity of the Rab9, Vps35, WASH and Serp tracheal tube-length phenotypes, and the colocalization of those in the Serp-containing endosomal-budding sites in S2 cells, suggest that Serp is one of the major cargoes of the retrograde trafficking mediated by Rab9, Vps35 and WASH. It was also found that association of WASH and actin filaments with retrograde cargo is Rab9 and Vps35 independent. Modular organization of machineries for cargo retrieval (retromer) and membrane remodelling (WASH) may allow flexibility in endosomal sorting systems. This work connects retrograde trafficking to luminal retention of the key rate-limiting protein required to restrict the tracheal tube length to the proper level and has identified a new regulatory process for tracheal tube size (Dong, 2013).
Previous works in mammalian cells revealed that recruitment of the cargo recognition subcomplex of retromer (Vps35, Vps29 and Vps26) to the endosomal membrane requires Rab5 and Rab7. Rab7 has been shown to bind directly to Vps35/29/26 complex, but no evidence is available for association of Rab5 or Rab9 with those components. This analysis of the Drosophila counterparts have shown that conversion of early to LE is accompanied by gradual reduction of relative abundance of GFP-Rab5 to RFP-Rab9 signals. Throughout the process, Vps35-mRFP is present as distinct puncta associated with endosomes. The following order of Rab9-Vps35 assembly is proposed. In early endosome, Rab9 recruits Vps35 to the endosomal membrane through physical binding. This process requires Rab5. After initial recruitment of Vps35, endosomal maturation proceeds with exchange of Rab5 with Rab7, which promotes retromer assembly and cargo concentration, followed by membrane scission induced by WASH and F-actin. Together, these results indicate that Rab5, Rab9 and Rab7 acts on retromer assembly at different stages of endosomal maturation and tubular membrane formation, where Serp and other cargos are retrieved for retrograde pathway (Dong, 2013).
In contrast to the tracheal diameter expansion, which is triggered by a cell autonomous burst of exocytosis, axial elongation is a slow and continuous process spanning stage 14–16. Therefore, any mechanism that restricts elongation is expected to be active throughout the elongation process. Previous studies showed that the two chitin deacetylases Serp and Verm act additively in tube length control and the overexpression of either one causes the same tube length defects, indicating that the amount of luminal chitin deacetylase must be precisely maintained. Furthermore, the expression of Serp mRNA in the trachea declines at stage 16, when luminal Serp protein is still abundant, implying that Serp synthesized in the previous stage must be effectively retained and reused. This study has shown that Serp is endocytosed and associates with GFP-Rab9. The requirement for Rab9 and Vps35 for the luminal retention of Serp suggests that the endocytosed Serp is sorted from LEs to the TGN for secretion into the lumen, and this recycling pathway actively maintains the steady-state level of Serp to optimize the tube elongation process. Moreover, the retrieval transport through the TGN might help promote the modification of the endocytosed Serp, to recover its activity or to bind to an adaptor for its subsequent polarized secretion (Dong, 2013).
In rab9, vps35 and wash mutants, Serp was mislocalized at the apical cortex of tracheal cells. This mislocalization may have been due to the trapping of Serp in endosomes marked with Rab7 or Rab11 which are apically localized in a Rab9-independent manner. Consistent with this idea, a recent report showed that the suppression of Vps35 expression in NLT cells and neurons causes the accumulation of its cargo protein β-secretase (BACE1) in endosomes. The second possibility is that the loss of Rab9 diverts the trafficking route of Serp from the endosomes to the cell surface, as observed in Rab9-deficient HeLa cells. In support of this idea, an enhancement of Serp accumulation at the surface of the epidermis was observed in rab9 mutants. The third possibility is that a defect in endocytosis arrested Serp at the apical membrane, because Vps35 has been shown to be involved in endocytosis in S2 cells. Further analysis will be required to understand the diverse influences of retrograde trafficking on endosomal dynamics (Dong, 2013).
The apical localization of Crb depends on the retromer complex, but it was unaffected in rab9 mutants. One possible explanation for this finding is that the alternative recycling pathway of Crb, which involves Rab11, compensates for the defect in retrograde trafficking in rab9 mutants. Another possibility is that Crb-retromer and Serp-retromer complexes use different trafficking pathways distinguished by sorting nexins, which organize endosomal membranes into distinct morphological and functional regions for transport to diverse destinations. Further study of the different sorting nexins should uncover the role of Rab9 in retromer cargo specificity (Dong, 2013).
The formin Dia and motor protein Myosin V (Didum) are required for the secretion of a number of markers, including 2A12 antigen, Pio and artificial ANF-GFP, into the tracheal lumen. On the other hand, the secretion of Serp and Verm and the cell-surface localization of Crb are normal in Dia mutants, indicating that a Dia-independent secretory pathway regulates these protein localizations in the trachea8. The spectrum of secreted protein localizations affected in the rab9 and vps35 mutants was nearly complementary to that of Dia, suggesting that the Dia-dependent and retrograde trafficking-dependent mechanisms are the two major apical secretory pathways in the trachea. Verm localization was not affected in the rab9 and vps35 mutants, suggesting that this putative chitin deacetylase behaves differently from Serp. The difference in cargo specificity for each pathway allows cells versatility in controlling the localization of different proteins according to distinct schedules, so that the secretory burst that triggers tube diameter expansion and the continuous recycling of chitin deacetylases in axial elongation are controlled separately. The results provide a molecular basis for the roles of distinct trafficking pathways in controlling tubule growth and geometry (Dong, 2013).
The Drosophila embryonic tracheal network is an excellent model to study tube size. The chitin-based apical luminal matrix and cell polarity are well known to regulate tube size in Drosophila trachea. Defects in luminal matrix and cell polarity lead to tube overexpansion. This study addressed the novel function of the rebuff (reb) gene, which encodes an evolutionarily conserved Smad-like protein. In reb mutants, tracheal tubes are moderately over-elongated. Despite the establishment of normal cell polarity, significantly reduced apical luminal matrix was observed in reb mutants. Among various luminal components, luminal Obstructor-A (ObstA) is drastically reduced. Interestingly, ObstA is localized in vesicle-like structures that are apically concentrated in reb mutants. To investigate the possibility that reb is involved in the endocytosis of ObstA, the co-localization of ObstA and endocytic markers was examined in reb mutants. It was observed that ObstA is localized in late endosomes and recycling endosomes. This suggests that in reb mutant trachea, endocytosed ObstA is degraded or recycled back to the apical region. However, ObstA vesicles are retained in the apical region and are failed to be secreted to the lumen. Taken together, these results suggest one function of reb is regulating the endocytosis of luminal matrix components (Chandran, 2018).
The apical extracellular matrix plays a central role in epithelial tube morphogenesis. In the Drosophila tracheal system, Serpentine (Serp), a secreted chitin deacetylase expressed by the tracheal cells plays a key role in regulating tube length. This study shows that the fly fat body, which is functionally equivalent to the mammalian liver, also contributes to tracheal morphogenesis. Serp is expressed by the fat body, and the secreted Serp is taken up by the tracheal cells and translocated to the lumen to functionally support normal tracheal development. This process is defective in rab9 and shrub/vps32 mutants and in wild-type embryos treated with a secretory pathway inhibitor, leading to an abundant accumulation of Serp in the fat body. Fat body-derived Serp reaches the tracheal lumen after establishment of epithelial barrier function and is retained in the lumen in a chitin synthase-dependent manner. These results thus reveal that the fat body, a mesodermal organ, actively contributes to tracheal development (Dong, 2014).
The specification of tissue identity during embryonic development requires precise spatio-temporal coordination of gene expression. Many transcription factors required for the development of organs have been identified and their expression patterns are known; however, the mechanisms through which they coordinate gene expression in time remain poorly understood.
This study shows that hormone-induced transcription factor Blimp-1 participates in the temporal coordination of tubulogenesis in Drosophila melanogaster by regulating the expression of many genes involved in tube maturation. In particular, Blimp-1 was shown to regulates the expression of genes involved in chitin deposition and F-actin organization. Blimp-1 was shown to be involved in the temporal control of lumen maturation by regulating the beginning of chitin deposition. Blimp-1 represses a variety of genes involved in tracheal maturation. Kinase Btk29A was shown to serve as a link between Blimp-1 transcriptional repression and apical extra-cellular matrix organization (Ozturk-Colak, 2018).
Specialized cellular functions and cell lineage fates are usually
regulated by only a few key instructive transcription factors required
to activate or repress specific patterns of gene expression. During the
development of multicellular organisms, these events must be precisely
timed. As a result, organogenesis requires an accurate spatio-temporal
regulation of gene expression over extended periods. While many
transcription factors required for the development of organs have been
identified and their expression pinpointed spatially, the mechanisms
through which they coordinate downstream gene expression in time remain
poorly understood. This study addressed part of this question during the
development of the Drosophila melanogaster tracheal system -- a model used
to study epithelial organ development (Ozturk-Colak, 2018).
The tracheal system of Drosophila is formed by a network of epithelial tubes that requires
tight temporal regulation of gene expression. Tracheal tube maturation
involves the timely and spatially regulated deposition of a chitinous
apical extracellular matrix (aECM), a process governed by downstream
effectors of the midembryonic ecdysone hormone pulse. One of these ecdysone response genes is the D. melanogaster B-lymphocyte inducing maturation protein-1 (Blimp-1). Blimp-1 is the homolog of human Prdm1 (positive
regulatory domain containing 1). Blimp-1/PRDM1 is a zinc
finger transcriptional repressor that belongs to the Prdm gene family,
and it was originally identified as a silencer of β-interferon gene
expression. Prdm family members contain a
conserved N-terminal domain, known as a positive regulatory (PR) domain.
This domain has been associated with the SET methyltransferase domain,
which is important for the regulation of chromatin-mediated gene
expression. In addition, Prdm family proteins
contain multiple zinc fingers that mediate sequence-specific DNA binding
and protein-–protein interactions. Prdm family
members modulate key cellular processes, including cell fate, and the
aberrant function of some members may lead to malignant transformation. During embryonic development, Blimp-1 controls a
plethora of cell-fate decisions in many organisms. In D. melanogaster, Blimp-1 serves as an
ecdysone-inducible gene that regulates ftz-f1 in pupal stages. By acting as a transcriptional repressor, Blimp-1 prevents the premature expression of ftz-f1, thereby influencing the temporal regulation of events that are crucial for insect development. The
expression level and stability of Blimp-1 is critical for the precise
timing of pupariation ( Ozturk-Colak, 2018 and references therein).
Blimp-1 exerts a function in tracheal system morphogenesis during
embryonic development. However, the question remains as to how this transcription factor
regulates tube maturation events downstream of the hormone ecdysone.
The role of Blimp-1 in the transcriptional regulation
of the regulation of tracheal tube maturation in D. melanogaster was examined in this study. Blimp-1 was found to be a transcriptional repressor of many genes involved
in tracheal development and its levels are critical for the precise
timing of luminal maturation and the final stages of tubulogenesis in
the embryo. The results indicate that Blimp-1, working downstream of
ecdysone, acts as a link of hormone action during tube maturation in
organogenesis (Ozturk-Colak, 2018).
This study found that Blimp-1 regulates multiple tracheal targets, thus
acting as a key gene in tracheal development.
Blimp-1 is an ecdysone response gene and, therefore, a link between the hormonal signal
and the timing of tracheal tube maturation in both embryos and larvae.
Blimp-1 was found to regulate the expression of many genes required for
tube maturation. Interestingly, in silico, four Blimp-1
binding sites were detected in Blimp-1 regulatory sequences using the parameters
described, which suggests that Blimp-1 may regulate its own expression.
This is in agreement with recent data showing that Blimp-1/PRDM1 is also
able to regulate its own expression in mammals.
Self-regulation of expression is consistent with the feedback loops in
which Blimp-1/PRDM1 participates, and also with its role in regulating
many developmental processes. Blimp-1 binding sites were found in the region of Tramtrack (Ttk),
another transcription factor involved in many features of tube
maturation (Ozturk-Colak, 2018).
Furthermore, Blimp-1 was observed to modulate the timing of the
expression of reb and exp, two genes involved in the genetic program
triggering timely chitin deposition. Untimely
chitin deposition was shown to disturb tube maturation, thereby
demonstrating that this process has to be tightly regulated during
tracheal development. Tracheal overexpression of reb leads to earlier
chitin deposition in all branches from stage 13 and sometimes chitin
appearance at stage 12. Accordingly, the results show that Reb is expressed
earlier in Blimp-1 embryos. This agrees with the
early chitin deposition phenotypes observed in Blimp-1 mutants. Furthermore, Blimp-1 also modulated knk expression during tube maturation stages. Knk is involved in directing chitin assembly in
the trachea and correct amounts of Knk at
specific times during metamorphosis are important for correct wing
cuticle differentiation and function. Taken together,
these in silico and in vivo results indicate that Blimp-1 is a
transcription factor that acts downstream of ecdysone, and that it is
involved in the correct timing of chitin synthesis and deposition during
embryonic development (Ozturk-Colak, 2018).
Blimp-1 binding sites were also found in the aPKC coding region. aPKC is
involved in the junction anisotropies that orient both actin rings and
taenidial ridges in the lumen of tracheal tubes. In
Blimp-1 mutants, both actin rings and taenidial ridges are either
undetectable or misoriented (Ozturk-Colak, 2016) --observations that
are consistent with changes in junction anisotropy (Ozturk-Colak, 2018).
Previous work has shown that Blimp-1 regulates chitin deposition levels and
architecture, and that, subsequently, the chitin aECM feeds back on the
cellular architecture by stabilizing F-actin bundling and cell shape via
the modulation of Src42A phosphorylation levels. However, this report provided no link between the
chitinous aECM and Src42A. Btk29A mutant larvae have an aECM phenotype,
which may be the result of their actin bundle phenotype. This study found that Btk29A removal can
partially rescue the Blimp-1 taenidial orientation and expansion
phenotype. In view of these results, it is proposed that the contribution of
Btk29A can be added to the feedback model for the generation of
supracellular taenidia put forward in Ozturk-Colak (2016). This study now
adds to a previous model by hypothesizing that Blimp-1 acts as a link
between the aECM and cells by regulating the levels of Btk29A. Btk29A and Src42A, together with the formin DAAM, have been shown to regulate the actin cytoskeleton. In agreement
with the current results, it is speculated that Btk29A might phosphorylate Src42A,
and that this phosphorylation event could be modulated by Blimp-1 and
DAAM (Ozturk-Colak, 2018).
To conclude, the results indicate that Blimp-1 is a key player in the
regulation of tracheal tube maturation, and, consequently, in the
feedback mechanism involved in the generation of supracellular taenidia (Ozturk-Colak, 2018).
During Drosophila metamorphosis, most larval cells die. Pupal and adult tissues form from imaginal cells, tissue-specific progenitors allocated in embryogenesis that remain quiescent during embryonic and larval life. Clonal analysis and fate mapping of single, identified cells show that tracheal system remodeling at metamorphosis involves a classical imaginal cell population and a population of differentiated, functional larval tracheal cells that reenter the cell cycle and regain developmental potency. In late larvae, both populations are activated and proliferate, spread over and replace old branches, and diversify into various stalk and coiled tracheolar cells under control of fibroblast growth factor signaling. Thus, Drosophila pupal/adult tissue progenitors can arise both by early allocation of multipotent cells and late return of differentiated cells to a multipotent state, even within a single tissue (Weaver, 2008).
Drosophila larval tissues are composed of differentiated larval cells and imaginal cells. Imaginal cells are pupal and adult tissue progenitors that reside in clusters embedded in or attached to larval tissue. They remain quiescent during embryogenesis and part or all of larval life, then proliferate and differentiate into pupal and adult tissues at metamorphosis. By contrast, larval cells cease dividing and differentiate early in development; however, they typically enlarge and become polyploid during larval life. At metamorphosis, most larval cells die. Although some larval neurons and muscles are retained in adult tissues, no differentiated cells are known to reenter the cell cycle and generate new cells and tissues. This study shows that tracheal (respiratory) system remodeling at metamorphosis is carried out by a classical imaginal cell population and another progenitor population that, like facultative stem cells in mammals, arises from differentiated cells (Weaver, 2008).
During embryogenesis, the tracheal system develops from segmentally repeated groups of ~80 cells that express Trachealess transcription factor and invaginate, forming sacs attached to epidermis by a stalk of spiracular branch (SB) cells. Branches bud from the sacs and cells diversify primarily under control of Branchless FGF (fibroblast growth factor), which activates Breathless FGFR (FGF Receptor) on tracheal cells. At metamorphosis, posterior tracheal segments Tr6 to Tr10 are lost; new branches form in Tr4 and Tr5 to supply posterior tissues and in Tr2 to supply flight muscle. Although most branches in Tr1 to Tr5 are retained, most of their cells are replaced by imaginal cells (Weaver, 2008)..
Previous work indicated that imaginal tracheal cells (tracheoblasts) compose the SB. Unlike other tracheal cells, SB cells express imaginal marker escargot (esg), remain small and quiescent during embryonic and early larval life, and do not form gas transport tubes. Bromodeoxyuridine (BrdU) incorporation studies showed that SB cells enter S phase at the beginning of the third larval period (L3) and divide 12 to 16 hours later. Proliferation continues for 24 hours, generating an expanding cluster of tracheoblasts at the SB-transverse connective (TC) junction. SB cells in the embryo and early larva express trachealess (trh) but, unlike most other tracheal cells, do not express the Trachealess target gene breathless (btl): The tracheal program is apparently arrested at this step. When activated in L3, they turn down esg and turn on btl as they proliferate and leave the SB (Weaver, 2008).
A 'molecular timer' strain was developed that highlights the burst of btl expression in activated tracheoblasts, which allowed distinguishing them from the larval tracheal cells they migrate over and replace. SB tracheoblasts followed stereotyped paths. In Tr4, they migrated along the TC onto the visceral branch; later, some differentiated into coiled tracheolar (CT) cells. Tracheoblasts respected specific boundaries, never spreading into neighboring tracheal segments or populating the dorsal trunk (DT). However, tracheoblasts were observed on the other side of the DT, along the dorsal branch (DB) of Tr4 and other anterior DBs (Weaver, 2008).
If DB tracheoblasts arise from SB tracheoblasts, they would have to move across the DT to reach the DB. However, tracheoblasts were never seen crossing the DT. To exclude this possibility, the fate of SB tracheoblasts was mapped using SB-specific flipase (FLP) recombinase to permanently label SB cells and their descendants. This labeled all tracheoblasts migrating out of the SB, but not DB tracheoblasts. Hence, DB tracheoblasts arise independently (Weaver, 2008).
To identify the source of DB tracheoblasts, early L3 larval DBs, which comprise five to seven cells were scrutinized, but no additional cells or cells were found with the distinctive small size and nuclear morphology of SB tracheoblasts. The positions and number of tracheoblast clones in a clonal analysis of larval DBs suggested that DB tracheoblasts arise from ~4 to 5 progenitors along the DB (Weaver, 2008).
Whether differentiated stalk cells (DB3 to DB7) might be the source of DB tracheoblasts was considered. To test this, a heat-inducible FLP transgene was used to permanently label and trace the fate of individual tracheal cells identified in live L2 larvae. This demonstrated that larval DB stalk cells are the source. Individual stalk cells displayed a range of proliferative capacities, giving rise to 2 to 22 tracheoblasts, although occasionally a labeled stalk cell failed to proliferate or degenerated, the standard fate of DB1 and DB2 cells (Weaver, 2008).
DB stalk cells have a complex morphology unlike typical progenitor or stem cells: They are tubular, with autocellular junctions, some (DB3 cells) forming Y-shaped tubes. Yet these cells become proliferating, migrating DB tracheoblasts while maintaining contacts with neighboring tracheal cells (Weaver, 2008).
Phosphohistone H3 staining showed that DB stalk cells in anterior segments begin dividing 14 to 16 hours after the second molt. Even before they divide, they have smaller nuclei than other larval tracheal cells, including DB stalk cells in posterior segments, which are otherwise indistinguishable but do not give rise to tracheoblasts. BrdU labeling of newly molted third-instar larvae showed that anterior DB stalk cells do not incorporate the label, implying that they do not endoreplicate and presumably remain diploid, unlike posterior DB cells and other differentiated larval cells, most of which endoreplicate and become polyploid (Weaver, 2008).
Although most DB tracheoblasts form multicellular stalks of pupal DBs, in Tr2 they form more elaborate structures. After proliferating and spreading along the DB, they aggregate, form secondary branches, and differentiate into multicellular stalks (MS), unicellular stalks (US), and Blistered (DSRF)-expressing CT cells with coiled intracellular lumens that unfurl on flight muscle. Fate mapping showed that single DB stalk cells in Tr2 routinely formed mixed clones containing MS, US, and CT cells. Thus, DB stalk cells in Tr2 transform into multipotent tracheoblasts that can proliferate and acquire different fates (Weaver, 2008).
Bnl/Btl signaling controls cell fate selection in the embryo. To test for function in pupal tracheoblasts, btl minus clones were generated. These rarely formed CT cells. Likewise, conditional expression of dominant-negative Btl in the L3 tracheal system reduced or eliminated secondary branches and CT cells. Constitutively active receptor induced ectopic secondary branches and CT cells throughout the tracheal system, including all anterior DBs. Thus, DB tracheoblasts in anterior segments can acquire new tracheal fates, and FGF signaling also plays a critical role in reselecting cell fates when DB stalk cells are reactivated (Weaver, 2008).
Anterior DB stalk cells are the first differentiated cells in Drosophila shown to reenter the cell cycle and regain developmental potency. They regain the same abilities to proliferate, spread, and differentiate into various tracheal cell types as SB tracheoblasts, classical imaginal cells that remain quiescent (blocked in tracheal outgrowth and cell diversification) during embryonic and most of larval life. This suggests that both types of tracheal progenitors arrive at a similar state, one by early developmental arrest (SB tracheoblasts), the other by late return to an earlier state (anterior DB stalk cells). The only known features that distinguish these cells from tracheal cells that lack progenitor potential (including posterior DB stalk cells that are otherwise indistinguishable from anterior DB stalk cells) are their small nuclear size and lack of endoreplication. These features may be part of a program that maintains, or allows cells to regain, the proliferative and diversification potential of early tracheal cells. This program is operative in imaginal tracheal cells and can apparently be implemented in other tracheal cells, independent of their differentiation program (Weaver, 2008).
The blurring of the distinction between imaginal and differentiated larval cells in Drosophila parallels a current debate about adult stem cells in mammals. Some mammalian tissues have dedicated stem cells maintained in a primitive state. However, other tissues may rely on facultative stem cells, differentiated cells that reenter the cell cycle to replenish lost cells. The current results show that progenitors with each of these features are present in a single Drosophila tissue and that both play crucial roles. This provides a tractable system for dissection of the arrest of a tissue-specific developmental program and reversal to an earlier, more plastic state, important steps in tissue engineering, repair, and cancer (Weaver, 2008).
Adult progenitor cells in the trachea of Drosophila larvae are activated and migrate out of niches when metamorphosis induces tracheal remodeling. In response to metabolic deficiency in decaying tracheal branches, signaling by the insulin pathway controls the progenitor cells by regulating Yorkie (Yki)-dependent proliferation and migration. Yki, a transcription coactivator that is regulated by Hippo signaling, promotes transcriptional activation of cell cycle regulators and components of the extracellular matrix in tracheal progenitor cells. These findings reveal that regulation of Yki signaling by the insulin pathway governs proliferation and migration of tracheal progenitor cells, thereby identifying the regulatory mechanism by which metabolic depression drives progenitor cell activation and cell division that underlies tracheal remodeling (Li, 2022).
Robust and controlled intestinal regeneration involves reciprocal interactions between the intestinal epithelium and its microenvironment. This study identifed signaling between enteroendocrine (EE) cells, vasculature-like trachea, and neurons, which drives regional and global stem cell proliferation during adult intestinal regeneration in Drosophila. Reactive oxygen species (ROS) from midgut cells promote production and secretion of diuretic hormone 31 (Dh31), from anterior midgut enteroendocrine (EE) cells. EE and neuronal Dh31 activate tracheal Dh31 receptor, leading to the production of the vascular endothelial growth factor (VEGF)- and platelet-derived-growth-factor (PDGF)-like ligand Pvf1. Pvf1 induces tracheal remodeling and intestinal stem cell (ISC) proliferation through autocrine and paracrine Pvr/mitogen-activated protein kinase (MAPK) signaling, respectively. While EE Dh31 exerts broad control of ISC proliferation throughout the midgut, effects of the neuronal source of the ligand appear restricted to the posterior midgut. Collectively,this work discovered an EE/neuronal/vascular signaling network, controlling global and domain-specific ISC proliferation during adult intestinal regeneration (Medina, 2025).
The tubular network of the tracheal system in the Drosophila embryo is created from a set of epithelial placodes by cell migration, rearrangements, fusions and shape changes. A designated number of cells is initially allocated to each branch of the system. The final cell number in the dorsal branches is not only determined by early patterning events and subsequent cell rearrangements but also by elimination of cells from the developing branch. Extruded cells die and are engulfed by macrophages. These results suggest that the pattern of cell extrusion and death is not hard-wired, but is determined by environmental cues (Baer, 2010).
In live studies of the tracheal system using embryos expressing GFP under the control of the breathless (btl) promoter GFP-expressing motile cells were observed that were not attached to the tracheal system. The btl gene, which encodes an FGF receptor homolog, is mainly expressed in the tracheal system, but also in glial cells and a few other cell type. However, it had not previously been described as being expressed in individual cells that were dispersed in the embryo. Since it was not clear what the individual cells outside the tracheal system were, other tracheal markers were used to determine whether they indeed derived from the tracheal system, or rather represented an as yet undiscovered cell type in which the btl promoter is active. When the tracheal system was marked with lacZ expressed under the control of the promoter of another tracheal gene, trachealess (trh-lacZ), it was observed that lacZ was also expressed not only in the tubular tracheal epithelia but also in single cells detached from the tracheal system (Baer, 2010).
In the live observations using GFP it was noticed that these cells appeared to be moving around in the embryo, preferentially along the dorsal trunk of the trachea. Previously hemocytes had been described to move along the tracheal system. Thus it was asked whether these cells might be a subpopulation of hemocytes that shared expression of some genes with the tracheal system. To assess their cell type, the cells were analyzed in embryos in which markers for the tracheal system and for hemocytes were simultaneously visualized. Hemocytes were visualized by using a croquemort-GAL4 transgene to control the expression of GFP. Croquemort is expressed in a subpopulation of hemocytes, the macrophages. For the tracheal system trh-lacZ was used (Baer, 2010).
It was found that the individual lacZ-expressing cells that are detached from the tracheal system also expressed the crq-controlled GFP. In addition, there were also many cells that showed only crq-GFP staining and no lacZ. There are two possible explanations for these findings. Either a subset of macrophages can activate transcription of the btl and trh promoters, or the doubly-stained objects are not single cells. Consistent with the latter explanation, it was noticed that the two fluorescent markers were often localized in non-overlapping parts of the cells, suggesting that lacZ-expressing cells or cell fragments might have been engulfed by macrophages. If this were the case, then the doubly-stained objects should contain two nuclei. Indeed, in triple stainings with the nuclear marker TOTO-3 two TOTO-3 signals were observed in the majority of the doubly-stained objects, indicating the presence of two nuclei. It is concluded therefore that the objects are hemocytes that have engulfed tracheal cells that have left the tracheal epithelium (Baer, 2010).
To test directly whether these single cells originate from the developing tracheal tree, more detailed live observations were performed. In previous experiments it was noticed that the disconnected cells can be preferentially found near the branches that undergo cellular rearrangements, for example, at the bases of dorsal branches. The areas around these sites were chosen for further studies. Live imaging of dorsal branches of the embryos expressing α-cat-GFP or sqh-GFP revealed that during the outgrowth of the dorsal branches individual tracheal cells detach from the tracheal system at, or near the site where the dorsal branches emerge from the dorsal trunk. The numbers of cells leaving the system from the base of branches and from the interbranch-regions of the dorsal trunk were compared in 12 videos of wildtype embryos. The 'base of the branch' is described as those cells that bordered at least on one side on a cell that was part of the dorsal branch. 42 branches were evaluated and cells were found leaving from the base in 20 cases, whereas in 40 interbranch-regions seven cells leaving the system were found. Thus, cells are almost threefold less likely to be extruded in the large interbranch area than from the small area at the base of the branch (Baer, 2010).
Since the detached cells appeared to be engulfed by macrophages and one function of macrophages is to clear up apoptotic cells, whether the trh-lacZ positive cells seen inside the hemocytes displayed markers for apoptosis was tested. It was found that some but not all of the trh-lacZ cells that were detached from the tracheal system gave a positive signal in a TUNEL reaction. Since the TUNEL assay only detects cells in a brief phase of apoptosis it was not surprising that only a subset of the detached cells were labelled. Thus this result is consistent with the notion that the engulfed cells are dying or dead cells. A further hallmark of apoptosis is the activation of caspase 3, which can be detected with an antibody that specifically recognizes the activated form. Trh-lacZ-expressing cells that were detached from the tracheal system also gave a positive signal when stained with antibodies against activated caspase3 (Baer, 2010).
Next, it was asked whether the detached cells undergo apoptosis because they have lost contact with the tracheal epithelium, or, conversely, whether they die within the epithelium and are subsequently extruded. This question was addressed by two approaches: tracing caspase activity in vivo and blocking apoptosis. To examine when the apoptotic program is induced in tracheal cells leaving the system an in vivo fluorescent sensor of caspase activity, the 'Apoliner' transgene (Bardet, 2008), was used. The Apoliner consists of two fluorophores, a membrane-anchored RFP and a GFP with nuclear localisation signal (NLS), which are linked by a caspase sensitive fragment of the DIAP1 protein. Upon caspase activation within the cell, the sensor is cleaved, resulting in translocation of GFP into the nucleus, whereas RFP stays at the membrane. The Apoliner was expressed using btl-GAL4 and the development of dorsal branches was followed in vivo. Nuclear GFP can be observed in individual cells in the tracheal system. Shortly after they begin to accumulate nuclear GFP these cells detach from the system. Thus apoptosis precedes cell removal from the tissue (Baer, 2010).
If death is a prerequisite for expulsion, then suppressing apoptosis should reduce the number of detached cells. To test this, the tracheal system was examined in embryos homozygous for a deficiency Df(3L)H99, which removes the three pro-apoptotic genes, grim, reaper and hid, and in which apoptosis therefore cannot occur. Since homozygous Df(3L)H99 embryos show strong overall developmental defects, the analysis was restricted to the dorsal branches in segments that do not show gross morphological abnormalities, i.e. segments 3-7. Whereas wild type stage 14/15 embryos show approximately 2-4 detached cells in segments 3-7 on each side, no detached cells were observed in embryos that were homozygous for Df(3L)H99. To exclude the possibility that this was due to a non-autonomous effect of the mutant phenotype of surrounding tissues in Df(3L)H99 mutant embryos, an alternative way of blocking apoptosis only in the tracheal system was used. The baculoviral inhibitor of apoptosis p35 was expressed in tracheal cells using the btl-GAL4 driver line. To confirm that apoptosis was blocked efficiently, the Apoliner was co-expressed together with p35. In these embryos no Apoliner signal was detected, indicating the absence of caspase activity, and no cells were seen to leave the tracheal system. This shows that the completion of the apoptotic program is necessary for detachment of the tracheal cells (Baer, 2010).
The fact that the detached cells were frequently found near the base of the dorsal branches of the tracheal system raises the question whether their leaving the epithelium might be associated with a specific developmental programme, such as the morphogenesis of the dorsal branches. Since each dorsal branch arises from six cells, but the final branch typically consists of only five cells, it has been suggested that the sixth cell migrates back to the dorsal trunk. The results point to an alternative possibility suggesting that apoptosis might be a mechanism that contributes to the determination of cell number in the dorsal branches. If the assumption that cell death is involved in determining the number of cells is correct, then dorsal branches should contain more cells if apoptosis cannot occur. To test this, cell numbers were counted in the dorsal branches of Df(3L)H99 mutant embryos and in embryos expressing p35 in the tracheal system. The embryos were stained with an antibody against Trachealess to mark specifically the nuclei of tracheal cells. The nuclei in dorsal branches were then counted under the microscope by focusing through the entire depth of the dorsal branch. Branches with an unclear course were not included in the analysis (Baer, 2010).
All tested genotypes showed variability in cell number in the dorsal branches, but there were clear differences between the mutant and the wild type embryos. 76.3% of the 93 dorsal branches evaluated in 10 wild type embryos consisted of 5 cells, 8.6% consisted of six cells and 15.1% consisted of 3 or 4 cells. This shows that branches with six cells are possible, but 5 cells are preferred. By contrast, embryos with blocked apoptosis had 51.6% (btl-Gal4, UAS-p35 embryos) or 56.6% (Df(3L)H99 mutant embryos) dorsal branches with six cells. These results indicate that removal of the cells by apoptosis indeed contributes to the determination of the final cell number in the dorsal branches (Baer, 2010).
It is unclear how the emigrating cell is determined, and why it is not determined in every one of the branches. The fact that this is so shows that, in contrast to many other situations in which apoptosis eliminates unwanted cells during development, cell death in this case is not the result of a hard-wired developmental programme. Instead it is more likely to be a response to unfavourable conditions in the cell's environment. Changes in the microenvironment have been shown to induce cell death in mammalian endothelial cells, with signals from the extracellular matrix influencing the balance between cell survival and apoptosis. Adhesion via integrins can protect cells from FAS mediated apoptosis, and integrins may act as mechano-transducers in this context. Thus it is possible that weakening of cell-cell junctions resulting from tissue remodelling in the tracheal system could trigger the apoptotic pathway. Junction rearrangement also affects cell death in the wing disc, where an imbalance in the junctional forces within the epithelium was found to be rebalanced by the elimination of cells. In this system, the initial irregularity arose through cell proliferation, but it is imaginable that in the tracheal system cell intercalation might affect local junctional forces, and cell extrusion can be triggered to obtain a geometrically optimal structure (Baer, 2010).
The fact that only half of the dorsal branches in the mutant embryos had six cells shows that apoptosis cannot be the only mechanism for the removal of supernumerary cells. There must be other ways of losing cells, perhaps by re-integration into the dorsal trunk, as previously suggested. Such cases were indeed observed. More stringently, even embryos in which a larger number of branches have six cells, namely btl-Gal4, UAS-p35 embryos, can develop normally and hatch. It is of course possible that subtle defects, for example a reduced efficiency in oxygen delivery, would not manifest themselves in easily measurable phenotypes in a laboratory setting (Baer, 2010).
In summary, the results are consistent with a scenario in which cells that find themselves in a sub-optimal epithelial context either move away, or die and leave the epithelium. If the apoptotic pathway is blocked, they may be forced to use the option of moving elsewhere, or induce neighbouring cells to rearrange further, or they remain in place, and the system is able to tolerate a sub-optimal structure (Baer, 2010).
The tracheal system of Drosophila has proven to
be an excellent model system for studying the development of branched
tubular organs. Mechanisms regulating the patterning and initial
maturation of the tracheal system have been largely worked out, yet
important questions remain regarding how the mature tubes inflate with
air at the end of embryogenesis, and how the tracheal system grows in
response to the oxygen needs of a developing larva that increases nearly
1000-fold in volume over a four day period. This study describes the
cloning and characterization of uninflatable (uif), a gene that
encodes a large transmembrane protein containing carbohydrate binding
and cell signaling motifs in its extracellular domain. Uif is highly
conserved in insect species, but does not appear to have a true ortholog
in vertebrate species. uif is expressed zygotically beginning in
stage 5 embryos, and Uif protein localizes to the apical plasma membrane
in all ectodermally derived epithelia, most notably in the tracheal
system. uif mutant animals show defects in tracheal inflation at
the end of embryogenesis, and die primarily as larvae. Tracheal tubes in
mutant larvae are often crushed or twisted, although tracheal patterning
and maturation appear normal during embryogenesis. uif mutant
larvae also show defects in tracheal growth and molting of their
tracheal cuticle (Zhang, 2009).
This study has described the
initial characterization of a novel gene, uninflatable, which
encodes a single-pass type I transmembrane protein with several
different carbohydrate binding motifs and epidermal growth factor
repeats in its extracellular domain. This gene is highly conserved in
insect species, but does not appear to have a true ortholog in mammals
or other vertebrate organisms. Uninflatable is expressed on the apical
surface of ectodermally-derived epithelial cells including the
epidermis, trachea, salivary gland and fore- and hindgut, although it is
most highly expressed in tracheal epithelia starting as the tracheal
placode invaginates and persisting throughout its development. Three mutant alleles of uif were isolated, and the
predominant defects associated with loss of uif were found to be defective
air inflation at the end of embryogenesis, and tracheal growth and
tracheal molting defects in larvae (Zhang, 2009).
The most obvious
defects observed in uif mutant late embryos and newly hatched
first instar larvae were incomplete inflation of the trachea coupled
with tracheal tubes that often appeared pinched or stretched. Live
imaging of individual uif mutant embryos and larvae revealed
that these tracheal inflation defects result from the inability to
completely inflate the trachea at the end of embryogenesis rather than a
defect in the maintenance of an inflated tracheal system after hatching.
Interestingly, tracheae in uif mutant embryos are normally
patterned and appear to mature in a manner indistinguishable to tracheae
in wild type animals. Thus this tracheal inflation defect likely results
from a requirement for uif function late during embryogenesis
(Zhang, 2009).
Several genes have been characterized whose defects
include incomplete tracheal inflation. Included in these genes are those
that function at the end of embryogenesis to clear the trachea of solid
luminal material. This process requires clathrin-mediated endocytosis
and includes the proteins clathrin heavy chain, the GTPase dynamin
(encoded by shibire), and a clathrin binding transmembrane
protein encoded by wurst. Notably, mutations in all of these
genes also result in elongated tracheal tubes suggesting a defect in
tracheal tube size control. Gas filling defects were also observed for
mutations in serp and verm, two putative matrix chitin
deacytlases that function to regulate tracheal tube length. Tracheal
maturation appears normal in uif mutant embryos, including the
secretion of chitin into the tracheal lumen and the subsequent uptake of
luminal solids near the end of embryogenesis. In addition, uif
mutant tracheae do not show diameter or length defects during
embryogenesis, suggesting that the luminal chitin cylinder forms
normally and that Serp and Verm are correctly functioning during the
maturation process. Taken together these results suggest that the air
filling defects observed in uif mutant embryos occurs through a
mechanism independent of that linked to endocytosis of solid luminal
material (Zhang, 2009).
A second class of proteins that likely
function in proper air inflation is the epithelial sodium channels
(ENaCs). In mammals, ENaC proteins are required to help remove liquid
from embryonic lung prior to birth. There are 16 ENaC genes
(also referred to as pickpocket or ppk genes) in the
Drosophila genome, of which 9 are expressed in the embryonic
tracheal system. It is thought that the influx of sodium through these
channels drives water from the lumen into the epithelial cells, and that
degassing of this liquid inflates the trachea. Although there are no
loss of function mutations in ENaCs that result in air filling defects
in Drosophila, RNA interference mediated knockdown of
ppk4 and ppk11 results in partially liquid-filled
tracheae in larvae. In addition, inhibiting these channels using
amiloride also results in fluid filled tracheae in larvae. Although both
of these examples affected air filling after a molt, it seems likely
that ENaC proteins may also contribute to air filling at the end of
embryogenesis. At this point it is not possible to exclude the
possibility that uif may regulate the expression or activity of
ENaC encoded proteins in tracheal epithelia (Zhang,
2009).
A third possibility, and one that is favored, is that
uif is playing primarily a structural role in embryonic
tracheal maturation. Two pieces of evidence support this notion. First,
in early stage 17 mutant embryos the tracheae are of normal length and
diameter and have a stereotypic appearance that is indistinguishable
from that of wild type animals, whereas in newly hatched mutant larvae
the tubes are often crushed or twisted. Second, while dissecting
tracheae from wild type and uif mutant third instar larvae it
was clear that the mutant tracheae were more brittle. Wild type tracheae
have an elastic property that makes them difficult to break, whereas we
had to be very careful dissecting uif tracheae in order to get
a section that included more than one metameric unit. Together these
observations suggested a model in which the mechanical properties of
uif mutant tracheae are compromised, thereby allowing the
tracheal cuticle to fail when the embryo initiates violent muscular
contractions prior to hatching. As noted in the live imaging, tracheal
inflation initiates well after the embryo begins these dramatic muscular
contractions. It is predicted that a combination of crushed tubes and
small cracks in the tracheal cuticle prevent complete inflation (Zhang,
2009).
uif mutant larvae that survive to second or third
instar show striking defects in tracheal growth and tracheal molting.
The tracheae in these uif mutant larvae have short dorsal
trunks that are well out of proportion to the body length of the animal.
It is difficult to accurately assess the diameter of the dorsal trunk in
these animals, however, because the mutant animals fail to completely
shed their tracheal cuticle, and often only the first instar lumen is
inflated. This tracheal molting defect was nearly completely penetrant
in all third instar uif2B7 and
uif1A15 mutant larvae examined, but the severity
seemed to vary along the anterior-posterior axis, with some middle and
anterior sections showing no molting defects, whereas the sections near
the posterior spiracles showed at least two and sometimes three tracheal
cuticles. A previous study carried out morphometric analyses of larval
tracheal growth and observed that tracheae lengthen in a continuous
fashion, whereas the diameter increases in a stepwise manner coincident
with the molt, suggesting that tube length is controlled independently
from tube diameter. The current results indicate that uif plays
a role in regulating the growth of the tracheae along their length
(Zhang, 2009).
How might uif regulate larval tracheal tube
length? One possibility is that Uif might regulate the interface between
the tracheal epithelium and the cuticle, possibly by providing
lubrication through its hyalin domains. Loss of uif would
therefore result in a situation where the epidermis is too tightly bound
to the overlying cuticle to allow for growth along the long axis of the
tracheae. This type of mechanism could also account for the tracheal
molting defects and thereby couple these phenotypes. An earlier study
identified mutations in the Matrix Metalloproteinase encoded by
Mmp1; similar defects were found in tracheal elongation. It was
speculated that the Mmp1 tracheal defects might be caused by
the inability of the tracheal epithelial cells to loosen their
attachment to the cuticle. Since uif2B7 mutant
larvae have no full length Uif protein, it is unlikely that Uif is a
primary target of MMP1 in tracheal cells. In addition, this study
observed no genetic interaction between uif and Mmp1
by second-site noncomplementation (Zhang, 2009).
An
alternative hypothesis is that Uif might serve as a receptor or
co-receptor for a systemic signal that couples tracheal growth with
larval growth. The extracellular domain of Uif contains multiple EGF
domains and a laminin G domain, and the cytoplasmic domain is remarkably
conserved in all insect species. A combination of structure/function
analysis and genetic and biochemical approaches to identify Uif
interacting proteins should help to shed light on this functions of
uif. Understanding how Uif couples tracheal growth to the
growth of the larva may serve as an important paradigm for similar
couplings of organ growth to organismal growth in other species (Zhang,
2009).
Finally, hypoxia may account for all the other larval
phenotypes associated with loss of uif, including early larval
lethality, slow growth, developmental arrest and failure to pupariate.
Most uif mutant larvae die as first instars, and those that die
early almost invariably have the most severe tracheal inflation defects.
uif mutant larvae that survive to second or third instar grow
much slower than their heterozygous siblings.
btl>uifRNAi larvae that have only lost uif
function in their tracheae show an identical slow growth phenotype. This
study consistently observed uif mutant larvae wandering away
from food, suggesting that they were experiencing hypoxia. Not
surprisingly these larvae accumulate less fat and have a transparent
appearance. Examination of the tracheae in these mutant animals revealed
that the inflated portion of the trachea was often just through the
first instar tracheal lumen, and therefore these animals were likely
oxygen starved as well. Consistent with this notion, additional tracheal
branching was observed in these mutant animals suggesting that the
hypoxia induced factor pathway had been engaged. Thus, hypoxia could
explain the growth defects observed in uif mutant larvae. These
growth defects may have then resulted in the observed developmental
delays. For example, mutant third instar larvae may not have reached a
critical weight threshold needed for pupariation, and thus would not
have been able to pupariate even if they experienced the metamorphic
pulse of ecdysone. Interestingly, many uif mutant animals even
failed to advance to third instar, but this did not reflect a defect in
epidermal molting or ecdysis, as no evidence was found for an extra set
of head skeleton or epidermal cuticle. Rather the animals just arrested
as first or second instars. It is possible that hypoxia-induced growth
defects contributed to this phenotype as well, since larvae have to
attain a critical size to be competent for molting, just as they do for
pupariation (Zhang, 2009).
