The Interactive Fly
Genes involved in tissue and organ development
Fertilization typically involves membrane fusion between sperm and eggs. In Drosophila, however, sperm enter eggs with membranes intact. Consequently, sperm plasma membrane breakdown (PMBD) and subsequent events of sperm activation occur in the egg cytoplasm. It has been proposed that mutations in the sneaky (snky) gene result in male sterility due to failure in PMBD. This study supports this proposal by demonstrating persistence of a plasma membrane protein around the head of snky sperm after entry into the egg. It is further shown that snky is expressed in testes and encodes a predicted integral membrane protein with multiple transmembrane domains, a DC-STAMP-like domain, and a variant RING finger. Using a transgene that expresses an active Snky-Green fluorescent protein fusion (Snky-GFP), it is shown that the protein is localized to the acrosome, a membrane-bound vesicle located at the apical tip of sperm. Snky-GFP also allowed following the fate of the protein and the acrosome during fertilization. In many animals, the acrosome is a secretory vesicle with exocytosis essential for sperm penetration through the egg coats. Surprisingly, the Drosophila acrosome is a paternally inherited structure. Evidence is presented that the acrosome induces changes in sperm plasma membrane, exclusive of exocytosis and through the action of the acrosomal membrane protein Snky. Existence of testis-expressed Snky-like genes in many animals, including humans, suggests conserved protein function. The characteristics of Drosophila Snky, acrosome function and sperm PMBD is related to membrane fusion events that occur in other systems (Wilson, 2006).
Studies of fertilization have revealed the universal importance of membrane events to prepare and coordinate the gametes and ensure their successful union. These events have been most extensively studied in sperm of selected marine invertebrate and mammalian species and include dynamic changes in membrane proteins and lipids during sperm capacitation, acrosome exocytosis, binding, adhesion and membrane fusion with the egg. The majority of molecules with confirmed roles in these processes are cell adhesion molecules, ion channels or signal transduction components with molecular mechanisms of action that have been studied in multiple cell types. There is considerable interest in discovering proteins that act specifically during fertilization. The characterization of these molecules will contribute to a better understanding of the specialized properties of gametes and to the practical goal of identifying new targets for contraceptives (Wilson, 2006).
A genetic approach using Drosophila was employed to identify novel sperm molecules required for fertilization. Previous studies of male sterile mutants indicated that those affecting fertilization might be relatively rare. Therefore, a large number of male-sterile mutations were isolated and categorized to find a subset that disrupt sperm-egg interactions or induce paternal effect defects (Fitch, 1998; Wakimoto, 2004). Previously described were mutations in a gene called sneaky (snky), which met genetic criteria of specifically affecting fertilization (Fitch, 1998). Mutations of snky showed detectable effects only on male fertility. Sperm produced by mutant males were competent to enter the egg but arrested before sperm nuclear decondensation and aster formation. It was proposed that this sperm activation defect is due to a failure in breakdown of the sperm plasma membrane, a step that normally occurs immediately after sperm entry into the egg in Drosophila (Wilson, 2006). P>This study provides phenotypic evidence of a role for Snky in affecting the integrity of the sperm plasma membrane during fertilization. The snky gene was molecularly identify and its transcript and predicted protein were characterized to investigate how the protein might function. The results indicate that Snky is an acrosomal membrane protein that is contributed to the early embryo. In addition to suggesting a possible molecular role for Snky, these studies have implications for the function and the fate of the acrosome during Drosophila fertilization (Wilson, 2006). <>Fertilization in Drosophila is unorthodox in that it does not involve a membrane fusion event between the sperm and egg. The mechanism used by the sperm to penetrate through the egg plasma membrane remains an interesting mystery. In ultrastructural studies of fertilization in D. melanogaster and D. montana, Perotti (1975) obsrved that the entire sperm enters the egg, including the excessively long tail surrounded by its plasma membrane. There was no evidence for 'extra' membranes that would support uptake into the egg by endocytosis. While these studies did not capture the sperm before nuclear decondensation, Perotti described one case in which a decondensing sperm nucleus was surrounded by what appeared to be remnants of the sperm plasma membrane. These studies support the idea that the Drosophila sperm head must undergo PMBD in the egg cytoplasm. This step is required for the sperm nucleus to gain acces to activating cytoplasmic factors that promote nuclear decondensation and replication, and for the centrosome to elaborate the sperm aster (Fitch, 1998). The current studies support this sequence of events and implicate Snky function in an event upstream of PMBD. The original proposal that snky mutant sperm fail in sperm activation because they arrest before PMBD was based on their poor accessibility to membrane impermeant chromatin dyes (Fitch, 1998). Further support was obtained by Ohsako (2003), who used an antibody to a proposed cell surface proteoglycan to demonstrate retention on the sperm head before and after insemination. In the current study, the rat CD2 protein was ectopically expressed on the sperm plasma membrane and it was showed that immunoreactivity to this specific integral membrane protein was retained on the head of snky1 sperm for at least 30 minutes after entry into the egg. These observations provide strong support for the proposal that snky sperm do not shed the plasma membrane surrounding the head. They also suggest that understanding the function of the Snky protein should provide clues about how PMBD is initiated or accomplished at the molecular level (Wilson, 2006).
olecular studies show that Snky is a member of a family of related membrane proteins that are present in animals but are apparently absent in other lineages. Conservation among Snky family members points to three domains of potential functional significance. The most striking motif is the C4-C4 RING finger, which has been noted in at least four other proteins. In one of these, hNOT4, the C4-C4 RING has been shown to fold and bind zinc in a cross-brace fashion similar to the more common C3HC4 RING finger, suggesting structural and functional similarity between these variants. Although RING finger proteins have been implicated in a broad variety of cellular functions, recent studies suggest that their unifying role is in mediating protein-protein interactions in large multiprotein assemblages. Thus, Snky may play a role in organizing and holding together macromolecular complexes at the membrane via its C4-C4 RING. The second Cys-containing region is the patterned 6 Cys motif, the functional significance of which is indicated by conservation and by the recovery of a male sterile snky allele that mutates the second Cys to a Ser. Cysteines in this region may form disulfide bonds to create a binding pocket or otherwise allow interactions with additional proteins. The third region is the DC-STAMP-like region, named after Dendritic cell specific transmembrane protein, a plasma membrane protein that was first described for its expression in human dendritic cells. Thus far, the function of only a few DC-STAMP-domain-containing proteins have been examined through the analysis of mutations or other knockdown strategies. The general classification of these proteins has been as members of a new class of putative receptors. Mouse DC-STAMP is upregulated in differentiating osteoclasts, and required for osteoclast fusion to form multinucleate cells. The C. elegans SPE-42 is required in sperm and is proposed to act at a step in fertilization just before or during sperm-egg membrane fusion (Kroft, 2005). Snky is the third DC-STAMP domain-containing protein to have its function studied and, like Spe-42, it has a role in sperm function. Although the precise molecular function of the DC-STAMP domain remains unknown, these three examples support a role in mediating membrane-membrane interactions (Wilson, 2006).
Th predicted structure of Snky, with its multiple transmembrane domains, and Snky-GFP localization studies provide evidence that Snky is an acrosomal membrane protein. This localization presents the intriguing question of how an acrosomal membrane protein is able to influence the integrity of the sperm plasma membrane. The findings also have broader implications for acrosome function. The acrosome is a Golgi-derived membrane-bound organelle found at the apical end of sperm, and its function has been extensively studied in marine invertebrates and mammals. In these organisms, the acrosome is best known as a specialized secretory vesicle that undergoes exocytosis. Like many secretory events, a rise in intracellular Ca2+ is required for acrosome exocytosis. This increase in Ca2+ requires influx into the sperm as well as efflux from internal stores. The acrosome has been identified as an internal source of Ca2+ and is believed to release Ca2+ to contribute to its own exocytosis (Walensky, 1995; Herrick, 2005). Ultrastructural studies show that exocytosis involves vesiculation of the outer acrosomal membrane and overlying plasma membrane. This results in the release of contents of the acrosome, which include hydrolytic enzymes and other components that facilitate binding and penetration through the egg coats. Exocytosis also exposes the inner acrosomal membrane, resulting in a new membrane patch and associated acrosomal molecules on the surface of the sperm. In marine invertebrates, this newly exposed region is the site of binding and membrane fusion with the egg, providing a direct physical link between the requirement for acrosome exocytosis and membrane fusion. In mammals, acrosome exocytosis is also a prerequisite for sperm to bind to and fuse with the egg. However, the plasma membrane that lies over the equatorial segment of the acrosome, and not the acrosome membrane itself, is believed to be the point of membrane fusion with the egg. Experimental studies of hamster sperm suggest that fusion competency of this specialized region of mammalian sperm requires changes that occur immediately before or during acrosome exocytosis, as well as the contents of the acrosome (Wilson, 2006 and references therein).
Conidering these known acrosome functions, it is interesting that acrosomes are not universal features of animal sperm. Acrosomes are not present in the amoeboid sperm of nematodes, and they are occasionally absent or reduced in species that otherwise possess the flagellated sperm typical of animals. For instance, the absence or reduction of an acrosome in mature sperm of teleosts and certain insect species is well documented and is generally considered a derived condition. Hence the requirement for the acrosome during fertilization has been eliminated or bypassed in some lineages during the course of evolution. Ultrastructural studies show that an acrosome is typical of insect sperm. However, few studies have examined the role of the insect acrosome during fertilization. Studies of fertilization in the house fly Musca domestica suggested 'loosening or loss' of the sperm plasma membrane before entry into the micropyle of the egg, followed by exocytosis of acrosomal contents during passage of the sperm through the micropyle (Wilson, 2006).
Thes studies revealed a different fate for the Drosophila acrosome. The observation that Snky-GFP was a paternally contributed molecule to the early egg was a surprising finding. The persistence of a single prominent Snky-GFP structure, with an intensity and shape in eggs that appeared similar to those seen in mature sperm, suggests the possibility that the inherited structure was an intact acrosome. This possibility was further supported by studies tracking GFPsecr, a soluble protein that is secreted into the extracellular space when expressed in other Drosophila cells. Its robust and identical appearance to Snky-GFP argues against exocytosis, at least until after prometaphase of the first embryonic cell cycle (Wilson, 2006).
Thesecytological observations of the fate of the acrosome during normal fertilization, combined with the defect in PMBD observed in snky- mutant sperm, suggest that the acrosome may be acting primarily as a signaling vesicle to elicit changes in the overlying sperm plasma membrane. This activity requires the Snky acrosomal membrane protein, but occurs without acrosome exocytosis. Snky may be serving as a receptor that permits communication between the acrosome and plasma membrane. Alternatively, Snky or its associated proteins may serve to initiate or maintain contact between the membranes, or modify membrane lipids or proteins in preparation for PMBD. In this sense, Snky, DC-STAMP and SPE-42 may share a common mechanism in promoting membrane interactions (Yagi, 2005; Kroft, 2005). However, in the case of Snky, the pathway would lead to breakdown of the overlying plasma membrane, rather than fusion between two membranes (Wilson, 2006).
In Droophila, Snky is required specifically for male fertility.
It will be interesting to determine whether Snky family members are required
for sperm function in zebrafish, which have sperm that lack acrosomes, and for
acrosome function in marine invertebrate and mammalian sperm. If human
Snky-like proteins are specifically required for sperm function, then they may
be potential targets for male contraceptives. More immediately, these studies of
Snky provide tools for further investigating how the membrane events that
occur during Drosophila fertilization compare to conventional views
of membrane dynamics during sperm activation and fertilization in animals.
Research on the acrosome has focused primarily on its function as a
specialized secretory or Ca2+ storage vesicle. These studies suggest
a primary role as signaling vesicle in Drosophila, with a newly
identified acrosomal membrane protein communicating directly or indirectly
with the plasma membrane to affect changes in membrane integrity. Comparisons
among species should continue to shed light on the intriguing ways in which
sperm structures and fertilization molecules, such as Snky, may be selected
for conservation or diversification during the course of evolution (Wilson, 2006).
Chromatin condensation is a typical feature of sperm cells. During mammalian
spermiogenesis, histones are first replaced by transition proteins and then by
protamines, while little of this process is known for Drosophila. This study characterizes
three genes n the fly genome, Mst35Ba, Mst35Bb, and
Mst77F. The results indicate that Mst35Ba and Mst35Bb
encode dProtA and dProtB, respectively. These are considerably larger than
mammalian protamines, but, as in mammals, both protamines contain typical
cysteine/arginine clusters. Mst77F encodes a linker histone-like protein
showing significant similarity to mammalian HILS1 protein. ProtamineA-enhanced
green fluorescent protein (eGFP), ProtamineB-eGFP, and Mst77F-eGFP carrying
Drosophila lines show that these proteins become the important
chromosomal protein components of elongating spermatids, and His2AvDGFP
vanishes. Mst77F mutants [ms(3)nc3] are
characterized by small round nuclei and are sterile as males. These data suggest
the major features of chromatin condensation in Drosophila
spermatogenesis correspond to those in mammals. During early fertilization
steps, the paternal pronucleus still contains protamines and Mst77F but regains
a nucleosomal conformation before zygote formation. In eggs laid by
sesame-deficient females, the paternal pronucleus remains in a
protamine-based chromatin status but Mst77F-eGFP is removed, suggesting that the
sesame gene product is essential for removal of protamines while Mst77F
removal is independent of Sesame (Raja, 2005).
For mammals, the somatic set of histones are modified, as these
are in part replaced by specific variants during meiotic prophase. After
meiosis, histones are replaced by major transition proteins TP1 and TP2
and subsequenly by highly basic protamines to ensure the
remodeling of chromatin to a typically highly condensed and transcriptionally
silent state of mature sperm. These replacements leads to a shift from
histone-based nucleosomal conformation to a radically different conformation,
resembling stacked doughnut structures containing protamines as major chromatin
condensing proteins and DNA. Some mammals have only one protamine gene,
while mice andhumans have two genes encoding two
different protamines, both of which are essential for fertility and are
haploinsufficient. HILS1 (spermatid-specific
linker histone H1-like protein) has been proposed to participate in chromatin
remodeling in mouse and human spermiogenesis.
The transition between histone removal and its replacement
by protamines in mice and humans is characterized by small 6- to 10-kDa
transition proteins acting as a short-term chromosomal proteins.
In mice, the transition proteins TP1 and TP2 are redundant
in function. In fishes and birds, transition proteins are missing and protamines
directly reorganize the chromatin. In annelids and echinoderms, the nucleosomal
configuration is maintained in sperms,
while protamine-like proteins have been described for
mussels. These protamine-like proteins lack the typical
high cysteine content necessary for disulfide bridges.
Therefore, a doughnut-type chromatin structure as in mammals is unlikely to
occur in mussels. It has been proposed that
the protamine-like proteins in mussels belong to the histone H1 family. The
sperm chromatin of mussels contain core histones and thus a nucleosomal
configuration, but histone H1 is replaced by protamine-like molecules which
organize the higher order structure of the chromatin (Raja, 2005).
For Drosophila melanogaster, chromatin reorganization after meiosis has
not been studied at the molecular level. At the light microscopic level,
the Drosophila spermatid nucleus is initially round after meiosis and
then is shaped to a thin needle-like structure with highly condensed chromatin,
so that the volume of the nucleus is condensed over 200-fold.
In mammals, thevolume of the nucleus is reduced over
20-fold. In the mature sperms of Drosophila, core
histones are not detectable by immunohistology. There is
histochemical evidence for the presence of very basic proteins in sperm,
but it still remins an open question whether histones are
replaced by protamine-like basic proteins in Drosophila. The analysis of
the Drosophila genome sequence
revealed that the proteins encoded by two genes show similarity to mammalian
protamines for which the male-specific transcripts Mst35Ba and
Mst35Bb have been found and have been proposed to
encode protamine-like proteins. Another male specifically
transcribed gene, Mst77F, is a distant relative of the histone H1/H5
(linker histone) family and has been proposed to play a role either as a
transition protein or as a replacement protein for compaction of the
Drosophila sperm chromatin. With enhanced green
fluorescent protein (eGFP) fusion for these abovementioned proteins, this study shows that Mst35Ba and Mst35Bb indeed encode protamines and
Mst77F encodes a linker histone-like protein. The expression pattern of
Mst77F overlaps the pattern of protamines as a chromatin component. Furthermore, during fertilization, the removal of protamines from the male
pronucleus requires the function of the maternal component, Sesame, but not for
the removal of Mst77F. It has been shown that sesame mutants cause
impairment of the entry of histones into the male pronucleus (Raja, 2005).
Mst35Ba and Mst35Bb are
present at cytological position 35B6 and 35B6-7, respectively, on the chromosome
arm 2L. These two genes are arranged in tandem, and both consist of three exons.
The 5'UTR, codingregion, and the 3'UTR of these
genes are highly identical; they probably arose from a
recent gene duplication. The encoded protamines show over 94% identity to each
other (Raja, 2005).
A remarkable feature of protamines is their ability to form intermolecular
disulfide bridges, which is reflected by the conserved cysteine residues within
mammalian protamines. The dProtA and dProtB are of 146
amino acids (aa) and 144 aa, respectively, and thus longer than even the human
and mouse Protamine-2, which are 102 aa and 107 aa, respectively.
Both Drosophila protamines contain 10 cysteines each
and show significant similarity, particularly with respect to a high cysteine,
lysine, and arginine content to mammalian protamines.
Human and mouse Prtamine-1 aligns to the
N-terminal half of the Drosophila protamines (from aa positions 27 to
82), and four cysteine residues are conserved and regularly spaced. In contrast,
Protamine-2 of huma and mouse
shows relatively high similarity to the C-terminal half of the Drosophila
protamines, with four cysteines in this region that are conserved and regularly
spaced, whereas one cysteine is shared with the mouse and human Protamine-1 (Raja, 2005).
Mst77F is present at the cytological position 77F on the chromosome arm 3L and
lies within the large intron of PKA-R1. Mst77F is also male specifically
transcribed, and the encoded protein has been proposed to be a linker histone
H1/H5 type, which could also play the role of a transition protein or a
protamine. The Mst77F protein shares a
significant similarity to the HILS1 protein of mouse
and human HILS1, whee the percentages of cysteine,
lysine, and arginine are similar to that of mHILS1 and hHILS1.
HILS1 protein has bee recently described as a component of
the mammalian sperm nucleus. Drosophila Mst77F
encodes a protein of 215 aa with a molecular mass of 24.5 kDa and with a pI of
9.86. mHILS1 is of 170 aa and shows 39% similarity to Mst77F.
Mst77F contains 10 cysine residues as in Drosophila
protamines, and mHILS1 contains eight cystine
residues, of which four residues are conserved (Raja, 2005).
As there are considerable differences between the mammalian protamines as well
as between the mammalian HILS1 proteins and the presumptive Drosophila
homologue Mst77F, additional experiments are essential to clarify if these
proteins are indeed involved in the condensation of sperm chromatin (Raja,
2005).
Drosophila protamine mRNAs are transcribed at the primary spermatocyte
stage, whereas in mammals protamine mRNAs are synthesized at the round
spermatid stage and translationally repressed until the
elongated stage, which is mediated by 3'UTR. The Drosophila
ProtamineA-eGFP and ProtamineB-eGFP constructs do not contain the 3'UTR of the
respective protamine genes. Nevertheless, the transgenic flies carrying
these constructs still show repression of translation. So, in Drosophila,
the region responsible for the translational repression is most likely in the
5'UTR. Deletion constructs of Mst35Bb and Mst77F 5'UTRs fused to
the reporter lacZ show that the translation repression element is indeed
present in the 5'UTR. This holds true also for the mRNA of the
Mst77F-eGFP fusion gene, as is the case for all mRNAs investigated
concerning translational repression so far in male germ lines of
Drosophila. In contrast to mammalian spermatogenesis, in Drosophila transcription
ceases already with the entry into meiotic divisions.
Since the protamines are made in the elongated spermatids, the transcriptional
silencing in Drosophila spermatogenesis seems to be independent of
protamines (Raja, 2005).
When primary amino acid sequences of Drosophila protamines are compared
to mammalian protamines, it is quite evident that Drosophila protamines
are relatively large. dProtA and dProtB are over 94% identical to each other.
This could explain that both the protamines may be functionally redundant. Human
and mouse Protamine-1 aligns with the N terminus of both Drosophila
protamines, and Protamine-2 aligns more to the C
terminus. It is possible that the Drosophila
protamines undergo posttranslational cleavage at the N terminus, as is known for
mammals. The cytoplasmic eGFP fused at the C terminus
shows clear nuclear localization, indicating that the tagged protamine is
functionally intact. Drosophila protamines each contain 10 cysteine
residues at identical positions, while over 4 of 10 cysteines at the N terminus
and the C terminus are conserved with human and mouse Protamine-1 and
Protamine-2, respectively. With
nine cysteines, the content is highest in Protamine-1 of mice. Inter- or
intra-disulfide bridges can be formed between the cysteine-rich protamines to
condense the DNA. For mice it is shown that mutation in
protamine-1 or protamine-2 is haploinsufficient and causes male
sterility. A haploid situation was analyzed for the Mst35Ba and
Mst35Bb genes with the deficiency Df(2L)Exel8033/+;
these flies are fertile males and show normal
spermatogenesis. The large amount of identity that both dProtA
and dProtB exhibit can contribute to the functional redundancy (Raja, 2005).
Chromatin reorganization is an essential feature during spermiogenesis. The
functional significance of chromatin compaction during spermiogenesis is still
unknown. The main explanation seems to be that compaction of the sperm nucleus
is an essential factor for its mobility as well as for the penetration of sperm
into the egg and genomic stability. In mammals, somatic histones are in part
replaced by spermatid-specific variants during meiotic prophase,
later by major transitin proteins TP1 and TP2,
and subsequently by highy basic protamines to ensure the
remodeling of chromatin to a typically highly condensed and transcriptionally
silent state of mature sperm. These replacements lead to a shift from
histone-based nucleosomal conformation to a radically different conformation,
resembling stacked doughnut structures containing major chromatin condensing
proteins and DNA in the nucleus (Raja, 2005).
In Drosophila, so far no proteins have been identified that are involved
in the packaging of the genome in the mature sperm nucleus. One observation,
that Histone3.3 variant and the somatic H3 isoform in Drosophila are
vanishing at the time of chromatin condensation, supports the view of histone
displacement, but it was still a
question of whether it is the real absence of histones at this stage in
Drosophila or whether the antibodies are not accessible to the mature
sperm due to the tight packaging of the chromatin. To
circumvent this problem, the GFP fusion approach was chosen, use was made of the
existing His2AvDGFP, and
Protamine-eGFP andMst77F-eGFP fusion transgenic flies were generated
in order to
analyze the situation in Drosophila. The results clearly show that
histone His2AvD is lost from the spermatid nuclei at the time of appearance of
protamines and Mst77F during later stages of spermatid differentiation. The
exact molecular mechanisms underlying the histone displacement, degradation, and
incorporation of protamines onto the chromatin are poorly understood.
For mammals, evidence has een obtained that histone H2A is
ubiquitinated in mouse spermatids around the developmental time period when
histones are removed from the chromatin.
The mammalian HR6B ubiquiti-conjugating enzyme is the
homologue of yeast RAD6, and both can ubiquitinate histones in vitro.
Thus far, the mechanism of hstone displacement and protamine
incorporation is unknown during spermiogenesis in Drosophila. In
flies as well as in mammals, many questions remain unanswered that need
to be addressed about these
underlying mechanisms of chromatin remodeling during spermiogenesis (Raja, 2005).
In mammals, transition proteins act as intermediates in the histone-to-protamine
transition. In mice, the onset of HILS1 and transition
proteins TP1 and TP2 (major forms) overlaps with the pattern of Protamine-1 and
later with Protamine-2 but HILS1 and the transition proteins are no longer present in the mature sperm.
Mice lacking both TP1 and TP2show normal transcriptional
repression, histone displacement, nuclear shaping, and protamine deposition but
show the loss of genomic integrity with large numbers of DNA breaks leading to
male sterility. In Drosophila,
histones are displaced with sychronous accumulation of protamines and Mst77F.
Mst77F, a distant relative of the histone H1/H5 (linker histone) family, has
been proposed to play a role either as a transition protein or as a protamine
for compaction of the Drosophila sperm chromatin.
Mst77F shows highest similarity to HILS1 with respect to the cysteines and basic
amino acid content but
not to mouse TP1, TP2, or H1t. Moreover, the results
show that the pattern of expression of Mst77F in the nucleus is similar to that
of mHILS1 in the nucleus, with the exception that Mst77F is also transiently
detected in the flagella and persists in mature sperm nuclei, unlike mHILS1. In
mammalian mature sperm nuclei, it is only the protamines that are the chromatin
condensing proteins which persist. This again raises the question of whether
Mst77F could also play the role of protamines. However, one additional copy of
dProtB (dProtA and dProtB showing 94% identity may be functionally redundant)
does not rescue the ms(3)nc3 phenotype, indicating that the
role of Mst77F may be completely or partially different from that of protamines
in the nucleus. However, a null mutation for Mst77F is required to answer
this question with respect to chromatin condensation. In
ms(3)nc3 mutants, the chromatin condensation with the
native protamines continues to take place. When a closer look was taken at the
deposition of ProtamineB-eGFP in
ms(3)nc3/Df(3L)ri-79c
trans-heterozygotes, it revealed that the condensed chromatin in the
tid-shaped nuclei is concentrated at the two
opposite ends, with a lightly stained chromatin spaced in the center. So the
chromatin condensation takes place but may not be complete with the
incorporation of the mutant Mst77F protein. The large amount of chromatin
compaction or condensation seen in Drosophila mature sperm when compared
to that of mouse and human sperm possibly could be the result of persistence of
Mst77F in the mature sperm nuclei. It remains to be clarified whether the sperm
nucleus contains further protamines that have not yet been properly annotated (Raja, 2005).
ms(3)nc3 is a second-site noncomplementation (nc) mutation
that was isolated in an ethylmethanesulfonate screen to identify interacting
proteins involved in microtubule function in Drosophila. This study shows
that ms(3)nc3 is a
single missense mutation from a T>A transition, causing the substitution of threonine instead of serine
at aa position 149. Mst77F shows a pattern of
expression similar to protamines in the nucleus and was also seen in the flagella until the
individualization stage. Since
ms(3)nc3 fails to complement class I alleles at the
ß2 tubulin locus, it is possible that Mst77F
has a dual role to play as a chromatin condensing protein in the nucleus and for
the normal nuclear shaping. Nuclear shaping is a microtubule-based event.
ms(3)nc3 leds to a tid-shaped nuclear
phenotype, where the nucleus fails to shape into a needle-like nuclei. Similar
defective nuclear shaping is seen with the few homozygous and heteroallelic
combinations of class I alleles of ß2 tubulin. The
incorporation of the defective subunit encoded by ms(3)nc3
may interfere with the function of the resulting complex. These data suggest the
involvement of an Mst77F (a linker histone variant) in the microtubule dynamics
during the nuclear shaping. This again complements the role of sea urchin
histone H1 in the stabilization of flagellar microtubules (Raja, 2005).
After the first steps in the fertilization process, the male gamete is still in
the highly compact protamine-based chromatin structure. In a wild-type egg, the
paternal pronucleus changes the shape from the needle-like to a spherical
structure. Furthermore, the male pronucleus acquires a nucleosome-based
structure before zygote formation and thus is transformed into a
replication-competent male pronucleus. sesame is a maternal effect
mutation in HIRA and had been mapped to 7C1.
