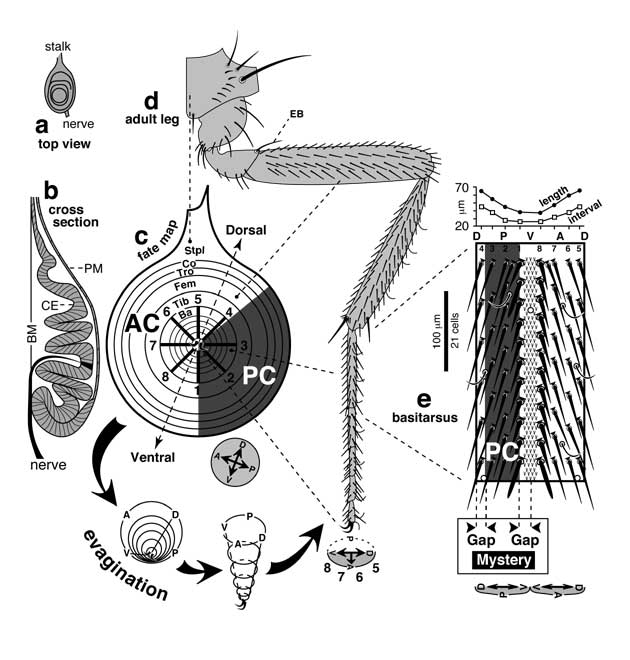
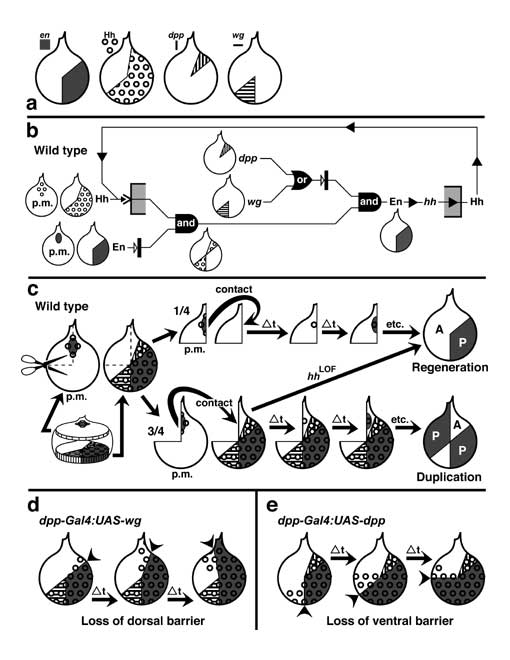
Select image to enlarge
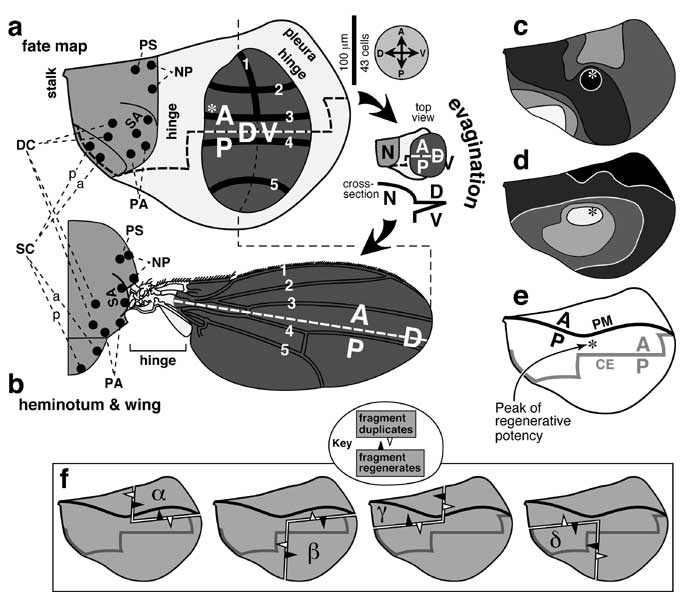
Figure 6.1
Fate map of the wing disc and regenerative potency of its fragments.
a. Fate map (abridged) of a mature right wing disc as per Bryant [524] except that dots are actual SOP sites [1925] and thick lines are pre-vein zones (1-5) [4189]. (See [834, 987, 1368, 3372, 4429] for further details.) The wing is darkly shaded, the heminotum (a.k.a. body wall) medium shaded, and the hinge and pleura lightly shaded. The wing disc is ~300 μm across X ~450 μm long [524] -- roughly twice the dimensions of a leg disc (cf. Fig. 5.1). Like the 1st- and 2nd-leg discs (vs. the 3rd [3422]) the wing disc has a neural connection to the CNS [2128], but the nerve fiber enters along with the trachea (not shown) [3565]. Based on cell diameters in the wing pouch (mean of 10 counts along 25 μm transects in Fig. 8b of [350]; cf. scale bar), the disc would be 130 cells across X 190 cells long, but these are underestimates since the epithelium is highly folded. Vein 1 is the bristled part of the anterior wing margin [1741] (thin dashed line = posterior margin). The A/P compartment boundary is drawn as a thick dashed line. Directions (A, anterior; P, posterior; D, dorsal; V, ventral) are given in the compass at right. Bristle names ('a' and 'p' = ant. or post. members): DC (dorsocentrals), NP (notopleurals), PA (post-alars), PS (presutural), SA (supra-alars), SC (scutellars). The layout of SOPs prefigures the adult bristle pattern, except that scutellar SOPs rearrange ('p' moves posterior to 'a'). In a and c-e, the asterisk marks the peak of regenerative potency as mapped surgically (cf. Fig. 4.5).
