Deformed
The six Drosophila proteins that belong to the antennapedia-type Homeobox subfamily are
Antennapedia (ANTP), Abdominal-A (ABD-A), Deformed (DFD), Proboscipedia (PB),
Sex combs reduced (SCR) and Ultrabithorax (UBX).
The ExPASy World Wide Web (WWW) molecular biology server of the Geneva University Hospital and
the University of Geneva provides extensive documentation for the 'Homeobox' antennapedia-type protein signature. See four paralogous Hox clusters of mammals for homologies of Deformed with the mammalian Hox clusters.
In C. elegans, the epithelial Pn.p cells adopt either a vulval precursor cell fate or fuse with the surrounding hypodermis (the F fate). Two pathways that control vulval precursor cell fate converge on the Hox gene lin-39: the Ras pathway, functioning downstream of the LET-23 Epidermal growth factor receptor and the Wnt pathway. LIN-39 is an Antennapedia class homeodomain, most similar to those of the Drosophila homeotic genes Deformed and Sex combs reduced. The Wnt signal is transduced through a pathway involving the beta-catenin homolog BAR-1 and controls whether P3.p through P8.p adopt the vulval precursor cell fate. In bar-1 mutants, P3.p through P8.p can adopt F fates instead of vulval precursor cell fates. The Wnt/bar-1 signaling pathway acts by regulating the expression of the Hox gene lin-39, since bar-1 is required for LIN-39 expression and forced lin-39 expression rescues the bar-1 mutant phenotype. LIN-39 activity is also regulated by the anchor cell signal/let-23 receptor tyrosine kinase/let-60 Ras signaling pathway. These genetic and molecular experiments show that the vulval precursor cells can integrate the input from the BAR-1 and LET-60 Ras signaling pathways by coordinately regulating activity of the common target, LIN-39 Hox (Eisenmann, 1998).
Vulval cell-fate determination in C. elegans requires the action of numerous gene products, including components of the Ras/Raf/MAPK signaling cascade and the hox gene lin-39, an Antennapedia class homeodomain, most similar to those of the Drosophila homeotic genes Deformed and Sex combs reduced. sem-4 encodes a zinc finger protein, homologous to Drosophila Spalt, with roles in the fate specification of sex myoblasts, coelomocytes, and multiple neuronal lineages in C. elegans. By characterizing three new alleles of sem-4, identified in a screen for vulval-defective mutants, it has been determined that loss of sem-4 activity results in abnormal specification of the secondary vulval cell lineages. sem-4 interactions with other genes involved in vulval differentiation were analyzed and it was determined that sem-4 does not function directly in the Ras-mediated signal transduction pathway but acts in close association with and upstream of lin-39 to promote vulval cell fate. sem-4 regulates lin-39 expression and it is proposed that sem-4 is a regulator of lin-39 in the vulval cell-fate determination pathway that may act to link lin-39 to incoming signals (Grant, 2000).
During larval development in C. elegans, some of the cells of the ventral epidermis, the Pn.p cells, fuse with the growing epidermal syncytium hyp7. The pattern of these cell fusions is regulated in a complex, sexually dimorphic manner. It is essential that some Pn.p cells remain unfused in order for some sex-specific mating structures to be generated. The pattern of Pn.p cell fusion is regulated combinatorially by two genes of the C. elegans Hox gene cluster: lin-39 and mab-5. Some of the complexity in the Pn.p cell fusion pattern arises because these two Hox proteins can regulate each others activities. A zinc-finger transcription factor, REF-2, is described that is required for the Pn.p cells to be generated and to remain unfused. ref-2 encodes a zinc-finger transcription factor of the odd-paired (opa)/Zic family. REF-2 functions with the Hox proteins to prevent Pn.p cell fusion. ref-2 may also be a transcriptional target of the Hox proteins (Alber, 2002).
The Notch signaling pathway controls growth, differentiation and patterning in divergent animal phyla; in humans, defective Notch signaling has been implicated in cancer, stroke and neurodegenerative disorders. Despite its developmental and medical significance, little is known about the factors that render cells to become competent for Notch signaling. This study shows that during vulval development in the nematode C. elegans the HOX protein LIN-39 and its EXD/PBX-like cofactor CEH-20 are required for LIN-12/Notch-mediated lateral signaling that specifies the 2° vulval cell fate. Inactivation of either lin-39 or ceh-20 resulted in the misspecification of 2° vulval cells and suppresses the multivulva phenotype of lin-12(n137) gain-of-function mutant animals. Furthermore, both LIN-39 and CEH-20 are required for the expression of basal levels of the genes encoding the LIN-12/Notch receptor and one of its ligands in the vulval precursor cells, LAG-2/Delta/Serrate, rendering them competent for the subsequent lin-12/Notch induction events. These results suggest that the transcription factors LIN-39 and CEH-20, which function at the bottom of the RTK/Ras and Wnt pathways in vulval induction, serve as major integration sites in coordinating and transmitting signals to the LIN-12/Notch cascade to regulate vulval cell fates (Takács-Vellai, 2007).
Convergent intercellular signals must be precisely coordinated in order to elicit specific biological responses. The C. elegans vulva provides an excellent experimental microcosm for studying how cell fate is specified according to the combined effects of different signaling pathways. This paper has studied the role of the Hox gene lin-39 and the Exd ortholog ceh-20 in vulval development. Genetic and molecular evidence is presented that the HOX protein LIN-39 and its putative cofactor CEH-20 are required for basal expression levels of lin-12 and lag-2 in the VPCs prior to vulval induction; this regulation may be important to render the VPCs competent for the subsequent lin-12/Notch induction events at the L3 larval stage. Identifying transcriptional regulators of lateral signaling in C. elegans vulval development will be essential for understanding how the Notch signaling pathway specifies cell fate in divergent animal species, and how compromised Notch signaling leads to human diseases (Takács-Vellai, 2007).
LIN-39 and CEH-20 are both required at the first larval stage to prevent fusion of the VPCs to the surrounding hypodermis. The data lead to the attractive possibility that LIN-39 and its putative cofactor CEH-20 regulate the competence of the VPCs to respond to any of the patterning signals during vulval formation. Along this line, it is challenging to speculate that, besides regulating lin-12 and lag-2 expression, they might also promote the expression of components of the inductive pathway (such as let-23) or other Notch pathway genes in the VPCs (Takács-Vellai, 2007).
It has been shown that CEH-20 binds in vitro, together with LIN-39, to the promoter of the twist transcription factor ortholog hlh-8 to regulate its expression in postembryonic mesodermal cells. ChIP experiments demonstrate that LIN-39 associates with the lag-2promoter, suggesting that the regulation of lag-2 expression by LIN-39 may be direct. It is proposed that LIN-39 forms a heterodimer with CEH-20 to promote the basal transcription of lag-2 and lin-12 in the VPCs. Based on their different expression pattern in the Pn.p lineages, ceh-20 is assumed to have some functions that are independent of lin-39. Indeed, mab-5 has been shown to be expressed in the descendants of the posterior VPCs, P7.p and P8.p, and to prevent them from adopting an induced vulval fate. Thus, it is possible that CEH-20 also interacts and functions with MAB-5 in controlling certain aspects of vulval fate specification. Furthermore, it is noted that ceh-20(ay9) mutant animals sometimes displayed a dual AC phenotype, whereas lin-39 mutants never did. RNAi-mediated depletion of mab-5 sometimes resulted in 2 ACs, suggesting that the correct AC specification requires the combined activity of mab-5 and ceh-20 (Takács-Vellai, 2007).
Finally, CEH-20 has been shown to be required as a cofactor for autoregulatory expression of the anterior Hox paralog (labial-like) ceh-13 in embryonic cells. Because ceh-13 is expressed all along the anteroposterior body axis in the ventral mid-line during the L1–L4 larval stages and a few percent of the ceh-13(sw1) mutant animals that are able to develop into fertile adults exhibit various defects in vulval formation, it is possible that CEH-13 acts with CEH-20 to control cell fate in the anterior VPC lineages. The future analysis of a potential role of ceh-13 in vulval development would help to establish the role of all of the major body Hox genes in this important process (Takács-Vellai, 2007).
Morphogenesis represents a phase of development during which cell fates are executed. The conserved hox genes are key cell fate determinants during metazoan development, but their role in controlling organ morphogenesis is less understood. This study shows that the C. elegans hox gene lin-39 regulates epidermal morphogenesis via its novel target, the essential zinc finger protein VAB-23. During the development of the vulva, the egg-laying organ of the hermaphrodite, the EGFR/RAS/MAPK signaling pathway activates, together with LIN-39 HOX, the expression of VAB-23 in the primary cell lineage to control the formation of the seven vulval toroids. VAB-23 regulates the formation of homotypic contacts between contralateral pairs of cells with the same sub-fates at the vulval midline by inducing smp-1 (semaphorin) transcription. In addition, VAB-23 prevents ectopic vulval cell fusions by negatively regulating expression of the fusogen eff-1. Thus, LIN-39 and the EGFR/RAS/MAPK signaling pathway, which specify cell fates earlier during vulval induction, continue to act during the subsequent phase of cell fate execution by regulating various aspects of epidermal morphogenesis. Vulval cell fate specification and execution are, therefore, tightly coupled processes (Pellegrino, 2011).