HIRA family of genes (named afteryeast HIR genes; HIR is an acronym for
'histone regulator') includes the yeast HIR1 and HIR2 repressors of histone gene
transcription in S. cerevisiae/em>, human TUPLE-1/HIRA, chicken HIRA, and mouse HIRA. In
Drosophila, HIRA is expressed in the female germ line and a high level of
HIRA mRNA is deposited in the egg. Human HIRA is
shown to bind to histone H2B and H4. The WD repeats
present at the N-terminal part of HIRA could probably function as a part of a
multiprotein complex. Xenopus HIRA proteins are
also known in promoting chromatin assembly that is independent of DNA synthesis
in vitro. The corresponding maternal effect mutant sesame, in which the sperm fertilizes the egg but no zygote is formed, has been analyzed. Although the shape change of the nucleus to the spherical structure occurs in these mutants, maternal histones are not incorporated into the male pronucleus, which
strengthens the function of HIRA in binding to the core histones. This study shows
that neither Drosophila protamine is removed from the male pronucleus
in sesame mutants. This leads to the proposal that the transport and
incorporation of histones onto the chromatin in some manner is coupled to the
removal of protamines in which HIRA could play an important role in the
multiprotein complex required in this chromatin reconstitution process.
Mst77F removal from the male pronuceus in contrast to protamines
is independent of HIRA (Raja, 2005).
During spermiogenesis, chromatin reorganization of the complete genome is an
essential feature for male fertility. This process leads to an extremely
condensed state of the haploid genome in the sperm and requires a reorganization
of the paternal genome in the male pronucleus during fertilization and before
zygote formation. With the characterization of the chromatin condensing proteins
in Drosophila, it would be possible to gain more insight into the
mechanisms of sperm chromatin reorganization during spermiogenesis and
fertilization (Raja, 2005).
Nuclear DNA in the male gamete of sexually reproducing animals is organized as sperm chromatin compacted primarily by sperm-specific protamines. Fertilization leads to sperm chromatin remodeling, during which protamines are expelled and replaced by histones. Despite increased understanding of the factors that mediate nucleosome assembly in the nascent male pronucleus, the machinery for protamine removal remains largely unknown. This study identified four Drosophila protamine chaperones that mediate the dissociation of protamine-DNA complexes: NAP-1, NLP, and nucleophosmin are previously characterized histone chaperones, and TAP/p32 has no known function in chromatin metabolism. This study showed TAP/p32 to be required for the removal of Drosophila protamine B in vitro, whereas NAP-1, NLP, and Nph share roles in the removal of protamine A. Embryos from P32-null females show defective formation of the male pronucleus in vivo. TAP/p32, similar to NAP-1, NLP, and Nph, facilitates nucleosome assembly in vitro and is therefore a histone chaperone. Furthermore, mutants of P32, Nlp, and Nph exhibit synthetic-lethal genetic interactions. In summary, this study identified factors mediating protamine removal from DNA and reconstituted in a defined system the process of sperm chromatin remodeling that exchanges protamines for histones to form the nucleosome-based chromatin characteristic of somatic cells (Emelyanov, 2014).
The DNA of metazoan somatic cells is packaged into a compact nucleoprotein complex termed chromatin. Chromatin fiber is comprised of highly conserved repetitive units (nucleosomes) that contain an octamer of four core histones and 145-147 base pairs (bp) of DNA wrapped around the octamer in 1.65 turns of a left-handed superhelix. Nucleosomes are assembled in vivo in an ATP-dependent fashion through a concerted and sequential action of core histone chaperones and motor proteins that belong to the Snf2 family of DNA-dependent ATPases. For instance, Drosophila ACF/CHRAC can mediate chromatin assembly in conjunction with histone chaperone NAP-1. Other known ATP-dependent chromatin assembly factors include RSF, CHD1, ATRX, and ToRC/NoRC (Emelyanov, 2014).
The most abundant chromatin component in male germline cells is protamines-small positively charged arginine- and cysteine-rich protein. During spermiogenesis, protamines replace 85%-95% of DNA-bound histones in the nucleus to achieve a higher density of sperm nuclear DNA. Crystalline-like sperm chromatin structure is sixfold more compact than metaphase chromosomes and renders sperm DNA enzymatically inert. At fertilization, the oocyte remodels the condensed sperm chromatin into a transcriptionally competent chromatin of the male pronucleus. During this process, protamines are expelled and replaced with oocyte-supplied histones, which are then organized into nucleosomes. Sperm chromatin remodeling (SCR) is controlled by biochemical activities in the early oocyte, but components of these activities remain largely unknown. However various protein factors have been implicated in SCR, including core histone chaperones from Xenopus and Drosophila (NAP-1, p22, DF31, HIRA, and Yemanuclein). In mammals, members of nucleoplasmin/nucleophosmin family proteins (NPM1-3) function in sperm chromatin decondensation in vitro. In addition, Npm2 knockout female mice exhibit fertility defects consistent with a role of NPM2 in nuclear and nucleolar chromatin organization. It was also suggested that sperm chromatin decondensation is ATP-dependent (Emelyanov, 2014).
Drosophila sperm cells contain two major protamines (A and B) encoded by male-specific transcripts Mst35Ba and Mst35Bb, respectively. The Drosophila maternal effect mutant sésame (ssm) prevents male pronucleus formation. ssm encodes the histone variant H3.3-specific chaperone HIRA, postulated to be required for replication-independent deposition of histones in the male pronucleus during sperm decondensation . In eggs from homozygous ssm females, maternal histones are not deposited in the chromatin of male pronuclei, preventing normal mitosis and resulting in the development of gynogenetic haploid embryos and embryonic stage lethality. A similar phenotype is observed in null mutants of the gene encoding ATP-dependent chromatin assembly factor CHD1. Thus, CHD1 and HIRA act cooperatively and are required for nucleosome assembly during SCR. Intriguingly, protamines are efficiently expelled from the DNA of nascent male pronuclei in Chd1 and ssm eggs, suggesting that protamine removal and histone deposition are functionally distinct steps (Emelyanov, 2014).
This study used a biochemical approach to identify specific protein components of the Drosophila egg machinery that promote the dissociation of protamine-DNA complexes of sperm chromatin. These factors turn out to be two known core histone chaperones (NAP-1 and NLP), a homolog of mammalian nucleophosmin, and a novel Drosophila histone chaperone (TAP/p32). These putative 'protamine chaperones' facilitate SCR independently of CHD1 and HIRA, which mediate nucleosome assembly in nascent male pronuclei. Of note, TAP/p32 is specifically required to expel Drosophila protamine B from sperm chromatin in vitro, whereas NAP-1, NLP, and nucleophosmin share roles in removal of protamine A. In vivo evidence is provided that TAP/p32 functions in Drosophila egg SCR. In conclusion, this study has characterized protein factors that mediate the first obligatory step of SCR (protamine dissociation) and reconstituted the complete SCR reaction (reorganization of protamine-containing sperm chromatin into core histone-containing nucleosome arrays) in a purified defined system in vitro (Emelyanov, 2014).
Although recent studies provide details of sperm chromatin composition (Hammoud 2009; Miller 2010) and assembly during spermiogenesis, relatively little is known about the protein machinery that mediates SCR during fertilization. In vivo analyses in Drosophila suggest that removal of protamines from sperm chromatin is biochemically uncoupled from subsequent nucleosome assembly because male pronucleus-specific nucleosome assembly factors CHD1 and HIRA are not required for protamine removal. This study demonstrates that, indeed, a separate set of protein factors (protamine chaperones) is required for protamine eviction. Using assay-based biochemical approaches, four Drosophila proteins were identified that are sufficient for unraveling of DNA-protamine complexes in vitro. Their biochemical activities and mechanisms of SCR were analyzed. Significantly, it was possible to recapitulate the entire process of SCR (protamine eviction and nucleosome assembly) in a defined purified system. Biological functions of TAP/p32 were further analyzed in Drosophila, and evidence was obtained of its proposed roles in SCR in vivo. Finally, it was discovered that all four proteins additionally share a function as core histone chaperones (Emelyanov, 2014).
Recent microarray analysis of mating-responsive genes in Drosophila revealed that CG6459/P32 expression is strongly activated in the female lower reproductive tract within 6 h of mating. In fact, CG6459/P32 exhibits the strongest response of all genes identified in the study. The up-regulation is transient and is reversed 24 h after mating. This temporal expression pattern of TAP/p32 further supports its proposed role during fertilization (Emelyanov, 2014).
Intriguingly, orthologs of protamine chaperones are expressed in unicellular organisms, such as S. cerevisiae, which do not express protamines and whose gametes do not undergo the chromatin reorganization characteristic of metazoan sperm cells. Evidence is provided that Mam33p, an S. cerevisiae ortholog of TAP/p32, is involved in chromatin remodeling and DNA compaction and/or repair. It is likely that protamine chaperone homologs in unicellular eukaryotes perform conserved functions of core histone chaperones and are involved in nucleosome assembly and remodeling. During metazoan evolution, however, their biochemical activities may have been harnessed for SCR owing to biochemical similarities (net charge and amino acid composition) and functional roles (DNA compaction) of histones and protamines (Emelyanov, 2014).
Although yeast TAP/p32 ortholog Mam33p is clearly involved in regulation of DNA integrity/repair in response to treatment with mutagens and genetically interacts with factors of chromatin remodeling and DNA compaction and repair, it is also required for metabolism of alternative carbon sources. Similarly, mammalian TAP/p32 has also been implicated in mitochondrial function. Furthermore, Drosophila TAP/p32 and NAP-1, although subject to nuclear translocation, are also efficiently recruited to mitochondria. Hence, it is interesting to consider the apparent dual role of TAP/p32 in mitochondrial function and nuclear DNA compaction. Mature sperm cells in Drosophila and vertebrates contain a stack of mitochondrial structures in the midpiece at the junction of the head and tail. (In Drosophila, the sperm mitochondria are depleted of DNA, and, in most metazoan species during fertilization, sperm mitochondria undergo rapid ubiquitination and degradation by autophagocytosis) Thus, the elevated affinity of TAP/p32 and NAP-1 to protein components of mitochondria may be used and adapted for rapid and specific recruitment of the TAP/p32 and NAP-1 to the sperm head, which would facilitate their loading onto sperm chromatin for its processing (Emelyanov, 2014).
It has been suggested that Xenopus nucleoplasmin is sufficient for the initial stage of SCR (decondensation of demembranated sperm and removal of sperm basic proteins SP1-6 in vitro). However, the removal of sperm proteins (and their replacement by histones) in the presence of nucleoplasmin does not appear complete/quantitative. Furthermore, the Xenopus sperm decondensation assay is prone to artifacts: It is frequently performed (and works) with heterologous proteins and extracts, including those from yeast. In contrast, the current analyses suggest that a family of several factors may share partially redundant roles in protamine removal, and their cooperative action is necessary and sufficient for complete protamine eviction from sperm chromatin substrates. On the other hand, considering poor evolutionary conservation of protamine number and identities, it is possible that species other than Drosophila use smaller or larger sets of factors for SCR (Emelyanov, 2014).
Despite being ~94% identical, protamines A and B require different chaperones for their removal. For instance, in the absence of TAP/p32, a mixture of NAP-1, NLP, and Nph is incapable of protamine B eviction. Protamine polypeptides are extremely evolutionarily divergent. In fact, it is rarely possible to assign a protamine function based on a sequence conservation search of related proteins in distinct metazoan species. For example, a closely related organism, Drosophila simulans, expresses one protein homologous to D. melanogaster protamines. It is more closely related to protamine B and shares with it only 77% identity. D. simulans also express orthologs of protamine chaperones. A high degree of functional/sequence specificity makes it unlikely that D. melanogaster protamine chaperones will be able to remodel MSC assembled from more divergent, evolutionarily distant protamines. This specificity may contribute to gametic isolation of distinct species. In the future, it will be interesting to analyze cross-reactivity of protamines and protamine chaperones from these species in MSC remodeling in vitro and in vivo (Emelyanov, 2014).
The Drosophila modulatory calcineurin-interacting protein (MCIP) sarah (sra) is essential for meiotic progression in oocytes. Activation of mature oocytes initiates development by releasing the prior arrest of female meiosis, degrading certain maternal mRNAs while initiating the translation of others, and modifying egg coverings. In vertebrates and marine invertebrates, the fertilizing sperm triggers activation events through a rise in free calcium within the egg. In insects, egg activation occurs independently of sperm and is instead triggered by passage of the egg through the female reproductive tract; it is unknown whether calcium signaling is involved. MCIPs [also termed regulators of calcineurin (RCNs), calcipressins, or DSCR1 (Down syndrome critical region 1)] are highly conserved regulators of calcineurin, a Ca2+/calmodulin-dependent protein phosphatase 1 and 2. Although overexpression experiments in several organisms have revealed that MCIPs inhibit calcineurin activity, their in vivo functions remain unclear. Eggs from sra null mothers are arrested at anaphase of meiosis I. This phenotype was due to loss of function of sra specifically in the female germline. Sra is physically associated with the catalytic subunit of calcineurin, and its overexpression suppresses the phenotypes caused by constitutively activated calcineurin, such as rough eye or loss of wing veins. Hyperactivation of calcineurin signaling in the germline cells resulted in a meiotic-arrestphenotype, which can also be suppressed by overexpression of Sra. All these results support the hypothesis that Sra regulates female meiosis by controlling calcineurin activity in the germline. This is the first unambiguous demonstration that the regulation of calcineurin signaling by MCIPs plays a critical role in a defined biological process (Takeo, 2006; Horner, 2006).
sarah mutation disrupts several aspects of egg activation in Drosophila. Eggs laid by sarah mutant females arrest in anaphase of meiosis I and fail to fully polyadenylate and translate bicoid mRNA. Furthermore, sarah mutant eggs show elevated cyclin B levels, indicating a failure to inactivate M-phase promoting factor (MPF). Taken together, these results demonstrate that calcium signaling is involved in Drosophila egg activation and suggest a molecular mechanism for the sarah phenotype. The conversion of the sperm nucleus into a functional male pronucleus is compromised in sarah mutant eggs, indicating that the Drosophila egg's competence to support male pronuclear maturation is acquired during activation. Despite its independence from a sperm trigger, egg activation in Drosophila involves calcium-mediated pathways that are likely to be analogous to those in other animals. It is intriguing that among these downstream events is the acquisition of the egg's competence to remodel the sperm nucleus into the male pronucleus (Horner, 2006).
To explore the in vivo function of the Drosophila MCIP Sra, a null mutation in this locus was created by gene targeting. Homozygotes of the null allele sraKO are semilethal during larval or pupal stages. In addition, sraKO females were sterile, and their ovulation is abnormal (Ejima, 2004). These phenotypes were rescued by either sarah transgenes. Taken together, these results unambiguously demonstrate that sra is responsible for the phenotypes associated with sraKO, which include developmental defects in both sexes and ovulation and sterility in females (Takeo, 2006).
There is no apparent morphological abnormality in ovarian development in sraKO females, but eggs from sraKO mothers failed to hatch. Wild-type eggs at 2 hr after deposition already have completed meiosis and undergo synchronous mitotic nuclear division. In contrast, eggs—hereafter referred to as sra eggs—from sraKO mothers had a localized DAPI-stained signal in the cortical region near the anterior pole, indicating that sra eggs are arrested during meiosis. To analyze this phenotype in detail, the pattern of chromosome segregation and the spindle shape of sra eggs were examined. Spindle microtubules were visualized with tubulin antibody staining. In wild-type females, mature oocytes are arrested at metaphase of meiosis I, during which the chromosomes are seen as a large mass of chromatin. After the release of meiotic arrest during ovulation, individual chromosome arms become visible and migrate toward the poles; the chromosomes subsequently undergo meiosis II, after which nuclear fusion and the mitotic divisions of the zygote take place. In sra eggs, the meiotic chromosomes were seen in between the metaphase plate and the poles. It was confirmed that the oocytes taken from sra mutants including sraGS3080 and sraGS3168 are arrested at metaphase I as in the wild-type. Therefore, sra eggs are arrested at anaphase I shortly after the meiotic resumption from the metaphase I arrest (Takeo, 2006).
To determine whether the meiotic-arrest phenotype of sra eggs is caused by loss of function in the germline or somatic cells of sra mothers, mutant germline clones were generated by the flippase-dominant female sterile (FLP-DFS) technique. All eggs laid by wild-type females completed meiotic divisions, whereas most eggs (98%) from sra mothers arrested at anaphase of meiosis I. sra germline clones were also arrested at anaphase I, reproducing the phenotype of sraKO. A few sra eggs, including germline clones, were arrested at meiosis II. In these eggs, the two spindles were perpendicular to each other, rather than in tandem as in the wild-type. These results demonstrate that the meiotic defects in sra eggs are specifically attributable to the loss of function of sra in the female germline. Consistent with this conclusion, sra is highly expressed in the female germline during oogenesis (Ejima, 2004) and in early embryos. Furthermore, the meiotic-arrest phenotype caused by sra mutations was almost fully rescued by nos-GAL4/UASp-sra transgenes. These results establish that sra is required in the germline for meiotic progression in Drosophila females (Takeo, 2006).
Sra is a Drosophila member of the modulatory calcineurin-interacting protein (MCIP) family of proteins, which are known to function as endogenous regulators of calcineurin. Calcineurin consists of two subunits, CnA and CnB. The Drosophila genome contains three genes encoding CnA subunits (CanA1, Pp2B-14D, and CanA-14F) and two genes encoding CnB subunits (CanB and CanB2). The functions of calcineurin have been poorly analyzed in Drosophila. It is hypothesized that sra functions as an endogenous regulator of calcineurin in Drosophila. Analyses of the expression pattern of Drosophila calcineurin genes by RT-PCR revealed that all three CnA and both CnB genes were expressed in larvae and adult females, but among these, only Pp2B-14D, CanA-14F, and CanB2 are expressed in early embryos and ovaries. Therefore, these three calcineurin subunits are candidates for interacting with Sra in the female germline (Takeo, 2006).
A constitutively active form of calcineurin can be created by truncating the C-terminal part of CnA (Sullivan, 2002; Gajewski, 2003). Misexpression of the active form of Pp2B-14D (Pp2B-14Dact) causes morphological abnormalities in eyes and wings (Sullivan, 2002). To determine the effects of sra on calcineurin signaling, whether activated calcineurin-dependent phenotypes can be modified by coexpression of sra was tested. Overexpression of sra alone in developing eyes by using GMR-GAL4 did not induce any phenotypic change. Flies misexpressing Pp2B-14Dact showed a mild rough-eye phenotype, which was completely suppressed by coexpression of sra. Similarly, misexpression of Pp2B-14Dact in the posterior compartment of developing imaginal discs by using en-GAL4 resulted in loss of wing veins and reduction of wing size. These wing phenotypes were also completely suppressed by coexpression of sra, whereas overexpression of sra alone had no effect on wing morphology. Furthermore, overexpression of sra rescued the lethality induced by the muscle-specific expression of Pp2B-14Dact by using 24B-GAL4. All these results clearly show that sra has an inhibitory effect on calcineurin signaling (Takeo, 2006).
If Sra acts as an inhibitor of calcineurin in vivo, it was speculated that calcineurin signaling is hyperactivated in sra mutants; that is, hyperactivation of calcineurin signaling might also affect the meiotic phenotype as in sra mutants. It was found that females carrying nos-GAL4 and UASp-Pp2B-14Dact had fully developed ovaries, but were sterile or semisterile, depending on the transgenic lines. The sterility or semisterility caused by nos>Pp2B-14Dact was effectively rescued by co-overexpression of sra, demonstrating that sra counteracts activated calcineurin (Takeo, 2006).
To characterize the meiotic phenotype caused by Pp2B-14Dact, eggs were stained from semisterile females expressing nos>Pp2B-14Dact with DAPI and tubulin antibody to visualize chromosomes and spindles, respectively. A total of 63 eggs were examined. Of these, 10% (6/63) developed normally, whereas 14% (9/63) had neither DAPI signaling nor tubulin antibody staining. The remaining 76% showed complex abnormalities that could be classified into three types: (1) dispersed chromatins with no obvious spindle (33%); (2) apparently normal chromosomes with an abnormal spindle (38%), and (3) a mass of chromatin with an apparently normal spindle (5%). Also the nuclei of mature oocytes taken from nos>Pp2B-14Dact females were observed to see whether meiotic arrest at metaphase I is normal. It was found that the majority had abnormal nuclei containing dispersed chromatins, and that the remaining were arrested at metaphase I or anaphase I. These results suggest that calcineurin signaling was activated to a greater extent in the germline of females constructed in this way than in sra mutants. Taken together, these results demonstrate that the regulation of calcineurin signaling is critical for female meiosis, and its regulator Sra/MCIP is essential for meiotic progression at the time of egg activation in the Drosophila female (Takeo, 2006).
In vertebrates whose meiotic arrest occurs at metaphase II, arrest is released at the time of fertilization. The mechanisms of meiotic arrest and resumption have been extensively studied in mice and frogs, and several key components have been identified, including Cdc2/Cyclin B (MPF) and MAP kinase. In addition, the so-called “Ca2+ transient” mediated by IP3 signaling has been linked to egg activation; this transient promotes completion of meiosis, ion-channel opening, and cortical granule exocytosis. Recent studies have revealed that Ca2+/calmoduin-dependent protein kinase II (CaMKII) is physiologically activated in mouse oocytes in response to fertilizing sperm. CaMKII is implicated in the regulation of the timing of re-entry into mitosis through the phosphorylation of Cdc25C, a phosphatase mediating G1/M transition by dephosphorylating MPF in Xenopus. More recently, CaMKII was shown to phosphorylate an anaphase-promoting complex/cyclosome (APC/C) inhibitor, Emi1-related protein (Erp1), resulting in its degradation and thereby releasing the brakes on the cell cycle from metaphase II in Xenopus eggs (Takeo, 2006).
Less is known about the mechanism of egg activation in Drosophila. Genetic screens for female-sterile mutations have identified several genes involved in female meiosis. For example, twine, a homolog of Drosophila cdc25, is required for arrest at metaphase I in mature oocytes. cortex (cort) and grauzone (grau) mutant eggs exhibit meiotic arrest at meiosis II with defects in cytoplasmic polyadenylation and translation of maternal bicoid mRNAs. cort encodes an APC/C activator protein Cdc20, suggesting that APC/C-Cdc20Cort-mediated inactivation of MPF is required for the translational control of poly(A)-dependent maternal mRNA. grau encodes a member of the C2H2-type zinc-finger protein family and activates transcription of cort to induce the completion of female meiosis. In addition, a recent study reported that a small cell-cycle regulator, Cks30A, plays an essential role in meiotic progression by associating with Cdk1 (Cdc2)/cyclin complexes and mediating Cyclin A degradation in the female germline. Therefore, cell-cycle regulators involved in meiotic progression are likely to be conserved between vertebrates and Drosophila (Takeo, 2006).
The involvement of calcineurin signaling in female meiosis has not previously been described in any organism. These studies on sra are the first to demonstrate that regulation of a Ca2+-dependent phosphatase is critical for the progression of female meiosis. Analyses of mutations in calcineurin genes and identification of the substrates in germline cells should facilitate further understanding of the role of calcineurin signaling in female meiosis (Takeo, 2006).
In the Drosophila oocyte, meiosis is arrested in the first division of metaphase: at this stage a tapered spindle forms, aligned parallel to the egg surface. The chromosomes are therefore located in the cortical region near the anterior pole, whereas fusion of parental complements occurs in the inner ooplasm. How does the female pronucleus reach the interior of the egg? The second meiotic spindles are arranged in tandem, end to end, and disposed perpendicular to the longitudinal axis of the egg with the innermost spindle carrying the female pronucleus. This pattern of spindle organization is probably involved in the migration of the female pronucleus deeper into the egg near the cytoplasmic domain of the male pronucleus. The precise time at which the mitotic spindle of Drosophila changes orientation is unknown. However, spindle rotation from a position parallel to the egg surface to a radial orientation presumably occurs during or shortly after the oocyte passes through the oviduct. How spindle orientation is achieved and maintained during meiosis is an intriguing question. Microtubules linking spindle poles to the oocyte surface have been implicted in the rotation and anchoring of the meiotic apparatus in Xenopus oocytes and in other organisms, but this does not seem to be the case in the Drosophila oocyte, since the meiotic spindles lack astral microtubels. However, the observation that a transient array of microtubules links the meiotic apparatus to discrete subcortical foci suggests that in Drosophila the orientation of the spindle also requires a functional interaction between the spindle and the oocyte cortex (Riparbelli, 1996 and references).
>The microtubule array of mitosis II observed between the twin spindles at metaphase, anaphase and telophase might be an intermediate between the anastral poles of the meiotic I spindles and the astral poles of the mitotic spindles in early embryos. A complex pathway of spindle assembly takes place during resumption of meiosis at fertilization, consisting of a transient array of microtubules radiating from the equatorial region of the spindle toward discrete foci in the egg cortex. A monastral array of microtubules is observed between twin metaphase II spindles in fertilized eggs. These microtubules originating from disc-shaped material stain with Rb188 antibody specific for an antigen asssociated with the centrosome of Drosophila embryos (DMAP190 or CP190). Therefore, the Drosophila egg contains a maternal pool of centrosomal components undetectable in mature inactivated oocytes. These components nucleate microtubues in a monastral array after activation, but are unable to organize bipolar spindles (Riparbelli, 1996).
The meiosis II spindle of Drosophila oocytes is distinctive in structure, consisting of two tandem spindles with anastral distal poles and an aster-associated spindle pole body between the central poles. Assembly of the
anastral:astral meiosis II spindle occurs by reorganization of the meiosis I spindle, without breakdown of the meiosis I
spindle. The unusual disc- or ring-shaped central spindle pole body forms de novo in the center of the elongated
meiosis I spindle, followed by formation of the central spindle poles. gamma-Tubulin transiently localizes to the
central spindle pole body, implying that the body acts as a microtubule nucleating center for assembly of the central
poles. The first step in formation of the central pole body is the appearance of puckers in the center of the the meiosis I spindle, followed by the pinching out from the spindle of a disc or ring of microtubules that becomes the central pole body. The manner in which the central spindle pole body forms suggests the involvement of a microtubule motor. If so, the motor involved is likely to be different from Ncd (Nonclaret disjunctional), since loss of Ncd function does not seem to prevent its formation. Following the formation of the central spindle pole body, the microtubules arrayed to either side of the central body narrow into poles, forming the mature meiosis II spindle. The central poles become more tapered during progression through meiosis II, and the central spindle pole body also changes in morphology: the disc or ring becomes asterlike, then enlarges into a ring that lies between the two central telophase II nuclei (Endow, 1998).
Localization of gamma-tubulin to the meiosis II spindle is dependent on the microtubule motor protein,
Ncd. Absence of Ncd results in loss of gamma-tubulin localization to the spindle and
destabilization of microtubules in the central region of the spindle. Likewise, during meiosis I, the minus-end motility of Ncd and its crosslinking activity are probably needed to focus microtubules into spindle poles for the correct functioning of meiosis I. Assembly of the anastral:astral meiosis II spindle
probably involves rapid reassortment of microtubule plus and minus ends in the center of the meiosis I spindle. This
can be accounted for by a model that also accounts for the loss of gamma-tubulin localization to the spindle and
destabilization of microtubules in the absence of Ncd (Endow, 1998).
Evidence is provided that a distinct midzone is present in the Drosophila melanogaster female meiosis I spindle. This region has the ability to bind the Pavarotti kinesin-like (PAV-KLP) and Abnormal spindle (Asp) proteins, indicating a correct organization of the central spindle microtubules. The core component centrosomal protein centrosomin (CNN) has been identified at an unexpected site within the anaphase I spindle, indicating a role for CNN during the biogenesis of the female meiotic apparatus. However, there are no apparent defects in the midzone organization of cnn oocytes, whereas defects occur later when the central aster forms. The primary mutant phenotype of cnn oocytes is the failure to form a developed central microtubule organizing center (MTOC), although twin meiosis II spindles usually do form. Thus the central MTOC may not be essential for the formation of the inner poles of twin meiosis II spindles, as generally proposed, but it might be involved in maintaining their proper spacing. The proposal is discussed that, in the presence of a central MTOC, a chromatin-driven mechanism of spindle assembly like that described during meiosis I may control the morphogenesis of the twin meiosis II spindles (Riparbelli, 2005).
