labial
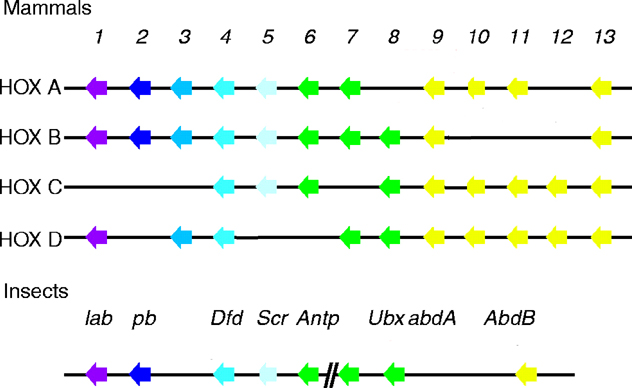
The labial homeodomain constitutes an independent family within the Antennapedia class of homeodomain proteins (see four paralogous Hox clusters of mammals). Drosophila Proboscipedia is a part of this family, as well as human Hoxa-1, Hoxa-2 and C. elegans ceh-13 (Gehring, 1994 and Frasch, 1995).
For information about Hox Clusters and Networks see the Homeobox Genes DataBase
The conservation of developmental functions exerted by Antp-class homeoproteins in protostomes and deuterostomes has suggested that homologs with related functions are present in diploblastic animals, in particular, in Hydra. Phylogenetic analyses show that Antp-class homeodomains belong either to non-Hox or to Hox/paraHox families. See Phylogenetic relationships among 200 Antp-class genes. Among the 13 non-Hox families, 9 reported here have diploblastic homologs: Msx, Emx, Barx, Evx, Tlx, NK-2, and Prh/Hex, Not, and Dlx. Among the Hox/paraHox, poriferan sequences are not found, and the cnidarian sequences form at least five distinct cnox families. Cnox-1 shows some affinity to paralogous group (PG) 1; this group includes Drosophila Labial. Cnox-2 is related to Drosophila Intermediate neuroblast defective. Cnox-3 and 5 show some affinity to PG9-10; this group includes Drosophila AbominalB. Cnox-4 has no counterparts in Drosophila or vertebrates. Intermediate Hox/paraHox genes (PG 3 to 8 and lox) do not have clear cnidarian counterparts. In Hydra, cnox-1, cnox-2, and cnox-3 are not found chromosomally linked within a 150-kb range and display specific expression patterns in the adult head. During regeneration, cnox-1 is expressed as an early gene whatever the polarity, whereas cnox-2 is up-regulated later during head but not foot regeneration. Finally, cnox-3 expression is reestablished in the adult head once the head is fully formed. These results suggest that the Hydra genes related to anterior Hox/paraHox genes are involved at different stages of apical differentiation. However, the positional information defining the oral/aboral axis in Hydra cannot be correlated strictly to that characterizing the anterior-posterior axis in vertebrates or arthropods (Gauchat, 2000)
Clusters of homeobox-containing HOM-C/hox genes determine the morphology of animal body plans and body parts and are thought to mediate positional information. A description is presented of the onset of embryonic expression of ceh-13, the Caenorhabditis elegans ortholog of the Drosophila labial gene, which is the earliest gene of the C. elegans Hox gene cluster to be activated in C. elegans development. At the beginning of gastrulation, ceh-13 is asymmetrically expressed in posterior daughters of anteroposterior divisions, first in the posterior daughter of the intestinal precursor cell E and then in all posterior daughters of the AB descendants ABxxx. Evidence is presented that supports position-independent activation of ceh-13 during early C. elegans embryogenesis, which integrates cell fate determinants and cell polarity cues. These findings imply that mechanisms other than cell-extrinsic anteroposterior positional signals play an important role in the activation and regulation of the C. elegans Hox gene ceh-13. During the first few embryonic divisions, differentially localized maternal gene products account for the various developmental potentials found in the first few embryonic cells. The data show that at least two of these proteins, SKN-1 and POP-1, influence ceh-13 expression in Ep (posterior progeny of E) and may function as cell-specific or cell-lineage-specific modulators of ceh-13 activation (Wittmann, 1997).
The C. elegans lin-39, mab-5 and egl-5 Hox genes, although not tightly clustered as are the Drosophila Hox genes, specify cell fates along the anterior-posterior body axis of the nematode during postembryonic development, but little is known about Hox gene functions during embryogenesis. The C. elegans labial-like gene, ceh-13, is expressed in cells of many different tissues and lineages and the rostral boundary of its expression domain is anterior to those of the other Hox genes. The CEH-13 protein shows structural features typical for labial class proteins, including the organization of exons and introns and the conservation of the homeodomain, as well as the presence of a conserved hexapeptide. By transposon-mediated mutagenesis, a zygotic recessive ceh-13 loss-of-function allele (sw1) was isolated that exhibits an embryonic sublethal phenotype. Lineage analyses and immunostainings reveal defects in the organization of the anterior lateral epidermis and anterior body wall muscle cells. The epidermal and mesodermal identity of these cells, however, is correctly specified. ceh-13(sw1) mutant embryos also show fusion and adhesion defects in ectodermal cells. This suggests that ceh-13 plays a role in the anterior organization of the C. elegans embryo and is involved in the regulation of cell affinities (Brunschwig, 1999).
ceh-13 is first expressed at the onset of gastrulation in the posterior daughters of the intestinal precursor cell E (Ep) and posterior daughters of AB descendants ABxxx (ABxxxp). The ceh-13 endodermal expression is maintained in all Ep descendants for at least two more cell divisions before fading out during morphogenesis. No staining in the intestine could be detected in larval and adult stages. During embryogenesis, in addition to the E lineage, ceh-13 is detected in many different cell types of AB, MS and D lineages. At the comma stage, ceh-13 is strongly expressed in the lateral hypodermal (epidermal) cells H2 and V1 and in cells of the prospective ventral nerve cord. Signals in anterior dorsal hypodermal cells as well as in some anterior body wall muscle cells are also observed. lin-39, mab-5 and egl-5 have been shown to be expressed in successive domains along the anterior-posterior body axis of 1.5-fold-stage embryos. At this stage, ceh-13 is mainly expressed anterior to the expression domain of lin-39. In L1 larvae, the expression of CEH-13 in the different ventral nerve cord cells and the lateral and dorsal hypodermal cells shows the same anterior boundary as in the comma-stage embryos, while the most anterior H2 lineage appears much more weakly stained. Thus, throughout development, the ceh-13 rostral boundary of expression is located more anterior than the domains of expression of the other members of the Hox cluster. However, CEH-13- expressing cells are not limited to the anterior part of C. elegans, but are found all along the body axis and in the male tail (Brunschwig, 1999).