Organ size typically increases dramatically during juvenile growth. This growth presents a fundamental tension, as organs need resiliency to resist stresses while still maintaining plasticity to accommodate growth. The extracellular matrix (ECM) is central to providing resiliency, but how ECM is remodeled to accommodate growth is poorly understood. This study investigated remodeling of Drosophila respiratory tubes (tracheae) that elongate continually during larval growth, despite being lined with a rigid cuticular ECM. Cuticle is initially deposited with a characteristic pattern of repeating ridges and valleys known as taenidia. For tubes to elongate, this study found that the extracellular protease Mmp1 is required for expansion of ECM between the taenidial ridges during each intermolt period. Mmp1 protein localizes in periodically spaced puncta that are in register with the taenidial spacing. Mmp1 also degrades old cuticle at molts, promotes apical membrane expansion in larval tracheae, and promotes tube elongation in embryonic tracheae. Whereas work in other developmental systems has demonstrated that MMPs are required for axial elongation occurring in localized growth zones, this study demonstrates that MMPs can also mediate interstitial matrix remodeling during growth of an organ system (Glasheen, 2010).
This study found that larval tracheal tubes elongate their apical matrix at discrete sites along the long axis of the tube. Immediately after a molt in wild-type animals, the new apical matrix (cuticle) is constructed with taenidial ridges at a fixed interval of ~ 0.8 µm. During intermolt tube elongation, this matrix expands the taenidial interval to ~ 1.6 µm. At molting, this fully expanded matrix is discarded and replaced with a new matrix with a taenidial interval again at 0.8 µm, which will again expand about two-fold. This precise remodeling of the taenidia is accomplished by the extracellular protease Mmp1, which is localized in discrete apical puncta, each associated with an individual taenidium. In Mmp1 mutants, the taenidial ridges do not expand, the taenidial interval remains fixed, and larvae cannot elongate their tracheae as their bodies elongate, causing stretched tubes that eventually break. In normal tube elongation, ECM expansion is coupled with cellular apical membrane expansion, and Mmp1 is required for the coordination of ECM and cellular expansion; Mmp1 is able to promote both aspects of tube expansion when overepressed in embryos. Mmp1 is also required for the other important tracheal cuticle remodeling event in larvae: degrading cuticle into pieces that can be discarded at each molt. Thus, Mmp1 tracheal tubes cannot elongate because they cannot remodel apical extracellular matrix either to expand it or to discard it (Glasheen, 2010).
On first inspection, it is difficult to account for the progressively deteriorating phenotype of Mmp1 mutants. Although the tracheal system appears morphologically normal at hatching in Mmp1 null mutants, within several hours, they develop taut stretched tracheal systems. As has been previously reported, this phenotype worsens throughout larval life: by second instar, many animals have broken dorsal trunks and some death occurs; by third instar, nearly all animals have tracheal breaks and all eventually die (Page-McCaw, 2003). This presents an apparent paradox as early third instar mutants, with shortened tracheal systems, have normal spacing of their taenidia. This paradox is resolved, however, when one considers that in each instar taenidial expansion is a requirement for tube elongation. Thus, at the start of second instar, although the taenidia are correctly spaced, they comprise a tube that is already shorter than wild-type and is virtually unexpandable, and thus, tubes frequently break as the animal grows. By third instar, although the taenidia are again deposited with normal spacing, the collective failures of elongation in the previous instars make the entire tracheal system very short and highly abnormal. Taken alone, the taenidial expansion data might suggest that the Mmp1 tubes fail to elongate at all. If Mmp1 mutant tubes were unable to achieve any elongation, then the tracheal system of third instar larvae would be ~ 1/8 the length of wild-type controls; it was observed that an Mmp1 tracheal segment is about 1/4 the length of wild-type. Thus, despite the lack of taenidial expansion, there appears to be some kind of other elongation at work in these mutant tubes. Possibilities include an aberrant brute-force stretching of unremodeled cuticle, or a burst of tube elongation during molts when the tube releases cuticle (Glasheen, 2010).
In electron micrographs of late Mmp1 mutants, the cuticle appears to separate from the epithelial layer in late Mmp1 samples, but not in wild-type; the explanation of artefactual sample fracturing is unlikely, as cell membranes appeared intact in TEMs from both genotypes. Two models were envisioned to explain this separation. One possibility is that matrix components are still secreted by the mutant cells, but they require matrix remodeling to become incorporated into cuticle, and so they accumulate between the cells and the unremodeled cuticle, creating a gap. They would have to be electron-transparent to be consistent with the images, and there are reports of electron-transparent cuticle layers in insects; indeed, the taenidial cores of the TEMs can appear electron-transparent. Alternatively, it is possible that Mmp1 is required for processing adhesion molecules that hold the cuticle to the cell layer, so that adhesion is lost in the absence of the Mmp1. The first model is favored since it accounts for the fate of the matrix components that should have been deposited in the cuticle in the absence of remodeling (Glasheen, 2010).
An important question arising from this study is how MMP localization and activation is controlled to produce uniform tube elongation. One simple model is that the cells can sense tension and respond by secreting or activating Mmp1. However, this model is not supported by the observation that Tubby (Tb) mutants, with slack in the tracheal tubes, still increase the intertaenidial distance during the intermolt period. Thus, the hypothesis is favored that tube elongation is under developmental regulation. The Tb mutant phenotype indicates that such a tube elongation program is independent of body elongation. One possible mechanism is that the developmental program could trigger Mmp1 secretion from cytoplasmic vesicles to the apical ECM; how MMP secretion is controlled is an open question. The protease appears to be localized in extracellular puncta precisely coordinated with the taenidia. This localization pattern may be ultimately directed by the actin cytoskeleton, which appears to pattern the taenidia during new cuticle secretion. Consistent with the possibility of actin localizing Mmp1, the actin cytoskeleton generally regulates apical secretion in the embryonic tracheal system; and in cultured neurons, it has been observed that MMP-containing vesicles traffic along microfilaments. Mmp1 mutants that lack a hemopexin domain, or dominant-negative mutants that interfere specifically with the Mmp1 hemopexin domain, are still able to elongate their tubes, indicating that the hemopexin domain is not required for secretion, localization, or activation of Mmp1 in these discrete puncta (Glasheen, 2010).
The interstitial remodeling of tracheal cuticle stands in contrast to other developmental mechanisms of ECM deposition and remodeling that occur during axial growth of rigid or load-bearing structures. During vertebrate long-bone growth, elongation takes place at the growth plates near the ends of the bones, concentrating ECM remodeling and deposition distally. In plants, axial elongation takes place at the meristem regions, which are located distally like growth plates, again confining ECM deposition to distal regions. These cases of spatially restricted remodeling occur in contexts where maintaining the integrity of a rigid ECM presents a structural hurdle to simultaneous remodeling. Interestingly, MMPs appear to be required for both these kinds of growth. Mouse MMP13 mutants cannot remodel the cartilage at the growth plate to make bone, and MMP9 is also required for normal long-bone growth. In Arabidopsis, the MMP mutant At2-mmp1 cannot extend shoots, and in the Loblolly pine, MMP expression is correlated with embryonic root (radicle) protrusion, which is hindered by an MMP inhibitor. In Drosophila larval tracheae, the cuticle is not fully sclerotized and so retains some plasticity, which might be expected as the cuticle does not provide rigidity along the axis but instead protects circumferentially from crushing forces. This different set of structural requirements probably explains why in tracheae deposition of matrix is not confined to distal regions or even to one region per segment, but rather is dispersed across the length of the tissue. The requirement for tube rigidity in the circumferential axis, but not along the body axis, also addresses why tracheal elongation can occur continually, modifying an existing cuticle. In contrast, tube circumferential expansion cannot occur continually but is limited to the molt, when the cuticle is completely replaced. The importance of MMPs in axial growth is underscored by the common requirement for MMPs during all these cases, despite the different structural contexts for matrix remodeling in bone elongation, plant growth, and tracheal elongation (Glasheen, 2010).
In Mmp1 mutants, where the cuticle remains unelongated, it is striking that elongation of the underlying cells and their apical membranes is also impaired. Although there is some excess of apical membrane in Mmp1 mutants, the excess is much less than expected had the cells underlying the cuticle expanded their membranes to the wild-type extent. These results suggest that either Mmp1 activity or cuticle elongation is required for cells and apical membranes to elongate normally (Glasheen, 2010).
How are Mmp1 activity and/or cuticle remodeling coordinated with membrane expansion? Although it is possible that matrix remodeling and membrane expansion represent distinct activities of Mmp1, the simplicity of having a direct causal relationship between matrix remodeling and membrane expansion is appealing. One example of a combined model is that extracellular matrix elongation, mediated by Mmp1, places cells under tension, and cells respond by elongating apical membrane. Another model is that Mmp1 proteolytic activity, which remodels cuticle, may simultaneously generate an inductive signal (or inactivate a negative regulator) that causes the underlying cells to elongate, thus coordinating the elongation of both the rigid cuticle and the underlying cells. Production of such a signal would be analogous to mammalian MMPs cleaving laminin-5 or collagen IV to unmask cryptic signaling sites that promote cell migration; this model would also be consistent with recent findings that release of a collagen IV domain by MT2-MMP is required for branching morphogenesis of the submandibular gland in mice. Consistent with Mmp1 activity generating a cuticle-derived signal, misexpression of Mmp1 in embryonic tracheae causes tube elongation only when cuticle is present (Glasheen, 2010).
These results show that extracellular matrix remodeling is a critical aspect of tube elongation in larval tracheae. Mmp1 mutants cannot remodel cuticle and cannot properly elongate their tubes or degrade unnecessary matrix material to be discarded at molts. These remodeling events are regulated separately from the initial deposition and patterning of cuticle, as Mmp1 mutants are able to secrete normally patterned cuticles. Additionally, Mmp1 appears to regulate cell elongation, perhaps directly by processing a signaling molecule, or indirectly by regulating the ECM. Hence, a matrix metalloproteinase appears to act as a critical coregulator of matrix and cellular growth. Finally, it is significant that tracheal remodeling but not initial tracheal morphogenesis requires a matrix metalloproteinase. This analysis of this remodeling phenotype reinforces the notion that matrix metalloproteinases are specialists for remodeling existing tissues, rather than forming tissues, likely because of the need to alter existing ECM that limits plasticity (Glasheen, 2010).
The Drosophila Air Sac Primordium (ASP) has emerged as an important structure where cellular, genetic and molecular events responsible for invasive behavior and branching morphogenesis can be studied. This report presents data which demonstrate that a Cathepsin-L encoded by the gene Cysteine proteinase-1 (CP1) in Drosophila is necessary for invasive behavior during ASP development. CP1 is expressed in ASP and knockdown of CP1 results in suppression of migratory and invasive behavior observed during ASP development. It was further shown that CP1 possibly regulates invasive behavior by promoting degradation of Basement Membrane. These data provide clues to the possible role of Cathepsin L in human lung development and tumor invasion, especially, given the similarities between human lung and Drosophila ASP development (Dong, 2015).
To characterize the cellular mechanisms of subcellular tube formation, this study refined methods of high pressure freezing/freeze substitution to prepare Drosophila larvae for transmission electron microscopic (TEM) analysis. Using these methods, it was found that subcellular tracheal tube formation may proceed through a previously undescribed multimembrane intermediate composed of vesicles bound within a novel subcellular compartment. Correlative light/TEM procedures were developed to identify labeled cells in TEM-fixed larval samples. Using this technique, it was found that the vacuolar ATPase (V-ATPase) and the V-ATPase regulator Rabconnectin-3 are required for subcellular tube formation, probably in a step resolving the intermediate compartment into a mature lumen. In general, these methods should be applicable to analyzing the many cell biological problems which can be addressed using Drosophila larvae (Nikolova, 2015).
Tubular networks are central to the structure and function of many organs, such as the vertebrate lungs or the Drosophila tracheal system. Their component epithelial cells are able to proliferate and to undergo complex morphogenetic movements, while maintaining their barrier function. Little is known about the details of the mitotic process in tubular epithelia. This study presents a comprehensive model of cellular remodeling and proliferation in the dorsal branches of third-instar Drosophila larvae. Through a combination of immunostaining and novel live imaging techniques, this study identified the key steps in the transition from a unicellular to a multicellular tube. Junctional remodeling precedes mitosis and, as the cells divide, new junctions are formed through several variations of what is refered to as 'asymmetric cytokinesis'. Depending on the spacing of cells along the dorsal branch, mitosis can occur either before or after the transition to a multicellular tube. In both instances, cell separation is accomplished through asymmetric cytokinesis, a process that is initiated by the ingression of the cytokinetic ring. Unequal cell compartments are a possible but rare outcome of completing mitosis through this mechanism. The Dpp signaling pathway was found to be required but not sufficient for cell division in the dorsal branches (Denes, 2015).
The terminal cells of the larval Drosophila tracheal system extend dozens of branched cellular processes, most of which become hollow intracellular tubes that support gas exchange with internal tissues. Previously, a forward genetic mosaic screen was undertaken to uncover the pathways regulating terminal cell size, morphogenesis, and the generation and maintenance of new intracellular tubes. This initial work identified several mutations affecting terminal cell size and branch number, and suggested that branch complexity and cell size are typically coupled but could be genetically separated. To deepen understanding of these processes, the molecular identities was undertaken of mutations in the genes sprout, denuded and asthmatic, that had been implicated in the initial screen. This paper reveals the molecular identity of these genes and describes their function in the context of the TOR and Hippo pathways, which are widely appreciated to be key regulators of cell and organ size (Burguete, 2019).
Seamless tubes form intracellularly without cell-cell or autocellular junctions (see Labarsky, 2003). Such tubes have been described across phyla, but remain mysterious despite their simple architecture. In Drosophila, seamless tubes are found within tracheal terminal cells, which have dozens of branched protrusions extending hundreds of micrometres. This study has found that mutations in multiple components of the dynein motor complex block seamless tube growth, raising the possibility that the lumenal membrane forms through minus-end-directed transport of apical membrane components along microtubules. Growth of seamless tubes is polarized along the proximodistal axis by Rab35 and its apical membrane-localized GAP, Whacked. Strikingly, loss of whacked (or constitutive activation of Rab35) leads to tube overgrowth at terminal cell branch tips, whereas overexpression of Whacked (or dominant-negative Rab35) causes formation of ectopic tubes surrounding the terminal cell nucleus. Thus, vesicle trafficking has key roles in making and shaping seamless tubes (Schottenfeld-Roames, 2013).
Three tube types -- multicellular, autocellular and seamless -- are found in the Drosophila trachea. Most tracheal cells contribute to multicellular tubes or make themselves into unicellular tubes by wrapping around a lumenal space and forming autocellular adherens junctions, but two specialized tracheal cell types, fusion cells and terminal cells, make 'seamless' tubes. How seamless tubes are made and how they are shaped are largely unknown. One hypothesis holds that seamless tubes are built by 'cell hollowing', in which vesicles traffic to the centre of the cell and fuse to form an internal tube of apical membrane, whereas an alternative model proposes that apical membrane is extended internally from the site of intercellular adhesion. In both models, transport of apical membrane would probably play a key role. As terminal cells make seamless tubes continuously during larval life, they serve as an especially sensitive model system in which to dissect the genetic program (Schottenfeld-Roames, 2013).
Tracheal cells are initially organized into epithelial sacs with their apical surface facing the sac lumen. During tubulogenesis, γ-tubulin becomes localized to the lumenal membrane of each tracheal cell, generating microtubule networks oriented with minus ends towards the apical membrane. Terminal and fusion cells are first selected as tip cells that undergo a partial epithelial-to-mesenchymal transition and initiate branching morphogenesis: they lose all but one or two cell-cell contacts and become migratory. Branchless-FGF signalling induces a subpopulation of tip cells to differentiate as terminal cells. During larval life, terminal cells ramify on tissues spread across several hundred micrometres, with branching patterns that reflect local hypoxia. A single seamless tube forms within each branched extension of the terminal cell (Schottenfeld-Roames, 2013).
How trafficking contributes to seamless tube morphogenesis is unknown. Despite clues that vesicle transport plays a role in the genesis of seamless tubes, the tube morphogenesis genes remain elusive. This study characterized the cytoskeletal polarity of larval terminal cells, shows that a minus-end-directed microtubule motor complex is required for seamless tube growth, and characterizes mutations in whacked (wkd) that uncouple seamless tube growth from the normal spatial cues. Sequence analysis indicates that wkd encodes a RabGAP, and it was shown that Rab35 is the essential target of Wkd, and that together, Wkd and Rab35 can polarize the growth of seamless tubes (Schottenfeld-Roames, 2013).
Apical-basal polarity and cytoskeletal organization was examined in mature larval terminal cells. The lumenal membrane was decorated by puncta of Crumbs, a definitive apical membrane marker. Actin filaments were found enriched in three distinct subcellular domains: surrounding seamless tubes, decorating filopodia and outlining short stretches of basolateral membrane. The microtubule cytoskeleton also seemed polarized, with γ-tubulin lining the seamless tubes and enriched at tube tips. These data are consistent with tracheal studies in the embryo. EB1::GFP analyses of growing (plus-end) microtubules demonstrated that some are oriented towards the soma and others towards branch tips. Stable acetylated microtubules ran parallel to the tubes and extended beyond the lumen at branch tips where they may template tube growth. Consistent with such a role, microtubule-tract-associated fragments of apical membrane were observed distal to the blind ends of the seamless tubes. Filopodia extended past the stable microtubules as expected. These data indicate that mature terminal cells maintain the polarity and organization described for embryonic terminal cells. On the basis of γ-tubulin localization, it is inferred that a subset of microtubules is nucleated at the apical membrane, and that apically targeted transport along such microtubules would require minus-end motor proteins. Indeed, homozygous mutant Lissencephaly-1. As γ-tubulin lines the entire apical membrane, growth through minus-end-directed transport might be expected to occur all along the length of seamless tubes, and indeed, a pulse of CD8::GFP (transmembrane protein tagged with GFP) synthesis uniformly labelled the apical membrane as it first became detectable (Schottenfeld-Roames, 2013).
The cytoplasmic dynein motor complex drives minus-end-directed transport of intracellular vesicles in many cell types; to test for its requirement in seamless tube formation, terminal cells were examined mutant for any of four dynein motor complex genes: Dynein heavy chain 64C (Dhc64C), Dynein light intermediate chain (dlic), Dynactin p150 (Glued) and Lis-1. Mutant terminal cells showed a cell autonomous requirement for these genes. Mutant terminal cells had thin cytoplasmic branches that lacked air-filling, and antibody staining revealed that seamless tubes did not extend into these branches although acetylated microtubules often did. It was also noted that formation of filopodia at branch tips is disrupted in dynein motor complex mutants, which may account for the decreased number of branches in mutant terminal cells. Ectopic seamless tubes that were not air-filled were detected near the nucleus, as described below. Interestingly, discontinuous apical membrane fragments (similar to those in Lis-1 embryos) were found in terminal branches lacking seamless tubes, and were associated with microtubule tracts. Whereas γ-tubulin was enriched on truncated tubes and on these presumptive seamless tube intermediates, diffuse γ-tubulin staining was detected throughout the mutant cells, indicating that assembly of apical membrane is required to establish or maintain γ-tubulin localization. Likewise, Crumbs seemed reduced and aberrantly localized. Reduced levels of acetylated microtubule staining in these cells may reflect loss of apical γ-tubulin. Importantly, these data show that stable microtubules extend through cellular projections that lack seamless tubes. Thus, without minus-end-directed transport, stable microtubules are insufficient to promote seamless tube formation, but stable cellular projections are formed and maintained in the absence of seamless tubes (Schottenfeld-Roames, 2013).
In contrast to these defects in seamless tube generation, mutations in wkd confer overly exuberant tube growth. Examination of wkd terminal cell tips revealed a 'U-turn'; phenotype in which seamless tubes executed a series of 180 degree turns - the possibility is entertained that branch retraction, similar to that observed in talin mutants, could contribute to the U-turn defect (Schottenfeld-Roames, 2013).
Homozygous wkd animals survived until pharate adult stages, and, other than the seamless tube defects, had normal tracheal tubes at the third larval instar. Mosaic analysis revealed a terminal cell autonomous requirement for wkd. Mutant clones in multicellular tubes, and in unicellular tubes that lumenize by making autocellular adherens junctions, were of normal morphology. Strikingly, fusion cells, which also form seamless tubes, were unaffected by loss of wkd (Schottenfeld-Roames, 2013).
To determine the molecular nature of wkd, a positional cloning approach was taken. Mapping techniques defined a candidate gene interval of ~ 75 kilobases (kb). Focused was placed on CG5344 as it encodes a protein containing a TBC (Tre2/Bub2/Cdc16) domain characteristic of Rab GTPase-activating proteins, and hence was likely to participate in vesicular trafficking, a process that could lie at the heart of seamless tube formation. A single nucleotide change was identified that resulted in mis-sense (PC24) and non-sense (220) mutations in the CG5344 coding sequence. Pan-tracheal knockdown of wkd by RNA-mediated interference (RNAi) caused terminal cell-specific U-turn defects (other defects characteristic of the ethyl methanesulfonate (EMS)-induced alleles of wkd were detected at a low frequency). A genomic rescue construct for CG5344 rescued wkd mutants, confirming gene identity. On the basis of these results, it is concluded that wkd is CG5344 and that it probably regulates vesicular trafficking during seamless tube morphogenesis (Schottenfeld-Roames, 2013).
To determine the Rab target(s) of Wkd regulation, whether tracheal expression of constitutively active 'GTP-locked'; Rab isoforms (henceforth, RabCA) might phenocopy wkd was investigated. RabCA for 31 of the 33 Drosophila Rabs were tested individually in the tracheal system. Rab35CA alone conferred terminal-cell-specific U-turns defects (Schottenfeld-Roames, 2013).
To evaluate Wkd overexpression, UAS-wkd was expressed in wild-type animals in a pan-tracheal pattern. Excess Wkd caused formation of ectopic seamless tubes surrounding the terminal cell nucleus. At higher levels of expression, small spheres of apical membrane were found adjacent to the nucleus and less abundantly at more distal sites. Consistent with Wkd regulation of vesicle trafficking by modulation of Rab35, expression of a dominant-negative Rab35 (henceforth, Rab35DN) caused formation of ectopic proximal tubules (Schottenfeld-Roames, 2013).
Attempts were made to determine whether Rab35 was the essential target of Wkd GAP activity. Wkd primary structure is equally conserved in three human RabGAPs. All three act as Rab35GAPs, although each has been proposed to have additional targets. To further determine if Wkd acts as a Rab35GAP, whether Rab35DN could suppress wkd mutants was examined; tracheal-specific expression of Rab35DN strongly suppressed the 'U-turn'; defects of wkd-null animals and, surprisingly, also rescued the lethality of wkd. Since mutant Rab35 isoforms phenocopy wkd gain and loss of function, Rab35DN bypasses the requirement for wkd and human Wkd orthologues are Rab35GAPs, it is concluded that the critical function of Wkd is as a GAP for Drosophila Rab35 (Schottenfeld-Roames, 2013).
In other systems Rab35 is implicated in polarized membrane addition to plasma membrane compartments - for example, immune synapse, cytokinetic furrow and so on - or, in actin regulation. A role for actin in fusion cell seamless tube formation has been proposed, so whether Wkd and Rab35 act by modulation of the terminal cell actin cytoskeleton was examined. As the actin-bundling protein Fascin (Drosophila singed) was recently identified biochemically as a Rab35 effector, a role of singed in terminal cell tubes was examined, but found no evidence was found for one. Furthermore, overexpression of Wkd, or of Rab35DN, did not significantly alter the terminal cell actin cytoskeleton, leading to the conclusion that actin regulation is not a primary function of Wkd/Rab35 during seamless tube morphogenesis (Schottenfeld-Roames, 2013).
The alternative model -- that Rab35 acts in polarized membrane addition -- was found to be attractive, because extra Rab35-GTP activity promoted seamless tube growth at branch tips whereas depletion of Rab35-GTP promoted tube growth at the cell soma. To test this model, advantage was taken of the information that expression of an activated Breathless-FGFR (lambdaBtl in terminal cells induces robust growth of ectopic seamless tubes surrounding the nucleus; whether growth of the ectopic tubes could be redirected from the soma to the branch tips was investigated by eliminating wkd. The activated FGFR phenotype was not altered in wkd heterozygotes, but in wkd mutant animals (or wkd-RNAi animals) the site of ectopic seamless tube growth was strikingly different. In some cells, extra tubes were found throughout the cell - in the soma and at branch tip - whereas in others extra tubes were present only at the branch tip. Thus, the position of seamless tube growth is dependent on Wkd activity, although Wkd itself is not essential for tube formation. These data provide evidence against branch retraction (as occurs in talin mutants) as the mechanism for generating a U-turn phenotype, because branch retraction would not redirect ectopic tube growth (Schottenfeld-Roames, 2013).
To better understand how Wkd and Rab35 determined the site of seamless tube growth, their subcellular distribution was examined. Pan-tracheal expression of mKate2-tagged Wkd (Wkd::mKate2) rescued wkd-null animals. The steady-state subcellular localization of Wkd::mKate2 was restricted to the lumenal membrane with higher accumulation at the growing tips of seamless tubes. At lower levels, cytoplasmic puncta of Wkd::mKate2 were noted that could reflect vesicular localization, as well as labeling of filopodia. It was found that YFP::Rab35 was distributed in a diffuse pattern throughout the terminal cell cytoplasm with some apical enrichment, and notable localization to filopodia. Substantial co-localization of Wkd::mKate2with YFP::Rab35 was found at the apical membrane, in cytoplasmic puncta, and in filopodia. Among endosomal Rabs, Rab35 seemed uniquely abundant within filopodia, and showed the greatest overlap with Wkd at the apical membrane. Substantial overlap was noted between Wkd/Rab35 and acetylated microtubules, including at positions distal to the blind end of seamless tubes. The enrichment of Wkd along seamless tubes indicates that Rab35 functions in an apical membrane trafficking event, leading to the speculation that recycling endosomes at filopodia might be targeted to the growing seamless tube by minus-end motor transport (Schottenfeld-Roames, 2013).
In a similar vein, it is speculated that vesicles might be transported from the soma towards branch tips in a process regulated by Wkd and Rab35. Disruption of such transport might explain why overexpression of Wkd leads to ectopic seamless tube growth in the soma. Whether Wkd::mKate2 localization was compromised in dynein motor complex mutants was examined. As these cells have branches that lack apical membrane/seamless tubes, disruption in the localization pattern of Wkd was anticipated, but it was wondered whether co-localization with acetylated tubulin would be intact, indicative of a microtubule association independent of dynein motor transport. It was found that Wkd::mKate2 is broadly distributed throughout the cytoplasm of dynein motor complex mutants, and does not show enrichment on acetylated microtubule tracts; indeed, substantial basal enrichment was detected of Wkd::mKate2. If Wkd/Rab35-dependent trafficking of apical vesicles was dynein motor complex dependent, ectopic seamless tubes would be expected in the soma of dynein motor complex mutants, similar to those seen with Wkd overexpression or expression of Rab35DN. In fact, such ectopic tubes were consistently found in the dynein motor complex mutants, consistent with dynein-dependent trafficking of Rab35 vesicles. It cannot be ruled out that these defects are due to dynein-dependent processes unrelated to Wkd and Rab35; however, whether the ectopic tubes could be redirected distally by expression of Rab35CA, or elimination of Wkd, was examined. The motor complex ectopic tube phenotype could not be altered, indicating that the phenotype does not arise as an indirect consequence of altered Wkd localization or Rab35 activity (Schottenfeld-Roames, 2013).
The roles of RabGAP proteins have started to become clear only in recent years. Historically, it has been difficult to determine which Rab proteins are substrates of specific RabGAPs. Tests of in vitro GAP activity produced conflicting results, and in some cases did not seem indicative of in vivo function. Indeed, the specificity of Carabin (also known as Wkd orthologue TBC1D10C) has been controversial: it was first shown to act as a RasGAP, whereas later studies indicate a Rab35-specific GAP activity. The in vivo genetic data for wkd, together with recent studies characterizing the function of all three human Wkd-like TBC protein, make a compelling case that this family of proteins acts as GAPs for Rab35. Furthermore, this study establishes a role for classical vesicle trafficking proteins in seamless tube growth. As seamless tubes, but not multicellular or autocellular tracheal tubes, are affected by mutations in wkd and Rab35, this study also establishes an in vivo cell-type-specific requirement for trafficking genes in tube morphogenesis (Schottenfeld-Roames, 2013).
It is concluded that Wkd and Rab35 regulate polarized growth of seamless tubes, and it is speculated that Wkd and Rab35 direct transport of apical membrane vesicles to the distal tip of terminal cell branches (when equilibrium is shifted towards active Rab35-GTP), or to a central location adjacent to the terminal cell nucleus (when equilibrium is shifted towards inactive Rab35-GDP). Analogous to its previously described roles in targeting vesicles to the immune synapse in T cells, the cytokinetic furrow in Drosophila S2 cells and the neuromuscular junction in motor neurons, Rab35 would promote transport of vesicles from a recycling endosome compartment to the apical membrane. It is further speculated that Breathless-FGFR activation at branch tips may couple terminal cell branching with seamless tube growth within that new branch (Schottenfeld-Roames, 2013).
The Drosophila tracheal system
is a branched tubular network that forms in the embryo
by a post-mitotic program of morphogenesis. In third instar larvae (L3),
cells constituting the second tracheal metamere (Tr2) reenter the cell
cycle. Clonal analysis of L3 Tr2 reveals that dividing cells in the dorsal
trunk, dorsal branch and transverse connective branches respect lineage
restriction boundaries near branch junctions. These boundaries
corresponded to domains of gene expression, for example where cells
expressing Spalt, Delta
and Serrate in the dorsal trunk meet
vein-expressing cells in the dorsal branch or transverse connective. Notch
signaling was activated to one side of these borders and was required for
the identity, specializations and segregation of border cells. These
findings suggest that Tr2 is comprised of developmental compartments and
that developmental compartments are an organizational feature relevant to
branched tubular networks (Rao, 2015).
Epithelial remodelling is an essential mechanism for organogenesis, during which cells change shape and position while maintaining contact with each other. Adherens junctions (AJs) mediate stable intercellular cohesion but must be actively reorganised to allow morphogenesis. Vesicle trafficking and the microtubule (MT) cytoskeleton contribute to regulating AJs but their interrelationship remains elusive. This study reports a detailed analysis of the role of MTs in cell remodelling during formation of the tracheal system in the Drosophila embryo. Induction of MT depolymerisation specifically in tracheal cells showed that MTs were essential during a specific time frame of tracheal cell elongation while the branch extended. In the absence of MTs, one tracheal cell
per branch overelongated, ultimately leading to branch break.
Three-dimensional quantifications revealed that MTs were crucial to
sustain E-Cadherin (Shotgun) and
Par-3 (Bazooka) levels at AJs.
Maintaining E-Cadherin/Par-3 levels at the apical domain required de
novo synthesis rather than internalisation and recycling from and to
the apical plasma membrane. However, apical targeting of E-Cadherin
and Par-3 required functional recycling endosomes, suggesting an
intermediate role for this compartment in targeting de novo
synthesized E-Cadherin to the plasma membrane. The apical enrichment
of recycling endosomes was dependent on the MT motor Dynein and essential for the
function of this vesicular compartment. In addition, E-Cadherin
dynamics and MT requirement differed in remodelling tracheal cells versus planar epithelial cells. Altogether, these results uncover an MT-Dynein-dependent apical restriction of recycling endosomes that controls adhesion by sustaining Par-3 and E-Cadherin levels at AJs during morphogenesis (Le Droguen, 2015).
This study has reveal the importance of the functional interplay between MTs, vesicular trafficking and the control of AJ dynamics in cells undergoing extensive remodelling through collective migration. MT depletion in tracheal cells induces the formation of intracellular dots containing E-Cad and Par-3. Interestingly, these intracellular accumulations are only seen in remodelling tracheal cells and not in planar epithelia. These dots are still detected when endocytosis is affected, showing that they are de novo synthesis route intermediates. Altogether, this suggests that maintaining the correct level of E-Cad/Par-3 at the apical domain requires a continuous supply of newly synthesized proteins, which could be essential for the intensive AJ reorganisation that occurs during cell intercalation and elongation of the tracheal branch (Le Droguen, 2015).
Using the photo-convertible E-Cad-EosFP in flat epithelium, a previous study showed that E-Cad that is engaged in homophilic interactions at the AJs forms very stable domains. This study has demonstratde that MT depletion does not affect the integrity of this 2D epithelium. In addition, it was shown that the pool of photo-converted E-Cad-EosFP is less stable in tracheal cells than in epidermal cells. Together with a FRAP assay suggesting that AJs are more dynamic in tracheal cells than in epithelial cells, the results highlight a specific fine-tuning of AJ components in tracheal cells undergoing cell movement and cell shape changes in 3D through cell intercalation, cell elongation and thereby organ formation. This fine-tuning is likely to cycle between internalisation, recycling, degradation and de novo synthesis, the latter being MT dependent. When the balance is altered in the absence of MTs, and thus when E-Cad or Par-3 are reduced at AJs, tracheal cells overelongate after completing intercalation. During branch elongation without MTs, the two migrating leading cells generate a pulling force on the following stalk cells, which display a critical reduction in AJ components. Either the tip cell or the base cell of the stalk becomes unable to maintain its integrity in response to this force. Consequently, this tracheal cell overelongates by a factor 1.8, preventing the remaining stalk cells from reaching their average size. As a result, DBs present a single overelongated cell and several underelongated cells (Le Droguen, 2015).
An overlap was detected between the E-Cad intracellular dots generated in the absence of MTs and recycling endosome vesicles. Interfering with Rab11 function in tracheal cells induces cell overelongation and affects E-Cad and Par-3 distribution at the AJs, as does MT depletion. These results illustrate that the E-Cad de novo synthesis pathway passes through the Rab11-positive recycling endosome compartment. The overlap of E-Cad and recycling endosome markers represents only a small proportion of the total intracellular E-Cad, suggesting a transient residence in this vesicular compartment for this newly synthesized protein. Recent studies conducted in different model systems have revealed that some newly synthesized apical plasma membrane proteins, such as E-Cad and Rhodopsin, leave the trans-Golgi network to cross Rab11-positive recycling endosome compartments before reaching the apical surface). This apical trafficking route is used specifically in tracheal cells and requires the MT network. Quantification of Rab11DN-associated defects upon tracheal branch formation reveals that impairing recycling endosome function has a similar effect to altering the distribution of E-Cad at AJs by depleting the MT network. Moreover, interfering with Rab11 function reduces Par-3 levels at the AJs of tracheal cells. Interestingly, Par-3 does not colocalise with the E-Cad cytoplasmic pool, indicating that functional recycling endosomes are required by Par-3 and E-Cad to assemble as a complex and to be targeted to the apical domain of tracheal cells. However, as E-Cad and Par-3 can be apically targeted in the absence of the other, this suggests that apical targeting of E-Cad and Par-3 can be independent in tracheal cells or that redundant pathways could sustain the localisation of each protein. For example, the Nectin protein Echinoid is required for Par-3 localisation at AJs in shg mutant cells. Moreover, the apical distribution of PI(4,5)P2 in the Drosophila follicular epithelium sustains Par-3 apical anchorage at the plasma membrane. Furthermore, Par-3-independent localisation of E-Cad has been observed (Le Droguen, 2015).
Dynamic MTs in the ectoderm locally upregulate AJ turnover through RhoA activity. RhoA stabilises cellular contacts through acto-myosin regulation. In tracheal cells, MT depletion does not alter actin distribution. Moreover, tracheal cells mutant for the Myosin light chain zipper (zip) and also expressing a dominant-negative form of Zip do not present obvious defects in E-Cad distribution at stage 14. By contrast, MT depletion induces the cytoplasmic accumulation of E-Cad and Par-3 in tracheal cells only and not in ectodermal cells at the same developmental stage. Thus far, cytoplasmic accumulation of E-Cad and Par-3 has only been observed after colchicine-induced MT depolymerisation during polarity establishment at embryo cellularisation, when AJs are extremely dynamic and vesicular trafficking is strongly active. This study demonstrated that the E-Cad distribution in tracheal cells is more sensitive to MT depolymerisation and to Rab11DN overexpression than that in the overlying ectodermal cells. The comparison of the maximum recovery of the E-Cad signal in a FRAP assay in tracheal cells and in ectodermal cells, together with the differences in stability of the photo-converted E-Cad-EosFP between these two tissues, suggest that AJs are more dynamic in tracheae. It is thus conceivable that tracheal cells, which undergo extensive cell shape changes through collective cell migration, require more dynamic AJs as sustained by an efficient targeting of E-Cad. These discrepancies between ectodermal and tracheal epithelia underline the importance of investigating MT function in different morphogenetic contexts under different constraints (Le Droguen, 2015).
This study has demonstrated that the MT minus-end motor Dynein is essential for the restricted localisation of recycling endosomes in a developing organism. The Dynein requirement for the apical enrichment of recycling endosomes is in agreement with the MT minus ends being anchored at the apical plasma membrane. The asymmetric distribution of recycling endosome vesicles has been observed in various differentiated cell types, especially during cell division. Vesicles are found enriched either in the apical domain or at the microtubule-organising centre (MTOC; i.e. the centrosome) during mitosis. Indeed, in vivo, Dynein physically interacts with Nuf. During metaphase, Dynein is required for the maintenance of Nuf at the centrosome. This study demonstrates that Dynein is also required for the apical distribution of recycling endosomes in non-dividing tracheal cells. In a context in which Dynein function is altered, Nuf-positive recycling endosome vesicles are dispersed but colocalise with E-Cad and Par-3 intracellular dots, indicating that the recycling endosome compartment remains functional for the assembly of such a complex (Le Droguen, 2015).
A previous study has characterised the relocalisation of the MTOC in tracheal cells; MTs are nucleated and anchored at the apical domain just above the AJs. It has also been demonstrated that such MT organisation is crucial for tracheal morphogenesis. Non-centrosomal MT organisation occurs in many differentiated cell types but the functional relevance of such an organisation is still poorly understood. It will be informative to investigate whether such non-centrosomal MT organisation provides a means to regulate epithelial remodelling by controlling the apical enrichment of recycling endosomes and thus AJ dynamics (Le Droguen, 2015).
Tubular organs exhibit a striking orientation of landmarks according to the physical anisotropy of the 3D shape, in addition to planar cell polarization. However, the influence of 3D tissue topography on the constituting cells remains underexplored. This study identified a regulatory network polarizing cellular biochemistry according to the physical anisotropy of the 3D tube geometry (tube cell polarization) by a genome-wide, tissue-specific RNAi screen. During Drosophila airway remodelling, each apical cellular junction is equipotent to establish perpendicular actomyosin cables, irrespective of the longitudinal or transverse tube axis. A dynamic transverse enrichment of atypical protein kinase C (aPKC) shifts the balance and transiently targets activated small GTPase RhoA, myosin phosphorylation and Rab11 vesicle trafficking to longitudinal junctions. It is proposed that the PAR complex translates tube physical anisotropy into longitudinal junctional anisotropy, where cell-cell communication aligns the contractile cytoskeleton of neighbouring cells (Hosono, 2015).
The vertebrate tight junction is a critical claudin-based cell-cell junction that functions to prevent free paracellular diffusion between epithelial cells. In Drosophila, this barrier is provided by the septate junction, which, despite being ultrastructurally distinct from the vertebrate tight junction, also contains the claudin-family proteins Megatrachea and Sinuous. This study identified a third Drosophila claudin, Kune-kune, that localizes to septate junctions and is required for junction organization and paracellular barrier function, but not for apical-basal polarity. In the tracheal system, septate junctions have a barrier-independent function that promotes lumenal secretion of Vermiform and Serpentine, extracellular matrix modifier proteins that are required to restrict tube length. As with Sinuous and Megatrachea, loss of Kune-kune prevents this secretion and results in overly elongated tubes. Embryos lacking all three characterized claudins have tracheal phenotypes similar to any single mutant, indicating that these claudins act in the same pathway controlling tracheal tube length. However, there are distinct requirements for these claudins in epithelial septate junction formation. Megatrachea is predominantly required for correct localization of septate junction components, while Sinuous is predominantly required for maintaining normal levels of septate junction proteins. Kune-kune is required for both localization and levels. Double- and triple-mutant combinations of Sinuous and Megatrachea with Kune-kune resemble the Kune-kune single mutant, suggesting that Kune-kune has a more central role in septate junction formation than either Sinuous or Megatrachea (Nelson, 2010).
The Drosophila genome encodes seven predicted claudin-family molecules. Two of these, Mega and Sinu, have previously been characterized and were shown to be required for SJ organization and function. Sequence comparisons indicated that, although all seven claudin-like molecules show a large sequence divergence, CG1298 is more closely related to Sinu and Mega than the other Drosophila claudin-family members. Therefore, although many Drosophila claudin-family members are not required for barrier function, it was reasoned that CG1298 may play a role in SJ paracellular barrier formation. Accordingly, a detailed analysis of CG1298, which was named kune-kune (Japanese for 'wiggling like a snake,' pronounced koon-eh koon-eh and abbreviated kune) was characterized for its tracheal phenotype (Nelson, 2010).
The kune locus contains a single exon that codes for a protein of 264 amino acids. As is characteristic for claudins, the TMpred transmembrane algorith predicts Kune to have four transmembrane domains with intracellular N and C termini, a large initial extracellular loop, and two smaller loops. Kune also contains a W-GLW-C-C motif in the large extracellular loop and a C-terminal PDZ-binding motif, features that are found in almost all claudin family members. Notably, the PDZ-binding motif in Kune is a better match to consensus PDZ-binding motifs than the motif in Sinuous. Furthermore, in contrast to Mega and Sinu, whose N termini are 28 and 38 aa respectively, Kune has a short N terminus of 9 aa that is more typical of vertebrate claudins. Thus, Kune has features that more closely resemble vertebrate claudins than do the so far characterized Drosophila claudins Sinu and Mega (Nelson, 2010).
To determine the expression pattern of Kune, anti-Kune sera were generated and wild-type (WT) embryos were immunized. As with Mega and Sinu, Kune is highly expressed in ectodermally derived tissues, including the epidermis, salivary gland, trachea, hindgut, and foregut beginning at embryonic stage 13. In these tissues, Kune colocalizes with the SJ protein Coracle (Cor) and localizes basal to the adherens junction marker, DE-cadherin (E-cad), suggesting that Kune is a SJ protein. As with many other SJ proteins, Kune is also expressed in glial cells (Nelson, 2010).
Since most SJ proteins show interdependence for correct localization and junction function, it was asked whether localization of Kune depends on the presence of other SJ proteins. Indeed, Kune is mislocalized to more basal positions in the primary epithelia of mega, sinu, cor, and Atpα null mutants, providing strong evidence that Kune is a SJ protein (Nelson, 2010).
To directly assess the function of Kune during development, a PiggyBac insertion, PBac{3HPy}C309, in the 5'-untranslated region (UTR) of kune was identified as a putative null mutation. Embryos homozygous for the kuneC309 chromosome fail to hatch and completely lack Kune protein as assessed by immunohistochemistry. Expression of a UAS-kune construct using the ubiquitous da-Gal4 driver at 28° rescued the embryonic lethality of kuneC309 embryos, demonstrating that lethality was due to loss of Kune. Further, embryos trans-heterozygous for kuneC309 and Df(2R)BSC696 (which deletes the kune locus and also eliminates Kune staining) or homozygous for Df(2R)BSC696 fail to hatch and display tracheal and septate junction phenotypes that are indistinguishable from kuneC309 homozygotes. These results indicate that kune is an essential gene and that kuneC309 is a null or strong loss-of-function allele of kune (Nelson, 2010).
To determine if Kune is required for SJ organization and function, the subcellular localization of several SJ proteins was examined in kune mutant epithelia. As is seen in other SJ mutants, kune epithelia show a reduction and/or mislocalization of the SJ components Cor, Mega, Sinu, Atpα, Discs large (Dlg), and NeurexinIV (Nrx) to more basal locations in all primary epithelia. This phenotype is also seen in animals that express kune-RNAi using the ubiquitous da-Gal4 driver, although the phenotype is less severe, presumably due to incomplete knockdown. Consistent with the immunohistological evidence of SJ defects, a 10-kDa fluorescent dye injected into the body cavity of kune animals readily diffused into the lumen of the trachea and salivary gland, indicating a loss of the paracellular barrier. Expression of the UAS-kune construct with da-Gal4 rescued Cor localization and improved the barrier function of kune mutants. Thus, Kune is an essential component of SJs in primary epithelia (Nelson, 2010).