Eighty years ago, the pioneering studies of Huettner (Huettner, 1924) on the maturation of the Drosophila melanogaster oocyte revealed that chromosome segregation during the first meiosis is supported by a peculiar spindle apparatus that transforms during the second meiosis into twin spindles arranged in tandem and disposed perpendicularly to the longitudinal axis of the egg. These spindles ensure the reductional divisions and the formation of the haploid complements, the innermost of which is the female pronucleus. Huettner first observed that the meiotic spindles are anastral and lack centrioles at their poles, though they are tapered. Despite the potential of these observations for clarifying acentrosomal pathways of microtubule (MT) organization, the female meiotic apparatus of Drosophila has received little attention for many years. During the last decade some researches have concentrated their attention on the morphogenesis and organization of the spindle during metaphase of the first meiotic division. Immunohistochemical analysis failed to reveal centrosomal components, such as CP60, CP190 and gamma-tubulin, at the spindle poles, confirming, in its essential aspects, the general description of Huettner. The role of gamma-tubulin during meiosis is, however, controversial since mutants for this gene have abnormal first meiotic spindles, but meiosis is not terminally arrested and appears mostly normal. Several lines of evidence suggest that meiotic spindle assembly in the Drosophila female begins with a chromosome-driven mechanism of MT organization. Mutational analysis indicates that, in the absence of centrosomes, MT bundling and bipolarity of anastral meiotic spindles requires kinesin-like proteins with minus-end-directed MT motor activity. Defects in the genes nonclaret disjunctional (ncd) and subito (sub), which encode kinesin-like proteins, lead to the formation of abnormal meiotic I spindles with frayed or undefined poles. The cooperative interaction between motor proteins and the products of the genes mini spindles (msps) and transforming acidic coiled-coil protein (tacc), localized at the acentrosomal poles of the first meiotic spindle, might be crucial for the bipolarity of the meiotic spindle. The MT minus-end-associated Abnormal spindle (Asp) protein, localized at the extremities of the meiotic spindle, could be also involved in the stabilization of focused poles (Riparbelli, 2005 and references therein).
Oocytes are activated to resume meiosis by passage through the oviduct. The spindle, positioned parallel to the cortex during metaphase arrest, reorients perpendicular to the oocyte surface during completion of meiosis I. The spindle elongates during anaphase-telophase of the first meiosis and transforms during meiosis II into two tandemly arranged spindles (Huettner, 1924). Although the meiotic II spindles are anastral, an unusual structure, from which an extensive array of MTs nucleates, forms between the two internal spindle poles (Riparbelli, 1996). It has been shown that this structure, in contrast to the poles of the meiotic spindles, contains several centrosomal proteins usually found at the mitotic spindle poles, such as gamma-tubulin, CP60 and CP190 and centrosomin (CNN) (Llamazares, 1999). However, although this central MTOC contains centrosomal components and is able to nucleate an astral array of MTs, it lacks centrioles, is unable to duplicate and disappears after the completion of meiosis (Riparbelli, 2005).
Whereas previous analyses provided insight into the mechanisms of spindle assembly and chromosome segregation during metaphase of the first meiosis, they provided no clear information on the kinetics of spindle morphogenesis during transition from meiosis I to II. This aspect was investigated by time-lapse analysis of ncd-gfp oocytes (Endow, 1998). The main event of the transforming of meiotic I spindle in twin tandem arranged meiotic II spindles is the formation in the elongated meiotic I spindle of lateral puckers, which are correlated with the organization of the central MTOC (Endow, 1998). However, despite the usefulness of this elegant in vivo approach, some aspects, such as the individualization of the twin spindles and the origin and function of the central MTOC, remain unclear. The findings presented in this report both expand and clarify earlier studies on the structural organization and dynamics of the female meiotic spindle, suggesting an alternative model of meiosis II spindle assembly (Riparbelli, 2005).
After oocyte activation, meiosis resumes and the metaphase-arrested spindle elongates and undergoes a pivoting movement to reorient perpendicularly to the surface (Endow, 1997). Two distinct populations of interpolar MTs, interior and peripheral, form a top-like anaphase spindle. The interior MTs run parallel to the main axis of the spindle to form longitudinal thick bundles, whereas the peripheral ones spread out from the poles to form opposite cone-shaped half spindles. The peripheral MTs, which form a cage around the interior bundles, intersect at the equatorial plane of the spindle and extend both deeply into and above this plane until they reach the cell cortex. Thus, the spindle appears to be anchored to the oocyte surface through the peripheral MT set that forms the inner half spindle. Some clusters of MTs appear through the equatorial region of the spindle where opposite MTs overlapped and/or interacted tangentially. As anaphase progresses the clustering of MTs is more evident through the spindle equator. Most of the interior MT bundles terminate in a small non-fluorescing area at the opposite poles. Co-labeling with propidium iodide indicated that these regions of lower MT density correspond to the chromosomes and that the bundles might therefore be kinetochore fibers. The kinetochore MTs of different chromosomes form separated bundles and do not focus on a common pole. Thus the spindle poles appear broad and frayed. Although the density of the MTs made it difficult to resolve the structure of the central region of the spindle, suitable optical sections showed that some MT bundles overlap in this region. During telophase a diffuse tubulin accumulation was observed at the middle of the spindle at the sites where the interior MTs overlap (Riparbelli, 2005).
Transition from meiosis I to meiosis II is marked by the expansion of the cage of MTs that surround the interior interpolar MTs to form an extensive network. Some of the peripheral MTs also contact the oocyte cortex. Astral arrays of MTs of various size become apparent at this time within the equator of the spindle and outside its boundaries. Intermediate to the formation of these structures might be the clusters of overlapping MTs observed during the first meiosis. These clusters become increasingly compact at the onset of second meiosis and could become centers from which MTs spread. A distinct tubulin aggregate was seen to bulge out at the middle of the interior MT bundles. Kinetochore MTs run almost parallel in each half spindle both outward and inwards to each chromosome set they hold. Thus, kinetochore fibers form distinct parallel 'minispindles' and the outer poles appear frayed. The inner poles cannot be distinguished at this time. Optical sections midway along the spindle show an irregular belt-shaped equatorial tubulin aggregate that could be derived from the tubulin accumulation observed during telophase of the first meiosis. Several MTs that span from this aggregate end at the cytoplasmic asters or mingle within the peripheral network (Riparbelli, 2005).
As meiosis progresses the peripheral MT array gradually disassembles and the MT asters become free in the cytoplasm or remain connected to the remnant of the peripheral MT framework or to the spindle by thin threads and the equatorial tubulin cluster is more evident and protruding. Free asters are transient structures that disappear over time and are no longer visible from anaphase II on. The outward extremities of the minispindles formed by the kinetochore fibers focus on common outer poles, whereas the inner kinetochore fibers still run almost parallel. Thus, the individualization of the meiosis II twin spindles occurs gradually and requires the formation of the internal poles that start to be organized during late prophase/early prometaphase when the inwards extremities of the minispindles have partially coalesced midway along the spindle. Short MTs form a loose cloud at the spindle equator. During prometaphase/early metaphase, the twin spindles begin to be distinguishable, though their inner poles are formed by three to four distinct bundles and appear broad. The MTs at the spindle equator are more prominent and form an extensive astral array, the 'central aster', which increases in size during subsequent metaphase and anaphase. At telophase the MTs of the central aster reach their maximum length, although they have decreased in density (Riparbelli, 2005).
Although female meiosis in Drosophila differs from male meiosis and mitosis in that spindle poles lack centrosomes and cytokinesis does not occur, this study demonstrates that the central region of the spindle has a common organization in these systems and that a structured midzone exists in the female meiotic spindle. At least two main events are involved in the formation of the spindle midzone in Drosophila cells (Inoue, 2004): the overlay of antiparallel MTs and the release of a subpopulation of MTs from the spindle poles. The central region of the female meiosis spindle during late anaphase I has the ability to bind PAV-KLP, the Drosophila ortholog of MKLP. This protein has been found in association with the putative MT plus ends at the midzone of anaphase spindles of mitotic cells and spermatocytes where it might function in regulating the dynamics and organization of the overlapping MTs. Thus the female meiotic spindle could have a structured midzone formed by overlapping antiparallel MTs. This is consistent with the finding, from sagittal optical sections, that opposite MT bundles overlap at the middle of the central spindle. The increased density of MTs in the central region of the spindle at telophase of the first meiosis might be consistent with the release of a subset of MTs from the opposite spindle poles. When this process occurs in mitotic cells and spermatocytes (Riparbelli, 2002), the Asp protein accumulates at the minus ends of the central spindle MTs. The Asp staining observed in the central region of the spindle during telophase of the first meiosis could be such a protein localization, at the predicted MT minus ends, suggesting that MTs of the central spindle undergo similar dynamics during female meiosis and in both mitosis and male meiosis (Riparbelli, 2005).
Morphological evidence indicates that several differences exist between the meiosis I and II spindles: the meiosis I spindle is anastral, whereas the twin meiosis II spindles have outer anastral poles and an unusual central aster between the inner poles (Riparbelli, 1996; Endow, 1997). These observations suggest that spindle assembly could require different mechanisms during meiosis I and II. Whereas the organization of the meiosis I spindle involves a chromosomal-dependent pathway of MT organization, the individualization of the twin spindles during meiosis II requires the formation of new poles in the center of the prophase II spindle. However, the mechanism by which the inner poles form is unclear. This issue has been difficult to address because the formation of the inner spindle poles occurs in a region of very high MT density, and thus any stage of this process is obscured from view. It has been proposed that the formation of the inner poles might require the reorganization of the central spindle in which the polarity of MT ends reverse (Endow, 1998). This hypothesis is supported by the presence of an unusual MTOC that has been postulated to play a major role in the process of MT nucleation for the formation of the inner half spindles during meiosis II (reviewed by Megraw, 2000). However, normal-looking twin bipolar spindles can form during meiosis II, both when the central aster is defective as in polo mutants (Riparbelli, 2000) and when it is very reduced as in wispy, KLP3A and cnn mutants (Riparbelli, 2005).
A new model of spindle assembly during meiosis II is suggested. Spindle elongation during anaphase/telophase of the first meiosis moves the homologous chromosomes to opposite poles that appear broad and frayed because the kinetochore fibers of different chromosomes do not focus on a common pole during later meiosis I stages. Starting from late prophase II, two opposite half spindles become evident, with outer focused poles where Asp accumulates. By contrast, inner poles are not evident at this point. However, the accumulation of Asp midway along the spindle suggests that the minus ends of opposite interpolar MTs are facing into the central region of the spindle. These observations are consistent with the finding, in each prophase II half spindle, of two opposite MT populations, with their plus ends near the chromatin and their minus ends outwards. The outward minus ends are focused on distinct poles, whereas the minus ends facing the spindle equator remain unbound. Control of spindle assembly could be achieved during meiosis II by the same chromosome-driven mechanisms of MT organization working in meiosis I. MTs in the vicinity of the condensed chromatin could be sorted into bundles and focused in stable poles by the cooperative interaction of molecular motors and cross-linking proteins. Loss of ncd function results, indeed, in the destabilization and fragmentation of both inner and outer spindle poles (Endow, 1998). Accordingly, the inner half spindles are formed during late prophase by parallel MTs that are first bundled at the spindle equator in distinct foci, and that then coalesce into a single pole. This model of meiotic spindle assembly is consistent with the formation of twin spindles in oocytes lacking normal-shaped central asters. It is proposed, therefore, that the acentrosomal pathway makes an essential contribution to spindle formation during meiosis II, even when a functionally active MTOC is present in the oocyte. An alternative mechanism might be that the inner half meiosis II spindles could be formed by a mixture of two MT populations: MTs nucleated in the vicinity of the chromosomes and MTs of the central aster. The gradual tethering together of these two MT populations could give rise to the tapered inner spindle poles, as proposed during the assembly of the mitotic spindle in cultured vertebrate cells. To what extent inner half spindles might be formed by two MT populations is difficult to determine, since individual MTs and their origin cannot be resolved within the assembling spindle. However, the Asp staining observed during transition from prophase to metaphase of the second meiosis suggests that the MTs radiating from the central aster contribute minimally, if at all, to the formation of the internal half spindles. The Asp staining is strong in the central region of the spindle, whereas it is weaker in the central aster, pointing to a lower MT density. This suggests that the contribution of the central MTOC is not essential for the higher MT density within the region where the inner poles will organize. Consistently, optical sections at the level of the spindle equator during prophase of the second meiosis reveal a continuity between the peripheral MT network and the ring-like tubulin aggregate, but no link is observed between this structure and the MTs of the central spindle. Finally, staining of Asp indicates that the minus ends of the inner half meiosis II spindle MTs start to coalesce into a single pole and, therefore, are completed before a distinct central aster is apparent. On the basis of these considerations the first hypothesis is favored in which the function of the central aster is redundant for the meiosis spindle organization (Riparbelli, 2005).
However, if the meiosis II spindles are assembled by an acentrosomal pathway, what is the role of the central aster? In the absence of a developed central aster, the twin spindles can be improperly spaced or oriented with respect to the long axis of the oocyte, resulting in the failure to correctly position the female pronucleus. The central aster could, therefore, be needed to keep the correct spacing between neighboring twin spindles. Accordingly, twin spindles are misaligned and at variable distances from each other when the central aster is defective as in polo mutants (Riparbelli, 2000). Mutation in cnn has been shown (Vaizel-Ohayan, 1999) to impair the spatial organization of mitotic spindles in the early Drosophila embryo (Riparbelli, 2005).
In vivo observations revealed that the precursor of the central spindle could be identified by a tubulin pucker midway along the central spindle at the end of meiosis I (Endow, 1998). This structure first appears as a tubulin aggregate to the spindle midzone, where antiparallel MTs overlap and recruit the PAV-KLP motor and CNN proteins. There are two possible mechanisms in the formation of the central aster. One possibility is that the MTs are nucleated by a discrete MTOC. The other possibility is that motor proteins may cross-link and organize randomly nucleated MTs into aster-like structures. The finding of several centrosomal proteins at the focus of the central aster during meiosis II (Megraw, 2000), strongly points to the first possibility. Since gamma-tubulin at the centrosome seems to be dependent on cnn function (Terada, 2003), the finding of CNN at the spindle midzone during anaphase I could indicate the prerequisite for the recruitment of gamma-tubulin at the central spindle and, therefore, for the nucleation of the central aster MTs. However, in cnn mutants that lack this protein at the spindle midzone in anaphase I and fail to accumulate gamma-tubulin, a faint astral array of MTs forms at the middle of the spindle during prometaphase/early metaphase II. In the absence of gamma-tubulin the central aster becomes poorly organized during subsequent meiotic stages and then disappears. It is therefore proposed (see Fig. 8 of Riparbelli, 2005 ) that the assembly of this structure might first rely on the cooperative interaction of motor proteins and acentrosomal MTs to form a polarized array of MTs that spread out from the equatorial region of the spindle. This process does not require ncd function, since mutants for the Ncd motor protein assemble an astral array of MTs at the beginning of meiosis II (Endow, 1998). Recruitment of material along these MTs might then contribute to the accumulation of centrosomal proteins, thus leading to the formation of a true MTOC that in turn could lead to further growth of the central aster. According to this model the presence of centrosomal material and other components, such as PAV-KLP, at its focus might be due to a fortuitous recruitment along its MTs. This is consistent with the finding that when the integrity of the central aster is affected in cnn mutant oocytes, there is little or no accumulation of PAV-KLP between twin spindles. By contrast, the motor protein is always found at the midzone of mutant spindles during both anaphase I and anaphase II, where it is needed for spindle dynamics. This is consistent with the observation that the accumulation of pericentrin and gamma-tubulin at the vertebrate centrosome is inhibited in the absence of tubulin or by microinjection of antibodies against cytoplasmic dynein. This two step mechanism could explain why gammaTub37CD mutants show an astral array of MTs at the beginning of metaphase II in the absence of detectable gamma-tubulin. The sperm aster is usually retained to be assembled by a true centrosome derived from both paternal and maternal sources. In particular the male gamete provides the centriole around which maternal components accumulate to form a mature centrosome able to nucleate MTs and to reproduce. Although rigorous conclusions are difficult to draw from negative results, the observation that cnn mutant oocytes have a developed sperm aster lacking CNN and gamma-tubulin, points to alternative mechanisms of sperm aster assembly. However, the possibility cannot be excluded that the amount of these proteins could be so much lower in mutant oocytes that they have escaped immunofluorescence analysis (Riparbelli, 2005).
Crossovers ensure the accurate segregation of homologous chromosomes from one another during meiosis. This study describes the identity and function of the Drosophila gene recombination defective (rec), which is required for most meiotic crossing over. rec encodes a member of the mini-chromosome maintenance (MCM) protein family. Six MCM proteins (MCM2-7) are essential for DNA replication and are found in all eukaryotes. REC (Matsubayashi, 2003) is the Drosophila ortholog of the recently identified seventh member of this family, MCM8. Phylogenetic analysis reveals the existence of yet another family member, MCM9, and shows that MCM8 and MCM9 arose early in eukaryotic evolution, though one or both have been lost in multiple eukaryotic lineages. Drosophila has lost MCM9 but retained MCM8, represented by REC. Genetic and molecular methods were used to study the function of REC in meiotic recombination. Epistasis experiments suggest that REC acts after the Rad51 ortholog SPN-A but before the endonuclease MEI-9. Although crossovers are reduced by 95% in rec mutants, the frequency of noncrossover gene conversion is significantly increased. Interestingly, gene conversion tracts in rec mutants are about half the length of tracts in wild-type flies. To account for these phenotypes, it is proposed that REC facilitates repair synthesis during meiotic recombination. In the absence of REC, synthesis does not proceed far enough to allow formation of an intermediate that can give rise to crossovers, and recombination proceeds via synthesis-dependent strand annealing to generate only noncrossover products (Blanton, 2005).
Faithful segregation of homologous chromosomes in meiosis requires crossovers, which, in concert with sister chromatid cohesion, form the chiasmata that hold and orient homologs on the meiotic spindle. Crossovers are distributed nonrandomly between chromosomes, along each chromosome arm, and relative to one another, indicating that meiotic recombination is tightly regulated. One aspect of this regulation is the process that determines whether a recombination event becomes a crossover or a noncrossover (Blanton, 2005).
Models of meiotic recombination must account for the production of both crossovers and noncrossovers. Current models are derived from the double-strand break (DSB) repair model. In this model, recombination is initiated with a DSB on one chromatid. Resection of the 5' ends leaves 3' single-stranded overhangs. One of these overhanging ends invades a homologous, non-sister duplex and primes repair DNA synthesis. The strand displaced by the migrating synthesis bubble is captured by the other 3' overhang, which primes synthesis using the displaced strand as a template. Ligation of the newly synthesized ends to the resected 5' ends generates an intermediate with two Holliday junctions. This double Holliday junction (DHJ) intermediate is resolved by an unknown endonuclease to form either crossover or noncrossover products (Blanton, 2005).
Recent data from yeast has resulted in modification of this model. Allers and Lichten (2001) physically monitored formation of recombination intermediates and products in Saccharomyces cerevisiae using an ectopic recombination system and found that noncrossover products appear before DHJ intermediates. They proposed that noncrossovers arise not through a DHJ intermediate, but through synthesis-dependent strand annealing (SDSA). In SDSA, the nascent strand dissociates from the template and anneals to the other resected end. Trimming of any overhangs and filling in of any gaps, followed by ligation, results in noncrossover products. Subsequent genetic tests of this model in S. cerevisiae are consistent with most noncrossovers coming from SDSA, while the remainder are derived from a DHJ intermediate [3] (Blanton, 2005).
These models also take into account the occurrence of gene conversionnonreciprocal transfer of information from one duplex to anotherthat can be associated with both crossovers and noncrossovers. Possible origins of gene conversion during SDSA are illustrated in this paper. Heteroduplex DNA (hDNA), in which the two strands are derived from different parental molecules, is produced by both invasion of a single-stranded overhang into a homologous template and annealing of a newly synthesized strand to the other single-stranded overhang. Sequence differences between homologous chromosomes result in base/base mismatches and insertion/deletion heterologies in hDNA, and these can be recognized and repaired by the mismatch repair system. The product contains a region of sequence derived from the homologous chromosome, referred to as a gene conversion tract. If heterologies are not repaired, each strand will convey different genetic information to the haploid product of meiosis. Upon the first round of DNA replication and mitosis after fertilization or germination, these strands separate, resulting in the post-meiotic segregation (PMS) of parental alleles. PMS results in a mosaic individual, or, for unicellular eukaryotes, a sectored colony (Blanton, 2005).
Though it is more difficult to physically observe intermediates formed during meiotic recombination in Drosophila, a wealth of evidence indicates that recombination is also initiated by DSBs in this organism. PubMed ID: MEI-W68, the Drosophila ortholog of Spo11, which catalyzes meiotic DSB formation in S. cerevisiae, is required to generate both crossovers and noncrossovers, and in mei-W68 mutants recombination is restored by treatment with ionizing radiation. Mutations in Drosophila genes required for strand invasion cause female sterility that is suppressed by mutation of mei-W68. Thus, the early steps in meiotic recombination appear to be similar between Drosophila and S. cerevisiae. In contrast, later stages of crossover production are different, since most crossovers in Drosophila require the XPF/Rad1 ortholog MEI-9, its binding partner ERCC1, and several novel proteins, including MUS-312 and MEI-218. In addition, it is not known whether noncrossovers in Drosophila are derived from a DHJ intermediate or SDSA, although SDSA is the most common pathway for repair of mitotic DSBs generated by transposable element excision (Blanton, 2005).
In Drosophila, mutagenesis screens have been used to identify many novel genes required for meiotic recombination. The gene recombination defective (rec) was identified more than 25 years ago by Grell (1984) in an ethyl methanesulfonate (EMS) screen for temperature-sensitive recombination mutants. Her preliminary characterization of two null alleles showed that rec mutants have high levels of chromosome nondisjunction and reduced fertility, both indicative of homologous chromosome segregation defects. Since these mutants are able to pair homologous chromosomes normally, but exhibit a severe reduction in crossing over, Grell concluded that rec is involved in generating meiotic crossovers (Blanton, 2005).
To gain insight into the function of the REC protein in meiotic recombination, rec was molecularly and it was found to encode the Drosophila ortholog of MCM8. The eukaryotic mini-chromosome maintenance (MCM) family of proteins contains six members (MCM2-7) that form a heterohexameric helicase required for replication. Though MCM2-7 are essential in all eukaryotes, rec mutants are viable, and no function has been found for REC outside of meiosis. To explore the defect in meiotic recombination further, the distribution of crossing over was examined in rec mutants and it was found that residual crossovers are distributed abnormally. This finding, coupled with epistasis analysis, suggests that REC might act at an intermediate step in recombination. Further insight into the function of REC comes from the finding that the frequency of noncrossovers is substantially increased in rec mutants, and that these noncrossover events have significantly shorter gene conversion tracts than those of wild-type females. Based on these phenotypes and the structural similarity between REC and MCM proteins, it is proposed that REC facilitates processive repair DNA synthesis, and is a prerequisite for formation of the DHJ intermediate during meiotic recombination. In the absence of REC, recombination proceeds through SDSA to generate noncrossovers (Blanton, 2005).
Understanding how crossovers form is crucial to understanding the mechanisms eukaryotes use to faithfully pass half of their genetic information to the next generation. In Drosophila, many components of the meiotic recombination pathway have been identified, but a complete picture of the process has yet to emerge. This paper describes the molecular and genetic characterization of an important participant in this pathway -- REC, the Drosophila homolog of MCM8, giving new insight into requirements for crossover formation (Blanton, 2005).
The data support a model in which REC acts at an intermediate step of meiotic recombination. REC is not required for pre-meiotic S phase because homologous chromosomes in rec mutant females form normal synaptonemal complex, indicative of complete replication of genomic DNA. The finding that rec mutant females have about twice the normal number of noncrossover gene conversions indicates that initiation of recombination is not impaired in rec mutants; rather, very few DSBs are repaired as crossovers. The data suggest that REC functions after strand invasion, since females mutant for both rec and spn-A, which encodes the Rad51 ortholog, phenocopy spn-A single mutants. Based on the distribution of residual crossovers in rec mutants and in mei-9; rec double mutants, it is likely that REC does not function with MEI-9 at resolution but acts at some previous step (Blanton, 2005).
Normally, some recombination events become crossovers and some become noncrossovers. An increase in noncrossovers would occur if the crossover pathway were blocked so that most or all events followed the noncrossover pathway. In the ry intragenic recombination assay, noncrossover gene conversions are recovered only if they span a mutant site and convert that site to the wild-type sequence. In contrast, a crossover can be recovered if it occurs anywhere between the two mutations, as long as it generates a wild-type chromosome. Based on conversion tract lengths and the distance between the two mutations, it is expected that many of the crossovers recovered would not be detected if they instead became noncrossovers, because they would not contain a conversion tract long enough to span a mutant site. The increase in noncrossovers that was observed in rec mutants, therefore, appears to be more than expected from this simple interpretation. A possible explanation for the increased frequency of noncrossovers in rec mutants comes from a hypothesis proposed by Bhagat (2004), who suggested that crossover distribution is disrupted as the result of a feedback mechanism that ensures one crossover per chromosome. The proposed feedback mechanism senses some intermediate in the crossover pathway (e.g., the DHJ structure). In mutants in which this intermediate does not form, a signal causes the cell to initiate additional recombination events to ensure that a crossover is obtained. These initiations may occur outside the normal constraints, leading to a disruption of the normal distribution and an apparent polar reduction in crossing over. According to this model, rec mutants are impaired in formation of some crucial intermediate leading to crossovers. As a result, more recombination events are initiated, but most of these still become noncrossovers. Thus, the frequency of noncrossovers is elevated, and the crossovers that are produced do not follow the normal distribution (Blanton, 2005).
The defect in rec mutants is not limited to an increased production of noncrossovers at the apparent expense of crossovers. Noncrossover gene conversion tract length is significantly reduced in rec mutants. This could result from a defect in generating hDNA or a defect in repairing hDNA. Defects in repair of hDNA result in PMS of markers within the heteroduplex tract. PMS was not detected in any of the events from wild-type or rec mutant females. Thus, rec mutants are not defective in repair of hDNA; rather, formation of hDNA may be compromised (Blanton, 2005).
The length of hDNA can be affected by the extent of strand invasion and the amount of repair synthesis. In S. cerevisiae, the Mer3 helicase has been shown in vitro to stimulate Rad51-mediated strand invasion (Mazina, 2004). As in rec mutants, mutations in the gene that encodes Mer3 cause a reduction in the frequency of crossovers and an increase in the frequency of noncrossovers (Nakagawa, 1999). However, in physical assays mer3 mutants are defective in the transition from DSB to strand invasion intermediate. The data suggest that Drosophila REC acts after strand invasion, so the notion is not favored that REC performs a function similar to that of Mer3. Furthermore, based on the similarity of REC to replicative MCMs, it is thought plausible that rec mutants have shorter conversion tracts because repair synthesis is diminished (Blanton, 2005).
What is the relationship between reduced repair synthesis and decreased crossing over in rec mutants? In S. cerevisiae, crossovers are believed to arise through resolution of the DHJ intermediate. Although this process can also give rise to noncrossovers, most noncrossovers are thought to arise through SDSA. There is evidence in S. cerevisiae that formation of a DHJ intermediate requires more repair synthesis than SDSA. If this is also the case in Drosophila, then decreased repair synthesis would increase the probability that a meiotic DSB will be repaired through SDSA instead of the DHJ pathway (Blanton, 2005).
A model is proposed in which REC drives crossover formation by acting at the repair synthesis step of meiotic recombination. In the absence of REC, synthesis does not proceed far enough to allow second-end capture and formation of the DHJ intermediate, resulting in a deficit of crossovers. Noncrossovers may still be formed through SDSA. There are two versions of this model. First, REC may facilitate repair synthesis at all sites of recombination. In this version, noncrossovers may normally arise through the DHJ pathway or the SDSA pathway, but in rec mutants the SDSA pathway is favored; the decrease in gene conversion tract length in rec mutants reflects an overall decrease in repair synthesis. Alternatively, REC may facilitate synthesis only at those recombination sites designated to become DHJ intermediates . In this version of the model, sites lacking REC in wild-type flies undergo SDSA. The decrease in mean tract length in rec mutants is due to loss of those noncrossovers that would have arisen via a DHJ intermediate (Blanton, 2005).