b. Derivatives of the wing disc (shaded as per fate map). During evagination (cartoon between a and b) the wing pouch expands, folds along the D/V line, and tucks its V side underneath ('N' = notum) [1311, 3374, 4189]. When the hinge region contracts [4509], its cells pack densely [3374] with some apoptosis [2849] to form a menagerie of elements [526, 1866, 4065]. This contraption allows the wing to twist so as to maximize the lift per stroke [1045, 1115, 4748]. Distal edge of the hinge is approximate [678]. The hinge region is genetically distinct from the blade per se since (1) it overgrows in response to excess Wg [459, 2252, 2254, 3088], and (2) it does not depend on Dpp signaling [157]. Indeed, wg expression in the hinge is induced by vestigial-ON cells in the blade [2570]. Bristles (except along front edge) are omitted.
c, d. Regenerative potency, assessed by γ-irradiation of young wing discs [3440]. c. Darker shading indicates higher frequencies (50-60, -70, -80, -90. -100%) of remaining markers in discs that exhibit deficiencies but no duplications. d. Darker shading denotes higher incidence (0-5, -10, -20, -60, -80%) of duplicated markers in discs that manifest duplications and deficiencies. Note that the same (central) region resists becoming deficient (c) or duplicated (d).
e. Path of the A/P compartment boundary in the peripodial membrane (PM, solid line) and columnar epithelium (CE, shaded line) as assessed by engrailed and patched expression [1133, 2072, 2216]. The regenerative peak (asterisk) falls between the A/P lines of the two layers (in top view). The D/V boundary in the peripodial membrane has not been charted so precisely [498]. The signaling role of this membrane in normal development is only starting to be explored [773, 1473, 3508].
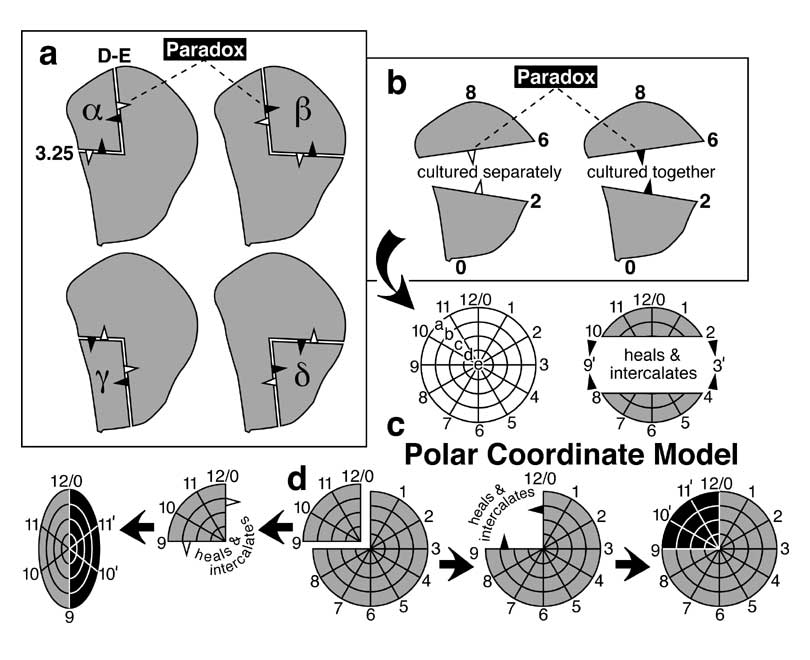
f. Superposition of the entire A/P boundary (e) upon the 1/4 (α, β, γ, δ) and 3/4 fragments whose regenerative behavior helped found the Polar Coordinate Model (cf. Fig. 4.6a). As indicated in the key (cf. Fig. 4.5), an unfilled arrowhead means that the fragment to its rear duplicates, while a solid arrowhead means that the fragment behind it regenerates. Given recent insights into leg disc regeneration (cf. Fig. 5.10c) it may be possible to figure out why quadrants duplicate and 3/4 pieces regenerate, but no unifying hypothesis is obvious based on geometry alone.
|
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}