In addition to their roles in epithelial tissues, SJs are also required to establish the blood-brain barrier in flies. In the central nervous system (CNS), surface glial cells completely ensheath the ventral nerve cord and form SJs at glia-glia contacts. This generates a tight paracellular seal that separates the K+-rich hemolymph from neural cells, which is essential for generation of action potentials. This study found that Kune is expressed in glial cells, which is most clearly seen at the central midline. Dye injections revealed that Kune, like Sinu and Mega, is required for the CNS glial barrier, since the dye penetrated into the nerve cord of kune but not WT embryos. Taken together, the above results all identify Kune as a critical SJ component in multiple tissue types (Nelson, 2010).
Since almost all characterized SJ proteins are required for tracheal tube size control, the tracheal system of kune embryos was examined. Staining with the 2A12 lumenal marker demonstrated that the length of the DT of stage 16 kune embryos was significantly increased over WT controls and appeared tortuous (thus the name kune-kune). This phenotype was identical in both kune/Df(2R)BSC696 embryos and embryos expressing kune-RNAi using the da-Gal4 driver (Nelson, 2010).
It has been established that Sinu, Mega, and other SJ proteins are required for apical secretion of the putative chitin deacetylases Verm and Serp, which restrict tracheal tube length. Therefore lumenal accumulation of Verm and Serp and the organization of the chitin-based lumenal matrix were examined at embryonic stage 16. As is typical for a SJ component, kune mutant embryos and embryos expressing kune-RNAi do not accumulate Verm or Serp in the tracheal lumen. This secretion defect could be largely rescued by expression of UAS-kune with the da-Gal4 driver, although not to WT levels. Additionally, staining with a fluorescent chitin binding probe showed that, while the lumen of WT trachea contains a dense, fibrilar chitin cable, kune trachea have a diffuse, disorganized lumenal matrix. kune trachea also lack the gap between the chitin cable and the apical surface of the cells that is found in WT trachea (Nelson, 2010).
Since Kune is closely related to both Mega and Sinu and all three localize to the SJ, it was asked if these claudins are partially redundant in junction organization or tube size control. To test this, the trachea of mega, kune, and sinu single, double, and triple mutants were examined. If the claudins have redundant functions in tube size control, the phenotypes should be worse when multiple claudins are missing. However, the tracheal length defects of mega; kune and kune; sinu embryos appear no more severe than in any of the single mutants. Strikingly, even embryos lacking all three Kune-related claudins do not appear to have more severe tracheal length defects than single mutants. In contrast, previous work has shown that embryos containing mutations in both sinu and the SJ gene varicose (vari) have trachea that are more tortuous than either single mutant. These results suggest that, although some SJ proteins have redundant functions in restricting tracheal tube length, the Drosophila claudins Kune, Sinu, and Mega all function in the same pathway of tracheal tube size control (Nelson, 2010).
Given that tracheal tube length is only a limited readout of SJ function, the effects of single-, double-, and triple-mutant combinations of kune, sinu, and mega on SJ organization were compared using the subcellular localization of Cor as an assay. Focused was placed on the hindgut and salivary gland, since SJ organization is clearest in these columnar cells. Interestingly, the levels and localization of Cor are strikingly different between the three claudin mutations, suggesting that different claudins have unique functions. For example, Cor is completely mislocalized to basal positions in the hindgut of mega embryos, but the levels are not dramatically lower than in WT. On the other hand, the hindguts of sinu embryos show lower overall levels of Cor, but retain significant apicolateral enrichment where the SJ is normally found. In kune mutants, Cor is both reduced and completely mislocalized. Similar, more pronounced effects are seen in the salivary glands where loss of mega causes only slight basolateral mislocalization of Cor, loss of sinu causes almost no Cor mislocalization, and loss of kune strongly mislocalizes and reduces Cor staining. The localization and levels of the SJ markers Dlg and Atpα were also more severely disrupted in kune mutants than in sinu or mega mutants, indicating that the effects were not specific to Cor. Interestingly, the levels and localization of Cor are not obviously different between the kune single mutant and the double and triple mutants, indicating that the kune phenotype is the most severe. Together, these results suggest that Kune has a more central role in SJ organization than either Sinu or Mega. This possibility is particularly intriguing in light of the greater similarity of Kune to vertebrate claudins than either Sinu or Mega. Perhaps a more central role for Kune in barrier junction formation has constrained its evolution and thus Kune more closely resembles ancestral claudins than do Sinu and Mega, which may have evolved more specialized functions (Nelson, 2010).
It is curious that multiple nonredundant claudins are required for SJ organization and barrier function. The exact reason for this is unclear, but perhaps each claudin interacts independently with specific junctional molecules to establish a SJ scaffold. This would be consistent with their divergent protein sequences and the differences in their N and C termini. Importantly, vertebrate claudins also have nonredundant roles in TJ function. For example, paracellular barrier function is compromised in the epidermis of mice lacking claudin-1 despite the presence of claudin-4 at the TJ (Nelson, 2010).
Like other SJ components, Kune does not appear to be required for establishment of apical-basal polarity since the levels and localization of the apical marker, Crumbs (Crb), and the adherens junction marker, E-cad, were normal in kune embryos. However, it was recently shown that some SJ components have a role in a newly identified phase of Drosophila epithelial polarity that occurs between stages 11 and 13. Because of redundancy between SJ components involved in this polarity phase and the SJ component yrt, SJ proteins required for polarity can be identified only in a yrt zygotic mutant background. For example, single zygotic mutations in either the SJ gene Atpα or yrt show normal apical localization of Crb at stage 12. Atpα, yrt double mutants on the other hand show severe mislocalization of Crb, indicative of a loss of polarity. In contrast, neither kune single mutants nor kune; yrt double mutants display any obvious polarity defects, demonstrating that kune is not required for either establishment or maintenance of epithelial polarity (Nelson, 2010).
Previous work has shown that neither mega nor sinu are required for establishment of apical-basal polarity or for maintenance of epithelial polarity at mid-embryogenesis. Together with the findings in this article, the available evidence suggests that Drosophila claudins are not required for epithelial polarity. This parallels the situation in Caenorhabditis elegans where mutations in the claudin-like proteins CLC1-4 disrupt barrier function, but not epithelial polarity. Similarly, claudins do not appear to be required for epithelial polarity in mammalian epithelial cells, since Eph4 cells can establish normal polarity even when lacking claudin complexes and tight junction strands due to elimination of ZO-1 and ZO-2. The absence of a role for claudins in polarity in any characterized species is consistent with the proposal that barrier junctions arose after polarity during the evolution of metazoans (Nelson, 2010).
These results show that Kune is an essential claudin that is required in all examined tissues for the organization and function of SJs. Kune expression and localization overlaps with the Drosophila claudins Mega and Sinu, but it was found that all three claudins play unique roles in SJ organization. Importantly, Kune more closely resembles vertebrate claudins than either Mega or Sinu and appears to play a more central role in SJ organization. Further work is needed to establish the complete molecular organization of SJs, but such work will be facilitated by the presented characterization of Kune and its interaction with other SJ components (Nelson, 2010).
Cells at the tips of budding branches in the Drosophila tracheal system generate two morphologically different types of seamless tubes. Terminal cells (TCs) form branched lumenized extensions that mediate gas exchange at target tissues, whereas fusion cells (FCs) form ring-like connections between adjacent tracheal metameres. Each tracheal branch contains a specific set of TCs, FCs, or both, but the mechanisms that select between the two tip cell types in a branch-specific fashion are not clear. This study shows that the ETS domain transcriptional repressor anterior open (aop) is dispensable for directed tracheal cell migration, but plays a key role in tracheal tip cell fate specification. Whereas aop globally inhibits TC and FC specification, MAPK signaling overcomes this inhibition by triggering degradation of Aop in tip cells. Loss of aop function causes excessive FC and TC specification, indicating that without Aop-mediated inhibition, all tracheal cells are competent to adopt a specialized fate. Aop plays a dual role by inhibiting both MAPK and Wingless signaling, which induce TC and FC fate, respectively. In addition, the branch-specific choice between the two seamless tube types depends on the tracheal branch identity gene spalt major, which is sufficient to inhibit TC specification. Thus, a single repressor, Aop, integrates two different signals to couple tip cell fate selection with branch identity. The switch from a branching towards an anastomosing tip cell type may have evolved with the acquisition of a main tube that connects separate tracheal primordia to generate a tubular network (Caviglia, 2013).
This work has investigated how the choice between the two types of specialized tip cells in the tracheal system is controlled. The transcriptional repressor Aop plays a key role in linking tracheal tip cell fate selection with branch identity. First, a novel tube morphogenesis phenotype is described in aop mutants, which is due to the massive mis-specification of regular epithelial cells into specialized tracheal tip cells. aop is specifically required for controlling tracheal cell fate, whereas aop, like pnt, is dispensable for primary tracheal branching, thus uncoupling roles of RTK signaling in cell fate specification and cell motility. The finding that tracheal branching morphogenesis proceeds normally in the presence of excess tip cell-like cells suggests that collective cell migration is surprisingly robust and that mis-specified cells apparently do not impede the guided migration of the tracheal primordium. Second, it was demonstrated that in the absence of inhibitors of MAPK signaling (aop and sty), all tracheal cells are competent to assume either TC or FC fate. The transcriptional repressor Aop globally blocks both TC and FC differentiation, but high-levels of MAPK signaling in tip cells relieve Aop-mediated inhibition, thus permitting differentiation. Third, the results suggest that in the DT region Aop limits FC induction through a distinct mechanism by antagonizing Wg signaling in addition to MAPK signaling. Conversely, in the other branches, Aop limits TC differentiation by blocking MAPK-dependent activation of Pnt. Fourth, it was shown that the region-specific choice between the two cell fates in the DT is determined by Wg signaling and by the selector gene salm. Based on these results, a model is proposed in which a single repressor, Aop, integrates MAPK and Wg signals to couple tip cell fate selection with branch identity. High levels of Bnl signaling trigger Pnt activation and Aop degradation in tracheal tip cells. It is proposed that in the DT, unlike in other tracheal cells, MAPK-induced degradation of Aop releases inhibition of Wg signaling. This is consistent with recent work showing an inhibitory effect of Aop on Wg signaling, possibly through direct interaction of Aop and Arm, or through Aop-mediated transcriptional repression of Wg pathway component. The current work extends the evidence for this unexpected intersection between two major conserved signaling pathways, suggesting that this function of Aop is likely to be more widespread than previously appreciated. The findings also provide an explanation for the puzzling observation that, in pnt mutants, TCs are lost, while FCs become ectopically specified. As pnt is required for expression of the feedback inhibitor sty, loss of pnt is expected to lead to MAPK pathway activation and consequently to increased Aop degradation. This would release Aop-mediated repression of Wg signaling, resulting in extra FCs, whereas TCs are absent because of the lack of pnt-dependent induction. This suggests that excessive FC specification in the DT of aop and sty mutants is mainly due to deregulated Wg signaling, rather than to de-repression of pnt-dependent MAPK target genes. Consistent with this notion, it was shown that pnt is not required for Delta and Dys expression in tracheal cells, although constitutively active AopACT represses their expression (Caviglia, 2013).
The results further show that salm function constrains the fate that is chosen by cells when released from the Aop inhibitory block. MAPK signaling triggers Aop degradation in all tip cells, but only in the absence of salm does this signal lead to TC induction. In salm-expressing cells, degradation of Aop releases Wg signaling, resulting in FC specification. Thus, salm biases the choice between two morphologically different types of seamless tubes. This is reminiscent of the role of salm in switching between different cell types in the peripheral nervous system and in muscles. salm expression is sufficient to repress TC formation. The genetic results, consistent with biochemical data showing that Salm acts as a transcriptional repressor, suggest that salm promotes FC fate by repressing genes involved in TC development. However, salm is not sufficient to overcome the requirement for Wg signaling in FC induction, indicating that Wg does not act solely via
salm to induce FC fate. Indeed, FC induction requires genes whose expression is independent of salm (esg, dys). In addition, it is proposed that a feedback loop between Wg signaling and salm expression maintains levels of Wg signaling in the DT sufficiently high to induce FC fate. Taken together, these results suggest that the default specialized tip cell fate, and possibly an ancestral tracheal cell state, is TC fate. Although FCs and TCs differ in their morphology, they share a unique topology as seamless unicellular tubes. FCs and TCs might therefore represent variations of a prototypical seamless tube cell type. Salm might modify cellular morphology by repressing TC genes, including DSRF, which mediates cell elongation and shape change. Intriguingly, Wg-dependent salm expression in the DT of dipterans correlates with a shift towards FC as the specialized fate adopted by the tip cells of this branch. This study has shown that salm expression inhibits TC fate, while promoting the formation of a multicellular main tracheal tube by inhibiting cell intercalation. It is therefore tempting to speculate that the salm-dependent switch from a branching towards an anastomosing tip cell type in the DT may have evolved with the acquisition in higher insects of a main tube that connects separate tracheal primordia to generate a tubular network. It will be of great interest to identify the relevant target genes that mediate the effect of Salm on tube morphology and tip cell fate (Caviglia, 2013).
The mechanisms of tip cell selection during angiogenesis in vertebrates are beginning to be understood at the molecular level. However, the signals that control the formation of vascular anastomoses by a particular set of tip cells are not known. Intriguingly, the development of secondary lumina in aop mutants is reminiscent of transluminal pillar formation during intussusceptive angiogenesis, which is thought to subdivide an existing vessel without sprouting. Although the cellular basis for this process is not understood, it is conceivable that specialized endothelial cells are involved in transluminal pillar formation. This work provides a paradigm for deciphering how two major signaling pathways crosstalk and are integrated to control cell fate in a developing tubular organ. It will be interesting to see whether similar principles govern tip cell fate choice during tube morphogenesis in vertebrates and invertebrates (Caviglia, 2013).
Dedicator of cytokinesis (DOCK) family genes are known as DOCK1-DOCK11 in mammals. DOCK family proteins mainly regulate actin filament polymerization and/or depolymerization and are GEF proteins, which contribute to cellular signaling events by activating small G proteins. Sponge (Spg) is a Drosophila counterpart to mammalian DOCK3/DOCK4, and plays a role in embryonic central nervous system development, R7 photoreceptor cell differentiation, and adult thorax development. In order to conduct further functional analyses on Spg in vivo, its localization was examined in third instar larval wing imaginal discs. Immunostaining with purified anti-Spg IgG revealed that Spg mainly localized in the air sac primordium (ASP) in wing imaginal discs. Spg is therefore predicted to play an important role in the ASP. The specific knockdown of Spg by the breathless-GAL4 driver in tracheal cells induced lethality accompanied with a defect in ASP development and the induction of apoptosis. The monitoring of ERK signaling activity in wing imaginal discs by immunostaining with anti-diphospho-ERK IgG revealed reductions in the ERK signal cascade in Spg knockdown clones. Furthermore, the overexpression of D-raf suppressed defects in survival and the proliferation of cells in the ASP induced by the knockdown of Spg. Collectively, these results indicate that Spg plays a critical role in ASP development and tracheal cell viability that is mediated by the ERK signaling pathway (Morishita, 2017).
The role of tip and rear cells in collective migration is still a matter of debate and their differences at the cytoskeletal level are poorly understood. This study analysed these issues in the Drosophila trachea, an organ that develops from the collective migration of clusters of cells that respond to Branchless (Bnl), a FGF homologue expressed in surrounding tissues. Individual cells in the migratory cluster were tracked and their features were characterized; two prototypical types of cytoskeletal organization were unveiled that account for tip and rear cells respectively. Indeed, once the former are specified, they remain as such throughout migration. Furthermore, it was shown that FGF signalling in a single tip cell can trigger the migration of the cells in the branch. Finally, specific Rac activation was found at the tip cells, and how FGF-independent cell features such as adhesion and motility act on coupling the behaviour of trailing and tip cells was analyzed. Thus, the combined effect of FGF promoting leading cell behaviour and the modulation of cell properties in a cluster can account for the wide range of migratory events driven by FGF (Lebreton, 2013).
Among the tracheal branches from each placode, two grow towards the ventral side of
the embryo, one in the anterior and the other in the posterior region of the segment,
the lateral trunk anterior (LTa) and the lateral trunk posterior (LTp) respectively. By a combination of migration, intercalation and elongation, the tip cell
of the LTp migrates towards the central nervous system (CNS), and the resulting
ganglionic branch (GB) connects the CNS to the main tracheal tube. Another cell from the LTp migrates towards the LTa of the adjacent
posterior metamere and makes a fusion branch that connects the two LT branches. This study focused on this branch (LTp/GB) because its complex morphology and pattern of migration make it
particularly appropriate for analysing the morphology and behaviour of the tip and
trailing cell during tracheal collective migration (Lebreton, 2013).
The FGF signalling pathway is involved in many
morphogenetic events requiring collective migration of cell clusters. However, it is
not entirely clear whether in these events FGF signalling is directly involved in
triggering cell migration, or alternatively if it is required for other processes such as
cell determination which only affect cell migration indirectly. Moreover, while FGF
might be required it is not clear either whether all the cells or just a subset of those
need to directly receive the signal to sustain the migration of the entire cluster. One
well-studied case is the role of FGF in the development of the zebra fish lateral line.
In that case, FGF appears to be produced by the leading cells which signal to the
trailing cells, the cells where FGF signalling is active. Restriction of FGF signalling is
thereafter required for the asymmetric expression of the receptors for the chemokines
that guide migration (Lebreton, 2013).
A very different scenario applies in the case of Drosophila tracheal migration. On the
one hand, FGF is expressed in groups of cells outside the migrating cluster. On the other hand the results in the LTp/GB
indicate that FGF signalling is required and sufficient in the leading cells, and not in
the trailing cells, for the migration of the whole cell cluster. Therefore, in spite of its
widespread involvement, the mechanisms triggered by FGF signalling in collective
migration appear to be quite different (Lebreton, 2013).
Rho inactivation produced breaks and
detachment in the LTp/GB cluster while its constitutive activation led these cells to
hold together impairing migration. Likewise, upon Cdc42 inactivation LTp/GB cells
were associated by thin extensions associated in some cases with breaks, while upon
its constitutive activation, the LTp/GB transient pyramidal organisation did not
evolve, or evolved much more slowly, towards branch elongation. However, the
phenotypes from each RhoGTPase mutants don't look alike and the detailed analysis
suggests that Rho impinges primarily on cell adhesion while Cdc42 does so on cell
motility (Lebreton, 2013).
These results are consistent with previous findings that show a role for Rho in
regulating adherens junctions stability and for Cdc42 as the main mediator of
filopodia formation. It is noted, however, that Cdc42 was found to exert in the LTp/GB an
opposite effect to the one identified in other systems, as Cdc42DN mutants showed
more protrusions and were more actin-enriched basally than wild-type cells and
Cdc42ACT mutants showed a reduced the motility of LTp/GB (Lebreton, 2013).
There is an increasing amount of data pointing to the different effect of RhoGTPases
in vitro versus in vivo models and also among various cell types. A unidirectional assignment between a
specific cellular process in vivo and a single RhoGTPase is probably an
oversimplification and this was not the aim of the current study. Rather the study relied on mutant
forms of the RhoGTPases to modulate cell features, either individually or collectively,
to assess their role in the overall behaviour of the cell cluster. In doing so, the results
point to a critical role for a balance between cell adhesion and cell motility for the
collective migration of a cell cluster (Lebreton, 2013).
The results support the following model for the specification, features and behaviour
of leading cells in the migration of the LTp/GB branch. Upon signalling
from the FGF pathway, tip cells reorganise their cytoskeleton features (actin
enrichment at the basal membrane, small apical surface and an apicobasal polarity
along the proximo-distal axis), thereby enabling them to acquire leading behaviour.
Indeed, FGF can induce migratory capacity to the whole cluster by signalling only the
tip cells, where a dynamic transition between states of Rac activity is needed to
acquire a leading role. How the behaviour of tip cells leads collective migration
thereafter depends on the features of the cells in the cluster, which are determined by
various regulators (among these, the RhoGTPases) which act, at least in part,
in an FGF-independent manner. Ultimately, the balance between individual cell
properties such as cell adhesion, motility and apicobasal polarity will (1) determine the
net movement of the overall cell bodies or alternatively changes in cell shape in terms
of elongation, (2) control the migratory speed and (3) define whether cells will migrate
individually or in clusters and whether clusters will bifurcate in different paths. The
combined effect of the changes promoting leading cell behaviour and modulation of
cell features is likely to be a widely exploited mechanism in collective migration. In
particular, the actual balance between these cell features may dictate the specifics of
each migratory process and, consequently, the final shape of the tissues and organs
they contribute to generate (Lebreton, 2013).
Segmented organisms have serially repeated structures that become specialized in some segments. The Drosophila corpora allata, prothoracic glands, and trachea are shown to have a homologous origin and can convert into each other. The tracheal epithelial tubes develop from ten trunk placodes, and homologous ectodermal cells in the maxilla and labium form the corpora allata and the prothoracic glands. The early endocrine and trachea gene networks are similar, with STAT and Hox genes inducing their activation. The initial invagination of the trachea and the endocrine primordia is identical, but activation of Snail in the glands induces an epithelial-mesenchymal transition (EMT), after which the corpora allata and prothoracic gland primordia coalesce and migrate dorsally, joining the corpora cardiaca to form the ring gland. It is proposed that the arthropod ectodermal endocrine glands and respiratory organs arose through an extreme process of divergent evolution from a metameric repeated structure (Sanchez-Higueras, 2013).
The endocrine control of molting and metamorphosis in
insects is regulated by the corpora allata (ca) and the prothoracic
glands (pg), which secrete juvenile hormone and
ecdysone, respectively. In Diptera, these glands and the
corpora cardiaca (cc) fuse during development to form a
tripartite endocrine organ called the ring gland.
While the corpora cardiaca is known to originate from the
migration of anterior mesodermal cells, the origin of the other
two ring gland components is unclear (Sanchez-Higueras, 2013).
The tracheae have a completely different structure consisting
of a tubular network of polarized cells. The tracheae are
specified in the second thoracic to the eighth abdominal segments
(T2-A8) by the activation of trachealess (trh) and ventral
veinless (vvl) (Sanchez-Higueras, 2013).
The enhancers controlling trh and vvl in the
tracheal primordia have been isolated and shown to be activated by JAK/
STAT signaling. While the trh enhancers are restricted to
the tracheal primordia in the T2-A8 segments, the vvl1+2
enhancer is also expressed in cells at homologous positions
in the maxilla (Mx), labium (Lb), T1, and A9 segments in a
pattern reproducing the early transcription of vvl. The fate of these nontracheal vvl-expressing cells was unknown, but it was shown that ectopic trh expression
transforms these cells into tracheae. To identify their
fate, vvl1+2-EGFP and mCherry constructs were made (Sanchez-Higueras, 2013).
Although the vvl1+2 enhancer drives expression transiently, the stability of the EGFP and mCherry proteins labels
these cells during development. It was observed that while the
T1 and A9 patches remained in the surface and integrated
with the embryonic epidermis, the patches in the Mx and Lb
invaginated just as the tracheal primordia did. Next, the Mx and Lb patches fused, and a group of
them underwent an epithelial-mesenchymal transition (EMT)
initiating a dorsal migration toward the anterior of the aorta,
where they integrate into the ring gland. To find out what controls the EMT, the expression
of the snail (sna) gene, a key EMT regulator, was studied. Besides
its expression in the mesoderm primordium, it was found that sna
is also transcribed in two patches of cells that become the
migrating primordium. Using
sna bacterial artificial chromosomes (BACs) with different
cis-regulatory regions, the enhancer activating
sna in the ring gland primordium (sna-rg). A sna-rg-GFP
construct labels the subset of Mx and Lb vvl1+2-expressing
cells that experience EMT and migrate to form the ring gland. Staining with seven-up
(svp) and spalt (sal) (also known as salm) markers,
which label the ca and the pg, respectively, showed that the sna-rg-GFP cells form these two endocrine glands. The sna-rg-GFP-expressing
cells in the Mx activated svp, and those in the
Lb activated sal before they coalesced, indicating
that the ca and pg are specified in different segments before
they migrate (Sanchez-Higueras, 2013).
To test whether Hox genes, the major regulators of anteroposterior
segment differentiation, participate in gland
morphogenesis, vvl1+2-GFP embryos were stained, and it was found that the Mx vvl1+2 primordium expressed Deformed (Dfd)
and the Lb primordium Sex combs reduced (Scr), while the
T1 primordium expressed very low levels of Scr.
Dfd mutant embryos lacked the ca, while Scr mutant embryos
lacked the pg. Dfd and Scr expression in the gland
primordia was transient, suggesting that they control their
specification. Consistently, in Dfd, Scr double-mutant embryos,
vvl1+2 was not activated in the Mx and Lb patches, and the same was true for vvl transcription. In these mutants, the sna-rg-GFP expression was almost absent, and the ca and pg did not form. In each case, Dfd controlled the expression of the Mx patch and Scr of the Lb patch (Sanchez-Higueras, 2013).
The capacity of different Hox genes to rescue the
ring gland defects of Scr, Dfd double mutants was tested. Induction of Dfd with the sal-Gal4 line in
these mutants restored the expression of vvl1+2
and sna-rg-GFP in the Mx and the Lb. However, in contrast
to the wild-type, both segments formed a
ca as all cells express Svp. Similarly, induction
of Scr also restored the vvl1+2 and sna-rg-GFP
expression, but both primordia formed a pg as they activate
Sal and Phantom, an enzyme required for ecdysone synthesis. The capacity of both Dfd and Scr to restore vvl expression, regardless of the segment, led to a test of whether other Hox proteins could have the same function. Induction of Antennapaedia (Antp), Ultrabithorax (Ubx), abdominal-A (abd-A), or Abdominal-B (Abd-B) restored
vvl1+2 expression in the Mx and Lb, but these cells formed
tubes instead of migratory gland primordia. These cephalic tubes are trachea, as they do not
activate sna-rg, they express Trh, and their nuclei
accumulate Tango (Tgo), a maternal protein that is only translocated
to the nucleus in salivary glands and tracheal cells,
indicating that the trunk Hox proteins can restore vvl expression
in the Mx and Lb but induce their transformation to
trachea (Sanchez-Higueras, 2013).
To investigate whether vvl and trh expression is normally
under Hox control in the trunk, focus was placed on Antp, which is
expressed at high levels in the tracheal pits. In
double-mutant Dfd, Antp embryos,
vvl1+2 was maintained in the Lb where Scr was present, while
the Mx, T1, and T2 patches were missing. In T3-A8, vvl1+2
expression, although reduced, was present, probably due to
the expression of Ubx, Abd-A, and Abd-B in the posterior
thorax and abdomen. Thus, Antp regulates vvl expression in
the tracheal T2 primordium. Surprisingly, in Dfd, Antp double
mutants, Trh and Tgo were maintained in the T2 tracheal pit, indicating that although Hox genes can activate ectopic trh expression, in the tracheal primordia they may be
acting redundantly with some other unidentified factor, explaining
why the capacity of Hox proteins to specify trachea
had not been reported previously (Sanchez-Higueras, 2013).
sna null mutants were studied to determine sna's requirement
for ring gland development, but their aberrant gastrulation precluded analyzing specific ring gland defects. To investigate sna function in the gland primordia, the sna
mutants were rescued with the sna-squish BAC, which drives normal
Sna expression except in the ring gland. These
embryos have a normal gastrulation and activate the sna-rg-
GFP; however, the gland primordia degenerate and disappear. To block apoptosis, these embryos
were made homozygous for the H99 deficiency, which removes three
apoptotic inducers. In this situation, the ca and pg
primordia invaginated and survived, but they did not undergo
EMT. As a result, the gland primordia maintain
epithelial polarity, do not migrate, and form small pouches
that remain attached to the epidermis.
Vvl is required for tracheal migration. In vvl mutant
embryos, sna-rg-GFP expression was activated, but the cells
degenerated. In vvl mutant embryos also mutant
for H99, the primordia underwent EMT and migrated up to the primordia coalescence; however, the later dorsal migration did not progress (Sanchez-Higueras, 2013).
This study has shown that the ca and pg develop from vvl-expressing
cephalic cells at positions where other segments
form trachea, suggesting that they could be part of a segmentally
repeated structure that is modified in each segment by the
activity of different Hox proteins. As the cephalic primordia are
transformed into trachea by ectopic expression of trunk Hox, tests were performed to see whether the trachea primordia could form gland cells. Ectopic expression of Dfd with arm-
Gal4 resulted in the activation of sna-rg-GFP on the ventral
side of the tracheal pits. These sna-rg-GFP0-expressing
cells also expressed vvl1+2 and Trh and had
nuclear Tgo, showing that they conserve tracheal characteristics.
These sna-rg-GFP-positive cells did not show EMT and
remained associated to the ventral anterior tracheal branch.
The strength of ectopic sna-rg-GFP expression increased
when ectopic Dfd was induced in trh mutant embryos. However, migratory behaviors in the sna-rg-GFP cells
were only observed if Dfd was coexpressed with Sal. Thus, sal is expressed several times in the gland primordia, first at st9-10 repressing trunk Hox expression in
the cephalic segments and second from st11 in
the prothoracic gland. It is uncertain whether the sal requirement
for migration is linked to the first function or whether it
represents an additional role (Sanchez-Higueras, 2013).
These results show that the endocrine ectodermal glands and
the respiratory trachea develop as serially homologous organs
in Drosophila. The identical regulation of vvl in the primordia of
trachea and gland by the combined action of the JAK/STAT
pathway and Hox proteins could represent the vestiges of an ancestral regulatory
network retained to specify these serially repeated structures,
while the activation of Sna for gland development and Trh and
Tgo for trachea formation could represent network modifications
recruited later by specific Hox proteins during the
functional specialization of each primordium. This
hypothesis or alternative possibilities should be confirmed
by analyzing the expression of these gene networks in various
arthropod species. The diversification of glands and respiratory
organs must have occurred before the split of insects
and crustaceans, as there is a correspondence between the
endocrine glands in both classes, with the corpora cardiaca
corresponding to the pericardial organ, the corpora allata to
the mandibular organ, and the prothoracic gland to the Y gland. Despite their divergent morphology, a correspondence between the insect trachea and the crustacean gills
can also be made, as both respiratory organs coexpress vvl
and trh during their organogenesis. Divergence between
endocrine glands and respiratory organs may have occurred
when the evolution of the arthropod exoskeleton required
solving two simultaneous problems: the need to molt to allow
growth, and the need for specialized organs for gas exchange (Sanchez-Higueras, 2013).
The cytoskeleton is a major determinant of cell-shape changes that drive the formation of complex tissues during development. Important roles for actomyosin during tissue morphogenesis have been identified, but the role of the microtubule cytoskeleton is less clear. This study shows that during tubulogenesis of the salivary glands in the fly embryo, the microtubule cytoskeleton undergoes major rearrangements, including a 90° change in alignment relative to the apicobasal axis, loss of centrosomal attachment, and apical stabilization. Disruption of the microtubule cytoskeleton leads to failure of apical constriction in placodal cells fated to invaginate. This failure is due to loss of an apical medial actomyosin network whose pulsatile behavior in wild-type embryos drives the apical constriction of the cells. The medial actomyosin network interacts with the minus ends of acentrosomal microtubule bundles through the cytolinker protein Shot, and disruption of Shot also impairs apical constriction (Booth, 2014).
Tubes are essential for nutrient transport and gas exchange in multicellular eukaryotes, but how connections between different tube types are maintained over time is unknown. In the Drosophila tracheal system, mutations in oak gall (okg) and conjoined (cnj) confer identical defects, including late onset blockage near the terminal cell-stalk cell junction and the ectopic extension of autocellular, seamed tubes into the terminal cell. It was determined that okg and cnj encode the E and G subunits of the vacuolar ATPase (vATPase); both the V0 and V1 domains are required for terminal cell morphogenesis. Remarkably, the ectopic seamed tubes running along vATPase-deficient terminal cells belonged to the neighboring stalk cells. All vATPase-deficient tracheal cells had reduced apical domains and terminal cells displayed mislocalized apical proteins. Consistent with recent reports that the mTOR and vATPase pathways intersect, this study found that mTOR pathway mutants phenocopied okg and cnj. Furthermore, terminal cells depleted for the apical determinants Par6 or aPKC had identical ectopic seamed tube defects. This study has thus identified a novel mechanism of compensatory branching in which stalk cells extend autocellular tubes into neighboring terminal cells with undersized apical domains. This compensatory branching also occurs in response to injury, with damaged terminal cells being rapidly invaded by their stalk cell neighbor (Francis, 2015).
Connection of tubules into larger networks is the key process for the development of circulatory systems. In Drosophila development, tip cells of the tracheal system lead the migration of each branch and connect tubules by adhering to each other and simultaneously changing into a torus-shape. This study shows that as adhesion sites form between fusion cells, myosin and microtubules form polarized bundles that connect the new adhesion site to the cells' microtubule-organizing centres, and that E-cadherin and retrograde recycling endosomes are preferentially deposited at the new adhesion site. It was demonstrated that microtubules help balancing tip cell contraction, which is driven by myosin, and is required for adhesion and tube fusion. Also, retrograde recycling and directed secretion of a specific matrix protein into the fusion-cell interface promote fusion. The study proposes that microtubule bundles connecting these cell-cell interfaces coordinate cell contractility and apical secretion to facilitate tube fusion (Kato, 2016).
Previous studies showed that F-actin-enriched cell protrusions form in the tip of migrating tracheal branches. This study showed that tracheal FCs form polarized microtubule bundles oriented towards the leading edge of the migrating cells. The function of these microtubules is twofold: to concentrate E-cadherin to the newly contacted cell interface and to initiate the formation of new adherens junctions. The E-cadherin that accumulated at the new cell interface is not recycled from the cell surface, but is instead drawn from a newly synthesized pool and recruited preferentially to the FC contact site, and not to existing adherens junctions between FCs and stalk cells. It is speculated that the forward reorientation of the MTOC and the polarization of the microtubule plus ends towards the leading edge underlie the preferential deposition of E-cadherin at the FC contact site. One possible mechanism of preferential deposition is considered, in which microtubules transport endosomes containing E-cadherin towards the contact site, and attempts were made to test this model by imaging vesicular trafficking of the complex containing E-cadherin-GFP and other adherens junction components. No definitive evidence for this model was found. However, the Golgi apparatus was found to shift forward, towards the FC contact site, and that an E-cadherin-GFP signal increased in the plasma membrane before becoming concentrated at the contact site. Based on these observations, a model is favored in which the relocalization of the Golgi apparatus near the leading edge of the FC provides a source for E-cadherin that is deposited locally in the plasma membrane and the trans association of E-cadherin between the FCs nucleates a further concentration of E-cadherin via cis clustering. MTOC components have been shown to be located apically in stalk cells and the microtubule function is required for the apical assembly of adherens junction proteins Par-3 and E-cadherin through regulation of recycling endosomes. This mechanism appears different from FCs, as assembly of new adherens junction occurs in the cell interface enriched with microtubule plus ends opposite to the centriole located in the proximal side (Kato, 2016).
A second microtubule function was discovered in this study, which was to equalize the contraction in FC pairs after contact. The coordinated contraction in FC pairs pulls two FC-stalk cell junctions simultaneously towards the FC contact site. The contractile force comes from a myosin-driven process; the microtubules may serve as a 'ratchet' to fix the length of the FCs after each round of contraction. When microtubules were inhibited, the FCs relaxed to their original length after contracting, which delayed the overall fusion process. When microtubules were destabilized, branch fusion proceeded, albeit with delays and imbalances, and fusion was eventually completed. Even in this condition, the conversion of the FC cells into a torus shape occurred simultaneously, suggesting that there is a mechanism to coordinate the fusion event in FC pairs. The proposed ratchet-like function of microtubule must in some way be linked to the contractile activity of myosin. One good candidate molecule for coupling the actomyosin contraction to the microtubule function is Short-stop (Shot), which belongs to the conserved spectraplakin family of cytoskeletal proteins, and was shown to be required for tracheal branch fusion. The involvement of microtubules and Shot in the ratchet-like mechanisms observed in several contraction-dependent morphogenetic events should be tested in the future (Kato, 2016).
It is interesting to note that balancing of the force applied to the E-cadherin conjugated cell interface via microtubule plus ends is similar in configuration to the mitotic spindle, where microtubule plus ends attached to the kinetochore of each of paired sister chromosomes applies pulling force to each spindle pole. The equal number of cadherin-catenin complex in each side of the FC interface associated with trans-conjugated E-cadherin pairs in the FC interface may provide a platform for microtubule plus end attachment for generating balanced contractile force (Kato, 2016).
When FCs are fully contracted, the plasma membrane of the two adherens junctions in each FC are connected in a single burst and cell pairs are converted simultaneously into a torus shape. This process requires ADP-ribosylation factor-like 3 (Arl3) GTPase, which associates with the microtubules and intracellular vesicles concentrated at the FC contact site. This study showed that Arl3 is required for directed Serp trafficking, and that GFP-Rab9 overexpression overrides the requirement for Arl3. It is proposed that the microtubule-dependent transport of the Golgi apparatus and endosomes facilitates the concentration of Rab9 and Arl3 at the FC contact site, where they act together to increase the concentration of Serp in the lumen. The deacetylation of chitin converts it to the more hydrophilic form chitosan. The increase in water absorption by chitosan would cause the luminal matrix gel to swell, simultaneously pushing the plasma membranes of the FC interface closer to the plasma membrane of the FC-stalk cell interface so that the membrane-fusion machinery triggers the conversion of the paired FCs into a torus shape (Kato, 2016).
A number of issues remain to be explained. Lumen formation in FCs was clearly detected in arl3 mutants, but not in the normal context. This is probably because very small lumen is sufficient to trigger fusion of wild-type FCs. Although Arl3 is absolutely required for fusion, Rab9 and Serp are not, suggesting that the proposed luminal-matrix swelling due to chitin deacetylation is not the sole mechanism of plasma membrane fusion, and additional Arl3-regulated process of fusion control must exist. Moreover, additional Rab9 cargo that acts together with Serp to rescue the arl3 mutants is predicted. To uncover the entire fusion process, it will be necessary to search for additional Arl3 and Rab9 targets, and to analyse FC-specific membrane trafficking and secretion (Kato, 2016).
Extracellular vesicles (EVs) comprise diverse types of cell-released membranous structures that are thought to play important roles in intercellular communication. While the formation and functions of EVs have been investigated extensively in cultured cells, studies of EVs in vivo have remained scarce. This study reports that EVs are present in the developing lumen of tracheal tubes in Drosophila embryos. Two distinct EV subpopulations are defined, one of which contains the Munc13-4 homologue Staccato (Stac) and is spatially and temporally associated tracheal tube fusion (anastomosis) events. The formation of Stac-positive luminal EVs depends on the tracheal tip-cell-specific GTPase Arl3, which is also required for the formation of Stac-positive multivesicular bodies, suggesting that Stac-EVs derive from fusion of Stac-MVBs with the luminal membrane in tip cells during anastomosis formation. The GTPases Rab27 and Rab35 cooperate downstream of Arl3 to promote Stac-MVB formation and tube fusion. It is proposed that Stac-MVBs act as membrane reservoirs that facilitate tracheal lumen fusion in a process regulated by Arl3, Rab27, Rab35, and Stac/Munc13-4 (Camelo, 2022).
This paper reports the presence of membranous EVs in the lumen of developing tracheal tubes in Drosophila embryos. A subset of these EVs is associated with tracheal tube fusion sites and carries the Munc13-4 ortholog Stac. Stac associates with lysosome-related organelles (LROs) that display features of MVBs, the formation of which, as well as of Stac-EVs, depends on the fusion-cell-specific GTPase Arl3. Moreover, we identified Rab27 and Rab35 as partially redundant regulators of Arl3-dependent Stac-MVB formation and tracheal tube fusion. It is proposed that tracheal anastomosis formation involves fusion of Stac-MVBs with the luminal membranes of FCs, resulting in release of Stac-EVs. Attempts to visualize tracheal EV exocytosis using the pH-sensitive fluorescent proteins pHluorin fused to the lysosomal transmembrane protein Lamp1 were unsuccessful. However, it was possible to detect EVs that emerged at and remained associated with anastomosis sites upon completion of dorsal trunk (DT) fusion. Moreover, the absence of Stac-EVs from fusion-defective tracheal metameres in shot mutant embryos supports the idea that Stac-EVs are released during tube fusion (Camelo, 2022).
How is MVB formation regulated in FCs? Consistent with the finding that Rab27 and Rab35 act downstream of Arl3 to promote Stac-MVB formation, Rab27a, Rab27b and Rab35 regulate MVB-dependent exosome secretion in mammalian cells. Moreover, the Stac homolog Munc13-4 is a key effector of Rab27a in LRO docking in hematopoietic cells and melanocytes, and regulates MVB maturation and exosome release in cancer cells. Whereas Rab27 and Rab35 are required for Stac-MVB formation or for anchoring Stac at MVBs, Rab39 is dispensable for FC-specific Stac-LRO formation, suggesting that it acts in a successive step, such as transport or plasma membrane docking of LROs. While the current findings define a core pathway of MVB maturation and EV release in tracheal cells, additional players, such as ESCRT complex components, are likely to be involved. MVBs also act as intermediates in intracellular lumen formation in tracheal terminal cells, which resemble FCs in forming seamless tubes. Interestingly, Rab35 directs transport of apical components to promote terminal cell lumen growth. Whether Rab35 also participates in MVB formation in tracheal terminal cells remains to be tested (Camelo, 2022).
Despite substantial progress in characterizing EVs and their mode of action in cultured cells, studies of physiological functions of EVs in vivo have remained scarce. Exosomes are released into the seminal fluid in Drosophila testis accessory glands and modulate female reproductive behavior. Glia-derived exosomes stimulate growth of motor neurons and tracheae in Drosophila larvae through exosomal microRNA-dependent gene regulation in target cells. Moreover, cardiomyocyte-derived EVs are taken up by macrophages and endothelial cells in zebrafish embryos, and may play a role in trophic support of endothelial cells during vasculogenesis. It will be exciting to explore possible functions of tracheal EVs, including potential roles in long-range intercellular communication across the tracheal lumen. However, the luminal EVs that this study discovered in late-stage embryos are unlikely to participate in such intercellular communication, as the presence at this stage of a luminal matrix appears to constrain EV mobility, and the cuticle covering the apical tracheal cell surface presumably prevents fusion of luminal EVs with the plasma membrane. Rather, Stac-EVs are likely to constitute remnants of MVB-plasma membrane fusion events during anastomosis formation. Stac-MVBs could serve as membrane reservoirs that facilitate lumen fusion. ILVs might back-fuse with the MVB limiting membrane, resulting in a rapid increase of membrane material available at the luminal membrane interfaces in FCs. Such a mechanism has been described in dendritic cells, where MVBs carrying major histocompatibility complex class II (MHC II) in ILVs are reorganized after stimulation, resulting in tubular membrane extensions and transport of MHCII to the plasma membrane. Recruitment of ILVs to the MVB limiting membrane will increase the surface area and may lead to surface exposure of ILV membrane proteins that could promote membrane fusion. Exploring how MVB exocytosis is regulated in tracheal cells, and whether similar mechanisms contribute to vascular anastomosis formation in vertebrates, will be exciting subjects for future studies (Camelo, 2022).
Insects are small relative to vertebrates, and were larger in the Paleozoic when atmospheric oxygen levels were higher. The safety margin for oxygen delivery does not increase in larger insects, due to an increased mass-specific investment in the tracheal system and a greater use of convection in larger insects. Prior studies have shown that the dimensions and number of tracheal system branches varies inversely with rearing oxygen in embryonic and larval insects. This study tested whether rearing in 10, 21, or 40 kPa atmospheric oxygen atmospheres for 5-7 generations affected the tracheal dimensions and diffusing capacities of pupal and adult Drosophila. Abdominal tracheae and pupal snorkel tracheae showed weak responses to oxygen, while leg tracheae showed strong, but imperfect compensatory changes. The diffusing capacity of leg tracheae appears closely matched to predicted oxygen transport needs by diffusion, perhaps explaining the consistent and significant responses of these tracheae to rearing oxygen. The reduced investment in tracheal structure in insects reared in higher oxygen levels may be important for conserving tissue oxygen partial pressure and may provide an important mechanism for insects to invest only the space and materials necessary into respiratory structure (Klok, 2016).