Homozygous ceh-13(sw1) animals exhibit a recessive variable abnormal morphology phenotype characterized by incompletely penetrant zygotic lethality. On average, about 97% of the ceh-13(sw1) homozygous animals arrest during embryogenesis or at early larval stages. Rare ceh-13(sw1) homozygotes that survive to adulthood (approximately 3%) show less severe morphogenetic defects than the arrested ceh-13 animals, but are smaller and develop as much as 6 times slower than wild-type animals. In wild-type C. elegans, when most of the embryonic cell divisions are completed, changes of cell shape and adhesion transform the ellipsoidal embryo into a cylindrical worm through the elongation process. ceh-13(sw1) mutants appear to be morphologically normal until the beginning of elongation. At this stage, mutant embryos start to elongate variably to some extent before again retracting. This results in very short animals with anterior, and occasionally more posterior, protuberances. In the most extreme mutant phenotypes, the hypodermis ruptures and the inner cells spill out of the embryo. In addition, ceh-13(sw1) mutants exhibit movement defects. Soon after the onset of elongation, wild-type embryos begin to twitch inside the eggshell; this twitching movement increases when animals reach the two-fold stage. ceh-13 animals, by contrast, either continue to twitch weakly or even remain paralyzed. Interestingly, the ceh-13(sw1) escapers move forward but are unable to move backward properly. This latter defect suggests that some ceh-13 activity is required in neurons since the same muscles are used for forward and backward movements. The putative affected neurons have not yet been identified (Brunschwig, 1999).
ceh-13 mutant embryos show adhesion defects but no lineage transformation. To determine whether the morphogenetic defects in ceh-13(sw1) mutants could be caused by lineage transformation, lineage analyses were performed on four mutant embryos by using a four-dimensional microscope and analysis software. No significant alterations in the division pattern up to the bean stage are noticed in the ceh-13(sw1) embryos, as compared to the naturally occurring variability observed in wild-type embryos. Furthermore, as determined by morphological criteria, the identity of all cells appear to be correctly specified in the mutants. However, subtle deviations could be observed in the positions of nuclei when reconstructed wild-type and ceh-13 mutant embryos were compared at the premorphogenetic stage. Moreover, in three out of four lineaged ceh-13 mutant embryos, individual cells lost contact with the rest of the embryo at the comma stage. In two cases, neuronal precursor cells detached from the embryos. In another ceh-13(sw1) mutant embryo, two hypodermal descendants were observed that partially lost contact with the embryo, leading to a rupture in the anterior hypodermis as the elongation proceeded. Altogether, these observations suggest that, while ceh-13 has no effect on the specification of the cell lineage, it is required for proper cell-cell adhesion in the embryo (Brunschwig, 1999).
Anterior epidermal and mesodermal cells are mislocalized in ceh-13 mutant embryos. In wild-type embryos, immediately after ventral body enclosure, the first fusions of dorsal hypodermal cells are detected anterior and posterior to the deirid sensillae, to form the hyp7 syncytium. At the same time, constrictions of circumferential filament bundles in the hypodermis squeeze the embryo into its final thin vermiform shape. In addition to the hypodermis, it has been shown that body wall muscles also play a crucial role in this morphogenetic transformation. In order to understand the basis of the severe morphogenetic defects in ceh-13(sw1) homozygotes, the organization of the hypodermal and muscle cells were examined in mutant animals. By using an antibody that stains the hypodermal adherens junctions, and another antibody that is specific for all non-neuronal ectodermal nuclei, it was shown that the fate of all hypodermal cells were correctly specified. This result is in agreement with the lineage analyses. However, lineage analyses and immunostainings reveal that cells are mislocalized in ceh-13(sw1) mutant embryos. In wild-type embryos, the seam cells form lateral rows, ten cells wide, arrayed in two groups of five cells (H0, H1, H2, V1-6 and T) one group on either side of the body along the anterior-posterior axis. In ceh-13(sw1) mutant embryos, from the bean stage onward, a mislocalization of V1, and in most cases also of H1 and H2, has been observed. These defects correlate with the strong expression of ceh-13 in H2 and V1. Moreover, at least one hypodermal cell remains unfused with the dorsal hypodermal syncytium in the ceh-13(sw1) mutant. In two-fold-stage mutant embryos, this cell is still unfused. This defective pattern of fusion is a typical feature of ceh-13(sw1) mutant embryos. Since ceh-13 expression is found in embryonic body wall muscles, the organization of these mesodermal cells was examined in mutant embryos. During the elongation process, the body wall muscle cells are spread longitudinally beneath the hypodermis in two dorsal and two ventral rows, or quadrants. Immunofluorescence staining of ceh-13(sw1) mutant embryos with the monoclonal antibody anti-myosin heavy chain A, mAb5-6, indicates that terminal mesodermal fate is achieved. The anterior body wall muscles are, however, disorganized in ceh-13(sw1) mutant embryos. At the bean stage in ceh-13(sw1) mutant embryos on which lineage analyses were performed, muscle cells of the D lineage are mislocalized in the region surrounding the seam cell V1. However, unlike the anterior seam cells, which are reproducibly mislocalized, the identity of the mispositioned body wall muscle cells differs in the various lineaged embryos (Brunschwig, 1999).
In contrast to the labial-like genes in other species, ceh-13 mutants do not appear to alter pattern formation or the fate of restricted sets of cells. Nonetheless, similarly to ceh-13, some genes of the labial class have also been suggested to control cell-cell interactions. The analysis of hindbrain in Hoxa-1 -/- mutant mice reveals that the remnants of hindbrain constrictions, termed rhombomeres 4 and 5, appear to be fused caudally with rhombomere 6 to form a single fourth rhombomeric structure, suggesting abnormal cellular adhesion at the rhombomere boundaries. Finally, in Drosophila labial mutants, the incomplete head involution appears to be due to a failure of the mandibular and maxillary lobes to fuse with the procephalic lobe, as well as the failure of cells to assimilate into the dorsal pouch (Brunschwig, 1999 and references).
C. elegans contains a set of six cluster-type homeobox (Hox) genes that are required during larval development. Some of them, (but unlike in flies, not all of them) are also required during embryogenesis. It has been suggested that the control of the embryonic expression of the worm Hox genes might differ from that of other species by being regulated in a lineal rather than a regional mode. Here, a trans-species analysis of the cis-regulatory region of ceh-13, the worm ortholog of the Drosophila labial and the vertebrate Hox1 genes has been performed; the molecular mechanisms that regulate its expression may be similar to what has been found in species that follow a regulative, non-cell-autonomous mode of development. Two enhancer fragments have been identified that are involved in different aspects of the embryonic ceh-13 expression pattern. Important features of comma-stage expression depend on an autoregulatory input that requires ceh-13 and ceh-20 functions. The data show that the molecular nature of Hox1 class gene autoregulation has been conserved among worms, flies, and vertebrates. The second regulatory sequence is sufficient to drive correct early embryonic expression of ceh-13. Interestingly, this enhancer fragment acts as a response element of the Wnt/WG signaling pathway in Drosophila embryos (Streit, 2002).
pMF1. containing 8.1 kb of upstream sequences as well as the first exon, the first
intron, and most of the second exon, mimics endogenous
ceh-13 expression in transgenic animals.
Comparative sequence analysis of pMF1 led to the identification
of a 10-bp-long sequence motif (TGATGGATGG)
in enh450 (starting at nucleotide position 26483 relative to
the ATG start codon of ceh-13) that is identical to that of
the HOXB1/PBX autoregulatory element of the mouse gene
Hoxb1 and differs by only one base pair substitution from that of the
LAB/EXD autoregulatory element of the Drosophila gene
labial. In order to assess the importance
of the 10-bp element and to test whether this sequence
represents an autoregulatory element similar to the
one described for the mouse and the fly, a
mutational analysis was performed in the context of the enh450 (a fragment able to drive most aspects of the normal ceh-13 in the expression pattern of comma-stage embryos). In Drosophila
and in mice, it has been shown that the activity of
the element is sensitive to point mutations in the core
binding sites of the LAB(HOXB1) and EXD(PBX) proteins. In analogy to
these findings, two separate 1-bp substitutions were intoduced in the 10-bp worm element that were expected to
disrupt the potential binding sites for CEH-13 and for a
putative worm ortholog of EXD/PBX. In the double-mutant construct mutE/L,
the point mutations in the two half sites of the element
were combined. The results of this single point mutation
analysis confirmed that the 10-bp sequence element is
absolutely required for the function of enh450 and strongly
suggest that it functions as a direct autoregulatory element
of ceh-13 (Streit, 2002).