To analyze how the silk glands of the lepidopteran Bombyx mori develop, two genes were cloned and identified that encode the homeodomain and its flanking regions identical to
the corresponding regions of Drosophila Deformed and Sex combs reduced. The Bombyx silk gland is assumed to be an evolutionally homologous organ to the Drosophila salivary gland because both structures are formed in the labial segment and share some similar functions. Bombyx Deformed is expressed in the mandibular and maxillary segments, whereas
expression of Bombyx Sex combs reduced is first limited to the labial segment and at later stages
extended to the anterior part of the prothoracic segment. The expression of Bombyx Sex combs
reduced then disappears from the invaginating placodes of silk glands where expression of Bombyx
fork head/SGF-1 follows. In the mutant embryos, which lack the 3' end region of Bombyx
Antennapedia, Bombyx Sex combs reduced is
expressed ectopically in the thoracic and abdominal regions, in addition to expression in the labial segment. Bombyx fork head/SGF-1 is also
ectopically expressed in the T1, T2, and T3 segments, resulting in the ectopic induction of the silk gland
invaginations. These results suggest that Bombyx homeobox genes such as the Bombyx Deformed and
Sex combs reduced are associated with determination of the segment identities and Bombyx Sex
combs reduced is involved in the induction of silk gland development (Kokubo, 1997).
Molecular developmental studies of fly and mouse embryos have shown that the identity of individual
body segments is controlled by a suite of homeobox-containing genes called the Hox cluster. To
examine the conservation of this patterning mechanism in other segmented phyla,
four Hox gene homologs isolated from glossiphoniid leeches of the genus Helobdella are described. Based on
sequence similarity and phylogenetic analysis, the leech genes Lox7, Lox6, Lox20, and Lox5 are
deemed to be orthologs of the Drosophila genes lab, Dfd, Scr, and Antp, respectively. Sequence
similarities between Lox5 and Antp outside the homeodomain and phylogenetic reconstructions suggest
that the Antennapedia family of Hox genes had already expanded to
include at least two discrete Antp and Ubx/abdA precursors prior to the annelid/arthropod divergence.
In situ hybridization reveals that the four Lox genes described in this study are all expressed at high
levels within the segmented portion of the central nervous system (CNS), with variable levels of
expression in the segmental mesoderm. Little or no expression is seen in peripheral ectoderm or
endoderm, or in the unsegmented head region (prostomium). Each Lox gene has a distinct anterior
expression boundary within one of the four rostral segments, and the anterior-posterior (AP) order of
these expression boundaries is identical to that reported for the orthologous Hox gene products in fly
and mouse. This finding supports the idea that the process of AP axis differentiation is conserved
among the higher metazoan phyla with respect to the regional expression of individual Hox genes along
that axis. One unusual feature of leech Hox genes is the observation that some genes are only
expressed during later development -- beginning at the time of terminal cell differentiation -- whereas
others begin expression at a much earlier stage; their RNA ceases to be detectable shortly after
the onset of expression of the 'late' Hox genes. The functional significance of this temporal disparity is
unknown, but it is noteworthy that only the two 'early' Hox genes display high levels of mesodermal
expression (Kourakis, 1997).
Sequence analysis of a newly isolated Hirudo medicinalis cDNA containing an Antennapedia
(Antp)-class homeobox suggests that the corresponding gene, Lox6, is an ortholog of the Drosophila
Deformed gene. In situ hybridization of whole-mounted preparations shows that the major sites
of Lox6 expression during embryogenesis are the central nervous system (CNS) and the peripheral
sensory system. Lox6 mRNA can be detected in a subset of neurons in each ganglion from the
subesophageal ganglion (RG2) to the most posterior ganglion, with the highest level of expression seen
in RG3. Peripherally, Lox6 is expressed principally in the primordia of the sensillae and in the eyes.
This pattern of expression of Lox6 suggests that one of its functions may be to contribute to the
diversification of neuronal phenotypes (Wong, 1998).
The Hox gene, Cnox-2, has been isolated from Hydractinia symbiolongicarpus, a
hydrozoan displaying division of labor. Cnox-2 is most closely related to anterior members of the Antennapedia gene
complex from Drosophila, with the greatest similarity to Deformed. Different patterns of aboral-to-oral Cnox-2 expression are found
among polyp polymorphs, and experimental conversion of one polyp type to another is
accompanied by concordant alteration in Cnox-2 expression. These results are consistent with the suggestion
that polyp polymorphism, characteristic of hydractiniid hydroids, arose via evolutionary modification of
proportioning of head to body column (Cartwright, 1999).
Expression patterns for five Hox genes were examined by whole-mount in situ hybridization in larvae of Chaetopterus, a
polychaete annelid with a tagmatized axial body plan. The polychaetes are generally thought to be the basal
taxon of the annelids and thus are a good group to sample
for developmental patterns more likely to represent ancestral
states than those of the more phylogenetically derived
leeches. Phylogenetic analysis demonstrates that these genes are orthologs of
the Drosophila genes labial, proboscipedia, zen, Deformed, and Sex combs reduced: they have been termed CH-Hox1, CH-Hox2,
CH-Hox3, CH-Hox4, and CH-Hox5, respectively. Expression studies reveal a biphasic expression pattern. In early larval
stages, well before any indications of segmental organization exist, a novel pattern of expression in bilateral posterior
proliferating cell populations, corresponding to the teloblasts, was detected for each of the genes, with CH-Hox1 and
CH-Hox2 expressed before the remaining three. In middle larval stages, all five genes are expressed in bilateral strips along
the ventral midline, corresponding with the developing ventral nerve cord. In addition, CH-Hox1 and CH-Hox2 show strong
expression at the foregut-midgut boundary. By late larval stages the expression is generally confined to the ventral CNS and
ectoderm of the anterior parapodia. Anterior boundaries of expression are 'colinear', at later larval stages, with CH-Hox2
expressed most rostrally, in the first segment, and anterior expression boundaries for CH-Hox1, CH-Hox3, CH-Hox4, and
CH-Hox5 in segments 2, 3, 4, and 5, respectively. Like vertebrates and spiders, but unlike insects, CH-Hox3 participates in
this colinear axial expression pattern. CH-Hox1 and CH-Hox2 have distinct posterior boundaries of expression in the ninth segment, which corresponds to a major morphological boundary, and may reflect a reorganization of Hox gene regulation
related to the evolutionary reorganization of the Chaetopterus body plan (Irvine, 2000).
The most striking aspect of Hox gene expression found in
this study is the early onset of expression in the posterior of
the larva. This early expression is localized to bilaterally
disposed cell populations located ventrolaterally, just anterior
to the pygidium. CH-Hox1 and CH-Hox2 are the first of
these genes to be expressed at detectable levels, exhibiting
robust in situ hybridization signals from early stage L1 (18
h). Expression of CH-Hox3, CH-Hox4, and CH-Hox5 is not
detected until stage L2 (some 24 h later), with an apparent
temporal gradient of expression onset from CH-Hox1 and 2
to CH-Hox3 and 4, with CH-Hox5 showing the latest
expression. The temporal sequence of Hox gene
expression is not an artifact, since expression of each of
these genes is detectable and persists in a bilaterally symmetrical
fashion in the prepygidial region throughout all
subsequent developmental stages. Although the expression
pattern follows the same sort of temporal sequence seen in
other metazoans, the expression of Hox genes in Chaetopterus
begins well before any indications of segmental organization
appear in the larval body plan (Irvine, 2000).
All five of the Hox genes examined are expressed at
these early stages in the same general region in the posterior
of the animal and may be turning on at nearly the same
times in many of the same cells. Expression initially occurs
in a largely overlapping domain and it is likely that Hox
gene transcription may be loosely regulated in early stages
to be refined later, either by translational regulation or by
differences in the timing of transcriptional downregulation
in a way similar to that seen in planarian flatworms (Irvine, 2000).