The data do not indicate whether noncrossovers in wild-type flies arise through SDSA, DHJ, or a combination of the two. In Drosophila, SDSA is a primary pathway for DSB repair in nonmeiotic cells. It may be that SDSA is the 'default' pathway for recombinational repair of DSBs, and that meiosis-specific modifications promote formation of DHJs to allow crossing over. REC does not appear to play a role in SDSA in nonmeiotic cells, and therefore REC may be a component of the meiosis-specific modifications to DSB repair in Drosophila. To better understand the role of REC and the process of meiotic recombination, it will be important to determine the source of noncrossover recombinants in wild-type females (Blanton, 2005).
Conventional centrosomes are absent from a female meiotic spindle in many animals. Instead, chromosomes drive spindle assembly, but the molecular mechanism of this acentrosomal spindle formation is not well understood. This study screened female sterile mutations for defects in acentrosomal spindle formation in Drosophila female meiosis. One of them, remnants (rem), disrupted bipolar spindle morphology and chromosome alignment in non-activated oocytes. It was found that rem encodes a conserved subunit of Cdc2 (Cks30A). Since Drosophila oocytes arrest in metaphase I, the defect represents a new Cks function before metaphase-anaphase transition. In addition, it was found that the essential pole components, Msps and D-TACC, are often mislocalized to the equator, which may explain part of the spindle defect. The second cks gene cks85A, in contrast, has an important role in mitosis. In conclusion, this study describes a new pre-anaphase role for a Cks in acentrosomal meiotic spindle formation (Pearson, 2005).
Spindle formation in female meiosis is unique in terms of the absence of conventional centrosomes. Instead, chromosomes have a central role in the assembly of spindle microtubules. This acentrosomal (also called acentriolar or anastral) spindle formation is common in female meiosis for many animals including mammals, insects and worms. Despite potential medical implications, this spindle formation is much less studied than centrosome-mediated spindle formation in mitosis (Pearson, 2005).
Drosophila provides a valuable tool to study the acentrosomal spindle formation in vivo. Unlike many other species, mature non-activated Drosophila oocytes arrest in metaphase of meiosis I until ovulation, which coincides with fertilization. This provides a unique opportunity to study spindle formation, without interference from chromosome segregation or meiotic exit (Pearson, 2005).
Two components of acentrosomal spindle poles, Msps and D-TACC, physically interact and are crucial for spindle bipolarity. Other studies have identified essential components for spindle formation, such as kinesin-like proteins (Ncd and Sub, γ-tubulin, and a membrane protein surrounding the spindle (Axs). Some of these spindle components are probably modulated by cell-cycle regulators, but knowledge of the regulation is limited. To identify essential components and regulators, a cytological screen was performed for mutants defective in acentrosomal spindle formation of non-activated oocytes (Pearson, 2005).
Through the screen, remnants was identified and identified as a mutant of a Drosophila Cks/Suc1 homologue, Cks30A. Cks is the third subunit of the Cdc2 (Cdk1)-cyclin B complex, but the role of Cks is less clearcut than that of other subunits of the complex. It is implicated in entry into mitosis/meiosis, metaphase-anaphase transition, exit from mitosis/meiosis and inactivation of Cdk inhibitors. This study shows that Cks30A is required for spindle morphogenesis and chromosome alignment in the metaphase I spindle in arrested mature oocytes. This requirement of a Cks before metaphase-anaphase transition represents a new function that has not previously been identified. Furthermore, it was found that essential spindle pole components Msps and D-TACC mislocalize in the mutant, which may be partly responsible for the spindle defects (Pearson, 2005).
For molecular analysis of the acentrosomal spindle in Drosophila female meiosis, female sterile mutants were screened for spindle defects in non-activated oocytes. Female sterile mutants on the second chromosome have previously been isolated. This study focused on classes of mutants that lay eggs that do not develop beyond the blastoderm stage. This category of mutants includes known meiotic mutants affecting spindle formation, such as fs(2)TW1 (γ-tubulin 37C) and subito (a kinesin-like protein (Pearson, 2005).
The identity of the remnants (rem) gene was identified by positional cloneing. The rem gene was previously mapped to 30A-C using a deficiency Df(2L)30AC (Schupbach, 1989). One missense mutation was identified in the gene CG3738 (cks, hereafter called cks30A; Finley, 1994). There were no other mutations within coding sequences and splicing junctions in the region. In addition, the amount and size of the transcripts that are known to be expressed in adult females was tested, and no differences were found between rem and wild type (Pearson, 2005).
Cks30A is one of two Drosophila homologues of Saccharomyces cerevisiae Cks1/Schizosaccharomyces pombe Suc1, a conserved subunit of the Cdc2 (Cdk1)/cyclin B complex, and has been shown to interact with Cdc2 (Finley, 1994). The mutation in rem1 results in a conversion of the 61st amino acid from proline to leucine. This proline is completely conserved among all Cks homologues, further confirming that the mutation is not a polymorphism. Crystal structure analysis has indicated that this residue forms part of the interaction surface with Cdc2 (Bourne, 1996). Immunoblots using an anti-human Cks1 antibody indicated that this mutation disrupts the stability of the Cks30A protein (Pearson, 2005).
To explain the role of Cks30A, focus was placed on the rem1 mutant in non-activated oocytes, which arrest in metaphase I. Non-activated oocytes were dissected from wild type and the rem1 mutant, and chromosomes and spindles were visualized by immunostaining (Pearson, 2005).
In wild type, non-activated mature oocytes contain a single bipolar spindle around chromosomes. Bivalent chromosomes align symmetrically with chiasmatic chromosomes at the equator and achiasmatic chromosomes that are located nearer the poles. The rem1 mutant was able to enter meiosis, condense chromosomes and assemble microtubules around chromosomes. However, only a minority of spindles showed normal spindle morphology and chromosome alignment (Pearson, 2005).
The most prominent defect in the rem1 mutant was chromosome misalignment. This defect was observed in about a half of the spindles. Even in the cases in which the spindle remained well organized, chiasmatic chromosomes often moved away from the equator and lost overall symmetrical distribution. The second class of defect in the rem1 mutant was abnormal spindle morphology. Although the abnormality varied from spindle to spindle in the rem1 mutant, the most typical defect was the formation of ectopic poles near the spindle equator. The focusing of spindle poles seemed to be unaffected (Pearson, 2005).
Further quantitative analysis showed no significant difference between the phenotypes of rem1 homozygotes (rem1/rem1) and hemizygotes (rem1/Df). This indicates that the rem1 mutation is genetically amorphic. A recent independent study (Swan, 2005a) has indicated that another weaker allele remHG24 shows similar abnormalities at a lower frequency. These results indicate that Cks30A is required before the metaphase-anaphase transition for spindle morphology and chromosome alignment (Pearson, 2005).
To gain an insight into the spindle defects in female meiosis, the localization of Msps was examined. Msps protein belongs to a conserved family of microtubule regulators, including XMAP215, and is the first protein identified at the acentrosomal poles in Drosophila. An msps mutation often leads to the formation of a tripolar spindle in female meiosis I (Pearson, 2005).
In wild type, Msps protein is accumulated at the acentrosomal poles of the metaphase I spindle in female meiosis, although the localization sometimes spreads to the spindle microtubules. In the rem1 mutant, although the Msps protein is still concentrated at the poles, it is often accumulated around the equator of the spindle. Mislocalization of this important pole protein to the equator in the rem1 mutant may sometimes lead to the formation of ectopic spindle poles near the equator (Pearson, 2005).
Msps localization is dependent on another pole protein D-TACC, which binds to Msps. To test whether D-TACC also mislocalizes, the localization of D-TACC was examined in the rem1 mutant. In wild type, D-TACC is highly concentrated at the acentrosomal pole. In the rem1 mutant, D-TACC often accumulates at the spindle equator, although it is still concentrated around the poles to some degree. In summary, Cks30A is required for correct localization of the essential pole proteins, Msps and D-TACC (Pearson, 2005).
To gain an insight into how the defect in the Cdc2 complex leads to Msps or D-TACC mislocalization to the spindle equator, the localization of cyclin B was examined. Cyclin B is considered to be the main determinant of the activity and cellular localization of the Cdc2 complex. Immunostaining in non-activated oocytes showed that cyclin B is localized to the metaphase I spindle, with a concentration around the spindle equator. This cyclin B localization could suggest a possible regulatory role of the Cdc2 complex in the transport of Msps and D-TACC from the spindle equator to the poles. The cyclin B localization is not affected in the rem mutant, suggesting that Cks30A mainly affects the substrate specificity of the Cdc2 complex, as shown in other systems (Pearson, 2005).
The Drosophila genome contains one more predicted cks homologue (CG9790), which is called cks85A. Although mammalian genomes also have two Cks genes, they are more similar in sequence to each other than to either of the two cks genes in Drosophila (Pearson, 2005).
The gene expression pattern of the two cks genes was examined during Drosophila development. RNAs were isolated from various stages of development and analysed by reverse transcription-PCR (RT-PCR) using primers that correspond to each of the cks genes. cks30A gave strong signals in adult females and embryos, whereas it gave only weak signals in adult males, larvae and pupae. This maternal expression pattern is consistent with the observed female sterile phenotype of the cks30A (rem1) mutant. In contrast, cks85A signals were obtained more uniformly throughout the development without sex specificity in adults. In S2 cultured cells, which originated from embryos, both genes were well expressed (Pearson, 2005).
To identify the Cks proteins, an anti-human Cks1 antibody was used for immunoblots of protein extracts from embryos and S2 cells. Although the antibody recognized many proteins, two bands were detected within a range of molecular weights consistent with the Cks proteins. In embryos laid by the rem1 mutant, the amount of the smaller band was greatly reduced. To further confirm their identity, S2 cells were subjected to RNA interference (RNAi) using doublestranded RNAs (dsRNAs) corresponding to the cks genes. It was found that both of the bands disappeared when both genes were simultaneously knocked down by RNAi. It indicated that, consistent with RT-PCR results, S2 cells produced both the Cks proteins and that RNAi effectively depletes them (Pearson, 2005).
Cytological analysis showed that cks85A RNAi results in a significant increase in chromosome misalignment/missegregation and spindle abnormality in mitosis after an extended time, whereas cks30A RNAi has a lesser impact on mitotic progression. About a half of anaphase or telophase cells had lagging chromosomes or chromosome bridges after cks85A RNAi. In some cases, spindles contained scattered chromosomes the sister chromatids of which were either attached or detached. The frequency of multipolar spindles was also increased. The genetic and RNAi results indicated that cks85A has an important function in mitotic progression, whereas cks30A mainly functions in female meiosis (Pearson, 2005).
This study has shown a new pre-anaphase function of a Cks protein in acentrosomal spindle formation during Drosophila female meiosis. Through a cytological screen, spindle defects in remnants among female sterile mutants. Cytological analysis showed that Cks30A is required for correct formation of the acentrosomal spindle and chromosome alignment in female meiosis I. The observation on mislocalization of the essential pole components, Msps and D-TACC, in the mutant provides a molecular insight into a role of Cks30A in spindle morphogenesis (Pearson, 2005).
Cks/Suc1 protein is the third subunit of the Cdc2-cyclin B complex, which is conserved across eukaryotes. Although it has been known to be essential for the cell cycle, the function seems to be less straightforward than that of the other subunits of the Cdc2 complex. One reason is that Cks also interacts with other Cdks and has Cdk-independent functions. Even if Cks is limited to roles in mitosis/meiosis, Cks proteins are implicated in entry into mitosis/meiosis, metaphase-anaphase transition and also exit from mitosis/meiosis. Furthermore, the roles of Cks were further complicated by the fact that animal genomes encode two Cks homologues (Pearson, 2005).
Studies in Caenorhabditis elegans and mice showed that one of two cks genes is required for female fertility. Similarly, the results indicated that one of two Drosophila cks homologues, cks30A, is expressed maternally and is required for female meiosis. Further analysis indicated that Cks30A is required for proper bipolar spindle formation and chromosome alignment in mature oocytes arrested in metaphase I. In C. elegans, depletion of one of the Cks proteins by RNAi results in a failure to complete meiosis I. Similarly, in mice, oocytes from a Cks2 knockout cannot progress past metaphase I and a small percentage of oocytes show chromosome congression failure. In both cases, the defects were interpreted mainly as post-metaphase defects. Since Drosophila non-activated oocytes are arrested in metaphase I until ovulation, pre-anaphase function of Cks30A can be distinguised from possible post-metaphase function. This study clearly showed that Drosophila Cks30A has a function in establishing metaphase I, in addition to later functions that have reported recently (Swan, 2005a; Pearson, 2005).
At the moment, it is not known how the cks30A mutation disrupts spindle formation and chromosome alignment in female meiosis. It has been thought that a loss of Cks function affects the Cdc2 activity towards certain substrates. It was found that the essential pole components, Msps and D-TACC, mislocalize to the spindle equator in the mutant. Previously, it was hypothesized that Msps is transported by the Ncd motor and anchored to the poles by D-TACC. D-TACC localizes to the poles independently from Ncd, but may also be transported from the spindle equator along microtubules by other motors. Cks30A-dependent Cdc2 activity may be required for activating the transport system at the onset of spindle formation in female meiosis. Consistently, it was found that cyclin B is concentrated around the equator of the metaphase I spindle. Msps is the XMAP215 homologue and belongs to a family of conserved microtubule-associated proteins. It is a major microtubule regulator, both in mitosis/meiosis and interphase. The mislocalization of this microtubule-regulating activity could lead to the disruption of spindle organization in the mutant (Pearson, 2005).
Meiosis in female oocytes lacks centrosomes, the microtubule-organizing centers. In Drosophila oocytes, meiotic spindle assembly depends on the chromosomal passenger complex (CPC). To investigate the mechanisms that regulate Aurora B activity, this study examined the role of protein phosphatase 2A (PP2A) in Drosophila oocyte meiosis. Both forms of PP2A, B55 and B56, antagonize the Aurora B spindle assembly function, suggesting that a balance between Aurora B and PP2A activity maintains the oocyte spindle during meiosis I. PP2A-B56, which has a B subunit encoded by two partially redundant paralogs, wdb and wrd, is also required for maintenance of sister chromatid cohesion, establishment of end-on microtubule attachments, and metaphase I arrest in oocytes. WDB recruitment to the centromeres depends on BUBR1, MEI-S332 and kinetochore protein SPC105R. Although BUBR1 stabilizes microtubule attachments in Drosophila oocytes, it is not required for cohesion maintenance during meiosis I. At least three populations of PP2A-B56 regulate meiosis are proposed, two of which depend on SPC105R and a third that is associated with the spindle (Zhang, 2021).
Meiosis is a highly specialized cell division that requires significant
reorganization of the canonical cell-cycle machinery and the use of
meiosis-specific cell-cycle regulators. The anaphase-promoting complex (APC, a machine for degrading proteins; see APC subunits Cdc27 and morula; for review see Acquaviva, 2006)
and a conserved APC adaptor/activator, Cdc20 (also known as Fizzy), are required for
anaphase progression in mitotic cells. The APC has also been implicated in
meiosis, although it is not yet understood how it mediates these non-canonical divisions. Cortex (Cort) is a diverged Fzy homologue that is expressed in the female germline of Drosophila, where it functions with the Cdk1-interacting protein Cks30A to drive anaphase in meiosis II. This study shows
that Cort functions together with the canonical mitotic APC adaptor Fzy to
target the three mitotic cyclins (A, B and B3) for destruction in the egg and
drive anaphase progression in both meiotic divisions. In addition to
controlling cyclin destruction globally in the egg, Cort and Fzy appear to
both be required for the local destruction of cyclin B on spindles. Cyclin B associates with spindle microtubules throughout meiosis I and
meiosis II, and dissociates from the meiotic spindle in anaphase II. Fzy and
Cort are required for this loss of cyclin B from the meiotic spindle. These
results lead to a model in which the germline-specific APCCort
cooperates with the more general APCFzy, both locally on the
meiotic spindle and globally in the egg cytoplasm, to target cyclins for
destruction and drive progression through the two meiotic divisions (Swan, 2007).
The cell divisions of female meiosis and the ensuing mitotic cycles of
early embryogenesis represent two examples of non-canonical cell cycles.
Meiosis differs from the typical mitotic cycle in several respects. Most
notably, two divisions occur in sequence without an intervening S-phase,
resulting in the production of four haploid gametes. Additionally, the first
meiotic division involves the segregation of homologous chromosomes and occurs
without sister chromatid segregation, whereas the second meiotic division
involves the segregation of sister chromatids, as occurs in mitosis. The
regulation of meiosis requires a significant reorganization of the canonical
cell-cycle machinery and the use of a number of meiosis-specific cell-cycle
regulators. One example is in the regulation of anaphase - the
coordinated series of events that results in the segregation of chromosomes to
produce two daughter nuclei. In mitotically dividing cells, anaphase
progression crucially depends on the inactivation of the mitotic kinase Cdk1
(also known as Cdc2) and on the subsequent release of sister chromatid
cohesion through the destruction of cohesin complexes. These events are
controlled by an E3 ubiquitin ligase -- the anaphase-promoting complex (APC) --
in association with an adaptor protein, Fzy, and this complex targets mitotic
cyclins and securin (potential Drosophila homolog; Pimples) for destruction (reviewed in
Peters, 2002). The role of the
APC in meiosis appears to be more complex than in mitotic cells. For example,
the APC only partially inhibits Cdk1 activity between meiotic divisions
and sister chromatid cohesion persists at centromeres through anaphase I. It is not yet clear how the activity of the APC is modified in these specialized
cell divisions (Swan, 2007).
In most eukaryotes, the meiotic cell cycle is followed by another atypical
cell cycle -- the cleavage divisions of early embryogenesis. In
Drosophila, these cleavage cycles occur as a series of synchronized,
rapid nuclear divisions and are referred to as syncytial divisions. The female
meiotic cell cycle is not only closely linked to the syncytial mitotic cell
cycle in time, but it also occurs within a shared cytoplasm -- that of the egg.
Therefore, these two distinct cell cycles share a common pool of cell-cycle
regulators, and may share common strategies for spatially and temporally
regulating cell-cycle progression within a syncytium (Swan, 2007).
One way in which the syncytial cell cycle is modified appears to be in the
limited destruction of mitotic cyclins in each cell cycle, apparently by
restricting their destruction to the area of the mitotic nuclei. Although
there is evidence that cyclin destruction is spatially regulated in somatic
cells, this strategy appears to be of particular importance in the syncytial embryo of Drosophila as a means to conserve mitotic cyclins for the duration of the rapid syncytial divisions. Several lines of evidence suggest that at least one cyclin, cyclin B, undergoes limited local destruction on mitotic spindles in the syncytial embryo. It is not yet known what mediates this local cyclin B destruction, and it is also not known whether this is unique to the syncytial mitotic cell cycle or if it occurs in the preceding meiotic divisions (Swan, 2007).
Drosophila represents an excellent model system for understanding
how the canonical cell-cycle machinery is developmentally modified, and how
novel cell-cycle regulators are used to control meiosis and syncytial
divisions. cortex (cort) encodes a Cdc20/Cdh1 (Cdh1 is also
known as Fzr and Rap)-related protein, that appears to be required
specifically in female meiosis (Chu, 2001; Lieberfarb, 1996; Page, 1996) and functions with a germline-specific Cks gene, Cks30A, to mediate the destruction of cyclin A (Swan, 2005a; Swan, 2005b). This study shows that the canonical APC adaptor Fzy functions together with Cort to target mitotic cyclins for destruction, and to drive anaphase in both meiosis I and meiosis II. Female meiosis, like the subsequent syncytial mitotic cell cycles, appears to involve the local destruction of cyclin B, and both Cort and Fzy were found to be required for this process (Swan, 2007).
In most cell types, in both Drosophila and in other metazoans, the
APCFzy drives anaphase progression by targeting mitotic cyclins and other mitotic proteins for destruction. This study shows that the female germline is an exception in that the APCFzy is not sufficient. A germline-specific APC adaptor, Cort, cooperates with Fzy to mediate cyclin destruction in meiosis (Swan, 2007).
The cort gene encodes a diverged member of the Fzy/Cdh1 family
(Chu, 2001). Fzy/Cdh1
homologues interact with the APC and with specific sequences (D-box, KEN box
or A-box) found on cyclins and on other APC targets. As such, Fzy/Cdh1
proteins act as specificity factors to target proteins for ubiquitination and
eventual destruction. Cort protein, like all Fzy/Cdh1-family proteins,
contains seven WD domains in the C-terminal-half of the protein, implicated in substrate recognition (Pfleger, 2001). Cort has an N-terminal C-box (amino
acids 482, 483) and a C-terminal IR tail (amino acids 54-60), both implicated
in binding to the APC. In addition to containing these conserved functional
domains, Cort displays a conserved ability to mediate cyclin destruction.
cort mutations result in the overaccumulation of cyclin A, cyclin B
and cyclin B3 in the egg (Swan, 2005a), whereas the ectopic expression of Cort in the wing disc leads to a reduction in the levels of these mitotic cyclins. Taken together, these results indicate that Cortex encodes a functional member of the Fzy/Cdh1 family (Swan, 2007).
Although the Drosophila genome has four genes that encode Fzy/Cdh1
proteins, only two of these proteins, Fzy and Cort, are expressed at
detectable levels in the female germline (Raff, 2002;
Jacobs et al., 2002; Chu, 2001). The role of these two APC adaptors has been studied both individually and in double
mutants, and it was found that they function together to promote anaphase in
both the first and second meiotic divisions of female meiosis. In most cell
types in Drosophila and other eukaryotes, a single APC complex,
APCFzy, is responsible for cyclin destruction and anaphase
progression. It is therefore surprising that, in the female germline of
Drosophila, two APC adaptors are necessary for meiotic progression.
In the case of meiosis I, Cort and Fzy appear to play largely redundant roles, since only removing both genes results in a significant block in meiosis I. The
two APC complexes may also be functionally redundant with respect to global
cyclin levels. Mutations in either fzy or cort result in an
increase in the levels of cyclin A, cyclin B and cyclin B3, whereas mutation
in both genes results in even-further increases in cyclin levels (Swan, 2007).
Although Cort and Fzy have overlapping roles in promoting anaphase I, both
are essential for meiosis II. This could simply reflect a greater quantitative
requirement for APC activity in meiosis II. Alternatively, the two APC
complexes could have distinct roles in the second meiotic division. Consistent
with this latter possibility, mutations in either cort or
fzy both result in arrest at different stages of meiosis II:
cort mutants arrest with the sister chromatids associated, and
therefore in metaphase, whereas fzy mutants almost invariably arrest
with separated sister chromatids, and are therefore in anaphase. cort
and fzy also result in different patterns of cyclin B stabilization
on the arrested spindles, suggesting roles in metaphase and anaphase,
respectively. Therefore, Cort may function to initiate sister chromatid
separation at the onset of anaphase II and Fzy may primarily function later,
in anaphase II. Alternatively, the later arrest observed in fzy could
simply reflect the fact that the fzy alleles that have been used are
not nulls, and it is possible that a complete loss of Fzy activity would also
result in a metaphase arrest, as seen in cort. However, comparing the
meiosis II phenotypes of fzy with Cks30A-null mutants
suggests that the later arrest in fzy is not simply due to residual
activity. Cks30A-null mutants have a weaker meiotic arrest than
fzy; they complete meiosis at high frequency
(Swan, 2005a), but they
display a higher frequency of metaphase arrest or delay. The fact that
fzy does not similarly cause a delay in metaphase of meiosis II
suggests that it is only required at anaphase. Therefore, it is possible that
Fzy is crucial at anaphase, whereas Cort is necessary for the metaphase to
anaphase transition (Swan, 2007).
The different temporal requirements for Cort and Fzy prior to and after
sister chromatid separation, respectively, could be related to their apparent
differences in substrate specificity. Western analysis
reveals that Cort is
more important for the destruction of cyclin A and cyclin B3, whereas Fzy
appears to play a greater role in cyclin B destruction in the egg. In mitotic
cells, cyclin destruction occurs sequentially. Cyclin A is destroyed first, in
prometaphase, and this is a prerequisite for sister chromatid separation.
Cyclin B destruction occurs at anaphase onset and is necessary for later
anaphase events, subsequent to sister chromatid separation.
Therefore, it is possible that Cort promotes the early stages of meiotic
anaphase by targeting cyclin A for destruction, whereas Fzy is more crucial
later, through its targeting of cyclin B for destruction (Swan, 2007).
The meiotic cell cycle differs in many respects from the standard mitotic
cycle. Whereas APC-mediated destruction of mitotic regulators appears to be
required for anaphase progression in most or all mitotic cells, the role of
the APC and cyclin destruction in meiosis is not as well-understood. This
analysis of the two APC adaptors Cort and Fzy has permitted an evaluation of
the role of the APC complex in female meiosis in Drosophila. The APC is required for anaphase progression in both meiotic divisions.
Correlating with its requirement for the completion of meiosis, the APC is
required for the destruction of mitotic cyclins. At least one of these
cyclins, cyclin B, is a crucial substrate in meiosis, because the expression
of a stabilized form of cyclin B disrupts this process. Therefore, APC
activity and cyclin destruction are required for anaphase progression in both
meiotic divisions, in addition to in mitosis. APC activity has been implicated
in both meiotic divisions in C. elegans and in
the mouse, and in the second, but not the first, meiotic division in
Xenopus. In yeast, two APC complexes, the mitotic APCFzy
and a meiosis-specific complex (APCAma1 in S. cerevisiae
and APCMfr1 in S. pombe) function together to mediate
protein destruction in meiosis. It now appears that Drosophila also uses two APC complexes in female meiosis, and this may turn out to be a common strategy in other eukaryotes (Swan, 2007).
Cks30A belongs to a highly conserved family of proteins that bind to and
stimulate the activity of the mitotic kinase Cdk1. In Xenopus, the
Cks30A homologue Xep9 stimulates the Cdk-dependent phosphorylation of APC
subunits, and thereby promotes the activation of the APCFzy complex. The current
results suggest that Cks30A may have a similar role in stimulating both the
APCFzy and APCCort in female meiosis in
Drosophila. (1) Cks30A, like cort and
fzy, is required for the completion of meiosis II and, like
fzy, it is required for the completion of the first mitotic division
of embryogenesis (Lieberfarb, 1996; Page, 1996; Swan,
2005). (2) Cks30A, as are Cort and Fzy, is necessary for
global cyclin destruction in the Drosophila egg and for local cyclin
B destruction on the meiotic spindle. Global levels of cyclin A
and cyclin B3 are elevated to a greater extent in Cks30A mutants than
in single mutants for cort or fzy, consistent with the idea
of Cks30A activating both Cort and Fzy. (3) Cks30A is
necessary for the activity of ectopically expressed Cort in the adult wing. Cks30A may also play a role in activating APCFzy in mitotic cells.
the temperature-sensitive fzy6 allele is lethal at all
temperatures in a Cks30A-null background, suggesting that the Cks30A-dependent activation of APCFzy becomes essential when Fzy activity is compromised (Swan, 2007).
Although Cks30A appears to promote the activity of the APCCort
and the APCFzy, these complexes seems to retain some activity in
the absence of Cks30A. Whereas cort and fzy cause an arrest
in meiosis II, Cks30A-null mutants are typically delayed only in
meiosis II (Swan,
2005a). Also, although cyclin A and cyclin B3 levels are elevated
more in Cks30A eggs than in either fzy or cort, their levels
are still not as high as in fzy; cort double mutants, indicating that
Fzy and Cort can destroy cyclin A and cyclin B3 to some degree in the absence
of Cks30A. Cyclin B destruction is even less dependent on Cks30A, because
cyclin B levels are affected less in Cks30A mutants than in either
cort or fzy single mutants. Therefore, Cks30A may be more
crucial for the activity of APCCort and APCFzy complexes
on cyclin A and cyclin B3, and less crucial for their activity on cyclin B.