The second enhancer element that has been identified in
the ceh-13 promoter region is a 740-bp fragment (enh740)
located at the downstream end of enh3.4. enh740 drives
early embryonic expression of a reporter gene in C. elegans
in a pattern indistinguishable from ceh-13. This fragment
does not contain candidate sequences for the kind of auto-regulatory
element discussed above. Nevertheless, it is able
to induce LacZ expression in groups of cells of all germ
layers in the Drosophila embryo around stage 15. Interestingly,
enh740 is activated by and dependent on Wg signaling
and its nuclear effector Pan in all three germ layers and therefore acts as a strong Wg sensor that shows little or no tissue specificity. This is in contrast to the Wg-dependent Drosophila enhancer elements characterized so far that are all specific for particular tissues (Streit, 2002).
Expression of lab also depends on the Wg signaling
pathway. In the embryonic midgut of Drosophila, low Wg
levels stimulate its transcription and that of another Hox
gene (Ubx), whereas high levels repress both genes. Analysis
of the Ubx midgut enhancer reveals that activation and
repression are mediated by different enhancer elements.
Whereas Wg-mediated transcriptional activation is conferred directly by binding of Pangolin (the Drosophila
LEF-1/TCF homolog) and its coactivator Armadillo, the
Drosophila ß-catenin equivalent, it
appears that repression in response to high Wg levels is
indirect and based on transcriptional activation of the
Teashirt (Tsh) repressor. enh740 may
contain only Wg activator elements but lacks Tsh repressor
sequences and therefore acts as an activator even at high Wg
concentrations. This may also explain why the expression
domain of enh740 was shifted toward the posterior compared
to endogenous Lab. Further biochemical and genetic
analyses are required to determine whether Pangolin binds
directly to the 740-bp element and how it activates reporter
gene expression (Streit, 2002).
The finding that enh740 acts as a strong Wg response
element in Drosophila raises the challenging question of
whether expression of enh740 in C. elegans also depends on
Wnt/Wg signaling. Indeed, genetic studies in C. elegans
have revealed that Wnt/Wg signaling acts very early during
nematode development. In four-cell-stage embryos, for example,
MOM-2/Wnt/Wg signaling specifies the production of endoderm.
Furthermore, several C. elegans Wnt/Wg pathway members
are involved in a embryo-wide polarity system that
generates differences in cell fate between many sister cells
generated by a/p divisions. The ß-catenin/Armadillo-related
protein WRM-1 and LIT-1, a serine/threonine protein
kinase homologous to the Drosophila tissue polarity protein
Nemo, are effectors of this signaling pathway. WRM-1
binds and activates LIT-1, which in turn phosphorylates
POP-1 and causes a reduction of its apparent nuclear levels in the posterior daughters of a/p dividing cells. In the AB lineage, for example, POP-1 asymmetry is present at each of several sequential a/p divisions and is first established during the a/p divisions of the ABxx cells (Streit, 2002).
Because of this a/p polarity system, POP-1 is also asymmetrically
distributed after the fourth AB division that
leads to the formation of the ceh-13 expressing ABxxxp.
Therefore, and given the fact that POP-1 has been proposed to
function as a transcriptional repressor in this pathway, it is tempting to speculate that high POP-1 levels may negatively regulate ceh-13 expression in the ABxxxa cells, the anterior sisters of the ABxxxp cells.
This view is supported by the complete absence of ceh-13::
gfp reporter gene expression in the AB lineage of lit-1
mutant animals, that lack POP-1 asymmetry and show high levels of POP-1 immunostaining in the nuclei of all ABxxxx cells. Surprisingly, however, a complete
absence of pMF1 reporter gene expression is found in all ABxxxx
cells of pop-1 (RNAi) animals that lack maternal and
embryonic POP-1 expression. Thus, it is possible
that POP-1, at low nuclear concentrations or upon phosphorylation
by LIT-1, may act as a transcriptional activator
of ceh-13. However, since mutations in pop-1 and lit-1
result in cell fate changes and start to act earlier in development than ceh-13 is
first expressed, other explanations for these results can not
be excluded. An argument for a direct involvement of
POP-1 in control of ceh-13 is the fact that enh740 does
contain several putative POP-1 binding sites and at least
one of them does bind POP-1 in vitro. Altogether, it is
thought that WNT signaling, or more specifically POP-1, is
very likely to be one of the spatiotemporal cues involved in
control of ceh-13 expression in the early embryo. However,
early embryonic ceh-13/enh740 expression cannot depend
on Wg mediated polarity cues only: POP-1 asymmetry in
the AB lineage is established already in the daughters of the
ABxx cells, but onset of ceh-13 expression occurs only one
cell division later. Regardless of the POP-1 distribution,
ceh-13 continues to be expressed in both the anterior and
posterior daughters of almost all of the ABxxxp cells. Furthermore, ceh-13 is not expressed in
the cells of the early C lineage. Thus, it is proposed that, in
addition to Wnt/Wg-mediated polarity cues, other yet
unidentified factors participate in the regulation of ceh-13/
enh740 early expression and provide the cells with temporal
and lineage-specific inputs. Further work will be required
to confirm the involvement of the worm Wnt/Wg
pathway and its nuclear effector POP-1 on enh740 and to
identify other regulatory proteins that bind to it (Streit, 2002).
Earlier experiments have suggested that the pattern of
expression of the worm Hox genes are determined, at least
in part, by mechanisms independent of the global position
of the cells along the a/p axis. This is in contrast to Drosophila
and vertebrates, where the Hox genes are controlled by
global positional mechanisms. Based on these observations,
it has been suggested that Hox gene regulation in C. elegans
may rely on different strategies than those previously
characterized in Drosophila and mammalians. Nevertheless,
it has been show in this study that the molecular nature of the mechanisms
that control Hox 1 class genes may be more conserved
between flies and worms than previously assumed (Streit, 2002).
Hox genes are transcriptional regulators of metazoan body regionalization along the anteroposterior axis that act by specifying positional identity in differentiating cells. ceh-13, the labial ortholog in C. elegans, is expressed both during embryogenesis and post- embryonic development. Using GFP reporter analysis and immunocytochemistry, a spatiotemporal pattern of gene expression was discovered in the male tail during the L3 and L4 larval stages that is TGF-ß pathway-dependent. Analysis of reporter activity in transgenic animals identified a distinct promoter region driving male tail-specific ceh-13 expression. The conservation between Caenorhabditis species of sequence motifs within this region is reported and it is speculated that, in the course of evolutionary diversification, ceh-13 may have acquired new functionality while conserving its homeotic role (Stoyanov, 2003).