The general model of annelid development is based on
the presence of a 'posterior growth zone' just anterior to
the pygidium. The dynamics of cell division and posterior
expansion in this larval region are completely unknown:
various authors have claimed historically that both definitive
ectodermal and mesodermal stem cells reside in this
region and give rise to the various adult tissues of the
posttrochal region. The extreme version
of this type of teloblastic growth is seen in the
clitellates in which highly stereotyped sets of five bilaterally
symmetrical teloblast stem cells can be found and
named and their descendents studied in detail. The condition
in the clitellates is most often interpreted as an
acceleration of the formation of the adult body plan, as
these animals have forsaken larval development to become
direct developers.
The locations of cells expressing Hox gene transcripts
during early development in Chaetopterus coincide with
the predicted location of the prepygidial growth zone, both
in the anterior-posterior level and in the bilateral localization
of expressing cell populations. This largely overlapping
pattern of expression in terminal cell populations at early
stages has not been reported in other taxa, including those
with teloblastic growth, such as molluscs, crustaceans, and
most notably, a related class of annelids, the leeches. In
flies, for example, Antp transcripts are detected at the
cellular blastoderm stage, but are already localized to the
primordia of segments T1 and T2. In contrast,
Chaetopterus transcripts for each of the
genes examined are present in the putative growth
zone before any segmental primordia are formed. The very
early appearance of Hox gene expression in the putative
Chaetopterus growth zone may relate to the possibility of
early specification of segmental fate within teloblast lineages (Irvine, 2000).
Cell transplantation experiments in leeches have
shown that segmental founder cells derived from both
ectodermal and mesodermal teloblasts are determined at
the time of their birth. If
this early fate specification is true in polychaetes, it could
explain the presence of Hox genes, acting in a fate specification
role, in the set of segmental founder cells. The absence of
Hox gene expression in determined segmental founder cells
in leech embryos suggests that Hox gene expression is not
involved in the initial establishment of segmental identity
in annelids, and that the early pattern in polychaetes has
some other function, such as in cell proliferation. If the
early Hox expression detected in Chaetopterus does indeed
have a role in the precocious specification of segmental fate,
the lack of expression in leeches suggests that other, possibly
upstream, genes serve in place of the early phase of Hox
expression seen in Chaetopterus (Irvine, 2000).
The Tribolium castaneum (Tc)
ortholog of the Drosophila homeotic gene Deformed
(Dfd) has been examined and its expression pattern during embryogenesis in this beetle has been determined. Tc Dfd is expressed
in the blastoderm and the condensing germ rudiment
in a region that gives rise to gnathal segments. During
germ band extension Tc Dfd is expressed in the mandibular
and maxillary segments, their appendages, and
the dorsal ridge. Comparison of insect Dfd protein sequences
reveals several highly conserved regions. To determine
whether common molecular features reflect conserved
regulatory functions, the Gal4 system was used to
express the Tribolium protein in Drosophila embryos.
When Tc Dfd is expressed throughout embryonic ectoderm
under the control of P69B, the beetle protein auto-regulates
the endogenous Dfd gene. In addition, the Drosophila
proboscipedia gene (a normal target of Dfd) is
ectopically activated in the antennal and thoracic segments.
The ability of the beetle and fly
proteins to rescue defects in Dfd minus mutants were compared by expressing
each throughout the embryonic during embryogenesis.
Both proteins rescue Dfd minus defects to the same extent in
that they each restore the development of mouth hooks
and cirri, as well as cause gain-of-function abnormalities
of posterior mouth parts. As before, pb is ectopically
activated in the antennal segment. This is the first demonstration
of the ability of a heterologous homeotic selector protein to directly regulate a target gene independent of an endogenous Drosophila autoregulatory loop (Brown, 1999).
Among insects, the genetic regulation of regional identities in the
postoral head or gnathal segments (mandibular, maxillary, and labial)
is best understood in Drosophila. In part, normal gnathal development depends on Deformed (Dfd) and Sex combs reduced (Scr), genes in the
split Drosophila homeotic complex. The gnathal segments
of Dfd and Scr mutant larvae are abnormal
but not homeotically transformed. In the red flour beetle, Tribolium castaneum, a loss-of-function
mutation of the Deformed ortholog has been isolated. Mutant larvae
display a strong transformation of mandibular appendages to antennae.
The maxillary appendages, normally composed of an endite and a
telopodite, develop only the telopodite in mutant larvae. Mutations in the beetle Scr and Antennapedia orthologs are known to cause the labial and thoracic appendages, respectively, to be transformed to antennae. Moreover, a
deficiency of most of the beetle homeotic complex causes all gnathal
(as well as thoracic and abdominal) segments to develop antennae. These
and other observations are consistent with the hypothesis that
ancestral insect homeotic gene functions have been modified
considerably during the evolution of the highly specialized maggot
head. One of the ancestral homeobox genes that arose close to the root
of the Eumetazoa appears to have given rise to Dfd, Scr,
and the Antennapedia homeobox-class homeotic genes. Evidence from both Tribolium and Drosophila suggests that this ancestral gene served to repress anterior development as well as confer a trunk-specific identity (Brown, 2000).
Loss of TcDfd function results in a strong homeotic transformation
of the mandibular appendages to antennae. Clearly, unlike in flies,
TcDfd is important for repressing anterior development in
the mandibular segment. It is not clear whether TcDfd also
acts to promote mandibular identity in that segment. The role of
TcDfd in the beetle maxilla appears to parallel that of its
fly ortholog. Beetles resemble flies in having two domains of embryonic
maxillary Distal-less expression: one dorsal and one ventral. In each
insect, dorsal expression is Dfd-independent, whereas
ventral expression is Dfd dependent. Further, mutation in
each insect results in the loss of structures in the ventral region
(the endite in the case of Tribolium). It has been suggested that the cirri, ventral organ, and
mouth hooks are all homologous to structures (lacinia or galea) forming
parts of the maxillary endite of some other insects. It appears that
the Dfd orthologs of both insects act to promote ventral
maxillary identity. The observation that
Agpin, an apparent TcDfd gain-of-function mutation, causes ectopic appearance of the galea in adults further supports the conclusion that the gene promotes ventral maxillary identity in beetles, as it does in flies (Brown, 2000 and references therein).
The Drosophila ANTC includes five homeotic selector
genes. Probable null mutant phenotypes have now been identified for
Tribolium orthologs of four of these genes [to date no
Tc labial (Tclab) variant has been isolated]. In
each case, the beetle larval mutant phenotype differs significantly
from that described in flies. Most
significantly, mutants of the Antp, Scr, and Dfd
orthologs include transformations of thoracic, labial, and mandibular
appendages, respectively, to antennae [it is likely that the ancestral
pb gene function became highly derived before the origin of
the insects. Moreover, in homozygous condition a deficiency of
most of the complex results in an antennal transformation of all
gnathal, thoracic, and abdominal segments. Given that anterior
development in Tribolium embryos is far less derived than
that in Drosophila, it is likely that the functions of the beetle homeotic genes are more ancestral. The following model is presented, based on the information to date. The first eumetazoans had a
complex of three genes. Conventional thought considers these
to represent head, trunk, and tail genes, although it seems likely that
anterior-most development did not depend on the Hox genes. Duplication
and divergence of the central gene ultimately gave rise to
Dfd, Scr, and the Antp-class genes
(Antp, Ubx, and abd-A) before the
arthropod radiation. It is proposed that the ancestral trunk gene and
its derivatives played two roles: suppression of genes resulting in
anterior development and (probably in an evolutionarily labile fashion)
determination of specialized trunk segment features. As noted, the
beetle Antp, Scr, and Dfd orthologs all perform
the former function in at least part of their expression domains [some
evidence suggests that Drosophila homeotic genes play
similar embryonic roles. However, it appears
that the Tribolium Scr and Ubx orthologs no
longer have this function in the thorax. That is, loss of function of
the Antp ortholog is sufficient to transform the thoracic
segments to antenna despite the normal expression of the Scr
and Ubx orthologs in the anterior and posterior thorax,
respectively. No data from flies or other arthropods speak to a possible ancestral role of lab in repressing anterior development in the intercalary
segment; further speculation awaits the isolation of a Tclab mutation (Brown, 2000 and references therein).
Specialized insect mouthparts, such as those of Drosophila, are derived from an ancestral mandibulate state, but little is known about the developmental genetics of mandibulate mouthparts. The metamorphic patterning of mandibulate mouthparts of the beetle Tribolium castaneum was studied RNA interference to deplete the expression of 13 genes involved in mouthpart patterning. These data were used to test three hypotheses related to mouthpart development and evolution. First, the prediction was tested that maxillary and labial palps are patterned using conserved components of the leg-patterning network. This hypothesis was strongly supported: depletion of Distal-less and dachshund led to distal and intermediate deletions of these structures while depletion of homothorax led to homeotic transformation of the proximal maxilla and labium, joint formation required the action of Notch signaling components and odd-skipped paralogs, and distal growth and patterning required epidermal growth factor (EGF) signaling. Additionally, depletion of abrupt or pdm/nubbin caused fusions of palp segments. Second, the hypotheses was tested for how adult endites, the inner branches of the maxillary and labial appendages, are formed at metamorphosis. The data reveal that Distal-less, Notch signaling components, and odd-skipped paralogs, but not dachshund, are required for metamorphosis of the maxillary endites. Endite development thus requires components of the limb proximal-distal axis patterning and joint segmentation networks. Finally, adult mandible development is considered in light of the gnathobasic hypothesis. Interestingly, while EGF activity is required for distal, but not proximal, patterning of other appendages, it is required for normal metamorphic growth of the mandibles (Angelini, 2012).