The Drosophila protein Jim Lovell (Lov) is a putative transcription factor of the BTB/POZ (Bric- a-Brac/Tramtrack/Broad/ Pox virus and Zinc finger) domain class that is expressed in many elements of the developing larval nervous system. It has roles in innate behaviors such as larval locomotion and adult courtship. In performing tissue-specific knockdown with the Gal4-UAS system this study identified a new behavioral phenotype for lov: larvae failed to burrow into their food during their growth phase and then failed to tunnel into an agarose substratum during their wandering phase. These phenotypes originate in a previously unrecognized role for lov in the tracheae. By using tracheal-specific Gal4 lines, Lov immunolocalization and a lov enhancer trap line, lov was stablished to be normally expressed in the tracheae from late in embryogenesis through larval life. Using an assay that monitors food burrowing, substrate tunneling and death lov tracheal knockdown was shown to result in tracheal fluid-filling, producing hypoxia that activates the aberrant behaviors and inhibits development. The role of lov in the tracheae that initiates this sequence of events was investigated. When lov levels are reduced, the tracheal cells are smaller, more numerous and show lower levels of endopolyploidization. Together these findings indicate that Lov is necessary for tracheal endoreplicative growth and that its loss in this tissue causes loss of tracheal integrity resulting in chronic hypoxia and abnormal burrowing and tunneling behavior (Zhou, 2016).
Centrosome amplification is a hallmark of cancer, although how this process affects tumorigenesis is still not understood. Besides the contribution of supernumerary centrosomes to mitotic defects, their
biological effects in the post-mitotic cell are not well known. This study
analyzed the effects of centrosome amplification in post-mitotic cells during single-cell branching. It was shown that Drosophila tracheal cells with extra centrosomes branch more than wild-type cells. Mutations in Rca1 and CycA affect subcellular branching, causing tracheal tip cells to form more than one subcellular lumen. Rca1 and CycA post-mitotic cells have supernumerary centrosomes and other mutant conditions that
increase centrosome number also show excess of subcellular lumen
branching. Furthermore, de novo lumen formation is impaired in mutant
embryos with fewer centrioles. The data presented define a requirement for
the centrosome as a microtubule-organizing center (MTOC) for the
initiation of subcellular lumen formation. The study proposes that
centrosomes are necessary to drive subcellular lumen formation. In
addition, centrosome amplification increases single-cell branching, a
process parallel to capillary sprouting in blood vessels. These results
shed new light on how centrosomes can contribute to pathology
independently of mitotic defects (Ricolo, 2016).
The architecture of the outer body wall cuticle is fundamental to protect arthropods against invading pathogens and numerous other harmful stresses. Such robust cuticles are formed by parallel running chitin microfibrils. Molting and also local wounding leads to dynamic assembly and disassembly of the chitin-matrix throughout development. However, the underlying molecular mechanisms that organize proper chitin-matrix formation are poorly known. A key region has been identified for cuticle thickening at the apical cell surface, the cuticle assembly zone, where Obstructor-A (Obst-A) coordinates the formation of the chitin-matrix. Obst-A binds chitin and the deacetylase Serpentine (Serp) in a core complex which is required for chitin-matrix maturation and preservation. This study presents evidence that Chitinase 2 (Cht2) could be essential for this molecular machinery. Cht2 is expressed in the chitin-matrix of epidermis, trachea, and the digestive system. There, Cht2 is enriched at the apical cell surface and the dense chitin-matrix. It was further shown that in Cht2 knockdown larvae the assembly zone is rudimentary preventing normal cuticle formation and pore canal organization. As sequence similarities of Cht2 and the core complex proteins indicate evolutionarily conserved molecular mechanisms, these findings suggest that Cht2 is involved in chitin formation also in other insects (Pesch, 2016).
Chitin is an aminopolysaccharide present in insects as a major structural component of the cuticle. However, current knowledge on the chitin biosynthetic machinery, especially its constituents and mechanism, is limited. Using three independent binding assays, including co-immunoprecipitation, split-ubiquitin membrane yeast two-hybrid assay, and pull-down assay, this study demonstrated that choline transporter-like protein 2 (Ctl2) interacts with krotzkopf verkehrt (kkv) in Drosophila melanogaster. The global knockdown of Ctl2 by RNA interference (RNAi) induced lethality at the larval stage. Tissue-specific RNAi to silence Ctl2 in the tracheal system and in the epidermis of the flies resulted in lethality at the first larval instar. The knockdown of Ctl2 in wings led to shrunken wings containing accumulated fluid. Calcofluor White staining demonstrated reduced chitin content in the first longitudinal vein of Ctl2 knockdown wings. The pro-cuticle, which was thinner compared to wildtype, exhibited a reduced number of chitin laminar layers. Phylogenetic analyses revealed orthologues of Ctl2 in different insect orders with highly conserved domains. These findings provide new insights into cuticle formation, wherein Ctl2 plays an important role as a chitin-synthase interacting protein (Duan, 2022).
The extracellular matrix (ECM), a structure contributed to and commonly shared by many cells in an organism, plays an active role during morphogenesis. This study used the Drosophila tracheal system to study the complex relationship between the ECM and epithelial cells during development. There is an active feedback mechanism between the apical ECM (aECM) and the apical F-actin in tracheal cells. Furthermore, cell-cell junctions are key players in this aECM patterning and organisation, and individual cells contribute autonomously to their aECM. Strikingly, changes in the aECM influence the levels of phosphorylated Src42A (pSrc) at cell junctions. Therefore, it is proposed that Src42A phosphorylation levels provide a link for the ECM environment to ensure proper cytoskeletal organisation (Ozturk-Colak, 2016).
This study examined the apical ECM (aECM) of Drosophila melanogaster trachea, the insect respiratory system. Once the different branches of the tracheal system have been established to cover the overall embryonic body, tracheal cells begin to secrete the components of a chitin-rich aECM that lines up the lumen of the tracheal tubes and can be visualised by the incorporation of chitin-binding probes. A distinctive feature of this aECM are taenidial folds, a series of cuticle ridges that compose a helical structure running perpendicular to the tube length along the entire lumen. Taenidia are believed to confer mechanical strength to the tubes and have been compared to a coiled spring within a rubber tube or to the corrugated hose of a vacuum cleaner. From the very first descriptions, it was noticed that taenidia are unaffected by the presence of cell boundaries, thereby indicating that they are a supracellular structure and suggesting a substantial degree of intercellular coordination. It has been reported that taenidial organisation correlates with that of the apical F-actin bundles in underlying cells—the formation of these bundles preceding the appearance of taenidia. However, the relationship between these bundles and taenidia is still poorly understood. In addition, physical modelling has recently revealed that the interaction of the apical cellular membrane and the aECM determines the stability of biological tubes, thus generating more questions about how this interaction occurs (Ozturk-Colak, 2016).
This study reports that there is a dynamic relationship between sub-apical F-actin and taenidial folds during tracheal lumen formation. Cell-cell junctions participate in organising F-actin bundles and the taenidial fold supracellular aECM and this chitinous aECM contributes to regulating F-actin organisation in a two-way regulatory mechanism (Ozturk-Colak, 2016).
The contribution of chitin deposition to the organisation of taenidia was examined. When studying the contribution of tracheal actin rings to this process, mutants were chosen that do not completely inhibit chitin deposition, as these mutants would probably heavily impair tracheal development, thus hindering specific analysis of the morphogenesis of taenidia. Thus, this investigation turned to Blimp-1, an ecdysone response gene that encodes the Drosophila homolog of the transcriptional factor B-lymphocyte-inducing maturation protein gene and whose mutants have been reported to have misshapen trachea almost completely devoid of taenidia (Ozturk-Colak, 2016).
Indeed, Blimp-1 mutant embryos were grossly inflated compared to the wild-type, a phenotype associated with weaker embryonic cuticles caused by mutations impairing the deposition or organisation of chitin. Consistent with this observation, Blimp-1 mutants showed a pale ectodermal cuticle with smaller denticles, although their phenotype is weaker than that of the kkv chitin synthase mutants. This observation suggests that, while chitin deposition is severely impaired, some still accumulated in the cuticle of Blimp-1 mutant embryos. In support of this hypothesis, lower levels of fluostain signal were detected in the trachea of Blimp-1 mutants compared to the wild-type. Thus, this study expected to find similarly less conspicuous taenidia, which was indeed the case. However, the most obvious abnormal feature of taenidia was their pattern, as they were not organised in folds perpendicular to the tube axis but instead ran parallel to it. Given the close correlation between taenidia and actin bundle organisation, actin arrangement was examined in Blimp-1 mutants, and it was found to be severely impaired. In most Blimp-1 mutants examined, no tracheal actin rings were observed. However, in the mutant embryos in which apical actin bundles were detected, these were oriented in parallel to the tube length like the chitin structures. Thus, as is the case for the other mutant genotypes examined so far, in Blimp-1 embryos the lack of a proper arrangement of taenidial folds correlates with either the absence or abnormal pattern of actin rings (Ozturk-Colak, 2016).
Detailed ultrastructural analysis by TEM confirmed the close interplay between actin and chitin in both tal/pri and Blimp-1 mutants. In wild-type embryos, each taenidium is formed by a plasma membrane protrusion and the taenidia have a regular shape. Arrangement of plasma membrane protrusions in tal/pri and Blimp-1 mutant tracheal cells is irregular. At the end of embryogenesis, whereas the breadth of these taenidia is very constant in wt animals, it is highly variable in tal/pri and Blimp-1 mutants. This result is in line with the finding that proper F-actin ring organisation and chitin deposition are necessary for taenidial morphogenesis (Ozturk-Colak, 2016).
The observation of an effect of a mutation in a gene required for proper chitin arrangement on actin bundling was unexpected. To assess whether the effect of Blimp-1 mutations on actin organisation was indeed a consequence of abnormal chitin deposition in the tracheal cuticle rather than the result of a direct and yet unknown role of Blimp-1 in F-actin bundling, tracheal actin organisation was examined in mutants for kkv, a gene required for chitin morphogenesis only. Surprisingly, kkv mutants also lacked actin rings, thereby indicating a feedback role of proper chitin-mediated tracheal cuticle in F-actin organisation. In addition, F-actin bundles formed normally and thereafter collapsed in kkv mutants. This finding indicates that a proper cuticle is not required for the establishment of the F-actin rings but instead for their maintenance. This implies that proper chitin deposition/organisation contributes to ensure the proper organisation and stability of the apical F-actin rings (Ozturk-Colak, 2016).
How could the apical chitin in the ECM influence actin bundling? It was observed that both kkv and Blimp-1 mutations had an effect on tracheal cell shape. In the wild-type trachea, the cells of the DT were organised such that the longest axis of their apical shape is parallel to the tube axis. However, in both Blimp-1 and kkv mutant trachea, the anteroposterior elongation of the cells of the DT was lost, causing cells to be more square shaped. Thus, it was hypothesised that the change in taenidial orientation in kkv and Blimp-1 mutants could be attributed to the alteration in the overall orientation or shape of the tracheal cells. Interestingly, a modification of cell shape/orientation also occurs in embryos mutant for the Src-family kinase Src42A. However, and as previously reported for F-actin, this study found taenidia to follow the same organisation in Src42A mutant embryos as the wild-type indicating that proper organisation of taenidia can be uncoupled from correct tracheal cell shape/orientation and thus that the former is not merely a consequence of the latter (Ozturk-Colak, 2016).
Having identified and characterised genes that specifically affect taenidial patterning, the individual cell contributions to this supracellular organisation was examined by impairing genetic functions in mosaics. It was not possible to generate mosaics by mitotic recombination since there are no cell divisions after tracheal invagination and RNAi-mediated knockdown often does not work in Drosophila embryogenesis. This was indeed the case upon expression of UAS-RNAi constructs for either tal/pri or Blimp-1 in the embryonic tracheal cells. Thus, alternative approaches were used to produce tracheal cellular chimeras (Ozturk-Colak, 2016).
First, advantage was taken of the effect of Blimp-1 overexpression on taenidial formation. To generate tracheal DTs with distinct cellular composition, an AbdB-Gal4 line was used that drives expression only in the posterior part of the embryo. This approach served as an internal control within the same embryo. Upon expression of UASBlimp-1 under these conditions, lower levels of chitin were detected in the posterior metameres. Thus, chitin deposition seems to be highly dependent on the levels of Blimp-1 activity as both loss-of-function mutations and overexpression of Blimp-1 induce low levels of chitin. It is also noted that overexpression of Blimp-1 gives rise to tracheal cells with a less elongated apical side, like that of Blimp-1 and kkv mutants. Then the trachea at the border of the AbdB-Gal4 domain were examined, finding a perfect correlation between the different physical appearance of taenidia and cells and their genotype, with either wild-type or increased levels of Blimp-1. Flip-out clones expressing Blimp-1 were generated in a wild-type background, and similar results were obtained in these clones. Thus, it is concluded that Blimp-1 regulates chitin accumulation in a cell-autonomous manner and that each cell contributes independently to the chitin deposition of their corresponding segments of the taenidial folds (Ozturk-Colak, 2016).
As a second approach to mosaic analysis, the same AbdBGal4 line was used to drive expression of tal/pri and Blimp-1 in tal/pri and in Blimp-1 loss-of-function mutant backgrounds, respectively. For both mutants, a rescuing effect was seen in the posterior tracheal metameres as taenidial folds became organised perpendicularly to the tube length. Using this approach, it was possible to generate borders of cells with and without tal/pri and Blimp-1 function and taenidia were analyzed in these conditions. In the case of the tal/pri rescue experiment, a difference was detected between the cells expressing the wild-type tal/pri gene and those with a wild-type phenotype, an observation consistent with the non-cell autonomous function of the Tal/Pri peptides. However, in the case of the Blimp-1 rescue experiment, taenidia tended to follow the orientation dictated by the genotype of their respective cells. Moreover, and due to the expression domain of the AbdBGAL4 driver not being completely continuous, single cells of one of the genotypes were observed surrounded by cells of the other and either mutant cells could be detected with a longitudinal arrangement of the taenidia or 'rescued' cells with a perpendicular arrangement; in this case, there was a correlation between the physical appearance of taenidia and the corresponding cell genotype. Interestingly, intermediate orientations between the prototypical longitudinal taenidia were also detected in the mutant domain and the perpendicular ones in the rescued domain. These results suggest that cells 'adapt' the orientation of 'their' segments of the taenidia to the global orientation of the segments of the taenidia contributed by neighbouring cells (Ozturk-Colak, 2016).
These results show that tracheal taenidia can form proper rings even when the neighbouring cells do not. This indicates that, to a certain degree, segments of taenidia can organise properly even in the absence of proper subjacent actin rings provided that the segments of taenidia contributed by the neighbouring cells are properly organised (Ozturk-Colak, 2016).
The role for the apical chitin ECM in tracheal actin organisation indicates a feedback mechanism to generate the supracellular taenidial structures. In the light of the above and previously published results, the following model is proposed for the formation of the taenidial folds that expand the overall diameter of the tracheal tube. On the one hand, actin polymerises in rings at the apical side of the tracheal cells in a tal/pri-dependent process; these actin rings are then required for the particular accumulation of the kkv chitin synthase and for the appearance of folds in the plasma membrane. In turn, kkv accumulation leads to a localised increased production and deposition of chitin along specific enriched stripes above the actin rings in a Blimp-1-mediated process. On the other hand, the cellular AJs are instrumental in ensuring that apical F-actin bundles from each cell follow a supracellular organ arrangement. It has to be noted that each cell appears to independently organise or maintain, to a certain degree, the proper orientation of their actin bundles, as determined by Blimp-1 clonal analysis and the disruption of cell adhesion by downregulation of α-Cat and, consequently, DE-Cad. These results further suggest cell polarity along the circumferential axis of the tracheal tube. Nevertheless, this is not an absolute value as cells also have the capacity to modify the orientation of their sections of the taenidia to keep the continuity of these structures along the tube. In this regard, cell adhesion is central to ensure the continuity of the intracellular actin bundles as a patterning element for the overall tube. Subsequently, the chitin aECM feeds back on to the cellular architecture by stabilising F-actin bundling and cell shape via the modulation of Src42A phosphorylation levels. The combination of all these phenomena explain just how it is that the cells of the tracheal epithelium can cooperate unconsciously so as to form a helicoid [chitinous] thickening continuous from one end of the trachea to another (Ozturk-Colak, 2016).
During sprouting angiogenesis in the vertebrate vascular system, and primary branching in the Drosophila tracheal system, specialized tip cells direct branch outgrowth and network formation. When tip cells lumenize, they form subcellular (seamless) tubes. How these seamless tubes are made, shaped and maintained remains poorly understood. This study characterize a Drosophila mutant called ichor (ich), and show that ich is essential for the integrity and shape of seamless tubes in tracheal terminal cells. Ich regulates seamless tubulogenesis via its role in promoting the formation of a mature apical extracellular matrix (aECM) lining the lumen of the seamless tubes. ich encodes a zinc finger protein (CG11966) that acts, as a transcriptional activator required for the expression of multiple aECM factors, including a novel membrane-anchored trypsin protease (CG8213). Thus, the integrity and shape of seamless tubes are regulated by the aECM that lines their lumens (Rosa, 2018).
Epithelia grow and shape into functional structures during organogenesis. Although most of the focus on organogenesis has been drawn to the building of biological structures, the disassembly of pre-existing structures is also an important event to reach a functional adult organ. Examples of disassembly processes include the regression of the Mullerian or Wolffian ducts during gonad development and mammary gland involution during the post-lactational period in adult females. To date, it is unclear how organ disassembly is controlled at the cellular level. This study follows the Drosophila larval trachea through metamorphosis and shows that its disassembly is a hormone-driven and precisely orchestrated process. It occurs in two phases: first, remodeling of the apical extracellular matrix (aECM), mediated by matrix metalloproteases and independent of the actomyosin cytoskeleton, results in a progressive shortening of the entire trachea and a nuclear-to-cytoplasmic relocalization of the Hippo effector Yorkie (Yki). Second, a decreased transcription of the Yki target, Diap1, in the posterior metameres and the activation of caspases result in the apoptotic loss of the posterior half of the trachea while the anterior half escapes cell death. Thus, this work unravels a mechanism by which hormone-driven ECM remodeling controls sequential tissue shortening and apoptotic cell removal through the transcriptional activity of Yki, leading to organ disassembly during animal development (Fraire-Zamora, 2021).
This study report how a functional organ, the larval trachea of Drosophila melanogaster, undergoes a hormone-driven disassembly during metamorphosis. The dorsal trunks of the trachea shorten modularly in two sequential phases: (1) an initial progressive phase of a controlled reduction in the trachea length, involving aECM remodeling and cellular shape changes and, as a consequence, (2) Yki inactivation results in a decreased transcription of its target gene (and apoptosis inhibitor) Diap1 in the posterior metameres, resulting in their loss through cell death (Fraire-Zamora, 2021).
It was found that the activation of apoptosis results in the disassembly and loss of only the posterior metameres (Tr6-Tr8). Why are anterior metameres (Tr3-Tr5) not affected? This is an intriguing observation because most of the signaling inputs occur along the whole dorsal trunk. These results suggest that the anterior metameres are 'protected' or 'desensitized' against the signals that occur along the dorsal trunks. The results on downregulation of AbdB suggest that Hox genes play a role in the differential response between anterior and posterior metameres during dorsal trunk disassembly. However, it cannot be excluded that other elements might also contribute, such as the proximity of pools of adult progenitor cells to the anterior metameres. Whether the anterior metameres are protected against MMP-1 activity and cell death through a compartmentalization of the trachea via Hox genes or through signals from the progenitor cells is a matter of future work (Fraire-Zamora, 2021).
The Hpo signaling pathway and its effector Yki/YAP are conserved both in invertebrates and vertebrates where they regulate organ size through the transcriptional control of genes related to proliferation and cell death. While work in mammalian cell cultures has shown that ECM-related inactivation of YAP can lead to an increase in apoptosis, most of the focus has concentrated on how the Hpo signaling pathway controls proliferation, with very few examples on the activation of apoptosis (Fraire-Zamora, 2021).
The current results unveil a new role of the Hpo pathway in the hormone-driven disassembly of an organ through an ECM-triggered inactivation of the Hpo effector Yki, resulting in the triggering of apoptosis on the posterior end of the trachea. Given the conservation of the Hpo pathway, it is speculated that its role in organ disassembly could be of general use in some of the widespread events leading to organ involution (i.e., the controlled regression or shrinkage of an organ) during embryonic development or in homeostatic processes during adult life or aging (Fraire-Zamora, 2021).
Intercalation allows cells to exchange positions in a spatially oriented manner in an array of diverse processes, spanning convergent extension in embryonic gastrulation to the formation of tubular organs. However, given the co-occurrence of cell intercalation and changes in cell shape, it is sometimes difficult to ascertain their respective contribution to morphogenesis. A well-established model to analyse intercalation, particularly in tubular organs, is the Drosophila tracheal system. There, fibroblast growth factor (FGF) signalling at the tip of the dorsal branches generates a 'pulling' force believed to promote cell elongation and cell intercalation, which account for the final branch extension. This study used a variety of experimental conditions to study the contribution of cell elongation and cell intercalation to morphogenesis and analysed their mutual requirements. Evidence is provided that cell intercalation does not require cell elongation and vice versa. The two cell behaviours are controlled by independent but simultaneous mechanisms, and cell elongation is sufficient to account for full extension of the dorsal branch, while cell intercalation has a specific role in setting the diameter of this structure. Thus, rather than viewing changes in cell shape and cell intercalation as just redundant events that add robustness to a given morphogenetic process, this study finds that they can also act by contributing to different features of tissue architecture (Casani, 2020).
Terminal differentiation of an organ is the last step in development that enables the organism to survive in the outside world after birth. Terminal differentiation of the insect tracheae that ends with filling the tubular network with gas is not fully understood at the tissue level. This study demonstrates that yet unidentified valves at the end of the tracheal system of the fruit fly Drosophila melanogaster embryo are important elements allowing terminal differentiation of this organ. Formation of these valves depends on the function of the zona pellucida protein Trynity (Tyn). The tracheae of tyn mutant embryos that lack these structures do not fill with gas. Additionally, external material penetrates into the tracheal tubes indicating that the tyn spiracles are permanently open. It is concluded that the tracheal endings have to be closed to ensure gas-filling. It is speculated that according to physical models closing of the tubular tracheal network provokes initial increase of the internal hydrostatic pressure necessary for gas generation through cavitation when the pressure is subsequently decreased (Wang, 2018).
This study characterized a Drosophila melanogaster cis-regulatory module (CRM) termed TT-CRM. The TT-CRM is 646 bp long and is located in one of the introns of CG32239 (RhoGEF64C) and resides about 3,500 bp upstream of CG13711 and about 620 bp upstream of CG12493. Analysis of 646 bp-lacZ lines revealed that TT-CRM drives gene expression not only to the larval, prepupal, and pupal tracheal system but also to the adult dorsal longitudinal muscles. The patterns of mRNA expression of the transgene and of the CGs that lie in the vicinity of TT-CRM were investigated both in dissected trachea and in adult thoraces. Through RT-qPCR it was observed that in the tracheal system the pattern of expression of 646 bp-lacZ is similar to the pattern of expression of CG32239 and CG13711, whereas in the thoracic muscles 646 bp-lacZ expression accompanies the expression of CG12493. Together, these results suggest new functions for two previously characterized D. melanogaster genes and also contribute to the initial characterization of a novel CRM that drives a dynamic pattern of expression throughout development (Wester, 2018).
The outer surface of insects is covered by the cuticle, which is derived from the apical extracellular matrix (aECM). The aECM is secreted by epidermal cells during embryogenesis. The aECM exhibits large variations in structure, function, and constituent molecules, reflecting the enormous diversity in insect appearances. To investigate the molecular principles of aECM organization and function, the role of a conserved aECM protein, the ZP domain protein Trynity, was studied in Drosophila melanogaster. trynity was first identified as an essential gene for epidermal barrier function. trynity mutation caused disintegration of the outermost envelope layer of the cuticle, resulting in small-molecule leakage and in growth and molting defects. In addition, the tracheal tubules of trynity mutants showed defects in pore-like structures of the cuticle, and the mutant tracheal cells failed to absorb luminal proteins and liquid. These findings indicated that trynity plays essential roles in organizing nano-level structures in the envelope layer of the cuticle that both restrict molecular trafficking through the epidermis and promote the massive absorption pulse in the trachea (Itakura, 2018).
Tube size control and how tubular anisotropy is translated at the cellular level are still not fully understood. This study investigated these mechanisms using the Drosophila tracheal system. The apical polarity protein Crumbs transiently accumulates anisotropically at longitudinal cell junctions during tube elongation. Evidence is provided indicating that the accumulation of Crumbs in specific apical domains correlates with apical surface expansion, suggesting a link between the anisotropic accumulation of Crumbs at the cellular level and membrane expansion. This study finds that Src42A is required for the anisotropic accumulation of Crumbs, thereby identifying the first polarised cell behaviour downstream of Src42A. The results indicate that Src42A regulates a mechanism that increases the fraction of Crb protein at longitudinal junctions, and genetic interaction experiments are consistent with Crb acting downstream of Src42A in controlling tube size. Collectively, these results suggest a model in which Src42A would sense the inherent anisotropic mechanical tension of the tube and translate it into a polarised Crumbs accumulation, which may promote a bias towards longitudinal membrane expansion, orienting cell elongation and, as a consequence, longitudinal growth at the tissue level. This work provides new insights into the key question of how organ growth is controlled and polarised and unveils the function of two conserved proteins, Crumbs and Src42A, with important roles in development and homeostasis as well as in disease, in this biological process (Olivares-Castineira, 2018).
Since Crb has been proposed to regulate tube length by promoting apical membrane growth, Crb accumulation was examined in the Dorsal Trunk (DT, the main tracheal trunk connecting to the exterior through the spiracles). Crb can localise to different subdomains of the apical membrane during tracheal development: the SubApical Region (SAR) and the Apical Free Region (AFR). The AFR corresponds to the most apical domain, free of contact with other epithelial cells and in direct contact with the lumen in the case of tubular organs like the trachea, while the SAR corresponds to the most apicolateral membrane domain of contact between neighboring epithelial cells. Previous work has shown that during the stages of higher longitudinal DT growth, stage 15 onwards, Crb accumulated strongly in the SAR, displaying a mesh-like pattern that identifies the apical junctional domain. Strikingly, Crb was anisotropically (not uniformly) distributed in the SAR of cell junctions. Cell junctions are classified as longitudinal cell junctions (LCJs), mainly parallel to the longitudinal axis of the tube, and transverse cell junctions (TCJs), perpendicular to the longitudinal axis. Crb accumulation was found to be more visible at LCJs than at TCJs; several examples were observed where accumulation of Crb at TCJs was almost absent. The accumulation of Crb (total fluorescence intensity/junctional length) was quantified at LCJs and TCJs; accumulation was found to be biased to LCJs, where levels were around 30% higher than at TCJs (average % of difference of Crb accumulation at LCJs and TCJs). To compare different embryos the LCJ/TCJ ratio was calculated of Crb accumulation, which showed an average of 1,5 (n = 15 embryos), indicating that Crb is anisotropically distributed, i.e. polarised. In contrast to Crb, DE-Cadherin (DE-cad), a core component of the Adherens Junctions (AJs), was equally distributed among all cell junctions. The ratio of accumulation in LCJ/TCJ was close to 1, indicating that the anisotropic distribution is not a general feature of all junctional proteins. These results indicated that a larger proportion of LCJs accumulate higher levels of Crb than TCJs (Olivares-Castineira, 2018).
To further investigate this observation time-lapse imaging was carried out in embryos carrying the viable and functional CrbGFP allele as the only source of functional Crb protein. Enrichments were observed of Crb protein at LCJs, and less conspicuous accumulations were found at TCJs, from late stage 15 and during stage 16 over a period of 1,30-2 hours. This correlated with an increase in tube length of around a 30% and a moderate increase in tube diameter of an 11% (Olivares-Castineira, 2018).
Altogether these results point to a polarised accumulation of Crb that correlates with an anisotropic growth along the longitudinal axis of the DT during stage 16. It is worth pointing out that anisotropies of Crb, like the one described in this study, or of other apical determinants, have important implications in morphogenesis (Olivares-Castineira, 2018).
Different molecular mechanisms could underlie the preferential accumulation of Crb at LCJs, such as specific Crb degradation at TCJs, specific stabilisation at LCJs, targeted intracellular trafficking, differential protein recycling, among others. To investigate the possible mechanism behind the anisotropic pattern of Crb accumulation FRAP analysis was performed at either LCJs or TCJs of embryos carrying the CrbGFP allele. The amount of fluorescent protein, relative to the pre-bleach value, mobilized during the experimental time (mobile fraction, Mf) was significantly higher at LCJs compared to TCJs, indicating a higher recovery of CrbGFP protein at LCJs. To assess the recovery kinetics the half-time (t1/2, time to reach half of the Mf) was calculated. The half-time was not significantly different at LCJs and TCJs, suggesting that the recovery rate is comparable at the differently oriented junctions. Kymographs of the bleached regions suggested that the recovery was not due to lateral diffusion. Altogether the results indicated a higher mobility of Crb protein at LCJs but a constant rate of incorporation in all junctions (Olivares-Castineira, 2018).
Hence, on the one hand higher levels of Crb accumulate at LCJs, and on the other, FRAP experiments show that Crb protein is more mobile at LCJs. These results could suggest the existence of two molecularly defined different pools of Crb in the junctions with different mobility: a basal level-pool with lower mobility and an enrichment-pool with higher mobility. The basal level-pool would be present in all junctions, while a mechanism acting specifically at LCJs would ensure also the presence of the enrichment-pool there. The increased mobility/instability of the enrichment-pool of Crb at LCJs would contribute to increase the total Crb mobility at LCJs. Further experiments will be required to test this possibility and to understand how the molecular mechanism underlying the increased accumulation of Crb at LCJs relates to the differential mobility of Crb protein that was documented (Olivares-Castineira, 2018).
It was next asked how the anisotropic distribution of Crb is regulated. To investigate this question attention was turned to Src42A, as it triggers one of the mechanisms regulating tube elongation, orienting membrane growth on the longitudinal axis. In conditions of Src42A loss of function, LCJs do not expand and tubes become shorter. Crb accumulation was analyzed in loss of function conditions for Src42A. In one case the Src42AF80 allele was used that lacks the distinct accumulation of phosphorylated Src42A (pSrc42A) at the apical junctional region but does not affect the stability or membrane localisation of the protein. This mutation renders a kinase non-activatable protein that was previously shown to strongly affect tracheal tube elongation. In contrast, a kinase-dead dominant negative form of Src42A (Src42DN) was expressed in the trachea, that was also previously shown to affect tube elongation. In both cases a more uniform distribution was observed of Crb at LCJs and TCJs. Quantification of Crb levels indicated that the differences between the accumulation of Crb at LCJs compared to TCJs were reduced. Analysis of the LCJ/TCJ ratio of Crb clearly showed a significant decrease when compared to the control, indicating a more uniform accumulation in Src42A loss of function conditions. DE-cad LCJ/TCJ ratio in Src42A loss of function conditions remained close to 1, indicating a homogeneous distribution. Altogether these results show that a decrease in Src42A activity leads to a decrease of the anisotropic accumulation of Crb (Olivares-Castineira, 2018).
To investigate whether Src42A promotes an increased accumulation of Crb protein at LCJs or a depletion at TCJs the total levels of protein accumulation were quantified at LCJs and TCJs and control (i.e. heterozygotes) and Src42A mutants (i.e. Src42AF80 homozygotes) were compared from the same experiment. While variability was observed within each genotypic group, different independent experiments indicated that in control embryos there is an increased accumulation of Crb protein at LCJs that is lost in Src42AF80 mutants. In Src42A mutant conditions the levels of Crb accumulation at LCJs and TCJs were similar to those of TCJs of control embryos, indicating that Src42A regulates a mechanism that increases the fraction of Crb protein at LCJs (Olivares-Castineira, 2018).
Consistent with a role for Src42A in regulating directly or indirectly Crb accumulation partial co-localisation was found of Crb and Src42A protein, and with pSrc42A at the SAR. However, no polarised accumulation was detected of the active pSrc42A fraction during tube elongation, as previously documented. While there may be transient anisotropies of pSrc42A accumulation that cannot be detected with the available antibodies, this result suggests that other factors (e.g. mechanical or chemical) modulate the activity of pSrc42A in the different junctions to regulate the anisotropic accumulation of Crb (Olivares-Castineira, 2018).
To further explore Src42A requirement FRAP experiments were performed in CrbGFP embryos in which Src42A was downregulated. Clear differences were found with respect to control: while in the control the Mf and recovery curves of LCJs and TCJs were clearly different, in Src42DN conditions the Mf and recovery curves of LCJs and TCJs were comparable. The Mf at the LCJs of Src42DN was significantly lower than the Mf at the LCJs in control embryos, and was similar to the Mf at TCJs in control and mutant embryos. The halftime recovery, t1/2, was comparable to that of control embryos, indicating a recovery rate similar in all cases (Olivares-Castineira, 2018).
Altogether these results indicate that Src42A contributes to Crb preferential enrichment at LCJs and that it increases Crb mobility there. The fact that Crb levels and Crb recovery are affected particularly at LCJs when Src42A is downregulated strongly suggests that Src42A is (more) active precisely at LCJs. The results are consistent with the proposed model in which a mechanism acting specifically at LCJs, that is now proposed to be mediated by Src42A, would ensure the accumulation of an enrichment-pool of Crb at LCJs with high protein mobility. In the absence of this Src42A-mediated activity, only the basal level-pool of Crb would be present at LCJs and TCJs, leading to comparable levels and mobility of Crb in all junctions. Src42A would not regulate the basal level-pool of Crb, and would instead be required to top up Crb at LCJs with an enrichment-pool of Crb. Future experiments addressing the molecular mechanism by which Src42A regulates Crb accumulation and its mobility will help to fully understand how it regulates the anisotropic accumulation of Crb at LCJs. Src42A-independent accumulation of Crb in tracheal cells together with other Src42A-independent mechanisms of apical membrane growth may be responsible for tube growth in the absence of Src42A (Olivares-Castineira, 2018).
Src42A was found to be required for the anisotropic accumulation of Crb at LCJs. It was then asked whether the overelongation of tubes observed in Src42A overactivation conditions (either overexpression of a wild type form of Src42A or expression of a constitutively active protein) was due to an increased accumulation of Crb at LCJs. The results did not support this expectation. Crb was found to be strongly decreased in the SAR of DT cells both in conditions of overexpression (UASSrc42A) or constitutive activation (UASSrc42ACA). In conditions of mild overexpression of wild type Src42A, rare cases (around 5-8% of embryos) were found where it was possible to detect some levels of Crb in the SAR, which accumulated preferentially at LCJs, as expected. These results suggested that the tube length defects produced by Src42A overexpression/overactivation were caused by a mechanism different than the one operating in physiological conditions. To investigate this possible mechanism the levels and distribution of the total Src42A protein and the pSrc42A active fraction were analyzed in both Src42A overexpression and overactivation conditions. Levels of Src42A protein were found to be increased but still enriched in the membrane region. Interestingly, pSrc42A was not restricted any more to the junctional apical region as in the wild type and instead it was expanded along the whole apicobasal membrane. The increase and expansion of pSrc42A accumulation observed in overexpression and in overactivation conditions indicate that Src42A activity is overactivated in both cases and may explain the similarity of phenotypes. Further analysis also indicated that Src42A overexpression/overactivation leads to a general loss of cell organisation and membrane polarity, as evidenced by the miss-localisation of markers of membrane polarity, like the Septate Junction protein Megatrachea. These results indicate that an unregulated accumulation of active pSrc42A leads to a generalised miss-organisation of the cell and prevents proper Crb accumulation (Olivares-Castineira, 2018).
To investigate the cause of tube overelongation found in Src42A overexpression /overactivation conditions other known tube length regulators were analyzed. One of them is Serp, which regulates the aECM organisation. In Src42A and Src42ACA overexpression conditions Serp was found to be lost from the luminal compartment, although, as in wild type, tracheal cells accumulate Serp at early stages. This result provides explanation for the tube elongation defects observed under these conditions, as Serp absence leads to tube overelongation. Interestingly, defects in Serp accumulation could not be detected in Src42 mutants or in Src42ADN conditions, as previously reported. These results suggested again that Src42A overactivation use a different mechanism than the one used in physiological conditions to drive tube elongation. Hence, the analysis of Src42A overactivation provides new results that allow to revisit and reinterpret previously published work (Olivares-Castineira, 2018).
Altogether the results indicate that an unregulated accumulation of active pSrc42A leads to a generalised miss-organisation of the cell and prevents proper accumulation of Crb and Serp. In addition, it was also observed that DE-cad was not properly localised either. Interestingly, it has been shown that the tracheal accumulation of these proteins depends on their recycling. Thus, the results could suggest a role of Src42A in protein trafficking. In this context, the loss of cell organisation and membrane polarity produced by mislocalisation of pSrc42A could interfere with protein trafficking. Roles for Src42A in protein trafficking have been proposed in different contexts. Src42A could regulate protein trafficking directly, or indirectly through the regulation of the actin cytoskeleton. The actin cytoskeleton plays a capital role in protein trafficking and Src42A acts as a regulator of the actin cytoskeleton. A disruption of actin organisation in Src42A overactivation could lead to defects in the sorting of different cargoes as well as defects in endosomal maturation. Further experiments will be required to investigate a possible involvement of Src42A in protein trafficking during tracheal development (Olivares-Castineira, 2018).
After identifying an anisotropic accumulation of Crb regulated by Src42A, it was asked how this mechanism relates to tube elongation. Crb has been proposed to promote apical membrane growth independently of its role in apicobasal polarity at late stages of epithelial differentiation. In the trachea Crb was proposed to mediate tube elongation by promoting apical membrane growth. Interestingly, the results show an enrichment of Crb in the SAR of LCJ during tube elongation. This observation raises the hypothesis that it is precisely this accumulation of Crb in the SAR of LCJs what favours or facilitates apical membrane expansion (either by membrane growth or membrane transformation leading to cell shape changes), orienting cell elongation and as a consequence the longitudinal growth of the tube. Crb recycling during tracheal development could favour the mobilisation of cellular and/or membrane components facilitating membrane growth or membrane transformation. To investigate this possibility Crb accumulation in the SAR was analyzed in different experimental conditions in which apicobasal polarity was unaffected (Olivares-Castineira, 2018).
Altogether the results confirm a role of EGFR in regulating the accumulation of Crb in the SAR or AFR, at least in tubular organs. EGFR regulates the trafficking of different cargoes, in particular Crb and Serp, raising the possibility that the regulation of the apical surface area depends on targets different than Crb. However, the fact that Serp is not present in the SGs and that Crb has already been proposed to promote apical membrane growth, strongly suggest that Crb is at least one of the targets downstream of EGFR regulating apical expansion. On the other hand, the results correlate apical cell expansion with Crb subcellular localisation in the SAR. It is suggested that Crb accumulation in the SAR of LCJs could promote their expansion facilitating the elongation of the cell along the longitudinal axis, in agreement with the proposed role of Crb promoting apical membrane expansion. Previous observations such as the expansion of the photoreceptor stalk membrane upon Crb overexpression support this hypothesis, indicating that this can be a general mechanism (Olivares-Castineira, 2018).
Crb was proposed to regulate tube size by promoting apical membrane growth. Accordingly, it was found that a weak overexpression of Crb in an otherwise wild type background caused a mild increase in DT dimensions (a significant 12% enlargement of DT and a non-significant 9% diameter expansion) without perturbing the epithelial integrity and polarity. Src42A was shown to control tube elongation through interactions with dDaam and the remodelling of AJs. This study is now showing that Src42A regulates Crb levels, suggesting that Src42A may control tube elongation at least in part through regulation of Crb. Thus, it was asked whether increased levels of Crb can bypass or compensate the requirement of Src42A in tube growth. To evaluate this possibility genetic interaction experiments were performed to test the ability of a weak Crb overexpression in a Src42A loss of function background. Interestingly, Crb overexpression was found to produced a partial but significant rescue of the short-DT phenotype of Src42A loss of function. This result indicates that Crb acts downstream or in parallel of Src42A. Because it was also observed that Src42A is required for Crb preferential enrichment at LCJs, the hypothesis is favored that Crb acts downstream of Src42A contributing to its function in tube elongation (Olivares-Castineira, 2018).
Remarkably, besides a rescue in DT length, an increase was also detected in the diameter of the tube when Crb was overexpressed in a Src42A loss of function background (a 25% expansion with respect to Src42ADN mutants). Under these conditions, the DT diameter was not perfectly smooth and often showed dilations that were not detected in Src42ADN or Crb overexpression conditions on their own. This isometric expansion of the DT along the diametrical and longitudinal directions is interpreted to be the result of an isotropic excess of Crb. Because in the absence of Src42A activity Crb accumulation is not properly polarised, this may promote a non-polarised increase of tube growth. To find support for this interpretation Crb accumulation was analyzed in conditions of weak Crb overexpression. High levels of Crb were detected in the whole apical domain and in vesicles that precluded a proper analysis of Crb localisation and a systematic quantification of Crb accumulation. However, it was possible to observe in examples in which a distinct accumulation of Crb could be detected that the overexpression of Crb in a wild type background leads to high enrichments of the protein particularly at LCJs. This result suggests that the activity of Src42A biases the increased accumulation of Crb to the LCJs, correlating with a preferential growth mainly along the longitudinal axis. In contrast, a more generalised pattern of Crb overexpression could be detected in a Src42A loss of function mutant background, consistent with the isometric tube growth observed. In summary, although it was not possible to directly test whether an anisotropic accumulation of Crb can exclusively compensate tube elongation in Src42A loss of function conditions, the results are consistent with the hypothesis that it is the anisotropic accumulation of Crb, regulated by Src42A, that mediates or promotes oriented tube growth along the longitudinal axis. Future experiments involving the generation of new tools designed to specifically localise Crb protein at desired subcellular domains will be needed to prove the current model and to confirm an instructive and causal role of the anisotropic accumulation of Crb in cell elongation and polarised tracheal tube growth (Olivares-Castineira, 2018).
To summarise, this study finds that Crb is transiently enriched in the SAR of DT cells in a polarised/anisotropic manner. This polarised distribution correlates with different dynamics or turnover of Crb protein, which appears to be more mobile and accumulate more at longitudinal junctions than at transverse ones. This polarised distribution also correlates with the anisotropic expansion of the apical membrane, axially-biased, that drives the longitudinal enlargement of the tracheal tubes. Interestingly it was also found that Src42A is required for this anisotropic accumulation of Crb. Src42A was already known to regulate tube growth along the longitudinal axis, and this study now proposes that it performs this activity at least in part by promoting a Crb anisotropic enrichment. Src42A was also proposed to act as a mechanical sensor, translating the polarised cylindrical mechanical tension (an inherent property of cylindrical structures) into polarised cell behaviour. Hence, it is proposed that Src42A would sense differential longitudinal/transverse tension stimuli and translate them into the cell by polarising Crb accumulation. It is likely that this Crb anisotropic accumulation in the SAR of LCJs mediates apical membrane expansion in the longitudinal direction, which would help to orient cell elongation and as a consequence longitudinal tube growth. A causal role for this Crb anisotropic accumulation in orienting cell elongation awaits definitive confirmation (Olivares-Castineira, 2018).
In light of the current results, the following model is now proposed. Different mechanisms operate to regulate tube growth. On the one hand secretion drives apical membrane growth along the transverse axis independently of Src42A. In addition, a basal level-pool of Crb accumulation independent of Src42A may promote or contribute to isotropic apical expansion. On the other hand the presence of a properly organised luminal aECM also controls tube growth by restricting tube elongation. A Src42A-dependent mechanism acts in coordination with these other mechanisms. Src42A would contribute to tube elongation through interactions with dDaam, the remodelling of AJs and topping up Crb accumulation at LCJs with an enrichment-pool of Crb. This increased accumulation of Crb at LCJs would bias the growth of the tube along the longitudinal axis, counteracting the restrictive activity of the aECM on tube elongation. In the absence of Src42A activity, the Src42A independent mechanism/s of membrane growth would still operate, and would favour a compensatory growth along the transverse axis as observed, as diametrical growth is not restricted by the aECM (Olivares-Castineira, 2018).
The regulation of size and shape of tubular organs is important for organ function, as evidenced by the fact that loss of regulation can lead to pathological conditions such as polycystic kidney disease (PKD), cerebral cavernous malformation (CCM) or hereditary hemorrhagic telangiectasia (HHT). Src proteins have been implicated in malformations like PKD, highlighting the importance of investigating the mechanisms underlying their activities. While Src42A was proposed to regulate polarised cell shape changes during tracheal tube elongation through interactions with dDaam and the remodelling of AJs, no polarised downstream effectors have been identified up to date. Hence, identifying that Crb anisotropy is one of the downstream effects of Src42A activity adds an important piece to the puzzle. Src42A and Crb are conserved proteins with important roles in development and homeostasis and are involved in different pathologies. This work provides an ideal model where to investigate the molecular mechanisms underlying their activities, their interactions, and their roles in morphogenesis (Olivares-Castineira, 2018).