Expression patterns for five Hox genes were examined by whole-mount in situ hybridization in larvae of Chaetopterus, a polychaete annelid with a tagmatized axial body plan. The polychaetes are generally thought to be the basal taxon of the annelids and thus are a good group to sample for developmental patterns more likely to represent ancestral states than those of the more phylogenetically derived leeches. Phylogenetic analysis demonstrates that these genes are orthologs of the Drosophila genes labial, proboscipedia, zen, Deformed, and Sex combs reduced: they have been termed CH-Hox1, CH-Hox2, CH-Hox3, CH-Hox4, and CH-Hox5, respectively. Expression studies reveal a biphasic expression pattern. In early larval stages, well before any indications of segmental organization exist, a novel pattern of expression in bilateral posterior proliferating cell populations, corresponding to the teloblasts, was detected for each of the genes, with CH-Hox1 and CH-Hox2 expressed before the remaining three. In middle larval stages, all five genes are expressed in bilateral strips along the ventral midline, corresponding with the developing ventral nerve cord. In addition, CH-Hox1 and CH-Hox2 show strong expression at the foregut-midgut boundary. By late larval stages the expression is generally confined to the ventral CNS and ectoderm of the anterior parapodia. Anterior boundaries of expression are 'colinear', at later larval stages, with CH-Hox2 expressed most rostrally, in the first segment, and anterior expression boundaries for CH-Hox1, CH-Hox3, CH-Hox4, and CH-Hox5 in segments 2, 3, 4, and 5, respectively. Like vertebrates and spiders, but unlike insects, CH-Hox3 participates in this colinear axial expression pattern. CH-Hox1 and CH-Hox2 have distinct posterior boundaries of expression in the ninth segment, which corresponds to a major morphological boundary, and may reflect a reorganization of Hox gene regulation related to the evolutionary reorganization of the Chaetopterus body plan (Irvine, 2000).
The most striking aspect of Hox gene expression found in this study is the early onset of expression in the posterior of the larva. This early expression is localized to bilaterally disposed cell populations located ventrolaterally, just anterior to the pygidium. CH-Hox1 and CH-Hox2 are the first of these genes to be expressed at detectable levels, exhibiting robust in situ hybridization signals from early stage L1 (18 h). Expression of CH-Hox3, CH-Hox4, and CH-Hox5 is not detected until stage L2 (some 24 h later), with an apparent temporal gradient of expression onset from CH-Hox1 and 2 to CH-Hox3 and 4, with CH-Hox5 showing the latest expression. The temporal sequence of Hox gene expression is not an artifact, since expression of each of these genes is detectable and persists in a bilaterally symmetrical fashion in the prepygidial region throughout all subsequent developmental stages. Although the expression pattern follows the same sort of temporal sequence seen in other metazoans, the expression of Hox genes in Chaetopterus begins well before any indications of segmental organization appear in the larval body plan (Irvine, 2000).
All five of the Hox genes examined are expressed at these early stages in the same general region in the posterior of the animal and may be turning on at nearly the same times in many of the same cells. Expression initially occurs in a largely overlapping domain and it is likely that Hox gene transcription may be loosely regulated in early stages to be refined later, either by translational regulation or by differences in the timing of transcriptional downregulation in a way similar to that seen in planarian flatworms (Irvine, 2000).
The general model of annelid development is based on the presence of a 'posterior growth zone' just anterior to the pygidium. The dynamics of cell division and posterior expansion in this larval region are completely unknown: various authors have claimed historically that both definitive ectodermal and mesodermal stem cells reside in this region and give rise to the various adult tissues of the posttrochal region. The extreme version of this type of teloblastic growth is seen in the clitellates in which highly stereotyped sets of five bilaterally symmetrical teloblast stem cells can be found and named and their descendents studied in detail. The condition in the clitellates is most often interpreted as an acceleration of the formation of the adult body plan, as these animals have forsaken larval development to become direct developers. The locations of cells expressing Hox gene transcripts during early development in Chaetopterus coincide with the predicted location of the prepygidial growth zone, both in the anterior-posterior level and in the bilateral localization of expressing cell populations. This largely overlapping pattern of expression in terminal cell populations at early stages has not been reported in other taxa, including those with teloblastic growth, such as molluscs, crustaceans, and most notably, a related class of annelids, the leeches. In flies, for example, Antp transcripts are detected at the cellular blastoderm stage, but are already localized to the primordia of segments T1 and T2. In contrast, Chaetopterus transcripts for each of the genes examined are present in the putative growth zone before any segmental primordia are formed. The very early appearance of Hox gene expression in the putative Chaetopterus growth zone may relate to the possibility of early specification of segmental fate within teloblast lineages (Irvine, 2000).
Cell transplantation experiments in leeches have shown that segmental founder cells derived from both ectodermal and mesodermal teloblasts are determined at the time of their birth. If this early fate specification is true in polychaetes, it could explain the presence of Hox genes, acting in a fate specification role, in the set of segmental founder cells. The absence of Hox gene expression in determined segmental founder cells in leech embryos suggests that Hox gene expression is not involved in the initial establishment of segmental identity in annelids, and that the early pattern in polychaetes has some other function, such as in cell proliferation. If the early Hox expression detected in Chaetopterus does indeed have a role in the precocious specification of segmental fate, the lack of expression in leeches suggests that other, possibly upstream, genes serve in place of the early phase of Hox expression seen in Chaetopterus (Irvine, 2000).
In the freshwater planarian Dugesia japonica, five cDNAs for HOM/HOX homeobox genes, (Plox or planarian HOM/HOX homeobox genes) were cloned and sequenced. Together with sequence data on platyhelminthes HOM/HOX homeobox genes that have been deposited in databases, comparison of the deduced amino acid sequences reveals that planarians have at least seven HOM/HOX homeobox genes, Plox1 to Plox7. Plox1, Plox3 and Plox7 show high similarity to the fly genes labial (82% amino acid identity in the homeodomain), Dfd (82%) and abd-a (88%), respectively. Plox4 may correspond to Sex combs reduced (Scr) rather than Dfd (75%), because of Plox4's high similarity with nematode mab5 (67%), leech Lox5-hro (77%), and paralog group 4 in vertebrates; planarian chich corresponds to Scr in the fly. Plox2 could not be classified. Both Plox5 and Plox6 resemble Antp in the fly. Whole-mount in situ hybridization and RT-PCR reveal that Plox4 (72% homologous to Sex combs reduced) and Plox5 (77% homologous to Scr) are increasingly expressed along a spatial gradient in the posterior region of intact animals. During regeneration, Plox5 is expressed only in the posterior region of regenerating body pieces, suggesting that the gene is involved in the anteroposterior patterning in planarians. Plox5 is not found to be expressed in a blastema-specific manner. X-ray irradiation experiments show that at a minimum, Plox5 is expressed in some cells other than neoblasts, but that the induction of Plox5 expression during regeneration might require neoblasts. Chromosome walking data are needed to understand the genomic organization and relationships of planarian HOM/HOX homeobox genes to those of other animal groups (Orii, 1999).