In D. melanogaster, Dll mutants lack maxillary structures and portions of the proboscis (i.e., labium), although Dll expression in the maxillary anlagen is weaker than in the leg or antennal discs. Paralleling the results for T. castaneum, in the horned beetle Onthophagus taurus distal regions of the adult mouthparts were deleted with larval Dll RNAi (Simonnet 2011). The embryonic and metamorphic functions of Dll in T. castaneum are also similar: the gene is required for the development of distal structures at both stages, and during embryogenesis Dll is expressed throughout the developing palps. Interestingly, removal of T. castaneum Dll expression earlier during larval life led to delayed metamorphosis, as well as changes in appendage morphology (Suzuki, 2009). Many insects delay molting after appendage loss to allow time for regeneration, and this dual role of Dll suggests a mechanism linking these processes (Angelini, 2012).
The data from T. castaneum provide evidence for a conserved gap gene role of dac during patterning of mouthparts and legs of this species. dachshund is not expressed in or required for development of the labial and maxillary anlagen of D. melanogaster. In T. castaneum embryos dac is expressed strongly in the proximal maxilla and part of the developing endite. Embryonic dac expression is weaker in the distal maxillary palp and the labium. The current data show a clear metamorphic requirement for dac in the intermediate regions of the maxillary and labial palps, as does a recent study of O. taurus (Simonnet, 2011). A function for dac in the development of an intermediate portion of the maxillary and labial appendages has so far only been observed in these two beetles, while data from two species with specialized mouthparts (the milkweed bug O. fasciatus and D. melanogaster) found that dac is not required for PD patterning of the mouthparts. Thus, comparative data from other species do not support the hypothesis that this mouthpart patterning role is ancestral. However, if mandibulate mouthparts evolved from leg-like structures similarities in the expression and function of genes patterning both legs and mouthparts are expected to be plesiomorphic. This hypothesis can be further tested by examining the role of dac in mouthpart development in additional insect orders, particularly those that retain mandibulate mouthparts, and in other arthropods (Angelini, 2012).
The effects of hth depletion are distinct in different species, but typically involve some degree of homeotic transformation. In D. melanogaster, hth is expressed in the labial discs, but without nuclear expression of its cofactor Extradenticle. Maxillary palps are retained in hth loss-of-function flies, but they may possess bristles typical of legs, indicating a partial proboscis-to-leg transformation. In the cricket Gryllus bimaculatus, which has mandibulate mouthparts, hth depletion causes transformation of proximal mouthpart structures towards antennal identity, with a loss of endites, while distal structures are transformed towards leg identity (Ronco, 2008). hth RNAi in T. castaneum transformed intermediate regions of the maxilla and labium towards distal mouthpart identity. Proximal regions also appeared transformed, but their identity could not be established, while distal regions appeared wild type. In the beetle O. taurus, proximal regions of the labium are transformed towards maxillary endite identity, but distal regions of the labium and the entire maxilla remain relatively unaffected (Angelini, 2012).
These results highlight the similarity between patterning of the maxilla, labium and legs in T. castaneum. Functional data from two species with highly derived mouthpart morphologies, D. melanogaster and the milkweed bug Oncopeltus fasciatus, suggest only limited similarity between mouthpart and leg patterning. One explanation for this low degree of conservation is that evolution of the ancestral patterning mechanism has occurred in concert with the functional and morphological diversification of these mouthparts. A correlation between generative mechanisms and structural morphology has been used as a common null hypothesis, although exceptions in which similar morphologies result from different developmental pathways are documented. Nevertheless, this hypothesis predicts that developmental patterning should be more highly conserved across appendage types in species that retain the ancestral mandibulate mouthpart morphology (Angelini, 2012).
The maxillary and labial palps are an interesting case of serial homology. Despite a difference in overall size, their shape and arrangement of sensillae are similar. The intermediate segments of each palp type are also similar, but differ in number, which suggests that segment number is regulated independently from other morphological traits. The RNAi depletion of pdm in T. castaneum caused the reduction and deletion of the third maxillary palp segment, producing a phenotype closely resembling the wildtype morphology of the labial palps. While a role for pdm in the labium cannot be excluded, the absence of observed labial phenotypes was significant compared to maxillary results. Therefore, it is hypothesized that the difference in the number of palp segments results from specific activation of pdm in the maxillary palp. Loss of function in the Hox gene Deformed during T. castaneum embryogenesis causes a transformation of the larval maxillae towards labial identity. Since Hox genes are the primary determinants of body segment identity, it is proposed that pdm is activated by Deformed, and repressed by the labial Hox gene Sex combs reduced. RNAi targeting pdm in another mandibulate insect, the cricket Acheta domesticus, generated defects in the antenna and legs, but no defects in the mouthparts, despite similar pdm expression in these appendages (Turchyn; 2011; Angelini, 2012).
Endites are a primitive component of arthropod appendages, and they are retained in insect mouthparts, as well as in the mouthparts and thoracic appendages of many crustaceans (Boxshall 2004). At least three hypotheses have been put forward for how endites are patterned, and these hypotheses are not mutually exclusive. The first hypothesis states that multiple PD axes result from redeployment of a PD axis patterning mechanism shared by palps and endites. A second hypothesis posits that endites and appendage segments form by the same mechanism, Notch-mediated in-folding of the cuticle. A third hypothesis states that dac expression initiates endite branching from the main appendage axis. The axis redeployment hypothesis predicts that depletion of genes involved in PD axis patterning will have similar effects on the development of palps and endites. Some support for this hypothesis comes from studies of endite morphogenesis and the expression and function of leg gap genes in the embryos of T. castaneum and the orthopteran Schistocerca americana, but not all data are consistent with it. The segmentation hypothesis predicts that endites will fail to differentiate if genes required for joints are depleted. This hypothesis was posed based on a comparative developmental study of segmented and phyllopodous crustacean limbs. Finally, the dac-mediated hypothesis predicts that depletion of dac will lead to reduced endites. This hypothesis emerged from the observation that dac expression is reiterated along the medial edges of larval endites in the crustacean Triops longicaudatus. Comparative expression data from the isopod Porcellio scaber are also consistent with the dac-mediated hypothesis (Angelini, 2012).
The current data are consistent with predictions of the axis redeployment and segmentation hypotheses but do not support a role for dac in endite metamorphosis. Adult endites were disrupted by depletion of Dll, Krn, the odd-related genes, and Notch signaling, and to a lesser degree hth. In the maxilla depletion of most of these genes led to the failure of the single larval endite to divide into two distinct branches, while in the labium, their depletion caused reduction of the ligula. Their requirement in the endites is consistent with the hypothesis that these structures are generated by redeploying appendage PD axis determinants. Depletion of Notch signaling components and the odd paralogs produced reductions and fusions between palp segments, between the palps and endites, and between the lacinia and galea. Thus, these data are compatible with both the hypothesis that a reiterated PD axis is used to pattern the endites and the hypothesis that endite formation is linked to joint formation. Normal endite development in dac-depleted specimens is inconsistent with the dac-mediated hypothesis (Angelini, 2012).
It is noteworthy that endite specification and the division of the single larval endite into the adult galea and lacinia appear to be separable functions. For example, Ser RNAi resulted in a single endite lobe with lacinia identity medially and galea identity laterally. In contrast, severe Dll RNAi individuals had a single endite that lacked also obvious lacinia identity (Angelini, 2012).
The mandibulate structure of Tribolium mouthparts is the pleisomorphic state for insects and is shared by a majority of insect orders. These mouthparts are characterized by robust mandibles, lacking segmentation. A classic debate in arthropod morphology concerns whether the mandibles of insects and myriapods are derived from a whole appendage or only from proximal appendage regions; the latter are called gnathobasic mandibles. Palps are retained on the mandibles of many crustaceans, making it clear that the biting regions of their mandibles are gnathobasic. Phylogenetic support for the gnathobasic hypothesis comes from phlyogenetic studies that place insects nested within crustaceans (Regier, 2010). The first developmental genetic support for the gnathobasic hypothesis came from the discovery that insect mandibles lack Dll expression. Furthermore, neither mutations in Dll nor its depletion through RNAi have been observed to alter mandible development in insects, including T. castaneum. This evidence has led to widespread acceptance of the gnathobasic hypothesis. Of the 13 genes depleted in this study, two (Krn and hth) produced results that would not be predicted by the most straightforward form of the gnathobasic hypothesis for mandible origins (Angelini, 2012).