The relatively weaker meiotic arrest in Cks30A mutants compared to
fzy; cort double mutants may also indicate that the APC has other
meiotic targets that can be destroyed in the absence of Cks30A (Swan, 2007).
Cyclin B undergoes local oscillations in its association with mitotic
spindles in syncytial embryos, appearing transiently along the full length of
the mitotic spindle in early metaphase and gradually disappearing from the
spindle starting at the centrosomes and ending at the kinetochores. The
timing of this loss of cyclin B from the spindle, at the onset of anaphase,
corresponds with the timing of cyclin B destruction in other cell types,
suggesting the possibility that cyclin B is locally destroyed on the spindle
in anaphase. This study shows that cyclin B is subject to similar local
oscillations in the female meiotic cycles, and that cyclin B
destruction is necessary for the completion of female meiosis. Importantly, the local loss of cyclin B from the spindle in meiosis is
dependent on the APC adaptors Cort and Fzy, and that the local loss of cyclin
B from the spindle in mitosis depends on Fzy. These results
strongly suggest that the local loss of cyclin B from the spindle in anaphase
of meiosis II and anaphase of mitosis is actually due to its local
destruction (Swan, 2007).
The pattern of accumulation and loss of cyclin B from the spindle in
meiosis differs in some respects compared to syncytial mitotic cycles. (1) In metaphase of mitosis, cyclin B initially accumulates throughout the spindle microtubules, whereas, in metaphase of the meiotic divisions, cyclin B first appears exclusively at the spindle mid-zone. This difference may reflect the fact that the meiotic spindle does not contain centrosomes and cyclin B may, therefore, not load onto spindles from centrosomes and progress along the spindles to the kinetochores, as has been proposed for mitosis.
(2) The timing of cyclin B destruction appears to be different between the meiotic and mitotic cycles. Most strikingly, there is no loss of cyclin B from the spindle in anaphase of meiosis I, implying that local cyclin B destruction is not necessary for the completion of the first meiotic division. In addition, the loss of cyclin B from the spindle following meiosis II occurs only late in anaphase rather than at the onset of anaphase, as occurs in the syncytial mitotic cycles. It is not yet known how cyclin B destruction is prevented in anaphase I and early in anaphase of meiosis II. One possibility is that the spindle-assembly checkpoint is locally active during these stages. This checkpoint is required for the proper completion of female meiosis in Drosophila, and it will be interesting to see if this requirement reflects a role in inhibiting either APCFzy or APCCort activity (Swan, 2007).
The specific accumulation of cyclin B at the spindle mid-zone in meiosis
may reflect the unique properties of the meiotic spindle. The mid-zone
microtubules or central spindle microtubules are a subset of spindle
microtubules that do not end in kinetochores, but instead overlap at the
mid-zone with microtubules from the other pole. In dividing cells, the central spindle is crucial for cytokinesis, but, in female meiosis, it appears to have a role in spindle assembly. Along with cyclin B, the chromosomal passenger proteins
Aurora B and Incenp are recruited to the spindle mid-zone. It will be of great interest to determine what these proteins do at the mid-zone and how cyclin B destruction at this site may be important for anaphase in meiosis. It will also be important to determine how the APCCort targets cyclin B at the spindle mid-zone. It has not been possible to detect any specific localization of GFP or HA-tagged Cortex in meiosis or in the syncytial embryo, but it is possible that its activity is
spatially regulated (Swan, 2007).
In conclusion, these results support a model in which two APC complexes,
APCFzy and APCCort, cooperate to mediate the destruction of meiotic cyclins and allow progression through female meiosis (Swan, 2007).
Centrosomes, the major organizers of the microtubule network in most animal cells (see The centrosome cycle in mammalian cells), are composed of centrioles embedded in a web of pericentriolar material (PCM). Recruitment and stabilization of PCM on the centrosome is a centriole-dependent function. Compared to the considerable number of PCM proteins known, the molecular characterization of centrioles is still very limited. Only a few centriolar proteins have been identified so far in Drosophila, most related to centriole duplication. asterless (asl) has been cloned and found to encode a 120 kD highly coiled-coil protein that is a constitutive pancentriolar and basal body component. Loss of asl function impedes the stabilization/maintenance of PCM at the centrosome. In embryos deficient for Asl, development is arrested right after fertilization. Asl shares significant homology with Cep 152, a protein described as a component of the human centrosome for which no functional data is yet available. The cloning of asl offers new insight into the molecular composition of Drosophila centrioles and a possible model for the role of its human homolog. In addition, the phenotype of asl-deficient flies reveals that a functional centrosome is required for Drosophila embryo development (Varmark, 2007).
Asl functions during the first stages of zygotic development. After sperm entry and activation, female meiosis is resumed and four haploid nuclei are produced in wild-type eggs. The most internal of these nuclei, which is the closest to the sperm nucleus, becomes the female pronucleus, while the others, usually clustered into a polar body, remain inactive and eventually disappear. Recruitment of PCM around the paternally contributed centriole results in the assembly of the first zygotic centrosome and in the organization of a prominent aster, which is thought to mediate male and female pronuclear fusion. Soon afterwards, the duplicated centrosomes migrate apart over the male pronucleus, and fusion with the female pronucleus takes place. The first mitotic spindle is then assembled, and repeated rounds of nuclear division cycles result in the exponential proliferation of syncytial nuclei. In embryos derived from asl1/Df(3R)ED1557 mothers, female meiosis proceeds normally: distinct polar bodies and female pronuclei can be identified, sperm entry takes place, and upon fertilization, all five nuclei are present and arranged in a seemingly wild-type configuration. Cnn accumulates at a point near the male pronucleus, presumably around the paternally provided basal body that is wild-type for Asl, but a functional MTOC is not organized, development is brought to a halt, and the first zygotic mitosis never occurs. Instead, nonfunctional, anastral spindle-shaped structures are organized around the chromatin. These spindles, which do not segregate chromosomes, persist in embryos aged for 1-2 hr, a stage at which wild-type embryos contain hundreds of nuclei. Thus, maternal Asl is needed to facilitate the centrosome function required for initiation of cleavage cycles in the fly. This terminal phenotype is indistinguishable from the phenotype of embryos derived from γTUBTW1 homozygous females, which lack the maternal γTUB37C gene (Varmark, 2007).
The oocyte is the only cell in Drosophila that goes through
meiosis with meiotic recombination, but several germ cells
in a 16-cell cyst enter meiosis and form synaptonemal
complexes (SC) before one cell is selected to become the
oocyte. Using an antibody that recognises a component of
the SC or the synapsed chromosomes, an analysis was carried out of how meiosis becomes restricted to one cell, in relation to
the other events in oocyte determination. Although Bicaudal-D
and egalitarian mutants both cause the development of cysts with
no oocyte, they have opposite effects on the behavior of
the SC: none of the cells in the cyst form SC in BicD null
mutants, whereas all of the cells do in egl and orb mutants.
Furthermore, unlike all cytoplasmic markers for the
oocyte, the SC still becomes restricted to one cell when the
microtubules are depolymerised, even though the BicD/Egl
complex is not localised. These results have lead to the proposal of a
model in which BicD, Egl and Orb control entry into
meiosis by regulating translation (Huynh, 2000).
In the course of a study on the role of inscuteable (insc) during
oogenesis, it has been found that an anti Insc antibody recognizes a nuclear
structure that is present in some of the germ cells in regions
2a to 3 of the germarium. However, this staining does
not disappear in germline clones of protein null allele insc 22,
ndicating that it is due to a cross-reaction of the antibody. Nevertheless, the staining pattern is very reminiscent of that expected for a component of the
synaptonemal complex (SC), and therefore the staining was analyzed
further, since this would be the first marker identified for the SC
structure in Drosophila. Several lines of evidence indicate that the antiserum does
indeed label the SC or a component associated with its formation. (1) The nuclear staining colocalizes with DNA, and has a morphology that corresponds exactly with the observed behaviour of the SC in electron micrographs. The staining is dotty in very early region 2a when the SC starts to form, becomes more thin and thread-like when the chromosomes are fully synapsed, and then becomes more compact in region 3, when the meiotic chromosomes condense to form the karyosome. (2) This structure
first appears at the stage when the cysts enter into meiosis. The mitotic cysts in region 1 of the germarium express Bam protein, but this disappears after the final division when the cysts move from region 1 to region 2a of the
germarium. The nuclear staining is only detectable in cysts that no longer show any Bam expression, indicating that it labels a postmitotic
structure. (3) The spatial distribution of the signal within the cyst precisely
follows that described for the SC at the EM level. The signal first appears in two cells in early region 2a and spreads to four cells per cyst in the middle of 2a, before it is restricted to two cells, and finally to one cell in region 2b. Ovaries from females that are mutant for C(3)G were examined, since these are the only characterized
mutants that completely abolish the formation of the
SC at the electron microscope level. C(3)G encodes the fly
homolog of yeast Zip1 and mammalian SCP1,
mponents of the transverse filament of the SC, and the effects
of the C(3)G mutation on the SC are therefore likely to be direct
(Szauter and Hawley, personal communication to Huynh, 2000). The nuclear
structure stained by this antibody is absent in C(3)G mutant cysts, even though the localization of Orb protein to the oocyte occurs normally. Thus, the
antibody acts as a marker for the formation of the SC, although
the molecular nature of the epitope recognized is not known (Huynh, 2000).
A detailed analysis of the behavior of the SC in comparison
to that of cytoplasmic markers for oocyte determination, such as
Orb and Bic-D proteins, reveals a number of distinct steps in the
restriction of oocyte fate to one cell. The SC first
appears in early region 2a cysts in the nuclei of two cells, which
are presumably the pro-oocytes. The punctate appearance of the
SC suggests that they are at the zygotene stage of meiotic
prophase 1. The next one or two cysts per germarium have four
cells in synapsis. Two of these cells have four ring canals (the
pro-oocytes) and contain an almost continuous SC, typical of the
pachytene stage, while the two cells on either side, presumably
the cells with three ring canals, contain a zygotene-like SC. In the
midle of region 2a, the SC disappears from the two cells with
three ring canals, but the two pro-oocytes still have complete
SCs, and accumulate Orb and Bic-D proteins. Soon afterwards,
Orb and Bic-D become concentrated in only one of these cells,
providing the first sign that this pro-oocyte has been selected to
become the oocyte. However, the SC still appears identical in
both pro-oocytes at this stage. The SC disappears from
one pro-oocyte as the cyst enters region 2b, and the cell that
remains in meiosis is always the one that has already
accumulated Orb or Bic-D. Finally, SC becomes more compact
in region 3 and a hole forms in its middle, before it disappears
soon after the cyst leaves the germarium.
This comparison of the behavior of nuclear and
cytoplasmic markers for the oocyte reveals two important
features about how oocyte fate becomes restricted to one cell.
(1) The two pro-oocytes are already different from the other
14 cells in the cyst in early region 2a, as they both start to form
SC at this stage. BicD and Orb only accumulate in these cells
in mid 2a, about two cysts further down the germarium.
(2) Orb and Bic-D become restricted to the oocyte
before any sign of oocyte identity can be deduced from the
behavior of the SC (Huynh, 2000).
A cyst can progress through the normal pattern of SC
localization to one cell in the presence of high concentrations
of colcemid, suggesting restriction of SC to one cell is not mediated by microtubules. Unlike the microtubules, BicD, orb and egl mutations disrupt
all steps in the restriction of the SC to one cell, and this leads
to two important conclusions: (1) BicD and Egl must have
a function that is independent of microtubules, even though
they are required for the establishment or maintenance of the
MTOC in the oocyte; (2) this
function of BicD, Egl and Orb does not depend on their own
localisation to the oocyte, since all three proteins are
completely delocalized after colcemid treatments, yet the SC
still becomes restricted to one cell.
Although both BicD and egl mutations give rise to cysts in
which all 16 cells appear identical, they have different effects
on the behavior of the SC itself. In BicD null germline clones,
none of the cells form a detectable SC, whereas all cells reach
the full pachytene stage in egl mutants (Huynh, 2000).
The discovery that the restriction of SC to one cell requires
neither microtubules nor the localization of BicD, Egl and Orb
raises the question of how this asymmetry arises. It has
previously been suggested that BicD and Egl function in the
transport of meiosis promoting factors and oocyte determinants
from the future nurse cells into the oocyte. Although this could
still be the case if this transport occurs either very early in
region 2a or along some non-microtubule cytoskeletal network,
such as actin, this model cannot easily explain why BicD and
egl mutations have opposite effects on SC formation. An alternative model is preferred in which BicD, Egl and
Orb are required to interpret a pre-existing asymmetry that is
set up in region 1 (Huynh, 2000 and references therein).
The divisions that give rise to the cyst are asymmetric with
respect to the fusome, and recent data strongly suggest that this
structure, or some factor associated with it, somehow marks
the future oocyte. If this is
correct, this unidentified mark could regulate the BicD/Egl
complex, so that it performs different functions in the different
cells of the cyst. For example, the Egl-dependent activity of the
complex could repress SC in the cells that do not inherit the
factor, and the BicD-dependent activity could enhance its
formation in the cells that do, thereby explaining the different
phenotypes of the null mutations in the two genes. It is
interesting to note that BicD protein is phosphorylated, and that
mutations that disrupt this phosphorylation give rise to egg
chambers with 16 nurse cells. Thus,
this post-translational modification could be responsible for the
spatial regulation of the activity of the BicD/Egl complex (Huynh, 2000).
Although these results suggest that BicD and Egl have
functions that are independent of the microtubules, the nature
of this activity is unclear. However, a number of lines of
evidence suggest that these proteins may be involved in
translational control. (1) BicD was originally identified
because two single amino acid changes in the gene produce a
dominant bicaudal phenotype in which Oskar mRNA is mis-expressed
at the anterior of the oocyte. Since Oskar translation is
normally repressed unless the RNA is localized to the posterior
pole, these mutant BicD proteins must not only trap Oskar RNA
at the anterior, but also relieve translational repression.
Mutations in egl suppress the BicD gain-of-function
phenotype, while extra copies of egl enhance it, indicating that
the ectopic translation of Oskar mRNA requires the formation
of the BicD/Egl complex. The second argument for a role of BicD and Egl in
translational control comes from the discovery that orb null
mutations give a very similar phenotype to egl mutants. Orb
protein, which contains two RNA-binding motifs, has recently
been shown to associate with the 3'UTR of Oskar mRNA, and
is required for its efficient translation. Similarly, the Xenopus Orb homolog,
CPEB,binds to elements in the 3'UTRs of a number of
mRNAs, and induces the polyadenylation and translational
activation of these mRNAs during oocyte maturation. Furthermore, the Spisula solidissima (clam)
homolog plays a role not only in translational activation, but
also in repression, since it binds to masking elements in the
3'UTRs of cyclin mRNAs to prevent their translation before
fertilization. Thus,
Orb functions as a regulator of translation, and can act as both
a repressor and an activator in other species. This raises the
possibility that the BicD/Egl complex exerts different effects
in the cells of the cyst by controlling the inhibitory and
activating functions of Orb. For example, Orb could repress the
translation of factors required for SC formation in the future
nurse cells, and activate their translation in the pro-oocytes and
oocyte. If this model is correct, the selection of the oocyte
would occur by a similar mechanism to the other asymmetries
that are generated later in oogenesis, which are also all based
on the translational regulation of asymmetrically localized
mRNAs, such as Bicoid, Gurken and Oskar (Huynh, 2000 and references therein).
The behavior of the SC indicates that the determination of the
oocyte occurs in two steps. The two pro-oocytes must have
been selected by early region 2a, because they already behave
differently from the other 14 cells of the cyst at this stage, but
the development of the cyst remains symmetric until the end
of 2a, when BicD and Orb disappear from the losing pro-oocyte.
It has been proposed that the choice between
the two pro-oocytes could depend on competition between
these cells as they progress through meiosis, with the cell that
is more advanced becoming the oocyte and then inhibiting its
neighbor. However, the results presented here argue against this model:
(1) cytoplasmic factors, such as BicD and Orb, are
concentrated in one cell before there is any visible difference
between the SCs in the two pro-oocytes; (2) the
cytoplasmic aspects of oocyte determination occur normally in
C(3)G mutants, which completely lack the SC, and
in meiW68 mutants, which fail to initiate
meiotic recombination. Thus, any
competition between these two cells must be independent of
SC formation and recombination (Huynh, 2000 and references therein).
Although meiosis is not required for oocyte determination,
it can clearly influence this process, as demonstrated by the
results on the spn genes. Several lines of evidence indicate that
mutations in spnB, C and D disrupt the repair of dsDNA breaks
during meiotic recombination, activating a checkpoint pathway
that inhibits Gurken mRNA translation and the formation of the
karyosome. The results presented here strongly suggest that this checkpoint also
inhibits the determination of the oocyte, since the SC becomes
restricted to one cell much later than in wild type in these
mutants. This phenotype also allows the
time when recombination occurs to be narrowed down. This process cannot begin
until the SC forms in early region 2a, but the double-strand
DNA breaks have to be repaired before the two cells with three
ring canals exit meiosis, since this stage is delayed in spnC
mutants, indicating that the checkpoint pathway has already
been activated (Huynh, 2000).
Activation of the meiotic checkpoint causes a change in the
mobility of Vasa protein, leading to the suggestion that the
patterning defects seen in spn mutants result from the
inhibition of Vasa by this pathway. The results presented here show that the SC becomes restricted to one
cell at the normal time in most vasa mutant cysts. Thus, the
delay in oocyte determination in spn mutants cannot be a
consequence of the inhibition of Vasa, suggesting that the
checkpoint pathway has additional targets that control oocyte
selection (Huynh, 2000).
One problem in the study of cyst development in region 2 has
been the difficulty in ordering the various developmental
events that occur in this region. Using this marker for the SC,
the behavior of this structure
relative to the localization of cytoplasmic factors like Orb and
BicD could be followed, and these could be correlated with the data from EM studies on
the behavior of the SC, and the centrioles. On the
basis of this comparison, a number of
distinct stages in the restriction of oocyte fate to one cell can be distinguished: (1) The
first cyst in region 2a shows no sign of SC, but
Bam protein has already disappeared. (2) The two pro-oocytes
reach the zygotene stage of meiosis in early region 2a, and
start to form SC. (3) Soon afterwards, the
two cells with three ring canals also form SC. The SC in the
pro-oocytes has reached its maximum length, indicating that
they have reached the pachytene stage. The dsDNA breaks
generated during recombination must have already been
repaired, since the meiotic checkpoint can arrest the pattern of
SC staining at this stage. EM data also suggest that
intracellular transport begins at this point, since the first signs
of the migration of the centrioles towards the pro-oocytes
can be seen when the two cells with three ring canals
are in meiosis, and this may correlate with the first appearance
of a focus of microtubules in the cyst in the middle of region
2a. (4) The SC disappears from the two
cells with three ring canals in the middle of region 2a, but the two
pro-oocytes still have complete SCs. Orb and Bic-D
start to accumulate in the pro-oocytes at this stage.
The centrioles have migrated to either side of the largest ring
canal, which connects the two pro-oocytes, and the first signs
of 'nutrient streaming' appear, since elongated mitochondria
can be seen inside the ring canals in electron micrographs. (5)
All of the steps in cyst development so far are symmetric,
relative the largest ring canal, and the first asymmetry
becomes evident in cysts numbers 5 and 6, when Orb and Bic-D
become concentrated in one cell. The centrioles also start
to move into the oocyte, and the largest ring canal is
presumably open, because mitochondria can now be seen
inside it. However, both pro-oocytes still contain an identical
intact SC at this stage. (6) As the cyst enters region 2b, one
pro-oocyte loses its SC and reverts to the nurse cell pathway
of development. The pro-oocyte that remains in
meiosis and becomes the oocyte is always the cell that has
already accumulated Orb and Bic-D. The cytoplasm of the
oocyte now contains all of the centrioles, BicD and Orb
proteins, and an obvious MTOC, which nucleates
microtubules that extend into the other 15 cells of the
cyst. Thus, both the nucleus and cytoplasm of the oocyte are
clearly different from the other cells of the cyst by this stage.
Immediately afterwards, the oocyte starts to behave differently
from the other cells in the cyst, as it moves to the posterior
during the transition between region 2b and region 3. At
the same time, the karyosome forms, and the SC becomes
more compact, before disappearing soon after the cyst leaves
the germarium (Huynh, 2000).
At the onset of meiosis, each chromosome needs to find its homologue and pair to ensure proper segregation. In Drosophila, pairing occurs during the mitotic cycles preceding meiosis.This study shows that germ cell nuclei undergo marked movements during this developmental window. Microtubules and Dynein are driving nuclear rotations and are required for centromere pairing and clustering. It was further found that Klaroid (SUN) and Klarsicht (KASH) co-localize with centromeres at the nuclear envelope and are required for proper chromosome motions and pairing. Mud (NuMA in vertebrates) was identified as co-localizing with centromeres, Klarsicht and Klaroid. Mud is also required to maintain the integrity of the nuclear envelope and for the correct assembly of the synaptonemal complex. These findings reveal a mechanism for chromosome pairing in Drosophila, and indicate that microtubules, centrosomes and associated proteins play a crucial role in the dynamic organization of chromosomes inside the nucleus (Christophorou, 2015).
Assembly of the synaptonemal complex (SC) in Drosophila depends on two independent pathways defined by the chromosome axis proteins C(2)M and ORD. Because C(2)M encodes a Kleisin-like protein and ORD is required for sister-chromatid cohesion, the hypothesis was tested that these two SC assembly pathways depend on two cohesin complexes. Through single- and double-mutant analysis to study the mitotic cohesion proteins Stromalin (SA) and Nipped-B (SCC2) in meiosis, evidence was provided that there are at least two meiosis-specific cohesin complexes. One complex depends on C(2)M, SA, and Nipped-B. Despite the presence of mitotic cohesins SA and Nipped-B, this pathway has only a minor role in meiotic sister-centromere cohesion and is primarily required for homolog interactions. C(2)M is continuously incorporated into pachytene chromosomes even though SC assembly is complete. In contrast, the second complex, which depends on meiosis-specific proteins SOLO, SUNN, and ORD is required for sister-chromatid cohesion, localizes to the centromeres and is not incorporated during prophase. Multiple cohesin complexes may provide the diversity of activities required by the meiotic cell (Gyuricza, 2016).
Based on similar mutant phenotypes and double-mutant analysis, it is proposed that SC assembly in Drosophila depends on two meiotic cohesin complexes. The first includes C(2)M, SA, and Nipped-B. The most important function of C(2)M/SA/Nipped-B is SC assembly, which is demonstrated by the more significant SC assembly defects observed in c(2)M mutants and SA or Nipped-B knockdowns compared to sunn, solo, or ord mutants. The cytological results suggest that, like C(2)M, SA has only a minor role in meiotic sister-centromere cohesion. Correlating with this difference is that C(2)M, SA, or Nipped-B are required for the accumulation of SMC proteins on the chromosome arms but not the centromeres. Furthermore, Nipped-B, like C(2)M, localizes to the chromosome arms but not the centromeres. These observations indicate a significant change in cohesin regulation. While SA and Nipped-B are required for sister-chromatid cohesion in mitotic cells, they have a new partner, C(2)M, for a non-cohesion function in meiosis. There are minor differences in the c(2)M and SA phenotypes, which has also been observed with solo and sunn, suggesting there could be additional minor complexes. SA and Nipped-B could be required to maintain sister-chromatid cohesion on the chromosome arms in late prophase, a function that C(2)M likely does not have (Gyuricza, 2016).
The second proposed meiotic cohesin complex includes SOLO, SUNN, and ORD, which are also highly diverged, making homology assignments difficult. Based on sequence features, SUNN may be a SA homolog, while SOLO has been shown to interact with SMC1 and to have sequence motifs similar to the SMC1 interaction domains of kleisins. The role of ORD in this context is unclear. It is possible that ORD is a positive regulator like Nipped-B. Genetic evidence shows that ord, sunn, and solo are required for sister-chromatid cohesion, which correlates with SMC1/3 and SC accumulation at the centromeres. In addition, there are elevated levels of sister-chromatid exchange and abnormal SC structure in ord and solo mutants (Gyuricza, 2016).
Surprisingly, this study found an important role for the meiotic cohesins SUNN and ORD in mitotic germline cells, which is consistent with prior observations that ORD localizes to centromeric foci in premeiotic cells and ord mutants have defects in mitotically dividing germline cells. Since Rec8 in C. elegans is also observed in premeiotic cells, it may be a conserved feature of meiotic cohesins required for sister-chromatid cohesion that they accumulate and function in premeiotic mitotic germline cells (Gyuricza, 2016).
A striking difference between C(2)M/SA and SOLO/SUNN is that C(2)M incorporation is continuous during pachytene even after the SC is fully assembled. This may result in a dynamic SC, which has been observed in budding yeast. Paradoxically, this study also found that C(2)M must load during a narrow window of early prophase in order to support crossover formation. Cohesins have been shown to be loaded during prophase in a number of systems including Drosophila. In contrast, centromeric SOLO/SUNN can only be loaded prior to meiotic prophase. Sister-chromatid cohesion in mitotic cells is established during S-phase and in mammals, Rec8 cohesin cannot be replenished and dissociates with age. Whereas in mitotic cells the dynamic and stable cohesin complexes involve the same four core subunits, in meiosis, there may be separate cohesin complexes that differ in their regulation and capacity to be replaced or replenished. These observations complement Weng (2014) who showed there was cohesin replenishment during meiotic prophase, although after pachytene and possibly not at the centromeres. Interestingly, mouse Nipbl (Nipped-B) and the meiosis-specific SMC1β show pronounced accumulation starting at leptotene, indicating that, as in Drosophila, some mouse cohesins are loaded during pachytene while other cohesins are stable (Gyuricza, 2016).
Meiosis-specific cohesin complexes appear to be a highly conserved feature of meiosis. While there is some variation in the constituents of each cohesin complex, the results suggesting two major pathways contributing to SC assembly help explain the results with coh-3/coh-4 and rec-8 in C. elegans or rad21l and rec8 in mouse. Only the double mutants in each case eliminate all SC. The role of the respective kleisins could also be conserved. Like C(2)M, Rad21L has been proposed to be primarily responsible for inter-homolog chromosome interactions. Multiple cohesin complexes may be required because some cohesins need to be loaded at a specific time (S-phase for cohesion) while others need to be exchanged during pachytene. A dynamic cohesin complex may be important to provide plasticity to the meiotic chromosomes and allow them to respond to double-stranded breaks (DSBs) and regulate crossover formation, crossover interference, and chromosome segregation. Alternatively, different cohesin complexes may accumulate at different locations. If meiotic cohesins, directly or indirectly, interact with SC proteins, they may have a strong influence on the pattern of SC assembly and influence the frequency and distribution of DSBs and crossovers (Gyuricza, 2016).
Formation of the synaptonemal complex (SC), or synapsis, between homologs in meiosis is essential for crossing over and chromosome segregation. How SC assembly initiates is poorly understood but may have a critical role in ensuring synapsis between homologs and regulating double-strand break (DSB) and crossover formation. This study investigated the genetic requirements for synapsis in Drosophila and found that there are three temporally and genetically distinct stages of synapsis initiation. In meiotic prophase 1 'early zygotene' oocytes, synapsis is only observed at the centromeres. It was also found that nonhomologous centromeres are clustered during this process. In 'mid-zygotene' oocytes, SC initiates at several euchromatic sites. The centromeric and first euchromatic SC initiation sites depend on the cohesion protein ORD. In 'late zygotene' oocytes, SC initiates at many more sites that depend on the Kleisin-like protein C(2)M. Surprisingly, late zygotene synapsis initiation events are independent of the earlier mid-zygotene events, whereas both mid and late synapsis initiation events depend on the cohesin subunits SMC1 and SMC3. It is proposed that the enrichment of cohesion proteins at specific sites promotes homolog interactions and the initiation of euchromatic SC assembly independent of DSBs. Furthermore, the early euchromatic SC initiation events at mid-zygotene may be required for DSBs to be repaired as crossovers (Tanneti, 2011).