Molecular developmental studies of fly and mouse embryos have shown that the identity of individual body segments is controlled by a suite of homeobox-containing genes called the Hox cluster. To examine the conservation of this patterning mechanism in other segmented phyla, four Hox gene homologs isolated from glossiphoniid leeches of the genus Helobdella are described. Based on sequence similarity and phylogenetic analysis, the leech genes Lox7, Lox6, Lox20, and Lox5 are deemed to be orthologs of the Drosophila genes lab, Dfd, Scr, and Antp, respectively. Sequence similarities between Lox5 and Antp outside the homeodomain and phylogenetic reconstructions suggest that the Antennapedia family of Hox genes had already expanded to include at least two discrete Antp and Ubx/abdA precursors prior to the annelid/arthropod divergence. In situ hybridization reveals that the four Lox genes described in this study are all expressed at high levels within the segmented portion of the central nervous system (CNS), with variable levels of expression in the segmental mesoderm. Little or no expression is seen in peripheral ectoderm or endoderm, or in the unsegmented head region (prostomium). Each Lox gene has a distinct anterior expression boundary within one of the four rostral segments, and the anterior-posterior (AP) order of these expression boundaries is identical to that reported for the orthologous Hox gene products in fly and mouse. This finding supports the idea that the process of AP axis differentiation is conserved among the higher metazoan phyla with respect to the regional expression of individual Hox genes along that axis. One unusual feature of leech Hox genes is the observation that some genes are only expressed during later development -- beginning at the time of terminal cell differentiation -- whereas others begin expression at a much earlier stage; their RNA ceases to be detectable shortly after the onset of expression of the 'late' Hox genes. The functional significance of this temporal disparity is unknown, but it is noteworthy that only the two 'early' Hox genes display high levels of mesodermal expression (Kourakis, 1997).
cDNA fragments of the homologs of the Drosophila head homeotic genes labial (lab), proboscipedia (pb), and Deformed (Dfd) have been isolated from the crustacean Porcellio scaber. P. scaber (order Isopoda) belongs to the subclass Malacostraca (higher crustaceans); relative to phylogenetically more basal groups, it is as derived as insects are in its body plan and tagmatization. Moreover, the interpretations of the expression patterns reported here are based on the assumptions that: (1) the Insecta is monophyletic, with the order Thysanura a basal group; (2) the Mandibulata are monophyletic with the Crustacea, a sister group of the Insecta, and (3) the Chelicerata (spiders and mites) is an outgroup in relation to the Insectan-Crustacean clade. Because the accumulation domains of the head homeotic complex (Hox) genes had not been previously reported for crustaceans, the expression patterns of these genes was examined in P. scaber embryos by using in situ hybridization. The P. scaber lab homolog is expressed in the developing second antennal segment and its appendages. This expression domain in crustaceans and in the homologous intercalary segment of insects suggests that the lab gene specified this metamere in the last common ancestor of these two groups. The expression domain of the P. scaber pb gene is in the posterior part of the second antennal segment. This domain, in contrast to that in insects, is colinear with the domains of other head genes in P. scaber; it differs from the insect pb gene expression domain in the posterior mouthparts, suggesting that the insect and crustacean patterns evolved independently from a broader ancestral domain similar to that found in modern chelicerates. P. scaber Dfd is expressed in the mandibular segment and paragnaths (a pair of ventral mouthpart structures associated with the stomodeum) and differs from insects, where expression is in the mandibular and maxillary segments. Thus, like pb, Dfd shows a divergent Hox gene deployment. It is concluded that homologous structures of the mandibulate head display striking differences in their underlying developmental programs related to Hox gene expression (Abzhanov, 1999).
To assign the P. scaber head appendages to specific segments, the monoclonal antibody Mab4D9, which recognizes EN, was used to indicate the posterior of the segmental borders. The antibody revealed six segments in the embryonic head of P. scaber: ocular, first antennal, second antennal, mandibular, first maxillary, and second maxillary. The most anterior region of the head, the labrum, develops as a pair of small appendage-like structures that fuse medially at the stage when development is about 65%-70% complete. In early embryos, the EN stripe of the ocular segment is not complete and is interrupted on the ventral side by the labrum. The labrum itself does not express EN and, in this respect, appears to be continuous with the stomodeum. The first antennal segment bears a pair of small uniramose antennae, which are reduced in the adult. The second antennae are the largest pair of the appendages on the head. The stomodeal opening protrudes at the level of the posterior first antennal (a1) segment and extends to the posterior of the second antennal (a2) segment: this results in a ventral interruption of the EN stripes of the a1 and a2 segments. The broad mandibular EN band is seen in the developing posterior mandibular appendages. EN is expressed in a similar fashion in the first and second maxillary segments (Abzhanov, 1999).
lab expression in the Drosophila head is in the intercalary segment, a small metamere that is devoid of appendages, located posterior to the antennal segment and anterior to the mandibular segment. The exact role of lab in this segment is unclear, because mutants do not show an obvious homeotic transformation. Nevertheless, the gene is important for the formation of the embryonic and adult head in Drosophila, because mutants do show defects in the development of cephalic structures. lab expression in the intercalary segment is conserved in all of the insect species examined thus far. Comparison of the P. scaber lab expression domain to that of insects reveals both conservation and change. P. scaber expression is restricted to the first postantennular metamere, i.e., the a2 segment, which, based on morphological and molecular (EN expression) data, is thought to be homologous to the intercalary. Moreover, innervation of this first postantennular segment is associated with the tritocerebral ganglion of the brain in all mandibulates. The fact that lab is expressed in a homologous segment in insects and crustaceans suggests that it was recruited for and might be involved in conserved and homologous developmental processes. However, in Porcellio, lab is expressed in the a2 appendages, whereas the adult insect intercalary segment is limbless. Interestingly, the embryos of some insects develop small transitory appendages on this segment, which might be atavisms suggestive to a more primitive state. It is not clear, however, whether lab is directly involved in appendage loss in the insects; mutations in this gene do not cause limb growth from the intercalary. Nevertheless, it is possible that in crustaceans, lab contributes to the unique morphology of the second, as compared with the first, pair of antennae. For example, in P. scaber, the first antennae are greatly reduced in size, whereas the second antennae are large and more leg-like (Abzhanov, 1999).
A comparison of the crustacean/insect and chelicerate (spiders and mites) patterns of Hox gene expression is made difficult by the uncertainty of segmental homologies between the two groups. The more traditional view, based on anatomy and patterns of innervation, concludes that the segment associated with the deuterocerebral ganglion of the central nervous system is greatly reduced or absent in modern chelicerates. If this conclusion is correct, the more anterior ocular/protocerebral segment would be homologous to the same segment in insects and crustaceans, whereas the next posterior segment in the spider would correspond to the intercalary/tritocerebral segment of insects and to the second antennal segment of crustaceans. Thus, the homolog of the insect antennal and the crustacean first antennal segment would be absent in chelicerates. More recently, this question of head-segment homology has been revisited by using the patterns of Hox gene expression as a basis for determining the presence or absence of the deuterocerebral segment in chelicerates. Using the spider Cupiennius salei, the mite Archegozetes longisetosus, and several head Hox genes as probes, it has been concluded that this segment is present.In chelicerates, lab is expressed anteriorly in the developing pedipalps (homologous to the appendages of the intercalary segment). This expression extends posteriorly, to the fourth pair of walking limbs: this location corresponds to the first pair of the thoracic appendages in insects and crustaceans (maxillipeds in P. scaber). Therefore, the anterior boundary of lab expression appears to be conserved among chelicerates, crustaceans, and insects. It is notable, however, that the posterior limit of expression is not conserved. The integumentary expression of lab is limited to the intercalary or second antennal segment in insects and crustaceans, respectively, whereas in chelicerates, expression extends posteriorly a further four full segments. In A. longisetosus, pb is expressed in a broad domain from the pedipalps to the third pair of walking legs, where it is accumulated in the appendages. This boundary is colinear with both lab and Dfd, and is thus similar to the relative expression domains of these genes in vertebrates, annelids, and Porcellio; but it is dissimilar to that seen in insects. Thus, the anterior boundary of pb expression in the a2 segment of P. scaber appears to resemble that seen in chelicerates rather than that in insects. However, it should also be noted that, as for lab, the expression domain of chelicerate pb extends further to the posterior than that in insects and crustaceans (Abzhanov, 1999).