Loss of EGF function in insects leads to distal appendage defects, including pretarsal or tarsal deletions. The role of EGF signaling in distal appendage regions is conserved in T. castaneum metamorphosis, since depletion of the EGF ligand Krn leads to reduction of the antennal flagellum, and maxillary and labial palps, as well as to deletion of the pretarsus and malformation of the tarsus. In light of the restriction of Krns role to distal appendage regions and regulation of distal EGF ligand expression by Dll in D. melanogaster, the gnathobasic hypothesis predicts that Krn should not be required for normal development of the mandible in T. castaneum. In contrast to this prediction, depletion of Krn produced a significant reduction in mandible length (Angelini, 2012).
The hypothesis of a gnathobasic mandible also predicts that hth depletion should produce effects in the mandible similar to those in the proximal regions of other appendage types. In T. castaneum, hth RNAi during metamorphosis caused homeotic transformation of proximal regions of the maxilla, labium and legs. However, the mandibles were not affected by hth depletion. In the beetle O. taurus, hth depletion slightly altered mandible shape, but also without apparent homeosis. In contrast, hth RNAi in embryos of the cricket G. bimaculatus transformed the mandible towards a leg-like structure distally and an antenna-like structure proximally, paralleling the transformation observed in other appendages. Because these results come from only two lineages and from different life stages, additional data are needed to determine whether a homeotic role for hth was present ancestrally in insect mandibles (Angelini, 2012).
These data must be weighed alongside other evidence bearing on the gnathobasic hypothesis. In T. castaneum, the lack of phenotypic effects on mandible metamorphosis of other genes in this study is consistent with the gnathobasic hypothesis. In particular, it was observed that mandible metamorphosis was normal following depletion of genes involved in distal growth and patterning or joint formation. Moreover, homology at one biological level, such as anatomy, does not preclude divergence at other levels, such as development. Nevertheless, since developmental genetic studies of Dll and other appendage-patterning genes have been used as strong support for the gnathobasic homology of the insect mandible, the findings of Krn function highlight the difficulties in establishing serial homology based solely on developmental data (Angelini, 2012).
This study provides a genetic model of adult mouthpart development in Tribolium castaneum based on 13 genes. While previous studies have examined patterning in species with derived mouthpart morphologies, T. castaneum retains the pleisomorphic, mandibulate state of insect mouthparts. These results demonstrate the conservation of many gene functions in the maxilla and labium, relative to the legs, thus supporting the interpretation of novel gene functions in groups with derived mouthpart morphology as indicative of their specialized morphogenetic roles in those species. Mandibulate mouthparts such as those of T. castaneum include medial maxillary and labial endites, and the current data are consistent with hypotheses of reiteration in the PD axis and specification by Notch signaling, but rule out a direct role for dac in branch generation or patterning at metamorphosis. Additionally these results demonstrate that a regulator of distal leg development, Krn, which encodes an EGF ligand, is required for normal mandible elongation. This finding underscores the complex relationship between homology at the levels of anatomy and developmental patterning (Angelini, 2012).
Hox genes are known to control the identity of
serially repeated structures in arthropods and vertebrates.
The expression pattern of the Hox genes Deformed, Sex combs reduced, Antennapedia, and Ultrabithorax/abdominal-A (Ubx/abd-A)
from the honey bee Apis mellifera were analyzed. A
cDNA with the complete coding region of the Antennapedia
gene from Apis was cloned. Comparison with Antp proteins
from other insect species revealed several regions of homology.
The expression patterns of the isolated Hox
genes from Apis show that the original expression patterns
of Dfd, Scr, and Antp appear between late blastoderm
and early germ band stage in a temporal and spatial
sequence. Each of them shows up as a belt, spanning approximately
two segment anlagen; Dfd in the anterior
gnathal region, Scr in the posterior gnathal and anterior
thoracic region, and Antp in the thoracic region. Following
expansion of the Antp domain in the abdomen as a
gradient towards the posterior, Ubx/abd-A expression appears
laterally in the abdomen. During gastrulation and
in the germ band stage the domains of strong expression
do not overlap any more, but touch each other. After gastrulation
the borders of the expression domains partly
correlate with parasegment and partly with segment
boundaries. Laterally, gaps between the domain of each
gene may show no expression of any of the genes examined (Walldorf, 2000).
In the blastoderm stage the expression domains of all
four genes are similar in fruit fly and honey bee embryos.
Certainly these genes play very similar roles in establishing
the general character of that part of the insect
body where they have their main expression domain.
Even in Chelicerata the orthologs show comparable
patterns. However, some differences should be mentioned. In the
fly, the borders of the Dfd domain are segmental. In
the bee, the anterior expression border is initially parasegmental
in the rudiment of the intercalary segment: this changes from segmental to parasegmental during
gastrulation in the ventral part of the maxillary segment. The steady Engrailed pattern shows that this change is not a result of cell movement. The same development of that pattern is reported for Tribolium. This is most
clearly seen ventrally in the parasegmental Scr pattern in
the germ band of the bee, corresponding to segmental in
that area in the fly. While some differences in the Dfd pattern are
consistently observed between the bee and the fly, the
complicated posterior borders of the Scr and the Antp
patterns of young and old embryos of flies and bees are identical. The
same Antp pattern is also found in Thermobia domestica. However, in Manduca sexta the
anterior border of Antp is the compartment border of T1. It may be that some of these differences are only the result of different sensitivity levels of
the stainings. Probably the ventral switch from segmental
to parasegmental is more general in insects but is not
seen very clearly in some cases, such as in the anterior
part of the fruit fly head, due to the subsequent head involution.
The resulting ventrolateral corners of the expression
borders of Dfd, Scr, and Antp correlate fairly
well with the position of the appendage anlagen and may
help to pin down their position (Walldorf, 2000).
cDNA fragments of the homologs of the Drosophila
head homeotic genes labial (lab),
proboscipedia (pb), and
Deformed (Dfd) have been isolated from
the crustacean Porcellio scaber. P. scaber (order Isopoda) belongs to the subclass Malacostraca (higher crustaceans); relative to phylogenetically more basal groups, it is as derived as insects are in its body plan and
tagmatization. Moreover, the interpretations of the expression patterns reported here are based on the
assumptions that: (1) the Insecta is monophyletic, with the order Thysanura a basal group; (2) the Mandibulata are monophyletic with the Crustacea, a sister
group of the Insecta, and (3) the Chelicerata (spiders and mites) is an outgroup in relation to the Insectan-Crustacean clade.
Because the
accumulation domains of the head homeotic complex (Hox) genes had not
been previously reported for crustaceans, the expression
patterns of these genes was examined in P. scaber embryos by using in situ hybridization. The P. scaber lab
homolog is expressed in the developing second antennal segment and
its appendages. This expression domain in crustaceans and in the
homologous intercalary segment of insects suggests that the
lab gene specified this metamere in the last common
ancestor of these two groups. The expression domain of the P.
scaber pb gene is in the posterior part of the second antennal
segment. This domain, in contrast to that in insects, is colinear with
the domains of other head genes in P. scaber; it
differs from the insect pb gene expression domain in the posterior mouthparts, suggesting that the insect and crustacean patterns evolved independently from a broader ancestral domain similar
to that found in modern chelicerates. P. scaber Dfd is expressed in the mandibular segment and paragnaths (a pair of ventral
mouthpart structures associated with the stomodeum) and differs from
insects, where expression is in the mandibular and maxillary segments.
Thus, like pb, Dfd shows a divergent Hox
gene deployment. It is concluded that homologous structures of the
mandibulate head display striking differences in their underlying
developmental programs related to Hox gene expression (Abzhanov, 1999).
To assign the
P. scaber head appendages to specific segments, the monoclonal antibody Mab4D9, which
recognizes EN, was used to indicate the posterior of the segmental borders. The antibody revealed six segments in the
embryonic head of P. scaber: ocular, first antennal, second antennal, mandibular, first maxillary, and second maxillary. The most anterior region of the head, the labrum, develops as a pair of small appendage-like structures that
fuse medially at the stage when development is about 65%-70% complete. In early embryos, the EN stripe of the ocular segment is not
complete and is interrupted on the ventral side by the labrum. The labrum itself does not express EN and, in this respect, appears to be continuous with the
stomodeum. The first antennal segment bears a pair of small uniramose antennae, which are reduced in the adult. The second antennae are the largest pair of
the appendages on the head. The stomodeal opening protrudes at the level of the posterior first antennal (a1) segment and extends to the posterior of the
second antennal (a2) segment: this results in a ventral interruption of the EN stripes of the a1 and a2 segments. The broad mandibular EN band is seen in
the developing posterior mandibular appendages. EN is expressed in a similar fashion in the first and second maxillary segments (Abzhanov, 1999).