Drosophila pro-oocytes develop within 16-cell cysts that are
arranged in temporal order within the ovary. Each ovary
contains several germaria, where pairs of pro-oocytes begin
their development and enter prophase in region 2a and a single
oocyte is selected by region 3. Oocytes are defined
by the presence of the synaptonemal complex (SC), which is
detected by antibodies to the transverse element C(3)G (Page, 2001),
a coiled-coil protein similar to proteins in budding yeast
(ZIP1), C. elegans (SYP-1, SYP-2), and mammals (SYCP1)
(Page, 2004; Watts, 2011). Zygotene pro-oocytes were identified by their patchy
C(3)G staining, as opposed to the thread-like staining typical
of pachytene. Furthermore, by comparing the amount of
synapsis to the relative positions of the pro-oocytes in the
wild-type germarium, three stages of zygotene were defined (Tanneti, 2011).
First, early zygotene pro-oocytes have one or two patches of
C(3)G that colocalize with CID, a centromere-specific histone H3. These pro-oocytes reside in the earliest
(most anterior) part of region 2a, indicating that synapsis initiates
at the centromeres before any other sites. These results
were confirmed by comparing CID localization to histone
modifications specific for the heterochromatin or euchromatin. Because there are four pairs of centromeres, the observation that most wild-type pro-oocytes
have one or two CID foci indicates that nonhomologous
centromeres cluster in meiotic prophase, confirming
previous observations using electron microscopy (Tanneti, 2011).
Second, mid-zygotene pro-oocytes have the centromeric
C(3)G staining plus approximately six additional sites in the euchromatin. Finally,
late zygotene pro-oocytes contain many C(3)G foci but lack
the continuous threadlike pattern of pachytene. Surprisingly,
the mid-zygotene patches do not appear to get longer.
Instead, there are more patches in late zygotene, suggesting
that the progression from mid- to late zygotene involves the
establishment of new SC initiation sites rather than polymerization
from the small number of sites in mid-zygotene. It is
suggested that the noncentromeric C(3)G sites in mid-zygotene
represent the first euchromatic sites to initiate synapsis. This study provides evidence that the mid-zygotene sites have features in common with centromere synapsis sites but are mechanistically distinct and genetically separable from the additional synapsis initiation sites observed in late zygotene (Tanneti, 2011).
C(2)M is a lateral element component and is a member of the Kleisen family that includes Rec8 and Rad21 homologs (Schleiffer, 2003). In wild-type, C(2)M colocalizes with C(3)G in most locations except at the centromeres. In
females lacking C(2)M, the first two stages of zygotene
appear to occur normally. Early zygotene pro-oocytes exhibit one or two foci of CID that colocalize with C(3)G, showing that C(2)M is not required for
centromere clustering or centromere synapsis. These results
confirm previous observations (Khetani, 2007) that C(2)M is not required
for centromere clustering in pachytene oocytes and are
consistent with the observation that C(2)M does not localize
to the centromeric regions. Early zygotene
in c(2)M mutants is followed by cysts with several
patches of euchromatic C(3)G staining that resemble wildtype
cells in mid-zygotene. Synapsis in a c(2)M mutant does not, however, progress beyond this point. Examination of histone modifications
in c(2)M mutants confirmed that synapsis is
blocked in mid-zygotene with a small number of euchromatin
initiation sites. Based on the similarities between wild-type
mid-zygotene and c(2)M mutants, it is suggested that synapsis
initiates in a c(2)M-independent manner at a small number
of specialized sites on the chromosomes, which include
approximately six euchromatic sites and the centromeres, and that C(2)M is required for additional initiation sites typical of late zygotene (Tanneti, 2011).
There is a striking similarity between the number of euchromatic
synapsis initiation sites (~6) during mid-zygotene and
the number of crossovers in Drosophila females. In
order to determine the relationship between SC initiation sites
and double-strand break (DSB) formation, c(2)M mutant
oocytes were stained for C(3)G and γ-H2AV.
DSBs in a c(2)M mutant are usually associated with a patch
of C(3)G staining (55/56 γ-H2AV foci were touching or overlapped
a patch of C(3)G). This experiment was also performed in
an okr mutant background (okr encodes the Drosophila homolog of Rad54) where the DSBs are not repaired and
γ-H2AV staining accumulates, allowing all DSBs to be counted. Most of the γ-H2AV foci in okr c(2)M mutant
germaria colocalized with a patch of C(3)G, suggesting
that the initiation of SC and recombination usually occur within
the same region in c(2)M mutants. Indeed, MEI-P22, a protein required for DSB formation, also colocalizes with the SC in
c(2)M mutant oocytes. It should be noted
that previous observations showed that DSB formation is
partially dependent on the SC. Indeed, the number of
γ-H2AV foci in the okr c(2)M double mutant in region 3 oocytes
was reduced compared to a okr single mutant. Overall, these results suggest that the SC, or a factor which stimulates SC formation, promotes
recruitment of proteins required for DSB formation (Tanneti, 2011).
To investigate whether there is a connection between early
SC initiation events and meiotic recombination, double
mutants with c(2)M were constructed. Unlike wild-type, where
γ-H2AV foci are not observed until pachytene, the block
in synapsis observed in c(2)M mutants allowed examination of
the relationship between SC initiation and DSB formation. By
double staining with CID, it was found that eliminating meiotic
DSBs with a mei-W68 mutation did not prevent formation of
either the centromere and euchromatic SC in a c(2)M mutant. The small decrease in the number of euchromatic SC sites in the c(2)M mei-W68
double mutant may indicate that the number of initiation sites
is sensitive to DSB formation. Furthermore, SC initiation is not
grossly affected by a reduction in crossing over (mei-218), an increase in crossing over (TM6), or a defect in DSB repair (okr). DSBs do not occur in the heterochromatin; thus, it is not surprising that centromere SC is independent of DSB formation. However, these results show that the initiation of euchromatic synapsis at mid zygotene does not depend on DSBs or crossovers (Tanneti, 2011).
Because DSBs or recombination are not required for synapsis in wild-type or c(2)M mutants, tests were performed to see whether structural components of the meiotic chromosomes regulate SC initiation. ORD is a meiosis-specific protein required for cohesion and crossover formation that may be a component of the SC
lateral elements. Although previous studies have shown that ord mutant oocytes generate threads of C(3)G staining that resemble pachytene, the effect of ord on zygotene progression has not been previously examined (Tanneti, 2011).
Consistent with previous results, this study found that centromere
clustering is defective and the association of
SC proteins with the centromeres is disrupted in ord mutant
oocytes. Furthermore, zygotene appeared abnormal;
rather than observing centromeric and euchromatic SC
initiation sites typical of mid-zygotene in early region 2a, it was
found that many ord mutant pro-oocytes with C(3)G staining only
around the nuclear DNA. Of the 108 pro-oocytes examined
in five germaria, 36 (33%) had no nuclear C(3)G. The remaining
pro-oocytes [72, (67%)] either had a number of C(3)G patches
that was more typical of late zygotene, usually in region 2a, or were in pachytene. It is concluded that the centromeric and euchromatic synapsis sites typical
of early and mid zygotene are absent in ord mutants, suggesting
that, in the absence of ORD, synapsis does not initiate normally (Tanneti, 2011).
Because ord mutants do eventually form threads of SC, it
was difficult to be sure that SC initiation was defective. To
test whether ord has a role in mid-zygotene synapsis, tests were performed to see whether the euchromatic patches of C(3)G in a c(2)M
mutant depend on ord. Even though both
single mutants exhibit at least some SC formation, most of the C(3)G staining in the c(2)M ord double mutant surrounded the DNA and within the nucleus.
This nonchromosomal C(3)G localization in the c(2)M ord double mutant was much more pronounced than in the ord single mutant. In
addition, C(3)G-staining ring-like structures were observed similar to what has been reported in some c(3)G missense mutants. All the nonchromosomal C(3)G staining may be due to polycomplex formation. c(2)M ord double mutant pro-oocytes were identified by the prominent C(3)G around the DNA, and the number of C(3)G patches on the chromosomes was found to be drastically reduced compared to wild-type zygotene or either single mutant (Tanneti, 2011).
These results demonstrate that ord is required
for the centromeric and euchromatic synapsis
sites observed in c(2)M mutants. Conversely,
C(2)M is required for the threadlike synapsis
observed in ord mutants. The synergistic
phenotype of the double mutant suggests that there are two
types of synapsis initiation - one depends on ORD (early and mid-zygotene) and the other depends on C(2)M (late zygotene) - and that these are independent events. In the absence of both types of synapsis initiation, C(3)G cannot load onto the chromosomes and accumulates in polycomplexes (Tanneti, 2011).
Like other Kleisin family members, C(2)M has been shown to
physically interact with the cohesin subunit SMC3 (Heidmann, 2004). To
determine whether C(2)M localization depends on an interaction
with cohesin, oocytes lacking SMC1 and
SMC3 were examined. To examine oocytes lacking SMC3 (encoded by cap), the recently developed short hairpin RNA (shRNA)
resource, which allows RNA interference (RNAi) knockdown
of gene expression in the Drosophila female germline, was used. Both the chromosomal
localization of C(3)G and C(2)M were absent when cap shRNA was expressed in the germline. Furthermore, SMC1 staining was eliminated, suggesting that the RNAi was effective at knocking out SMC3 function. Like the c(2)M ord double mutant, most C(3)G staining accumulated around the periphery of the DNA,
suggesting that the function of SMC3 in synapsis occurs
through at least two independent interactions with C(2)M and
ORD. Unlike the c(2)M ord double mutant, however, it was not possible to distinguish the pro-oocytes from the nurse cells because C(3)G staining was evenly distributed among the cells in each germarium cyst. Importantly, oocyte selection was not perturbed because one cell in each cyst accumulated ORB
protein, a cytoplasmic marker for the oocyte. Thus, the loss of SMC3 may have a more severe phenotype than the c(2)M ord double mutant (Tanneti, 2011).
These results were confirmed with the analysis of SMC1 mutant germline clones. As with cap RNAi, there was an absence of nuclear C(2)M and C(3)G threads in oocytes lacking SMC1, indicating a complete block in synapsis. Also similar to cap RNAi, the accumulation of ORB in one cell indicated that an oocyte was established. The only difference compared to cap RNAi was that there was much less C(3)G staining around the periphery of the DNA. It is not known whether this minor difference is due to the different methods (RNAi versus germline clone) or distinct functions of the two SMC proteins. Nevertheless, the results of these two experiments demonstrate that SMC1 and SMC3 are required for synapsis (Tanneti, 2011).
It is concluded that synapsis initiation during zygotene in
Drosophila females occurs in three stages. In early zygotene, the centromeres are the first sites to accumulate the transverse filament protein C(3)G. Indeed, cohesion proteins SMC1, SMC3, and ORD are detected at the centromeres before meiotic prophase (prior to or during premeiotic S phase), which could explain why synapsis is first observed at the centromeres. Interestingly, the SC also
forms first at the centromeres in budding yeast and depends on cohesion proteins. In mid-zygotene, synapsis initiates at a small number of euchromatic sites. These first two steps depend on the ORD protein. Finally, in late zygotene, synapsis initiates at a larger number of euchromatic sites. This stage requires C(2)M and appears to occur through a new set of initiation events rather than extending synapsis, or 'zipping up,' from the mid-zygotene initiation sites. Indeed, the synapsis initiation events in mid and late zygotene are independent and genetically separable, supporting a model where synapsis occurs through two independent waves of initiation events. In the absence of ORD, early and mid-zygotene synapsis events are skipped and the late zygotene initiation events occur with normal kinetics. This is not without
consequence, however, because at the electron microscopy
level, this synapsis is abnormal and tripartite SC is not visible. Both waves of synapsis initiation depend on the cohesin proteins SMC1 and SMC3, which may interact independently with C(2)M and ORD (Tanneti, 2011).
In addition to its role in centromere synapsis, ORD and the
SMC proteins are required for the pairing and clustering of
centromeres, whereas the SC components C(2)M or C(3)G
are not. Thus, cohesion proteins may be able to function in
a pairing role independent of DSBs, as Rec8 does in budding
yeast for centromere coupling. It is suggested that the first
euchromatic sites to initiate SC assembly in Drosophila are
in regions where cohesion proteins are most abundant. This
model is attractive because it provides a mechanism for SC
initiation in the absence of DSBs. Interestingly, the number
of euchromatic initiation sites in mid-zygotene or in c(2)M
mutants approximates the number of crossovers in the
genome. Not only do these mid-zygotene sites depend on ORD, but in ord mutants, crossing over is reduced to less than 10% of wild-type, even though DSBs occur normally. It is suggested that the reduction in crossing over in
ord mutants is due to the absence of the synapsis initiation
sites at mid-zygotene. Whether the synapsis initiation sites
actually correspond to crossover sites awaits further study (Tanneti, 2011).
ORD may have a function similar to yeast Rec8 because it is required
for synapsis at the centromeres and a subset of euchromatic
sites. Interestingly, the findings with C(2)M, which is not an
ortholog of Rec8, are also probably relevant to other species.
Several recent studies have revealed Non-Rec8 Kleisin
homologs in mouse and C. elegans (COH-3 and COH-). These parallels between the synapsis pathway in flies and that of organisms that depend on DSBs
for synapsis could reflect the existence of a conserved underlying
mechanism of synapsis. If synapsis initiation sites can
be marked prior to DSB formation in a process involving cohesion
proteins, and if proteins like Zip3 can be recruited in the
absence of DSBs, as is true in C. elegans and likely in
Drosophila, the timing of the DSB then becomes less of a determining
factor in the process of synapsis (Tanneti, 2011).
Sister centromere fusion is a process unique to meiosis that promotes co-orientation of the sister kinetochores, ensuring they attach to microtubules from the same pole during metaphase I. This study found that the kinetochore protein SPC105R/KNL1 and Protein Phosphatase 1 (PP1-87B) regulate sister centromere fusion in Drosophila oocytes. The analysis of these two proteins, however, has shown that two independent mechanisms maintain sister centromere fusion. Maintenance of sister centromere fusion by SPC105R depends on Separase, suggesting cohesin proteins must be maintained at the core centromeres. In contrast, maintenance of sister centromere fusion by PP1-87B does not depend on either Separase or WAPL. Instead, PP1-87B maintains sister centromeres fusion by regulating microtubule dynamics. This study has demonstrated that this regulation is through antagonizing Polo kinase and BubR1, two proteins known to promote stability of kinetochore-microtubule (KT-MT) attachments, suggesting that PP1-87B maintains sister centromere fusion by inhibiting stable KT-MT attachments. Surprisingly, C(3)G, the transverse element of the synaptonemal complex (SC), is also required for centromere separation in Pp1-87B RNAi oocytes. This is evidence for a functional role of centromeric SC in the meiotic divisions, that might involve regulating microtubule dynamics. Together, this study proposes that two mechanisms maintain co-orientation in Drosophila oocytes: one involves SPC105R to protect cohesins at sister centromeres and another involves PP1-87B to regulate spindle forces at end-on attachments (Wang, 2019).
The necessity of sister kinetochores to co-orient toward the same pole for co-segregation at anaphase I differentiates the first meiotic division from the second division. A meiosis-specific mechanism exists that ensures sister chromatid co-segregation by rearranging sister kinetochores, aligning them next to each other and facilitating microtubule attachments to the same pole]. This process is referred to as co-orientation, in contrast to mono-orientation, when homologous kinetochores orient to the same pole. Given the importance of co-orientation in meiosis the mechanism underlying this process is still poorly understood, maybe because many of the essential proteins are not conserved across phyla (Wang, 2019).
Most studies of co-orientation have focused on how fusion of the centromeres and kinetochores is established. In budding yeast, centromere fusion occurs independently of cohesins: Spo13 and the Polo kinase homolog Cdc5 recruit a meiosis-specific protein complex, monopolin (Csm1, Lrs4, Mam1, CK1) to the kinetochore. Lrs4 and Csm1 form a V-shaped structure that interacts with the N-terminal domain of Dsn1 in the Mis12 complex to fuse sister kinetochores. While the monopolin complex is not widely conserved, cohesin-independent mechanisms may exist in other organisms. A bridge between the kinetochore proteins MIS12 and NDC80 is required for co-orientation in maize. In contrast, cohesins are required for co-orientation in several organisms. The meiosis-specific cohesin Rec8 is indispensable for sister centromere fusion in fission yeast and Arabidopsis. Cohesin is localized to the core-centromere in fission yeast and mice. In Drosophila melanogaster oocytes, cohesins (SMC1/SMC3/SOLO/SUNN) establish cohesion in meiotic S-phase and show an enrichment that colocalizes with centromere protein CID/CENP-A. Like fission yeast and mouse, Drosophila may require high concentrations of cohesins to fuse sister centromeres together for co-orientation during meiosis (Wang, 2019).
In mice, a novel kinetochore protein, Meikin, recruits Plk1 to protect Rec8 at centromeres. Although poorly conserved, Meikin is proposed to be a functional homolog of Spo13 in budding yeast and Moa1 in fission yeast. They all contain Polo-box domains that recruit Polo kinase to centromeres. Loss of Polo in both fission yeast (Plo1) and mice results in kinetochore separation, suggesting a conserved role for Polo in co-orientation. In fission yeast, Moa1-Plo1 phosphorylates Spc7 (KNL1) to recruit Bub1 and Sgo1 for the protection of centromere cohesion in meiosis I. These results suggest the mechanism for maintaining sister centromere fusion involves kinetochore proteins recruiting proteins that protect cohesion. However, how centromere cohesion is established prior to metaphase I, and how sister centromere fusion is released during meiosis II, still needs to be investigated (Wang, 2019).
Previous work has found that depletion of the kinetochore protein SPC105R (KNL1) in Drosophila oocytes results in separated centromeres at metaphase I, suggesting a defect in sister centromere fusion. Thus, Drosophila SPC105R and fission yeast Spc7 may have conserved functions in co-orientation (Radford, 2015). This study has identified a second Drosophila protein required for sister-centromere fusion, Protein Phosphatase 1 isoform 87B (PP1-87B). However, sister centromere separation in SPC105R and PP1-87B depleted Drosophila oocytes occurs by different mechanisms, the former is Separase dependent and the latter is Separase independent. Based on these results, a model is proposed for the establishment, protection and release of co-orientation. Sister centromere fusion necessary for co-orientation is established through cohesins that are protected by SPC105R. Subsequently, PP1-87B maintains co-orientation in a cohesin-independent manner by antagonizing stable kinetochore-microtubule (KT-MT) interactions. The implication is that the release of co-orientation during meiosis II is cohesin-independent and MT dependent. A surprising interaction was found between PP1-87B and C(3)G, the transverse element of the synaptonemal complex (SC), in regulating sister centromere separation. Overall, these results suggest a new mechanism where KT-MT interactions and centromeric SC regulate sister kinetochore co-orientation during female meiosis (Wang, 2019).
The synaptonemal complex (SC) is a conserved meiotic structure that regulates the repair of double-strand breaks (DSBs) into crossovers or gene conversions. The removal of any central-region SC component, such as the Drosophila melanogaster transverse filament protein C(3)G, causes a complete loss of SC structure and crossovers. To better understand the role of the SC in meiosis, CRISPR/Cas9 was used to construct 3 in-frame deletions within the predicted coiled-coil region of the C(3)G protein. Since these 3 deletion mutations disrupt SC maintenance at different times during pachytene and exhibit distinct defects in key meiotic processes, they allowed definition the stages of pachytene when the SC is necessary for homolog pairing and recombination during pachytene. These studies demonstrate that the X chromosome and the autosomes display substantially different defects in pairing and recombination when SC structure is disrupted, suggesting that the X chromosome is potentially regulated differently from the autosomes (Billmyre, 2019).
Several facets of meiosis ensure the faithful inheritance of chromosomes from parents to offspring. During the creation of eggs and sperm the genome must be reduced to a haploid state containing a single set of chromosomes. The failure to properly segregate chromosomes results in gametes with an incorrect number of chromosomes. Indeed, errors in meiotic chromosome segregation are the leading cause of miscarriage and aneuploidy in humans, which can result in chromosomal disorders such as Down syndrome and Turner syndrome (Billmyre, 2019).
Proper segregation of chromosomes during meiosis relies on the formation of programmed double-strand breaks (DSBs), which are initiated by the evolutionarily conserved type II DNA topoisomerase-like protein Spo11 (Mei-W68 in Drosophila). These DSBs are then repaired as crossover or gene conversion events. Crossovers mature into chiasmata, which physically hold homologous chromosomes together from nuclear envelope breakdown until homolog separation at anaphase I, thus ensuring proper segregation of chromosomes. The placement of crossover events is highly nonrandom and is strictly regulated by multiple processes. First, crossover interference prevents 2 crossovers from occurring in close proximity to each other. Second, crossovers are excluded from the heterochromatin. Third, as a result of the centromere effect, crossing over is also reduced in those euchromatic regions that lie in proximity to the centromeres. Finally, even within the medial and distal euchromatin, crossing over is substantially higher toward the middle of the chromosome arms. These constraints do not affect the frequency or distribution of gene conversion events, which appear to be randomly distributed throughout the euchromatin. Thus, the control of crossover distribution may act at the level of DSB fate choice, rather than in determining the position of DSBs (Billmyre, 2019).
Previous studies have demonstrated that the synaptonemal complex (SC), a large protein structure that forms between homologous chromosomes, plays a role in controlling crossover distribution. The SC is a highly conserved tripartite structure, with 2 lateral elements and a central region. The central region is composed of transverse filament and central element proteins, while the lateral element proteins connect the central region to the chromosome axes. The known proteins that make up the Drosophila central region include the main transverse filament protein C(3)G, the transverse filament-like protein Corolla, and the central element protein Corona (CONA) (Billmyre, 2019).
Work in Caenorhabditis elegans has shown that the SC functions to monitor crossover placement by preventing additional crossover designation in a region adjacent to an existing crossover precursor. Furthermore, there is evidence in Saccharomyces cerevisiae that Zip1, a transverse filament protein, has 2 separable functions-one in building the SC and the other in recombination. Lastly, in rice, there is evidence that a partial loss of the SC results in increased crossing over and crossover proximity similar to what was reported in C. elegans. Based on what is known in other model systems, it is likely that the Drosophila SC is also playing a role in regulating the fate of DSBs and monitoring crossover placement (Billmyre, 2019).
In Drosophila females, ~24 DSBs are formed in early pachytene. Unlike in many other organisms where DSBs occur prior to SC formation, in Drosophila DSBs are formed in the context of fully formed SC. In the absence of the central region of the SC, DSB formation is substantially reduced, but not eliminated. Nonetheless, even in the presence of a substantial number of residual DSBs (37% of wild type), the loss of these SC proteins results in a complete loss of crossover formation. The abolishment of the central region of the SC also results in a high frequency of unpaired homologs during pachytene. In addition to disrupting meiotic pairing, the loss of any of the known central-region components in the (premeiotic) mitotic region of the ovaries also impairs mitotic pairing of the second and third chromosomes (Billmyre, 2019).
Since the vast majority of SC mutants in Drosophila are null mutants and therefore fail to form any SC structure, it is difficult to investigate the interactions of the wild-type versions of these proteins at the protein level or discover how the SC is involved in DSB repair and fate choice. In Drosophila, the study of transgenes carrying in-frame deletions of either the N- or C-terminal globular domains of C(3)G has shown that both of these regions are required for proper SC assembly and crossover formation. However, these defects were too severe to allow investigation of the function of the SC in crossover placement and formation. One domain which has not been tested is the large predicted coiled-coil domain in C(3)G. Coiled-coil domains are a key conserved feature of transverse filament proteins across many organisms and are known to be important for protein-protein interactions (Billmyre, 2019).
This study has characterize 3 in-frame deletion mutations in the coiled-coil domain of the Drosophila melanogaster c(3)G, all of which cause a partial loss of SC function at different stages in early meiosis. Advantage was taken of the different stages of SC loss to examine when the SC is necessary for multiple meiotic events such as pairing and recombination. Unlike any previously characterized Drosophila meiotic mutants, the effects of these mutants on X chromosome recombination is different from their effects on autosomal recombination. It is inferred from this observation that chromosomes can respond differently to a failure in SC maintenance. It was also shown that the SC in early pachytene is important for the maintenance of euchromatic pairing, especially in the distal euchromatin (in relation to the centromere) regions of the chromosome arms. The maintenance of X chromosome pairing is more sensitive to SC defects than is pairing maintenance on the autosomes, suggesting there may be additional chromosome-specific processes that mediate pairing. These mutants allowed us to examine the temporal requirement for the synaptonemal complex in crossover placement and maintenance of pairing (Billmyre, 2019).
Both the regulation of SC assembly and disassembly, and its maintenance after assembly, is poorly understood. Work in other organisms has shown that posttranslational modifications are important in SC structure and function. It is known that SUMOylation and N-terminal acetylation promote assembly of the SC while phosphorylation or dephosphorylation promote disassembly of the SC with modifications occurring on multiple SC proteins. Thus far, no posttranslationally modified sites have been identified on C(3)G. However, it is likely that these sites do exist, and it is speculated that sites promoting SC assembly, maintenance, and disassembly may be disrupted in these mutants (Billmyre, 2019).
Another possibility is that the deletions described in this study could destabilize protein-protein interaction sites between C(3)G and other central-region proteins, resulting in an unstable SC that is difficult to maintain. It is noted that the mutant with the smallest deletion, c(3)GccΔ3, exhibited the strongest SC defect. While this deletion was predicted to only disrupt a single coil, the best explanation for the more severe phenotype is that it actually disrupts the coiled coil. This may have caused a large disruption in the rest of the coiled-coil structure. In the future, it will be important to further dissect these domains to better understand the regulation of SC assembly and disassembly (Billmyre, 2019).
A surprising result from these studies was the ability of these deletions to allow the progressive loss of homologous euchromatic pairing through pachytene. The mechanism behind establishing and maintaining homolog pairing is a long-standing, unanswered question in the meiosis field. Previous work in Drosophila has shown that in the complete absence of the central-region proteins C(3)G and CONA, euchromatic pairing is significantly reduced in early to mid and mid pachytene (Billmyre, 2019).
Partial loss-of-function mutations have allowed testing of the importance of C(3)G in maintaining pairing throughout pachytene when the SC is present in early pachytene (unlike previous studies of null mutants in which the SC is always absent). From these mutants, a timeline is now available of when the SC is necessary to maintain pairing and recombination on the X chromosome and the autosomes. By comparing these mutants, it is hypothesized that the X chromosome needs a full-length SC earlier in pachytene for proper maintenance of pairing and recombination while the autosomes are likely capable of placing crossovers as late as mid pachytene, resulting in a proximal euchromatin shift in crossovers where pairing is maintained (Billmyre, 2019).
In both c(3)GccΔ1 and c(3)GccΔ3 mutants, distal pairing of the X chromosome and the autosomes was most strongly reduced. One likely explanation for this stronger effect on distal regions of the chromosome arms is that normally the disassembly of the SC is initiated on the euchromatic chromosome arms with the centromeric region being removed last. Since the loss of the SC in c(3)GccΔ1 and c(3)GccΔ3 mutants occurs in a manner similar to wild-type SC disassembly, the distal regions of the chromosome may be affected earlier and more strongly than the proximal euchromatin regions. The proximal euchromatin region contains a large amount of heterochromatin that could be mediating pairing interactions and stabilizing pairing in the absence of the SC. Furthermore, examination of centromere pairing suggests that the centromeres are still paired and could be facilitating the proximal euchromatic pairing. This idea is supported by the higher levels of proximal euchromatic pairing compared with distal pairing in c(3)Gnull (Billmyre, 2019).
