Select image to enlarge

Figure 8.3
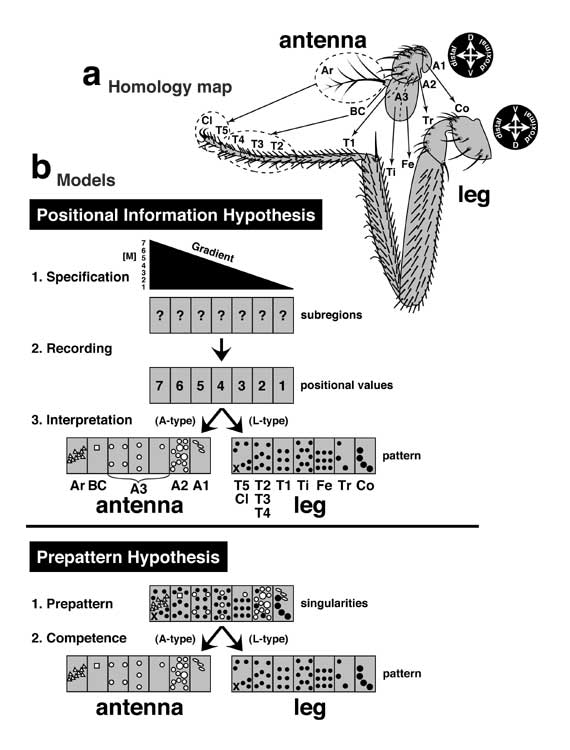
Our current understanding of what makes an antenna different from a leg. This problem was discussed earlier with regard to the 1969 debate between positional information (PI) and prepatterns (cf. Fig. 4.3). It is being revisited here to see how much progress has been made since then. The answer, in short, is 'a little'.
a. The old problem posed by the 1:1 correspondence of antennal and leg regions. These homologies were deduced from AntpGOF phenotypes [3445] and confirmed in other mutants (e.g., babGOF [1516]; cf. Fig. 4.3).
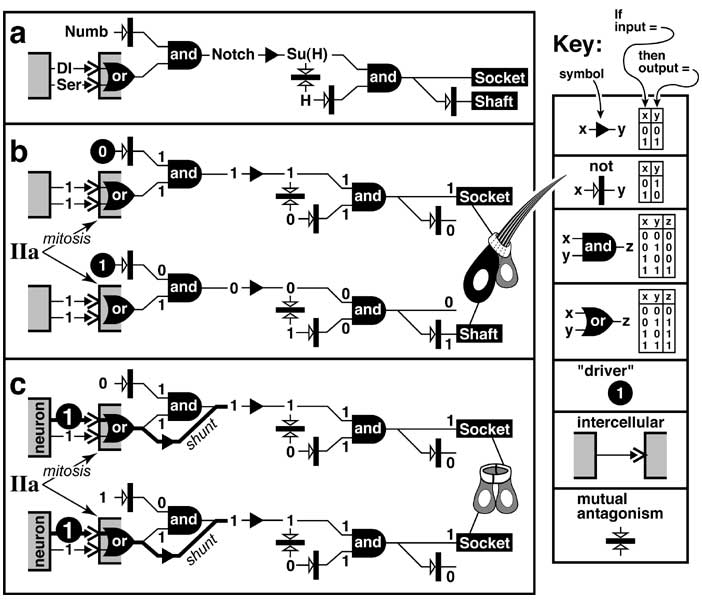
b. The 'Either Duo Hypothesis' for why antennae develop differently from legs. Antennae and legs are both thought to use an unknown morphogen (M) to specify radial positions relative to the future tip (cf. Fig. 5.4). In keeping with Wolpert's model (cf. Fig. 4.3), cells may convert this scalar signal into digital states ('positional values'), but those states are not propagated by cell lineage [544, 3446, 4575], nor are they stabilized by 'memory' genes of the Pc-G or Trx-G (see text). Thus, no 'recording' phase is interposed between specification and interpretation (A and L denote antennal vs. leg types of development). Eventually the antennal and leg rudiments turn ON the same region-specific genes but express some of them differently. Horizontal bars denote expression domains; gray shading means weak or incomplete expression; checkered shading indicates heterogeneity. (N.B.: the axis is reversed relative to Fig. 5.11.) One code is well documented: the combination 'hth-ON AND Dll-ON' (1) turns ON spalt and (2) evokes ectopic antennae in other discs (via Gal4 drivers) [1085]. Also, hthLOF or DllLOF clones turn OFF spalt and cause antennal-to-leg homeosis [677, 1085, 3226, 3380, 3716]. The implication is that antennal identity is encoded by the Hth-Dll overlap, which leg discs lack (until late 3rd instar) [4760]. If so, then how do cells outside A2-A3 avoid making leg tissue? Distally this role may be filled by ss since ssLOF clones exhibit the same homeosis as hthLOF or DllLOF [564, 1119, 1166, 2933, 4150, 4508]. However, ssGOF cannot induce antennae outside the head (except for a minor claw-to-arista switch in the leg) [1119], so an unidentified factor 'X' (in antenna but not leg) probably cooperates with Ss as Hth works with Dll. Either of these pairs of agents could enable antennal cells to deviate from the leg program in the same way that Ubx lets haltere cells deviate from the wing program (cf. Fig. 8.2). The logic is summarized in the circuit diagram (cf. Fig. 2.7 for symbols), which also shows how exogenous Hox proteins (e.g., Antp) cause homeosis by stifling Hth [4806] or Ss [1119]. The final stage of development involves creating patterns of small cuticular elements (e.g., bristles and sensilla, rendered abstractly here). We know a fair amount about how this is done [1653, 2055, 3531, 3548, 4125] (see Chs. 3 and 5 for the leg), but the links between the genes that act locally and the ones act regionally remain elusive. Those links may involve an intermediate level of prepatterns (cf. Fig. 6.14), so Stern's hypothesis is still as relevant as Wolpert's (cf. Fig. 4.3). Overall, this system is murkier than our picture of embryo segmentation (cf. Fig. 4.2). Remaining questions include: (1) what is the initial bias (different tip morphogens?) that steers tissue into an A-type vs. L-type mode of interpretation? and (2) is A1 actually a 'coxa in disguise' (miniaturized?), or does it use a third duo of genes to veer away from a leg fate? Black or white symbols in the antennal and leg schematics are abstract renditions of the the actual patterns.
Genes (DO = details omitted): al (aristaless) [618, 881]Δ, bab (bric à brac) [1516], dac (dachshund) [4761], Dll (Distal-less; DO: expression above the tibia only appears in late 3rd instar) [1085, 3242, 4761]Δ, hth (homothorax) [1085, 4761]Δ, spalt [1085]Δ, ss (spineless; DO: expression in leg disc shifts to periphery in late 3rd instar; cf. tango) [1166]Δ. In the leg disc, spalt is expressed in 'isolated cells scattered around the proximal region' [220].
N.B.: During early antennal development, hth is expressed throughout the primordium [677], and ss expression is dependent upon hth AND Dll (I. Duncan, pers. comm.).
The diagram in a is redrawn from [3445], and domains of gene expression (b) are modified from [3242] based on above references.
|
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}