In D. melanogaster, the expression domain of Dfd
includes the mandibular and maxillary segments and appendages. It has
been shown to be required genetically for the normal development of both mandibles and maxillae in Drosophila. Dfd expression has been found to be very similar in
all insect groups studied, including the basal insect T. domestica, and the mandibles and maxillae of insects are homologous to the mandibles and first maxillae (maxillulae) of
crustaceans. Comparison of the Dfd expression domains in P. scaber and insects reveals that the crustacean domain is smaller. P. scaber Dfd is
expressed strongly in the paragnaths and the mandibular segment, but
not the mandibular appendages. The paragnaths are associated with the
stomodeum, but their exact embryonic origin is obscure. Some authors
have concluded that these structures are sternal protrusions of the
mandibular segment associated with the mouth, reduced appendages
associated with the mandibles, or even structures homologous to the
insect hypopharynx. Whatever their allegiance, paragnaths
are found in a diversity of crustaceans. Thus, P. scaber
Dfd is not expressed in the mandibles or maxillae (only mesodermal expression is detected in mandibles), where Dfd function is
required in insects, suggesting that Dfd has a different
developmental function in the crustacean head. It is not known which,
if any, selector gene is expressed in the ectoderm of the crustacean
mandibles proper (Abzhanov, 1999).
A
comparison of the crustacean/insect and chelicerate (spiders and mites) patterns of Hox
gene expression is made difficult by the uncertainty of segmental
homologies between the two groups. The more traditional view, based on
anatomy and patterns of innervation, concludes that the segment
associated with the deuterocerebral ganglion of the central nervous
system is greatly reduced or absent in modern chelicerates. If
this conclusion is correct, the more anterior ocular/protocerebral
segment would be homologous to the same segment in insects and
crustaceans, whereas the next posterior segment in the spider would
correspond to the intercalary/tritocerebral segment of insects and to
the second antennal segment of crustaceans. Thus, the homolog of the
insect antennal and the crustacean first antennal segment would be
absent in chelicerates. More recently, this question of
head-segment homology has been revisited by using the patterns of Hox
gene expression as a basis for determining the presence or absence of
the deuterocerebral segment in chelicerates. Using the spider Cupiennius salei, the mite
Archegozetes longisetosus, and several head Hox genes as
probes, it has been concluded that this segment is present.In chelicerates, lab
is expressed anteriorly in the developing pedipalps (homologous to the
appendages of the intercalary segment). This expression extends
posteriorly, to the fourth pair of walking limbs: this location corresponds
to the first pair of the thoracic appendages in insects and crustaceans
(maxillipeds in P. scaber). Therefore, the anterior
boundary of lab expression appears to be conserved among
chelicerates, crustaceans, and insects. It is notable, however, that
the posterior limit of expression is not conserved. The integumentary
expression of lab is limited to the intercalary or second
antennal segment in insects and crustaceans, respectively, whereas in
chelicerates, expression extends posteriorly a further four full
segments. In A. longisetosus, pb is expressed in a broad
domain from the pedipalps to the third pair of walking legs, where it
is accumulated in the appendages. This boundary is colinear with
both lab and Dfd, and is thus similar to the
relative expression domains of these genes in vertebrates, annelids,
and Porcellio; but it is dissimilar to that seen in
insects. Thus, the anterior boundary of pb expression in
the a2 segment of P. scaber appears to resemble that seen in
chelicerates rather than that in insects. However, it should also be
noted that, as for lab, the expression domain of chelicerate
pb extends further to the posterior than that in insects and
crustaceans (Abzhanov, 1999).
Dfd expression has been examined in both C. salei
and A. longisetosus. In C. salei, Dfd
is expressed in all walking legs. The mite shows a similar
expression domain in the L1-L4 legs, with additional accumulation
covering all opistosomal (abdominal) segments except the most terminal
ones. The anterior boundary of Dfd accumulation appears
to be conserved and located in the mandibular (insects) and homologous
L1 (chelicerates) segments. If one assigns the
paragnaths to the mandibular segment, the anterior boundary of
Dfd expression then appears to be similar, albeit not
identical, in P. scaber vis-a-vis insects and chelicerates. However, both the chelicerate and insect Dfd domains are
clearly broader than those seen in the crustacean (Abzhanov, 1999).
The extended and broadly overlapping expression domains in
chelicerates are reminiscent of those in vertebrates and are probably
closer to an ancestral state. In contrast, the expression domains in
insects and crustaceans are more resolved and segment-specific. Based on morphological and recent molecular evidence, the Crustacea
belongs to the monophyletic group Mandibulata, which is a close sister
group to the Insecta. Thus, crustaceans represent an ideal
case for study of the evolution of the homologous head Hox gene
expression patterns and possible functions in the homologous structures
of insects (Abzhanov, 1999).
Comparison of the expression patterns of the crustacean and insect Hox
genes demonstrates that there is conservation of segment affinity
(e.g., lab) and spatial colinearity (e.g., lab,
pb, Dfd, and Scr) of expression. In addition, the anterior boundaries
of the lab and Dfd genes appear to be conserved
in insects, crustaceans, and chelicerates. However, there is also
divergence of the observed expression domains (e.g., pb and
Dfd). Consequently, substantial
variation in the deployment of the Hox genes, and presumably in the
developmental processes regulated by them, can be seen in homologous
and morphologically similar crustacean and insect head structures.
Genes involved in the development of mandibles and posterior mouthparts
in insects are expressed in novel, though still colinear domains. For
example, in insects, the maxillary and labial mouthparts express
pb, whereas in P. scaber, the homologous
appendages both express and probably depend on Scr, a
different head homeotic gene (Abzhanov, 1999).
It is hypothesized that the mandibulate head evolved prior to the
establishment of the defined head Hox gene expression domains which
have been recruited independently to their current regions and developmental functions in crustaceans and insects. This model involves an intermediate hypothetical
mandibulate ancestor that did not have segment-specific expression
domains and probably resembled the pattern of expression seen in modern
chelicerates. The specification of individual segments and mouthparts
in such an animal would depend on the redundant and/or fractional
functions of multiple Hox genes, and would be facilitated by the
subsequent evolution of more distinct expression domains. That is, the head Hox genes would have functioned in a manner analogous to the
genes of the D. melanogaster Bithorax complex. To test this model and to better understand the evolution of the Hox genes and head
structures, further studies across different crustacean and myriapod
groups will be required (Abzhanov, 1999).
Insects have evolved a large variety of specialized feeding
strategies, with a corresponding variability in mouthpart
morphology. In the Hemiptera, the mandibular and
maxillary segments give rise to two similar pairs of long, thin
stylets. The paired maxillary appendages form
channels for liquid flow and the piercing mandibles lie on
either side. These four interlocked stylets run down a groove
in the long, fused labium, which provides support. Although
in typical mandibulate insects the maxillary and labial
appendages are very similar, in the Hemiptera, it is
the mandibular and maxillary appendages that share a highly
unusual morphology very different from the labium.
These specialized mouthparts represent an important
evolutionary innovation that allows this order of insects to
feed by extracting fluids from other organisms.
There is little understanding of the
developmental mechanisms that underlie mouthpart diversity.
Until recently it was difficult to perform any analysis of
gene function outside of the genetic model insects
Drosophila melanogaster and Tribolium castaneum. In this
paper, the use of dsRNA-mediated interference
(RNAi) is described to dissect gene function in the development of the
milkweed bug Oncopeltus fasciatus, which has specialized
suctorial mouthparts. The Hox genes Deformed (Dfd),
proboscipedia (pb) and Sex combs reduced (Scr) have
previously been shown to be expressed in the gnathal
appendages of this species. Strikingly, the milkweed bug
was found to have an unusual expression pattern of pb.
Here, by analyzing single and combination RNAi
depletions, it has been found that Dfd, pb and Scr are used in the
milkweed bug to specify the identity of the mouthparts. The
exact roles of the genes, however, are different from what
is known in the two genetic model insects. The maxillary
appendages in the bug are determined by the activities of
the genes Dfd and Scr, rather than Dfd and pb as in the fly and beetle. The mandibular appendages are specified by
Dfd, but their unique morphology in Oncopeltus suggests
that Dfd's target genes are different. As in flies and beetles,
the labium is specified by the combined activities of pb and
Scr, but again, the function of pb appears to be different.