Finally, it is speculated that the ability of the c(3)GccΔ1 mutants to exhibit a distal euchromatic pairing defect that is more severe than the defect seen in c(3)G null mutants results from the residual proximal crossovers that do form in c(3)GccΔ1 mutants. Previous work has shown that crossovers can preserve synapsis but only in their vicinity. Perhaps the stresses that provoke separation become more concentrated on the distal regions that lack crossovers. For example, it is possible that the untethered distal regions could experience a higher mechanical stress due to nuclear movements than the pericentric regions containing a crossover. The lack of a strong pairing defect in c(3)GccΔ2 mutants is probably due to the persistence of a full-length SC until mid pachytene. Together, these data support a role for the SC in maintaining euchromatic pairing during early to mid prophase (Billmyre, 2019).
The autosomal increase in proximal euchromatin crossovers displayed in these mutants mimics the interchromosomal effect. The interchromosomal effect has been reported in flies that are heterozygous for chromosome aberrations that suppress exchange in trans to a wild-type chromosome. Thus, the absence of crossover formation on one chromosome promotes increased recombination on the other chromosomes, with more crossovers placed in the proximal euchromatin regions. The mechanism that controls the interchromosomal effect in balancer heterozygotes is poorly understood. Additionally, the interchromosomal effect has been reported in C. elegans mutants with defective synapsis, further supporting this possibility (56). It is possible that the interchromosomal effect is partially responsible for the increase in proximal euchromatin crossovers in c(3)GccΔ1 and c(3)GccΔ3 mutants due to the loss of X chromosome recombination (Billmyre, 2019).
However, the interchromosomal effect cannot explain the increase in proximal euchromatin recombination in c(3)GccΔ2 mutants since X recombination appears normal. In theory, this phenotype could be explained by crossover homeostasis, which functions to control the number of crossovers so the appropriate number is placed (reviewed in ref. 57). In many organisms, when there is a deficit of crossovers by the end of early to mid pachytene, the cell will continue to place crossovers in alternative locations to maintain an appropriate number. Such a process could result in crossovers being placed later than normal, which could be an issue when the SC is breaking down prematurely and homolog pairing is lost. However, Mehrotra and McKim (22) provide evidence that crossover homeostasis is unlikely to occur in Drosophila females. It is unknown how much of a role the SC plays in the repair of DSBs into crossover versus non-crossover events. It is possible the SC must be present to interact with factors necessary for regulating the placement of crossovers. For example, Vilya, a pro-crossover factor, localizes to the SC and DSBs prior to being recruited to recombination nodules. If DSB repair on the autosomes does not occur until early to mid pachytene and the SC is necessary for the determination of a crossover fate, it follows that loss of the SC in the euchromatin would result in a shift of crossover formation toward proximal euchromatin regions where the SC may still be present. This mechanism could also be increasing proximal euchromatin recombination in c(3)GccΔ1 and c(3)GccΔ3 flies. Alternatively, SC-independent heterochromatic pairing may be holding the proximal euchromatin region in close proximity, allowing for crossing over in that region. In addition to interacting with pro-crossover factors, the SC may be interacting with a currently unknown protein which regulates crossover placement differently on the X chromosome versus the autosomes (Billmyre, 2019).
This set of mutants represents a unique tool to investigate not only the temporal requirements of the SC but also the differences in crossover placement between the X chromosome and the autosomes. Since c(3)GccΔ2 mutants do not display defects in X chromosome recombination, it is concluded that a full-length SC throughout early to mid pachytene is sufficient for X chromosome crossover placement but not for normal distribution of autosomal crossovers. Examining autosomal recombination in all 3 mutants suggests that a full-length SC is necessary in mid pachytene for proper crossover distribution on the autosomes. There are multiple explanations for the recombination differences between the X chromosome and the autosomes (Billmyre, 2019).
The first of these hypotheses is that there might exist a timing difference in either synapsis or crossover placement between the X chromosome and the autosomes. Work in C. elegans has provided evidence for timing differences between the sex chromosomes and the autosomes. For example, the X chromosome initiates premeiotic DNA replication later than the autosomes. This could be significant, as replication timing has been shown to impact crossover designation in barley. Additionally, in C. elegans, the X chromosome and the autosomes pair at the same time, but synapsis of the X chromosome is delayed and the X chromosome has lower levels of DSB formation compared with the autosomes. Thus, the timing of when each chromosome is fully synapsed could be critical to ensure normal crossover placement, and the premature disruption of synapsis may affect the activity of pro-crossover factors. For example, in C. elegans, the XND-1 protein is required for genome-wide crossover placement and is important for normal rates of DSBs on the X chromosome. Currently, it is unknown in Drosophila if there are differences in the timing of DSB repair or synapsis of the X chromosome as compared with the autosomes, and the data suggest this as a possibility (Billmyre, 2019).
A second, but not mutually exclusive, explanation for the differences between the chromosomes may be a structural one. The X chromosome is acrocentric (the centromere is near the end of the chromosome), while the autosomes are both metacentric (the centromere is near the center of the chromosome) and, perhaps, these structural differences mean that the X chromosome is more sensitive to loss of the SC. The data suggest that loss of SC maintenance disrupts the maintenance of euchromatic homolog pairing more severely on the X chromosome than on the autosomes. It is unknown if metacentric chromosomes are different in terms of synapsis and recombination as compared with acrocentric chromosomes, and further investigation is needed to determine if structural differences affect these processes (Billmyre, 2019).
It is clear from decades of research that the regulation of recombination requires many factors and precise timing. This study shows that the SC plays a vital role in maintaining homolog pairing and proper crossover distribution in Drosophila female meiosis. Many differences between sex chromosomes and autosomes have been documented in a multitude of organisms, and our data are consistent with these differences extending into the processes that control chromosome pairing and recombination. With this set of mutants, a system has been established to examine X chromosome and autosome biology in Drosophila meiosis that will allow future work to unravel the mechanism behind meiotic chromosomal differences (Billmyre, 2019).
During meiosis, each chromosome must selectively pair and synapse with its own unique homolog to enable crossover formation and subsequent segregation. How homolog pairing is maintained in early meiosis to ensure synapsis occurs exclusively between homologs is unknown. This study aimed to further understand this process by examining the meiotic defects of a unique Drosophila mutant, Mcm5A7. Mcm5A7 mutants are proficient in homolog pairing at meiotic onset yet fail to maintain pairing as meiotic synapsis ensues, causing seemingly normal synapsis between non-homologous loci. This pairing defect corresponds with a reduction of SMC1-dependent centromere clustering at meiotic onset. Overexpressing SMC1 in this mutant significantly restores centromere clustering, homolog pairing, and crossover formation. These data indicate that the initial meiotic pairing of homologs is not sufficient to yield synapsis exclusively between homologs and provide a model in which meiotic homolog pairing must be stabilized by centromeric SMC1 to ensure proper synapsis (Hatkevich, 2019).
Accurate segregation of homologous chromosomes during the first meiotic division is essential to reestablish the diploid genome upon sexual fertilization. To ensure faithful meiosis I chromosomal segregation, homologs must become physically connected in part through crossover formation. To enable homolog crossover events, a series of chromosomal and cellular events occur in early meiotic prophase I (Hatkevich, 2019).
During or just prior to the onset of meiosis, homologous chromosomes pair along their entire lengths. Between paired homologs, synapsis, the formation of the synaptonemal complex (SC), ensues. The SC is a tripartite scaffold built between homologs extending the length of the chromosomes and consists of a central region that is nestled between two lateral elements, which are successors of cohesin-based chromosome axes formed between sister chromatids. Coincident with synapsis, DSBs are formed and repaired using a homologous template via homologous recombination (HR), resulting in crossover formation between homologs (Hatkevich, 2019).
Perhaps the most enigmatic event within early meiosis is the mechanism by which a meiotic chromosome selectively pairs and synapses with its unique homologous partner. Initial homolog pairing is believed to be facilitated through early meiotic chromosome movement and telomere or the centromere clustering. However, how homologous pairing is maintained during synapsis to ensure the SC is formed exclusively between homologs is unknown (Hatkevich, 2019).
The model organism Drosophila melanogaster has been used to uncover meiotic mechanisms for over a century. In Drosophila, prior to meiosis, chromosomes enter the germline unpaired; throughout the pre-meiotic region, homologous chromosomes gradually pair. In the nuclei at the last mitotic division prior to meiotic onset (in the 8-cell cyst), centromere-directed chromosomal movements occur, presumably ensuring complete homologous pairing. Also during pre-meiotic mitotic cycles, several proteins, including the cohesin SMC1, are enriched at the centromere. The onset of meiotic prophase I occurs in the 16-cell cyst. At zygotene, the first cytologically resolved stage of prophase, centromeres are clustered into 1 or 2 groups, and the SC nucleates in patches along chromosome arms. As zygotene proceeds into early pachytene, the SC extends between paired chromosomes, yielding full-length SC exclusively between homologs. How these early meiotic events, particularly SMC1 enrichment at the centromere and centromere clustering, contribute to meiotic homologous pairing and synapsis in Drosophila is largely unknown (Hatkevich, 2019).
This study used the Drosophila early meiotic program and a unique genetic mutant to investigate how homolog pairing is maintained during meiotic synapsis. Meiotic homologs in a previously described Drosophila mutant, Mcm5A7, initially pair, but are unable to maintain pairing during synapsis, suggesting that initial meiotic pairing must be subsequently stabilized by an unknown mechanism to ensure proper synapsis. Using Mcm5A7 as a genetic tool to interrogate pairing stabilization mechanism(s), it was found that SMC1 localization at the centromere is compromised, correlating with a severe defect in meiotic centromere clustering and a decrease of crossover formation. However, arm cohesion and SC structure appear unperturbed in these mutants. By overexpressing SMC1, this study shows that the defects in centromere clustering, meiotic homolog pairing, homosynapsis, and crossing over in Mcm5A7 mutants are caused by a lack of centromeric SMC1 localization at meiotic onset. From these results, a model for proper synapsis is suggested in which initial meiotic pairing must be stabilized by centromere clustering, a meiotic event produced by SMC1-enrichment at the centromere and dynamic chromosome movements (Hatkevich, 2019).
At the beginning of this study, it was hypothesized that the crossover defect in Mcm5A7 mutants was due to a homolog pairing deficiency. FISH results support this hypothesis and revealed that homolog pairing can be lost during synapsis, resulting in seemingly normal SC between heterologous sequences. Centromere-directed chromosome movements occur in Mcm5A7 mutants, presumably to promote initial chromosome arm pairing; however, centromere pairing and clustering are perturbed. SMC1 enrichment at the centromere is decreased in Mcm5A7 mutants, while arm cohesion appears normal. Overexpression of SMC1 rescues centromeric-SMC1 localization and downstream meiotic defects, including centromere clustering, pairing, crossover formation, and segregation. From these data, it is proposed that centromeric SMC1 stabilizes initial homolog pairing through centromere clustering, securing meiotic pairing, ensuring homosynapsis and promoting crossover formation (Hatkevich, 2019).
Prior to the onset of meiosis, cohesins are loaded onto centromeres, and homologous chromosomes pair, with arm pairing preceding centromere pairing. Initial homolog pairing is achieved in part by centromere-directed movements in the division prior to meiotic onset.
A model in which initial homologous chromosomal pairing is stabilized throughout early meiosis by SMC1-dependent centromere clustering (Hatkevich, 2019).
According to this model, the enrichment of SMC1 at the centromere combined with chromosome movements in pre-meiotic stages yield centromere clustering at meiotic onset. While chromosome arms and centromeres enter meiosis paired, heterologous centromere clustering and/or centromere pairing are required to stabilize pairing during SC assembly. As initial euchromatic SC patches extend along the arms of paired homologs, DSBs are formed and subsequently repaired via HR to yield crossovers, which promote accurate disjunction at the end of meiosis (Hatkevich, 2019).
In Mcm5A7 mutants, coordinated pre-meiotic centromere-directed movements occur, but a sufficient amount of SMC1 is not localized at the centromere to yield centromere clustering. Thus, at meiotic onset, arms are paired, but centromeres are not clustered. As euchromatic SC nucleation occurs, the stabilization provided by centromere clustering is absent, and homologous loci become unpaired. As synapsis extends, the SC is able to form between nearby chromosomes, regardless of homology, yielding heterosynapsis (intrachromosomal and/or interchromomosomal). DSBs made within regions of heterosynapsis are not repaired via HR due to the absence of an available homologous template. Therefore, crossovers are reduced, and nondisjunction occurs at high frequency in Mcm5A7 mutants (Hatkevich, 2019).
The SMC1-dependent centromere clustering pairing model highlights the finding that initial meiotic pairing is not sufficient to yield complete homosynapsis. Rather, centromeric SMC1-dependent stabilization must occur after pairing and during synapsis. The inherent requirement of pairing stabilization for proper synapsis suggests that there is a force that opposes homolog alignment during synapsis. Perhaps the SC assembly process itself creates an opposing force that can push paired homologs away from one another in the absence of stabilization; a similar hypothesis was previously proposed in C. elegans. An alternative hypothesis is that recombination, which coincides temporally with synapsis assembly, creates a destabilizing force. However, when meiotic DSBs are eliminated in Mcm5A7 mutants (as shown through mei-P22 Mcm5A7 double mutants), homologs are unpaired at a frequency similar to Mcm5A7 mutants, indicating that the opposing force is independent of recombination. Regardless of the origin of the force, it is proposed that SMC1-dependent centromere clusters act as anchors at the nuclear envelope to maintain the proximity of homologous axes (Hatkevich, 2019).
Although meiotic pairing programs vary among organisms, the SMC1-dependent centromere clustering pairing model may be broadly applicable. In Drosophila and C. elegans, meiotic pairing is independent of meiotic recombination. In contrast, meiotic pairing in organisms such as yeast, plants, and mice require DSB formation (although recombination-independent alignment is required for pairing in these organisms). In DSB-dependent pairing programs, homologs are considered paired at ~400 nm, where DSB-mediated interhomolog interactions can be visualized as bridges. However, contemporaneous with DSB formation, centromeres are coupled or clustered. It is speculated that these centromere interactions stabilize the DSB-dependent arm pairing to ensure synapsis exclusively between homologs in many sexually-reproducing organisms (Hatkevich, 2019).
This study reveals the interesting phenomenon of stable heterosynapsis in Drosophila. Extensive heterosynapsis has been previously reported in C. elegans and yeast with variable SC integrity. Though SC aberrations in Mcm5A7 mutants cannot be ruled out, the data reveal no structural defects, supporting the notion that 'normal' synapsis is largely homology-independent in Drosophila, as has been observed in C. elegans (Hatkevich, 2019).
In Drosophila, synapsis along the arms initiates as patches during zygotene. In Mcm5A7 mutants, synapsis initiation between paired homologs appears normal in zygotene but SC elongation fails to be limited to homologous regions. Thus, initiation of synapsis may require homology, but elongation may not. Because this study examined only specific loci and not whole chromosomes, future studies determining the degree of heterosynapsis in Mcm5A7 mutants may provide more insight into how synapsis and homology interact in flies (Hatkevich, 2019).
While Mcm5A7 has proven to be a valuable genetic tool, how this particular mutation affects SMC1 localization at the meiotic centromere is unknown. Mcm5A7 mutants do not affect centromere clustering and pairing in a manner similar to that of mutants that disrupt centromere integrity, such as cal1 Cenp-C double heterozygotes. However, the results do not exclude a role for MCM5 in centromere function or integrity (Hatkevich, 2019).
The canonical role of MCM5 is to function within the replicative helicase, MCM2-7, unwinding double-stranded DNA ahead of the replication fork during S-phase. Because of its important replication role, Mcm5 is an essential gene in every proliferating cell. Numerous studies have shown that replication is required for cohesion localization and establishment, but examining a direct role for any MCM protein in cohesin deposition is difficult since MCMs are essential for replication, which in turn is required for establishing cohesion (Hatkevich, 2019).
Because MCM5 functions within the MCM2-7 replicative helicase, it is tempting to speculate that the Mcm5A7 mutation may directly perturb SMC1 localization, either through defects in replication or cohesin deposition. No replication defect in Mcm5A7 mutants has been detected, in either a mitotic or meiotic context. In the future, when individual pre-meiotic nuclei can be isolated from cysts, higher-resolution replication assays may determine whether replication is subtly disrupted in Mcm5A7 mutants. At this point, however, it seems more likely that the Mcm5A7 cleanly separates the replication role of MCM5 from a role in meiotic cohesin deposition (Hatkevich, 2019).
The TORC regulator GATOR1/SEACIT controls meiotic entry and early meiotic events in yeast. However, how metabolic pathways influence meiotic progression in metazoans remains poorly understood. This study examined the role of the TORC regulators GATOR1 and GATOR in the response to meiotic double-stranded breaks (DSB) during Drosophila oogenesis. In mutants of the GATOR component mio, meiotic DSBs trigger the constitutive downregulation of TORC activity and a permanent arrest in oocyte growth. Conversely, in GATOR mutants, high TORC activity results in the delayed repair of meiotic DSBs and the hyperactivation of p53. Unexpectedly, it was found that GATOR inhibits retrotransposon expression in the presence of meiotic DSBs in a pathway that functions in parallel to p53. Thus, these studies have revealed a link between oocyte metabolism, the repair of meiotic DSBs and retrotransposon expression (Wei, 2019).
Metabolism impacts meiotic progression during oogenesis. Target of Rapamycin Complex 1 (TORC1) is a multi-protein complex that functions as a master regulator of metabolism. In the presence of adequate nutrients and positive upstream growth signals, TORC1, which contains the serine/threonine kinase Target of Rapamycin, becomes active and functions to stimulate growth and inhibit catabolic metabolism through the phosphorylation of down-stream effector proteins. The Seh1 Associated Complex Inhibits TORC1 (SEACIT), originally identified in yeast, inhibits TORC1 activity in response to amino acid limitation. SEACIT, known as the GAP Activity Towards Rags complex 1 (GATOR1) in metazoans, is comprised of three highly conserved proteins Npr2/Nprl2, Npr3/Nprl3 and Iml1/Depdc5. In Drosophila and mammals, depleting any of the three GATOR1 components results in increased TORC1 activity and growth, as well as a reduced response to amino acid starvation. Thus, the role of the SEACIT/GATOR1 complex in the regulation of TORC1 activity is highly conserved in eukaryotes (Wei, 2019).
The multi-protein GATOR2 complex, known as Seh1 Associated Complex Activates TORC1 (SEACAT) in yeast, inhibits the activity of GATOR1 and thus functions to activate TORC1 (see Mio prevents the constitutive downregulation of TORC1 activity in response to meiotic DSBs). In metazoans, the GATOR2 complex functions in multiple amino acid sensing pathways. In tissue culture cells, depleting GATOR2 components results in the constitutive activation of GATOR1 and the permanent downregulation of TORC1 activity. However, genetic studies of the role of individual GATOR2 components in Drosophila, indicate that the requirement for the GATOR2 complex is more nuanced when examined in the context of a multicellular animal. For example, mutations in the GATOR2 component mio, result in a block to oocyte growth and differentiation, due to the constitutive downregulation of TORC1 activity in the female germline. However, mio is not required to maintain TORC1 activity in most somatic tissues of Drosophila (Wei et al., 2016). Why there is a tissue specific requirement for mio in the female germline of Drosophila is currently unknown (Wei, 2019).
In single celled eukaryotes, nutrient limitation often facilitates meiotic entry. In the yeast Saccharomyces cerevisiae, the down-regulation of TORC1 by SEACIT/GATOR1 in response to amino acid stress promotes both meiotic entry and early meiotic progression. Surprisingly, as is observed in yeast, during Drosophila oogenesis the GATOR1 complex promotes meiotic entry. These data raise the intriguing possibility that in Drosophila the GATOR1 complex and low TORC1 activity may be critical to the regulation of additional events of the early meiotic cycle (Wei, 2019).
This study reports that the GATOR complex is critical to the response to meiotic DSB during Drosophila oogenesis. Restraining TORC1 activity via a pathway that involves both GATOR1 and the Tuberous sclerosis complex (TSC) promotes the timely repair of meiotic DSBs and prevents the hyperactivation of p53 in the female germline. Notably, the delayed repair of meiotic DSBs in GATOR1 mutants is due, at least in part, to the hyperactivation of the TORC1 target S6K. Conversely, the data indicate that the GATOR2 component Mio opposes the activity of GATOR1 in the female germline, thus preventing the constitutive downregulation of TORC1 activity and allowing for the growth and development of the oocyte in later stages of oogenesis. Thus, this study has identified a regulatory loop required to modulate TORC1 activity in response to meiotic DSBs during Drosophila oogenesis. Finally, during the course of these studies, it was observed that the GATOR1 complex prevents the derepression of retrotransposon expression in the presence of meiotic DSBs (Wei, 2019).
Previous work has shown that in Drosophila, mutations in the GATOR2 component mio, result in the constitutive activation of the GATOR1 pathway in the female germline but not in somatic tissues. This study demonstrates that the tissue specific requirement for mio during oogenesis is due, at least in part, to the generation of meiotic DBSs during oogenesis. In Drosophila, only the female germline undergoes meiotic recombination and thus experiences the genotoxic stress associated with developmentally programmed DSBs. This study shows that in mio mutants, blocking the formation of meiotic DSBs prevents the constitutive downregulation of TORC1 activity thus allowing for the growth and development of the oocyte. These data are consistent with the model that meiotic DSBs trigger the activation of a TORC1 inhibitory pathway that must be opposed and/or attenuated by the GATOR2 component Mio (see A working model for the role of the GATOR complex in the response to meiotic DSBs) (Wei, 2019).
While there are several possible models that might explain these data, it is believed the most parsimonious explanation for these results is that the TORC1 inhibitory pathway activated by meiotic DSBs, involves both GATOR1 and TSC. This model is consistent with the ability of both GATOR1 and TSC depletions to rescue the mio mutant phenotype. Additionally, recent reports indicate that GATOR1 and TSC act in a common pathway to downregulate TORC1 activity in response to multiple upstream inhibitory inputs. Previously, it was determined that in Drosophila, amino acid starvation induces a dramatic GATOR1/TSC dependent decrease in TORC1 activity in somatic tissues, that far exceeds any reduction in TORC1 activity observed in GATOR2 null mutants. This observation strongly suggests that, in addition to the removal of the GATOR2 inhibition of GATOR1, there is an activation step that is required to fully potentiate the GATOR1/TSC pathway (Wei, 2019).
Thus, based on these data the following model is proposed. Meiotic DSBs activate, or are required to maintain, a GATOR1/TSC dependent pathway that downregulates TORC1 activity in the female germline. The GATOR2 component Mio is required to oppose or turnoff this pathway to prevent the constitutive downregulation of TORC1 activity in later stages of oogenesis. While it is believed that the data support the role of the GATOR1/TSC pathway, it is conceded that an alternative regulator of TORC1 activity may also be critical to the downregulation of TORC1 activity in response to meiotic DSBs (Wei, 2019).
Hyperactivation of TORC1 has been linked to defects in the DNA damage response in single celled and multicellular organisms. The observation that meiotic DSBs likely promote the GATOR1 dependent downregulation of TORC1 activity during Drosophila oogenesis, suggested that limiting TORC1 activity may be important to the regulation of meiotic DSB repair. In previous work, it was found that GATOR1 mutant ovaries had TORC1 activity levels approximately three times higher than those observed in wild-type ovaries. This study demonstrates that GATOR1 mutant ovaries exhibit multiple phenotypes consistent with the misregulation of meiotic DSB repair including, an increase in the steady state number of Mei-W68/Spo-11 induced DSBs, the retention of meiotic DSBs into later stages of oogenesis and the hyperactivation of p53. Importantly, RNAi depletions of Tsc1 partially phenocopied the GATOR1 ovarian defects. Thus, the misregulation of meiotic DSBs observed in GATOR1 mutant oocytes are due to high TORC1 activity and not to a TORC1 independent function of the GATOR1 complex (Wei, 2019).
Epistasis analysis between the GATOR1 component nprl3 and the Rad51 homolog spnA, strongly suggest that GATOR1 impacts the repair, rather than the generation, of meiotic DSBs. This study determined that double mutants of nprl2 and the Rad51 homolog spnA, which is required for the repair of meiotic DSBs, have approximately the same number of DSBs as spnA single mutants. These data are consistent with GATOR1 and spnA influencing the common process of DNA repair and are inconsistent with GATOR1 mutants producing supernumerary breaks (Wei, 2019).
These observations on the role of the GATOR1 complex during Drosophila oogenesis are particularly intriguing in light of similar meiotic defects observed in a npr3 mutants in Saccharomyces cerevisiae. In the sporulation proficient strain SK1, npr3 mutant cells enter meiosis and express the transcription factor and master regulator of gametogenesis IME1 with wild-type kinetics. Subsequently, npr3 mutants exhibit a mild delay in the generation of meiotic DNA breaks, but a substantial delay in the repair of meiotic DSBs. Thus, yeast and Drosophila SEACIT/GATOR1 mutants share a common meiotic phenotype, the delayed repair of meiotic DSBs. These results raise the intriguing possibility that low TORC1 activity may be a common feature of the early meiotic cycle in many organisms (Wei, 2019).
Notably, the data indicate that the delay in the repair of meiotic DSBs in GATOR1 mutants is due to the hyperactivation of the TORC1 downstream target S6K. S6K is a critical downstream effector of TORC1 that impacts multiple essential cellular processes including, but not limited to cell growth, energy balance and aging. Intriguingly, in mammals, S6K has been implicated in the regulation of the DNA damage response with hyperactivation of the TORC1-S6K pathway resulting in the accumulation of unrepaired DSBs and genome instability. Thus, similar to what is reported in mammals, the data are consistent with the model that the hyperactivity of the TORC1/S6K axis delays the repair of DSBs in Drosophila (Wei, 2019).
Finally, it was determined that GATOR1 mutants have a diminished response to DSBs outside the female germline in somatic tissues of Drosophila. Similar to what is observed in TSC mutant cells in humans that have increased levels of TORC1 activity, this study find that GATOR1 mutant embryos have a reduced ability to survive low levels of γ-irradiation. Moreover, in the somatic follicle cells of the ovary a delay was observed in the repair of DSBs after adult females are exposed to low levels of γ-irradiation. Thus, in Drosophila inappropriately high TORC1 activity delays the repair of DSBs in both the germline and somatic tissues (Wei, 2019).
The initiation of homologous recombination through the programmed generation of DNA double-stranded breaks (DSBs) is a universal feature of meiosis. DSBs represent a dangerous form of DNA damage that can result in dramatic and permanent changes to the germline genome. To minimize this destructive potential, the generation and repair of meiotic DSBs is tightly controlled in space and time . The activation of transposable elements represents an additional threat to genome integrity in germ line cells. Genotoxic stress, resulting from DNA damage, has been implicated in the deregulation of transposons in multiple organisms. Thus, germ line cells may be at an increased risk for transposon derepression due to the genotoxic stress associated with meiotic recombination. Consistent with this hypothesis, germ line cells have evolved extensive surveillance systems to detect and silence transposons beyond the pathways present in most somatic tissues (Wei, 2019).
Previous studies have shown that DNA damage promotes the deregulation of retrotransposon in multiple organisms, including Drosophila. In line with these studies, this study found that in GATOR1 mutants, the DSBs that initiate meiotic recombination trigger the deregulation of retrotransposon expression. Similarly, p53 mutant females derepress retrotransposon expression during oogenesis, but as observed in GATOR1 mutants, primarily in the presence of meiotic DSBs. Double mutants of nprl3, p53 exhibit a dramatic increase in retrotransposon expression relative to either p53 or nprl3 single mutants, implying that p53 and GATOR1 act through independent pathways to repress retrotransposon expression in the female germline. One possibility is that both GATOR1 and p53 independently impact genome stability. Thus, disabling both pathways may have an additive effect on both genome stability and retrotransposon expression. Consistent with the hypothesis that genome instability drives retrotransposon expression, this study found that mutants in spnA/Rad51, which fail to repair meiotic DSBs, also exhibit increased transcription of multiple retrotransposons. Intriguingly, the SpnA homolog Rad51, as well as other genes required for DNA repair, was recently identified in a high throughput screen for genes that suppress (Long Interspersed Element-1) LINE1 expression in mammalian tissue culture cells (Wei, 2019).