Hippo-like pathways are ancient signaling modules first identified in yeasts. The best-defined metazoan module forms the core of the Hippo pathway, which regulates organ size and cell fate. Hippo-like kinase modules consist of a Sterile 20-like kinase, an NDR kinase, and non-catalytic protein scaffolds. In the Hippo pathway, the upstream kinase Hippo can be activated by another kinase, Tao-1. This study delineate a related Hippo-like signaling module that Tao-1 regulates to control tracheal morphogenesis in Drosophila melanogaster. Tao-1 activates the Sterile 20-like kinase GckIII by phosphorylating its activation loop, a mode of regulation that is conserved in humans. Tao-1 and GckIII act upstream of the NDR kinase Tricornered to ensure proper tube formation in trachea. This study reveals that Tao-1 activates two related kinase modules to control both growth and morphogenesis. The Hippo-like signaling pathway delineated by this study has a potential role in the human vascular disease cerebral cavernous malformation (Poon, 2018).
Most organs are composed of epithelial or endothelial cells organized as tubes; junctional complexes maintain the intercellular or autocellular connections ('seams') that seal the cells into selectively permeable tubes. Should these connections be compromised, the consequences for organ function can be catastrophic. For example, individuals with the vascular disease, familial cerebral cavernous malformation (CCM), may suffer from seizures and strokes as a consequence of dilated leaky tubes (Poon, 2018).
Three architecturally distinct tube types have been described, and all three are found in the Drosophila tracheal system. These include multicellular seamed tubes (with intercellular junctions), seamed tubes formed by single cells (with autocellular junctions), and seamless tubes formed within single cells (no junctions). Most seamless tubes are thought to form intracellularly, although they may also form by fusion of membrane along autocellular junctions, converting autocellular tubes into seamless ones (Poon, 2018).
Tubes with single seams (autocellular) and seamless tubes form relatively late during tracheal development. The first tubes of the tracheal system are large multicellular sacs generated by invagination from the embryonic ectoderm. The tracheal epithelial cells are polarized along their apical-basolateral axis, with the apical membrane of each cell facing the lumen of the tracheal sac to which it belongs. Cells are next recruited to the distinct primary branches that migrate away from the sacs toward Branchless-FGF chemoattractant cues. Many of the primary branches initially form short wide tubes that lengthen and narrow over time, as the cells that comprise them intercalate, changing the underlying tubular architecture from multicellular to autocellular. Tip cells are required for, and lead, the migration of the new branches. Ultimately, tip cells assume specialized cell fates (terminal cell or fusion cell), and initiate secondary branch formation by targeting apical membrane internally to form seamless tube. The precise mechanism by which this occurs remains subject to debate and may differ between terminal and fusion cells (Poon, 2018).
Intriguingly, Drosophila terminal cells contain both autocellular and seamless tubes. Transition from one tube type (autocellular) to the other (seamless) occurs within the terminal cell at a location proximal to the terminal cell nucleus. This observation raises a number of questions including: how do the two tube types connect to each other, how do they match each other in diameter, and which pathways are required to regulate and execute these processes? To begin to address these questions, this study has taken a genetic approach and have screened through a large collection of terminal cell mutants to identify those that display tube morphogenesis defects within the region of the terminal cell wherein the autocellular-to-seamless tube transition occurs (hereafter, the 'transition zone') (Poon, 2018).
This study reports the identification and characterization of two mutants that disrupt lumen morphology in the transition zone in strikingly different ways. The first mutant, lotus, shows a transition zone gap in gas-filled tube, and more precise analysis shows that the gap corresponds to a local absence of tube, although tube is present proximal and distal to the gap. The second mutant, wheezy, shows dramatic tube dilation in the transition zone. These mutants may thus have opposing effects on the addition of apical membrane within the transition zone: too little, resulting in a gap, in lotus mutants; and too much, resulting in a dilation, in wheezy mutants. Lotus encodes N-ethylmaleimide sensitive factor 2 (NSF2), a protein required for SNARE recycling. This implies an especially stringent requirement for vesicle traffic in connecting the two tube types. Significantly, this study found that the second mutant, wheezy, encodes Germinal center kinase III (GCKIII). The human orthologs of GCKIII are effectors of the protein encoded by the human vascular disease gene, Cerebral cavernous malformation 3 (Ccm3), and are putative Golgi-resident kinases. Patients suffering from cerebral cavernous malformations show gross dilations of cerebral capillaries, thus suggesting that CCM3/GCKIII play a conserved role in tubulogenesis in limiting diametric tube expansion. Mutations in lotus and in wheezy both point to a crucial role of apical membrane delivery in tube morphogenesis (Poon, 2018).
The study goes on to show that Drosophila Ccm3 has a loss of function phenotype identical to GckIII, and that it interacts genetically with GckIII. Interestingly, the results stand in contrast to a previous report suggesting a requirement for the genes in tracheal lumen formation. Further, ectopic localization was found of junctional proteins in Ccm3-depleted or GckIII mutant cells, and an overaccumulation of the apical determinant, Crumbs, in GckIII mutant cells. In contrast, although Crumbs also accumulates in lotus/Nsf2 mutant cells, it does not appear to be enriched in the lumenal membrane. Overexpression of Crumbs by itself induces formation of multiple tiny dilations throughout the terminal cell, but not a large transition zone tube dilation. Nevertheless, knockdown of crumbs in GckIII mutants strongly suppresses the large local dilation defect. Similarly, mutations in lotus (Nsf2), that cause mislocalization of Crumbs, were found to epistatic to GckIII. It was also found that knockdown of the essential septate junction scaffold, Varicose (a MAGUK protein orthologous to human PALS-2), suppresses the transition zone dilation defect associated with GckIII mutants. These data suggest that it is the inappropriate increase of septate junctions in combination with overaccumulation of Crumbs that confers a transition zone dilation defect. These studies of transition zone tube morphogenesis have direct relevance to human development and disease, and provide important mechanistic insight into the basic biological question of how tubes of differing architecture connect to each other (Poon, 2018).
In the terminal cell 'transition zone,' seamed and seamless tubes must connect. This study found that during development, remodeling of cell junctions reduced tube type complexity in the transition zone, such that by third instar only seamless tubes are still present in the majority of terminal cells. How the tubes connect initially has been an outstanding question in the field for more than a decade, and whether, or how, transition zone tubes remodel over developmental time has not previously been addressed. This study describes several mutations that disrupt tube morphogenesis in the terminal cell transition zone. The mutants fall into two classes that appear to act in opposing manners: in the case of mutants like lotus, there appears to be a deficit of apical membrane addition to the growing tube within the transition zone, most frequently resulting in a failure to maintain a continuous tube. In the case of mutants such as wheezy, too much apical membrane is added to the transition zone tube, resulting in a grossly dilated appearance. Molecular genetic characterization of the genes is consistent with this hypothesis as lotus encodes a factor required for membrane traffic and wheezy encodes a putative Golgi-resident kinase that is required for restricting the accumulation of apical and junctional proteins. In this context, it is important to consider that secretion of specific cargo into the tube lumen during embryonic tracheal development is dependent upon septate junctions; perhaps an increase in transition zone septate junctions promotes enhanced targeting of vesicles to the lumenal membrane. However, the existence of septate junctions in the transition zone by itself does not appear to lead to tube dilation because ~4% of wild-type third instar terminal cells have septate junctions in the transition zone, but do not have dilations. This, together with the inability of Crumbs overexpression to cause transition zone dilations, and with the suppression of such dilations by knockdown of crumbs or of varicose, leads to the proposal that both ectopic septate junctions and increased accumulation of apical polarity proteins are required for transition zone tube dilation. This study provides genetic and molecular insight into the process of junctional remodeling and tubulogenesis in the transition zone (Poon, 2018).
This study has determined that wheezy encodes GCKIII, the only known downstream effector of CCM3, a protein mutated in human vascular disease characterized by grossly dilated, leaky capillaries. It therefore becomes important to ask whether the insights into the cellular basis of GCKIII/CCM3 deficiency are compatible with what is known from patient samples and vertebrate CCM models. In excised CCMs, staining of apical polarity proteins and junctional proteins has not been reported, but ultrastructural analyses suggest that endothelial cells lack functional tight junctions, and in mouse models, lymphatic vessels show gaps between endothelial cells or show shorter intercellular junctions. On the other hand, EM analysis of vessels from endothelial-specific Ccm3 knockout mice showed tight junctions were present and did not detect any gaps between endothelial cells. It is not clear how to reconcile these data, although it is possible that loss of CCM1 and CCM2 cause tube dilation by a mechanism that is distinct from that seen with loss of CCM3, as suggested by Chan (2011). In the Drosophila tracheal system, a clear and dramatic increase is seen in apical polarity protein accumulation, and a convincing defect in remodeling of septate junctions; this suggests that a reexamination of endothelial tight junctions and an initial study of apical polarity determinants in vertebrate CCM models should be a high priority. Indeed, extant data hint that such studies could be of great value. For example, mutations in the zebrafish orthologs of Stardust and Yurt (physical interactors and regulators of Crumbs in Drosophila) have cardiovascular defects. Moreover, remodeling of endothelial cell junctions during angiogenesis, as cells change neighbors and as the tubes they comprise change architectures, is likely to be critical to the formation of functional tubes. That terminal cells are the only tracheal cell type affected by loss of CCM3 or GCKIII activity is striking, and may be directly relevant to human disease given that ∼30% of cells comprising capillaries in the cerebral cortex have an analogous cellular architecture, the so-called 'seamless' endothelial cells). In addition, it is noted that the cellular architecture of endothelial tip cells that lead outgrowth of the zebrafish intersomitic vessels is dynamic: these cells often form seamless tubes, but sometimes appear to make tubes of mixed autocellular and seamless character (like fly terminal cells), and at other times appear to contribute to multicellular tubes. Given the dynamic nature of tight junctions during angiogenesis, defects in junction remodeling such as those seen with wheezy mutants in the Drosophila tracheal system, might be expected to give rise to leaky vessels composed of endothelial cells that are not properly attached to each other (Poon, 2018).
The precise function of CCM3 and GCKIII during angiogenesis is not clear. another study has suggested a requirement for the CCM3 pathway in lumen formation, whereas a third has suggested that the pathway regulates cell junctions via phosphorylation of Moesin. The current data conclusively show that neither model is consistent with the observed effects of loss of the CCM3/GCKIII pathway in the fly tracheal system; each model is discussed in the context of the data below (Poon, 2018).
Chan (2011) is built upon the outcome of two sets of experiments: tubulogenesis assays of CCM3- or GCKIII-depleted endothelial cells and RNAi studies in the Drosophila tracheal system. The tubulogenesis assays have been used to great effect, allowing identification of a number of genes required for endothelial lumen formation; however, it is important to appreciate that the endothelial cells in this assay must do more than simply lumenize, they must also migrate, remodel the matrix, and organize and reorganize cell:cell contacts. Disruption in any of these preconditions for lumenization would also compromise tubulogenesis. Thus, should the junctional remodeling defects observed in the fly tracheal system also pertain to endothelial cells depleted for CCM3 or GCKIII, those remodeling defects might be sufficient to block lumenization in the in vitro assays. Additionally, the fact that patient CCMs are dilated capillaries rather than lumenless capillaries would also seem to argue against this model (Poon, 2018).
It is noted that the data for GckIII and Ccm3 in the Drosophila tracheal system stand in contrast to the findings of Chan (2011). Unlike Chan, this study found tube dilation to be the primary consequence of loss of CCM3 or GCKIII function. The fully penetrant transition zone defectobserved in wheezy mutant flies is compelling, and is directly supported by RNAi experiments targeting either GckIII or Ccm3, including with RNAi transgenes employed by Chan. Consistent with the findings of Chan, no defects are observed in terminal cell gas-filling (but not in other tracheal cell tubes by mosaic analysis using a null allele of GckIII). However, the lack of observable gas-filled tubes is not due to failure in tube formation, as suggested by Chan, but rather due to a failure in liquid clearance. Indeed, liquid-filled tubes were readily detected in the knocked-down cells. Importantly, the data argue that Ccm3 and GckIII are not required for making a lumen, but instead are required to prevent inappropriate trafficking of junctional and apical proteins (Poon, 2018).
This study also addressed the model that phosphorylation of Moesin is the essential function of the CCM3/GCKIII pathway during tubulogenesis. Moesin is the only ERM (ezrin-radixin-moesin) family member in Drosophila, and this study found that phosphorylation of Moesin in the tracheal system is not compromised by loss of CCM3 or GCKIII (in fact it is elevated). These data suggest that the essential target of GCKIII kinase activity is not Moesin, but another protein yet to be identified. Going forward, it will be of great interest to determine what proteins are regulated by GCKIII kinase activity in the tracheal system, and to test whether vertebrate orthologs of these factors are required during angiogenesis (Poon, 2018).
Most organs of multicellular organisms are built from epithelial tubes. To exert their functions, tubes rely on apico-basal polarity, on junctions, which form a barrier to separate the inside from the outside, and on a proper lumen, required for gas or liquid transport. This study has identified apnoia (apn; CG15887), a novel Drosophila gene required for tracheal tube elongation and lumen stability at larval stages. Larvae lacking Apn show abnormal tracheal inflation and twisted airway tubes, but no obvious defects in early steps of tracheal maturation. apn encodes a transmembrane protein, primarily expressed in the tracheae, which exerts its function by controlling the localization of Crumbs (Crb), an evolutionarily conserved apical determinant. Apn physically interacts with Crb to control its localization and maintenance at the apical membrane of developing airways. In apn mutant tracheal cells, Crb fails to localize apically and is trapped in retromer-positive vesicles. Consistent with the role of Crb in apical membrane growth, RNAi-mediated knockdown of Crb results in decreased apical surface growth of tracheal cells and impaired axial elongation of the dorsal trunk. It is concluded that Apn is a novel regulator of tracheal tube expansion in larval tracheae, the function of which is mediated by Crb (Skouloudaki, 2019).
Animal organs consist of epithelial tissues, which form the boundaries between internal and external environment. During development, epithelia are instrumental to shape the various organs. Many epithelial tissues form tubular organs, such as the gut, the kidney or the respiratory system. A fundamental feature of epithelial tubes and sheets is to keep the balance between the maintenance of structural integrity and tissue rigidity during organ growth and morphogenesis. To understand how this balance is achieved during rapid, temporally regulated developmental transitions from juvenile to adult body shapes, several studies in various animal models have focused on elucidating how cell proliferation, cell polarity, cell shape changes and trafficking contribute to the formation of the tubular lumen length and diameter. The correct coordination of these processes is crucial for normal organ function. This is reflected in the fact that several human diseases are linked to defects in epithelial tube formation and maintenance, such as polycystic kidney disease or cystic fibrosis (Skouloudaki, 2019).
The developing tracheae of Drosophila melanogaster, a network of branched epithelial tubes that ensure oxygen supply to the cells of the body, has emerged as an ideal system to study cell fate determination and morphogenesis of epithelial tubes. The available genetic tools, as well as the ease to image the tracheal system in the fly embryo, has provided detailed insights into the developmental processes required to form tubular structures with defined functional lumens and have contributed to elucidating the interplay between tissue growth, differentiation and cell polarity (Skouloudaki, 2019).
The stereotypically branched tracheal system of Drosophila is set up at mid-embryogenesis. Once a continuous tubular network has formed, the tube expands to accommodate an increased oxygen supply to all tissues during animal growth. Tube expansion occurs by growth along the diameter and along the anterior-posterior axis. Growth is accompanied by the formation of a transient cable, comprised of a chitinous apical extracellular matrix (aECM), which fills the lumen of the tube. The generation of this cable requires the secretion of chitin and chitin-modifying enzymes. Mutations in genes affecting secretion or organization of the chitin cable result in excessively elongated tracheal tubes or tubes with irregular diameter (with constricted and swollen areas along the tube). Axial growth, on the other hand, depends on the proper elongation of tracheal cells along the anterior-posterior axis. At later stages of embryogenesis, the lumen becomes cleared and filled with air (Skouloudaki, 2019).
After hatching, the larvae undergo two molts, a process during which animals rapidly shed and replace their exoskeleton with a new one, bigger in size. For this, a new chitinous aECM is secreted apically, thus surrounding the old tube. Remodeling of this aECM permits tissue growth between larval molts. The molting process is initiated by the separation of the old aECM from the apical surface of the epithelial cells and the secretion of chitinases and proteinases, which partially degrade the old cuticle. The remnants of the old cuticle in each metamer are shed through the spiracular branches. This process, called ecdysis, is followed immediately by clearance of the molting fluid and air filling. Interestingly, while the diameter of the dorsal trunk only increases at each molt, tube length increases continuously throughout larval life, particularly during intermolt periods. Despite the importance of tube expansion and elongation for larval development, the underlying mechanisms that control tracheal growth at this stage remain poorly understood (Skouloudaki, 2019).
A well-established regulator of apical domain size in developing epithelia is Crumbs (Crb). Crb is a type I transmembrane protein, which acts as an apical determinant of epithelial tissues. It has a large extracellular, a single transmembrane and a short cytoplasmic domain. Loss- and gain-of-function experiments have shown that apical levels of Crb are important for proper cell polarity, tissue integrity and growth. For instance, absence of Crb in embryonic epithelia results in loss of apical identity and disruption of epithelial organization. In contrast, overexpression of Crb triggers apical membrane expansion, which leads to a disordered epithelium, abnormal expansion of tracheal tubes and/or tissue overgrowth. These results underscore the importance of Crb levels for epithelial development and homeostasis (Skouloudaki, 2019).
Several mechanisms have been uncovered that ensure proper levels of apical Crb. These include: stabilization of Crb at the membrane, mediated through interactions of its cytoplasmic domain with scaffolding proteins, e.g. Stardust (Sdt), or by homophilic interactions between Crb extracellular domains, regulation of Crb trafficking, including endocytosis by AP-2, Rab5 or Avalanche, membrane delivery by Rab11, recycling by the retromer and endocytic sorting by the ESCRT III component Shrub/Vps32 (Skouloudaki, 2019).
To gain further insight into the molecular mechanisms that regulate Crb and its activity during epithelial growth, this study set out to identify novel interacting partners of Crb using the yeast two-hybrid system. One of the candidates identified, CG15887, encodes a transmembrane protein, which localizes to the apical surface of tracheal tubes. This study found that the CG15887 protein physically interacts with Crb. Based on the phenotype of mutations in CG15887, which is characterized by defects in tracheal growth and inflation during larval stages, this gene was named apnoia (apn). apn mutant animals die as second instar larvae with dorsal trunks displaying reduced axial growth and impaired apical surface area expansion, resulting in shorter tubes. This phenotype is correlated with the absence of Crb from the apical surface. RNAi knock-down of crb phenocopies the apn mutant phenotype of impaired longitudinal growth. These results identify Apn as a novel regulator of tracheal tube growth in the larvae, which acts through Crb to control axial tube expansion (Skouloudaki, 2019).
This work identifies Apn as an essential protein for airway maturation in Drosophila larval stages. Apn is localized apically in tracheal epithelial cells, where it co-localizes and physically interacts with Crb. apn1 mutant larvae exhibit loss of tracheal tissue structure, manifested by tube size defects and impaired gas filling, resulting in body size reduction and lethality at second instar. At the cellular level, exclusion of Crb from the apical membrane in apn1 mutant larval tracheae goes along with apical cell surface reduction and an overall tracheal tube shortening. Absence of apn leads to Crb inhibition and accumulation in enlarged, Vps35/retromer-positive vesicles (Skouloudaki, 2019).
Elongation of the tracheal tube has been extensively studied in embryos where it has been shown to rely on different mechanisms, such as the organization of the aECM and cell shape changes. Anisotropic growth of the apical plasma membrane is an additional mechanism to achieve proper longitudinal tube expansion. However, only few proteins have been described so far to regulate this process. One of these, the protein kinase Src42A, is required for the expansion of the cells in the axial direction, and loss of Src42A function results in tube length shortening, which is associated with an increased tube diameter. Src42A has been suggested to exert its function, at least in part, by controlling DEcad recycling and hence adherens junctions remodeling and/or by its interaction with the Diaphanous-related formin dDAAM (Drosophila Dishevelled-Associated Activator of Morphogenesis), loss of which results in reduced apical levels of activated pSrc42A. More recently, Src42A has been suggested to control axial expansion by inducing anisotropic localization of Crb preferentially along the longitudinal junctions. However, this study never observed any anisotropic distribution of Crb in wild type larval tracheal cells, making it unlikely that, at this developmental stage, axial expansion is regulated by a Src42A-dependent mechanism. This assumption is corroborated by the observation that, unlike in Src42A mutants, the lack of longitudinal expansion in apn1 mutant larval tubes is not associated with circumferential expansion. Another protein regulating tube elongation in the embryo is the epidermal growth factor receptor, EGFR. Expressing a constitutively active EGFR results in shortened tracheal tubes with smaller apical cell surfaces, but with increased diametrical growth. In this condition, Crb shows altered apical distribution. This phenotype differs from the apn1 phenotype, where apical localization of Crb is almost completely lost and only longitudinal tube growth is affected. This suggests that Apn executes tube length expansion by a different mechanism (Skouloudaki, 2019).
How does decrease in tubular growth lead to loss of tracheal structure? During development, the larval body, including the tracheal tissue, elongates about 8-fold. Impaired axial tracheal cell growth in apn1 mutants thus may affect the balance between the forces exerted by apical membrane growth on the one hand and the resistance provided by the luminal aECM on the other, an important mechanism described previously to control tube shape in the embryo. This could lead to physical rupture of tubes mutant for apn1, allowing fluid entry. The presence of fluid would, in turn, disrupt proper gas filling, resulting in hypoxia and, consequently, in impaired body growth (Skouloudaki, 2019).
Several studies have shown that, in some tissues, Crb accumulation on the apical membrane is mediated by the retromer complex, which controls either the retrograde transport of Crb to the trans-Golgi or the direct trafficking from the endosomes to the plasma membrane. The physical interaction of Apn and Crb, the functional requirement of Apn for Crb apical localization and the fact that in apn1 mutants Crb is trapped in Vps35-positive/retromer vesicles all suggest that Apn is required for trafficking and/or maintenance of Crb at the apical membrane (Skouloudaki, 2019).
However, the increase in the size of Vps35-positive vesicles in apn1 mutant cells, which is, to some extent, due to the accumulation of Crb, suggests defects in retromer function, which may prevent Crb lysosomal degradation. Further studies will help to elucidate at which level Apn controls Crb trafficking in larval tracheae (Skouloudaki, 2019).
Branching morphogenesis underlies organogenesis in vertebrates and invertebrates, yet is incompletely understood. This study shows that the sarco-endoplasmic reticulum Ca(2+) reuptake pump (SERCA) directs budding across germ layers and species. Clonal knockdown demonstrated a cell-autonomous role for SERCA in Drosophila air sac budding. Live imaging of Drosophila tracheogenesis revealed elevated Ca(2+) levels in migratory tip cells as they form branches. SERCA blockade abolished this Ca(2+) differential, aborting both cell migration and new branching. Activating protein kinase C (PKC) rescued Ca(2+) in tip cells and restored cell migration and branching. Likewise, inhibiting SERCA abolished mammalian epithelial budding, PKC activation rescued budding, while morphogens did not. Mesoderm (zebrafish angiogenesis) and ectoderm (Drosophila nervous system) behaved similarly, suggesting a conserved requirement for cell-autonomous Ca(2+) signaling, established by SERCA, in iterative budding (Truong, 2017).
SERCA performs diverse regulatory functions, ranging from roles in periodic contractility in muscle to ER stress and protein folding. The current findings reveal a new function for SERCA, as a conserved controller of iterative budding. The initiation of new buds and encoding of the timing of formation of these buds has been proposed to be controlled by growth factor morphogens. Specifically, FGF10 acting on airway epithelial FGFR2b (in mammals) or Branchless acting on Breathless (in Drosophila) are required for proper branching of mammalian lungs or Drosophila trachea, respectively. Unidentified morphogens have also been proposed to act as a 'branching clock' that work with FGF signaling to coordinate the branching program. In contrast to this hypothesis that unidentified growth factor morphogens serve as the 'clock' to direct the timing of branching, this study showsthat SERCA is a central organizer that directs the onset and rate of budding. Morphogens must operate upstream of SERCA, because SERCA blockade stalls the branching program, while supply of exogenous morphogens (e.g. FGFs) is insufficient to overcome this blockade. Thus, it is proposed that SERCA must integrate inputs from morphogens like FGF and establishes a differential in Ca2+ levels at branching tips to indicate the timing for directed cell migration and branch formation (Truong, 2017).
This novel role of SERCA as a central organizer of branching seems highly conserved, as branching in both invertebrates and vertebrates, as well as tissues from all germ layers, requires SERCA. In all these systems, branch iteration rate is determined by the level of SERCA function; these effects are mediated by controlling cell migration. SERCA's effects are not mediated by altering cell shape, and do not require alterations in proliferation. Live Ca2+ imaging in Drosophila reveals that SERCA directs cell migration at branch points by establishing a local Ca2+ differential, where the Ca2+ level is higher in the leading cell that migrates to form a new branch. The cells trailing behind it maintain comparatively lower Ca2+ levels. Loss of this local Ca2+ differential halts migration and branching. Reinstatement of this local Ca2+ differential, whether by lifting of SERCA blockade or by PKC activation, restores cell migration and branching (Truong, 2017).
Beyond the Ca2+ differential revealed by light-sheet imaging of Drosophila embryos, episodic Ca2+ impulses were observed to propagate through the tracheal epithelium as the cells migrate and fuse to form their branched network. These propagating Ca2+ waves have been predicted by computational modeling, yet they do not appear to be important for branch iteration, raising the question as to their function. A recent publication on tracheal tube anastomosis did not implicate these whole-cell Ca2+ impulses in membrane fusion. The increase in frequency of these impulses upon fusion of cells from adjacent segments suggests they may be a response to cell-cell contact, which could in turn modulate cell membrane machinery. Similar Ca2+ impulses have been described in other cell types, such as in fungi following contact with a pathogen. The remarkable similarity of these Ca2+ impulses from animals to fungi suggests that they are highly conserved and may have been adapted by evolution to suit each specific cellular environment. The function of these impulses may, alternatively, relate to maintenance or elongation of the branched network that has formed. Indeed, in mammalian lung, periodic Ca2+ waves course through airway smooth muscle, inducing waves of contractility. These waves are thought to mechano-regulate branching morphogenesis, whereby abolishing the Ca2+ waves impairs airway growth and elongation (Truong, 2017).
The results consistently demonstrate that SERCA instructs budding across germ layers, tissue types, and species, suggesting that the role of SERCA may be more broadly generalizable. A conserved regulator simplifies understanding of how a vast array of branched tissues could arise from one platform, and specialize based on local morphogen inputs. Thus, the current findings may unite disparate observations of Ca2+ signaling involvement in other types of branching, such as axonal pathfinding, plant gravitotropism, angiogenesis, and endothelial wound healing. A centralized control of branching also holds implications for understanding a range of disease mechanisms. Regarding the lung, the significance of reduced epithelial SERCA function has been highlighted in human and animal studies of asthma as well as in other burgeoning diseases such as cystic fibrosis, lung fibrosis and lung cancer. This study suggests that these oft intractable pulmonary challenges may feature SERCA-related lesions of cell migration. Examples include airway remodeling in asthma, alveolar remodeling in fibrosis, or lung cancer invasiveness. More generally, altered SERCA expression or function has been associated with numerous cancers, and changes in SERCA expression have been reported during cell lineage differentiation. Therefore, a wider opportunity may lie in determining how SERCA-mediated Ca2+ switching helps cells find not just their route, but also their fate (Truong, 2017).
Branching morphogenesis helps increase the efficiency of gas and liquid transport in many animal organs. Studies in several model organisms have highlighted the molecular and cellular complexity behind branching morphogenesis. To understand this complexity, computational models have been developed with the goal of identifying the "major rules" that globally explain the branching patterns. These models also guide further experimental exploration of the biological processes that execute and maintain these rules. This paper introduces the tracheal gills of mayfly (Ephemeroptera) larvae as a model system to study the generation of branched respiratory patterns. First, the gills of the mayfly Cloeon dipterum are described, and the geometry of its branching trachea is quantitatively characterized. This characterization was extended to those of related species to generate the morphospace of branching patterns. Then, it was shown how an algorithm based on the "space colonization" concept (SCA) can generate this branching morphospace via growth towards a hypothetical attractor molecule (M). SCA differs from other branch-generating algorithms in that the geometry generated depends to a great extent on its perception of the "external" space available for branching, uses few rules and, importantly, can be easily translated into a realistic "biological patterning algorithm". A gene was identified in the C. dipterum genome (Cd-bnl) that is orthologous to the fibroblast growth factor branchless (bnl), which stimulates growth and branching of embryonic trachea in Drosophila. In C. dipterum, this gene is expressed in the gill margins and areas of finer tracheolar branching from thicker trachea. Thus, Cd-bnl may perform the function of M in our model. Finally, this general mechanism is discussed in the context of other branching pattern-generating algorithms (Ruiz-Sobrino, 2020).
Biological tubes are essential for animal survival, and their functions are critically dependent on tube shape. Analyzing the contributions of cell shape and organization to the morphogenesis of small tubes has been hampered by the limitations of existing programs in quantifying cell geometry on highly curved tubular surfaces and calculating tube-specific parameters. This study describes QuBiT (Quantitative Tool for Biological Tubes) and used it to analyze morphogenesis of embryonic Drosophila trachaea (airway). In the main tube, previously unknown anterior-to-posterior (A-P) gradients were found of cell apical orientation and aspect ratio and periodicity in the organization of apical cell surfaces. Inferred cell intercalation during development dampens an A-P gradient of the number of cells per cross-section of the tube, but does not change the patterns of cell connectivity. Computationally "unrolling" the apical surface of WT trachea and the hindgut reveals previously unrecognized spatial patterns of the apical marker Uninflatable and a non-redundant role for the Na(+)/K(+) ATPase in apical marker organization. These unexpected findings demonstrate the importance of a computational tool for analyzing small diameter biological tubes (Yang, 2019).
Tracheal cells use Rab GTPases to organize their internal membrane transport, resulting in the specific localization or secretion of proteins on the apical or basal membrane compartments. Some contributions of Rabs to junctional remodelling and governance of tracheal lumen contents are known, but it is reasonable to assume that they play important further roles in morphogenesis. This pertains in particular to terminal tracheal cells, specialized branch-forming cells that drastically reshape both their apical and basal membrane during the larval stages. A loss-of-function screen was performed in the tracheal system, knocking down endogenously tagged alleles of 26 Rabs by targeting the tag via RNAi. This revealed that at least 14 Rabs are required to ensure proper cell fate specification and migration of the dorsal branches, as well as their epithelial fusion with the contralateral dorsal branch. The screen implicated four Rabs in the subcellular morphogenesis of terminal cells themselves. Further tests suggested residual gene function after knockdown, leading to a discussion the limitations of this approach. It is concluded that more Rabs than identified here may be important for tracheal morphogenesis, and that the tracheal system offers great opportunities for studying several Rabs that have barely been characterized so far (Best, 2020).
Branching morphogenesis and seamless tube formation in Drosophila melanogaster are essential for the development of vascular and tracheal systems, and instructive in studying complex branched structures such as human organs. Zipper is a myosin II's actin-binding heavy chain; hence, it is important for contracting actin, cell proliferation, and cell sheet adhesion for branching of the tracheal system in post-larval development of the D. melanogaster. Nevertheless, the specific role of Zipper in the larva is still in question. This paper intended to investigate the specific role of Zipper in branching morphogenesis and lumenogenesis in early developmental stages. It did so by checking the localization of the protein in the cytoplasm of the terminal cells and also by analyzing the morphology of zipper RNAi loss-of-function mutants in regard to branching and lumen formation in the terminal cells. A rescue experiment of RNAi mutants was also performed to check the sufficiency of Zipper in branching morphogenesis. Confocal imaging showed the localization of Zipper in the cytoplasm of the terminal cells, and respective quantitative analyses demonstrated that zipper RNAi terminal cells develop significantly fewer branches. Such a result hinted that Zipper is required for the regulation of branching in the terminal cells of D. melanogaster. Nevertheless, Zipper is not significantly involved in the formation of seamless tubes. One hypothesis is that Zipper's contractility at the lateral epidermis' leading edge allows cell sheet movement and respective elongation; as a result of such an elongation, further branching may occur in the elongated region of the cell, hence defining branching morphogenesis in the terminal cells of the tracheal system (Shin, 2021).
Grainy head (Grh) is a conserved transcription factor (TF) controlling epithelial differentiation and regeneration. To elucidate Grh functions, embryonic Grh targets were identified by ChIP-seq and gene expression analysis. Grh was shown to control hundreds of target genes. Repression or activation correlates with the distance of Grh binding sites to the transcription start sites of its targets. Analysis of 54 Grh-responsive enhancers during development and upon wounding suggests cooperation with distinct TFs in different contexts. In the airways, Grh repressed genes encode key TFs involved in branching and cell differentiation. Reduction of the POU-domain TF, Vvl, (ventral veins lacking) largely ameliorates the airway morphogenesis defects of grh mutants. Vvl and Grh proteins additionally interact with each other and regulate a set of common enhancers during epithelial morphogenesis. It is concluded that Grh and Vvl participate in a regulatory network controlling epithelial maturation (Yao, 2017).
Grh controls epithelial development and regeneration in multiple organisms. ChIP-seq data provide a broad view of Grh-binding to its targets in all Grh-expressing tissues. The analysis of Grh-dependent regulatory sequences indicates that the majority of the 5599 peaks that include the consensus Grh-binding sequence identify true Grh targets. Hitherto, analysis of Grh targets in development focused on proteins involved in epidermal barrier formation, adhesion molecules and junctional proteins. Identification of functional Grh targets in the airways adds large groups of proteins involved in lipid metabolism, cell signaling and TFs. This suggests additional functions of Grh in tubulogenesis that might explain several of its additional roles. For example, the phenotype of grh mutants in the airways includes the selective expansion of the epithelial apical membranes, a phenotype that has not been detected in any of the mutants affecting junctional proteins or the formation and modification of the apical extracellular barrier. Definition of new Grh targets during airway maturation provides a rich resource for future studies addressing how Grh controls epithelial morphogenesis (Yao, 2017).
A prevalent group of Grh targets in the epidermis and airways includes genes involved in innate immune responses ranging from pattern recognition receptors to effectors. Interestingly, several putative GRHL2 targets in human bronchial epithelial cells, such as serpins and chitinase 3-like proteins, have been implicated in immune responses. Analysis of PGRP-LC reveals a direct role of Grh in endowing epithelial cells the ability to combat infections. Although the PGRP-LC reporter expression was not inducible by wounding or bacterial injection, it remains possible that Grh also directly controls the activation of epithelial immune responses upon infection. Indeed, partial inactivation of Grh by RNAi in adult flies resulted in increased morbidity and mortality upon bacterial infection (Yao, 2017).
Analysis of 47 new Grh-activated enhancers in epithelial development suggests the presence of distinct, tissue-specific Grh co-factors in the control of target genes in different epithelial cell types. The activation of some of these reporters upon injury expands the repertoire of Grh-activated enhancers and is in line with previous models proposing wound-induced interactions of Grh with other factors. These interactions could be induced by post-translational modifications of Grh or its co-factors by Rolled and other kinases downstream of Stit receptor kinase signaling and might facilitate the activation of transcription by Grh pre-bound to chromatin (Yao, 2017).
The ChIP-seq and gene expression analysis also reveal a potential role for Grh as a repressor. Such a repressor function of Grh is consistent with previous studies addressing Grh function on individual targets in flies and mammals. A higher correlation of PcG-binding sites and repressive chromatin marks were found around the Grh-binding sites of repressed targets as compared with the binding sites of activated genes. The positioning of Grh-binding sites relative to the TSS of repressed versus activated or unaffected genes also differs: Grh-binding sites are usually further from the TSS in repressed target genes. This observation is supported by the analysis of vvl ds3 and vvl 1.8, the only two identified repressible enhancers, which are located more than 2 kb from the vvl TSS. The difference in the structure of the repressed and activated Grh enhancers suggests that Grh repression might require chromatin looping and involve co-repressors. Further work is needed to elucidate a potential direct function of Grh in transcriptional repression (Yao, 2017).
A characteristic group of Grh targets in the airways includes TFs involved in epithelial cell differentiation. This resembles the complex regulatory functions of Grh during neuronal specification. For instance, in neuroblasts, Grh demarcates the last time window for TF expression by repressing Castor. In intermediate neural progenitors (INPs), Grh is detected in the 'middle-aged' INPs and overlaps with the expression of the TFs Dichaete and Eyeless. The three TFs cross-regulate each other. Similar cross-talk between Grh and its TF targets might specify and maintain epithelial differentiation. Since reduction of vvl in grh mutants largely ameliorates the tube elongation defects, the direct or indirect repression of genes encoding TFs is likely to be a crucial function of Grh in the airways. The shared expression pattern of Vvl and Grh, their binding to a set of common enhancers and their ability to form complexes suggest that they collectively control tube growth during airway maturation. Given their co-expression in other contexts, they might also co-operate during neural cell specification and epithelial immune responses (Yao, 2017).
The use of adult Drosophila melanogaster as a model for hematopoiesis or organismal immunity has been debated. Addressing this question, an extensive reservoir of blood cells (hemocytes) was identified at the respiratory epithelia (tracheal air sacs) of the thorax and head. Lineage tracing and functional analyses demonstrate that the majority of adult hemocytes are phagocytic macrophages (plasmatocytes) from the embryonic lineage that parallels vertebrate tissue macrophages. Surprisingly, no sign of adult hemocyte expansion was observed. Instead, hemocytes play a role in relaying an innate immune response to the blood cell reservoir: through Imd signaling and the Jak/Stat pathway ligand Upd3, hemocytes act as sentinels of bacterial infection, inducing expression of the antimicrobial peptide Drosocin in respiratory epithelia and colocalizing fat body domains. Drosocin expression in turn promotes animal survival after infection. This work identifies a multi-signal relay of organismal humoral immunity, establishing adult Drosophila as model for inter-organ immunity (Sanchez Bosch, 2019).
Drosophila melanogaster has greatly promoted understanding of innate immunity and blood cell development, but the capacity of the adult animal as a model remains a matter of debate. Most studies reported lack of new blood cell production and increasing immunosenescence, while one publication claimed continued hematopoietic activity in adult Drosophila (Sanchez Bosch, 2019).
Drosophila blood cells, or hemocytes, emerge from two lineages that persist into the adult, showing parallels with the two myeloid systems in vertebrates. First, hemocytes originating in the embryo parallel vertebrate tissue macrophages, as they quickly differentiate into plasmatocytes (macrophage-like cells), and subsequently proliferate extensively, mainly in the hematopoietic pockets (HPs) of the larva (Gold, 2014; Gold, 2015; Makhijani, 2011; Makhijani, 2012). At least some of these plasmatocytes can further differentiate into other blood cell types such as crystal cells and, under immune challenge, lamellocytes. Second, hemocytes originating in the lymph gland (LG) also give rise to plasmatocytes, crystal cells, and lamellocytes, yet in the lymph gland they are predominantly generated from blood cell progenitors (prohemocytes). At the beginning of metamorphosis, hemocytes from both the hematopoietic pockets and the lymph gland enter the open circulatory system and intermix. The subsequent fate and capacity of adult blood cells has remained largely unclear. Accordingly, this study comprehensively investigated the hematopoietic capacity of the blood cell system in adult Drosophila. A second part of this study focused on the role of adult blood cells in the humoral immune response, identifying a system of organismal innate immunity that centers on the respiratory epithelia in Drosophila (Sanchez Bosch, 2019).
Historically, Drosophila has been instrumental in the discovery of innate immunity and Toll like receptor (TLR) signaling. Toll and the related immune deficiency (Imd) signaling are evolutionary conserved NFκB family pathways, studied in detail regarding their upstream activation by pathogens and other inputs, and downstream signal transduction components and mechanisms. Targets include antimicrobial peptides (AMPs), which have been investigated for their transcriptional gene regulation and functional properties. TLR signaling has been well established also in vertebrate systems for its roles in infection and inflammation. However, it has been far less understood how multiple tissues or organs communicate with each other to elicit local innate immune responses (Sanchez Bosch, 2019).
Addressing these questions, this study clarifies basic principles of the blood cell system in adult Drosophila and its role in multi-tissue organismal immunity. An extensive blood cell reservoir was identified at the respiratory epithelia and fat body, its dynamics were investigated and probed for various signs of hematopoiesis. A key role of adult blood cells is demonstrated as sentinels of bacterial infection that trigger a humoral response in their reservoir, i.e., the respiratory epithelia and colocalizing domains of the fat body. This response culminates in the expression of the AMP gene Drosocin, which is shown to be significant for animal survival after bacterial infection. This work identifies Imd signaling and Upd3 expression in hemocytes as required steps in this relay of organismal immunity, laying the foundation for the use of adult Drosophila to dissect additional mechanisms of multi-tissue innate immunity in the future (Sanchez Bosch, 2019).
This study discovered a central role for an extensive blood cell reservoir at the respiratory epithelia and fat body of adult Drosophila. The reservoir serves as major receptacle of blood cells and foreign particles, and in addition executes a local humoral immune response of Drosocin expression that promotes animal survival after bacterial infection. Both functions are tied together by hemocytes acting as sentinels of infection, that signal through the Imd pathway and Upd3 to induce Drosocin expression in the tissues of their surrounding reservoir, i.e., the respiratory epithelia and colocalizing domains of the fat body (Sanchez Bosch, 2019).
Historic literature on Drosophila and other insects focused on the adult heart as the site of hemocyte accumulation. It described clusters of hemocytes at the ostia of the heart as 'immune organ', locations where hemocytes and bacteria accumulate. More recently, adult blood cell production at the heart was proposed (Ghosh, 2015). Some studies described functions of hemocytes in other locations, such as at the ovaries or along the gut of adult flies. Taking a more global cryosectioning approach afforded the identification of the largest reservoir of hemocytes in adult Drosophila, which surrounds the respiratory epithelia and is lined by fat body of the thorax and head. It is concluded that hemocytes and particles are delivered to these areas by the streaming hemolymph, even though the detailed anatomy of the open circulatory system remains to be mapped in more detail. Hemocytes may be physically caught in these locations, or in addition may engage in active adhesion. The intimate relationship of hemocytes with the respiratory epithelia, hemolymph, and adjacent fat body may serve interconnected roles, (1) guarding the respiratory epithelia as a barrier to the environment through functions of hemocytes in both phagocytosis and the induction of humoral immunity, and (2) facilitating gas exchange of hemocytes and nearby immune tissues, which in turn may again benefit defense functions. The former may be particularly advantageous in the defense against fungal pathogens that invade Drosophila via the tracheal system as primary route of infection, such as the entomopathogenic fungus B. bassiana. Regarding the latter, a study in caterpillars described the association of hemocytes with trachea, proposing a function for the respiratory system to supply hemocytes with oxygen (Sanchez Bosch, 2019).
Drosophila adult blood cells derive from two lineages: one that originates in the embryo and resembles vertebrate tissue macrophages, and another that produces blood cells in the lymph gland through a progenitor-based mechanism. It is estimated that more than 60% of adult hemocytes derive from the embryonic lineage is surprising, considering past views that the majority of adult hemocytes would derive from the lymph gland. It places more importance on the Drosophila embryonic lineage of hemocytes and suggests additional parallels with tissue macrophages in vertebrates, which persist into adulthood and form a separate myeloid system independent of the progenitor-derived monocyte lineage. Future research will show whether the relative contribution of the two hemocyte lineages to the adult blood cell pool will be the same or different under conditions of stress and immune challenges (Sanchez Bosch, 2019).
Given that embryonic-lineage plasmatocytes are highly proliferative in the hematopoietic pockets of the larva, and lymph gland hemocyte progenitors and some lymph gland plasmatocytes proliferate during larval development, the absence of hemocyte proliferation in the adult may be surprising. Nevertheless, combining the broad evidence supporting lack of significant hematopoietic activity in adult Drosophila, and evidence that Srp in adult Drosophila is not a progenitor marker, the findings robustly contradict an adult hematopoiesis model. The findings further reveal important differences to embryonic development, where Srp is required for the specification of undifferentiated prohemocytes. This study shows that during maturation of the adult animal, hemocytes relocate to the respiratory epithelia and the heart, upon completion of cytolysis of larval fat body cells, thereby refuting claims of new blood cell production at the heart. Similarly, seemingly increased numbers of fluorescently labeled hemocytes following bacterial infection are likely based on infection-induced upregulation of hemocyte-specific genes and their respective enhancers including the reporter HmlΔ-GAL4, UAS-GFP. Enhanced hemocyte expression of Hemolectin (Hml) and other hemocyte-specific markers post-infection has been described previously (Sanchez Bosch, 2019).
