|
| DB Cinema Main Page
|
Dynamics of Thin Filopodia |
|
| After work published in Jeffrey Miller, Scott E. Fraser and David McClay (1995), Dynamics of thin filopodia during sea urchin gastrulation, Development 121: 2501-2511. | |
|
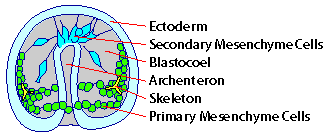
Background Information Sea urchins are favorite organisms to study gastrulation because their early embryos are transparent, and we can watch what's going on during this important phase of development. Gastrulation begins in earnest with the ingression of primary mesenchyme cells in the center of the vegetal plate. Once inside the blastocoel, they migrate by extending filopodia. After a period of migration, they organize into a ring in the vegetal half of the blastocoel; they fuse to form a syncytium that secretes the skeleton of spicules. When primary mesenchyme cells begin to migrate, remaining cells of the vegetal plate invaginate to form the archenteron (future gut). The archenteron elongates by convergent extension. While archenteron elongation is still under way, another group of mesenchyme cells at the tip of the archenteron becomes motile and extends filopodia. These are the secondary mesenchyme cells; their filopodia contact and pull the archenteron to the ectoderm of the future mouth. The dimensions of the filopodia of migrating primary mesenchyme cells and secondary mesenchyme cells pulling the tip of the archenteron are 1 �m, or more, in diameter. The authors used Nomarski differential interference contrast microscopy and electronic image enhancement to capture the following sequences. These sequences show that there are thin filopodia (0.2 - 0.4 �m in diameter) extending from, and to, primary mesenchyme cells, ectodermal cells, and secondary mesenchyme cells. These thin filopodia don't seem to be used for locomotion. Rather, they appear to assess their environment like the thin filopodia seen at the tips of axon growth cones. |
|
Click on the "thumbnail" image or the QuickTime link to watch a sequence. You need QuickTime to watch these sequences; all sequences are silent. |
|
|
| DB Cinema Main Page | ||
|
|
||
| Labeled diagram of
sea urchin embryo at midgastrula stage for reference when you view sequences. After Fig. 1 in Miller, J., et al., Development 121: 2501-2511 (1995). |
||
|
Images collected at 1 frame/second. GENERAL FILOPODIAL DYNAMICS |
||
 |
Primary mesenchyme cells in subequatorial
ring send thin filopodia along the basal surface of ectodermal cells.
QuickTime (359 K) |
|
|
FILOPODIAL GROWTH |
||
 |
Primary mesenchyme cells in subequatorial
ring extending filopodia. A middle primary mesenchyme cell extends a filopod
which kinks several times then appears to begin retracting.
QuickTime (495 K) |
|
 |
Primary mesenchyme cell in subequatorial
ring extending a filopod.
QuickTime (258 K) |
|
 |
Primary mesenchyme cell extending
a thin filopod. Notice the membrane flow which moves toward the cell body.
A migrating pigment cell displays blebbling behavior (lower left).
QuickTime (413 K) |
|
|
FILOPODIAL RETRACTION |
||
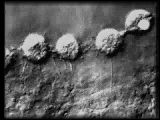 |
Primary mesenchyme cell in subequatorial
ring resorbs a thin filopod. Notice how the filopod kinks just prior to
and during retraction.
QuickTime (259 K) |
|
 |
Filopod of a primary mesenchyme
cell retracting.
QuickTime (287 K) |
|
|
CELL-CELL INTERACTIONS MEDIATED BY THIN FILOPODIA PRIMARY MESENCHYME CELL MIGRATION |
||
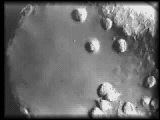 |
Primary mesenchyme cells during
their migratory phase extend small, short-lived filopodia that may contact
the ectoderm or other primary mesenchyme cells as they assemble into the
subequatorial ring.
QuickTime (527 K) |
|
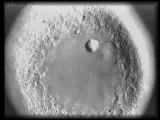 |
Migrating primary mesenchyme
cell sends out two filopodia that contact ectodermal cells. Primary mesenchyme
cells may use filopodia to gather positional information that will determine
the site(s) of spiculogenesis.
QuickTime (645 K) |
|
|
PRIMARY MESENCHYME CELL-ECTODERM INTERACTIONS |
||
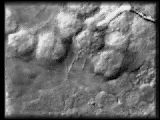 |
Primary mesenchyme cell-ectoderm
interactions occur after the subequatorial ring forms, during the time the
spicules grow. The filopodia may enable primary mesenchyme cells to survey
their environment, gathering positional information that specifies skeletal
morphology.
QuickTime (699 K) |
|
 |
More primary mesenchyme cell-ectoderm
interactions. This sequence also shows an ectodermal cell extending a short,
stubby filopod (lower center). This behavior can also be seen in some of
the other sequences.
QuickTime (391 K) |
|
 |
A very long primary mesenchyme
cell filopod contacting and ectodermal cell at the animal pole. The amazing
length of the thin filopod allows a primary mesenchyme cell to interact
with cells at any other position within the embryo.
QuickTime (521 K) |
|
|
SECONDARY MESENCHYME CELL-ECTODERM INTERACTIONS |
||
 |
A secondary mesenchyme cell extends
a filopod that rapidly becomes a lamellapod. Secondary mesenchyme cells
display a much wider range of filopod/lamellopod morphologies than primary
mesenchyme cells.
QuickTime (402 K) |
|
|
PRIMARY MESENCHYME CELL-SECONDARY MESENCHYME CELL INTERACTIONS |
||
 |
A primary mesenchyme cell extends
a filopod that contacts a secondary mesenchyme cell, bends and then retracts.
QuickTime (468 K) |
|
 |
Extensive cell-cell interactions
between secondary mesenchyme cells (to the left) and primary mesenchyme
cells (to the right) occur throughout gastrulation. These direct interactions
may be used to communicate cell fate information that suppresses the ability
of secondary mesenchyme cells to convert to primary mesenchyme cell fate.
QuickTime (1.6 MB) |
|
Large QuickTime
Movies of "Dynamics of Thin Filopodia" for classroom use. Coming
soon!
|
| We welcome your comments and suggestions for the Developmental Biology Cinema. |
 SOCIETY
FOR DEVELOPMENTAL BIOLOGY SOCIETY
FOR DEVELOPMENTAL BIOLOGY
SDB Webmaster |