However, the current data also suggest that the GATOR1 complex may influence retrotransposon expression independent of the regulation of TORC1 activity. While both GATOR1 and TSC are required for the efficient repair of meiotic DSBs, in contrast to GATOR1 mutant ovaries, little to no increase was observed in retrotransposon expression in the Tsc1 depleted ovaries. We believe this reflects the incomplete depletion of Tsc1 by RNAi resulting in a reduced retention of meiotic DSBs relative to GATOR1 mutants. However, a second possibility is that the GATOR1 complex inhibits retrotransposon expression independent of TORC1 inhibition. As is observed with spnA the depletion of GATOR1 components, but not TSC components result in the activation of LINE1 expression in HeLa cells. Taken together, these data hint that the GATOR1 complex may impact retrotransposon expression in the germline via two independent pathways: First by promoting the repair of meiotic DSBs through the downregulation of TORC1 activity and second via a pathway that functions independent of TORC1 inhibition (Wei, 2019).
Genes encoding components of the GATOR1 complex are often deleted in cancers. As is observed in GATOR1 mutants, cancer cells frequently have increased TORC1 activity, increased genomic instability and increased retrotransposon expression. Thus, in the future it will be important to identify the molecular mechanism by which the GATOR1 complex influences both the response to genotoxic stress and the expression of retrotransposons under both normal and pathological conditions (Wei, 2019).
In all species, new chromosomal inversions are constantly being formed by spontaneous rearrangement and then stochastically eliminated from natural populations. In Drosophila, when new chromosomal inversions overlap with a pre-existing inversion in the population, their rate of elimination becomes a function of the relative size, position, and linkage phase of the gene rearrangements. These altered dynamics result from complex meiotic behavior wherein overlapping inversions generate asymmetric dyads that cause both meiotic drive/drag and segmental aneuploidy. In this context, patterns in rare inversion polymorphisms of a natural population can be modeled from the fundamental genetic processes of forming asymmetric dyads via crossing-over in meiosis I and preferential segregation from asymmetric dyads in meiosis II. A mathematical model was developed of crossover-dependent female meiotic drive and was parameterized with published experimental data from Drosophila melanogaster laboratory constructs. This mechanism is demonstrated to favor smaller, distal inversions and accelerate the elimination of larger, proximal inversions. Simulated sampling experiments indicate that the paracentric inversions directly observed in natural population surveys of Drosophila melanogaster are a biased subset that both maximizes meiotic drive and minimizes the frequency of lethal zygotes caused by this cytogenetic mechanism. Incorporating this form of selection into a population genetic model accurately predicts the shift in relative size, position, and linkage phase for rare inversions found in this species. The model and analysis presented in this study suggest that this weak form of female meiotic drive is an important process influencing the genomic distribution of rare inversion polymorphisms (Koury, 2023).
The global replacement of histones with protamines in sperm chromatin is widespread in animals, including insects, but its actual function remains enigmatic. In the Drosophila paternal effect mutant paternal loss (pal), sperm chromatin retains germline histones H3 and H4 genome wide without impairing sperm viability. However, after fertilization, pal sperm chromosomes are targeted by the egg chromosomal passenger complex and engage into a catastrophic premature division in synchrony with female meiosis II. pal encodes a rapidly evolving transition protein specifically required for the eviction of (H3-H4)(2) tetramers from spermatid DNA after the removal of H2A-H2B dimers. This study thus reveals an unsuspected role of histone eviction from insect sperm chromatin: safeguarding the integrity of the male pronucleus during female meiosis (Dubruille, 2023).
Sperm chromatin is generally characterized by a high level of DNA compaction, which reduces nuclear volume and contributes to the shape and hydrodynamic properties of the sperm head. In many animal species, tight packaging of sperm DNA follows the replacement of nucleosomal histones with sperm nuclear basic proteins (SNBPs, see Protamine A), such as the well-characterized mammalian protamines. This distinct, global chromatin remodeling process known as the histone-to-protamine transition occurs during spermiogenesis, the differentiation of post-meiotic spermatids. Although the histone-to-protamine transition is generally assumed to be essential for the formation of functional sperm, not all animals use SNBPs for sperm chromatin assembly. For example, whereas mammalian sperm DNA is mostly packaged with protamines, many other vertebrates maintain a full nucleosome-based sperm chromatin. Even within groups of related species (e.g., teleost fishes), sperm chromatin can vary between protamine based and histone based. These repeated transitions between nucleosomal and nucleoprotamine sperm chromatin have left the function of histone replacement by SNBPs unclear (Dubruille, 2023).
Despite their enormous diversity and ancient evolutionary history, all major orders of insects consistently encode "histone-free" packaging of sperm DNA. As the best characterized example, the model species Drosophila melanogaster has ultracompact sperm chromatin entirely organized with protamine-like SNBPs, except for centromeres that retain the centromeric histone H3 (CenH3). This indicates that a stringent selective requirement for SNBP-based sperm chromatin must exist in insects. This work discovered that the replacement of histones with SNBPs is functionally linked to a postfertilization process through the characterization of a Drosophila paternal effect mutant named paternal loss (pal) Paternal effect mutants represent invaluable genetic tools to investigate the influence of spermiogenesis and sperm chromatin composition on the fate of paternal chromosomes at fertilization. In Drosophila, the molecular characterization of Characterization of pal first reveals the extraordinary plasticity of insect sperm chromatin composition. Although Drosophila spermiogenesis is sensitive to the loss of particular SNBPs, the global retention of H3 and H4 histones observed in pal mutants has no detectable impact on sperm differentiation or function despite substantial changes in nuclear morphology. It suggests that major diversification of sperm chromatin composition in animals could result from minimal changes in genes involved in the histone-to-protamine transition (Dubruille, 2023).
In pal sperm, it was hypothesized that (H3-H4)2 tetramers are distributed throughout the genome, reflecting the original position of nucleosomes before the removal of H2A and H2B at the histone-to-protamine transition. These tetrasomes must then coexist with SNBPs that are subsequently deposited, although less abundantly than in normal sperm chromatin. Incidentally, this study also establishes that histone elimination during Drosophila spermiogenesis is a sequential process, with the removal of H2A-H2B and H3-H4 being temporally and functionally distinct. Notably, during mouse spermiogenesis, the replacement of H2A and H2B with testis-specific histone variants prepares the final replacement of nucleosomes with protamines, whereas in Xenopus, only H2A-H2B dimers are partially replaced with SNBPs, leading to the retention of H3 and H4 in sperm. The sequential elimination of histones in spermatids could thus be widespread in animals (Dubruille, 2023).
The recurrent packaging of sperm DNA with nonhistone proteins is a longstanding mystery in chromatin biology and evolution. Main hypotheses on this specialization include sperm nuclear compaction, transcriptional shutdown in spermatids, and sperm DNA protection against damages. The current work suggests that the timing of egg fertilization with respect to female meiosis progression is a previously unexplored, key determinant of sperm chromatin evolution in animals. In insects, early fertilization (metaphase of meiosis I) inevitably exposes the decondensing sperm nucleus to egg cytoplasmic cues that initiate the second meiotic division of maternal chromosomes. It is proposed that the global and invariable replacement of histones with SNBPs in insects establishes an epigenetic identity of paternal chromosomes that prevents their deleterious interaction with the egg CPC. In vertebrate eggs, the relatively late oocyte fertilization timing (metaphase of meiosis II) may offer a more permissive environment to the fertilizing sperm nucleus during its transformation into a pronucleus. In support of this possibility, premature chromosome condensation of sperm chromosomes, a defect highly reminiscent of the pal phenotype, is frequently observed when in vitro fertilization or intracytoplasmic sperm injection is performed with immature mammalian oocytes. The temporal uncoupling of female meiosis II initiation and male pronuclear formation in vertebrates could have thus favored the extreme diversification of sperm chromatin types, including histone-based sperm chromatin. Still, other animals with an early type of fertilization, such as the model nematode Caenorhabditis elegans, maintain histone-based sperm chromatin, which suggests the evolution of additional, yet unknown mechanisms to protect the paternal genome during the formation of the diploid zygote (Dubruille, 2023).
References Acquaviva, C. and Pines, J. (2006). The anaphase-promoting complex/cyclosome: APC/C. J. Cell Sci. 119(Pt 12): 2401-4. PubMed ID: PubMed ID: 16763193
Allers, T. and Lichten, M. (2001) Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 106: 47-57. PubMed ID: PubMed ID: 11461701
Bhagat, R., Manheim, E. A., Sherizen, D. E. and McKim, K. S. (2004). Studies on crossover-specific mutants and the distribution of crossing over in Drosophila females. Cytogenet Genome Res 107: 160-171. PubMed ID: PubMed ID: 15467361
Billmyre, K. K., Cahoon, C. K., Heenan, G. M., Wesley, E. R., Yu, Z., Unruh, J. R., Takeo, S. and Hawley, R. S. (2019). X chromosome and autosomal recombination are differentially sensitive to disruptions in SC maintenance. Proc Natl Acad Sci U S A. PubMed ID: 31570610
Blanton, H. L., Radford, S. J., McMahan, S., Kearney, H. M., Ibrahim, J. G. and Sekelsky, J. (2005). REC, Drosophila MCM8, drives formation of meiotic crossovers. PLoS Genet. 1(3): e40. PubMed ID: PubMed ID: 16189551
Christophorou, N., Rubin, T., Bonnet, I., Piolot, T., Arnaud, M. and Huynh, J. R. (2015). Microtubule-driven nuclear rotations promote meiotic chromosome dynamics. Nat Cell Biol 17: 1388-1400. PubMed ID: 26458247
Chu, T., Henrion, G., Haegeli, V. and Strickland, S. (2001). Cortex, a Drosophila gene required to complete oocyte meiosis, is a member of the Cdc20/fizzy protein family. Genesis 29: 141-152. PubMed ID: PubMed ID: 11252055
Dubruille, R., Herbette, M., Revel, M., Horard, B., Chang, C. H., Loppin, B. (2023). Histone removal in sperm protects paternal chromosomes from premature division at fertilization. Science, 382(6671):725-731 PubMed ID: 37943933
Emelyanov, A. V., Rabbani, J., Mehta, M., Vershilova, E., Keogh, M. C., Fyodorov, D. V. (2014) Drosophila TAP/p32 is a core histone chaperone that cooperates with NAP-1, NLP, and nucleophosmin in sperm chromatin remodeling during fertilization. Genes Dev 28: 2027-2040. PubMed ID: 25228646
Endow, S. A. and Komma, D. J. (1998). Assembly and dynamics of an anastral:astral spindle: the meiosis II spindle of Drosophila oocytes. J. Cell Sci. 111, 2487-2495. PubMed ID: 9701548
Fitch, K. R. and Wakimoto, B. T. (1998). The paternal effect gene ms(3)sneaky is required for sperm activation and the initiation of embryogenesis in Drosophila melanogaster. Dev. Biol. 197: 270-282. PubMed ID: PubMed ID: 9630751
Grell, R. F. (1984). Time of recombination in the Drosophila melanogaster oocyte. III. Selection and characterization of temperature-sensitive and -insensitive recombination-deficient alleles in Drosophila. Genetics 108: 435-443
Gyuricza, M. R., Manheimer, K. B., Apte, V., Krishnan, B., Joyce, E. F., McKee, B. D. and McKim, K. S. (2016). Dynamic and stable cohesins regulate synaptonemal complex assembly and chromosome segregation. Curr Biol 26(13):1688-1698. PubMed ID: 27291057
Hatkevich, T., Boudreau, V., Rubin, T., Maddox, P. S., Huynh, J. R. and Sekelsky, J. (2019). Centromeric SMC1 promotes centromer clustering and stabilizes meiotic homolog pairing. PLoS Genet 15(10): e1008412. PubMed ID: 31609962
Horner, V. L., et al. (2006). The Drosophila calcipressin Sarah is required for several aspects of egg activation. Curr. Biol. 16(14): 1441-6. PubMed ID: PubMed ID: 16860744
Huettner, A. F. (1924). Maturation and fertilization in Drosophila melanogaster. J. Morphol. Physiol. 39: 249-265
Huynh, J.-R. and St Johnston, D. (2000). The role of BicD, Egl, Orb and the microtubules in the restriction of meiosis to
the Drosophila oocyte. Development 127: 2785-2794. PubMed ID: PubMed ID: 10851125
Inoue, Y. H., Savoian, M. S., Suzuki, T., Mathe, E., Yamamoto, M. T. and Glover, D. M. (2004). Mutations in orbit/mast reveal that the central spindle is comprised of two microtubule populations, those that initiate cleavage and those that propagate furrow ingression. J. Cell Biol. 166: 49-60. PubMed ID: 15240569
Jacobs, H., Richter, D., Venkatesh, T. and Lehner, C. (2002). Completion of mitosis requires neither fzr/rap nor fzr2, a male germline-specific Drosophila Cdh1 homolog. Curr. Biol. 12: 1435-1441. PubMed ID: PubMed ID: 12194827
Jang, J. K., Gladstein, A. C., Das, A., Shapiro, J. G., Sisco, Z. L. and McKim, K. S. (2021). Multiple pools of PP2A regulate spindle assembly, kinetochore attachments and cohesion in Drosophila oocytes. J Cell Sci 134(14). PubMed ID: 34297127
Koury, S. A. (2023). Female meiotic drive shapes the distribution of rare inversion polymorphisms in Drosophila melanogaster. Genetics. PubMed ID: 37616566
Lieberfarb, M. E., Chu, T., Wreden, C., Theurkauf, W., Gergen, J. P. and Strickland, S. (1996). Mutations that perturb poly(A)-dependent maternal mRNA activation block the initiation of development. Development 122: 579-588. PubMed ID: PubMed ID: 8625809
Mazina, O. M., et al. (2004). Saccharomyces cerevisiae Mer3 helicase stimulates 3'-5' heteroduplex extension by Rad51; implications for crossover control in meiotic recombination. Cell 117: 47-56. PubMed ID: PubMed ID: 15066281
Matsubayashi, H., Yamamoto, M. T. (2003). REC, a new member of the MCM-related protein family, is required for meiotic recombination in Drosophila. Genes Genet. Syst. 78: 363-371. PubMed ID: PubMed ID: 14676427
Megraw, T. L. and Kaufmann, T. C. (2000). The centrosome in Drosophila oocyte development. In The Centrosome in Cell Replication and Early Development (ed. R. E. Palazzo and G. P. Schatten), pp. 385-407. San Diego: Academic Press.
Merker, J. D., Dominska, M. and Petes, T. D. (2003) Patterns of heteroduplex formation associated with the initiation of meiotic recombination in the yeast Saccharomyces cerevisiae. Genetics 165: 47-63. PubMed ID: PubMed ID: 14504217
Nakagawa, T. and Ogawa, H. (1999). The Saccharomyces cerevisiae MER3 gene, encoding a novel helicase-like protein, is required for crossover control in meiosis. EMBO J. 18: 5714-5723. PubMed ID: PubMed ID: 10523314
Page, A. W. and Orr-Weaver, T. L. (1996). The Drosophila genes grauzone and cortex are necessary for proper female meiosis. J. Cell Sci. 109: 1707-1715. PubMed ID: PubMed ID: 8832393
Pearson, N. J., Cullen, C. F., Dzhindzhev, N. S. and Ohkura, H. (2005). A pre-anaphase role for a Cks/Suc1 in acentrosomal spindle formation of Drosophila female meiosis. EMBO Rep. 6(11): 1058-63. PubMed ID: PubMed ID: 16170306
Peters, J. M. (2002). The anaphase-promoting complex: proteolysis in mitosis and beyond. Mol. Cell 9: 931-943. PubMed ID: PubMed ID: 12049731
Pfleger, C. M., Lee, E., Kirschner, M. W. (2001). Substrate recognition by the Cdc20 and Cdh1 components of the anaphase-promoting complex. Genes Dev. 15(18): 2396-407. PubMed ID: 11562349
Raff, J. W., Jeffers, K. and Huang, J.-y. (2002). The roles of Fzy/Cdc20 and Fzr/Cdh1 in regulating the destruction of cyclin B in space and time. J. Cell Biol. 157: 1139-1149. PubMed ID: 12082076
Raja, S. J. and Renkawitz-Pohl, R. (2005). Replacement by Drosophila melanogaster protamines and Mst77F of histones during chromatin condensation in late spermatids and role of Sesame in the removal of these proteins from the male pronucleus. Molec. Cell. Biol. 25: 6165-6177. PubMed ID: PubMed ID: 15988027
Riparbelli, M. G. and Callaini, G. (1996). Meiotic spindle organisation in fertilized Drosophila oocyte: presence of centrosomal components in the meiotic apparatus. J. Cell Sci. 109: 911-918. PubMed ID: 8743938
Riparbelli, M. G., Callaini, G. and Glover, D. M. (2000). Failure of pronuclear migration and repeated divisions of polar body nuclei associated with MTOC defects in polo eggs of Drosophila. J. Cell Sci. 113: 3341-3350. PubMed ID: 10954431
Riparbelli, M. G., Callaini, G., Glover, D. M. and Avides, M. C. (2002). A requirement for the Abnormal spindle protein to organise microtubules of the central spindle for cytokinesis in Drosophila. J. Cell Sci. 115: 913-922. PubMed ID: 11870210
Riparbelli, M. G. and Callaini, G. (2005). The meiotic spindle of the Drosophila oocyte: the role of centrosomin and the central aster. J. Cell. Sci. 118(Pt 13): 2827-36. PubMed ID: 15976443
Swan, A., Barcelo, G. and Schupbach, T. (2005a). Drosophila Cks30A interacts with Cdk1 to target Cyclin A for destruction in the female germline. Development 132: 3669-3678. PubMed ID: PubMed ID: 16033797
<>Swan, A. and Schupbach, T. (2005b). Drosophila female meiosis and embryonic syncytial mitosis use specialized Cks and CDC20 proteins for cyclin destruction. Cell Cycle 4: 1332-1334. PubMed ID: PubMed ID: 16138012
Tanneti, N. S., Landy, K., Joyce, E. F. and McKim, K. S. (2011). A pathway for synapsis initiation during zygotene in Drosophila oocytes. Curr. Biol. 21(21): 1852-7. PubMed ID: 22036181
Terada, Y., Uetake, Y. and Kuriyama, R. (2003). Interaction of Aurora-A and centrosomin at the microtubule-nucleating site in Drosophila and mammalian cells.
J. Cell Biol. 162(5): 757-63. PubMed ID: 12939255
Vaizel-Ohayon, D. and Schejter, E. D. (1999). Mutations in centrosomin reveal requirements for centrosomal function during early Drosophila embryogenesis.
Curr. Biol. 9(16): 889-98. PubMed ID: 10469591
>Varmark, H., et al. (2007). Asterless is a centriolar protein required for centrosome function and embryo development in Drosophila. Curr. Biol. 17(20): 1735-45. PubMed ID: PubMed ID: 17935995
Wkimoto, B. T., Lindsley, D. and Herrera, C. (2004). Toward a comprehensive genetic analysis of male fertility in Drosophila melanogaster. Genetics 167: 207-216. PubMed ID: PubMed ID: 15166148
Wang, L. I., Das, A. and McKim, K. S. (2019). Sister centromere fusion during meiosis I depends on maintaining cohesins and destabilizing microtubule attachments. PLoS Genet 15(5): e1008072. PubMed ID: 31150390
Wei, Y., Bettedi, L., Ting, C. Y., Kim, K., Zhang, Y., Cai, J. and Lilly, M. A. (2019). The GATOR complex regulates an essential response to meiotic double-stranded breaks in Drosophila. Elife 8. PubMed ID: 31650955
Wilson, K. L., Fitch, K. R., Bafus, B. T. and Wakimoto, B. T. (2006). Sperm plasma membrane breakdown during Drosophila fertilization requires sneaky, an acrosomal membrane protein. Development 133(24): 4871-9. PubMed ID: PubMed ID: 17092953
Genes involved in tissue and organ development
Home page: The Interactive Fly © 1995, 1996 Thomas B. Brody, Ph.D.
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
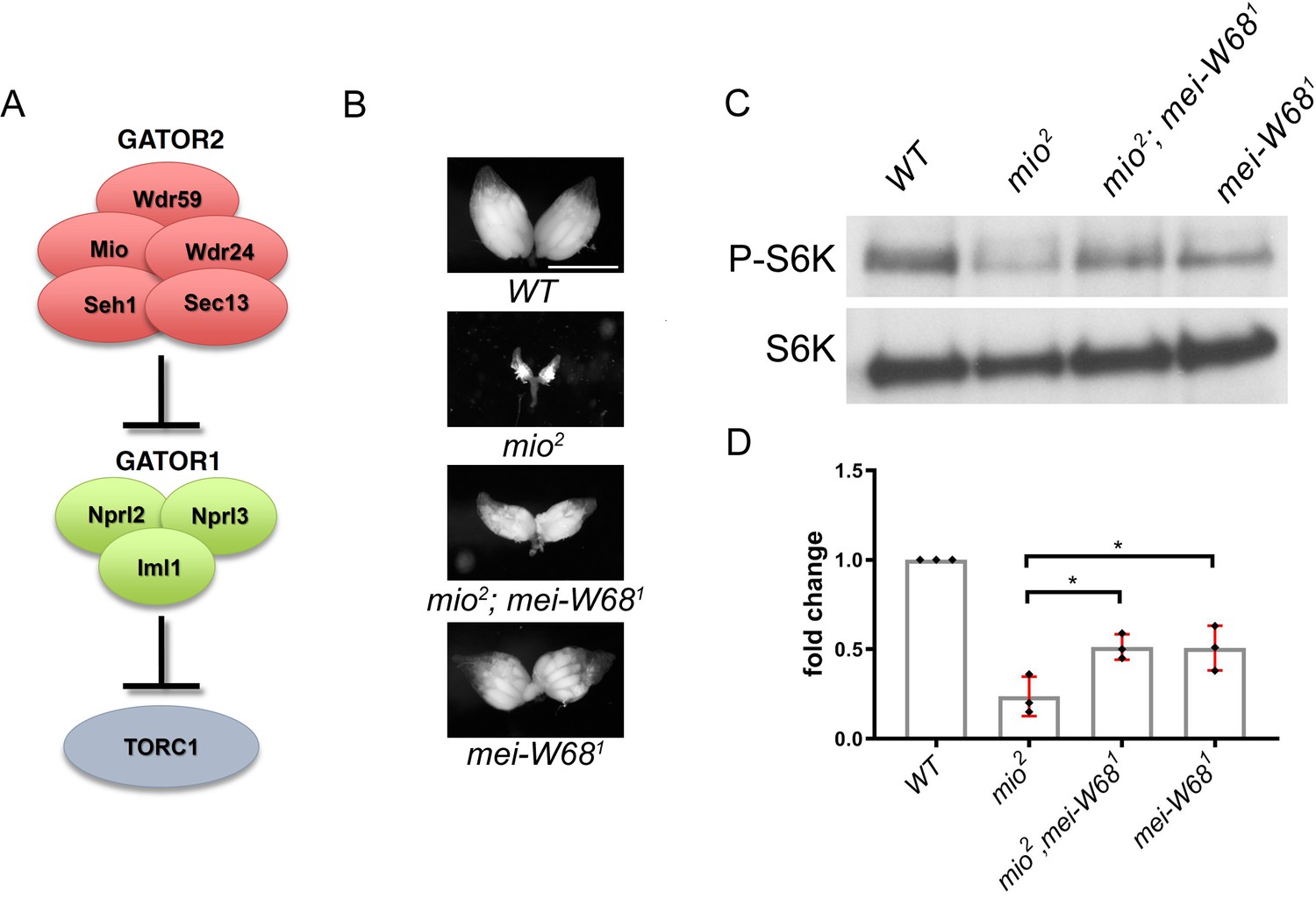{kind=link}
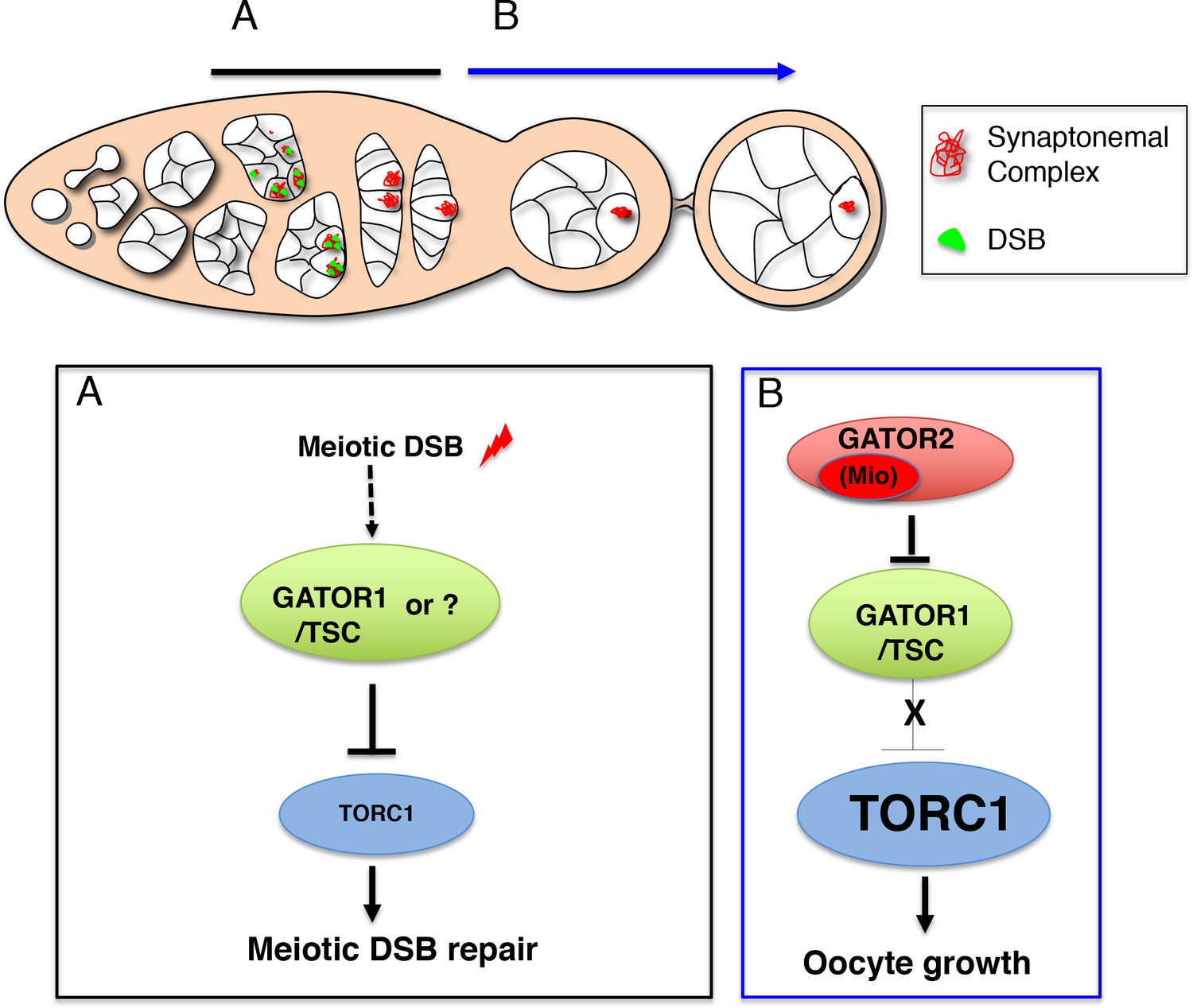{kind=link}