Taken together, this broad evidence speaks to a lack of significant hematopoietic capacity of the blood cell system in adult Drosophila. The findings are in agreement with other studies that have reported a lack of hemocyte proliferation in adult Drosophila and functional immunosenescence in aging flies. Despite the scope of conditions tested, the possibility cannot be excluded that some other specific immune challenge or stress might exist that would be potent enough to trigger proliferation- or differentiation-based blood cell production in adult Drosophila. Likewise, it cannot be excluded that adult Drosophila may possess small numbers of proliferation- and/or differentiation-competent progenitors that may have persisted e.g. from the lymph gland posterior lobes; such cells might give rise to new differentiated hemocytes, although according to the current data they would remain insignificant in number (Sanchez Bosch, 2019).
Taking into account the short reproductive phase and relatively short life span of Drosophila, the adult fly may be sufficiently equipped with the pool of hemocytes that is produced in the embryo and larva. In fact, hemocytes do not seem essential for the immediate survival of adult flies: Drosophila ablated of hemocytes, and mutants devoid of hemocytes, survive to adulthood although they are more prone to, and succumb more rapidly to infection. A model is proposed that places emphasis on larval development as the sensitive phase for the expansion and regulation of the adult blood cell pool. In the larva, hemocytes of both the embryonic and lymph gland lineage integrate signals from a variety of internal and external stimuli to adapt to existing life conditions (Sanchez Bosch, 2019).
This work reveals a role for hemocytes in a local humoral immune response of the fat body and respiratory epithelia. Previous studies on hemocyte-ablated flies have reported increases in Defensin and IM1 expression. In contrast, this study finds a positive role for hemocytes in the induction of Drosocin in tissues that form the hemocyte reservoir, i.e., the respiratory epithelia and fat body domains of the head and thorax. The concept of hemocytes promoting AMP expression in other tissues is well established. A role for AMP expression in surface epithelia that interface with the environment was reported in a previous study, and Drosocin expression was described in embryonic and larval trachea and the abdominal tracheal trunks of adult Drosophila, albeit not in the respiratory epithelia (Sanchez Bosch, 2019).
In adult Drosophila, hemocytes tightly localize between the respiratory epithelia and fat body tissue that occupies the space toward to the cuticle exoskeleton. It is proposed that this close anatomical relationship facilitates rapid local signaling. Consistent with previous knowledge that Drosocin expression is lost in imd mutant backgrounds, this study found that hemocyte-autonomous Imd signaling is required, albeit not sufficient, to trigger the infection-induced Drosocin response. Likewise, the Imd pathway upstream receptor PGRP-LC is required in hemocytes, suggesting that DAP-type peptidoglycan recognition and initiation of Imd signaling are a critical step in triggering the Drosocin response. Transcriptional induction of upd3 by Imd signaling is supported by putative Rel binding sites identified in the upd3 genomic region, two of which are even fully conserved across seven Drosophilids including Drosophila melanogaster. The data suggest roles for hemocyte-expressed upd3, and corresponding Jak/Stat signaling in cells of the fat body and respiratory system, all of which are required albeit not sufficient. Overactivation of the pathway paradoxically suppresses Drosocin expression, and even temporally restricted expression of activated hopTumL in trachea was largely lethal, possibly indicating leaky expression of the transgene. Overall, it can only be speculated that the unexpected effects of Jak/Stat overactivation might be due to activation of some negative feedback loop or other complex signaling changes that remain a matter of future investigation (Sanchez Bosch, 2019).
Several reports provide precedent for a role of hemocyte-expressed Upd3 in the induction of immune responses in other target tissues. Following septic injury, upregulation of upd3 in hemocytes triggers induction of stress peptide genes of the turandot family including totA in fat body. Similarly, in response to injury, hemocyte-produced Upd3 induces Jak/Stat signaling in the fat body and gut. Under lipid-rich diet, upd3 is induced in hemocytes, causing impaired glucose homeostasis and reduced lifespan in adult Drosophila. In the larva, hemocyte-derived Upd2 and -3 activate Jak/Stat signaling in muscle, which are required for the immune response against parasitic wasps. However, in the Drosocin response around the reservoir of hemocytes, the data predict that additional signal/s and/or signaling pathway/s are needed to initiate Drosocin expression and potentially restrict its expression to defined fat body domains of the head and thorax. Additional events may include signaling through Toll or other signaling pathways in hemocytes and/or other tissues including the respiratory epithelia and fat body. Likewise, other types of signals may be required, such as reactive oxygen species (ROS) or nitric oxide (NO), which play roles in the relay of innate immune responses to infection and stress, or non-peptide hormones including ecdysone, which confers competence in the embryonic tracheal Drosocin response to bacterial infection and enhances humoral immunity under conditions of dehydration. Lastly, there could be requirement for additional processing to make bacterial ligands accessible for receptors in other tissues, as has been reported for Psidin, a lysosomal protein required in blood cells for degradation of engulfed bacteria and expression of Defensin in the fat body, although this mechanism may not be universal in all systems (Sanchez Bosch, 2019).
This work reveals an active role of endogenous Drosocin expression in survival after bacterial infection. Since the cloning of Drosocin and its classification as inducible antibacterial peptide, Drosocin has been studied for its transcriptional regulation, illustrating its induction under a variety of bacterial and other immune challenges. Drosocin structure and antimicrobial function have been studied in vitro and by overexpression from transgenes in Drosophila and in heterologous vertebrate systems. Consistent with the current findings, a recent study on CRISPR-based Drosocin null mutants reached similar conclusions regarding the requirement of endogenous Drosocin expression for animal survival following E. cloacae infection (Hanson, 2019). Expanding from these findings, this study reveals the anatomical features of Drosocin expression and its unique path of induction. In addition to Drosocin's role in animal survival after bacterial infection, the data suggest contribution of Drosocin to animal survival after injury through PBS injection. Injury has emerged as a factor that affects survival, a phenomenon for which the molecular mechanisms still remain to be determined. Alternatively, considering that the fly surface and living conditions are not sterile and survival experiments are performed over extended periods of time, it cannot be ruled out that PBS injections may have led to inadvertent infection with some low level contaminating microbes. A role for endogenous Drosocin levels in the antimicrobial response is strongly supported by independent data in the literature. Specifically, the minimum inhibitory concentration (MIC) of Drosocin against E. coli and E. cloacae was determined to be well within the range or below the endogenous concentration of Drosocin in the Drosophila hemolymph (MIC is 1 or 2 μM for the glycosylated forms, and 8 or 10 μM for the unglycosylated form, respectively, compared to 40 μM Drosocin in the Drosophila hemolymph (Sanchez Bosch, 2019).
In conclusion, this study revokes the use of adult Drosophila as effective model to study hematopoiesis, and establishes it as promising system for organismal immunity centering on the immune signaling relay at the reservoir of blood cells. At the evolutionary level, this model shows parallels with vertebrate immune cells of the lung and innate immune responses to bacterial infection. The Drosophila model opens countless avenues for exciting future research, e.g., to investigate additional molecular and cellular mechanisms in the immune signaling relay, the role and regulation of the system in the defense against pathogens that invade the trachea as natural route of infection, the use of the same axis by gram-positive or non-bacterial pathogens, and the induction of other AMPs and immune effector genes in the same axis of regulation (Sanchez Bosch, 2019).
Tango1 enables ER-to-Golgi trafficking of large proteins. Loss of Tango1, in addition to disrupting protein secretion and ER/Golgi morphology, causes ER stress and defects in cell shape. The previously observed dependence of smaller cargos on Tango1 is a secondary effect. If large cargos like Dumpy, which this study identifies as a Tango1 cargo, are removed from the cell, nonbulky proteins reenter the secretory pathway. Removal of blocking cargo also restores cell morphology and attenuates the ER-stress response. Thus, failures in the secretion of nonbulky proteins, ER stress, and defective cell morphology are secondary consequences of bulky cargo retention. By contrast, ER/Golgi defects in Tango1-depleted cells persist in the absence of bulky cargo, showing that they are due to a secretion-independent function of Tango1. Therefore, maintenance of ER/Golgi architecture and bulky cargo transport are the primary functions for Tango1 (Rios-Barrera, 2017).
The endoplasmic reticulum (ER) serves as a major factory for protein and lipid synthesis. Proteins and lipoproteins produced in the ER are packed into COPII-coated vesicles, which bud off at ER exit sites (ERES) and then move toward the Golgi complex where they are sorted to their final destinations. Regular COPII vesicles are 60-90 nm in size, which is sufficient to contain most membrane and secreted molecules. The loading of larger cargo requires specialized machinery that allows the formation of bigger vesicles to accommodate these bulky molecules. Tango1 (Transport and Golgi organization 1), a member of the MIA/cTAGE (melanoma inhibitory activity/cutaneous T cell lymphoma-associated antigen) family, is a key component in the loading of such large molecules into COPII-coated vesicles. Molecules like collagens and ApoB (apolipoprotein B)-containing chylomicrons are 250-450 nm long and rely on Tango1 for their transport out of the ER, by physically interacting with Tango1 or Tango1 mediators at the ERES (Rios-Barrera, 2017).
Tango1 is an ER transmembrane protein that orchestrates the loading of its cargo into vesicles by interacting with it in the ER lumen. The interaction of Tango1 with its cargo then promotes the recruitment of Sec23 and Sec24 coatomers on the cytoplasmic side, while it slows the binding of the outer layer coat proteins Sec13 and Sec31 to the budding vesicle. This delays the budding of the COPII carrier. Tango1 also recruits additional membrane material to the ERES from the Golgi intermediate compartment (ERGIC) pool, thereby allowing vesicles to grow larger. It also interacts directly with Sec16, which is proposed to enhance cargo secretion. A shorter isoform of mammalian Tango1 lacks the cargo recognition domain but nevertheless facilitates the formation of megacarrier vesicles (Rios-Barrera, 2017).
Apart from bulky proteins, some heterologous, smaller proteins like secreted horseradish peroxidase (ssHRP, 44 kDa) and secreted GFP (27 kDa) also depend on Tango1 for their secretion . Unlike for collagen or ApoB, there is no evidence for a direct interaction between Tango1 and ssHRP or secreted GFP. It is not clear why Tango1 would regulate the secretion of these molecules, but it has been proposed that in the absence of Tango1, the accumulation of nonbulky proteins at the ER might be due to abnormally accumulated Tango1 cargo blocking the ER; however, this has not been tested experimentally (Rios-Barrera, 2017).
Drosophila Tango1 is the only member of the MIA/cTAGE family found in the fruit fly, which simplifies functional studies. Like vertebrate Tango1, the Drosophila protein participates in the secretion of collagen. And as in vertebrates, ssHRP, secreted GFP, and other nonbulky molecules like Hedgehog-GFP also accumulate in the absence of Tango1. These results have led to the proposal that Tango1 participates in general secretion. However, most of the evidence for these conclusions comes from overexpression and heterologous systems that might not reflect the physiological situation (Rios-Barrera, 2017).
This study describes a tango1 mutant allele that was identified in a mutagenesis screen for genes affecting the structure and shape of terminal cells of the Drosophila tracheal system. Tracheal terminal cells form highly ramified structures with branches of more than 100 μm in length that transport oxygen through subcellular tubes formed by the apical plasma membrane. Their growth relies heavily on membrane and protein trafficking, making them a very suitable model to study subcellular transport. Terminal cells were used to study the function of Tango1, and loss of Tango1 was found to affect general protein secretion indirectly, and it also leads to defects in cell morphology and in the structure of the ER and Golgi. The defects in ER and Golgi organization of cells lacking Tango1 persist even in the absence of Tango1 cargo (Rios-Barrera, 2017).
These studies led to an explanation of why, in the absence of Tango1, nonbulky proteins accumulate in the ER despite not being direct Tango1 cargos. These cargos are retained in the ER as a consequence of nonsecreted bulky proteins interfering with their transport. However, the effect of loss of Tango1 on ER/Golgi morphology can be uncoupled from its role in bulky cargo secretion (Rios-Barrera, 2017).
This study has described a role of Tango1, which was initially identified through its function in tracheal terminal cells and other tissues in Drosophila embryos, larvae, and pupae. Due to their complex shapes and great size, terminal cells are a well-suited system to study polarized membrane and protein trafficking, with the easily scorable changes in branch number and maturation status providing a useful quantitative readout that serves as a proxy for functional membrane and protein trafficking machinery. Moreover, this analyses are conducted in the physiological context of different tissues in the intact organism (Rios-Barrera, 2017).
The loss-of-function allele tango12L3443 has a stop codon eight amino acids downstream of the PRD domain and eliminates the 89 C-terminal amino acids of the full-length protein. It is unlikely that the mutation leads to a complete loss of function. First, terminal cells expressing an RNAi construct against tango1 show stronger defects, with fewer branches per cell than homozygous tango12L3443 cells. Second, the mutant protein appears not to be destabilized nor degraded, but instead is present at apparently normal levels, albeit at inappropriate sites. Predictions of the deleted fragment of the protein suggest it is disorganized and that it contains an arginine-rich domain that has no known interaction partners and that is not present in human Tango1. In homozygous mutant terminal cells, the mutant tango12L3443 protein fails to localize at ERES. In mammalian Tango1, the Sec16-interacting region within the PRD domain is necessary for the localization of Tango1 to the ERES and for its interaction with Sec23 and Sec16, but since this domain is fully present in tango12L3443, the results mean that either the missing 89 C-terminal amino acids contain additional essential localization signals, or that the PRD domain is structurally affected by the truncation of the protein. The latter is considered less likely, as a truncation of eight amino acids downstream of the PRD domain is unlikely to destabilize the polyproline motifs, especially as the overall stability of the protein does not seem to be affected. Furthermore, this region shows a high density of phosphoserines (Ser-1345, Ser-1348, Ser-1390, and Ser-1392), suggesting it might serve as a docking site for adapter proteins or other interactors (Rios-Barrera, 2017).
Terminal cells lacking Tango1 have fewer branches than control cells and are often not properly filled with air. This loss-of-function phenotype is not due to a direct requirement for Tango1, as it is suppressed by the simultaneous removal of Dumpy (Dpy), an extracellular protein involved in epidermal-cuticle attachment, aposition of wing surfaces and trachea development. It also cannot be explained by the individual loss of crb, Piopio (Pio) a zona pellucida (ZP) domain protein that mediates the adhesion of the apical epithelial surface and the overlying apical extracellular matrix, or dpy, since knocking down any of these genes has no effect on cell morphology. Instead, it is proposed that the cell morphological defect is due at least in part to the activation of the ER stress response, since expression of Xbp1 is sufficient to recapitulate the phenotype. Xbp1 regulates the expression of genes involved in protein folding, glycosylation, trafficking, and lipid metabolism. It is possible that one or a small number of specific genes downstream of Xbp1 are responsible for defective branch formation or stability, but the phenotype could also be a secondary consequence of the physiological effects of the ER stress response itself, for example, a failure to deliver sufficient lipids and membrane from the ER to the apical plasma membrane (Rios-Barrera, 2017).
Collagen, with a length of 300 nm, and ApoB chylomicrons with a diameter of > 250 nm, have both been biochemically validated as Tango1 cargos. These molecules are not expressed in terminal cells, and therefore it was clear that Tango1 must have a different substrate in these cells. Given that Tango1 is known for the transport of bulky cargo, that Dpy is the largest Drosophila protein at 800 nm length, and that Dpy vesicles are associated with Tango1 rings in tracheal cells, it is proposed that Dpy is a further direct target of Tango1. Colocalization of Tango1 with its cargo has also been observed in other tissues: with collagen in Drosophila follicle cells and with ApoB in mammalian cell lines (Rios-Barrera, 2017).
No regions of sequence similarity that could represent Tango1-binding sites have been found in Tango1 cargos. There are several possible explanations for this. First, these proteins may contain binding motifs, but the motifs are purely conformational and not represented in a linear amino acid sequence. There is no evidence for or against this hypothesis, but it would be highly unusual, and there is support for alternative explanations. Thus, as a second possibility, all three proteins may require Tango1 for their secretion, but variable adapters could mediate the interactions. In vertebrates, Tango1 can indeed interact with its cargo through other molecules; for instance, its interaction with collagen is mediated by Hsp47. However, in Drosophila, there is no Hsp47 homolog. In the case of ApoB, it has been suggested that microsomal triglyceride transfer protein (MTP) and its binding partner, protein disulphide isomerase (PDI), might associate with Tango1 and TALI to promote ApoB chylomicrons loading into COPII vesicles. Evidence supporting this is that the lack of MTP leads to ApoB accumulation at the ER. It is not known if secretion of other Tango1 cargos like collagen or Dpy also depends on MTP and PDI, but PDI is known also to form a complex with the collagen-modifying enzyme prolyl 4-hydroxylase. Previous work has shown that terminal cells lacking MTP show air-filling defects and fail to secrete Pio and Uninflatable to the apical membrane, and that loss of MTP in fat body cells also affects lipoprotein secretion, as it does in vertebrates. Since cells lacking MTP or Tango1 have similar phenotypes, it is plausible that the MTP function might be connected to the activity of Tango1 (Rios-Barrera, 2017).
The data is interpreted to mean that in the absence of Tango1, primary cargo accumulates in the ER, and in addition, there are secondary, indirect effects that can be suppressed by reducing the Tango1 cargo that overloads the ER. The secondary effects include activation of the ER stress response and intracellular accumulation of other trafficked proteins like Crb, laminins, and overexpressed proteins and probably also the accumulation of heterologous proteins like secreted HRP or GFP in other systems (Rios-Barrera, 2017).
The data suggest that primary and secondary cargo reach the ERES but fail to be trafficked further along the secretory pathway. In this model, primary cargo, probably recruited by adaptors, would be competing with other secondary cargo for ERES/COPII availability, creating a bottleneck at the ERES. This is consistent with recent experiments that show that in tango1-knockdown HeLa cells, VSVG-GFP trafficking does not stop completely, but is delayed. Furthermore, in these experiments, VSVG-GFP is mostly seen in association with Sec16 and Sec31, supporting the clogging model (Rios-Barrera, 2017).
It is not immediately clear why cargo accumulation in terminal cells lacking Tango1 affects the secretion of Crb but not of βPS integrin. While steady states are looked at in this analyses, Maeda (2016) measured the dynamics of secretion and found that loss of Tango1 leads to a reduced rate of secretion of VSVG-GFP, an effect that would have been missed for any proteins the current study classified as not affected by loss of Tango1. Irrespective, a range of mechanisms can be thought of that might be responsible for this difference, including alternative secretion pathways and differences in protein recycling. Alternative independent secretory pathways have been reported in different contexts. For instance, while both αPS1 and βPS integrin chains depend on Sec16 for their transport, the αPS1 chain can bypass the Golgi apparatus and can instead use the dGRASP-dependent pathway for its transport. It would be possible then that in terminal cells, βPS integrin is also trafficked through an alternative pathway that is not affected by loss of Tango1. Similarly, tracheal cells lacking Sec24-CD accumulate Gasp, Vermiform, and Fasciclin III, but not Crb, supporting a role for alternative secretion pathways for different proteins, as has been already proposed. Following this logic, overexpressed βPS integrin would then also be trafficked through a different route from that of the endogenous βPS integrin, possibly because of higher expression levels or because of the presence of the Venus fused to the normal protein (Rios-Barrera, 2017).
Drosophila Tango1 was initially found to facilitate collagen secretion in the fat body. More recently, the accumulation of other nonbulky proteins at the ER in the absence of Tango1 has led to the proposal of two models to explain these results: one in which Tango1 regulates general secretion, and the second one where Tango1 is specialized on the secretion of ECM components, since loss of Tango1 leads to the accumulation of the ECM molecules SPARC and collagen. The current results suggest a third explanation, where cargo accumulation in the ER might not necessarily be a direct consequence of only the loss of Tango1. Instead, in addition to depending on Tango1, some proteins of the ECM appear also to depend on each other for their efficient secretion. This is the case for laminins LanB1 and LanB2, which require trimerization before exiting the ER, while LanA can be secreted as a monomer. Loss of collagen itself leads to the intracellular accumulation of ECM components in fat body cells, such as the laminins and SPARC. Conversely, SPARC is required for proper collagen and laminin secretion and assembly in the ECM. Furthermore, intricate biochemical interactions take place between ECM components. Hence, due to the complex genetic and biochemical interactions between ECM components, the dependence of any one of them on Tango1 is difficult to determine without further biochemical evidence. The concept of interdependent protein transport from the ER as such is not new, as it has also been observed in other systems, for instance in immune complexes. During the assembly of T-cell receptor complexes and IgM antibodies, subunits that are not assembled are retained in the ER and degraded (Rios-Barrera, 2017).
Nevertheless, these observations in glial cells, which express laminins but not collagen, allow at least these requirements to be partly separate. This study found that laminins are accumulated due to general ER clogging and not because they rely on Tango1 for their export. This is based on observations that once the protein causing the ER block is removed, laminin secretion can continue in the absence of Tango1. It is still unclear why glial cells can secrete laminins in the absence of collagen whereas fat body cells cannot, but presumably laminin secretion can be mediated by different, unidentified cargo receptors expressed in glial cells (Rios-Barrera, 2017).
This study found that Sec16 forms aberrant aggregates in cells lacking Tango1, as in mammalian cell lines, and that the number of Sec16 particles is reduced. Other studies have shown that Tango1 overexpression produces larger ERES, and that Tango1 and Sec16 depend on each other for localization to ERES. In addition, lack of Tango1 also affects the distribution of Golgi markers. Thus, Tango1 influences not only the trafficking of cargos, but also the morphology of the secretory system (Rios-Barrera, 2017).
It had been suggested that the disorganization of ER and Golgi apparatus in cells lacking Tango1 might be an indirect consequence of the accumulation of Tango1 cargo. The work of Maeda (2016) has provided a possible explanation for the molecular basis, and proposed that Tango1 makes general secretion more efficient, but it has not formally excluded the possibility that the primary cause for the observed defects is secretory protein overload. This study has now shown that this is not the case: In the absence of Tango1, an aberrant ER and Golgi morphology is still observed even after the main primary substrates of Tango1 were removed and, thereby, secretion of other molecules was restored and the ER stress response was prevented (Rios-Barrera, 2017).
ERGIC53 accumulates at the ERES in the absence of Tango1, and this can be partly reversed by removing dpy. This is in apparent contradiction to findings in mammalian cells where Tango1 was necessary for the recruitment of membranes containing ERGIC53 to the budding collagen megacarrier vesicle. However, ERGIC53 also has Tango1-independent means of reaching the ER. The current results indicate that its retrieval from the ER to the ERGIC compartment depends directly or indirectly partly on Tango1. As a cargo receptor for glycoproteins, ERGIC53 may be retained at the ERES as a consequence of the accumulation of its own cargo at these sites. This would mean that it cannot be delivered back to the ERGIC or the cis-Golgi apparatus for further rounds of retrograde transport, which may, in turn, be an explanation for the enlargement of the Golgi matrix protein 130 kD (GM130) compartment seen after Tango1 knockdown (Rios-Barrera, 2017).
The finding that Tango1-depleted cells have a functional secretory pathway despite the ER-Golgi disorganization was unexpected. Stress stimuli like amino acid starvation (but not ER stress response itself) lead to Sec16 translocation into Sec bodies and inhibition of protein secretion. However, uncoupling of ER-Golgi organization from functional secretion has also been observed in other contexts. Loss of Sec23 or Sec24-CD leads to a target peptide sequence KDEL appearing in aggregates of varying sizes and intensities similar to those observed for Sec16 and for KDEL-RFP in cells lacking tango1. Also, GM130 is reduced in Sec23 mutant embryos. Despite these structural problems, these embryos do not show generalized secretion defects and also do not affect the functionality of the Golgi apparatus, as determined by glycosylation status of membrane proteins (Rios-Barrera, 2017).
Thus, Tango1 appears to have an important structural function in coordinating the organization of the ER and the Golgi apparatus, and this, in turn, may enhance vesicle trafficking. This fits with the role of Tango1 in recruiting ERGIC membranes to the ERES, and also with the effects of loss of Tango1 in the distribution of ER and Golgi markers. It has been proposed that the ER and Golgi apparatus in insects, which unlike in mammalian cells is not centralized but spread throughout the cytoplasm, is less efficient for secretion of bulky cargo than mammalian cells that can accommodate and transport it more efficiently through the Golgi ribbon. This difference could explain why tango1 knockout mice seem to have only collagen secretion defects and die only as neonates. However, a complete blockage of the ER might also be prevented by the activity of other MIA3/cTAGE5 family homologs. In mammalian cell culture experiments, even if loss of tango1 affects secretion of HRP, the secretion of other overexpressed molecules like alkaline phosphatase is not affected. This could also be because of the presence of other MIA3/cTAGE5 family homologs. By contrast, because there are no other MIA3/cTAGE5 family proteins in Drosophila, loss of tango1 may lead to the accumulation of a wider range of overexpressed proteins and more overt mutant phenotypes than in mammals (Rios-Barrera, 2017).
The fly trachea is the equivalent of the mammalian lung and is a useful model for human respiratory diseases. However, little is known about the molecular mechanisms underlying tracheal air filling during larval development. This study discovered that PTPMT1 has a function in tracheal air filling. PTPMT1 is a widely conserved, ubiquitously expressed mitochondrial phosphatase. To reveal PTPMT1's functions in genetically tractable invertebrates and whether those functions are tissue specific, a Drosophila model of PTPMT1 depletion was generated. Fly PTPMT1 mutants showed impairments in tracheal air filling and subsequent activation of innate immune responses. On a cellular level, these defects are preceded by aggregation of mitochondria within the tracheal epithelial cells. This work demonstrates a cell-type-specific role for PTPMT1 in fly tracheal epithelial cells to support air filling and to prevent immune activation. The establishment of this model will facilitate exploration of PTPMT1's physiological functions in vivo (Papakyrikos, 2020).
Plasma membranes fulfil many physiological functions. In polarized cells, different membrane compartments take on specialized roles, each being allocated correct amounts of membrane. The Drosophila tracheal system, an established tubulogenesis model, contains branched terminal cells with subcellular tubes formed by apical plasma membrane invagination. This study shows that apical endocytosis and late endosome-mediated trafficking are required for membrane allocation to the apical and basal membrane domains. Basal plasma membrane growth stops if endocytosis is blocked, whereas the apical membrane grows excessively. Plasma membrane is initially delivered apically and then continuously endocytosed, together with apical and basal cargo. An organelle is described carrying markers of late endosomes and multivesicular bodies (MVBs) that is abolished by inhibiting endocytosis and which is suggested to act as transit station for membrane destined to be redistributed both apically and basally. This is based on the observation that disrupting MVB formation prevents growth of both compartments (Mathew, 2020).
Coordination of stem cell function by local and niche-derived signals is essential to preserve adult tissue homeostasis and organismal health. The vasculature is a prominent component of multiple stem cell niches. However, its role in adult intestinal homeostasis remains largely understudied. This study has uncover a previously unrecognised crosstalk between adult intestinal stem cells in Drosophila and the vasculature-like tracheal system, which is essential for intestinal regeneration. Following damage to the intestinal epithelium, gut-derived reactive oxygen species activate tracheal HIF-1α and bidirectional FGF/FGFR signalling, leading to reversible remodelling of gut-associated terminal tracheal cells and intestinal stem cell proliferation following damage. Unexpectedly, reactive oxygen species-induced adult tracheal plasticity involves downregulation of the tracheal specification factor trachealess (trh) and upregulation of IGF2 messenger RNA-binding protein (IGF2BP2/Imp). These results reveal an intestine-vasculature inter-organ communication programme that is essential to adapt the stem cell response to the proliferative demands of the intestinal epithelium (Perochon, 2021).
The Drosophila trachea, as the functional equivalent of mammalian blood vessels, senses hypoxia and oxygenates the body. This study shows that the adult intestinal tracheae are dynamic and respond to enteric infection, oxidative agents and tumours with increased terminal branching. Increased tracheation is necessary for efficient damage-induced intestinal stem cell (ISC)-mediated regeneration and is sufficient to drive ISC proliferation in undamaged intestines. Gut damage or tumours induce HIF-1α (Sima in Drosophila), which stimulates tracheole branching via the FGF (Branchless (Bnl))-FGFR (Breathless (Btl)) signalling cascade. Bnl-Btl signalling is required in the intestinal epithelium and the trachea for efficient damage-induced tracheal remodelling and ISC proliferation. Chemical or Pseudomonas-generated reactive oxygen species directly affect the trachea and are necessary for branching and intestinal regeneration. Similarly, tracheole branching and the resulting increase in oxygenation are essential for intestinal tumour growth. This study has identified a mechanism of tracheal-intestinal tissue communication, whereby damage and tumours induce neo-tracheogenesis in Drosophila, a process reminiscent of cancer-induced neoangiogenesis in mammals (Tamamouna, 2021).
Extensive remodeling of the airways is a major characteristic of chronic inflammatory lung diseases such as asthma or chronic obstructive pulmonary disease (COPD). To elucidate the importance of a deregulated immune response in the airways for remodeling processes, a matching Drosophila model was established. Here, triggering the Imd (immune deficiency) pathway in tracheal cells induced organ-wide remodeling. This structural remodeling comprises disorganization of epithelial structures and comprehensive epithelial thickening. These structural changes do not depend on the Imd pathway's canonical branch terminating on nuclear factor κB (NF-κB) activation. Instead, activation of a different segment of the Imd pathway that branches off downstream of Tak1 and comprises activation of c-Jun N-terminal kinase (JNK) and forkhead transcription factor of the O subgroup (FoxO) signaling is necessary and sufficient to mediate the observed structural changes of the airways. These findings imply that targeting JNK and FoxO signaling in the airways could be a promising strategy to interfere with disease-associated airway remodeling processes (Wagner, 2021).
Membrane trafficking plays many roles in morphogenesis, from bulk membrane provision to targeted delivery of proteins and other cargos. In tracheal terminal cells of the Drosophila respiratory system, transport through late endosomes balances membrane delivery between the basal plasma membrane and the apical membrane, which forms a subcellular tube, but it has been unclear how the direction of growth of the subcellular tube with the overall cell growth is coordinated. This study shows that endosomes also organize F-actin. Actin assembles around late endocytic vesicles in the growth cone of the cell, reaching from the tip of the subcellular tube to the leading filopodia of the basal membrane. Preventing nucleation of endosomal actin disturbs the directionality of tube growth, uncoupling it from the direction of cell elongation. Severing actin in this area affects tube integrity. These findings show a new role for late endosomes in directing morphogenesis by organizing actin, in addition to their known role in membrane and protein trafficking (Rios-Barrera, 2022).
Biological tubes are fundamental units of most metazoan organs. Their defective morphogenesis can cause malformations and pathologies. An integral component of biological tubes is the extracellular matrix, present apically (aECM) and basally (BM). Studies using the Drosophila tracheal system established an essential function for the aECM in tubulogenesis. This study demonstrates that the BM also plays a critical role in this process. BM components are deposited in a spatial-temporal manner in the trachea. Laminins, core BM components, control size and shape of tracheal tubes and their topology within the embryo. At a cellular level, laminins control cell shape changes and distribution of the cortical cytoskeleton component α-spectrin. Finally, the BM and aECM act independently-yet cooperatively-to control tube elongation and together to guarantee tissue integrity. These results unravel key roles for the BM in shaping, positioning, and maintaining biological tubes (Klussmann-Fricke, 2022).
Early life environmental influences such as exposure to cigarette smoke (CS) can disturb molecular processes of lung development and thereby increase the risk for later development of chronic respiratory diseases. Among the latter, asthma and chronic obstructive pulmonary disease (COPD) are the most common. The airway epithelium plays a key role in their disease pathophysiology but how CS exposure in early life influences airway developmental pathways and epithelial stress responses or survival is poorly understood. Using Drosophila melanogaster larvae as a model for early life, this study demonstrates that CS enters the entire larval airway system, where it activates cyp18a1 which is homologues to human CYP1A1 to metabolize CS-derived polycyclic aromatic hydrocarbons and further induces heat shock protein 70. RNASeq studies of isolated airways showed that CS dysregulates pathways involved in oxidative stress response, innate immune response, xenobiotic and glutathione metabolic processes as well as developmental processes (BMP, FGF signaling) in both sexes, while other pathways were exclusive to females or males. Glutathione S-transferase genes were further validated by qPCR showing upregulation of gstD4, gstD5 and gstD8 in respiratory tracts of females, while gstD8 was downregulated and gstD5 unchanged in males. ROS levels were increased in airways after CS. Exposure to CS further resulted in higher larval mortality, lower larval-pupal transition, and hatching rates in males only as compared to air-exposed controls. Taken together, early life CS induces airway epithelial stress responses and dysregulates pathways involved in the fly's branching morphogenesis as well as in mammalian lung development. CS further affected fitness and development in a highly sex-specific manner (Sirocko, 2022).
Larval terminal cells of the Drosophila tracheal system generate extensive branched tubes, requiring a huge increase in apical membrane. Terminal cells were compromised for apical membrane expansion - mTOR-vATPase axis and apical polarity mutants - were invaded by the neighboring stalk cell. The invading cell grows and branches, replacing the original single intercellular junction between stalk and terminal cell with multiple intercellular junctions. This study characterize disjointed, a mutation in the same phenotypic class. disjointed encodes Drosophila Archease, required for RNA ligase (RtcB) function critical for tRNA maturation and ER stress-regulated nonconventional splicing of Xbp1 mRNA. The steady-state subcellular localization of Archease is principally nuclear, and dependent upon TOR-vATPase activity. In tracheal cells mutant for Rheb or vATPase, Archease localization shifted dramatically from nucleus to cytoplasm. Further, this study found that blocking tRNA maturation by knockdown of tRNAse z also induced compensatory branching. Taken together, these data suggest that the TOR-vATPase axis promotes apical membrane growth in part through nuclear localization of Archease, where Archease is required for tRNA maturation (Francis, 2022).
The Drosophila tracheal system consists of an interconnected network of branched tubes. The system originates from ten pairs of epithelial sacs composed of roughly 80 cells each. During embryogenesis, these sacs, which have invaginated from the surface epithelium, undergo successive rounds of branching. In primary branching, tip cells actively migrate and pull trailing cells along behind them; in this manner, typical sacs give rise to six primary branches. The largest tubes are multicellular and smaller tubes are autocellular (surrounding a lumenal space and sealing by formation of autocellular junctions) or seamless (elaborating an internal apical membrane that extends through the branches of stellate-shaped terminal cells). Secondary and tertiary branching are accompanied by specification of 'fusion' cells that mediate anastomoses with neighboring branches, and terminal cells extend blind-ended seamless tubes to target tissues. Embryonic tracheal terminal cells are the ultimate cells in tracheal branches and are connected to the network via intercellular junctions with their neighboring autocellular stalk cells. Terminal cells initially form a single intracellular blind-ended tube, but, subsequently, during larval life, sprout dozens of long, highly branched 'seamless' tubes. These tubes are initially liquid-filled, but undergo gas-filling at molts. During each instar, new larger tubes are constructed around the pre-existing gas-filled tubes, and become gas-filled themselves at the molt. In terminal cells, the new tubes are not only larger, but include new branches targeting additional tissues (Francis, 2022).
The apical (lumenal) membrane domain of terminal cells must expand dramatically to permit new tubes to ramify adequately on internal tissues, thus permitting gas exchange. Mutations that restrict the generation of apical membrane in terminal cells, such as vATPase and Rheb mutations, or that directly affect the generation of apical polarity, such as aPKC and par-6 mutations, cause compensatory growth of the neighboring wild-type stalk cells, which lengthen, invade and branch within the mutant terminal cells. Mutations in two vATPase genes were isolated based on the presence of a gas-filling defect in what appeared to be the terminal cell tube, adjacent to the position where the intercellular junction connecting terminal (seamless tubes) and stalk (autocellular tube) cells would normally be found. In fact, the tube lacking gas-filling is actually that of the invading stalk cell - the failure to gas-fill may be related to the remodeling that occurs as an autocellular tube replaces the seamless tube, or may reflect the fact that liquid clearance typically occurs during molts and newly made tubes remain liquid filled in-between molts. Because the autocellular tubes of the stalk cell penetrate dozens of microns into the compromised terminal cell, and branch, the result is pairs of terminal and stalk cells with multiple intercellular junctions marking the connections of stalk cell-derived autocellular tubes - surrounded by terminal cell cytoplasm - and terminal cell-derived seamless tubes (Francis, 2022).
In a previously described forward genetic screen, mutations were identified in four loci that caused related gas-filling defects. Upon further characterization, one of these loci, lotus (Nsf2), was distinct, preventing the formation of a stable connection between autocellular and seamless tubes, whereas mutations in the other three loci induced compensatory growth from neighboring wild-type stalk cells, as described above. Two of those loci were identified as components of the vATPase: oak gall (Vha26) and conjoined (Vha13). This study identified the remaining locus, disjointed, as encoding the Drosophila Archease. Although evolutionarily ancient, relatively few studies have focused on Archease. Compelling work has established a requirement for Archease as a co-factor for the RNA ligase, RtcB. Together, Archease and RtcB play an essential role in transfer RNA (tRNA) maturation and in nonconventional splicing of X-box binding protein 1 (Xbp1) mRNA (Francis, 2022).
Nascent tRNAs undergo three processing events to become mature tRNAs ready to function in translation: (1) removal of the 3' extension sequences, (2) intron splicing and ligation and (3) aminoacylation. Archease and RtcB are required for the second of these tRNA maturation steps. Unlike mRNA splicing, tRNA splicing is not mediated by the spliceosome, but is instead carried out by complexes containing only protein. Not all tRNAs contain an intron, but throughout the animal kingdom roughly 6-7% of tRNA genes (slightly higher in zebrafish and frogs, and closer to ~25% in budding and fission yeast) contain an intron (Schmidt and Matera, 2020). The tRNA splicing endonuclease (SEN/TSEN) complex initiates the splicing process by cleaving the tRNA intron (Schmidt and Matera, 2020). Plants and fungi differ from metazoans in using a 'healing and sealing' ligation pathway, rather than a direct ligation pathway. The tRNA ligase complex consists of the core tRNA ligase (HSPC117/RtcB) and ASW (also known as C2orf49), CGI-99 (also known as C14orf166 or RTRAF), FAM98B and the DEAD-box helicase DDX1. Archease (ARCH or ZBTB8OS) is a co-factor for RtcB and is required for full activity of the tRNA ligase; however, its interaction with RtcB is transient. Unlike the 'healing and sealing' pathway, the direct ligation pathway produces not only a spliced tRNA, but also a circular intronic RNA, tricRNA, of poorly understood function. In yeast, splicing of tRNAs occurs in the cytoplasm, in association with mitochondria, whereas in eukaryotes pre-tRNA splicing occurs in the nucleoplasm (Francis, 2022).
The tRNA ligase complex and Archease also are required for the nonconventional splicing of a transcription factor in the cytoplasm. Xbp1 is a transcription factor that induces a suite of unfolded protein response (UPR) genes under conditions of endoplasmic reticulum (ER) stress. In the absence of ER stress, Xbp1 mRNA encodes for a nonfunctional protein, Xbp1m. Upon ER stress, the nuclease activity of the Inositol-requiring enzyme 1 (IRE1) stress sensor is activated, and cleaves the Xbp1 mRNA. The mRNA fragments are then ligated by Archease/RtcB, resulting in a change in the reading frame of the transcript, and consequent translation of a functional Xbp1s transcription factor. Nonconventional splicing of the Xbp1 mRNA is thought to be cytoplasmic, as the mRNA is localized to the ER membrane, where IRE1 cleaves it to initiate nonconventional splicing (Francis, 2022).
Taken together, requirements for RtcB/Archease at the cytoplasmic face of the ER, and in the nucleus, implies that the tRNA ligase complex/Archease can shuttle between the nucleus and the cytoplasm. Indeed, the tRNA ligation complex is known to shuttle, and has been found both in the nucleus and in association with cytoplasmic RNA granules. Published data have described a broad cytoplasmic distribution of Archease, at least in cell culture. This study characterized the requirement for Archease in tracheal terminal cell morphogenesis, identify a primarily nuclear localization for Archease, and uncovered regulation of Archease subcellular location by TOR-vATPase pathway activity (Francis, 2022).
In this and previous work, it was determined that most mutations that impair tracheal terminal cell apical membrane growth, and, in particular, mutations in components of the TOR pathway, trigger induction of a compensatory growth program in neighboring wild-type tracheal stalk cells. As part of this compensatory program, stalk cells, which form autocellular tubes, are induced to grow, branch, and extend into the terminal cell. These two cells, which normally share a single intercellular junction, instead share two, three or more intercellular junctions. Moreover, these junctions are no longer located at the initial point of cell-cell contact, but instead the stalk autocellular tubes and their surrounding cytoplasm invade the terminal cell and become enveloped by the cytoplasm of the terminal cell. One fascinating and unresolved question is whether this invasion of the terminal cell requires the generation of a signal from the terminal cell to the stalk cell, or whether the altered mechanical properties of the terminal cell are sufficient to trigger compensatory growth (Francis, 2022).
The relationship between core TOR pathway components, the vATPase and Archease is complex. In prior work, it was determined that vATPase is epistatic to (downstream of) TOR in regulating apical membrane growth. However, the genetic relationships are not quite so straightforward, as vATPase activity also regulates mTOR. There had been no previous indication in the literature that TOR would regulate Archease; however, given the role of TOR in promoting translation, and the role of Archease in tRNA splicing, such regulation has an appealing logic (Francis, 2022).
It was found that mutations in Rheb as well as vATPase components Vha26 and Vha13 had profound effects on Archease subcellular localization. In wild-type tracheal cells, Archease appeared greatly enriched in nuclei - this distribution of Archease in Drosophila was found to be consistent across multiple tissues, including salivary gland epithelia and neurons. This stands in contrast to published reports of Archease localization in HeLa cells, where Archease is reported to be largely cytoplasmic. This discrepancy may reflect differences between post-mitotic and actively cycling cells, cells in culture compared with in vivo, or other factors. Importantly, in vATPase and Rheb mutant cells, there was a dramatic redistribution of Archease from the nucleus to the cytoplasm. The subcellular localization of Archease is likely important, as its two best characterized functions, splicing of tRNA introns and splicing of Xbp1 mRNA, are thought to occur in the nucleus and the cytoplasm, respectively. This would suggest that conditions required for induction of the stress response by Xbp1s might correspond with a general decrease in tRNA availability, and, likewise, that robust mTOR-driven cell growth might limit the ability of the cell to generate the Xbp1s isoform. A less-well-characterized role of the tRNA ligase complex and Archease is in the production of tricRNAs - circularized tRNA introns that are stable and abundant, but of unknown function. A role for tricRNAs in compensatory growth cannot be ruled out (Francis, 2022).
In addition to implications for cellular growth and the stress response, regulation of Archease subcellular localization may also be of interest from the perspective of neuronal regeneration. Genetic disruption of the XBP1 arm of the UPR impedes axonal regeneration. This would suggest that cytoplasmic localization of Archease, where it would be required for productive splicing of Xbp1 mRNA, might contribute to regeneration. However, depletion of the mTOR pathway inhibitors PTEN and TSC1 (Tuberous Sclerosis 1/Hamartin) significantly enhances axonal regeneration, and would be predicted to shift Archease to the nucleus. It may be that under these conditions sufficient Archease is present in the cytoplasm to promote Xbp1 splicing, or, alternatively, it may be that regeneration could be further enhanced by upregulation of Archease expression together with activation of TOR signaling (Francis, 2022).
Polyploidy is an extended phenomenon in biology. However, its physiological significance and whether it defines specific cell behaviors is not well understood. Polyploidy connection to macroautophagy/autophagy was examined, using the larval respiratory system of Drosophila as a model. This system comprises cells with the same function yet with notably different ploidy status, namely diploid progenitors and their polyploid larval counterparts, the latter destined to die during metamorphosis. An association was identified between polyploidy and autophagy and higher endoreplication status was found to correlate with elevated autophagy. Finally, it is reported that tissue histolysis in the trachea during Drosophila metamorphosis is mediated by autophagy, which triggers the apoptosis of polyploid cells (Pino-Jimenez, 2023).