Comparison of the expression patterns of the crustacean and insect Hox genes demonstrates that there is conservation of segment affinity (e.g., lab) and spatial colinearity (e.g., lab, pb, Dfd, and Scr) of expression. In addition, the anterior boundaries of the lab and Dfd genes appear to be conserved in insects, crustaceans, and chelicerates. However, there is also divergence of the observed expression domains (e.g., pb and Dfd). Consequently, substantial variation in the deployment of the Hox genes, and presumably in the developmental processes regulated by them, can be seen in homologous and morphologically similar crustacean and insect head structures. Genes involved in the development of mandibles and posterior mouthparts in insects are expressed in novel, though still colinear domains. For example, in insects, the maxillary and labial mouthparts express pb, whereas in P. scaber, the homologous appendages both express and probably depend on Scr, a different head homeotic gene (Abzhanov, 1999).
It is hypothesized that the mandibulate head evolved prior to the establishment of the defined head Hox gene expression domains which have been recruited independently to their current regions and developmental functions in crustaceans and insects. This model involves an intermediate hypothetical mandibulate ancestor that did not have segment-specific expression domains and probably resembled the pattern of expression seen in modern chelicerates. The specification of individual segments and mouthparts in such an animal would depend on the redundant and/or fractional functions of multiple Hox genes, and would be facilitated by the subsequent evolution of more distinct expression domains. That is, the head Hox genes would have functioned in a manner analogous to the genes of the D. melanogaster Bithorax complex. To test this model and to better understand the evolution of the Hox genes and head structures, further studies across different crustacean and myriapod groups will be required (Abzhanov, 1999).
The chicken ortholog Hoxb-1expressed under control of Drosophila regulatory sequences can rescue Drosophila labial null mutants. This indicated that the chicken protein can regulate labial target genes. Labial and chicken proteins are divergent except for a short stretch of amino acids terminal to the homeodomain (Lutz, 1996).
The Strongylocentrotus purpuratus genome contains a single ten-gene Hox complex >0.5 megabase in length. This complex was isolated on overlapping bacterial artificial chromosome and P1 artificial chromosome genomic recombinants by using probes for individual genes and by genomic walking. Echinoderm Hox genes of paralog groups (PG) 1 and 2 are reported. The cluster includes genes representing all paralog groups of vertebrate Hox clusters, except that there is a single gene of the PG4-5 types and only three genes of the PG9-12 types. The echinoderm Hox gene cluster is essentially similar to those of the bilaterally organized chordates, despite the radically altered pentameral body plans of these animals (Martinez, 1999).
The cephalochordate amphioxus has a single Hox gene cluster. The genomic
organization of four adjacent amphioxus genes, AmphiHox-1 to AmphiHox-4, together with analysis of their spatiotemporal expression patterns, is described. These genes obey temporal colinearity; three of the genes also obey spatial colinearity in the developing neural tube. AmphiHox-1, AmphiHox-3, and AmphiHox-4 show segmental modulation of their expression levels, a two-segment phasing of spatial colinearity, and, at least for AmphiHox-4, asymmetrical expression. AmphiHox-2 is unlike other amphioxus Hox genes: it does not obey spatial colinearity and it has no positional expression in the neural tube. AmphiHox-2 is expressed in the preoral pit of larvae, from which the homolog of the anterior pituitary develops. It is suggested that the ancestral role of chordate Hox genes was primarily in the neural tube and that chordate Hox genes can functionally diverge in a manner analogous to that of Drosophila ftz or zen(Wada, 1999).
Genes of the Hox cluster are restricted to the animal kingdom and play a central role in axial patterning in divergent animal phyla. Despite its evolutionary and developmental significance, the origin of the Hox gene cluster is obscure. The consensus is that a primordial Hox cluster arose by tandem gene duplication close to animal origins. Several homeobox genes with high sequence identity to Hox genes are found outside the Hox cluster and are known as 'dispersed' Hox-like genes; these genes may have been transposed away from an expanding cluster. Three of these dispersed homeobox genes form a novel gene cluster in the cephalochordate amphioxus. AmphiCdx, AmphiXlox and AmphiGsx are in respectively the Caudal, Zerknullt, and Gsh (related to Labial and Proboscipedia) families of homeodomain proteins. The finding that amphioxus Gsx, Xlox and Cdx class genes form a novel homeobox cluster challenges the idea that these homeobox gene classes are 'dispersed' Hox genes. Instead it is argued that this 'ParaHox' gene cluster is an ancient paralogue (evolutionary sister) of the Hox gene cluster. ParaHox and Hox gene clusters arose by duplication of a ProtoHox gene cluster containing Gsx, Xlox and Cdx genes representing an anterior, 'group three' and posterior subfamily. Amphioxus ParaHox genes have co-linear developmental expression patterns in anterior, middle and posterior tissues. It is proposed that the origin of distinct Hox and ParaHox genes by gene-cluster duplication facilitated an increase in body complexity during the Cambrian explosion (Brooke, 1998).
Ascidians (phylum Urochordata) and vertebrates belong to the Phylum Chordata and both have dorsal tubular central nervous systems. The structure of the ascidian neural tube is extremely simple, containing less than 400 cells, among which less than 100 cells are neurons. Recent studies suggest that despite its simple organization, the mechanisms patterning the ascidian neural tube are similar to those of the more complex vertebrate brain. However, identification of homologous regions between vertebrate and ascidian nervous systems remains to be resolved. This study describes the expression of the HrPax-258 gene (Drosophila homolog: Sparkling), an ascidian homolog of vertebrate Pax-2, Pax-5 and Pax-8 genes. Molecular phylogenetic analyses indicate that HrPax-258 is descendant from a single precursor gene that gave rise to the three vertebrate genes. The expression pattern of HrPax-258 suggests that this subfamily of Pax genes has conserved roles in regional specification of the brain. Comparison with expression of ascidian Otx (Hroth) and a Hox gene (HrHox1) by double-staining in situ hybridizations indicates that the ascidian brain region can be subdivided into three parts: an anterior region marked by Hroth, and probably homologous to the vertebrate forebrain and midbrain; a middle region, marked by HrPax-258 and probably homologous to the vertebrate anterior hindbrain (and maybe also the midbrain), and a posterior region marked by Hox genes, which is homologous to the vertebrate hindbrain and spinal cord. Later expression of HrPax-258 in atrial primordia implies that basal chordates such as ascidians have already acquired a sensory organ that develops from epidermal thickenings (placodes) and expresses HrPax-258; this organ may be homologous to the vertebrate ear. The atrial primordia develop as a pair of ectodermal invaginations that fuse to form one atrial siphon. This mode of formation is strikingly similar to that of vertebrate placodes. The atrium of adult ascidians develops ciliated sensory cells in cupular organs resembling those of the vertebrate acoustico-lateralis system. Therefore, placodes are not likely to be a newly acquired feature in vertebrates, but may have already been possessed by the earliest chordates. It is suggested that the atrial primordia of ascidian larvae are homologous to the vertebrate inner ear (otic system). HrPax-258 is also expressed in the primordial pharynx. This organ develops into the oral siphon of adult ascidians. The oral siphon of adult ascidians can be regarded as a mouth. The similarity of cell lineage strongly supports the homology between the ascidian and vertebrate mouth (Wada, 1998).