Additionally, the regulatory control of pb by the other two
genes seems to be different in the bug than in either of the
other species. These novelties in Hox function, expression
pattern and regulatory relationships may have been
important for the evolution of the unique Hemipteran head (Hughes, 2000).
Dfd is the sole gene responsible for mandibular identity. This conforms with the expression of Dfd in the mandibular segment. The single depletion of Dfd transforms the mandibular appendages to distal antennal identity, rather than the long thin stylets normally formed. Thus Dfd is necessary for proper mandibular development. In contrast, the depletion of pb leaves the mandibular appendages untouched. Thus pb is not necessary for mandibular development. The phenotype of the Scr depletion is more difficult to interpret, since the mandibular stylets generally fail to grow, and are merely short bristles in a mass of undifferentiated tissue. Since Scr is not expressed in the mandibular segment, it is suspected that this is an indirect effect of the Scr phenotype on other segments of the head. Because head development is integrated to some degree, non-local, indirect effects are often seen in Drosophila Hox mutants, particularly labial and Deformed. In the case of milkweed bug Scr, disruption of the proximal labium may be interfering with the normal development of the adjacent mandibular and maxillary appendages. The Dfd Scr double depletion corroborates this view. The mandibles in the double form distal antennae, indistinguishable from either the Dfd depletion alone or the triple. Therefore it can be inferred that, when present, Scr is not acting in the mandibular appendage to directly specify any identity over that of the default state (Hughes, 2000).
The inferred activity of Dfd, but not pb and Scr, in the mandibular appendages matches the predictions based on expression patterns. While Dfd is expressed strongly in the mandibular appendages, pb and Scr are not. The role of Dfd as the sole Hox gene regulating mandibular identity also matches the situation in Tribolium, where Dfd mutants transform Mn structures, while maxillopedia (mxp, the pb homolog) and Cephalothorax (Cx, the Scr homolog) mutants leave the Mn unaffected. The effect of Hox mutations on the reduced mandibular structures of Drosophila, however, is more difficult to determine. In the embryo, Dfd mutations disrupt the dorsolateral papillae of the terminal sense organ, which are thought to derive from the Mn segment. In the Drosophila adult, Dfd mutations disrupt parts of the head capsule. Mutations in pb and Scr do not appear to affect these presumed mandibular structures. Although both beetles and milkweed bugs use Dfd to specify the mandibular segment, the resulting appendages are very different. In contrast to the chewing mandibles of the beetle, the bug mandibles are very thin stylets nearly as long as the body. Again, these are very different from the mandibular appendages in Drosophila, which are internal structures in the embryo, and are either missing or incorporated into the head capsule of the adult. Thus it is concluded that while Dfd's basic role in the mandible may be conserved, the developmental module driven by Dfd is extremely labile (Hughes, 2000).
In the maxillary segment, Dfd depletion results in only a partial transformation of the maxilla to antennal identity. However, the Dfd;Scr double depletion results in complete transformation of the maxillae to antennae. Thus it can be concluded that Dfd acts in concert with Scr in the maxillary segment. The curled phenotype of the Dfd depletion maxillary appendage suggests that Scr may be repressing growth of the transformed limb in its posterior domain; this repression is released in the Dfd;Scr double depletion. The phenotypes in the maxillary segment are somewhat in conflict however, regarding the role of pb. Two results suggest that pb is not acting to specify maxillary identity. (1) Depletion of pb alone leaves the maxillary segment unaffected, therefore pb is not necessary for wild-type maxillary development. (2) The phenotype of the Dfd;Scr double is the same as the triple depletion (i.e., antennae), so it can be inferred that pb is not acting to confer any identity over that of the default. There is, however, a subtle difference between the phenotypes of the Dfd single and Dfd;pb double depletion. While the Dfd depletion produces short, curled antenna-like appendages, the Dfd;pb double produces straight antenna-like appendages, often with pretarsal claws. This suggests that in the absence of Dfd, pb can affect the maxillary appendages. Whether this activity derives from its small dorsal maxillary domain of accumulation, or from an expanded domain of pb expression in the Dfd depletion, is not yet clear. Nevertheless, based on the pb depletion, it is concluded that, in wild-type embryos, pb is not active in the specification of the maxillary stylet (Hughes, 2000).
This lack of pb function in determining the maxillary stylet matches the prediction made based on the expression pattern of pb in the bug. Thus far this is the only insect known not to use pb to determine maxillary identity. In Drosophila and Tribolium, pb/mxp mutations cause transformation of the maxillary palps, which become reduced in Drosophila, and transform to legs in Tribolium. In the bug, it would appear that Scr assumes much of this role. This is in contrast to Drosophila or Tribolium, where the Scr/Cx mutation does not affect the maxillary appendage. While Dfd and Scr work together to specify maxillary identity in the bug, it seems that Dfd has the principal role. Perhaps by activating a similar set of target genes as in the mandible, Dfd may induce a similar stylet identity in the maxillary appendage. This contrasts with mandibulate insects, in which the maxillary morphology is most similar to the labium, probably due to the activity of pb in both of those appendages (Hughes, 2000).
Since the triple depletion animals should lack all Hox activity in the gnathal segments, the default state of these appendages in the absence of Hox function can be inferred. Antennae have been thought to represent the default appendage state in the absence of Hox activity, since a large deletion of much of the Hom-C in the beetle was found to produce a larva bearing a series of antennae instead of mouthparts and legs. Likewise, in the adult fly, the pb and Scr double mutant combination in the adult labium gives rise to antennae. Triple depletion indicates that, in the milkweed bug as well, antennae represent the no-Hox state for the gnathal appendages. In the triple depletion, the maxillary and labial segments form full-length antennae. Since wild-type antennae are the only paired appendages that lack Hox expression, it is reasonable that this morphology is produced in the absence of Hox function. Furthermore, this result indicates that the genetic program that specifies the presence of an appendage is independent of Hox expression, and that the function of the Hox gene products is to confer a specific morphology onto a generic appendage program. In the absence of Hox activity, this generic appendage turns on the antennal differentiation pathway (Hughes, 2000).
In the absence of Hox activity, the mandibular appendages, however, each form only a distal antenna on a short base. This suggests that a portion of the generic appendage is missing. Previous work has suggested that insect mandibles, which lack Distal-less (Dll) expression, are gnathobasic (lack distal portions of the appendage). In Drosophila and Tribolium, Dll is repressed by Dfd in the mandibular segment. In the milkweed bug, therefore, depletion of Dfd is likely associated with Dll derepression. Consistent with this hypothesis, the morphology of the transformed mandibular appendage is primarily to distal antenna. Interestingly, as noted, the basal portions of the appendage are distorted or missing, compared to the maxillary or labial default appendages. This suggests that some aspects of the reduced structure of the insect mandible are independent of the Dfd/Dll pathway (Hughes, 2000).
In this study, evidence has been found of remodeling in the role of Hox genes in head appendage development. This change in the role of the Hox genes is due to three mechanisms: (1) change in expression patterns; (2) change in regulatory relationships, and (3) change in function. Previous work had shown that pb possesses a unique expression pattern in the milkweed bug. The RNAi analysis confirms that the unusual expression pattern does in fact reflect an unusual domain of activity. Unlike Drosophila, Tribolium and probably most other insects, pb is not necessary for development of the maxillary appendages in the milkweed bug. The pb gene is also regulated differently in the bug, where it appears to be independent of the activity of Dfd and Scr. This contrasts strongly with the regulatory interactions described for the fly and the beetle. Studies of more distant insects are needed, however, before the ancestral state and the direction of change for these regulatory interactions can be confidently inferred. Lastly, evidence was found of novel functions in the bug that probably reflects differences in the suite of target genes activated by the Hox genes. For example, the function of pb is different in the bug, where it induces a mixed leg-like identity rather than a maxillary palp. Even in the mandibular segment, where Dfd is the conserved controller of identity, the resulting mandibular appendage is strikingly different from the mandibles of other insects, indicating a very different developmental module at work downstream of the Hox signal. It is not known, however, whether the repertoire of target genes is different, whether the orthologous target genes themselves have different functions or if both possibilities are correct (Hughes, 2000).