Knowing the molecular makeup of an organ system is required for its in-depth understanding. This analyzed the molecular repertoire of the adult tracheal system of the fruit fly Drosophila melanogaster using transcriptome studies to advance knowledge of the adult insect tracheal system. Comparing this to the larval tracheal system revealed several major differences that likely influence organ function. During the transition from larval to adult tracheal system, a shift in the expression of genes responsible for the formation of cuticular structure occurs. This change in transcript composition manifests in the physical properties of cuticular structures of the adult trachea. Enhanced tonic activation of the immune system is observed in the adult trachea, which encompasses the increased expression of antimicrobial peptides. In addition, modulatory processes are conspicuous, in this case mainly by the increased expression of G protein-coupled receptors in the adult trachea. Finally, all components of a peripheral circadian clock are present in the adult tracheal system, which is not the case in the larval tracheal system. Comparative analysis of driver lines targeting the adult tracheal system revealed that even the canonical tracheal driver line breathless (btl)-Gal4 is not able to target all parts of the adult tracheal system. This study have uncovered a specific transcriptome pattern of the adult tracheal system and provide this dataset as a basis for further analyses of the adult insect tracheal system (Bossen, 2023).
Extracellular matrix (ECM) assembly and remodelling is critical during development and organ morphogenesis. Dysregulation of ECM is implicated in many pathogenic conditions, including cancer. The type II transmembrane serine protease matriptase and the serine protease prostasin are key factors in a proteolytic cascade that regulates epithelial ECM differentiation during development in vertebrates. This study shows by rescue experiments that the Drosophila proteases Notopleural (Np) and Tracheal-prostasin (Tpr) are functional homologues of matriptase and prostasin, respectively. Np mediates morphogenesis and remodelling of apical ECM during tracheal system development and is essential for maintenance of the transepithelial barrier function. Both Np and Tpr degrade the zona pellucida-domain (ZP-domain) protein Dumpy, a component of the transient tracheal apical ECM. Tpr zymogen and the ZP domain of the ECM protein Piopio are cleaved by Np and matriptase in vitro. These data indicate that the evolutionarily conserved ZP domain, present in many ECM proteins of vertebrates and invertebrates, is a novel target of the conserved matriptase-prostasin proteolytic cascade (Drees, 2019).
This paper reports that the vertebrate matriptase-prostasin proteolytic cascade, which is crucial for extracellular matrix differentiation and tissue homeostasis, is conserved in Drosophila. Np acts as a functional Drosophila homologue of matriptase, and tpr mediates prostasin function in the Drosophila tracheal system. Cleavage targets of these conserved extracellular proteolytic pathways are the ZP domains, present in many extracellular proteins of both vertebrates and invertebrates. The Np-mediated protease cascade controls at least three distinct cellular processes during tracheal development, i.e. morphogenesis of the taenidial folds, degradation of the tracheal Dpy cable in the tracheal lumen, and maintenance of the transepithelial barrier function (Drees, 2019).
In vertebrates, proteolysis by matriptase plays a key role in regulating epithelial differentiation. Ectopic expression of human matriptase in the developing tracheal system of Np mutant embryos rescues all aspects of the Np mutant phenotype, i.e. degradation of the Dpy luminal cable, taenidial folds formation, and gas filling of the tubes. The fact that matriptase can functionally substitute for the lack of Np activity indicates that the two proteins share similar functions. Furthermore, similar to matriptase, Np is differentially localized in tissue and stage-dependent patterns at the apical plasma membrane and the apical extracellular space. These findings suggest processing of Np by ectodomain shedding similar to what has been described for matriptase. However, the processing of human matriptase involves not only ectodomain shedding but also the transient interaction of the stem region with its cognate inhibitor HAI-2. Absence of HAI-2 prevents cell surface translocation of matriptase and causes its accumulation in the Golgi compartment (Lai, 2015). Ectopic HAI-2 expression in combination with matriptase facilitates secretion of matriptase in the Drosophila tracheal system. This finding and the lack of a Drosophila HAI-2 homologue indicate different regulatory mechanisms for the translocation and ectodomain shedding of Np and matriptase. This assumption is also consistent with the lack of conserved regulatory stem regions of Np and matriptase. Therefore, different protein processing mechanisms of the two otherwise functionally equivalent proteins are proposed. However, the apparent diverse regulation of both proteins provides the possibility to establish an in vivo experimental system to analyse aspects of matriptase regulation and processing in Drosophila (Drees, 2019).
The matriptase-prostasin proteolytic cascade is initiated by rapid matriptase autoactivation as shown by an in vitro cell-free system as was observed with Np. Thus, the proteolytic cascade involved in aECM formation and maturation in Drosophila is likely to be initiated by Np autoactivation. Once activated, it acts on Tpr, a direct downstream target of Np in the developing tracheal system. This conclusion is based on the observation that in vitro-purified Np is able to cleave Tpr at the zymogen activation site, implying a direct activation of Tpr zymogen by Np in vivo. Furthermore, the tracheal aECM phenotype of tpr mutant embryos is less pronounced than the Np phenotype, since taenidial folds are wild-type like in tpr mutant embryos, while Np mutant embryos lack taenidial folds. The observation that human matriptase also cleaves Tpr at the same zymogen activation site provides additional support for the functional identity of matriptase and Np (Drees, 2019).
Ectopic tracheal expression of human prostasin, together with human HAI-2 in tpr mutant embryos, rescues the defects of the aECM and the LC phenotype of tpr mutants. Thus, Tpr is a functional homologue of human prostasin in the developing trachea. Also, human HAI-2 is required for prostasin secretion into the tracheal lumen, as has been reported for prostasin in vertebrate tissues [43]. LC defects, as observed in tpr mutants, are often caused by an impaired transepithelial barrier function. However, tpr mutant embryos develop a normal barrier and, thus, it is supposed that the LC defects are likely caused by hampered degradation of luminal material and/or Tpr might affect epithelial sodium channels (ENaCs). ENaCs are located in the apical membrane of tracheal cells and are critical for tracheal gas fillin. In vertebrates, prostasin activates ENaCs by inducing proteolytic cleavage of the gamma subunit. It will be interesting to see whether Tpr plays a similar role for ENaC activation in Drosophila (Drees, 2019).
Vertebrate prostasin is widely expressed in ectodermal tissue and most functional aspects of human matriptase are mediated via prostasin in the various tissues. However, the functional relationship of matriptase and prostasin remains to be clarified since matriptase activation and shedding is prostasin-dependent in specific tissues. In contrast to vertebrate prostasin, Drosophila Tpr expression is confined to the tracheal system and not detectable in other ectodermal tissues. Thus, while vertebrate matriptase and prostasin are co-expressed in most tissues, a different scenario is proposed in Drosophila. Tpr belongs to a small group of seven very similar proteases in Drosophila. The corresponding genes are differentially expressed in specific spatial patterns in various ectodermal tissues.It is proposed that such protease zymogens represent putative Np cleavage targets that mediate Tpr-like functions in the different ectodermal tissues that express Np including salivary glands, hindgut, and epidermis (Drees, 2019).
Drosophila tracheal development is a paradigm for the generation of branched tubular systems. Early steps of tracheal maturation, notably tube formation and tubular network assembly, develop independently of Np, while tracheal aECM formation and the transepithelial barrier function during late embryonic tracheogenesis depend on Np. Main differentiation events of the aECM, such as taenidial folds morphogenesis and degradation of luminal protein matrix, are controlled by Np and Tpr (Drees, 2019).
The taenidial folds of the tracheal aECM mainly consist of chitin running perpendicular to the tracheal tube length along the lumen. Their main function is to provide stiffness combined with concurrent flexibility of the tube. Taenidial folds formation is severely affected in Np mutant embryos. The outmost taenidial structure, the hydrophobic envelope, is not detectable and the chitin strands of taenidial folds are highly disorganized. Chitin-interacting proteins, involved in chitin organization, may represent putative targets of Np activity. The aECM phenotype of tpr mutant embryos suggests that tpr function is more specific and confined to establish a proper adhesion between the apical side of tracheal cells and the overlaying taenidial folds of the aECM (Drees, 2019).
Both Np and Tpr are also involved in the degradation of tracheal luminal Dpy, a large ZP domain-containing protein. Luminal Dpy is part of a chitin-proteinous matrix within the tracheal lumen and is essential for normal tracheal network and tube formation. The luminal matrix is degraded and removed from the tracheal lumen during formation of the tracheal taenidial folds and the subsequent gas filling of the tracheal system. Np appears to be the key factor in Dpy cable degradation since Np-deficient embryos completely lack Dpy degradation. Some degradation, however, is mediated via Tpr proteolysis because tpr mutants display remaining Dpy material in the tracheal lumen during late embryogenesis. Thus, the combination of Np and Tpr accomplish luminal Dpy degradation. Alternatively or in addition, Np activates unknown proteases that mediate complete Dpy degradation prior to the gas filling of the tracheal tubes (Drees, 2019).
The transepithelial barrier function is established by the septate junction (SJ) protein complexes, localized at the apico-lateral membrane of epithelial cells [50]. The lack of bona fide SJ proteins like the Drosophila claudin Mega causes a disruption of the ladder-like SJ ultrastructure and a barrier function defect [32]. In Np mutants, the ladder-like ultrastructure of SJs and the barrier function appear to be properly established, but the barrier function collapses during the end of embryogenesis. Thus, Np is essential for the maintenance of the transepithelial barrier function mediated by SJs. This function of Np is reminiscent of matriptase function in mammals. Tracer injection experiments into the dermis of matriptase-deficient mice indicate impaired epidermal tight junction function in such animals [18]. In intestinal epithelial model cell layers and hypomorphic matriptase mice, the essential tight junction component Claudin-2 is deregulated. This observation suggests that reduced barrier integrity was caused, at least in part, by an impaired claudin-2 protein turnover [51]. Furthermore, matriptase cleaves EpCAM, which in turn decreases EpCAM ability to associate with claudin-7 followed by lysosomal degradation of claudin-7 [52]. Thus, it is speculated that Np may also control the maintenance of the epithelial barrier in Drosophila by regulating the function or turnover of claudins in SJs, the invertebrate analogue of the vertebrate tight junction (Drees, 2019).
The aECM protein Dpy is an in vivo downstream target of both Np and human matriptase proteolytic activity. This observation was puzzling since Dpy is not conserved in vertebrates. However, Dpy contains a conserved region, the ZP domain. The ZP domain defines a conserved family of aECM proteins, originally identified in the zona pellucida coat surrounding the mammalian oocyte. The 260 amino acids long ZP domain is proposed to act as a module promoting polymerization of proteins into threads and matrices essential for the organization of highly specialized apical extracellular structures. In fact, the ZP domain was confirmed as a target of Np and matriptase by data showing that Pio, an aECM protein containing a ZP domain, is cleaved by Np and matriptase. Both cleave the ZP domain of Pio within the short linker region, which separates ZP-N and ZP-C, the two subdomains of the ZP domain. Pio is secreted apically in the tracheal lumen and establishes together with Dpy, possibly via ZP-domain polymerization, a structural matrix in the tracheal lumen that is essential for the formation of an interconnected branched network. Cleavage of ZP domains within a meshwork of Dpy and Pio filaments may facilitate rapid degradation of the luminal extracellular matrix, the prerequisite for normal gas filling of the tracheal system. This conclusion is supported by phenotypic analysis of Np mutant embryos, which exhibit a stable, undegraded luminal Dpy cable and lack tracheal gas filling. ZP-domain proteins also play crucial roles in development of embryonic epidermal cuticle, an aECM that protects the animal against the external milieu. Eight ZP-domain proteins are required for the localized reorganization of epidermal cells and to sculpture the actin-rich apical extensions, the denticles. The observation that Np mutants exhibit reduced and rudimentary denticles in the epidermis underlines the possibility that the epidermal ZP-proteins are also targets of Np protease activity (Drees, 2019).
The results showing that human matriptase cleaves the ZP domain of Pio open future directions to explore novel targets of the matriptase-prostasin catalytic pathway. In vertebrates, ZP-domain proteins are involved in remodelling apical extracellular structures, such as ZP1-ZP3, important in the mammalian ovary for fertilization and uromodulin, which is released into the tubular kidney lumen where it polymerizes in a gel-like matrix that controls salt transport and urine concentration. Also, mutations in genes encoding ZP-domain proteins cause human diseases such as deafness, triggered by mutations in alpha- and beta-tectorin. The tectorins are components of the tectorial membrane, an aECM necessary for sound transmission to neural cells in the cochlea. These examples already demonstrate the importance of ZP-domain proteins for mammalian physiology. Based on the results reported in this study, it is proposed that ZP-domain cleavage by the matriptase-prostasin proteolytic cascade may represent a conserved process to control ZP-domain protein functions, which are crucial for apical matrix remodelling during development, wound repair, and differentiation (Drees, 2019).
Membrane expansion integrates multiple forces to mediate precise tube growth and network formation. Defects lead to deformations, as found in diseases such as polycystic kidney diseases, aortic aneurysms, stenosis, and tortuosity.
This study identified a mechanism of sensing and responding to the membrane-driven expansion of tracheal tubes. The apical membrane is anchored to the apical extracellular matrix (aECM) and causes expansion forces that elongate the tracheal tubes. The aECM provides a mechanical tension that balances the resulting expansion forces, with Dumpy being an elastic molecule that modulates the mechanical stress on the matrix during tracheal tube expansion. The zona pellucida (ZP) domain protein Piopio was shown to interact and cooperates with the ZP protein Dumpy at tracheal cells. To resist shear stresses which arise during tube expansion, Piopio undergoes ectodomain shedding by the Matriptase homolog Notopleural, which releases Piopio-Dumpy-mediated linkages between membranes and extracellular matrix. Failure of this process leads to deformations of the apical membrane, tears the apical matrix, and impairs tubular network function. Conserved ectodomain shedding was shown of the human TGFβ type III receptor by Notopleural and the human Matriptase, providing novel findings for in-depth analysis of diseases caused by cell and tube shape changes (Drees, 2023).
Tracheal tube lumen expansion requires mechanical stress regulation at apical cell membranes and attached aECM. This involves the proteolytic processing of proteins that set local membrane-matrix linkages. Thus, the membrane microenvironment exhibits critical roles in regulating tube and network functionality (Drees, 2023).
ZP domain proteins organize protective aECM in the kidney, tectorial inner ear, and ZP, as well as in Drosophila epidermis, tendon cells, and appendages. Drosophila ZP domain proteins link the aECM to actin and polarity complexes in epithelial cells. Dumpy establishes force-resistant filaments for anchoring tendon cells to the pupal cuticle (Drees, 2023).
This study found that ZP protein-mediated microenvironmental changes increase the flexibility of membrane-matrix association, resulting from the activity of ZP domain proteins. Shear stress stimulates the activity of membrane-anchored proteases and potentially also apical transmembrane protease Np since the misdistribution of the tracheal cytoskeleton was not observed when blisters arise. The dynamic membrane-matrix association control is based on findings that loss of Np prevents Pio ectodomain shedding at the apical cell membrane resulting in immobile localization of Pio at the membrane and Dpy localization within the matrix. Direct interaction and overlapping subcellular localization at the cell surface showed that both proteins form a ZP matrix that potentially attaches membrane and ECM. Deregulation of Pio shedding blocks ZP matrix rearrangement and release of membrane-matrix linkages under tube expansion and subsequent shear stress. This destabilizes the microenvironment of membranes, causing blister formation at the membrane due to ongoing membrane expansion. Additionally, Pio could be part of a force-sensing signal transduction system destabilizing the membrane and matrix. The observation that the membrane deformations are maintained in Np mutant embryos supports the postulated Np function to redistribute and deregulate membrane-matrix associations in stage 16 embryos when tracheal tube length expands. In contrast, Np overexpression potentially uncouples the Pio-Dpy ZP matrix membrane linkages resulting very likely in unbalanced forces causing sinusoidal tubes (Drees, 2023).
The membrane defects observed in both Pio and Np mutants indicate errors in the coupling of the membrane matrix due to the involvement of Pio. In pio mutants, gaps appear between the deformed membrane and the apical matrix. These changes in apical cell membrane shape are consistent with increased cell and tube elongation in pio mutant embryos because the matrix is uncoupled from the membrane in such mutants. In contrast to pio mutants, the large membrane bulges in Np mutants affect the membrane and the apical matrix. Since apical Pio is not cleaved in Np mutants, the matrix is not uncoupled from the membrane as in pio mutant embryos but is likely more intensely coupled, which leads to tearing of the matrix axially along the membrane bulges, when the tube expands in length. If apical Pio detachment reduces coupling between the matrix and apical membrane, then it is likely that Np mutant embryos may exhibit a reduced tube length phenotype. In Np mutant embryos, average tracheal dorsal trunk length tends to be reduced compared to wt embryos, suggesting that Pio shedding is critical in controlling tracheal tube lumen length (Drees, 2023).
Is Pio ectodomain shedding in response to tension? Tension was not measured directly. However, the developmental profile of mechanical tension during tracheal tube length elongation in stage 16 embryos is consistent with the profile of Pio shedding. Np cleaves apical Pio during stage 16 when tube length expands. In contrast, Pio shedding decreases sharply at early stage 17 when tube elongation is completed. The model, therefore, predicts that loss of Pio or increased Pio secretion at stage 16 may reduce the coupling of the membrane matrix so that increased tracheal tube elongation is maintained until the end of stage 16, which is found in pio mutants and upon Np overexpression. Unknown proteases may likely be involved in Pio processing since cleaved mCherry::Pio is also detectable in inactive NpS990A cells. Previously a mutation at the Pio ZP domain (R196A) resistant to NP cleavage in cell culture experiments (Drees, 2019). Establishing a corresponding mutant fly line would be essential in determining whether the observed phenotype resembles the phenotype of the Np mutant embryos. In addition, unknown mechanisms, such as distinct membrane connections during development and emerging links to the developing cuticle, may also influence tension at the apical membrane during tube length control (Drees, 2023).
Indeed, the anti-Pio antibody, which detects all different Pio variants, showed a punctuate Pio pattern overlapping with the apical cell membrane markers Crb and Uninflatable (Uif) at the dorsal trunk cells of stage 16 embryos. Additionally, Pio antibody also revealed early tracheal expression from embryonic stage 11 onward, and due to Pio function in narrow dorsal and ventral branches, strong luminal Pio antibody staining is detectable from early stage 14 until stage 17, when airway protein clearance removes luminal contents. In the pio5m and pio17c mutants, Pio stainings were strongly reduced although some puncta were still detectable in the trachea. Similarly, Pio antibody staining is intracellular in the trachea of stage 11 pio2R-16 point mutation embryos. Interestingly, also dpy mutants showed strongly reduced and intracellular Pio antibody staining (Drees, 2023).
mCherry::Pio was generated as a tool for in vivo Pio expression and localization pattern analysis during tube lumen length expansion. The mCherry::Pio resembled the Pio antibody expression pattern from early tracheal development onward. However, luminal mCherry::Pio enrichment occurs specifically during stage 16, when tubes expand. The stage 16 embryos showed mCherry::Pio puncta accumulating apically in dorsal trunk cells. Moreover, mCherry::Pio puncta partially overlapped with Dpy::YFP and chitin at the taenidial folds, forming at apical cell membranes. Supported by several observations, such as antibody staining, video monitoring, FRAP experiments, and western blot studies, these findings indicate that Pio may play a significant role at the apical cell membrane and matrix in dorsal trunk cells of stage 16 embryo (Drees, 2023).
Furthermore, it was shown that Np mediates Pio ZP domain cleavage for luminal release of the short Pio variant during ongoing tube length expansion. The luminal cleaved mCherry::Pio is enriched at the end of stage 16 and finally internalized by the subsequent airway clearance process during stage 17 after tube length expansion. Such rapid luminal Pio internalization is consistent with a sharp pulse of endocytosis rapidly internalizing the luminal contents during stage 17. Wurst is required to mediate the internalization of proteins in the airways. Consistencly, during stage 17, luminal Pio antibody staining fades in control embryos but not in Wurst deficient embryos (Drees, 2023).
Nevertheless, Pio and its endocytosis depend on its interaction with the chitin matrix and the Np-mediated cleavage. In stage 16 wurst and mega mutant embryos, Pio antibody staining at the chitin cable, suggesting that Pio is cleaved and released into the dorsal trunk tube lumen. Also, the Cht2 overexpression did not prevent the luminal release of Pio. However, reduced wurst, mega function, and Cht2 overexpression caused an enrichment of punctuate Pio staining at the apical cell membrane and matrix . Although the three proteins are involved in different subcellular requirements, they all contribute to the determination of tube size by affecting either the apical cell membrane or the formation of a well-structured apical extracellular chitin matrix, indicating that changes at the apical cell membrane and matrix in stage 16 embryos affect the Pio pattern at the membrane. It also shows that local Pio linkages at the cell membrane and matrix are still cleaved by the Np function for luminal Pio release, which explains why those mutant embryos do not show pio mutant-like membrane deformations and Np-mutant-like bulges. This is in line with observations that tracheal Pio overexpression cannot cause tube size defects as the Np function is sufficient to organize local Pio linkages at the membrane and matrix. Therefore, it is unlikely that tracheal tube length defects in wurst and mega mutants as well as in Cht2 misexpression embryos are caused by the apical Pio density enrichment (Drees, 2023).
Nevertheless, oversized tube length due to the misregulation of the apical cell membrane and adjacent chitin matrix may cause changes to local Pio set linkages and the need for Np-mediated cleavage. Strikingly, a lack of Pio release was observed in Np mutants. This shows that Pio density at the membrane versus lumen depends predominantly on Np function. The molecular mechanisms that coordinate the Np-mediated Pio cleavage are unknown and will be necessary for understanding how tubes resist forces that impact cell membranes and matrices. On the other hand, Pio is required for the extracellular secretion of its interaction partner Dpy. At the same time, Dpy is needed for Pio localization at the cell membrane and its distribution into the tube lumen. Consistently, in vivo, mCherry::Pio, and Dpy::eYFP localization patterns overlap at the apical cell surface and within the tube lumen. These observations support the model that Pio and Dpy interact at the cell surface where Np mediates Pio cleavage to support luminal Pio release by the large and stretchable matrix protein Dpy (Drees, 2023).
Taenidial organization prevents the collapse of the tracheal tube. Therefore, cortical (apical) actin organizes into parallel-running bundles that proceed to the onset of cuticle secretion and correspond precisely to the cuticle’s taenidial folds. Mutant larvae of the F-actin nucleator formin DAAM show mosaic taenidial fold patterns, indicating a failure of alignment with each other and along the tracheal tubes. In contrast, pio mutant dorsal tracheal trunks contained increased ring spacing. Fusion cells are narrow doughnut-shaped cells where actin accumulates into a spotted pattern. Formins, such as Diaphanous, are essential in organizing the actin cytoskeleton. However, dorsal trunk tube fusion defects were not found as found in the presence of the activated Diaphanous (Drees, 2023).
On the other hand, ectopic expression of DAAM in fusion cells induces changes in apical actin organization but does not cause any phenotypic effects. DAAM is associated with the tyrosine kinase Src42A, which orients membrane growth in the axial tube dimension. Src42 overexpression elongates tracheal tubes due to flattened axially elongated dorsal trunk cells and AJ remodeling. Although flattened cells and tube overexpansion are similar in pio mutant embryos, mislocalization of AJ components was not observed, as found upon constitutive Src42 activation. Instead, an unusual stretched appearance of AJs was detected at the fusion cells of pio mutant dorsal trunks, which has not been observed before and may play a role in regulating axial taenidial fold spacing and tube elongation (Drees, 2023).
Self-organizing physical principles govern the regular spacing pattern of the tracheal taenidial folds. The actomyosin cortex and increased actin activity before and turnover at stage 16 drive the regular pattern formation. However, the cell cortex and actomyosin are in frictional contact with a rigid apical ECM. The Src42A mutant embryos contain shortened tube length but increased taenidial fold period pattern due to decreased friction. In contrast, the chitinase synthase mutant kkv1 has tube dilation defects and no regular but an aberrant pearling pattern caused by zero fiction (Drees, 2023).
In contrast, pio mutant embryos do not contain tube dilation defects or shortened tubes but increased tube length. Furthermore, chitin-binding probe (cbp) and antibody stainings reveal the presence of a luminal chitin cable and a solid aECM structure in pio mutant stage 16 embryos. In addition, apical actin enrichment in tracheal cells of pio mutant embryos appeared wt-like. Nonetheless, pio mutant embryos show an increased taenidial fold period compared with wt, indicating a decreased friction. Thus, it is proposed that the lack of Pio reduces friction. Reasons might be subtle defects of actomyosin constriction or chitin matrix, which was not detected in the pio mutant tracheal cells. Further reasons for lower friction might also be the loss of Pio set local linkages between apical cortex and aECM in stage 16 embryos, which are modified by Np, as proposed in in a model (see Model of apical Pio and Dpy matrix at the apical cell surface and Pio proteolysis and release. (Drees, 2023).
Heterozygous and homozygous pio mutant embryos generally do not show tubal collapse. However, the loss of Pio and accompanying lack of Dpy secretion in stage 17 pio mutant embryos led to the loss of a Pio/Dpy matrix, impacting the late embryonic maturation and differentiation of a normal chitin matrix at the apical cell surface. TEM images reveal reduced dense chitin matrix material at taenidial folds and misarranged taenidial fold pattern, suggesting impaired taenidial function prevents tube lumen from collapsing after tube protein clearance. wurst knockdown and mutant embryos do not show general tube collapse, but luminal chitin fiber organization is disturbed in stage 17 embryos. Therefore, transheterozygous wurst;pio mutant embryos may combine both defects and suffer from maturation deficits of the chitin/ZP matrix at the apical cell surface and within the tube lumen, which finally causes a high number of embryos with incomplete gas-filling due to tube collapse. These maturation deficits are even more dramatic in the wurst;pio double mutants, which show no gas-filling (Drees, 2023).
These studies on human Matriptase provide evidence for a mechanistic conservation of ZP domain protein as a substrate for ectodomain shedding. The upregulation of Matriptase activity and increased TGFβ receptor density affect human and mouse model idiopathic pulmonary fibrosis cells on pulmonary fibrogenesis. Furthermore, the human Matriptase induces the release of proinflammatory cytokines in endothelial cells, which contribute to atherosclerosis and probably also to abdominal aortic aneurysms. The membrane bulges arising in the Drosophila model during tracheal tube elongation upon Np loss of function showed analogy to the appearance of artery aneurysms. Bulges with varying phenotypic expression in different organs can lead to aortic rupture due to fragile artery walls or degeneration of layers in responses to stimuli, such as shear stresses. Indeed, aneurysm development is forced by alterations in the ECM and is characterized by extensive ECM fragmentation caused by shedding of membrane-bound proteins (Drees, 2023).
This study identified a dynamic control of matrix proteolysis, very likely enabling fast and site-specific uncoupling of membrane-matrix linkages when tubes expand. Such a scenario has not yet been studied in angiogenesis. It may represent a new starting point for genetic studies to decipher the putative roles of ZP domain proteins and Matriptase in clinically relevant syndromes, including the formation of aneurysms caused by membrane deformation and defects in size determination of airways and vessels (Drees, 2023).
The evolutionary origins of complex morphological structures such as the vertebrate eye or insect wing remain one of the greatest mysteries of biology. Recent comparative studies of gene expression imply that new structures are not built from scratch, but rather form by co-opting preexisting gene networks. A key prediction of this model is that upstream factors within the network will activate their preexisting targets (i.e., enhancers) to form novel anatomies. This study shows how a recently derived morphological novelty present in the genitalia of D. melanogaster employs an ancestral Hox-regulated network deployed in the embryo to generate the larval posterior spiracle. This study demonstratea how transcriptional enhancers and constituent transcription factor binding sites are used in both ancestral and novel contexts. These results illustrate network co-option at the level of individual connections between regulatory genes and highlight how morphological novelty may originate through the co-option of networks controlling seemingly unrelated structures (Glassford, 2015).
This study has shown how a gene regulatory network (GRN) underlying a novel structure, the posterior lobe, is composed of components that are active in the embryonic posterior spiracle, an ancestral Hox-regulated structure that was present at the inception of this novelty (see Model Depicting the Co-option of Genes, Enhancers, and Transcription Factor Binding Sites during the Origination of the Novel Posterior Lobe). These findings confirm previous speculation that network co-option proceeds through the re-use of individual transcription factor binding sites within enhancer sequences. Further, the data help calibrate expectations concerning the degree of physical similarity between novel and ancestral structures during co-option events. Below, it is discussed how the architecture of the posterior spiracle network may have predisposed it for co-option in the genitalia, and the general implications of these findings are explored with regard to the origins of morphological novelty (Glassford, 2015).
While these results illustrate the downstream consequences of co-option, the upstream causative events await characterization. It is suspected that some number of high-level regulators of the posterior spiracle network recently evolved novel genital expression patterns through alterations within their regulatory regions. Currently, Unpaired represents the best candidate upstream factor, as it is positioned near the top of the spiracle network, differs in expression greatly between lobed and non-lobed species (unlike Spalt and Abd-B), and is the only high-level factor in the spiracle network for which a shared lobe and spiracle enhancer has yet to be identified. Indeed, a reporter screen of the 30 kb of regulatory DNA immediately surrounding the upd gene identified a posterior spiracle enhancer that is not deployed in the posterior lobe, marking an important point of divergence separating the posterior spiracle and posterior lobe networks. However, the identification of enhancers controlling upd expression in the posterior lobe will be required to resolve its role in this structure’s origination (Glassford, 2015).
The architecture of the posterior spiracle network may have shaped the possible developmental contexts in which it could be co-opted. The Hox factor Abd-B has a deeply conserved role in the insect abdomen and genitalia. The top-level factors of the posterior spiracle network depend upon Abd-B for activation in the embryo. This regulation by Abd-B extends to lower tiers of the network, such as Poxn. The tight integration of Abd-B with multiple tiers of the posterior spiracle network may have limited this network’s re-deployment to posterior body segments that express Abd-B. Indeed, several components of this network (Poxn, ems, upd) are activated early during genital development in the presumptive cleavage furrow separating the lateral plate from the clasper. This may represent the aftereffect of multiple waves of re-deployment in Abd-B expressing tissues. Examination of additional examples of network co-option at the level of constituent regulatory sequences could reveal general rules that govern and bias network redeployment (Glassford, 2015).
Historically, the identification of co-option events has relied upon comparative analyses of gene co-expression. The first examples of co-option were diagnosed by finding novel gene expression patterns near zones of ancestral function, such as the deployment of the posterior wing patterning circuit within novel butterfly eyespots. Subsequently, many examples of co-option have involved educated guesses of the types of networks that contribute to the novelty, such as the role of the appendage specification network within beetle horns or the sharing of the biomineralization network between adult and larval skeletons of sea urchins. The data suggest that tracing the evolutionary origins of individual enhancers provides a less biased path for connecting novelties to their ancestral beginnings, as any of the seven enhancers that this study has characterized in the posterior lobe would have led to the spiracle network. Further, this approach is likely to illuminate the underlying cellular mechanisms by which the co-option of a network is translated into a novel developmental outcome (Glassford, 2015).
Rather than generating a serial homolog of the posterior spiracle, the co-option event forming the posterior lobe resulted in an epithelial outgrowth, likely owing to the deployment of only a portion of the spiracle network in the genitalia. This is reflected by the absence of the Cut transcription factor and downstream genes that control the spiracular chamber’s development. Of the 10 genes this study has identified in both networks, 9 are active in the stigmatophore, the outer sheath of the posterior spiracle that protrudes from the body through a process that involves convergent extension. Collectively, these findings imply that similar morphogenic processes are activated by this shared network in the novel setting of the posterior lobe. It is proposed that the inspection of enhancers underlying other novel three-dimensional structures may reveal similar networks that have been used over and over again to generate "unique styles of architecture" within developing tissues (Glassford, 2015).
Insect respiration has long been thought to be solely dependent on an elaborate tracheal system without assistance from the circulatory system or immune cells. This study describes that Drosophila crystal cells-myeloid-like immune cells called haemocytes-control respiration by oxygenating Prophenoloxidase 2 (PPO2) proteins. Crystal cells direct the movement of haemocytes between the trachea of the larval body wall and the circulation to collect oxygen. Aided by copper and a neutral pH, oxygen is trapped in the crystalline structures of PPO2 in crystal cells. Conversely, PPO2 crystals can be dissolved when carbonic anhydrase lowers the intracellular pH and then reassembled into crystals in cellulo by adhering to the trachea. Physiologically, larvae lacking crystal cells or PPO2, or those expressing a copper-binding mutant of PPO2, display hypoxic responses under normoxic conditions and are susceptible to hypoxia. These hypoxic phenotypes can be rescued by hyperoxia, expression of arthropod haemocyanin or prevention of larval burrowing activity to expose their respiratory organs. Thus, it is proposed that insect immune cells collaborate with the tracheal system to reserve and transport oxygen through the phase transition of PPO2 crystals, facilitating internal oxygen homeostasis in a process that is comparable to vertebrate respiration (Shin, 2024).
This study presents evidence for the critical role of Drosophila haemocytes, specifically crystal cells, in insect respiration through the phase transition of PPO2. Crystal cells orchestrate the dynamic shuttling of haemocytes between the haematopoietic pocket and the circulation. This movement is essential for generating in cellulo crystals of PPO2, which is primarily contained within crystal cells and undergoes phase transitions according to oxygen levels, intracellular pH and copper concentrations. As a result, the ambient oxygen level is a key determinant of the status of PPO2 crystals and substantially influences the duration of crystal cell adherence to the haematopoietic pocket. The phase transition of PPO2 enables it to either be oxygenated and crystallize, serving as a potential oxygen reservoir, or deoxygenate and dissolve to support the respiratory process. Notably, this process mainly targets the fat body-a critical regulator of animal growth and survival that is highly dependent on oxygen availability with insufficient tracheal innervation. Consequently, larvae lacking crystal cells or PPO2 or expressing a mutant form of PPO2 incapable of binding to copper ions display hypoxic responses under normoxic conditions and become intolerant to hypoxia. Notably, these results demonstrate that the expression of haemocyanin II (Hc2;) from Limulus polyphemus in crystal cells effectively restores the respiratory function of PPO2, underscoring the evolutionary conservation and significance of this mechanism in respiration (Shin, 2024).
The presence of membraneless organelles, mainly characterized by liquid-liquid phase separation, has become increasingly recognized in various cellular processes. Building on these discoveries, the current study reveals the reversible phase transition of PPO2 within crystal cells as a critical regulator of animal respiration. Under normoxic conditions, crystal cell relocation and PPO2 phase transition probably occur while maintaining dynamic equilibrium. This process facilitates the respiration of tissues that lack tracheal innervation or exhibit high oxygen demand, including but not limited to the fat body. While cytosolic PPO1 is known for its role in the main phenoloxidase activities, and crystallized PPO2 for its role in the sequestration of enzymatic phenoloxidase functions, this study indicates an additional function of PPO2 protein in reversible oxygen collection. Notably, these findings demonstrate that haemocyanin from the horseshoe crab co-constitutes crystals with PPO2. Moreover, haemoglobin solutions have been observed to undergo phase separation under physiological pH, ionic strength and haemoglobin concentrations. These observations suggest that respiratory protein condensates may function as a universally conserved respiratory hub for tunable gas exchange. Although the current findings provide specific evidence for the presence of PPO2 protein in in cellulo crystals, additional factors or phase-transition steps, ranging from liquid-to-gel or gel-to-solid states, may be involved in their formation. Moreover, the mechanisms underlying the acquisition of metastability in the solid-like state within a short time in vivo remain unclear. Future biochemical and biophysical studies of PPO2 crystals, complementing genetic and cellular analyses, will shed light on the mechanisms of the PPO2 phase separation and the kinetics of crystal assembly in vivo (Shin, 2024).
This study highlights that larval habitat, whether plastic vials in laboratory conditions or rotten fruits in the wild, inherently exposes Drosophila larvae to hypoxia. This environmental challenge forces the larvae to find a balance between fitness and immunity. While digging deeper provides better protection from predators, it potentially exposes them to hypoxia. Among the two major immune cell types in Drosophila, crystal cells have been recognized for their distinctive function in wound healing and melanization. However, the current study suggests that crystal cells offer a unique dual-protection mechanism through both innate immune responses and respiratory support, providing insights into the evolutionary origins of respiratory immune cells in animals. From a haematopoiesis perspective, crystal cells exhibit convergent functions resembling both platelets and erythrocytes, which are functionally analogous to megakaryocyte-erythrocyte lineage myeloid cells. Furthermore, in other insects, haemocytes are often observed surrounding tracheal branches, and other insects possess crystal cell equivalents known as oenocytoids. These observations suggest that the association between haemocytes and the tracheal system for respiration and oxygen responsiveness may be conserved across insects and not limited to Drosophila (Shin, 2024).
Tracheal tube maturation starts with an apical secretion pulse that deposits extracellular matrix components to form a chitin-based apical luminal matrix (aECM). This aECM is then cleared and followed by the maturation of taenidial folds. Finally, air fills the tubes. Meanwhile, the cellular junctions are maintained to ensure tube integrity. The Osiris (Osi) gene family is located at the Triplo-lethal (Tpl) locus on chromosome 3R 83D4-E3 and exhibits dosage sensitivity. This study shows that three Osi genes (Osi9, Osi15, Osi19), function redundantly to regulate adherens junction (AJ) maintenance, luminal clearance, taenidial fold formation, tube morphology, and air filling during tube maturation. The localization of Osi proteins in endosomes (Rab7-containing late endosomes, Rab11-containing recycling endosomes, Lamp-containing lysosomes) and the reduction of these endosomes in Osi mutants suggest the possible role of Osi genes in tube maturation through endosome-mediated trafficking. Tube maturation was examined in zygotic rab11 and rab7 mutants, respectively, to determine whether endosome-mediated trafficking is required. Interestingly, similar tube maturation defects were observed in rab11 but not in rab7 mutants, suggesting the involvement of Rab11-mediated trafficking, but not Rab7-mediated trafficking, in this process. To investigate whether Osi genes regulate tube maturation primarily through the maintenance of Rab11-containing endosomes, rab11 was overexpressed in Osi mutant trachea. Surprisingly, no obvious rescue was observed. Thus, increasing endosome numbers is not sufficient to rescue tube maturation defects in Osi mutants. These results suggest that Osi genes regulate other aspects of endosome-mediated trafficking, or regulate an unknown mechanism that converges or acts in parallel with Rab11-mediated trafficking during tube maturation (Scholl, 2023).
Larval tracheae of Drosophila harbour progenitors of the adult tracheal system (tracheoblasts). Thoracic tracheoblasts are arrested in the G2 phase of the cell cycle in an ATR (mei-41)-Checkpoint Kinase1 (grapes, Chk1) dependent manner prior to mitotic re-entry. This study investigated developmental regulation of Chk1 activation. This study reports Wnt signaling is high in tracheoblasts and this is necessary for high levels of activated (phosphorylated) Chk1. Canonical Wnt signaling facilitates this by transcriptional upregulation of Chk1 expression in cells that have ATR kinase activity. Wnt signaling is dependent on four Wnts (Wg, Wnt5, 6,10) that are expressed at high levels in arrested tracheoblasts and are downregulated at mitotic re-entry. Interestingly, none of the Wnts are dispensable and act synergistically to induce Chk1. Finally, this study shows that downregulation of Wnt signaling and Chk1 expression leads to mitotic re-entry and the concomitant upregulation of Dpp signaling, driving tracheoblast proliferation (Kizhedathu, 2020).
Cell growth is well defined for late (postembryonic) stages of development, but evidence for early (embryonic) cell growth during postmitotic morphogenesis is limited. This study reports early cell growth as a key characteristic of tubulogenesis in the Drosophila embryonic salivary gland (SG) and trachea. A BTB/POZ domain nuclear factor, Ribbon (Rib), mediates this early cell growth. Rib binds the transcription start site of nearly every SG-expressed ribosomal protein gene (RPG) and is required for full expression of all RPGs tested. Rib binding to RPG promoters in vitro is weak and not sequence specific, suggesting that specificity is achieved through cofactor interactions. Accordingly, this study demonstrates Rib's ability to physically interact with each of the three known regulators of RPG transcription. Surprisingly, Rib-dependent early cell growth in another tubular organ, the embryonic trachea, is not mediated by direct RPG transcription. These findings support a model of early cell growth customized by transcriptional regulatory networks to coordinate organ form and function (Loganathan, 2022).
How signaling proteins generate a multitude of information to organize tissue patterns is critical to understanding morphogenesis. In Drosophila, FGF produced in wing-disc cells regulates the development of the disc-associated air-sac-primordium (ASP). This study shows that FGF is Glycosylphosphatidylinositol-anchored to the producing cell surface and that this modification both inhibits free FGF secretion and promotes target-specific cytoneme contacts and contact-dependent FGF release. FGF-source and ASP cells extend cytonemes that present FGF and FGFR on their surfaces and reciprocally recognize each other over distance by contacting through cell-adhesion-molecule (CAM)-like FGF-FGFR binding. Contact-mediated FGF-FGFR interactions induce bidirectional responses in ASP and source cells that, in turn, polarize FGF-sending and FGF-receiving cytonemes toward each other to reinforce signaling contacts. Subsequent un-anchoring of FGFR-bound-FGF from the source membrane dissociates cytoneme contacts and delivers FGF target-specifically to ASP cytonemes for paracrine functions. Thus, GPI-anchored FGF organizes both source and recipient cells and self-regulates its cytoneme-mediated tissue-specific dispersion (Du, 2022).
Drosophila trachea is a classical model for analyzing epithelial, especially tubular epithelial biology. This study identified lateral E-cadherin mediated junctions that encircle the cells just basal to the zonula adherens in the larval trachea. The lateral junction is associated with downstream adapters, including catenins, and has a distinct junctional actin cortex. The lateral cortex contributes to the development of a supracellular actomyosin mesh in the late larvae. Establishment of this cytoskeletal structure depends on lateral junction associated Rho1 and Cdc42 GTPases, and Arp and WASP pathways. The supracellular network takes the character of stress fibers along the AP axis in the early hours of pupation. It contributes to the shortening of the epithelial tube albeit in a manner redundant to ECM-mediated compression mechanism. In conclusion, this study shows the in vivo existence of functional lateral adherens junction and suggest a role for it in mediating dynamic cytoskeletal events during tissue scale morphogenesis (Pradhan, 2023).
Germline mutations upregulating RAS signaling are associated with multiple developmental disorders. A hallmark of these conditions is that the same mutation may present vastly different phenotypes in different individuals, even in monozygotic twins. This study demonstrates how the origins of such largely unexplained phenotypic variations may be dissected using highly controlled studies in Drosophila that have been gene edited to carry activating variants of MEK, a core enzyme in the RAS pathway. This allowed measurement of the small but consistent increase in signaling output of such alleles in vivo. The fraction of mutation carriers reaching adulthood was strongly reduced, but most surviving animals had normal RAS-dependent structures. These results were rationalized using a stochastic signaling model and support it by quantifying cell fate specification errors in bilaterally symmetric larval trachea, a RAS-dependent structure that allows isolation the effects of mutations from potential contributions of genetic modifiers and environmental differences. It is proposed that the small increase in signaling output shifts the distribution of phenotypes into a regime, where stochastic variation causes defects in some individuals, but not in others. These findings shed light on phenotypic heterogeneity of developmental disorders caused by deregulated RAS signaling and offer a framework for investigating causal effects of other pathogenic alleles and mild mutations in general (Marmion, 2023).
The re-use of genes in new organs forms the base of many evolutionary novelties. A well-characterised case is the recruitment of the posterior spiracle. gene network to the Drosophila male genitalia. This study found that this network has also been co-opted to the testis mesoderm where is required for sperm liberation, providing an example of sequentially repeated developmental co-options. Associated to this co-option event, an evolutionary expression novelty appeared, the activation of the posterior segment determinant Engrailed to the anterior A8 segment controlled by common testis and spiracle regulatory elements. Enhancer deletion shows that A8 anterior Engrailed activation is not required for spiracle development but only necessary in the testis. This study presents an example of pre-adaptive developmental novelty: the activation of the Engrailed transcription factor in the anterior compartment of the A8 segment where, despite having no specific function, opens the possibility of this developmental factor acquiring one. It is proposed that recently co-opted networks become interlocked, so that any change to the network because of its function in one organ, will be mirrored by other organs even if it provides no selective advantage to them (Molina-Gil, 2023).
It has been observed that genes playing particular roles during organ development can be recruited to perform novel functions in other organs. The re-use, or co-option, of developmental genes in new organs, is the base of many evolutionary novelties. An example of this is the highly transparent crystallin proteins that refract light in the eye lens. In all vertebrates, α-crystallin evolved from the co-option of a small heat shock protein to the eye; while in birds δ-crystallin evolved from the co-option of a different protein, the Arginosuccinase lyase involved in arginine biosynthesis (Molina-Gil, 2023).