In the invertebrate chordate amphioxus, as in vertebrates, retinoic acid (RA) specifies position along the anterior/posterior axis with elevated RA signaling in the middle third of the endoderm setting the posterior limit of the pharynx. AmphiHox1 is also expressed in the middle third of the developing amphioxus endoderm and is activated by RA signaling. Knockdown of AmphiHox1 function with an antisense morpholino oligonucleotide shows that AmphiHox1 mediates the role of RA signaling in setting the posterior limit of the pharynx by repressing expression of pharyngeal markers in the posterior foregut/midgut endoderm. The spatiotemporal expression of these endodermal genes in embryos treated with RA or the RA antagonist BMS009 indicates that Pax1/9, Pitx and Notch are probably more upstream than Otx and Nodal in the hierarchy of genes repressed by RA signaling. This work highlights the potential of amphioxus, a genomically simple, vertebrate-like invertebrate chordate, as a paradigm for understanding gene hierarchies similar to the more complex ones of vertebrates (Schubert, 2005 ).
The vertebrate Hox genes have been shown to confer regional identity along the anteroposterior axis of the developing embryo, especially within the central nervous system (CNS) and the paraxial mesoderm. The notochord has been shown to play vital roles in patterning adjacent tissues along both the dorsoventral and mediolateral axes. However, the notochord's role in imparting anteroposterior information to adjacent structures is less well understood, especially since the notochord shows no morphological distinctions along the anteroposterior axis and is not generally described as a segmental or compartmentalized structure. Four zebrafish hox genes (hoxb1, hoxb5, hoxc6 and hoxc8) are regionally expressed along the anteroposterior extent of the developing notochord. Notochord expression for each gene is transient, but maintains a definite, gene-specific anterior limit throughout its duration. The hox gene expression in the zebrafish notochord is spatially colinear with those genes lying most 3' in the hox clusters having the most anterior limits. The expression patterns of these hox cluster genes in the zebrafish are the most direct molecular evidence for a system of anteroposterior regionalization of the notochord in any vertebrate studied to date (Prince, 1998).
The vertebrate hindbrain is divided into rhombomeres (r); serially homologous segments. Pbx and Hox proteins are hypothesized to form heterodimeric, DNA binding transcription complexes which specify rhombomere identities. Eliminating zebrafish Lazarus (Lzr/Pbx4) and Pbx2 function prevents hindbrain segmentation and causes a wholesale anterior homeotic transformation of r2-r6, to r1 identity. Pbx proteins interact with Hox paralog group 1 proteins to specify segment identities broadly within the hindbrain, and this process involves the Pbx:Hox-1-dependent induction of Fgf signals in r4. It is proposed that in the absence of Pbx function, r2-r6 acquire a homogeneous ground state identity, that of r1, and that Pbx proteins, functioning primarily with their Hox partners, function to modify this ground state identity during normal hindbrain development (Waskiewicz, 2002).
Interactions between Pbx and Hox-1 paralogs (hoxa1 and hoxb1 in the mouse, hoxb1a and hoxb1b in the zebrafish) directly specify r4 identity via essential Hox-1/Pbx binding sites in the regulatory regions of target genes. In mouse Hoxa1-/-;b1-/- double mutants, r4 identity is not specified. In the zebrafish, morpholino knockdown of hoxb1a and hoxb1b do not prevent r4 specification; however, this is likely due to the failure of the morpholinos to completely eliminate gene function rather than due to a basic difference in the mechanism of hox-1 function in mouse and zebrafish, since reducing hoxb1a and hoxb1b function in a zygotic lzr/pbx4 background prevents r4 specification. Furthermore, reducing pbx activity uncovers a role for hox-1 genes in specifying pattern more broadly in the hindbrain, since krox20 expression in r3 and r5 and val/mafB expression in r5 and r6 are also virtually eliminated in Zlzr; hox-1 MO-injected embryos (Waskiewicz, 2002).
These observations suggest that Hox-1 proteins, together with their Pbx partners, function at the top of the hindbrain patterning hierarchy, possibly specifying rhombomere identities through the regulation of signals derived from r4. Indeed, previous work in the mouse hasdemonstrated that hoxa1, which is never expressed in r3, nevertheless contributes non-cell autonomously to its specification. fgf3 and fgf8 are expressed in r4 in zebrafish and are required for the specification of r5 and r6. In zebrafish embryos lacking pbx2 and pbx4 function, upregulation of Fgf signals (Fgf3 and Fgf8) in r4 fails to occur. hox-1 genes are both required and sufficient to drive fgf3 expression in r4. These observations, together with previous work in the zebrafish and mouse, led to a model for hindbrain patterning in the zebrafish. In this model, Pbx interacts with Hox-1 genes to specify r4 identity including the upregulation of hoxb1a, hoxa2, fgf3, and fgf8. Fgf3 and Fgf8 in turn signal to the surrounding rhombomeres, contributing to the establishment of r3, r5, and r6 identity (Waskiewicz, 2002).
It is important to note, however, that Pbx function in hindbrain development is not limited to driving the expression of r4-derived patterning signals. If this were the case, mutant cells would be able to acquire r2-r6 identities in a wild-type host. However, mosaic analysis demonstrates unambiguously that Pbx is required cell autonomously in r2-r6 since mutant cells cannot contribute to, and as a result are excluded from, these rhombomeres in a wild-type host. Thus it is proposed that Pbx is required at multiple levels of the hindbrain patterning hierarchy, first with Hox-1 proteins in r4 but then subsequently with other Hox or as yet unidentified non-Hox partners to specify aspects of r2-r6 identity in a cell-autonomous manner. Consistent with this, Hox-3 expression in r5 and r6 has been shown to be maintained by a Pbx-dependent autoregulatory loop, and Hoxa2, which requires Pbx function in zebrafish has been shown to contribute to r2 and r3 specification in the mouse (Waskiewicz, 2002).
In contrast to the strong effects seen on patterning in r2-r6 in MZlzr; pbx2 MO embryos, patterning of the posterior-most vagal region of the hindbrain is relatively mildly affected. The anterior limit of hoxb4 expression is diffuse; however, levels of hoxb4 expression are normal. Vagal motor neurons characteristic of the posterior hindbrain are also present, although reduced in number. Most strikingly, cells lacking Pbx function are able to contribute normally to the hindbrain posterior to the r6/r7 boundary, suggesting that they are able to acquire the cell surface characteristics of the posterior-most hindbrain. The observation that posterior hindbrain identities are less strongly affected by loss of Pbx function is unexpected, given that the Hox genes that are expressed in this region are strongly auto- and cross-regulated, and the required autoregulatory elements contain essential Pbx:Hox binding sites. However, it is noted that in a cross-species reporter assay designed to test the Pbx dependence of such a Hoxb4 autoregulatory element, reporter expression was only partially suppressed in Exd mutant flies, suggesting that Hox-4 may be partially Pbx independent in vertebrates. Other critical Hox-4 targets may be similarly Pbx independent. Another explanation is that aspects of posterior hindbrain patterning may not be strictly dependent on Hox function. The vagal hindbrain is outside of the meristic series, forming a nonsegmented transition zone between the segmental region of the hindbrain and the spinal cord. The robust ability of MZlzr; pbx2 MO cells to contribute to this region in genetic mosaics may reflect a reduced importance of cell sorting for the maintenance of identity in this region of the hindbrain (Waskiewicz, 2002).
In this work, an important distinction was made between r1 and r0, with r1 being the narrow, ephA4a, fgfr3-expressing domain immediately anterior to r2, and r0 being the larger engrailed-expressing domain between r1 and the mid-hindbrain junction. A similar distinction has been hypothesized based on morphological criteria in the chick, and
molecular analysis has suggested that the r1-r0 distinction may indeed be a common feature of vertebrate embryos since fgfr3 is expressed in a similar domain in the chick. Importantly, r1 is the posterior-most region of the neural tube which does not express any hox genes; however, it is competent to respond to Hox-encoded patterning information. Taken together with the strong evidence presented (physical and genetic interactions between Pbx and Hox proteins) it is proposed that the primary function of Pbx genes during hindbrain patterning is to facilitate Hox function, and that r1 is a hindbrain ground state: the default fate established in the absence of Pbx:Hox activity (Waskiewicz, 2002).