During Drosophila segmentation, gap genes function as short-range
gradients that determine the boundaries of pair-rule stripes. A classical
example is Drosophila Krüppel (Dm'Kr) which is
expressed in the middle of the syncytial blastoderm embryo. Patterning defects
in Dm'Kr mutants are centred symmetrically around its bell-shaped
expression profile. The role of Krüppel was examined in the
short-germ beetle Tribolium castaneum where the pair-rule stripes
corresponding to the 10 abdominal segments arise during growth stages
subsequent to the blastoderm. The previously described mutation
jaws is an amorphic Tc'Kr allele. Pair-rule gene expression
in the blastoderm is affected neither in the amorphic mutant nor in
Tc'Kr RNAi embryos. Only during subsequent growth of the germ band
does pair-rule patterning become disrupted. However, only segments arising
posterior to the Tc'Kr expression domain are affected, i.e., the
deletion profile is asymmetric relative to the expression domain. Moreover,
stripe formation does not recover in posterior abdominal segments, i.e., the
Tc'Krjaws phenotype does not constitute a gap in segment
formation but results from a breakdown of segmentation past the 5th
eve stripe. Alteration of pair-rule gene expression in
Tc'Krjaws mutants does not suggest a direct role of
Tc'Kr in defining specific stripe boundaries as in
Drosophila. Together, these findings show that the segmentation
function of Krüppel in this short-germ insect is fundamentally
different from its role in the long-germ embryo of Drosophila. The
role of Tc'Kr in Hox gene regulation, however, is in better
accordance to the Drosophila paradigm (Cerny, 2005).
The most obvious differences between the phenotypes of Krüppel
in Tribolium and Drosophila are the homeotic transformations
in Tc'Krjaws and Tc'Kr RNAi larvae that are not
evident in Dm'Kr mutants. Such transformations are not entirely
unexpected given that in Drosophila the expression boundaries of Hox
genes are also set by gap genes, including Dm'Kr. However, in
Drosophila gap mutants all segments that would be transformed because
of misregulation of homeotic genes usually also suffer segmentation defects
and fail to develop. By contrast, Tribolium segment primordia
anterior of, and within, the Krüppel expression domain do
differentiate, such that homeotic transformations can manifest themselves in
the differentiated larva (Cerny, 2005).
The expression of homeotic genes in Tc'Krjaws embryos
is consistent with the morphological transformations observed. The results with
Tc'Dfd, Tc'Scr, Tc'Antp and Tc'Ubx confirm and extend
earlier findings for Tc'pb and Tc'UBX/Tc'ABD-A expression.
Notably, the complementary double-segmental expression of Dfd and
Scr in Tc'Krjaws embryos explains the phenotype
of alternating maxillary and labial segments. These expression
patterns indicate that the posterior limit of Tc'Dfd and
Tc'Scr domains is set through inhibition by Tc'Kr. In this
respect, Tc'Kr fulfils a function similar to Drosophila gap
genes (Cerny, 2005).
The homeotic phenotype of Tc'gt RNAi embryos
could suggest a similar function in Hox regulation for Tc'gt. Indeed
Tc'Antp anteriorly expands and gnathal Hox genes
(Tc'Scr) repress in Tc'gt RNAi embryos, consistent with
the expansion of thoracic fates found in differentiated Tc'gt RNAi
larvae. These transformations are just opposite to those of
Tc'Krjaws larvae. Interestingly, in embryos that lack
Tc'Kr and at the same time have reduced Tc'gt activity, the
homeotic effect of Tc'Krjaws clearly is epistatic. This shows that the
ectopic Tc'gt stripes in the Tc'Kr mutant do not contribute
to the Tc'Kr phenotype. However, this experiment suggests that the
homeotic transformation of gnathal segments into thorax in Tc'gt RNAi
embryos is indeed an indirect effect and comes about through misregulation of
Tc'Kr in these embryos. This interpretation is supported by the finding that the Tc'Kr expression domain expands anteriorly in
Tc'gt RNAi embryos. Evidently, it is
expansion of Tc'Kr that results in repression of gnathal Hox genes in
maxilla and labium of Tc'gt RNAi embryos, not loss of gnathal Hox
gene activation. Similarly, expansion of Tc'Antp in Tc'gt
RNAi larvae could be due to activation by anteriorly expanded Tc'Kr.
However, as Antp is not significantly reduced in
Tc'Krjaws, it seems more likely that Tc'gt acts
directly to define the anterior boundary of the Tc'Antp domain (Cerny, 2005).
In addition to gap gene input, Drosophila Hox genes also receive
input from pair-rule genes. The near-pair-rule pattern of Tc'Dfd and
Tc'Scr in Tc'Krjaws embryos reveals an important
role of pair-rule genes also in defining Tribolium Hox domain
boundaries. It seems likely that regulation of Tc'Dfd and
Tc'Scr by pair-rule genes is responsible for the precision of their
expression boundaries in wild-type Tribolium embryos, while input
from gap genes defines the broad region where a particular Hox gene can become
active (Cerny, 2005).
In Drosophila, Krüppel is expressed in a bell-shaped profile
centered over the primordia of segments T2 to A3. In
the Tribolium blastoderm, only one such gradient is present; the
Tc'Kr domain covers the posterior pole. When
the germ rudiment has formed, the Tc'Kr domain retracts from the
posterior end and forms a distinct domain overlapping the three thoracic
segment primordia. At
this stage, therefore, the Tc'Kr domain covers more anterior segment
primordia (and more anterior pair-rule stripes) than does its
Drosophila counterpart (Cerny, 2005).
The results for Tc'Kr deviate from those obtained for
Krüppel in Oncopeltus fasciatus. In
this short-germ insect, knock-down of Kr also results in
mis-expression of Hox genes, although the effects are more limited as only one
ectopic Of'Dfd domain is detected. Interestingly, expression of
Of'en in such embryos seems to indicate a clear gap phenotype, i.e.
perfect segmental stripes reappear posterior to a region of segmental
disruption. Incomplete inactivation of Of'Kr could be responsible for
this difference; it is noted, however, that weak Tc'Kr RNAi situations do
not result in obvious gap phenotypes. Rather, in such embryos the segmentation process simply breaks down somewhat later than in Tc'Krjaws, i.e., the additional segments present in weak Tc'Kr RNAi embryos appear to represent anterior abdominal rather than posterior (post-gap) abdominal segments.
Oncopeltus is sometimes denoted an intermediate-germ insect, because
a few more segments are formed already in the blastoderm than, for example, in
Tribolium. It will be interesting to see if the 'next posterior' gap
gene in Oncopeltus will also display a 'gap' phenotype, and to find
out whether pair-rule gene expression in Of'Kr RNAi embryos indicates
a role in the regulation of specific stripes boundaries (Cerny, 2005).
Hox genes play a fundamental role in the establishment of
chordate body plan, especially in the anteroposterior
patterning of the nervous system. Particularly interesting
are the anterior groups of Hox genes (Hox1-Hox4) since
their expression is coupled to the control of regional
identity in the anterior regions of the nervous system,
where the highest structural diversity is observed.
Ascidians (phylum Urochordata) are considered a good model to investigate evolution of Hox gene organization,
regulation and function. CiHox3, a Ciona intestinalis anterior
Hox gene homologous to the paralogy group 3 genes has been cloned and the
expression pattern characterized. In situ
hybridization at the larva stage reveals that CiHox3
expression is restricted to the visceral ganglion of the
central nervous system. The presence of a sharp posterior
boundary and the absence of transcript in mesodermal
tissues are distinctive features of CiHox3 expression when
compared to the paralogy group 3 in other chordates. The regulatory elements underlying
CiHox3 neural-specific expression have been characterized and, using transgenic
analysis, an 80 bp enhancer
responsible for CiHox3 activation in the central nervous
system (CNS) has been isolated. A comparative study between mouse and
Ciona Hox3 promoters demonstrates that divergent
mechanisms are involved in the regulation of these genes in
vertebrates and ascidians. Mouse Hoxa3 and Hoxb3 rhombomere-specific regulatory
elements tested in Ciona embryos were unable to reproduce
any neural-specific expression. Conversely, the smallest fragment that
drives nervous system expression in Ciona was negative in
mice (Locascio, 1999).
Continued: Deformed Evolutionary homologs part 2/3
|part 3/3
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Deformed :
Biological Overview
| Regulation
| Protein Interactions
| Developmental Biology
| Effects of Mutation
| References
Society for Developmental Biology's Web server.
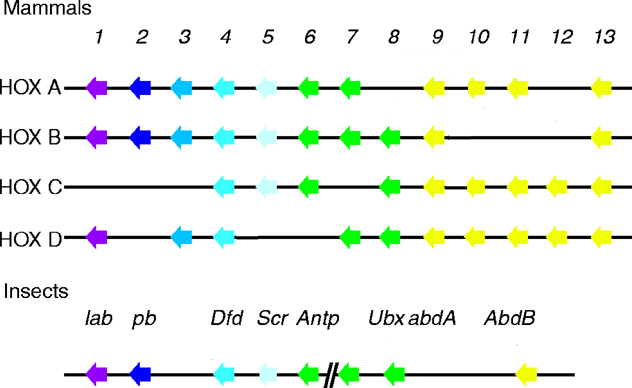{kind=link}