Although there is abundant research on single-gene co-option, few studies have considered the functional consequences of full-gene network co-option. One of the best-characterised co-option cases is the recruitment of the appendage-forming gene network to form the eye-spots that decorate butterfly wings in several species. Similarly, in the Drosophila melanogaster subgroup, the larval respiratory posterior spiracles and the adult male genitalia share the expression of numerous genes due to the recent co-option into the male genital disc primordium of a pre-existing gene network controlling the formation of the external larval respiratory organs. The co-option of this gene network to the male genitalia resulted in the formation of the posterior lobe, a structure present in D. simulans and D. mauritiana, closely related to Drosophila melanogaster, but not in the more distant D. biarmipes or D. ananassae species (Molina-Gil, 2023).
The formation of the posterior spiracles and the posterior lobes have been well-studied in D. melanogaster. Posterior spiracle organogenesis is regulated by a gene network activated in the eighth abdominal larval segment (A8) by the Hox protein Abdominal-B (Abd-B). The internal spiracular chamber is formed from A8 anterior compartment cells (A8a) when Abd-B activates in the dorsal ectoderm the transcription of the JAK/STAT signalling pathway ligand Unpaired (Upd) as well as the Empty spiracles (Ems) and the Cut (Ct) transcription factors. The external protruding stigmatophore is formed from both anterior and posterior compartment A8 cells when Abd-B activates the Spalt (Sal) transcription factor, which in turn activates engrailed (en) transcription in a unique A8 pattern. These primary factors activate the RhoGAP Cv-c and RhoGEF64C cytoskeletal regulators, the cell polarity gene crumbs (crb) and various Cadherins6 Abd-B also modulates the expression of wingless and the EGF regulator rhomboid genes generating A8 specific segmental information distinct from that in more anterior segments (Molina-Gil, 2023).
The posterior lobe is a hook-shaped structure used by the male to grasp the female during mating, which may act as a prezygotic reproductive isolation barrier facilitating speciation. In D. melanogaster, ten genes of the spiracle gene network are required for the formation of the posterior lobe, with their activation in at least seven cases being regulated in both structures by the same cis-regulatory elements (CRE). The study of two of these enhancers, revealed that the same DNA-binding sites activate the CRE's expression in both organs, making this one of the best-characterised cases of whole gene network co-option (Molina-Gil, 2023).
The recruitment of a gene network to a new organ exposes it to different selective pressures that may accelerate the appearance of novelties. One such novelty is the expression of the posterior segment determinant engrailed in the anterior compartment cells of the A8 segment. Engrailed is crucial during segmentation, and its expression in the anterior compartment is surprising given that En has been localised to the posterior cells all along arthropod evolution (Molina-Gil, 2023).
Drosophila segmentation results from the activation of the segment-polarity genes engrailed (en), hedgehog (hh) and wingless (wg) in periodic stripes of cells along the antero-posterior axis of the embryo. Once activated, the segment-polarity genes engage in cross-regulatory interactions that maintain their expression. As a result, En and its direct target hh become expressed throughout development in the posterior compartment of every segment, where they regulate cell tension and adhesive characteristics that prevent posterior cells from mixing with anterior compartment cells, generating stable signalling boundaries. Mutant embryos for either en or hh result in an almost complete fusion of segments. As the posterior spiracle is one of the few circumferential organs in the embryo, en activity in anterior A8 cells could be required for establishing the circumferential information pattern necessary for spiracle organogenesis that contributed to the evolution of the protruding posterior spiracles characteristic of dipteran larvae (Molina-Gil, 2023).
This study investigated when en was recruited to the anterior A8 cells (A8a) and the cis and trans regulatory elements responsible for it. A8a expression appeared in Diptera before the evolution of the posterior lobe, and evidence is presented that this is associated with a previous posterior spiracle gene network co-option event to the testis mesoderm. Working with Drosophila melanogaster, this study showed that Engrailed expression in the A8a compartment is not required for the spiracle's development and that the engrailed CRE controlling spiracle expression is required in the testis cyst cells for spermiation. This work presents an example of repeated sequential gene network co-option events involving tissues of different germ layers, and shows how this resulted in the generation of a bona fide pre-adaptive developmental expression novelty: the activation of the En transcription factor in the anterior compartment of the embryonic A8 segment where, despite having no specific function, it opens the possibility of this important developmental factor acquiring one in the future. The expression of en in the anterior compartment of the A8 segment, was likely caused by the regulatory interlocking of the co-opted networks. It is proposed that gene network interlocking occurs as the result of the use of the same gene network in several organs, so that any change to the network because of its functionality in one organ, will be mirrored in all organs even if it has no selective advantage in some of them (Molina-Gil, 2023).
Previous work showed that the ectodermally expressed posterior spiracle gene network had been co-opted to the ectodermal male genitalia in the Drosophila melanogaster clade. This study shows the same gene network has also been co-opted to the mesodermal head cyst cells (HCCs) in Drosophila. Part of the posterior spiracle gene network becomes activated in the HCCs using the same enhancers driving expression in the posterior spiracle (Molina-Gil, 2023).
Although Abd-B is expressed in the somatic testis cells during embryogenesis, it is not expressed in the adult HCCs. Two alternative explanations are suggested. The first possibility is that one of the spiracle primary targets, many of which encode transcription factors, becomes expressed in the testis HCC independently of Abd-B regulation, and this results in the activation of the other spiracle genes due to cross-regulatory network interactions. Alternatively, the embryonic Abd-B expression could be speculated to epigenetically modify the spiracle gene network leaving it in a poised state that later could become activated in the absence of Abd-B (Molina-Gil, 2023).
The finding that the posterior spiracle gene network has been co-opted twice to different organs, suggests that the coordinated activation of several transcription factors and signalling molecules of the network does not necessarily cause detrimental effects. The reported effects of experimentally inducing the ectopic activation of the eye gene regulatory network in D. melanogaster, a situation akin to what may happen during co-option, may help in understanding this. eyeless activation in the imaginal primordia results in the formation of ectopic eyes and causes morphological alterations leading to lethality. However, it has been noted that ectopic Eyeless expression in the wing only induces eye development in proximal cells where Dpp is also expressed, and similar results were observed with Hh43,44 showing that the activation of a gene network inducer does not cause developmental transformations in all cells where it is expressed. Thus, when the co-option of a gene network causes developmental transformations reducing the animal's fitness, it will be lost. However, if the co-option had no influence on local developmental processes, it could be tolerated giving the opportunity to the gene network elements to interact with allelic variants present in the population whose interaction could result in selective pressures fixing the trait. Such a series of steps may have led to the co-option of other developmental gene networks such as the activation of the appendage gene network in the butterfly wings that resulted in the formation of novel wing spot patterns (Molina-Gil, 2023).
Expression of Engrailed in posterior metameric stripes is characteristic of arthropods, but is also observed in Onycophorans and in certain worms indicating an ancient origin12,46,47. In flies, anterior En activation has rarely been reported48, and in D. melanogaster, anterior expression is the exception. En activation in A8a associated to the posterior spiracle is present in D. virilis, but not in E. balteatus, suggesting it originated in the higher Diptera (Brachicera). This is supported by analyses showing that in other Diptera like Bactrocera dorsalis (Tephritidae) and Lucilia sericata (Calliphoridae), that present well-developed larval posterior spiracles, engrailed expression is restricted to the posterior A8 segment. The A8a engrailed spiracle expression does not depend on the segmentation gene network, but is regulated by the posterior spiracle network through Abd-B, Sal and the JAK/STAT signalling pathway. Despite its complex regulation, this en enhancer has no function in Drosophila melanogaster's spiracle organogenesis. Taken together, Engrailed expression in the anterior compartment of A8 was acquired after functional posterior spiracles already existed in dipteran larvae and its origin had to do with testis evolution. Future experiments should establish if this occurred due to the de novo appearance of the enD CRE, or to a silent enD cryptic enhancer becoming activated due to the recruitment of a new transregulatory factor in the spiracles/testis gene network or to changes in chromatin accessibility (Molina-Gil, 2023).
Most insects have a pair of non-protruding spiracle openings in each trunk segment (see Interlocking of co-opted gene networks). In the hemimetabolous bug Oncopeltus fasciatus nymph or the holometabolous beetle Tribolium castaneum larva these are formed by an internal spiracular chamber that expresses the Ct protein,; however, in Oncopeltus, Ct expression is not regulated by Hox proteins as it is in Drosophila, but depends on the tracheal protein Trachealess. Although in Tribolium, Spalt is expressed in the lateral ectoderm, it is not associated with the spiracles. Similarly, Engrailed is restricted to a posterior stripe of cells in the segment that does not surround the spiracular opening. Analysis of E. balteatus embryos suggests Sal was recruited in Diptera to the protruding stigmatophore before engrailed was expressed in A8a. Analysis of Sal and En expression does not reveal any activation in E. balteatus testis, indicating the spiracle network is not expressed in the gonads of all Diptera. This situation changes in Drosophila species where both Sal and En are expressed in A8a associated with the spiracles as well as in the testes. The co-option of the posterior spiracle cascade to the testis and the recruitment of Engrailed expression may have been the result of the major sperm size and gonad morphology changes occurring at some point after the divergence of Episyrphus and Drosophila that could have required the selection of new genetic variants to maintain fertility. As the enD enhancer has no apparent function in the posterior spiracles, but is required for male spermiation in D. melanogaster, it is proposed that originally this CRE was selected for its testicular function and that expression in the posterior spiracles was a side effect caused by the interlocked regulation of the co-opted gene networks in both organs which was not eliminated because it had no deleterious effects. Finally, after the divergence from the virilis group, a second co-option event of the spiracle gene network in the male genital disc of D. melanogaster and closely related species led to the evolution of the posterior lobe. Given the extreme variability of the posterior lobe sizes between various D. melanogaster genotypes it is unclear whether the enD enhancer has a direct function on lobe development or if its expression there is aphenotypic, and only the result of its interlocking with other functional elements of the co-opted gene network (Molina-Gil, 2023).
These results suggest that the selection of a new trait in a co-opted gene network in one organ, results in its activation in all organs where the network is expressed. This implies a slower acquisition of new traits in co-opted gene networks as novelties would become discarded if they had detrimental effects in any of the interlocked organs. On the other hand, if the new trait (for example the activation of a novel transcription factor) was selected for its function in one organ, this would lead to the fixation of non-functional pre-adaptive traits with functional potential in all interlocked organs. Thus, it is proposed that co-option can lead to sensu stricto pre-adaptation cases, as opposed to the associated term of exaptation. While in exaptation the co-opted character had a previous selective function that has been recruited to perform a novel function (i.e., heat shock proteins recruited to form the eye crystallin, or feathers that could have served for heat regulation or sexual display before being co-opted for flight) cases like the expression of Engrailed in the posterior spiracle provide no selective advantage but could conceivably acquire it in the future, as has happened in the anterior wing of several Diptera where En has acquired a new role in wing pigmentation (Molina-Gil, 2023).
Tight barriers are crucial for animals. Insect respiratory cells establish barriers through their extracellular matrices. These chitinous-matrices must be soft and flexible to provide ventilation, but also tight enough to allow oxygen flow and protection against dehydration, infections, and environmental stresses. However, genes that control soft, flexible chitin-matrices are poorly known. This study investigated the genes of the chitinolytic glycosylhydrolase-family 18 in the tracheal system of Drosophila melanogaster. Five chitinases and three chitinase-like genes were found to organize the tracheal chitin-cuticles. Most of the chitinases degrade chitin from airway lumina to enable oxygen delivery. They further improve chitin-cuticles to enhance tube stability and integrity against stresses. Unexpectedly, some chitinases also support chitin assembly to expand the tube lumen properly. Moreover, Chitinase2 plays a decisive role in the chitin-cuticle formation that establishes taenidial folds to support tube stability. Chitinase2 is apically enriched on the surface of tracheal cells, where it controls the chitin-matrix architecture independently of other known cuticular proteins or chitinases. It is supposed that the principle mechanisms of chitin-cuticle assembly and degradation require a set of critical glycosylhydrolases for flexible and not-flexible cuticles. The same glycosylhydrolases support thick laminar cuticle formation and are evolutionarily conserved among arthropods (Behr, 2020).
Hedgehog (Hh) is an evolutionarily conserved signaling protein that has essential roles in animal development and homeostasis. This study investigated Hh signaling in the region of the Drosophila wing imaginal disc that produces Hh and is near the tracheal air sac primordium (ASP) and myoblasts. Hh distributes in concentration gradients in the anterior compartment of the wing disc, ASP and myoblasts, and activates genes in each tissue. Some targets of Hh signal transduction are common to the disc, ASP and myoblasts, whereas others are tissue-specific. Signaling in the three tissues is cytoneme-mediated and cytoneme-dependent. Some ASP cells project cytonemes that receive both Hh and Branchless (Bnl), and some targets regulated by Hh signaling in the ASP are also dependent on Bnl signal transduction. It is concluded that the single source of Hh in the wing disc regulates cell type-specific responses in three discreet target tissues (Hatori, 2020).
In many animals, progression of developmental stages is temporally controlled by steroid hormones. In Drosophila, the level of ecdysone titer oscillates and developmental stage transitions, such as larval molting and metamorphosis, are induced at each of ecdysone peaks. Ecdysone titer also peaks at the stage of mid-embryogenesis and the embryonic ecdysone is necessary for morphogenesis of several organs, although the regulatory mechanisms of embryonic organogenesis dependent on ecdysone signaling are still open questions. This study found that absence or interruption of embryonic ecdysone signaling caused multiple defects in the tracheal system, including decrease of luminal protein deposition, uneven dilation of the dorsal trunk and loss of terminal branches. It was also revealed that an ecdysone-inducible gene polished rice (pri) is essential for tip cell fate decision in dorsal branches. As overexpression of pri can restore the defects caused by disturbance of ecdysone biosynthesis, pri functions as one of the major mediators of embryonic ecdysone signal in tracheogenesis. These results demonstrate that ecdysone and its downstream target pri play essential roles in tracheal development by modulating cell fate decision (Taira, 2021).
Insulin signaling in Drosophila has a significant role in regulating growth, metabolism, fecundity, stress response, and longevity. The molecular mechanism by which insulin signaling regulates these vital processes is dependent on the nutrient status and oxygen availability of the organism. In a genetic screen to identify novel genes that regulate Drosophila insulin signalling, Lumens interrupted (lint), a gene that has previously been shown to act in tracheal development, was discovered. The knockdown of lint gene expression using a Dilp2Gal4 driver which expresses in the neuronal insulin-producing cells (IPCs), led to defects in systemic insulin signaling, metabolic status and growth. However, this analysis of lint knockdown phenotypes revealed that downregulation of lint in the trachea and not IPCs was responsible for the growth phenotypes, as the Gal4 driver is also expressed in the tracheal system. We found various tracheal terminal branch defects, including reduction in the length as well as number of branches in the lint knockdown background. This study reveals that substantial effects of lint downregulation arose because of tracheal defects, which induced tissue hypoxia, altered systemic insulin/TOR signaling, and resulted in effects on developmental growth regulation (Pathak, 2021).
Progenitors of the thoracic tracheal system of adult Drosophila (tracheoblasts) arrest in G2 during larval life and rekindle a mitotic program subsequently. G2 arrest is dependent on ATR-dependent phosphorylation of Chk1 that is actuated in the absence of detectable DNA damage. This study looks into in the mechanisms that activate ATR/Chk1. Levels of reactive oxygen species (ROS) are high in arrested tracheoblasts and decrease upon mitotic re-entry. High ROS is dependent on expression of Duox, an H2O2 generating-Dual Oxidase. ROS quenching by overexpression of Superoxide Dismutase 1, or by knockdown of Duox, abolishes Chk1 phosphorylation and results in precocious proliferation. Tracheae deficient in Duox, or deficient in both Duox and regulators of DNA damage-dependent ATR/Chk1 activation (ATRIP/TOPBP1/Claspin), can induce phosphorylation of Chk1 in response to micromolar concentrations of H2O2 in minutes. The findings presented reveal that H2O2 activates ATR/Chk1 in tracheoblasts by a non-canonical, potentially direct, mechanism (Kizhedathu, 2021).
Abrams, E. W. and Andrew, D. J. (2005). CrebA regulates secretory activity in the Drosophila salivary gland and epidermis. Development 132: 2743-2758. PubMed ID: 15901661
Baer, M. M., Bilstein, A., Caussinus, E., Csiszar, A., Affolter, M. and Leptin, M. (2010). The role of apoptosis in shaping the tracheal system in the Drosophila embryo. Mech. Dev. 127(1-2): 28-35. PubMed ID: 19995601
Bardet, P. L. et al. (2008). A fluorescent reporter of caspase activity for live imaging. Proc. Natl. Acad. Sci. 105: 13901-13905. PubMed ID: 18779587
Behr, M. and Riedel, D. (2020). Glycosylhydrolase genes control respiratory tubes sizes and airway stability. Sci Rep 10(1): 13377. PubMed ID: 32770153
Best, B. T. and Leptin, M. (2020). Multiple Requirements for Rab GTPases in the Development of Drosophila Tracheal Dorsal Branches and Terminal Cells. G3 (Bethesda). PubMed ID: 31980432
Booth, A. J., Blanchard, G. B., Adams, R. J. and Roper, K. (2014). A dynamic microtubule cytoskeleton directs medial actomyosin function during tube formation. Dev Cell 29: 562-576. PubMed ID: 24914560
Bossen, J., Prange, R., Kuhle, J. P., Kunzel, S., Niu, X., Hammel, J. U., Krieger, L., Knop, M., Ehrhardt, B., Uliczka, K., Krauss-Etschmann, S. and Roeder, T. (2023). Adult and Larval Tracheal Systems Exhibit Different Molecular Architectures in Drosophila. Int J Mol Sci 24(6). PubMed ID: 36982710
Bunt, S., Hooley, C., Hu, N., Scahill, C., Weavers, H. and Skaer, H. (2010). Hemocyte-secreted type IV collagen enhances BMP signaling to guide renal tubule morphogenesis in Drosophila. Dev Cell 19: 296-306. PubMed ID: 20708591
Burguete, A. S., Francis, D., Rosa, J. and Ghabrial, A. (2019). The regulation of cell size and branch complexity in the terminal cells of the Drosophila tracheal system. Dev Biol. PubMed ID: 30735663
Camelo, C., Korte, A., Jacobs, T. and Luschnig, S. (2022). Tracheal tube fusion in Drosophila involves release of extracellular vesicles from multivesicular bodies. J Cell Sci. PubMed ID: 35019140
Casani, S., Casanova, J. and Llimargas, M. (2020). Unravelling the distinct contribution of cell shape changes and cell intercalation to tissue morphogenesis: the case of the Drosophila trachea. Open Biol 10(11): 200329. PubMed ID: 33234070
Caviglia, S. and Luschnig, S. (2013). The ETS domain transcriptional repressor Anterior open inhibits MAP kinase and Wingless signaling to couple tracheal cell fate with branch identity. Development 140: 1240-1249. PubMed ID: 23444354
Chan, A. C., Drakos, S. G., Ruiz, O. E., Smith, A. C., Gibson, C. C., Ling, J., Passi, S. F., Stratman, A. N., Sacharidou, A., Revelo, M. P., Grossmann, A. H., Diakos, N. A., Davis, G. E., Metzstein, M. M., Whitehead, K. J. and Li, D. Y. (2011). Mutations in 2 distinct genetic pathways result in cerebral cavernous malformations in mice. J Clin Invest 121(5): 1871-1881. PubMed ID: 21490399
Chandran, R. R., Scholl, A., Yang, Y. and Jiang, L. (2018). rebuff regulates apical luminal matrix to control tube size in Drosophila trachea. Biol Open 7(9). PubMed ID: 30185423
Denes, A. S., Kanca, O. and Affolter, M. (2015). A cellular process that includes asymmetric cytokinesis remodels the dorsal tracheal branches in Drosophila larvae. Development 142: 1794-1805. PubMed ID: 25968315
Dong, B., Kakihara, K., Otani, T., Wada, H. and Hayashi, S. (2013). Rab9 and retromer regulate retrograde trafficking of luminal protein required for epithelial tube length control. Nat Commun 4: 1358. PubMed ID: 23322046
Dong, B., Miao, G. and Hayashi, S. (2014). A fat body-derived apical extracellular matrix enzyme is transported to the tracheal lumen and is required for tube morphogenesis in Drosophila. Development 141: 4104-4109. PubMed ID: 25336738
Dong, Q., Brenneman, B., Fields, C. and Srivastava, A. (2015). A Cathepsin-L is required for invasive behavior during air sac primordium development in Drosophila melanogaster. FEBS Lett 589(20 Pt B):3090-7. PubMed ID: 26341534
Drees, L., Konigsmann, T., Jaspers, M. H. J., Pflanz, R., Riedel, D., Schuh, R. (2019). Conserved function of the matriptase-prostasin proteolytic cascade during epithelial morphogenesis. PLoS Genet, 15(1):e1007882 PubMed ID: 30601807
Drees, L., Schneider, S., Riedel, D., Schuh, R., Behr, M. (2023). The proteolysis of ZP proteins is essential to control cell membrane structure and integrity of developing tracheal tubes in Drosophila. Elife, 12 PubMed ID: 37872795
Du, L., Sohr, A., Li, Y. and Roy, S. (2022). GPI-anchored FGF directs cytoneme-mediated bidirectional contacts to regulate its tissue-specific dispersion Nat Commun 13(1): 3482. PubMed ID: 35710780
Duan, Y., Zhu, W., Zhao, X., Merzendorfer, H., Chen, J., Zou, X. and Yang, Q. (2022). Choline transporter-like protein 2 interacts with chitin synthase 1 and is involved in insect cuticle development. Insect Biochem Mol Biol 141: 103718. PubMed ID: 34982980
Estella, C., Rieckhof, G., Calleja, M. and Morata, G. (2003). The role of buttonhead and Sp1 in the development of the ventral imaginal discs of Drosophila. Development 130: 5929-5941. PubMed ID: 14561634
Fraire-Zamora, J. J., Tosi, S., Solon, J. and Casanova, J. (2021). Control of hormone-driven organ disassembly by ECM remodeling and Yorkie-dependent apoptosis. Curr Biol. PubMed ID: 34666006.
Franch-Marro, X., Martin, N., Averof, M. and Casanova, J. (2006). Association of tracheal placodes with leg primordia in Drosophila and implications for the origin of insect tracheal systems. Development 133(5): 785-90. PubMed ID: 16469971
Francis, D. and Ghabrial, A. S. (2015). Compensatory branching morphogenesis of stalk cells in the Drosophila trachea. Development 142: 2048-2057. PubMed ID: 25977367
Francis, D., Burguete, A. S. and Ghabrial, A. (2022). Regulation of Archease by the mTOR-vATPase axis. Development. PubMed ID: 36111596
Glasheen, B. M., et al. (2010). A matrix metalloproteinase mediates airway remodeling in Drosophila. Dev. Biol. 344(2): 772-83. PubMed ID: 20513443
Glassford, W. J., Johnson, W. C., Dall, N. R., Smith, S. J., Liu, Y., Boll, W., Noll, M., Rebeiz, M. (2015). Co-option of an Ancestral Hox-Regulated Network Underlies a Recently Evolved Morphological Novelty. Dev Cell, 34(5):520-531 PubMed ID: 26343453
Gold, K. S. and Bruckner, K. (2014). Drosophila as a model for the two myeloid blood cell systems in vertebrates. Exp Hematol 42(8): 717-727. PubMed ID: 24946019
Gold, K. S. and Bruckner, K. (2015). Macrophages and cellular immunity in Drosophila melanogaster. Semin Immunol 27(6): 357-368. PubMed ID: 27117654
Hanson, M. A., Dostalova, A., Ceroni, C., Poidevin, M., Kondo, S. and Lemaitre, B. (2019). Synergy and remarkable specificity of antimicrobial peptides in vivo using a systematic knockout approach. Elife 8. PubMed ID: 30803481
Hatori, R. and Kornberg, T. B. (2020). Hedgehog produced by the Drosophila wing imaginal disc induces distinct responses in three target tissues. Development 147(22). PubMed ID: 33028613
Hosono, C., Matsuda, R., Adryan, B. and Samakovlis, C. (2015). Transient junction anisotropies orient annular cell polarization in the Drosophila airway tubes. Nat Cell Biol 17: 1569-1576. PubMed ID: 26551273
Itakura, Y., Inagaki, S., Wada, H. and Hayashi, S. (2018). Trynity controls epidermal barrier function and respiratory tube maturation in Drosophila by modulating apical extracellular matrix nano-patterning. PLoS One 13(12): e0209058. PubMed ID: 30576352
Kamei, M., et al. (2006). Endothelial tubes assemble from intracellular vacuoles in vivo. Nature 442: 453-456. PubMed ID: 16799567
Kato, K., Dong, B., Wada, H., Tanaka-Matakatsu, M., Yagi, Y. and Hayashi, S. (2016). Microtubule-dependent balanced cell contraction and luminal-matrix modification accelerate epithelial tube fusion. Nat Commun 7: 11141. PubMed ID: 27067650
Kizhedathu, A., Kunnappallil, R. S., Bagul, A. V., Verma, P. and Guha, A. (2020). Multiple Wnts act synergistically to induce Chk1/Grapes expression and mediate G2 arrest in Drosophila tracheoblasts. Elife 9. PubMed ID: 32876044
Kizhedathu, A., Chhajed, P., Yeramala, L., Sain Basu, D., Mukherjee, T., Vinothkumar, K. R. and Guha, A. (2021). Duox generated reactive oxygen species activate ATR/Chk1 to induce G2 arrest in Drosophila tracheoblasts. Elife 10. PubMed ID: 34622778
Klok, C. J., Kaiser, A., Socha, J. J., Lee, W. K. and Harrison, J. F. (2016). Multigenerational effects of rearing atmospheric oxygen level on the tracheal dimensions and diffusing capacities of pupal and adult Drosophila melanogaster. Adv Exp Med Biol 903: 285-300. PubMed ID: 27343104
Klussmann-Fricke, B. J., Martin-Bermudo, M. D. and Llimargas, M. (2022). The basement membrane controls size and integrity of the Drosophila tracheal tubes. Cell Rep 39(4): 110734. PubMed ID: 35476979
Kondo, T. and Hayashi, S. (2013). Mitotic cell rounding accelerates epithelial invagination. Nature 494: 125-129. PubMed ID: 23334416
Lebreton, G. and Casanova, J. (2013). Specification of leading and trailing cell features during collective migration in the Drosophila trachea. J Cell Sci. 127(Pt 2): 465-74. PubMed ID: 24213534
Le Droguen, P.M., Claret, S., Guichet, A. and Brodu, V. (2015). Microtubule-dependent apical restriction of recycling endosomes sustains adherens junctions during morphogenesis of the Drosophila tracheal system. Development 142: 363-374. PubMed ID: 25564624
Li, Y., Dong, P., Yang, Y., Guo, T., Zhao, Q., Miao, D., Li, H., Lu, T., Xia, F., Lyu, J., Ma, J., Kornberg, T. B., Zhang, Q. and Huang, H. (2022). Metabolic control of progenitor cell propagation during Drosophila tracheal remodeling. Nat Commun 13(1): 2817. PubMed ID: 35595807
Loganathan, R., Levings, D. C., Kim, J. H., Wells, M. B., Chiu, H., Wu, Y., Slattery, M. and Andrew, D. J. (2022). Ribbon boosts ribosomal protein gene expression to coordinate organ form and function. J Cell Biol 221(4). PubMed ID: 35195669
Lubarsky, B. and Krasnow, M. A. (2003). Tube morphogenesis: making and shaping biological tubes. Cell 112: 19-28. PubMed ID: 12526790
Makhijani, K., Alexander, B., Tanaka, T., Rulifson, E. and Bruckner, K. (2011). The peripheral nervous system supports blood cell homing and survival in the Drosophila larva. Development 138(24): 5379-5391. PubMed ID: 22071105
Makhijani, K. and Bruckner, K. (2012). Of blood cells and the nervous system: hematopoiesis in the Drosophila larva. Fly (Austin) 6(4): 254-260. PubMed ID: 23022764
Marmion, R. A., Simpkins, A. G., Barrett, L. A., Denberg, D. W., Zusman, S., Schottenfeld-Roames, J., Schupbach, T. and Shvartsman, S. Y. (2023). Stochastic phenotypes in RAS-dependent developmental diseases. Curr Biol. PubMed ID: 36706752
Martin-Belmonte, F., et al. (2007). PTEN-mediated apical segregation of phosphoinositides controls epithelial morphogenesis through Cdc42. Cell 128: 383-397. PubMed ID: 17254974
Mathew, R., Rios-Barrera, L. D., Machado, P., Schwab, Y. and Leptin, M. (2020). Transcytosis via the late endocytic pathway as a cell morphogenetic mechanism. Embo j: e105332. PubMed ID: 32657472
Medina, A. B., Perochon, J., Tian, Y., Johnson, C. T., Holcombe, J., Ramesh, P., Polcownuk, S., Yu, Y., Cordero, J. B. (2025). Neuroendocrine control of intestinal regeneration through the vascular niche in Drosophila. Dev Cell, 60(22):3085-3101 e3086 PubMed ID: 40695286
Molina-Gil, S., Sotillos, S., Espinosa-Vazquez, J. M., Almudi, I., Hombría, J. C. (2023). Interlocking of co-opted developmental gene networks in Drosophila and the evolution of pre-adaptive novelty. Nat Commun, 14(1):5730 PubMed ID: 37714829
Morishita, K., Anh Suong, D. N., Yoshida, H. and Yamaguchi, M. (2017). The Drosophila DOCK family protein Sponge is required for development of the air sac primordium. Exp Cell Res [Epub ahead of print]. PubMed ID: 28341448
Nelson, K. S., Furuse, M., Beitel, G. J. (2010). The Drosophila Claudin Kune-kune is required for septate junction organization and tracheal tube size control. Genetics 185(3): 831-9. PubMed ID: 20407131
Nikolova, L. S. and Metzstein, M. M. (2015). Intracellular lumen formation in Drosophila proceeds via a novel subcellular compartment. Development [Epub ahead of print]. PubMed ID: 26428009
Olivares-Castineira, I. and Llimargas, M. (2018). Anisotropic Crb accumulation, modulated by Src42A, is coupled to polarised epithelial tube growth in Drosophila. PLoS Genet 14(11): e1007824. PubMed ID: 30475799
Olver, R. E., Walters, D. V. and Wilson, S. M. (2004). Developmental regulation of lung liquid transport, Annu. Rev. Physiol. 66: 77-101. PubMed ID: 14977397
Ozturk-Colak, A., Moussian, B., Araujo, S. J. and Casanova, J. (2016). A feedback mechanism converts individual cell features into a supracellular ECM structure in Drosophila trachea. Elife 5. PubMed ID: 26836303
Ozturk-Colak, A., Stephan-Otto Attolini, C., Casanova, J. and Araujo, S. J. (2018). Blimp-1 mediates tracheal lumen maturation in Drosophila melanogaster. Genetics. PubMed ID: 30082278
Papakyrikos, A. M., Kim, M. J. and Wang, X. (2020). Drosophila PTPMT1 Has a Function in Tracheal Air Filling. iScience 23(7): 101285. PubMed ID: 32629421
Pathak, H., Maya, A. V., Tanari, A. B., Sarkar, S. and Varghese, J. (2021). Lint, a transmembrane serine protease, regulates growth and metabolism in Drosophila. Genetics. PubMed ID: 33693655
Pereanu, W., Spindler, S., Cruz, L. and Hartenstein, V. (2006). Tracheal development in the Drosophila brain is constrained by glial cells. Dev. Biol. 302(1): 169-80. PubMed ID: 17046740
Perens, E. A. and Shaham S. (2005). C. elegans daf-6 encodes a patched-related protein required for lumen formation. Dev. Cell 8(6): 893-906. PubMed ID: 15935778
Perochon, J., Yu, Y., Aughey, G. N., Medina, A. B., Southall, T. D. and Cordero, J. B. (2021). Dynamic adult tracheal plasticity drives stem cell adaptation to changes in intestinal homeostasis in Drosophila. Nat Cell Biol 23(5): 485-496. PubMed ID: 33972729
Pesch, Y. Y., Riedel, D. and Behr, M. (2016). Drosophila Chitinase 2 is expressed in chitin producing organs for cuticle formation. Arthropod Struct Dev [Epub ahead of print]. PubMed ID: 27832982
Pino-Jimenez, B., Giannios, P. and Casanova, J. (2023). Polyploidy-associated autophagy promotes larval tracheal histolysis at Drosophila metamorphosis. Autophagy: 1-10. PubMed ID: 37424089
Poon, C. L. C., Liu, W., Song, Y., Gomez, M., Kulaberoglu, Y., Zhang, X., Xu, W., Veraksa, A., Hergovich, A., Ghabrial, A. and Harvey, K. F. (2018). A Hippo-like signaling pathway controls tracheal morphogenesis in Drosophila melanogaster. Dev Cell 47(5): 564-575. PubMed ID: 30458981
Pradhan, R., Kumar, S. and Mathew, R. (2023). Lateral adherens junctions mediate a supracellular actomyosin cortex in Drosophila trachea. iScience 26(4): 106380. PubMed ID: 37009223
Rao, P.R., Lin, L., Huang, H., Guha, A., Roy, S. and Kornberg, T.B. (2015). Developmental compartments in the larval trachea of Drosophila. Elife [Epub ahead of print]. PubMed ID: 26491942
Ricolo, D., Deligiannaki, M., Casanova, J. and Araújo, S.J. (2016). Centrosome amplification increases single-cell branching in post-mitotic cells. Curr Biol [Epub ahead of print]. PubMed ID: 27693136
Rios-Barrera, L. D., Sigurbjornsdottir, S., Baer, M. and Leptin, M. (2017). Dual function for Tango1 in secretion of bulky cargo and in ER-Golgi morphology. Proc Natl Acad Sci U S A 114(48): E10389-E10398. PubMed ID: 29138315
Rios-Barrera, L. D. and Leptin, M. (2022). An endosome-associated actin network involved in directed apical plasma membrane growth. J Cell Biol 221(3). PubMed ID: 35061016
Rosa, J. B., Metzstein, M. M. and Ghabrial, A. S. (2018). An Ichor-dependent apical extracellular matrix regulates seamless tube shape and integrity. PLoS Genet 14(1): e1007146. PubMed ID: 29309404
Ruiz-Sobrino, A., Martin-Blanco, C. A., Navarro, T., Almudi, I., Masiero, G., Jimenez-Caballero, M., Buchwalter, D. B., Funk, D. H., Gattolliat, J. L., Lemos, M. C., Jimenez, F. and Casares, F. (2020). Space colonization by branching trachea explains the morphospace of a simple respiratory organ. Dev Biol. PubMed ID: 32109442
Sanchez-Higueras, C., Sotillos, S. and Castelli-Gair Hombria, J. (2013). Common origin of insect trachea and endocrine organs from a segmentally repeated precursor. Curr Biol. 24(1):76-81. PubMed ID: 24332544
Sanchez Bosch, P., Makhijani, K., Herboso, L., Gold, K. S., Baginsky, R., Woodcock, K. J., Alexander, B., Kukar, K., Corcoran, S., Jacobs, T., Ouyang, D., Wong, C., Ramond, E. J. V., Rhiner, C., Moreno, E., Lemaitre, B., Geissmann, F. and Bruckner, K. (2019). Adult Drosophila lack hematopoiesis but rely on a blood cell reservoir at the respiratory epithelia to relay infection signals to surrounding tissues. Dev Cell. PubMed ID: 31735669
Scholl, A., Ndoja, I., Dhakal, N., Morante, D., Ivan, A., Newman, D., Mossington, T., Clemans, C., Surapaneni, S., Powers, M. and Jiang, L. (2023). The Osiris family genes function as novel regulators of the tube maturation process in the Drosophila trachea. PLoS Genet 19(1): e1010571. PubMed ID: 36689473
Schottenfeld-Roames, J. and Ghabrial, A. S. (2012). Whacked and Rab35 polarize dynein-motor-complex-dependent seamless tube growth. Nat Cell Biol 14: 386-393. PubMed ID: 22407366
Shin, J. H. and Jeong, C. W. (2021). Zipper Is Necessary for Branching Morphogenesis of the Terminal Cells in the Drosophila melanogaster's Tracheal System. Biology (Basel) 10(8). PubMed ID: 34439961
Shin, M., Chang, E., Lee, D., Kim, N., Cho, B., Cha, N., Koranteng, F., Song, J. J., Shim, J. (2024). Drosophila immune cells transport oxygen through PPO2 protein phase transition Nature, 631(8020):350-359 PubMed ID: 38926577
Sirocko, K. T., Angstmann, H., Papenmeier, S., Wagner, C., Spohn, M., Indenbirken, D., Ehrhardt, B., Kovacevic, D., Hammer, B., Svanes, C., Rabe, K. F., Roeder, T., Uliczka, K. and Krauss-Etschmann, S. (2022). Early-life exposure to tobacco smoke alters airway signaling pathways and later mortality in D. melanogaster. Environ Pollut: 119696. PubMed ID: 35780997
Skouloudaki, K., Papadopoulos, D. K., Tomancak, P. and Knust, E. (2019). The apical protein Apnoia interacts with Crumbs to regulate tracheal growth and inflation. PLoS Genet 15(1): e1007852. PubMed ID: 30645584
Taira, Y., Wada, H., Hayashi, S. and Kageyama, Y. (2021). polished rice mediates ecdysone-dependent control of Drosophila embryonic organogenesis. Genes Cells. PubMed ID: 33621395
Tamamouna, V., Rahman, M. M., Petersson, M., Charalambous, I., Kux, K., Mainor, H., Bolender, V., Isbilir, B., Edgar, B. A. and Pitsouli, C. (2021). Remodelling of oxygen-transporting tracheoles drives intestinal regeneration and tumorigenesis in Drosophila. Nat Cell Biol 23(5): 497-510. PubMed ID: 33972730
Truong, T. V., Bower, D. J., Featherstone, N. C., Connell, M. G., Al Alam, D., Frey, M. R., Trinh, L. A., Fernandez, G. E., Warburton, D., Fraser, S. E., Bennett, D. and Jesudason, E. C. (2017). SERCA directs cell migration and branching across species and germ layers. Biol Open 6(10): 1458-1471. PubMed ID: 28821490
Tsarouhas, V., et al. (2007). Sequential pulses of apical epithelial secretion and endocytosis drive airway maturation in Drosophila. Dev. Cell 13: 214-225. PubMed ID: 17681133
Wagner, C., Uliczka, K., Bossen, J., Niu, X., Fink, C., Thiedmann, M., Knop, M., Vock, C., Abdelsadik, A., Zissler, U. M., Isermann, K., Garn, H., Pieper, M., Wegmann, M., Koczulla, A. R., Vogelmeier, C. F., Schmidt-Weber, C. B., Fehrenbach, H., Konig, P., Silverman, N., Renz, H., Pfefferle, P., Heine, H. and Roeder, T. (2021). Constitutive immune activity promotes JNK- and FoxO-dependent remodeling of Drosophila airways. Cell Rep 35(1): 108956. PubMed ID: 33826881
Wang, Y., Berger, J. and Moussian, B. (2018). Trynity models a tube valve in the Drosophila larval airway system. Dev Biol [Epub ahead of print]. PubMed ID: 29518377
Weaver, M. and Krasnow, M. A. (2008). Dual origin of tissue-specific progenitor cells in Drosophila tracheal remodeling. Science 321(5895): 1496-9. PubMed ID: 18669822
Wester, J., Lima, C. A. C., Machado, M. C. R., Zampar, P. V., Tavares, S. S. and Monesi, N. (2018). Characterization of a novel Drosophila melanogaster cis-regulatory module that drives gene expression to the larval tracheal system and adult thoracic musculature. Genesis: e23222. PubMed ID: 30096221
Wilk, R., Weizman, I. and Shilo, B-Z. (1996). trachealess encodes a bHLH-PAS protein that is an inducer of tracheal cell fates in Drosophila. Genes and Devel. 10: 93-102. PubMed ID: 8557198
Yang, R., Li, E., Kwon, Y. J., Mani, M. and Beitel, G. J. (2019). QuBiT: a quantitative tool for analyzing epithelial tubes reveals unexpected patterns of organization in the Drosophila trachea. Development. PubMed ID: 30967427
Yao, L., Wang, S., Orzechowski-Westholm, J., Dai, Q., Matsuda, R., Hosono, C., Bray, S., Lai, E. C. and Samakovlis, C. (2017). Genome-wide identification of Grainy head targets in Drosophila reveals regulatory interactions with the POU-domain transcription factor, Vvl. Development 144: 3145-3155. PubMed ID: 28760809
Zhang, L. and Ward, R. E. t. (2009). uninflatable encodes a novel ectodermal apical surface protein required for tracheal inflation in Drosophila. Dev Biol 336: 201-212. PubMed ID: 19818339
Zhou, F., Qiang, K. M. and Beckingham, K. M. (2016). Failure to burrow and tunnel reveals roles for jim lovell in the growth and endoreplication of the Drosophila larval tracheae. PLoS One 11: e0160233. PubMed ID: 27494251
genes expressed in trachea and spiracles
Genes involved in organ development
Home page: The Interactive Fly © 2025 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
{kind=link}
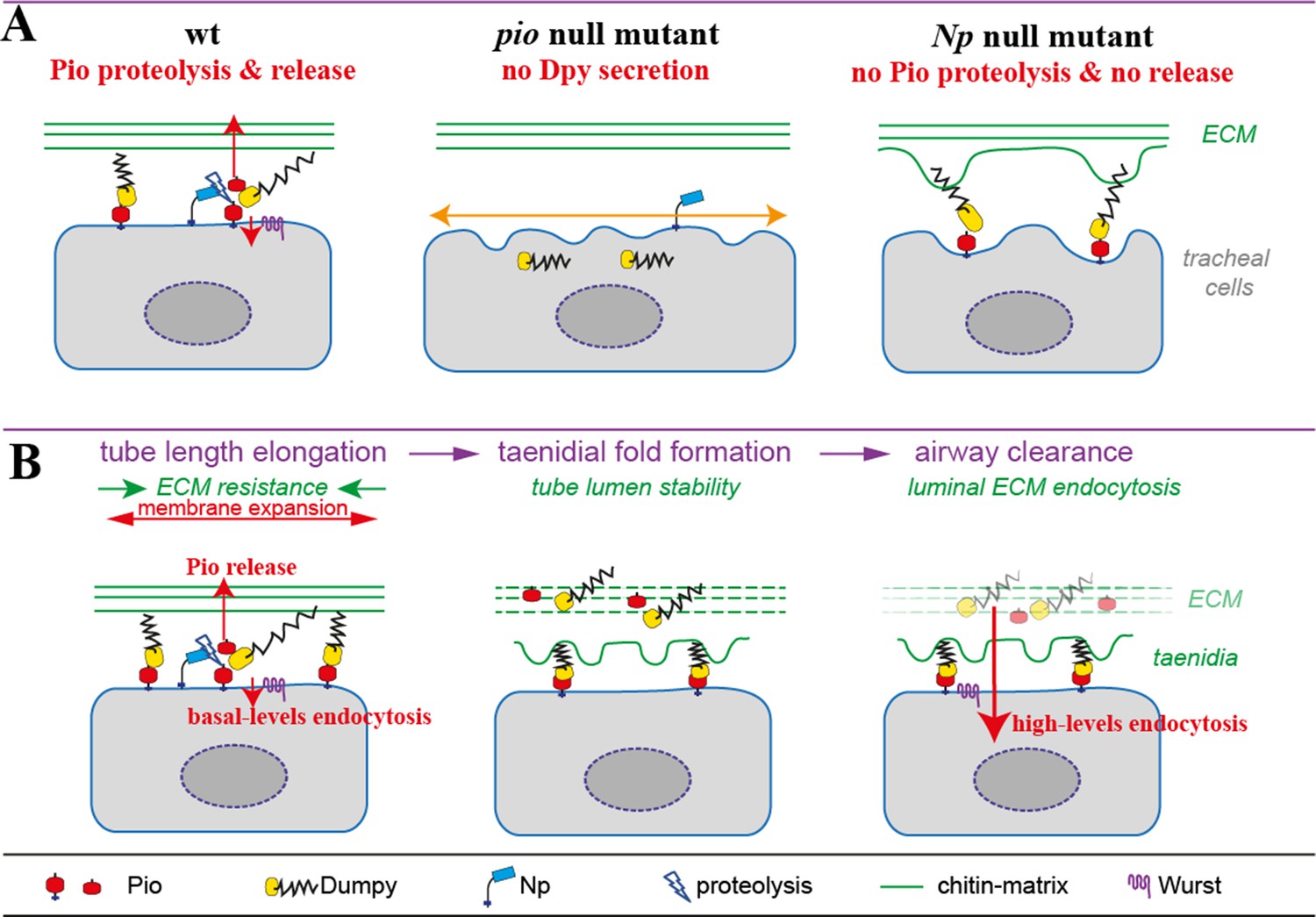{kind=link}
{kind=link}