Hox genes are classically ascribed to function in patterning the anterior-posterior axis of bilaterian animals; however, their role in directing molecular mechanisms underlying morphogenesis at the cellular level remains largely unstudied. This study has unveil a non-classical role for the zebrafish hoxb1b gene, which shares ancestral functions with mammalian Hoxa1, in controlling progenitor cell shape and oriented cell division during zebrafish anterior hindbrain neural tube morphogenesis. This is likely distinct from its role in cell fate acquisition and segment boundary formation. Without affecting major components of apico-basal or planar cell polarity, Hoxb1b regulates mitotic spindle rotation during the oriented neural keel symmetric mitoses that are required for normal neural tube lumen formation in the zebrafish. This function correlates with a non-cell-autonomous requirement for Hoxb1b in regulating microtubule plus-end dynamics in progenitor cells in interphase. It is proposed that Hox genes can influence global tissue morphogenesis by control of microtubule dynamics in individual cells in vivo (Zigman, 2014).
Gene duplication of Labial homologs in vertebrates As a result of a whole genome duplication event in the lineage leading to teleosts, the zebrafish has seven clusters of Hox patterning genes, rather than four, as described for tetrapod vertebrates. To investigate the consequences of this genome duplication, a detailed comparison of genes from a single Hox paralog group, paralog group (PG) 1 was carried out. The sequences, expression patterns and potential functions of all four of the zebrafish PG1 Hox genes were examined, and these data were compared with that available for the three mouse genes. Since the basic functions of Hox genes appear to be tightly constrained, comparison with mouse data has allowed specific changes to be identified in the developmental roles of Hox genes that have occurred during vertebrate evolution. Variation in expression patterns, amino acid sequences within functional domains, and potential gene functions were identified both within the PG1 genes of zebrafish, and in comparison to mouse PG1 genes. Novel expression patterns were observed in the midbrain, such that zebrafish hoxa1a and hoxc1a are expressed anterior to the domain traditionally thought to be under Hox patterning control. The hoxc1a gene shows significant coding sequence changes in known functional domains, which correlate with a reduced capacity to cause posteriorizing transformations. Moreover, the hoxb1 duplicate genes have differing functional capacities, suggesting divergence after duplication. An intriguing function, 'shuffling', has occurred between paralogs, such that one of the zebrafish hoxb1 duplicates, hoxb1b, performs the role in hindbrain patterning played in mouse by the non-orthologous Hoxa1 gene (McClintock, 2001).
A morpholino-based knockdown approach has been used to investigate the functions of a pair of zebrafish Hox gene duplicates, hoxb1a and hoxb1b, which are expressed during development of the hindbrain. The zebrafish hoxb1 duplicates have equivalent functions to mouse Hoxb1 and its paralogue Hoxa1. Thus, a 'function shuffling' has been revealed among genes of paralog group 1 during the evolution of vertebrates. Like mouse Hoxb1, zebrafish hoxb1a is required for migration of the VIIth cranial nerve branchiomotor neurons from their point of origin in hindbrain rhombomere 4 toward the posterior. By contrast, zebrafish hoxb1b, like mouse Hoxa1, is required for proper segmental organization of rhombomere 4 and the posterior hindbrain. Double knockdown experiments demonstrate that the zebrafish hoxb1 duplicates have partially redundant functions. However, using an RNA rescue approach, it has been revealed that these duplicated genes do not have interchangeable biochemical functions: only hoxb1a can properly pattern the VIIth cranial nerve. Despite this difference in protein function, evidence is provided that the hoxb1 duplicate genes were initially maintained in the genome because of complementary degenerative mutations in defined cis-regulatory elements (McClintock, 2002).
Transactivational domain structure of Labial homologs Hox proteins control developmental patterns and cell differentiation in vertebrates by acting as positive or negative regulators of still unidentified downstream target genes. The homeodomain and other small accessory sequences encode the DNA-protein and protein-protein interaction functions that ultimately dictate target recognition and functional specificity in vivo. The effector domains responsible for either positive or negative interactions with the cell transcriptional machinery are unknown for most Hox proteins, largely due to a lack of physiological targets on which to carry out functional analyses. The transcriptional activation domains have been identified for three human Hox proteins, HOXB1, HOXB3, and HOXD9, that interact in vivo with the autoregulatory and cross-regulatory enhancers of the murine Hoxb-1 and human HOXD9 genes. Activation domains have been defined both in a homologous context, i.e., within a HOX protein binding as a monomer or as a HOX-PBX heterodimer to the specific target, and in a heterologous context, after translocation to the yeast Gal4 DNA-binding domain. Transfection analysis indicates that activation domains can be identified in different regions of the three HOX proteins depending on the context in which they interact with the DNA target. These results suggest that Hox proteins may be multifunctional transcriptional regulators, interacting with different cofactors and/or components of the transcriptional machinery, depending on the structure of their target regulatory elements (Vigano, 1998).
A conventional deletion analysis on the 270-aa N terminus of HOXD9 shows that the first 75 residues contain a potential transcriptional activator when tested in the context of a Gal4 chimeric protein. In contrast, this region is dispensable when the activity of the protein is tested on the HCR (Hox control region, a 90-bp, ATTA-rich autoregulatory element identified in the HOXD9 locus), a context in which most of the activating function appears to be located within residues 76 to 264. The regions identified by the two alternative assays share no obvious characteristics with canonical eukaryotic activator domains and are only loosely conserved among different vertebrate species. The activation domain of another posteriorly expressed Hox protein, HOXD8, can be localized to a similar sub-N-terminal region. HOXD8 and HOXD9 bind the multiple ATTA-containing sites within the HCR as monomers in a noncooperative fashion, while Gal4-DBD chimeras bind the Gal4-responsive element (UAS) as a homodimer, a context that could force the HOXD9 N-terminal region to assume a different structural conformation and unmask a potential activating function in the N-terminal 75 residues. For the HOXB1-PBX heterodimer, the analysis carried out on the natural autoregulatory element identifies a transcriptional activation domain in a Ser-Pro-rich, 52-residue sub-N-terminal region. This region also contains most of the HOXB1 transcriptional activity when tested as a Gal4-DBD chimera, a possible indication that the 52-residue region assumes a similar conformation or activates transcription by a similar mechanism, either in the context of a homodimer or in that of a HOX-PBX heterodimer. The activity of HOXB3 was tested in three different contexts, i.e., upon binding DNA as a monomer to an ATTA-containing element: as a HOX-PBX heterodimer to a bipartite HOX-PBX core element, and as a Gal4-DBD chimeric homodimer to the Gal4-responsive element. Although in the context of a monomer the transcriptional activity is spread over the entire protein sequence, only the C terminus contains a potent activator domain in the context of a Gal4 homodimer or of a PBX heterodimer. The 71-residue C terminus is relatively highly conserved in the mammalian group 3 Hox proteins (Vigano, 1998). Interaction of Labial homologs with PBX Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
labial: Biological Overview | Regulation | Developmental Biology | Effects of Mutation | References
Society for Developmental Biology's Web server.
{kind=link}
{kind=link}